Embed Size (px)
DESCRIPTION
3 Mitocondrias
Citation preview

MITOCONDRIAS

Mitocondrias
Las mitocondrias son organelos que
se encuentran en todas las células
eucarióticas ocupando una parte
importante del volumen celular.
Estos organelos proporcionan el
andamiaje sobre el cual se ubican la
gran variedad de moléculas que
participan en las reacciones de
transferencia de energía desde las
moléculas de nutrientes hasta el
ATP.

Presentan formas granulares o filamentosas, pero en general
tienen forma cilíndrica.
Su tamaño es también variable, con un promedio de 3.5 um de
largo por 0.5 um de ancho.
Su número varía según el tipo celular. En hepatocitos hay entre
1.000 y 2.000 mitocondrias. En células musculares el número es
mucho mayor.

Aunque su distribución dentro de la célula es generalmente
uniforme, existen numerosas excepciones.
Pueden desplazarse de una región a otra del citoplasma
donde se necesite energía.Los microtúbulos y proteínas motoras
del citoesqueleto participan en esos movimientos
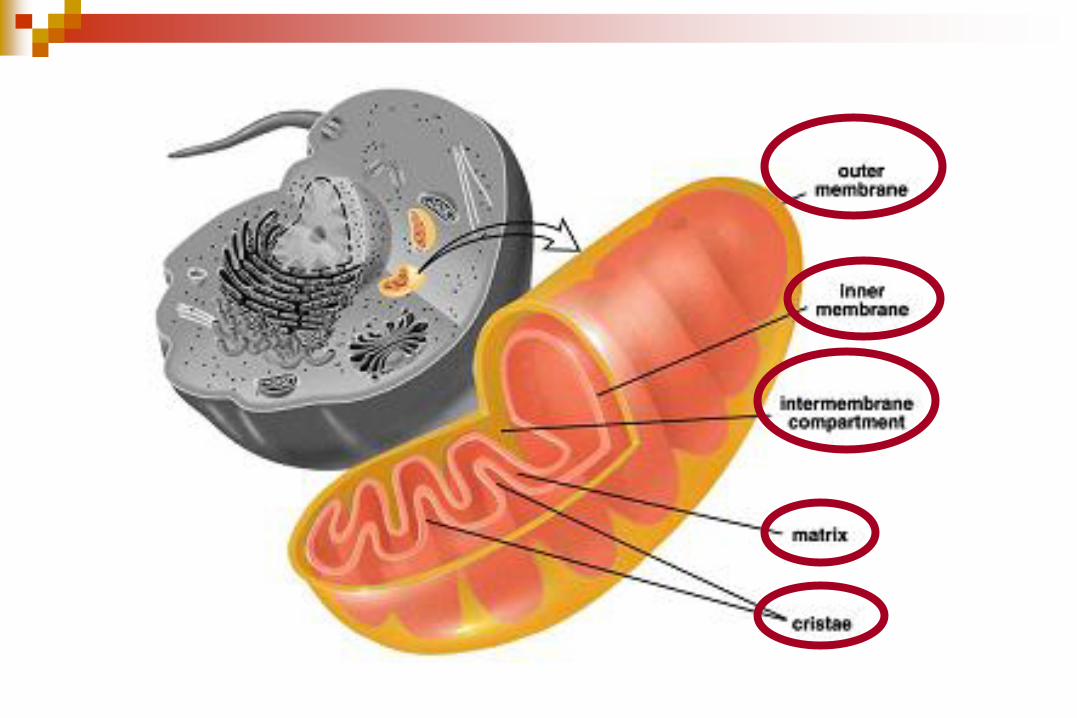
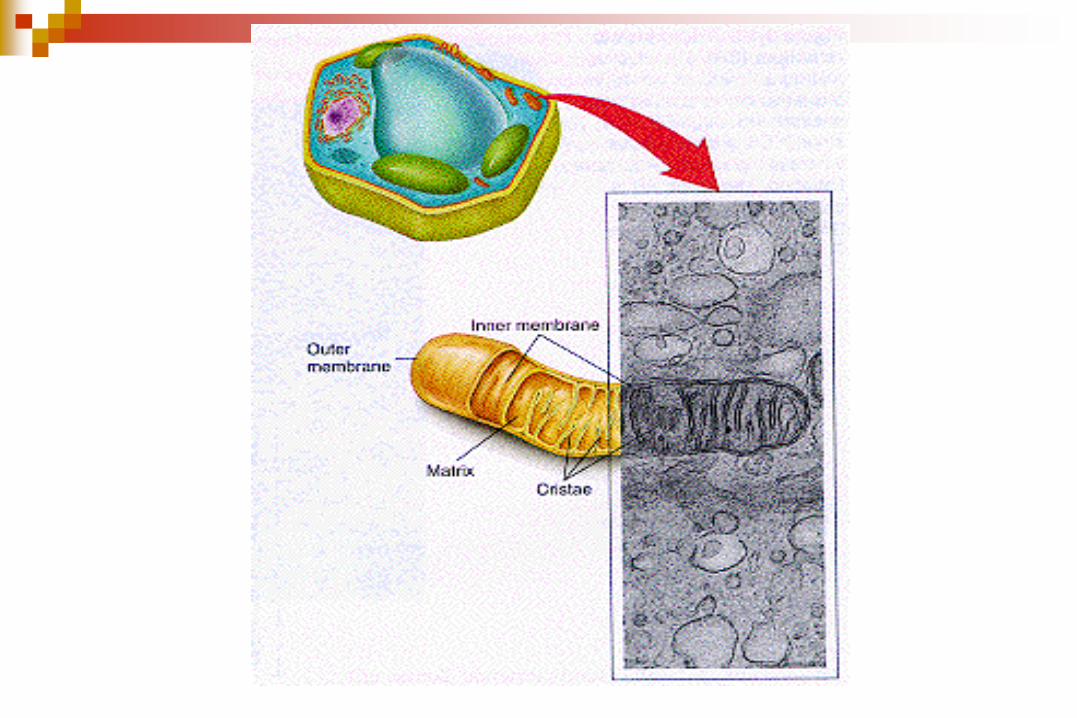
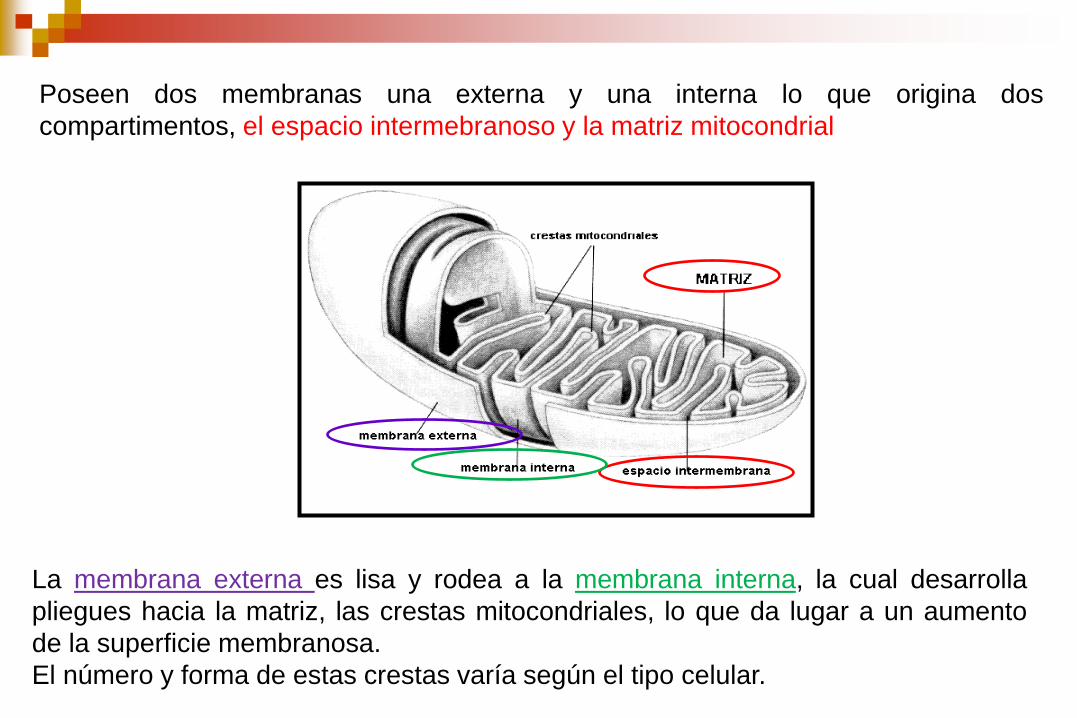
Poseen dos membranas una externa y una interna lo que origina dos
compartimentos, el espacio intermebranoso y la matriz mitocondrial
La membrana externa es lisa y rodea a la membrana interna, la cual desarrolla
pliegues hacia la matriz, las crestas mitocondriales, lo que da lugar a un aumento
de la superficie membranosa.
El número y forma de estas crestas varía según el tipo celular.
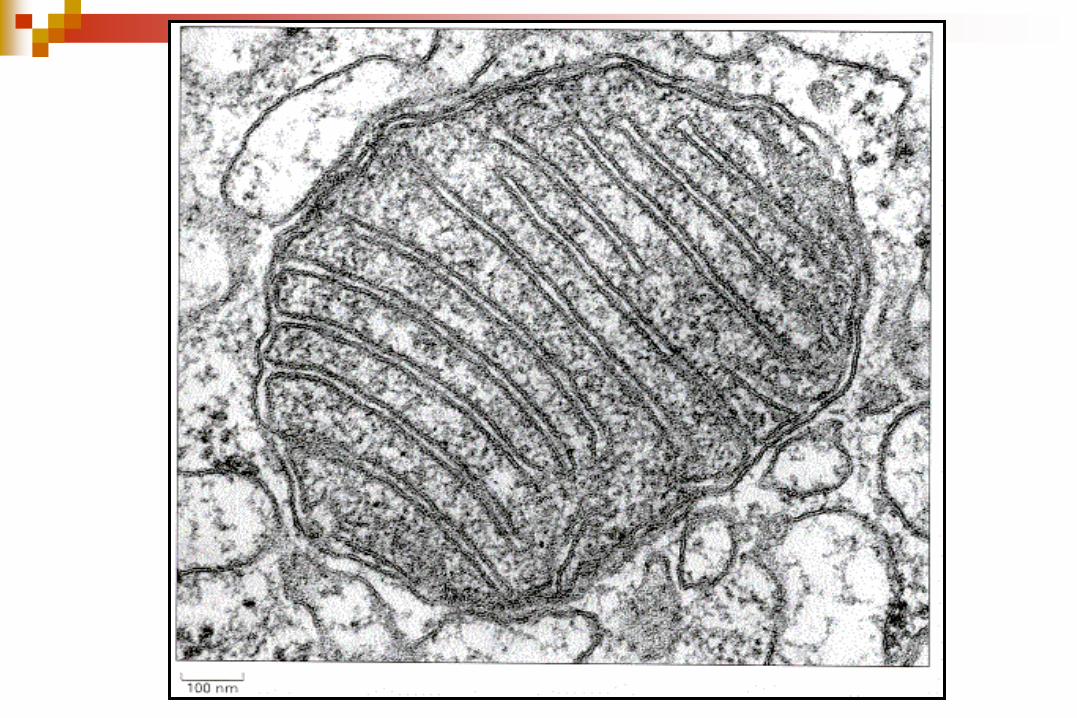
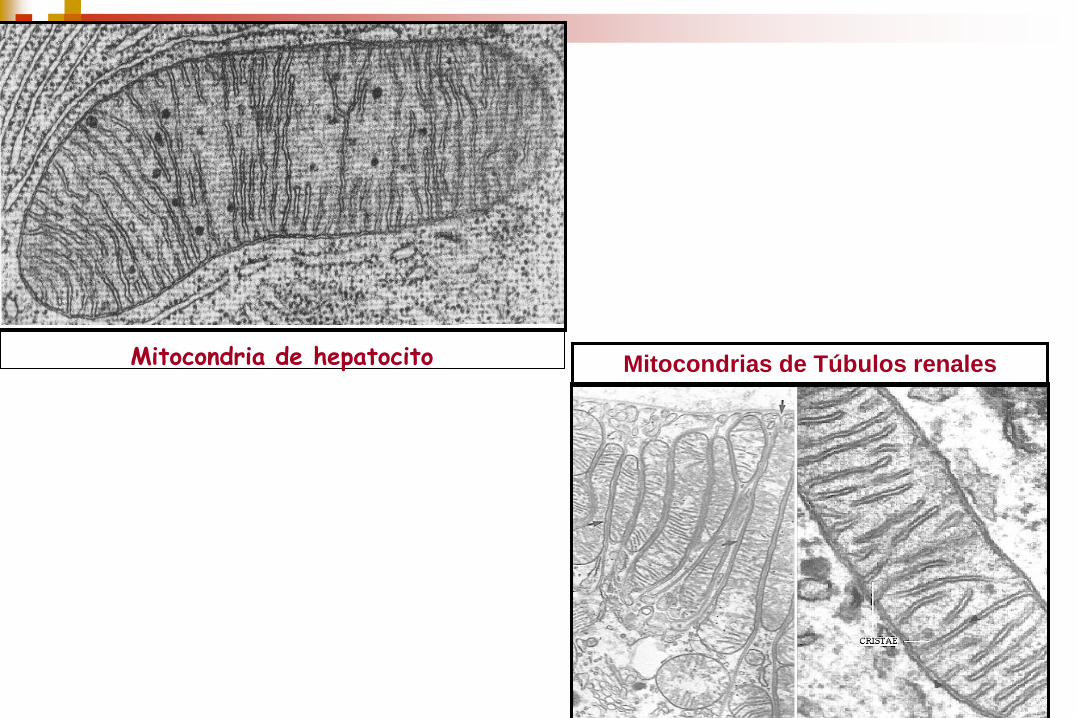
Mitocondrias de Túbulos renales Mitocondria de hepatocito
Una sonrisa para la Foto

CARACTERÍSTICAS Y COMPOSICIÓN DE LOS
COMPARTIMIENTOS MITOCONDRIALES
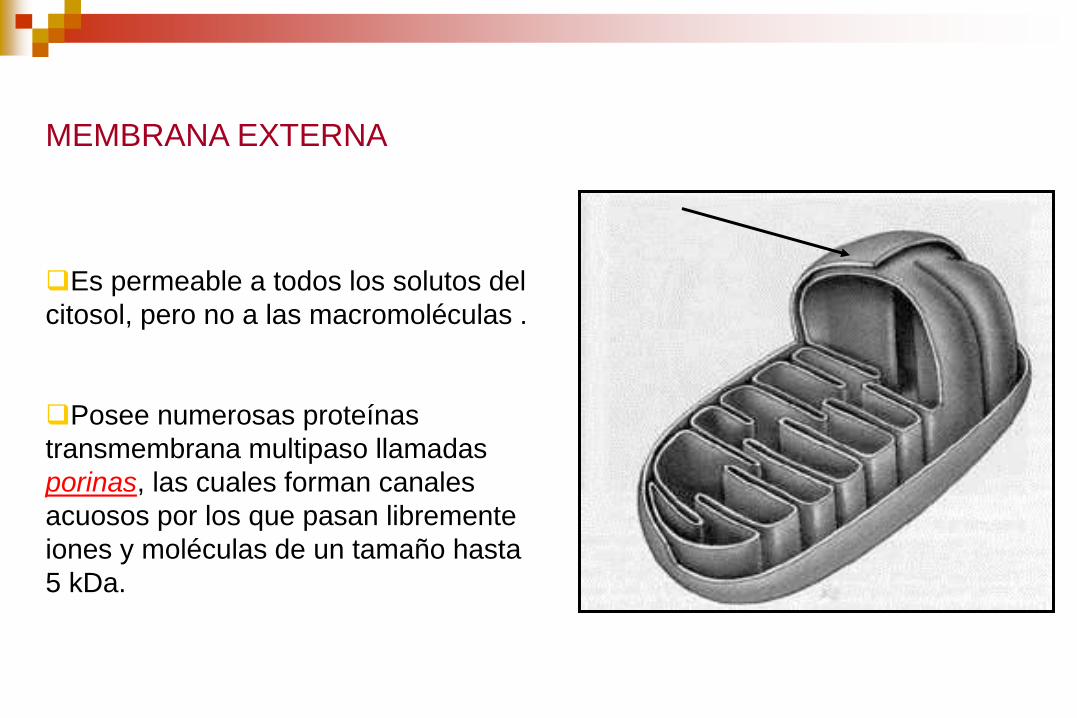
MEMBRANA EXTERNA
Es permeable a todos los solutos del
citosol, pero no a las macromoléculas .
Posee numerosas proteínas
transmembrana multipaso llamadas
porinas, las cuales forman canales
acuosos por los que pasan libremente
iones y moléculas de un tamaño hasta
5 kDa.
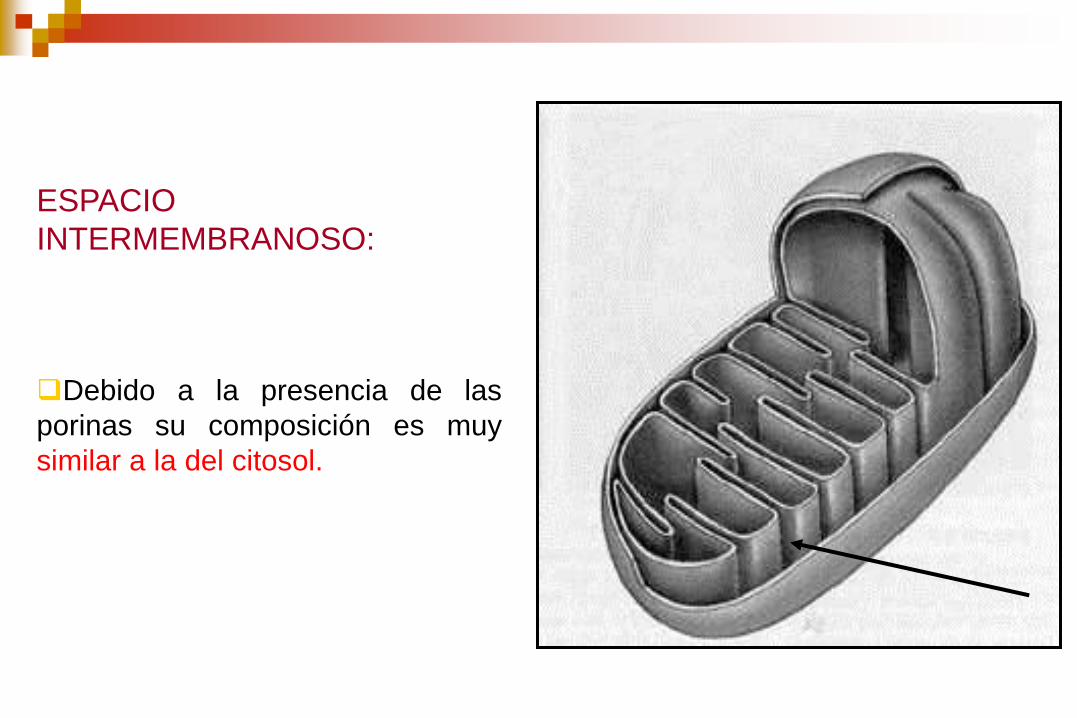
ESPACIO
INTERMEMBRANOSO:
Debido a la presencia de las
porinas su composición es muy
similar a la del citosol.
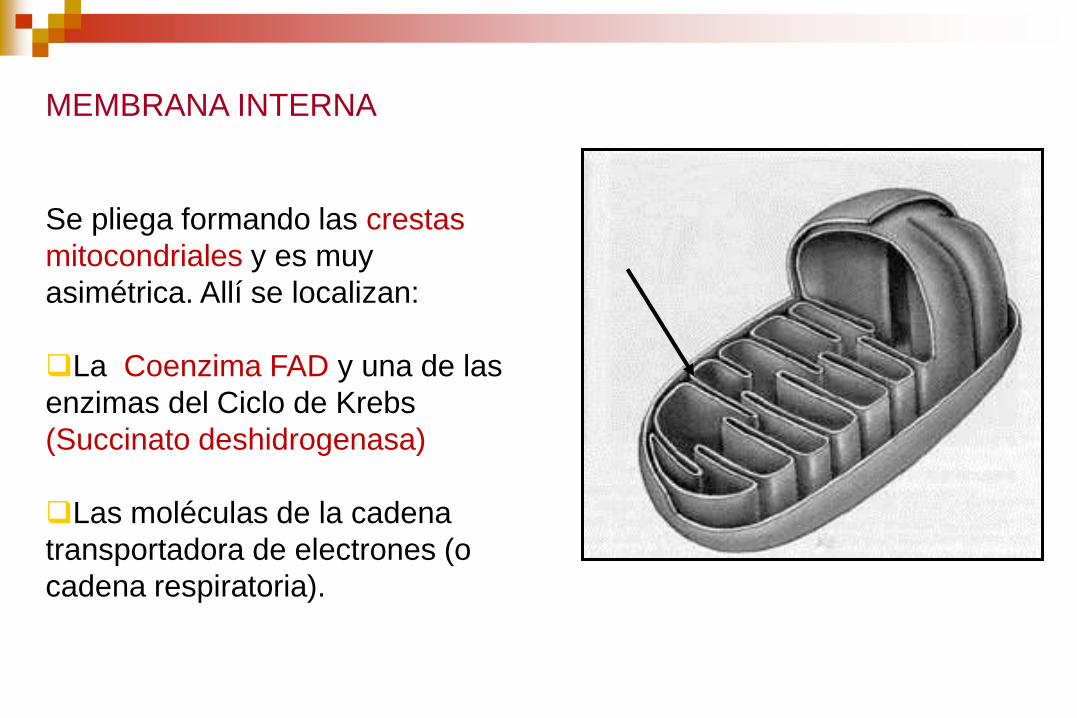
MEMBRANA INTERNA
Se pliega formando las crestas
mitocondriales y es muy
asimétrica. Allí se localizan:
La Coenzima FAD y una de las
enzimas del Ciclo de Krebs
(Succinato deshidrogenasa)
Las moléculas de la cadena
transportadora de electrones (o
cadena respiratoria).

Además allí se ubican:
La ATP sintetasa. Complejo proteico que presenta dos sectores:
uno transmembrna (F0) que tiene un canal protónico y otro orientado
hacia la matriz (F1) , este último cataliza la formación de ATP a partir
de ADP y Fosfato (Fosforilación Oxidativa)
La Cardiolipina un fosfolípido doble que impide el paso de
cualquier soluto a través de la bicapa , excepto O2, CO2, H2O y NH3
Diversos canales iónicos y permeasas, que permiten el pasaje
selectivo de iones y moléculas desde el espacio intermembranoso a la
matriz y viceversa

MATRIZ MITOCONDRIAL
Allí se encuentran numerosas moléculas entre
ellas:
Varias copias de ADN circular (13)
Trece(13) tipos de ARNm sintetizados a
partir de otros tantos genes de ese ADN.
Dos tipos de ARNr (ribosómico).
Veintidos (22) tipos de ARNt (de
transferencia).

ADEMÁS
Gránulos de distintos tamaños compuestos principalmente por Ca2+
El complejo enzimático piruvato deshidrogenasa
Las enzimas de la -oxidación de los ácidos grasos.
Las enzimas del Ciclo de Krebs, excepto la succinato deshidrogenasa.
La Coenzima Acetilante, (Acetil CoA), la coenzima NAD+, ADP Fosfato
y O2

Las mitocondrias se dividen para reemplazar a las que
desaparecen o son degradadas cuando ya han
envejecido. También su número se duplica antes de
cada división celular.
La reproducción se produce por la división de
mitocondrias preexistentes.
Este proceso al igual que la división de las bacterias se
denomina fisión binaria.
No todas ellas se dividen, de tal manera que algunas
deben dividirse varias veces para compensar este
hecho.
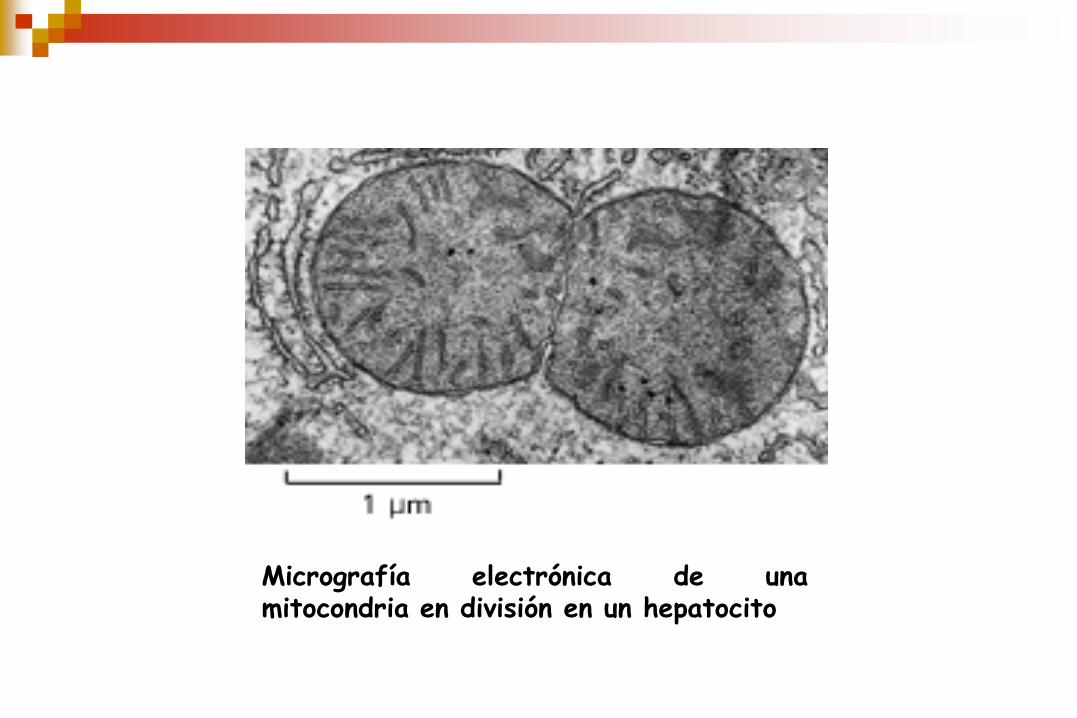
Micrografía electrónica de una mitocondria en división en un hepatocito
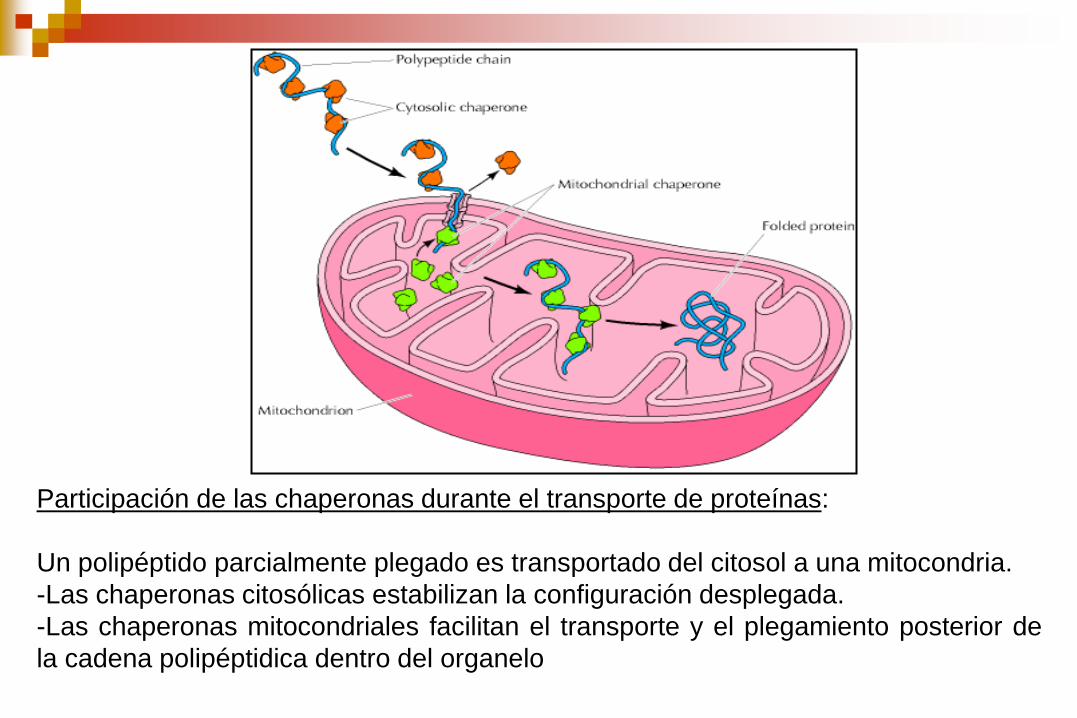
Participación de las chaperonas durante el transporte de proteínas:
Un polipéptido parcialmente plegado es transportado del citosol a una mitocondria.
-Las chaperonas citosólicas estabilizan la configuración desplegada.
-Las chaperonas mitocondriales facilitan el transporte y el plegamiento posterior de
la cadena polipéptidica dentro del organelo
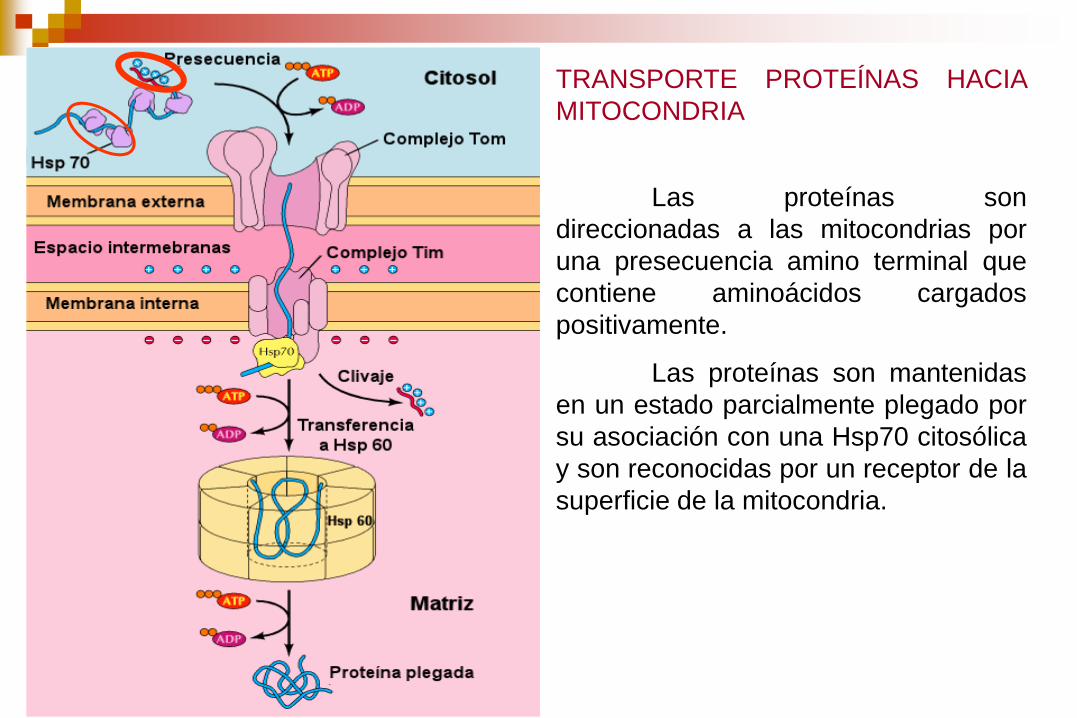
TRANSPORTE PROTEÍNAS HACIA
MITOCONDRIA
Las proteínas son
direccionadas a las mitocondrias por
una presecuencia amino terminal que
contiene aminoácidos cargados
positivamente.
Las proteínas son mantenidas
en un estado parcialmente plegado por
su asociación con una Hsp70 citosólica
y son reconocidas por un receptor de la
superficie de la mitocondria.
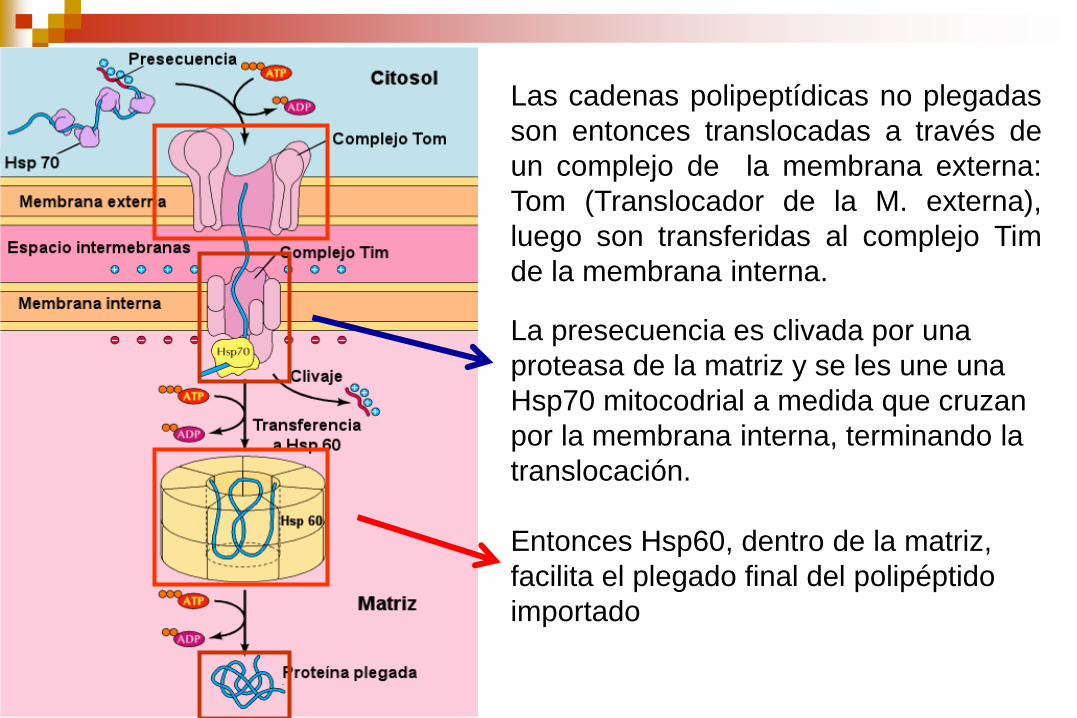
Las cadenas polipeptídicas no plegadas
son entonces translocadas a través de
un complejo de la membrana externa:
Tom (Translocador de la M. externa),
luego son transferidas al complejo Tim
de la membrana interna.
La presecuencia es clivada por una
proteasa de la matriz y se les une una
Hsp70 mitocodrial a medida que cruzan
por la membrana interna, terminando la
translocación.
Entonces Hsp60, dentro de la matriz,
facilita el plegado final del polipéptido
importado
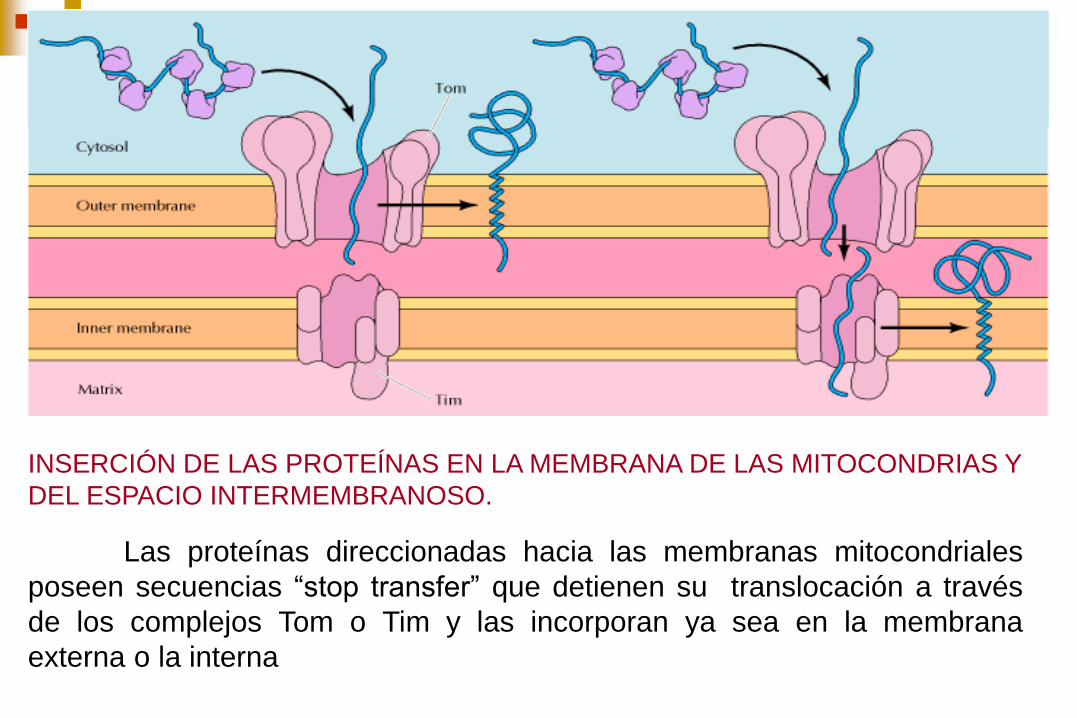
INSERCIÓN DE LAS PROTEÍNAS EN LA MEMBRANA DE LAS MITOCONDRIAS Y
DEL ESPACIO INTERMEMBRANOSO.
Las proteínas direccionadas hacia las membranas mitocondriales
poseen secuencias “stop transfer” que detienen su translocación a través
de los complejos Tom o Tim y las incorporan ya sea en la membrana
externa o la interna
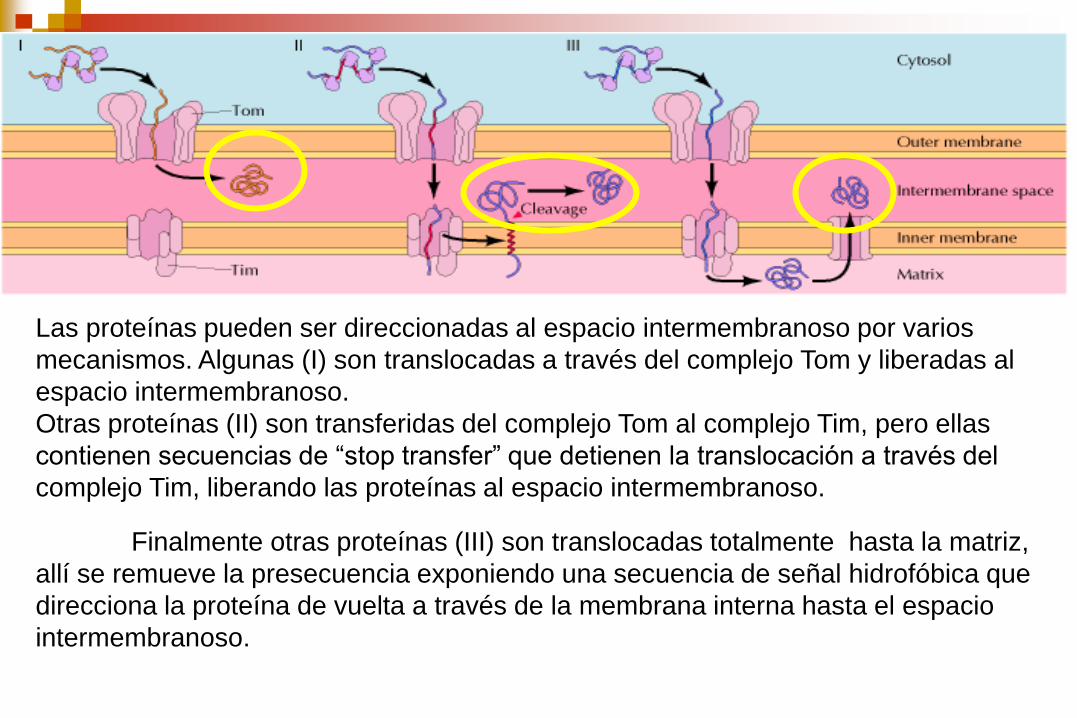
Las proteínas pueden ser direccionadas al espacio intermembranoso por varios
mecanismos. Algunas (I) son translocadas a través del complejo Tom y liberadas al
espacio intermembranoso.
Otras proteínas (II) son transferidas del complejo Tom al complejo Tim, pero ellas
contienen secuencias de “stop transfer” que detienen la translocación a través del
complejo Tim, liberando las proteínas al espacio intermembranoso.
Finalmente otras proteínas (III) son translocadas totalmente hasta la matriz,
allí se remueve la presecuencia exponiendo una secuencia de señal hidrofóbica que
direcciona la proteína de vuelta a través de la membrana interna hasta el espacio
intermembranoso.

LA FUNCION PRINCIPAL DE LAS MITOCONDRIAS ES
GENERAR ATP.
Se necesita energía para realizar los siguientes trabajos:
Sintetizar y degradar moléculas.
Transportar solutos en contra de gradientes electroquímicos a
través de sus membranas.
Incorporar macromoléculas y partículas desde el exterior.
Secretar macromoléculas al medio extracelular.
Trasladarse o contraerse
Desplazar sus propios componentes.
Multiplicarse.
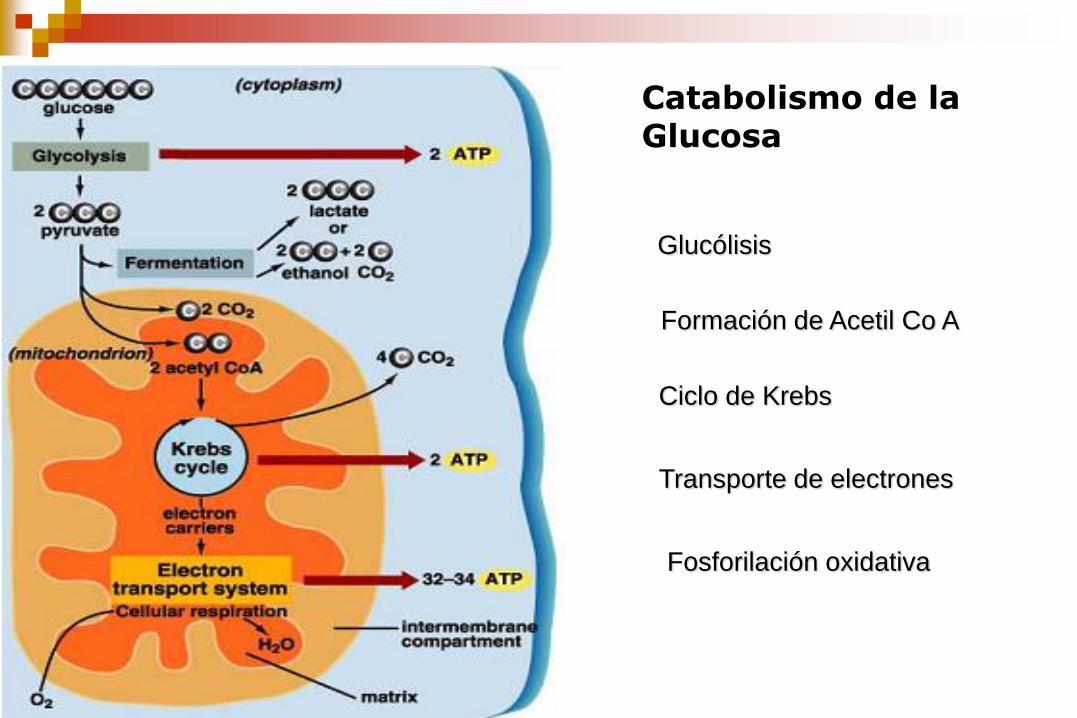
Catabolismo de la Glucosa
Glucólisis
Formación de Acetil Co A
Ciclo de Krebs
Transporte de electrones
Fosforilación oxidativa

CITOESQUELETO

Es una red filamentosa de proteínas que se extiende en
el citoplasma de todas las células eucariotas.
Proporciona un armazón estructural a la célula
determinando la forma y la organización general del
citoplasma.
Es una estructura dinámica y se reorganiza
continuamente conforme las células se mueven
GENERALIDADES

Compuesto por tres tipos principales de filamentos protéicos:
Filamentos de actina (microfilamentos) 7 nm
Filamentos Intermedios 10 nm
Microtúbulos 25 nm
se unen a la membrana plasmática, a los orgánulos y entre sí
mediante proteínas adaptadoras.
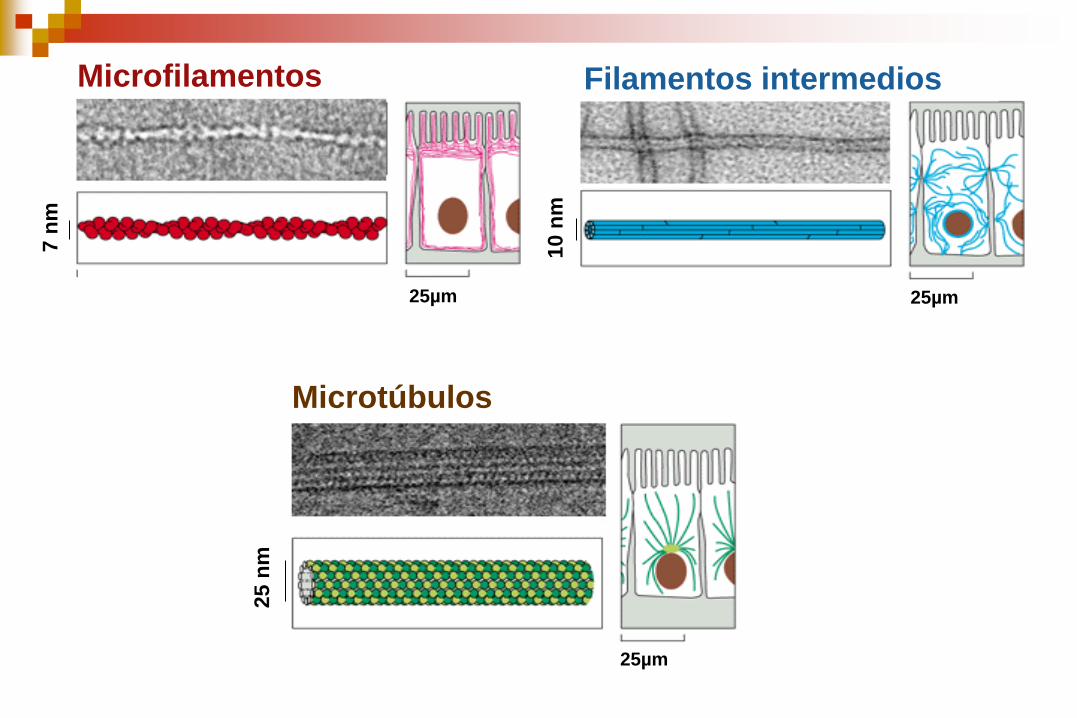
Microtúbulos
Microfilamentos Filamentos intermedios
25 n
m
10 n
m
7 n
m
25µm 25µm
25µm
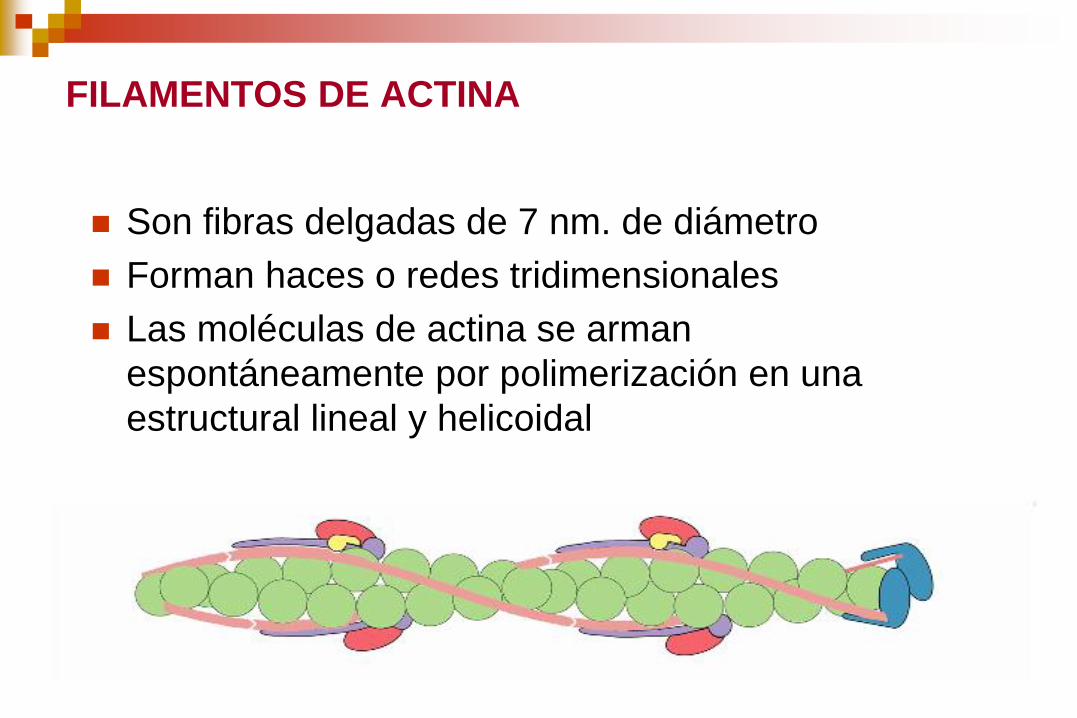
FILAMENTOS DE ACTINA
Son fibras delgadas de 7 nm. de diámetro
Forman haces o redes tridimensionales
Las moléculas de actina se arman
espontáneamente por polimerización en una
estructural lineal y helicoidal
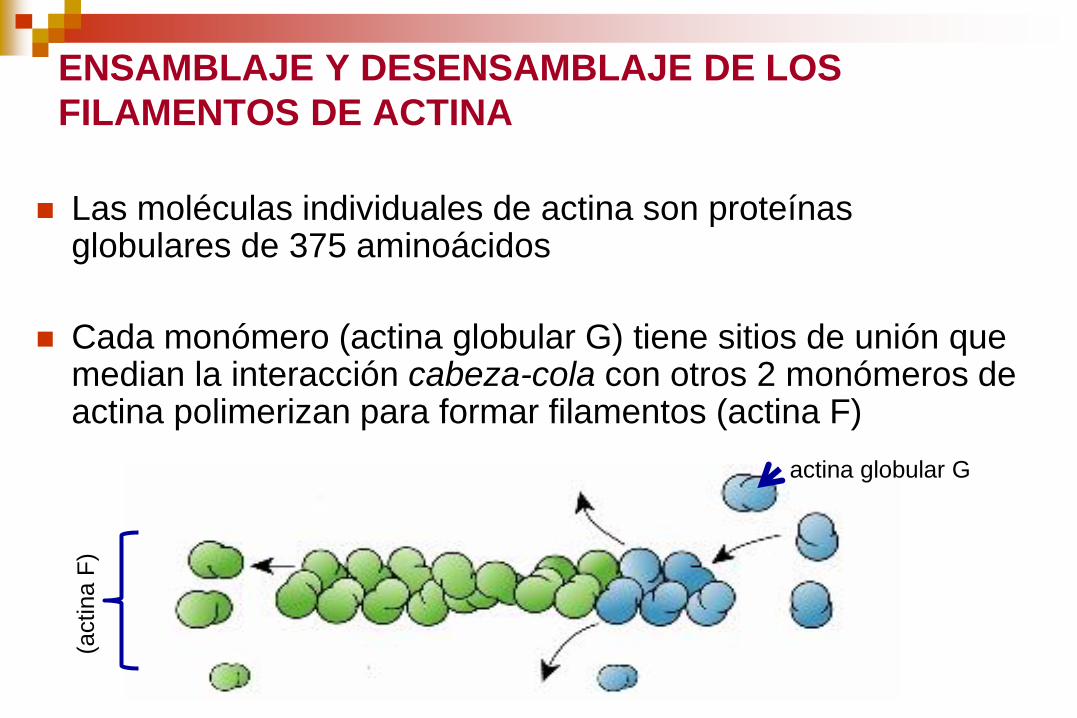
ENSAMBLAJE Y DESENSAMBLAJE DE LOS
FILAMENTOS DE ACTINA
Las moléculas individuales de actina son proteínas globulares de 375 aminoácidos
Cada monómero (actina globular G) tiene sitios de unión que median la interacción cabeza-cola con otros 2 monómeros de actina polimerizan para formar filamentos (actina F)
(actina F
)
actina globular G
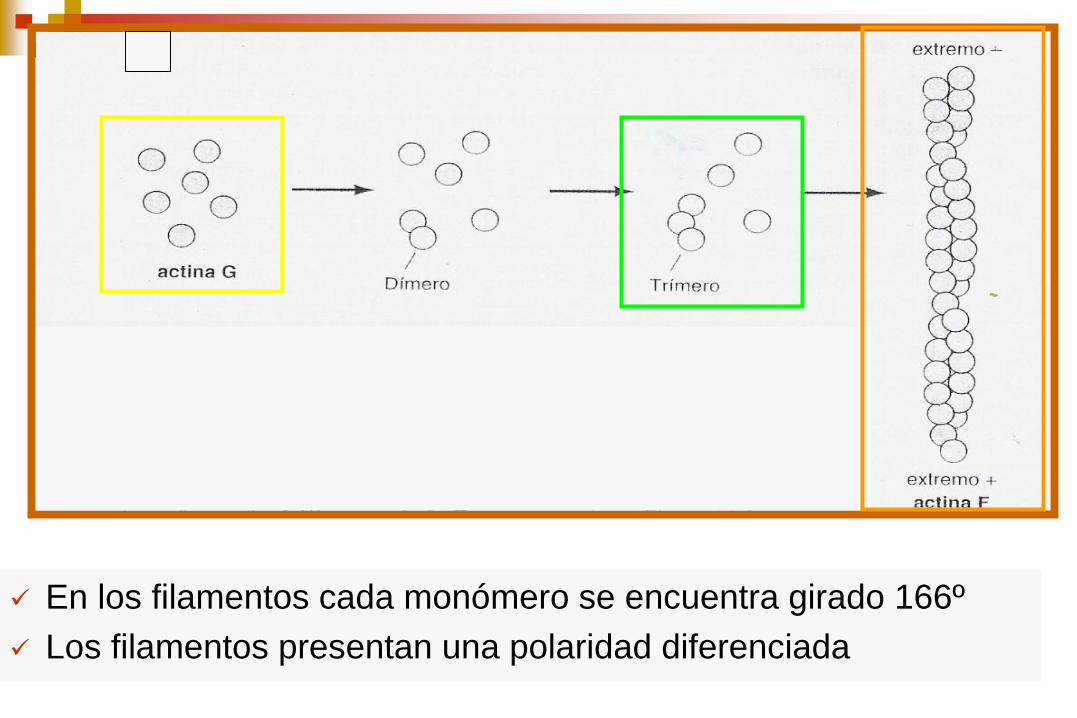
En los filamentos cada monómero se encuentra girado 166º
Los filamentos presentan una polaridad diferenciada

El primer paso en la polimerización denominado
nucleación es la formación de un pequeño
agregado constituido por tres monómeros de
actina.
Los monómeros de actina están unidos a ATP, el
cual se hidroliza a ADP tras el ensamblaje.
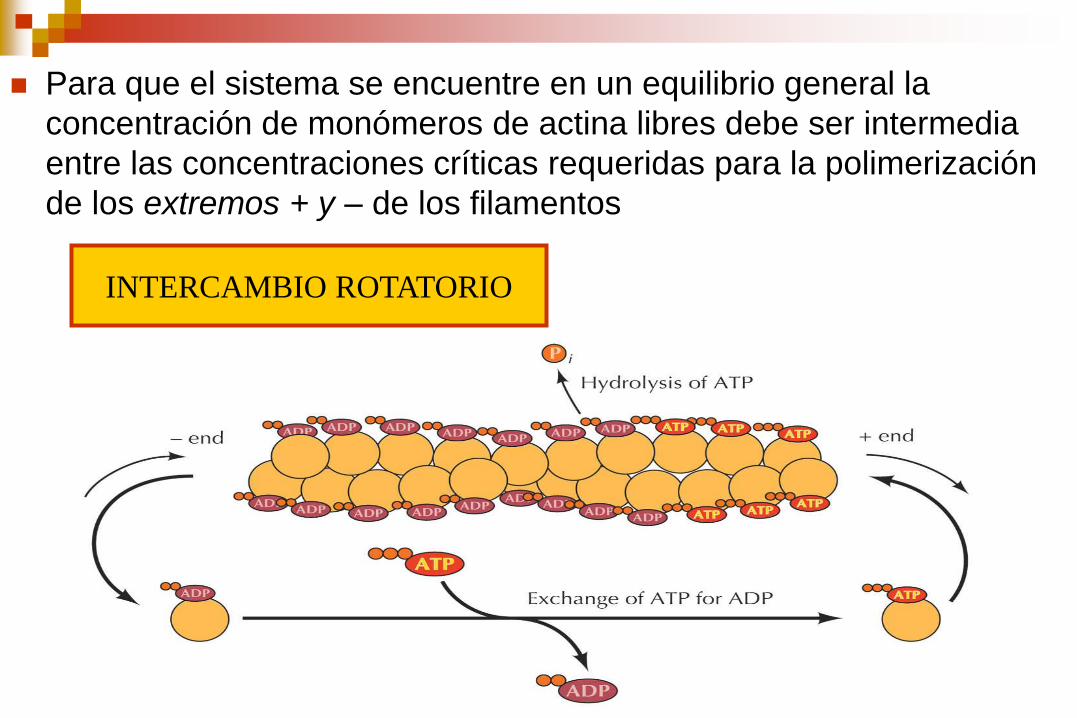
Para que el sistema se encuentre en un equilibrio general la
concentración de monómeros de actina libres debe ser intermedia
entre las concentraciones críticas requeridas para la polimerización
de los extremos + y – de los filamentos
INTERCAMBIO ROTATORIO

EFECTOS DE LAS PROTEÍNAS DE UNIÓN A LA ACTINA
SOBRE EL RECICLAJE DE LOS FILAMENTOS
La cofilina al unirse a los filamentos de actina aumenta la velocidad de disociación de los monómeros unidos al ADP. Además puede cortar los filamentos y generar mas extremos
La profilina puede estimular el intercambio de ADP unido por ATP
Además se pueden formar nuevos filamentos nucleados por las proteínas Arp2/3
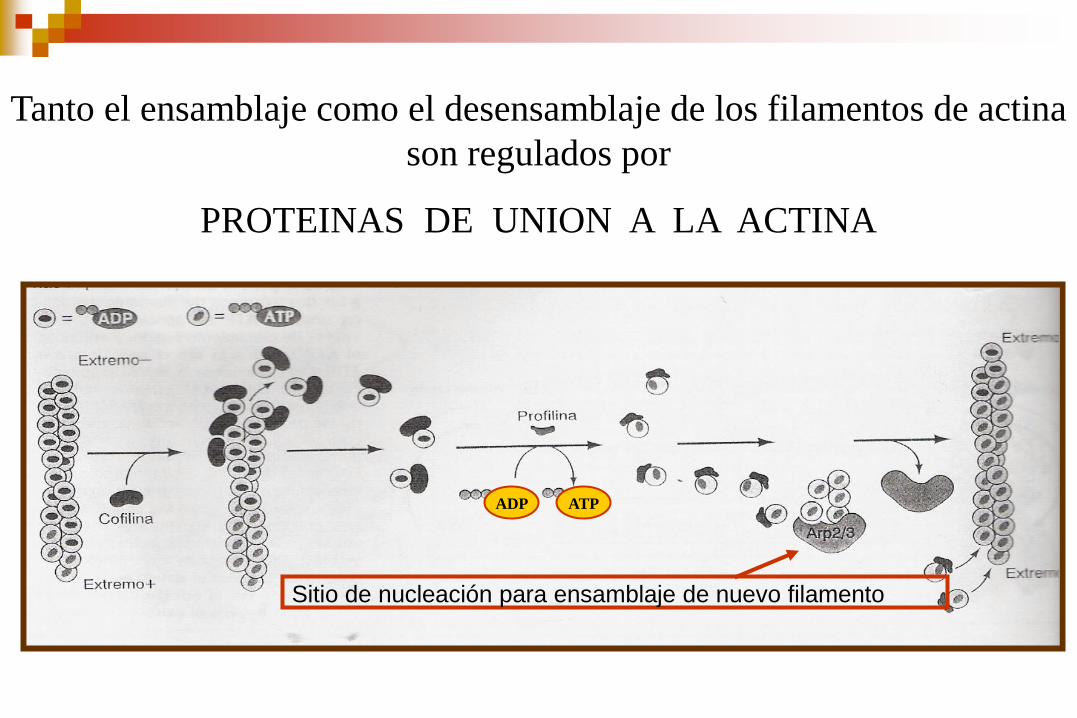
Sitio de nucleación para ensamblaje de nuevo filamento
ADP ATP
Tanto el ensamblaje como el desensamblaje de los filamentos de actina
son regulados por
PROTEINAS DE UNION A LA ACTINA
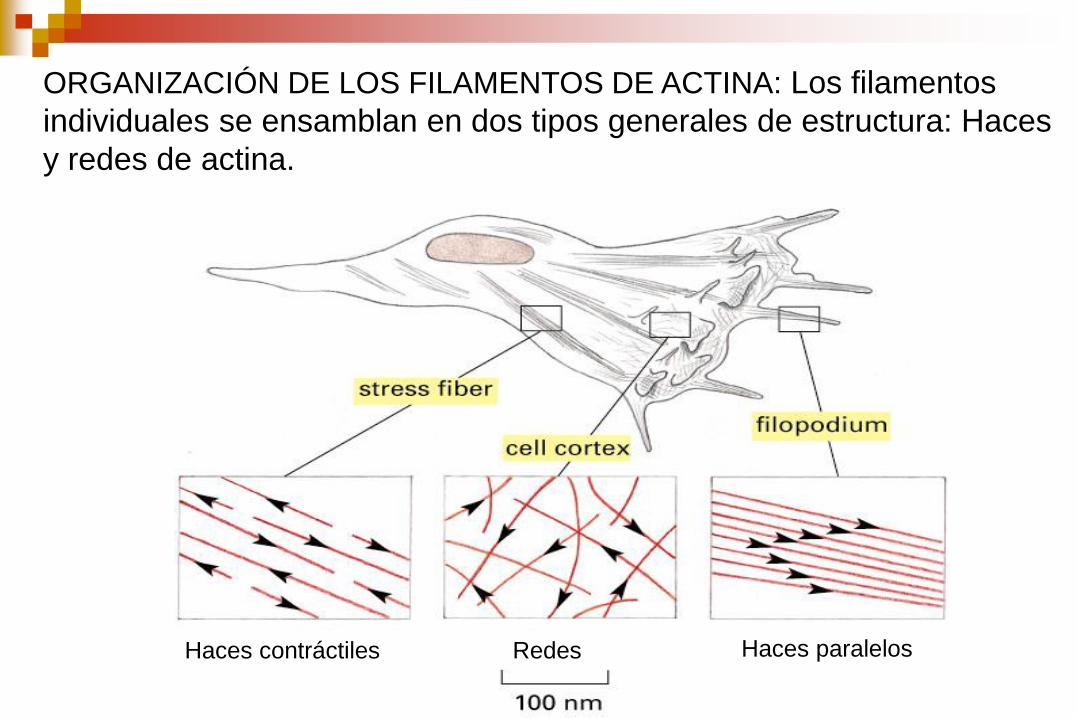
ORGANIZACIÓN DE LOS FILAMENTOS DE ACTINA: Los filamentos
individuales se ensamblan en dos tipos generales de estructura: Haces
y redes de actina.
Haces contráctiles Redes Haces paralelos
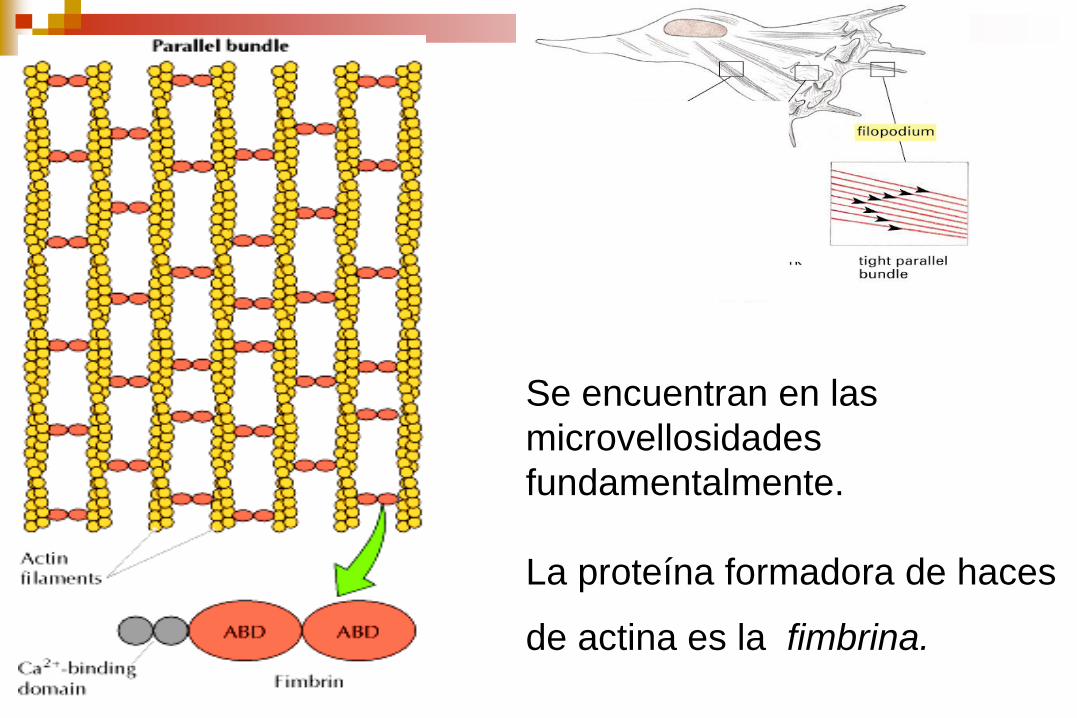
Se encuentran en las
microvellosidades
fundamentalmente.
La proteína formadora de haces
de actina es la fimbrina.
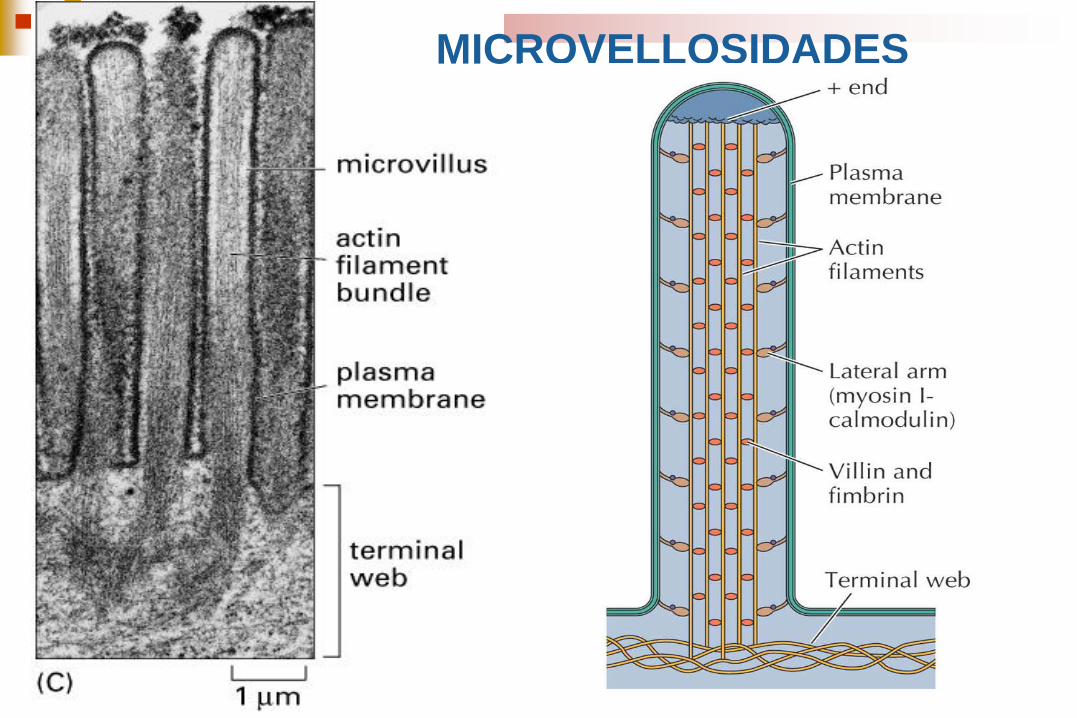
MICROVELLOSIDADES
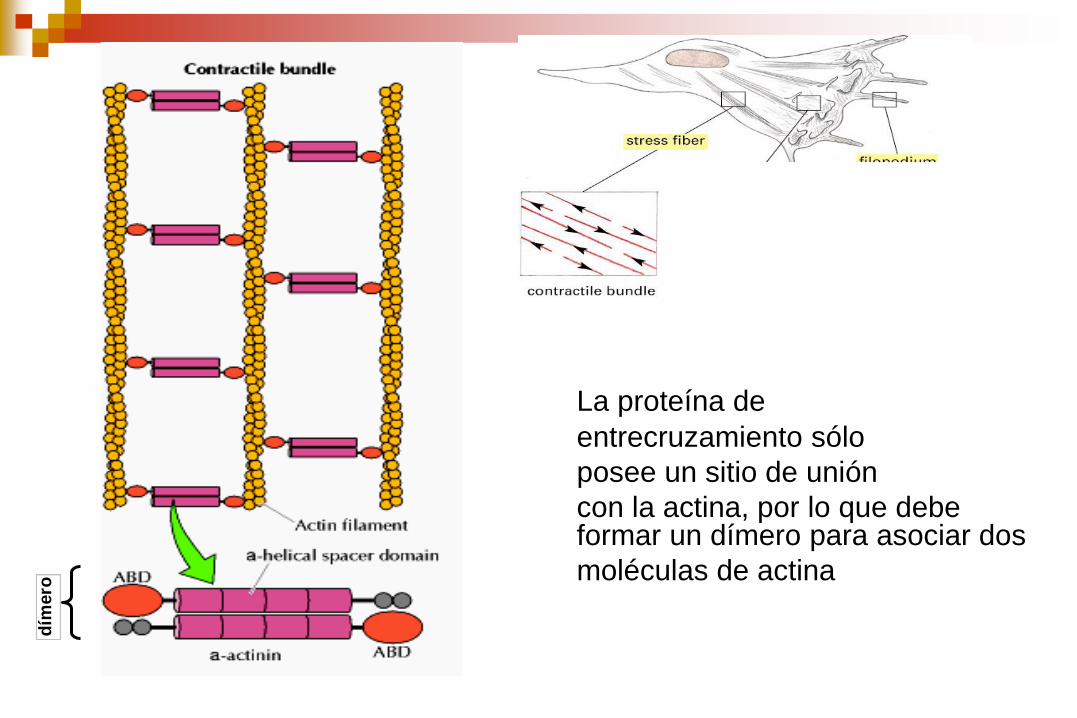
La proteína de
entrecruzamiento sólo
posee un sitio de unión
con la actina, por lo que debe formar un dímero para asociar dos
moléculas de actina
dím
ero
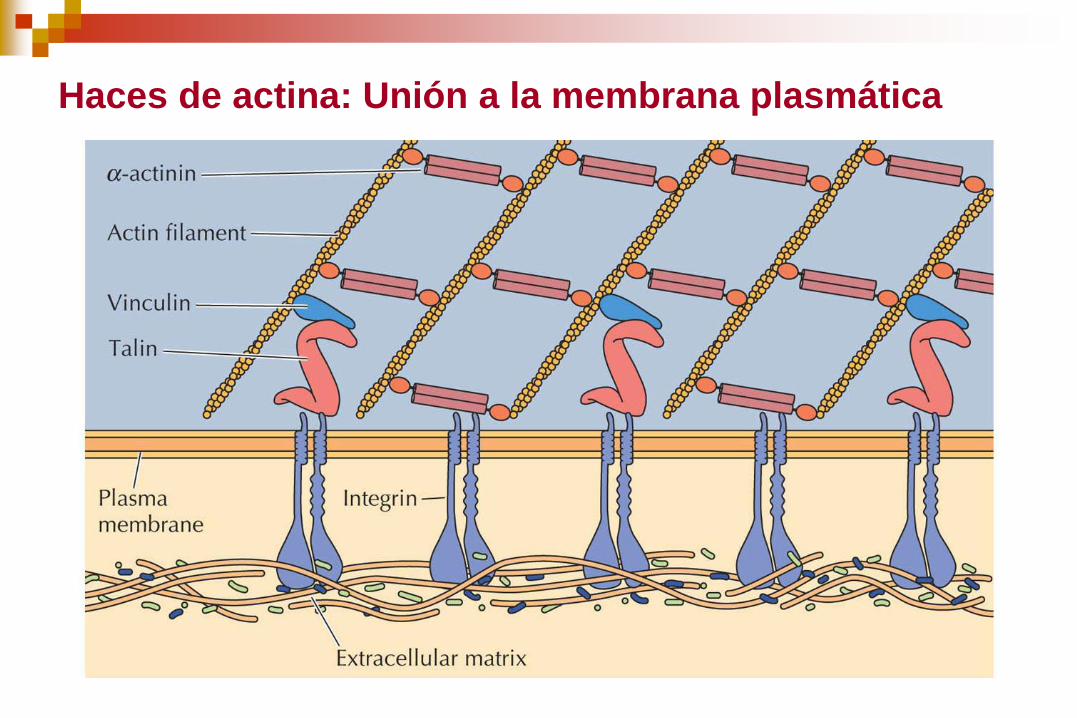
Haces de actina: Unión a la membrana plasmática
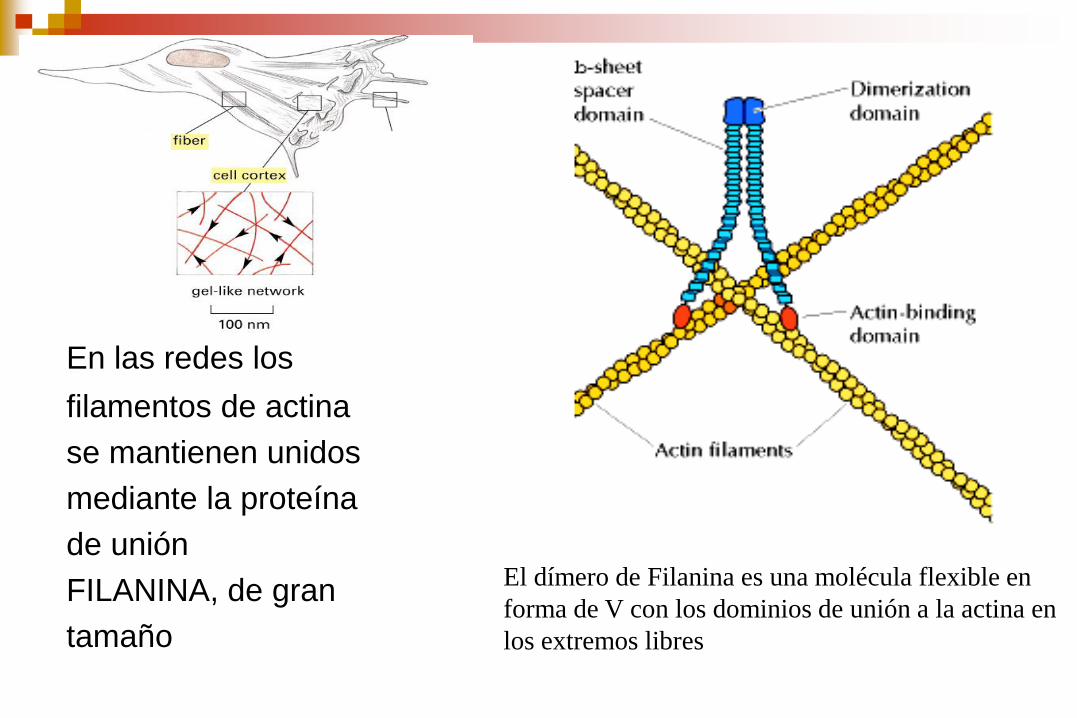
En las redes los
filamentos de actina
se mantienen unidos
mediante la proteína
de unión
FILANINA, de gran
tamaño
El dímero de Filanina es una molécula flexible en
forma de V con los dominios de unión a la actina en
los extremos libres

FUNCIONES
Anclaje y movimiento de proteínas de la membrana:
Se distribuye en forma de redes tridimensionales por
todas las células y se utilizan como estructuras de anclaje
en las uniones especializadas
Formación del núcleo estructural de microvellosidades
El velo terminal de filamentos de actina en la región apical
de la célula sirve para distribuir tensiones debajo de la
superficie celular

Locomoción celular
Se logra por la fuerza que ejercen los filamentos de actina al polimerizarse a la altura de sus extremos de crecimiento. Las extensiones del borde de avance de una célula reptante se denominan lamelipodios
Emisión de prolongaciones celulares: exhiben finas protrusiones llamadas filopodios ubicadas alrededor de la superficie celular
Contracción Celular
En interacción con la miosina generan fuerza y movimiento

FILAMENTOS INTERMEDIOS
Tienen un diámetro de 10 nm.
No están directamente
implicados en los
movimientos celulares
Desempeñan un papel
estructural proporcionando
resistencia mecánica a las
células y tejidos
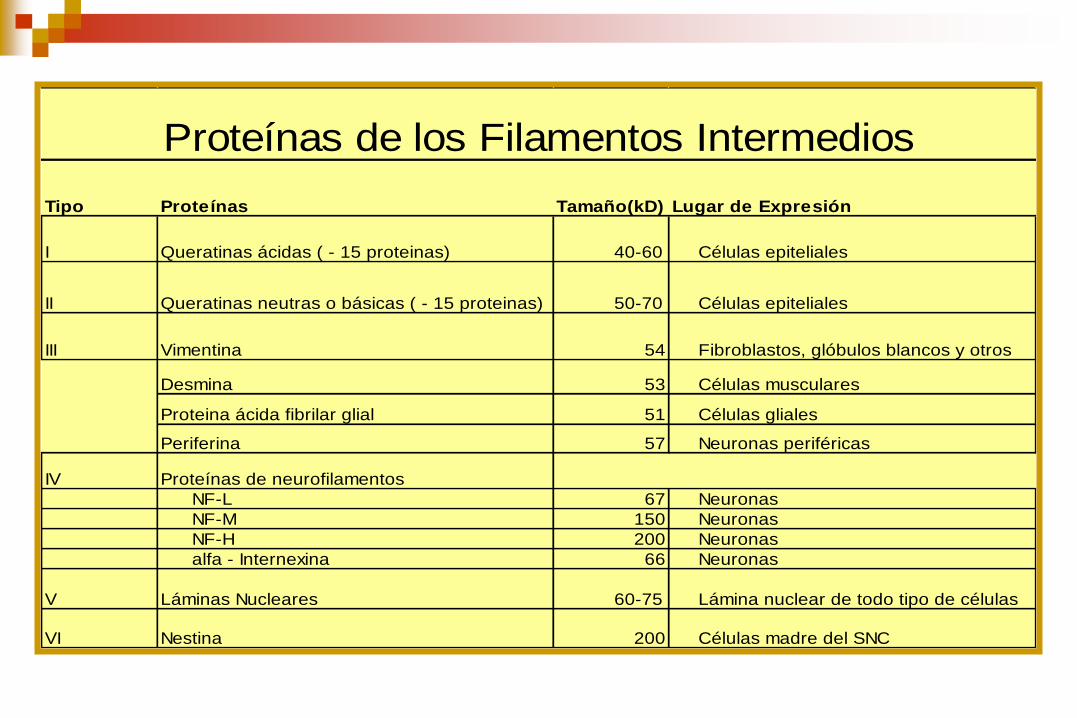
Tipo Proteínas Tamaño(kD) Lugar de Expresión
I Queratinas ácidas ( - 15 proteinas) 40-60 Células epiteliales
II Queratinas neutras o básicas ( - 15 proteinas) 50-70 Células epiteliales
III Vimentina 54 Fibroblastos, glóbulos blancos y otros
Desmina 53 Células musculares
Proteina ácida fibrilar glial 51 Células gliales
Periferina 57 Neuronas periféricas
IV Proteínas de neurofilamentos
NF-L 67 Neuronas
NF-M 150 Neuronas
NF-H 200 Neuronas
alfa - Internexina 66 Neuronas
V Láminas Nucleares 60-75 Lámina nuclear de todo tipo de células
VI Nestina 200 Células madre del SNC
Proteínas de los Filamentos Intermedios

ORGANIZACIÓN ESTRUCTURAL COMÚN
Todos tienen un dominio en alfa hélice con eje central de
aproximadamente 310 aminoácidos flanqueado por
dominios amino y carboxilo terminales que varían entre
las diferentes proteínas de filamentos intermedios en
tamaño, secuencia, estructura.
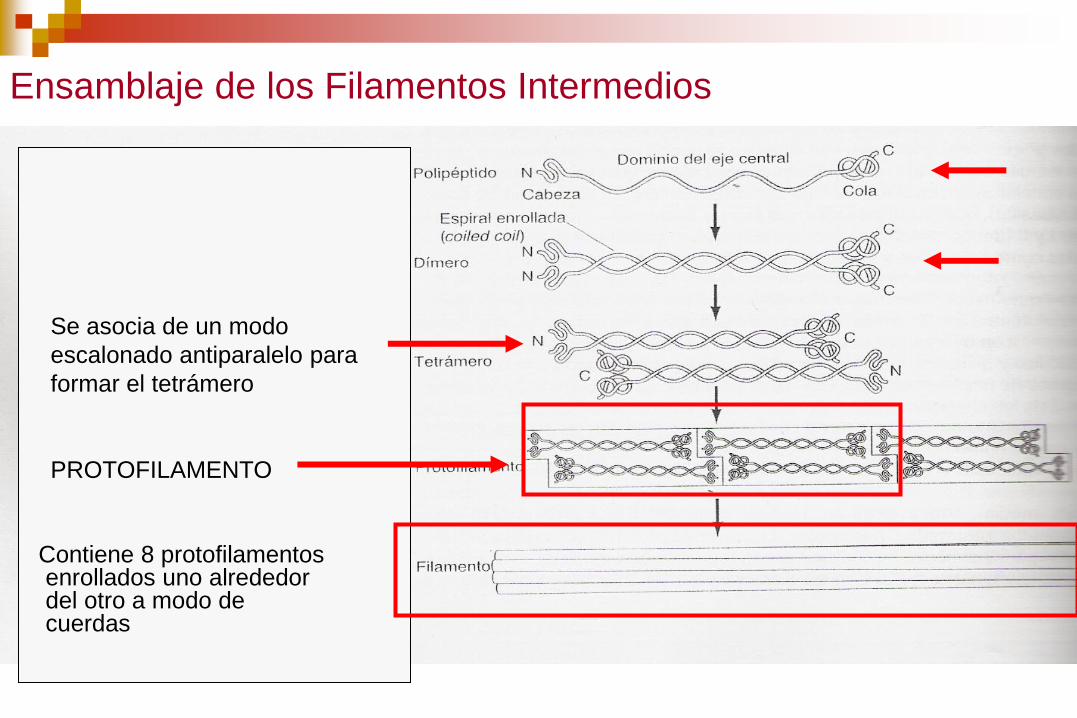
Ensamblaje de los Filamentos Intermedios
Se asocia de un modo
escalonado antiparalelo para
formar el tetrámero
PROTOFILAMENTO
Contiene 8 protofilamentos enrollados uno alrededor del otro a modo de cuerdas

Los filamentos intermedios son apolares, no tienen
diferenciados un extremo + y –
Son más estables. No exhiben el comportamiento
dinámico asociado a otros componentes del
citoesqueleto.
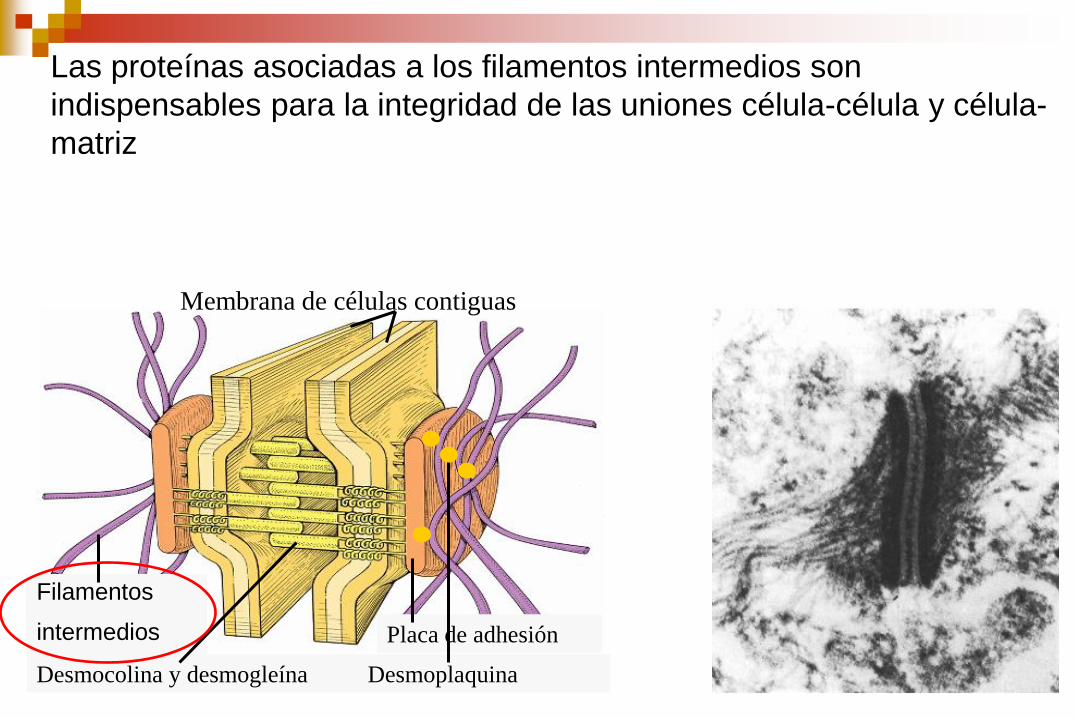
Las proteínas asociadas a los filamentos intermedios son
indispensables para la integridad de las uniones célula-célula y célula-
matriz
Membrana de células contiguas
Filamentos
intermedios Placa de adhesión
Desmocolina y desmogleína Desmoplaquina

MICROTÚBULOS
Varillas rígidas y huecas de 25 nm de diámetro
Pueden desensamblarse con rapidez en un sitio y
ensamblarse en otro
Crean un sistema de conexiones dentro de la célula
que guía el movimiento vesicular
Son estructuras polares con 2 extremos diferenciados
+ y -

Estructura, ensamblaje e inestabilidad dinámica de los
microtúbulos
Consiste en 13 protofilamentos
de moléculas globulares
diméricas de tubulina dispuestos
en forma circular.
La molécula alfa de un dímero
se une a la molécula beta del
dímero siguiente en un molde
que se repite, y el polímero
formado se llama
protofilamento
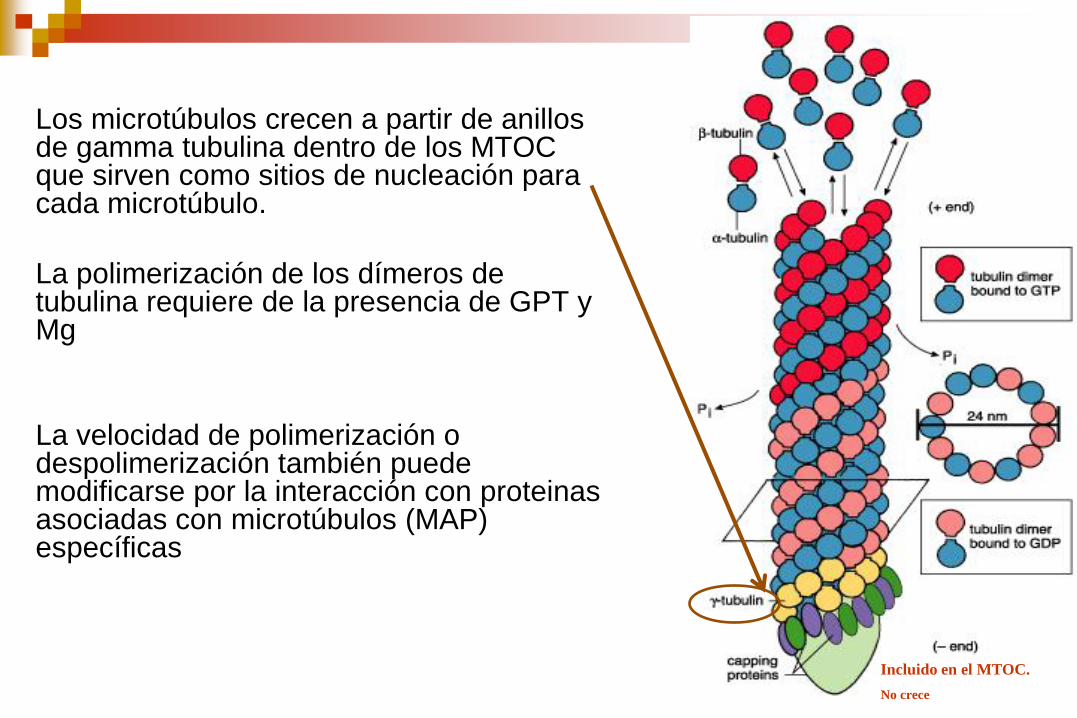
Los microtúbulos crecen a partir de anillos de gamma tubulina dentro de los MTOC que sirven como sitios de nucleación para cada microtúbulo.
La polimerización de los dímeros de tubulina requiere de la presencia de GPT y Mg
La velocidad de polimerización o despolimerización también puede modificarse por la interacción con proteinas asociadas con microtúbulos (MAP) específicas
Incluido en el MTOC.
No crece
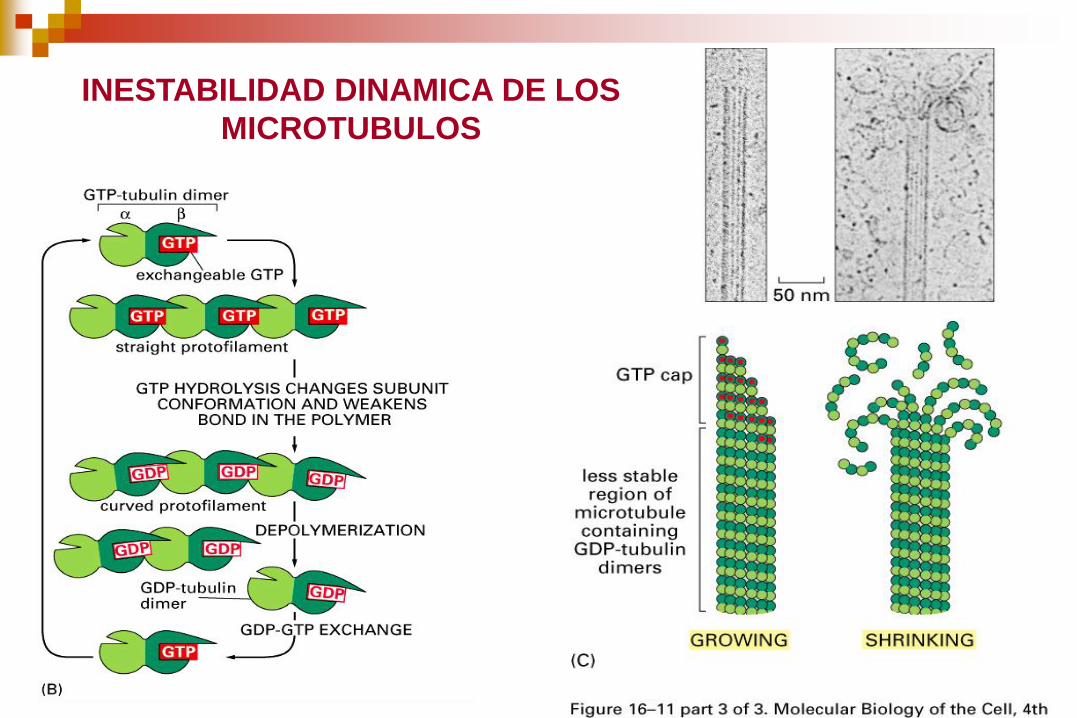
INESTABILIDAD DINAMICA DE LOS
MICROTUBULOS

La Longitud de los microtúbulos cambia dinámicamente
conforme se añaden o extraen dímeros de tubulina en el
fenómeno llamado inestabilidad dinámica
El MTOC o centrosoma
(constituido por los centríolos y
el material pericentriolar denso
y amorfo) controla la cantidad,
la dirección y la organización
de los microtúbulos

FUNCIONES
Transporte vesicular intracelular . Ej.: movimiento de vesículas de secreción, lisosomas
Movimientos de cilios y flagelos
Fijación de los cromosomas al huso mitótico y su movimiento durante la mitosis y la meiosis
Alargamiento y movimiento de las células
Mantenimiento de la forma celular, en particular de su asimetría
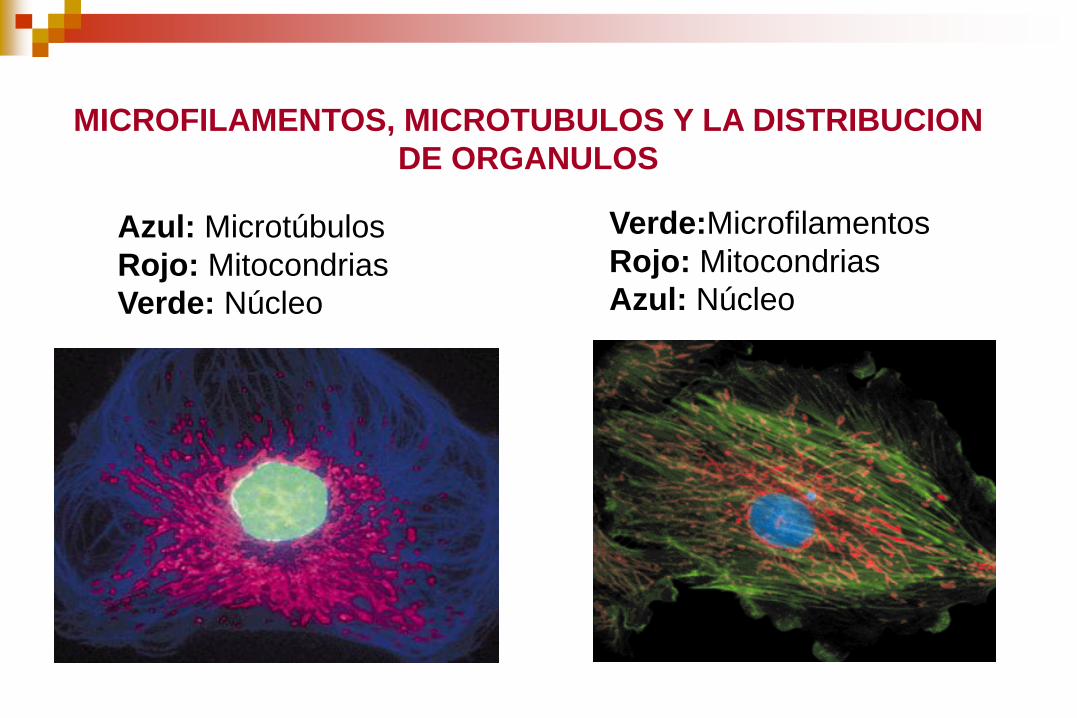
Azul: Microtúbulos
Rojo: Mitocondrias
Verde: Núcleo
Verde:Microfilamentos
Rojo: Mitocondrias
Azul: Núcleo
MICROFILAMENTOS, MICROTUBULOS Y LA DISTRIBUCION
DE ORGANULOS

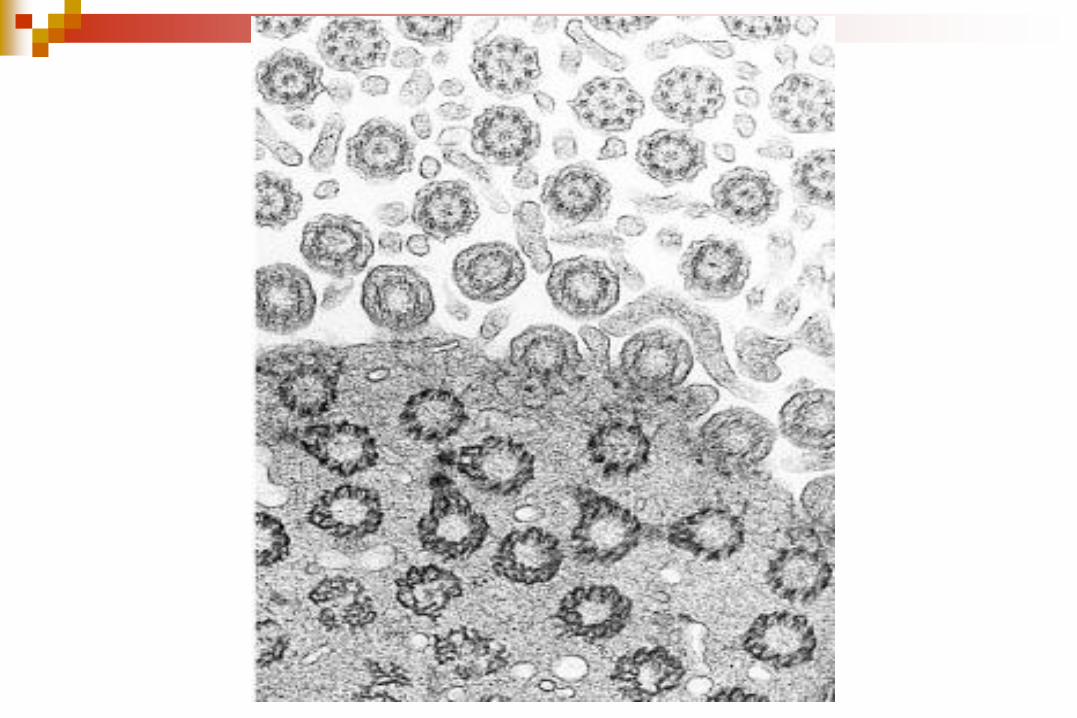
CILIOS Y FLAGELOS
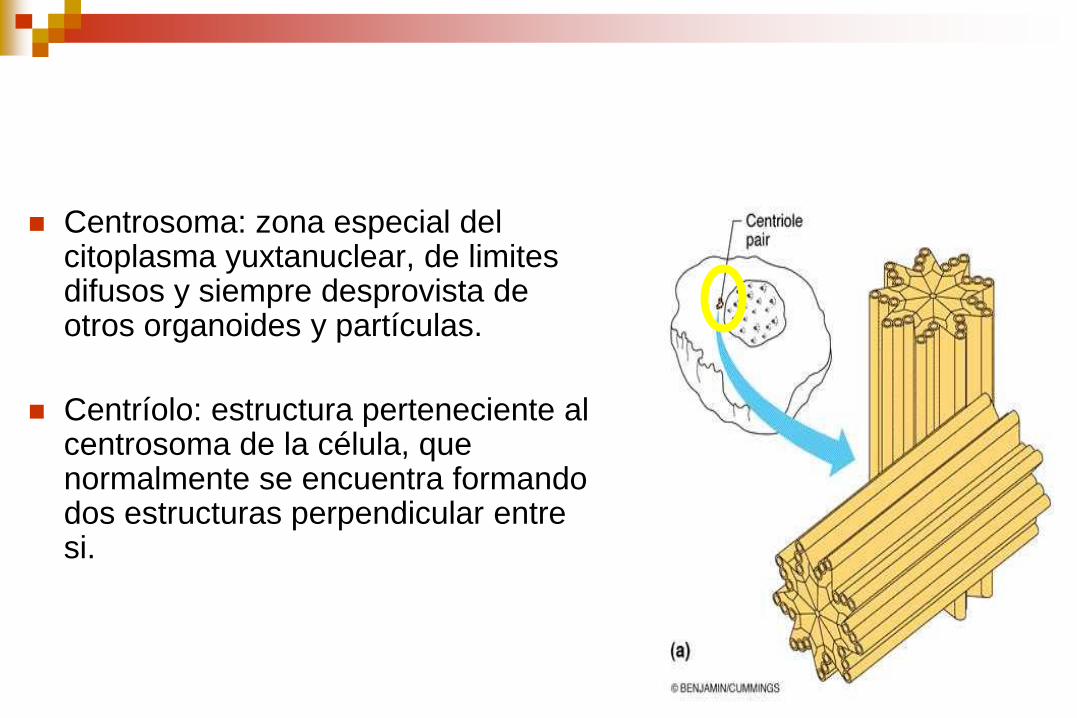
Centrosoma: zona especial del citoplasma yuxtanuclear, de limites difusos y siempre desprovista de otros organoides y partículas.
Centríolo: estructura perteneciente al centrosoma de la célula, que normalmente se encuentra formando dos estructuras perpendicular entre si.
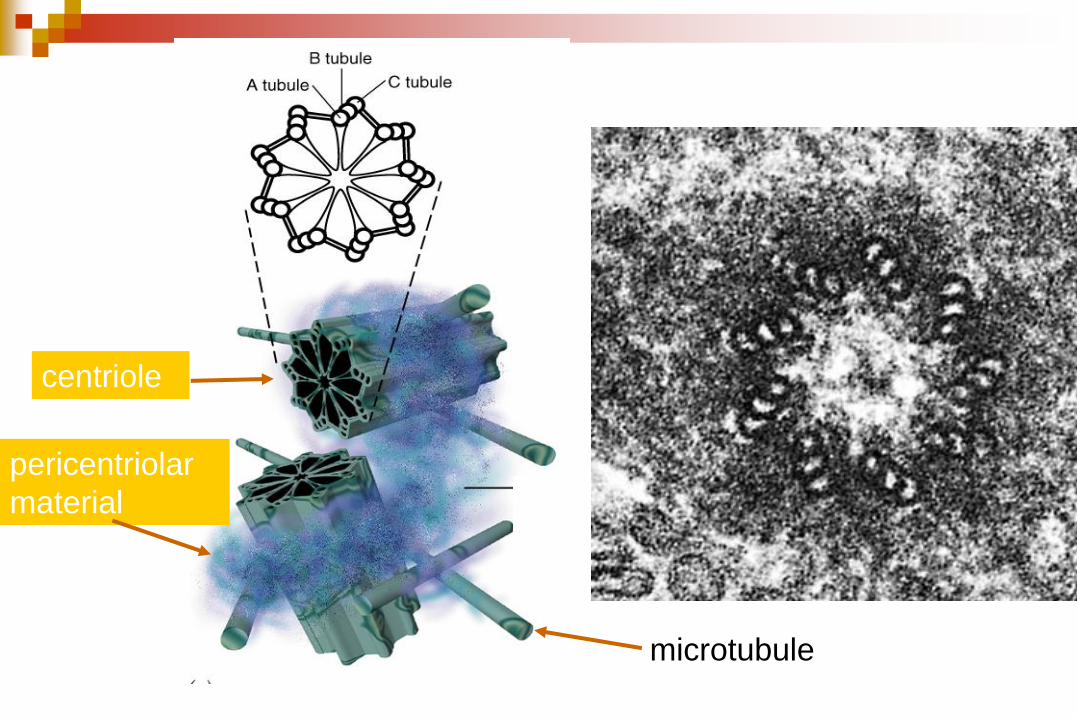
centriole
pericentriolar
material
microtubule
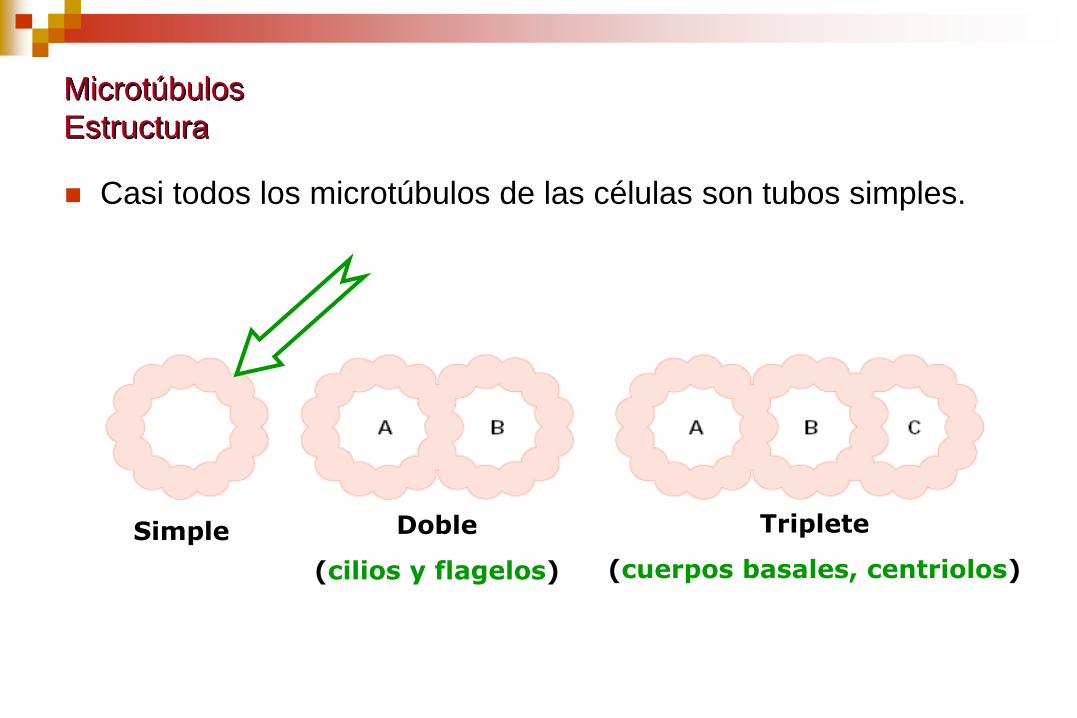
Microtúbulos
Estructura
Casi todos los microtúbulos de las células son tubos simples.
Simple Doble
(cilios y flagelos)
Triplete
(cuerpos basales, centriolos)

Microtúbulos
Estructura
En casos aislados se observan:
Microtúbulos dobles: cilias y flagelos
Microtúbulos triples: centriolos y cuerpos basales.
Contienen:
Un microtúbulo completo de 13 protofilamentos (túbulo A).
Uno o dos túbulos adicionales (B y C) compuestos por 10
protofilamentos.
1 2 3 4 5
6 7 8 9
10 11 12
13 1 2 4
3
5 6
8 9 10
1 2 3
5 6
8 9 10
4
7 7
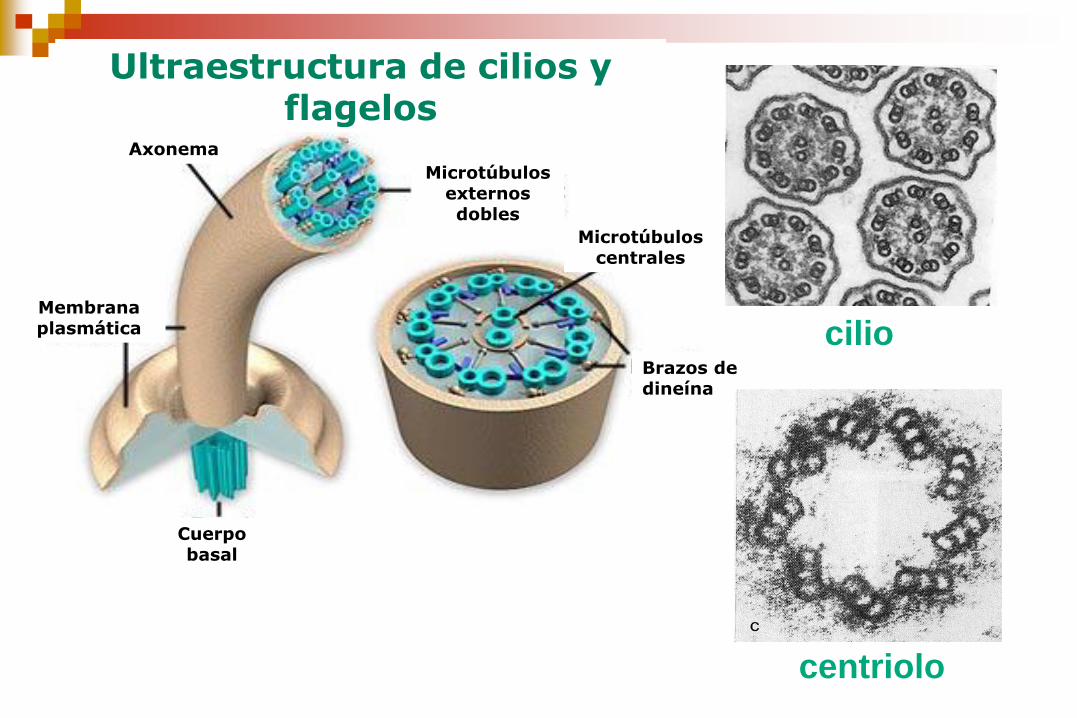
cilio
centriolo
Cuerpo basal
Membrana plasmática
Axonema
Microtúbulos externos dobles
Microtúbulos centrales
Brazos de dineína
Ultraestructura de cilios y flagelos

Centrosoma
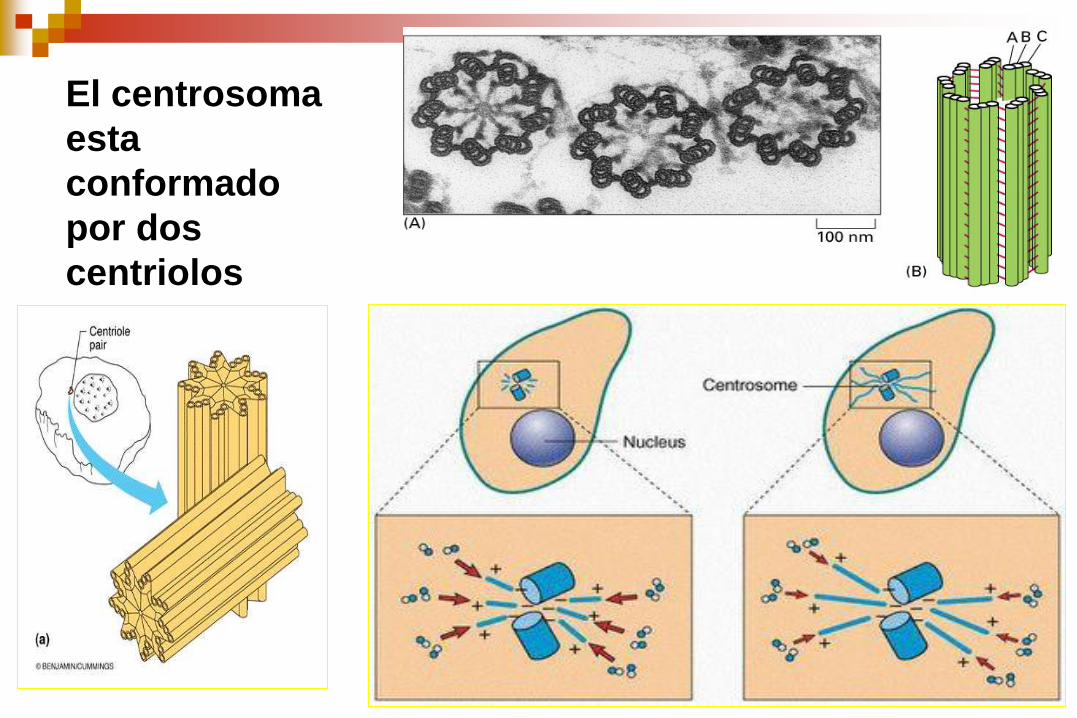
El centrosoma
esta
conformado
por dos
centriolos

Centrosoma
Estructura
El centrosoma contiene 2 centríolos.
Cada centríolo esta estructurado por microtúbulos triples dispuestos perpendicularmente.
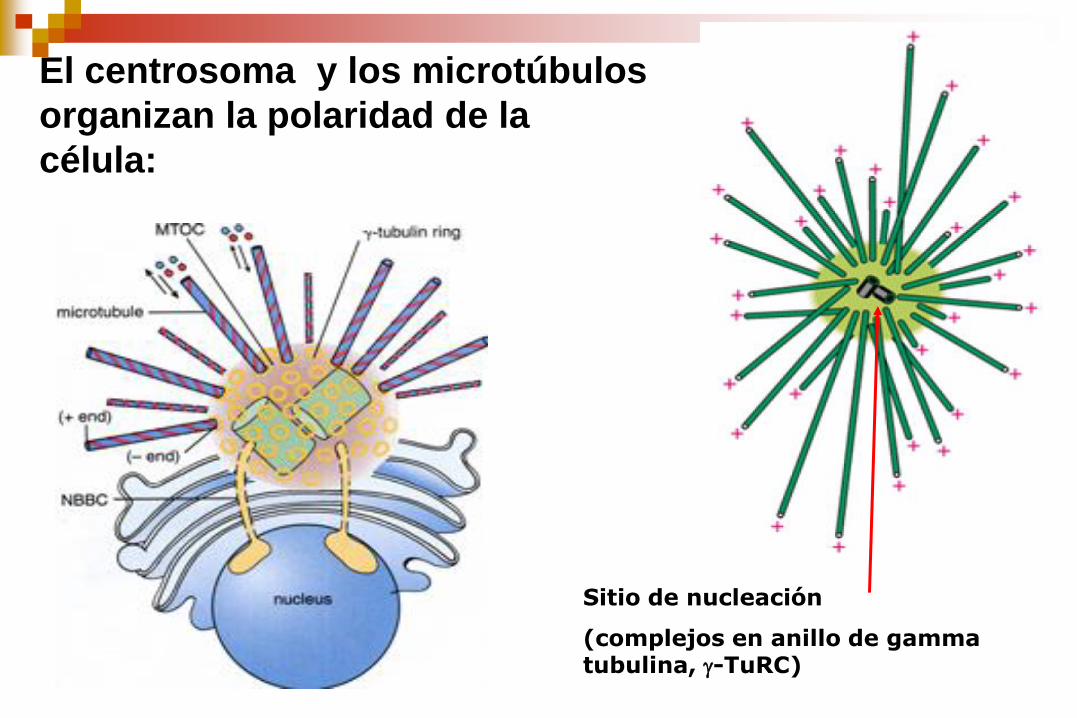
El centrosoma y los microtúbulos
organizan la polaridad de la
célula:
Sitio de nucleación
(complejos en anillo de gamma tubulina, -TuRC)

Presente en todas las células
eucariotas; excepto, plantas
superiores.
Participa en :
Organización de microtúbulos.
Coordinación movimientos de
cilios y flagelos.
Centrosoma
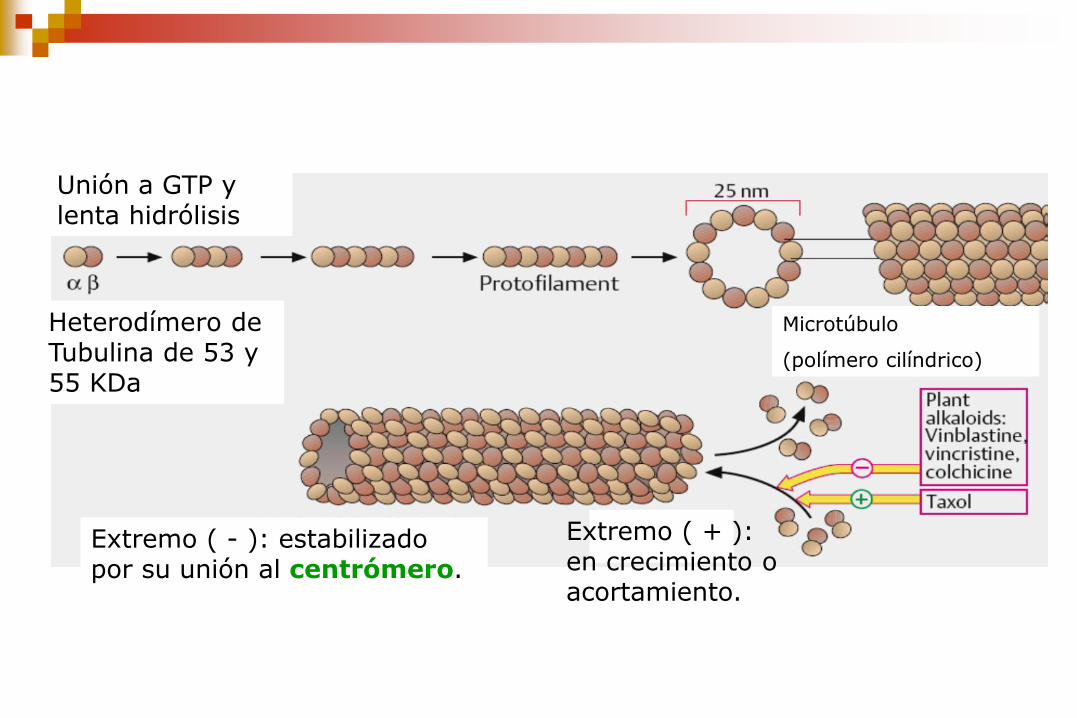
Extremo ( - ): estabilizado por su unión al centrómero.
Heterodímero de Tubulina de 53 y 55 KDa
Extremo ( + ): en crecimiento o acortamiento.
Unión a GTP y lenta hidrólisis
Microtúbulo
(polímero cilíndrico)

Cilios y flagelos

Cilios y flagelos
Varias células eucariotas tienen prolongaciones de su
citoplasma que les permite el movimiento en un medio
líquido:
Cilios
Flagelos

Cilios y flagelos
Los cilios y los flagelos eucarióticos tienen una estructura muy similar:
Diámetro: 0,25 μm.
Axonema: constituído por microtúbulos y sus proteínas asociadas.
Muchas bacterias también tienen flagelos de estructura diferente carentes de microtúbulos.
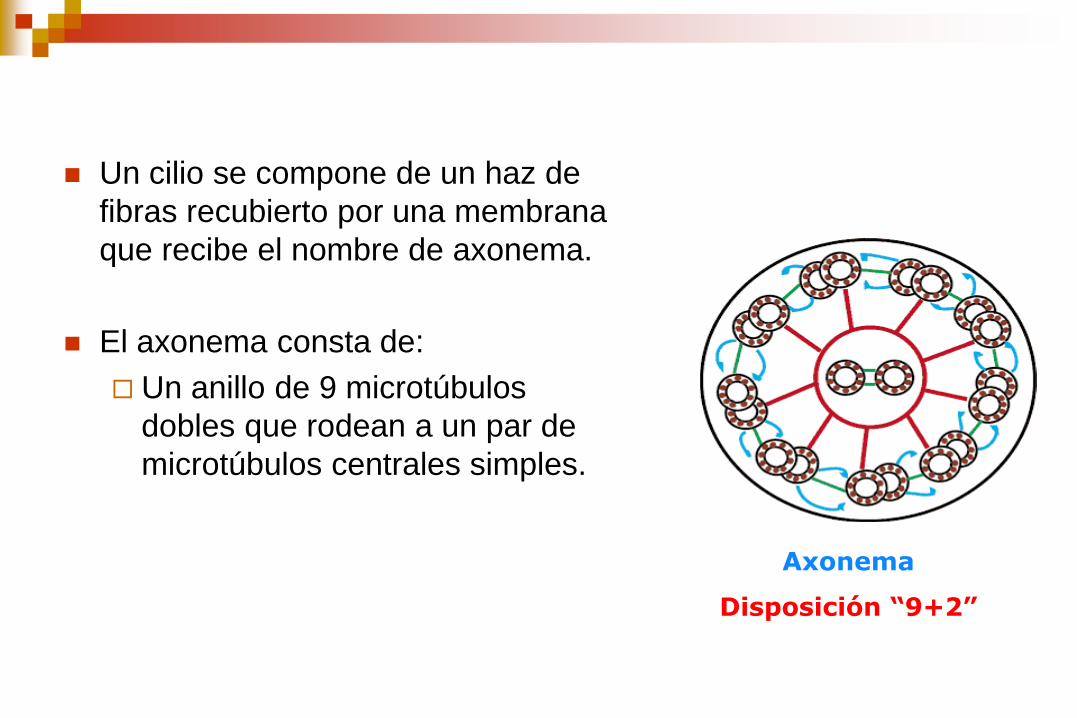
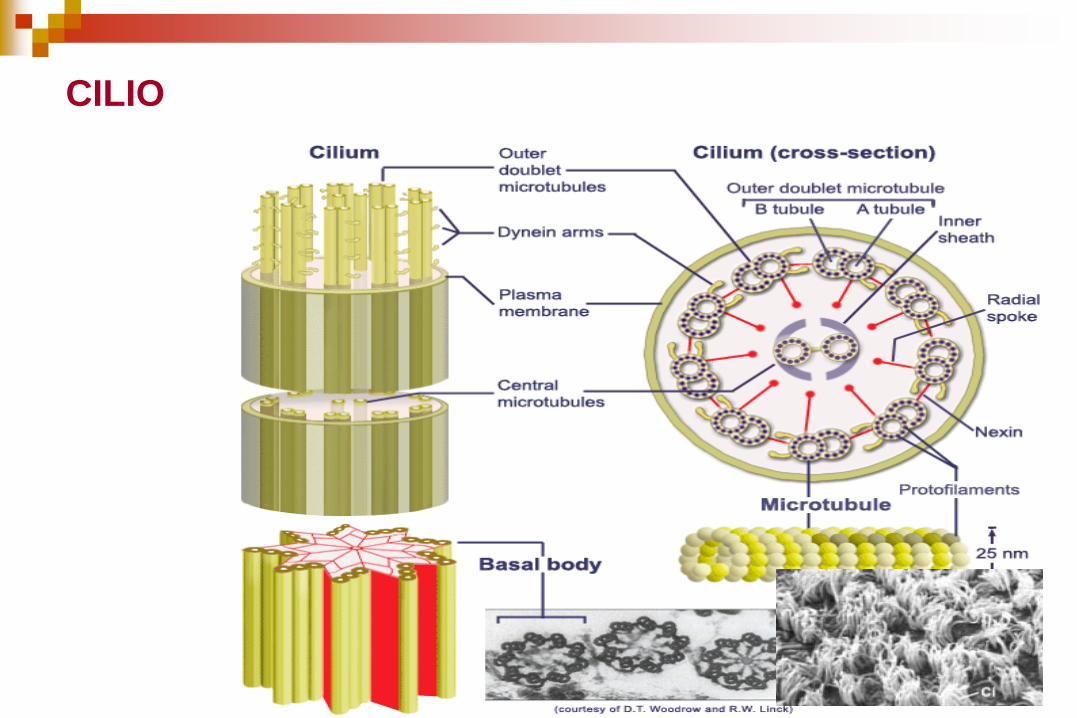
Un cilio se compone de un haz de
fibras recubierto por una membrana
que recibe el nombre de axonema.
El axonema consta de:
Un anillo de 9 microtúbulos
dobles que rodean a un par de
microtúbulos centrales simples.
Axonema
Disposición “9+2”
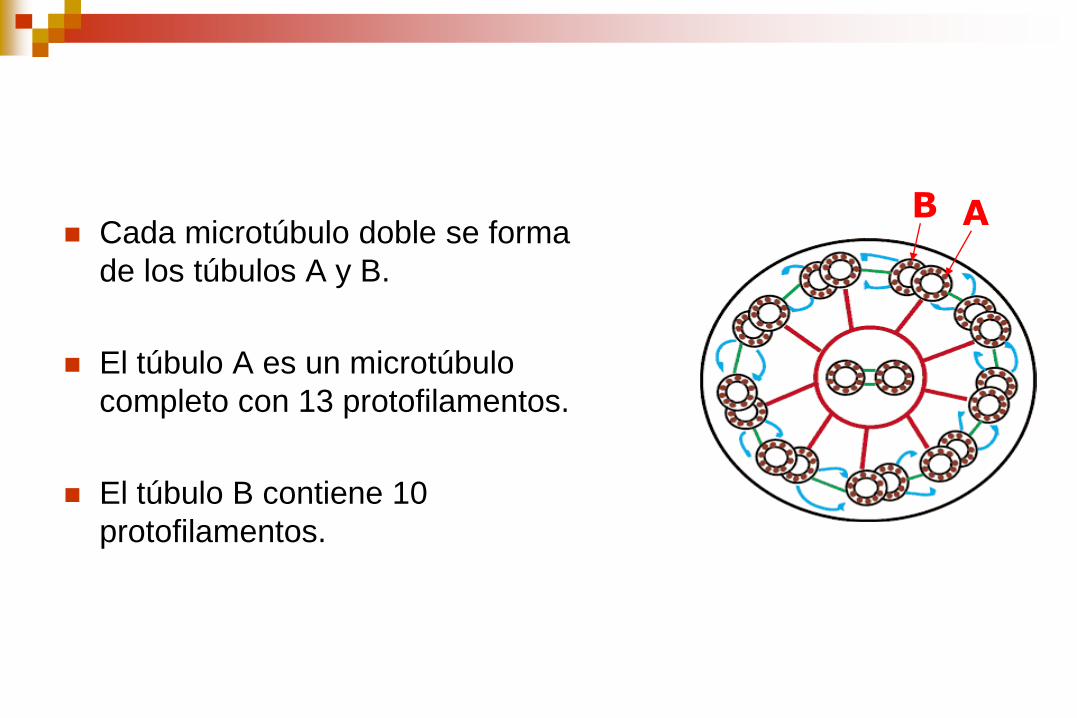
Cada microtúbulo doble se forma
de los túbulos A y B.
El túbulo A es un microtúbulo
completo con 13 protofilamentos.
El túbulo B contiene 10
protofilamentos.
A B
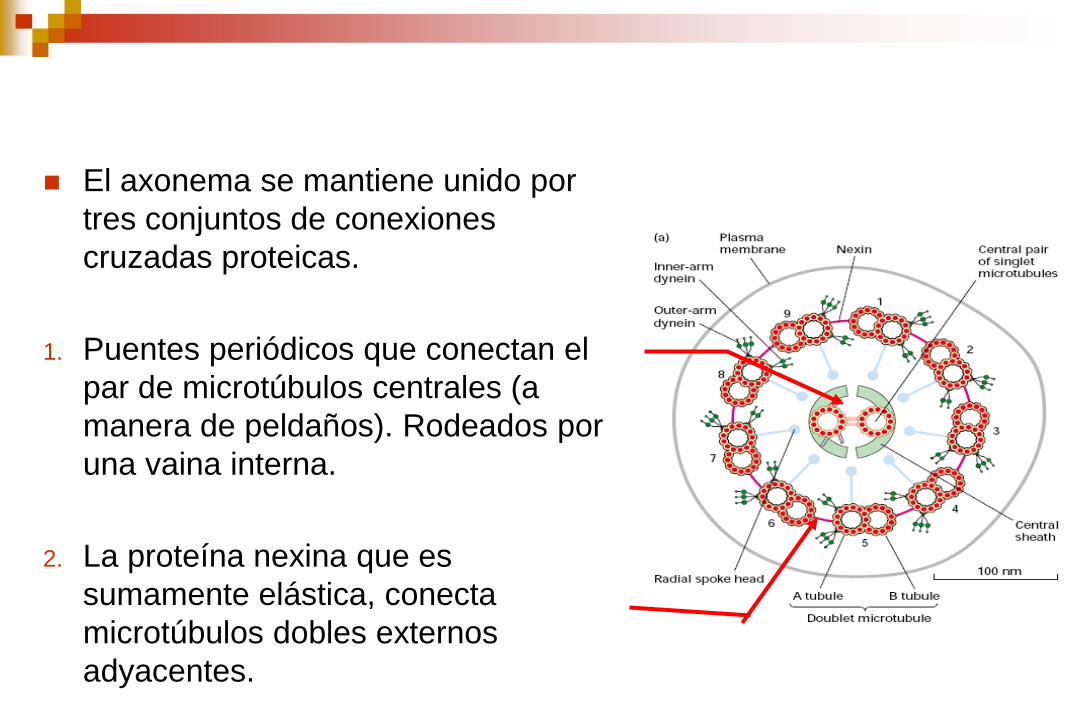
El axonema se mantiene unido por
tres conjuntos de conexiones
cruzadas proteicas.
1. Puentes periódicos que conectan el
par de microtúbulos centrales (a
manera de peldaños). Rodeados por
una vaina interna.
2. La proteína nexina que es
sumamente elástica, conecta
microtúbulos dobles externos
adyacentes.
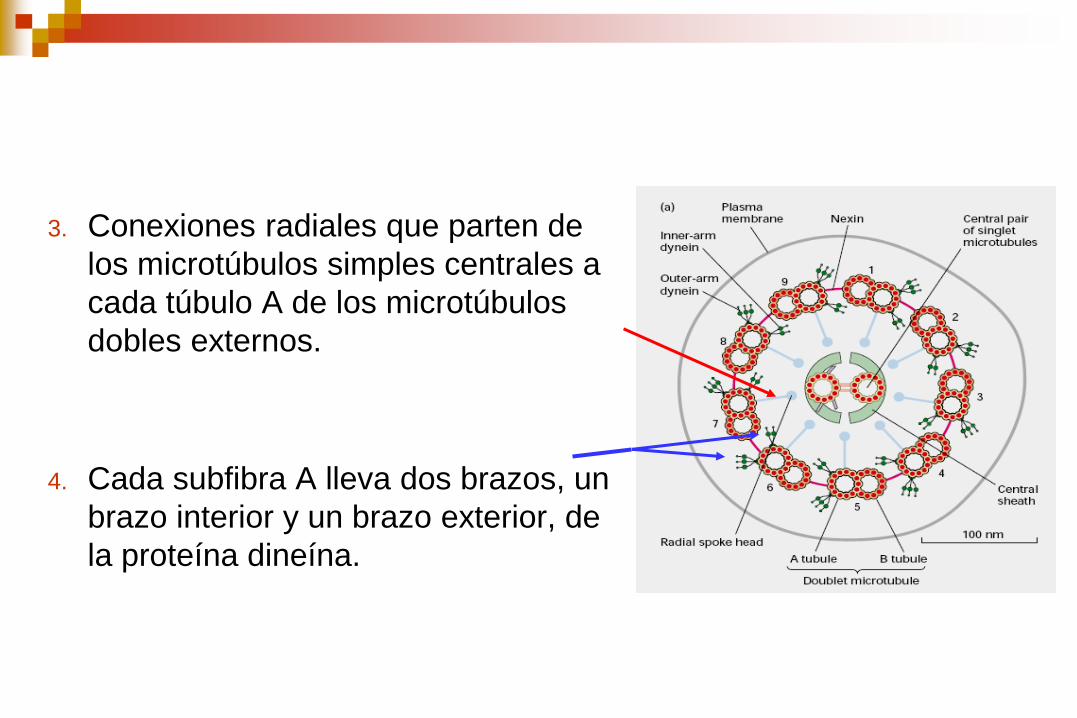
3. Conexiones radiales que parten de
los microtúbulos simples centrales a
cada túbulo A de los microtúbulos
dobles externos.
4. Cada subfibra A lleva dos brazos, un
brazo interior y un brazo exterior, de
la proteína dineína.
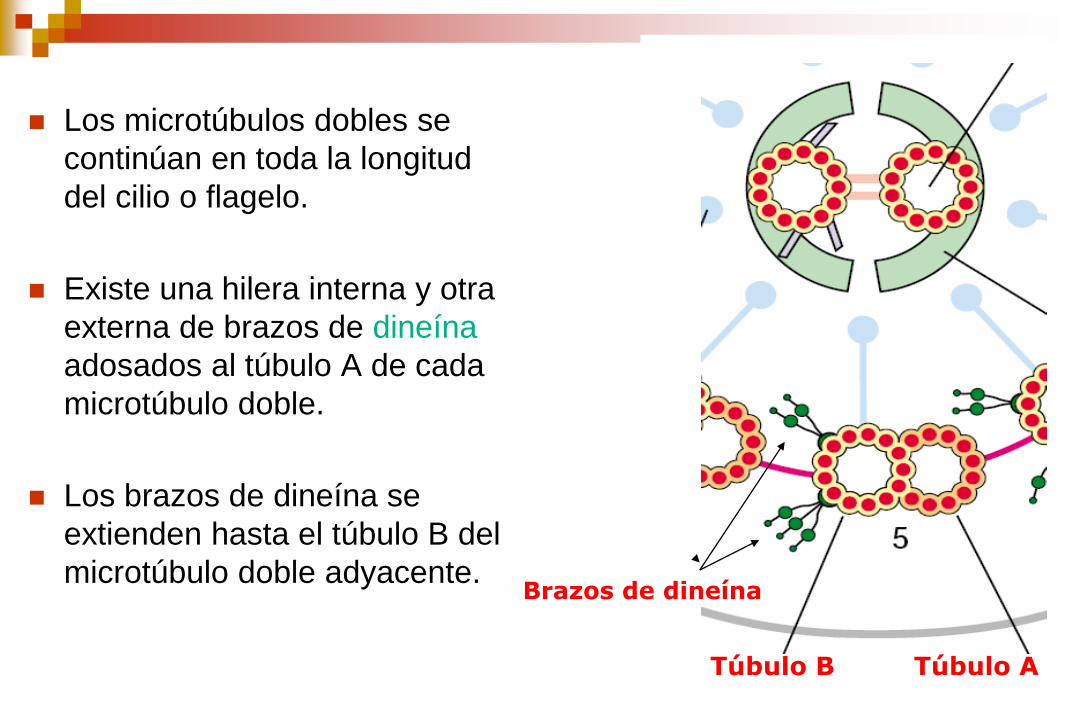
Los microtúbulos dobles se
continúan en toda la longitud
del cilio o flagelo.
Existe una hilera interna y otra
externa de brazos de dineína
adosados al túbulo A de cada
microtúbulo doble.
Los brazos de dineína se
extienden hasta el túbulo B del
microtúbulo doble adyacente.
Túbulo B Túbulo A
Brazos de dineína
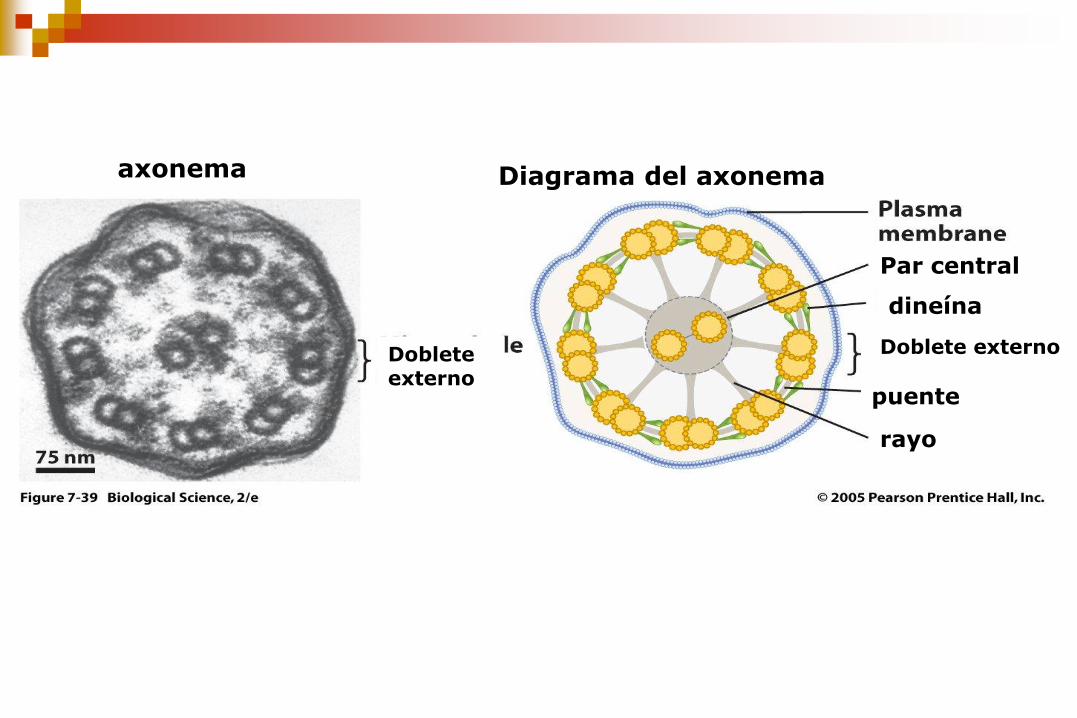
rayo
puente
dineína
Par central
Doblete externo
Diagrama del axonema axonema
Doblete externo
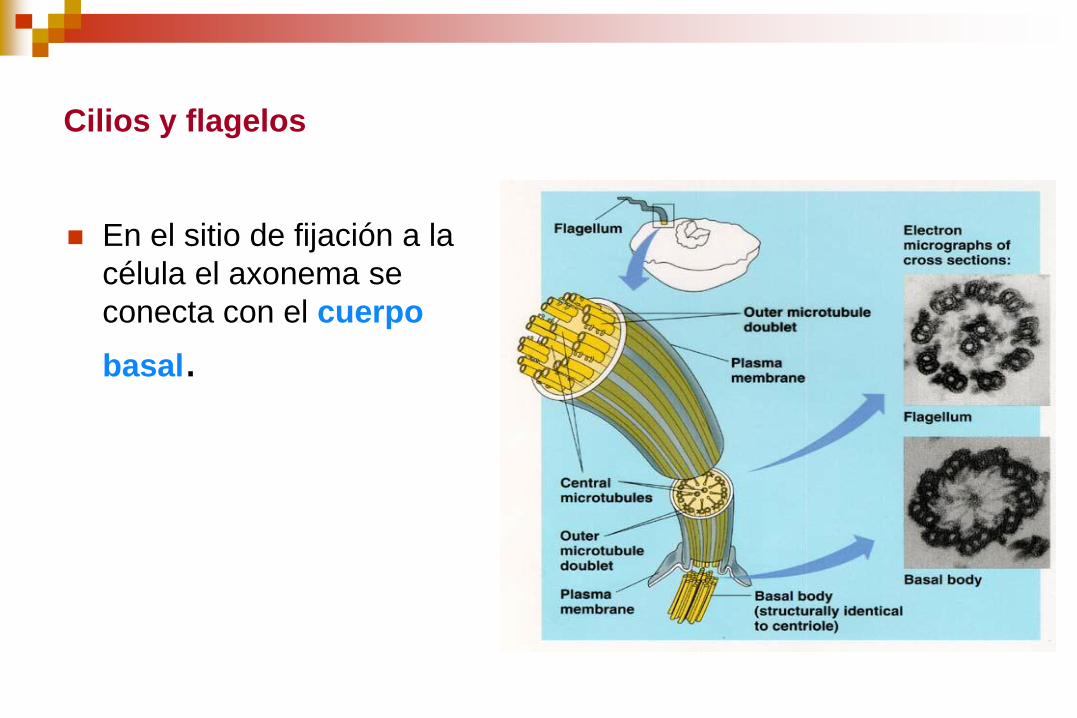
Cilios y flagelos
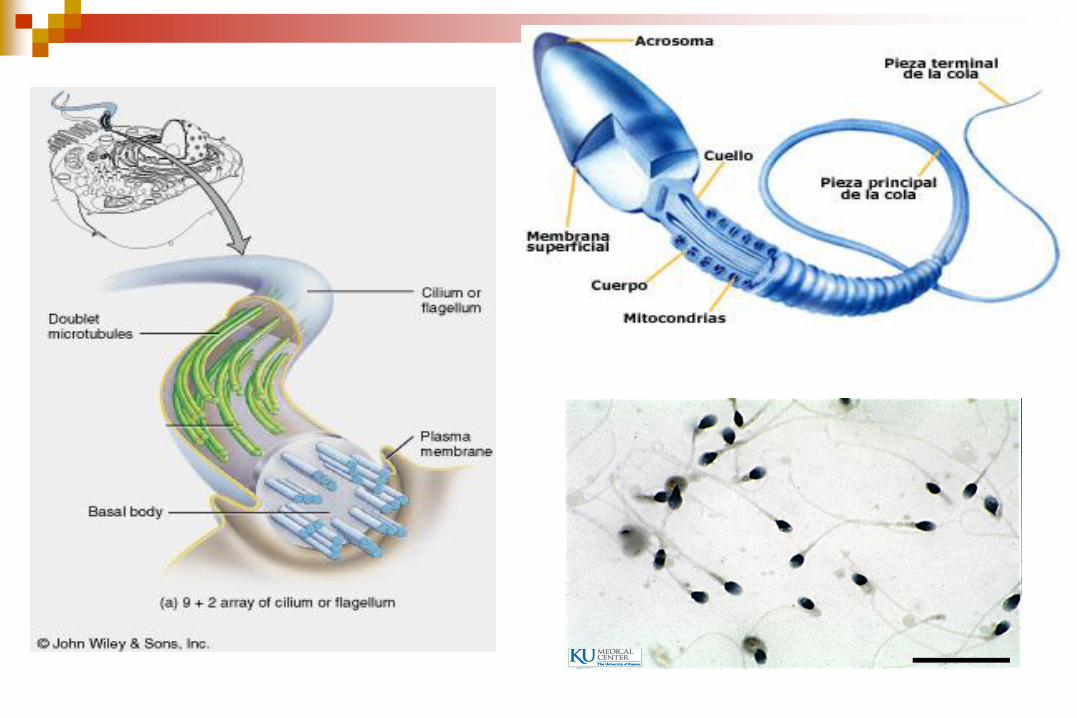
En el sitio de fijación a la
célula el axonema se
conecta con el cuerpo
basal.
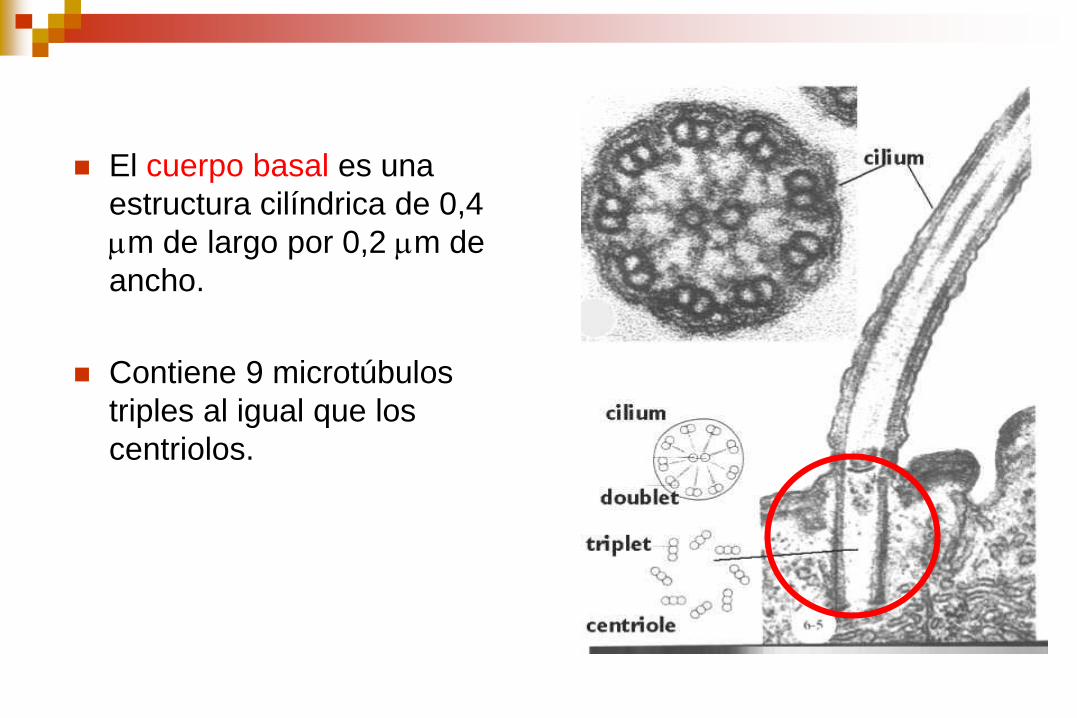
El cuerpo basal es una
estructura cilíndrica de 0,4
m de largo por 0,2 m de
ancho.
Contiene 9 microtúbulos
triples al igual que los
centriolos.
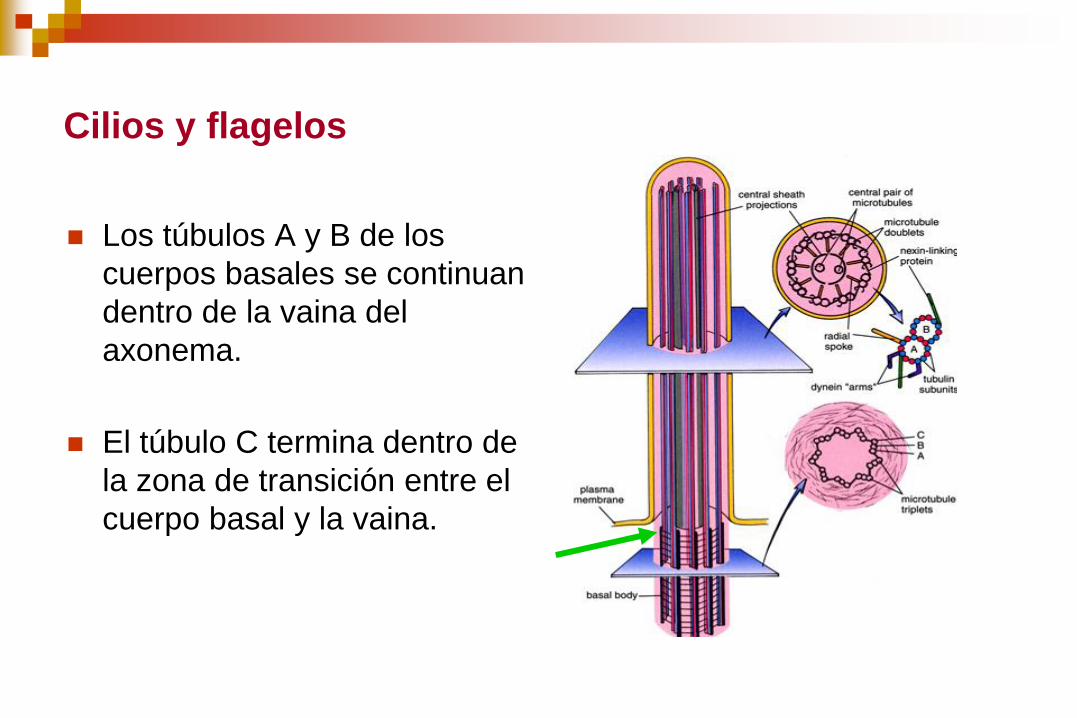
Cilios y flagelos
Los túbulos A y B de los
cuerpos basales se continuan
dentro de la vaina del
axonema.
El túbulo C termina dentro de
la zona de transición entre el
cuerpo basal y la vaina.
CILIO

Cilios
Son orgánulos de apariencia capilar en las superficies de
muchas células animales y vegetales.
Funciones:
Mueven fluido sobre la superficie de la célula.
Impulsan a «remo» células simples a través de un fluido.
En los seres humanos, por ejemplo, las células epiteliales
que recubren el tracto respiratorio tienen cada una unos 200
cilios que pulsan en sincronía para impulsar la mucosidad
hacia la garganta para su eliminación.
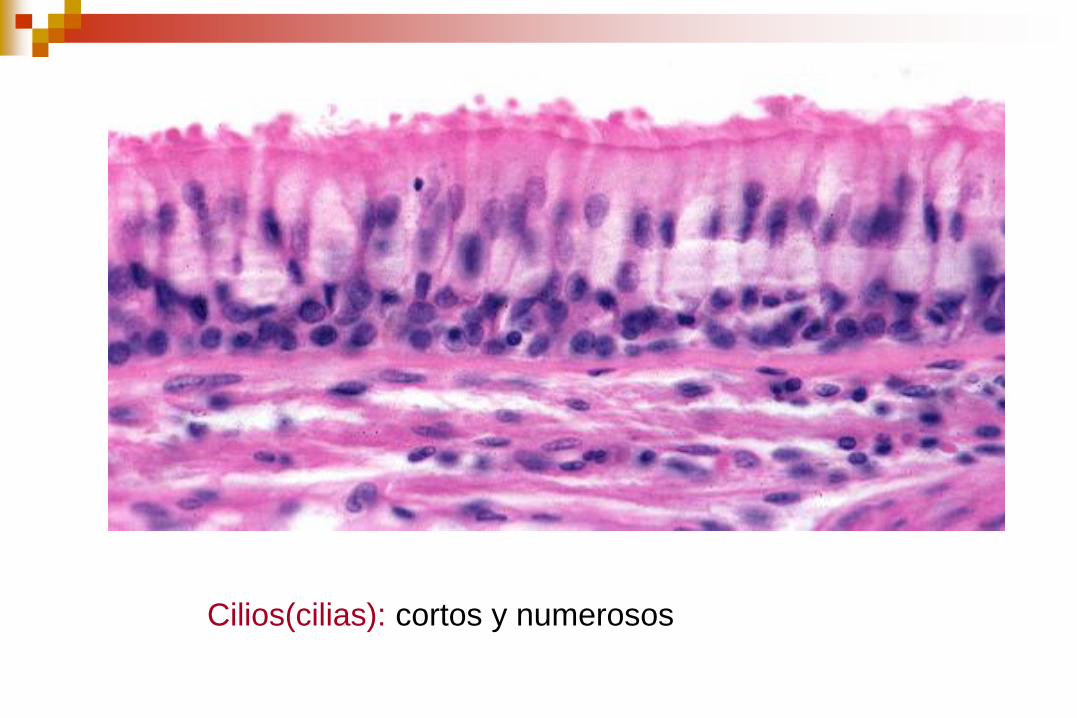
Cilios(cilias): cortos y numerosos
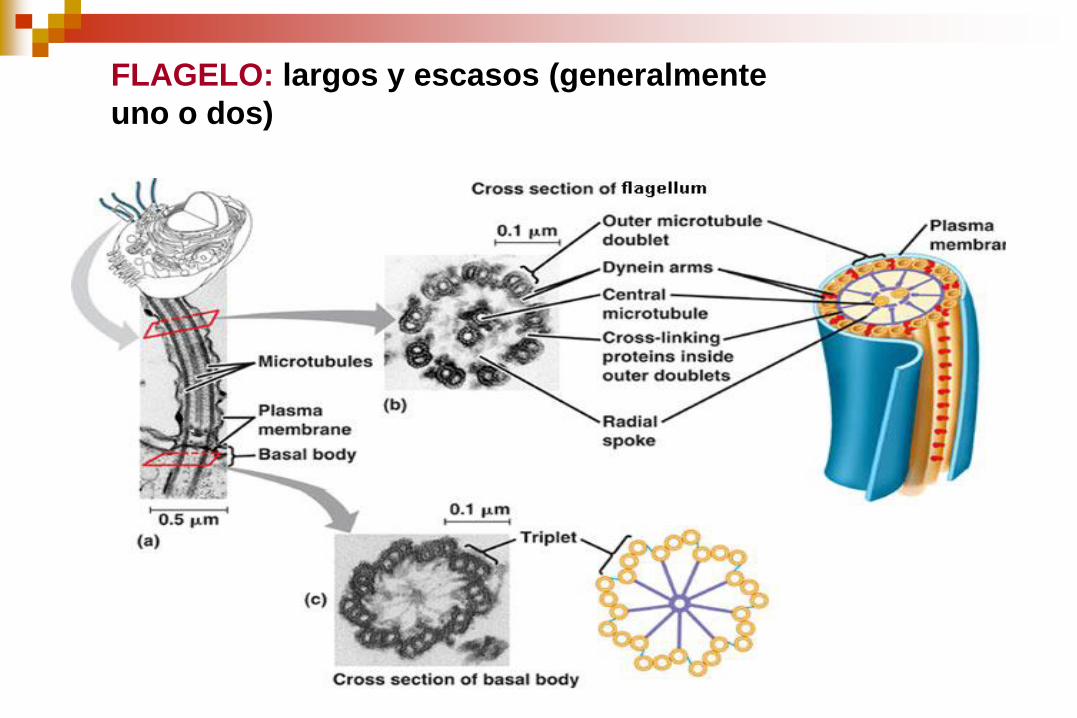
FLAGELO: largos y escasos (generalmente
uno o dos)

Organelas fibrilares - Amanzo
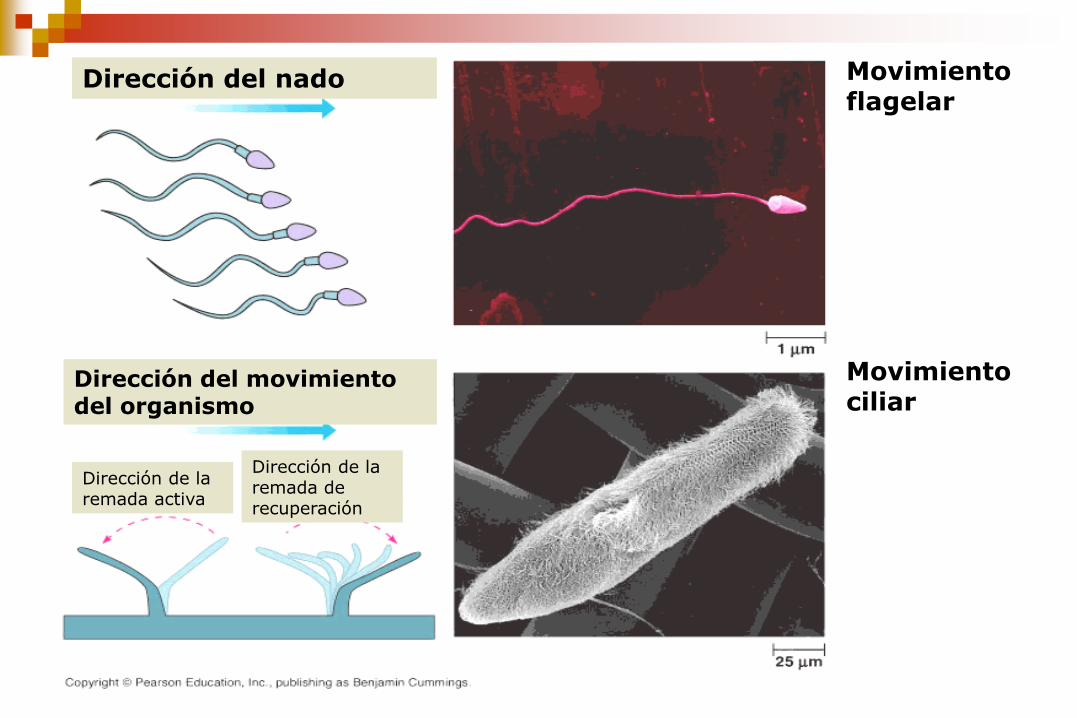
Movimiento flagelar
Movimiento ciliar
Dirección del nado
Dirección del movimiento del organismo
Dirección de la remada activa
Dirección de la remada de recuperación

FIN….





























