Embed Size (px)
Citation preview

Journal of Controlled Release 142 (2010) 70–77
Contents lists available at ScienceDirect
Journal of Controlled Release
j ourna l homepage: www.e lsev ie r.com/ locate / j conre l
GENEDELIVERY
A dual-functioning adenoviral vector encoding both transforming growthfactor-β3 and shRNA silencing type I collagen: Construction andcontrolled release for chondrogenesis
Feng Zhang 1, Yongchang Yao 1, Jinghua Hao, Ruijie Zhou, Chengzheng Liu, Yihong Gong, Dong-An Wang ⁎Division of Bioengineering, School of Chemical and Biomedical Engineering, Nanyang Technological University, Singapore 637457, Republic of Singapore
Abbreviations: TGF-β3, transforming growth factorAd-sh, recombinant adenovirus that specifically exprcollagen; Ad-T, recombinant adenovirus that specifigrowth factor-β3; Ad-dual, recombinant adenovirucollagen-targeting shRNA and TGF-β3; MOI, multiplichairpin RNA; MV, Malignant rabbit fibroma virus.⁎ Corresponding author. Division of BioEngineering, Sc
Engineering, Nanyang Technological University, 70Singapore 637457, Republic of Singapore. Tel.: +65 631
E-mail address: [email protected] (D.-A. Wang).1 The two authors have equal contributions to this re
0168-3659/$ – see front matter © 2009 Elsevier B.V. Adoi:10.1016/j.jconrel.2009.09.027
a b s t r a c t
a r t i c l e i n f oArticle history:Received 25 July 2009Accepted 29 September 2009Available online 12 October 2009
Keywords:AdenovirusDual-functioningCartilage regenerationTransforming growth factorRNA interference
Hyaline articular cartilage degeneration is a common clinical syndrome globally, whereas cell-based therapyhas proved to be a good solution to such problems. Given that transforming growth factor (TGF-β3) is helpfulin maintaining chondrocytic phenotype or stimulating chondrogenic differentiation of stem cells, vectorscontaining TGF-β3 expression cassette can be delivered to therapeutic cells. One problem involved in theapplication of therapeutic cells in chondrogenesis is the undesirable production of type I collagen in suchcells as chondrocytes and synovial mesenchymal stem cells during ex vivo culture, which undermines themechanical strength of engineered cartilage. RNA interference (RNAi) strategy can be used to knock down itsexpression to allow better biological and mechanical functions in artificial tissues. In this study, for the firsttime we report the construction of an adenoviral vector that can express both TGF-β3 to promotechondrogenesis and short hairpin RNA (shRNA) targeting type I collagen to block its production. This dual-functioning vector (Ad-dual) was found to function well in three model cell types: human fibroblast,osteoblast and porcine chondrocyte in terms of the release of TGF-β3 protein and down-regulation of type Icollagen production. Besides, we also tested its efficacy in porcine synovial mesenchymal stem cells,highlighting its potential applications in cell-based therapy for the treatment of articular cartilagedegeneration.
-β3; Ad-null, null adenovirus;esses shRNA targeting type Ically expresses transformings that encodes both type Iity of infection; shRNA, short
hool of Chemical & BiomedicalNanyang Drive, N1.3-B2-13,6 8890; fax: +65 6791 1761.
search article.
ll rights reserved.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Hyaline articular cartilage trauma and degeneration cause painand affect life quality of human beings worldwide [1]. Although manyremedial strategies have been developed for its treatment, it stillpresents a challenge in clinical trials. Among the various techniques,regenerative medicine provides a powerful strategy for cartilageregeneration and repair. Regenerative medicine is a discipline thataims to restore the structure and functions of adult body parts bytreatment with regenerative drugs, injection of stem/progenitor cells
or transplantation of artificial organs or tissues that have beendeveloped in vitro [2,3]. For articular cartilage regeneration, chon-drocytes or stem/progenitor cells are obtained from patients ordonors and amplified for injection or transplantation. This treatment,particularly if coupled with gene therapy, has borne great promises inrestoring the functions of articular cartilage for the great number ofpatients [4,5]. Gene manipulations applied to cells will improve cellfunctions and overcome the problems that may occur during thetreatment.
Growth factors have long been delivered to cells to promote cellgrowth, proliferation or differentiation, working as an importantregulator of cells. Among the various growth factors delivered forarticular regeneration, TGF-β3 was substantiated to help in main-taining a round or polygonal shape of chondrocytes and stimulatetotal collagen synthesis, which is crucial in maintaining the cartilagefunctions [6,7]. Recombinant TGF-β3 protein has also been deliveredto stem cells such as mesenchymal stem cells for in vitro chondrogen-esis [8–12]. However, such methods suffer from the lack of sustainedrelease of growth factors. Gene therapy has enabled researchers toachieve a sustained and effective supply of the protein by incorpo-rating the protein-encoding gene into a vector, which is thentransfected or transduced into cells for expression. For instance,

71F. Zhang et al. / Journal of Controlled Release 142 (2010) 70–77
GENEDELIVERY
delivery of TGF-β3 to rat bonemarrowmesenchymal stem cells via anadenoviral vector has been demonstrated to elicit BMSC differentia-tion into chondrocytes [13].
In articular cartilage, chondrocytes are located in cavities calledcartilage lacunae and surrounded by cartilage matrix, which is mainlycomposed of type II collagen and proteoglycans [14]. The introductionof other components such as type I collagen leads to fibrocartilageformation and alters the mechanical strength of cartilage, thusundermines its performance. However, as chondrocytes are passagedand amplified ex vivo to obtain a sufficient population for transplan-tation, type I collagen will be inevitably expressed as a result [15].Even the freshly-isolated synovial mesenchymal stem cells,which are currently acknowledged to have excellent chondrogeniccapabilities, express type I collagen [16]. Therefore, it is critical toblock its expression in the therapeutic cells in order to maintainthe mechanical strength of articular cartilage. RNA interference(RNAi) [17,18], which is a mechanism of small interfering RNA(siRNA) mediated gene silencing, has provided effective tools tomeet this end. Such strategy has been applied to treat some diseases[19].
To achieve the above two objectives simultaneously, we selectedadenovirus [20,21] as a model recombinant dual-functioning adeno-viral vector (Ad-dual) for investigation. This Ad-dual can both pro-mote TGF-β3 synthesis and suppress the expression of type I collagenby the release of corresponding short hairpin RNA (shRNA). Thesefunctions were assessed in the following cell types: fibroblast,osteoblast, chondrocyte and synovial mesenchymal stem cells.Quantitative RT-PCR (qRT-PCR) and flowcytometry have revealedthe effect of various multiplicity of infections (MOIs) on cell viability,infection efficiency, type I collagen inhibition and TGF-β3 expression,and appropriate MOIs were selected for subsequent investigation as atool to control the release of TGF-β3 and shRNA. qRT-PCR andimmunofluorescent staining demonstrated the successful down-regulation of type I collagen, while qRT-PCR and ELISA showed therelease of TGF-β3 both at the mRNA and protein levels.
2. Materials and methods
2.1. Construction and production of null and all other recombinantadenoviral vector
Schematic diagrams of various recombinant adenoviral vectorswere given in Fig. 1. These adenoviral vectors include null adenovirus(Ad-null), recombinant adenovirus that specifically expresses shRNAtargeting type I collagen (Ad-sh), recombinant adenovirus that
Fig. 1. Schematic diagrams of the produced adenoviral vectors. These vectors includenull adenovirus (Ad-null), recombinant adenovirus that specifically expresses shRNAtargeting type I collagen (Ad-sh), recombinant adenovirus that specifically expressestransforming growth factor-β3 (Ad-T) and recombinant adenovirus that encodes bothtype I collagen-targeting shRNA and TGF-β3 (Ad-dual).
specifically expresses transforming growth factor-β3 (Ad-T) andrecombinant adenovirus that encodes both type I collagen-targetingshRNA and TGF-β3 (Ad-dual). Ad-sh was produced using Adeno-XViraTrak ZsGreen1 Promoterless Expression System 2 (ClontechLaboratories, Mountain View, CA, USA) by incorporating annealedshRNA-encoding double-strand sequence with BamHI and EcoRI over-hangs at the two ends (Sense: 5′-GATCCGCAATCACCTGCGTACAGAATT-CAAGAGATTCTGTACG-CAGGTGATTGTTTTTTACGCGTG-3′; Antisense:5′AATTCACGCGTAAAAAACA-ATCACCTGCG-TACAGAATCTCTT-GAATTCTGTACGCAGGTGATTGCG-3′). All the other three recombinantadenoviruses were produced using Adeno-X ViraTrak ZsGreen1Expression System 2 (Clontech Laboratories, Mountain View, CA,USA). Ad-T was constructed by our labmates [14]. Ad-dual was con-structed by sequentially incorporating SalI/PstI-digested TGF-β3 codingsequence amplified from hTGF-β3-pCMV6-XL5 plasmid DNA (OrigeneTechnologies, Rockville, MD, USA) (Primers forward: ACGCGTCGACAT-GAAGATGCACTT; reverse: TGCACTGCAGTCAGCTACATTTAC), ApaI/XhoI-digested human U6 promoter amplified from human fibroblastgenome (Primers forward:ATTTGCGGGCCCGCAGGAAGAGGGCCTAT;reverse: CC-GCTCGAGTCGTCCTTTCCACAAG), and annealed shRNA-en-coding double-strand sequencewithXhoI and PstI overhangs at the twoends (Sense: TCGAGCAATCACCT-GCGTACAGAATTCAAGAGATTCTG-TACGCAGGTGATTGTTTTTTACGCGTCTGCA; Antisense: GACGCGTAAAA-AACAATCACCTGCGTACAGAATCTCTTGAATTCTGTACGCAGGTGATTGC).Recombinant pDNR plasmid and recombinant adenoviral vector wereenzymatically digested to prove successful recombination. All therecombinant plasmids were amplified in 5α, competent Escherichia colicells (New England Biolabs, Beverly, MA, USA) and purifiedwith QIAfilterplasmid extraction kit (Qiagen, Hilden, Germany) The above recombinantadenoviral plasmid was linearized by PacI digestion and transfected intoHEK 293 cells (ATCC, Manassas, VA) using Lipofectamine reagents(Invitrogen, Carlsbad, CA, USA) for viral packaging. The packaged adeno-virus was then purified using Adeno-XTM Virus Purification Kit (ClontechLaboratories, Mountain View, CA, USA) and titrated using Adeno-XTM
Rapid Titer Kit (Clontech Laboratories, Mountain View, CA, USA) for sub-sequent experiments.
2.2. Cell transduction
Human fibroblast and human foetal osteoblast were purchasedfrom American Type Culture Collections (ATCC, Manassas, VA, USA).Porcine chondrocyte and synovial mesenchymal stem cells wereisolated according to previously published papers [22,23]. The cellswere cultured and seeded in 24-well or 6-well plates at a density of5×104 cells/cm2. The next day recombinant adenoviruses werediluted in DMEM or DMEM/F12 medium (for osteoblast culture,Invitrogen, Carlsbad, CA, USA) without FBS and added to the cellsat certain MOIs. Two hours later, the medium was removed andreplaced with fresh medium. Cells without transduction were usedas negative controls (Neg). Additionally, to compare the transduc-tion of Ad-dual and co-transduction of Ad-sh and Ad-T, synovialmesenchymal stem cells were either transduced with Ad-dual atMOI 200, or co-transduced with Ad-sh and Ad-T (Ad-sh/Ad-T co-transduction) at MOI 200 for each recombinant adenovirus, toensure that cells in these two groups are exposed to the same dosageof both genes.
2.3. Flowcytometry analysis
72 h after infection, cells were trypsinized, washed with PBS andstained with propidium iodide (50 μg/mL, BD, Heidelberg, Germany).The cells were then subjected to FACSCalibur flow cytometer andCellQuest software (BD, Heidelberg, Germany) to determine thepercentages of dead cells (stained in red), infected cells (with greenfluorescence emission) and viable infected cells. 5000 cells werecounted in each measurement.
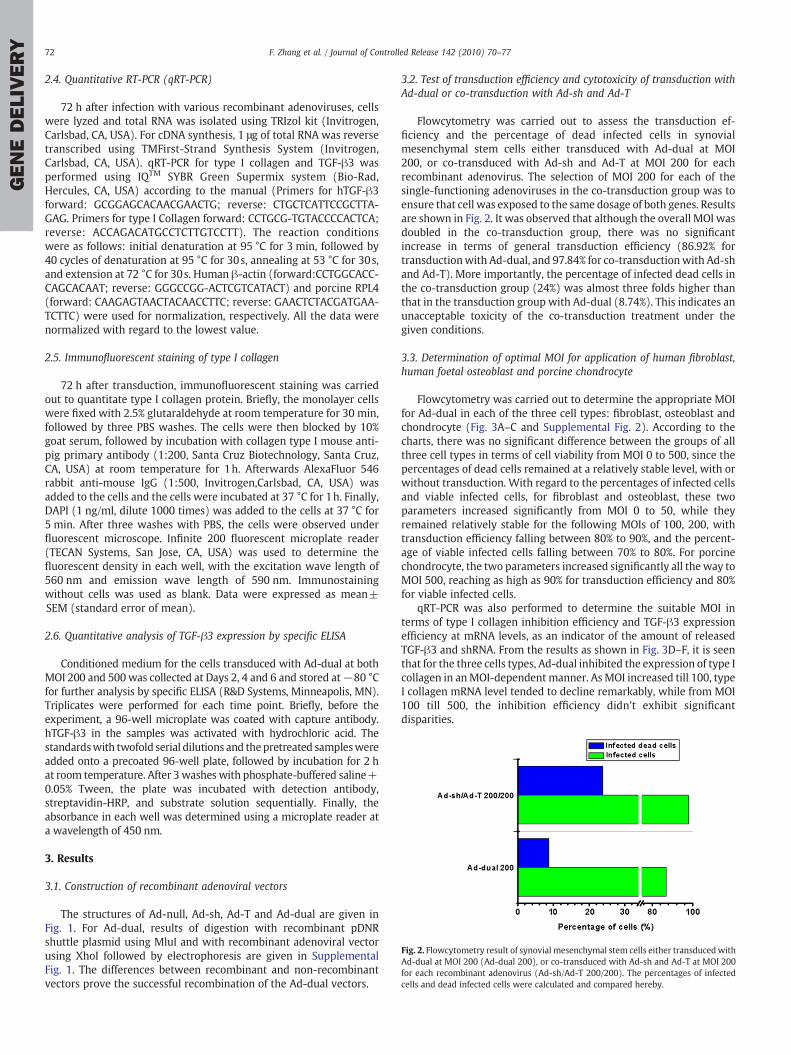
Fig. 2. Flowcytometry result of synovial mesenchymal stem cells either transduced withAd-dual at MOI 200 (Ad-dual 200), or co-transduced with Ad-sh and Ad-T at MOI 200for each recombinant adenovirus (Ad-sh/Ad-T 200/200). The percentages of infectedcells and dead infected cells were calculated and compared hereby.
72 F. Zhang et al. / Journal of Controlled Release 142 (2010) 70–77
GENEDELIVERY
2.4. Quantitative RT-PCR (qRT-PCR)
72 h after infection with various recombinant adenoviruses, cellswere lyzed and total RNA was isolated using TRIzol kit (Invitrogen,Carlsbad, CA, USA). For cDNA synthesis, 1 μg of total RNA was reversetranscribed using TMFirst-Strand Synthesis System (Invitrogen,Carlsbad, CA, USA). qRT-PCR for type I collagen and TGF-β3 wasperformed using IQTM SYBR Green Supermix system (Bio-Rad,Hercules, CA, USA) according to the manual (Primers for hTGF-β3forward: GCGGAGCACAACGAACTG; reverse: CTGCTCATTCCGCTTA-GAG. Primers for type I Collagen forward: CCTGCG-TGTACCCCACTCA;reverse: ACCAGACATGCCTCTTGTCCTT). The reaction conditionswere as follows: initial denaturation at 95 °C for 3 min, followed by40 cycles of denaturation at 95 °C for 30s, annealing at 53 °C for 30s,and extension at 72 °C for 30s. Human β-actin (forward:CCTGGCACC-CAGCACAAT; reverse: GGGCCGG-ACTCGTCATACT) and porcine RPL4(forward: CAAGAGTAACTACAACCTTC; reverse: GAACTCTACGATGAA-TCTTC) were used for normalization, respectively. All the data werenormalized with regard to the lowest value.
2.5. Immunofluorescent staining of type I collagen
72 h after transduction, immunofluorescent staining was carriedout to quantitate type I collagen protein. Briefly, the monolayer cellswere fixed with 2.5% glutaraldehyde at room temperature for 30 min,followed by three PBS washes. The cells were then blocked by 10%goat serum, followed by incubation with collagen type I mouse anti-pig primary antibody (1:200, Santa Cruz Biotechnology, Santa Cruz,CA, USA) at room temperature for 1h. Afterwards AlexaFluor 546rabbit anti-mouse IgG (1:500, Invitrogen,Carlsbad, CA, USA) wasadded to the cells and the cells were incubated at 37 °C for 1h. Finally,DAPI (1 ng/ml, dilute 1000 times) was added to the cells at 37 °C for5 min. After three washes with PBS, the cells were observed underfluorescent microscope. Infinite 200 fluorescent microplate reader(TECAN Systems, San Jose, CA, USA) was used to determine thefluorescent density in each well, with the excitation wave length of560 nm and emission wave length of 590 nm. Immunostainingwithout cells was used as blank. Data were expressed as mean±SEM (standard error of mean).
2.6. Quantitative analysis of TGF-β3 expression by specific ELISA
Conditioned medium for the cells transduced with Ad-dual at bothMOI 200 and 500 was collected at Days 2, 4 and 6 and stored at−80 °Cfor further analysis by specific ELISA (R&D Systems, Minneapolis, MN).Triplicates were performed for each time point. Briefly, before theexperiment, a 96-well microplate was coated with capture antibody.hTGF-β3 in the samples was activated with hydrochloric acid. Thestandardswith twofold serial dilutions and thepretreated sampleswereadded onto a precoated 96-well plate, followed by incubation for 2 hat room temperature. After 3 washes with phosphate-buffered saline+0.05% Tween, the plate was incubated with detection antibody,streptavidin-HRP, and substrate solution sequentially. Finally, theabsorbance in each well was determined using a microplate reader ata wavelength of 450 nm.
3. Results
3.1. Construction of recombinant adenoviral vectors
The structures of Ad-null, Ad-sh, Ad-T and Ad-dual are given inFig. 1. For Ad-dual, results of digestion with recombinant pDNRshuttle plasmid using MluI and with recombinant adenoviral vectorusing XhoI followed by electrophoresis are given in SupplementalFig. 1. The differences between recombinant and non-recombinantvectors prove the successful recombination of the Ad-dual vectors.
3.2. Test of transduction efficiency and cytotoxicity of transduction withAd-dual or co-transduction with Ad-sh and Ad-T
Flowcytometry was carried out to assess the transduction ef-ficiency and the percentage of dead infected cells in synovialmesenchymal stem cells either transduced with Ad-dual at MOI200, or co-transduced with Ad-sh and Ad-T at MOI 200 for eachrecombinant adenovirus. The selection of MOI 200 for each of thesingle-functioning adenoviruses in the co-transduction group was toensure that cell was exposed to the same dosage of both genes. Resultsare shown in Fig. 2. It was observed that although the overall MOI wasdoubled in the co-transduction group, there was no significantincrease in terms of general transduction efficiency (86.92% fortransductionwith Ad-dual, and 97.84% for co-transductionwith Ad-shand Ad-T). More importantly, the percentage of infected dead cells inthe co-transduction group (24%) was almost three folds higher thanthat in the transduction group with Ad-dual (8.74%). This indicates anunacceptable toxicity of the co-transduction treatment under thegiven conditions.
3.3. Determination of optimal MOI for application of human fibroblast,human foetal osteoblast and porcine chondrocyte
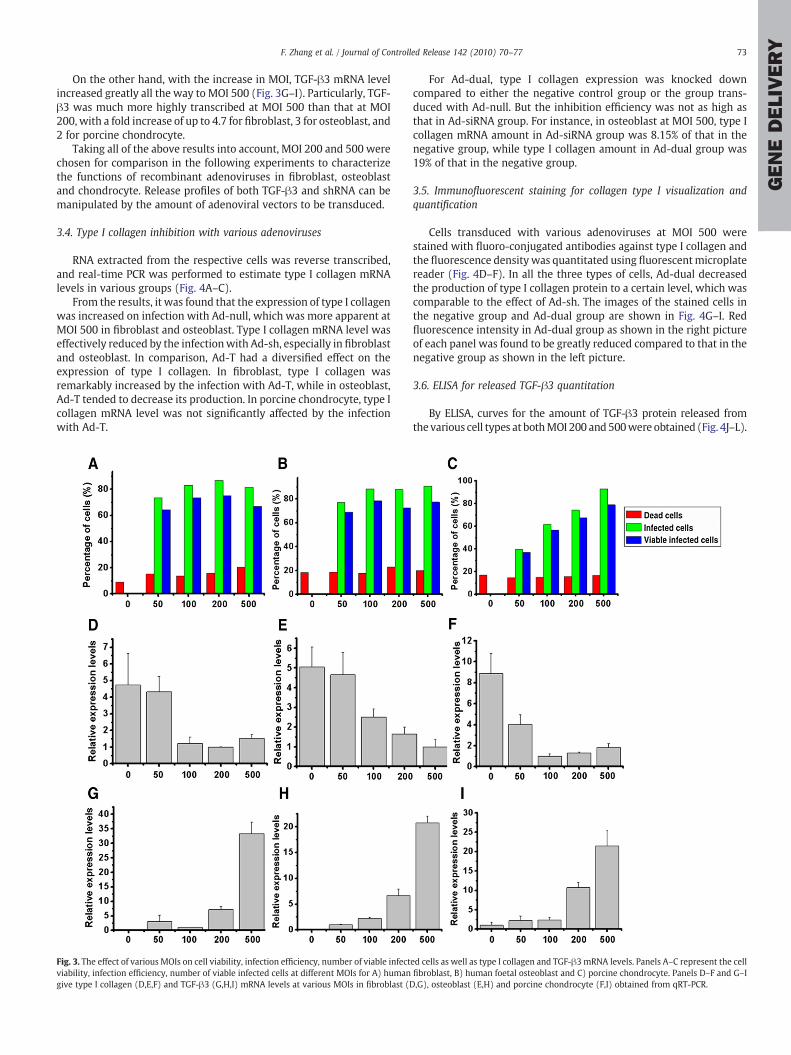
Flowcytometry was carried out to determine the appropriate MOIfor Ad-dual in each of the three cell types: fibroblast, osteoblast andchondrocyte (Fig. 3A–C and Supplemental Fig. 2). According to thecharts, there was no significant difference between the groups of allthree cell types in terms of cell viability from MOI 0 to 500, since thepercentages of dead cells remained at a relatively stable level, with orwithout transduction. With regard to the percentages of infected cellsand viable infected cells, for fibroblast and osteoblast, these twoparameters increased significantly from MOI 0 to 50, while theyremained relatively stable for the following MOIs of 100, 200, withtransduction efficiency falling between 80% to 90%, and the percent-age of viable infected cells falling between 70% to 80%. For porcinechondrocyte, the two parameters increased significantly all the way toMOI 500, reaching as high as 90% for transduction efficiency and 80%for viable infected cells.
qRT-PCR was also performed to determine the suitable MOI interms of type I collagen inhibition efficiency and TGF-β3 expressionefficiency at mRNA levels, as an indicator of the amount of releasedTGF-β3 and shRNA. From the results as shown in Fig. 3D–F, it is seenthat for the three cells types, Ad-dual inhibited the expression of type Icollagen in anMOI-dependent manner. AsMOI increased till 100, typeI collagen mRNA level tended to decline remarkably, while from MOI100 till 500, the inhibition efficiency didn't exhibit significantdisparities.
73F. Zhang et al. / Journal of Controlled Release 142 (2010) 70–77
GENEDELIVERY
On the other hand, with the increase in MOI, TGF-β3 mRNA levelincreased greatly all the way to MOI 500 (Fig. 3G–I). Particularly, TGF-β3 was much more highly transcribed at MOI 500 than that at MOI200, with a fold increase of up to 4.7 for fibroblast, 3 for osteoblast, and2 for porcine chondrocyte.
Taking all of the above results into account, MOI 200 and 500 werechosen for comparison in the following experiments to characterizethe functions of recombinant adenoviruses in fibroblast, osteoblastand chondrocyte. Release profiles of both TGF-β3 and shRNA can bemanipulated by the amount of adenoviral vectors to be transduced.
3.4. Type I collagen inhibition with various adenoviruses
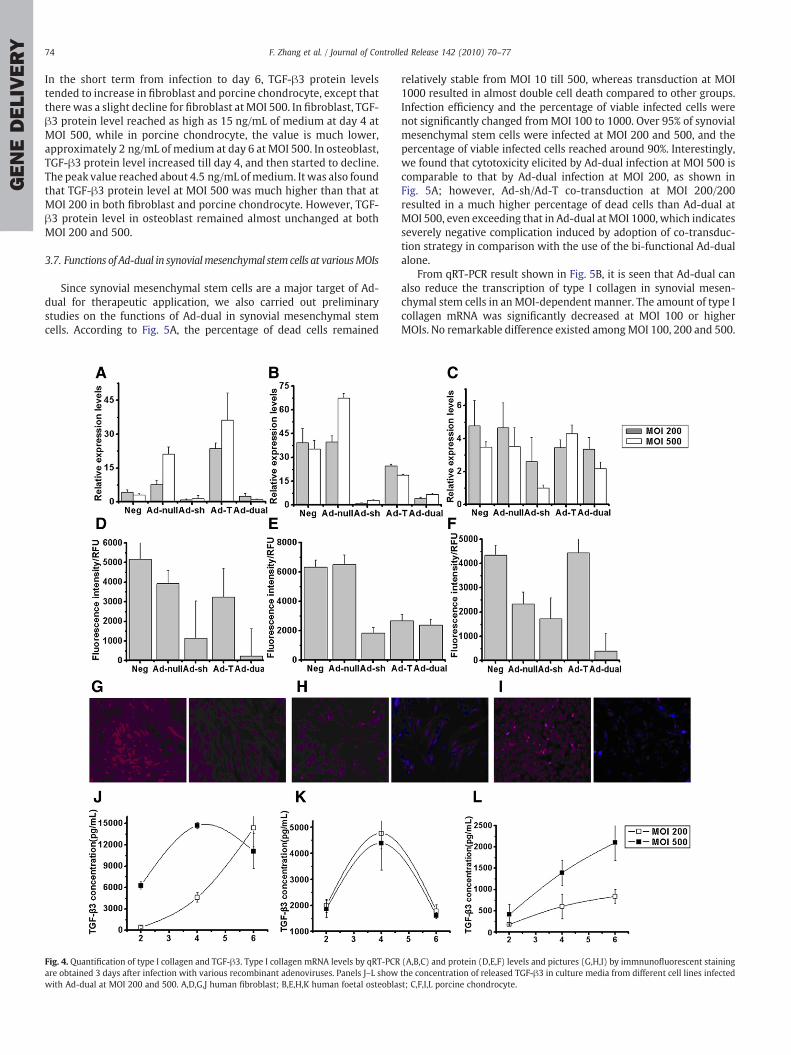
RNA extracted from the respective cells was reverse transcribed,and real-time PCR was performed to estimate type I collagen mRNAlevels in various groups (Fig. 4A–C).
From the results, it was found that the expression of type I collagenwas increased on infection with Ad-null, which was more apparent atMOI 500 in fibroblast and osteoblast. Type I collagen mRNA level waseffectively reduced by the infectionwith Ad-sh, especially in fibroblastand osteoblast. In comparison, Ad-T had a diversified effect on theexpression of type I collagen. In fibroblast, type I collagen wasremarkably increased by the infection with Ad-T, while in osteoblast,Ad-T tended to decrease its production. In porcine chondrocyte, type Icollagen mRNA level was not significantly affected by the infectionwith Ad-T.
Fig. 3. The effect of various MOIs on cell viability, infection efficiency, number of viable infectviability, infection efficiency, number of viable infected cells at different MOIs for A) humangive type I collagen (D,E,F) and TGF-β3 (G,H,I) mRNA levels at various MOIs in fibroblast (D
For Ad-dual, type I collagen expression was knocked downcompared to either the negative control group or the group trans-duced with Ad-null. But the inhibition efficiency was not as high asthat in Ad-siRNA group. For instance, in osteoblast at MOI 500, type Icollagen mRNA amount in Ad-siRNA group was 8.15% of that in thenegative group, while type I collagen amount in Ad-dual group was19% of that in the negative group.
3.5. Immunofluorescent staining for collagen type I visualization andquantification
Cells transduced with various adenoviruses at MOI 500 werestained with fluoro-conjugated antibodies against type I collagen andthe fluorescence density was quantitated using fluorescentmicroplatereader (Fig. 4D–F). In all the three types of cells, Ad-dual decreasedthe production of type I collagen protein to a certain level, which wascomparable to the effect of Ad-sh. The images of the stained cells inthe negative group and Ad-dual group are shown in Fig. 4G–I. Redfluorescence intensity in Ad-dual group as shown in the right pictureof each panel was found to be greatly reduced compared to that in thenegative group as shown in the left picture.
3.6. ELISA for released TGF-β3 quantitation
By ELISA, curves for the amount of TGF-β3 protein released fromthe various cell types at bothMOI 200 and500were obtained (Fig. 4J–L).
ed cells as well as type I collagen and TGF-β3mRNA levels. Panels A–C represent the cellfibroblast, B) human foetal osteoblast and C) porcine chondrocyte. Panels D–F and G–I,G), osteoblast (E,H) and porcine chondrocyte (F,I) obtained from qRT-PCR.
74 F. Zhang et al. / Journal of Controlled Release 142 (2010) 70–77
GENEDELIVERY
In the short term from infection to day 6, TGF-β3 protein levelstended to increase in fibroblast and porcine chondrocyte, except thatthere was a slight decline for fibroblast at MOI 500. In fibroblast, TGF-β3 protein level reached as high as 15 ng/mL of medium at day 4 atMOI 500, while in porcine chondrocyte, the value is much lower,approximately 2 ng/mL of medium at day 6 at MOI 500. In osteoblast,TGF-β3 protein level increased till day 4, and then started to decline.The peak value reached about 4.5 ng/mL ofmedium. It was also foundthat TGF-β3 protein level at MOI 500 was much higher than that atMOI 200 in both fibroblast and porcine chondrocyte. However, TGF-β3 protein level in osteoblast remained almost unchanged at bothMOI 200 and 500.
3.7. Functions of Ad-dual in synovialmesenchymal stem cells at variousMOIs
Since synovial mesenchymal stem cells are a major target of Ad-dual for therapeutic application, we also carried out preliminarystudies on the functions of Ad-dual in synovial mesenchymal stemcells. According to Fig. 5A, the percentage of dead cells remained
Fig. 4. Quantification of type I collagen and TGF-β3. Type I collagen mRNA levels by qRT-PCRare obtained 3 days after infection with various recombinant adenoviruses. Panels J–L showwith Ad-dual at MOI 200 and 500. A,D,G,J human fibroblast; B,E,H,K human foetal osteobla
relatively stable from MOI 10 till 500, whereas transduction at MOI1000 resulted in almost double cell death compared to other groups.Infection efficiency and the percentage of viable infected cells werenot significantly changed fromMOI 100 to 1000. Over 95% of synovialmesenchymal stem cells were infected at MOI 200 and 500, and thepercentage of viable infected cells reached around 90%. Interestingly,we found that cytotoxicity elicited by Ad-dual infection at MOI 500 iscomparable to that by Ad-dual infection at MOI 200, as shown inFig. 5A; however, Ad-sh/Ad-T co-transduction at MOI 200/200resulted in a much higher percentage of dead cells than Ad-dual atMOI 500, even exceeding that in Ad-dual at MOI 1000, which indicatesseverely negative complication induced by adoption of co-transduc-tion strategy in comparison with the use of the bi-functional Ad-dualalone.
From qRT-PCR result shown in Fig. 5B, it is seen that Ad-dual canalso reduce the transcription of type I collagen in synovial mesen-chymal stem cells in an MOI-dependent manner. The amount of type Icollagen mRNA was significantly decreased at MOI 100 or higherMOIs. No remarkable difference existed amongMOI 100, 200 and 500.
(A,B,C) and protein (D,E,F) levels and pictures (G,H,I) by immnunofluorescent stainingthe concentration of released TGF-β3 in culture media from different cell lines infectedst; C,F,I,L porcine chondrocyte.
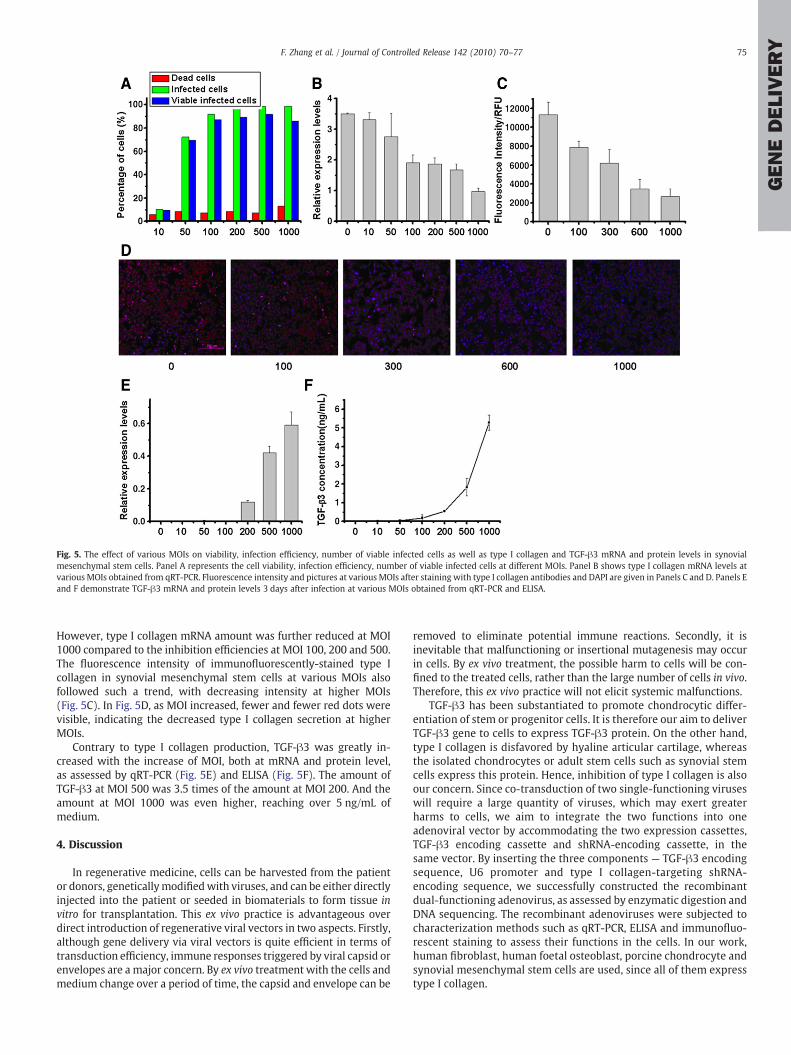
Fig. 5. The effect of various MOIs on viability, infection efficiency, number of viable infected cells as well as type I collagen and TGF-β3 mRNA and protein levels in synovialmesenchymal stem cells. Panel A represents the cell viability, infection efficiency, number of viable infected cells at different MOIs. Panel B shows type I collagen mRNA levels atvarious MOIs obtained from qRT-PCR. Fluorescence intensity and pictures at various MOIs after staining with type I collagen antibodies and DAPI are given in Panels C and D. Panels Eand F demonstrate TGF-β3 mRNA and protein levels 3 days after infection at various MOIs obtained from qRT-PCR and ELISA.
75F. Zhang et al. / Journal of Controlled Release 142 (2010) 70–77
GENEDELIVERY
However, type I collagen mRNA amount was further reduced at MOI1000 compared to the inhibition efficiencies at MOI 100, 200 and 500.The fluorescence intensity of immunofluorescently-stained type Icollagen in synovial mesenchymal stem cells at various MOIs alsofollowed such a trend, with decreasing intensity at higher MOIs(Fig. 5C). In Fig. 5D, as MOI increased, fewer and fewer red dots werevisible, indicating the decreased type I collagen secretion at higherMOIs.
Contrary to type I collagen production, TGF-β3 was greatly in-creased with the increase of MOI, both at mRNA and protein level,as assessed by qRT-PCR (Fig. 5E) and ELISA (Fig. 5F). The amount ofTGF-β3 at MOI 500 was 3.5 times of the amount at MOI 200. And theamount at MOI 1000 was even higher, reaching over 5 ng/mL ofmedium.
4. Discussion
In regenerative medicine, cells can be harvested from the patientor donors, geneticallymodifiedwith viruses, and can be either directlyinjected into the patient or seeded in biomaterials to form tissue invitro for transplantation. This ex vivo practice is advantageous overdirect introduction of regenerative viral vectors in two aspects. Firstly,although gene delivery via viral vectors is quite efficient in terms oftransduction efficiency, immune responses triggered by viral capsid orenvelopes are a major concern. By ex vivo treatment with the cells andmedium change over a period of time, the capsid and envelope can be
removed to eliminate potential immune reactions. Secondly, it isinevitable that malfunctioning or insertional mutagenesis may occurin cells. By ex vivo treatment, the possible harm to cells will be con-fined to the treated cells, rather than the large number of cells in vivo.Therefore, this ex vivo practice will not elicit systemic malfunctions.
TGF-β3 has been substantiated to promote chondrocytic differ-entiation of stem or progenitor cells. It is therefore our aim to deliverTGF-β3 gene to cells to express TGF-β3 protein. On the other hand,type I collagen is disfavored by hyaline articular cartilage, whereasthe isolated chondrocytes or adult stem cells such as synovial stemcells express this protein. Hence, inhibition of type I collagen is alsoour concern. Since co-transduction of two single-functioning viruseswill require a large quantity of viruses, which may exert greaterharms to cells, we aim to integrate the two functions into oneadenoviral vector by accommodating the two expression cassettes,TGF-β3 encoding cassette and shRNA-encoding cassette, in thesame vector. By inserting the three components — TGF-β3 encodingsequence, U6 promoter and type I collagen-targeting shRNA-encoding sequence, we successfully constructed the recombinantdual-functioning adenovirus, as assessed by enzymatic digestion andDNA sequencing. The recombinant adenoviruses were subjected tocharacterization methods such as qRT-PCR, ELISA and immunofluo-rescent staining to assess their functions in the cells. In our work,human fibroblast, human foetal osteoblast, porcine chondrocyte andsynovial mesenchymal stem cells are used, since all of them expresstype I collagen.

76 F. Zhang et al. / Journal of Controlled Release 142 (2010) 70–77
GENEDELIVERY
Before testing the functions of the recombinant adenoviruses,we assessed the performance of (co-)transduction with Ad-dual orAd-sh/Ad-T combination. For the same amount of genes to be dosed tothe cells, when MOI 200 was applied for the transduction with Ad-dual, co-transduction should be carried out at MOI 200 for both Ad-shand Ad-T, which means that doubled amount of adeno-virions wasused for co-transduction in comparison with the Ad-dual transduc-tion. However, according to the result, doubled MOI in co-transduc-tion samples didn't significantly increase the general transductionefficiency (by counting the percentage of infected cell) but elevatedthe toxicity up to nearly 275%. Further taking account that tissueengineering or therapeutic regeneration generally demands largenumbers of cells — and consequently large amount of viral vectors,the doubled cost and tripled toxicity along with the strategy of Ad-sh/Ad-T co-transductionmake it infeasible and unacceptable for practicalapplications in contrast with the Ad-dual strategy.
For viral application, optimum MOI needs to be determined foreach cell type. Here flowcytometry was performed to assess the effectof various MOIs on cell viability and transduction efficiency. Atappropriate MOI, a large majority, if not all, of the cells should beinfected with the virus. At various MOIs the percentages of dead cellsdidn't change significantly, indicating that the recombinant dual-functioning adenovirus does not affect cell viability significantly.Transduction efficiency can reach up to 80–90%, while the viableinfected cells account for 70–80% of total at both MOI 200 and 500.The percentages are large enough for application or cell sorting. qRT-PCR for TGF-β3 and type I collagen mRNA levels was also performed.No significant difference was found between MOI 200 and 500 interms of type I collagen mRNA level, whereas TGF-β3 mRNA level wasmuch higher at MOI 500. Therefore, MOI 500 seems to be optimal forall the cell types. However, for discretion, we tried both MOI 200 and500 in the following experiments. If the adenoviral vector performswell at MOI 200, we may as well reduce its amount of usage to MOI200 in order to minimize its possible side effect, rather than simplyapplying MOI 500.
For type I collagen inhibition efficiency, we used all the single-functioning adenoviruses (Ad-sh andAd-T) aswell as Ad-null to comparetheir effect on type I collagen expression. It was found that even Ad-nullcan raise the expression of type I collagen mRNA to some extent. Othershave reported that Malignant rabbit fibroma virus (MV)was able to elicitthe increase of type I collagenmRNA level in vitro [24]. Here interestinglywe observed that null adenoviruses also have such effect to raise type Icollagen mRNA levels. It is to our expectation that Ad-sh has stronginhibitive effect on type I collagen expression. However, Ad-T seems tohave diversified effects depending on cell types. It raised type I collagenmRNA level in fibroblast while reducing mRNA level in osteoblast. Inporcine chondrocyte, Ad-T does not exert remarkable effect. Thisdiscovery differs from previous studies, which showed that TGF-β3could decrease type I collagen amount and improves scar-free woundhealing [25]. Ad-dual reduced type I collagen mRNA level significantlycompared to either the negative control or Ad-null group especially infibroblast andosteoblast.However, its inhibitiveefficiencywasnot ashighasAd-sh. This indicates that the expressionof shRNA is less efficient by theinfectionwithAd-dual than thatwithAd-sh. However, conclusions can bedrawn thatAd-dual didhaveanobvious inhibitive effect on type I collagenexpression. To improve the silencing efficacy, the vector can beconstructed in the manner that the two expression cassettes (TGF-β3encoding cassette and type I collagen-targeting shRNA-encoding cassette)are located a certain distance away from each other. Besides, compared toAd-T group, no matter whether Ad-T raises or reduces type I collagenmRNA production, type I collagen mRNA level in the Ad-dual group wasalways below that in Ad-T group. This demonstrates that even if TGF-β3can down-regulate type I collagen production in a specific cell type, Ad-dual works better due to the dual effects. It is therefore of significance toconstruct such a dual-functioning adenovirus even if TGF-β3 can inhibittype I collagen production.
It should be noted that expression of type I collagen is onlypartially blocked. Since adenovirus works in an episomal fashion, theexpression of delivered genes will be attenuated after each cycle ofcell replication. Hence, the function of type I collagen inhibition willbe undermined after a certain period of time. Therefore, improvementcan be made by applying the strategies in this model dual-functioningadenoviral vector to a dual-functioning retroviral vector, which isintegrative and thus can almost completely block type I collagenproduction.
It is also notable that for porcine chondrocyte, type I collagenknock-down efficiency was not as high as that in fibroblast andosteoblast. The underlying reason might be that there are two base-pair mismatches between the designed siRNA sequence specific tohuman type I collagen α1 chain and porcine type I collagen genesequence, which can reduce the blocking efficiency significantly [26].
Through qRT-PCR and ELISA, it is clearly evidenced that TGF-β3 isexpressed in the three cell types both at mRNA and protein levels. Atprotein level, TGF-β3 expression at MOI 500 was much higher thanthat at MOI 200 in fibroblast and chondrocyte. However, for osteoblastTGF-β3 protein released at MOI 500 did not differ significantly fromthat at MOI 200. This might be due to the short half-life of TGF-β3mRNA in osteoblast before its translation into proteins. It is also notedthat TGF-β3 protein level increased up to six days after transduction infibroblast and porcine chondrocytes, while the amount started todecrease at day 4 for osteoblast. This indicates that TGF-β3 is moretransiently expressed in osteoblast than the other two cell types. Inporcine chondrocytes, the amount of released TGF-β3 is relativelylower. In order to reach the amount that has therapeutic effect, cellpopulation can be increased for therapeutic applications.
Since synovial mesenchymal stem cells are a possible source forchondrogenesis, we carried out preliminary studies on the efficacy ofAd-dual in this cell type. Results have shown that after infection withAd-dual, type I collagen production was inhibited while TGF-β3 wasexpressed, indicating the successful functioning of Ad-dual in synovialmesenchymal stem cells. Therefore, the construction of Ad-dual hasprovided a potential tool to promote type I collagen suppressedchondrogenesis using synovial mesenchymal stem cells.
In conclusion, we developed amodel adenoviral co-vector that cansimultaneously deliver TGF-β3 and siRNA that targets type I collagento cells toward type I collagen suppressed chondrogenesis. Therecombinant adenovirus was subjected to characterization methodssuch as qRT-PCR, ELISA and immunofluorescent staining to assess itsfunctions in the cells. Human fibroblast, human foetal osteoblast,porcine chondrocyte and synovial mesenchymal stem cells were used.Results have all demonstrated that our recombinant adenovirus canperform the two functions as expected. It can both efficiently down-regulate the expression of type I collagen and produce TGF-β3. Par-ticularly, as synovial mesenchymal stem cells are one of our targetcells, hereby we have provided preliminary results showing that Ad-dual can function well in this cell type. Therefore, this adenoviralvector, when applied to therapeutic cells, has great potential topromote type I collagen suppressed chondrogenesis and cartilagerepair.
Acknowledgements
This researchwas financially supported by AcRF Tier 1 Grant RG64/08, Ministry of Education (MoE), Singapore. We acknowledge RohanRajeev Varshney for his kind technical support for our work.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.jconrel.2009.09.027.

77F. Zhang et al. / Journal of Controlled Release 142 (2010) 70–77
GENEDELIVERY
References
[1] J.A. Buckwalter, Articular cartilage injuries, Clin. Orthop. Relat. Res. 404 (2002)21–37.
[2] L. Stoick-Cooper, R.T. Cristi, G. Moon, Advances in signaling in vertebrateregeneration as a prelude to regenerative medicine, Genes Dev. 21 (2007)1292–1315.
[3] K. Muneoka, C.H. Allan, X. Yang, J. Lee, M. Han, Mammalian regeneration andregenerative medicine, Birth Defects Res. C Embryo Today 84 (2008) 265–280.
[4] A.F. Steinert, U. Noth, R.S. Tuan, Concepts in gene therapy for cartilage repair.Injury, Int. J. Care Injured 39 (2008) S97–S113.
[5] M.H. Porteus, J.P. Connelly, S.M. Pruett, A look to future directions in gene therapyresearch for monogenic diseases, PLoS Genet 2 (2006) 1285–1292.
[6] K. Yun, H.T. Moon, Inducing chondrogenic differentiation in injectable hydrogelsembedded with rabbit chondrocytes and growth factor for neocartilage formation,J. Biosci. Bioeng. 105 (2008) 122–126.
[7] K. Na, S. Kim, D.G. Woo, B.K. Sun, H.N. Yang, H.M. Chung, K.H. Park, Synergisticeffect of TGFbeta-3 on chondrogenic differentiation of rabbit chondrocytes inthermo-reversible hydrogel constructs blended with hyaluronic acid by in vivotest, J. Biotechnol. 128 (2007) 412–422.
[8] C.K. Chuang, L.Y. Sung, S.M. Hwang, W.H. Lo, H.C. Chen, Y.C. Hu, Baculovirus as anew gene delivery vector for stem cell engineering and bone tissue engineering,Gene Ther. 14 (2007) 1417–1424.
[9] L. Meinel, S. Hofmann, O. Betz, R. Fajardo, H.P. Merkle, R. Langer, C.H. Evans, G.Vunjak-Novakovic, D.L. Kaplan, Osteogenesis by human mesenchymal stem cellscultured on silk biomaterials: comparison of adenovirus mediated gene transferand protein delivery of BMP-2, Biomaterials 27 (2006) 4993–5002.
[10] T.T. Tang, B. Lu, B. Yue, X.H. Xie, Y.Z. Xie, K.R. Dai, J.X. Lu, J.R. Lou, Treatment ofosteonecrosis of the femoral head with hBMP-2-gene-modified tissue-engineeredbone in goats, J. Bone Joint Surg. Br. 89 (2007) 127–129.
[11] T. Zachos, A. Diggs, S. Weisbrode, J. Bartlett, A. Bertone, Mesenchymal stem cell-mediated gene delivery of bone morphogenetic protein-2 in an articular fracturemodel, Mol. Ther. 15 (2007) 1543–1550.
[12] H. Aslan, Y. Zilberman, V. Arbeli, D. Sheyn, Y. Matan, M. Liebergall, J.Z. Li, G.A. Helm,D. Gazit, Z. Gazit, Nucleofection-based ex vivo nonviral gene delivery to humanstem cells as a platform for tissue regeneration, Tissue Eng. 12 (2006) 877–889.
[13] J.H. Hao, Y.C. Yao, R.R. Varshney, L.C. Wang, C. Prakash, H. Li, D.A. Wang, Genetransfer and living release of transforming growth factor-β3 for cartilage tissueengineering applications, Tissue Eng: Part C 14 (2008) 273–280.
[14] A.M. Bhosale, J.B. Richardson, Articular cartilage: structure, injuries and review ofmanagement, Brit. Med. Bull. 87 (2008) 77–95.
[15] R. Mayne, M.S. Vail, P.M. Mayne, E.J. Miller, Changes in type of collagensynthesized as clones of chick chondrocytes grow and eventually lose divisioncapacity, Proc. Natl. Acad. Sci. U. S. A. 73 (1976) 1674–1678.
[16] A. Yokoyama, I. Sekiya, K. Miyazaki, S. Ichinose, Y. Hata, T. Muneta, In vitrocartilage formation of composites of synovium-derived mesenchymal stem cellswith collagen gel, Cell Tissue Res. 322 (2005) 289–298.
[17] Y.C. Yao, C.M. Wang, R.R. Varshney, D.A. Wang, Antisense makes sense inengineered regenerative medicine, Pharm. Res. 26 (2008) 263–275.
[18] S. Brantl, Antisense-RNA regulation and RNA interference, BBA-Gene Struct. Expr.1575 (2002) 15–25.
[19] A. Inoue, K.A. Takahashi, O. Mazda, R. Terauchi, Y. Arai, T. Kishida, M. Shin-Ya, H.Asada, T. Morihara, H. Tonomura, S. Ohashi, Y. Kajikawa, Y. Kawahito, J. Imanishi,M. Kawata, T. Kubo, Electro-transfer of small interfering RNA ameliorated arthritisin rats, Biochem. Biophys. Res. Commun. 336 (2005) 903–908.
[20] L.L. Southwood, D.D. Frisbie, C.E. Kawcak, C.W. McIlwraith, Delivery of growthfactors using gene therapy to enhance bone healing, Vet. Surg. 33 (2004) 565–578.
[21] J.A. Spector, B.J. Mehrara, J.S. Luchs, J.A. Greenwald, P.J. Fagenholz, P.B. Saadeh, D.S.Steinbrech, M.T. Longaker, Expression of adenovirally delivered gene products inhealing osseous tissues, Ann. Plast. Surg. 44 (2000) 522–528.
[22] W.B. Tsai, C.H. Chen, J.F. Chen, The effects of types of degradable polymers onporcine chondrocyte adhesion, proliferation and gene expression, J. Mater. Sci.:Mater. Med. 17 (2006) 337–343.
[23] J.B. Fan, R.R. Varshney, L. Ren, D.Z. Cai, D.A. Wang, Synovium-derived mesenchy-mal stem cells: a new cell source for musculoskeletal regeneration, Tissue Eng:Part B 15 (2009), doi:10.1089/ten.teb.2008.0586.
[24] D.S. Strayer, Effect of virus infection on levels of transcripts for cellular genes, VirusRes. 30 (1993) 259–269.
[25] R. Hosokawa, K. Nonaka, M. Morifuji, L. Shum, M. Ohishi, TGF-β3 decreases type Icollagen and scarring after labioplasty, J. Dent. Res. 82 (2003) 558–564.
[26] S.E. Martin, N.J. Caplen, Mismatched siRNAs downregulate mRNAs as a function oftarget site location, FEBS Lett 580 (2006) 3694–3698.





























