Embed Size (px)
Citation preview

--~~-----------------------------~~
Freisetzung eines Botenstoffes der Entziindung(TNF-a) aus Lebermakrophagen in Perfusions-Zellkultur
Thomas Hartung, Stephan Hermening, Achim Sauer und AlbrechtWendelBiochemische Pharmakologie, Universitat Konstanz
ZusammenfassungTumor Nekrose Faktor-a ist ein zentraler Botenstoff derEntziindung, d.h. das Protein wird von Zellen desImmunsystemes bei Infektionen, Yerletrungen oderAutoimmunerkrankungen freigesetzt und unterhdlt bzw.verstdrkt den Entziindungsvorgang. Die Messung solcherBotenstoffe im Versuchstier bei experimentellen Schddi-gungen ist eine Methode, um die Wirksamkeit vonmoglichen Pharmaka zu testen. Versuche, alternativeModelle der Freisetzung solcher Botenstoffe aus Zellkul-turen zu. verwenden, sind dadurch limitiert, daft erhebli-che Unterschiede im Zeitverlauf der Bildung zwischenVersuchstier und der Kultur bestehen.Es wird die These aufgestellt, dafJ dies im wesentlichenauf die fehlende Elimination der einmal freigesetztenBotenstoffe zuruckzufuhren ist. Untersucht wurde dasModell der Bakterienendotoxin-induzierten Freisetzungvon Tumor-Nekrose-Faktor-a aus Lebermakrophagender Ratte. Die bekannte Diskrepan: zwischen derFreisetzung in vivo, hier der Ratte im Endotoxin-Schock,und der Produktion unter iiblichen Kulturbedingungenaus den wichtigsten Produzenten, den Kupfferzellen derRatte, wurde gezeigt. Durch Ubergang von der stati-schen Inkubation zur Perfusionsrellkultur wurde eineElimination in dem Modell erzeugt, d.h. ein stdndigerErsatz des Zelliiberstandes durchfrisches Medium.Tatsachlicb ergab die Modellierung einer Elimination inder Zellkultur durch kontinuierliche Perfusion einen derin-vivo-Situation ahnlichen Verlauf der Freisetzung desMediators. Damit erlaubt die Perfusionskultur eineAnndherung an die in vivo Situation, die die Relevanzder Zellkultur fur die Testung von pharmakologischenWirkstoffen erhoht.
Summary: The formation of a mediator of inflammation(TNF-a) by rat liver macrophages in perfusion culture
Tumor necrosis factor-a (TNF-a) represents a centraldistal mediator of inflammation. It is a protein releasedupon infections, traumatic lesions or autoimmunedisorders from immunocompetent cells and thus main-tains or enhances the inflammatory reaction. The deter-mination of such mediators in experimentally challengedanimals is a mean for testing the efficacy of putativedrugs. Alternative attempts for the assessment of media-tor release from cell cultures are limited by profounddifferences in the time course of mediator release invitro.We checked the hypothesis that these kinetic differencesare due to the lack of elimination of mediators formedand released in vitro. We used the release ofTNF-afrom liver macrophage cultures stimulated with thebacterial cell wall component endotoxin as a model. Thediscrepancy between the in vivo release of the cytokineduring endotoxic shock in the rat and the in vitro releasefrom Kupffer cells was confirmed. By using a continuousopen perfusion system instead of a static culture, thesimulation of an elimination resulted in a mediatorrelease that closely resembled the kinetics seen in vivo.Perfusion cultures appear to be suitable for relevant invitro screening models in drug development and testing.
Keywords: endotoxin, Kupffer cells, cytokines, kinetics,in vitro pharmacology
Die heutige Entziindungsforschungkonzentriert sich auf die Bildungkorpereigener Mediatoren als Ver-mittler und Verstarker der durch
Verletzung, Infektion oder Autoim-munerkrankungen ausgelosten Pri-rnarschadigung (Gallin et al., 1992).Ein zentraler Ansatz zum Studiumder Bildung dieser Botenstoffe be-steht in einer experimentellen Scha-
digung des Versuchstieres und an-schliefsender Messung der Freiset-zung von Mediatoren (Neugebauer etal., 1993; Beutler, 1993). Aufgrundseiner Kornplexitat ist das Netzwerkaus Zytokinen, Lipidmediatoren und
1 Elnleltung
ALTEX 13, 1/96 17

HARTUNG ET AL. .r;-> ~------------------------------~~~--~
vielen anderen Mediatoren nochweitgehend unverstanden. Als re-duktionistischer Ansatz sind dabeiZellkulturexperimente hilfreich, indenen die Bildung solcher Substan-zen z.B. auf bakterielle Stimuli hindurch Leukozyten untersucht werdenkann. Gegenuber dem Tierversuchhaben diese in vitro Methoden je-doch auch Nachteile. Sowohl dieStimuli als auch die daraufhin freige-setzten Mediatoren sind dauerhaft inder Zellkultur prasent und wirkenweiter auf die Leukozyten, wahrendsie im Versuchstier einer standigenElimination unterliegen und nur kurzihre Wirkung entfalten konnen, DiePharmakologie kennt den erhebli-chen Unterschied zwischen einerkurzzeitigen Gabe (Bolus) und derDauerinfusion von Wirkstoffen. Tat-sachlich unterscheidet sich auch dieMediatorbildung in vitro deutlichvon der in vivo:
• Durch die dauerhafte Anwesenheiteines Stimulus kommt es im allge-meinen zu grofseren freigesetztenMediatormengen pro Zelle.
• Statt einem schnell erreichten Ma-ximum ("peak") im Versuchstierfindet man in vitro meist einelangfristige Produktion, die ofterst durch das Absterben der Leu-kozyten in der Kultur endet.
• Die Palette freigesetzter Mediato-ren in vitro ist meist breiter, dajedes Zellprodukt ruckwirken undseinerseits weitere Faktoren frei-setzen kann.
Aus diesen Grunden kann die Zell-kulltur heute den Tierversuch zumStudium der Bildung von Botenstof-fen der Entzundung und ihrer Modu-lation durch pharmakologische MaB-nahmen nicht ersetzen.Der Einsatz der Perfusionszellkul-
tur stellt eine Moglichkeit dar, eineAngleichung an die in vivo Situationvorzunehmen. Seit wenigen Jahrenist ein ZeIlkultursystem standardi-siert verfUgbar (Minusheet -Technik),das die Kultur yon beliebigen Zellenim DurchtluB ermoglicht. Anders alsunter den ublichen statischen Kultur-bedingungen fuhrt hier die kontinu-
18
ierliche Entfernung aller freigesetz-ten Zellprodukte aus der Kultur zuVerhaltnissen, die eher der in vivoSituation entsprechen. Der Vorteildieser Kulturmethode fur die Mor-phologie yon Zellen ist vielfach be-legt (Minuth et al., 1992; Aigner etaI., 1994; Gebhardt et aI., 1979;Jauregui et aI., 1994; Farghali et al.,1994).Aus den genannten Uberlegungen
heraus wurde deshalb diese Technikauf eine stimulierbare Leistung derZellen, namlich die Freisetzung vonEntzundungsmediaroren, angewen-det.Die Erwartungshaltung besteht
darin, durch eine Annaherung derBedingungen der in vitro Freisetzungvon Entziindungsmediatoren an diein vivo Situation einen erheblichenTeil dieser Phanomene in vitro stu-dierbar zu machen. Gerade die tier-experimentellen Modelle der Ent-ziindungsforschung sind fur Ver-suchstiere sehr belastend. Die aufdem Weg der Perfusionskultur er-wartete Aufwertung der ZeUkultur-methodik soli hier einen teilweisenUbergang zu in vitro Methoden er-moglichen, und damit eine Voraus-wahl von Wirkstoffen in der phar-makologischen Forschung bereitsvor dem Tierversuch moglich ma-chen.
2 Problemstellung und Auswahlyon TNF -o als Me8parameter
Als Modell wurde die Endotoxin-induzierbare Freisetzung von TumorNekrose Faktor-o; (TNF-a) aus Le-bermakrophagen (Kupffersche Zel-len) der Ratte gewahlt (siehe FotoTitelseite): Endotoxine sind derHauptzell wandbestandteil gramne-gativer Bakterien. Endotoxine sindLipopolysaccharide (LPS) der aulie-ren Zellmembran. LPS ist ein zentra-ler Ausloser der Akut-Phase-Reakti-on und fuhrt zur Aktivierung korper-eigener Funktionen (Komplement-,Gerinnungs- und Kininsystem). Esstimuliert Monozyten und Makro-phagen zur Freisetzung endogenerPyrogene (Interleukin 1, Interleukin
6, TNF-a), die eine Fieberreaktionverrnitteln. Hohe LPS-Dosen losenim Versuchstier durch eine Uberakti-vierung des unspezifischen Immun-systems den Endotoxin-Schock aus,der ein weit verbreiteter Tierversuchist. Dabei handelt es sich urn eineGanzkorper-Entziindungsreaktion,die die grundlegende Symptomatikdes Septischen Schocks beim Patien-ten widerspiegelt.Durch die experimentelle Auslo-
sung dieses Schadens werden Ma-krophagen aktiviert, denen als ZelJender unspezifischen Abwehr eineSchliisselrolle zukommt. Kupfferzel-len der Leber stellen mit 80% diegrobte Population aller sefshaftenMakrophagen im Organismus (La-skin, 1990) und machen 30% dernichtparenchymalen Zellen (NPC) inder Leber aus. Die Lebermakropha-gen bilden eine protektive Barrierefur die systemische Zirkulation, in-dem sie potentielle Noxen (Endoto-xine, Mikroorganismen, Immunkom-plexe, Tumorzellen) beseitigen, diedie Leber uber das Pfortaderbluterreichen. Nach Stimulation sezer-nieren Kupfferzellen eine Reihe bio-aktiver Mediatoren, zum einen vonder Arachidonsaure abstammendeLipidmediatoren (Prostaglandine,Leukotriene), zum anderen Zytokine(Interleukin l,Interleukin 6,Interleu-kin 12, TNF-a). Diese Faktoren sindwichtige Regulatoren der Immunant-wort.TNF-a ist einer der zentralen Me-
diatoren der Entzundung. Er ist einPolypeptid mit einem Molekularge-wicht yon 17 kD und wird haupt-sachlich yon Monozyten und Makro-phagen, aber auch von T-Lymphozy-ten, Neutrophilen und NK-Zellen ge-bildet (Tracy et al., 1989). EineReihe yon Effekten von LPS lassensich durch die Gabe von TNF simu-lieren (Beutler et al., 1985a; Tiegs etaI., 1989) und durch Antikorper ge-gen TNF-a hemmen (Beutler et al.,1985b; Tiegs et aI., 1990). Gerade imModell des Endotoxin-Schocks wur-de die zentraJe Rolle yon TNF alsterminalem Mediator wiederholt ge-zeigt.
ALTEX 13, 1/96

wr:"'\ HARTUNG ET AL.--~~~------------------------------¥C~
3 Versuchstiere, Material undMethoden
3.1 VersuchstiereFur die Experimente wurden mannli-che Fischer-Ratten (F344, CharlesRiver, Sulzfelden) mit einem Ge-wicht von 200-300 g verwendet. DieHaltung erfolgte in der Tierfor-schungsanlage Konstanz fur minde-stens eine Woche. Die Tiere hattenfreien Zugang zu Futter und Wasserund wurden unter natiirlichen Licht-verhaltnissen (l2h Tag/Nacht-Rhythmus, lOO Lux) bei 55% Luft-feuchtigkeit gehalten.
3.2 TNF-Freisetzung in Endoto-xin-behandelten Ratten in vivoLPS (von Salmonella abortus equi,Sigma, Deisenhofen) wurde in PBSgelost und den Ratten iiber dieSchwanzvene i.v. injiziert (50mg/kg). Nach verschiedenen Zeiten wur-de den Tieren Schwanzblut durchInzision zur Bestimmung yon TNFabgenommen. In dem Versuch wur-den zwei Gruppen von je drei Tierenverwendet, denen abwechselnd Se-rumproben entnommen wurden.Es handelt sich bei dies em Ver-
such um ein mehrere Jahre zuruck-liegendes, nicht publiziertes Experi-ment. Zu dies em Zeitpunkt war einRatten-TNF-Elisa noch nicht verfug-bar, deshalb erfolgte die Messungseinerzeit im Bioassay. Es wurde aufeine Wiederholung verzichtet, da ei-nerseits eine gute Korrelation vonBioassay und ELISA bei den in vitroExperimenten bestand und anderer-seits der Vergleich der Kinetik keinequantitativen Werte erfordert.
3.3 KupfferzellpraparationDie Praparation der Rattenleberzel-len wurde nach der Methode vonSeglen (1976), modifiziert nach Har-tung (1991), durchgefiihrt. Die Leberwurde dabei in zwei Schritten erstmit EGT A-haltiger Salzlosung unddann mit einer Kollagenase-Losungrezirkulierend perfundiert. Die nachAusschutteln gewonnene Primarzell-suspension wurde mittels Gaze (100urn PorengroBe, Eckert, Waldkirch/Brsg.) grob gereinigt, und die ver-
ALTEX \3, \/96
schiedenen Zellpopulationen wurdendurch Differentialzentrifugation ge-trennt. Die nichtparenchymalen Zel-len (NPC) wurden durch Zentrifuga-tion (100 g, 3 min) yon den Hepato-zyten getrennt. Urn die Zellen vonZelldebris zu trennen, schlossen sichvier Zentrifugationsschritte (400 g, 7min.) an. Fur die Versuche wurdenNPC mit einer Vitalitat >95% (Try-pan-Blau-Ausschluss) verwendet.Die ZelIen wurden bei 37DC, 20% O2und 5% CO2 kulti viert.
3.4 Kollagenbeschichtung desTragermaterialsMinuthsheets (Minucells and Minu-tissue, Bad Abbach) wurden mitGlasdeckglaschen bestuckt (Abb.la)und 10 Minuten in 70% Ethanolgelegt. Nach Uberfuhren der Sheetsin Greiner-24well-Kulturplattenfolgten zwei Waschschritte mit steri-lem PBS (jeweils 10 Minuten schut-
a) Aufbau eines Minusheet
302®10~~~
§~~.•• 5
b) Perfusionskultur in der Minuthkammer untereinfachen experimentellen Bedingungen(Peristaltikpumpe. Heizplatte)
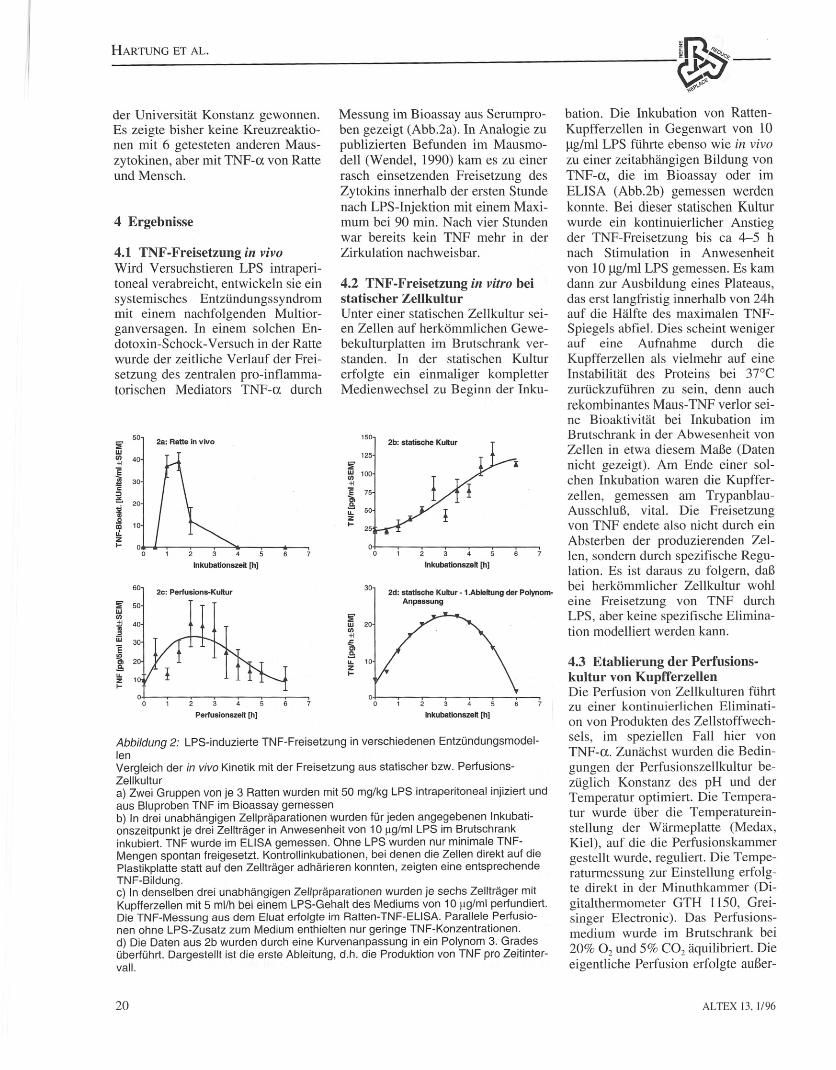
Abbildung 1: Zusammensetzen der Zell-Trager •.Minusheets" (a) und Aufbau derPerfusionszellkultur (b)Die Abbildung zeigt das Zusammenset-zen eines Zellkulturtraqers (Minusheet):Es wurden Deckqlaschen von 13 mmDurchmesser verwendet, die sparer mitKollagen beschichtet wurden. DieRatten-Lebermakrophagen wurden aufdie Trager aufgebracht. nach Adharenzin eine Medium-gefUlite Perfusionskam-mer OberfOhrtund unter kontinuierlicherDurchspGlung perfundiert.
teln). Nach dem Waschen wurdendie Vertiefungen mit 500 III Kolla-gen (0,2 mg/ml Kollagen in 0,1%Essigsaure) gefullt und fur minde-stens zwei Tage bei 37DC ruhengelassen. Die Sheets wurden an-schlieBend einmal mit EDT A-halti-ger HANKS-Salzlosung und dreimalmit reiner HANKS-Salzlosung ge-spult (pro Waschgang zwei Minutenschiitteln). Bis zur endgiiltigen Ver-wendung lieBen sich die Minuths-heets lichtgeschiitzt bei 4DCJagern.
3.5 Inkubation der KupfferzellenIn die Vertiefungen einer 24weIl-Kulturplatte kamen jeweils ein Mi-nuthsheet und 500 !1l Zellsuspensionmit 2106 NPC/ml. Die Zellen wur-den iiber Nacht vorkultiviert undanschlieBend in bereits mit vorge-warmtem Medium (HEPES-gepuf-fert, 37DC) gefi.illte Minuth-Kam-mern (Minucells and Minutissue, BadAbbach) uberfuhrt (pro Kammer 6Sheets, Abb.l b). Die Perfusion erfolg-te durch eine Mehrkanalschlauchpum-pe (Peristaltikpumpe IPC-8, IsmatecLaboratoriumstechnik, Wertheim-Mohnfeld). Die Perfusionsgeschwin-digkeit betrug 5 mllh. Alle 30 minwurden Aliquots entnommen und biszur Messung bei -70°C weggefroren.Parallel wurden entsprechend mit Zel-len besiedelte Minusheets in eine neue24well-Kulturplatte mit 500,ul Medi-um uberfuhrt (statische Kulturbedin-gungen).
3.6 TNF-BestimmungDie TNF-Bestimmung erfolgte ent-weder im kommerziellen Ratten-TNF-ELISA (Biosource, Laboserv,Marburg) oder im TNF-Bioassay.Dieser wurde mit der TNF-sensitivenFibrosarkom-Zellinie WEHI 164Klon 13 nach der Methode vonEspevik und Nissen-Meyer (1986)durchgefiihrt. Ais Standard dientehier rekombinantes Maus- TNF-a,ein Geschenk yon Dr. A. Adolf,Bender & Co., Wien. Die Spezifitatder Messung wurde mit einem kreuz-reagierenden Schaf-anti-Maus- TNF-a-Antiserum gezeigt. Das Antiserumwurde durch Immunisierung einesSchafes in der Tierforschungsanlage
19
H__A_R_TU_N_G_E_T_A_L_. ~~----
¥v~
50 2a: Ratte in vivo150
!i 2b: statische Ku~u,W 125en 40-H !:EE w 100U! 30 en.., -H
" i 75~:!!.20~
'"U. 50 t.2 z
m 10 l-Ii.zI-
2 3 4 5
InkubalionszeH [hI InkubalionszeH [hI
60 2c: Perfusions-KuRur 302d: slalische Ku~u, - 1.AbleHung de, Polynom-
!:E 50Anpassung
wIII !:E;!j 40 W 20.. en::riii
-H
E ~'g, oSoSu.ZI-
00 2 3 4 5
PerfusionszeH [hI InkubalionszeH [hI
der Universitat Konstanz gewonnen.Es zeigte bisher keine Kreuzreaktio-nen mit 6 getesteten anderen Maus-zytokinen, aber mit TNF-a von Ratteund Mensch.
4 Ergebnisse
4.1 TNF-Freisetzung in vivoWird Versuchstieren LPS intraperi-toneal verabreicht, entwickeln sie einsystemisches Entzundungssyndrommit einem nachfolgenden Multi or-ganversagen. In einem soJchen En-dotoxin-Schock- Versuch in der Rattewurde der zeitliche Verlauf der Frei-setzung des zentralen pro-inflamma-torischen Mediators TNF-a durch
Messung im Bioassay aus Serumpro-ben gezeigt (Abb.2a). In Analogie zupublizierten Befunden im Mausmo-dell (Wendel, 1990) kam es zu einerrasch einsetzenden Freisetzung desZytokins innerhalb der ersten Stundenach LPS-Injektion mit einem Maxi-mum bei 90 min. Nach vier Stundenwar bereits kein TNF mehr in derZirkulation nachweisbar.
4.2 TNF-Freisetzung in vitro beistatischer ZellkulturUnter einer statischen Zellkultur sei-en Zellen auf herkommlichen Gewe-bekulturplatten im Brutschrank ver-standen. In der statischen Kulturerfolgte ein einmaliger kompletterMedienwechsel zu Beginn der Inku-
Abbildung 2: LPS-induzierte TNF-Freisetzung in verschiedenen Entzundunqsrnodel-lenVergleich der in vivo Kinetik mit der Freisetzung aus statischer bzw. Perfusions-Zellkultura) Zwei Gruppen von je 3 Ratten wurden mit 50 mg/kg LPS intraperitoneal injiziert undaus Bluproben TNF im Bioassay gemessenb) In drei unabhanqiqen Zellpraparationen wurden fUr jeden angegebenen Inkubati-onszeitpunkt je drei Zelltraqer in Anwesenheit von 10 f.lg/ml LPS im Brutschrankinkubiert. TNF wurde im ELISA gemessen. Ohne LPS wurden nur minimale TNF-Mengen spontan freigesetzt. Kontrollinkubationen, bei denen die Zellen direkt auf diePlastikplatte statt auf den Zelltraqer adharieren konnten, zeigten eine entsprechendeTNF-Bildung.c) In denselben drei unabhangigen Zeupraparationen wurden je sechs Zeutraqer mitKupfferzellen mit 5 mllh bei einem LPS-Gehalt des Mediums von 10 J-lg/ml perfundiert.Die TNF-Messung aus dem Eluat erfolgte im Ratten-TNF-ELISA. Parallele Perfusio-nen ohne LPS-Zusatz zum Medium enthielten nur geringe TNF-Konzentrationen.d) Die Daten aus 2b wurden durch eine Kurvenanpassung in ein Polynom 3. Gradesubertuhrt. Dargestellt ist die erste Ableitung, d.h. die Produktion von TNF pro Zeitinter-vall.
20
bation. Die Inkubation von Ratten-Kupfferzellen in Gegenwart von 10ug/ml LPS fuhrte ebenso wie in vivozu einer zeitabhangigen Bildung vonTNF-a, die im Bioassay oder imELISA (Abb.2b) gemessen werdenkonnte. Bei dieser statischen Kulturwurde ein kontinuierlicher Anstiegder TNF-Freisetzung bis ca 4-5 hnach Stimulation in Anwesenheitvon 10 ug/rnl LPS gemessen. Es kamdann zur Ausbildung eines Plateaus,das erst langfristig innerhalb von 24hauf die Halfte des maximal en TNF-Spiegels abfiel. Dies scheint wenigerauf eine Aufnahme durch dieKupfferzellen als vielmehr auf eineInstabilitat des Proteins bei 37°Czuruckzufuhren zu sein, denn auchrekombinantes Maus- TNF verlor sei-ne Bioaktivitat bei Inkubation imBrutschrank in der Abwesenheit vonZellen in etwa diesem MaJ3e (Datennicht gezeigt). Am Ende einer sol-chen Inkubation waren die Kupffer-zellen, gemessen am Trypanblau-AusschluB, vital. Die Freisetzungyon TNF endete also nicht durch einAbsterben der produzierenden ZeJ-len, sondern durch spezifische Regu-lation. Es ist daraus zu folgern, daBbei herkommlicher Zellkultur wohleine Freisetzung von TNF durchLPS, aber keine spezifische Elimina-tion modelliert werden kann.
4.3 Etablierung der Perfusions-kultur yon KupfferzellenDie Perfusion von Zellkulturen fiihrtzu einer kontinuierlichen Eliminati-on von Produkten des Zellstoffwech-sels, im speziellen Fall hier vonTNF-a. Zunachst wurden die Bedin-gungen der Perfusionszellkultur be-ziiglich Konstanz des pH und derTemperatur optimiert. Die Tempera-tur wurde uber die Temperaturein-stellung der Warrneplatte (Medax,Kiel), auf die die Perfusionskammergestellt wurde, reguliert. Die Tempe-raturmessung zur Einstellung erfolg-te direkt in der Minuthkammer (Di-gitalthermometer GTH 1150, Grei-singer Electronic). Das Perfusions-medium wurde im Brutschrank bei20% O2 und 5% CO2 aquilibriert. Dieeigentliche Perfusion erfolgte auBer-
ALTEX 13, 1I96

wl-"\ HARTUNG ET AL.--~~~---------------------------~~~
halb des Brutschranks, d.h. unterUmgebungsluft mit niedrigeremCO2-Gehalt. Unter diesen Bedingun-gen erwies sich dieser in statischenSystemen eingesetzte Bicarbonat-Puffer als unzureichend, der pH ver-schob sich in den alkalischen Be-reich (pH 8,4) durch Entweichen yonCO2 aus dem an sich geschlossenenPerfusionssystem. Durch den Einsatzyon HEPES gepufferten RPMI 1640Medium (Biochrom) konnte del' pHiiber den erforderlichen Zeitraumyon sechs Stunden bei pH 7,4 ken-stant gehalten werden.Das Kammervolumen betrug ca. 3
m!. DiePerfusionsgeschwindigkeitwurde auf 5 rnl/h festgesetzt, urneine ausreichende Durchspiilung undden damit verbundenen Stoffaus-tausch zu gewahrleisten, Sie darfgleichzeitig nicht zu groB gewahltwerden, darnit die zu messendenBotenstoffe (hier TNF) nicht zu starkverdiinnt werden und sich damit del'Messung entziehen.
4.4 TNF-Freisetzung aus Kupffer-zellen in PerfusionszellkulturenUrn die LPS-induzierte TNF-Freiset-zung aus Kupfferzellen unter in vivoahnlichen Voraussetzungen zu stu-dieren, wurden die Zellen kontinu-ierlich mit frischem Medium perfun-diert und das sezernierte TNF konti-nuierlich aus den Kulturen entfernt.Es wurden die Kinetiken der LPS-induzierten TNF-Freisetzung in Per-fusionskultur und statischer Zellkul-tur iiber einen Zeitraum yon 6 Stun-den verglichen. LPS war iiber dengesamten Versuchszeitraum demMedium zugesetzt, d.h. auch daskontinuierlich eingespulte frischeMedium enthielt 10 ug/rnl LPS.In den perfundierten Zellkulturen
kam es nach einer anfanglichen Er-hohung del' TNF-Freisetzung mit ei-nem Maximum bei zwei Stunden zueiner Reduktion, und die TNF-Frei-setzung fiel nach sechs Stunden aufden Kontrollwert ab (Abb. 2c). Da-mit kommt die Kinetik der TNF-Freisetzung eher den in vivo Verhalt-nissen nahe als dem unter statischenBedingungen in vitro erhaltenenVeri auf.
ALTEX 13, 1/96
5 Diskussion
Schwerpunkt der Entziindungsfor-schung del' letzten 20 Jahre war dieUntersuchung yon sogenannten Me-diatoren, d.h. korpereigenen Signal-stoffen, die den Entziindungsvor-gang steuern. Nachdern zunachst dieEicosanoide (Thromboxan, Leu-kotriene, Prostacyclin und die Pros-taglandine) im Zentrum des Interes-ses standen, sind es heute vor allemProteine wie die Zytokine (Interleu-kine, Interferone, TNF-a, Wachs-tumsfaktoren). Das Gebiet hat sichjedoch extrem diversifiziert; insge-sarnt sind uber 100 Substanzen heuteGegenstand der Forschung (darunterreaktiver Sauerstoff, Stickoxid, Pro-teasen, Plattchen-aktivierender-Fak-tor, Komplementfaktoren, etc.).Praktisch jeder dieser Mediatorenwird als rnoglicher pharmakologi-scher Eingriffspunkt mit entspre-chenden Synthese-Inhibitoren, Ant-agonisten oder Agonisten diskutiertund untersucht. Der Zugang ist dabeiim wesentlichen tierexperimentell,d.h. eine dem klinischen Bild ahneln-de Schadigung wird dem Versuchs-tier beigebracht und die Bildungdiesel' Botenstoffe wird studiert (Bei-spiele fiir TNF: Ha et al., 1983,1987; Kiener et al., ]988; Fong et aI.,1989; Ertel et al., 1991; Smith et a!.,1993). Danach werden die Testsub-stan zen im Tierversuch sowohl aufdie Hemmung der Mediatorbildung/-wirkung als auch auf das Endergeb-nis des Tierversuchs hin untersucht(Schwellung, Entziindung, Letalitat,etc.; Beispiele fiir TNF: Badger etal., 1989; Schade et al., 1989; Har-tung et al., 1992).Im allgemeinen sind die Zellen, die
den jeweiligen Mediator im Patien-ten oder im Versuchstier bilden,bekannt. Dennoch kann man wegeneiniger grundlegender Schwierigkei-ten wie dem Fehlen einer Eliminati-on nur naherungsweise die Freiset-zung in vitro studieren. Die Ergeb-nisse in Zellkultur unterscheiden sichnach Menge, Kinetik und Muster derfreigesetzten Mediatoren erheblichvorn Versuchtier. Hier wurde amBeispiel yon TNF gezeigt, daB sich
tatsachlich die Freisetzung im Ver-suchstier (Abb.2a) und aus den furdie Produktion hauptsachlich verant-wortlichen Kupfferzellen der Leberwesentlich unterscheidet. Ein ober-flachlicher Vergleich zeigt, daf derhochste Spiegel (peak) in vitro erstmehrere Stunden sparer beobachtetwird. In vivo unterliegt TNF jedocheiner raschen Elimination: 1m Nage-tier wurden Eliminationshalbwert-zeiten yon II bis 27 Minuten gemes-sen (Waage, 1987; Flick et aI.,1985). Es wurde die These verfolgt,daf sich die Unterschiede zwischender Kinetik der TNF-Bildung in vivound in vitro nicht auf eine Insuffizi-enz del' Zellkultur, sondern lediglichauf den Mangel einer Eliminationzuriickfiihren laBt. Dazu wurde einekontinuierliche Elimination des ge-messenen TNF durch Perfusion derKultur in dem Modell erzeugt. Diegewahlte willkiirliche Elirninations-halbwertzeit (Fluf 5 ml/h bei 3 mlKammervolumen) betrug 36 min.Bereits diese grobe Annaherung andie Situation im Tier fuhrte zu einerKinetik, die derjenigen im Versuchs-tier weitgehend ahnelt. Bei der Wahlder FluBrate handelt es sich um einenKompromiB aus den in vivo beob-achtbaren Eliminationsraten und derunvermeidbaren Verdiinnung des zumessenden TNF. Die grofste Streu-ung der TNF-Messung in Perfusions-kultur verglichen mit der statischenKultur hangt u.a. damit zusammen,daB bereits an der Nachweisgrenzedes ELiSAs gearbeitet werden muB-te. Das Maximum der TNF-Produk-tion lag hier bei 2 h, wahrend es imVersuchstier bei 1.5 h lag. Der ver-bleibende Unterschied konnte auf diegeringere Temperatur (37°C imBrutschrank im Vergleich zu 40-42°C in der Ratte) zuriickzufiihrensein.Diese Annaherung an die in vivo
Verhaltnisse ist kaurn durch die Be-seitigung autokrin stimulierenderFaktoren der Kupfferzellen erklar-bar, sondern resultiert wahrschein-lich aus den unterschiedlichen Ver-fahren der Probengewinnung: In derherkornrnlichen statischen Kultur ak-kumuliert das gebildete TNF: Es
21

HARTUNG ET AL. ~r;)~---------------------------h~'--
~c~..~.wird deshalb zu jedem Zeitpunktnicht die aktuelle Produktion, son-dern das Integral der bisherigen Bil-dung gemessen. Urn dies zu veran-schaulichen, wurde in Abb. 2d eineKurvenanpassung fur die TNF-Bil-dung unter statischen Kulturbedin-gungen vorgenommen (Polynom 3.Grades). Dargestellt ist die entspre-chende erste Ableitung, die die je-weilige Neuproduktion pro Zeitinter-vall darstellt. Auch auf diese Weiseerhalt man eine Produktion, die derin vivo Situation angenahert ist. AI-lerdings Iiegt das Maximum del' Pro-duktion jetzt erst bei 3 h. Ob dies aufeine unphysiologische Ruckwirkunggebildeter, stimulierender Produkte(wie TNF selbst) zuruckzufuhren ist,bleibt zu klaren.Die Ergebnisse zeigen aber, daB
bezuglich der TNF-Freisetzung zwi-schen beiden Zellkultursystemen tat-sachlich nur geringe Unterschiedebestehen. Auch die pro Zelle freige-setzte TNF-Menge unterschied sichnicht signifikant. Bezuglich der be-obachteten Kinetik wurden beimVergleich zu in vivo jedoch bishersprichwortlich "Apfel und Birnen"verglichen. Die Perfusionszellkulturenthalt jedoch die Komponente del'ansonsten fehlenden Elimination, diebei solchen Botenstoffen, die einerstarkeren autokrinen Regulation un-terliegen, einen noch deutlicherenEinfluB auf ihre Kinetik haben sollte.Hinzu kommt im Vergleich zumTierversuch, daB del' Stimulus LPSbei den ZeJlkultursystemen standigprasent war, wahrend er in vivo einerraschen Elimination unterliegt. ImGegensatz zur statischen Kultur lieBesich dies jedoch in der Perfusions-kultur ebenfalls modellieren.Erweiterte Anwendungen der Me-
thodik ergeben sich bei der Untersu-chung yon pharmakologischen Inhi-bitoren: Hier kann neben der Dauer-infusion des Wirkstoffes erstmalsauch die Bolus-Gabe, d.h. einmaligeVerabreichung des Wirkstoffes ent-sprechend z.B. einer Injektion imVersuchstier, in Zellkultur model-liert werden.Die Vorteile der Perfusionskultur
bei Dauerkultivierung yon ZelJen
22
bezuglich der Morphologie und Dif-ferenzierung der Zellen ist vielfachgezeigt (Minuth et al., 1992; Aigneret al., 1994; Gebhardt et aI., 1979;Jauregui et al., 1994; Farghali et al.,1994). Hier wurden nun erstmals inKurzzeitkulturen Effekte del' Perfusi-onskultur studiert. Mit del' dadurcherreichten Annaherung an die in vivoVerhaltnisse wird eine wichtige Dis-krepanz zwischen Ergebnissen ausTierversuchen und aus Zellkulturenabgebaut. Diese war bisher ein we-sentlicher Grund dafur, daB die Ent-zundungsforschung fast ausschliels-lich auf Tierversuchen basierte.
DanksagungDie Autoren dank en ElisabethSchmied und Gregor Pin ski fur ex-zeJlente technische Unterstutzung.Prof. W. Minuth (Universitat Re-gens burg, Anatomie) hat durch kriti-sche Diskussion und hilfreiche Rat-schlage zu diesem Projekt beigetra-gen. Die Arbeit wurde unterstutztdurch das Land Baden- Wiirtternbergim Rahmen der Forderung der Ent-wicklung von Alternativmethodenzur Vermeidung von Tierversuchen.
Literatur
Aigner, 1., Kloth, S., Kubitza, M.,Kashgarian, M., Dermietzel, R. undMinuth, W. W. (1994). Maturationof renal collecting duct cells in vivoand under peri fusion culture. Epith.Cell. BioI. 3,70-78.
Badger, A. M., Olivera, D., Talmadge,J. E. und Hanna, N. (1989). Protecti-ve effect of SK&F 86002, a noveldual inhibitor of arachidonic acidmetabolism, in murine models ofendotoxin shock: inhibition of tumornecrosis factor as a possible mecha-nism of action. Circ. Shock 27, 51-61.
Beutler, B., Milsark, I. W. und Cerami,A. (1985a). Cachectin/tumor necro-sis factor: production, distribution,and metabolic fate in vivo. 1. lmmu-nol. 135,3972-3977.
Beutler, B., Milsark, I. W. und Cerami,A. (l985b). Passive immunizationagainst cachectin/tumor necrosis
factor protects mice from lethal ef-fect of endotoxin. Science 229, 869-871.
Beutler, B. (1993). Tumor necrosisfactors: the molecules and theiremerging role in medicine. NewYork: Raven Press.
Ertel, W., Horrison, M. H., Wang, P.,Zheng, F., Ayala, A., und Chaudry,1. H. (1991). The complex pattern ofcytokines in sepsis. Ann. Surg. 214,141-148.
Espevik, T. und Nissen-Meyer, J.(1986). A highly sensitive cell line,WEHr 164 clone 13, for measuringcytotoxicity factor/tumor necrosisfactor from human monocytes. 1.lmmunol. Meth. 95,99-105
Farghali, H., Kamenikova, L. und Hy-nie, S. (1994). The concept of appli-cation of immobilized and perfusedmammalian cells (a bioreactor mo-del) in biomedical research. Physiol.Res. 43, 117-120.
Flick, D. A. und Gifford, G. E. (1985).Tumor necrosis factor. In P.F. Tor-rence (Hrsg.), Biological responsemodifiers (171-218). Orlando: Aca-demic Press.
Fong, Y., Tracey, K. J., Moldawer, L.L., Hesse, D. G., Manogue, K. B.,Kenney, J. S., Lee, A. T., Kuo, G.c. Allison, A. c. Lowry, S. F. undCerami, A. (1989). Antibodies tocachectinltumor necrosis factor re-duce interleukin IB and interleukin 6apperance during lethal bacteremia.1. Exp. Med. 170, 1627-1633.
Galin, J. 1., Goldstein, I. M. undSnyderman, R. (1992). Inflammation- Basic principles and clinical cor-relates. New York: Raven Press.
Gebhardt, R. und Mecke, D. (J 979).Peri fused monolayer cultures of rathepatocytes as an improved in vitrosystem for studies on ureogenesis.Exp. Cell Res. 124,349-359.
Ha, D. K. K., Gardner, 1. D. undLawton, J. W. (1983). Characteriza-tion of macrophage function in My-cobacterium lepraemurium-infectedmice: sensitivity of mice to endoto-xin and release of mediators andlysosomal enzymes after endotoxintreatment. Parasite Immunol. 5,513-526.
Ha, D. K. K., Cheng, C. P., Fung, K.P., Choy, Y. M. und Lee, C. Y.(1987). Effect of anti-inflammatorydrugs on the production of tumor
ALTEX 13, 1196

wr:"\ HARTUNG ET AL.--~~<--------------------------------¥0~
necrosis factor and lipopolysacchari-de induced mortality in mice.Cancer Lett. 34, 291-296.
Hartung, T., und Wendel, A., (1991).Endotoxin-inducible cytotoxicty inliver cell cultures I. Biochem. Phar-macol. 42, 1129-1135.
Hartung, T, Tiegs, G. und Wendel, A.(1992). The role of leukotriene D4 inseptic shock models. Eicosanoids 5,S42-S44.
Jauregui, H. 0., Naik, S., Santangini,H., Pan, J., Trenkler, D. und Mullon,C. (1994). Primary cultures of rathepatocytes in hollow fiber cham-bers. In. Vitro Cell Dev. Bioi. 30A,23-29.
Kiener, P. A., Marek, F., Rodgers, G.,Lin, P-F., WaIT, G. und Desiderio, J.(1988). Induction of tumor necrosisfactor, lFN-y, and acute lethality inmice by toxic and non-toxic forms oflipid A. 1. Immunol. 141,870-874.
Laskin, D. L. (1990). Nonparenchymalcells and hepatotoxicity. Sem. LiverDisease 10,293-304.
Neugebauer, E. A. und Holaday, J. W.(1993). Handbook of mediators in
septic shock. Ann Arbor: CRe Press.Minuth, W., Derrnietzel, R. , Kloth, S.und Hennerkes, B. (1992). A newmethod culturing renal cells underpermanent superfusion and produ-cing a luminal-basal medium gradi-ent. Kidney International 41, 215-219.
Schade, U. F., Ernst, M., Reinke, M.und Wolter, D. T., (1989). Lipoxy-genase inhibitors suppress formationof tumor necrosis factor in vitro andin vivo. Biochem. Biophys. Res.Commun. 159,748-754.
Seglen, P. O. (1976). Preparation ofisolated rat liver cells. Meth. CellBiol. 13, 29-83.
Smith, S. R., Calzetta, A., Bankowski,J., Kenworthy-Bott, L. und Termi-nelli, C. (1993). Lipopolysacchari-de-induced cytokine production andmortality in mice treated withCorynebacterium parvum. 1. Leu-koc. Biol. 54, 23-29.
Tiegs, G., Wolter, M. und Wendel, A.(1989). Tumor necrosis factor is aterminal mediator in D-galactosami-ne/endotoxin induced hepatitis in
mice. Biochem. Pharmacol. 38,627-631.
Tiegs, G., Niehorster, M. und Wendel,A. (1990). Leukocyte alterations donot account for hepatitis induced byendotoxin or TNF-a in galactosami-ne-sensitized mice. Biochem. Phar-macol. 40, 1317-1322.
Tracy, K. J., Vlassara, H. und Cerami,A. (1989). Cachectin/tumour necro-sis factor. Lancet 20, 1122- J 126.
Waage, A. (1987). Production andclearance of tumor necrosis factor inrats exposed to endotoxin and dexa-methasone. Clin. Immunol. Immun-opathol. 45, 348-355.
Wendel, A. (1990). Biochemical Phar-macology of inflammatory liver in-jury in mice. Meth. Enzymol. 186,675-680.
KorrespondenzadresseDr. Dr. med. Thomas HartungBiochemische PharrnakologieUniversitat KonstanzPostfach 5560D-78434 Konstanz
ALTEX 13, 1/96 23





























