Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO PARÁ
PRÓ-REITORIA DE PESQUISA E PÓS-GRADUAÇÃO
DIRETORIA DE PESQUISA
PROGRAMA INSTITUCIONAL DE BOLSAS DE INICIAÇÃO CIENTÍFICA
RELATÓRIO TÉCNICO - CIENTÍFICO
Período: 01/08/2014 a 01/082015
( ) PARCIAL
(x) FINAL
IDENTIFICAÇÃO DO PROJETOTítulo do Projeto de Pesquisa (ao qual está vinculado o Plano de Trabalho): Controle do comportamento reprodutivo de pássaros sub-Oscines equatoriais: implicações para estudos da biodiversidade.
RESUMO DO PROJETO
Nome do Orientador: Maria Luisa da Silva
Titulação do Orientador: pós-doutoramento na área de comportamento de aves
Faculdade: Ciências Biológicas
Unidade: Instituto de Ciências Biológicas
Laboratório: de Ornitologia e Bioacústica (LoBio)
Título do Plano de Trabalho: Estudo do repertório vocal de contexto comportamental de Myiozetes cayanensis (Aves, Tyrannidae).
Nome do Bolsista: Felipe Matheus dos Reis Castro
Tipo de Bolsa: (x) PIBIC/CNPq
() PIBIC/UFPA
() PIBIC/INTERIOR
() PIBIC/FAPESPA
() PRODOUTOR
() PARD – renovação
() PIBIC/PIAD
() PIBIC/AF-CNPq
() PIBIC/AF-UFPA

INTRODUÇÃO
A bioacústica é uma importante ferramenta baseada no estudo de sons produzidos por animais. O advento de novas tecnologias pós anos 70, essa nova ferramenta vem sendo utilizada em várias linhas de pesquisa como a ecologia, filogenia, etologia e neurociências (VIELLIARD & SILVA 2007).
Para haver comunicação são necessários três fatores, que são: a presença de um emissor, um receptor, e um sinal que possa transmitir uma informação desejada de forma eficiente. Entre os vertebrados, a utilização do sinal acústico está presente em mamíferos, anfíbios e aves adequando este sinal às exigências de propagação do meio onde cada espécie se encontra (VIELLIARD, 2004). Nas aves, a produção de um sinal acústico depende do fluxo do ar expirado nos pulmões até o aparelho fonador que é a siringe (SILVA & VIELLIARD, 2010). As emissões sonoras emitidas por esse mecanismo, as vocalizações, podem ser identificadas como canto e chamados ou gritos.
O canto é considerado o sinal sonoro de uma determinada espécie que é responsável pela transmissão da informação de reconhecimento específico, sendo adaptado ao meio em que essa espécie se encontra e utilizado na busca de um indivíduo da mesma espécie. Isto se deve à necessidade de manutenção da espécie e o indivíduo que desvia do padrão de uma determinada espécie, não será reconhecido como membro da mesma e, como consequência, acabará sendo eliminado por seleção natural (VIELLIARD 2004). Já os chamados são todas as vocalizações que apresentam todas as outras funções biológicas, com exceção da função de reconhecimento de espécie, como por exemplo, defesa territorial e corte (VIELLIARD 2011).
A ordem dos passeriformes encontra-se dividida entre as duas SubOrdens, que são a dos Suboscines, que apresentam cantos estereotipados e simples e dos Oscines, que geralmente emitem cantos variados e complexos indivíduo (SILVA & VIELLIARD 2010, SILVA 1995). Os cantos simples dos SubOscines são esperados serem inatos, pois não necessitam de um modelo para que sejam funcionais, os filhotes podem ser criados em isolamento acústico na ausência de audição do canto da espécie. O que canto dos Oscines, que se apresenta sob uma forma mais complexa, pois é mediado por um mecanismo que se encontra presente em alguns taxa de aves e mamíferos, que é a aprendizagem. Logo, essa estratégia necessita de um modelo que possa desempenhar um papel de “tutor” para os jovens, objetivando com que eles possam aprender o canto de reconhecimento da espécie (VIELLIARD 2004).
Na etologia, ciência que estuda o comportamento dos animais, a Bioacústica se tornou necessária para o entendimento de sinais sonoros desses animais. E uma abordagem prática envolvendo a relação do comportamento com o entendimento dos sinais sonoros dos animais, é a descrição do repertório vocal. Segundo Vielliard & Silva (2007), o repertório vocal consiste em quantificar os sinais sonoros de uma espécie e buscar associá-los a funções biológicas de seu comportamento. É importante ressaltar que o tamanho desse repertório pode variar de espécie para espécie dependendo do nível de complexidade de seu sistema de comunicação.
Mediante a essa ideia, existe a hipótese da complexidade social, a qual afirma que as espécies com uma estrutura social complexa tendem a desenvolver mecanismos de comunicação mais complexos, sendo essa hipótese comprovada no trabalho de Grieves e seus colaboradores(2015), que encontrou 11 chamados em contextos diferentes e uma vocalização em grupo para a espécie Anu preto (Crotophaga ani).
Uma técnica bastante utilizada pelos ornitólogos com o objetivo de entender o repertório vocal de uma espécie é o playback, que segundo Vielliard e Silva (2010), consiste em reproduzir por meio de uma fonte sonora (e. g. caixa acústica) o canto de uma determinada espécie, tendo como objetivo incitar uma resposta (tanto vocal quanto aproximação e observação visual). Esse estímulo sonoro pode desencadear ações de territorialismo e/ou uma série de outros comportamentos na espécie a ser estudada.

No caso do Myiozetetes cayanensis, objeto deste estudo, pertencente à família Tyrannidae, é uma das espécies de bentevis reconhecida por apresentar os lados da cabeça bem anegrados e pelas bordas das rêmiges (penas das asas) e retrizes (penas da cauda) com a coloração ferrugínea, características que lhe dão o nome de Bentevizinho-de-asa-ferrugínea. Esta espécie pertence à SubOrdem Suboscine, logo apresenta o canto de reconhecimento especifico considerado estereotipado. Partindo deste pressuposto, este trabalho buscou descrever o repertório vocal do Myiozetetes cayanensis.
JUSTIFICATIVA
A descrição do repertório vocal é de fundamental importância para o entendimento da diversidade funcional dos chamados das aves, entendimento este que nos proporciona uma melhor compreensão sobre o comportamento delas (GRIEVES et al. 2015). O estudo do repertório vocal é fundamental para a análise do repertório de como se dá as relações intraespecíficas, pois revela algumas estratégias de vida que se mantiveram mediante à necessidade da emissão de sinais sonoros durante o decorrer do processo evolutivo desta espécie (VIELLIARD 2007).
A descrição do repertório vocal é fundamental para dar suporte à hipótese de que a complexidade social direciona a evolução da complexidade de um sistema de comunicação (FREEBERG et al. 2012). Ou seja, espécies que apresentam complexidade social na relação entre os indivíduos de um dado agrupamento, tendem a desenvolver mecanismos de comunicação mais complexos, que no caso do presente estudo está baseada na comunicação vocal.
A ontogênese do canto dos passeriformes da subordem Tyranni ou Suboscine se apresenta sob a forma estereotipada, simples e de repertórios pequenos com variações geográficas mínimas (SILVA 1995). No entanto, mediante a grande diversidade dessa subordem e de poucos estudos com enfoque no comportamento vocal da mesma, é imprudente fazer generalizações. Alguns estudos feitos com Tiranídeos, como o Pitangus sulphuratus, mostraram que seu repertório vocal na área urbana de Santiago de Cali, na Colômbia, apresentou pequenas variações individuais (MENDONZA & ARCE 2012). Diante do exposto, torna-se importante estudo com espécies pertencentes a essa subordem para uma melhor compreensão de seu comportamento vocal, gerando conhecimento acerca da evolução dos passeriformes.
OBJETIVO:
Descrever o repertório vocal relacionando a contextos comportamentais e definir estes contextos através do registro da frequência das vocalizações relacionadas ao comportamento realizado na hora da emissão da vocalização de Myiozetetes cayanensis.
MATERIAL E MÉTODOS:
- Objeto de estudo:
Classificação taxonômica da espécie:
Reino: Animalia
Filo: Chordata
Subfilo: Vertebrata
Classe: Aves
Ordem: Passeriformes
Subordem: Tyranni
Família: Tyrannidae
Gênero: Myiozetetes
Espécie: cayanensis (Linnaeus)

Nome popular: Bentevizinho-de-asa-ferrugínea
- Características:
Segundo Sick (1997), M. cayanensis pertence à família Tyrannidae que apresenta um total de 413 espécies e possui a sua maior concentração na região tropical, constituindo 18% das espécies de Passeriformes da América do Sul, e essa família se adaptou aos mais variados nichos ecológicos, sendo que na Amazônia Brasileira, encontram-se quatro espécies do gênero Myiozetetes. Semelhante ao bem-te-vi (Pitangus sulphuratus), porém de tamanho menor, o bentevizinho-de-asa-ferrugínea possui em torno de 17,5 cm de comprimento. O M. cayanensis é uma das várias espécies de bentevis pequenos cuja identificação não é fácil; sendo possível reconhece-lo pelos lados bem anegrados da cabeça, pela faixa amarela ou alaranjada no píleo e, sobretudo, pelas bordas nitidamente ferrugíneas das rêmiges e das retrizes (SICK 1997).
Seu canto de reconhecimento específico é em dueto que segundo Farabaugh (1982), envolve a coordenação temporal de duas ou mais vocalizações de aves pertencentes a um mesmo grupo familiar, geralmente um casal, podendo ser acompanhado por exibições visuais em algumas espécies. No caso do Myiozetetes cayanensis, este dueto é acompanhado às vezes por uma exibição visual (display) na qual o macho e a fêmea ao vocalizar batem suas asas simultaneamente.
Figura 1: Sonograma de um dueto do Myiozetetes cayanensis após a utilização do playback. Os retângulos marcados abaixo das vocalizações representam o ruído do som do bater das asas que é o display da espécie.
- Distribuição Geográfica:
Myiozetetes cayanensis se distribui no norte e Oeste da Colômbia e Oeste do equador (sul para El Oro); Guianas, Venezuela, Leste da Colômbia, Leste do equador (poucos registros), Leste da Amazônia e Brasil central (a Amazônia, tanto quanto a foz do Rio Negro, e embora ao Sul do Pará, Maranhão, e Piauí para Minas Gerais, Rio de Janeiro, e Mato Grosso do Sul), Norte da Bolívia (Sul para Cochabamba e Norte para Santa Cruz), e Adjacente ao Sudeste do Peru (Reserva Tambopata em Madre de Dios) (RIDGELY & TUDOR 1994).

Figura 2: Distribuição geográfica do Myiozetetes cayanensis
- Área de Estudo:
1. Universidade Federal do Pará
O estudo foi desenvolvido na Universidade Federal do Pará, localizada no bairro do Guamá em Belém, PA. Esta área é formada por uma amostra de ecossistema Amazônico costeiro. Apesar de este local estar bastante impactado por ação antrópica, ele apresenta uma avifauna bastante diversificada. O Myiozetetes cayanensis é uma das espécies presentes e foi possível fazer seu estudo comportamental em meio natural através da observação visual e/ou sonora. A área foi dividida em quatro pontos (A, B, C e D), sendo A, B e C localizados no Campus básico e o ponto D localizado no Campus do profissional, como mostra a (figura 3).
Figura 3: mapa da área de estudo na Universidade Federal do Pará com a marcação dos pontos A, B, C e D.

Repertório Vocal
As vocalizações foram registradas através do gravador TASCAM DA-P1 e microfone unidirecional Sennheiser ME67. Os registros das vocalizações do objeto de estudo foram realizadas nos quatro pontos selecionados (ver figura 3) em um total de dez dias de esforço amostral até o presente momento. O programa utilizado para a análise das gravações foi o Adobe Audition 1.5. As observações comportamentais foram anotadas no caderno de campo ou foram narradas no decorrer das gravações. Essas vocalizações foram divididas em categorias mediante ao contexto comportamental observado, categorias essas reconhecidas como canto 1, e chamados 1, 2, 3 e 4.
Baseado no método preestabelecido por Ferraz (2008), os registros das vocalizações foram realizados em duas etapas. A primeira consiste na captura dos sinais sonoros emitidos e na caracterização preliminar do contexto comportamental, relacionando-o com cada vocalização capturada. Na fase seguinte deu-se continuidade ao registro vocal buscando a captura de novos componentes vocais para o repertório da espécie, além de buscar a contextualização das vocalizações através do uso da técnica de playback.
Todas as vocalizações analisadas no programa Adobe audition 1.5, foram colocadas em uma planilha do Microsoft Excel 2010, organizadas na ordem de sua coleta durante os dias de gravação. E cada uma dessas vocalizações, recebeu um código formado pela letra G (significa gravação) seguida de um número que corresponde a gravação em que essa vocalização se encontra mais uma letra V (significa vocalização) seguida de um número que corresponde a vocalização na ordem das coletas, por exemplo, G1V12 é a vocalização encontrada na gravação 1 e na ordem de coleta ela é 12ª vocalização. A análise das vocalizações foram feitas no plano melódico do som, demonstração gráfica (sonogramas) que apresenta a variação da frequência ao longo do tempo (frequência em Hz no eixo das ordenadas e tempo em segundos no eixo das abscissas).
Cada vocalização apresenta um conjunto de notas que foram nomeadas com as letras do alfabeto mediante a ordem na qual foram encontradas nos dias de amostragem. Cada nota apresenta seu formato especifico com suas características físicas definidas. Todas as vocalizações foram representadas por sonogramas (gráficos onde os sons estão representados).
Os registros das vocalizações, amostras deste estudo, ocorreram durante o período de um ano e o esforço amostral ocorreu durante dez dias de coleta (um pela manhã e nove pela tarde) no decorrer deste período. A cada visita, utilizou-se um ou dois pontos marcados na área de estudo para a busca da espécie objeto deste, pois alguns dias, existiam pontos que não apresentavam nenhum contato visual ou auditivo, logo buscamos o ponto seguinte na ordem alfabética. Quando a espécie era encontrada, fizemos as observações sobre seu comportamento associado às amostras vocais (quando possível), comportamentos estes que eram anotados em um caderno de campo ou descritos através das narrações no decorrer das gravações.
Playback:
Para realização dos experimentos de playback a metodologia utilizada está baseada em Ferraz (2008), porém com algumas adaptações. As vocalizações oriundas da primeira etapa da coleta de registros vocais (vocalizações 1, 2, 3, 4, 5 e 6) presentes no arquivo sonoro do Laboratório de Ornitologia e Bioacústica (LOBio) foram utilizadas durante o playback. O playback foi realizado com o auxilio de um celular modelo LG-L4 e caixa amplificadora B-betron KB-45. Foram salvos os arquivos contendo as vocalizações no celular, e quando iniciado o teste, estas eram reproduzidas através da caixa amplificadora para que os indivíduos do local possam escutar e emitir uma resposta seja visual e/ou auditva. Esta técnica foi dividida em três momentos que são:
- Pré-playback: Após a visualização e identificação dos indivíduos e seus territórios é registrado o número de destes e seu estado fisiológico (se estavam forrageando ou nidificando, por exemplo). Quando os indivíduos não se encontrarem em comportamento de disputa por território, por exemplo, ou outros comportamentos que pudessem desviar sua atenção ao playback, prosseguem com os testes. Por um período de 2 minutos serão observados os tipos de vocalizações emitidas

espontaneamente e suas frequências de ocorrência, além de informações referentes ao comportamento dos indivíduos avistados. No caso de situação adversa no início do playback, a exemplo da ausência de indivíduo no ponto amostral, dificuldade de visualização ou barulho demasiado, amostrou-se o ponto seguinte.
- Playback: O playback será constituído por cinco sessões de exposição às vocalizações, cada uma com duração de um minuto, intercaladas por trinta segundos de silêncio. Para a realização do playback será utilizadas diferentes vocalizações, que serão sorteadas para cada ponto amostrado distante pelo menos 150 metros um do outro.
- Pós-playback: Durante este fase serão observadas as mesmas informações de comportamento verificadas durante o pré-playback.
RESULTADOS:
Foram encontrados no Campus da Universidade Federal do Pará, no período da manhã e a tarde um total de 375 vocalizações (amostras). O repertório vocal do Myiozetetes cayanensis apresentou 5 categorias de vocalizações principais (um canto e 4 chamados) com seus devidos contextos comportamentais estabelecidos mediante as observações. Essas 5 grandes categorias de vocalizações apresentaram variantes dentro de cada uma delas, devido a variação na ordem das notas presentes em cada vocalização. O repertório também apresentou as combinações dessas vocalizações (canto e chamados), sendo todas apresentadas na forma de sonogramas juntamente com seus respectivos comportamentos observados em campo.
A vocalização 1 que é considerada neste estudo como chamado 1 (ver figura 4), e é composta por uma nota A (frequência máx. de 3320 Hertz Hz e mín. de 2530 Hz); uma nota E (frequência máx. 4450 Hz e mín. 2850 Hz), e 5 notas D (frequência máx.5060 Hz e mín. 2810 Hz). Esta vocalização apresenta algumas variações que dependem das quantidades e ordem das notas D, E ou F. O intervalo entre a nota A e a E apresenta duração de 527 milissegundos (ms); o intervalo entre nota E e a primeira nota D é de 176 ms; o intervalo entre a primeira e a segunda nota D apresenta um intervalo de 202 ms e o intervalo entre as notas D tende a aumentar progressivamente (a terceira e quarta nota D o intervalo foi de 261 ms). A vocalização apresenta uma duração de 3 segundos e 653 ms.
Esta vocalização foi emitida como resposta a vocalizações de indivíduos da mesma espécie que se encontravam longe por volta de uns 100 metros ou perto cerca de 5 metros, em que este indivíduo emitia essa vocalização sozinho. Quando excitado pela vocalização de playback da mesma espécie, o indivíduo responsável por este tipo de vocalização sobrevoava bem próximo à fonte sonora, chegando a passar bem próximo do observador. Por conta desse comportamento que foi visto em locais próximos do ninho, onde um indivíduo sozinho sobrevoava bem próximo a fonte sonora e ao observador, o chamado 1 foi considerado mediante a seu contexto comportamental como um chamado de territorial e apresenta algumas variações que serão mencionadas a seguir.

Figura 4: Sonograma da Vocalização 1 (V1G1) composta pelas notas A, E e D, formando AEDDDDD.
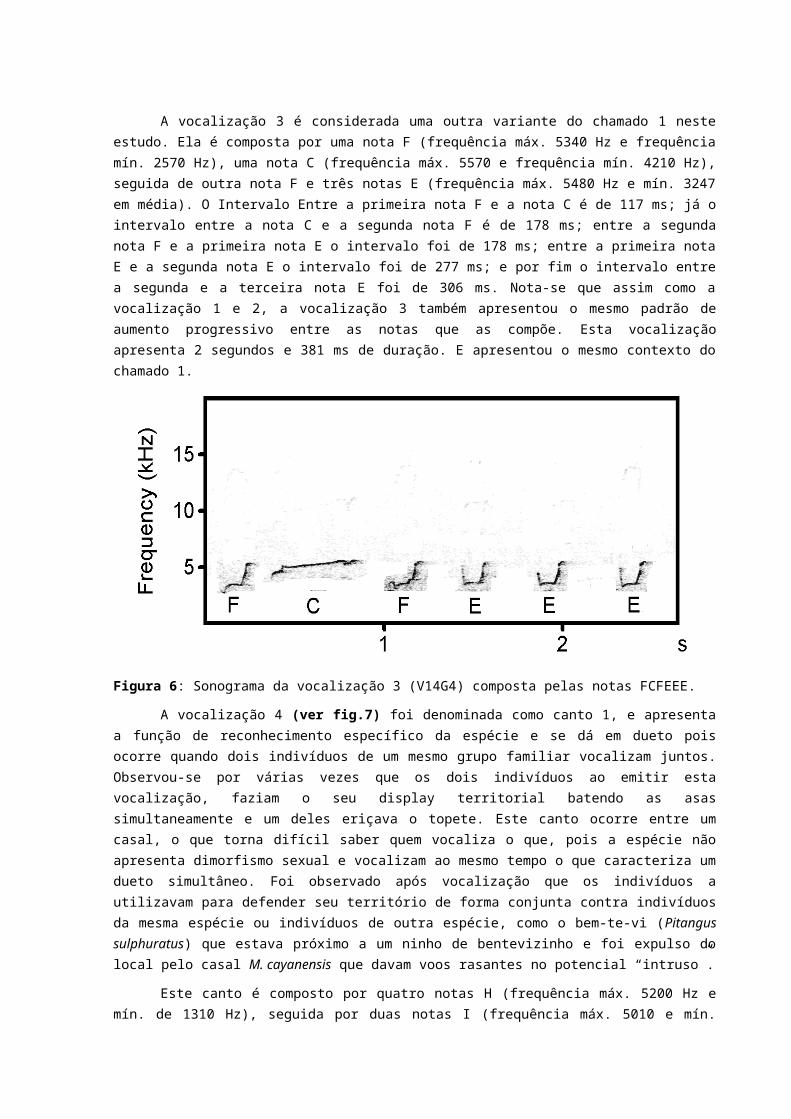
A vocalização 2 (ver fig. 2) é considerada uma variação do chamado 1, formada por uma nota B (frequência máx. 4820 Hz e mín. 3560 Hz); seguida de uma nota E (frequência máx.4820 Hz e mín. 2710 Hz), seguida de uma nota D (frequência máx. 4960 Hz e mín. 2710 Hz), seguida de 4 notas E, e por fim 3 notas G (frequência máx. 4170 Hz e mín. 2710 Hz). O intervalo entre a nota B e a primeira nota E é de 258 ms; já o intervalo entre a primeira nota E e a nota D é de 184 ms; já o intervalo entre a quinta nota E e a primeira nota G é de 357 ms e, assim como a vocalização 1, essa vocalização tende a aumentar o intervalo entre as notas progressivamente também. A vocalização apresenta 4 segundos e 613 ms de duração. Esta apresenta o mesmo contexto comportamental do chamado 1.
Figura 5: Sonograma da vocalização 2 (V12G3) compostas pelas notas BEDEEEEGGG.
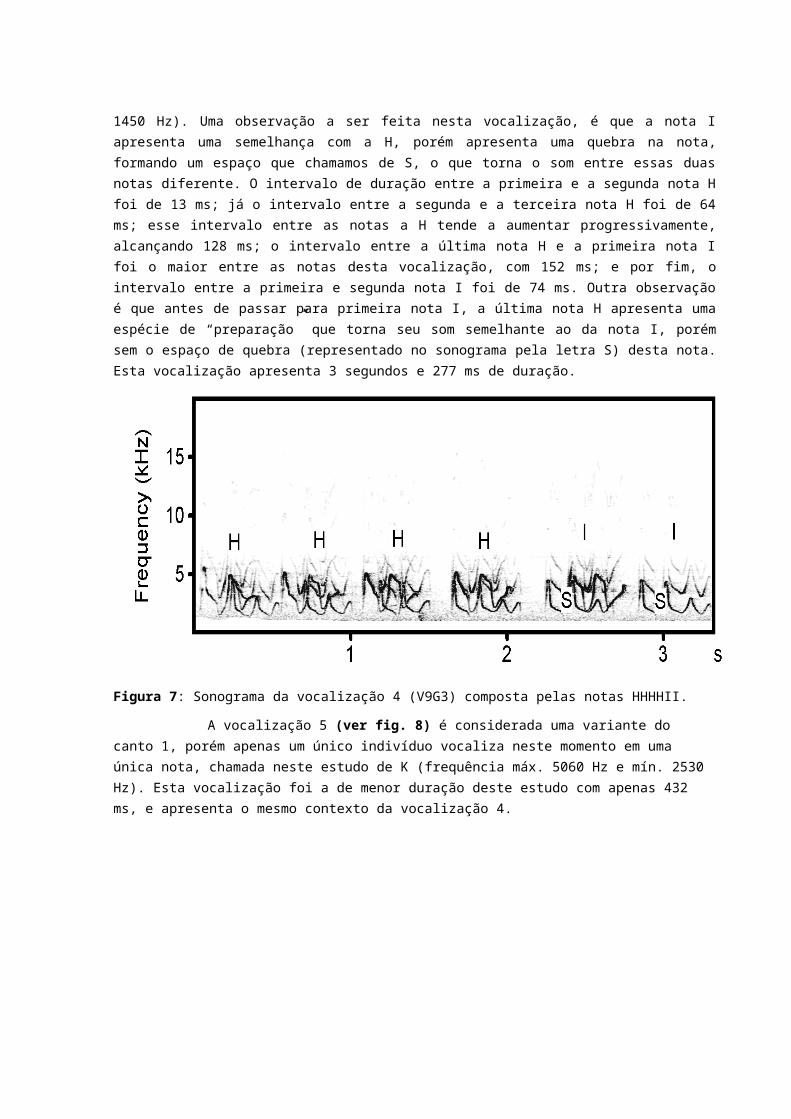
A vocalização 3 é considerada uma outra variante do chamado 1 neste estudo. Ela é composta por uma nota F (frequência máx. 5340 Hz e frequência mín. 2570 Hz), uma nota C (frequência máx. 5570 e frequência mín. 4210 Hz), seguida de outra nota F e três notas E (frequência máx. 5480 Hz e mín. 3247 em média). O Intervalo Entre a primeira nota F e a nota C é de 117 ms; já o intervalo entre a nota C e a segunda nota F é de 178 ms; entre a segunda nota F e a primeira nota E o intervalo foi de 178 ms; entre a primeira nota E e a segunda nota E o intervalo foi de 277 ms; e por fim o intervalo entre a segunda e a terceira nota E foi de 306 ms. Nota-se que assim como a

vocalização 1 e 2, a vocalização 3 também apresentou o mesmo padrão de aumento progressivo entre as notas que as compõe. Esta vocalização apresenta 2 segundos e 381 ms de duração. E apresentou o mesmo contexto do chamado 1.
Figura 6: Sonograma da vocalização 3 (V14G4) composta pelas notas FCFEEE.
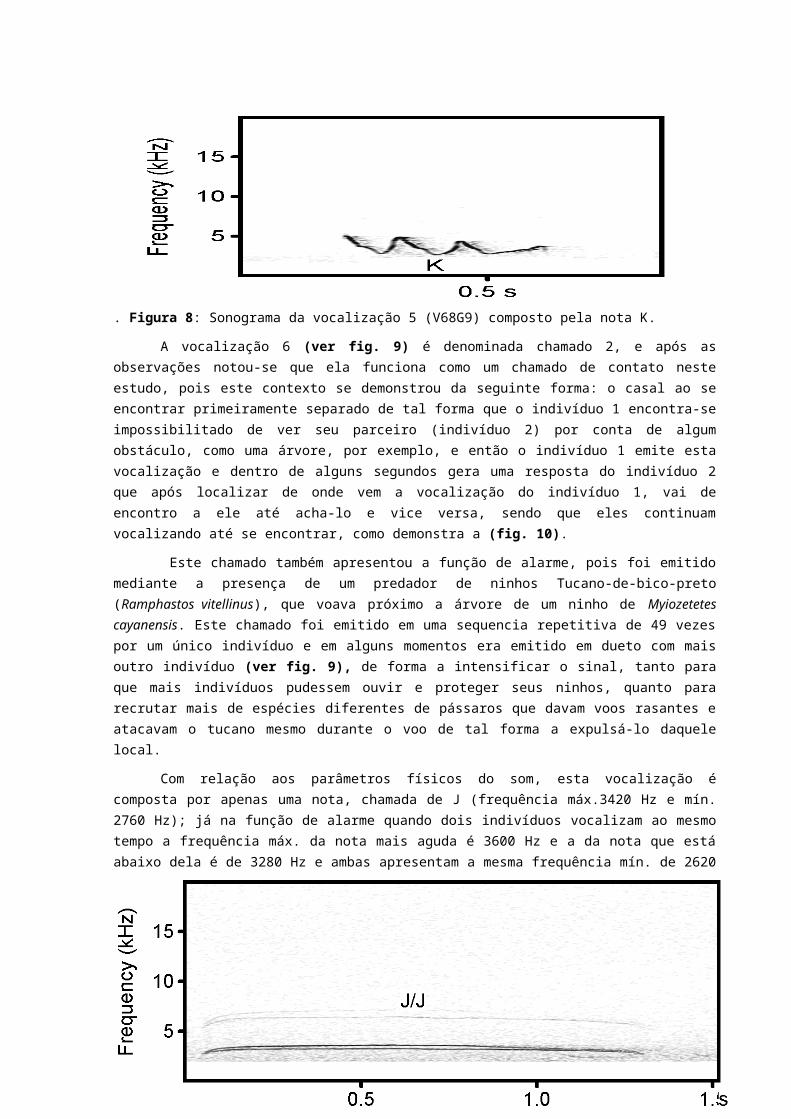
A vocalização 4 (ver fig.7) foi denominada como canto 1, e apresenta a função de reconhecimento específico da espécie e se dá em dueto pois ocorre quando dois indivíduos de um mesmo grupo familiar vocalizam juntos. Observou-se por várias vezes que os dois indivíduos ao emitir esta vocalização, faziam o seu display territorial batendo as asas simultaneamente e um deles eriçava o topete. Este canto ocorre entre um casal, o que torna difícil saber quem vocaliza o que, pois a espécie não apresenta dimorfismo sexual e vocalizam ao mesmo tempo o que caracteriza um dueto simultâneo. Foi observado após vocalização que os indivíduos a utilizavam para defender seu território de forma conjunta contra indivíduos da mesma espécie ou indivíduos de outra espécie, como o bem-te-vi (Pitangus sulphuratus) que estava próximo a um ninho de bentevizinho e foi expulso do local pelo casal M. cayanensis que davam voos rasantes no potencial “intruso”.
Este canto é composto por quatro notas H (frequência máx. 5200 Hz e mín. de 1310 Hz), seguida por duas notas I (frequência máx. 5010 e mín. 1450 Hz). Uma observação a ser feita nesta vocalização, é que a nota I apresenta uma semelhança com a H, porém apresenta uma quebra na nota, formando um espaço que chamamos de S, o que torna o som entre essas duas notas diferente. O intervalo de duração entre a primeira e a segunda nota H foi de 13 ms; já o intervalo entre a segunda e a terceira nota H foi de 64 ms; esse intervalo entre as notas a H tende a aumentar progressivamente, alcançando 128 ms; o intervalo entre a última nota H e a primeira nota I foi o maior entre as notas desta vocalização, com 152 ms; e por fim, o intervalo entre a primeira e segunda nota I foi de 74 ms. Outra observação é que antes de passar para primeira nota I, a última nota H apresenta uma espécie de “preparação” que torna seu som semelhante ao da nota I, porém sem o espaço de quebra (representado no sonograma pela letra S) desta nota. Esta vocalização apresenta 3 segundos e 277 ms de duração.
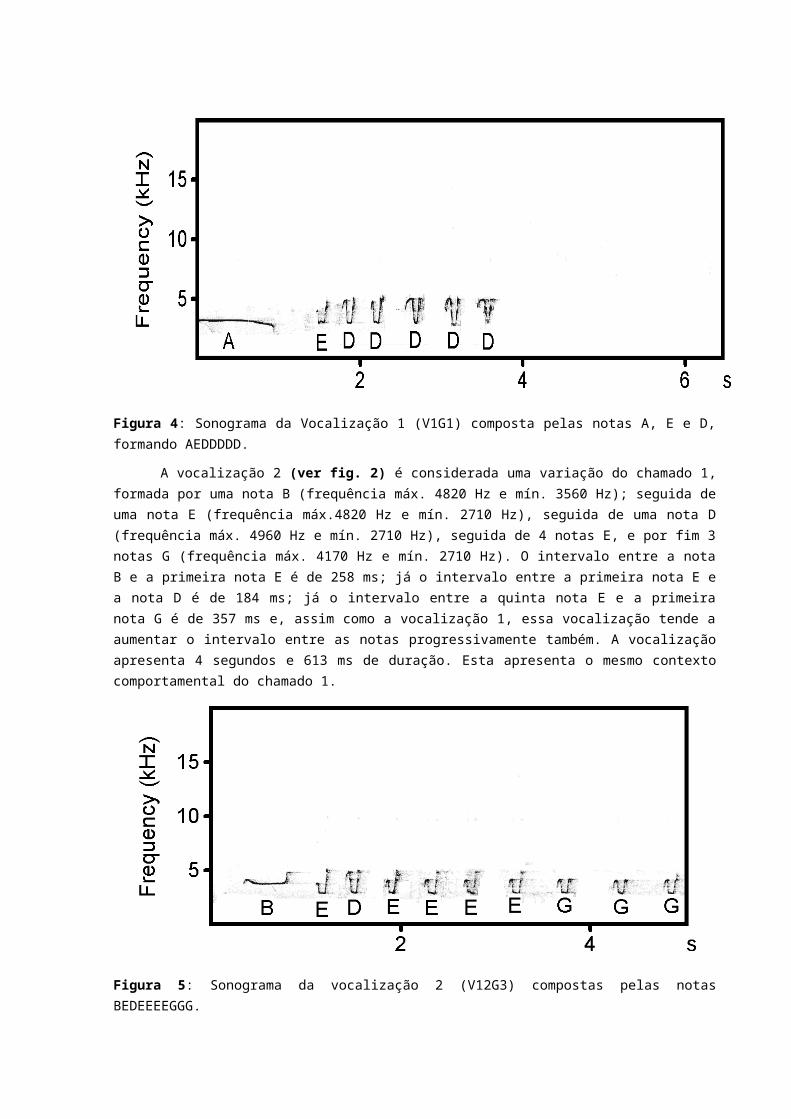
Figura 7: Sonograma da vocalização 4 (V9G3) composta pelas notas HHHHII.
A vocalização 5 (ver fig. 8) é considerada uma variante do canto 1, porém apenas um único indivíduo vocaliza neste momento em uma única nota, chamada neste estudo de K (frequência máx. 5060 Hz e mín. 2530 Hz). Esta vocalização foi a de menor duração deste estudo com apenas 432 ms, e apresenta o mesmo contexto da vocalização 4.
. Figura 8: Sonograma da vocalização 5 (V68G9) composto pela nota K.
A vocalização 6 (ver fig. 9) é denominada chamado 2, e após as observações notou-se que ela funciona como um chamado de contato neste estudo, pois este contexto se demonstrou da seguinte forma: o casal ao se encontrar primeiramente separado de tal forma que o indivíduo 1 encontra-se impossibilitado de ver seu parceiro (indivíduo 2) por conta de algum obstáculo, como uma árvore, por exemplo, e então o indivíduo 1 emite esta vocalização e dentro de alguns segundos gera uma resposta do indivíduo 2 que após localizar de onde vem a vocalização do indivíduo 1, vai de encontro a ele até acha-lo e vice versa, sendo que eles continuam vocalizando até se encontrar, como demonstra a (fig. 10).
Este chamado também apresentou a função de alarme, pois foi emitido mediante a presença de um predador de ninhos Tucano-de-bico-preto (Ramphastos vitellinus), que voava próximo a árvore de um ninho de Myiozetetes cayanensis. Este chamado foi emitido em uma sequencia repetitiva de 49 vezes por um único indivíduo e em alguns momentos era emitido em dueto com mais outro indivíduo (ver fig. 9), de forma a intensificar o sinal, tanto para que mais indivíduos pudessem ouvir e proteger seus ninhos, quanto para recrutar mais de espécies diferentes de pássaros que davam voos rasantes e atacavam o tucano mesmo durante o voo de tal forma a expulsá-lo daquele local.
Com relação aos parâmetros físicos do som, esta vocalização é composta por apenas uma nota, chamada de J (frequência máx.3420 Hz e mín. 2760 Hz); já na função de alarme quando dois indivíduos vocalizam ao mesmo tempo a frequência máx. da nota mais aguda é 3600 Hz e a da nota que está abaixo dela é de 3280 Hz e ambas apresentam a mesma frequência mín. de 2620 Hz. Observamos que em média o tempo que um indivíduo leva para responder ao outro é de 5,33 segundos. Esta vocalização apresenta 805 ms de duração sendo a segunda vocalização de menor duração do repertório deste estudo.
Figura 9: sonograma da vocalização 6 (V330G30) composto por J/J. Contexto de alarme.
Figura 10: Sonograma da vocalização 6 (V32G6) composta pela nota J- um indivíduo vocaliza com a primeira nota J e o outro responde com a segunda nota J. Contexto de chamado de contato.
A vocalização 7 (ver fig. 11) é considerada como um chamado 3 e não foi descrito no repertório vocal das 14 vocalizações do Myiozetetes cayanensis encontrado por (FERRAZ, 2008). Esta vocalização nova foi observada no momento em que dois indivíduos da mesma espécie estavam confronto durante o voo, logo foi definida como um chamado agonístico. Esta vocalização foi observada em um mesmo contexto, porém entre espécies diferentes, onde o M. cayanensis dava voos rasantes e atacava com seu bico de forma a expulsar de seu território um Tucano-grande-de-papo-branco (Ramphastos tucanus) que voava próximo a uma árvore que onde havia um ninho de Myiozetetes cayanensis.
A vocalização 7 apresenta vários harmônicos (fig. 11), e por isso foi levado em consideração apenas a nota de maior intensidade da vocalização (nota que se apresenta mais escurecida no
sonograma) que apresenta a frequência máxima de 6420 Hz e a mínima de 4680 Hz. Já a duração desta vocalização foi de 1 segundo e 385 ms.
Figura 11: Sonograma da vocalização 7 com a nota de maior intensidade indicada pela seta vermelha.
A vocalização 8 (ver fig. 12) foi denominada como chamado 4, e foi observada em vários momentos em que indivíduos a emitiam tanto dentro quanto fora do ninho. Observou-se o seguinte comportamento dos dois indivíduos: um ficava fora e outro ficava dentro do ninho e geralmente após a emissão dessa vocalização pelo indivíduo que estava dentro do ninho, o outro que estava fora não entrava no ninho e, em alguns momentos, o indivíduo que estava fora também vocalizava e observou-se que a partir disso o indivíduo que estava dentro, saia do ninho e depois de um curto período o outro indivíduo entrava no ninho, logo se classificou esse esta vocalização como chamado de vigilância do ninho, pois o casal se alterna para vigiar o ninho.
Esta vocalização se apresenta sobre duas notas de forma diferentes que são: as notas M (frequência máx. 3090 Hz e mín. 2390 Hz) e as notas L (frequência máx. 3980 Hz e mín. 2250 Hz) que estão ambas em uma série sequencial. Acredita-se que estejam associadas ao indivíduo que está fora e a outra nota associada ao indivíduo que está dentro do ninho, pois as notas apresentam frequências e formas diferentes. A vocalização apresentou um intervalo de tempo de 1 segundo e 581 ms.
Figura 12: sonograma da vocalização 8 (V302G28) composta pelas notas MMMMMLLLLLLL.
A vocalização 9 (ver fig.13) foi considerada uma combinação entre o chamado 1 (chamado territorial) e o canto 1 (canto em dueto). Essa combinação foi observada em quatro ocasiões, em que os M. cayanensis responderam ao playback da mesma espécie de tal modo que o indivíduo dava voos rasantes bem próximos a fonte sonora de forma a defender seu território e fazia o display territorial, batendo as asas durante a parte que emitia o canto 1. Está vocalização apresentou a seguinte composição: uma nota B (frequência máx. 3980 Hz e mín. 2150 Hz), seguida de 4 notas H (frequência máx. 5200 Hz e mín. de 1310 Hz). A vocalização apresenta duração de 3 segundos e 218 ms.
Figura 13: sonograma da vocalização 9 (V7G3) composta pelas notas BHHHH.
A vocalização 10 (ver fig. 14) é considerada uma combinação entre o chamado 1 (chamado territorial), canto 1 (um único indivíduo vocalizando) e canto 1 (em dueto). Essa vocalização foi observada uma única vez, e foi emitida como resposta a emissão do play back da vocalização V7G3 (de M. cayanensis). O contexto comportamental dessa vocalização foi a aproximação desses dois indivíduos que voaram próximo a fonte sonora e ao observador, pousando logo após em uma árvore que estava a aproximadamente 10 metros do observador e fizeram o display territorial batendo as asas. A vocalização apresentou a seguinte composição: uma nota F (frequência máx. 4030 Hz e mín. 2100 Hz), uma nota D (frequência máx. 5390 Hz e mín. 2710 Hz), uma nota E (frequência máx. 4070 Hz e mín. 2290 Hz), uma nota K (frequência máx. 5760 Hz e mín. 2250 Hz), uma nota F, uma nota K, duas notas G (frequência máx. 4210 e mín. 2430 Hz), uma nota K, uma nota G, uma nota K e uma D e três notas H (frequência máx. 5760 Hz e mín. 1920 Hz). A vocalização tem duração de 4 segundos e 741 ms.
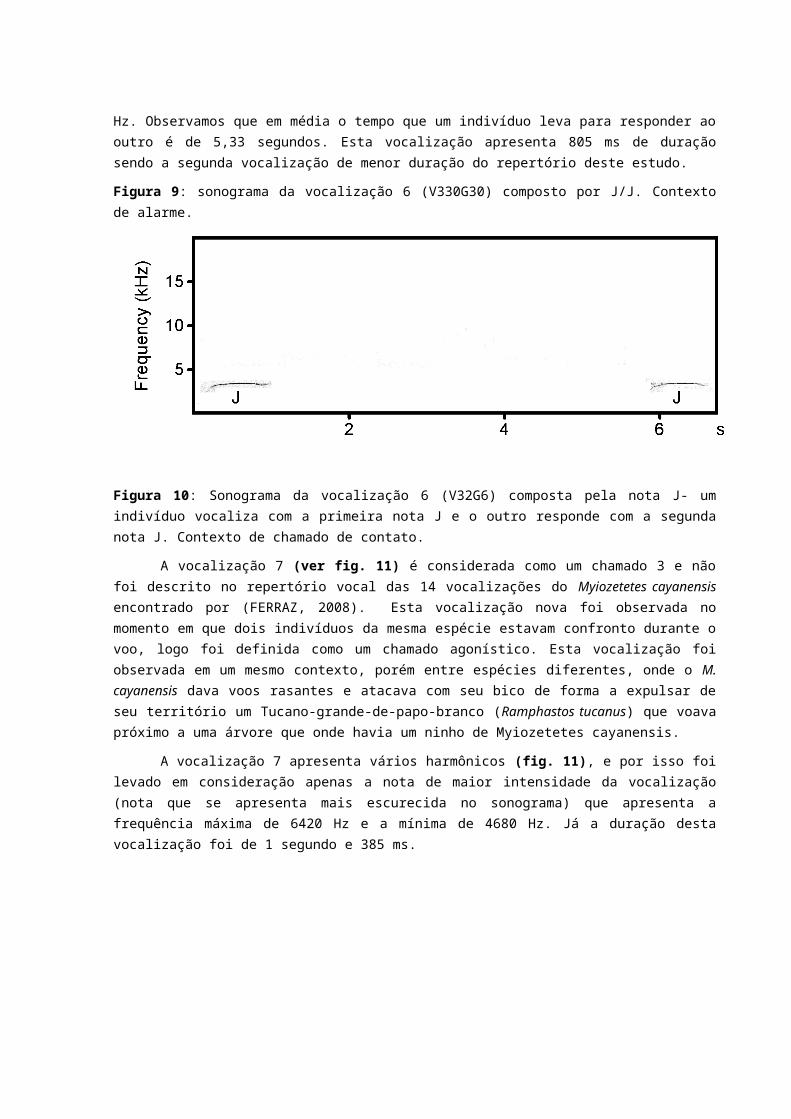
Figura 14: sonograma da vocalização 10 (V320G29) composta pelas notas FDEKFKGGKGKDHHH.
A vocalização 11 (ver fig.15) é considerada uma combinação de chamados ou um chamado misto entre o chamado 2 (contato a longa distancia ou alarme) e o chamado 1 (territorial). Este chamado foi emitido em dois momentos e apresentou o seguinte contexto comportamental: o indivíduo respondeu ao play back de forma a defender seu território passando próximo à fonte sonora através de um voo rasante e pousando próximo ao observador (cerca de uns 10 metros). Acredita-se que a primeira parte desse chamado misto que é o chamado 2 (contato a longa distância ou alarme), fora utilizada para recrutar o outro indivíduo que estava longe daquele local para que assim pudessem defender o território juntos com um objetivo de ter um melhor sucesso contra a uma provável ameaça para o território deles, que neste caso foi o observador, porém como o outro não estava lá ainda, ele emitiu o chamado territorial para tentar defender seu território sozinho naquele momento.
O chamado misto é composto pelas seguintes notas: uma nota J (frequência máx. 2950 Hz e mín. 2340 Hz), uma nota B (frequência máx. 4540 Hz e mín. 2760 Hz) e três notas E (frequência máx. 4870 Hz e mín. 2710 Hz). A vocalização apresenta uma duração de 3 segundos e 792 centésimos.
Figura 15: sonograma da vocalização 11 (V155G14) composta pelas notas JBEEE.
A vocalização 12 (ver fig.16) é uma combinação entre dois chamados, ou seja, também é um chamado misto entre o chamado 4 (vigilância do ninho) e o chamado 1 (territorial). Este chamado foi catalogado em apenas uma ocasião em que não foi observado um contexto comportamental, porém

mediante a suas funções já preestabelecidas, deduz-se que um bentevizinho provavelmente fora do ninho, ao se comunicar com o outro que estava dentro, observou que existia uma potencial ameaça ao seu território nas proximidades no ninho e acabou reagindo com o chamado 1 (territorial)·
A vocalização é composta por 5 notas M (frequência máx. 3510 Hz e mín. 1960 Hz.) e uma nota B (frequência máx. 4680 Hz e mín.2850 Hz). A vocalização apresentou uma duração de 1 segundo e 60 ms.
Figura 16: sonograma da vocalização 12 (V141G13) composta por MMMMMB.
DISCUSSÃO:
Um dado bastante observado e muito interessante no repertório vocal do Myiozetetes cayanensis foi a forma com que a defesa de território estava fortemente correlacionada a seu comportamento vocal, pois observou-se nas análises das vocalizações a existência de dois casos relacionados a isso.
No primeiro caso, notou-se que os chamados mistos se apresentavam em combinações de chamados em que pelo menos um deles era o chamado territorial, por exemplo, chamado misto entre chamado territorial mais dueto; chamado de contato a longa distância mais chamado territorial; chamado de vigilância de ninho mais o chamado territorial e o maior dentre os chamados mistos que foi o chamado territorial mais canto na forma individual mais canto na forma de dueto. O segundo caso, é o fato de a espécie apresentar um canto e um chamado com a mesma função de defesa territorial.
Observou-se que as combinações do bentevizinho apresentavam em geral uma forma de defender território contra potenciais ameaças de tal forma a expulsá-las do território, sendo este o mesmo resultado encontrado por Fandiño-Mariño (1989), na descrição do repertório vocal do anu branco (Guira guira).
A família Tyrannidae, como o próprio nome pode nos dar ideia, é um espécie que defende fortemente seu território sendo bastante brigões contra indivíduos da mesma ou de espécies diferentes (SICK, 1997). Vivem em agrupamentos com poucos indivíduos (3 a 5 indivíduos em média observados nos dias de amostragem deste estudo). Esta família também é a mais numerosa dentre as aves do Brasil (CBRO, 2014). Segundo Ferraz (2008), a espécie M. cayanensis apresentou-se bastante abundante no campus da Universidade Federal do Pará, corroborando com o status de espécie comum. Já o período reprodutivo da espécie se distribuiu ao longo de todo o ano a partir da observação de ninhos tanto no segundo semestre de 2014 como no primeiro de 2015 (Novembro e Abril, respectivamente).
Acredita-se que essas condições mencionadas no parágrafo anterior atuem como uma pressão para o bentevizinho utilizar uma estratégia comportamental que pudesse garantir recursos alimentares e reprodutivos nessas áreas em que vivem várias espécies de Tiranideos ocupando nichos semelhantes e vários Myiozetetes cayanesis ocupando o mesmo nicho. Logo, a estratégia comportamental para contornar este problema, foi justamente o desenvolvimento de um agressivo comportamento de defesa territorial, como observado em seu canto e na maioria dos chamados que são voltados para este contexto comportamental, em especial, o chamado agonístico que foi observado quando o bentevizinho atacava agressivamente indivíduos da mesma espécie ou de espécies diferentes, como o Ramphastos vitelinus que de alguma forma ameaçavam seu território principalmente quando estavam próximos a áreas com ninhos de M. cayanesis.
Dentre as formas de comunicação vocal em aves podemos ter 2 casos para comparar com o repertório do Bentevizinho, dentre eles estão: espécies com grande repertório vocal e canto de reconhecimento especifico inato (não aprendido); espécies com pequeno repertório vocal e canto de reconhecimento aprendido.
No primeiro caso, a espécie anu preto (Crothophaga ani) é um ótimo modelo espécie de avifauna pertencente aos não passeriformes e que apresenta um canto inato, porém possui uma larga complexidade social no que se relaciona a seu complexo sistema cooperativo de reprodução, sendo esta complexidade social responsável para explicar os 11 chamados diferentes e um chamado grupal descrito para esta espécie que corrobora com a hipótese da complexidade social (GRIEVES et al., 2015). Logo, o repertório vocal do primeiro caso é considerado um repertório grande em comparação com o repertório vocal do bentevizinho, pois Crothophaga ani apresenta laços sociais envolvidos nesse sistemas cooperativo de reprodução que favorecem esse a complexidade no sistema de comunicação desta espécie.
No segundo caso, podemos citar o beija-flor-cinza (Aphantochroa cirrochloris) pertencente a família Trochilidae, como um modelo de ave que apresenta um canto aprendido e repertório vocal pequeno A. cirrochloris apresentou apenas 3 chamados em seu repertório vocal e todos voltados para a defesa de território (FERREIRA, 2006). Embora o beija-flor-cinza apresente o canto aprendido e o Myiozetetes cayanensis apresente o canto inato, essas duas espécies compartilham uma característica em comum que é a territorialidade. Tanto a família Trochilidae como a Tyrannidae apresentam a característica defesa territorial contra indivíduos, independentemente do tamanho físico, pois são tão brigões que atacam indivíduos de espécies diferentes e também os que pertencem a mesma espécie (SICK, 1997).
Os Aphantochroa cirrochloris como a maioria dos beija-flores, são geralmente indivíduos mais solitários podendo ser encontrados de um a dois indivíduos no inicio do período reprodutivo, e no fim deste somente a mãe cuida da prole (SICK, 1997). Quando se encontram em casal, defendem seu território de forma separada com macho defendendo o território longe da fêmea (Cody,1968).
Já o bentevizinho apesar de não ser uma espécie que vive em grupos grandes, como o anu branco (Guira guira), ele vive em grupos um pouco maiores do aquele com apenas dois indivíduos de Aphantochroa cirrochloris, pois o bentevizinho foi encontrado geralmente em grupos de 3 a 5 indivíduos nesse presente estudo. Além do mais, os casais de bentevizinhos defendem o território e cuidam da prole juntos (SICK, 1997). Essa defesa de território em conjunto foi observada de uma forma interessante, onde o grupo familiar cantava em trio (3 indivíduos cantando simultaneamente), fazendo o display de defesa territorial juntos e passando próximo a fonte de emissão sonora do play back como apontado por (FERRAZ, 2008).
Por conta dessas formas comportamentais de A. cirrochloris e M. cayanensis serem diferentes, a hipótese da complexidade social pode explicar muito bem essa diferença no número de chamados, pois o bentevizinho atua muito mais em conjunto na forma de defender território e na criação da prole do que beija-flor-cinza, logo o bentevizinho apresenta relações sociais mais complexas quando comparado com Aphantochroa cirrochloris, relações essas que acabam refletindo
em um mecanismo de comunicação mais complexo do bentevizinho que apresenta mais chamados do que A. cirrochloris.
A vocalização de dentro do ninho apresenta um contexto semelhante ao encontrado por (DRAGONETTI et al., 2013), pois os indivíduos utilizam apenas durante o período reprodutivo como foi observado que um indivíduo que fica dentro do ninho se comunica com um outro que fica fora do ninho com este mesmo tipo de vocalização, indicando um contexto como se fosse o de alternância de quem fica no ninho para que o outro que esta fora fique de vigilância. E este chamado é fundamental para um melhor sucesso na sobrevivência da prole, pois é muito mais vantajoso para aquelas espécies em que casal cuida desta prole e defende o território juntos quando comparados aquelas espécies em que um único indivíduo cuida da prole, pois o gasto de energia acaba sendo dividido entre o casal ao longo do desenvolvimento da prole (SICK, 1997).
Segundo Gill (2007), o número de vocalizações distintas para a maioria das aves, varia de 5 a 14 vocalizações. Como o Myiozetetes cayanensis apresentou um repertório com 9 vocalizações distintas, ele é considerado uma espécie com repertório vocal intermediário entre 5 e 14 vocalizações diferentes. Mas é importante ressaltar que o bentevizinho corrobora com a hipótese da complexidade social, pois ele não é uma espécie muito sociável com vários indivíduos da mesma espécie, o que acredita-se refletir em sua complexidade de comunicação, que apresentou apenas 4 chamados diferentes.
CONCLUSÃO:
Myiozetetes caynensis apresenta um canto de reconhecimento especifico estereotipado e o esperado para espécies da subordem Tyranni que deveria apresentar um repertório vocal restrito, apresentando 4 categorias chamados distintos. Entretanto registramos 9 tipos diferentes de vocalizações das 14 encontradas por (FERRAZ, 2008), incluindo uma vocalização nova (chamado agonistico), desta forma o repertório da espécie apresenta 15 vocalizações registradas, número que pode aumentar durante mais anos de observações.
Com relação aos contextos comportamentais, observou-se que o bentevizinho corroborou com a hipótese da complexidade social, pois esta espécie que mantém poucas relações sociais com outros indivíduos da mesma espécie, uma vez que os ataca para expulsá-los de seu território, apresentando pouca complexidade em seu sistema de comunicação que possui apenas 4 chamados com contextos comportamentais definidos.
Apesar de apresentar apenas 4 categorias de chamados, ele possui uma alta capacidade de combinar estes chamados e o canto, mantendo uma única função biológica, além de uma capacidade de mudar a sequencias das notas do chamado territorial, produzindo uma gama de variantes desse chamado, porém mantendo também a função biológica deste chamado. No caso da estratégia de vida adotada pelo bentevizinho que é a de defender fortemente o território, essa mudança na ordem das notas lhe garante um leque de vocalizações variantes do chamado territorial o que lhe fornece uma vantagem na intimidação e/ou expulsão de uma potencial ameaça a seu território.
REFERÊNCIAS BIBLIOGRÁFICAS
CODY, M. L. 1968. Condor 70:270-71. (território). COMITÊ BRASILEIRO DE REGISTROS ORNITOLÓGICOS (CBRO). Listas das Aves do
Brasil. 11.ed. Florianópolis. CBRO, 2014. 41 p. DRAGONETT, MARCOS, et al. The Vocal Repertoire of the Eurasian Stone-Curlew (Burhinus
oedicnemus). v125, n.1, p.34–49, 2013. FARABAUGH, S. M. (1982). The social and ecological significance of duetting. In: Kroodsma,
D. E. & Miller, E. H. (eds). Acoustic Communication in birds (Vol. 2). New York: Academic press.
FERRAZ, Dnilson Oliveira. Estudo do Bentevizinho-de-asa-ferrugínea Myiozetetes cayanensis (AVES, TYRANNIDAE). 2008. 88p. Monografia (graduação em bacharelado em ciências biológicas)- Faculdade de Biologia, Universidade Federal do Pará, Belém, 2008.
FREEBERG, T. M. 2006. Social complexity can drive vocal complexity. Psychological Science 17: 557–561.
GILL, F. B. 2007.Ornithology.W. H. Freeman,NewYork,NY. FANDIÑO-MARIÑO, J.H. A comunicação sonora do Anu-branco Guira guira. Campinas:
Editora Unicamp, 1989. 302p. GRIEVES, L.A. et al. (2015). Vocal Repertoire of Cooperatively Breeding Smooth-billed Anis.
J. Field Ornithol. V. 86, n. 2, p. 130-143, Jan. 2015. MENDOZA, A.M. & ARCE, M.I. (2012). Aproximación al impacto de la pertubación urbana en
las vocalizaciones de Pitangus sulphuratus (TYRANNIDAE) en Santiago de Cali, Valle del Cauca (Colombia). Revista de ciências. 16: 19-29.
RIDGELY, R.S & TUDOR, Guy. The birds of the South America: The Suboscine Passerine. ed. 1. Austin: University of the Texas, 1994.
SEPLAN. Novo Município Paraense, N°33. Santa Bárbara do Pará. Governo do Pará. Belém. CEJUP. 1993.
SICK, Helmut. Ornitologia Brasileira. Rio de Janeiro: Nova fronteira, 1997. SILVA, M. L. (1995). - Estereotipia e versatilidade nos cantos das aves: os padrões de canto
em sabiás e outras aves. Anais de Etologia 13: 133-147. SILVA, M. L. & VIELLIARD, J. M. E. A aprendizagem vocal em aves: evidências
comportamentais e neurobiológicas. In: Grauben Assis, Regina Brito e William Lee Martin (Org.). Estudos do Comportamento II. Belém: Editora da UFPA, (2011).
VIELLIARD, J. M. E. A diversidade de sinais e sistemas de comunicação sonora na fauna brasileira. In: Seminário Música Ciência Tecnologia, 1, 2004. São Paulo. Acústica Musical. São Paulo: USP, 2004. p. 145-152.
VIELLIARD, J. M. E. & SILVA, M. L. da. A Bioacustica como ferramenta de pesquisa em Comportamento animal. In: Estudos do Comportamento II. Grauben Assis, Regina Brito e William Lee Martin (eds.). Belém: Editora da UFPA, 2007.
VIELLIARD, J. M. E. & SILVA, M. L da. Bioacústica- Bases teóricas e regras práticas de uso em ornitologia. In: MATTHER, Sandro Von; et.al. Ornitologia e conservação: ciência aplicada, técnicas de pesquisa e levantamento. ed. 1. Rio de Janeiro: Technical books, 2010. cap. 13, p. 315-326.
DIFICULDADES
A principal dificuldade do estudo foi conseguir observar os indivíduos por muito tempo, pois geralmente eles conseguem se esconder em árvores ou vão embora do local sem que se pudesse encontrá-los novamente, o que prejudicou as filmagens dos contextos comportamentais dos mesmo que foram feitas no período de março a junho.
PARECER DO ORIENTADOR: O aluno Felipe Castro teve um desempenho excelente, é motivado para observar aves em trabalho de campo de alto grau de dificuldade e escreve relativamente bem, o que é uma qualidade rara.





























