Embed Size (px)
Citation preview

1
UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS FARMACÊUTICAS
A RETIRADA DE CETAMINA PROMOVE EFEITO ANSIOGÊNICO,
DÉFICIT DE MEMÓRIA DE CURTO PRAZO E ALTERAÇÕES NO
COMPORTAMENTO DEPRESSIVO EM RATAS ADOLESCENTES.
Sabrina de Carvalho
BELÉM – PA
2016

2
UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS FARMACÊUTICAS
A RETIRADA DE CETAMINA PROMOVE EFEITO ANSIOGÊNICO,
DÉFICIT DE MEMÓRIA DE CURTO PRAZO E ALTERAÇÕES NO
COMPORTAMENTO DEPRESSIVO EM RATAS ADOLESCENTES.
Autor: Sabrina de Carvalho
Orientadora: Profª. Dra. Cristiane do Socorro Ferraz Maia
Dissertação apresentada ao Programa de Pós-graduação em
Ciências Farmacêuticas, área de concentração: Fármacos e
Medicamentos, do Instituto de Ciências da Saúde da
Universidade Federal do Pará como requisito para a obtenção do
título de Mestre em Ciências Farmacêuticas.
BELÉM – PA
2016

3
Dados Internacionais de Catalogação-na-Publicação (CIP)
Biblioteca do Instituto de Ciências da Saúde / UFPA
Carvalho, Sabrina de.
A retirada de cetamina promove efeito ansiogênico, déficit de memória de
curto prazo e alterações no comportamento depressivo em ratas adolescentes
/ Sabrina de Carvalho; orientadora, Cristiane do Socorro Ferraz Maia. –
2016.
Dissertação (Mestrado) - Universidade Federal do Pará, Instituto de
Ciências da Saúde, Programa de Pós-Graduação em Ciências Farmacêuticas,
Belém, 2016.
1. Cetamina. 2. Alterações Neurocomportamentais. 3. Ansiedade. 4.
Depressão. 5. Memória. I. Título.
CDD: 22. ed. : 615.1

4
SABRINA DE CARVALHO
A RETIRADA DE CETAMINA PROMOVE EFEITO ANSIOGÊNICO, DÉFICIT DE
MEMÓRIA DE CURTO PRAZO E ALTERAÇÕES NO COMPORTAMENTO
DEPRESSIVO EM RATAS ADOLESCENTES.
Dissertação apresentada ao Programa de Pós-graduação em
Ciências Farmacêuticas, do Instituto de Ciências da Saúde da
Universidade Federal do Pará para obtenção do título de Mestre.
Área de concentração: Fármacos e Medicamentos
Aprovado em ___/___/___
Banca Examinadora
____________________________________________
Prof. Dr. Cristiane do Socorro Ferraz Maia, PPGCF/UFPA (Orientadora)
___________________________________________
Prof. Dr. Enéas Andrade Fontes Junior, PPGCF/UFPA
____________________________________________
Prof. Dr. Rui Daniel Schröder Prediger, UFSC

5
DEDICATÓRIA
Dedico este trabalho a Deus, por direcionar meus
passos nesta jornada árdua e brilhante. Às minhas mães
(Deize, Tereza e Dione) e irmãos (Anderson e Juliana)
pelo carinho, incentivo e amor. Ao meu esposo Jessé
Cartágenes, por seu companheirismo ao longo da
elaboração deste trabalho.

6
AGRADECIMENTOS
Inicio meus agradecimentos por Deus, já que Ele colocou pessoas tão especiais ao
meu lado. Sou grata, pelo fôlego de vida e pelas lindas oportunidades que me
proporcionou até este momento.
A minhas mães, Tereza, Deize, Dione e Doralice, meu infinito agradecimento.
Sempre acreditaram em minha capacidade. Obrigada pelo amor incondicional!
A meu querido esposo, Jessé, por ser tão importante na minha vida. Sempre ao meu
lado, me pondo para cima e me fazendo acreditar que posso mais que imagino.
Devido a seu companheirismo, amizade, paciência, compreensão, apoio, alegria e
amor, este trabalho pôde ser concretizado. Obrigada por ter feito do meu sonho o
nosso sonho!
A meus irmãos, Juliana e Anderson, pois, a seu modo, sempre se orgulharam de
mim e confiaram em meu trabalho. Obrigada pela confiança!
A meus tios, tias, primos e primas, especialmente tia Dayse e primas Larissa, Lays e
Bianca que vibraram comigo, desde a aprovação. Obrigada pela força!
À minha amiga, Josiane, por só querer o meu bem e me valorizar tanto como
pessoa. Obrigada pela amizade!
A meus amigos do mestrado, pelos momentos maravilhosos, especialmente à
Luanna, que esteve braço a braço nessa caminhada, compartilhando momentos de
muito conhecimento. Obrigada por dividir comigo as angústias e alegrias!
A todos os alunos de iniciação científica que me ajudaram ativa ou passivamente
neste projeto.
Ao professor Enéas pelos momentos de muito conhecimento transmitido e de muita
sabedoria.
À orientadora Cristiane Maia, que sempre acreditou em meu potencial de uma forma
a que eu não acreditava ser capaz de corresponder, sou grata pela construção do
meu conhecimento durante esses sete anos de convívio, que Deus continue lhe
abençoando em conhecimento e sabedoria.

7
“Que os vossos esforços desafiem as impossibilidades,
lembrai-vos de que as grandes coisas do homem foram
conquistadas do que parecia impossível”.
Charles Chaplin

8
RESUMO
CARVALHO, S. A retirada de cetamina promove efeito ansiogênico, déficit de
memória de curto prazo e alterações no comportamento depressivo em ratas
adolescentes. 66f. Dissertação (Mestrado) – Faculdade de Farmácia, Universidade
Federal do Pará, Belém, 2016.
O uso de substâncias psicotrópicas é frequente na população mundial. O consumo
dessas drogas está relacionado ao desejo do homem em explorar suas emoções e
sentidos através do prazer, bem como, para aliviar sentimentos dolorosos. Por muito
tempo, havia o predomínio do uso de substâncias estimulantes pelos adictos,
principalmente entre os adolescentes. No entanto, drogas com efeitos alucinógenos,
como a cetamina, têm sido consumidas para esses fins. O presente estudo
investigou alterações comportamentais após retirada de cetamina em ratos fêmeas
adolescentes. Este projeto foi aprovado pelo comitê de ética em pesquisas
envolvendo animais da UFPA (BIO-CEPAE-UFPA: BIO 224-14). Foram utilizados
ratos Wistar fêmeas do 35º ao 37º dia pós-natal (n=20, n=10 por grupo) que
receberam por via intraperitoneal cetamina na dose de 10 mg/kg/dia, durante três
dias consecutivos. Três horas após a última administração, os animais foram
submetidos sequencialmente aos ensaios do campo aberto, labirinto em cruz
elevado, nado forçado e esquiva inibitória do tipo stem-down. A retirada de cetamina
promoveu efeito ansiogênico, déficit de memória de curta duração e comportamento
do tipo depressivo nos roedores. Não foi observado prejuízos na atividade
locomotora espontânea. Em conjunto, os resultados do presente estudo demonstram
que a retirada de doses subanestésicas de cetamina promove alterações
neurocomportamentais em ratos fêmeas adolescentes.
Palavra-chave: Adolescente, ansiedade, cetamina, depressão, memória.

9
ABSTRACT
CARVALHO, S. Ketamine withdrawal promotes anxiogenic effects, short-term
memory deficit and alterations in depressive behavior in rats adolescents.
2016. 66f. Dissertation (Master degree) – Faculdade de Farmácia, Universidade
Federal do Pará, Belém, 2016.
The use of drugs is common in the world population. The consumption of these drugs
is related to the desire of man to explore your emotions and senses through
pleasure. For a long time, there was a predominance of stimulants by addicts,
especially among adolescents. However, drugs with hallucinogenic effects, such as
ketamine, have been consumed for these purposes. The present study investigated
the behavioral changes after ketamine withdrawal in rats female adolescents. This
project was approved by the ethics committee on research involving animals UFPA
(BIO-CEPAE-UFPA: BIO 224-14). Female Wistar rats of 35 to 37 day postnatal
(n=20, n=10 per group) received by intraperitoneal ketamine at dose of 10
mg/kg/day, for three consecutive days. Three hours after last administration, the
animals were subjected sequentially to the open field test, elevated plus maze,
forced swimming and step-down inhibitory avoidance test. The withdrawal of
ketamine, promoted anxiogenic effect, memory deficit and short-depressive type
behavior in rodents. There was no damage in spontaneous locomotor activity.
Together, the results of this study demonstrate that the removal of subanesthetic
doses of ketamine promotes neurobehavioural changes in rats adolescent females.
Keyword: Adolescent, anxiety, ketamine, depression, memory.

10
LISTA DE FIGURAS
Figura 1: Fórmula estrutural enantiomérica da cetamina..........................................20
Figura 2: Diagrama esquemático do receptor de NMDA...........................................22
Figura 3: Campo Aberto............................................................................................31
Figura 4: Labirinto em Cruz Elevado.........................................................................32
Figura 5: Nado Forçado.............................................................................................33
Figura 6: Esquiva Inibitória........................................................................................34
Figura 7: Efeitos da administração de cetamina na atividade locomotora
espontânea, avaliada no teste do campo aberto.......................................................37
Figura 8: Efeitos da administração de cetamina na atividade locomotora
espontânea, no teste do Labirinto em cruz elevado...................................................38
Figura 9: Efeitos da administração de cetamina para respostas do tipo ansiedade,
no teste do labirinto em cruz elevado.........................................................................39
Figura 10: Efeitos da administração de cetamina no comportamento depressivo, no
teste do Nado forçado................................................................................................40
Figura 11: Efeitos da administração de cetamina na memória de curta duração, no
teste da Esquiva inibitória...........................................................................................40

11
LISTA DE QUADRO
Quadro 1: Descrição dos grupos experimentais...................................................... 30

12
LISTA DE ABREVIATURAS E SIGLAS
AMPA alfa-amino-3-hidroxi-metil-5-4-isoxazolpropiónico
ANOVA Análise de variância
CPF Córtex pré-frontal
DA Dopamina
e.p.m erro padrão médio
I.P Intraperitoneal
LAFICO Laboratório de Farmacologia da Inflamação e do
Comportamento
LCE Labirinto em Cruz Elevado
mGluR Receptor metabotrópico de glutamato
NMDA N-metil D-aspartato
PCP Fenciclidina
pKa Potencial de Ionização
R(-) Isômero dextrogiro da cetamina
S(+) Isômero levogiro da cetamina
S.S Solução Salina
SNC Sistema nervoso central
UNODOC Escritório das Nações Unidas sobre Drogas e Crime

13
SUMÁRIO
I. INTRODUÇÃO.......................................................................................................15
1.1. A adolescência e o Sistema Nervoso Central..........................................16
1.2. Substâncias psicotrópicas na adolescência.............................................17
1.3. Cetamina..................................................................................................19
1.3.1. QUÍMICA E FARMACOCINÉTICA DA CETAMINA..............................19
1.3.2. VIAS DE ADMINISTRAÇÃO.................................................................21
1.3.3. MECANISMO DE AÇÃO DA CETAMINA.............................................22
1.3.4. CETAMINA E O USO RECREATIVO...................................................23
1.3.5. NEUROTOXICIDADE DA CETAMINA.................................................24
II. OBJETIVOS..........................................................................................................27
2.1. Geral.......................................................................................................28
2.2. Específicos.............................................................................................28
III. METODOLOGIA.................................................................................................29
3.1. Animais e grupos experimentais............................................................30
3.2. Tratamento com cetamina.....................................................................31
3.3. Ensaios comportamentais.....................................................................31
3.3.1. TESTE DA AIVIDADE LOCOMOTORA ESPONTÂNEA...................31
3.3.2. TESTE DO LABIRINTO EM CRUZ ELEVADO.................................32
3.3.3 TESTE DO NADO FORÇADO...........................................................33
3.3.4. TESTE DA ESQUIVA INIBITÓRIA....................................................34
3.4. Análise estatística.................................................................................35
IV. RESULTADOS.................................................................................................36

14
4.1. Atividade Locomotora Espontânea......................................................37
4.2. Resposta do tipo ansiedade................................................................38
4.3. Teste do Nado Forçado.......................................................................39
4.4. Teste da Esquiva Inibitória..................................................................40
V. DISCUSSÃO.....................................................................................................41
VI. CONCLUSSÃO................................................................................................46
VII. REFERÊNCIAS...............................................................................................48
VIII. ANEXOS........................................................................................................66

15
I. INTRODUÇÃO

16
1.1. A adolescência e o desenvolvimento do Sistema Nervoso Central (SNC)
O termo “adolescência” é geralmente usado para descrever um período de
transição entre a infância e a idade adulta (SPEAR, 2000; CREWS et al. 2007), é
nessa fase que o indivíduo sofre transformações físicas, psicossociais e cognitivas
(PICKLES et al. 1998).
A faixa etária de 10 aos 20 anos de idade compreende a fase da adolescência
em humanos, segundo a Organização Mundial de Saúde - OMS (1965). Esse
também é o critério adotado pelo Ministério da Saúde do Brasil (Brasil, 1999a) e pelo
Instituto Brasileiro de Geografia e Estatística – IBGE (Brasil, 1998). Em roedores, o
período que engloba a adolescência corresponde à faixa etária entre 28-42 dias pós-
natais. Nesta faixa etária ocorrem mudanças comportamentais nos animais em
relação ao adolescente juvenil e tardio, associadas aos picos no crescimento
corpóreo, abertura vaginal nas fêmeas e aumento da maturidade espermática nos
túbulos seminíferos dos machos (SPEAR, 2000).
O encéfalo adolescente passa por um importante processo de transição. Pode
ser observado alterações progressivas e regressivas no SNC, fornecendo uma base
biológica para os comportamentos específicos da adolescência, associadas às
mudanças no comportamento durante a maturação para a vida adulta (CREWS et al.
2007). Importantes mudanças estruturais e funcionais na plasticidade sináptica e
conectividade neural ocorrem durante o desenvolvimento e maturação do SNC, a fim
de que o cérebro refine suas conexões gerando maior comunicação entre as regiões
cerebrais, possibilitando melhor interação e aumento da complexidade em funções
específicas (GUERRI e PASCUAL, 2010).
Podem ser observadas alterações tanto em componentes corticais e subcorticais
no cérebro adolescente, nos quais simultaneamente o volume da substância
cinzenta diminui enquanto o volume da substância branca aumenta (GIEDD, 2008).
Este evento está associado com um aumento na produção de axônios e sinapses no
início da puberdade e com uma rápida “poda” na adolescência tardia demarcando a
restruturação e refinamento de conexão entre neurônios e glia (CREWS et al. 2007;
GUERRI e PASCUAL, 2010).

17
Em humanos e roedores adolescentes, uma das transformações mais relevantes
durante a maturação do SNC é a reorganização das regiões de córtex pré-frontal e
áreas cerebrais do sistema límbico, que incluem: amígdala, hipocampo, núcleo
accumbens, hipotálamo e córtices frontais, pré-frontal e órbito-frontal (SPEAR, 2000;
CREWS et al. 2007; GUERRI e PASCUAL, 2010). Estas regiões são responsáveis
por mediar funções executivas - como controle do impulso, atenção, inibição da
resposta e planejamento de tarefas - e respostas comportamentais complexas, tais
como aprendizagem e memória (PASCUAL et al. 2011).
Desta forma, as mudanças no remodelamento destas circuitarias neurais estão
associadas com modificações funcionais cognitivas e com a aquisição de funções
executivas (CREWS et al. 2007; GUERRI e PASCUAL, 2010). Vale ressaltar que o
córtex pré-frontal é a última região encefálica em que ocorre o processo de
maturação e pode ser dotado da capacidade de resposta a fatores ambientais.
Como consequência, esta área está sujeita à maior vulnerabilidade a efeitos nocivos
do abuso de drogas e estresse (CREWS et al. 2007).
Portanto, exposição a drogas psicotrópicas durante as fases críticas de
desenvolvimento nos adolescentes, podem perturbar os processos de maturação e
plasticidade cerebral acarretando em déficits comportamentais e cognitivos
(GUERRI e PASCUAL, 2010). Além disso, podem também alterar áreas de
recompensa sensíveis a agentes estressores gerando aumento da ansiedade e
maior propensão à iniciação de consumo de drogas (SPEAR, 2000; GUERRI e
PASCUAL, 2010).
1.2. Substâncias psicotrópicas na adolescência
O consumo de substâncias psicotrópicas é uma prática universal e milenar
(SILVA et al. 2013). Desde tempos remotos, essas sustâncias eram utilizadas para
fins religiosos, culturais e medicinais (FERREIRA FILHO et al. 2003; MARANGONI e
OLIVEIRA, 2013; PRATTA e SANTOS, 2006). Porém, esse consumo, que antes era
restrito a determinadas situações como cerimônias e festas, aos poucos começa a
se disseminar, principalmente entre os adolescentes (MARANGONI et al. 2013).

18
De acordo com o VI Levantamento Nacional sobre o Consumo de Drogas
Psicotrópicas entre Estudantes do Ensino Fundamental e Médio das Redes Pública
e Privada das 27 Capitais Brasileiras, 22,5% relataram uso de alguma droga (exceto
álcool e tabaco), 10,6% referiram uso no último ano e 5,5% referiram uso no mês da
entrevista. Entre os que relataram algum consumo, embora a maioria tivesse idade
maior de 16 anos, também foram observados relatos na faixa entre 10 e 12 anos
(ANDRADE et al. 2010).
Segundo Feinstein et al (2012), indivíduos que iniciam o uso de qualquer
substância psicotrópica antes dos 15 anos, possuem uma probabilidade 6,5 vezes
maior de desenvolverem transtornos relacionados ao uso de drogas que aqueles
que iniciam após 21 anos (28,1% versus 4,3%). Isso se deve às modificações
estruturais que ocorrem nessa fase da vida, que vão desde transformações
moleculares até alterações no volume absoluto do córtex pré-frontal (WILLIS et al.
1994).
De uma forma geral, o álcool e o tabaco são as drogas mais consumidas
entre os adolescentes. Seus consumos são favorecidos por serem drogas lícitas, de
custo reduzido, amplamente disponível e de fácil acesso (JOHNSTON et al. 2009).
Aliado a estes fatores, estão o contexto familiar e de saúde mental. De acordo com
Malta et al. (2012), o efeito protetor da supervisão familiar associado aos aspectos
de solidão, insônia e ausência de amigos, aumenta a probabilidade do consumo de
tabaco, álcool e outras drogas.
Outro ponto importante abordado por Guerreito et al. (2011) é a prevalência
de substâncias em “club drugs”, ou seja, drogas frequentemente utilizadas por
adolescentes e jovens adultos no contexto de saídas à noite em discotecas, raves e
bares. Essas substâncias apresentam efeitos psicotrópicos diversos, estando
associadas à uma variedade de níveis de toxicidade, dependência e efeitos
adversos. As principais drogas observadas neste contexto são a
metilenodioximetanfetamina (MDMA), metanfetamina, ácido lisérgico dietilamida
(LSD), gamma-hidroxibutirato (GHB), flunitrazepam e cetamina.

19
1.3. Cetamina
O cloridrato de Cetamina foi sintetizado pela primeira vez em 1962, pelo
farmacêutico americano Calvin Stevens no laboratório Parke & Davis, com intuito de
substituir a fenciclidina (PCP) (SMITH et al. 2002; MORGAN et al. 2011). Quando
introduzida pela primeira vez para o uso clínico, a cetamina foi considerada uma
droga anestésica ideal e completa, uma vez que fornece todos os componentes
necessários de anestesia cirúrgica: alívio da dor, imobilidade, amnésia e perda de
consciência (ANNETTA et al. 2005; CORAZZA et al. 2013).
Na prática clínica é utilizada para indução de anestesia em pacientes com
choque hemodinâmico; indução da anestesia em pacientes com asma ativa;
sedação intramuscular em pacientes não cooperativos, particularmente em crianças;
sedação em unidades de tratamento intensivo; e na troca de curativos em pacientes
queimados (SILVA et al. 2010).
Alguns estudos relatam um potencial efeito antidepressivo da droga,
principalmente em pacientes resistentes ao tratamento do transtorno depressivo
maior (TDM) (OWOLABI et al. 2014). Além disso, a cetamina é capaz de
restabelecer psicoses em pacientes esquizofrênicos remitentes, através da
avaliação dos sintomas positivos e negativos da esquizofrenia humana tais como os
déficits auditivos e visuais; alterações nas regiões límbicas corticais, formação
hipocampal, núcleo accumbens, amigdala basolateral; aumento cerebral do fluxo
sanguíneo no córtex cingulado anterior e redução cerebral do fluxo sanguíneo no
hipocampo e córtex visual primário (BECKER et al. 2003; BRESSAN e PILOWSKV,
2003; KEILHOFF et al. 2004).
1.3.1. QUÍMICA E FARMACOCINÉTICA DA CETAMINA
A cetamina é uma arilciclohexamina, que apresenta um núcleo quiral
originando dois isômeros ópticos. Possui pKa 7,5, elevada lipossolubilidade, sendo
também solúvel em água (SILVA et al. 2010; LI et al. 2011). É comercialmente
utilizada com uma mistura racêmica dos dois isômeros opticamente ativos
(enantiômeros), um isômero levogiro S(+) e um isômero dextrogiro R(-). O isômero

20
S(+) da cetamina possui potência analgésica e anestésica superiores às do isômero
R(-) e da própria mistura racêmica (SILVA et al. 2010). (Figura 1)
De acordo com os estudos em animais de Plenninger, Durieux e
Himmelseheir (2002), foi demonstrado que a cetamina-S(+) apresenta
aproximadamente quatro vezes mais afinidade para a área de ligação da fenciclidina
no receptor N-metil D-Aspartato (NMDA), quando comparada à cetamina-R(+)
(LAURIETTI et al. 2000; JACSCH et al. 2002 apud SILVA et al. 2010). Sua alta
lipossolubilidade confere rápida e curta duração de ação após uma única dose,
possui baixa ligação às proteínas plasmáticas (27%), resultando em grande volume
de distribuição de aproximadamente 3-5L/Kg. Seu metabolismo envolve
principalmente a enzima citocromo P450 (CYP)2B6 (HIJAZI e BOULIEU, 2002 apud
KALSI et al. 2011; LI et al. 2011).
A biotransformação da cetamina por N-desmetilação leva à formação de
norcetamina e a oxidação do anel ciclohexanona leva à produção de
dehidronorcetamina. Ambos metabólitos possuem atividade farmacológica e são
eliminados na urina. Além do fígado, o metabolismo da cetamina ocorre em menor
proporção nos rins, intestino e pulmões (KALSI et al. 2011).
O clearance de eliminação desta substância é relativamente alto (1000 a 1600
mL.min-1), o que confere uma meia-vida de eliminação em torno de 2 a 3 horas. A
norcetamina, principal metabólito da cetamina, tem uma atividade farmacológica
Figura 1: Fórmula estrutural enantiomérica da cetamina
R(-) -Cetamina S(+) -Cetamina

21
avaliada em torno de 20% da molécula inicial, ou seja, 1/5 da potência da droga
original e surge no plasma dentro de 2 a 3 minutos após a administração
intravenosa. No processo de biotransformação, além da N-desalquilação (metabólito
I) e hidroxilação do anel ciclohexanona – (metabólito III e IV), ocorre também a
conjugação com ácido glicurônico e des-hidratação dos metabólitos hidroxilados,
formando os derivados ciclohexenos – (metabólito II – dehidronorcetamina). Apenas
4% da cetamina é excretada por via renal na forma inalterada. Portanto, uma
possível inadequação na eliminação renal tem pouco efeito cumulativo sobre
cetamina no organismo humano (JANSEN, 2002; LI et al. 2011; SILVA et al. 2010).
1.3.2. VIAS DE ADMINISTRAÇÃO
A cetamina pode ser administrada por várias vias, incluindo intratecal,
intraperitoneal, intranasal e retal. No entanto, geralmente as vias intravenosa e
intramuscular são as mais utilizadas. A administração do fármaco por via oral é
bastante errática. Apesar da facilidade de atravessar as biomembranas, a droga
atinge uma biodisponibilidade muito limitada (17-24%), devido ao intenso
metabolismo hepático de primeira passagem (FANTA et al. 2015; KHARASCH e
LABROO, 1992). Curiosamente, na via oral, os efeitos podem ser prolongados por 4-
6 horas (QUIBELL et al. 2011).
A via intravenosa produz biodisponibilidade máxima e rápido início de efeitos
(menos de 1 min). Na administração intramuscular, ocorre uma rápida absorção,
atingindo uma biodisponibilidade de até 93%, com um pico em cerca de 5 a 15
minutos da administração (CLEMENTS, NIMMO, e GRANT, 1982; MION &
VILLEVIEILLE, 2013).
A insuflação nasal é uma via intimamente ligada ao abuso desta droga. Isso
pode ser explicado, pelo menos em parte, pela rápida absorção, com a
biodisponibilidade de aproximadamente 45%. Por esta via, os efeitos cerebrais são
atingidos em cerca de 5 minutos e está associado à meia-vida curta, o que contribui
para a estimulação de administrações repetidas (YANAGIHARA et al. 2003).

22
Lado Extracelular
Lado Citoplasmático
Fenciclidina (PCP)
Dizocilpina
Cetamina
Glutamato
Glicina (GLY) D-serina
D-cicloserina
1.3.3. MECANISMO DE AÇÃO DA CETAMINA
A cetamina interage com alguns receptores, tais como, receptores
glutamatérgicos NMDA e não-NMDA do tipo α-amino-3-hidroxi-5-metil-isoxazol-4-
propionato (AMPA); bem como, com receptores opioides, gabaérgicos e
serotoninérgicos; além de agir, direta ou indiretamente, sobre as monoaminas:
acetilcolina, noradrenalina e dopamina (OLIVEIRA et al. 2004; GARCIA et al. 2008).
O mecanismo primário de ação da cetamina dá-se pelo bloqueio dos
receptores de NMDA, no interior do canal iônico, no sítio de ligação da fenciclidina
(PCP) (SHIMTH et al. 2004; LI et al. 2011). Esses receptores de glutamato NMDA
(expressos primariamente no hipocampo, no córtex cerebral e na medula espinhal)
são receptores ionotrópicos ativados pelo glutamato, principal neurotransmissor
excitatório do SNC. O glutamato é importante na regulação da plasticidade sináptica,
no aprendizado e memória (GARCIA et al. 2008). A ativação desse receptor exige a
ligação simultânea de glutamato e glicina (co-agonista obrigatório), que promovem a
abertura do canal permitindo o efluxo de K+, bem como o influxo de Na+ e Ca2+ (LI et
al. 2011) (Figura 2).
Figura 2: Diagrama esquemático do receptor de N-metil-D-aspartato (NMDA). A fenciclidina (PCP),
cetamina, e a dizocilpina ligam-se no interior do receptor de NMDA. O receptor de NMDA está
bloqueado por Mg2+
de uma forma sensível à voltagem. A ativação do receptor de NMDA por meio da
ligação de glutamato e glicina resultam na abertura do canal, permitindo o influxo de íons Na+ e
pequenas quantidades de íons de Ca2+
e efluxo de K+. Fonte: Li et al. 2011.
A cetamina não apresenta atividade analgésica sobre os receptores opioides
(HUSTVEIT et al. 1995 apud SILVA et al. 2010), visto que, a naloxona, antagonista

23
opioide, não é capaz de reverter seu efeito analgésico. Isto pode ser explicado por
uma afinidade 10 a 20 vezes inferior pelos receptores opioides do que pelo receptor
NMDA. No entanto, sugere-se que os efeitos adversos psicomiméticos da
substância estariam relacionados com os receptores opioides kappa, pois os
agonistas kappa produzem efeitos similares (RAEDER et al. 2000).
Outra interação observada ocorre indiretamente sobre a neurotransmissão
colinérgica. Em concentrações clínicas, este fármaco inibe a liberação de acetilcolina
mediada pelo receptor NMDA. Essa inibição não apresenta relevância clínica, porém
os efeitos adversos comportamentais podem estar relacionados também a este
efeito (DURVAL, 2004 apud SIILVA et al. 2010).
A captação de dopamina também é inibida pela cetamina, o que pode levar a
um aumento da atividade dopaminérgica central, podendo estar relacionado aos
fenômenos de dependência química, além de precipitar sintomas psicóticos
relacionados à esquizofrenia. Essas evidências levaram diversos autores a
pesquisarem modelos propícios para maiores estudos na área, como um possível
modelo de esquizofrenia (VASCONCELOS et al. 2005).
1.3.4. CETAMINA E O USO RECREATIVO
Os primeiros relatos do consumo recreativo da cetamina aconteceram na
década de 1960, porém nos anos de 1980 e 1990 as notificações se tornaram mais
significativas (MORGAN et al. 2011; TRUJILLO et al. 2011; CORAZZA et al. 2013).
Entre os usuários, a cetamina é conhecida como “Special K”, “vitamina K”, “valium
gato” ou “K". Encontra-se disponível comercialmente como um líquido injetável, mas
geralmente é utilizada como droga de abuso na forma de pó (DILLON et al. 2003;
FREESE et al. 2002).
Geograficamente, o uso recreativo da cetamina é evidenciado mundialmente,
porém, é mais comum no Leste e no Sudeste da Ásia, devido ao seu preço
relativamente baixo em comparação com outras drogas psicomiméticas (UNODC,
2010; KALSI et al. 2011; CHEN et al. 2014). Ela também está se tornando popular
no Reino Unido, Austrália, Hong Kong, e da China Continental (UNODC, 2013).

24
Relatórios globais de apreensão de cetamina demonstraram aumento de
quantidades insignificantes apreendidas em 1999 para mais de 11 toneladas em
2007, com quase tudo isso no Leste e no Sudeste da Ásia, onde as apreensões de
cetamina excederam a de heroína (KALSI et al. 2011). A prevalência exata do uso
recreativo da droga ainda é muito escasso; estudos isolados em determinados
países sugerem que as taxas de utilização da cetamina são baixas, entre 0,1 e 4%
(CHEN et al. 2014).
Adictos afirmam que a droga é gratificante e pode produzir uma variedade de
efeitos psicoativos. De acordo com seus relatos, doses baixas da substância são
capazes de promovem estimulação, excitação, euforia, distorções sensoriais e
embriaguez lúcida, que é intensificada por sentimentos de empatia (DILLON et al.
2003; JANSEN e DARRACOT-ČANKOVIĆ, 2001).
Segundo Dillon et al (2003), doses mais elevadas de cetamina são capazes
de produzir um estado de alucinação referido como "K-furo", uma experiência
dissociativa intensa que inclui visões e distorção de tempo, sentido e de identidade.
Às vezes experiências fora do corpo, perto da morte, ou experiências de
renascimento podem ocorrer.
1.3.5. EFEITOS NEUROTÓXICOS DA CETAMINA
Postula-se um dos efeitos neurotóxicos da cetamina ao período de
sinaptogênese (ZOU et al. 2009). A maior vulnerabilidade do cérebro em
desenvolvimento ocorre no momento da sinaptogênese rápida, também conhecido
como surto de crescimento do cérebro, que ocorre imediatamente após
diferenciação e migração dos neurônios (ZOU et al. 2009). Nos seres humanos,
começa a partir do terceiro trimestre e continua 2 a 3 anos após o nascimento (BAI
et al. 2013), nos roedores ocorre durante as três primeiras semanas pós-natais, com
um pico durante a segunda semana (SEMPLE et al. 2013).
A sinaptogênese é caracterizada pelas alterações morfológicas e bioquímicas
em elementos pré e/ou pós-sinápticos, tais como, as proteínas vesiculares pré-
sinápticas sinaptofisina e sinapsina que estão envolvidas na formação e plasticidade
das sinapses (KIM et al. 2003., ZHAO et al. 2000). Em várias espécies de

25
mamíferos, neurônios presentes no nascimento, são submetidos a um período de
superprodução de arborização e contatos sinápticos, com intuito de aumentar a
densidade sináptica, seguido por uma fase de eliminação ou poda de refinamento
(SEMPLE et al., 2013). Essa poda contribui para a plasticidade e permiti o
processamento mais eficiente da cognição na fase adulta. É uma estratégia de
desenvolvimento básico, comum à maioria das regiões do sistema nervoso central
de mamíferos, incluindo seres humanos, primatas e roedores (ANDERSEN, 2003).
Adicionado a isso, a exposição de cetamina induz neuroapoptose no cérebro
de roedores (IKONOMIDOU et al, 2011; JEVTOVIC-TODOROVIC et al, 2005),
através da desregulação de receptores de NMDA (SLIKKER et al., 2007; WANG et
al., 2005, 2006, ZOU et al, 2009).
Tem sido postulado que a exposição contínua de cetamina desenvolve morte
celular seletiva por um mecanismo que envolve uma regulação positiva das
subunidades compensatórias do receptor de NMDA (WANG et al., 2005, 2006). Esta
regulação positiva inicia com a cascata de influxo de cálcio (Ca+2), que produz um
aumento de espécies reativas de oxigênio (ROS), que parece ser originadas na
mitocôndria (SLIKKER et al., 2007). Além disso, reduz o potencial de membrana e
perturba o transporte de elétrons, levando ao maior aumento da produção de ROS
(SLIKKER et ai, 2007; WANG et al, 2006).
Os componentes centrais da apoptose neuronal ocorre através de um grupo
de enzimas proteolíticas chamadas caspases, que podem ser ativadas por
diferentes tipos de estimulação. Perda do potencial de membrana mitocondrial e
liberação de citocromo c das mitocôndrias são eventos chaves no envolvimento
mitocondrial na apoptose. O citocromo c liberado ativa caspase 9, que
consequentemente induz a ativação da caspase 3, resultando numa clivagem de
várias proteínas celulares, conduzindo finalmente as alterações típicas relacionadas
com a apoptose celular (BAI et al. 2013)
Atribuído a esses eventos a cetamina altera a neurogênese de células
progenitoras do tronco neural no cérebro em desenvolvimento (IKONOMIDOU et al.
1999).
A neurogênese envolve a produção de novos neurônios a partir da
proliferação de células progenitoras, seguido da migração neuronal, maturação e
integração de redes neurais (MENINI, 2010; MING e SONG, 2005).

26
A neurogênese de células progenitoras do tronco neural (neural stem
progenitor cells-NSPCs) determina a quantidade e formação de neurônios,
astrócitos, oligodendróglia e outras linhagens neurais (BAI et al. 2013). Enquanto
isso, as NSPCs exibem duas características, primeiramente, a capacidade de auto
renovação através da divisão celular, em seguida a capacidade de geração de tipo
de célula especializada através da diferenciação (MENINI, 2010).
É sabido que o consumo de cetamina altera a proliferação das células
progenitoras do tronco neural, bem como, “perturba” a expressão normal dos
receptores NMDA no processo de maturação, gerando pré-maturação dos
receptores NMDA funcionais (DONG e ANAND, 2013). Neste cenário, NSPCs têm
menor capacidade de proliferar e de diferenciar em neurônios no início do estágio de
desenvolvimento. Esses neurônios pré-amadurecidos podem tornar-se susceptíveis
à morte celular induzida, como resultado, os padrões normais de desenvolvimento
do cérebro são repostos para um estágio de desenvolvimento mais precoce (BAI et
al. 2013).
Outro fator importante é a atuação do glutamato no processo de diferenciação
neural das NSPCs, pois promove crescimento e diferenciação neuronal. Os
receptores NMDA parecem estar envolvidos na determinação da direção da
diferenciação em NSPCs. O influxo de cálcio tem papel importante na diferenciação
neuronal do NSPCs a exposição à cetamina induz a expressão de subunidades dos
receptores NMDA, formando receptores NMDA mais funcionais, assim que a
diferenciação continua (BAI et al. 2013).
Diante do exposto acima, para testar possíveis hipóteses das alterações
neurocomportamentais após retirada de cetamina, o presente estudo padronizou-se
um modelo que mimetize o consumo recreativo, a fim de investigar os efeitos na
atividade locomotora espontânea, no comportamento do tipo ansiedade,
comportamento depressivo e memória de curto-prazo.

27
II. OBJETIVOS

28
2.1. Objetivo geral:
Investigar os efeitos neurocomportamentais em ratos fêmeas adolescentes
após administração de cetamina por três dias consecutivos.
2.2. Objetivos específicos:
Investigar os efeitos neurocomportamentais gerados pela suspensão da
cetamina em ratas adolescentes após três dias de administração consecutivos,
sobre:
Atividade locomotora espontânea, no teste do Campo Aberto;
Emocionalidade, no teste do Labirinto em Cruz Elevado;
Comportamento do tipo depressivo, no teste do Nado Forçado;
Cognição (memória de curto prazo), no teste da Esquiva Inibitória.

29
III. METODOLOGIA

30
3.1. Animais e Grupos experimentais
Este estudo foi inicialmente aprovado pelo comitê de ética em pesquisa com
animais de experimentação da Universidade Federal do Pará (UFPA), sob parecer
CEPAE_UFPA 224-14, obedecendo aos critérios e as normas estabelecidas por
Guias de Cuidado e Uso de Animais Laboratoriais.
Foram utilizados ratos Wistar, fêmeas (n=20), com 35 dias de vida, e média
de peso entre 60-90g, provenientes do Biotério da UFPA. Estes animais foram
encaminhados para Faculdade de Farmácia (UFPA), no qual foram mantidos em um
biotério de experimentação em condições padronizadas de temperatura à 25ºC,
exaustão, ciclo de luz claro/escuro de 12 horas (7:00- 19:00 horas), água e comida
ad libitum. Nas salas onde ocorreram os experimentos comportamentais utilizou-se
lâmpadas fluorescentes para iluminação (12 lux), e houve controle de temperatura e
ruído.
Os ensaios comportamentais foram realizados no Laboratório de
Farmacologia da Inflamação e do Comportamento (LAFICO), da Faculdade de
Farmácia, do Instituto de Ciências da Saúde da UFPA.
Para este estudo os 20 animais foram divididos em dois grupos experimentais
(Quadro 1):
Quadro 1: Descrição dos grupos experimentais.
GRUPO DESCRIÇÃO NÚMERO
Controle Animais tratados por via I.P. com S.S. por três dias consecutivos 10
Cetamina Animais tratados por via I.P. com 10mg/Kg/dia de cetamina por três dias
consecutivos
10
TOTAL 20
Legenda: S.S= Solução salina; I.P.= intraperitoneal.

31
3.2. Tratamento com Cetamina
Após a divisão dos grupos experimentais os animais foram mantidos em
caixas próprias em grupos de 5 (cinco) animais para evitar o estresse pelo
isolamento. Foi administrado por via intraperitoneal (I.P.) solução salina (S.S) ou
cloridrato de dextrocetamina (Cristália, Brasil) na dose de 10mg/kg/dia, por três dias
consecutivos (HUANG et al. 2012) do 35º ao 37º dia de vida, o que corresponde ao
início da adolescência em modelo animal (SPEAR, 2000; 2004). A cetamina foi
solubilizada em S.S momentos antes da administração por via intraperitoneal em um
volume final de 0,1 mL para cada 100 g de peso do animal (KOS et al. 2010).
3.3. Ensaios Comportamentais
Os ensaios comportamentais foram iniciados 3 horas após a última
administração de cetamina (EIDE et al. 1994; SIGTERMANS et a., 2009; NIESTERS
et al. 2013), não excedendo o horário de 18 horas a fim de não interferir no ciclo
circadiano do animal.
No dia dos ensaios comportamentais, os animais foram encaminhados para o
LAFICO e conduzidos à sala do teste por um período de, no mínimo, uma hora para
aclimatação e habituação ao ambiente.
.
3.3.1. TESTE DA ATIVIDADE LOCOMOTORA ESPONTÂNEA/ CAMPO ABERTO
Neste estudo, foi utilizada uma arena em madeira (100 x 100 x 40 cm),
pintada com material não permeável na cor preta (Figura 3).
Figura 3: Campo Aberto

32
O objetivo deste teste foi avaliar atividade locomotora espontânea
(PANDOLFO et al. 2007), de acordo com dois parâmetros diferentes: a distância
total percorrida (em metros) e o número de levantamento (“rearing”), executado
pelos animais, durante 5 minutos. O primeiro parâmetro foi analisado pelo software
ANY-maze (San Diego, CA) e o segundo foi registrado com o uso de um contador
manual. O ato de levantar foi considerado quando o animal se apoiou nas 2 patas
trazeiras mantendo as 2 patas dianteiras levantadas (BRENES et al. 2009). Estes
procedimentos foram realizados para avaliar possíveis prejuízos locomotores
causados pela suspensão de cetamina.
3.3.2 TESTE DO LABIRINTO EM CRUZ ELEVADO (LCE)
O LCE é um dos modelos validados para verificar comportamento do tipo
ansiogênico em ratos (PELLOW et al. 1985). É baseado em respostas
incondicionadas aos ambientes potencialmente perigosos, tais como a aversão
natural de roedores aos espaços abertos (TREIT et al. 1993). Os índices principais
de ansiedade no teste compreendem medidas espaço-temporais de fuga dos braços
abertos (ACEVEDO et al. 2014).
O LCE é um equipamento em madeira, na forma de cruz, elevado 50 cm do
chão, com dois braços fechados (50 x 10 x 40 cm) e dois abertos (50 x 10 x 1 cm),
opostos entre si (HANDLEY e MITHANI, 1984). Nos braços abertos há uma proteção
de 1 cm de altura circundando-os à fim de impedir a queda dos animais do aparato
(Figura 4).
Figura 4: Labirinto em Cruz Elevado.

33
Após o teste do campo aberto, cada animal foi posicionado no centro do
LCE com a face voltada para um dos braços fechados, no qual foi permitida
exploração espontânea do animal por 5 minutos. Um observador, que não teve
conhecimento prévio do grupo a que pertencia cada animal, registrou o número de
entradas nos braços abertos (EBA) e fechados (EBF), assim como, o tempo de
permanência nos braços abertos. O percentual de EBA (%EBA) e de TBA (%TBA)
foram calculadas de acordo com as fórmulas: (EBA / EBA + EBF) x 100; (TBA / TBA
+ TBF) x 100 (PELLOW; FILE, 1986).
Foi possível observar resposta do tipo ansiedade através da diminuição na
entrada e/ou tempo de permanência nos braços abertos. O parâmetro EBF foi
utilizado para avaliar prováveis alterações motoras nos animais.
3.3.3 TESTE DO NADO FORÇADO
O teste do nado forçado consiste em um cilindro Plexiglass (30 cm de
diâmetro e 50 cm de altura), contendo 40 centímetros de volume de água a uma
temperatura de 23 ± 1 °C (Figura 5). É um teste preditivo para screening de
fármacos antidepressivos (ALCARO et al. 2002; PETIT-DEMOULIERE et al. 2005).
Os roedores geralmente exibem quatro tipos de comportamentos durante o
teste: fuga; imobilidade contínua (permanecer flutuando, mantendo somente os
Figura 5: Nado Forçado.

34
movimentos mínimos necessários para manter a cabeça fora da água); nado
(circulação em todo o cilindro, nadando de um quadrante para o outro); e climbing
(“escalada” é definido como movimentos ascendentes ao longo dos lados do
cilindro).
O teste consistiu em posicionar o animal no centro do equipamento,
permitindo sua livre exploração no aparato por um período de 5 minutos, nos dois
primeiros minutos não foi mensurado nenhum parâmetro, visto que esta fase é
considerada a habituação ao teste. Nos três últimos minutos foi mensurado o tempo
de imobilidade, parâmetro que assumem um comportamento do tipo depressivo,
quando os animais demostram níveis elevados de imobilidade no teste.
3.3.4 TESTE DA ESQUIVA INIBITÓRIA
Após o teste do nado forçado e secagem dos animais, foi realizado o teste da
esquiva inibitória do tipo step-down. Este teste baseia-se num modelo de memória,
que utiliza estímulo aversivo como fator para a obtenção da resposta
comportamental (IZQUIERDO; MEDINA, 1997). O teste foi realizado em uma caixa
de alumínio (50 x 50 x 35 cm), com o assoalho constituído 15 barras de cobre
paralelas, distribuídas com uma distância de 1 cm entre si, conectadas a um
estimulador elétrico. Em posição lateral ao assoalho de barras encontra-se uma
plataforma que não é conectada ao estimulador elétrico (Figura 6).
Figura 6: Esquiva Inibitória.

35
O protocolo utilizado foi adaptado por Costa e Tomaz (1998). O teste
consistiu em uma sessão de habituação de 180 segundos, no qual foi permitido a
exploração interna no aparato. Após 24 horas, os animais foram reexpostos a
plataforma segura com a face voltada de forma oposta ao observador e
imediatamente após o animal descer da plataforma e colocar as quatros patas sobre
as grades, foi aplicado um choque elétrico de 0,4 mA durante 1 segundo (estímulo
aversivo). A medida da retenção da memória de curta duração foi avaliada em uma
sessão de teste realizada 1,5 hora (uma hora e trinta minutos) após o estímulo
aversivo. Para isto, foi realizado o mesmo procedimento, mas omitindo-se o choque,
sendo novamente cronometrado o tempo em que o animal levou para descer da
plataforma. Foi estabelecido um tempo limite de 180 segundos para a execução do
trabalho.
Neste teste, a retenção da memória consistiu em o animal não descer da
plataforma, ou fazê-lo após um período de tempo maior do que aquele observado na
sessão de treino (ROESLER et al. 1999).
3.4. Análise Estatística
Foi realizado o teste de distribuição gaussiana, para cada grupo experimental
pelo teste de Kolmogorov-Smirnov. Após a observação da homogeneidade dos
dados, foi realizado o teste estatístico t de Student e os dados foram expressos pela
média ± erro padrão da média (e.p.m.) de 10 animais de cada grupo experimental,
no qual a probabilidade utilizada como existência de diferença significativa foi
p<0,05. A construção gráfica e análise estatística foi realizada no programa
GraphpPad Prism 5.0.
No teste da esquiva inibitória do tipo step-down foi realizada análise não
paramétrica onde foram apresentadas as medianas e intervalo interquartis das
latências de descida da plataforma e estes foram analisados no teste Wilcoxon (um
teste não paramétrico equivalente ao teste T pareado), e estes foram analisados
estatisticamente através do teste Many Whitney.

36
IV. RESULTADOS

37
4.1. Atividade Locomotora Espontânea
A figura 7 ilustra a atividade locomotora espontânea no teste do campo
aberto, através dos parâmetros distância total percorrida e número de “rearings”
(exploração vertical). Os grupos controle (n=10) e cetamina (n=10), avaliados 3
horas após administração, não apresentaram diferença estatística nas distâncias
percorridas em metros (Fig. 7A) quando avaliadas pelo teste t de Student [t = 0,048;
P = 0,96]. Com relação ao número de levantamentos (Fig. 7B), o teste t de Student [t
= 0,567; P = 0,57] também não indicou diferença estatística entre os grupos,
sugerindo que o grupo tratado com cetamina (10 mg/kg, I.P) não apresentou
comprometimento motor quando comparado ao grupo controle.
A suspensão de cetamina após três dias de administração consecutivos não
comprometeu a atividade motora no teste do campo aberto quando comparado ao
grupo controle [t=0,113; p= 0,91] (Figura 8).
A B
Figura 7: Efeitos observados após 3 horas da última administração de cetamina (10 mg/Kg/dia
durante 3 dias, I.P) e S.S sobre a atividade locomotora espontânea de ratos (fêmeas) adolescentes,
avaliados por 5 minutos no teste do campo aberto. Os resultados estão expressos como a média ±
e.p.m da distância total percorrida em metros (A) e do número de rearing (B) de 10 animais por grupo
(Teste t de Student).
Controle Cetamina0
5
10
15
20
Dis
tân
cia
To
tal P
erc
orr
ida (
m)
Controle Cetamina0
5
10
15
Nº
de levan
tam
en
tos

38
4.2. Respostas do tipo ansiedade
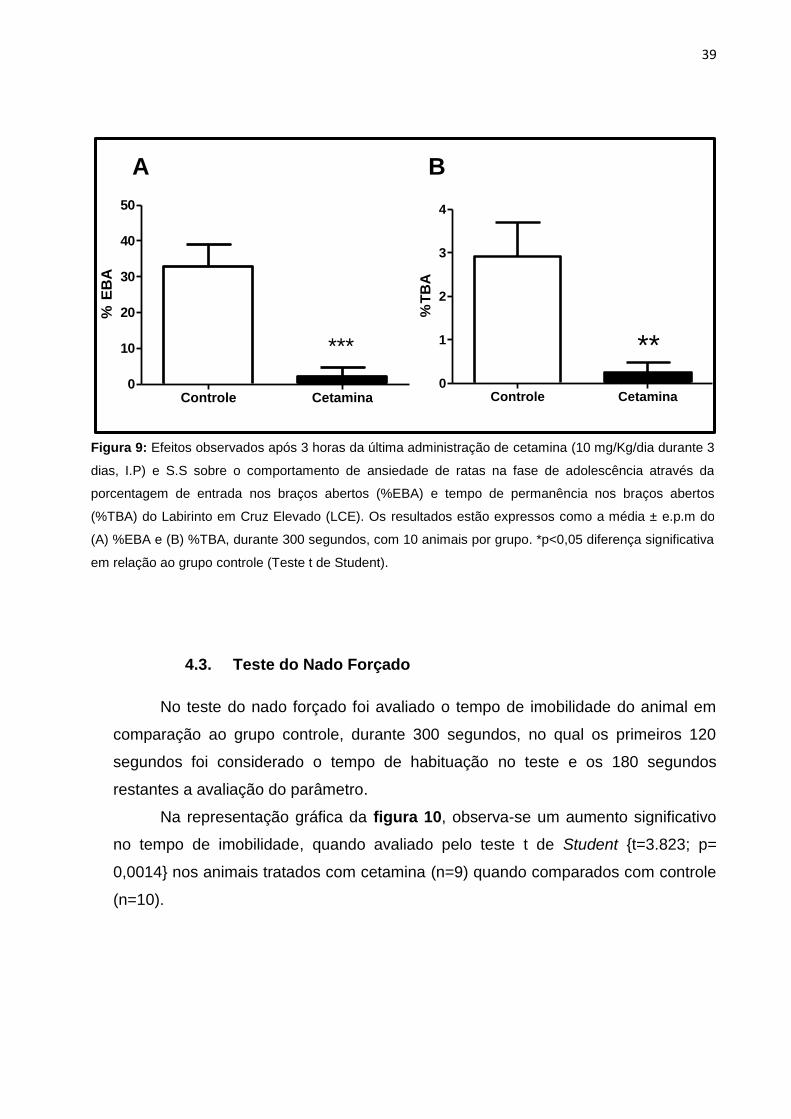
Na figura 9 é possível observar os resultados obtidos no teste do LCE,
através dos parâmetros relacionados à ansiedade (%EBA e %TBA) durante 5
minutos no equipamento. Como pode ser observado, o teste t de Student
demonstrou uma redução de %EBA (Figura 9A) e %TBA (Figura 9B) nos animais
tratados quando comparados ao grupo controle [(t=4,840; p<0,0001), (t=3,391;
p=0,0031), respectivamente], indicativo de possível efeito ansiogênico promovido
pela suspensão da droga.
Controle Cetamina0
1
2
3
4
Nº
En
trad
as b
raço
fech
ad
o
Figura 8: Efeitos observados após 3 horas da última administração de cetamina (10
mg/Kg/dia durante 3 dias, I.P) e S.S sobre a atividade motora espontânea de ratos (fêmeas)
adolescentes, através do número de entradas nos braços fechados do labirinto em cruz
elevado (LCE). Os resultados estão expressos como a média ± e.p.m, durante 5 minutos,
com 10 animais por grupo (Teste t de Student).
39
4.3. Teste do Nado Forçado
No teste do nado forçado foi avaliado o tempo de imobilidade do animal em
comparação ao grupo controle, durante 300 segundos, no qual os primeiros 120
segundos foi considerado o tempo de habituação no teste e os 180 segundos
restantes a avaliação do parâmetro.
Na representação gráfica da figura 10, observa-se um aumento significativo
no tempo de imobilidade, quando avaliado pelo teste t de Student {t=3.823; p=
0,0014} nos animais tratados com cetamina (n=9) quando comparados com controle
(n=10).
A B
Figura 9: Efeitos observados após 3 horas da última administração de cetamina (10 mg/Kg/dia durante 3
dias, I.P) e S.S sobre o comportamento de ansiedade de ratas na fase de adolescência através da
porcentagem de entrada nos braços abertos (%EBA) e tempo de permanência nos braços abertos
(%TBA) do Labirinto em Cruz Elevado (LCE). Os resultados estão expressos como a média ± e.p.m do
(A) %EBA e (B) %TBA, durante 300 segundos, com 10 animais por grupo. *p<0,05 diferença significativa
em relação ao grupo controle (Teste t de Student).
Controle Cetamina0
1
2
3
4
**
%T
BA
Controle Cetamina0
10
20
30
40
50
***
% E
BA

40
Figura 10: Efeitos observados após 3 horas da última administração de cetamina (10 mg/Kg/dia
durante 3 dias, I.P) e S.S sobre o tempo de imobilidade no teste do nado forçado em ratas
adolescentes. Os resultados estão expressos como a média ± e.p.m do tempo de imobilidade, no
teste do nado forçado, durante 300 segundos, n= 10 animais por grupo (Teste T de Student).
4.4. Teste da Esquiva Inibitória
A Figura 11 ilustra os resultados obtidos da suspensão de cetamina sobre a
memória de curto prazo avaliada na esquiva inibitória. Os resultados obtidos indicam
prejuízo na capacidade de retenção da memória de curto-prazo, pelo grupo tratado
com cetamina [P<0,05].
Controle Cetamina0
20
40
60
80
100 **
Tem
po
Im
ob
ilid
ad
e (
seg
)
0
30
60
90
120
150
180
Controle Cetamina
Treino
Teste 1,5 h
***
**
##
Latê
ncia
(s)
Figura 11: Efeitos observados após suspensão de cetamina (10 mg/Kg/dia durante 3 dias, I.P) e S.S
sobre a retenção da memória de curto-prazo na tarefa de esquiva inibitória do tipo step-down em
ratos fêmeas adolescentes. Os resultados estão expressos com valores de mediana interquartil dos
tempos de latência para descida da plataforma nas sessões de treino (branco) e teste (cinza) (n= 10
animais por grupo). *p≤0,05 comparado à sessão de treino do mesmo grupo (teste de Wilcoxon).
#p≤0,05 comparado à sessão de treino do grupo controle (teste de Man Whitney)

41
V. DISCUSSÃO

42
No modelo experimental, foi testada a hipótese de possíveis danos
neurocomportamentais ocasionados pela suspensão de cetamina em ratos fêmeas
adolescentes. Para elucidar esses eventos, foram utilizados testes comportamentais
no campo aberto, LCE, nado forçado e esquiva inibitória do tipo Step-down. De
maior destaque, os resultados do presente estudo sugerem: 1) Efeito do tipo
ansiogênico após suspensão da droga; 2) Comportamento do tipo depressivo nos
roedores; 3) Prejuízos na memória de curto-prazo. É importante ressaltar que não
foram observadas alterações na atividade locomotora espontânea dos animais, em
ambos os parâmetros de distância total percorrida e atos de levantar.
É sabido que o mecanismo primário da cetamina dá-se pelo bloqueio dos
receptores de glutamato do tipo NMDA e consequentemente acúmulo desse
neurotransmissor na fenda sináptica (MAENGE, ZARATE, 2007). Esse
neurotransmissor é essencial para o desenvolvimento e envelhecimento cerebral,
bem como, é importante no processo de aprendizado e memória (SEGOVIA et al.
2001; STEVENS, 2008). Além disso, a hiperestimulação desses receptores, é
extremamente prejudicial ao cérebro (DONG et al. 2009; TZINGOUNIS &
WALDICHE, 2007), principalmente na adolescência (CREWS, 2000).
Nos adolescentes a alteração ocasionada pelo excesso de
neurotransmissores, envolve modificações estruturais e fisiológicas no córtex pré-
frontal, amígdala e hipocampo (CREWS, 2000; SLAWECK et al. 2004).
Desequilíbrios nessas regiões durante o processo de maturação contribuem para a
maior propensão à adição de drogas na fase da adolescência (GUERRI e
PASCUAL, 2010). Da mesma forma, o desenvolvimento relativamente tardio da
circuitaria do córtex pré-frontal envolvida no julgamento e controle inibitório pode
gerar a propensão dos adolescentes a uma maior impulsividade e fazê-lo ignorar as
consequências negativas do seu comportamento (CREWS et al. 2007; GUERRI e
PASCUAL, 2010).
A partir da análise comportamental, foi possível observar que a atividade
locomotora espontânea não foi prejudicada após administração de cetamina em
ratos fêmeas adolescentes. Esses resultados foram obtidos, através da avaliação do
número de rearing (atos de levantar) combinada à avaliação da distância total

43
percorrida no campo aberto, bem como, o número de entradas nos braços fechados
no aparato do LCE.
Estudos demonstram que administração aguda de antagonistas de receptores
de NMDA promove aumento da atividade locomotora (KEILHOFF et al. 2004;
MCLEAN e WOLLEY, 2009). Esses achados foram evidenciados com a
administração única de 100 mg/kg (CHATTERJEE et al. 2010) e 10mg/kg (ARRUDA
et al. 2008) de cetamina. Esse achado é atribuído à ação agonista da droga sobre os
receptores dopaminérgicos-D2 no estriado (IRIFUNE et al. 1991; SEEMAN et al.
2009). Nossos achados não indicam alteração na exploração espontânea, dessa
forma, sugere-se nesse modelo experimental que a retirada de cetamina não causa
alterações sustentadas e persistentes nessa região do SNC.
Com o intuito de abranger o estudo sobre os aspectos da
ansiedade/emocionalidade, foram utilizados os ensaios comportamentais no teste do
LCE. Os resultados demonstraram redução nos percentuais de entrada e tempo nos
braços abertos, que é um comportamento característico de efeito ansiogênico.
Este comportamento pode estar relacionado às modificações nas regiões
dorsal e mediana do núcleo da rafe, que consistem em duas grandes vias
ascendentes de receptores serotoninérgicos no prosencéfalo (BABAR et al. 2001).
Estas circuitarias são responsáveis pela regulação dos diferentes tipos de funções
cognitivas e comportamentos emocionais como memória e ansiedade (IVERSEN,
1984; HANDLEY e MCBLANE, 1993; GRAEFF et al. 1996; MELIK et al. 2000; TREIT
et al. 2001).
Para avaliar os possíveis efeitos depressivos nos animais foi empregado o
teste do nado forçado, que consiste num modelo tradicional para o estudo da
depressão em roedores, pois apresentam alto valor preditivo devido à resposta aos
medicamentos antidepressivos existentes (PORSOLT et al. 1978). Conforme o
objetivo do nado forçado em avaliar postura de imobilidade (OLOWABI et al. 2014),
nosso estudo demonstrou um aumento no tempo de imobilidade após retirada de
cetamina, o que caracteriza comportamento do tipo depressivo.
De acordo com Murakami et al (2000), níveis aumentado de glutamato são
descritos em regiões corticais de animais submetidos ao teste do nado forçado, após

44
uso de drogas antagonista de MNDAR. Esse fato parece se correlacionar
positivamente com o desenvolvimento do comportamento do tipo ansioso em
roedores (LI et al, 2010; TORDERA et al, 2007).
Outra hipótese para tal achado comportamental, refere-se a níveis de BDNF
diminuídos. Estudos em animais demonstraram que expressão do fator neurotrófico
derivado do cérebro (BDNF) pode sofrer desregulação por stress. Em animais,
podem ser utilizados várias formas de estímulos estressores, tais como estresse de
imobilização, choques nos pés, derrota social, privação materna precoce e
administração ou retirada brusca de drogas (LEE e KIM, 2010). Esses eventos
diminuem significativamente a expressão BDNF no hipocampo (especialmente no
giro dentado) e córtex pré-frontal (DUMAN e VOLETI, 2014; ROCERRI et al. 2002).
O BDNF é um fator neurotrófico importante, visto que é um membro da família
do fator de crescimento mais abundante cérebro (DUMAN e VOLETI, 2014). Sua
função é mediada através da sua ligação a receptores específicos, tais como o
receptor de tirosina quinase B (TrkB) entre a família do receptor quinase da
tropomiosina (Trk) (CASTRÈN e RANTAMAKI, 2009; LEE e KIM, 2010). Este
mediador desempenha um papel importante durante a adolescência (período de
desenvolvimento), na orientação, função e sobrevivência dos neurônios, mas
também quando expresso em níveis elevados no cérebro adulto desempenha um
papel importante na função neuronal, plasticidade e sobrevivência (DUMAN e
VOLETI, 2014).
Para sustentar a hipótese das alterações de BDNF aos efeitos deletérios da
depressão, estudos pós-morte de vítimas de suicídio ou doentes com transtorno
depressivo maior (TDM) foram encontrados níveis baixos de BDNF e TrkB no soro
de pacientes e em regiões do hipocampo, respectivamente (CASTRE'N et al. 2007;
CASTRE'N e RANTAMAKI, 2010; THOMPSON et al. 2011).
Outra correlação neuroanatômica interessante é a relação entre hipertrofia da
amígdala e BDNF (FRODL et al. 2003). Embora não esteja claro em pacientes
humanos, o BDNF parece estar envolvido nesta anormalidade estrutural que foi
observado em estudos pré-clínicos que apresentavam níveis elevados de BDNF na
amígdala em resposta ao estresse (YU e CHEN, 2011). Desta forma, na depressão

45
o BDNF parece estar aumentado na amígdala e nucleus accumbens e diminuído no
hipocampo e no córtex pré-frontal (DUMAN e VOLETI, 2014).
A fim de abranger o estudo sobre o aspecto do processo mnemônico, foi
utilizado o teste da esquiva inibitória do tipo step-down, que consiste em avaliar o
aprendizado e memória (IZIQUIERDO, 1998). Neste estudo, os animais expostos à
cetamina apresentaram prejuízo na memória de curta duração, que foi caracterizado
pela diminuição do tempo de latência na descida da plataforma segura do aparato.
É importante ressaltar que o uso recreacional da cetamina está associado
com alterações cognitivas e elevação de sintomas patológicos (CURRAN et al. 2000;
ABEL et al. 2003). A hipótese de que alterações nos níveis de BDNF promovem
prejuízos cognitivos induzidos após administração de cetamina repetidas
(GOULART et al. 2010).
Os mecanismos neuroquímicos associados com comprometimento da
memória de curto prazo são escassos, mas pode estar relacionada com alterações
na sinalização mediada pelo BDNF (GOULARD et al. 2010). Tem sido atribuído as
três vias de sinalização intracelular diferentes: fosfatidil-inositol 3-cinase/proteína
cinase B (PI3K/Akt), fosfolipase C (PLC) e ERK-cinase ativada por mitógeno
(ERK/MAPK).
O BDNF tem papel importante na plasticidade sináptica (MURER et al. 2001),
é evidenciado que administrações subcrônicas de cetamina diminui a expressão de
mRNAs para neurotrofinas, incluindo BDNF, no cérebro de roedores (BECKER et al.
2008), essa diminuição prejudica o reforço e estabilização das conexões sinápticas
no hipocampo. Desta forma, o BDNF exerce papel importante na formação e
manutenção (armazenamento) da memória no hipocampo e amígdala
(BEKINSCHTEIN et al. 2014).

46
VI. CONCLUSÃO

47
A partir dos ensaios comportamentais sobre os efeitos da cetamina na forma
recreativa em ratos fêmeas adolescentes, os resultados obtidos demonstram que a
exposição à droga interferiu em alguns parâmetros comportamentais tais como,
aumento do comportamento do tipo ansiogênico e depressivo, assim como, prejuízo
na memória de curta no teste da esquiva inibitória do tipo step-down. No entanto, a
retirada da cetamina após três dias de consumo não alterou a atividade locomotora
espontânea.
Dessa forma, é necessário a utilização de métodos neuroquímicos e
bioquímicos para elucidar as respostas comportamentais encontradas neste estudo.

48
VII. REFERÊNCIAS

49
ACEVEDO, M.B; NIZHNIKOV, M.E; MOLINA, J.C; PAUTASSI, R.M. Relationship
between ethanol-induced activity and anxiolysis in the open field, elevated
plus maze, light-dark box, and ethanol intake in adolescent rats.
Behavioural Brain Research. P. 203–215, 2014.
ADAMS B.W.; MOGHADDAM B. Effect of clozapine, haloperidol, or M100907 on
phencyclidine-activated glutamate efflux in the prefrontal cortex. Biol
Psychiatry, v. 50, p. 750-757, 2001.
adolescence: introduction to part I. Ann NY Acad Sci, v. 1021, p. 27-32, 2004.
AGUIAR, M.S.S. Análise do Comportamento Defensivo Induzido pela
Microinjeção do Neuropeptídeo Substância P na Matéria Cinzenta
Periaquedutal Dorsal de Ratos. Departamento de Psicologia e Educação,
Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto. Universidade de São
Paulo, 1995.
ALMEIDA, R.N. Psicofarmacologia fundamentos práticos. Guanabara, 2006.
ALVES, A.S; et al. Emprego do antiinflamatório não esteróide ketoprofeno na
analgesia preemptiva em cães. Ciência Rural, Santa Maria, v. 31, n. 3, p. 439-444,
2001.
ANDRADE, A.G; DUARTE, P.C.A; OLIVEIRA, L.G. I Levantamento Nacional sobre
Uso de álcool, tabaco e outras drogas entre universitários das 27 capitais
brasileiras. Secretaria Nacional de Políticas sobre Drogas, Brasília, 2010.
Andersen, S.L. Trajectories of brain development: point of vulnerability or
window of opportunity? Neuroscience & Biobehavioral Reviews, v.27, p.3–18,
2003.
ANNETTA, M.G; IEMMA, D; GARISTO, C; TAFANI, C; PROIETTI, R. Ketamine:
new indications for an old drug. Current Drug Targets, P. 789–794, 2005.
ARAIN, M; HAQUE, M; JOAHL, L; MATHUR, P; NEL, W; RAIS, A; SANDHU, R;
SHARMA, S. Maturation of the adolescent brain. Journal Neuropsychiatric
Disease and Treatment. P. 449-461, 2013.

50
ARGIRIADOU, H; et al. Improvement of pain treatment after major abdominal
surgery by intravenous S(+)-ketamine. Journal Anesth Analg, v. 98, p. 1413-8,
2004.
ARRUDA, M.O.V, et al. Activies of the antipsychotic drugs haloperidol and
risperidone on behavioural effects induced by ketamine in mice. Sci. Pharm, v.
76, p. 673-687, 2008.
BABAR, E; et al. Effects of ketamine on different types of anxietyrfear and
related memory in rats with lesions of the median raphe nucleus. European
Journal of Pharmacology, v. 431, p. 315–320, 2001.
Bai, X; et al. Ketamine enhances human neural stem cell proliferation and
induces neuronal apoptosis via reactive oxygen species-mediated
mitochondrial pathway. Anesth Analg, v. 116, p. 869-80, 2013.
BECKER, A. et al. Ketamine-induced changes in rat behavior: a possible animal
model of schizophrenia. Prog Neuro-Psychoph, v. 27, p. 687– 700, 2003.
BERTI, M; BACIARELLO, M; TROGLIO, R; FANELLI, G. Clinical Uses of Low–
Dose Ketamine in Patients Undergoing Surgery. Current Drug Targets, V. 10, N.
8, 2009.
BLAKEMORE, S.J. Teenage kicks: cannabis and the adolescent brains. Lancet,
v. 29, n. 12, p. 615-785, 2012.
BRASIL. Ministério da Saúde. Assistência ao Pré-natal: Normais e manuais
técnicos. 2ªed. Brasília. Ministério da Saúde. 1998.
BRASIL. Ministério de Saúde. A adolescência grávida e os serviços de saúde do
município. Brasília. Ministério da Saúde. 1999a.
BRESSAN, R.A; PILOWSKV, L.S. Hipótese glutamatérgica da esquizofrenia. Rev
Bras Psiquiatr, v. 25, p. 177-83, 2003.
BRESSAN, R.A; PILOWSKY, L.S. Hipótese glutamatérgica da esquizofrenia.
Revista Brasileira de Psiquiatria, 2003.

51
CARLINI, E.A; et al. II Levantamento domiciliar sobre o uso de drogas
psicotrópicas no Brasil: estudo envolvendo as 108 maiores cidades do país –
2005. Brasília, Secretaria Nacional Antidrogas, 2006.
CARLINI, E.A; et al. VI Levantamento Nacional sobre o Consumo de Drogas
Psicotrópicas entre Estudantes do Ensino Fundamental e Médio da Rede
Pública e Privada de Ensino nas 27 Capitais Brasileiras. SENAD - Secretaria
Nacional de Políticas sobre Drogas, Brasília, 2010.
CASRTREN, E; RANTAMAKI, T. The Role of BDNF and Its Receptors in
Depression and Antidepressant Drug Action: Reactivation of Developmental
Plasticity. Developmental Neurobiology, 2009.
CHAKRABORTY, K; NEOGI, R; BASU, D. Club drugs: review of the ‘rave’ with a
note of concern for the Indian scenario. Journal Indian Med Res, v. 6, p. 594-604,
2011.
CHATTERJEE, M; GANGULY, S; SRIVASTAVA, M; PALIT, G. Effect of ‘chronic’
versus ‘acute’ ketamine administration and its ‘withdrawal’ effect on
behavioural alterations in mice: Implications for experimental psychosis.
Behav. Brain Res, v. 216, n. 1, p. 247-254, 2010.
CHEN, W.I; et al. Gender differences in subjective discontinuation symptoms
associated with ketamine use. Substanceabusepolicy, v. 9, n. 39, p. 1-7, 2014.
CHILMONCZYK, Z. 5-HT1A receptors. their role in anxiety and depression,
mechanism of activation. Lectures, 2002.
CLEMENTS, J.A; NIMMO, W.S; GRANT, I.S. Bioavailability, pharmacokinetics,
and analgesic activity of ketamine in humans. Journal of Pharmaceutical
Sciences, P. 539–542, 1982.
CORAZZA, O; ASSI, S; SCHIFANO, F. From “Special K” to “Special M”: The
Evolution of the Recreational Use of Ketamine and Methoxetamine. CNS
Neuroscience & Therapeutics, V. 19, P. 454–460, 2013.
COSTA, J.C; TOMAZ, C. Posttraining administration of substance P and its N-
terminal fragment block the amnestic effects of diazepam. Neurobiol Learn Mem
v. 69, p. 65-70, 1998.

52
COUNOTTE, D.S; SMIT, A.B; PATTIJ, T; SPIJKER, S. Development of the
motivational system during adolescence, and its sensitivity to disruption by
nicotine. Rev Cogn Neurosci, v. 1, n. 4, p. 430-43, 2011.
CREWS, F. T.; BRAUN, C.J.; HOPLIGHT, B.; SWITZER, R. C.; KNAPP, D.J. Binge ethanol consumption causes differential brain damage in young adolescent rats compared with adult rats. Alcoholism-Clinical and Experimental Research. 24:1712–1723, 2000. CREWS, F; HE, J; HODGE, C. Adolescent cortical development: A critical period
of vulnerability for addiction. Pharmacology Biochemistry and Behavior, v. 86, n. 2,
p.189-199, 2007.
DAYAN, J; BERNARD, A; OLLIAC, B; MAILHES, A; KERMARREC, S. Adolescent
brain development, risk-taking and vulnerability to addiction. Journal of
Physiology, v. 104, n. 5, p. 279-286, 2010.
DILLON P, COPELAND J, JANSEN K. Patterns of use and harms associated with
non-medical ketamine use. Drug Alcohol Depend, v. 69, p. 23-28, 2003.
DONG C, ANAND KJS. Developmental neurotoxicity of ketamine in pediatric
clinical use. Toxicology Letters, V. 220, P.53-70, 2013.
DUMAN, R.S. Neurobiology of stress, depression, and rapid acting
antidepressants: remodeling synaptic connections. Depression and anxiety, p.
291–296, 2014.
DUMAN, R.S; VOLETI, B. Signaling pathways underlying the pathophysiology
and treatment of depression: novel mechanisms for rapid-acting agents.
Neuropsychiatric Disorders, V. 35, 2012.
DUVAL, N.G.F. Anestésicos Venosos. In: Manica J Anestesiologia Princípios e
Técnicas, 3ª ed, p. .560-97, Porto Alegre: Artmed, 2004.
E. BABAR, T. OZGUNEN, E. MELIK, S. POLAT, H. AKMAN. Effects of ketamine on
different types of anxiety. Eur. J. Pharmacol. P. 315–320, 2001.
EIDE, P.K; JORUM, E; STUBHAUG, A; BREMNES, J; BREIVIK, H. Relief of post-
herpetic neuralgia with the N-methyl-D-aspartic acid receptor antagonist
ketamine: a double-blind, cross-over comparisonwith morphine and placebo.
Pain, v. 58, p. 347–54, 1994.

53
FANTA, S; KINNUNEN, M; BACKMAN, J.T; KALSO, E. Population
pharmacokinetics of S-ketamine and norketamine in healthy volunteers after
intravenous and oral dosing. European Journal of Clinical Pharmacology, P. 441–
447, 2015.
FARBER,N.B; et al. Antiepileptic drugs and agents that inhibit voltage-gated
sodium channels prevent NMDA antagonist neurotoxicity. Mol Psychiatry, v. 7,
p. 726-733, 2002a.
FEISTEIN, E.C; RICHTER L; FOSTER, S. E. Addressing the Critical Health
Problem of Adolescent Substance Use Through Health Care, Research, and
Public Policy. Journal of Adolescent Health, v. 50, n. 5, p. 431-36, 2012.
FERREIRA FILHO, O.F; et al. Perfil sociodemográfico e de padrões de uso entre
dependentes de cocaína hospitalizados. Rev Saúde Pública, São Paulo, v. 37, n.
6, p.751-9, 2003.
FOCCHI, G.R.A. Dependência de drogas: uma abordagem para leigos. Psychiatr
on line bras, v. 9, n. 9, p, 1-15, 2004.
FREESE, T.E; MIOTTO, K; REBACK, C.J. The effects and consequences of
selected club drugs. J Subst Abuse Treat, v. 23, p. 151-156, 2002.
FRODL T, MEISENZAHL EM, ZETZSCHE T, BORN C, JAGER M, GROLL C, et al.
Larger amygdala volumes in first depressive episode as compared to recurrent
major depression and healthy control subjects. Biol Psychiatry, 53: p. 338–44,
2003.
GALEF, B. Mechanisms for the social transmission ofacquired food
preferences from adult to weanling rats. J. Comparative and Psychology, 1997.
GARCIA, L.S; et al. Acute administration of ketamine induces antidepressant-
like effects in the forced swimming test and increases BDNF levels in the rat
hippocampus. Prog Neuropsychopharmacol Biol Psychiatry, v. 32, n. 1, p. 140-4,
2008.
GOLDMAN-RACKIC, P. Prefrontal cortical dysfunction in schiophenia: the
relevance of working memory. Pysychopathology and te brain, v. 1, n. 23, 1992.

54
GOULART, B.K; LIMA, M.N.M; FARIAS, C.B, REOLON, G.K; et al. Ketamine
impairs recognition memory consolidation and prevents learning-induced
increase in hippocampal brain-derived neurotrophic factor levels. Neuroscience,
p. 969-973, 2010.
GUERREIRO, D.F; CARMO, A.L; SILVA, J.A; NAVARRO, R; GOIS, C. Um Novo
Perfil de Abuso de Substâncias em Adolescentes e Jovens Adultos. Acta Med
Port. P. 739-756, 2011.
GUERRI, C.; PASCUAL, M. Mechanisms involved in the neurotoxic, cognitive,
and neurobehavioral effects of alcohol consumption during adolescence.
Alcohol. V. 44, v. 15-26, 2010.
HANDLEY, S.L; MITHANI, S. Effects of alpha-adrenoceptor agonists and
antagonists in a maze-exploration model of “fear” – motivated behavior.
Naunyn- Schmiedeberg’s Archives of Pharmacology, v. 327, p. 1-5, 1984.
HIJAZI,Y; BOULIEU, R. Contribution of CYP3A4, CYP2B6, and CYP2C9 isoforms
to N-demethylation of ketamine in human liver microsomes. Drug Metab Dispos,
v. 30, p. 853-8, 2002.
HUANG, L; LI, Y; JIN, W; JI, X; DONG, Z. Ketamine potentiates hippocampal
neurodegeneration and persistent learning and memory impairment through
the PKCγ–ERK signaling pathway in the developing brain. Brain Research, P.
164-171, 2012.
HUSTVEIT, O; MAURSET, A; OYE, I. Interaction of the chira1 forms of ketamine
with opioid, phencyclidine, and muscarinic receptors. Pharmacol Toxicol, n. 77,
p. 35-8, 1995.
IKONOMIDOU, C; BOSCH, F; MIKSA, M; BITTIGAU, P; VOCKLER, J; DIKRANIAN,
K. Blockade of NMDA receptors and apoptotic neurodegeneration in the
developing brain. Science, v.4, p.283:70, 1999.
IKONOMIDOU, C; KAINDL, A.M. Neuronal death and oxidative stress in the
developing brain. Antioxidants & Redox Signal, v.14, p.1535–1550, 2011.

55
IMRE, g; et al. Dose-response characteristics of ketamine effect on locomotion,
cognitive function and central neuronal activity. Brain Research Bulletin, v. 69, p.
338-345, 2006.
IRIFUNE, T; SHIMIZU, M; NOMOTO, H. Ketamine-induced hyperlocomotion
associated with alteration of presynaptic components of dopamine neurons in
the nucleus accumbens of mice. Pharmacol. Biochem. Behav. P. 399–407, 1991.
IRWIN SA, IGLEWICZ A. Oral Ketamine for the Rapid Treatment of Depression
and Anxiety in Patients Receiving Hospice Care. Journal of Palliative Medicine, v.
13, n. 7, p. 903-8, 2010.
IZQUIERDO, I; MEDINA, J.H. Memory formation: the sequence of biochemical
events in the hippocampus and its connection to activity in other brain
structures. Neurobiol Learn Mem, v. 68, p. 285-316, 1997.
IZQUIERDO, L.A; BARROS, D.M; VIANNA, M.R; COITINHO, A; DEDAVID E SILVA,
T; CHOI, H; MOLETTA, B; MEDINA, J.H; IZQUIERDO, I. Molecular
pharmacological dissection of short- and long-term memory. Cell Mol Neurobiol,
v. 3, p. 269-87, 2002.
IZQUIERDO, L.A; et al. Molecular pharmacological dissection of short and long-
term memory. Cellular and molecular Neurobiology, v. 22, p. 269-287, 2000.
JAKSCH, W; et al. Perioperative small-dose S(+)-ketamine has no incremental
beneficial effects on postoperative pain when standard-practice opioid
infusions are used. Anesthesiol analg, v.94, n. 4, p. 981-6, 2002.
JANSEN K.L. A review of the nonmedical use of ketamine: Use, users and
consequences. Jounal Psychoactive Drugs, v. 32, p. 419-33, 2000.
JANSEN, K.L. Non-medical use of ketamine. BMJ, V. 2, P. 601-2, 1993.
JANSEN, K.L; DARRACOT-CANKOVIC, R. The nonmedical use of ketamine, part
two: A review of problem use and dependence. J Psychoact Drugs, v. 33, p. 151-
158, 2001.

56
JEVTOVIC-TODOROVIC, V., & CARTER, L. B. The anesthetics nitrous oxide and
ketamine are more neurotoxic to old than to young rat brain. Neurobiology of
Aging, v.26, p. 947–956, 2005.
JODO, E. et al. Activation of medial prefrontal cortex by phencyclidine is
mediated via a hippocampo prefrontal pathway. Cerebral Cortex, v. 15, p. 663–
669, 2005.
JOHNSON, S.B; ROBERT, W; BLUM, M.D; GIEDD, J.N. Adolescent Maturity and
the Brain: The Promise and Pitfalls of Neuroscience Research in Adolescent
Health Policy. J Adolesc Health. P. 216–221, 2009.
KALSI, S.S; WOOD, D.M; DARGAN, P.I. The epidemiology and patterns of acute
and chronic toxicity associated with recreational ketamine use. Journal
emerging Health Threats, v. 4, 2011.
KAPCZINSKI, F; et al. Bases biológicas dos transtornos psiquiátricos: uma
abordagem translacional. Artmed, 3 ed. Porto Alegre, 2011.
KEILHOFF, G., BECKER, A., GRECKSCH, G., WOLF, G., BERNSTEIN, H.G.
Repeated application of ketamine to rats induces changes in the hippocampal
expression of parvalbumin, neuronal nitric oxide synthase and cFOS similar to
those found in human schizophrenia. Neuroscience, V.126, P. 591-8, 2004.
KELLEY, A.E; SCHOCHET, T; LANDRY, C.F. Risk taking and novelty seeking in
KHALSA, J.H; TREISMAN, G; MCCANCE-KATZ, E; TEDALDI, E. Medical
consequences of drug abuse and co-occurring infections: research at the
national institute on drug abuse. Subst Abus, v. 29, p. 5-16, 2008.
KHARASCH, E.D; LABROO, R. Metabolism of ketamine stereoisomers by human
liver microsomes. Anesthesiology, P. 1201–1207, 1992.
KIM, J; UDO, H; LI, L.H; YOUN, T.Y; KANDEL, E.R; BAILEY, C.H. Presynaptic
activation of silent synapses and growth of new synapse contribute to
intermediate and long-term facilitation in Alysia. Neuron, v.40, n.1, p. 151-165,
2003.

57
KOOB, G.F. Neurobiological Substrates for the Dark Side of Compulsivity in
Addiction. Journal Neuropharmacology, v. 7, n. 43, 2008.
KOOB, G.F. Theoretical frameworks and mechanistic aspects of alcohol
addiction: alcohol addiction as a reward deficit disorder. Curr Top Behav
Neurosci, v.13, p. 3-30, 2013.
KOOB, G.F; et al. Drug Addiction, Dysregulation of Reward, and Allostasis. Rev
neuropsychopharmacology, v. 24, n 2, 2001.
KOS, T., NIKIFORUK, A., RAFA, D., POPIK, P. The effects of NMDA receptor
antagonists on attentional set-shifting task performance in mice.
Psychopharmacology, v. 4, p. 911–921, 2010.
KRANK, M; STEWART, S.H; O’CONNOR, R; WOICIKI, P.B; WALL, A. CONROD,
P.J. Structural, concurrent, and predictive validity of the Substance Use Risk
Profile Scale in early adolescence. Addictive Behaviors, v. 36, n. 1-2, p. 37-46,
2011.
KRYSTAL, J.H. et al. NMDA receptor antagonist effects, cortical glutamatergic
function, and schizophrenia: toward a paradigm shift in medication
development. Psychopharmacology, v. 169, p. 215–233, 2003.
LAURIETTI, G.R; et al. Clinical evaluation of hemodynamic, analgesic,
psychedelic and neuromuscular block effects of racemic ketamine versus S(+)
ketamine. Rev Bras Anestesiol, v. 56, n. 5, p. 357-62, 2000.
LEE, B.H; KIM, Y.K. The Roles of BDNF in the Pathophysiology of Major
Depression and in Antidepressant Treatment. Psychiatry Investig , p. 231-235,
2010.
LI, J.H; et al. To use or not to use: an update on licit and illicit ketamine use.
Substance Abuse and Rehabilitation, v. 2, 2011.
LISTER, R. The use of a plus-maze to measure anxiety in the mouse.
Psychopharmacology, n. 92, p. 180-185., 1987.
LOPES, G.M; et al. Use of psychoactive substances by adolescents: curret
panorama. Rev Bras psiquiatr, v. 35, p. 51-61, 2013.

58
LORRAIN, D.S. et al. Effects of ketamine and N-methyl-D-aspartate on glutamate
and dopamine release in the rat prefrontal cortex: modulation by a group II
selective metabotropic glutamate receptor agonist LY379268. Neuroscience, v.
117, p. 697–706, 2003.
LUFT, A; MENDES, F.F. S(+) Cetamina em Baixas Doses: Atualização. Rev Bras
Anestesiol, v. 55, p. 460- 9, 2005.
MACHADO-VIEIRA R, SALVADORE G, DIAZGRANADOS N, ZARATE CA.
Ketamine and the next generation of antidepressants with a rapid onset of
action. Pharmacol Ther, v. 123, n. 2, p. 143–50, 2009.
MAENG, S. Cellular mechanisms underlying the antidepressant effects of
ketamine: Role of alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid
receptors. Biol Psychiatry, v. 63, p. 349-352, 2008.
MAENG, S; ZARATE, JR, C.A. The role of glutamate in the mood disorders:
results from the ketamine in the major depression study and the presumed
cellular mechanism underlying its antidepressant effects. Current Psychiatry
Reports, v.9, p. 467-74, 2007.
MALTTA, D.C; et al. Uso de substâncias psicoativas, contexto familiar e saúde
mental em adolescentes brasileiros, Pesquisa Nacional de Saúde dos
Escolares (PeNSE 2012). Rev Bras epidemiol, p. 46-61, 2014.
MANDRYK, M. et al. Participation of adenosine system in the ketamine-induced
motor ctivity in mice. Pharmacol Rep, v. 57, p. 55-60, 2005.
MARANGONI, S.R; et al. Fatores desencadeantes do uso de drogas de abuso em
mulheres. Rev enfermagem, v. 22, p. 662-70, 2013.
MATHÉ, J.M. et al. Differential actions of dizocilpine (MK-801) on the
mesolimbic and mesocortical dopamine systems : role of neuronal activity.
Neuropharmacology, v. 38, p. 121–128, 1999.
MENINI, A. The neurobiology of olfaction. Boca Raton (FL): CRC Press, 2010.
Chapter 11. Frontiers in Neuroscience.

59
MING, G.L; SONG, H. Adult neurogenesis in mammalian central nervous
system. Annu Rev Neuroscience, v.28, p. 223-250, 2005.
MION, G; VILLEVIEILLE, T. Ketamine Pharmacology: An Update
(Pharmacodynamics and Molecular Aspects, Recent Findings). CNS
Neuroscience and Therapeutics, 2013.
MORGAN, C.J.A; CURRAN, H.V. Ketamine use: a review. Addiction, v. 107, p. 27–
38, 2011.
MORGAN, C.J; CURRAN, H.V. Acute and chronic effects of ketamine upon
human memory: A review. Psychopharmacology (Berl), v. 188, p. 408-24, 2006.
MORGAN, C.J; et al. Semantic priming after ketamine acutely in healthy
volunteers and following chronic self-administration in substance users. Biol
Psychiatry, v. 59, p. 265-72, 2006.
MURROUGH JW. Ketamine as a Novel Antidepressant: From Synapse to
Behavior. CLINICAL PHARMACOLOGY & THERAPEUTICS,v. 91, n. 2, p. 2003-12,
2012.
NIESTERS, M; MARTINI, C; DAHAN, A. Ketamine for chronic pain: risks and
benefits. British Journal of Clinical Pharmacology, v. 77, n. 2, p. 357-367, 2013.
O’DONNELL P, GRACE A.A. Phencyclidine interferes with hipocampal gating of
nucleus accumbens neuronal activity in vivo. Neuroscience, v.87, p. 823-30,
1998.
OLIVEIRA, C.M.B; et al. Cetamina e Analgesia Preemptiva. Rev Bras Anestesiol,
v. 54, p. 739-52, 2004.
Organização Mundial da Saúde. Vivendo a adolescência: Fase da vida? Faixa
etária? Construção social? Afinal, o que é adolescência? 2013. Disonível em:
<www.adolescencia.org.br/site-pt-br/adolescencia.com> Acesso em 25/08/2016.
Organização Mundial de Saúde. Classificação de transtornos mentais e de
comportamento da CID-10. Ed Artes Medicas Sul, 2004.

60
OWOLABI, R.A; AKANMU, M.A; ADEYEMI, O. I. Effects of ketamine and N-
methyl-d-aspartate on fluoxetine-induced antidepressant-related behavior
using the forced swimming test. Neuroscience Letters, P.172–176, 2014.
PANDOLFO, P; PAMPLONA, F.A; PREDIGER, R.D.S; TAKAHASHI, R.N. Increased
sensitivity of adolescent spontaneously hypertensive rats, an animal model of
attention deficit hyperactivity disorder, to the locomotor stimulation induced by
the cannabinoid receptor agonist WIN 55,212-2. European Journal of
Pharmacology, p. 141–148, 2007.
PASCUAL, A; FREITAS, C, OBERMAN, L; HORVATH, J.C; HALKO, M; ELDAIEF,
M; BASHIR, S; VERNET, M; SHAFI, M; WESTOVER, B; VAHABZADEH-HAGH,
A.M; ROTENBERG, A. Characterizing Brain Cortical Plasticity and Network
Dynamics Across the Age-Span in Health and Disease with TMS-EEG and TMS-
fMRI. Brain Topogr. V. 24, p. 302–315, 2011.
PELLOW, S; CHOPIN, P; FILE, S.E; BRILEY, M. Validation of open: closed arm
entries in an elevated plus-maze as a measure of anxiety in the rat. J Neurosci
Methods, v. 4, p. 149-67, 1985.
PICKLES, A; PICKERING, K; SIMONOFF, E; SILBERG, J; MEYER, J; MAES, H.
Genetic "Clocks" and "Soft" Events: A Twin Model for Pubertal Development
and Other Recalled Sequences of Developmental Milestones, Transitions, or
Ages at Onset. Behavior Genetics, v. 28, n. 4, 1998.
PIETRASZEK, M. Significance of dysfunctional glutamatergic transmission for
the development of psychotic symptoms. Journal Pharmacol, v. 55, p. 133-54,
2003.
PLENNINGER, E.G; DURIEX, M.E; HIMMELSEHER, S. Cognitive impairment after
small-dose Ketamine isomrs in comparison to equianagesic racemic ketamine
in human volunteers. Anesthesiol, v. 96, n. 2, p. 357-66, 2002
PORSOLT, R. D; ANTON, G; BLAVET, N; JALFRE, M. Behavioural despair in rats:
a new model sensitive to antidepressant treatments. European Journal of
Pharmacology, n. 47, p. 379-391, 1978.

61
PRATTA, E.M.M; et al. Levantamento dos motivos e dos responsáveis pelo
primeiro contato de adolescentes de Ensino Médio com substâncias
psicoativas. Rev Saúde mental álcool e drogas, v. 2, n. 2, 2006.
QUIBELL, R; PROMMER, E.E; MIHALYO, M; TWYCROSS, R; WILCOCK, A.
Ketamine*. Journal of Pain and Symptom Management, 2011.
RAEDER, J.C; STENSETH, L.B. Ketamine: a new look at an old drug. Curr opin
Anesthesiology, v. 13, p. 463-8, 2000.
RAZOUX, F; GARCIA, R; LÉNA, I. Ketamine, at a dose that disrupts motor
behavior and latent inhibition, enhances prefrontal cortex synaptic efficacy and
glutamate release in the nucleus accumbens. Neuropsychopharmacology, v. 3, p.
729-27, 2007.
REYNOLDS, E.W; BADA, H.S. Pharmacology of drugs of abuse. Obstetrics and
Gynecology Clinics of North America, v. 30, n. 3, p. 501-522, 2003.
RIBEIRO L, BUSNELLO JV, KAPCZINSKI F. Neurofisiologia dos transtornos da
ansiedade. IN: BERNIK MA. Benzodiazepínicos: quatro décadas de experiência.
São Paulo: Editora da Universidade de São Paulo, p. 45-57, 1999.
RICAURTE, G.A; MCCANN, U.D. Recognition and management of complications
of new recreational drug use. Lancet, p. 2137-45, 2005.
ROBINSON, T.E; BERRIDGE, K.C. Addiction. Annu Rev Psychol, V. 54, P. 25-53,
2003.
RODVELT, K.R; KRACKE, G.R; SCHACHTMAN, T.R; MILLER, D.K. Ketamine
induces hyperactivity in rats and hypersensitivity to nicotine in rat striatal
slices. Pharmacol Biochem Behav, v. 1, p. 71-6, 2008.
ROESLER, R; REOLON, G.K; LUFT, T; MARTINS, M.R; SCHRÖDER, N; VIANNA,
M.R; QUEVEDO, J. NMDA receptors mediate consolidation of contextual
memory in the hippocampus after context preexposure. Neurochem Res, n. 30,
v. 11, p. 1407-11, 2005.

62
RUTHERFORD, H.J; MAYES, L.C; POTENZA, M.N. Neurobiology of adolescent
substance use disorders: implications for prevention and treatment. Child
Adolesc Psychiatr Clin N Am, v. 19, n. 3, p. 479-92, 2010.
SEEMAN, H.C; GUAN, H. HIRBEC. Dopamine D2 High receptors stimulated by
phencyclidines, lysergic acid diethylamide, salvinorin A, and modafinil.
Synapse, p. 698–704, 2009.
SEMPLE, B.D; BLOMGRENB, K.C.D; GIMLINA, K; FERRIEROE, D.M; NOBLE-
HAEUSSLEINA, L.J. Brain development in rodents and humans: Identifying
benchmarks of maturation and vulnerability to injury across species. Prog
Neurobiol, P. 1–16, 2013.
SHIBUTA, S; MORITA, T; KOSAKA, J; KAMIBAYASHI, T; FUJINO, Y. Only extra-
high dose of ketamine affects l-glutamate-induced intracellular Ca2+ elevation
and neurotoxicity. Neuroscience Research, 98, 9–16, 2015.
SIGTERMANS, M.J; VAN HILTEN, J.J; BAUER, M.C.R; ARBOUS, M.S; MARINUS,
J; SARTON, E.Y; DAHAN A. Ketamine produces effective and long-term pain
relief in patients with Complex Regional Pain Syndrome Type 1. Pain, v. 145, p.
304–11, 2009.
SILVA, F.C.C, et al. Behavioral alterations and pro-oxidant effect of a single ketamine
administration to mice. Brain Research Bulletin, v. 83, p. 9–15, 2010.
SILVA, F.C.C. Alterações comportamentais e envolvimento de estresse
oxidativo provocados por diferentes tratamentos com cetamina. 2012. 128f.
Dissertação (Mestrado em Farmacologia) – Faculdade de Medicina, Universidade
Federal do Ceará, Ceará, 2012.
SILVA. J.L; et al. Uso de substâncias psicoativas “drogas”: uma revisão de
literatura Psychoactive substance use "drugs": a review. Rev Piauiense de
saúde, v.1, n. 2, p 02-08, 2013.
SILVESTRE, J.S; NADAL, R; PALLARÉS, M; FERRÉ, N. Acute effects of ketamine
in the holeboard, the elevated-plus maze, and the social interaction test in
Wistar rats. Depress Anxiety, v. 5, p. 29-33, 1997.

63
SLIKKER, W; ZOU, X; HOTCHKISS, C.E; DIVINE, R.L; SADOVOVA, N; TWADDLE,
N.C; WANG, C. Ketamine-induced neuronal cell death in the perinatal rhesus
monkey. Toxicological Sciences, v.98, p.145–158, 2007.
SMITH, K.M; et al. Club drugs: methylenodioximethamphetamine, funitrazepam,
ketamine, hydrochloride and hydroxybutyrate. Journal Health Syst Pharm, v. 59, p.
1067-76, 2002.
SPEAR, L.P. Adolescent brain development and animal models. Ann NY Acad Sci. 1021: 23 – 26, 2004. SPEAR, L.P. The adolescent brain and age-related behavioral manifestations.
Neurosci Biobehav Rev, v. 24, p. 427-63, 2000.
SQUIRE, L.R. Memory and the hippocampus: a synthesis from findings with
rats, monkeys, and humans. Psychol Rev, v. 99, n. 2, p. 195-231, 1992.
TORGALSBØEN, A.K; RUND B.R. "Full recovery" from schizophrenia in the long
term: a ten-year follow-up of eight former schizophrenic patients. Rev
Psychiatry, v. 61, p. 20-34, 1998.
TREIT, D; MENARD, J; ROYAN, C. Anxiogenic Stimuli in the Elevated Plus-
Maze. Pharmacology Biochemistry and Behavior, n. 44, p. 463-469, 1993.
TRUJILLO, K.A; et al. The Neurobehavioral Pharmacology of Ketamine:
Implications for Drug Abuse, Addiction, and Psychiatric Disorders. Journal
ILAR, v. 52, n. 3, 2011.
TRUJILLO, K.A; ZAMORA, J.J; WARMOTH, K.P. Increased Response to
Ketamine Following Treatment at Long Intervals: Implications for Intermittent
Use. J. Biopsych, v. 53, p. 178-83, 2008.
UNODC, United Nations Office on Drugs and Crime. World drug report. Vienna:
2014.
UNODC. United Nations office on drugs and crime: World drug report. Vienna: United
Nations Publications; 2010.
VASCONCELOS, S.M.M; et al. Cetamina: aspectos gerais e relação com a
esquizofrenia. Rev Psiq Clín, V. 10, N. 6, P. 32-10, 2005.

64
VERGE D., DAVAL G., PATEY A., GOZLAN H., EL MESTIKAWY S., HAMON M.
Presynaptic 5-HT autoreceptors on serotonergic cell bodies and/or dendrites
but not terminals are of the 5-HT. Eur. J. Pharmacol, 1985.
VERMA, A; MOGHADDAM, B. NMDA receptor antagonist impair prefrontal
cortex function as assessed via spatial delayed alternation performance in
rats: Modulation by dopamine. J. Neurosci, V. 16, P. 373-9, 1996.
WANG, C. Ketamine-induced neuronal cell death in the perinatal rhesus
monkey. Toxicological Sciences, 98(1), 145–158, 2006.
WANG, H; RAMAKRISHNAN, A; FLETCHER, S; PROCHOWNIK, E.V; GENETICS,
M. HHS Public Access, v.2, p. 145–155, 2005.
WANG, J.H; FU, Y; WILSON, F.A.W; MA,Y.Y. Ketamine affects memory
consolidation: Differential effects in T-maze and passive avoidance paradigms
in mice. Neuroscience, v.140, p.993–1002, 2006.
WHO, Expert Committee on Drug Dependence, thirty-four report. Geneva. 2006.
WILLS, T.A; VACCARO, D; MCNAMARA, G. Novelty seeking, risk taking, and
related constructs as predictors of adolescent substance use: an application of
Cloninger's theory. J Subst Abuse. P.1–20, 1994.
WITT, E.D. Research on alcohol and adolescent brain development:
opportunities and future directions. Alcohol, v. 44, n. 1, p. 119-124, 2010.
YANAGIHARA, Y; KARIYA, S; OHTANI, M; UCHINO, K; AOYAMA, T; YAMAMURA,
Y; IGA, T. Involvement of CYP2B6 in N-demethylation of ketamine in human
liver microsomes. Drug Metabolism and Disposition, 29(6), 887–890, 2001.
YU, H; CHEN, Z. The role of BDNF in depression on the basis of its location in
the neural circuitry. Acta Pharmacologica Sinica, 32: p. 3–11, 2011.
ZACNY, J.P., GALINKIN, J.L. Psychotropic drugs used in anesthesia. Practice,
abuse, liability and epidemiology of abuse. Anesthesiology, V. 90, p. 269-88,
1999.
ZHAO, W; CAVALLARO, S; GUSEV, P; ALKON, D.L. Nonreceptor tyrosine protein
kinase pp60c-src in spatial learming: synapse-specific changes in its gene

65
expression, tyrosine phosphorylation, and protein-protein interaction.
Proccedings of the National Academy of science of the United States of America, n.
14, v.97, p. 8098-8103, 2000.
ZOU, X. A; TUCKER, A; PATTERSON, A; REBECCA, L; DIVINE, B; NATALYA S.B;
XUAN, Z.A; JOSEPH, P; HANIG, C; MERLE, G; PAULE, A; WILLIAM, S; CHENG,
W. Prolonged exposure to ketamine increases neurodegeneration in the
developing monkey brain. Int. J. Devl Neuroscience, v.27, p. 727–731, 2009.

66
VIII. ANEXOS

67





























