Sistema Nervioso
SISTEMA NERVIOSO.
(Resumen clases)
Asignatura de Neurofisiologa
Carrera de Kinesiologa
Docente Dr. Joel Snchez C.
El estudio del sistema nervioso, al menos en los constituyentes
anatmicos del encfalo, data de pocas muy antiguas en las que los
constituyentes de la masa cerebral despertaron la curiosidad de
aquellos incipientes cientficos. Algunos nombres designados, por
ejemplo, por los griegos, se conservan todava, aunque la tendencia
moderna de la neuroanatoma es la nomenclatura funcional de ms fcil
comprensin.
I.- UNA ESQUEMATIZACIN TIL.-
Si se tratara de resumir en su ms breve expresin el estudio del
sistema nervioso, habra que hacer una distincin entre la morfologa,
es decir el aspecto anatmico, y el funcionamiento, la fisiologa del
sistema.
Hay que precisar de inmediato que ambas ramas, la neuroanatoma y
la neurofisiologa, deben terminar siendo estudiadas en un todo
integrado que permitir la comprensin global de este importante
sistema de relacin del organismo humano. Sin embargo, podemos
separar sus campos brevemente para facilitar la posterior
profundizacin del tema.
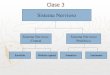
Anatoma.- El sistema se divide morfolgicamente en Sistema
Nervioso Central (SNC) y Sistema Nervioso Perifrico (SNP). Las
estructuras que integran a ambos segmento pueden ser visualizadas
con relativa facilidad en una diseccin completa del Sistema. El SNC
comprende los hemisferios cerebrales, el tronco enceflico (tallo) y
la mdula. En tanto que el SNP incluye a todos los cordones de
nervios que emergen o llegan al SNC sacando o llevando impulsos
nerviosos. Estos cordones nerviosos comprenden globalmente los 12
pares de nervios craneales que se encargan de inervar cabeza,
cuello y regin toraco-abdominal, y los 31 pares de nervios raqudeos
que emergen de la mdula para inervar desde la regin del trax y el
abdomen hasta la punta de los pies.
Fisiologa.- La fisiologa no es posible visualizarla en una
diseccin. Corresponde al comportamiento, a la forma como va a
funcionar el sistema para cumplir su cometido. Desde este punto de
vista el sistema nervioso se divide en Sistema Nervioso Sensitivo
(SNS) y Sistema Nervioso Motor (SNM). El primero est formado por
neuronas sensitivas o aferentes que se encargan de captar la
informacin que proporcionan los estmulos del medio tanto interno
como externo para conducirla hasta el SNC. Este sistema, el
sensitivo, puede ser conciente o inconsciente, segn si la
sensibilidad es captada por la conciencia del individuo, es decir
se hace percepcin, o si esa sensibilidad llega slo hasta ciertas
regiones del SNC donde es procesada sin que el individuo tenga
nocin de ella. El SNM lo forman las neuronas motoras o eferentes
encargadas de conducir la respuesta del SNC ante los estmulos del
medio (interno o externo). Se subdivide en Sistema Nerviosos Motor
Voluntario (SNMV) y Sistema Nervioso Motor Autnomo (SNMA) llamado
tambin neurovegetativo. El primero, el SNMV, trasmite las ordenes
generadas por la voluntad conciente del individuo hasta los
efectores que corresponden a msculo voluntarios esquelticos de
fibra estriada (la clula muscular voluntaria presenta en su
citoplasma lneas longitudinales, miofibrillas, que le dan el nombre
de miocito estriado). Por su parte el SNMA inerva musculatura lisa
(miocitos sin miofibrillas aparentes) involuntaria, adems de la
musculatura del miocardio que, siendoestriada, es sin embargo
autnoma. Este sistema nervioso, el autnomo, se subdivide en
simptico y parasimptico de accin antagnica, mientras uno estimula
un rgano, el otro inhibe.
II.- ORIGEN EMBRIONARIO DEL SISTEMA NERVIOSO.-
Se seala a continuacin, tal como se hizo en clases, un breve
recordatorio de embriologa.
Luego de la anfimixis que es la fusin de los gametos femenino y
masculino para formar el cigoto, es decir la primera clula del
nuevo individuo comienza un proceso intenso de mitosis que, de
manera general, se puede dividir en 7 etapas: la segmentacin,
compactacin, blastulacin, anexos embrionarios, gastrulacin,
neurulacin y organogensis.
La segmentacin es un proceso de aumento extraordinario del nmero
de clulas todava indiferenciadas que forman la mrula. Se contina
con la compactacin que implica diferenciar un medio interno del
embrin del medio externo por medio de capas de clulas trofoblsticas
destinadas a formar el corion y el llamado macizo celular interno
que dar origen al individuo propiamente tal. La blstulacin forma a
la blstula que es una estructura que en su interior presenta ya una
cavidad y como tal llega a implantarse en las paredes del tero. Los
anexos embrionarios son la formacin de la placenta y el cordn
umbilical. La gastrulacin es cuando el embrin presenta ya tres
capas, ecto, meso y endodermo, y la neurulacin es lo que ms
interesa en este resumen y que corresponde a la aparicin de sistema
nervioso central. La organognesis es el desarrollo de todos los
otros rganos y sistemas del nuevo individuo.
Desarrollo del Sistema Nervioso.
Este sistema es uno de los primeros en comenzar su formacin
luego de la blastulacin embrionaria que posibilita el implante del
huevo en la pared uterina. La migracin de las clulas del macizo
celular interno para formar el amnios y el saco vitelino conduce a
la aparicin de un embrin bilaminar donde es posible distinguir la
capa celular denominada ectodermo (epiblasto) formada del piso del
amnios, y el llamado endodermo (hipoblasto) que nace del techo del
saco vitelino.
El sistema nerviosos central (SNC) se deriva de la porcin dorsal
media engrosada del ectodermo por delante de la fosita primitiva,
la cual, es conocida como placa neural, la que aparece al inicio de
la tercera semana de la concepcin (o sea 5ta. semana de gestacin en
el lenguaje obsttrico)
Pero para que se construya la placa neural es necesario recordar
que las clulas del epiblasto (ectodermo) convergen hacia la lnea
media del disco embrionario formando un surco, es decir la lnea
primitiva delimitando un eje longitudinal de simetra bilateral
alrededor del cual se alinearn las estructuras embrionales y sus
rganos. Desde este momento, el embrin tendr una regin rostral
(ceflica) y caudal (cola) as como lado izquierdo y derecho; y
superficies dorsal (atrs) y ventral (adelante).
Saco gestacional observado al 16avo da post concepcin
(ecocografa del saco de 7 milmetros de dimetro).
El extremo rostral ceflico de la lnea primitiva termina en una
pequea fosita rodeada por una elevacin de clulas en circulo el
ndulo o nodo primitivo.
Este ndulo marca el sitio donde las clulas del epiblasto se
invaginan para formar la capa media (mesodermo) que se sita entre
el epiblasto e hipoblasto (disco bilaminar) denominndose a la
formacin de las tres capas o disco trilaminar como gastrulacin.
Casi de manera paralela con la gastrulacin que agrega el mesodermo
creando un embrin trilaminar a los 16 das de desarrollo
intrauterino comienza la migracin de clulas del ectodermo dorsal
para formar la placa neural que experimenta luego levantamientos
(pliegues) del neuroectodermo que crean un surco central que luego
se convierte en un tubo, el tubo neural, al unirse los pliegues
entre ellos ms o menos a las tres semanas de crecimiento del
embrin.
Durante el proceso de gastrulacin una estructura llamada
notocorda se origina de las clulas mesodrmicas cilndricas la cual
se extiende a lo largo de la lnea media (rostral y caudalmente),
esta estructura induce a que clulas del ectodermo que las recubre
se diferencien a clulas neurales precursoras, las cuales se
organizan en una estructura llamada placa neural. Este proceso que
da origen a esta placa se llama neurulacin.Las clulas de los
mrgenes laterales de la placa neural crecen y se acumulan formando
los pliegues neurales entre los cuales se encuentra el surco neural
(Foto #2). Todos estos elementos se encuentran baados por el lquido
amnitico que les provee nutricin hasta que se establezca el sistema
vascular primitivo.
Imgenes de la placa neural.
Tal como se vio en clases, de este tubo neural se formar el
Sistema Nervioso Central (SNC). Por lo tanto, la mitad superior ser
el sitio de desarrollo del encfalo (hemisferios cerebrales) y del
tronco enceflico, y la mitad inferior dar origen a la mdula. Hay
que destacar que tanto las paredes celulares del tubo, como la
cavidad central, van a jugar un papel primordial en la anatoma y la
fisiologa del SNC.
Embrionariamente el tubo neural en su mitad superior se
subdivide, para su estudio, en tres regiones, cada una con un
desarrollo diferente de sus paredes celulares siendo ellas el
presencfalo, el mesencfalo y el rombencfalo.
Cavidades embrionarias del tubo neural
La primera cavidad superior del tubo, llamada como se dijo
prosencfalo, tendr un aumento espectacular de las clulas de sus
paredes alcanzando un volumen de gran tamao que obliga a varios
repliegues sobre s misma dando nacimiento posteriormente a los
hemisferios cerebrales. Este prosencfalo, al desarrollarse en el
embrin maduro, se convierte en las estructuras llamadas telencfalo
y diencfalo. Las otras dos regiones embrionarias inferiores de la
mitad superior del tubo neural, el mesencfalo y el rombencfalo,
pasarn a la etapa madura del sistema formando tambin estructuras
propias. El mesencfalo embrionario para al estado adulto con el
mismo nombre, en tanto que el rombencfalo dar origen al metencfalo
y el mielencfalo, este ltimo limitando con la mdula.
En resumen, las estructuras finales del sistema nervioso ya
desarrollado, sern el telencfalo, el diencfalo, el mesencfalo, el
metencfalo y el mielencfalo.III.- CONDUCCIN NERVIOSA.-
El sistema nervioso esta formado no slo por tejido nervioso
(neuronas) sino que tambin por clulas no nerviosas llamadas gliales
(clulas de neuroglia) que cumplen variadas funciones de sostn a las
neuronas, y que se vern ms adelante durante el desarrollo del
curso.
Se considera a la neurona como la clula ms larga del organismo
teniendo, adems, la capacidad de conducir impulsos nerviosos bajo
la forma de conduccin elctrica, funcin centrada principalmente en
las membranas neuronales.
Potenciales bioelctricos.- Se le llama as a la diferencia
elctrica que se establece en la membrana de la neurona entre su
cara interna y la superficie exterior y que permite la conduccin
del impulso nervioso. En l participan de manera activa los cationes
Na+ y K+
El sodio es un catin de vida normal extracelular, es decir se
encuentra ms concentrado en el medio que rodea a la clula que en
interior, en tanto que el catin potasio se ubica a la inversa, es
decir est ms concentrado dentro del citoplasma neuronal que en el
exterior. La concentracin aproximada para ambos cationes es: Na+
137 meq. (miliequivalentes) fuera de la membrana y 10 meq. dentro
de la neurona. Para el K+ es de 141 meq. dentro de la clula y de
slo 4 meq en el medio extracelular.
Cuando la neurona esta en estado de reposo, es decir no est
conduciendo impulsos, su membrana se encuentra polarizada, siendo
positiva por fuera al acumularse los cationes sodios contra la cara
membranal externa, y negativa por dentro debido a la presencia de
aniones (iones de carga elctrica negativa), entre otros cloruros
(Cl-), fosfatos (PO4=), sulfatos (SO4=) y iones bicarbonato
(HCO3-). Es lo que se conoce como el Potencial de Reposo con un
valor aproximado de -60 milivoltios.
Si la neurona es estimulada por alguna forma de energa (lumnica,
qumica, sonora, temperatura, entre otras) la membrana responde en
el sitio del estmulo despolarizando la membrana en ese lugar. Se
produce la apertura de los canales de difusin del Na+ que ingresan
masivamente a la clula en tanto que en menor cantidad se produce la
salida de los iones potasio. Se crea as el llamado Potencial de
Accin en el que la clula lleva su carga a 0 (cero) en ese lugar
pudiendo incluso hacerse por breves momentos hasta ms positiva
dentro que fuera de la membrana, lo que se conoce como el potencial
de espiga o sobreexcitacin. El estmulo debe tener un potencial
energtico igual o superior a 7 milivoltios (estmulo umbral) para
que la neurona responda. De lo contrario el impulso no se va a
propagar y no habr conduccin nerviosa. Si el estmulo es suficiente,
la neurona responde a su mxima capacidad. Es lo que se conoce como
la ley del todo o nada del Sistema Nervioso.
Finalmente, la velocidad del impulso depender del grosor de la
fibra y de su condicin de mielnica o amielnica (con o sin vaina de
mielina). La mayor velocidad la presentan las fibras ms gruesas y,
sobre todo, las fibras mielnicas que conducen mediante el fenmeno
denominado conduccin saltatoria.
IV.- SINAPSIS.
Bsicamente se designa como sinapsis a un tipo de enlace que se
establece entre clulas con el fin de comunicarse informacin. De
manera particular este trmino se aplica a la forma como el sistema
nervioso transmite y propaga la informacin nerviosa tanto entre
neuronas como de stas a los efectores. En fenmeno, en realidad, es
ms amplio que el concepto generalizado de que la sinapsis slo sirve
para asegurar la continuidad de los impulsos nerviosos a travs de
la red neuronal que recorre los tejidos. En rigor, la sinapsis es
el mecanismo que interconecta a las neuronas entre s y con los
efectores, pero el resultado puede ser muy variado dependiendo de
los objetivos del fenmeno fisiolgico. Esto quiere decir que la
sinapsis puede servir para asegurar la transmisin de la informacin,
para modularla e incluso para detenerla de acuerdo a la regulacin
que hace la propia clula sobre la funcin como se ver ms
adelante.
Tipos de sinapsis.
En el caso neuronal se conocen dos formas de comunicacin: la
sinapsis elctrica y la sinapsis qumica.
A.- Sinapsis elctrica.-
En este caso hay un paso directo del potencial bioelctrico de
una neurona a otra a travs de las llamadas gap junctions, uniones
comunicantes. El flujo de la corriente es continuo sin retraso
sinptico, no existiendo variaciones del potencial que se trasmite
inalterado entre clulas.
Ha sido posible conocer, a travs de la microscopa electrnica, la
disposicin de las membranas pre y post sinpticas en este tipo de
uniones. Existen conjuntos proteicos en las membranas dispuestos de
manera simtrica conteniendo seis subunidades polipeptdicas llamadas
conexinas y que rodean a un canal central que coincide en ambas
membranas. Estas estructuras reciben el nombre de conexones y estn
ensambladas de tal forma que permiten el paso, por el canal
central, de molculas hidrosolubles e iones entre una clula y
otra.
En la sinapsis elctrica el espacio que separa a ambas membranas
es apenas de dos o tres nanmetros, comparado con la sinapsis qumica
en la que la cavidad sinptica puede alcanzar hasta los 18 o 20
nanmetros. Los canales de conexones pueden abrirse o cerrarse
dependiendo de las necesidades del fenmeno y obedecen a los
potenciales bioelctricos de membrana y las concentraciones de Ca++
e H+ en ambas clulas que establecen este tipo de sinapsis.
Entre las caractersticas especiales de las sinapsis elctricas
est que la transmisin puede hacerse en ambos sentidos, hacia y
desde el soma neuronal, aunque puede haber una transmisin ms rpida
en un sentido que en otro. Por ltimo, hay que sealar que este tipo
de sinapsis es caracterstica en el SNC, en el tejido muscular liso,
en ciertos reflejos que requieren de una transmisin rpida y, sobre
todo, en rganos que necesitan una sincronizacin simultanea
expandida en estereo, como por ejemplo el tejido del miocardio que
responde al unsono a los estmulos del tejido nodal.
B.- Sinapsis qumica.
Es la ms numerosa en el sistema nervioso. Se caracteriza porque
los terminales axonales se encuentran separados de la membrana post
sinptica por un espacio, la cavidad sinptica, que puede medir hasta
20 nanmetros de superficie, como se seal ms arriba. Adems de
presentar retraso sinptico, requiere de la presencia de sustancias
qumicas llamadas neurotransmisores que se vierten a la cavidad
sinptica con el fin de actuar sobre la membrana de la clula
siguiente, ya sea para asegurar la continuidad del potencial
bioelctrico, en algunos casos para modularlo o simplemente para
inhibirlo segn el fenmeno requerido.
Entre las propiedades de este tipo de sinapsis esta su
dependencia del Ca++ que al ingresar por difusin a los terminales
nerviosos, determinan la cantidad de neurotransmisores que se
liberan al medio. A su vez, la apertura de los canales de difusin
del Ca++ es dependiente de voltaje, es decir depende de los
potenciales de accin que llegan a los botones sinpticos.
Si bien los tipo, cantidades y tiempo de accin de los
neurotransmisores liberados desde la membrana presinptica tiene
gran importancia, ser la membrana post sinptica la que determinar
la accin del neurotransmisor a travs de los receptores que posee
para su captacin. Es por eso que un mismo neurotransmisor puede
producir diferentes efectos en la membrana post sinptica, sea sta
una neurona o un efector muscular, es decir puede provocar un
potencial excitatorio, denominado PIPS (Potencial Excitatorio Post
Sinptico) o una inhibicin de la membrana post sinptica evitando la
transmisin del potencial bioelctrico. En este caso ocurre un PIPS
(Potencial Inhibitorio Post Sinptico).
En el primer caso, el del PEPS, la accin del neurotransmisor ser
provocar un estmulo umbral en la membrana post sinptica que se
convertir en un potencial de accin propagado que puede ser ms rpido
o ms lento segn lo determine el tipo de canales de difusin que se
abran. En el segundo caso, cuando los receptores post sinpticos as
lo determinan y se provoca un PIPS, la accin del neurotransmisor
ser activar receptores que provocarn una hiperpolarizacin de la
membrana post sinptica hacindola refractaria al estmulo y, por lo
tanto, frenando la transmisin del impulso nervioso.
Neurotransmisores peptdicos y no peptdicos.- Los trasmisores
peptdicos (aquellos que presentan cadenas proteicas de aminocidos),
son sintetizados en el soma neuronal, en RER (retculo endoplsmico
rugoso) y de ah enviados a los terminales sinpticos en vacuolas
pulstiles, previo almacenamiento en el aparato de Golgi. Los no
peptdicos se sintetizan en el mismo terminal con participacin
activa de enzimas especficas, siendo almacenados en el
compartimiento endosmico sinptico en el mismo terminal desde donde
se liberan vesculas con el neurotransmisor que se fusionan con la
membrana del botn sinptico.
Control de los neurotransmisores.- Ya se dijo que los receptores
de la membrana post sinptica juegan un papel fundamental en la
regulacin de la transmisin. Sin embargo, existen formas de
controlar de manera directa al neurotransmisor con el fin de
regular la cantidad y el tiempo de su permanencia en la cavidad
sinptica y, por lo tanto, controlar el efecto sobre la clula post
sinptica. Generalmente los neurotransmisores peptdicos son
degradados por enzimas proteolticas que hidrolizan los enlaces
peptdicos de los aminocidos. Esto ocurre lejos de la membrana post
sinptica y sus componentes no se reciclan. En cambio aquellos no
peptdicos se degradan en la misma cavidad sinptica y sus
componentes suelen ser rescatados hacia la neurona presinptica por
transporte activo y en muchos casos reciclados para sintetizar ms
neurotransmisor. Tal es el ejemplo del neurotransmisor
acetilcolina, que se ver en detalle ms adelante, que es degradado
por la acetilcolinesterasa.
V.- UNIN NEUROMUSCULAR.-
La transmisin nerviosa no slo se efecta entre neuronas de la
manera como ya se describi ms arriba, es decir mediante sinapsis
elctrica o qumica, sino que tambin se realiza sobre el efector que
es el tejido muscular. Este tejido, como se recordar, est formado
por dos tipos de clula: clulas musculares lisas y estriadas. Las
clulas musculares lisas son tpicas del tejido muscular autnomo
manejado por el S.N.M. Autnomo (neurovegetativo) tanto simptico
como parasimptico, en tanto que los miocitos estriados son
caractersticos del tejido muscular voluntario (con excepcin del
tejido muscular cardiaco que, siendo fibras estriadas, es sin
embargo autnomo). Este tejido muscular voluntario es manejado por
el S.N.M. Voluntario.
El S.N.M. Autnomo descarga sobre el efector muscular dos tipos
de neurotransmisores: adrenalina (derivados catecolamnicos) en el
caso del S.N.M.A. Simptico, y acetilcolina en el caso del S.N.M.A.
Parasimptico.
El S.N.M. Voluntario descarga sobre el tejido muscular
voluntario acetilcolina. Sus terminaciones son, en este caso, slo
colinrgicas. Los receptores presentes en la membrana postsinaptica
de los miocitos, tanto en el msculo autnomo como voluntario se
describen a continuacin y son receptores sensibles a la
acetilcolina (colinrgicos) o a la adrenalina (adrenrgicos).
RECEPTORES COLINRGICOS.-
Estos receptores para la acetilcolina presentes en la membrana
post sinptica tanto de neuronas como de miocitos, son dependientes
de voltaje. Son macromolculas transmembranales con un poro
generalmente central para permitir el paso de iones pudiendo ser
altamente selectivos.
La accin de la acetilcolina se efecta tanto sobre la musculatura
estriada esqueltica, como tambin en otros sectores del SNC. La
acetilcolina resulta de la unin del cido actico, proveniente en su
mayora de la degradacin por gliclisis de la glucosa y que origina
al cido pirvico que luego es convertido en cido actico por
descarboxilacin oxidativa del piruvato.
La colina, por su parte, es un alcohol nitrogenado
(trimetilaminocefanol) que proviene de la fosfatidilcolina, un
fosfolpido aportado en su mayora de manera exgena, es decir por los
alimentos. La enzima acetilcolintransferasa une a ambas molculas
por deshidratacin para formar la acetilcolina.
Los receptores a la acetilcolina se dividen en los receptores
nicotnicos y los muscarnicos.Receptores nicotnicos Este receptor a
la acetilcolina es un pentmero proteico formado por cinco
subunidades: 2 alfa, 1 beta, 1epsiln-gamma y 1 delta. La unin de la
acetilcolina con estos receptores se efecta con las dos subunidades
alfa, en las interfaces alfa-gamma y alfa-delta. El punto de unin
con el ligando es la regin N-terminal de las subunidades alfa. La
activacin del receptor nicotnico se produce cuando dos molculas de
acetilcolina se unen a las subunidades alfa del pentmero proteico,
lo que induce la apertura del poro o canal inico permitiendo el
paso de los cationes Na, K y Ca, bloquendose la entrada de otro
tipo de cationes o aniones. El tiempo de accin de la acetilcolina
es de uno o dos milisegundos, despus de los cual el neurotransmisor
se disocia del receptor cerrndose el canal. A pesar de que la
acetilcolina permanezca en la cavidad sinptica ms all del tiempo
sealado, el poro permanece cerrado producindose una
desensibilizacin cuyo mecanismo no est an bien aclarado.
Se han identificado al menos dos tipos de receptores nicotnicos:
Los N1 que estn presentes en los ganglios del sistema nervioso
autnomo, y los N2 presentes en la placa neuromuscular. Estos
receptores se caracterizan por una respuesta rpida que significa la
depolarizacin de la membrana postsinptica cuando se activan los
canales de sodio. Se le considera un receptor ionotrpico. Existen
varios antagonistas de la acetilcolina para los receptores
nicotnicos, entre ellos la alfa-bungarotoxina que est presente en
el veneno de las vboras y que se une de manera irreversible a las
subunidades alfa de estos receptores.
Agonistas de los receptores nicotnicos.- Son compuestos de
colina que estn unidos a otros compuestos carboxlicos no siendo muy
importantes. Tambin la nicotina que le da el nombre a estos
receptores.
Antagonistas de los receptores nicotnicos.- Existen algunos
frmacos que inhiben a estos receptores en los ganglios
(mecamilamina), pero su accin generalizada como inhibidores de la
fibra preganglionar parasimptica y simptica los hace poco prcticos
prefirindose usar bloqueadores beta-adrenrgicos (por ejemplo en el
tratamiento de la hipertensin arterial).
Los receptores nicotnicos neuromusculares son bloqueados por la
tubocurarina (curare) y la succinilcolina, ambos bloqueadores que
impiden la accin de la acetilcolina sobre los msculos.
Debido a la accin de la acetilcolina en la memoria y el
aprendizaje, se ha relacionado el Alzheimer con la prdida de
receptores colinrgicos nicotnicos a nivel del SNC. Tratamiento con
Nicotina, agonista de la acetilcolina, tiene efectos positivos en
esta enfermedad, pero con efectos colaterales. Hoy se usa un
compuesto nuevo, el ABT-418, que ha surtido un buen resultado como
estimulador de los receptores nicotnicos del SNC en el tratamiento
del Alzheimer.
Receptores muscarnicos.
Son predominantes en el cerebro relacionndose con la memoria y
el aprendizaje. Son glucoprotenas cuyas funciones se realizan por
medio de la protena G (dependiente de GTP, de ah su nombre) de
membrana.
Se conocen cinco subtipos de receptores muscarnicos designados
con la letra M. Ellos son:
Receptor M1, abunda en el encfalo (aumenta el IP3 y calcio)
Receptor M2, abunda en el corazn (disminuye el AMPc)
Receptor M3, se encuentra en el tejido glandular y el msculo
liso
Receptor M4, abunda en el pncreas y el pulmn
Receptor M5, se cree que acta a nivel de las glndulas salivales
y el msculo ciliarLos ms estudiados son el M1 y el M2. Todos estos
receptores son de respuesta ms lenta, es decir son de un PEPS ms
tardo y actan a travs de un segundo mensajero que es el GMPc por lo
que se les denomina como metabotrpico. La respuesta de la clula
ante el estmulo de los receptores muscarinos es inhibir la accin de
la adenilciclasa que se forma como respuesta a los estmulos de
catecolamnicos adrenrgicos, por ejemplo la dopamina. La adicin de
agonistas colinrgicos provoca la disminucin del AMPc formado como
respuesta a los catecolamnicos. El receptor muscarino inhibe a la
adenilciclasa liberando una protena inhibitoria que se encuentra
adherida a GTP. Esta protena inhibitoria compite con la protena G
cuando esta es activada por estimuladores de la adenilciclasa.
Agonistas de los receptores muscarinos.- Su funcin es estimular
la accin de los segundos mensajeros. Entre estos estn metacolina,
carbacol, betanecol, pilocarpina y la muscarina.
Antagonistas de los receptores muscarinos.- Se pueden calificar
como antiespasmdico y parasimpaticolticos. Su accin es antagnica a
las funciones del parasimptico en especial las de tipo secretor
(saliva, lagrimales, bronquiales, gstricas). Entre estos
antagonistas estn:
Atropina y escopolamina. Son antagonistas de los M1 y los M2.
Son antiespamdicos en diarreas y clicos. Son antdoto de los efectos
antocolinestersicos.
Pirenzepina. Antagonista de los M1. Es un anticolinrgico puro
sin efectos colaterales.
RECEPTORES ADRENRGICOS.-
El neurotransmisor de la primera sinapsis del simptico es la
acetilcolina (Ach) y los receptores son nicticos y en la segunda
sinapsis el neurotransmisor es la noradrenalina o norepinefrina que
van actuar en los receptores adrenrgicos, estos pueden ser: Alfa1:
estn en todos los vasos sanguneos, excepto en los coronarios y los
de la musculatura esqueltica. Su estimulacin produce
vasoconstriccin Alfa2: se localizan en la membrana presinptica de
las fibras postganglionares del simptico. Su estimulacin produce
inhibicin de la liberacin del neurotransmisor Beta1: estn solo en
el miocardio. Su estimulacin produce un aumento de la
contractilidad, frecuencia, conduccin y excitabilidad Beta2: en los
vasos coronarios, de la musculatura esqueltica, musculatura lisa
bronquial, tero y rganos digestivos. Su estimulacin produce
relajacin, por lo que produce vasodilatacin coronaria, de la
musculatura esqueltica, tero, broncodilatador y disminucin de la
motilidad intestinalVI.- SISTEMA NERVIOSO CENTRAL.-
La estructura adulta del SNC, como se dijo, comprende de manera
global al encfalo, el tronco enceflico y la mdula, todos los
cuales, repetimos, son capaces de interpretar los estmulos del
medio externo e interno, y elaborar respuesta de acomodacin.
Encfalo.- Se origina del prosencfalo embrionario y comprende al
telencfalo y al diencfalo en el estado adulto. El primero de ellos,
el telencfalo, comprende a los hemisferios cerebrales, al
rinencfalo y a los ncleos profundos o basales.
Hemisferios cerebrales.-
Como se seal en clases, en la especie humana el desarrollo
(intensa mitosis) de las capas neuronales del extremo superior del
prosencfalo es de tal magnitud, que la masa cerebral debe doblarse
varias veces sobre s misma para ser comprendida dentro de la caja
craneana. Esto origina surcos, las cisuras y circunvoluciones,
hendiduras profundas, medianas y menores, que han permitido una
geografa de las funciones que se desarrollan en esta masa
enceflica. De manera muy resumida, ya que esta materia pertenece a
la nauroanatoma, las grandes cisuras, central (Rolando), lateral
(Silvio) y la parieto-occipital, han delimitado cuatro grandes
fracciones cerebrales llamadas lbulos y se designan con el nombre
del hueso craneal debajo del cual se encuentran.
Hay que recordar tambin que las funciones conscientes de la
persona, sensibilidad consciente (percepcin) y respuesta motora
voluntaria, se localizan principalmente en la llamada sustancia
gris de las masa cerebral, ms conocida como corteza cerebral.
A.- Corteza Cerebral.
La corteza cerebral forma un revestimiento completo de los
hemisferios cerebrales. Est compuesto por sustancia gris y contiene
aproximadamente 10.000 millones de neuronas. El espesor vara de 1,5
a 4,5 mm. Es ms gruesa sobre la cresta de una circunvolucin y ms
delgada en la profundidad del surco. La corteza cerebral al igual
que la sustancia gris de cualquier otro sitio del SNC consiste en
una mezcla de clulas nerviosas, fibras nerviosas, neuroglia y vasos
sanguneos.
CLULAS NERVIOSA DE LA CORTEZA CEREBRAL.-
Clulas piramidales: llevan ese nombre por su forma. La mayora
tienen un dimetro de 10 a 50 nm pero tambin hay clulas piramidales
gigantes conocidas como clulas de Betz cuyo dimetro puede ser hasta
de 120 nm. Se encuentran en la circunvolucin precentral motora.
Los vrtices estn orientados hacia la superficie pial de la
corteza. Una gruesa dendrita va hasta la piamadre y emite ramas
colaterales. Las neuritas poseen espinas dendrticas para las
sinapsis con otras neuronas. El axn nace de la base del cuerpo
celular y termina en las capas ms profundas o entra en la sustancia
blanca como fibra de proyeccin, asociacin o comisural.
Clulas estrelladas: a veces llamadas granulosas, son pequeas, 8
mm y tienen forma poligonal. Poseen mltiples dendritas y un axn
relativamente corto que termina en una neurona cercana.
Clulas fusiformes: tienen su eje longitudinal vertical a la
superficie y estn concentrados principalmente en las capas
corticales ms profundas. Las dendritas se originan en cada polo del
cuerpo celular, mientras que la dendrita superior asciende hacia la
superficie de la corteza y se ramifica en las capas superficiales.
El axn se origina en la parte inferior del cuerpo celular y entra
en la sustancia blanca como fibra de proyeccin, asociacin o
comisural.
Clulas horizontales de Cajal: son pequeas clulas fusiformes
orientadas horizontalmente que se hallan en las capas ms
superficiales de la corteza. Se origina una dendrita a cada lado
del axn corre paralelamente a la superficie de la corteza haciendo
contacto con las dendritas de las clulas piramidales.
Clulas de Marinotti: son pequeas clulas multiformes presentes en
todos los niveles de la corteza. La clula tiene dendritas cortas
pero el axn se dirige hacia la piamadre de la corteza, donde
termina en una capa ms superficial, en general, en la ms
superficial.
1.- CAPAS DE LA CORTEZA CEREBRALSe dividen por densidad y
disposicin de las clulas en:Capa molecular (capa plexiforme): es la
ms superficial. Consiste en una red densa de fibras nerviosas
orientadas tangencialmente. Estas derivan de dendritas de clulas
piramidales y fusiformes, los axones de clulas estrelladas y de
Martinotti. Tambin hay fibras aferentes que se originan en el
tlamo, de asociacin y comisurales. Entre las fibras nerviosas hay
algunas clulas de Cajal. Por ser la capa ms superficial se
establecen gran cantidad de sinapsis enter diferentes neuronas.
Capa granular externa: contiene un gran nmero de pequeas clulas
piramidales y estrelladas. Las dendritas de stas clulas terminan en
la capa molecular y los axones entran en las capas ms
profundas.
Capa piramidal externa: esta capa est compuesta por clulas
piramidales. Su tamao aumenta desde el lmite superficial hasta el
lmite ms profundo. Las dendritas pasan hasta la capa molecular y
los axones hasta la sustancia blanca como fibras de proyeccin,
asociacin o comisurales.
Capa granular interna: esta capa est compuesta por clulas
estrelladas dispuestas en forma muy compacta. Hay una gran
concentracin de fibras dispuestas horizontalmente conocidas en
conjunto como la banda externa de Baillarger.
Capa ganglionar (capa piramidal interna): esta capa contiene
clulas piramidales muy grandes y de tamao mediano. Entre las clulas
piramidales hay clulas estrelladas y de Martinotti. Adems hay un
gran nmero de fibras dispuestas horizontalmente que forman la banda
interna de Baillger. En las zonas motoras de la circunvolucin
precentral, las clulas de proyeccin de Betz dan origen
aproximadamente al 3% de las fibras de proyeccin del haz
corticoespinal.
Capa multiforme (capa de clulas polimrficas): aunque la mayora
de las clulas son fusiformes, muchas son clulas piramidales
modificadas cuyo cuerpo celular es triangular u ovoideo. Las clulas
de Martinotti tambin son conspicuas en esta capa. Hay muchas fibras
nerviosas que entran en la sustancia blanca subyacente.
No todas las reas de la corteza cerebral poseen seis capas.
Aquellas reas de la corteza en las cuales no puede reconocerse las
seis capas bsicas se denominan heterotpicas en oposicin a la mayora
que es homotpica.
2.- LOBULOS
Como se dijo, son 4 grandes masas neuronales en las que se
ubican las funciones primordiales de la actividad nerviosa
consciente. A ello se agrega un quinto lbulo que no es visible en
la parte externa del cerebro. De manera muy abreviada (no olvidarse
que este es un resumen de las clases para ayudar al estudio) la
fisiologa de los lbulos es la siguiente: Lbulo frontal.- Cumple
mltiples funciones, siendo principalmente la de organizar las
rdenes motoras armonizando la respuesta voluntaria. Se puede
reconocer en este lbulo:
Area Motora Primaria: se extiende sobre le limite superior del
lobulillo paracentral. Si se estimula produce movimientos aislados
en el lado opuesto del cuerpo y contraccin de grupos musculares
relacionados con la ejecucin de un movimiento especfico. Las reas
del cuerpo estn representadas en forma invertida en la
circunvolucin precentral. Comenzando desde abajo hacia arriba:
deglucin, lengua, maxilares, labios, laringe, prpado y cejas,
dedos, manos, mueca, codo, hombro y tronco etc.
La funcin del rea motora primaria consiste en llevar a cabo los
movimientos individuales de diferentes partes del cuerpo. Como
ayuda para esta funcin recibe numerosas fibras aferentes desde el
rea premotora, la corteza sensitiva, el tlamo, el cerebelo y los
ganglios basales. La corteza motora primaria no es responsable del
diseo del patrn de movimiento sino la estacin final para la
conversin del diseo en la ejecucin del movimiento, es decir la
elaboracin del movimiento propiamente tal se origina en otros
sectores muy complejos que reciben el nombre de reas de
asociacin.
Area Pre-motora: no tiene clulas gigantes de Betz. La
estimulacin elctrica de esta zona produce movimientos similares a
los del rea motora primaria pero se necesita estimulacin ms intensa
para producir el mismo grado de movimiento. Recibe numerosas
aferencias de la corteza sensitiva, tlamo y ganglios basales. La
funcin de sta rea es almacenar programas de actividad motora
reunidos como resultado de la experiencia pasada; es decir programa
la actividad motora primaria armonizando dicha respuesta.
Area Motora Suplementaria: se ubica en la circunvolucin frontal
medial y por delante del lobulillo paracentral. La estimulacin de
esta rea da como resultado movimientos de las extremidades
contralaterales, pero es necesario un estmulo ms fuerte que el
necesario en la zona primaria. La eliminacin de sta rea no produce
una prdida permanente de movimiento sino problemas de
coordinacin.
Campo Ocular Frontal: No corresponde a la percepcin visual
(lbulo occipital) sino al movimiento del receptor visual que es el
ojo. Se extiende hacia delante desde el rea facial de la
circunvolucin precentral hasta la circunvolucin frontal media. La
estimulacin de esta rea produce movimientos conjuntos de los ojos
en especial en el lado opuesto. Controla los movimientos de
seguimiento voluntario de los ojos y es independiente de los
estmulos visuales. El seguimiento involuntario ocular de los
objetos en movimiento comprende el rea visual en la corteza
occipital, que est conectada al campo ocular frontal por fibras de
asociacin.
Area Motora del Lenguaje de Broca: est ubicada en la
circunvolucin frontal inferior entre las ramas anterior y
ascendente y las ramas ascendente y posterior de la cisura lateral.
En la mayora de los individuos esta rea es importante en el
hemisferio izquierdo o dominante y su ablacin da como resultado
parlisis del lenguaje. La ablacin de la regin en el hemisferio no
dominante no tiene efectos sobre el lenguaje. Produce la formacin
de palabras por sus conexiones con las reas motoras adyacentes,
msculos de la laringe, boca, lengua etc.
Corteza Pre-frontal: ocupa la mayor parte de las
circunvoluciones frontal superior, media e inferior. Est vinculada
con la constitucin de la personalidad del individuo. Regula la
profundidad de los sentimientos y est relacionada con la
determinacin de la iniciativa y el juicio del individuo.ORGANIZACIN
GENERAL DEL SISTEMA NERVIOSO CENTRALLa mayor parte de las
actividades del S.N.C dependen de impulsos sensoriales que derivan
de campos receptores externos (piel) e internos (vsceras,
receptores musculares, etc.) adems de los campos correspondientes a
la visin, audicin, gusto y olfato. Las descargas de impulsos
ingresan al S.N.C a travs de los nervios raqudeos y craneanos,
hacen estacin en ncleos sensitivos a nivel de la mdula, el bulbo,
el cerebelo, el puente, el mesencfalo, el tlamo y finalmente
terminan en reas sensoriales primarias de la corteza cerebral.
Elaboracin de la informacin sensorial:Una de las principales
funciones del S.N.C es analizar la informacin que llega de manera
que se produzcan los efectos deseados, despus de una adecuada
integracin en los diferentes ncleos que participan en diferentes
procesos centrales de modulacin, control y coordinacin de las
respuestas motoras.
Conservacin de la informacin-memoria:Una gran parte de la
informacin que ingresa al S.N.C. es almacenada para su utilizacin
posteriormente en futuras acciones motoras, o para ser aprovechadas
en procesos mentales.
Destino de la informacin que ingresa a la mdula:
Parte de la informacin que llega a la mdula a travs de los
nervios raqudeos en integrada y traducida a respuestas motoras en
forma de reflejos medulares tipo somtico y autnomo. Otra parte es
conducida hacia el encfalo por vas nerviosas o cordones, entre los
cuales, los ms importantes son:
Cordones Posteriores (Gracilis y cuneatus): que conducen
impulsos procedentes de campos receptores de la sensibilidad
epicrtica de gran discriminacin (tacto fino, sensibilidad
vibratoria, sensibilidad, etc). Toda esta informacin llega a los
ncleos de Gall y Burdach (gracilis y cuneatus) que se encuentran en
el bulbo. Despus de hacer estacin en estos ncleos, los impulsos
pasan al lado contrario (decusacin sensitiva) y ascienden hasta el
tlamo ptico por la va del lemnisco medio. En el tlamo hace la
segunda estacin en el ncleo ventroposterolateral. Desde este ncleo
los impulsos se dirigen a la corteza cerebral siguiendo las
radiaciones sensitivas y se distribuyen a lo largo de la
circunvalacin parietal post-central o post-rolndica. Esta zona se
denomina rea somatosensorial primaria.
Cordn o haz espinotalmico ventral: esta va conduce impulsos que
son generados al receptores de presin, de tacto y de
mecanorreceptores no discriminativos. La informacin que llega, como
el caso anterior, parte de ella se integra para dar lugar a
respuestas motoras medulares somticas y autonmicas en forma de
reflejos. La otra parte de la informacin, despus de hacer estacin
en el asta posterior de la mdula cruza al lado contrario y asciende
siguiendo el haz espinotalmico ventral, que a nivel del tallo
cerebral forma el lemnisco lateral, que conduce los impulsos hacia
el ncleo ventropostero lateral del tlamo, donde se realiza el
segundo relevo o estacin. Desde este ncleo hasta la corteza, los
impulsos siguen el mismo camino y destino que los del caso
anterior.
c) Cordn o haz espinotalmico lateral: La distribucin de los
impulsos es igual que en el caso anterior. Las descargas provienen
de receptores de dolor y de termorreceptores perifricos. Los
impulsos que ingresan a la mdula, hacen estacin en el asta
posterior, cruzan al lado opuesto y ascienden por el haz
espinotalmico lateral. Siguiendo la misma va del lemnisco lateral
hasta llegar al ncleo ventroposterolateral, donde hacen estacin
para despus tener el mismo destino que los impulsos de los cordones
anteriores
d) Cordones o haces espinocerebelosos: Estos haces conducen
impulsos generados en campos propioceptores que comprenden
receptores tendinosos, capsulares, husos musculares, etc. Todas
estas descargas sensitivas forman parte de la sensibilidad
inconsciente, debido a que no alcanzan la corteza cerebral, y slo
llegan a la corteza cerebelosa formando parte de la informacin que
ser utilizada en los procesos de control y coordinacin motora que
realiza el cerebelo. Los impulsos ingresan a la mdula y hacen
estacin en la sustancia gris medular. Desde aqu una parte cruza la
lnea media y asciende por el haz espinocerebeloso ventral. Estos
impulsos ingresan al cerebelo por el pednculo cerebeloso superior.
Otra parte de las descargas ascienden por el haz espinocerebeloso
dorsal o directo, e ingresan al cerebelo por el pednculo cerebeloso
inferior del mismo lado.
rea somatosensorial primaria I:Toda la informacin que hace
estacin en el ncleo ventropostero lateral del tlamo llega en su
mayor parte a esta rea. Una parte menor llega al rea
somatosensorial primaria II.
El rea I se halla situada en la circunvalacin post-central de la
corteza cerebral humana.
A este nivel hay una orientacin espacial neta para la recepcin
de impulsos procedentes de las diversas partes del cuerpo. Las
dimensiones de estas reas son directamente proporcionales al nmero
de receptores especializados en cada una de las respectivas zonas
perifricas del cuerpo. En labios y pulgares, por ejemplo, hay gran
nmero de receptores y su representacin en la corteza, por lo tanto
es muy amplio. Lo contrario ocurre con las zonas del tronco que
poseen pocos receptores.
Funcin area sensorial primaria I: La extirpacin o destruccin de
esta rea provoca la prdida de los siguientes tipos de juicio
sensorial:
La persona no puede localizar bien las diferentes sensaciones en
las diversas partes del cuerpo con finura, pero puede localizar en
forma burda lo que indica participacin del tlamo en esta ltima
sensibilidad.
No puede juzgar cambios crticos de presiones ejercidas contra su
cuerpo.
No puede juzgar con exactitud el peso de los objetos.
Es incapaz de determinar las formas de los objetos, es decir, se
pierde la estereognosis.
No aprecia la textura de los materiales.
Es incapaz de percibir grados finos de temperaturas.
No puede reconocer la orientacin relativa de las diferentes
partes del cuerpo en relacin de unas con otras.
El dficit de funciones sensoriales producido por destruccin del
rea I puede ser recuperada en parte por las funciones del rea
somatosensorial II y del tlamo ptico.
La informacin que se genera en los campos de los telerreceptores
(ojo, oido, olfato y gusto) son conducidos por vias diferentes
(nervio ptico, nervio acstico, etc.). Hacia el tlamo ptico donde
hacen estacin en ncleos diferentes a los de la sensibilidad somtica
(cuerpos geniculados medio y lateral). Despus de este relevo los
impulsos viajan hacia reas sensoriales primarias especficas (rea
visual primaria, auditiva primaria, etc).
Movimientos voluntarios y posturales:La corteza cerebral recibe
gran cantidad de informacin en sus reas sensoriales primarias I
yII, reas visuales, auditivas, etc. Una parte de esta actividad
pasa a las reas de asociacin de la corteza cerebral (parietal,
occipital, temporal).
Toda la informacin se canaliza finalmente en un rea integrativa
general donde llega la informacin de las reas primarias,
secundarias o de asociacin, adems del aporte que puede llegar desde
los depsitos de memoria. En base a esta informacin se configura un
programa o engrama motor .
Este engrama se va a transformar en rea motora cuando se
traslada al rea motora y premotora en donde las neuronas que van a
ejecutar el programa motor configurado con anterioridad, descargarn
impulsos nerviosos que bajarn a los centros motores subcorticales y
medulares a travs de dos grandes vas: el sistema piramidal y el
sistema extrapiramidal. El primero inicia la actividad voluntaria
dando lugar a movimientos finos y hbiles con un alto grado de
precisin. El segundo se encarga de ajustar la postura del cuerpo,
provee un fondo estable para el movimiento fino y coordina la accin
de los diferentes msculos para hacer que los movimientos sean
suaves y precisos.
Sistema piramidal:Los axones de las neuronas motoras del rea 4
principales, descienden por la cpsula interna desde su origen en
las diferentes partes de la circunvalacin frontal ascendente, hasta
llegar al bulbo donde forman las pirmides bulbares, cuyas fibras, a
este nivel, se cruzan (decusacin de las pirmides). La decusacin
comprende el 80% de las fibras.
El 20% restante contina descendiendo por el mismo lado. El
primero constituye el haz piramidal cruzado y el segundo el haz
piramidal directo, cuyas fibras cruzan al lado opuesto poco antes
de llegar a su destino.
La mayor parte de las fibras terminan en neuronas inter calares
a travs de los cuales se activan las motoneuronas alfa del asta
anterior de la mdula.
Por medio de experimentos ha sido posible delimitar el rea
cortical donde se origina el sistema piramidal.
Las diferentes partes del cuerpo estn representadas en la
circunvalacin frontal ascendente. Los pies ocupan la parte superior
y la cara la inferior. La figura que se puede lograr se llama
homnculo motor casi igual al homnculo sensorial que se obtiene en
el rea sensorial.DOLOR.-Una sensibilidad que se destaca por su
importancia como sistema de alarma para informar sobre la accin de
cualquier estmulo que produzca lesin celular, es la sensibilidad
dolorosa. Los receptores del dolor estn constituidos por
terminaciones nerviosas libres que se hallan dispersas en las capas
superficiales de la piel y tejidos internos como periostio, paredes
arteriales, superficies articulares, lminas menngeas, etc.
En las vsceras los receptores estn muy distanciados, de manera
que para producir dolor se necesita el compromiso de una amplia
zona visceral para producir una sumatoria adecuada de impulsos que
abran la compuerta medular.
Si se aplica un estmulo trmico. La temperatura en el punto
estimulado debe llegar a 45C para que empiece a sentir dolor. Esta
es la T que comienza a producir lesin en los tejidos. La isquemia
tisular y las contracciones musculares mantenida (espasmos) son
estmulos que provocan dolor. El bloqueo de circulacin en el msculo
cardaco produce intensos dolores caractersticos del infarto
cardiaco.
Una vez que el campo receptor es estimulado, los impulsos
producidos ingresan a la mdula y penetran a la sustancia gris del
asta posterior, donde se encuentra un mecanismo llamado compuerta
del dolor. Este mecanismo est destinado a controlar el paso de
impulsos hacia el haz espinotalmico lateral, que es la va que los
conduce hacia el encfalo. El control es ejercido por un proceso
inhibitorio local originado por impulsos de ramas colaterales de
las fibras de los cordones posteriores y por un sistema central
(sistema Rafe Espinal) que conduce impulsos inhibitorios desde los
ncleos del rafe y sustancia periacueductal. Este sistema de
respuesta puede impedir el paso de impulsos de dolor cuando est
activado al mximo. Los impulsos de dolor se conducen desde campos
receptores a la mdula por fibras mielnicas del subgrupo delta
(dolor rpido) y por fibras amielnicas de baja velocidad de
conduccin (dolor lento). El dolor rpido del tipo pinchazo es bien
localizado (dolor epicrtico), en cambio el dolor lento del tipo
quemadura es un dolor no localizado (dolor proptoptico). El dolor
epicrtico se integra en la corteza cerebral, en cambio el dolor
protoptico puede integrarse en el tlamo. La extirpacin completa de
las reas sensitivas de la corteza cerebral no suprime la capacidad
de percibir dolor.Reacciones al dolor:El dolor produce reacciones
motoras y secretoras reflejas, adems de reacciones psquicas.
Algunas de las acciones reflejas ocurren directamente en la mdula
espinal iniciando reflejos de retraccin o de retiramiento que
alejan el miembro o el cuerpo del estmulo lesivo. Las reacciones,
adems de somticas, pueden ser autonmicas y se manifiestan como
aumento de la presin arterial, taquicardia y sudoracin en algunos
casos. En cambio, en otros puede producirse hipertensin y
braquicardia. Las reacciones psquicas al dolor pueden ser ms
sutiles y se manifiestan como angustia, ansiedad, depresin, aumento
de la excitabilidad muscular e irritabilidad.
Dolor referido:Este dolor se percibe en partes alejadas de los
tejidos en donde se ha desencadenado el dolor. Frecuentemente el
dolor referido se genera en una vscera, pero la sensacin dolorosa
se proyecta a una zona superficial del organismo. En el infarto
cardaco, la vscera lesionada es el corazn, sin embargo, el dolor se
proyecta al borde interno del brazo y a la zona esternal. En
lesiones de la articulacin de la cadera, el dolor se siente en la
rodilla. Como estos dos casos, hay innumerables ejemplos de dolor
referido. El dolor referido se ha explicado por un mecanismo de
convergencia a nivel medular, es decir, que los impulsos que vienen
de la superficie externa se juntan con los impulsos que proceden de
las vsceras de tal manera que el haz espinotalmico lateral pasa a
ser una va comn.Como a lo largo de la vida del individuo los
dolores se han generado casi siempre en las superficies externas,
cuando una viscera se enferma y descarga impulsos, la corteza
cerebral que recibe dicha informacin la proyecta hacia la
periferia.
El dolor visceral puede ser ocasionado por isquemia, irritacin,
inflamacin, compresin, estimulacin qumica, espasmos del msculo
liso, distencin, traccin de ligamentos, etc.RECEPTORES
NERVIOSOS.Resumen de clases.
Son neuronas encargadas de detectar los cambios ambientales,
tanto del medio ambiente interno como externo del individuo, e
informar al Sistema Nerviosos Central para que este elabore una
respuesta de acomodacin. Se requiere para ello de una neurona
primaria que, por lo general, es la encargada de recibir
directamente el estmulo y, mediante relevos, transportar la
informacin que, en la mayora de los casos, llegar hasta el tlamo,
en la regin del diencfalo, y de ah hasta alcanzar el destino que
corresponda.
La mecnica puede tener variantes. Por ejemplo, la clula
receptora del estmulo puede no ser una neurona, sino de otro
tejido, como ocurre con las papilas gustativas que estn formadas
por tejido epitelial. Sin embargo, igualmente la sensacin termina
siendo captada por una neurona que se encuentra adyacente a este
tipo de receptores. De la misma forma es bueno recordar que no
siempre es el tlamo la estacin intermediaria para todo tipo de
sensibilidad, pudiendo serlo otros ncleos del encfalo como sucede
con la sensacin olfatoria que remata en el rinencfalo. Por ltimo, y
siempre a manera de repaso de la materia ya vista, hay que tener
presente que, por lo general, la sensibilidad inconsciente se
reparte desde el tlamo hacia otros ncleos sensitivos del tronco
enceflico, donde se elabora una respuesta neurovegetativa que
desciende por va simptica y/o parasimptica. La sensibilidad
consciente, en cambio, alcanza las regiones corticales respectivas
transformndose en percepcin.
Clasificacin de los receptores.-
Desde un punto de vista de su ubicacin fsica, se pueden
clasificar en interorreceptores, sensibles a los cambios del medio
interno del individuo, y los exterorreceptores, influenciados por
el medio externo. De manera ms precisa, sin embargo, se ha optado
por clasificar a estos receptores de acuerdo al tipo de energa que
los estimulas. Se tiene entonces:
a.- Mecanorreceptores.- Responden a fenmenos fsicos, como el
tacto y el odo que son exterorreceptores. Entre los
interorreceptores mecnicos se considera a aquellos que detectan
cambios de presin sangunea, postura y balance, y los de contraccin
y estiramiento muscular.
b.- Termorreceptores.- Son estimulados por cambios de
temperatura. La piel est dotada de exterorreceptores altamente
sensibles al fro y al calor, en tanto que el llamado sndrome de
adaptacin general, controlado por el hipotlamo, es un
interorreceptor de temperatura.
c.- Fotorreceptores.- Son sensibles a las longitudes de las
ondas lumnicas. Se contemplan aqu slo exterorreceptores que
corresponden a los pigmentos visuales, conos y bastones, no
habindose reportado hasta ahora receptores internos de este
tipo.
d.- Quimiorreceptores.- Reaccionan a cambios qumicos del
ambiente. El gusto y el olfato son exterorreceptores, en cambio se
consideran dentro de lo interorreceptores a los receptores a
hormonas, a cambios de pH, a neurotrasmisores, a cambios
osmticos.
e.- Electrorreptores.- Se estimulan por cambios de conductancia
elctrica. No se han reportado en el caso humano, aunque hay
animales que los presentan, como anguilas, manta rayas y otros.
A.- MECANO RECEPTORES.
Como se dijo, el tacto y el odo son los principales
representantes de los exterorreceptores mecnicos.
1.- Tacto.-
Se considera como tal a un grupo importante de receptores que se
ubican en la piel clasificndolos de acuerdo al grado de intensidad
del estmulo sobre la superficie de este rgano. Es as que se conocen
los llamados del tacto fino, los de la presin, que puede ser
moderada o profunda, y los de la vibracin que puede ser de baja o
alta frecuencia.
El tacto fino se refiere a roces suaves sobre la piel que ponen
en accin los llamados discos de Merkel que se presentan abundantes
en la yema de los dedos y en los labios. El roce ligero es captado
por estos receptores y tambin por los vellos de la piel que al
moverse deforman la dendrita ubicada en la raz del pelo provocando
la depolarizacin de la membrana neuronal y, por lo tanto, un
potencial de accin propagado que lleva la sensacin a la
corteza.
La presin sobre la piel, como se seal, puede ser moderada siendo
captada en este caso por los llamados corpsculos de Meissner,
capaces de percibir tambin las vibraciones de baja frecuencia y los
roces del pelo sealados ms arriba compartiendo la sensibilidad con
los discos de Merkel.
Las vibraciones de alta frecuencia estimulan a los llamados
corpsculos de Vater Pacini que, adems, comparten con los de
Meissner la sensacin de presin moderada.
La presin profunda, es decir la presin fuerte provocada sobre la
piel y que hace que sta se deforme, es captada por los corpsculos
de Ruffini que actan tambin en sensaciones trmicas como se seala ms
abajo.
De manera especial se incluyen ac a los propioceptores o
propiorreceptores que captan la postura de msculos y tendones, en
lo que participan el rgano de Golgi y el huso neuromuscular
(sensibilidad fina de los msculos). La informacin, llamada de la
postura y el balance, sube por fascculos de la regin anterior de la
mdula hasta ncleos cerebelosos y de la base (ncleos profundos) que
elaboran una respuesta extrapiramidal de acomodacin.
En el caso de los mecanorreceptores internos del organismo, se
encuentran aquellos que pueden ser captados por el individuo como
reflejos carenciales, por ejemplo el hambre cuyos interorreceptores
se ubican en las paredes del estmago y se ponen en funcionamiento
al contraerse estas paredes por falta de alimentos, aunque en este
fenmeno participan tambin otros factores ms bien de tipo qumico,
como la cada de la tasa metablica y el descenso de la glicemia
(cantidad de azcar en la sangre).
Tambin la sed se activa por interorreptores en el fondo de las
fauces que pierden humedad informando de esta anomala a los centros
enceflicos. Lo mismo ocurre con la nausea que se manifiesta por
interorreceptores que informan desde las paredes del tracto
digestivo acerca del sentido los movimientos peristlticos de avance
de los alimentos. Un retroceso anmalo activa a estos receptores que
provocan la nausea desde los centros superiores.
ANEXO
Informacin complementaria de los mecanorreceptores para
integrarla con la anterior
ExteroceptoresSon los que se encuentran en estructuras derivadas
del ectodermo (como la piel).
Existen diferentes tipos:
1. Terminaciones libres:
1.1. Terminaciones libres amielnicas
1.2. Terminaciones de los folculos pilosos (o Terminacin libre
relacionada con el tacto)
2. Terminaciones Encapsuladas:
2.1. Discos de Merkel
2.2. Corpsculos de Meissner
2.3. Bulbo de Krause (fro) y Bulbo de Ruffini (calor)
2.4. Corpsculos de Vater Paccini
1.1. Terminaciones libres amielnicasSon fibras de tipo C.
Se ubican en el interior de las capas superficiales de la
epidermis.
Son receptores de dolor (nociceptor).
El dolor significa potencial o real destruccin tisular, porque
las terminaciones nerviosas libres se activan cuando se rompe una
clula, ya que cuando se rompe, sale potasio, serotonina, histamina,
es decir, una serie de sustancias que activan las terminaciones
libres.
1.2. Terminaciones de los folculos pilosos (o Terminacin libre
relacionada con el tacto)
Se ubica alrededor del bulbo del folculo piloso.
Corresponden a fibras amielnicas que se arrollan alrededor de un
bulbo piloso, de tal manera que nos permiten la sensacin tctil.
Son activadas con el movimiento del pelo.
2.1. Discos de MerkelEstn ubicados intraepitelialmente.
Se relacionan con tacto protoptico o grosero, es decir, aqul
tacto que nos permite discriminar en forma burda acerca de las
caractersticas fsicas de alguna estructura (por ejemplo si es duro
o blando).
2.2. Corpsculos de MeissnerEs una terminacin encapsulada, se
encuentran en gran cantidad en los pulpejos de los dedos.
Se encuentran entre epidermis y dermis.
Se relacionan con el tacto y la vibracin de baja frecuencia (30
a 40 Hz) de la piel sin pelo, sobretodo en la palma de las
manos.
Ayudan a discriminar en una superficie dura si estamos frente a
madera, vidrio o cemento, por ejemplo (tacto epicrtico).
2.3. Bulbo de Krause (fro) y Bulbo de Ruffini (calor) Son
receptores de temperatura.
Son encapsulados.
2.4. Corpsculos de Vater Paccini Se encuentran en las capas ms
profundas de la piel (hipodermis).
Son mecanorreceptores sensibles a la vibracin, en el fondo, son
fibras amielnicas rodeadas por clulas aplanadas que son
fibroblastos y lpidos.
Se descargan fundamentalmente ante estmulos de presin.
-La teora especfica seala que para cada modalidad de sensacin
existe un determinado receptor. Pero hay otra teora que indica que
cualquier receptor puede desencadenar otra sensacin, por ejemplo,
dolor, cuando la intensidad del estmulo pasa cierto nivel.
PropioceptoresSe ubican en las estructuras derivadas del
mesodermo, como son por ejemplo, las del aparato
musculoesqueltico.
Informan acerca del movimiento del cuerpo, es decir, de la
posicin de los segmentos corporales y acerca del movimiento o
cinestesia.
Estn representados por:
-Husos Neuromusculares Se ubican en el interior de los vientres
de la musculatura estriada.
-rganos Musculotendneos u rganos de GolgiSe ubican en la unin
entre los tendones y los vientres musculares.
Todas estas estructuras son las responsables de monitorear el
movimiento y posicin de los miembros, de las articulaciones, de los
ligamentos.
2.- Odo y equilibrio conciente.-
Se ubican ambos en el canal auditivo que se interna perforando
el hueso mastoideo en la regin temporal del crneo. Ambos receptores
se encuentran en la parte ms profunda de este canal en el llamado
odo interno. Ah se encuentra un rgano llamado laberinto que se
reparte entre la regin vestibular destinada al equilibrio, y la
regin coclear que contiene a la audicin.
Audicin:- Comprende los tres segmentos del canal auditivo. El
odo externo presenta el pabelln de la oreja destinados a capturar y
desviar hacia el interior a las ondas sonoras y el conducto
auditivo externo protegido por vellosidades y cerumen y que termina
en la primera membrana, el tmpano. El odo medio presenta una cadena
de pequeos huesos, martillo, yunque y estribo en ese mismo orden de
afuera hacia adentro, que unen al tmpano con la ventana oval, una
membrana que separa al odo medio del interno. Tambin el odo medio
comprende a la trompa de Eustaquio que une esta cavidad con la
faringe.
La regin coclear del odo interno la forman tres conductos
paralelos que se enrollan para dar origen al caracol en cuyo
extremo distal se ubica el verdadero receptor de la audicin que es
el rgano de Corti. Esta estructura est formada por cinco filas de
neuronas (unas 25 mil clulas) con dendritas de diferente longitud y
grosor que conectas a las dos membranas que conforman el rgano de
Corti, la membrana tectorial y la basilar.
El laberinto completo esta flotando en un lquido llamado
perilinfa que se extiende por detrs de membrana ventana oval y se
contina por el conducto ms interno del caracol llamado escala o
rampa vestibular para retornar por el canal ms externo, la rampa
timpnica y concluir en la quinta membrana, la ventana redonda cuya
cara externa da hacia el odo medio. El conducto central del caracol
llamado rampa coclear, contiene un lquido llamado endolinfa que
llena interiormente todo el laberinto, incluida la regin
vestibular.
Fisiolgicamente las ondas sonoras hacen vibrar el tmpano luego
de penetrar por el conducto auditivo externo. La vibracin se
trasmite a la cadena de pequeos huesos que la traspasan a su vez a
la ventana oval. Como la cara interna de la ventana oval esta baada
por la perilinfa, la vibracin se convierte en onda lquida que
recorre la rampa vestibular para retornar por la rampa timpnica. La
cara externa de la membrana basilar que forma el rgano de Corti est
baada por esta perilinfa lo que provoca su vibracin induciendo
potenciales de accin a las neuronas de este rgano que se
transportan por el octavo par craneal hasta la regin temporal de la
corteza. La diferente longitud y grosor de las dendritas de las
neuronas del rgano de Corti determinan los diferentes tonos y
timbres de los sonidos.
Finalmente la onda lquida se contina por la rampa timpnica para
detenerse totalmente en la cara interna de la membrana ventana
redonda.
DECIBELES
Efecto del sonido sobre el organismo humano
Dentro del estudio de los sonidos y la forma como estos son
captados por los receptores auditivos humanos, hubo que disear una
unidad de medicin que permitiera graficar las diferencias del
sonido, lo que ha servido en el campo de la medicina para detectar
las anomalas que pueda presentar la audicin.
A principios del siglo 20 se cre una unidad de sonido que recibi
el nombre de Unidad de Transmisin (TU en ingls) que reflejaba
experiencias realizadas antes con las variaciones de potencia que
ocurran en el desplazamiento del sonido a travs de un cable. Diez
de estas unidades formaban lo que se design como un BEL en honor a
Alejandro Graham Bell hasta entonces considerado como el inventor
del telfono.
Con el objeto de facilitar las conversiones se decidi fijar a la
dcima parte de un BEL como la unidad definitiva y que era lo que ms
se acercaba en su valor a la anterior Unidad de Transmisin (TU).
Por lo tanto se estableci el deciBEL (la dcima parte deun BEL) como
valor unitario del sonido.
Como la emisin de un sonido es producto de la potencia de alguna
forma de energa, se estableci una frmula para calcular los BEL de
un sonido que corresponde al logaritmo en base 10 de la relacin
entre la potencia medida y la de referencia:
log(P/Pref) = cantidad de BEL
P= Potencia con la que se transmite un sonido
P ref= unidad de energa con la cual se hace el cculo
Por ejemplo, una emisin de sonido que tenga 100 watts de
potencia, aplicando la frmula tendr un valor de 2 BEL, es decir 20
decibeles:
Log 100W = log 100 = log 100 = 2 es decir, 20 decibeles 1W
1Equilibrio.- Como se dijo se ubica en la regin vestibular del
laberinto en el odo interno. Est formado por los canales
semicirculares que se orientan en los tres sentidos del espacio
estando, adems, llenos del lquido endolinfa. Esto canales
desembocan en una bolsa membranosa llamada utrculo que contiene
pequeos grnulos sueltos de carbonato clcico llamados otolitos.
Tanto la desembocadura de los canales semicirculares, las ampullas,
como las paredes del utrculo, estn tapizadas de neuronas ciliadas
cuyos cilios flotan en la endolinfa. Los axones de estas neuronas
pasan a integrar tambin el VIII par craneal dirigindose a las
regiones corticales del encfalo.
La fisiologa del equilibrio conciente indica que los movimientos
horizontales del sujeto, a los lados y al frente, son detectados
por el canal semicircular respectivo. Se mueve entonces la
endolinfa de ese canal barriendo los cilios de la ampulla lo que
provoca el potencial de accin que se informa a la corteza. Los
movimientos verticales los detecta la agitacin de los otolitos que
se posan en el centro de gravedad del movimiento aplastando los
cilios de ese sector estimulando a las neuronas de la misma forma
que los canales semicirculares.
B.- TERMORRECEPTORES.
Captan cambios de temperatura externos para lo cual existen
receptores en la piel que actan paralelos a los del tacto. Los
corpsculos de Ruffini son sensibles al calor, adems de activarse
con la presin profunda. El fro, en cambio, es detectado por los
llamados corpsculos de Krausse. Se agrega en estos
termorreceptores, el llamado sndrome de adaptacin general ya citado
ms arriba. Se trata de la respuesta del organismo a los
requerimientos de los cambios trmicos externos que pueden influir
en la temperatura corporal que debe mantenerse independizada del
medio (homeotermia), y que requiere fenmenos de adaptacin interna.
Por ejemplo, y entre otros, est la mayor o menor eliminacin de agua
corporal (transpiracin), mayor o menor irrigacin sangunea
perifrica, disminucin o elevacin de la tasa metablica expresada en
caloras segn requerimientos, regulacin del metabolismo basal
productor de ATP, etc.
C.- FOTORRECEPTORES.
Su papel es convertir la energa lumnica, que se expresa en ondas
llamadas quantos, fotones o exitones en energa qumica de electrones
para luego estimular la membrana neuronal y crear potenciales
bioelctricos.
La luz del sol se expresa en ondas de diferente longitud cuya
energa es inversamente proporcional a su largo, es decir mientras
ms corta es la longitud de la onda, cuyo smbolo es lambda (), mayor
es la energa que posee. De esta variedad de ondas, las que van de
una longitud de 360 nanmetros a 750 constituyen el llamado espectro
de luz visible. Esto quiere decir que los receptores nerviosos
visuales pueden percibir slo los rangos de luz comprendidos en esos
parmetros. Es posible calcular la energa de las ondas lumnicas
mediante la frmula:
E. lumnica = h x c/donde h es la constante de Plank = 1,58 x
10-34 caloras por segundo y c es la velocidad de la luz en el vaco
igual a 300 x 103 kilmetros por segundo Los rangos del espectro de
luz visible pueden descomponerse en los colores del arcoiris
mediante un prisma. El desglose es el siguiente:
360 a 430 nanmetros: luz violeta
430 a 500 : luz azul
500 a 560 : luz verde
560 a 600 : luz amarilla
600 a 650 : luz naranja
650 a 750 : luz roja
Los accesorios del fotorreceptor humano se encuentran en el ojo
cuya estructura fue descrita en clases y se muestran en el esquema
adjunto.
Fisiolgicamente la luz ingresa regulada por la pupila que puede
abrirse (midriasis) o cerrarse (miosis) por accin neurovegetativa
que acciona los msculos radiales del iris, atravesando luego la
lente o cristalino que concentra los rayos lumnicos a la manera de
una lupa en la fovea, una depresin de la retina donde se acumulan
los conos, fotoclulas nerviosas encargadas de la visin aguda y la
visin en colores. Las ondas lumnicas que convergen en la fovea son
las que emite el objeto que se est mirando, segn se explic en
clases. En cambio las ondas que emiten los objetos de la periferia
estimulan a los bastones, fotoclulas que estn dispersas en la
retina. De ah que estos ltimos se encarguen de la llamada visin
borrosa (reojo), adems de la visin nocturna que es la adaptacin
bioqumica de los receptores y la adaptacin mecnica del iris a la
oscuridad.Bioqumica de la visin.-Los pigmentos visuales (rodopsina)
son los componentes qumicos de la visin capaces de reaccionar ante
los estmulos de la enega lumnica (fotones o quantos). Estas
molculas se ubican en los extremos de las fotoclulas (conos y
bastones) en el lmite con la membrana retina, siendo molculas
proteicas transmembranales de las fotoclulas. La molcula consta de
dos partes: una proteica llamada opsina que posee cerca de 350
aminocidos repartidos en siete cadenas polipeptdicas de estructura
-hlice, y la otra es un pigmento rojizo derivado de la vitamina A
(vitamina de origen lipdico) llamado 11-cis-retinal que se une a
las cadenas polipeptdicas de la opsina de manera perpendicular a la
membrana del receptor, lo que otorga estabilidad a toda la
estructura. La luz que traspasa la retina es absorvida por el
pigmento visual provocando la ruptura del enlace del 11-cis-retinal
que cambia a su forma trans (11-trans-retinal) que es una molcula
ms rgida y lineal que el ismero cis. Se forma, entonces, una
estructura diferente entre la opsina y el 11-trans-retinal llamada
batorrodopsina. Estos movimientos espaciales del pigmento se
traspasan a las clulas nerviosas de relevo provocando un potencial
de accin que es llevado a la corteza occipital por el 2 par craneal
ptico. La rodopsina en los bastones se sintetiza a mayor velocidad
en ausencia de la luz, ms o menos en los rangos de 500 nanmetros
(luz verde azulada). Eso le permite a los bastones adaptarse mejor
a la oscuridad posibilitando la llamada visin nocturna adems de la
visin borrosa o de reojo.
Los conos, por su parte, pueden discriminar entre las diferentes
longitudes de onda de la luz visible, es decir determinar colores,
gracias a ismeros de la opsina que se sensibilizan a los colores
azul, rojo y verde, permitiendo la variacin de tonalidades en
combinaciones neuronales que ocurren en la trayectoria del par
craneal visual hacia la regin occipital de la corteza.CAMPOS
VISUALES.-El camino que sigue el nervio ptico hacia la corteza
tiene caractersticas especiales, como se muestra en el grfico y se
explic en clases. Hay que considerar aqu que las diferentes vas
nerviosas se originan en sectores diferentes del lobo ocular
designados como campos visuales nasales (derecho e izquierdo) que
son los que limitan con la nariz, y campos visuales temporales que
son los adyescentes al hueso temporal. Estos ltimos cruzan hacia
los hemisferios contrarios.
La visin que abarca cada uno de estos nervios depende de la
forma cncava que tiene la retina en el fondo de cada ojo y que se
explica por el camino recto que siguen los rayos lumnicos.
ALGUNAS ANOMALAS DE LA VISIN.- GLAUCOMA.-
Un glaucoma se produce cuando se eleva la presin del lquido
interior del ojo lesionando al nervio ptico provocando la prdida de
la visin que puede llevar inclusive hasta la ceguera. El glaucoma
es una de las causas ms comunes de ceguera en pases como los
Estados Unidos y se dividen en cuatro: glaucoma de ngulo cerrado,
glaucoma de ngulo abierto, glaucoma congnito y el glaucoma
secundario. Estos cuatro tipos tienen como caracterstica principal
una presin elevada en el globo ocular y por ende todos pueden
causar daos de forma progresiva al nervio ptico.
Esta presin se eleva cuando el lquido del interior del ojo no es
drenado de manera apropiada, la presin oprime la unin del nervio
ptico y la retina contra la parte posterior del ojo, causando as
una reduccin en el suministro sanguneo al nervio ptico, que es el
que lleva la informacin visual desde el ojo hasta el cerebro. Dicha
prdida del suministro sanguneo implica que las clulas nerviosas
individuales mueran de forma progresiva y a medida que el nervio
ptico va deteriorndose se generan puntos ciegos en el campo de
visin. Lo primero que se afecta es la visin perifrica (visin
lateral), seguida de una disminucin en la visin central (frontal),
si estos sntomas no son tratados el glaucoma puede ocasionar
ceguera.
El glaucoma de ngulo cerrado: se presenta por lo general en
personas que nacieron con un ngulo estrecho entre el iris y la
crnea, y es una situacin muy comn en ojos hipermtropes (las
personas que ven de mejor forma objetos a distancia que aquellos
que estn cerca), el irirs puede desplazarse hacia delante y cerrar
la salida del humor acuoso, generndose luego un aumento repentino
de la presin intraocular. Los sntomas que este glaucoma presente
son: dolor, enrojecimiento, nuseas, prdida de la visin, en el caso
de otras personas, el glaucoma puede darse por el uso de gotas que
dilatan los ojos.El glaucoma de ngulo abierto: es el tipo ms comn
de glaucoma, aqu el iris no bloquea el ngulo de drenaje, en lugar
de esto, los delgados canales de salida del lquido al interior de
la pared ocular se estrechan con el paso del tiempo. Este mal
afecta a ambos ojos y en unos aos la elevacin de la presin va
daando al nervio ptico. Este tipo de glaucoma no genera sntomas a
corto plazo, ya que la prdida de la visin ocurre de forma gradual,
la nica forma de diagnosticar este tipo de glaucomas es haciendo
exmenes mdicos rutinarios de los ojos.El glaucoma secundario: es
ocasionado por otras enfermedades como uvetis, enfermedades
sistemticas, o el uso de medicamentos como los corticosteroides.El
glaucoma congnito: es aquel que se presenta al momento de nacer,
generado por defectos en el desarrollo de los canales oculares de
flujo de lquido y para curarlo se requiere ciruga, ste tipo de
glaucoma es generalmente hereditario.
CATARATAS
La catarata es la prdida de trasparencia del cristalino. El
cristalino es una lente transparente que tenemos detrs de la pupila
y que nos sirve para enfocar ntidamente los objetos. Por una serie
de circunstancias, enfermedades o ms frecuentemente debido al paso
de los aos, el cristalino puede ir perdiendo su natural
transparencia y convertirse en una lente opaca. Por tanto una
catarata ser ms o menos avanzada dependiendo de si la disminucin de
transparencia es mayor o menor. Cuanto mayor es la prdida de
transparencia del cristalino (o ms avanzada es la catarata) mayor
ser la disminucin de visin.
CAUSAS:
Edad avanzada
Diabetes
Infecciones o traumatismos oculares
Tendencia familiar
Las cataratas son una parte normal del envejecimiento. Cuanto
mayor es la persona, mayor es la probabilidad de aparicin y
progresin de las cataratas. Sin embargo hay ocasiones en que las
cataratas aparecen en edades ms precoces, sobre todo en personas
que padecen determinadas enfermedades como la diabetes,
inflamaciones intraoculares y alta miopa entre otras. Tambin pueden
aparecer cataratas tras un traumatismo ocular o en algunas familias
predispuestas. Casi tres de cada cuatro personas mayores de 75 aos
padecen de cataratas, la mayora de las personas que tienen
cataratas, las tienen en los dos ojos. Sin embargo, uno de los ojos
puede estar peor que el otro, porque cada una de las cataratas se
desarrolla con diferente rapidez.
D.- QUIMIORRECEPTORES.
Se considera en este rango a los receptores del gusto y el
olfato que actan en interdependencia. En ambos casos se requiere
que las sustancias estimuladoras estn disueltas en agua.
A.- Sentido Del Gusto
El sentido del gusto slo percibe cuatro sabores bsicos: dulce,
salado, cido y amargo; cada uno de ellos es detectado por un tipo
especial de papilas gustativas.La lengua posee casi 10.000 papilas
gustativas que estn distribuidas de forma desigual en la cara
superior de la lengua, donde forman manchas sensibles a clases
determinadas de compuestos que inducen las sensaciones del gusto.
Por lo general, las papilas sensibles a los sabores dulce y salado
se concentran en la punta de la lengua, las sensibles al cido
ocupan los lados y las sensibles al amargo estn en la parte
posterior. Los compuestos qumicos de los alimentos se disuelven en
la humedad de la boca y penetran en las papilas gustativas a travs
de los poros de la superficie de la lengua, donde entran en
contacto con clulas sensoriales. Cuando un receptor es estimulado
por una de las sustancias disueltas, enva impulsos nerviosos al
cerebro. La frecuencia con que se repiten los impulsos indica la
intensidad del sabor; es probable que el tipo de sabor quede
registrado por el tipo de clulas que hayan respondido al
estmulo.
Captacin del gusto
Dulce: en la punta de la lengua.Amargo: en las papilas
caliciformes (forma de calix)Salado y cido: en la punta y parte
anterior de los bordes de la lengua. Tambin hay receptores
sensibles a los sabores cidos en la mucosa de los labios, y
sensibles a todos los sabores en el velo del paladar.No hay
captacin gustativa en la regin sublingual y en la cara inferior de
la lengua.
La Lengua
La lengua es un rgano musculoso de la boca y es el asiento
principal del gusto y parte importante en la fonacin y en la
masticacin y deglucin de los alimentos. La lengua est cubierta por
una membrana mucosa, y se extiende desde el hueso hioides en la
parte posterior de la boca hacia los labios. La cara superior, los
lados y la parte anterior de la cara inferior son libres, solo el
resto est unido a la cavidad bucal, lo que permite muchos y
diversos movimientos. Los principales msculosque la forman son: el
lingual superior, que levanta la punta; el lingual inferior, que
baja la punta de la lengua, y el lingual transverso, que al
contraerse la dobla en forma de canaleta. Toda su superficie est
recubierta por una piel en la que se encuentran formaciones
especiales llamadas papilas. Estos son los verdaderos rganos
sensitivos. Segn forma y funcin estas pueden ser caliciformes,
filiformes y las fungiformes.
Papilas.-Son relieves de formas diferentes, las misma pueden
ser:Calciformes (en forma de cliz) Son las ms grandes. En numero de
9 a 11 forman la V lingual.Se encuentran por delante y alguna por
detrs de la V lingual. Estn mezcladas con las pailas filiformes. Su
mayor numero se localiza en los bordes y en la punta de la
lengua.
Fungiformes (con forma de sombrero de hongo)
Filiformes (con forma de filamentos) Estn distribuidas por toda
la cara superior de la lengua. Se disponen en hileras paralelas a
los brazos de la V lingual.Son filamentosas, cilndricas, y como
hilos (de ah su nombre).
Corpsculos del gusto.-
Los impulsos nerviosos gustativos se originan en los corpsculos
del gusto o bulbos del gusto.Estos corpsculos tienen el aspecto de
pequeos botellones cuya boca se abre en la superficie del epitelio,
de la boca y de la garganta. Su mayor numero se encuentra en la
cara superior de la lengua, en las papilas calciformes y
fungiformes.Cada corpsculo est formado por clulas de sostn que se
disponen como los gajos de una naranja. Estas clulas encierran y
protegen a las clulas gustativas. Las clulas de sostn y gustativas
son clulas epiteliales modificadas.
Deterioro Del Sentido Del GustoExisten dos clases de desordenes
del gusto, y son:
La "Hipogeusia": que consiste en la prdida de la habilidad de
saborear o distinguir entre lo dulce, lo salado, lo agrio, etc.
La "Ageusia", que es la prdida casi total de detectar
sabores.
Causas comunes:
En algunos casos sta condicin se presenta desde el nacimiento,
pero en la mayora de los casos se desarrolla despus de una lesin o
enfermedad.
Resfriado comn.
Infeccin nasal debido a una infeccin (como las infecciones de
las gladulas salivales), plipos, etc.
Gripe
Faringitis viral
Sequedad de la boca
Envejecimiento (el nmero de papilas gustativas disminuye con la
edad)
Fumar (en especial el fumar con pipa) dado que genera la
sequedad de la boca
Deficiencia de vitaminas (vitamina B12) o de minerales (zinc en
la dieta)
Lesiones en la boca, la nariz o la cabeza
Gingivitis
Sndrome de Sjogren (Siegren) Falta de humedad.
Faringitis estreptoccica (infeccin de la garganta por
estreptococos)
B.- Sentido Del Olfato
La cavidad nasal presenta tres zonas:- el vestbulo: es la parte
ms anterior e inferior de la nariz, y est tapizada por piel;- la
llamada "regin respiratoria": que se contina con el vestbulo y se
comunica con la nasofaringe, y est tapizada por una membrana mucosa
de tipo respiratorio;- la regin olfatoria: ubicada en el cornete
superior y tercio superior del tabique nasal, donde llegan filetes
nerviosos que atraviesan los orificios de un hueso llamado
etmoides, cuyo conjunto origina el nervio olfatorio, que alcanza el
llamado bulbo olfatorio, que se contina con las llamadas cintillas
olfatorias, cuyas fibras llevan los estmulos al cerebro y
cerebelo.
El olfato se localiza en el epitelio nasal. El epitelio
olfatorio est ubicado en el techo de la cavidad nasal, el epitelio
contiene cerca de 20 millones de clulas olfatorias especializadas,
con axones que se extienden hacia arriba, como fibras de los
nervios olfatorios. Esas fibras penetran la delgadsima placa
cribada del hueso etmoides, situado en el piso del crneo, a travs
de los poros de aqul hueso. El extremo de cada clula olfatoria de
la superficie epitelial ostenta varios vellos olfatorios que, al
parecer, reaccionan a los olores (sustancias qumicas) presentes en
el aire.A diferencia de los bulbos gustativos, que slo son
sensibles a unas cuantas categoras de sabores, el epitelio
olfatorio reacciona segn se cree a muchas sustancias. Los estmulos
olfativos se llaman odorantes. Las mezclas de esas sensaciones
olfatorias primarias generan el amplio espectro de olores que el
humano es capaz de percibir. Los rganos olfatorios reaccionan a
cantidades notablemente pequeas de sustancias. Por ejemplo la
ionona, que es el sustituto artificial del aroma de las violetas,
puede detectarse por casi todas las personas cuando su concentracin
en el aire es de apenas una parte por ms de 30 000 millones de
partes de aire.
En muchos casos resulta que la presencia de un tomo particular
en la molcula de la sustancia olorosa es la que le da su olor
peculiar. Por ejemplo, est el caso del agua, cuya molcula tiene un
tomo de oxgeno y dos de hidrgeno, H2O. Si se sustituye el tomo de
oxgeno por uno de azufre se obtiene el sulfito de hidrgeno, H2S.
Esta ltima sustancia despide el olor de huevos podridos. Vemos que
la sustitucin de un tomo de oxgeno por uno de azufre lleva a
cambios notablemente desagradables en el olor de las sustancias.Por
otro lado, existen algunas evidencias de que no es nada ms la
composicin qumica especfica de las molculas olorosas, sino tambin
su forma la que hace que reaccionemos a su olor. En efecto, existen
sustancias cuyas molculas tienen composiciones qumicas diferentes
pero con formas muy parecidas, y sentimos que tienen olores muy
parecidos.Sin embargo, la explicacin desde el punto de vista
molecular de cmo olemos sigue sin respuesta definitiva.
A pesar de su sensibilidad, el olfato es quizs el sentido que se
adapta con mayor rapidez. Los receptores olfatorios se adaptan en
un 50% durante el primer segundo de estmulo, de modo que hasta los
ms desagradables olores presentes en el aire dejan de ser
percibidos despus de unos cuantos minutos. Parte de la adaptacin
ocurre, segn se piensa, en el SNC.
Un asiento secundario de la quimiorrecepcin olfatoria es el
epitelio del rgano vomeronasal, estructura tubular que se abre en
la parte ventral del tabique nasal. Las neuronas sensoriales
situadas en el rgano vomeronasal detectan las feromonas, que
representan seales qumicas no voltiles que en los mamferos
inferiores inician comportamientos innatos y estereotipados de
carcter reproductor y social, as como cambios neuroendocrinos. Las
neuronas de este rgano se proyectan en los bulbos olfatorios
accesorios y no en el bulbo olfatorio principal, y tambin lo hacen
en el neuroepitelio olfatorio. No sabemos con certeza si el ser
humano utiliza el rgano vomeronasal para detectar y responder a las
seales qumicas de otros congneres. Al parecer, el desarrollo del
sistema olfatorio y vomeronasal es necesario para la maduracin
sexual normal.La sensacin olfatoria se inicia con la introduccin de
una sustancia odorante en los cilios de la neurona bipolar. La
mayor parte de esas sustancias son hidrfobas; a medida que pasan
desde la fase area de la cavidad nasal a la fase acuosa de la
mucosa olfatoria, son transportadas hacia los cilios por pequeas
protenas hidrosolubles denominadas protenas de fijacin de
sustancias odorantes y se unen de manera reversible a los
receptores de la superficie de los cilios. La unin origina cambios
de conformacin en la protena receptora y la generacin de
potenciales de accin en las neuronas primarias. Al parecer, la
intensidad es codificada por el nmero de impulsos que llegan a las
neuronas aferentes.En el ser humano se han identificado 300 a 1 000
genes de receptores olfatorios que pertenecen a 20 familias
diferentes, situadas en conglomerados, en ms de 25 sitios de
cromosomas diferentes. Al parecer, cada neurona olfatoria expresa
slo un gen del receptor o, a lo sumo, slo unos cuantos, y as se
establece la base molecular de la diferenciacin de los olores.
Anomalas del sentido del olfato.-
Los trminos relativos a los trastornos del olfato son: anosmia,
o falta de la capacidad para oler; hiposmia, o disminucin de dicha
capacidad; hiperosmia (aumento de la sensibilidad frente a un
olor); disosmia (distorsin en la percepcin de un olor); fantosmia,
percepcin de un olor cuando no existe ninguno; y agnosia o
incapacidad para clasificar, contrastar o identificar verbalmente
las sensaciones olfatorias, aunque pueda ser normal la capacidad
para diferenciar distintos olores o para reconocerlos.
Causas
Los desrdenes del olfato pueden tener muchas causas. La mayora
de las personas que desarrollan est condicin han tenido alguna
enfermedad o lesin, que son los desencadenantes de infecciones
respiratorias superiores y daos en la cabeza.
Algunas causas pueden ser plipos en las cavidades nasales,
sinusitis, desrdenes hormonales, o problemas dentales.
Exposicin a ciertos qumicos como insecticidas y solventes, y
algunos medicamentos.
Dos Enfermedades Que Afectan Al Sentido Del Olfato: rinitis y
sinusitis.Rinitis:Se denomina Rinitis a la inflamacin de la mucosa
nasal. Ataca tanto a adultos como a nios.Produce congestin,
insuficiencia respiratoria nasal, picazn nasal, rinorrea y en
algunos casos estornudos.
La causa de las rinitis pueden ser:a.- Infecciosa ( virales,
bacterianas, especficas), puede ser aguda (no ms de tres semanas de
duracin) o crnica (ms de tres semanas de duracin) o alrgica. En
este ltimo caso los sntomas se desencadenan ante la presencia de un
alrgeno (agente productor de la alergia)