Embed Size (px)
Citation preview

Universidad CEU Cardenal Herrera Departamento de Medicina y Cirugía Animal
Actualización de los sistemas de inducción de celos y ovulaciones en ovino
TESIS DOCTORAL
Presentada por:
Paula Martínez Ros
Dirigida por:
Dra. Susana Astiz Blanco
Dra. Empar García Roselló
Dr. Antonio González de Bulnes López
VALENCIA
2017


LosdoctoresSusanaAstizBlancoyAntonioGonzálezdeBulnesLópez, investigadores
del InstitutoNacional de Investigación y Tecnología Agraria y Alimentaria (INIA) y la
doctora Empar García Roselló, profesora del Departamento de Medicina y Cirugía
AnimaldelaFacultaddeVeterinariadelaUniversidadCEUCardenalHerrera
CERTIFICAN
Quelapresentememoriatitulada“Actualizacióndelossistemasdedeteccióndecelosy
ovulaciones en ovino”, ha sido realizada por Dña. Paula Martínez Ros, licenciada en
Veterinariapor laUniversidadCEUCardenalHerrera,bajosudirecciónyconstituye la
Memoria de Tesis para optar al título de Doctor por la Universidad CEU Cardenal
Herrera.
Yparaqueasíconstealosefectosoportunos,firmanelpresentecertificadoenMoncada,
a8demarzode2017.
SusanaAstizBlancoEmparGarcíaRosellóAntonioGonzálezdeBulnesLópez


“Nosirvelaciencia
sinoleacompañaelpoder”
DeLasmilyunanoches(Anónimo)


Amisovejas,quetantomeenseñaron.
Amipadre,quesupotransmitirme
lapasiónquesientoporestanobleespecie.
AJavi,
lasuertedemivida.


AGRADECIMIENTOS


Agradecimientos
11
AGRADECIMIENTOSQuiero manifestar mi más sincero agradecimiento a cuantas personas me hanayudadoaculminarlaelaboracióndelapresentetesis,sincuyaparticipaciónellonohubierasidoposible.Sontantas,queesposiblequemeolvidedealguien,siesasí,pidodisculpasdeantemano.
Amisdirectores,Dra.SusanaAstizBlanco,Dra.EmparGarcíaRosellóyDr.AntonioGonzálezdeBulnesLópez,porenseñarme,orientarme,ayudarmeyconfiarenmí.Graciaspor vuestro tiempo,dedicacióny sobre todo, por la inagotablepacienciaquemehabéisdispensadoalolargodeestosaños, inclusoenlosmomentosmásdifíciles,graciasporesperarme.Ydeunaformamuyespecial,Susana,graciasportu implicación,queha idomásalláde loprofesional.Graciaspor tuconstanciaysaber hacer, por tu apoyo y tu ánimo constante y, sobre todo, por tu gran valíaprofesionalypersonal.
Amispadres,porquetodocuantosoyoslodebo.Nopodemosescogerquiénnostraeaestemundo,perosiagradecer la inmensasuertedetenerunospadrestanmaravillososcomovosotros.Graciasportodoelesfuerzo,elapoyoyelcariñoquehabéisvolcadoenmíyenmieducación.Mamá,graciasporhabersidoparamíunejemplodeesfuerzo, luchayconstancia.Yatipapá,graciasportodalasabiduríaque me has transmitido y por compartir conmigo mis ilusiones, porque sin tuayudanolohubieselogrado.Acostumbrarmeanotenertecadadíaamiladoeslomásdifícilquehehechoenmivida,graciasporentenderqueeranecesariohacerlopara poder seguir mi camino. De no haberlo hecho esta tesis hoy no sería unarealidad.Graciasportodoloquehemoscompartidojuntos.
Al restodemi familia, Rosi, Pepe, Carmen, Sergio,Ruti, Gema,Roger, Tavi,MarcBernad, Júlia,Vio,Sonia,MarcBoixymi teteÓscar.Simplemente,graciasporserpartedemivida,especialmenteaRuti,porquenopodríaentendertodolovividosinellaamilado.
APaco,aPepeyamibisa,porhabersidoejemployfuentedeinspiraciónparamí.Porque sin estar ami lado nunca estuvisteis ausentes. Espero que, desde dondeestéis,podáissentirosorgullososdemí.Nuncadejéisdecuidarme.
AMario y Tere, gracias por entender la pasión que siento pormi trabajo y quedebidoaeso,nopuedacompartirconvosotrostodoeltiempoquemegustaría.
ANoniy Jaime,por compartir conmigo la ilusiónpormisproyectos.Graciasporalegrarospormiséxitosycompartirlosconmigo,perosobretodo,graciasporestarahíenlosmalosmomentos,queenestosúltimostresañosnohansidopocos.

Agradecimientos
12
ADarío y Paula, por entender que cuandohay que terminar una tesis, se debenaparcar los esquís, y además, saber hacerlo con una sonrisa. Gracias chicos porconvertir unos días de duro trabajo en unas mini vacaciones inolvidables, osquieromucho.Frank,atígraciasportupacienciaytuhospitalidad.
Un agradecimiento muy especial a los becarios de la Granja Docente y deInvestigación Veterinaria de la Universidad CEU Cardenal Herrera Marc, Eva y,muy especialmente, a Alejandro, el niño de mis ojos. También a mis queridosalumnosBorja,Pepe,Luis,Carla,Helena,Quique,…graciasporvuestraimplicaciónduranteaquellaslargasnochesdetomademuestrasyvuestraayudacuandooshenecesitado. Sois un gran equipo con una extraordinaria calidad humana yprofesional.Mismássincerosagradecimientosporvuestroapoyofísicoymoral.
AlprofesorCarlosBuxadé,porayudarmeaentenderque,pormuchoqueduela,enla vida es necesario cerrar algunas puertas, de lo contrario, no se pueden abrirotras.Graciasportusánimosytumotivaciónenlarealizacióndeestatesis.Graciasamigoportusconsejos,quetantomeayudansiempreenelámbitoprofesionalyenelpersonal.
Al profesor Adolfo Rodríguez Montesinos, por tirar de mí cada vez que me hevenidoabajoyayudarmeaverlaluzalfinaldeltúnel,porqueresultaquesíhabíaluzalfinaldeltúnel.Graciasporhabermantenidolaconstanciaylapacienciacadavezqueyolaperdía.
A los compañerosde la FacultaddeVeterinaria y de laUniversidadquemehanprestadosuapoyoysuayudacuandoloshenecesitado.Emilio,Meri,Jordi,ytantosotros de los que no me olvido, gracias por vuestros ánimos y empuje. Mónica,gracias de todo corazón por tus consejos y tu ayuda. Y muy especialmente aEstrella,graciasportuapoyomoral,porayudarmeaarrancarconlasgráficas,perosobretodo,graciasporestarahísiemprequetehenecesitado.
A los compañerosde la FacultaddeVeterinaria y de laUniversidadquemehanprestadosuapoyoysuayuda,graciasporvuestrosánimosyempujecuandoloshenecesitado.Emilio,Meri,Jordi,ytantosotros,graciasporlosánimos.aunquenoosnombreatodos,nomeolvidodevosotros.Graciasporestarahísiemprequeoshenecesitado.
A Carla y Laura, gracias por acogerme en vuestra casa siempre que lo henecesitado, durante mi estancia en el INIA y en mis posteriores visitas. Y a lapequeñaOliviaporhacermásfelicesmisestanciasenMadrid.
ATeresaSegarra,graciasporacompañarmedurantetodoestetiempo,contodoloqueellohasupuesto.

Agradecimientos
13
AGemi,portodoloquemehasenseñadodeovejas,porpodersentirtetancercacadadíaestandotanlejosyaunenladistanciadarmefuerzasparaseguiradelanteencadaunodemispasos,yporquesiempreconsiguesarrancarmeunasonrisaenlos peores momentos, en los que nadie más en el mundo es capaz de hacerlo.Graciaspornohabermefalladonunca.
Porúltimo,peronoporellomenosimportante,aJavi.Noencuentropalabrasparaexpresarelregaloqueerescadadíaparamí,asíque,simelopermites,lastomaréprestadasdelgranBarryWhite:“Youaremyfirst,mylast,myeverything”.Graciasporhabertecruzadoenmicamino.
…atodoscuantosmeapoyaronyalentaron,GRACIAS.


ÍNDICE


Indice
17
ÍNDICE1 Revisiónbibliográfica.............................................................................................43
1.1 Laimportanciadelganadoovino................................................................................43
1.2 Elganadoovinoenelmundo.........................................................................................44
1.3 ElganadoovinoenlaUniónEuropea.......................................................................46
1.4 ElganadoovinoenEspaña............................................................................................48
1.5 Descripcióndelasrazasdeinterésparaelpresentetrabajo..........................51
1.6 Fisiologíadelareproducción.Cicloestralycontrolneurodendocrino......53
1.6.1 Fasesdelcicloestral.................................................................................................................56
1.6.2 Foliculogénesis...........................................................................................................................57
1.6.3 Dinámicafolicularenlaoveja..............................................................................................60
1.6.4 Ovulación,crecimientoydesarrollodelcuerpolúteo...............................................61
1.6.5 Ciclosexualyestacionalidadenlaoveja.........................................................................63
1.7 Manejoycontroldelareproducción..........................................................................66
1.7.1 Progesterona,susanálogos(progestágenos)yeCG...................................................68
1.7.2 ProstaglandinaF2αysusanálogos...................................................................................71
1.7.3 Protocoloshormonalesmásutilizadosenprogramasdemontanatural.........72
2 Justificaciónyobjetivos..........................................................................................77
3 MaterialyMétodos...................................................................................................83
3.1 Animales:baseracial........................................................................................................83
3.2 Entornoexperimental:instalacionesymanejo.....................................................83
3.3 Diseñoexperimental..........................................................................................................85
FaseExperimental1.Efectodeladuracióndeltratamientodesincronizacióndeceloscondispositivosintravaginalesliberadoresdeprogestágenoseinfluenciadelaépocareproductivaydelarazaovina...........................................................................................................85
FaseExperimental2.Efectodeladuracióndeltratamientodesincronizacióndeceloscondispositivos intravaginales liberadoresdeprogestágenossobre la funcionalidadhipofisariayováricaenovejas,enfotoperiododecreciente..................................................90
Fase Experimental 3. Efecto del tipo de dispositivo progestativo intravaginal(progesterona natural o CIDR versus progestágeno sintético o FGA) sobre el

Indice
18
porcentaje de gestación en ovejas tratadas con un protocolo de sincronización deceloslargo(14días)enfotoperiododecreciente........................................................................93
Fase Experimental 4. Efecto del tipo de dispositivo progestativo intravaginal(progesterona natural o CIDR versus progestágeno sintético o FGA) sobre lafuncionalidad hipofisaria y ovárica en ovejas tratadas con un protocolo desincronizacióndeceloscorto(7días)einfluenciadelaépocareproductiva................95
3.4 Análisisestadístico.............................................................................................................97
4 Resultados................................................................................................................101
4.1 Fase experimental 1. Efecto de la duración del tratamiento desincronización de celos con dispositivos intravaginales liberadores de progestágenos einfluenciadelaépocareproductivaydelarazaovina....................................................................101
4.1.1 Experimento 1.1 Influencia del tipo de tratamiento sobre el porcentaje degestaciónduranteelfotoperiododecreciente...........................................................................102
4.1.2 Experimento 1.2 Influencia del tipo de tratamiento sobre el porcentaje degestaciónduranteelfotoperiodocreciente................................................................................105
4.1.3 Influenciadelfotoperiodosobrelastasasdegestaciónyprolificidad...........108
4.1.4 Influenciadelarazasobreelporcentajedegestaciónyprolificidad..............110
4.2 Fase experimental 2. Efecto de la duración del tratamiento desincronizacióndeceloscondispositivosintravaginalesliberadoresdeprogestágenossobrelafuncionalidadhipofisariayováricaenovejas,enfotoperiododecreciente........................111
4.2.1 Influenciadeltipodetratamientosobreelporcentajedepresentacióndeceloymomentodeaparicióndelcelo.....................................................................................................112
4.2.2 Influenciadeltipodetratamientosobreladinámicadecrecimientofolicular 115
4.2.3 Influenciadeltipodetratamientosobrelaliberacióndeestradiol.................119
4.2.4 InfluenciadeltipodetratamientosobreelmomentodeaparicióndelpicodeLHysobrelaconcentraciónmáximadeLH................................................................................121
4.2.5 Influenciadeltipodetratamientosobrelatasadeovulaciónyfuncionalidaddecuerposlúteos....................................................................................................................................123
4.3 Fase experimental 3. Efecto del tipo de dispositivo progestativointravaginal(progesteronanaturaloCIDRversusprogestágenosintéticooFGA)sobreelporcentaje de gestación en ovejas tratadas con un protocolo de sincronización de celoslargo(14días)enfotoperiodocreciente.................................................................................................124
4.4 Fase experimental 4. Efecto del tipo de dispositivo progestativointravaginal(progesteronanaturaloCIDRversusprogestágenosintéticooFGA)sobrela

Indice
19
funcionalidadhipofisariayováricaenovejastratadasconunprotocolodesincronizacióndeceloscorto(7días)einfluenciadelaépocareproductiva........................................................126
4.4.1 Experimento4.1Fotoperiodocreciente.......................................................................126
Influenciadeltipodetratamientosobreelporcentajedepresentacióndecelo,momentodeaparicióndelcelouagrupacióndeloscelos...........................................................................126
Influenciadeltipodetratamientosobreel...................................................................................129
Influenciadeltipodetratamientosobre........................................................................................130
Influenciadeltipodetratamientosobrela...................................................................................131
4.4.2 Experimento4.2Fotoperiododecreciente..................................................................132
5 Discusión..................................................................................................................139
5.1 Efecto de la duración del tratamiento de sincronización de celos condispositivos intravaginales liberadores de progestágenos e influencia de la épocareproductivaydelarazaovina...................................................................................................................139
5.2 Efecto de la duración del tratamiento de sincronización de celos condispositivos intravaginales liberadores de progestágenos sobre la funcionalidadhipofisariayováricaenovejas,enfotoperiododecreciente...........................................................146
5.3 Efecto del tipo de dispositivo progestativo intravaginal (progesteronanaturaloCIDRversusprogestágenosintéticooFGA)sobreelporcentajedegestaciónenovejastratadasconunprotocolodesincronizacióndeceloslargo(14días)enfotoperiodocreciente 155
5.4 Efecto del tipo de dispositivo progestativo intravaginal (progesteronanaturaloCIDRversusprogestágenosintéticooFGA)sobrelafuncionalidadhipofisariayovárica en ovejas tratadas con un protocolo de sincronización de celos corto (7 días) einfluenciadelaépocareproductiva..........................................................................................................160
6 Conclusiones............................................................................................................167
7 Bibliografía..............................................................................................................171


Indice
21
ÍNDICEDEFIGURASFigura1.Distribucióncontinentaldelcensoovino(FAOSTAT,2015).................................................................44Figura 2. Distribución, en Europa de las explotaciones y censo de distintos rumiantes entre zonasclasificadasonocomodesfavorecidas(RICA,2003).....................................................................................................46Figura3.DistribucióndelcensodeovinoporpaísesenlaUE-28enel2015(EUROSTAT,2016)..........47Figura4.DistribucióndelcensodeovinoenEspañaporComunidadesAutónomas(SITRAN,2016)..49Figura5.EvolucióndelcensodeovinoenEspañaentrelosaños2006y2015(MAGRAMA,2016).......50Figura6.EvolucióndelnúmerodeexplotacionesdeovinoenEspañaentreelaño2007yelaño2016(SITRAN,2016)..............................................................................................................................................................................50Figura 7. Esquema de activación/inhibición de la ciclicidad en la oveja mediante la melatonina.Fuente:Pathwaystopregnancyandparturation.Senger,P.L.2005...................................................................64Figura8.Protocolosdesincronizacióndeceloenbaseadispositivosintravaginalesdeprogestágenosde duración “clásica” (grupoControl) o “corta” (gruposPG-Corto y Corto-PG), utilizados en la FaseExperimental1...............................................................................................................................................................................86Figura9.Distribuciónracialdelasovejasporgruposdetratamiento(Grupocontrol:tratamiento14días + eCG a la retirada, Grupo PG-Corto: tratamiento 7 días + PGF2α al inicio + eCG a la retirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)...........................................................................88Figura10.Distribuciónracialdelasovejasporgruposdetratamiento(Grupocontrol:tratamiento14días + eCG a la retirada, Grupo PG-Corto: tratamiento 7 días + PGF2α al inicio + eCG a la retirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)...........................................................................89Figura11.ProtocolosdesincronizacióndeceloenbaseadispositivosintravaginalesdeprogestágenosGrupo FGA o progesterona Grupo CIDR, utilizados en la Fase Experimental 3. Protocolo desincronizaciónlargo,de14días.............................................................................................................................................95Figura 12. Protocolos de sincronización de celo en base a dispositivos intravaginales deprogestágenosGrupoFGAoprogesteronaGrupoCIDR,utilizadosenlaFaseExperimental4.Protocolocortodesincronizaciónde7días...........................................................................................................................................96Figura 13. Porcentaje de ovejas gestantes y no gestantes según grupo experimental durante elfotoperiodo decreciente (Grupo control: tratamiento 14 días + eCG a la retirada, Grupo PG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada).......................................................................................................................................................................103Figura 14. Porcentaje de ovejas que paren 1, 2, 3, 4 ó 5 corderos por parto dependiendo de laduración del tratamiento administrado en fotoperiodo decreciente (Grupo control: tratamiento 14días + eCG a la retirada, Grupo PG-Corto: tratamiento 7 días + PGF2α al inicio + eCG a la retirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)........................................................................104Figura15.Prolificidadmediayerrorestándarenlosdistintosgruposdetratamientoadministradosenfotoperiododecreciente(Grupocontrol:tratamiento14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada).......................................................................................................................................................................104Figura 16. Porcentaje de ovejas gestantes y no gestantes dependiendo del tipo de tratamientoadministradoenfotoperiodocreciente(Grupocontrol:tratamiento14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)...................................................................................................................................................105Figura 17. Porcentaje de ovejas que paren 1, 2, 3, 4 ó 5 corderos por parto dependiendo de laduracióndeltratamientoadministradoenfotoperiodocreciente(Grupocontrol:tratamiento14días+eCGa laretirada,GrupoPG-Corto: tratamiento7días+PGF2αal inicio+eCGa laretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)......................................................................................106Figura18.Prolificidadmediayerrorestándarenlosdistintosgruposdetratamientoadministradosen fotoperiodo creciente (Grupo control: tratamiento 14 días + eCG a la retirada, Grupo PG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada).......................................................................................................................................................................107

Indice
22
Figura19.Porcentajedeovejasgestantesynogestantesdependiendodeladuracióndeltratamientoadministradoenfotoperiodocreciente(GrupoLargo:tratamiento14días+eCGalaretirada,GrupodetratamientosCortos:tratamiento7días+PGF2αaliniciooalaretirada+eCGalaretirada).......107Figura 20. Porcentaje de ovejas que parieron 1, 2, 3, 4 ó 5 corderos por parto dependiendo de laduracióndeltratamientoadministradoenfotoperiodocreciente(GrupoLargo:tratamiento14días+eCGalaretirada,GrupodetratamientosCortos:tratamiento7días+PGF2αaliniciooalaretirada+eCGalaretirada).......................................................................................................................................................................108Figura 21. Efecto del fotoperiodo (creciente vs. decreciente) sobre el porcentaje de gestación enovejas sincronizadas con diferentes protocolos de sincronización basados en la inserción dedispositivosintravaginalesliberadoresdeprogestágenos......................................................................................109Figura 22. Efecto del fotoperiodo (creciente vs. decreciente) sobre la prolificidadmedia en ovejassincronizadas con diferentes protocolos de sincronización basados en la inserción de dispositivosintravaginalesliberadoresdeprogestágenos...............................................................................................................109Figura23.Porcentajedehembrasgestantesynogestantessegúnraza(lacaune, segureñaoF1delcrucedeéstasconromanov)sincronizadascondiferentesprotocolosdesincronizaciónbasadosenlainsercióndedispositivosintravaginalesliberadoresdeprogestágenos............................................................110Figura24.Porcentajedeovejascon1,2,3,4o5corderosporpartosegúnrazas(lacaune,segureñaoF1delcrucedeéstasconromanov)sincronizadascondiferentesprotocolosdesincronizaciónbasadosenlainsercióndedispositivosintravaginalesliberadoresdeprogestágenos................................................111Figura 25. Porcentaje de ovejas de cada grupo que mostraron celo en las horas posteriores a larestiradade lasesponjas. (Grupocontrol: tratamiento“largo”de14días+eCGa laretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)...................................................................................................................................................114Figura 26. Porcentaje de ovejas de cada grupo que salen en celo en las horas posteriores a larestiradade lasesponjas. (Grupocontrol: tratamiento“largo”de14días+eCGa laretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)...................................................................................................................................................115Figura27.Númeromediodefolículos≥a4mm,divididosenmedianos(4-5mm;barraroja)ygrandes(≥6mm; barra azul), durante la fase folicular de ovejas pertenecientes a tres grupos distintosexperimentales (Grupocontrol: tratamiento “largo”de14días+eCGa laretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)......................................................................................................................................................................116Figura 28.Diámetromediodelmayor folículo en tamaño (F1; líneaazul), del segundomayor (F2;línearoja)ydelrestodefolículos(resto;líneaverde)durantelafasefoliculardeovejaspertenecientesa tres grupos distintos de tratamiento (Grupo control: tratamiento “largo” de 14 días + eCG a laretirada,GrupoPG-Corto: tratamiento7días+PGF2αal inicio+eCGa la retirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)...........................................................................................................117Figura 29. Número medio de folículos mayores o iguales a 4mm, divididos en medianos (4-5mm;barra roja) y grandes (≥6mm;barraazul), durante la fase folicular de ovejas pertenecientes a tresgruposdistintosde tratamiento (Grupocontrol: tratamiento “largo”de14días+eCGa la retirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)......................................................................................................................................119Figura30.Nivelesdeestradiolmedios(pg/ml)encadagrupodetratamientoenlashorasposterioresalaretiradadelasesponjas.(Grupocontrol:tratamiento“largo”de14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada).a,bindicandiferenciassignificativasentregruposencadamomentodemuestreo,conP<0,001.............................................................................................................................................................120Figura31.Porcentajeabsoluto(superior)yacumulado(inferior)deovejasquepresentarondescargaprevulatoriadeLH (ng/ml) en cadagrupoexperimental en lashorasposterioresa laaparicióndelcelo(Grupocontrol:tratamiento“largo”de14días+eCGalaretirada,GrupoPG-Corto:tratamiento7

Indice
23
días + PGF2α al inicio + eCG a la retirada, Grupo Corto-PG: tratamiento 7 días + PGF2αy eCG a laretirada).........................................................................................................................................................................................122Figura 32. Número de CL y niveles medios de progesterona (ng/ml) en los distintos grupos detratamiento11díasdespuésdelaretiradadelasesponjas.(Grupocontrol:tratamiento“largo”de14días + eCG a la retirada, Grupo PG-Corto: tratamiento 7 días + PGF2α al inicio + eCG a la retirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)........................................................................123Figura33.Tiempomedio(enhoras)transcurridoentrelaretiradadelosdispositivosintravaginalesyelmomento en que salieron en celo las ovejas pertenecientes a los distintos grupos de tratamiento(grupo FGA, grupo CIDR), con los máximos y mínimos observados en cada grupo. Grupo FGA:tratamientode7díasconesponjasdeFGA+eCG+PGF2αalaretirada,GrupoCIDR:tratamientode7díascondispositivodeprogesterona+eCG+PGF2αalaretirada).......................................................................127Figura 34. Porcentaje de ovejas de cada grupo que presentaron celo en las horas posteriores a larestiradadelosdispositivosintravaginales(GrupoFGA:tratamientode7díasconesponjasdeFGA+eCG+PGF2αalaretirada,GrupoCIDR:tratamientode7díascondispositivodeprogesterona+eCG+PGF2αalaretirada)...................................................................................................................................................................128Figura 35. Porcentaje de ovejas de cada grupo que presentaron celo en las horas posteriores a laretiradade losdispositivos intravaginales (GrupoFGA: tratamientode7días conesponjasdeFGA+eCG+PGF2αalaretirada,GrupoCIDR:tratamientode7díascondispositivodeprogesterona+eCG+PGF2αalaretirada)...................................................................................................................................................................129Figura 36. Porcentaje absoluto (superior) y acumulativo (inferior) de ovejas que presentarondescarga provulatoria de LH (ng/ml) en cada grupo de tratamiento en las horas posteriores a laaparicióndelcelo(GrupoFGA:tratamientode7díasconesponjasdeFGA+eCG+PGF2αalaretirada,GrupoCIDR:tratamientode7díascondispositivodeprogesterona+eCG+PGF2αalaretirada).......130Figura37.Nivelesdeestradiolmedios(pg/ml)encadagrupodetratamientoenlashorasposterioresa la retirada de los dispositivos intravaginales (Grupo FGA: tratamiento de 7 días con esponjas deFGA+eCG+PGF2αalaretirada,GrupoCIDR:tratamientode7díascondispositivodeprogesterona+eCG+PGF2αalaretirada);aybindicandiferenciassignificativasentregruposencadamomentodemuestreo,conP<0,0001..........................................................................................................................................................131Figura 38. Número de CL y niveles medios de progesterona (ng/ml) en los distintos grupos detratamiento11díasdespuésdelaretiradadelosdispositivosintravaginales.(GrupoFGA:tratamientode 7 días con esponjas de FGA+ eCG + PGF2α a la retirada, Grupo CIDR: tratamiento de 7 días condispositivodeprogesterona+eCG+PGF2αalaretirada);aybindicandiferenciassignificativasentrelosdosgrupos(P<0,0001)......................................................................................................................................................132Figura 39. Tiempo medio transcurrido entre la retirada de los dispositivos intravaginales y elmomento de presentación de celo en ovejas pertenecientes a los distintos grupos experimentales(grupo FGA, grupo CIDR), con los máximos y mínimos observados en cada grupo (Grupo FGA:tratamientode7díasconesponjasdeFGA+eCG+PGF2αalaretirada,GrupoCIDR:tratamientode7díascondispositivodeprogesterona+eCG+PGF2αalaretirada).......................................................................133Figura40.Porcentajedeovejasdecadagrupoquemostróceloenlashorasposterioresalaretiradadelosdispositivosintravaginales(GrupoFGA:tratamientode7díasconesponjasdeFGA+eCG+PGF2αa la retirada,GrupoCIDR: tratamientode7díascondispositivodeprogesterona+eCG+PGF2αa laretirada).........................................................................................................................................................................................134Figura41.Porcentajedeovejasdecadagrupoquemostróceloenlashorasposterioresalaretiradadelosdispositivosintravaginales(GrupoFGA:tratamientode7díasconesponjasdeFGA+eCG+PGF2αa la retirada,GrupoCIDR: tratamientode7díascondispositivodeprogesterona+eCG+PGF2αa laretirada); a y b indican diferencias significativas entre los dos grupos con P=0,002; c y d indicandiferenciassignificativasentrelosdosgruposconP=0,019...................................................................................135


Indice
25
ÍNDICEDETABLASTabla1.Distribuciónracialdelasovejasporgruposdetratamiento(Grupocontrol:tratamiento14días + eCG a la retirada, Grupo PG-Corto: tratamiento 7 días + PGF2α al inicio + eCG a la retirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)...........................................................................87Tabla2.Distribuciónracialdelasovejasporgruposdetratamiento(Grupocontrol:tratamiento14días + eCG a la retirada, Grupo PG-Corto: tratamiento 7 días + PGF2α al inicio + eCG a la retirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)...........................................................................89Tabla 3. Número y porcentaje de animales por raza que se incluyeron en el estudio de distintostratamientos de sincronización del celo basados en el uso de dispositivos intravaginales deprogestágenosenfotoperiododecreciente.....................................................................................................................101Tabla 4. Número y porcentaje de animales por raza que se incluyeron en el estudio de distintostratamientos de sincronización del celo basados en el uso de dispositivos intravaginales deprogestágenosenfotoperiodocreciente..........................................................................................................................102Tabla5.Porcentajedepresentacióndecelosegúngrupoexperimental.........................................................112Tabla6.Tiempomediotranscurrido(h)entrelaretiradadelasesponjasintravaginalesyelmomentoenquesalieronencelolasovejaspertenecientesalosdistintosgruposexperimentales..........................113Tabla7.MediaydesviacióntípicadelmomentodeaparicióndelpicodeLH,enhoras,yconcentraciónmáximadeLH,enng/ml,enlasovejaspertenecientesalosdistintosgruposexperimentales..............121Tabla8.NúmerodeanimalesderazaLacaunedeaptitudlecheraqueseincluyeronencadagrupodetratamiento,valoresmediosdelaedad,condicióncorporalynúmerodelactaciones..............................125Tabla9.Porcentajedeovejasgestantesyprolificidadmediaencadagrupoexperimentaldependiendodeltipodetratamientoadministrado..............................................................................................................................125Tabla 10. Duración del anestro en distintas razas ovinas pertenecientes a países de la Cuenca delMediterráneo...............................................................................................................................................................................141


Indice
27
ÍNDICEDEABREVIATURAS°C gradoscentígrados
°N gradoslatitudnorte
AUC áreabajolacurva
BMP-15 proteínamorfogenéticaósea15
CC condicióncorporal
CC.AA. ComunidadesAutónomas
CIDR dispositivointernodeliberacióncontrolada
CL cuerpolúteo
cord cordero
d días
E2 estradiol
eCG gonadotropinacoriónicaequina
EGF factordecrecimientoepidermal
ELISA ensayoporinmunoadsorciónligadoaenzimas
F1 híbridodeprimerageneración
F1 tamañodelfolículomásgrande
F2 tamañodelsegundofolículomásgrande
Fresto tamañomediodelrestodelosfolículos
FAO OrganizacióndelasNacionesUnidasparalaAgriculturayla Alimentación
FAOSTAT basededatosestadísticosdelaOrganizacióndelasNacionesUnidas paralaAgriculturaylaAlimentación
FEAGAS FederaciónEspañoladeAsociacionesdeGanadoSelecto
FGA acetatodeflugestona
FGF factordecrecimientofibroblástico
FSH hormonafolículoestimulante
GnRH hormonaliberadoradegonadotropinas
GRA folículosgrandes
GRACREC folículosgrandesencrecimiento
hCG gonadotropinacoriónicahumana
IATF inseminaciónartificialatiempofijo

Indice
28
IGF factordecrecimientoinsulínico
IGF-I factordecrecimientoinsulínicotipoI
INIA InstitutoNacionaldeInvestigaciónyTecnologíaAgrariay Alimentaria
LH hormonaluteinizante
MED folículosmedianos
MEDCREC folículosmedianosencrecimiento
MGA acetatodemelengestrol
Mhz megaherzios
MPA acetatodemedroxiprogesterona
msnm metrossobreelniveldelmar
P4 progesterona
PAC políticaagrariacomunitaria
PDGF factordecrecimientoderivadodelasplaquetas
PGE2 prostaglandinaE2
PGF2α prostaglandinaF2α
PGI2 prostaciclina
SITRAN sistemaintegraldetrazabilidadanimal
TGF-β factordecrecimientotransformanteβ
UE UniónEuropea
Vs. versus

RESUMEN


Resumen
31
RESUMENLa administración de progestágenos es el método más utilizado como
tratamiento para la sincronización de celos y ovulaciones en ganado ovino. Enesencia,suaplicacióndeprimelasecrecióndeLHy,conello,impideelcrecimientoterminalylaovulacióndefolículospreovulatorios;unavezretiradoeltratamiento,seproducelaaparicióndelcelo,elpicodeLHylaovulacióndeformacontroladaysincronizada. Sin embargo, y a pesar de su amplio uso, los tratamientosprogestativos presentan varios inconvenientes. Por una parte, han sidoidentificados como agentes depresores de la fertilidad; por otro, su uso seencuentra cada vez más limitado debido, tanto a regulaciones sobre residuoshormonalesdediferentespaíses,comoacriteriosdebienestaranimal.
Es por ello por lo que en el presente trabajo de tesis doctoral se planteacomo objetivo la identificación y desarrollo de protocolos de sincronización decelosyovulacionesquetenganenconsideraciónlamejoradelbienestaranimalyla disminución de residuos hormonales, investigando los efectos obtenidos alreducireltiempodeexposiciónalosprogestágenosenlosprotocoloshormonalesy la utilización de otro tipo de dispositivo intravaginal basado en progesteronanatural(CIDR).
Enunaprimerafaseexperimentalseevaluóelefectodelareduccióneneltiempo de tratamiento con progestágenos (acetato de fluorogestona o FGA),concretamente,de14a7días,y laposibleinfluenciadelfotoperiodo, larazaylaaplicacióndePGF2αalinicioofinaldeltratamientoenelcasodelostratamientoscortos.Paraellosedeterminóelporcentajedegestacióny laprolificidaden tresgrupos de tratamiento (Grupo control: tratamiento 14 días + eCG a la retirada,Grupo PG-Corto: tratamiento 7 días + PGF2α al inicio + eCG a la retirada, GrupoCorto-PG:tratamiento7días+PGF2αyeCGa laretirada).Duranteel fotoperiododecreciente,elporcentajedegestacionesfuesemejanteenlostresgrupos(grupocontrolotratamientolargo76,7%,grupoCorto-PG90,0%ygrupoPG-Corto80%).En loreferentea laprolificidadtampocohubodiferenciassignificativasentre lostresgrupos(grupocontrol2,45±1,06,grupoPG-Cortoy2,76±1,05ygrupoCorto-PG 2,30±1,07). En el fotoperiodo creciente, los resultados tampoco difirieronsignificativamenteentregrupos,enfertilidad(grupocontrol80%,grupoPG-Corto79,2% y grupo Corto-PG, 59,1%), aunque la reducción de fertilidad en eltratamientoCorto-PGenel fotoperiodocreciente, respectodeldecrecientesí fuesignificativa.Laprolificidadtampocovariósignificativamenteenestefotoperiodo(grupocontrol2,42±1,10,grupoPG-Corto2,53±1,12ygrupoCorto-PG2,15±1,14).Por lo tanto, la aplicación de tratamientos con dispositivos intravaginalesimpregnadosconprogestágenospuedeserreducidadelos14díasutilizadosenlostratamientosclásicosa7días,siemprequeseacompañendelaadministracióndeprostaglandina,yaqueseobtienensimilarestasasdegestaciónyprolificidad.

Resumen
32
Enunasegunda faseexperimentalsecompararon losmismosmétodosdesincronizacióndecelosqueen la faseexperimentalanterior,peroprofundizandoenlosaspectosmásfisiológicosreferentesalafuncionalidadovárica,hipofisariayrespuestaaltratamientodesincronizacióntraslaimplementacióndelosdistintosprotocolos hormonales en la época favorable reproductiva (fotoperiododecreciente).
En cuanto al porcentaje de animales que salieron en celo en las primeras72htraslaretiradadeldispositivovaginalobservamosun100%(9/9),vs.63,6%(7/11) vs.90,0% (10/11) para los grupos control o largo, PG-Corto y Corto-PG,respectivamente, sin diferencias significativas, pero con tendencia en el caso delgrupoconpeorresultado(PG-Corto;P=0,069).Eltiempomediodesalidaencelomedidoenhorasfuesimilarentrelosgrupos(35,1hvs.43,4hy48htraslaretiradade los dispositivos vaginales), mientras que el rango de tiempos en el que lasovejassalíanencelo(indicadordeagrupación)eramenoreneltratamientoCorto-PG(48h)respectodeltratamientoPG-Corto(62h).
Encuantoa ladinámicafolicular,seobservóprincipalmente,queelgrupode tratamiento Corto-PG daba lugar a un mayor reclutamiento y desarrollofolicular que resultaba enmás folículosmedianos al final de la fase folicular, encomparaciónconelotrogrupodetratamientocorto(PG-Corto).Enloquerespectaa la fase luteal y el porcentaje de ovulación no se observaron diferenciasestadísticas en cuanto al número medio de cuerpos lúteos /oveja, aunque ladistribución del número de cuerpos lúteos sí que varió significativamente entregrupos,conmásanimalescon3CLentrelosgruposcortos(57,1%enCorto-PGy43,5% en PG-Corto) respecto del grupo control o largo (11,1%); a cambio, seobservaronmenosanimalescon4CLenlosgruposdeprotocoloscortos(0versus22.2%). Finalmente, también se observó una menor concentración de P4 en elgrupoPG-Corto(1,02±1,06ng/ml)respectodelosnivelesobservadosenelgrupoCorto-PG (2,11±2,82 ng/ml), o grupo control o tratamiento largo (2,01±1,82ng/ml).
Por lo tanto, el estudio en profundidad de la funcionalidad ovárica ehipofisaria en fotoperiodo decreciente demostró patrones endocrinos y decrecimiento y funcionalidad folicular similares en los tratamientos clásicosbasados en 14 días de tratamiento progestativo y los basados en 7 días conaplicacióndeprostaglandinaalaretiradadelprogestágeno(protocoloCorto-PG)y,enamboscasos,mejoresqueenlostratamientosbasadosen7díasconaplicaciónde prostaglandina a la inserción del progestágeno (protocolo PG-Corto). Por elcontrario, en fotoperiodo creciente, el protocolo Corto-PG se relacionó con unamenor tasa de salida en celo; este hallazgo pone de manifiesto la necesidad defuturosestudiosperopodríarelacionarsecon lapresenciadecuerpos lúteosconbajarespuestaaprostaglandinasenanestro.

Resumen
33
Enlatercerafaseexperimentalseestudiólautilizacióndedosdispositivoscondiferentesprogestativos(progestágenosenesponjasintravaginalesconFGAycon progesterona en dispositivos CIDR) aplicados en un protocolo clásico de 14días, con eCG a la retirada de los mismos durante la estación reproductiva(fotoperiodo decreciente). El porcentaje de gestación no difirió entre grupos(61,7%vs.51,6%paraelgrupoFGAyCIDR,respectivamente;P>0,05),ratificandola adecuación del nuevo dispositivo probado en base a progesterona natural.Además,secomprobóunmenorporcentajedepérdidasdeldispositivo(4,4%vs.1,9%paraelgrupoFGAyCIDR,respectivamente;P>0,05)yunprácticamentenuloporcentaje de dispositivos CIDR que producían secreciones malolientes (1%),respectodelasclásicasesponjasintravaginales,conmásdeun70%deellasdondesíseproducen.
En la última fase experimental se valoraron las posibles diferencias en larespuesta con dos dispositivos con diferentes progestativos (progestágenos enesponjasintravaginalesdeFGAyprogesteronaendispositivosCIDR)aplicadosenelprotocoloCorto-PG.Enépocadefotoperiodocrecientesedemostróunaeficaciasemejante del CIDR respecto a las esponjas, en cuanto a porcentaje depresentacióndecelos(100%enamboscasos),yalritmodepresentacióndeceloyfuncionalidad ovárica e hipofisaria (liberación de LH y pico de LH). Durante elfotoperiodo decreciente tampoco observamos diferencias en el porcentaje deovejasquesalieronencelo,nienel tiempomedioenmostrarlo.Sinembargo,seobservóunagrupamientomayor en lapresentacióndel celo enel grupo tratadoconeldispositivoCIDR,perounamenorliberacióndeestradiolalprincipiodelafase folicularyunmenorniveldeprogesteronadurante la fase luteal (5,94±1,42ng/ml frentea4,34±1,96ng/ml, respectivamente), sin estar éstospordebajodelosnivelesfisiológicosesperadosenlascorrespondientesfasesdelciclo.
Endefinitiva,elanálisisconjuntodelosexperimentosanterioresnosllevaacorroborar la idoneidadde lautilizacióndeprotocolosde sincronizacióndecelocortos de 7 días, siempre que se combinen con la administración deprostaglandina.Además,enelcasodelosdispositivosCIDR,habiendocorroboradounasimilitudenlacapacidaddesincronizaciónyfertilidad,sesumanlasventajasde lamejoraenelbienestaranimal, la reduccióndepérdidasde losdispositivosporpartedelosanimalessincronizadosylaposibilidaddesureutilizacióntrassuesterilización, por lo que este protocolo podría convertirse en una herramientaalternativa cuya incorporación en elmanejo reproductivo de los rebaños ovinosfavoreceríalosresultadosdefertilidaddeunaformanatural,sincostoexcesivoysiendodefácilaplicaciónporpartedeproductoresnotecnificados.


SUMMARY


Summary
37
SUMMARYProgestogen administration is themost frequently usedmethod as estrus
and ovulation synchronization treatment in sheep. In essence, its applicationdepresses the secretion of LH and, thereby, prevents the terminal growth ofpreovulatory follicles and their ovulation; once the intravaginal dispensers arewithdrawn,theonsetofestrus,LHpeakandovulationoccur.However,despiteitswide use, progestagen treatments have several disadvantages. On the one hand,theyhavebeenidentifiedasfertilityreducers;Ontheotherhand,itsuseisbeingincreasinglylimitedduetobothhormoneadministrationreductionandtoanimalwelfareregulationsinAnimalProduction.
Therefore, the present work had as objective the identification anddevelopment of protocols of estrus and ovulation synchronization in sheep thattakeintoconsiderationtheimprovementoftheanimalwelfareandthereductionof hormonal residues, focusing on the specific effects observed by reducing theexposuretimetoprogestogentreatmentsandotherintravaginalreleasesbasedonnaturalprogesterone(CIDR).
In a first experimental phase, the effects of the reduction in the time oftreatmentwithprogestagen(fluorogestonacetateorFGA)intravaginaldispensers,namely, during 14 (long treatment) versus 7 days (short treatment), and thepossibleinfluenceofphotoperiod,breedandapplicationofPGF2αatthebeginningorattheendoftreatmentinthecaseoftheshorttreatmentswereexplored.Thepercentageofgestationandprolificacywereevaluatedinthreetreatmentgroups(Controlgroup:progestagentreatmentduring14days+eCGatwithdrawalofthedispensers, Group PG-Short: treatment during 7 days + PGF2α at dispenserapplication + eCG at the dispensers withdrawal and the Group Short- PG withtreatmentduring7days+PGF2αandeCGatthedispenserswithdrawal.Duringthedecreasing photoperiod, the percentage of gestations was similar in the threegroups(controlgroupor long treatment:76.7%,groupShort-PG:90.0%andPG-Short: 80%). Similarly to gestation percentage, there were no significantdifferencesbetweenthethreegroupsintheprolificacy(controlgroup2.45±1.06,PG-Short2.76±1.05andShort-PGgroup2.30±1.07lambs/ewe).
During the growing photoperiod, the results did not differ significantlybetween groups either in fertility or prolificacy (control group 80%, group PG-Short79.2%andgroupShort-PG,59.1%),althoughfertilityreductionintheshort-PGtreatmentduringthegrowingphotoperiod,withrespecttothefertilityinthisgroupduringthedecreasingphotoperiodwassignificant.
Prolificacy neither changed significantly during this photoperiod (controlgroup 2.42 ± 1.10, PG-Short group 2.53 ± 1.12 and Short-PG group 2.15 ± 1.14lambs/ewe).

Summary
38
Therefore, the application of treatments with progestagen-impregnatedintravaginalreleasemaybereducedfromtheconventional14daystoalengthof7days, only if they are accompaniedby the administrationof prostaglandin, sincesimilarratesofgestationandprolificacyareachieved.
Inasecondexperimentalphase,thesameestrussynchronizationprotocolsas in the previous experimental phase were applied during the decreasingphotoperiod,with the aimatdeepening in themorephysiological aspects of theovarianandpituitaryfunction.
Thepercentageofewesthatshowedheatduringthefirst72hoursaftertheremovalofthereleaseweresimilaramonggroups[100%(9/9)vs.63.6%(7/11)vs.90.0%(10/11) for the longgroups,PG-ShortandPG-Short, respectively].Themeantimetoshowestruswasalsosimilaramongthethreegroups(35.1hvs.43.4hand 48h after the withdrawal of the vaginal release for the three groups,respectively),while the time range inwhich the sheep showedheatwas shorteraftertheshort-PGtreatment(48h)thanafterthePG-shorttreatment(62h).
Regardingthefolliculardynamic,weobservedthattheShort-PGtreatmentgroupshowedagreaterrecruitmentandfolliculardevelopmentratethatresultedinmoremediumfolliclesattheendofthefollicularphase,comparedtotheothershort treatment group (PG-Short). Regarding the luteal phase and the ovulationpercentage,nostatisticaldifferenceswereobservedinthemeannumberofcorpuslutea (CL)/sheep, although the distribution of the number of Cls did varysignificantly among groups, with more animals with 3 CL in the short groups(57.1%inShort-PGand43.5%inShort-PG)whencomparedtotheanimalsofthelonggroup(11.1%);Incontrast,feweranimalshad4CLsintheshortgroups(0%versus22.2%).Finally,alowerconcentrationofprogesterone(P4)inthePG-Shortgroup(1.02±1.06ng/ml)wasobservedwithrespecttothelevelsobservedintheShort-PGgroup(2.11±2.82ng/ml)or in thecontrolgroup(2.01±1.82ng/ml).
Therefore, the study of the ovarian and pituitary function during thedecreasing photoperiod demonstrated a similar endocrine and follicular growthpatternsandfunctionalityaftertheconventional14daysprogestagentreatmenttothoseobservedafterthe7daystreatmentwithapplicationofprostaglandinatthedispenserremoval(Short-PG).Moreover,resultswerebetterinthelonggroupandthe Short-PG group than after the short treatmentwith the prostaglandin at thebeginning of the protocol (PG-Short). In contrast, during the increasingphotoperiod, the Short-PG protocol was associated with a lower rate of estrusdetection; this fact couldbe related to thepresenceofCLswith lowresponse toprostaglandinsduringanestrusbutfuturestudiesonthisissueareneeded.
In the third experimental phase the differences in fertility after a classicprotocolof14days,usingtwodifferentintravaginaldevices(FGA-progestogensin

Summary
39
the intravaginal sponges and progesterone in CIDR releases) with eCG at thewithdrawal of the releases during the reproductive season were explored. Thepercentage of gestation did not differ between groups (61,7% vs.51,6% for theFGAandCIDRgroup, respectively,P>0.05), confirming the suitabilityof thenewnaturalprogesteronedevicetested.Inaddition,alowerpercentageoflostdeviceswereobserved(4,4%vs.1,9%fortheFGAandCIDRgroup,respectively,P>0.05)and a very reduced percentage of CIDR releases that caused malodor (1%),compared to the classic intravaginal FGA-sponges,withmore than 70%of themimpregnatedwithmalodoroussecretions.
Inthelastexperimentalphase,thedifferencesinthereproductiveoutcomesaftertheShort-PGprotocolusingtwodifferentintravaginaldevices(progestogensin the intravaginal sponges and progesterone in CIDR releases) were explored.During the increasing photoperiod, a similar efficacy was shown for the CIDRgroupinrelationtosponges,intermsofpercentageofestrus(100%inbothcases),in terms of mean time to estrus onset, and regarding the ovarian functionality(estradiolconcentrationduringthefollicularphase)andthepituitaryfunction(LHreleaseandpeakofLH).
Duringthedecreasingphotoperiod,wealsodidnotobservedifferences inthepercentageofestrus,or inthemeantimeforshowingheat.However,amoreintensegroupingeffectwasobservedinthegrouptreatedwithCIDRdevices,butalower release of estradiol at the beginning of the follicular phase and a lowersecretionofprogesteroneduringthelutealphase(5.94±1.42ng/mlvs.4.34±1.96ng/ml, respectively). However, these decreased values were not below thephysiologicallevelsexpectedateachcorrespondingcyclestage.
Finally, the jointanalysisof theseexperiments leadsustocorroboratetheappropriatenessoftheuseoftheshortsynchronizationprotocolsof7daysbasedon progestogen/progesteron intravaginal release, as long as they include theadministration of prostaglandin. In addition, in the case of CIDR release, oncedemonstrated their similar synchronizationand fertilitycapacity, theadvantagesof improved animal welfare, the reduction of losses of the release and thepossibility of their reuse after sterilization should be added. Therefore, such aprotocol couldbecomeanoption in sheepherds, improvingmaintaining fertilityoutcomes in a more natural way, without excessive cost and being easy to beappliedbynon-specializedproducers.


REVISIÓNBIBLIOGRÁFICA


RevisiónBibliográfica
43
1 Revisiónbibliográfica
1.1 Laimportanciadelganadoovino
Lamayoríadelostratadistascoincidenenseñalarqueladomesticacióndelaespecieovina(Ovisaries)tuvolugarenAsiaMenoryAsiaCentralenelperiodoNeolítico, extendiéndose posteriormente a Europa y África del Norte. A laPenínsulaIbéricalaovejallegaría,probablemente,haciaelquintomilenioa.C.porlascostasmediterráneas,europeayafricana,accediendoposteriormenteovinosdelana largaygruesa,procedentesdeEuropaCentral,aportadospor la invasióndelosceltasalrededordelsigloVIIa.C.(Buxadé,1996;DazaAndrada,2002).
Los ovinos domésticos han sido explotados desde la antigüedad para laproduccióndelanaypielesparaelvestidoydelecheycarneparaelalimento.Elinterés zootécnico de la especie se basa en su eficacia para aprovechar zonasáridas y semiáridas de bajas disponibilidades forrajeras, zonas de monte detopografía accidentada y subproductos y residuos de cultivos agrícolas. Lacapacidad de pastoreo y un reducido formato corporal son caracteres quepermiten a los ovinos satisfacer sus necesidades nutritivas más fácilmente queotras especies demayor tamaño en superficies de escasos recursos alimenticiosy/odedifícilacceso(Buxadé,1996;DazaAndrada,2002).
La explotación ovina se justifica, además, pormotivosmedioambientales,comercialesysociológicos(DazaAndrada,2002).Elpastoreoracionaldelaszonasde monte es un instrumento eficaz para prevenir incendios, y el aporte de lasdeyecciones al suelo lo enriquece enmateria orgánica potenciando el desarrollovegetalyevitandofenómenosdeerosión.Nocabeduda,porlotanto,queelganadoovino, explotado bajo fórmulas extensivas o semiextensivas de producción,contribuye al mantenimiento de los ecosistemas (Daza Andrada, 2002; Mollard,2002). Por otro lado, es una especie con razas perfectamente adaptadas a laproducción animal intensiva que permiten una gran rentabilidad de dichossistemas, ofreciendo estabilidad en oferta de mano de obra en zonas rurales(Hernandezetal.,2011;Elviraetal.,2013).
Los productos que genera son de extraordinaria calidad alcanzando en elmercado precios elevados (corderos lechales, quesos artesanales e industriales,prendas para vestir de las industrias textil y peletera), por lo tanto, es derelevancia en todoelmundo: en lospaísesenvíasdedesarrollo esunelementofundamental en las economías de subsistencia y, en los desarrollados, frena ladespoblaciónhumanadelcampoayudandoalmantenimientodelmedioruralyladevastacióndelcampoporincendios(DazaAndrada,2002).
En el siguiente apartado vamos a repasar, brevemente, la situación delsector ovino en el contexto mundial, en la Unión Europea y en España, como

RevisiónBibliográfica
44
introducción a la revisión bibliográfica dedicada a la reproducción en ovino queintegraestetrabajo.
1.2 Elganadoovinoenelmundo
SegúnlosúltimosdatosoficialesdelaFAO(FAOSTAT,2015),elcensoovinomundialestáen tornoa los1.200millonesdecabezas,desigualmenterepartidassegún continente (Figura 1). De éstas, alrededor del 70% se destinan a laproduccióncárnica,mientrasqueel30%sonovejaslecheras.Seestimaportantoque laproducción cárnica es laprincipal enesta especie, dedicándosea este fin,casilastrescuartaspartesdelasreproductorasovinasexistentes.
Figura1.Distribucióncontinentaldelcensoovino(FAOSTAT,2015)
Las máximas fracciones censales corresponden a Asia, que cuenta con elmayor censo a nivel mundial (45%), tanto para la producción cárnica, comolechera. El principal país productor es China, con 134 millones de ovejasdestinadasa laproduccióncárnicay43,8millonesdeovejas lecheras(FAOSTAT,2015).
El segundo continente productor de ganado ovino es África, donde seencuentra el 28%del censo total. Lamayor parte de la producción ovina en elcontinente africano se sitúa en Nigeria y Sudán, que cuentan con 40,5 y 39,8millonesdeovejas,respectivamente(FAOSTAT,2015).
Europaeseltercercontinenteencensodeovejas(11%).Lamayorpartedelcenso se destina a la producción cárnica (69,1%). Reino Unido es el mayorproductorcon23,8millonesdecabezas,seguidodeEspañacon16,5millonesdecabezasyRumanía,con10millonesdecabezas(FAOSTAT,2015).
África28%
América7%
Asia45%
Europa11%
Oceanía8%
Distribucióndelcensomundialovinoporcon4nentes
Con4nente Cabezas(enmillones)
Asia 549,6
África 340,5
Europa 130,3
Oceanía 102,4
América 87,1

RevisiónBibliográfica
45
Oceaníapresentaun8%delcensomundialtotal,explotandosóloovinodecarne, repartiéndose casi la totalidad de los efectivos por igual entre Australia(70%) y Nueva Zelanda (29%); mientras que la mínima presencia censal seproduce en América, siendo Brasil (17,6millones de cabezas) y Argentina (14,7millonesdecabezas)losmayoresproductoresdecarneovina(FAOSTAT,2015).
Los pequeños rumiantes participan en gran medida en la denominada“economíadesubsistencia”,queesfundamentalenlospaísesenvíasdedesarrollo,ya que cuentan con un 65% del censo mundial ovino. En estos países suimportancia no es sólo la que deriva de sus datos censales, sino también de surelevanciasocial,yaquelospequeñosrumiantes(ovinoycaprino)tienenescasosrequerimientos y una gran capacidadpara transformar subproductos y residuosagrícolas y domésticos en carne, lana, pieles y leche (Gutierrez-A, 1985;Buxadé,1996).
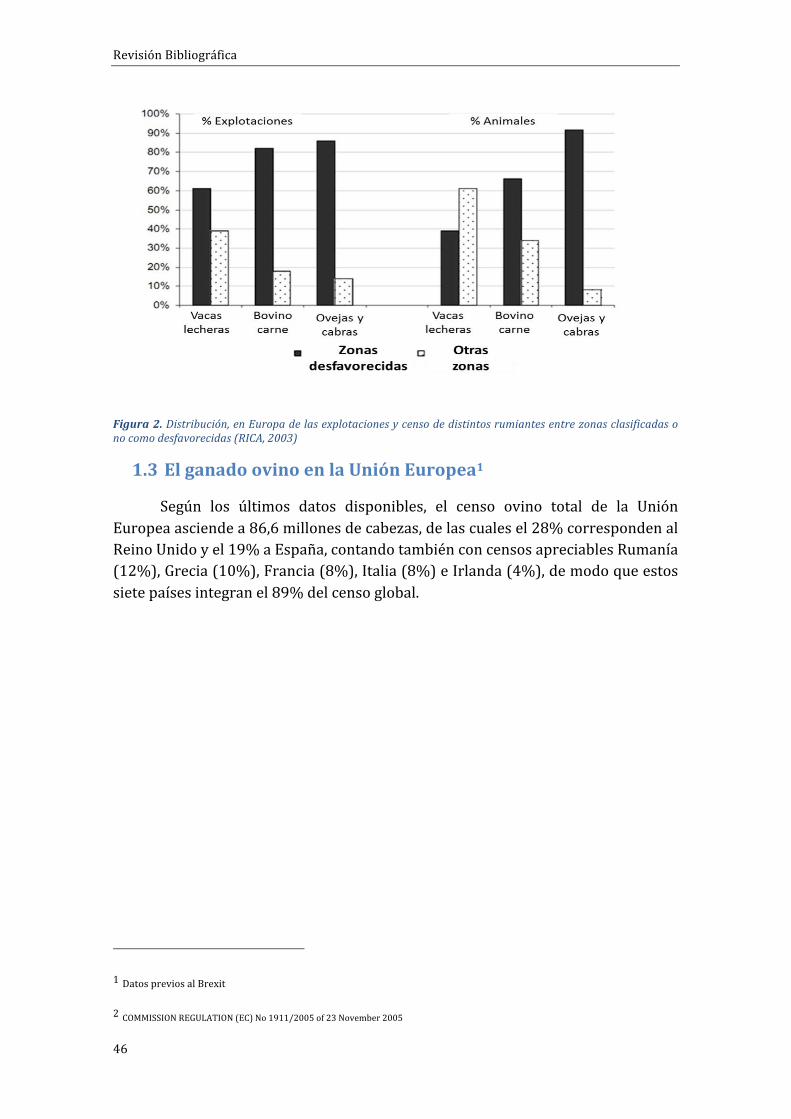
También en Europa se considera que el ganado ovino tiene un papelrelevante en la fijación de mano de obra y población humana en regionesdesfavorecidas,dadoquesuscaracterísticas fisiológicasydemanejopermitensuexplotaciónenmediosmásdesfavorablesparaotrosrumiantesmásselectoscomolosbovinos.Losovinossoncapacesdepastarymejorarlasformacionesvegetalesde la estepa de matorral mediterráneo; cuentan con un periodo de lactanciareducida (3-4meses) lo que aminora las necesidades hídricas, permitiendo unamejor adaptación a climas áridos. Tienen asimismo la capacidad de realizardiariamentedesplazamientoslargossindisminuirsuproductividad(Meuret,2006;Pouxetal.,2009).Dehecho, talycomosepuedeapreciaren la figura2,másdel90%delcensodeovinoenEuropasesitúaenzonasdesfavorecidas.
RevisiónBibliográfica
46
Figura2.Distribución,enEuropadelasexplotacionesycensodedistintosrumiantesentrezonasclasificadasonocomodesfavorecidas(RICA,2003)
1.3 ElganadoovinoenlaUniónEuropea1
Según los últimos datos disponibles, el censo ovino total de la UniónEuropeaasciendea86,6millonesdecabezas,delascualesel28%correspondenalReinoUnidoyel19%aEspaña,contandotambiénconcensosapreciablesRumanía(12%),Grecia(10%),Francia(8%),Italia(8%)eIrlanda(4%),demodoqueestossietepaísesintegranel89%delcensoglobal.
1DatospreviosalBrexit
2COMMISSIONREGULATION(EC)No1911/2005of23November2005

RevisiónBibliográfica
47
Figura3.DistribucióndelcensodeovinoporpaísesenlaUE-28enel2015(EUROSTAT,2016)
Elrespetoalmedioambienteesunodelospilaresdeldesarrollosostenible,yunadelasprioridadesdelaUEenlabúsquedadeunaagriculturamultifuncionalal servicio de sus ciudadanos, siendo además un elemento clave para la luchacontraelcambioclimático.Laaplicacióndeprincipiosdegestiónsostenibledelosrecursos naturales y los residuos generados por la actividad primaria, enparticular, la ganadería, es la respuesta a esta creciente demanda ambiental(Martinetal.,2004).
Esta situación se concreta con la creciente exigencia por parte de losconsumidoresdequelosproductoresdealimentosdeorigenanimalrespetenuncódigo ético, y lleven a cabo prácticas de manejo no contaminantes, lo que seconocecomoproductos“clean,greenandethical”(Martin&Kadokawa,2006).
El desarrollo de una política medioambiental comunitaria en el ámbitoganadero nace como consecuencia del crecimiento de la población y de lanecesidad de garantizar la seguridad alimentaria a la sociedad, que provocaronimportantescambiosenlossistemasdeproducciónagraria(Macrietal.,2006).
En el sector ganadero, las explotaciones extensivas evolucionaron asistemasdeproducciónintensivos,endondeseincrementólacargaganadera.Estatransformaciónde la ganadería tambiénha idoacompañadadeun cambiode sumapa de distribución, apareciendo en ciertas zonas una alta concentraciónganadera, que constituye una causa de determinados problemasmedioambientales(MAGRAMA,2015).
DISTRIBUCIÓN DEL CENSO DE OVINO POR PAÍSES EN LA UE-28
ReinoUnido28%
España19%
Rumanía12%
Grecia10%
Italia8%
Francia8%
Irlanda4%Portugal2%
Alemania2%Bulgaria2%
Hungría1%Holanda1%
Resto3%
País
Cabezas(en
millones)
ReinoUnido 23,9
España 16,8
Rumanía 10,06
Grecia 8,9
Italia 7,1
Francia 7,05
Irlanda 3,3
Portugal 2,02
Alemania 1,6
Bulgaria 1,3
Hungría 1,1
Holanda 1,03
Resto 2,5

RevisiónBibliográfica
48
A nivel comunitario, la normativa medioambiental actual ya incluye laganaderíadentrodesuregulación,especialmente lasexplotaciones intensivasdealgunasespeciesy,dentrodeestas, las granjasdemayor tamaño.Asímismo, endistintos Convenios Internacionales, como los Protocolos de Kyoto y deGotemburgo, se contempla al sector ganadero en general como una actividadpotencialmentecontaminanteyportantosujetaaregulación(MAGRAMA,2015).
Estasnormativas representanparael sectorganaderoel cumplimientodeuna serie de compromisos que tienen un marcado carácter técnico y para suaplicaciónnecesitanundesarrollo tecnológicoespecificoanivelde lasdiferenteszonasagro-climáticas,teniendoencuenta,además, loscondicionantesespecíficosde nuestros sistemas productivos. Por otra parte el carácter difuso de lasemisiones ganaderas dificulta su medición en las granjas, que también se velimitadoporcondicionantesdetiposanitario(MAGRAMA,2015).
1.4 ElganadoovinoenEspaña
La llegada de la oveja doméstica procedente de Asia a nuestra penínsulatuvoprobablementelugardesdeÁfrica.Lasprimerasreferenciasaovinosconunaantigüedadde3.700añoslasencontramosenlosgrabadosmuralesdelValledelasBatuecas de Salamanca, y ya en la época de los fenicios y romanos la lanaprocedente de las ovejas de nuestra península era muy apreciada (CamberoMuñoz, 1999). Según diversos datos históricos recopilados por Daza-Andrada(2002),elcensoovinoespañolocupódesdelaAltaEdadMediaunlugardestacadoen el contexto europeo, siendo el máximo productor y exportador de lana finadesdefinalesdelsigloXIIIhastalasegundamitaddelsigloXIX.
Posteriormente,elcensosefuereduciendodrásticamentedebidoafactorespolíticos,sociológicos, técnicosycomercialesentre losquecabedestacar,porunlado, la reducción de la superficie de pastos y cultivos forrajeros derivada de lapolíticaproteccionistade lapostguerrahacia los cultivoscerealistas;porotro, larevolución industrial que provocó la emigración de jóvenes pastores a las áreasindustrialesyfinalmente,lacaídadelpreciodelalanaoelaumentoprogresivodelcoste de la mano de obra (Daza Andrada, 2002). Sin embargo, la entrada en laUniónEuropea,deficitariaencarnedeovino,provocóun incrementocensalantelasbuenasprevisionescomercialesquetambiénocasionóunincrementodelcensodeovejasproductorasdeleche(Sánchez-Rodríguez,2011).
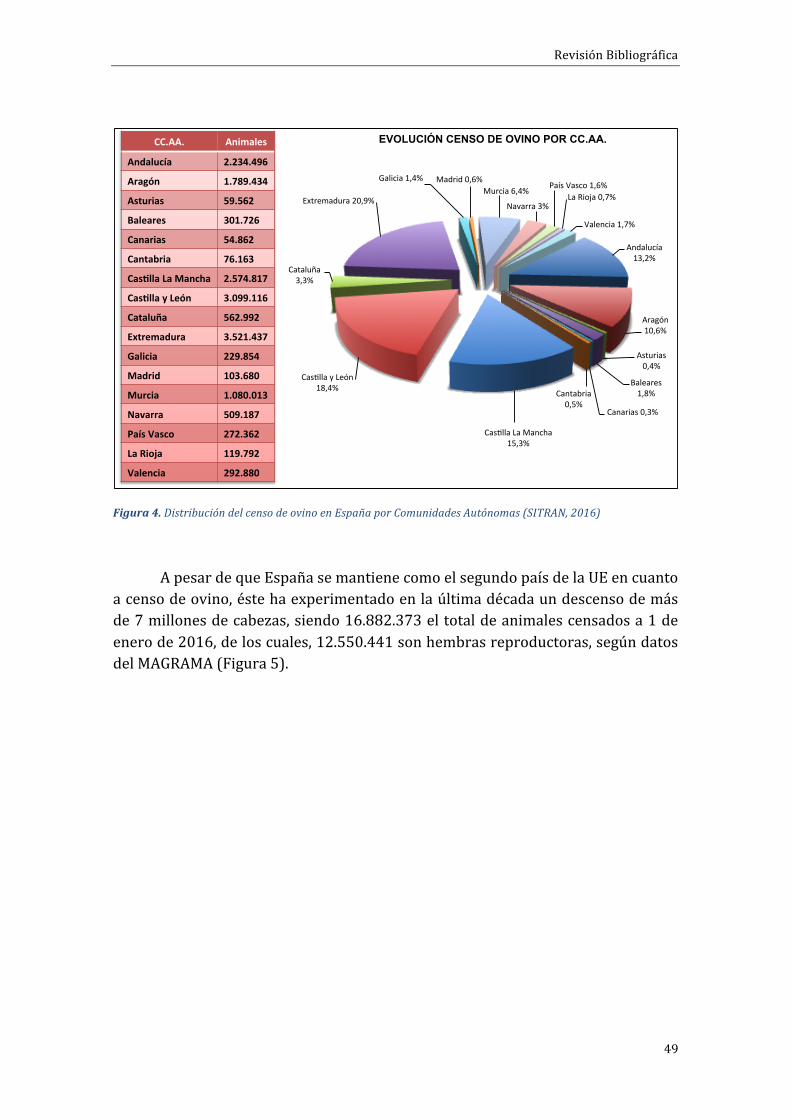
La cabaña ganadera ovina española es una de las más importantes de laUniónEuropea.Ocupaelsegundolugarenproduccióndecarneylanayeseltercerpaísproductordelechedeoveja,inmediatamentepordetrásdeGreciayRumanía.Las cinco comunidades autónomas con más censo de esta especie engloban el78,4%delmismoyson:Extremadura(20,9%),CastillayLeón(18,4%),CastillaLaMancha(15,3%),Andalucía(13,2%)yAragón(10,6%)(Figura4).
RevisiónBibliográfica
49
Figura4.DistribucióndelcensodeovinoenEspañaporComunidadesAutónomas(SITRAN,2016)
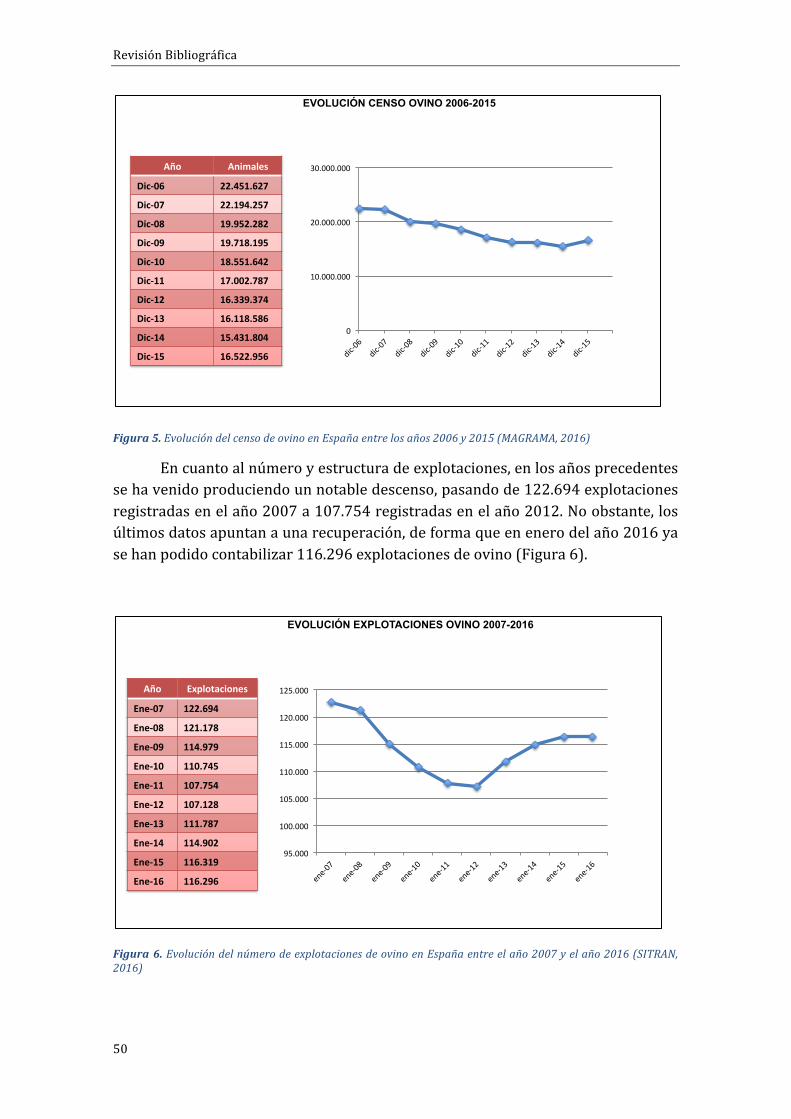
ApesardequeEspañasemantienecomoelsegundopaísdelaUEencuantoacensodeovino,éstehaexperimentadoenlaúltimadécadaundescensodemásde7millonesdecabezas,siendo16.882.373eltotaldeanimalescensadosa1deenerode2016,deloscuales,12.550.441sonhembrasreproductoras,segúndatosdelMAGRAMA(Figura5).
EVOLUCIÓN CENSO DE OVINO POR CC.AA.
Andalucía13,2%
Aragón10,6%
Asturias0,4%
Baleares1,8%
Canarias0,3%
Cantabria0,5%
Cas>llaLaMancha15,3%
Cas>llayLeón18,4%
Cataluña3,3%
Extremadura20,9%
Galicia1,4% Madrid0,6%Murcia6,4%
Navarra3%
PaísVasco1,6%LaRioja0,7%
Valencia1,7%
CC.AA. Animales
Andalucía 2.234.496
Aragón 1.789.434
Asturias 59.562
Baleares 301.726
Canarias 54.862
Cantabria 76.163
Cas@llaLaMancha 2.574.817
Cas@llayLeón 3.099.116
Cataluña 562.992
Extremadura 3.521.437
Galicia 229.854
Madrid 103.680
Murcia 1.080.013
Navarra 509.187
PaísVasco 272.362
LaRioja 119.792
Valencia 292.880
RevisiónBibliográfica
50
Figura5.EvolucióndelcensodeovinoenEspañaentrelosaños2006y2015(MAGRAMA,2016)
Encuantoalnúmeroyestructuradeexplotaciones,enlosañosprecedentessehavenidoproduciendounnotabledescenso,pasandode122.694explotacionesregistradasenelaño2007a107.754registradasenelaño2012.Noobstante,losúltimosdatosapuntanaunarecuperación,deformaqueenenerodelaño2016yasehanpodidocontabilizar116.296explotacionesdeovino(Figura6).
Figura6.EvolucióndelnúmerodeexplotacionesdeovinoenEspañaentreelaño2007yelaño2016(SITRAN,2016)
EVOLUCIÓN CENSO OVINO 2006-2015
Año Animales
Dic-06 22.451.627
Dic-07 22.194.257
Dic-08 19.952.282
Dic-09 19.718.195
Dic-10 18.551.642
Dic-11 17.002.787
Dic-12 16.339.374
Dic-13 16.118.586
Dic-14 15.431.804
Dic-15 16.522.956
0
10.000.000
20.000.000
30.000.000
dic-06
dic-07
dic-08
dic-09
dic-10
dic-11
dic-12
dic-13
dic-14
dic-15
EVOLUCIÓN EXPLOTACIONES OVINO 2007-2016
Año Explotaciones
Ene-07 122.694
Ene-08 121.178
Ene-09 114.979
Ene-10 110.745
Ene-11 107.754
Ene-12 107.128
Ene-13 111.787
Ene-14 114.902
Ene-15 116.319
Ene-16 116.296
95.000
100.000
105.000
110.000
115.000
120.000
125.000
ene-07
ene-08
ene-09
ene-10
ene-11
ene-12
ene-13
ene-14
ene-15
ene-16

RevisiónBibliográfica
51
El sector de los pequeños rumiantes en el Estado Español representa el8,6%delaproducciónfinalganadera.Estesectortieneunimportantepapelenlavertebracióndel territorio,aprovechamientode los recursos naturales,así comoun papel esencial en asegurar la cohesión del tejido rural y el uso sostenible dehábitatsenlaszonasdondeseasienta.
En consecuencia, el sector ovino en España experimenta un descensoacusadoengeneral, que sehacemáspatenteenel casodelovinode carne.Estacontinua crisis se ha agravado aún más si cabe, en los últimos años, por lasdrásticassubidasdepreciosdelasmateriasprimasutilizadasparalaalimentacióndelganado,elreducidoniveldepreciosdelosproductosquesufreelproductor,elescaso consumode carnedeovinoporhabitante en continuodeclive, la faltaderelevogeneracionalenestesectorylosproblemassanitarios,engeneral,comolalenguaazuloelscrapie,queafectandemaneradirectaeindirectaalaproducción,inclusoa lademandadeproductosanimales,porpartede lapoblación(Romero-Gonzálezetal.,2008;Lahozetal.,2010).
Aesto,hayquesumarlelabajaproductividadnumérica,entendiendocomotalelnúmerodecorderosproducidosporovejayañoyloscambiosrespectoalaPolítica Agraria Comunitaria (PAC), que trata de incentivar los sistemas deproducción extensivos, por ser, tradicionalmente, la forma más habitual deexplotacióndeeste tipodeganadoyporsunotable incidenciasocioeconómicaymedioambiental(GarcíaÁlvarez-Coque,2004).Sibienestoúltimoescierto,noloesmenosqueestossistemasdeproducciónanimalpresentanescasarentabilidad,lo que hace peligrar la viabilidad de las explotaciones de este tipo y susostenibilidad(Santamaríaetal.,1998).
1.5 Descripcióndelasrazasdeinterésparaelpresentetrabajo
La raza lacaune es una raza de origen francés. Con un censo de 1,29millonesdeovejas(loquesuponeun20%delacabañaovinafrancesa),ocupalaprimera posición entre las razas ovinas de Francia. Está dividida en dos ramasprincipalescondistintasaptitudes:produccióndelecheyproduccióndecarne,conun 65% y un 35% de hembras, respectivamente. Sin embargo, las hembras delsector lácteo han sido utilizadas durante mucho tiempo para la reposición dehembrasdelsectorcárnico(Perretetal.,1995).
En1975,apartirdelaexperienciaenmejoragenéticaparalaproduccióndeleche de la Cooperativa de Inseminación Artificial OVI-TEST, y a petición de lossocios especializados en la producción de carne, comenzó un programa deselecciónymultiplicación,gestionadopor laUPRA-LACAUNE(UnitéNationaledesélection et de promotion de la race Lacaune), con el principal objetivo deaumentar la prolificidad, dejando la producción de leche como un carácteradicional(Barilletetal.,2001).

RevisiónBibliográfica
52
Elestablecimientodeunesquemadeselecciónparamejorarlaprolificidadde las ovejas hizo aparecer un determinismo genético de carácter “prolificidad”basado en la segregación de una mutación con un fuerte efecto dentro de lapoblación lacaune. La creación de familias experimentales a partir de ovejassupuestamente portadoras de esta mutación, confirmó la existencia de un genmayor con efectos sobre la tasa de ovulación y la prolificidad, el BMP-15 (BoneMorphogenetic Protein 15), relacionado con la superfamilia del TGF-β(TransformingGrowthFactorβ)(Bodinetal.,2007).
Es a partir de los años ochenta y noventa, mediante este esquema deselección y multiplicación, cuando esta raza empieza a difundirse y a llegar amuchospaíses,entreellosEspaña,dondesudifusiónhasidoprogresivadebidoasu buena adaptabilidad a los medios secos y calurosos peninsulares y a lossistemas productivos españoles. Igualmente, posee una reconocida calidad en laproducción cárnica con la obtención de corderos con un alto índice detransformaciónencebaderoymuybuenacalidaddelacanal(Jounouetal.,2004).
Esta raza agrupa a animales de color predominantemente blanco, conindividuos que pueden presentar coloraciones en negro, de perfil plano, tamañograndeyconelvellónqueseextiendeportodalasuperficiecorporal,dejandoaldescubierto las extremidades, la parte inferior del tronco y la cabeza. Los pesosmediosdelashembrasoscilanentre70y80kgylosdelosmachosentre95y110kg(Fuentesetal.,2000).
LarazasegureñaesoriginariadelaszonasserranasdelaVegadelSegura,en las provincias de Albacete, Jaén, Granada y Almería. Se extiende por todo elsureste de España (Valencia, Alicante, Murcia, Sur de Almería y Málaga). Suelevado censo la constituye como una de las razas ovinas nacionales másimportantes(FEAGAS,2010).
Son ovinos demarcado dimorfismo sexual, perfil subconvexo, acornes enambossexos, tamañomedioyproporciones ligeramentealargadas,conunvellónsemicerrado de lana entrefina y color blanco uniforme, de reducida extensión(Fuentesetal.,2000).
Esta raza es de aptitud cárnica, pero se encuentra estrechamenteemparentada con lamanchega, de aptitud láctea, lo cual le confieremuybuenascualidadeslecherasparacriardoscorderos.Destacaporsuelevadarusticidad,quelepermitevivirenunmedioinaccesibleaotrosanimales.Estáadaptadaaunclimacontinental riguroso y extremo, y una pluviometría próxima a los 350 l/m²anuales,peromaldistribuidaalolargodelaño,quehacequeelperiodovegetativodelospastosquedereducidoaunospocosmeses(FEAGAS,2010).

RevisiónBibliográfica
53
LarazaromanovesoriginariadeRusia,delacuencaaltadelríoVolga,alnoroestedeMoscú,yadiferenciadelamayoríadelasrazaseuropeasyamericanasque son producto de cruzamientos y selección, ésta es una raza genéticamentepura,locualhacequetransfieraunvigorhíbridomuchomayorquecualquierotra(Fuentesetal.,2000).
Es una raza con prolificidad, vitalidad y habilidad materna muysobresalientes.Dehechoseconsideralarazamásprolíficaqueexiste.Además,losovinos romanov demuestran una rusticidad, vigorosidad sexual y productividadexcepcional(Fuentesetal.,2000).
Sonanimalespequeñosydecolorgrisonegrosucio.LacolaescortayestecaráctersetransmiteasuF1enloscruzamientos.Esacornetantoenmachoscomoenhembrasytienelasorejaslevantadasypuntiagudas(Fuentesetal.,2000).
LaF1provenientedelosrespectivoscrucesentremachosromanovyovejasde razas lacaune y segureña tiene como características principales un marcadoincremento de la prolificidad y de las características maternales sin perder larusticidadylaadaptaciónalmedioenelquesedesenvuelven(Flox&Daza,2000).
1.6 Fisiología de la reproducción. Ciclo estral y controlneurodendocrino
El ciclo estral está asociado a una secuencia de sucesos endocrinosinterrelacionados y que están principalmente regulados por el eje hipotálamo-hipófisis-gonadal.Elhipotálamosecretalahormonaliberadoradegonadotropinas(GnRH). La hipófisis produce hormona foliculoestimulante (FSH), hormonaluteinizante (LH), prolactina y oxitocina. Los folículos ováricos antrales secretanestrógeno e inhibina y el CL secreta progesterona y oxitocina, mientras que elendometrio uterino se encarga de la producción de prostanglandina F2α(Scaramuzzietal.,1993).
El desarrollo y maduración de los folículos ováricos, así como laesteroidogénesis,ovulaciónyformacióndelCL,estáncontroladasprincipalmentepor las gonadotropinas hipofisarias, que a su vez, dependen de la secreción deGnRH. La regulación de la secreción y biodisponibilidad de estas hormonasdepende de una compleja interacción entre varios factores internos y externos(Bartlewski et al., 2011). Entre los principales factores internos se encuentranaminoácidosproducidos localmente,esteroidesováricos,hormonastalescomolainhibina, activina y folistatina, agentes neurotransmisores y neuromoduladores.Los factores externos principales son las señales fotoperiódicas, las feromonasmasculinas, así como la nutrición y el estrés, también conocidos por afectar lafuncióndelejehipotálamo-hipófisis-gonadal(Menassoletal.,2012).Laregulaciónse puede lograr directamente, a través de los efectos de la secreción de GnRHdesdeelhipotálamohipofisario,o indirectamente,poralteraciónde lacapacidad

RevisiónBibliográfica
54
de respuesta de la hipófisis a la GnRH o de la sensibilidad del ovario a lasgonadotropinas(Bartlewskietal.,2011).
La GnRH u hormona liberadora de gonadotropinas, es un decapéptidosintetizado por neuronas especializadas del hipotálamo. Esta hormona essintetizada y almacenada en gránulos que son transportados por axones que sedirigen hacia la zona externa de la eminenciamedia (Seeburg etal., 1987; Fink,1988).Posteriormenteesliberadaenpulsossincronizadoshaciaelsistemaportalhipofisario y su secreción está regulada por las concentraciones de esteroidesgonadales tales como progesterona y estradiol (Clarke & Pompolo, 2005). Estospulsos estimulan la biosíntesis y secreción de FSH y LHpor parte de las célulasgonadotropasdelaadenohipófisis(Fink,1988).
Las gonadotropinas hipofisarias (FSH y LH) son hormonas de naturalezaglicoproteica,resultantesdelaasociacióndedossubunidades(αyβ)sintetizadasporlascélulasgonadotropasdellóbuloanteriordelahipófisis(Childs,2006).
LaFSHeslaencargadadeestimulareldesarrollofolicularovárico,actuandoaniveldelascélulasdelagranulosadelfolículoyestimulandolaactivacióndelasaromatasas, las cuales terminan el proceso de síntesis desde androstenedionahastaestradiol(Hsuehetal.,1984).
EnelcasodelaLH,existendospatronesfuncionalesdistintosdesecreciónenlaoveja,controlandocadaunodeellosdiferentesaspectosdelafunciónovárica(Geschwind&Dewey,1968;Arthur&Pearson,1989).
La secreción de LH pulsátil o tónica ocurre durante todo el ciclo enrespuestaalaliberacióndeGnRHyesimportanteparalaesteroidogénesisovárica,estimulando la síntesis de androstenediona por parte de las células de la tecainterna durante el desarrollo folicular. La descarga preovulatoria de LH, que seproduce inmediatamente después del celo, es la responsable de inducir laovulaciónylaformaciónymantenimientodelCL(Niswenderetal.,1981;Hansel&Convey,1983a;Arthur&Pearson,1989;Rawlings&Cook,1993;Goodman,1994).
LaaparicióndelpicopreovulatoriodeLHvaacompañadadeunadescargade FSH, produciéndose una segunda descarga entre las 20 y las 36 horassiguientes. Durante el ciclo estral también se produce una elevación de lasconcentracionesdeFSHcada4-5días,asociadaalaaparicióndeondasfoliculares(Souzaetal.,1997;Bartlewskietal.,2000;Evansetal.,2000;Gintheretal.,2000).
Lashormonasesteroideasováricassonlosproductosdesecreciónfolicularmejorconocidos.Haytrestiposdeesteroidesproducidosenlosfolículosováricos,todos ellos derivados del colesterol: las progestinas, los andrógenos y losestrógenos.

RevisiónBibliográfica
55
La pregnenolona es la progestina más importante del folículo, al ser laprecursoradelrestodehormonasesteroides.Sinembargo, lamásconocidaes laprogesterona(P4), intermediariaen lasíntesisdeotrosesteroidesyproducidaanivelováricopor lateca internadel folículoenelperiodoperiovulatorioyporelCL en el post-ovulatorio. Su función principal es preparar el útero para laimplantacióndel embrióny elmantenimientode la gestación (Hansel&Convey,1983a;Gore-Langton&Armstrong,1994;Reynolds&Redmer,1999).
El folículo es una fuente significativa de andrógenos ováricos, demaneraque la androstenediona y la testosterona son precursores inmediatos de losesteroidesestrogénicos,estronayestradiol,respectivamente.
Losestrógenos(estradiolyestrona),sonlosesteroidesmásimportantesanivelfisiológico,ycuyasíntesisserealizaenlascélulasdelagranulosa,atravésdelaaromatizacióndelaandrostenediona.Elestradiolinducelafasedereceptividadsexual o celo durante el ciclo sexual de las hembras de los mamíferos (Gore-Langton&Armstrong,1994).Asimismo,controlalasecrecióndelaFSHpormediodeunmecanismoderetroalimentación(Padmanabhan,2002).
La activina, la inhibina y la folistatina son péptidos producidos,principalmente, en las células de la granulosa del folículo ovárico. Regulan elcrecimientofolicularcontrolandolaliberacióndeFSH.Laactivinaylainhibinasondímerosqueconstandeunasubunidadαydosβ.Lainhibinatienecomofunciónsuprimir la producción y la secreción de FSH, mientras que la activina tiene elefectocontrario(Ying,1988).
Lafolistatina,esunmonómeroqueseunealasubunidadβdelaactivinayde la inhibina, modulando su acción sobre la secreción y liberación de FSH(Robertson,1992;Phillips&deKretser,1998).
Entre los principales factores de crecimiento implicados en la actividadováricadestacanlossiguientes:elfactordecrecimientoepidermal(EGF),elfactorde crecimiento fibroblástico (FGF), el factor de crecimiento derivado de lasplaquetas (PDGF), el factorde crecimiento transformadorβ (TGF-β), los factoresdecrecimientohematopoyético(citoquinas)yelfactordecrecimientosimilaralainsulina(IGF).Estosfactores,estándirectamenteinvolucradosenlaregulacióndela proliferación y diferenciación celular (Monget&Monniaux, 1995;Webbetal.,1999).
Las prostaglandinas son hormonas derivadas del ácido araquidónico. LamásimportanteparaelmantenimientodelaciclicidadsexualeslaprostaglandinaF2α(PGF2α).Suprincipalfuentedeproduccióneselendometriouterino(Thatcheretal.,1995).
Enlasetapasfinalesdelcicloestral,seproduceunasíntesisdereceptoresendometriales, mientras que la oxitocina, proveniente del CL, estimula la

RevisiónBibliográfica
56
produccióndePGF2α (Flintetal.,1986;Thatcheretal.,1995).Posteriormente, laPGF2αsintetizadaporlasglándulasendometrialesuterinasestransportadaporunmecanismodecontracorriente,medianteanastomosis,desdelavenauterinahastala arteria ovárica para ejercer su acción luteolítica por disminución del flujosanguíneo local, con la consiguiente disminución de la producción de P4(Knickerbockeretal.,1988;Stellflugetal.,1997;Juengel&Niswender,1999).
1.6.1 Fasesdelcicloestral
Elcicloestralcomúnmentesedivideendosfasesatendiendoalosprocesosquetienenlugarenelovario:lafaselutealylafasefolicular.Lafasefoliculartieneuna duración de 2-3 días y se caracteriza por la exhibición del comportamientosexual y la receptividad almacho (periodode24-40horas denominado celo), elpico preovulatorio de la LH y la ovulación. La transición de la fase folicular a lalutealsemarcaporlaovulacióneincluyeeldesarrollodelCLqueproduceP4.Estafasetambiénincluyelaluteolisis,propiciadaporlaprostaglandinaF2α(PGF2α).Sinembargo, la transiciónde la fase luteal a la folicularesmáscompleja. Si laovejaquedagestante,elCLpersistirádurantetodalagestación(Gordon,1996).
FasefolicularLa fase folicular incluye proestro y estro. El proestro sería el periodo
inmediatamente anterior al comportamiento estral y se caracteriza por ladisminuciónde lasconcentracionesséricasdeP4,aconsecuenciade laregresióndelCL(Senger,2003)yunaumentosignificativodeestradiol(E2),producidoporlos folículos en desarrollo, además de un marcado incremento en la actividadsecretora de todo el sistema reproductivo. El útero se alarga, el endometriocomienza a edematizarse y las glándulas uterinas incrementan su número. Lamucosavaginalsevuelvehiperémica,elnúmerodecapascelularesenelepiteliovaginalaumentaylacapasuperficialsecornifica(Abecia&Forcada,2011).
Elestrosecorresponderíaconelperiodoinmediatamenteposterior,conelperiododeaceptacióndelmacho,dereceptividadsexual,deaproximadamente36-40horas.Lasglándulassecretorasvaginales,cervicalesyuterinassecretanmayorcantidaddemucus,elendometrioylavaginasecongestionanyelcérvixserelaja.En la oveja, la ovulación ocurre normalmente de forma inmediata y espontánea,antesdelfinaldeestafase,alrededordelas24y48horasdeliniciodelcelo,siendomástardíaenrazasprolíficas(Goodman,1994),pudiendollegaraoscilarentrelas18y las72horas,conunamediade36horas,en funciónde laedad, larazay laestación(Goodman,1994).
En la fase folicular tiene lugar el crecimiento final y lamaduraciónde losfolículosovulatorios.Comoyasehaindicado,losfolículospreovulatoriossecretanaltascantidadesdeE2(Arthur&Pearson,1989),hormonaqueestimulaelcentrohipotalámico generador de pulsos de la GnRH. El aumento de la liberación de

RevisiónBibliográfica
57
GnRHinducirá,asuvez,unincrementoenlafrecuenciadeliberacióndepulsosdeLH,hastaalcanzarunpicodeliberación,conocidocomodescargapreovulatoriadeLH, que actúa como desencadenante de la ovulación. Este pico preovulatorioprovoca cambios en las paredes del folículo, determinando su ruptura y laconsiguienteliberacióndeloocito,18-24horasmástarde(Chemineauetal.,2010;Bartlewskietal., 2011). Laovulaciónmarca la transiciónde la fase folicular a laluteal(Gordon,1996).
FaselutealLafaselutealincluyemetaestroydiestro.Lafasedemetaestroseproduce
inmediatamentedespuésdelestroyeselperiododuranteelcuallascélulasdelagranulosa y la teca del folículo roto proliferan y se transforman en célulasluteínicasque llenanel antrodel folículo (luteogénesis), enunprocesoquedaráorigenauncuerpohemorrágicoy,finalmente,alCL(Keyes&Wiltbank,1988).ElestímulodelaLHesnecesarioparaelcrecimientoinicialdelCLyladiferenciacióncelular.Enestafaseseproduceunamarcadareduccióndelaactividadsecretoradela mucosa vaginal, cervical y uterina (Bartlewski et al., 1999; Niswender et al.,2000).
EldiestroeselperiododelcicloestraldondeelCLmaduracompletamenteyconstituyeuntejidoglandulartotalmentefuncional,secretandoaltascantidadesdeP4.Entre3y4díasdespuésde laovulación,elCLtieneundiámetrode6a8mm,alcanzandosumáximodiámetrode11a14mm,6díasdespués.Esenestemomentocuando la secrecióndeP4porpartedelCLalcanzanivelesmáximosypermanece en valores altos durante toda la gestación, en caso de que ésta seproduzca. La progesterona, junto con los estrógenos basales, mantendrá unfeedbacknegativo sobre lospulsos liberadoresdeGnRHy sobre la liberacióndegonadotropinas(McNeilly,1984).Encasodenoproducirselagestación,entrelosdías 12 y 15 después de la ovulación, las glándulas endometriales uterinassecretanunagenteluteolítico, laprostaglandinaF2α,queestransportadaatravésde lavenauterinay losvasos linfáticoshasta laarteriaovárica,determinando lapérdidadeactividadbiológicaylaatrofialutealdeformaabrupta,produciéndoseunanueva fase folicular (Knickerbockeretal.,1988;Gordon,1996;Bartlewskietal.,1999;Einer-Jensen&Hunter,2005).
1.6.2 Foliculogénesis
Lafoliculogénesiseselprocesoporelqueseproducenelcrecimientoy lamaduración de los folículos, desde su estado primordial (con unos 100 μm dediámetro)hastaelovulatorio(>4ó5mm,dependiendodelatasadeovulación)yque en la oveja tiene una duración aproximada de 180 días. La función de cadafolículo es proporcionar el soporte necesario para que el oocito que contienealcancesumáximopotencialypuedaunirseconelespermatozoideparaproducirun embrión viable. Para ello, las células somáticas del folículo son vitales en el

RevisiónBibliográfica
58
crecimiento y desarrollo del mismo, así como en el control de la maduraciónnuclear y citoplasmática del oocito en los folículos seleccionados para ovular(Cahill&Mauleón,1981).
En el ovario de las ovejas jóvenes existe una reserva de entre 40.000 y300.000 folículos primordiales con 5-6 células de pregranulosa y sin red capilarsanguíneaalguna,quepermanecenenunestadodequiescenciaalaesperadeundesarrolloposterior.Unavezqueseiniciaelcrecimientoyanohayvueltaatrásynopuedenretornaralestadodequiescencia,deformaqueelsiguienteestadiodedesarrollosonlosfolículosprimarios,formadosporoocitosrodeadosdeunacapade células de la granulosa (Driancourt, 1991). Existe una evolución continua defolículos primordiales a folículos primarios y posteriormente, a secundarios opreantrales,queyapresentande2a3capasdecélulasdegranulosa(Driancourt,1991;Fortune,2003).Tambiénseproduceenellosladiferenciacióndelascélulasde la tecaapartirde lascélulasdelestromaovárico,demaneraque formanunacapa de células aplanadas alrededor de la membrana basal del folículo. En unaoveja existenunos4.000 folículos secundarios comprometidos simultáneamente,que tienen diámetros que oscilan entre 0,03 y 0,1 mm; por esta razón, sudesarrollonodependedelasgonadotropinashipofisarias.
Durante esta fase de desarrollo las células de la granulosa adquierenreceptoresFSHylasdelatecareceptoresLH,conloquelaactividaddesíntesisdeesteroidespareceseradquiridaenlafasepreantraldeldesarrollofolicularaunqueno se expresa hasta la fase antral. El desarrollo de estos folículos se basa en unestímulo de la división celular donde, algunos factores de crecimiento, enparticular el IGF y el EGF, producidos en las células de la granulosa y la teca,respectivamente,jueganunpapelmuyactivo(Abecia&Forcada,2011;Bartlewskietal.,2011).
Elsiguienteestadiodedesarrollofoliculareseldefolículosconrespuestaalas gonadotropinas, conocidos como folículos terciarios o antrales, y cuyodesarrollo se ve estimulado por la LH y FSH (Campbell et al., 1995). Estánsometidos a una cierta atresia, de manera que hay a la vez unos 25 folículosantrales,conuntamañode1a2,5mmdediámetro,enunaoveja.Estosfolículostienen receptoresLHen las célulasde la teca, las cualesproducenandrógenosapartir del colesterol, mientras que las células de la granulosa tienen receptoresFSHyproducenestradiolapartirdelosandrógenossintetizadosporlascélulasdelatecaporunprocesodearomatización(Abecia&Forcada,2011;Bartlewskietal.,2011).
Los folículos, a partir de este estadio de desarrollo, se convierten en lafuentemásimportantedesíntesisdeesteroidesováricosduranteelciclosexual,enrespuestaalaLHyalaFSH.Estacapacidadsintéticaaumentaconformelohaceeltamañoylafuncionalidaddelosfolículos.Losfactoresdecrecimiento,asícomola

RevisiónBibliográfica
59
activina y la inhibina producidas en las células de la granulosa influyen en lafuncionalidaddelosfolículosantrales.LaIGFylaactivinapotencianlaacciónFSHen la aromatización,mientras que la EGF la deprime. Paralelamente, la IGF y lainhibinapotencianlaacciónLHde lascélulasde latecamientrasque laEGFy laactivinaladeprimen(Abecia&Forcada,2011;Bartlewskietal.,2011).
Al avanzar el desarrollo folicular, los folículos antrales se transforman enfolículospreovulatorioso folículosdeGraaf,mediante la formacióncompletadelantro y la diferenciación funcional y pasando a convertirse en folículosdependientesdegonadotropinas.Unaovejapuedepresentarentre1y8folículospreovulatorios cuyo tamaño oscila entre 2 y 5 mm de diámetro. Los folículospreovulatorios tienen elevados requerimientos de FSH y esto hace que sea elestadio folicular más vulnerable a la atresia, que se produce cuando lasconcentracionesplasmáticasdeFSHsonmenoresde1ng/mlocuandolasdeLHsonelevadassuperandolos5ng/ml.Enlafasefinaldeldesarrollodelosfolículospreovulatoriosquenosufrenatresia,tienelugarlaexpresióndelosreceptoresLHtambiénenlascélulasdelagranulosadondeLHyFSHactúandemanerasinérgicaenlaactividadaromatasaparaproducirgrandescantidadesdeestradiolenlafasefoliculardelcicloestral(Abecia&Forcada,2011;Bartlewskietal.,2011).
ElnúmerodereceptoresLHenlascélulasdelatecaaumentaparaproducirel sustrato androgénico de la aromatización. En este sentido, los folículospreovulatoriossonlasvíctimasdelaactividaddelfolículoovulatorio,muysensiblealaFSHyconunaaltasecrecióndeE2(quedalugaralcomportamientodeceloyqueprecedealpicopreovulatoriodeLH)ydeinhibina(Webb&Engeland,1982).Esta última en particular ejerce una fuerte retroalimentación negativa a nivelhipofisario,induciendounareduccióndelasíntesisdeFSHpordebajodelumbralcríticodesupervivenciarequeridopor los folículospreovulatorios.Elnúmerodefolículosquealcanzanesteestadiodedesarrolloenelciclosexualdelaoveja,conuntamañodeunos6mmdediámetro,estádeterminadoporlatasadeovulaciónintrínseca de la raza, que habitualmente oscila entre 1 y 2 pero que puedeaumentarconsiderablementeen lasrazasprolíficas(Campbelletal.,1995;Webbetal.,1999).Estefenómenoporelqueelfolículoovulatorioimpideeldesarrollodelosfolículospreovulatoriosseconoceconelnombrede“dominanciafolicular”.
Este folículo ovulatorio puede ser capaz de ovular, siempre y cuando seencuentreenfasefolicularyseproduzca,paralelamente,unpicopreovulatoriodeLH(Driancourtetal.,1988;Price&Webb,1989);encasocontrario,este folículoentraenprocesodeatresia,dandopasoaldesarrollodeotrofolículoreclutadodelgrupodefolículosquerespondenalasgonadotropinas,quereemplazalaanterior(Campbelletal.,1995).

RevisiónBibliográfica
60
1.6.3 Dinámicafolicularenlaoveja
Ladinámicafolicularesunprocesocontinuodecrecimientoyregresióndefolículosantrales,quefinalizaenlaovulaciónoenlaatresia(Lucyetal.,1992).
Eldesarrollode laecografía transrectaladaptadapara laespecieovinahapermitido observar que la selección y crecimiento de los folículos se realiza deforma continua a lo largo del ciclo sin evidencia firme de presentar un patrónorganizado de crecimiento y regresión durante el mismo (Schrick et al., 1993;Lopez-Sebastian etal., 1997; Adams, 1999). Estudios posteriores han descrito eldesarrollodeondasprimarias y secundarias, que se sucedende forma continua.Lasondasprimariassonaquellasenlasqueseevidenciaunfolículodominanteytienen lugaren la fase luteal tempranayen la fase foliculardel cicloy lasondassecundarias, aquellasdondenohaypresenciadeun folículodominanteyque seproduciríandeformacontinuadurantelafaselutealmedia(Adams,1999;Vinoleset al., 1999; Evans et al., 2000; Evans, 2003). Cada una de estas ondas estácompuesta por tres etapas: reclutamiento, selección y dominancia (Goodman &Hodgen,1983).
Elreclutamientoeslacapacidaddeungrupodefolículospararesponderala acción de las gonadotropinas hipofisarias, en particular, los patrones desecreción de FSH son los que determinan los procesos de crecimiento yreclutamientofoliculares(Pictonetal.,1990;Driancourt,1991;Adamsetal.,1992;Webbetal.,1999).
Laseleccióneselprocesoporelcualalgunosdelosfolículosreclutadossoncapaces de continuar su crecimiento hasta estadios preovulatorios mientras lamayoríaentranenatresia(Webbetal.,1999).Esteprocesoloinicianlosfolículosencrecimiento,quesecretanE2,inhibinayfolistatina,provocandoundescensoenla liberación de FSH (Gibbons et al., 1997; Gibbons et al., 1999; Padmanabhan,2002).
Ladominanciaeslaetapaenlaqueculminaelprocesodeselección,dondeunoounospocosfolículosadquierenmayortamañoycaracterísticasfuncionalesquepermitenaumentarsupropiocrecimientoalavezqueinhibenelcrecimientodelosrestantes(Webbetal.,1999).
Esta etapa puede ser ejercida a dos niveles. El primero se denominasistémico,indirectoopasivo,yseejerceatravésdelareduccióndelosnivelesdeFSH hasta niveles muy bajos; los folículos no dominantes entran en atresia ydesaparecen. Este efecto se produce por un mecanismo de retroalimentaciónnegativa que ejercen la inhibina y el E2 secretados por los folículos de mayortamaño (Martin et al., 1988; Driancourt, 1991). El segundo se denomina local,directooactivo,yconsisteenunefectoinhibitoriodirectosobreelcrecimientode

RevisiónBibliográfica
61
los folículos menos desarrollados, a través de la secreción de sustancias en lasangrequereducenlasensibilidaddeestosfolículosalaFSH(Gonzalez-Bulnesetal.,2004).
Las oleadas de crecimiento folicular aparecen durante el ciclo estral,durante el anestro estacional y durante la gestación. Normalmente la ovejapresenta tres oleadas de crecimiento y 4 oleadas en la cabra, aunque estánpublicadas variaciones entre ciclos (Menchaca et al., 2010). En este sentido,Gintheretal.,en1995,señalanunnúmeromediode4ondasfolicularesenovejasde la razaPolipay;otrosestudios comoeldeLeyvaetal., en1998,describenunnúmeromenordeondas(2enel21%delosanimalesy3enel79%)enovejasdelarazaSuffolk(Gintheretal.,1995;Leyvaetal.,1998).
CadaoleadadecrecimientofolicularestáprecedidadeunincrementoenlaconcentracióndeFSH.Traselreclutamiento,normalmenteunoyavecesdosotresfolículos continúan creciendo y adquieren un diámetro ≥5mm,mientras que elrestode folículosmedianos(4-5mm)ypequeños(2-4mm)entranenatresia.Elfolículo/smásgrandeovulaoregresayunanuevaoleadadecrecimientoemergeconunintervaloentreoleadade5días(rango3-7d)(Menchacaetal.,2010).
1.6.4 Ovulación,crecimientoydesarrollodelcuerpolúteo
Laovulaciónesunprocesoconsistenteenlarupturadelapareddelfolículoovulatorioy la liberacióndesucontenido, incluyendoeloocitomaduro,paradarlugaraunCL(Pineda,1989).
Enelfolículoovulatoriopuedendistinguirsecincocapascelulares,queson,desdelamásexternaalamásinterna:lasuperficieepitelial,latúnicaalbugínea,latecaexterna,latecainternayelestratogranuloso,queconstadeunascincoasietecapas de células, excepto en donde las células forman una base para lasustentación del oocito denominada cumulus oophorus. Asimismo, existe unamembranabasalqueseparaalascélulasdelagranulosadelatecainterna(Espey,1994).
Eloocitomadurosecretaunfactorquepermitelaexpansióndelascélulasdel cumulus oophorus y precede al proceso de ovulación propiamente dicho.Paralelamente las células de la granulosa empiezan a sufrir cambios funcionalesque reducen considerablemente su capacidad para producir E2. Esto a su vezestimula la expresión de enzimas con capacidad para degradar el colágeno, laelastina,losproteoglucanos,etc.,queparticipanactivamenteenlarupturafoliculary,portanto,en laovulación.Estoscambiosson inducidos,principalmente,por laLH hipofisaria. El inicio de estos pasos previos a la ovulación varía entre lasdiferentes especies demamíferos (Espey, 1994). En el caso de la oveja su inicioocurreentre21y33horasdespuésdelcomienzodelcelo(Stellflugetal.,1997).

RevisiónBibliográfica
62
Encasodequeel folículodominantealcance laovulación,dará lugaraunCL,enunprocesocontinuoeininterrumpidodeluteinización(Smithetal.,1994)querequieredelarápidaconversióndeunfolículopredominantementeproductorde estrógenos a un CL predominantemente productor de P4 (Bowen-Shauver &Telleria, 2003), hormona esencial para la implantación del blastocisto y elmantenimientodelagestación(Keyes&Wiltbank,1988).ElCLsedesarrollaenunproceso en el que las células foliculares de la teca se transforman en célulaslutealespequeñas,mientrasquelasdelagranulosapasanaserlascélulaslutealesgrandes. Además, el CL secreta, en pequeñas cantidades, péptidos tales comorelaxina,oxitocina,vasopresinaeinhibina(Field,1991).
Encasodequenoseproduzcagestación,elCLsufriráregresiónydarápasoa una nueva ovulación. Por ello, la dinámica de crecimiento del CL se describecomounpatróndecrecimientoyregresión(Jablonka-Shariffetal.,1993;Reynoldsetal.,1994;Reynolds&Redmer,1999).Enlaoveja,elCLtieneunpesoinicialde30a40mg,inmediatamentedespuésdelaovulación,yalcanzaunpesode750mgen,aproximadamente,12díasdespuésdelaovulación(Baserga,1985;Reynoldsetal.,1994).Engeneral,elpesodelCLestárelacionadopositivamenteconeltamañocorporalenlasdiferentesespeciesdemamíferos,pero,porelcontrario,existeunarelacióninversaentreeltamañodelCLyelnúmerodeCLs(Reynoldsetal.,1994).
Elmantenimientodelafunciónlutealsedebeprincipalmentealaaccióndela LH y la hormona de crecimiento (GH); sin embargo, existen otras sustanciasinvolucradas, tales comoel IGF-I, laPGE2y laPGI2, que también incrementan laproduccióndeP4(Juengel&Niswender,1999).
Cuando no se establece la gestación, desde el endometrio se inician losprocesosqueconducenalalisisdelCL.EstaseproduceporlaaccióndelaPGF2α,lacualsesintetizayliberaenelendometriouterino.LaproduccióndePGF2αes,asuvez,inducidaporlaoxitocinasintetizadaporelCL,lacualejercesuacciónatravésde receptores localizados a nivel del endometrio materno (Wathes & Lamming,1995).
Estosprocesosque tienen lugardurante la luteolisis coincidencon la fasede crecimiento de una onda folicular, el conjunto de ambas constituyen la fasefoliculardelciclosexual.EstasecaracterizaporunadisminucióndelosnivelesdeP4, que dan lugar a una mayor liberación de LH; necesaria para el desarrollo,maduraciónyovulacióndelfolículoovulatorio.Porelcontrario,sinoseproducelaluteolisis, los altos niveles de P4 mantienen en niveles basales (pulsos de bajafrecuencia)lasconcentracionesdeLH;enconsecuencia,elcrecimientodelfolículoseleccionado se desacelera, sufriendo un proceso de atresia que conduce a laaparicióndeuna fasede reclutamiento folicular yunanuevaondadedesarrollofolicular(Lopez-Sebastianetal.,1997;Adams,1999).

RevisiónBibliográfica
63
1.6.5 Ciclosexualyestacionalidadenlaoveja
En el proceso reproductivo intervienen tres elementos básicos: elhipotálamo,lahipófisisylasgónadas.Estastresestructurasserelacionanentresídemaneramuyestrechaparaasegurarlosdiferentesacontecimientosquetienenlugarenelciclosexualde laoveja.Elobjetivodelciclosexuales la formacióndeoocitosmadurosylapreparacióndelaparatogenitalparainstaurarlagestaciónsise produce la fecundación. Todo ello se consigue mediante el gobiernohipotalámico y la modulación que ejercen otros factores externos, tanto elfotoperiodocomoelnivelnutricional,oincluso,loscondicionantessocialesdentrodelrebaño(Abecia&Forcada,2011).
La mayoría de los mamíferos en la vida silvestre expresan variacionesestacionalesensuscaracterísticasreproductivas,adaptándosealascondicionesyciclos de su medio natural, distribuyendo los nacimientos de una forma más omenos marcada a lo largo del año. Según especies esta denominada“estacionalidad”esdediferente intensidad,siendoespecialmentemarcadaen lasovejasylascabras(Chemineauetal.,2008).
Ladomesticacióndeestasespeciesnohaimplicadounamodificacióndelospatrones estacionales de reproducciónmostrados por estos animales en la vidasalvaje,cuyoobjetivoesqueloscorderosyloscabritosnazcanalfinaldelinviernoo principio de la primavera, el periodomás favorable del añopara garantizar lasupervivencia de la progenie (Chemineau et al., 2008; Abecia et al., 2012). Estocondiciona,yadentrodelossistemasproductivos,cambiosenladisponibilidaddelosproductosfrescosderivadosdelovinoycaprino(carne,lecheyqueso),locualexplica la relevancia que tiene dentro de la producción animal, la característicaestacionalidadreproductivaenambasespecies.
Precisamenteunodelosrasgosmásimportantesdelareproducciónovinaeslaestacionalidad.Lospatronesdeactividadreproductivaenlaovejaadulta,nogestante,estándominadospordosritmosdistintos.Unodeellosesuncicloestralde16-17díasdeduración,conunamediade16,8±0,3días(Acritopoulouetal.,1977). El otro es un ritmo anual de ciclicidad ovárica caracterizado por el cese(anestro) y restauración (estación reproductiva) de ciclos ováricos ovulatorios(Legan&Karsch,1979;Bartlewskietal.,2011).
Peseaelloenelperiododetransiciónentreelanestroylaactividadsexual(finaldeverano), loscicloscortossonbastantecomunes.Estoesdebidoaqueelcuerpolúteo(CL),confrecuencia,involucionaprematuramente5o6díasdespuésdesuformación(Rosa&Bryant,2003).
Deestaforma,lareproducciónenlasovejassigueunpatrónestacional,esdecir,existeunaalternanciaentrelosperiodosdeanestroydeactividadsexual.Laestacionalidadestáreguladaporelfotoperiodooduracióndeluzdiaria,demanera

RevisiónBibliográfica
64
quelareduccióndeladuracióndeldíaestimulalaactividadsexualysuaumentoinduceelanestro(Legan&Karsch,1980).
Las ovejas son capaces de “monitorizar” por símismas los cambios en elfotoperiododiariomediantelasecrecióncircadianademelatoninaporpartedelaglándulapineal.
Lamelatoninaesunahormona,unadecuyas funcioneses la transducciónde la información fotoperiódica al control de las funciones estacionales,particularmente la reproducción (Malpaux et al., 1996). Esta fotorrecepción estransmitida desde la retina, a través de las neuronas, a un área del hipotálamoconocidacomonúcleosupraquiasmático.Deahíalgangliocervicalsuperior,desdedondeseproduceunasinapsiscon lasneuronas inhibitorias,quehacencontactocon los pinealocitos, que son las células de la glándula pineal (Figura 7). Lospinealocitos secretan la melatonina, que es sintetizada y liberada solamentedurantelanoche(Legan&Karsch,1979;Karschetal.,1984).
Figura 7. Esquema de activación/inhibición de la ciclicidad en la oveja mediante la melatonina. Fuente:Pathwaystopregnancyandparturation.Senger,P.L.2005(Senger,2005).
La producción de melatonina está regulada por el fotoperiodo,encontrándoseconcentracioneselevadasdeestahormonaensangresólodurantelashorasdeoscuridad.Lascaracterísticasdelpatróncircadianodelasecrecióndemelatoninavaríanconloscambiosenelciclodeluz-oscuridadalolargodelaño,loque permite que el animal “reconozca” los cambios en la proporción de luz-oscuridad(Malpaux,etal.,1996).Losperiodoslargosdesecrecióndemelatoninaestimulan y regulan la secreción pulsátil de GnRH hipotalámica, induciendo loscorrespondientes cambiosen la secrecióndeLHypromoviendoasí la ciclicidad.

RevisiónBibliográfica
65
Por tanto, es la GnRH la encargada demodular la liberación de gonadotropinashipofisarias y éstas, a su vez, controlan la actividad reproductora estacional(Kennawayetal.,1987;Malpauxetal.,1999).
Aunqueelfotoperiodoeselprincipaldeterminantedelaestacionalidadhayotros factores que pueden influir sobre los patrones reproductivos, como lagenética, las prácticas de manejo y las interacciones sociales (Rosa & Bryant,2003).
Laestaciónreproductivadeestaespecieesunasucesióndeciclosestrales,los cuales empiezan normalmente en verano o a principios de otoño, con unaduracióndecrecientedehorasdesol,yfinalizanalfinaldelinviernooprincipiodeprimavera, cuando los días comienzan a crecer (Abecia et al., 2011). Laresponsabledeestoscambiosesunacomplejacombinacióndeunritmocircanualendógenoconducidoysincronizadopor la luzy lamelatonina, lacualcontrola laactividadpulsátildelasneuronasdeGnRHenelhipotálamopreóptico-mediobasal(Chemineauetal.,2010).
Laduracióndelaestaciónreproductivatieneunaclarainfluenciaracial.Enlamayoríadelasrazasdelhemisferionorteexisteunpicodefertilidadafinalesdelotoño (octubre-noviembre). Por tanto, los mayores porcentajes de partos seregistran a finales de marzo y en abril, el periodo más favorable del año paragarantizar la supervivencia de la progenie (Chemineauetal., 2008;Abeciaetal.,2012).
Las razas de latitudes intermedias, como la merino australiana o lasmediterráneas,presentanunanestrobreve,duranteelcualunaciertaproporciónde las ovejas ovula espontáneamente. En los entornos tropicales y subtropicaleslasovejasnosonestacionales,osonintermitentementepoliéstricas,dependiendolaactividadsexual,deladisponibilidadydelacalidaddelalimento(Rosa&Bryant,2003).
Durante la estación no reproductiva (anestro), no se observan ciclosestralescomotales.Aunquelossignosdecomportamientodelestroylaovulaciónestánausentes,sedannoobstante,cambiosdinámicosenelcrecimientofolicularováricoysuregresiónalolargodelaestaciónnoreproductiva.Elanestrosedebea la imposibilidad de los folículos antrales para crecer ymadurar, lo cual suelesuceder en la fase preovulatoria del ciclo estral. Sin embargo, el desarrolloposterior de estos folículos puede estimularse artificialmente, lo que permite lareproducción durante el anestro o los periodos de transición (O'Callaghan etal.,1994).
Laestacionalidadafectatambiénalaactividadsexualdelosmoruecosque,aunquepuedenaparearseencualquiermomentodelaño,sufrenundescensodelaeficaciareproductivafueradelaestaciónnaturaldebidoalafaltadelibidoyauna

RevisiónBibliográfica
66
menor cantidad y calidad del eyaculado durante el periodo no reproductivo(Gerlach&Aurich,2000).
Este patrón de reproducción estacional, tiene como resultado un claroperiodo concentrado de partos y, si los animales están en ordeño, un patrónestacional de producción de leche y carne. Tal situación causa, lógicamente, unpatrónestacionaldepreciosdelosproductos,quesonmuybajoscuandolaofertadecarne/lecheesalta(generalmente,desdefinaldeprimaverahastaprincipiosdeotoño)yviceversa.
1.7 Manejoycontroldelareproducción
Laplanificaciónreproductivaesfundamentalenlaexplotaciónovinayaquetodoslosdemásfactoresproductivosydemanejovanadependerdelamismaysevan a organizar en función del calendario reproductivo que se elabore. Laprevisión del mismo va a permitir adelantarse a la compra de materias primasparalaalimentacióndelosanimales,preverlacontratacióndemanodeobraextra,conocerdeantemanolosmomentosdeventay,porlotanto,controlarlaspartidasdeingresosygastosalolargodelaño(DazaAndrada,2002).
Deaquí,esobviodeducirquesetratadeunaoportunidaddemejoradelarentabilidad en este tipo de explotaciones, ya que la homogeneización,consiguiendo producciones “fuera de estación”, permite que se puedan obtenerprecios más altos durante el invierno. En consecuencia, aquellos ganaderoscapacesdeponerenelmercadoproductos“fueradeestación”,sebeneficiandelospreciosmásaltosquerigenduranteel invierno,aumentandoasí surentabilidad,objetivo que se consigue minimizando o revirtiendo los efectos de laestacionalidad reproductiva mediante el uso estratégico de tecnologíasreproductivas y, en concreto,mediante la inducciónde ciclos estralesdurante laestacióndeanestro(Chemineauetal.,2008;Abeciaetal.,2012).Es tambiénunaherramientaesencialparalaintensificacióndelaproducciónovina(Hernándezetal.,2011).
Elmanejoreproductivoenelovinopuedeclasificarseennatural(mediantela alteración del fotoperiodo, el flushing o el “efecto macho”) o farmacológico(usando distintos productos hormonales, como melatonina, prostaglandinas oprogestágenos)(Haresign,1981).
La alteración del fotoperiodo implica la exposición de las ovejas a unaduracióndeldíareducidaartificialmentetrasunperiododeduraciónprolongada.Siseusasoloaceleraráel iniciodelperiodoreproductivo,aunqueconresultadosvariablesyconunaextensión impredecibleconrespectoal iniciode laciclicidad(Legan&Karsch,1980).

RevisiónBibliográfica
67
Elflushingconsisteenaumentarelplanodenutricióndelasovejas(ingestadeproteínayenergía)aproximadamente3-4semanasantesdelinicioplaneadodela estación reproductiva. Las ovejas con una condición corporal que estámejorandosebeneficiandeunamayortasadeovulacióny,portanto,deunamejorfertilidad.Porello,suelenrespondermejorcuandotienenunacondicióncorporalintermedia (2,5-3,5). Este método se utiliza normalmente para mejorar laprolificidad y la fecundidad, pero no con el objetivo de inducir y sincronizar elestro(Haresign,1981).
El efecto macho consiste en la introducción de moruecos a un grupo deovejasquehanestadoapartadasdelosmachosvariassemanasantes(almenos4semanas), este efecto mediado por las influencias sociales (quimiosensoriales,táctiles o visuales) tienen efectos potentes sobre la función reproductiva envariedad de especies. Los moruecos pueden estimular la secreción degonadotropinasy laovulaciónen laovejaenanestroa travésdeéstosestímulos(Delgadilloetal.,2009).
Encuantoalmanejofarmacológicodelcicloreproductivo,éstesebasaenlaadministracióndehormonasexógenasquemodifican lospatrones reproductivos(Thimonier,1981;Abeciaetal.,2011),bienmanteniendolafaselutealmedianteeluso de P4 y/o sus análogos progestágenos (Vilarino et al., 2013), bieninterrumpiéndolaconPGF2αoanálogossintéticos(Pope&Cárdenas,2004),obienmodificando el patrón anual de reproducción con melatonina (Malpaux et al.,1997).
Lafasefoliculartambiénpuedesermodificadacongonadotropinacoriónicaequina(eCG)administradaaltiempoquecesalaexposiciónalaP4,loqueaumentala velocidad del desarrollo folicular y la ovulación,mejorando así los índices defertilidad(Blaschietal.,2014).
Los protocolos basados en hormonas han sido ampliamente empleadospara incrementar el rendimiento reproductivo y la productividad en la oveja. Eluso de la inseminación artificial a tiempo fijo (IATF) permite asimismo lasincronizacióndepartos, laorganizaciónde lotesdecorderos según lademandadelmercado,uniniciomástempranodelapubertad,yunamejoradelafertilidad(Abeciaetal.,2012;Blaschietal.,2014).
Elmétodonaturalesmásbarato,perodalugaraunasincronizaciónmenosexacta y sólo es de utilidad en ciertas condiciones. En cambio, los métodosfarmacológicossoneficacespara lasincronizaciónexactadelestroen lamayoríade las situaciones, pero con la desventaja del coste del producto y suadministración.Detodasformas,alasegurarunasbuenascifrasdeproducción,sepuederentabilizarsiseutilizanparahacerunacubriciónenprimaverayasítenerlaposibilidaddevender loscorderosa finalesdeaño,queescuandoalcanzanelpreciomásalto(Abeciaetal.,2012).

RevisiónBibliográfica
68
1.7.1 Progesterona,susanálogos(progestágenos)yeCG
Losmétodosqueutilizanprogesteronaosusanálogosestánbasadosensusefectossobrelafaselutealdelciclo,simulandolaaccióndelaprogesteronanaturalproducida por el CL, es decir, controlar la secreción de LH desde la pituitaria(Hansel&Convey,1983b).
Losprimerostratamientosparacontrolarelciclosexualdelaovejafueronpublicadosafinalesdeladécadadelos40(Dutt&Casida,1948)yconsistíanenlaadministración diaria de inyecciones subcutáneas de 10 mg de progesterona,diluidosen2mldeaceitedemaíz,durante14días.Eltratamientoredujoelrango,dentro del cual, los animales se apareaban a los 8 días (O'Mary etal., 1950). EninvestigacionesposterioresseadministróeCGygonadotropinacoriónicahumana(hCG) además del tratamiento con progesterona. El control óptimo se alcanzócuandolaeCGfuedadaenelmomentodelaúltimainyeccióndeprogesteronaylahCG 24 horasmás tarde. Con este tratamiento lamayoría de las ovulaciones seprodujeron entre 20 y 28 horas después de la inyección de hCG. Sin embargo,ningunadelasovejasmostrócomportamientodeceloenestemomento(Bradenetal.,1960).
En1962,Southcottetal.usaronconéxitounanálogodelaprogesterona,6-methyl-17-acetoxyprogesterona,para inducir y sincronizar el estro enovejas. Suhipótesis era que un agente progestágeno cuya actividad cayese de manerarelativamente brusca después de finalizar el tratamiento debería dar mejoresresultados,yaquelostrabajosanterioresteníanbajasfertilidadesdebidoalefectopersistentedelaprogesteronaenelambienteuterinoyoviductal(Southcottetal.,1962). Además, se descubrió la vía de administración intravaginal de laprogesterona o sus análogos para facilitar la supresión inmediata de estashormonas(Abeciaetal.,2011).
El grado de sincronización obtenido y el intervalo entre el final deltratamientoyeliniciodelestrodependendelproductousadoy,engeneral,sonlosmásutilizadosenépocasdeanestro.Losprogestágenospuedenadministrarsededistintas formas (esponjas, implantes, etc.), por distintas vías (intravaginal,intramuscular,subcutánea)yadistintasdosis(Henderson&Robinson,2007).
Los dispositivos intravaginales impregnados con progestágenos, se hanusado desde principios de los años 60 para sincronizar ciclos estrales en ovejas(Robinson,1965).Existendosmétodosusadoscomercialmenteparaeste tipodeadministración: lasesponjasdepoliuretanoimpregnadasconprogestágenos;yeldispositivo interno de liberación controlada de fármacos (CIDR; ControlledInternalDrugRelease),diseñadoenNuevaZelandaafinalesdelosaños80yquecontiene0.30gdeprogesteronanatural(Welchetal.,1984;Abeciaetal.,2012).

RevisiónBibliográfica
69
Lasesponjasvaginalessemantienenencontactoconelcérvixpredisponenaunincrementodelamicrobiotabacterianaquepuedecausarvaginitisyformaradherencias que ocasionen problemas reproductivos en el rebaño (Amiridis &Cseh, 2012). Sin embargo, con el uso del CIDR, fabricado con un elastómero desilicona inerte, la presentación de vaginitis e infecciones vaginales son casiinexistentes(Tondello,2010).
Enovejas,lasesponjasintravaginalesolosCIDRssemantienenduranteunperiodo de 12-14 días. Las ovejas entran en estro aproximadamente 48 horasdespuésde retirar el dispositivo (Wheatonetal., 1993). Paraque el tratamientoprogestágeno sea efectivo es necesario que haya suficiente gonadotropinadisponible para iniciar los eventos preovulatorios, incrementando los niveles degonadotropinas endógenas con FSH exógena proporcionada a partir de laadministraciónde eCG en elmomentode la retiradadel producto (Powelletal.,1996).
Una posible alternativa paraminimizar el tiempo de permanencia de losdispositivosvaginales,es lasincronizacióndelestroy laovulaciónmedianteunamenor exposición al tratamiento progestágeno (Ungerfeld & Rubianes, 1999).EstostratamientosconsistenenlainsercióndeesponjasoCIDRs,impregnadosenprogestágenos,durante6días.EsteperiodoesmáscortoquelavidamediadeunposibleCLenelovario,porlotanto,esnecesarioaplicarunadosisúnicadePGF2α,osusanálogos,simultáneamentea la insercióndelaesponja,parainducir la lisisdel CL. La respuesta a este tratamiento es similar a la obtenida con lostratamientosdelargaduración(Letelieretal.,2009).
Elúnicodispositivo intravaginal comercializadoenEspañasepresentaenforma de esponjas impregnadas en progestágenos. Estas esponjas contienenacetatodemedroxiprogesterona(MPA)oacetatodeflugestona(FGA),ambosmáspotentesquelaprogesteronanatural.Enelcasodelacetatodeflugestona(FGA)ladosis empleada es de 20 ó 30 mg/esponja, mientras que en el acetato demedroxiprogesterona(MPA)seemplean60mg/esponja(Letelieretal.,2009).
El FGA es aproximadamente 20 veces más potente que la P4 y exhibeactividad progestágena pero no propiedades anti-progestágenas, anti-androgénicaso androgénicas junto conunabaja actividadglucocorticoide.Comoresultado de su unión a receptores de progesterona, el FGA actúa porretroalimentación negativa sobre el eje hipotálamo-hipofisario, suprimiendo laliberación de gonadotropinas desde la pituitaria y por tanto, el crecimientofolicularterminaly laovulación(Abeciaetal.,2011).En laactualidad,sepuedenencontraresponjasdeFGAadosdosisdiferentes:Sincropart®(30mgacetatodeflugestona,CevaSaludAnimal,Barcelona,España)yChronogest®(20mgacetatodeflugestona,MSDAnimalHealth,Salamanca,España).

RevisiónBibliográfica
70
ElMPAesaproximadamente10vecesmáspotentequelaP4,sumecanismode acción, al igual que el FGA, es la inhibición de los factores liberadores degonadotropinas hipofisarias a nivel hipotalámico. La consecuencia es unainhibiciónde la liberacióndeLHyFSH,sumadoa ladisminuciónde larespuestaováricadelasmismas,dandocomoresultadoelefectoanovulatorio(Abeciaetal.,2011). En la actualidad sólo existe una forma comercial de esponjas de MPA,Esponjavet®(60mgacetatodemedroxiprogesterona,Hipra,Girona,España).
Los niveles sanguíneos de progestágenos son elevados durante lapermanencia de la esponja, normalmente durante 12-14 días, para imitar laduracióndeunCLfuncionalnormal.Trassuretirada,lacaídabruscadelniveldeprogestágeno pone enmarcha los acontecimientos hormonales que conducen alceloy laovulación,siempreque lasovejasseencuentrensexualmenteactivas.Sinoestánsexualmenteactivas,eltratamientoestimulanteconprogestágenosdebeser reforzado con tratamientos estimulantes para los folículos, como eCG, en elmomento de la retirada de las esponjas, para inducir el crecimiento folicular, elestroylaovulación(Abeciaetal.,2011).
Elacetatodemelengestrol(MGA),otroprogestágenosintético,tambiénhasido usado para inducir el estro en ovejas y cabras. Se han propuesto algunosprotocolosparasuadministración,perotodosloincluyenenlaalimentación,bienmezclado en la ración total o como suplemento alimenticio cada 8-12 horas,normalmenteduranteunperiodode8-14días.Ladosisdiariamásusadaes0.25mg/animal/día,lamayoríadeloscasos,dosvecespordía(0.125mgx2)(Powelletal.,1996).Enunestudiorealizadoenlosaños70porinvestigadoresnoruegos,selogróqueel89%delasovejasentraranenceloenlos6díassiguientesyel74%deellasquedaranpreñadasy,aunquelosautoresmanteníanqueestosresultadoseran mucho mejores que la mayoría de los anteriores, incluyendo las esponjasintravaginales,variasconsideraciones ligadasa ladosificaciónoral, talescomoeltiempo, el trabajo, y el coste, hacen que la aplicación de estos métodos seacomercialmenteimposible(Velle&Helle,1979).
La eCG es una glicoproteína compleja, que, desde el punto de vistafarmacodinámico, presenta una alta actividad tanto FSH como LH. Gracias a suactividadFSH,laeCGestimulaelcrecimientodelascélulasintersticialesdelovario,así como el crecimiento y maduración de los folículos. Por su actividad LH estambién capaz de desencadenar la ovulación actuando directamente sobre elovario. En el macho, la eCG estimula el desarrollo del tejido intersticial de lostestículos y de las glándulas sexuales anejas. La eCG, aplicada después de untratamientoconprogestágenos,favoreceeliniciodeunafasefolicular,laaparicióndelceloyeldesencadenamientodeovulacionessincronizadas.
LadosisdeeCGvaríadesde250a500UI,dependiendodelaedad(250-300UI en corderas, 350-500UI en ovejas adultas), la época del año (400-500UI en

RevisiónBibliográfica
71
anestro,300-350UIdurantelaépocadereproducción),ylaraza(dosismásbajasenrazasprolíficas).Paraelapareamientodelosanimales,elratiomacho:hembrarecomendadoes1:10enépocareproductiva,aunquepuedesermásbajo (1:5-7)fuera de temporada. La inseminación puede realizarse desde las 47 horas, en elcasodeinseminaciónintrauterina,hastalas55horas,enelcasodeinseminacióncervical,despuésderetirareldispositivo(Naim,2009).
Los factores más importantes a tener en cuenta antes de decidir quémétodousar son: el gradode sincronizaciónnecesario, la estacióny los factoreseconómicosydemercado.Sinembargo,elresultadodelascubricionesdependeráde diversos factores, en particular de la habilidad y fertilidad de los machosdurante la época de anestro. Por eso es conveniente, fuera de la época dereproducción, utilizar implantes de melatonina en los machos, para que losresultados en cuantoa las tasasde concepciónydepartosno se veanafectados(DeNicoloetal.,2008).
Lasovejasqueseencuentranenanestro,alceloinducidolesiguesolootrocelo,16-18díasdespuésdelaretiradadeldispositivo,despuésdeesteyanotienenlugarmásceloshastalaépocareproductiva(Abeciaetal.,2011).
1.7.2 ProstaglandinaF2αysusanálogos
La prostaglandina F2α y sus análogos sintéticos han sido estudiadosampliamentedesdesudescubrimientoen1970comounpotenteagenteluteolíticoenrumiantes(McCrackenetal.,1972).
La administración exógena de PGF2α o sus análogos de síntesis se utilizapara inducir una luteolisis controlada, de manera que puede usarse parasincronizar el estro en las ovejas cíclicas. El efecto luteolítico da lugar a laregresión del CL y a un descenso de las concentraciones de P4 en sangre. Elincremento en la cantidad de gonadotropinas hipofisarias estimula el desarrollofolicular.Elceloaparecealos2-3díasylaovulaciónunas24horasmástarde.Losanálogos de prostaglandinas más comúnmente usados en veterinaria son elcloprostenolyelluprostiolyunprotocolohabitualenovejasycabrases125μgdecloprostenolo7,5mgdeluprostiol.Sinembargo,paraserefectivo,debeaplicarseen presencia de CL, de forma que los animales que estén en anestro noresponderánaltratamiento(McCrackenetal.,1970).
Los tratamientos conPGF2α tienen la ventaja de ser efectivosmediante laadministraciónporvía intramuscular,mejorandoelbienestaranimal (Douglas&Ginther, 1973). Además son de rápida y casi completa metabolización en lospulmonesy,porconsiguiente,noseacumulanenlostejidos(Piperetal.,1970).
ElprincipalinconvenientedelusodePGF2αesque,paraquelahormonaseaefectiva,esnecesarioaplicarlaenpresenciadeunCLactivoquerespondaalefectodePGF2α(Rubianesetal.,2003).LaadministracióndePGF2αentrelosdías3y14

RevisiónBibliográfica
72
delcicloinduceluteolisis,seguidadeestroyovulación(Acritopoulouetal.,1977;Acritopoulou & Haresign, 1980; Rubianes et al., 2003). Teniendo en cuenta laimposibilidaddeconocerlafasedelcicloestralenungrupodehembras,sehacenecesarialaadministracióndedosinyeccionesdePGF2αconunintervalode9-10días.Deesamanera,casitodoslosanimalesdelgrupoestaránenmitaddelafaselutealalasegundadosisyresponderánaltratamiento(Abeciaetal.,2011;Abeciaetal.,2012).
OtrodelosinconvenientesdeesteprotocolodedobleinyeccióndePGFesque la fertilidad de las ovejas en la primera monta es solamente del 70%,significativamente más baja que la obtenida con los tratamientos conprogestágenos (Boland et al., 1978; Godfrey et al., 1999). Este problema puededeberse a que el intervalo de9-10días asegura la presencia deCL a la segundainyecciónenlamayoríadelosanimales,perosupresenciaalteralafuncionalidad,la maduración final de los folículos preovulatorios y la luteogénesis normal.También es debido a la variabilidad en el tiempo de ovulación después de laluteolisis inducida (Barrett et al., 2002; Gonzalez-Bulnes et al., 2005). De igualformalaexistenciadealtasconcentracionesdeP4durantelafaselutealmediadelciclodisminuyelasecrecióndeLH.Estadisminuciónesnecesariaparacrecimientofinal y la maduración de los folículos preovulatorios (Campbell et al., 1995;Gonzalez-Bulnesetal.,2004).
1.7.3 Protocolos hormonales más utilizados en programas de montanatural
Prostaglandina+efectomachoEl tratamiento consiste en dos dosis de PGF2α o sus análogos,
administrándolos con 10 días de diferencia, y exponiendo a las ovejas al efectomachocoincidiendoconlaaplicacióndelasegundadosis.Elratiomacho:hembrarecomendadoes1:10.Lasovejasestaránenceloparasercubiertasporelmachoobienparalainseminaciónaproximadamente48horasdespuésdelasegundadosisde PGF2α. La inseminación puede realizarse entre 48 y 55 horas después de lasegunda inyección pudiéndose obtener ratios de fertilidad entre 44% y 62,5%(Abeciaetal.,2011).
LacombinacióndePGF2αenlafaselutealtempranacombinadaconelefectomachopuedeserunaalternativaadecuadapara lasincronizacióndelestroantesde la inseminación sin la previa detección del celo. El efecto macho se usacomúnmente para inducir un pico de LH, estro y ovulación durante la época detransición, sin embargo, también induce un incremento en la secreción de LHduranteelperiodoovulatorio,enovejascíclicas,tratadasconprogestágenosyconprostaglandinas (Evans et al., 2004; Hawken et al., 2007; Contreras-Solis et al.,2009).

RevisiónBibliográfica
73
Progesterona+eCGLos protocolos basados en el uso de progestágenos son los más
comúnmenteusadosentodoelmundo(Whitley&Jackson,2004).
El más habitual, en ovino, es la aplicación de esponjas intravaginalesinsertadasdurante12ó14días.Elestroseproducirá,aproximadamente,48horasdespuésdelaretiradadelosdispositivos.EltratamientoincluyelaadministracióndeeCG,coincidiendoconlaretiradadelasesponjas;ladosisdeeCGvaríade250a500UIdependiendodelaedad(250-300UIencorderasy350-500UIenovejas),la estación (400-500 UI durante el anestro y 300-350 durante la estaciónreproductiva)y laraza(menosdosisenrazasprolíficas).Durante lacubrición,elratiorecomendadoesdeunmachoporcada10ovejasenépocareproductiva,perodeberíasermásbajo(1:5-7)en“contraestación”.Lainseminaciónartificialpuedeser llevada a cabo desde las 47h (intrauterina) hasta las 55h (cervical) tras laretiradadelosdispositivos(Abeciaetal.,2011).
Progesterona+ProstaglandinasUnaestrategiaparareducirlaexposicióndelosanimalesalosdispositivos
intravaginalesasí comoa los tratamientos conprogestágenoses, evidentemente,acortarelperiododetratamiento.ParaqueeltratamientoconP4oprogestágenossea efectivo, éste debe ser más largo que la fase luteal. Así, durante el periodoovulatorio, es necesario eliminar el CL para reducir el tiempo de exposición.NormalmenteseempleaPGF2αosusanálogosparaesefin(Abeciaetal.,2011).
Se inserta el dispositivo liberador de progestágeno (esponja o dispositivosubcutáneo),sintéticoonatural,con30a60mg,puedeserdeMAPodeFGA.Eldíade la inserción del dispositivo es considerado el día 0. En caso de usar undispositivo subcutáneo, debe permanecer durante 9 días. La esponja o el CIDRpermaneceen lavaginaentre9y14díasen lugarde18,mientras seaplicaunasola dosis dePGF2α el día 9. El estro se presenta entre las 36 a 48horas tras laaplicación de PG y la ovulación 10 horas más tarde. La fertilidad tras lainseminación con semen congelado es mejor si se compara con los protocolosclásicos (61% ante un 57%). En el presente, el tiempo de permanencia de losdispositivos intravaginales puede reducirse más, de 5 a 7 días. El efecto en elovario es similar a los protocolos clásicos y la fertilidad puede verse mejorada(Peralta,2007;Tondello,2010;Abeciaetal.,2011).
Progesterona+Prostaglandina+eCGEn este protocolo de sincronización el dispositivo con el progestágeno se
utilizadelmismomodoqueenlosdosprotocolosanteriores.Dosdíasantes(48h)deretirareldispositivo,seadministrandosisúnicasdeeCGyPGF2α(250a400UIy10mg,respectivamente)ambasporvíaIM.Estopermiteunperiododeactividadmáslargoyunmejorreclutamientoymaduracióndelosfolículosylosoocitos.El

RevisiónBibliográfica
74
estro se presentará en 24-48 horas y la ovulación de 10 a 12 horas después deiniciadoslossignosdeestro(Suárez,2010).

JUSTIFICACIÓNYOBJETIVOS


JustificaciónyObjetivos
77
2 Justificaciónyobjetivos
La eficiencia reproductiva es un punto clave en la eficiencia productivaglobaldelasexplotacionesovinas,comoenelrestodeexplotacionesganaderas,yaque facilita lamejora de las producciones por selección de reproductores y poraumentode la fertilidadyprolificidadde losrebaños.Porello,esabsolutamentenecesariounadecuadomanejoreproductivo.Concretamente,enelganadoovino,las actuaciones fundamentales son el control de los periodos de monta y laaplicación de inseminación artificial. El control de los periodos de monta tienecomoobjetivoproducirlotesdecorderoshomogéneosparaabastecerlademandadel mercado, superando la limitación estacional en la oferta del producto yaumentandosuvaloreconómico.Porotrolado,lainseminaciónartificialfacilitalaintroducciónydifusióndemachosmejorantesylainterconexiónentrerebaños.
En la consecución de estos objetivos se hace necesaria la aplicación deprotocolosdereproducciónasistidaqueconllevanconfrecuencialautilizacióndetratamientos de inducción y sincronización de celos y que están basados,generalmente,enlautilizacióndehormonasexógenas.
Lasincronizacióndelcelo,enelcasodelospequeñosrumiantes,serealizahabitualmente mediante la manipulación de la fase luteal del ciclo sexual. Laprogesterona, o sus análogos, son las hormonas más utilizadas para lasincronización de estros en pequeños rumiantes. Habitualmente, se utilizantratamientos con progestágenos, que se aplican mediante dispositivosintravaginalesconunaduracióndeaproximadamentedossemanas,yaquedebensuperarlavidamediadeunposiblecuerpolúteoenelovario.
En la actualidad, el uso de estos protocolos se ve condicionado por unamayorsensibilidadgeneralporpartedelasociedad,respectodelbienestardelosanimales empleados en producción, que percibe de forma negativa el uso deesponjasvaginalesduranteperiodosdetiempolargos.Aestohabríaquesumarleque los consumidores, anivelmundial, están comenzandoademandar, cadavezmás, alimentos libres de residuos farmacológicos. De hecho, cada día crece elnúmero de estados que exigen a sus productores, importadores y exportadores,querespetenprácticassosteniblessinusodehormonassintéticas.Porello,en laactualidad, el empleo de progestágenos para sincronización del celo en diversasespeciesdomésticasestásiendoobjetoderevisión.EnelcasoconcretodelaUniónEuropeaseestá limitandoelusodeestas sustanciasporgenerar residuosen lascanales de animales (Unión Europea, 2010) (Reglamento (UE) 37/2010 de laComisión,de22dediciembrede2009).
Estos condicionantes implican lanecesidadde llevar a caboprácticasqueminimicenoevitencompletamenteelusode tratamientoshormonales,evitando,así, riesgos potenciales para la salud humana, originados por los residuos

JustificaciónyObjetivos
78
presentesenproductosdeorigenanimal,ymejorando,enloposible,elbienestaranimal.
Enbaseaestasconsideraciones,eldesarrolloexperimentalde lapresenteMemoria de Tesis Doctoral tiene como objetivo general la identificación ydesarrollodeprotocolosde sincronizaciónde celosyovulacionesque tenganenconsideración la mejora del bienestar animal y la disminución de residuoshormonales,investigandolosefectosobtenidosalreducireltiempodeexposicióna los progestágenos en los protocolos hormonales y la utilización de un nuevodispositivo intravaginal de progesterona natural (CIDR). La consecución de esteestudio pretende minimizar los riesgos para la salud de los consumidores y, almismotiempo,potenciarlasaludyelbienestardelosanimales,adaptándonosalacrecienteexigenciadelosconsumidores,conproductos“clean,greenandethical”.
Para alcanzar este objetivo general, se plantean los siguientes objetivosespecíficos:
1. Estudiarlarespuestareproductiva(actividadhipofisariayovárica,tasadepresentación de celos, fertilidad y prolificidad) frente a la aplicación deprogestágenos(FGA)enprotocolosdecortaolargaduración.
2. Estudiarlarespuestareproductiva(actividadhipofisariayovárica,tasadepresentación de celos, fertilidad y prolificidad) tras la utilización de dostipos diferentes de dispositivos intravaginales (progesterona natural oCIDRyprogestágenosoFGA),tantoenprotocoloslargos,comocortos.
EstosobjetivossecorrespondenconlassiguientesFasesExperimentales:
1. Fase Experimental 1. Efecto de la duración del tratamiento desincronización de celos con dispositivos intravaginales liberadores deprogestágenoseinfluenciadelaépocareproductivayraza.
2. FaseExperimental2.Evaluacióndelafuncionalidadhipofisariayovárica,así como tasa de presentación de celos, en ovejas tratadas con diferentesprotocolos de sincronización de celo con dispositivos intravaginalesliberadoresdeprogestágenos.
3. Faseexperimental3:Efectodedostratamientosdesincronizacióndeceloclásicos de 14 días de duración, con dispositivos intravaginales conprogesterona natural (CIDR) o progestágenos sintéticos (FGA-esponjas)durantelaépocareproductiva.
4. FaseExperimental4.Evaluacióndelafuncionalidadhipofisariayovárica,asícomotasadepresentacióndecelos,enovejastratadasconunprotocolocorto de 7 días, comparando dos tipos de dispositivos intravaginales deprogestativos, uno en base a progesterona natural (CIDR) y el clásico en

JustificaciónyObjetivos
79
baseaprogestágenossintéticos(FGA),durantelaépocareproductivayencontraestación.


MATERIALYMÉTODOS


MaterialyMétodos
83
3 MaterialyMétodos
3.1 Animales:baseracial
Los animales utilizados procedían en su origen de explotacionescomerciales.Lasfasesexperimentales1,2y4queintegranelpresenteestudiosehanrealizadoutilizandoanimalesdeaptitudcárnica,siendountotalde25ovejasderazalacaune,16ovejasderazasegureñay98ovejaspertenecientesalaF1delcrucedeovejasde lasdosrazasantescitadasconsementalesde larazaprolíficaromanov.Lafaseexperimental3seefectuócon303ovejasadultas,derazalacaunelechera.
3.2 Entornoexperimental:instalacionesymanejo
La Fase Experimental 1, se realizó en condiciones reales de unaexplotacióncomercialdeganadoovinoutilizandounasubpoblaciónde90ovejasintegrada en una población de 600 ovejas de distintas edades (1–8 años) y endistintafasedeproducción(vacías,gestantesyenlactación).
Laexplotaciónreferidaseencuentraubicadaenelnortede laComunidadValenciana, en la comarca de l’Alcalatén (latitud 40,70°N, altitud 360msnm). Setratadeunaexplotacióndeganadoovinoconorientaciónproductivacárnica.Estaganadería desarrolla su actividad en régimen semiextensivo, de forma que losanimales pernoctan en el aprisco y pastan en torno a 8 horas diarias,alimentándoseabasedelospastosdelazona.
Las instalaciones cuentan con un aprisco para los animales que salen alcampo durante el día y para las ovejas que se encuentran en el últimomes degestación,yunanavequealojaalasmadresduranteelamamantamiento.Ambosedificiostienenunasuperficiede1000m²cadaunoytambiénexisteunalmacénde500m²queseparaelapriscodelanave.Asimismo,cuentaconparquesalairelibre y tolvas para administrar el alimento, así como bebederos de canal de 1,5metros, demanera que los animales allí alojados puedan disponer de comida yaguaa libredisposición.Todos losanimalesde laexplotaciónestán identificadosconelsistemahomologadovigentemediantemarcasauriculares(crotales)ybolosruminaleselectrónicos.
El clima en esta zona del interior de la provincia de Castellón es del tipomediterráneoseco,semiárido,yseconsideraunclimadetransiciónentreel tipomediterráneoyeldesértico.Julioeselmesqueregistramenosprecipitaciones,tansólo15mm,mientrasqueoctubreeselmáslluviosocon66mm.Enreferenciaalas temperaturas, el mes más caluroso del año es agosto, con un promedio de23,8°C,mientrasqueelmásfríoesenero,conunamediade8,7°C(climate-data.org

MaterialyMétodos
84
14/1/16). La vegetación espontánea que sirve de alimento a los animales estádeterminadaporestaescasezdeprecipitacionesyporlasequedadestival.
Apartirde los100-150metrosdealtitud, lavegetacióndominanteesunaasociación entre encinar y carrascal (Quercetum rotundifoliae) y pinomediterráneo (Pinus halepensis) y matorral leñoso de porte arbustivo conocidocomocoscoja(Quercuscoccifera).Juntoconlacoscojasedanotrasespecies,talescomo el lentisco (Pistacia lentiscus), el acebuche (Olea europaea), algarrobosilvestre (Ceratonia siliqua), brezo (Erica multiflora), romero (Rosmarinusofficinalis), junza (Cyperus longus), coscollina (Rhamnus alaternus), palmito(Chamaeropshumilis)ytomillo(Thymusvulgaris).
Enlaszonasbajasnohayningúntipodecultivo,exceptuandoelolivo(Oleaeuropea)yelalmendro(Prunusdulcis),porloqueabundavegetaciónadventiciatalcomo lechetrezna (Euphorbia helioscopia), cardo (Carduus nutans), lisones(Sonchus oleraceus), correhuela (Convolvulus arvensis), verdolaga (Portulacaoleracea), bledos (Amaranthus retroflexus), grama (Cynodon dactylon), ballueca(Avena fatua), cañota (Sorghum halepensis), rabaniza (Diplotaxis erucoides),conejitos (Fumaria officinalis), amapola silvestre (Papaver rhoeas) e hinojo(Foeniculumvulgare).
LasFasesExperimentales2,3y4serealizaronenlaUnidaddeOvinodelaGranjaDocenteyde InvestigaciónVeterinariade laUniversidadCEUCardenalHerrera,ubicadaenlapoblacióndeNáquera(latitud39,62°N,altitud242m).
Losanimalesutilizadosenlafasesexperimentales2y4(n=49)pertenecíanalaF1deloscrucesanteriormentedescritosyestabanidentificadosconcrotaleshomologadosybolosruminaleselectrónicos.Paraestasdosfasesexperimentalesseutilizóunaproporción1:1(macho:hembra).
Enlafaseexperimental3seincluyeron303ovejasderazalacaunelechera,cedidasporunaexplotacióncomercialyelratiomacho:hembrasfuede1:5.
Las instalacionesde laUnidaddeOvinode laGranjaDocentecuentanconuna única nave para el rebaño, dividida en dos parques con separacioneseventuales para los grupos experimentales. Ambos parques tienen una zonacubiertayunazonaalairelibreycuentanconcomederosybebederosautomáticospara la administración ad libitum de agua y alimento. Además, en la zona decomederos se dispone de cornadizas autotrabantes y otras instalaciones ymaquinariaparafacilitarelmanejodelosanimales.
El climade esta zona es un clima subhúmedo seco, conunaprecipitaciónmediade570mm,quealcanzaunaclaromáximootoñalcorrespondientealmesdeoctubre.Latemperaturamediaanualde16,7°C,fluctuandoentreunatemperatura

MaterialyMétodos
85
mínimade-4,5°Cregistradaenelmesdeeneroyunamáximade39,0°Cregistradaenelmesdeagosto(climate-data.org14/1/16).
El tipo de explotación es intensivo con estabulación libre. La base de sualimentación está integrada por alfalfa y paja a libre disposición, sin salida apastos.
3.3 Diseñoexperimental
FaseExperimental1.Efectodeladuracióndeltratamientodesincronizaciónde celos con dispositivos intravaginales liberadores de progestágenos einfluenciadelaépocareproductivaydelarazaovina
Elobjetivoesencialdeestaprimerafaseexperimentalfueevaluarelefectode la reducción en el tiempo de tratamiento con progestágeno intravaginal;concretamente,de14a7díasysusposiblesdiferenciasdebidasalaépocadelaño(fotoperiodo)enqueseaplicabaeltratamientoyalaraza.
Paraello,secompararontresmétodosdistintosdesincronizacióndecelos,uno con tratamiento tradicional o “largo” y dos con tratamiento “corto”, en dosreplicados que se diferenciaban en la época del año en que se desarrollaron. Elprimerexperimentoserealizóenfotoperiododecreciente(experimento1.1;épocareproductiva natural) y el segundo en fotoperiodo creciente (experimento 1.2;“contra-estación”reproductiva).
En cada experimento se dividieron los animales, demanera aleatoria, entres lotes distintos, que constituyeron los grupos experimentales,correspondientesacadaunodelostratamientosdesincronizacióndecelos.
Gruposexperimentales:
• GrupoControl (n=30):El tratamientodesincronizaciónempleadofueelprotocolo“clásico”o“largo”,consistenteenlacolocacióndeesponjasintravaginales impregnadas con 20 mg de acetato de fluorogestona(Chronogest®,MSDAnimalHealth,Madrid,España)duranteunperiodode14días.Además,enelmomentodelaretiradadeldispositivo,seadministróuna única dosis de gonadotropina coriónica equina o eCG por víaintramuscular (Foligon®,MSDAnimalHealth,Madrid,España); ladosis seajustósegúnraza.Así,alasovejasderazasegureñaselesadministró400UIyalrestounadosismenor(350UI),porserderazasprolíficasoprocederdecrucesconlasmismas(figura8).• GrupoPG-Corto(n=30)eltratamientoconsistióenlacolocacióndelasesponjasintravaginales(delmismotipopreviamentedescrito)duranteunperiodo “corto”de7días.Esteprotocolohacenecesaria laeliminaciónde un posible CL y, para ello, se administró una dosis de 5 mg de

MaterialyMétodos
86
prostaglandina F2α por vía i.m (Dinolytic®, Zoetis, Madrid, España) en elmomentodelacolocacióndelosdispositivosintravaginales.• GrupoCorto-PG(n=30)eltratamientoconsistióenlacolocacióndelasesponjasintravaginales(delmismotipopreviamentedescrito)duranteunperiodo “corto”de7días.Esteprotocolohacenecesaria laeliminaciónde un posible CL y, para ello, se administró una dosis de 5 mg deprostaglandina F2α por vía i.m (Dinolytic®, Zoetis, Madrid, España) en elmomentodelaretiradadelaesponja.
Finalmente,enambosgrupossellevóacabolaadministracióndeunadosisde eCG en el momento de retirada del tratamiento (mismo producto y mismasdosispreviamentedescritasqueenelgrupocontrol;figura8).
La aplicación del tratamiento progestativo en los grupos de tratamientocortoserealizóunasemanadespuésqueenelgrupocontrol,conelfinderealizarlaretiradadelasesponjasintravaginalesenlostresgruposalmismotiempoyqueasí las condiciones ambientales y de manejo fuesen las mismas durante laaparicióndelceloylaovulación(figura8).
Figura 8. Protocolos de sincronización de celo en base a dispositivos intravaginales de progestágenos deduración“clásica”(grupoControl)o“corta”(gruposPG-CortoyCorto-PG),utilizadosenlaFaseExperimental1
Enelmomentodelaretiradadelasesponjas,seintrodujeronlosmachosenelrebañodeovejas,conunaproporciónde1machoporcada3hembras,yéstossemantuvieron conellasdurante48horas,momentoenque se retiraronparaquereposaran hasta la cubrición del siguiente lote. Los machos se estimularonpreviamente, cada uno de ellos, con tres implantes subcutáneos de melatonina(Melovine®,CevaSaludAnimal,Barcelona,España),60díasantesdeempezarlascubriciones. El porcentaje de gestación se valoró mediante ecografíatransabdominal entre los días 35-45 de la supuesta gestación, utilizando un
Día1
Día2
Día3
Día4
Día5
Día6
Día7
Día8
Día9
Día10
Día11
Día12
Día13
Día14
insertar esponjas
intravaginales
retirar esponjas
intravaginales
administrar eCG +
insertar esponjas
intravaginales
retirar esponjas
intravaginales +administrar
eCG +administrar PGF2α
insertar esponjas
intravaginales
retirar esponjas
intravaginales +administrar
eCG +administrar PGF2α
GRUPO CONTROL GRUPO PG-CORTO GRUPO CORTO-PG
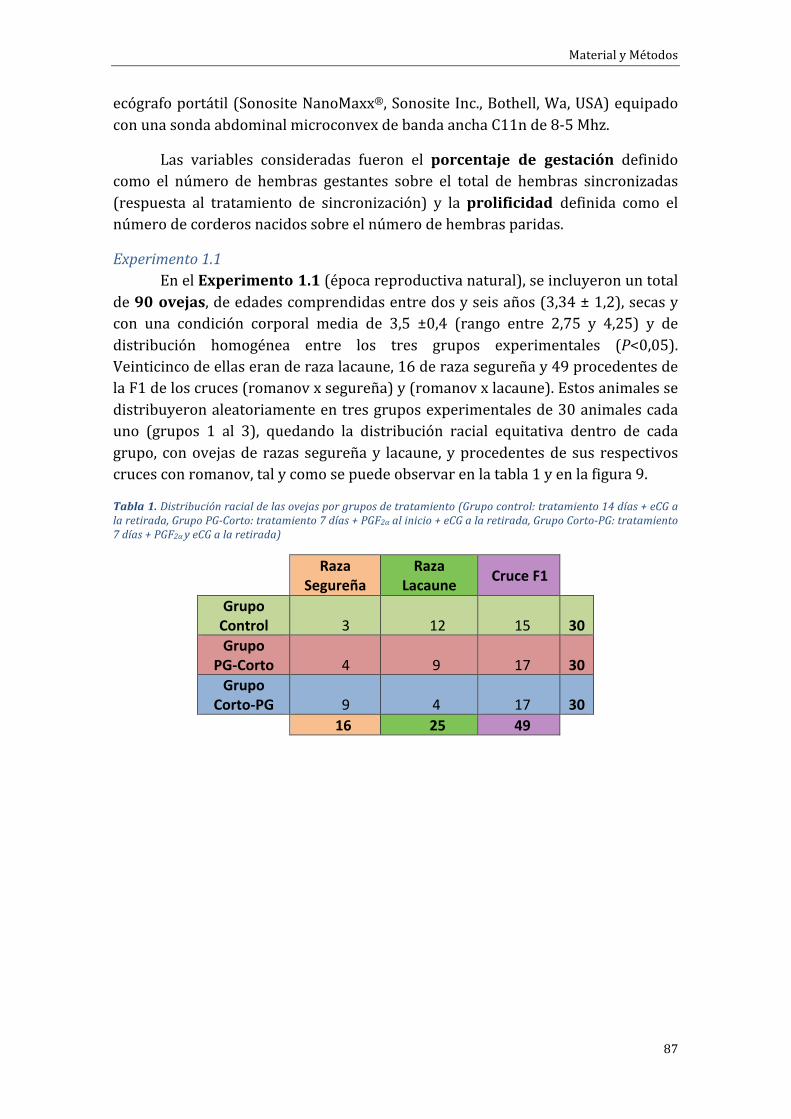
MaterialyMétodos
87
ecógrafoportátil(SonositeNanoMaxx®,SonositeInc.,Bothell,Wa,USA)equipadoconunasondaabdominalmicroconvexdebandaanchaC11nde8-5Mhz.
Las variables consideradas fueron el porcentaje de gestación definidocomo el número de hembras gestantes sobre el total de hembras sincronizadas(respuesta al tratamiento de sincronización) y la prolificidad definida como elnúmerodecorderosnacidossobreelnúmerodehembrasparidas.
Experimento1.1EnelExperimento1.1(épocareproductivanatural),seincluyeronuntotal
de90ovejas,deedadescomprendidasentredosyseisaños(3,34±1,2),secasycon una condición corporal media de 3,5 ±0,4 (rango entre 2,75 y 4,25) y dedistribución homogénea entre los tres grupos experimentales (P<0,05).Veinticincodeellaseranderazalacaune,16derazasegureñay49procedentesdelaF1deloscruces(romanovxsegureña)y(romanovxlacaune).Estosanimalessedistribuyeronaleatoriamenteentresgruposexperimentalesde30animalescadauno (grupos 1 al 3), quedando la distribución racial equitativa dentro de cadagrupo, conovejasde razas segureñay lacaune,yprocedentesde sus respectivoscrucesconromanov,talycomosepuedeobservarenlatabla1yenlafigura9.
Tabla1.Distribuciónracialdelasovejasporgruposdetratamiento(Grupocontrol:tratamiento14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
RazaSegureña
RazaLacaune CruceF1
GrupoControl 3 12 15
330
GrupoPG-Corto 4 9 17
330
GrupoCorto-PG 9 4 17
330
16 25 49
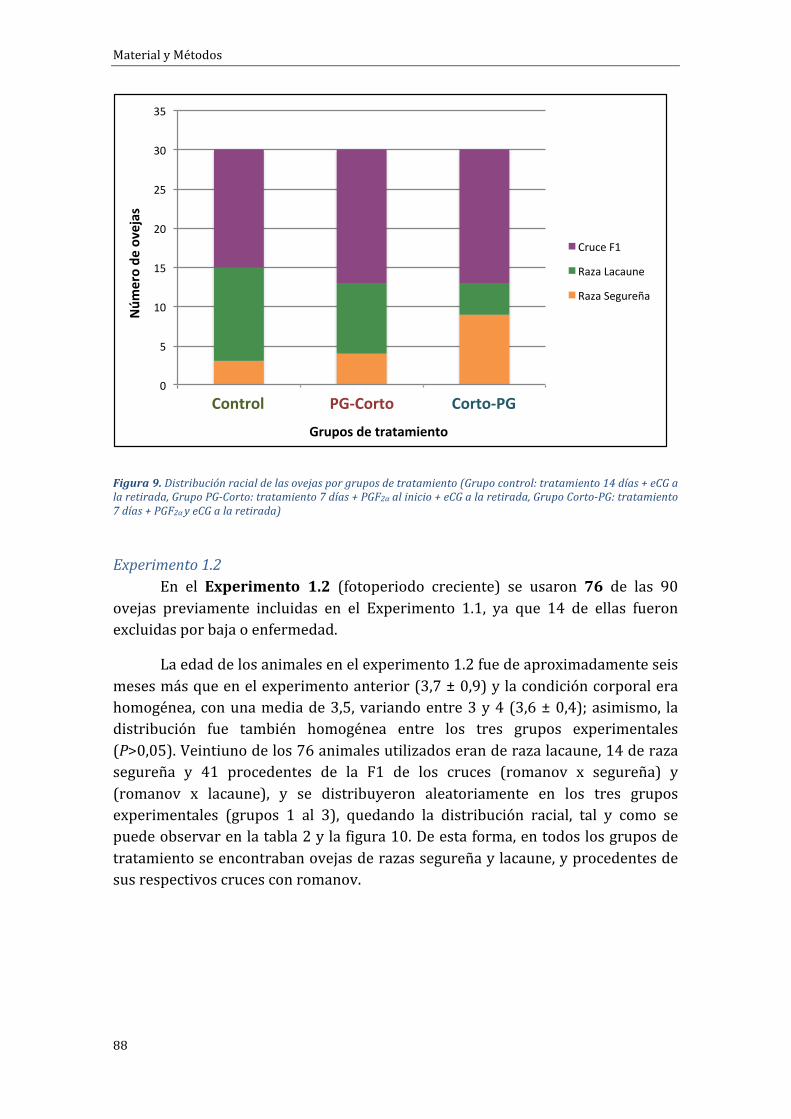
MaterialyMétodos
88
Figura9.Distribuciónracialdelasovejasporgruposdetratamiento(Grupocontrol:tratamiento14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
Experimento1.2
En el Experimento 1.2 (fotoperiodo creciente) se usaron 76 de las 90ovejas previamente incluidas en el Experimento 1.1, ya que 14 de ellas fueronexcluidasporbajaoenfermedad.
Laedaddelosanimalesenelexperimento1.2fuedeaproximadamenteseismesesmásqueenelexperimentoanterior(3,7±0,9)ylacondicióncorporalerahomogénea,conunamediade3,5,variandoentre3y4 (3,6±0,4);asimismo, ladistribución fue también homogénea entre los tres grupos experimentales(P>0,05).Veintiunodelos76animalesutilizadoseranderazalacaune,14derazasegureña y 41 procedentes de la F1 de los cruces (romanov x segureña) y(romanov x lacaune), y se distribuyeron aleatoriamente en los tres gruposexperimentales (grupos 1 al 3), quedando la distribución racial, tal y como sepuedeobservarenlatabla2ylafigura10.Deestaforma,entodoslosgruposdetratamientoseencontrabanovejasderazassegureñaylacaune,yprocedentesdesusrespectivoscrucesconromanov.
0
5
10
15
20
25
30
35
Grupo1 Grupo2 Grupo3
Núm
erode
ovejas
Gruposdetratamiento
CruceF1
RazaLacaune
RazaSegureña
ControlPG-CortoCorto-PG
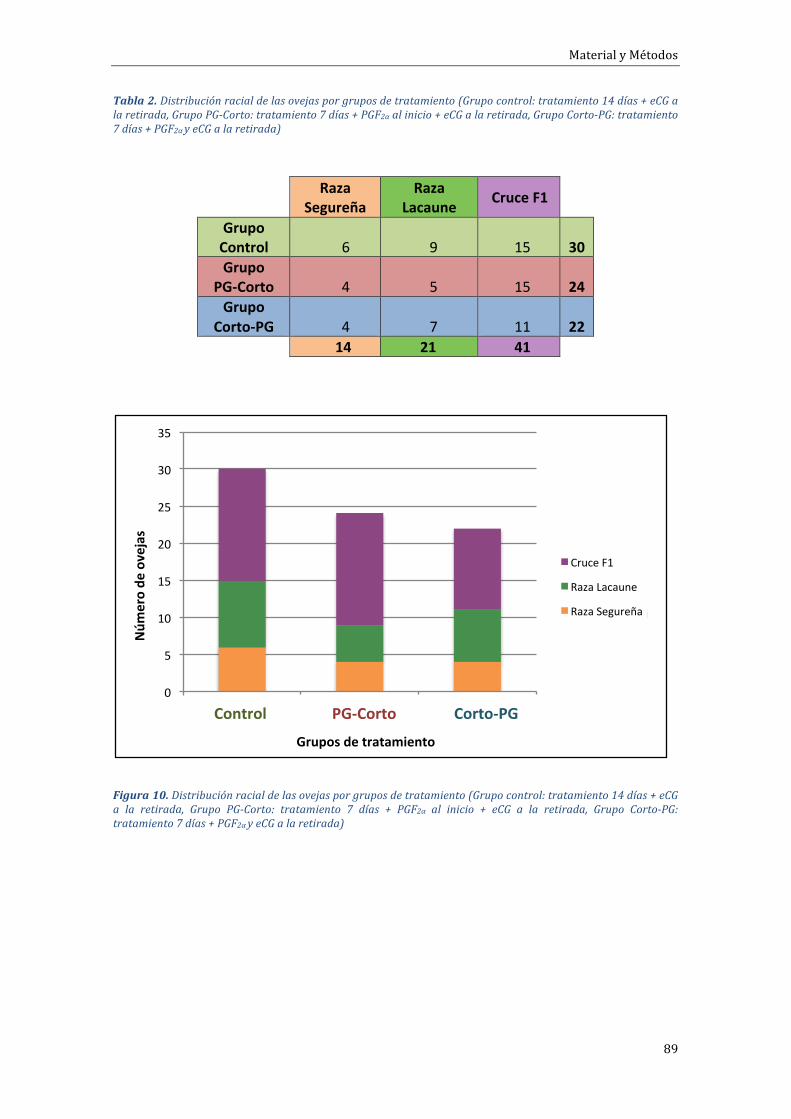
MaterialyMétodos
89
Tabla2.Distribuciónracialdelasovejasporgruposdetratamiento(Grupocontrol:tratamiento14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
RazaSegureña
RazaLacaune CruceF1
GrupoControl 6 9 15
330
GrupoPG-Corto 4 5 15
324
GrupoCorto-PG 4 7 11
322
14 21 41
Figura10.Distribuciónracialdelasovejasporgruposdetratamiento(Grupocontrol:tratamiento14días+eCGa la retirada, Grupo PG-Corto: tratamiento 7 días + PGF2α al inicio + eCG a la retirada, Grupo Corto-PG:tratamiento7días+PGF2αyeCGalaretirada)
0
5
10
15
20
25
30
35
Grupo1 Grupo2 Grupo3
Núm
erode
ovejas
Gruposdetratamiento
CruceF1
RazaLacaune
RazaSegureña
Control PG-Corto Corto-PG 0
5
10
15
20
25
30
35
Grupo1 Grupo2 Grupo3
Núm
erode
ovejas
Gruposdetratamiento
CruceF1
RazaLacaune
RazaSegureña
ControlPG-CortoCorto-PG0
5
10
15
20
25
30
35
Grupo1 Grupo2 Grupo3
Núm
erode
ovejas
Gruposdetratamiento
CruceF1
RazaLacaune
RazaSegureña
ControlPG-CortoCorto-PG0
5
10
15
20
25
30
35
Grupo1 Grupo2 Grupo3
Núm
erode
ovejas
Gruposdetratamiento
CruceF1
RazaLacaune
RazaSegureña
ControlPG-CortoCorto-PG0
5
10
15
20
25
30
35
Grupo1 Grupo2 Grupo3
Núm
erode
ovejas
Gruposdetratamiento
CruceF1
RazaLacaune
RazaSegureña
ControlPG-CortoCorto-PG

MaterialyMétodos
90
FaseExperimental2.Efectodeladuracióndeltratamientodesincronizacióndeceloscondispositivosintravaginalesliberadoresdeprogestágenossobrelafuncionalidadhipofisariayováricaenovejas,enfotoperiododecreciente
En esta fase se complementó la información obtenida en la Faseexperimental1medianteelestudiopormenorizadodelafuncionalidadhipofisariay ovárica relativa a los distintos métodos de sincronización. Por esta razón, elnúmerode animales utilizados fuemenor, de lamisma raza y durante lamismaépoca del año. Los animales se mantuvieron controlados en condicionesexperimentales, en la Granja Docente y de Investigación Veterinaria de laUniversidadCEUCardenalHerrera.
En total, se utilizaron 32 ovejas, todas de raza F1 (cruce realizado consementalesderazaromanovyovejasderazassegureñaylacaune)alcomprobarseenelexperimento1que larespuestaal tratamientodesincronizaciónnoseveíaafectadapor larazaovina.Laedadoscilóentre1y4añosdeedad(3,19±1,148años) y la condición corporalmedia fue de 3,5 ± 0,460 (rango entre 2,5 y 4). Elfotoperiodoenestafaseexperimentalfuedecreciente.
Los animales sedistribuyerondenuevoen tres grupos experimentales,segúnelprotocolodesincronizacióndeceloaplicadoydescritopreviamenteparala Fase Experimental 1 (Figura 8); Grupo Control (n=9), con el tratamiento“clásico o largo” de 14 días; Grupo PG-Corto (tratamiento “corto” conprostaglandinaaliniciodeltratamiento(n=12)yelGrupoCorto-PG(tratamiento“corto” y prostaglandina a la retirada del dispositivo vaginal (n=11). Todos losanimales presentaban edades y CC de distribución homogénea entre gruposexperimentales(P=0,815paraedadyP=0,745paraCC).
Losparámetrosevaluadosenestafaseexperimentalfueronlossiguientes:
• Porcentaje de hembras que mostraron celo, momento medio deaparicióndelmismoyagrupacióndeloscelosdesdelas12hastalas80htraslaretiradadelosdispositivos.
• Funcionalidadhipofisariayováricamediante:o Realizacióndemapasdecrecimientofolicular.o MedicióndelashormonasLH,E2circulanteyP4.
Ladeteccióndelcelofuellevadaacabomedianteinspecciónvisualdirectausandomoruecosentrenados, cadacuatrohorasdesde las12horashasta las80horas posteriores a la retirada de los dispositivos intravaginales cada 4 horas,considerándoseelmomentodeaparicióndelcelolaprimeravezenlaquelaovejaaceptabalamontadelmacho.

MaterialyMétodos
91
Lafuncionalidadováricadelasovejasseestudiómediantelavaloración,por ultrasonografía transrectal con ecógrafo portátil (Aloka SSD-500, Ecotron,Madrid,S;sondatransrectalde7.5MHz)delospatronesdecrecimientofolicularenelperiodoprevioalceloy,posteriormente,delatasadeovulaciónylacalidadmorfológicade losCLs; estosdatos se complementaron con ladeterminacióndelosnivelesplasmáticosdeE2yP4en fase folicular y luteal, respectivamente. Lafuncionalidad hipofisaria se valoró mediante la determinación de lascaracterísticasymomentodeaparicióndelpicopreovulatoriodeLH.
Las observaciones se efectuaron fijando la oveja en posición de decúbitodorsal.Lashecesseextraíanmanualmentedelrectoyposteriormenteseaplicabaungelhidrosoluble,conlafinalidaddeevitardañosaniveldelamucosarectalymejorar la transmisión de la imagen ecográfica. Seguidamente, la sonda secolocaba dentro de la ampolla rectal, orientando el transductorperpendicularmente hacia la pared abdominal. Los folículos fueron identificadoscomoestructurasanecoicasysumediciónfuerealizadacolocandoloscalibradoreselectrónicosenellímiteubicadoentrelaparedfolicularyelestromaovárico.LosCLssecaracterizabancomoestructurashipoecoicasyhomogéneas;susdiámetrosmayorymenor semidieron colocando, en formaperpendicular, los calibradoreselectrónicosdelecógrafoenlainterfaseubicadaentreeltejidolutealyelestromaovárico.
Losdatosecográficosseorganizaronparacaracterizarlospatronesdecadaondadecrecimientofolicularysusefectossobrelapoblaciónfolicular,duranteelintervalointerovulatorio.
Los folículos ovulatorios presentes en los ovarios se clasificaron por sudiámetro en cuatro grupos según su diámetro: folículos totales (mayores de 2mm), folículos pequeños (2,0-3,0 mm), folículos medianos (4,0-5,0 mm) yfolículosgrandes(mayorde5,0mm).Además,durantelafasefolicular,tambiénse tuvieron en cuenta el número de folículos nuevos (no detectados el díaanterior),folículosencrecimiento(aquellosenlosqueseproduceunaumentodetamañoconrespectoaldíaanterior)yfolículosenregresión(aquellosenlosque,bien se produce una disminución en el diámetro con respecto al día anterior, obienhayandesaparecido).Porúltimo,seanotóeltamañodelfolículomásgrande(F1), del segundo folículo más grande (F2) y el tamaño medio del resto de losfolículos(Fresto).
En el mismo periodo de fase folicular, para la evaluación de los nivelesplasmáticosdeE2 seobtuvieronmuestrasde sangrea las0, 12,24,36,48y60horas, y para la evaluación de los niveles plasmáticos de LH se obtuvieronmuestras a las 32, 36, 40, 44, 48, 52, 56, 60, 64, 68, 72, 76 y 80 horas. Estasmuestrasseobtuvieron,mediantepuncióndelavenayugular,entubosvacutainerheparinizados de 5 ml. Inmediatamente tras la extracción, se centrifugaron las

MaterialyMétodos
92
muestras durante 15 minutos a una velocidad de 3500 rpm. Seguidamente, sealmacenó el equivalente a 2ml de plasma/muestra, en criotubo de 3ml, a unatemperaturade-20ºChastasuposteriorprocesamiento.
La valoración de número y la calidad morfológica y funcional de lasovulaciones se llevó a cabo 11 días después de la retirada del tratamientoprogestativo. Esta observación se realizó mediante ecografía transrectal ymediantelavaloracióndelosnivelesplasmáticosdeprogesteronaenunamuestrade5mldesangreobtenidaentubosvacutainerheparinizados;estasmuestrasdesangrefueronprocesadasdelaformadescritaparalasmuestrasanteriores.
LaevaluacióndelasconcentracionesplasmáticasdeE2yP4serealizóenelDepartamento de Fisiología de la Facultad de Medicina de la UniversidadComplutense deMadrid,mientras que la determinación de los niveles de LH serealizó en el laboratorio de Fisiología Comparada del Departamento deReproducción Animal del INIA (Instituto Nacional de Investigación y TecnologíaAgrariayAlimentaria).
• La concentración plasmática de E2 se determinó con un kit deradioinmunoensayo (ESTR-US-CTUltrasensible, IBAMolecular,Madrid), según lametodologíadescritaporRomeuetal.(1995)yadaptadapara suusoenplasmaovinoporGonzález-Bulnesetal.(2003)(Romeuetal.,1995;Gonzalez-Bulnesetal.,2003).Lasensibilidaddelapruebafuede5pg/mlyloscoeficientesdevariaciónintraeinterensayofueron5,0y9,7%,respectivamente.
• LasconcentracionesplasmáticasdeLH fueronevaluadasusandounkitdeELISAbasadoenelprincipiodeunióncompetitiva(LHDETECTforovineTBMReproPharm,Nouzilly,France),cuyasensibilidadfuede0,01ng/ml.Paralasfasesexperimentales 2 y 4, los coeficientes de variación intra-ensayo fueron de 5,0 y3,4%y losdel inter-ensayofueron6,4y9,6%,respectivamente.Enrelacióna lascaracterísticas de la onda preovulatoria de LH, el inicio y final de ésta fuerondefinidoscomoelvaloranterioryposterior,respectivamente,alincrementoenun10%, de las concentraciones basales de LH. Las concentraciones basales de LH,fuerondeterminadoscalculandoelpromediodelasconcentracionesdeLHantesydespués del pico preovulatorio de LH. La duración del pico de LH fue definidacomoladiferenciaentreelinicioyelfinaldelaondapreovulatoria.Eláreabajolacurva (AUC), fue calculado usando el método del paralelepípedo descrito porRowlanden1989(Rowland,1989).
• Las concentraciones plasmáticas deP4 se determinaron con un kit deradioinmunoanálisis usando kits en fase sólida (PROG-CTRIA progesterona, IBAMolecular, Madrid). La sensibilidad del análisis y el coeficiente de variaciónintraensayoen la faseexperimental2 fueron0,3ng/mly3,1%,respectivamente.En los análisis realizados durante la fase experimental 4, la sensibilidad y loscoeficientes de variación intra e interensayos fueron 0,1 ng/ml, 4,8 y 10,0%,

MaterialyMétodos
93
respectivamente. La presencia de un CL activo fue definida cuando lasconcentracionesplasmáticasdeP4fueronigualesosuperioresa0,5ng/ml.
FaseExperimental3.Efectodeltipodedispositivoprogestativointravaginal(progesteronanaturaloCIDRversusprogestágenosintéticooFGA)sobreelporcentaje de gestación en ovejas tratadas con un protocolo desincronizacióndeceloslargo(14días)enfotoperiododecreciente
Esta parte del trabajo se centró en la comparación de la eficiencia deprotocolos de sincronización basados en la utilización de dispositivosintravaginalescondosprogestativosdiferentesenunprotocolo largo tradicionalde 14 días con administración de eCG a la retirada de los dispositivosintravaginales(figura11)durantefotoperiododecreciente.
Para esta fase experimental 3 se utilizaron un total de 303 ovejas,multíparas,de raza lacaune lechera, cedidasporunaexplotación comercialde lazona.Elexperimentoserealizóen4réplicasde75ovejasparaelprimerloteyde76paracadaunodelostresrestantes,entrelosmesesdeseptiembreadiciembre.En cadaunodeestos gruposhabíaun10%deovejasqueno se encontrabanenlactación(8enelprimergrupoyenelcuartoy7enelsegundoyeneltercero).Laedaddelosanimalesutilizadosfuede3,1±1,55años(valoresmínimoymáximode1,1y8,3años)yde3,0±1,93lactaciones/oveja;lacondicióncorporalentre1y3,8,conunpromediode2,4±0,51.Deellas,untotalde186(46ovejasencadaunade las dos primeras réplicas y 47 ovejas en cada una de las dos últimas) seasignarondemaneraaleatoriaparasusincronizaciónconesponjasintravaginalesimpregnadascon20mgdeacetatodeflugestona“FGA”(Chronogest®,MSDAnimalHealth,Madrid,España)y117(29ovejasencadaunadelastresprimerasréplicasy30enlaúltimadeellas)condispositivotipo“CIDR”(EAZI-BREED™CIDR®SheepInsert, Zoetis, Parsippany, New Jersey, EE.UU.), impregnado con 0,3 g deprogesterona, insertándolo durante 14 días, aplicando 400 U.I. eCG por vía i.m.(Foligon®,MSD Animal Health,Madrid, España) en elmomento de la retirada yprocediendoalaobservacióndecelotraslaretirada.
Elanálisisestadísticodelosdatosserealizósobreelconjuntodetodoslosanimales al finalizar la fase experimental. Dentro del grupo sincronizado condispositivos de FGA un total de 7 animales se perdieron por diferentes razonesantes de comenzar el protocolo de sincronización (eliminación, error deseparaciónenelmomentodecolocacióndelasesponjasymortalidad),porloquefinalmente,recibieroneldispositivodeFGA179ovejasadultas.Deéstos,duranteel tiempo de sincronización se observó un total de 8 pérdidas del dispositivo(4,469%). En elmomento del diagnóstico de gestación se perdieron 4 animalesadicionalespor lasmismasrazonesantesdescritas,por loque la “n” finalqueseconsideróenelestudiodentrodelgrupoFGAfuede167animales.Laedaddelosanimalesutilizadosenestegrupofuede3,1±1,31años(valoresmínimoymáximo

MaterialyMétodos
94
de1,2y8,2años)yde3,0±1,70lactaciones/oveja;lacondicióncorporalentre1y3,8,conunpromediode2,4±0,50.
En el caso del grupo CIDR un total de 12 animales se perdieron pordiferentesrazones(eliminación,errordeseparaciónenelmomentodecolocaciónde losCIDRymortalidad),por loquesesincronizóuntotalde105animalescondispositivosCIDR.Duranteeltiempodesincronizaciónseobservarondospérdidasdedispositivo(1,905%).Enelmomentodeldiagnósticodegestaciónseperdieron8 animales adicionalespor lasmismas razones antesdescritas, por loque la “n”finalqueseconsideróenelestudiofuede95animalesenelgrupoCIDR.Laedaddelosanimalesutilizadosenestegrupofuede2,9±1,90años(valoresmínimoymáximode1,1y8,3años)yde2,9±2,31lactaciones/oveja;lacondicióncorporalentre1y3,8,conunpromediode2,4±0,56.
Los datos de edad media, número de lactaciones por oveja y condicióncorporalmedia no difirieron significativamente entre los grupos experimentales(P=0,361;P=0,864yP=0,837,respectivamente).
Losmachos,utilizadosenunaproporción1:5tambiéneranderazalacaunede aptitud lechera. Se efectuaron diagnósticos de gestación mediante ecografíatransabdominal para determinar el porcentaje de gestación y la prolificidadestimada,conecógrafoportátil (SonoSiteMturbo,Madrid,S;sondamicroconvexC11de5a8MHz).
Gruposexperimentales:
• Grupo FGA: las hembras recibieron esponjas intravaginales impregnadacon 20 mg de acetato de flugestona “FGA” (Chronogest®, MSD Animal Health,Madrid,España).• Grupo CIDR: estas ovejas se trataron con el dispositivo tipo “CIDR”
impregnadocon0,3gdeprogesterona(EAZI-BREED™CIDR®SheepInsert,Zoetis,Parsippany,NewJersey,EE.UU.).
Enambosgruposseadministraron400U.I.eCGporvíai.m.(Foligon®,MSDAnimalHealth,Madrid,España)enelmomentode la retiradade losdispositivos(Figura11).

MaterialyMétodos
95
Figura11.ProtocolosdesincronizacióndeceloenbaseadispositivosintravaginalesdeprogestágenosGrupoFGAoprogesteronaGrupoCIDR,utilizadosenlaFaseExperimental3.Protocolodesincronizaciónlargo,de14días
FaseExperimental4.Efectodeltipodedispositivoprogestativointravaginal(progesteronanaturaloCIDRversusprogestágenosintéticooFGA)sobrelafuncionalidadhipofisaria y ovárica enovejas tratadas conunprotocolodesincronizacióndeceloscorto(7días)einfluenciadelaépocareproductiva
Para esta cuarta fase experimental se eligió el tratamiento corto quemejores resultados había inducido en las fases anteriores 1 y 2: el tratamientoCorto-PG.
Asípues,estapartedeltrabajosecentróenlacomparacióndelaeficienciade protocolos de sincronización basados en la utilización de dispositivosintravaginales con dos progestativos diferentes en el protocolo corto conadministración de la prostaglandina en el momento de la retirada de losdispositivosintravaginales(figura12).Paraellosellevaronacabo,deigualformaque en la primera fase experimental, dos experimentos consecutivos paradeterminarposiblesefectosdel fotoperiodo.Así,elexperimento4.1serealizóenfotoperiodocreciente,yelexperimento4.2,enfotoperiododecreciente.
Paraestafaseexperimental4seutilizóuntotalde49ovejas,pertenecientesalaF1deloscrucesrealizadosconsementalesderazaromanovyovejasderazassegureña y lacaune. La edadde los animalesutilizadososciló entre los2 y los5años (3.48±1.154) y la condición corporal entre 3.25 y 4, con un promedio de3.83±0,286).
Gruposexperimentales:
• Grupo FGA: las hembras recibieron esponjas intravaginalesimpregnadacon20mgdeacetatodeflugestona“FGA”(Chronogest®,MSDAnimalHealth,Madrid,España).
• Grupo CIDR: estas ovejas se trataron con el dispositivo tipo “CIDR”impregnadocon0,3gdeprogesterona(EAZI-BREED™CIDR®SheepInsert,Zoetis,Parsippany,NewJersey,EE.UU.).
insertar esponjas
intravaginales
GRUPO CIDR
retirar esponjas
intravaginales
administrar eCG +
insertar CIDR
retirar CIDR
administrar eCG
+
GRUPO FGA
Día1
Día2
Día3
Día4
Día5
Día6
Día7
Día8
Día9
Día10
Día11
Día12
Día13
Día14

MaterialyMétodos
96
Enambosgruposseadministraron5mgdeprostaglandinaF2α.(Dinolytic®,Zoetis, Madrid, España) y 400 U.I. eCG (Foligon®, MSD Animal Health, Madrid,España) por vía i.m. ambas, en el momento de la retirada de los dispositivos(Figura11).
Figura 12. Protocolos de sincronizaciónde celo enbasea dispositivos intravaginales deprogestágenosGrupoFGAoprogesteronaGrupoCIDR,utilizadosenlaFaseExperimental4.Protocolocortodesincronizaciónde7días
Experimento4.1Fotoperiodocreciente
En el Experimento 4.1 (fotoperiodo creciente) los animales sedistribuyeronaleatoriamenteendosgruposexperimentales:GrupoFGA(n=24)yGrupo CIDR (n=25). La distribución tanto de edad como de CC resultó serhomogéneaentrelosdosgruposexperimentales(P<0,05).
Lasvariablesconsideradasfueron:
• Porcentaje de hembras que mostraban celo, momento medio deaparicióndelmismoyagrupaciónde loscelos,desde las12hasta las80htraslaretiradadelosdispositivos• Funcionalidadhipofisariayováricamediante:
o DeterminacióndelashormonasLH,E2yP4.
LaevaluacióndelasconcentracionesplasmáticasdeE2yP4serealizóenelDepartamento de Fisiología de la Facultad de Medicina de la UniversidadComplutense deMadrid,mientras que la determinación de los niveles de LH serealizó en el laboratorio de Fisiología Comparada del Departamento deReproducción Animal del INIA (Instituto Nacional de Investigación y TecnologíaAgrariayAlimentaria).
• LaconcentracionesplasmáticasdeE2yP4sedeterminaronconunkit de radioinmunoensayoRIA igual que en la fase experimental 2.
Día1
Día2
Día3
Día4
Día5
Día6
Día7
insertar esponjas
intravaginales
GRUPO CIDR
retirar esponjas
intravaginales
administrar eCG + +administrar
PGF2α
insertar CIDR
retirar CIDR
administrar eCG
administrar PGF2α +
+
GRUPO FGA

MaterialyMétodos
97
loscoeficientesdevariaciónintraeinterensayoparaE2fueron5,0y9,7%,respectivamente.
• LasconcentracionesplasmáticasdeLHfueronevaluadasusandounkitdeELISAal igualqueen la faseexperimental2.Loscoeficientesdevariaciónintra-ensayofueronde5,0y3,4%ylosdelinter-ensayofueron6,4y9,6%,respectivamente.
Experimento4.2FotoperiododecrecienteEn el Experimento 4.2 (fotoperiodo decreciente) los animales se
distribuyeronenlosmismosgruposexperimentalespreviamentedescritosparaelexperimento4.1.,peroestavezsecruzaron losgrupos,demaneraqueelGrupoFGAincluíalos25animalesquepreviamenteerandelgrupoCIDRyelGrupoCIDRincluía las 24 ovejas previamente en el grupo FGA. La distribución tanto de CCresultóser,denuevo,homogéneaentrelosdosgruposexperimentales(P<0,05).
Enestaúltimafaseexperimentaltansólosevaloróelporcentajedesalidaen celo, elmomentomediodelmismoy la agrupaciónde los celos,desde las12hastalas80htraslaretiradadelosdispositivos.
3.4 Análisisestadístico
Todos los datos recopilados se resumieron en tablas excel para sudepuraciónycomprobaciónprevioalprocesadoestadístico.ElprogramautilizadofueSPSS®22.0(IBMCorporation,NuevaYorkNY,EE.UU.).
Se comprobó la homogeneidad de los grupos experimentales en cuanto afactoresdeconfusióntalescomoraza,edad,condicióncorporalyfotoperiodo.Lasvariablesdependientesfueronlarespuestaaltratamiento(fertilidadyprolificidadencasode lasFasesExperimentales1y3), tasasdepresentacióndecelos(fasesexperimentales2y4)yestudiodelafaseluteal(fases2y4).Enlosestudiosdeladinámica folicular (fase experimentales 2) las variables fueron el número defolículospresentes,lapresenciadefolículodominanteylaevidenciadeovulacióno no. Las determinaciones hormonales (fases experimentales 2 y 4) seconsiderarontambiénvariablescontinuasydependientes,yderelevanciaenestasmismasfasesexperimentales.
Las diferencias estadísticas en los parámetros continuos (prolificidad, CC,edadyparámetroshormonales)paramásdedos grupos se estimaronmedianteanálisis de varianza (ANOVA), pero también se realizaron las pruebas Student-Newman-Keuls,ydeDuncanposthocparacontrastarlasdiferenciasdentrodelosgrupos.

MaterialyMétodos
98
Para comparar las medias entre dos grupos en el caso de variablescontinuas(prolificidad,CC,edadyparámetroshormonales)seutilizólapruebadet de Student. Para las variables con varianza no esférica, se utilizó un nivel designificacióndeGreenhouse.
La comparación entre las frecuencias de las variables categóricas (raza,respuestaaltratamiento,salidaenceloaintervaloshorarios)seanalizómediantepurebasdeChicuadrado,aligualquelarespuestaaltratamientoofertilidad,queseconsiderócomovariabledicotómica(sí/no).Adicionalmente,seexploraronlassignificanciasdelefectodelostratamientosexperimentales,simultáneamenteconel efecto de factores que pudieran ejercer un efecto adicional y/o de confusión(edad, CC, raza y fotoperiodo) mediante procedimientos de regresión logísticabinaria, por pasos prospectivos basados en estadísticos deWald con criterio deP>0.10,incluyendolasinteraccionesdeprimergrado.
Los datos ecográficos fueron valorados usando un análisis de la varianza(ANOVA)deuna sola vía.Asimismo, se llevó a cabouna correlacióndePearson,paradeterminarlaasociaciónentreeldiámetroyelárea(conosincavidad)delCLylosnivelesdeP4enplasma.

RESULTADOS


Resultados
101
4 Resultados
4.1 Faseexperimental1.Efectodeladuracióndeltratamientode sincronización de celos con dispositivos intravaginalesliberadores de progestágenos e influencia de la épocareproductivaydelarazaovina
En el experimento 1.1 se utilizaron un total de 90 ovejas con una edadmediade3,36±1,2; de ellas, 25 erande raza lacaune (27,8%), 16 erande razasegureña (17,8%)y49eranel resultadode laF1de loscrucesde lasanteriorescon romanov (54,4%). Tal y como se ha descrito en el apartado material ymétodos, la distribución de razas fue homogénea entre grupos experimentales(P>0,05).
Tabla3.Númeroyporcentajedeanimalesporrazaqueseincluyeronenelestudiodedistintostratamientosdesincronización del celo basados en el uso de dispositivos intravaginales de progestágenos en fotoperiododecreciente
Raza %(N)
CruceF1 54,4(49)
Lacaune 27,8(25)
Segureña 17,8(16)
Enelexperimento1.2seutilizaron76ovejasdelas90queseutilizaronenelexperimento1.1.Delas14ovejasrestantes,12seencontrabangestantesydoscausaronbajapormuerte.Una,acausadeunatoxemiadegestaciónylaotraporunmeteorismo.
Las 76 hembras participantes en el experimento 1.2 teníanunamedia de3,7±0,9añosdeedadydeellas21pertenecíanalarazalacaune(27,63%),14alaraza segureña (18,42%) y 41 a la F1 de los cruces de las razas anteriormentecitadasconlarazaromanov(53,95%),talycomosepuedeobservarenlatabla4.Igualmente, ladistribuciónderazas fuehomogéneaentregruposexperimentales(P>0,05).

Resultados
102
Tabla4.Númeroyporcentajedeanimalesporrazaqueseincluyeronenelestudiodedistintostratamientosdesincronizacióndelcelobasadosenelusodedispositivosintravaginalesdeprogestágenosenfotoperiodocreciente
Raza %(N)
CruceF1 54,0(41)
Lacaune 27,6(21)
Segureña 18,4(14)
Enprimerlugar,describiremoslosresultadosseparadosporexperimentos1.1(fotoperiododecreciente)y1.2(fotoperiododecreciente),parapasaramostrarresultadoscongruposagrupadosencasodepermitirlolosresultadosestadísticosyeldiseñometodológico.
4.1.1 Experimento1.1Influenciadeltipodetratamientosobreelporcentajedegestaciónduranteelfotoperiododecreciente
Esteexperimentoserealizósobreuntotalde90animales,divididosentresgrupos experimentales distintos: Grupo control o de tratamiento “largo”(tratamiento 14 días + eCG a la retirada), GrupoPG-Corto (tratamiento 7 días +PGF2αalinicio+eCGalaretirada)y(GrupoCorto-PG:tratamiento7días+PGF2αyeCG a la retirada), con una edad media de 3,36 ± 1,21 años, sin diferenciassignificativasentrelostresgrupos(P=0,210),siendolamediaenelgrupocontrolde3,08±1,10años,de3,23±1,06añosenelgrupoPG-Cortoyde3,76±1,38añosenelgrupoCorto-PG.
La condición corporal no fue significativamente diferente entre los tresgrupos(P=0,462),conunamediatotalde3,51±0,40,siendoestamediade3,61±0,34enelgrupocontrolo“largo”,3,50±0,44enelgrupoPG-Cortoy3,42±0,40enelgrupoCorto-PG.
Los resultados de porcentaje de gestación en función del tratamiento sepueden observar en la figura 13. El grupo de animales que mostró un mayorporcentaje de gestación durante el fotoperiodo decreciente fue el tratamiento“corto” con prostaglandina F2α en el momento de la retirada del progestágeno(grupo Corto-PG: 90,0%), seguido del otro grupo corto (PG-Corto: 80%) y,finalmente, del grupo control o tratamiento largo (76,7%); sin embargo, estasdiferenciasnuméricasnofueronsignificativasenningúncaso(figura13).
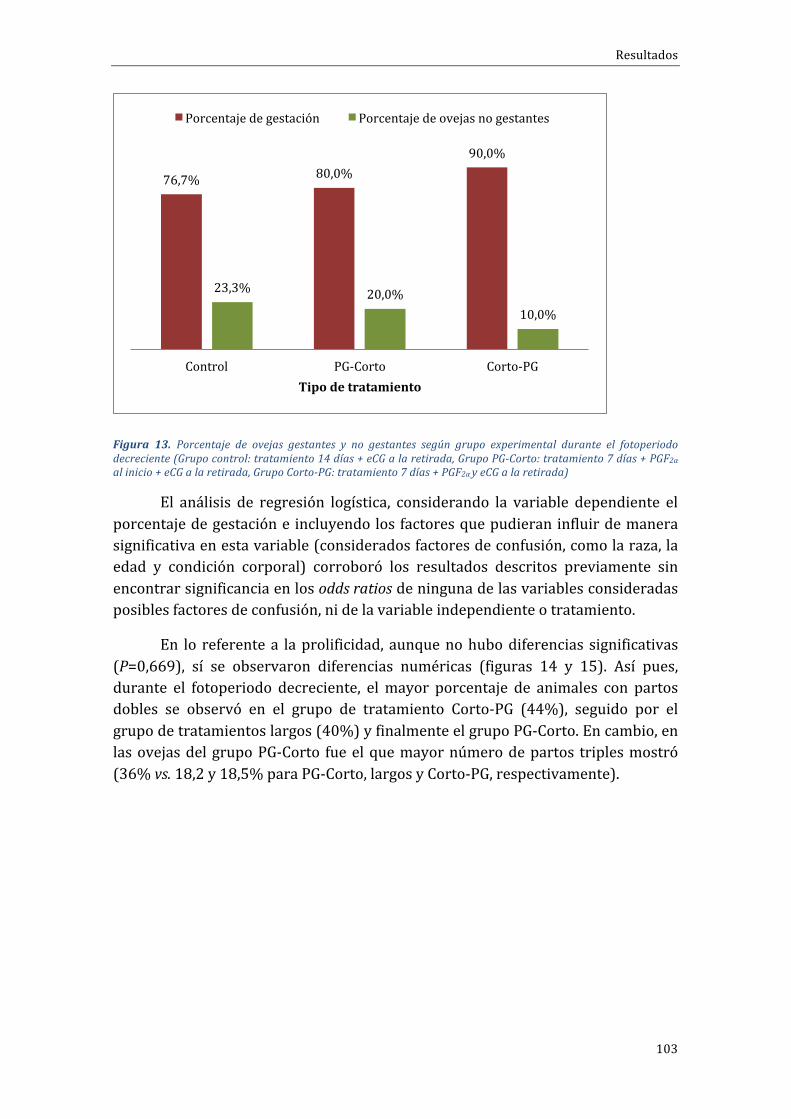
Resultados
103
Figura 13. Porcentaje de ovejas gestantes y no gestantes según grupo experimental durante el fotoperiododecreciente(Grupocontrol:tratamiento14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
El análisis de regresión logística, considerando la variable dependiente elporcentajedegestacióne incluyendolosfactoresquepudieraninfluirdemanerasignificativaenestavariable(consideradosfactoresdeconfusión,comolaraza,laedad y condición corporal) corroboró los resultados descritos previamente sinencontrarsignificanciaenlosoddsratiosdeningunadelasvariablesconsideradasposiblesfactoresdeconfusión,nidelavariableindependienteotratamiento.
En lo referentea laprolificidad, aunquenohubodiferencias significativas(P=0,669), sí se observaron diferencias numéricas (figuras 14 y 15). Así pues,durante el fotoperiodo decreciente, elmayor porcentaje de animales con partosdobles se observó en el grupo de tratamiento Corto-PG (44%), seguido por elgrupodetratamientoslargos(40%)yfinalmenteelgrupoPG-Corto.Encambio,enlasovejasdelgrupoPG-Corto fueelquemayornúmerodepartos triplesmostró(36%vs.18,2y18,5%paraPG-Corto,largosyCorto-PG,respectivamente).
76,7% 80,0%90,0%
23,3% 20,0%10,0%
Control PG-Corto Corto-PGTipodetratamiento
Porcentajedegestación Porcentajedeovejasnogestantes
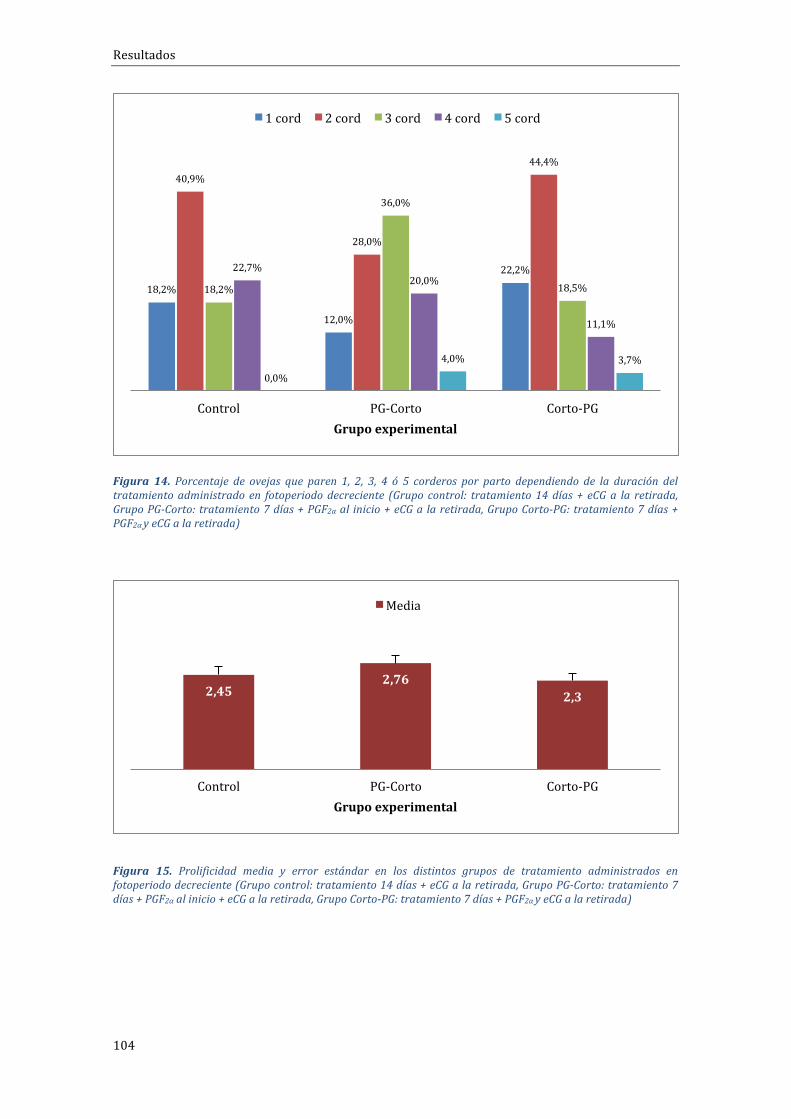
Resultados
104
Figura 14. Porcentaje de ovejas que paren 1, 2, 3, 4 ó 5 corderos por parto dependiendo de la duración deltratamientoadministrado en fotoperiododecreciente (Grupo control: tratamiento14días + eCGa la retirada,GrupoPG-Corto:tratamiento7días+PGF2αal inicio+eCGa laretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
Figura 15. Prolificidad media y error estándar en los distintos grupos de tratamiento administrados enfotoperiododecreciente(Grupocontrol:tratamiento14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
18,2%
12,0%
22,2%
40,9%
28,0%
44,4%
18,2%
36,0%
18,5%
22,7%20,0%
11,1%
0,0%
4,0% 3,7%
Control PG-Corto Corto-PGGrupoexperimental
1cord 2cord 3cord 4cord 5cord
2,452,76
2,3
Control PG-Corto Corto-PGGrupoexperimental
Media
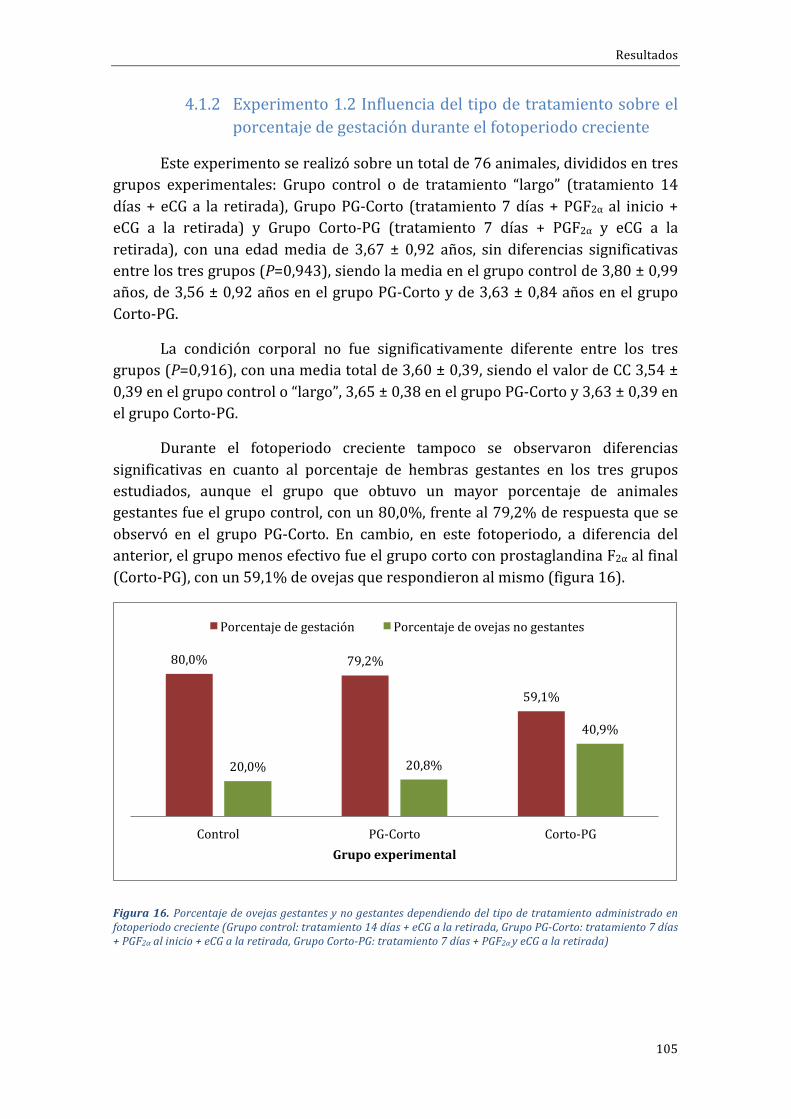
Resultados
105
4.1.2 Experimento1.2Influenciadeltipodetratamientosobreelporcentajedegestaciónduranteelfotoperiodocreciente
Esteexperimentoserealizósobreuntotalde76animales,divididosentresgrupos experimentales: Grupo control o de tratamiento “largo” (tratamiento 14días+eCGa la retirada),GrupoPG-Corto (tratamiento7días+PGF2α al inicio+eCG a la retirada) y Grupo Corto-PG (tratamiento 7 días + PGF2α y eCG a laretirada), con una edadmedia de 3,67 ± 0,92 años, sin diferencias significativasentrelostresgrupos(P=0,943),siendolamediaenelgrupocontrolde3,80±0,99años,de3,56±0,92añosenelgrupoPG-Cortoyde3,63±0,84añosenelgrupoCorto-PG.
La condición corporal no fue significativamente diferente entre los tresgrupos(P=0,916),conunamediatotalde3,60±0,39,siendoelvalordeCC3,54±0,39enelgrupocontrolo“largo”,3,65±0,38enelgrupoPG-Cortoy3,63±0,39enelgrupoCorto-PG.
Durante el fotoperiodo creciente tampoco se observaron diferenciassignificativas en cuanto al porcentaje de hembras gestantes en los tres gruposestudiados, aunque el grupo que obtuvo un mayor porcentaje de animalesgestantesfueelgrupocontrol,conun80,0%,frenteal79,2%derespuestaqueseobservó en el grupo PG-Corto. En cambio, en este fotoperiodo, a diferencia delanterior,elgrupomenosefectivofueelgrupocortoconprostaglandinaF2αalfinal(Corto-PG),conun59,1%deovejasquerespondieronalmismo(figura16).
Figura16.Porcentajedeovejasgestantesynogestantesdependiendodeltipodetratamientoadministradoenfotoperiodocreciente(Grupocontrol:tratamiento14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
80,0% 79,2%
59,1%
20,0% 20,8%
40,9%
Control PG-Corto Corto-PGGrupoexperimental
Porcentajedegestación Porcentajedeovejasnogestantes
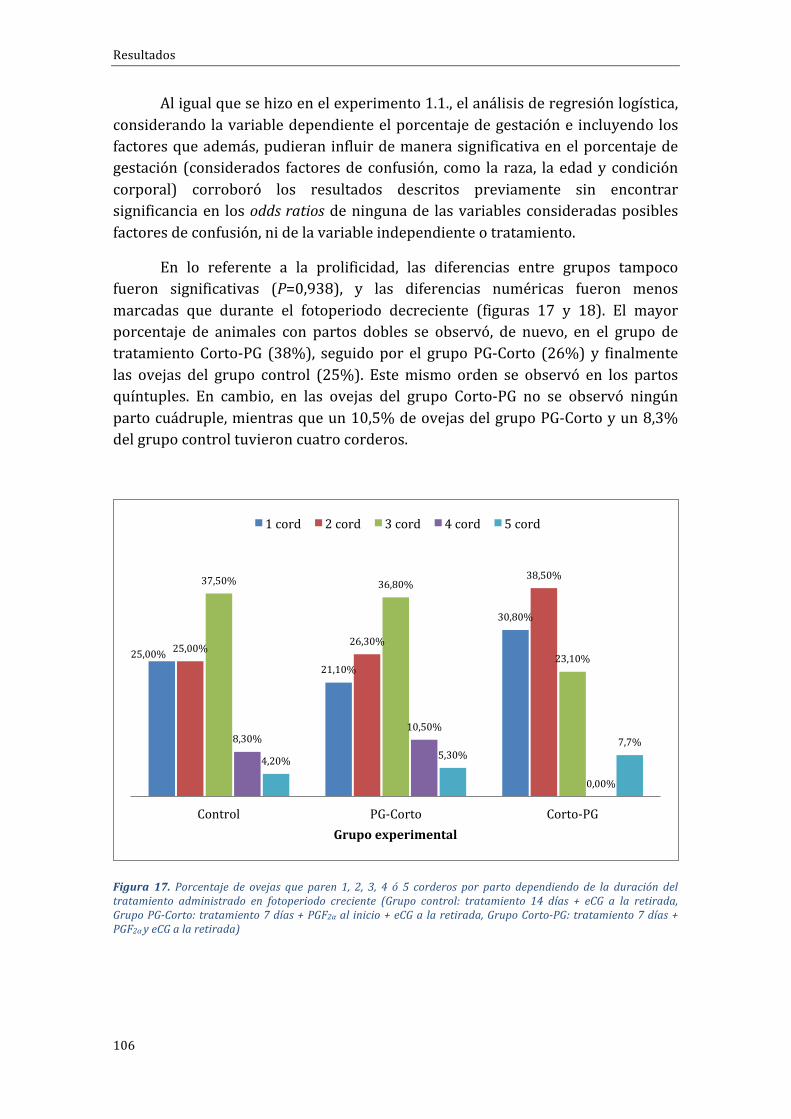
Resultados
106
Aligualquesehizoenelexperimento1.1.,elanálisisderegresiónlogística,considerandolavariabledependienteelporcentajedegestacióne incluyendolosfactoresqueademás,pudieraninfluirdemanerasignificativaenelporcentajedegestación (considerados factoresde confusión, como la raza, la edady condicióncorporal) corroboró los resultados descritos previamente sin encontrarsignificanciaen losoddsratiosdeningunade lasvariablesconsideradasposiblesfactoresdeconfusión,nidelavariableindependienteotratamiento.
En lo referente a la prolificidad, las diferencias entre grupos tampocofueron significativas (P=0,938), y las diferencias numéricas fueron menosmarcadas que durante el fotoperiodo decreciente (figuras 17 y 18). El mayorporcentaje de animales con partos dobles se observó, de nuevo, en el grupo detratamiento Corto-PG (38%), seguido por el grupoPG-Corto (26%) y finalmentelas ovejas del grupo control (25%). Estemismo orden se observó en los partosquíntuples. En cambio, en las ovejas del grupo Corto-PG no se observó ningúnpartocuádruple,mientrasqueun10,5%deovejasdelgrupoPG-Cortoyun8,3%delgrupocontroltuvieroncuatrocorderos.
Figura 17. Porcentaje de ovejas que paren 1, 2, 3, 4 ó 5 corderos por parto dependiendo de la duración deltratamiento administrado en fotoperiodo creciente (Grupo control: tratamiento 14 días + eCG a la retirada,GrupoPG-Corto:tratamiento7días+PGF2αal inicio+eCGa laretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
25,00%21,10%
30,80%
25,00% 26,30%
38,50%37,50% 36,80%
23,10%
8,30%10,50%
0,00%
4,20% 5,30%7,7%
Control PG-Corto Corto-PGGrupoexperimental
1cord 2cord 3cord 4cord 5cord
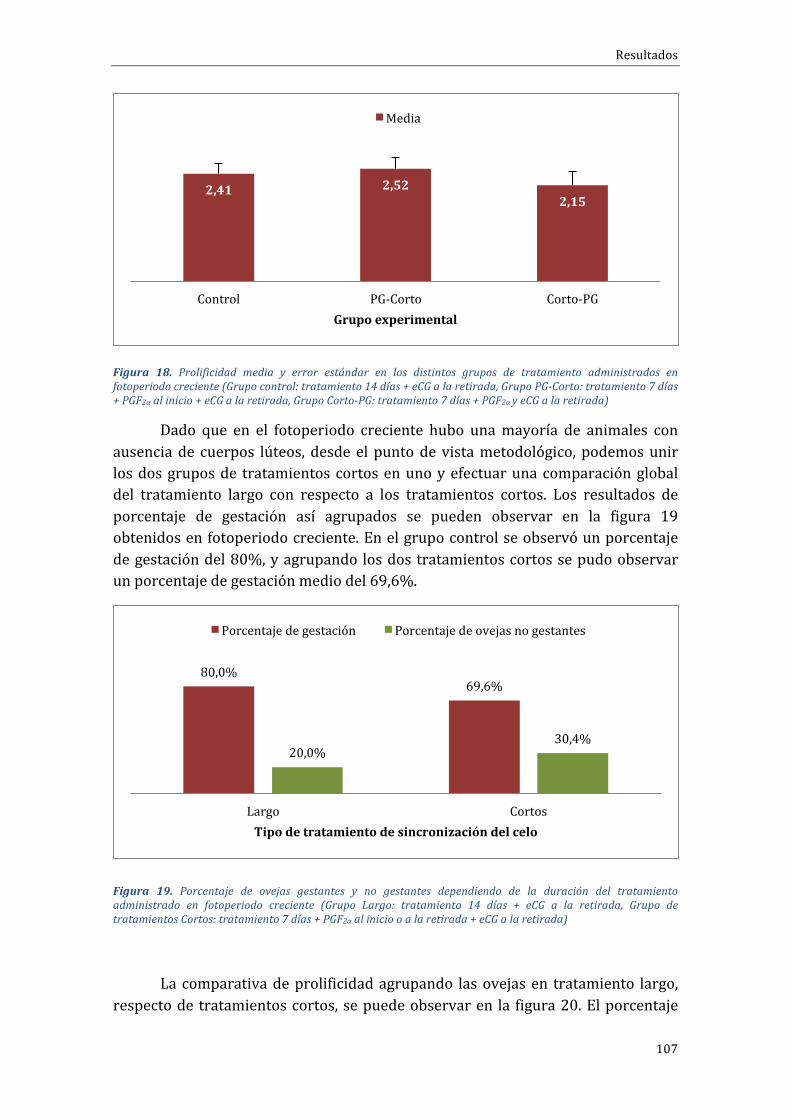
Resultados
107
Figura 18. Prolificidad media y error estándar en los distintos grupos de tratamiento administrados enfotoperiodocreciente(Grupocontrol:tratamiento14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
Dado que en el fotoperiodo creciente hubo unamayoría de animales conausenciade cuerpos lúteos,desdeelpuntodevistametodológico,podemosunirlosdosgruposde tratamientoscortosenunoyefectuarunacomparaciónglobaldel tratamiento largo con respecto a los tratamientos cortos. Los resultados deporcentaje de gestación así agrupados se pueden observar en la figura 19obtenidosenfotoperiodocreciente.Enelgrupocontrolseobservóunporcentajedegestacióndel80%,yagrupandolosdostratamientoscortossepudoobservarunporcentajedegestaciónmediodel69,6%.
Figura 19. Porcentaje de ovejas gestantes y no gestantes dependiendo de la duración del tratamientoadministrado en fotoperiodo creciente (Grupo Largo: tratamiento 14 días + eCG a la retirada, Grupo detratamientosCortos:tratamiento7días+PGF2αaliniciooalaretirada+eCGalaretirada)
Lacomparativadeprolificidadagrupando lasovejasen tratamiento largo,respectodetratamientoscortos,sepuedeobservaren la figura20.Elporcentaje
2,41 2,522,15
Control PG-Corto Corto-PGGrupoexperimental
Media
80,0%69,6%
20,0%30,4%
Largo CortosTipodetratamientodesincronizacióndelcelo
Porcentajedegestación Porcentajedeovejasnogestantes
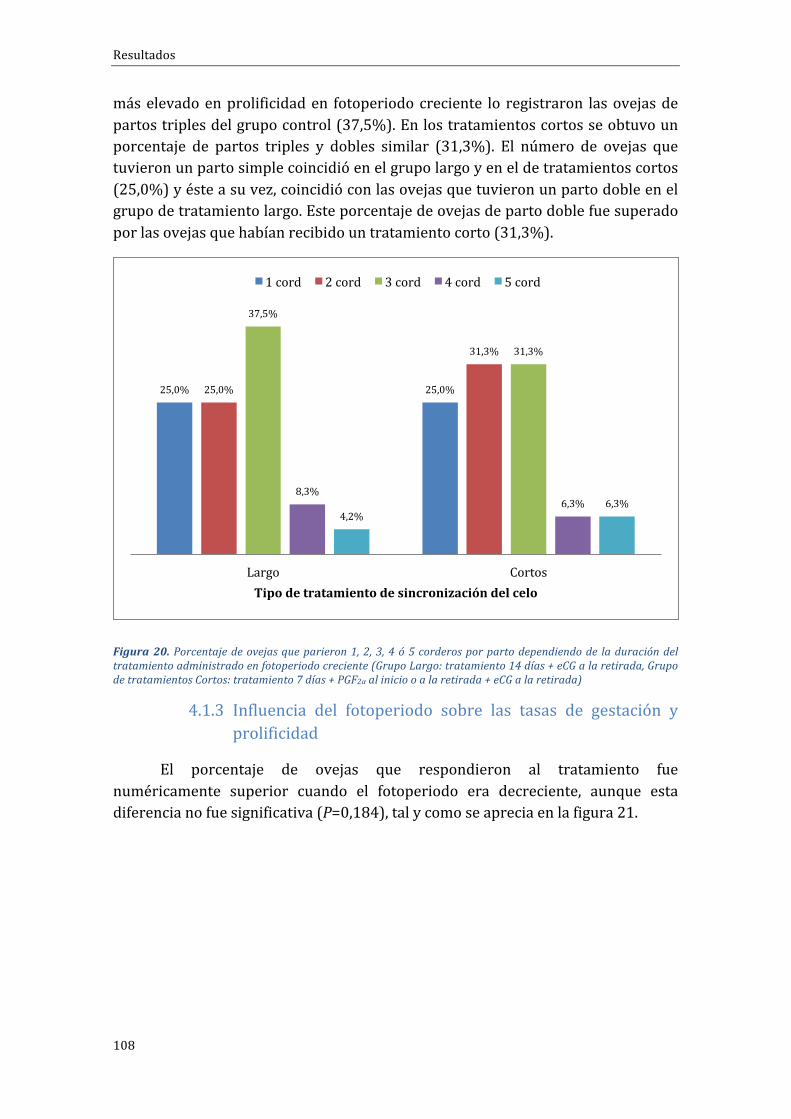
Resultados
108
máselevadoenprolificidaden fotoperiodocreciente lo registraron lasovejasdepartostriplesdelgrupocontrol(37,5%).Enlostratamientoscortosseobtuvounporcentaje de partos triples y dobles similar (31,3%). El número de ovejas quetuvieronunpartosimplecoincidióenelgrupolargoyeneldetratamientoscortos(25,0%)yésteasuvez,coincidióconlasovejasquetuvieronunpartodobleenelgrupodetratamientolargo.Esteporcentajedeovejasdepartodoblefuesuperadoporlasovejasquehabíanrecibidountratamientocorto(31,3%).
Figura20.Porcentajedeovejasqueparieron1,2,3,4ó5corderosporpartodependiendode laduracióndeltratamientoadministradoenfotoperiodocreciente(GrupoLargo:tratamiento14días+eCGalaretirada,GrupodetratamientosCortos:tratamiento7días+PGF2αaliniciooalaretirada+eCGalaretirada)
4.1.3 Influencia del fotoperiodo sobre las tasas de gestación yprolificidad
El porcentaje de ovejas que respondieron al tratamiento fuenuméricamente superior cuando el fotoperiodo era decreciente, aunque estadiferencianofuesignificativa(P=0,184),talycomoseapreciaenlafigura21.
25,0% 25,0%25,0%
31,3%
37,5%
31,3%
8,3%6,3%
4,2%6,3%
Largo CortosTipodetratamientodesincronizacióndelcelo
1cord 2cord 3cord 4cord 5cord
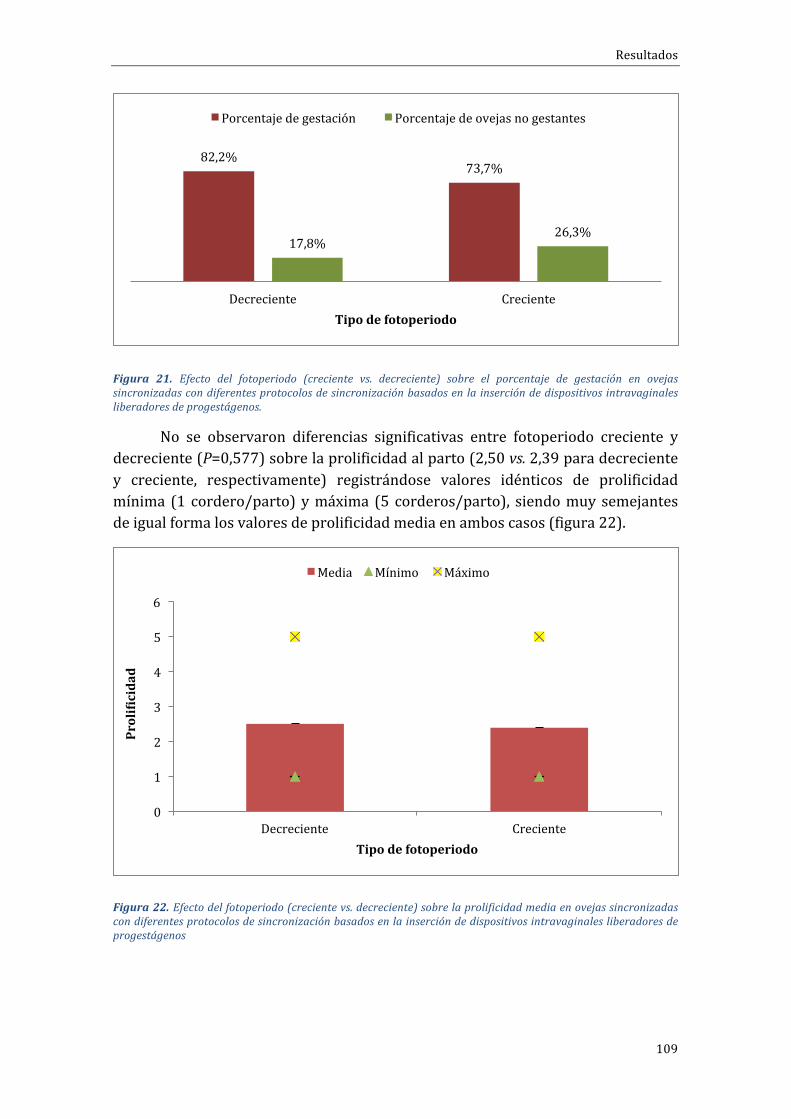
Resultados
109
Figura 21. Efecto del fotoperiodo (creciente vs. decreciente) sobre el porcentaje de gestación en ovejassincronizadascondiferentesprotocolosdesincronizaciónbasadosenlainsercióndedispositivosintravaginalesliberadoresdeprogestágenos.
No se observaron diferencias significativas entre fotoperiodo creciente ydecreciente(P=0,577)sobrelaprolificidadalparto(2,50vs.2,39paradecrecientey creciente, respectivamente) registrándose valores idénticos de prolificidadmínima (1 cordero/parto)ymáxima (5 corderos/parto), siendomuysemejantesdeigualformalosvaloresdeprolificidadmediaenamboscasos(figura22).
Figura22.Efectodelfotoperiodo(crecientevs.decreciente)sobrelaprolificidadmediaenovejassincronizadascondiferentesprotocolosdesincronizaciónbasadosenlainsercióndedispositivosintravaginalesliberadoresdeprogestágenos
82,2%73,7%
17,8%26,3%
Decreciente CrecienteTipodefotoperiodo
Porcentajedegestación Porcentajedeovejasnogestantes
0
1
2
3
4
5
6
Decreciente Creciente
Proliaicidad
Tipodefotoperiodo
Media Mínimo Máximo
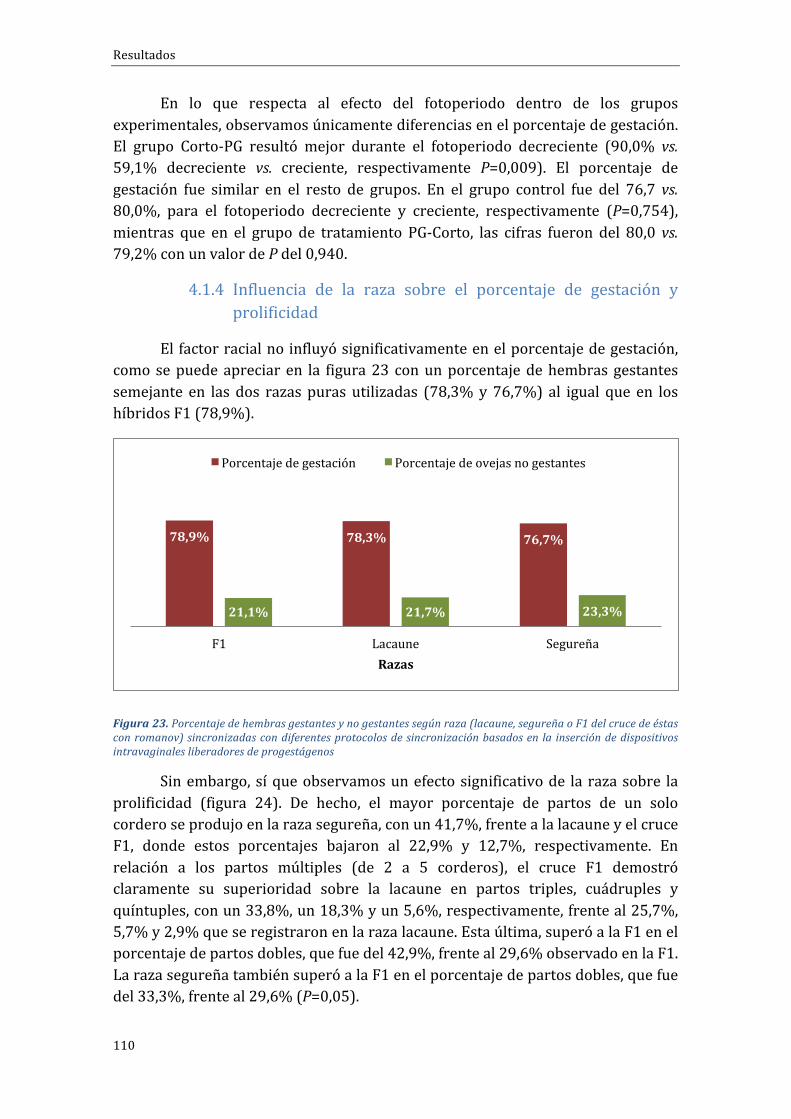
Resultados
110
En lo que respecta al efecto del fotoperiodo dentro de los gruposexperimentales,observamosúnicamentediferenciasenelporcentajedegestación.El grupo Corto-PG resultó mejor durante el fotoperiodo decreciente (90,0% vs.59,1% decreciente vs. creciente, respectivamente P=0,009). El porcentaje degestación fue similar en el resto de grupos. En el grupo control fue del 76,7 vs.80,0%, para el fotoperiodo decreciente y creciente, respectivamente (P=0,754),mientras que en el grupo de tratamiento PG-Corto, las cifras fueron del 80,0 vs.79,2%conunvalordePdel0,940.
4.1.4 Influencia de la raza sobre el porcentaje de gestación yprolificidad
Elfactorracialnoinfluyósignificativamenteenelporcentajedegestación,comosepuedeapreciar en la figura23 conunporcentajedehembrasgestantessemejante en las dos razaspuras utilizadas (78,3%y76,7%) al igual que en loshíbridosF1(78,9%).
Figura23.Porcentajedehembrasgestantesynogestantessegúnraza(lacaune,segureñaoF1delcrucedeéstasconromanov)sincronizadascondiferentesprotocolosdesincronizaciónbasadosen la insercióndedispositivosintravaginalesliberadoresdeprogestágenos
Sinembargo,síqueobservamosunefectosignificativode larazasobre laprolificidad (figura 24). De hecho, el mayor porcentaje de partos de un solocorderoseprodujoenlarazasegureña,conun41,7%,frentealalacauneyelcruceF1, donde estos porcentajes bajaron al 22,9% y 12,7%, respectivamente. Enrelación a los partos múltiples (de 2 a 5 corderos), el cruce F1 demostróclaramente su superioridad sobre la lacaune en partos triples, cuádruples yquíntuples,conun33,8%,un18,3%yun5,6%,respectivamente,frenteal25,7%,5,7%y2,9%queseregistraronenlarazalacaune.Estaúltima,superóalaF1enelporcentajedepartosdobles,quefuedel42,9%,frenteal29,6%observadoenlaF1.LarazasegureñatambiénsuperóalaF1enelporcentajedepartosdobles,quefuedel33,3%,frenteal29,6%(P=0,05).
78,9% 78,3% 76,7%
21,1% 21,7% 23,3%
F1 Lacaune SegureñaRazas
Porcentajedegestación Porcentajedeovejasnogestantes
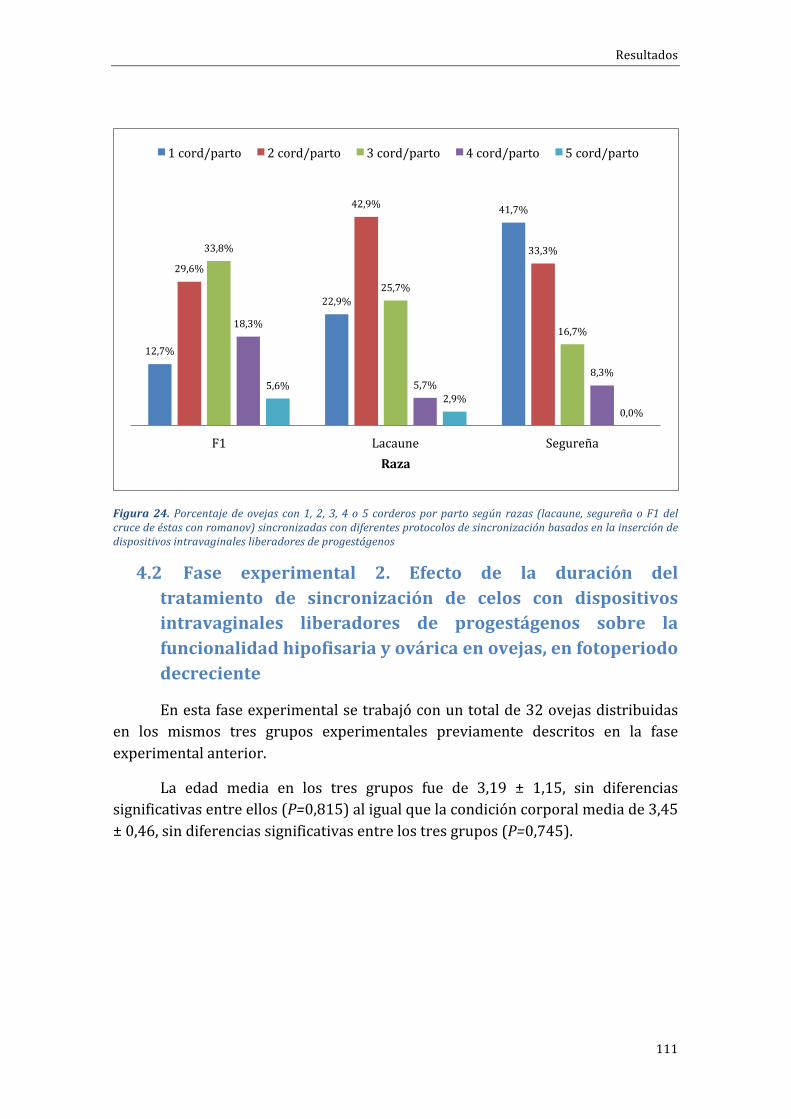
Resultados
111
Figura 24.Porcentajedeovejas con1,2,3,4o5 corderosporparto segúnrazas (lacaune, segureñaoF1delcrucedeéstasconromanov)sincronizadascondiferentesprotocolosdesincronizaciónbasadosenlainsercióndedispositivosintravaginalesliberadoresdeprogestágenos
4.2 Fase experimental 2. Efecto de la duración deltratamiento de sincronización de celos con dispositivosintravaginales liberadores de progestágenos sobre lafuncionalidadhipofisariayováricaenovejas,enfotoperiododecreciente
Enestafaseexperimentalsetrabajóconuntotalde32ovejasdistribuidasen los mismos tres grupos experimentales previamente descritos en la faseexperimentalanterior.
La edad media en los tres grupos fue de 3,19 ± 1,15, sin diferenciassignificativasentreellos(P=0,815)aligualquelacondicióncorporalmediade3,45±0,46,sindiferenciassignificativasentrelostresgrupos(P=0,745).
12,7%
22,9%
41,7%
29,6%
42,9%
33,3%33,8%
25,7%
16,7%18,3%
5,7%8,3%
5,6%2,9%
0,0%
F1 Lacaune SegureñaRaza
1cord/parto 2cord/parto 3cord/parto 4cord/parto 5cord/parto

Resultados
112
4.2.1 Influencia del tipo de tratamiento sobre el porcentaje depresentacióndeceloymomentodeaparicióndelcelo
La determinación del momento de aparición del celo, realizada cada 4hdesdelas12hhastalas80hdespuésdelaretiradadelasesponjas,mostróqueelporcentaje de hembras que salieron en celo fue del 87,66%. Los porcentajes derespuestaentregrupos fueron100%enelgrupocontrol,63,6%enelgrupoPG-Cortoydel90,9%enel grupoCorto-PG.Estasdiferenciasnuméricas,mostrabanunatendenciaestadísticamentesignificativa(P=0,065;Tabla5),queratificabaqueenelgrupoPG-Cortoseobtuvounamenorrespuestaaltratamientoenrelaciónalosotrosdosgrupos(controlyCorto-PG).
Tabla5.Porcentajedepresentacióndecelosegúngrupoexperimental
Grupodetratamiento Respuestaaltratamiento
Control 100,0%
PG-Corto 63,6%
Corto-PG 90,9%
(Grupocontrol:tratamiento“largo”de14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
El análisis de regresión logística, considerando la variable dependiente lapresentacióndecelocorroborólosresultadosdescritospreviamentesinencontrarsignificancia en las odds ratio de ninguna de los posibles factores de confusión(edad ni condición corporal) ni de la variable independiente o tratamiento, convaloresP>0,05entodosloscasos.
Eltiempo(T)mediodesalidaencelofuede42,31±14,88h,noexistiendodiferencias significativas entre los tres grupos (P=0,165; tabla 6). En el grupocontrol, el tiempomedio fue de 35,11 ± 5,20 h; en el grupo PG-Corto, de 43,43±19,24h;yengrupoCorto-PG,de48,0±16,0h.
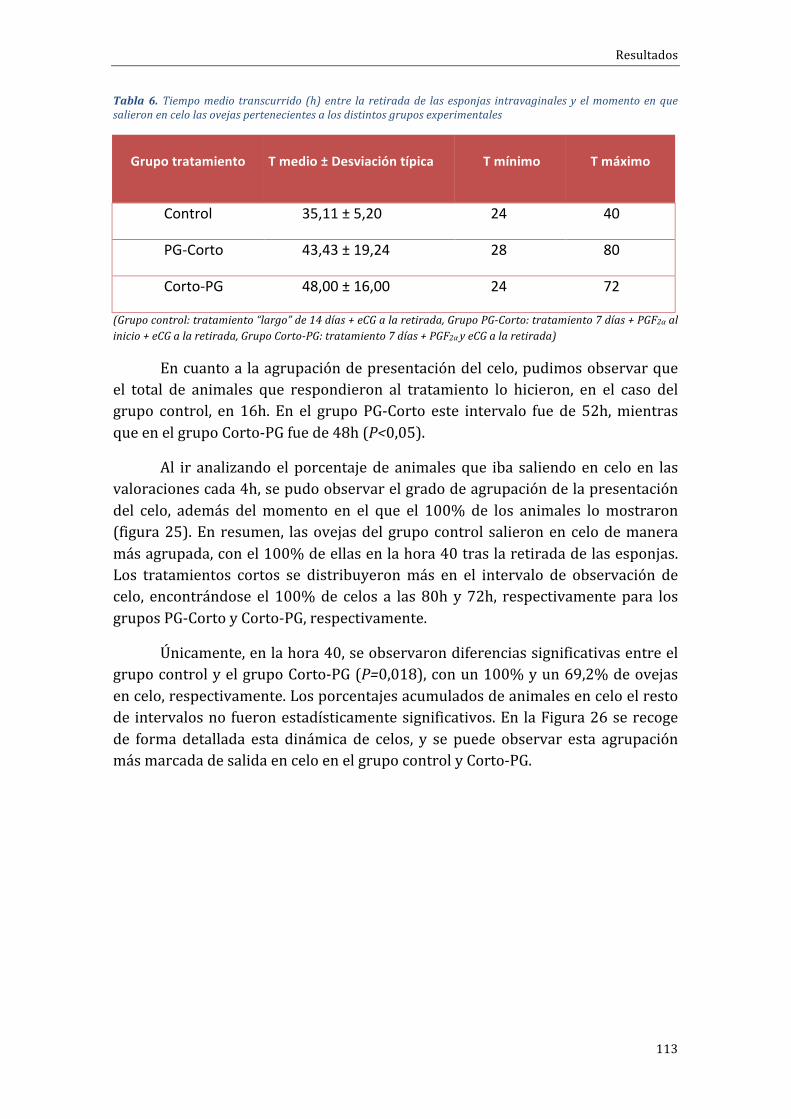
Resultados
113
Tabla 6.Tiempomedio transcurrido (h)entre la retiradade las esponjas intravaginalesy elmomentoenquesalieronencelolasovejaspertenecientesalosdistintosgruposexperimentales
Grupotratamiento Tmedio±Desviacióntípica Tmínimo Tmáximo
Control 35,11±5,20 24 40
PG-Corto 43,43±19,24 28 80
Corto-PG 48,00±16,00 24 72
(Grupocontrol:tratamiento“largo”de14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
Encuantoalaagrupacióndepresentacióndelcelo,pudimosobservarqueel total de animales que respondieron al tratamiento lo hicieron, en el caso delgrupo control, en 16h. En el grupoPG-Corto este intervalo fue de 52h,mientrasqueenelgrupoCorto-PGfuede48h(P<0,05).
Al ir analizandoelporcentajede animalesque iba saliendoen celo en lasvaloracionescada4h,sepudoobservarelgradodeagrupacióndelapresentacióndel celo, además delmomento en el que el 100% de los animales lomostraron(figura25).Enresumen, lasovejasdelgrupocontrol salieronencelodemaneramásagrupada,conel100%deellasenlahora40traslaretiradadelasesponjas.Los tratamientos cortos se distribuyeronmás en el intervalo de observación decelo, encontrándose el 100%de celos a las 80h y72h, respectivamentepara losgruposPG-CortoyCorto-PG,respectivamente.
Únicamente,enlahora40,seobservarondiferenciassignificativasentreelgrupocontrolyelgrupoCorto-PG(P=0,018),conun100%yun69,2%deovejasencelo,respectivamente.Losporcentajesacumuladosdeanimalesenceloelrestode intervalosno fueronestadísticamentesignificativos.En laFigura26serecogede forma detallada esta dinámica de celos, y se puede observar esta agrupaciónmásmarcadadesalidaenceloenelgrupocontrolyCorto-PG.
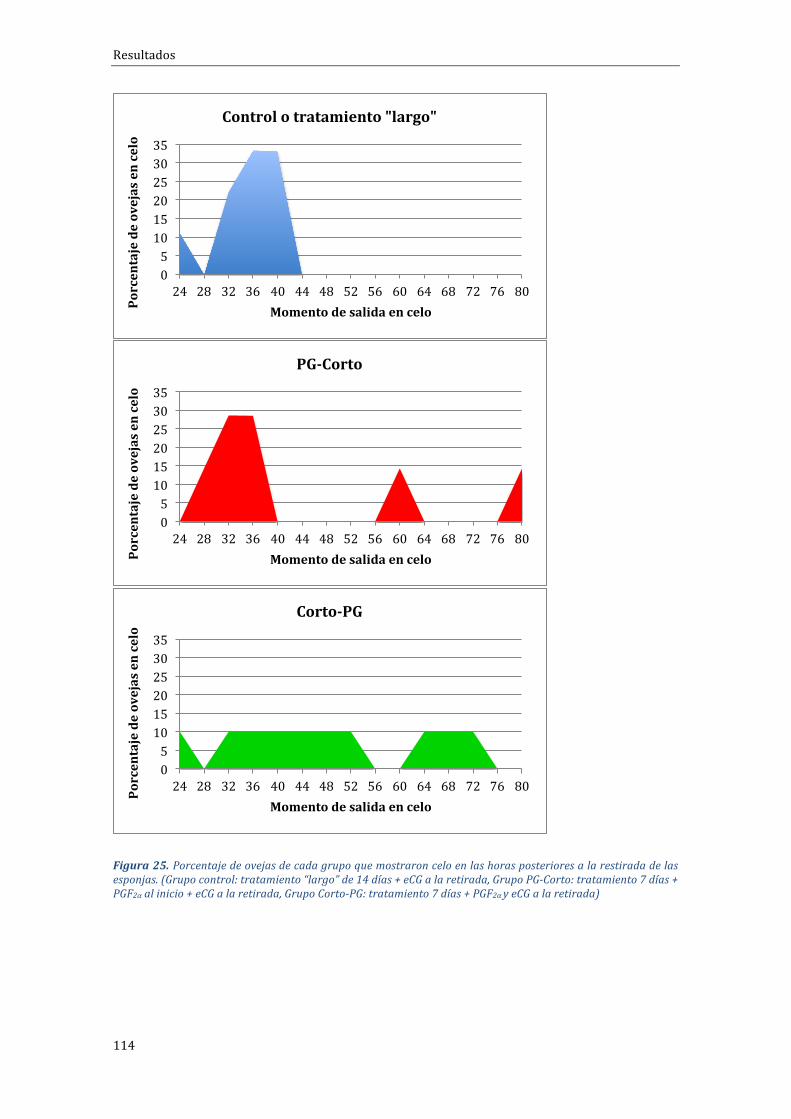
Resultados
114
Figura25.Porcentajedeovejasdecadagrupoquemostraronceloenlashorasposterioresalarestiradadelasesponjas.(Grupocontrol:tratamiento“largo”de14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
05101520253035
24 28 32 36 40 44 48 52 56 60 64 68 72 76 80
Porcentajedeovejasencelo
Momentodesalidaencelo
Controlotratamiento"largo"
05101520253035
24 28 32 36 40 44 48 52 56 60 64 68 72 76 80
Porcentajedeovejasencelo
Momentodesalidaencelo
PG-Corto
05101520253035
24 28 32 36 40 44 48 52 56 60 64 68 72 76 80Porcentajedeovejasencelo
Momentodesalidaencelo
Corto-PG
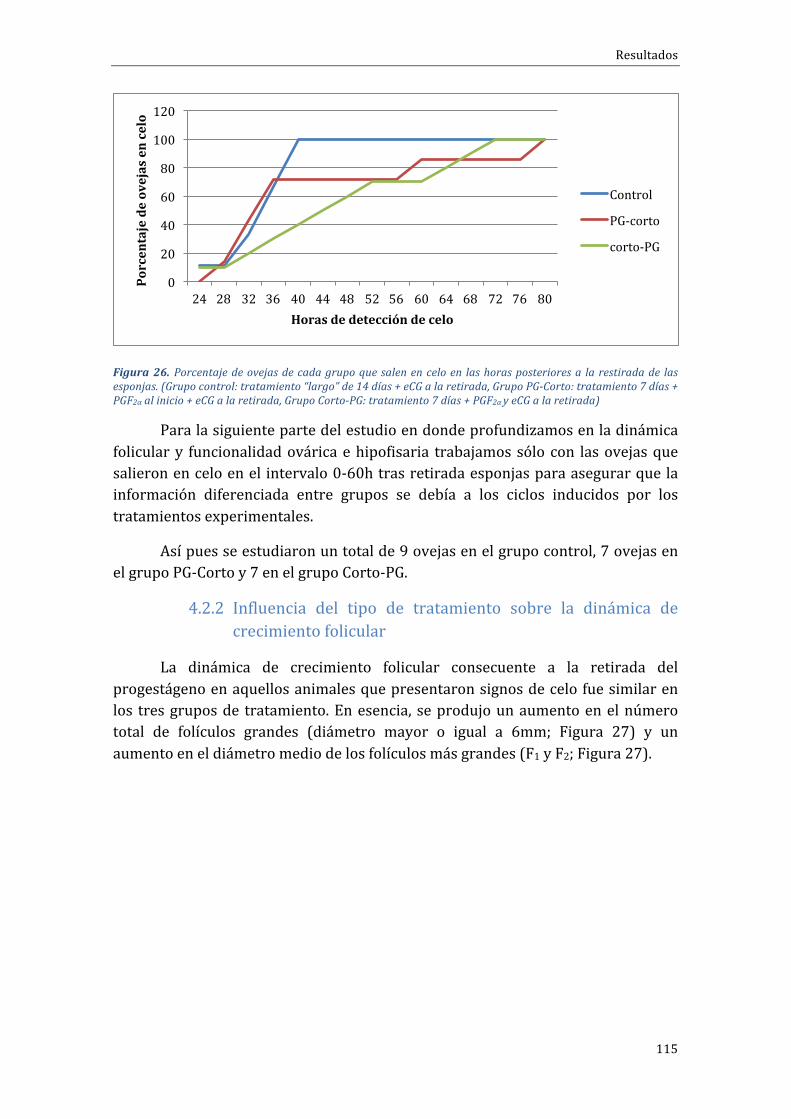
Resultados
115
Figura26.Porcentajedeovejasdecadagrupoquesalenenceloen lashorasposterioresa larestiradade lasesponjas.(Grupocontrol:tratamiento“largo”de14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
Paralasiguientepartedelestudioendondeprofundizamosenladinámicafoliculary funcionalidadováricaehipofisaria trabajamossólocon lasovejasquesalieronenceloenel intervalo0-60htrasretiradaesponjasparaasegurarquelainformación diferenciada entre grupos se debía a los ciclos inducidos por lostratamientosexperimentales.
Asípuesseestudiaronuntotalde9ovejasenelgrupocontrol,7ovejasenelgrupoPG-Cortoy7enelgrupoCorto-PG.
4.2.2 Influencia del tipo de tratamiento sobre la dinámica decrecimientofolicular
La dinámica de crecimiento folicular consecuente a la retirada delprogestágenoenaquellosanimalesquepresentaronsignosdecelofuesimilarenlos tresgruposde tratamiento.Enesencia, seprodujounaumentoenelnúmerototal de folículos grandes (diámetro mayor o igual a 6mm; Figura 27) y unaumentoeneldiámetromediodelosfolículosmásgrandes(F1yF2;Figura27).
0
20
40
60
80
100
120
24 28 32 36 40 44 48 52 56 60 64 68 72 76 80
Porcentajedeovejasencelo
Horasdedeteccióndecelo
Control
PG-corto
corto-PG
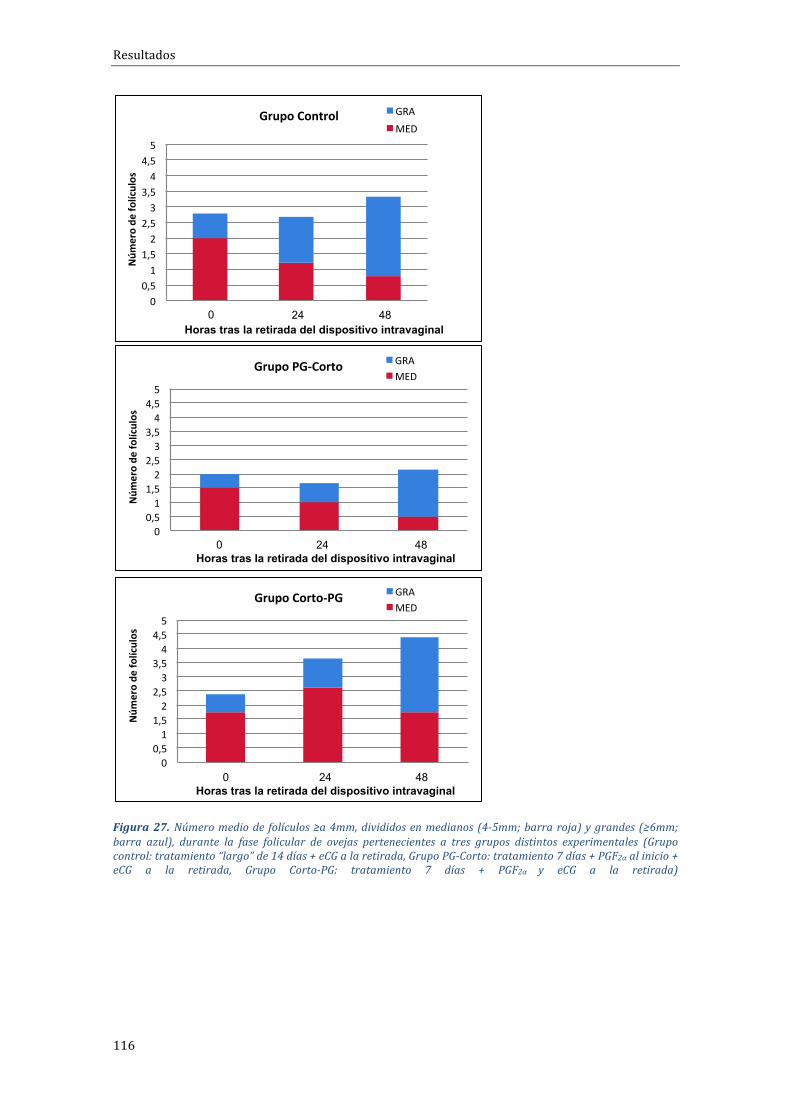
Resultados
116
Figura27.Númeromediode folículos≥a4mm,divididosenmedianos (4-5mm;barraroja)ygrandes (≥6mm;barra azul), durante la fase folicular de ovejas pertenecientes a tres grupos distintos experimentales (Grupocontrol:tratamiento“largo”de14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCG a la retirada, Grupo Corto-PG: tratamiento 7 días + PGF2α y eCG a la retirada)
00,51
1,52
2,53
3,54
4,55
1 2 3
Núm
erode
folículos
GrupoControl GRAMED
Horas tras la retirada del dispositivo intravaginal 0 24 48
00,51
1,52
2,53
3,54
4,55
1 2 3
Núm
erode
folículos
GrupoPG-Corto GRAMED
0 24 48 Horas tras la retirada del dispositivo intravaginal
00,51
1,52
2,53
3,54
4,55
1 2 3
Núm
erode
folículos
GrupoCorto-PG GRAMED
0 24 48 Horas tras la retirada del dispositivo intravaginal
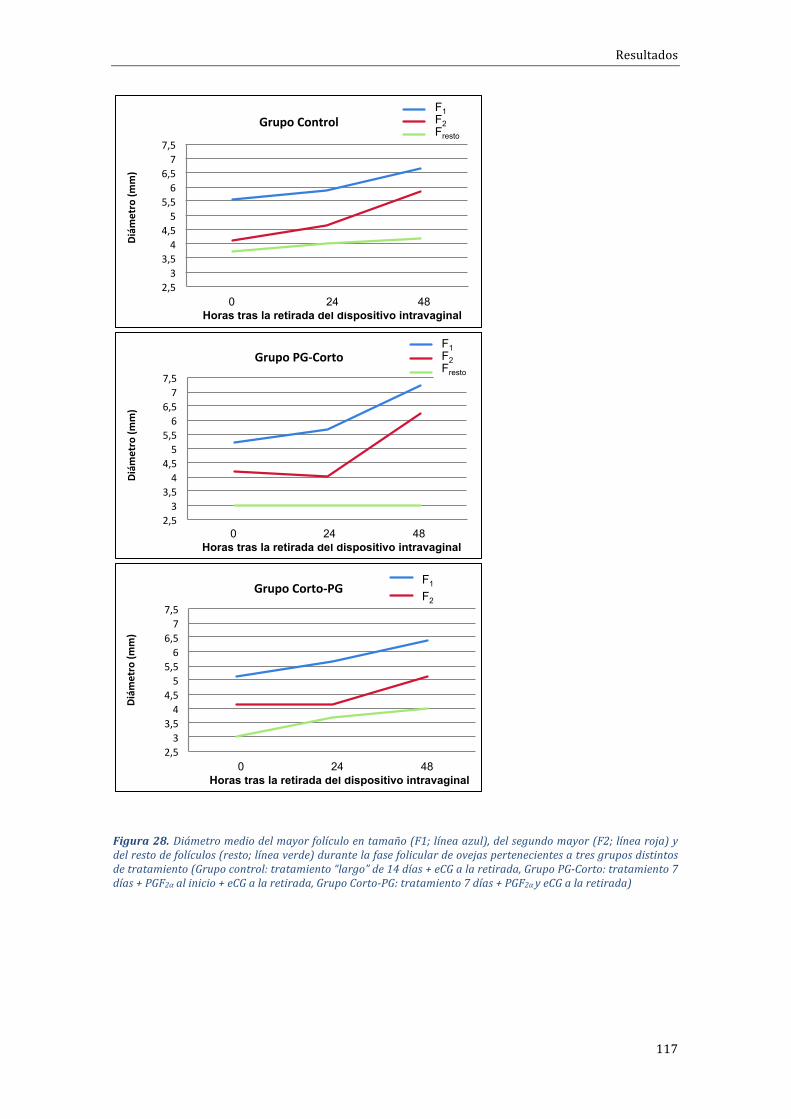
Resultados
117
Figura28.Diámetromediodelmayorfolículoentamaño(F1;líneaazul),delsegundomayor(F2;línearoja)ydelrestodefolículos(resto;líneaverde)durantelafasefoliculardeovejaspertenecientesatresgruposdistintosdetratamiento(Grupocontrol:tratamiento“largo”de14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
2,53
3,54
4,55
5,56
6,57
7,5
1 2 3
Diám
etro(m
m)
GrupoControlF1F2Resto
Horas tras la retirada del dispositivo intravaginal 0 24 48
F1 F2 Fresto
2,53
3,54
4,55
5,56
6,57
7,5
1 2 3
Diám
etro(m
m)
GrupoPG-CortoF1F2Resto
Horas tras la retirada del dispositivo intravaginal 0 24 48
F1 F2 Fresto
2,53
3,54
4,55
5,56
6,57
7,5
1 2 3
Diám
etro(m
m)
GrupoCorto-PGF1F2
Horas tras la retirada del dispositivo intravaginal 0 24 48
F1
F2

Resultados
118
Sin embargo, si bien los perfiles de crecimiento de los folículos fueronsimilares, hubodiferencias significativas enelnúmerode folículos entre los tresgrupos. Observamos que el número de folículos medianos descendió desde elmomentoderetiradadelprogestágenohasta48hdespuésenlosgruposcontrolyPG-Corto;porelcontrario,enelgrupoCorto-PGaumentóentrelas0hy24h,paradescender entre las 24h y 48h, aunque las diferencias no fueron significativas(P=0,231 yP=0,371, para la comparativa 0-24h y 24-48h, respectivamente). Sinembargo,apesardeestafaltadesignificancia,estadiferencianuméricaresultó,alas24h,enunnúmerodefolículosmedianos(4-5mm)mayorenelgrupoCorto-PGqueenlosotrosdosgrupos(P<0,05).Estasdiferenciassehicieronmásevidentesalconsiderarsólolosfolículosqueseencontrabanencrecimientoencadaunadelas observaciones (Figura 28); ya que se observó prácticamente el triple defolículosmedianosenelgrupoCorto-PGrespectoa losotrosgrupos(P<0,05).Encuanto a la evolución de la cantidad de folículos grandes en los diferentesmomentosencadagrupodetratamiento,observamosunaumentodelosmismosde0a24hentodoslosgrupos,sindiferenciassignificativasentremomentosparaningunode losgrupos(P=0,111;P=0,611yP=0.442).Sinembargo,enelpasode24 a 48h tras la retirada de los dispositivos, los grupos de tratamientos cortosmostraron tendencia y significancia estadística en la elevación del número defolículosgrandesdelahora24ala48(de0,67a1,67;P=0,076enPG-Cortoyde1,00a2,62;P=0,003enPG-Corto).
Enlafigura28sepuedeapreciartambiénqueelnúmeromediodefolículosgrandesencrecimientoalas48h(esdecir,losfolículosovulatorios)fuemayorenlosgruposquerecibieronuntratamientocorto(PG-CortoyCorto-PG),respectodelgrupo control, en que se llevó a cabo un tratamiento largo, lo que refleja estecrecimiento significativo previamente descrito de los folículos en estos grupos“cortos”.
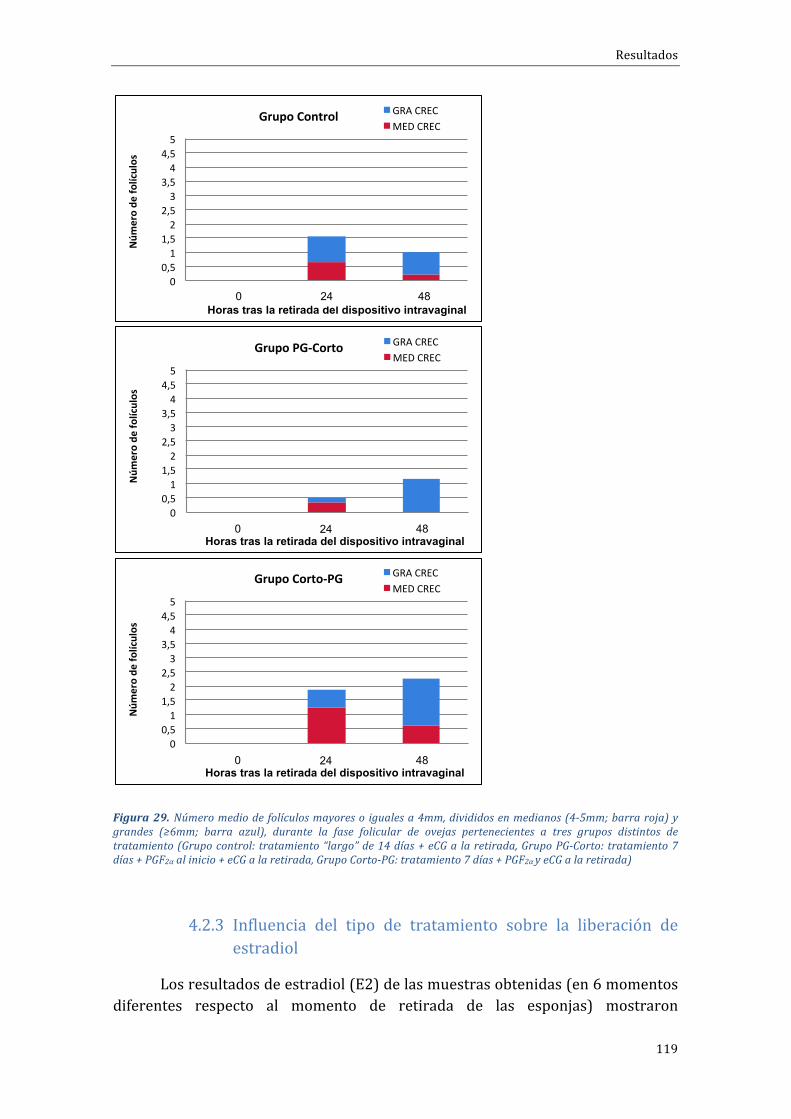
Resultados
119
Figura29.Númeromediode folículosmayoreso igualesa4mm,divididosenmedianos(4-5mm;barraroja)ygrandes (≥6mm; barra azul), durante la fase folicular de ovejas pertenecientes a tres grupos distintos detratamiento(Grupocontrol: tratamiento“largo”de14días+eCGa laretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
4.2.3 Influencia del tipo de tratamiento sobre la liberación deestradiol
Losresultadosdeestradiol(E2)delasmuestrasobtenidas(en6momentosdiferentes respecto al momento de retirada de las esponjas) mostraron
00,51
1,52
2,53
3,54
4,55
1 2 3
Núm
erode
folículos
GrupoControl GRACRECMEDCREC
Horas tras la retirada del dispositivo intravaginal 0 24 48
00,51
1,52
2,53
3,54
4,55
1 2 3
Núm
erode
folículos
GrupoPG-Corto GRACRECMEDCREC
0 24 48 Horas tras la retirada del dispositivo intravaginal
00,51
1,52
2,53
3,54
4,55
1 2 3
Núm
erode
folículos
GrupoCorto-PG GRACRECMEDCREC
0 24 48 Horas tras la retirada del dispositivo intravaginal
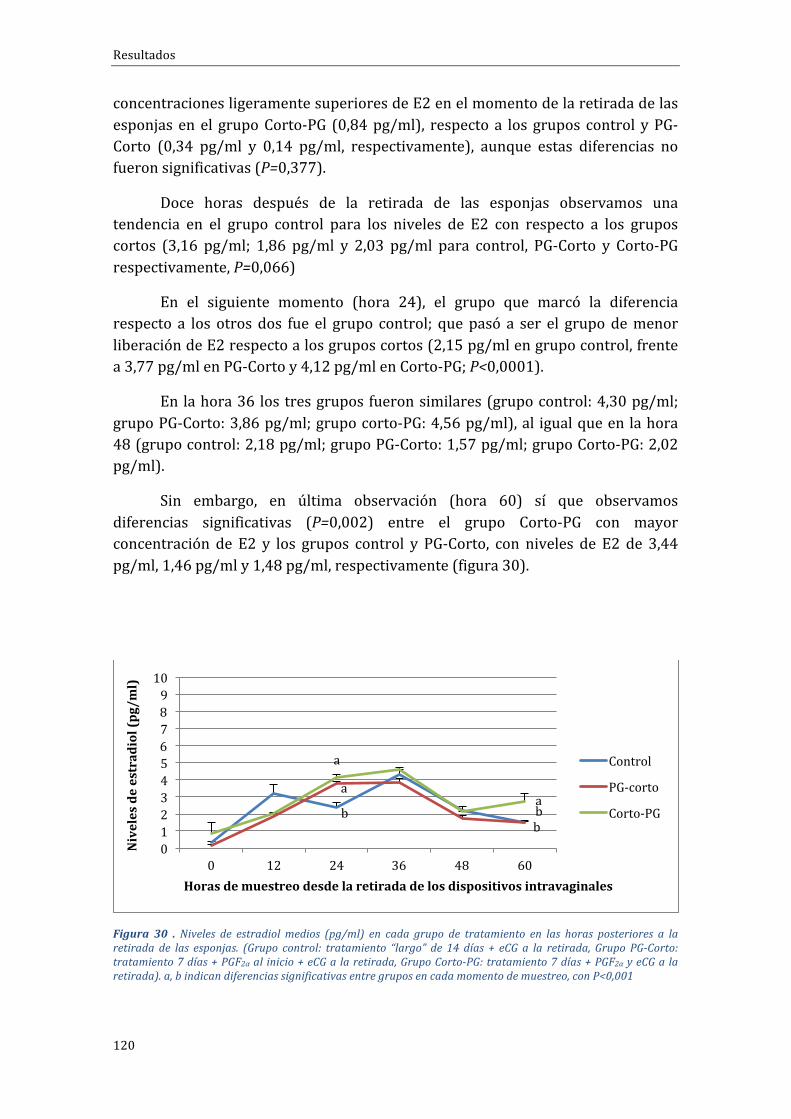
Resultados
120
concentracionesligeramentesuperioresdeE2enelmomentodelaretiradadelasesponjas enel grupoCorto-PG (0,84pg/ml), respectoa los grupos control yPG-Corto (0,34 pg/ml y 0,14 pg/ml, respectivamente), aunque estas diferencias nofueronsignificativas(P=0,377).
Doce horas después de la retirada de las esponjas observamos unatendencia en el grupo control para los niveles de E2 con respecto a los gruposcortos (3,16 pg/ml; 1,86 pg/ml y 2,03 pg/ml para control, PG-Corto y Corto-PGrespectivamente,P=0,066)
En el siguiente momento (hora 24), el grupo que marcó la diferenciarespecto a los otros dos fue el grupo control; quepasó a ser el grupodemenorliberacióndeE2respectoalosgruposcortos(2,15pg/mlengrupocontrol,frentea3,77pg/mlenPG-Cortoy4,12pg/mlenCorto-PG;P<0,0001).
En lahora36 lostresgruposfueronsimilares(grupocontrol:4,30pg/ml;grupoPG-Corto:3,86pg/ml;grupocorto-PG:4,56pg/ml),al igualqueenlahora48(grupocontrol:2,18pg/ml;grupoPG-Corto:1,57pg/ml;grupoCorto-PG:2,02pg/ml).
Sin embargo, en última observación (hora 60) sí que observamosdiferencias significativas (P=0,002) entre el grupo Corto-PG con mayorconcentración de E2 y los grupos control y PG-Corto, con niveles de E2 de 3,44pg/ml,1,46pg/mly1,48pg/ml,respectivamente(figura30).
Figura 30 . Niveles de estradiol medios (pg/ml) en cada grupo de tratamiento en las horas posteriores a laretirada de las esponjas. (Grupo control: tratamiento “largo” de 14 días + eCG a la retirada, Grupo PG-Corto:tratamiento7días+PGF2αal inicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada).a,bindicandiferenciassignificativasentregruposencadamomentodemuestreo,conP<0,001
bb
a
b
a
a
012345678910
0 12 24 36 48 60
Nivelesdeestradiol(pg/m
l)
Horasdemuestreodesdelaretiradadelosdispositivosintravaginales
Control
PG-corto
Corto-PG

Resultados
121
4.2.4 Influencia del tipo de tratamiento sobre el momento deaparición del pico de LH y sobre la concentraciónmáximadeLH
En relación a las concentraciones de LH observamos que en todas lashembras en celo, independientemente del grupo experimental, tuvo lugar unadescarga preovulatoria de LH posterior a la retirada de los dispositivos. Noobservamos diferencias en elmomento de aparición de la descarga de LH entregrupos(tabla7),sibienseobservóunmayoragrupamientoenelcasodelgrupoCorto-PG(figura31),conaproximadamenteel90%de lashembrascondescargadeLHenlas8horasposterioresalaaparicióndelcelo.
Tabla7.MediaydesviacióntípicadelmomentodeaparicióndelpicodeLH,enhoras,yconcentraciónmáximadeLH,enng/ml,enlasovejaspertenecientesalosdistintosgruposexperimentales
GrupoControl
Grupo
PG-Corto
Grupo
Corto-PG
MomentoapariciónpicoLH(h) 10,67±4,90 12,00±5,66 8,00±3,70
ConcentraciónmáximapicoLH(ng/ml) 38,95±27,66 12,40±4,40 47,13±30,51
(Grupocontrol:tratamiento“largo”de14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
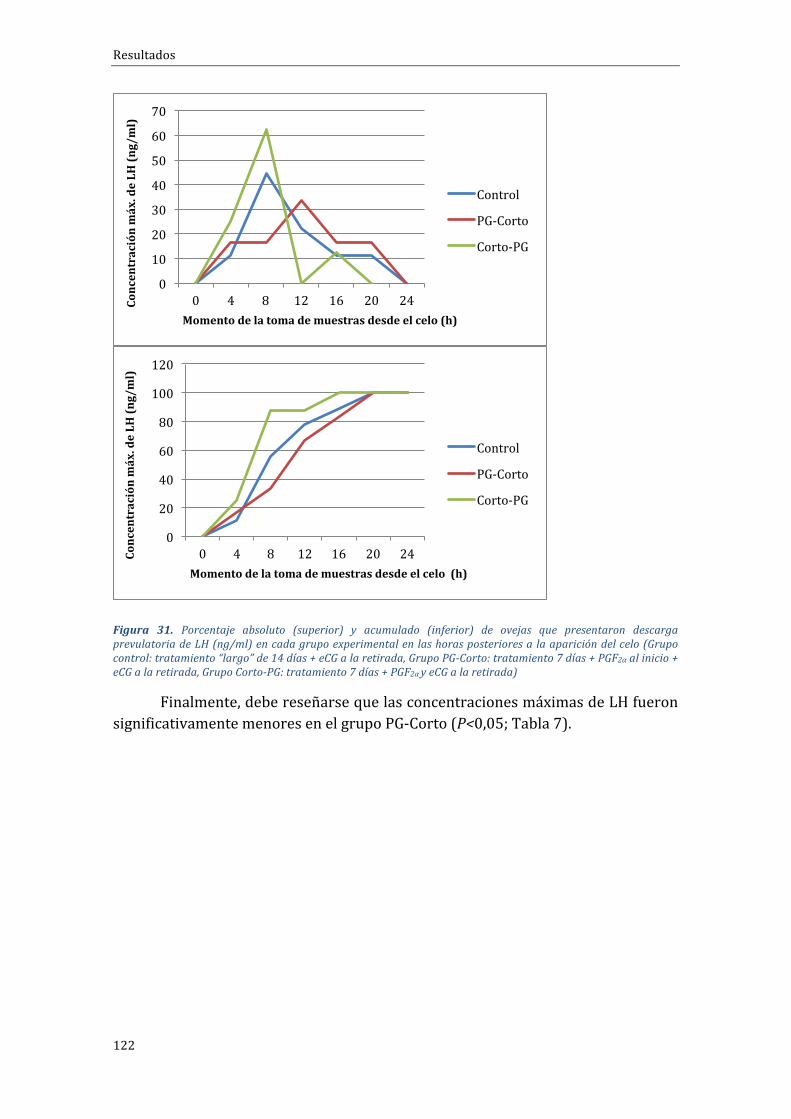
Resultados
122
Figura 31. Porcentaje absoluto (superior) y acumulado (inferior) de ovejas que presentaron descargaprevulatoriadeLH(ng/ml)encadagrupoexperimentalenlashorasposterioresalaaparicióndelcelo(Grupocontrol:tratamiento“largo”de14días+eCGalaretirada,GrupoPG-Corto:tratamiento7días+PGF2αalinicio+eCGalaretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
Finalmente,debereseñarsequelasconcentracionesmáximasdeLHfueronsignificativamentemenoresenelgrupoPG-Corto(P<0,05;Tabla7).
0
10
20
30
40
50
60
70
0 4 8 12 16 20 24Concentraciónmáx.deLH(ng/ml)
Momentodelatomademuestrasdesdeelcelo(h)
Control
PG-Corto
Corto-PG
0
20
40
60
80
100
120
0 4 8 12 16 20 24Concentraciónmáx.deLH(ng/ml)
Momentodelatomademuestrasdesdeelcelo(h)
Control
PG-Corto
Corto-PG
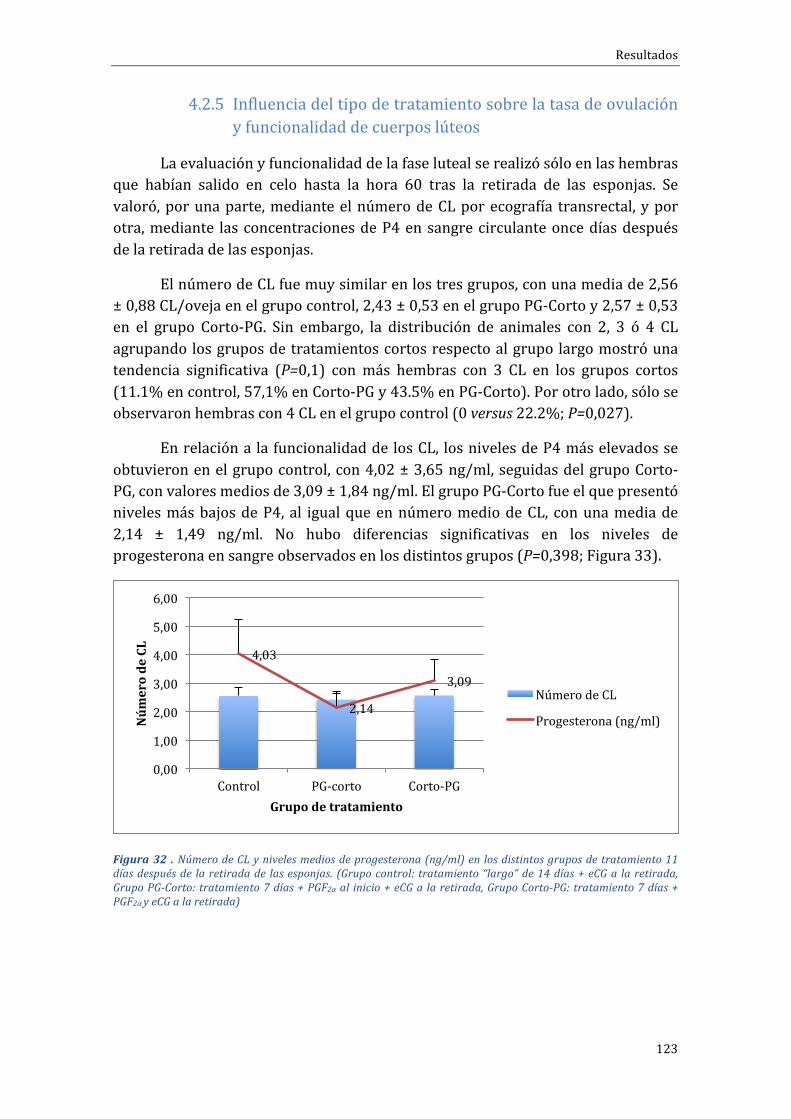
Resultados
123
4.2.5 Influenciadeltipodetratamientosobrelatasadeovulaciónyfuncionalidaddecuerposlúteos
Laevaluaciónyfuncionalidaddelafaselutealserealizósóloenlashembrasque habían salido en celo hasta la hora 60 tras la retirada de las esponjas. Sevaloró,porunaparte,medianteelnúmerodeCLporecografía transrectal,yporotra,mediante lasconcentracionesdeP4ensangrecirculanteoncedíasdespuésdelaretiradadelasesponjas.
ElnúmerodeCLfuemuysimilarenlostresgrupos,conunamediade2,56±0,88CL/ovejaenelgrupocontrol,2,43±0,53enelgrupoPG-Cortoy2,57±0,53en el grupo Corto-PG. Sin embargo, la distribución de animales con 2, 3 ó 4 CLagrupandolosgruposdetratamientoscortosrespectoalgrupolargomostróunatendencia significativa (P=0,1) conmás hembras con 3 CL en los grupos cortos(11.1%encontrol,57,1%enCorto-PGy43.5%enPG-Corto).Porotrolado,sóloseobservaronhembrascon4CLenelgrupocontrol(0versus22.2%;P=0,027).
Enrelacióna lafuncionalidaddelosCL, losnivelesdeP4máselevadosseobtuvieronenelgrupocontrol,con4,02±3,65ng/ml,seguidasdelgrupoCorto-PG,convaloresmediosde3,09±1,84ng/ml.ElgrupoPG-CortofueelquepresentónivelesmásbajosdeP4,al igualqueennúmeromediodeCL, conunamediade2,14 ± 1,49 ng/ml. No hubo diferencias significativas en los niveles deprogesteronaensangreobservadosenlosdistintosgrupos(P=0,398;Figura33).
Figura32 .NúmerodeCLynivelesmediosdeprogesterona(ng/ml)enlosdistintosgruposdetratamiento11díasdespuésde laretiradade lasesponjas. (Grupocontrol: tratamiento“largo”de14días+eCGa laretirada,GrupoPG-Corto:tratamiento7días+PGF2αal inicio+eCGa laretirada,GrupoCorto-PG:tratamiento7días+PGF2αyeCGalaretirada)
4,03
2,14
3,09
0,00
1,00
2,00
3,00
4,00
5,00
6,00
Control PG-corto Corto-PG
Núm
erodeCL
Grupodetratamiento
NúmerodeCL
Progesterona(ng/ml)

Resultados
124
4.3 Fase experimental 3. Efecto del tipo de dispositivoprogestativo intravaginal (progesterona natural o CIDRversusprogestágenosintéticooFGA)sobreelporcentajedegestación en ovejas tratadas con un protocolo desincronización de celos largo (14 días) en fotoperiodocreciente
En esta fase experimental se profundizó en la utilización de un nuevodispositivointravaginalconprogesteronanatural(CIDR®)encomparaciónconlasesponjasdeprogestágenoshabitualesenbaseaFGA,queseutilizaronenlasfasesexperimentalesprevias,1y2.
El tratamientode sincronización aplicadopara la comparación fue elmásampliamente difundido en la actualidad: el tratamiento de 14 días demantenimientode losdispositivos intravaginalmenteyadministraciónde400UIde eCG i.m. a la retiradade losmismos.El fotoperiodoen el que se efectuó esteexperimentofueeldecreciente.
Inicialmente, losgruposexperimentalesFGAyCIDRestabanformadospor186y117ovejas,respectivamente,ydeellasseperdieron7animalesenelprimergrupoy12enelsegundo.Estasbajasfueronprovocadasporeliminación,porerrordeseparacióndelotesenelmomentodecolocarlosdispositivosypormortalidad.Debido a estas pérdidas en ambos grupos, FGA y CIDR, se insertaron179 y 105dispositivos,respectivamente.
Estos animales se distribuyeron aleatoriamente, quedando los animalesrepartidos homogéneamente entre los dos grupos en lo que a edad y condicióncorporalserefiere.
Tal y como se detalla en el apartado dematerial ymétodos del presenteexperimento, las pérdidas de dispositivo vaginal observadas en los dos gruposexperimentalesfuerondel4,469%(8/179)enelgrupoFGAydel1,905%(2/105)en el grupo CIDR. Estos diferentes porcentajes numéricos no fueronestadísticamentesignificativos(P=0,332).
Porlotanto,enelmomentodelaretiradadelosdispositivos,elgrupoFGAhabíaquedadoreducidoa171animalesyelgrupoCIDRa103animales.40díasdespués,enelmomentodehacereldiagnósticodegestación,otras4ovejashabíandesaparecidodelgrupoFGA,bienporeliminación,porerrordecambioaotroloteopormortalidad.Porestasmismasrazonesdesaparecieron8animalesdelgrupoCIDR.
Finalmente, en el estudio de fertilidad, quedaron 262 ovejas,: 167 sesincronizaron y diagnosticaron con esponjas FGA mientras que 95 se
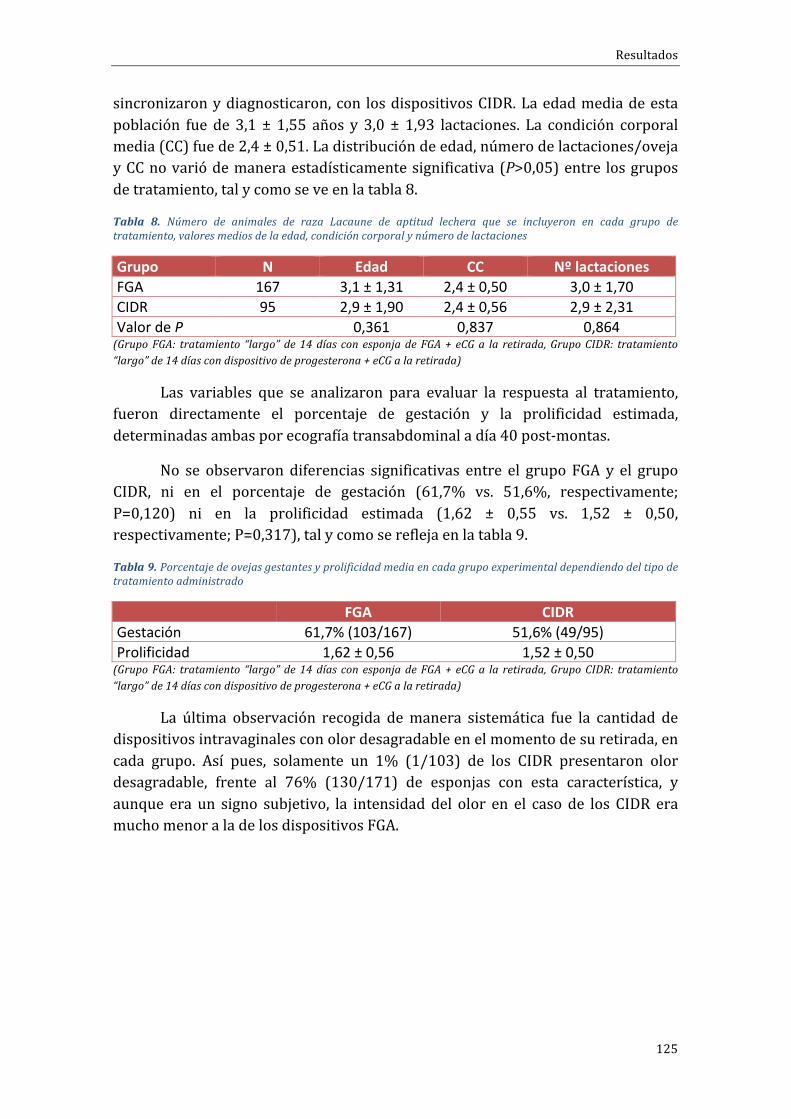
Resultados
125
sincronizaronydiagnosticaron,con losdispositivosCIDR.Laedadmediadeestapoblación fue de 3,1 ± 1,55 años y 3,0 ± 1,93 lactaciones. La condición corporalmedia(CC)fuede2,4±0,51.Ladistribucióndeedad,númerodelactaciones/ovejayCCnovariódemaneraestadísticamentesignificativa(P>0,05)entre losgruposdetratamiento,talycomoseveenlatabla8.
Tabla 8. Número de animales de raza Lacaune de aptitud lechera que se incluyeron en cada grupo detratamiento,valoresmediosdelaedad,condicióncorporalynúmerodelactaciones
Grupo N Edad CC NºlactacionesFGA 167 3,1±1,31 2,4±0,50 3,0±1,70CIDR 95 2,9±1,90 2,4±0,56 2,9±2,31ValordeP 0,361 0,837 0,864(GrupoFGA: tratamiento “largo”de14días con esponjadeFGA+ eCGa la retirada,GrupoCIDR: tratamiento“largo”de14díascondispositivodeprogesterona+eCGalaretirada)
Las variables que se analizaron para evaluar la respuesta al tratamiento,fueron directamente el porcentaje de gestación y la prolificidad estimada,determinadasambasporecografíatransabdominaladía40post-montas.
No se observarondiferencias significativas entre el grupo FGA y el grupoCIDR, ni en el porcentaje de gestación (61,7% vs. 51,6%, respectivamente;P=0,120) ni en la prolificidad estimada (1,62 ± 0,55 vs. 1,52 ± 0,50,respectivamente;P=0,317),talycomosereflejaenlatabla9.
Tabla9.Porcentajedeovejasgestantesyprolificidadmediaencadagrupoexperimentaldependiendodeltipodetratamientoadministrado
FGA CIDR
Gestación 61,7%(103/167) 51,6%(49/95)Prolificidad 1,62±0,56 1,52±0,50(GrupoFGA: tratamiento “largo”de14días con esponjadeFGA+ eCGa la retirada,GrupoCIDR: tratamiento“largo”de14díascondispositivodeprogesterona+eCGalaretirada)
La última observación recogida demanera sistemática fue la cantidad dedispositivosintravaginalesconolordesagradableenelmomentodesuretirada,encada grupo. Así pues, solamente un 1% (1/103) de los CIDR presentaron olordesagradable, frente al 76% (130/171) de esponjas con esta característica, yaunque era un signo subjetivo, la intensidad del olor en el caso de los CIDR eramuchomenoraladelosdispositivosFGA.

Resultados
126
4.4 Fase experimental 4. Efecto del tipo de dispositivoprogestativo intravaginal (progesterona natural o CIDRversusprogestágenosintéticooFGA)sobre la funcionalidadhipofisariayováricaenovejas tratadasconunprotocolodesincronizacióndeceloscorto(7días)einfluenciadelaépocareproductiva
En esta fase experimental 4 se abordaron los experimentos 4.1 y 4.2,comparandodos tratamientosde sincronizaciónde celosmedianteprogesteronanatural (dispositivosCIDR) yprogestágenos sintéticos (esponjasFGA)durante7díasyadministracióndePGF2αalaretiradadelosmismos(loquecorrespondeconelprotocoloCorto-PGdelasfasesexperimentalesprevias1y2).
4.4.1 Experimento4.1Fotoperiodocreciente
La condición corporal fue similar en ambos grupos donde la media y ladesviacióntípica fueron3,89±0,20enelcasodelgrupoFGAy3,77±0,33enelgrupo CIDR (P=0,125). La edadmedia de los animales resultó similar en ambosgrupos,siendolamediaenelgrupoFGAde3,45±0,84añosyenelgrupoCIDRde3,50±1,40años(P=0,901).
Influencia del tipo de tratamiento sobre el porcentaje de presentación de celo,momentodeaparicióndelcelouagrupacióndeloscelos
Larespuestaglobalaltratamientoexpresadacomopresentacióndeceloenlosanimalesfuedel100%enambosgruposantesdelas40horastraslaretiradade los dispositivos vaginales, por lo que no hubo diferencias entre gruposexperimentales.
El momento de aparición de celo fue más temprano en el grupo FGA;concretamentea las16h.EnelgrupoCIDR, tardaronunpocomás,alcanzandoelmínimoen24h.Elmáximofue40hparalosdosgrupos(figura34).Enbaseaello,el tiempo (T)mediode salidaenceloenel grupoFGA fuede32,00±6,01h,noexistiendodiferenciassignificativasconelgrupoCIDR,dondelasovejastardaronensalirencelo,demedia,33,76±4,01h(P=0,233).
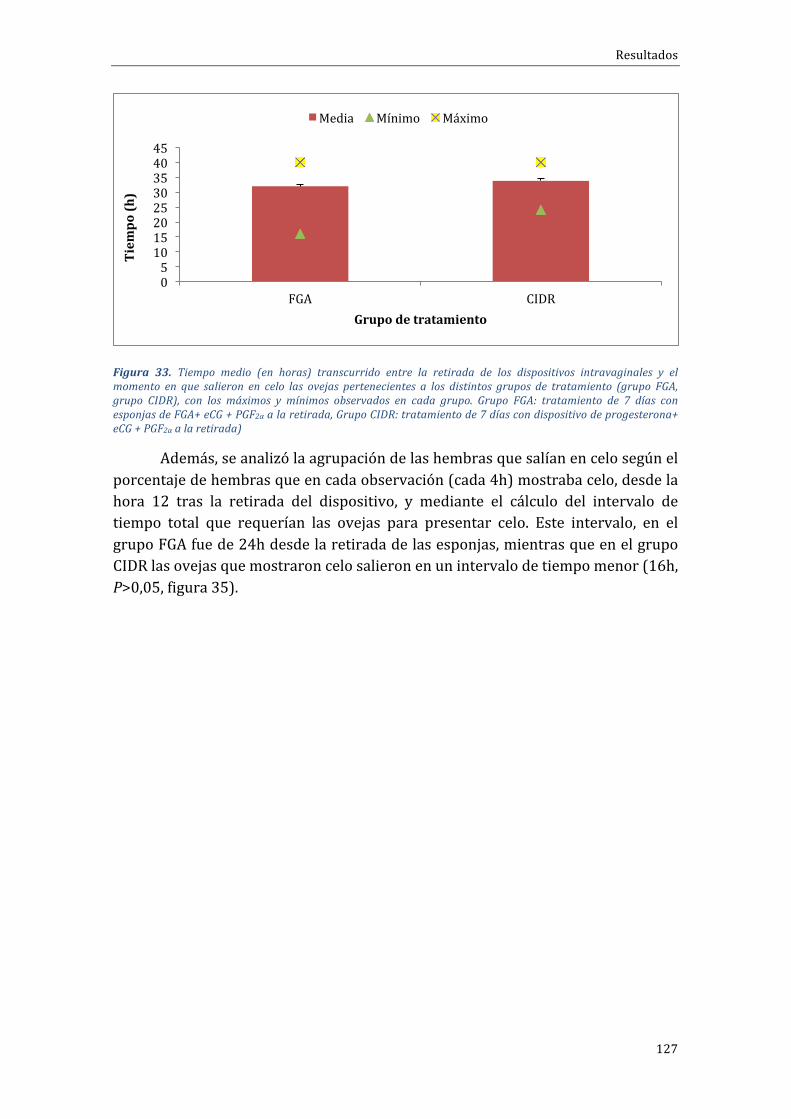
Resultados
127
Figura 33. Tiempo medio (en horas) transcurrido entre la retirada de los dispositivos intravaginales y elmomento en que salieron en celo las ovejas pertenecientes a los distintos grupos de tratamiento (grupo FGA,grupo CIDR), con los máximos y mínimos observados en cada grupo. Grupo FGA: tratamiento de 7 días conesponjasdeFGA+eCG+PGF2αalaretirada,GrupoCIDR:tratamientode7díascondispositivodeprogesterona+eCG+PGF2αalaretirada)
Además,seanalizólaagrupacióndelashembrasquesalíanencelosegúnelporcentajedehembrasqueencadaobservación(cada4h)mostrabacelo,desdelahora 12 tras la retirada del dispositivo, y mediante el cálculo del intervalo detiempo total que requerían las ovejas para presentar celo. Este intervalo, en elgrupoFGAfuede24hdesdelaretiradadelasesponjas,mientrasqueenelgrupoCIDRlasovejasquemostraroncelosalieronenunintervalodetiempomenor(16h,P>0,05,figura35).
051015202530354045
FGA CIDR
Tiem
po(h)
Grupodetratamiento
Media Mínimo Máximo
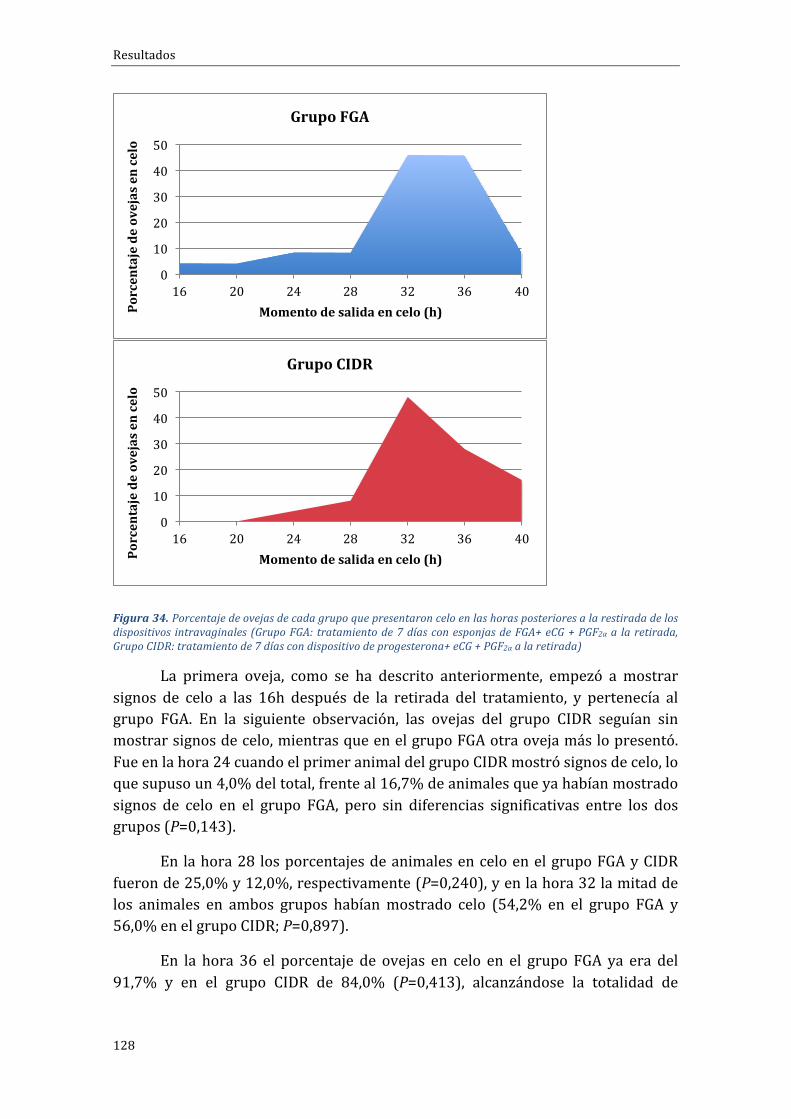
Resultados
128
Figura34.Porcentajedeovejasdecadagrupoquepresentaronceloenlashorasposterioresalarestiradadelosdispositivos intravaginales (GrupoFGA: tratamientode7díasconesponjasdeFGA+eCG+PGF2αa laretirada,GrupoCIDR:tratamientode7díascondispositivodeprogesterona+eCG+PGF2αalaretirada)
La primera oveja, como se ha descrito anteriormente, empezó a mostrarsignos de celo a las 16h después de la retirada del tratamiento, y pertenecía algrupo FGA. En la siguiente observación, las ovejas del grupo CIDR seguían sinmostrarsignosdecelo,mientrasqueenelgrupoFGAotraovejamáslopresentó.Fueenlahora24cuandoelprimeranimaldelgrupoCIDRmostrósignosdecelo,loquesupusoun4,0%deltotal,frenteal16,7%deanimalesqueyahabíanmostradosignos de celo en el grupo FGA, pero sin diferencias significativas entre los dosgrupos(P=0,143).
En lahora28 losporcentajesdeanimalesenceloenelgrupoFGAyCIDRfueronde25,0%y12,0%,respectivamente(P=0,240),yenlahora32lamitaddelos animales en ambos grupos habíanmostrado celo (54,2% en el grupo FGA y56,0%enelgrupoCIDR;P=0,897).
En la hora 36 el porcentaje de ovejas en celo en el grupo FGA ya era del91,7% y en el grupo CIDR de 84,0% (P=0,413), alcanzándose la totalidad de
0
10
20
30
40
50
16 20 24 28 32 36 40
Porcentajedeovejasencelo
Momentodesalidaencelo(h)
GrupoFGA
0
10
20
30
40
50
16 20 24 28 32 36 40
Porcentajedeovejasencelo
Momentodesalidaencelo(h)
GrupoCIDR
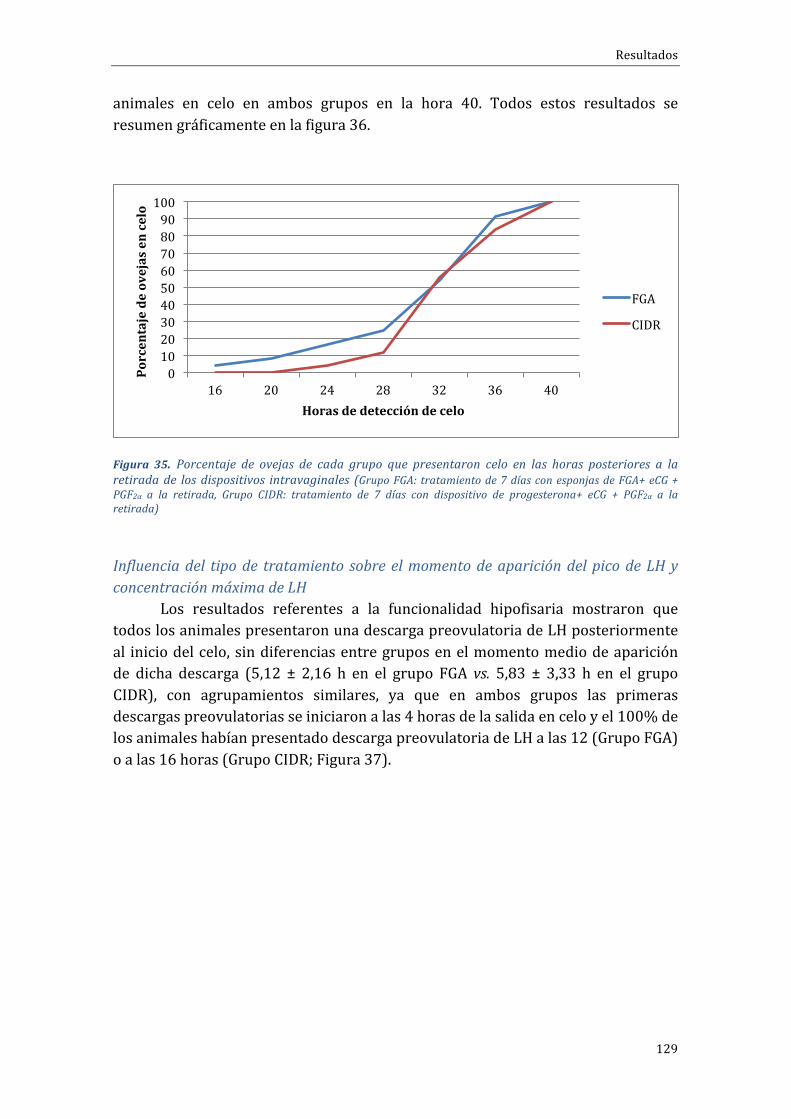
Resultados
129
animales en celo en ambos grupos en la hora 40. Todos estos resultados seresumengráficamenteenlafigura36.
Figura 35. Porcentaje de ovejas de cada grupo que presentaron celo en las horas posteriores a laretiradade losdispositivos intravaginales (GrupoFGA: tratamientode7díasconesponjasdeFGA+eCG+PGF2α a la retirada, Grupo CIDR: tratamiento de 7 días con dispositivo de progesterona+ eCG + PGF2α a laretirada)
Influenciadel tipode tratamiento sobre elmomentodeaparicióndelpicodeLHyconcentraciónmáximadeLH
Los resultados referentes a la funcionalidad hipofisaria mostraron quetodoslosanimalespresentaronunadescargapreovulatoriadeLHposteriormenteal iniciodelcelo,sindiferenciasentregruposenelmomentomediodeapariciónde dicha descarga (5,12 ± 2,16 h en el grupo FGA vs. 5,83 ± 3,33 h en el grupoCIDR), con agrupamientos similares, ya que en ambos grupos las primerasdescargaspreovulatoriasseiniciaronalas4horasdelasalidaenceloyel100%delosanimaleshabíanpresentadodescargapreovulatoriadeLHalas12(GrupoFGA)oalas16horas(GrupoCIDR;Figura37).
0102030405060708090100
16 20 24 28 32 36 40
Porcentajedeovejasencelo
Horasdedeteccióndecelo
FGA
CIDR
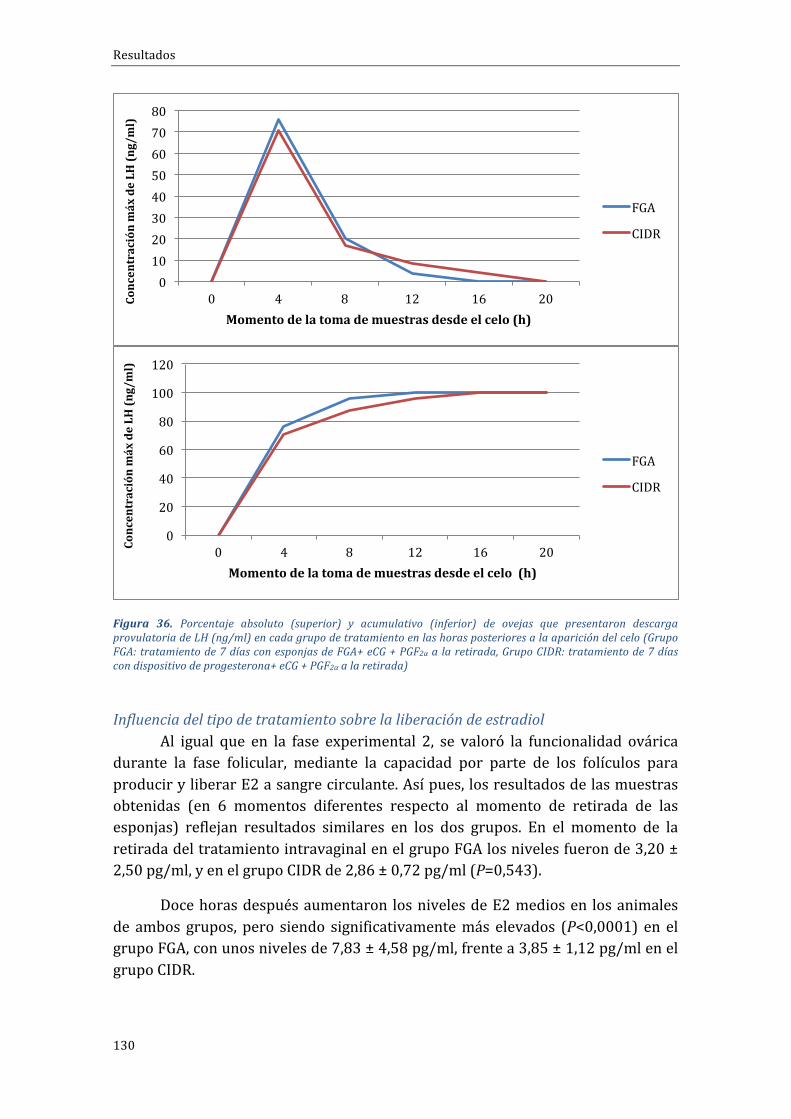
Resultados
130
Figura 36. Porcentaje absoluto (superior) y acumulativo (inferior) de ovejas que presentaron descargaprovulatoriadeLH(ng/ml)encadagrupodetratamientoenlashorasposterioresalaaparicióndelcelo(GrupoFGA:tratamientode7díasconesponjasdeFGA+eCG+PGF2αalaretirada,GrupoCIDR:tratamientode7díascondispositivodeprogesterona+eCG+PGF2αalaretirada)
Influenciadeltipodetratamientosobrelaliberacióndeestradiol
Al igual que en la fase experimental 2, se valoró la funcionalidad ováricadurante la fase folicular, mediante la capacidad por parte de los folículos paraproduciryliberarE2asangrecirculante.Asípues,losresultadosdelasmuestrasobtenidas (en 6 momentos diferentes respecto al momento de retirada de lasesponjas) reflejan resultados similares en los dos grupos. En el momento de laretiradadeltratamientointravaginalenelgrupoFGAlosnivelesfueronde3,20±2,50pg/ml,yenelgrupoCIDRde2,86±0,72pg/ml(P=0,543).
Docehorasdespuésaumentaron losnivelesdeE2mediosen losanimalesde ambos grupos, pero siendo significativamentemás elevados (P<0,0001) en elgrupoFGA,conunosnivelesde7,83±4,58pg/ml,frentea3,85±1,12pg/mlenelgrupoCIDR.
01020304050607080
0 4 8 12 16 20ConcentraciónmáxdeLH(ng/ml)
Momentodelatomademuestrasdesdeelcelo(h)
FGA
CIDR
0
20
40
60
80
100
120
0 4 8 12 16 20ConcentraciónmáxdeLH(ng/ml)
Momentodelatomademuestrasdesdeelcelo(h)
FGA
CIDR
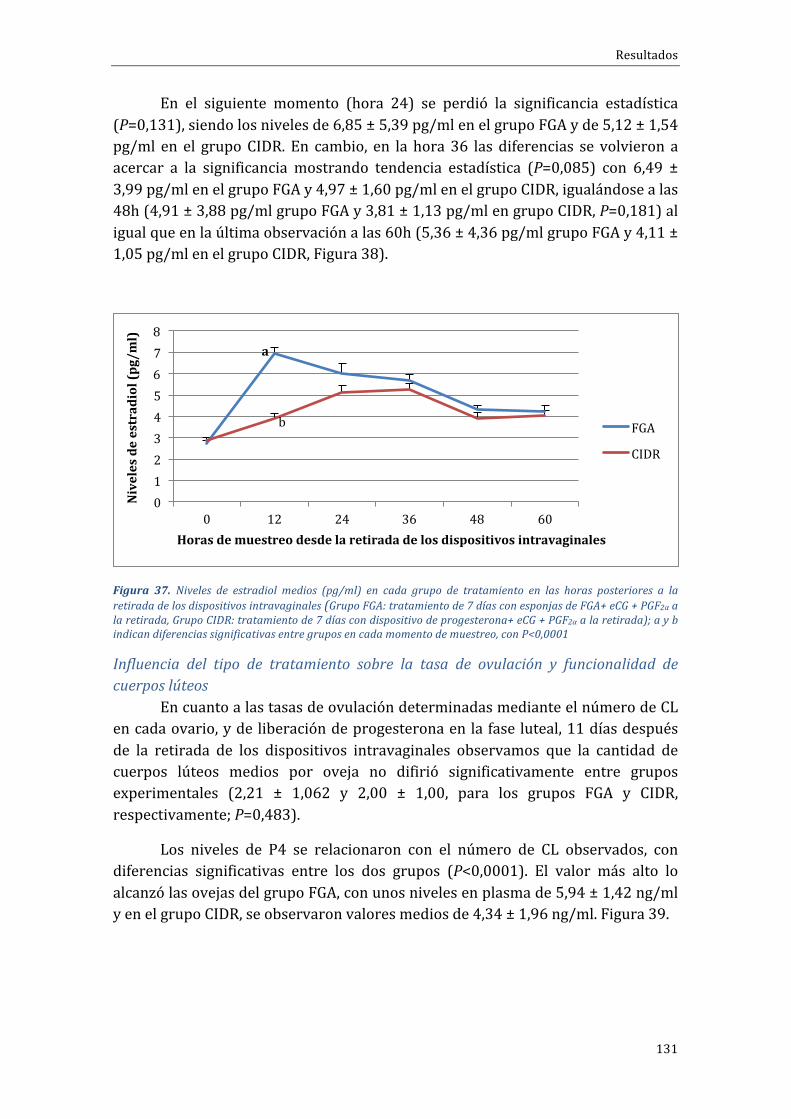
Resultados
131
En el siguiente momento (hora 24) se perdió la significancia estadística(P=0,131),siendolosnivelesde6,85±5,39pg/mlenelgrupoFGAyde5,12±1,54pg/ml en el grupoCIDR. En cambio, en la hora36 las diferencias se volvieron aacercar a la significancia mostrando tendencia estadística (P=0,085) con 6,49 ±3,99pg/mlenelgrupoFGAy4,97±1,60pg/mlenelgrupoCIDR,igualándosealas48h(4,91±3,88pg/mlgrupoFGAy3,81±1,13pg/mlengrupoCIDR,P=0,181)aligualqueenlaúltimaobservaciónalas60h(5,36±4,36pg/mlgrupoFGAy4,11±1,05pg/mlenelgrupoCIDR,Figura38).
Figura 37. Niveles de estradiol medios (pg/ml) en cada grupo de tratamiento en las horas posteriores a laretiradadelosdispositivosintravaginales(GrupoFGA:tratamientode7díasconesponjasdeFGA+eCG+PGF2αalaretirada,GrupoCIDR:tratamientode7díascondispositivodeprogesterona+eCG+PGF2αalaretirada);aybindicandiferenciassignificativasentregruposencadamomentodemuestreo,conP<0,0001
Influencia del tipo de tratamiento sobre la tasa de ovulación y funcionalidad decuerposlúteos
EncuantoalastasasdeovulacióndeterminadasmedianteelnúmerodeCLencadaovario,ydeliberacióndeprogesteronaenlafaseluteal,11díasdespuésde la retirada de los dispositivos intravaginales observamos que la cantidad decuerpos lúteos medios por oveja no difirió significativamente entre gruposexperimentales (2,21 ± 1,062 y 2,00 ± 1,00, para los grupos FGA y CIDR,respectivamente;P=0,483).
Los niveles de P4 se relacionaron con el número de CL observados, condiferencias significativas entre los dos grupos (P<0,0001). El valor más alto loalcanzólasovejasdelgrupoFGA,conunosnivelesenplasmade5,94±1,42ng/mlyenelgrupoCIDR,seobservaronvaloresmediosde4,34±1,96ng/ml.Figura39.
a
b
012345678
0 12 24 36 48 60
Nivelesdeestradiol(pg/m
l)
Horasdemuestreodesdelaretiradadelosdispositivosintravaginales
FGA
CIDR
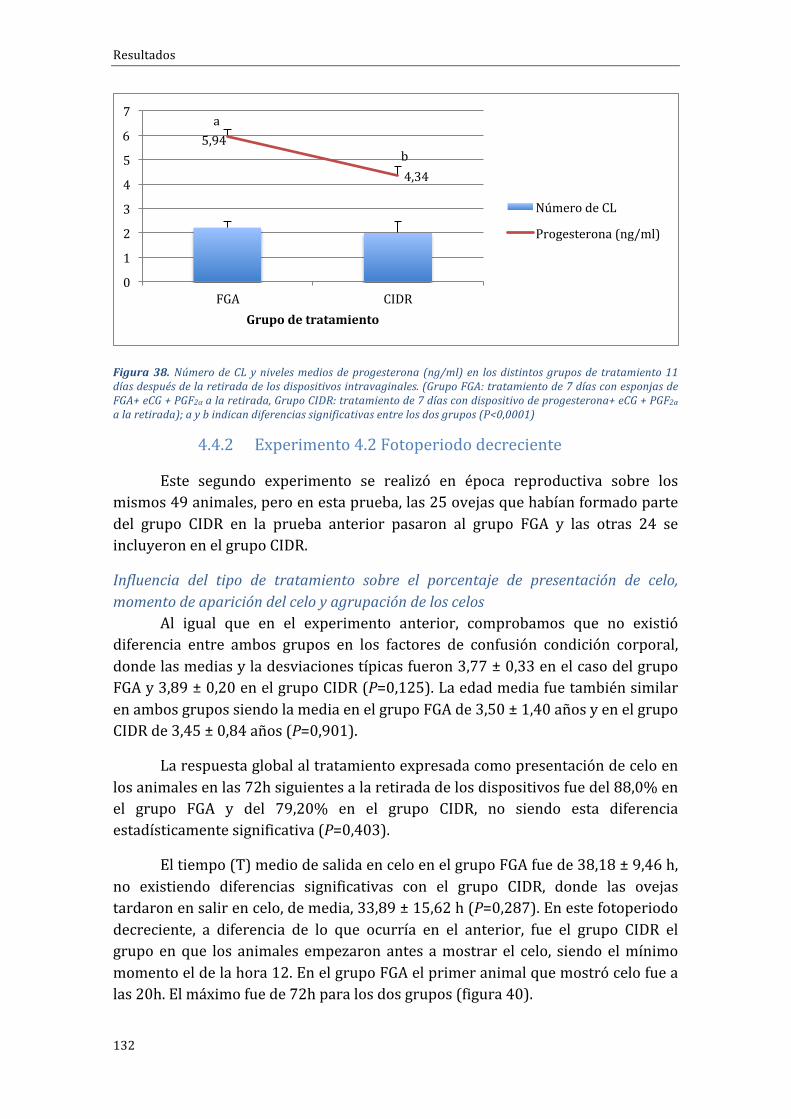
Resultados
132
Figura 38.NúmerodeCLynivelesmediosdeprogesterona(ng/ml)en losdistintosgruposde tratamiento11díasdespuésdelaretiradadelosdispositivosintravaginales.(GrupoFGA:tratamientode7díasconesponjasdeFGA+eCG+PGF2αalaretirada,GrupoCIDR:tratamientode7díascondispositivodeprogesterona+eCG+PGF2αalaretirada);aybindicandiferenciassignificativasentrelosdosgrupos(P<0,0001)
4.4.2 Experimento4.2Fotoperiododecreciente
Este segundo experimento se realizó en época reproductiva sobre losmismos49animales,peroenestaprueba,las25ovejasquehabíanformadopartedel grupo CIDR en la prueba anterior pasaron al grupo FGA y las otras 24 seincluyeronenelgrupoCIDR.
Influencia del tipo de tratamiento sobre el porcentaje de presentación de celo,momentodeaparicióndelceloyagrupacióndeloscelos
Al igual que en el experimento anterior, comprobamos que no existiódiferencia entre ambos grupos en los factores de confusión condición corporal,dondelasmediasyladesviacionestípicasfueron3,77±0,33enelcasodelgrupoFGAy3,89±0,20enelgrupoCIDR(P=0,125).LaedadmediafuetambiénsimilarenambosgrupossiendolamediaenelgrupoFGAde3,50±1,40añosyenelgrupoCIDRde3,45±0,84años(P=0,901).
Larespuestaglobalaltratamientoexpresadacomopresentacióndeceloenlosanimalesenlas72hsiguientesalaretiradadelosdispositivosfuedel88,0%enel grupo FGA y del 79,20% en el grupo CIDR, no siendo esta diferenciaestadísticamentesignificativa(P=0,403).
Eltiempo(T)mediodesalidaenceloenelgrupoFGAfuede38,18±9,46h,no existiendo diferencias significativas con el grupo CIDR, donde las ovejastardaronensalirencelo,demedia,33,89±15,62h(P=0,287).Enestefotoperiododecreciente, a diferencia de lo que ocurría en el anterior, fue el grupo CIDR elgrupoenque los animales empezaronantes amostrar el celo, siendoelmínimomomentoeldelahora12.EnelgrupoFGAelprimeranimalquemostrócelofuealas20h.Elmáximofuede72hparalosdosgrupos(figura40).
a
b5,94
4,34
0
1
2
3
4
5
6
7
FGA CIDRGrupodetratamiento
NúmerodeCL
Progesterona(ng/ml)
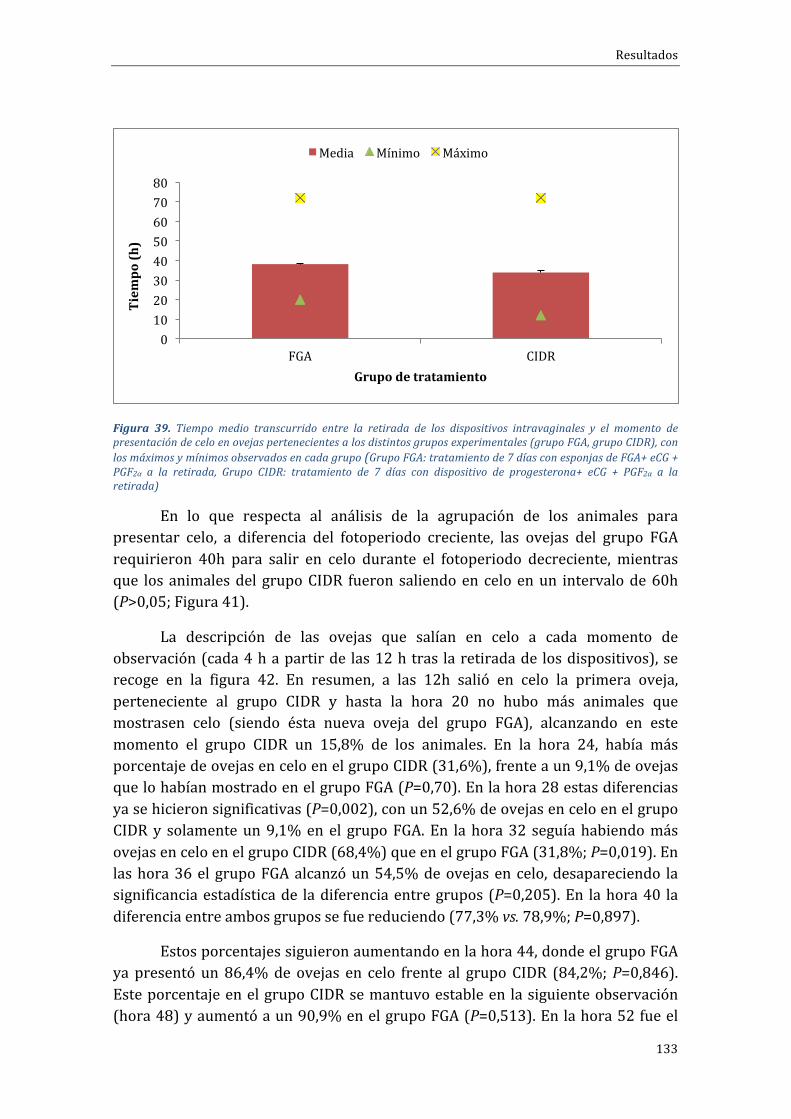
Resultados
133
Figura 39. Tiempo medio transcurrido entre la retirada de los dispositivos intravaginales y el momento depresentacióndeceloenovejaspertenecientesalosdistintosgruposexperimentales(grupoFGA,grupoCIDR),conlosmáximosymínimosobservadosencadagrupo(GrupoFGA:tratamientode7díasconesponjasdeFGA+eCG+PGF2α a la retirada, Grupo CIDR: tratamiento de 7 días con dispositivo de progesterona+ eCG + PGF2α a laretirada)
En lo que respecta al análisis de la agrupación de los animales parapresentar celo, a diferencia del fotoperiodo creciente, las ovejas del grupo FGArequirieron 40h para salir en celo durante el fotoperiodo decreciente, mientrasque losanimalesdel grupoCIDR fueron saliendoen celoenun intervalode60h(P>0,05;Figura41).
La descripción de las ovejas que salían en celo a cada momento deobservación(cada4hapartirde las12htras laretiradade losdispositivos),serecoge en la figura 42. En resumen, a las 12h salió en celo la primera oveja,perteneciente al grupo CIDR y hasta la hora 20 no hubo más animales quemostrasen celo (siendo ésta nueva oveja del grupo FGA), alcanzando en estemomento el grupo CIDR un 15,8% de los animales. En la hora 24, había másporcentajedeovejasenceloenelgrupoCIDR(31,6%),frenteaun9,1%deovejasquelohabíanmostradoenelgrupoFGA(P=0,70).Enlahora28estasdiferenciasyasehicieronsignificativas(P=0,002),conun52,6%deovejasenceloenelgrupoCIDRy solamenteun9,1%enel grupoFGA.En lahora32 seguíahabiendomásovejasenceloenelgrupoCIDR(68,4%)queenelgrupoFGA(31,8%;P=0,019).Enlashora36elgrupoFGAalcanzóun54,5%deovejasencelo,desapareciendo lasignificanciaestadísticade ladiferenciaentregrupos(P=0,205).En lahora40 ladiferenciaentreambosgrupossefuereduciendo(77,3%vs.78,9%;P=0,897).
Estosporcentajessiguieronaumentandoenlahora44,dondeelgrupoFGAya presentó un86,4%de ovejas en celo frente al grupoCIDR (84,2%;P=0,846).EsteporcentajeenelgrupoCIDRsemantuvoestableen lasiguienteobservación(hora48)yaumentóaun90,9%enelgrupoFGA(P=0,513).Enlahora52fueel
01020304050607080
FGA CIDR
Tiem
po(h)
Grupodetratamiento
Media Mínimo Máximo
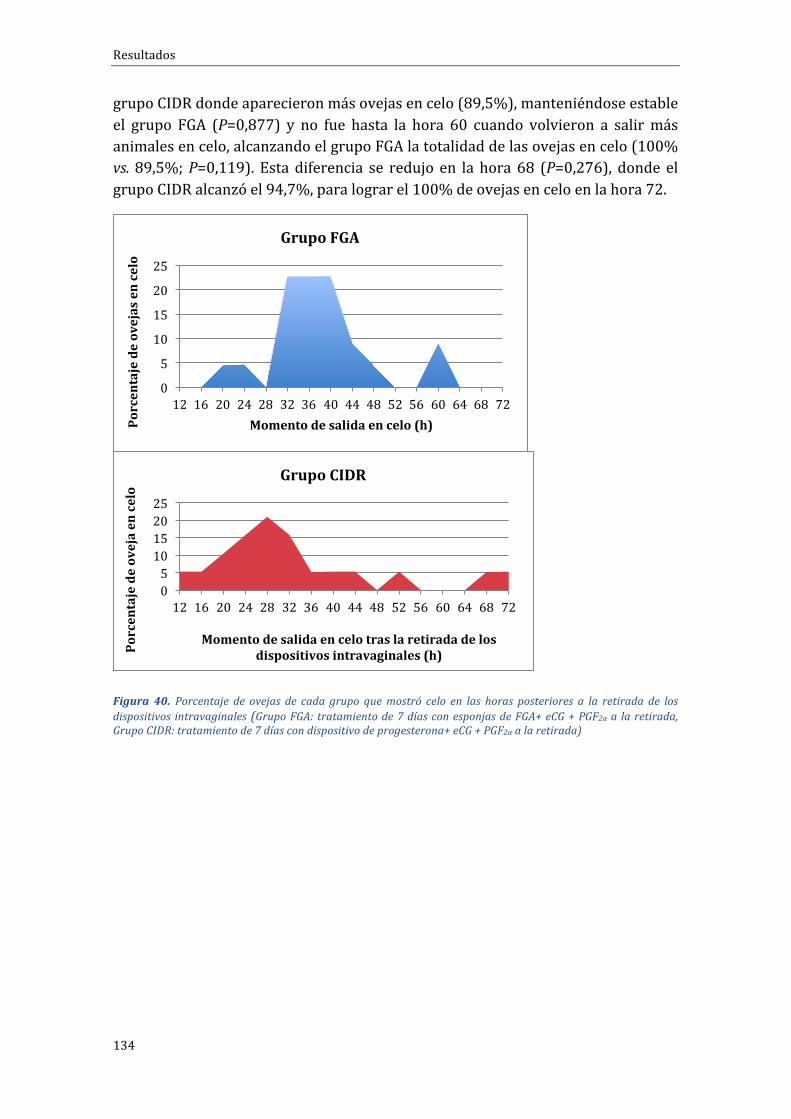
Resultados
134
grupoCIDRdondeaparecieronmásovejasencelo(89,5%),manteniéndoseestableel grupo FGA (P=0,877) y no fue hasta la hora 60 cuando volvieron a salirmásanimalesencelo,alcanzandoelgrupoFGAlatotalidaddelasovejasencelo(100%vs.89,5%;P=0,119). Esta diferencia se redujo en la hora68 (P=0,276), donde elgrupoCIDRalcanzóel94,7%,paralograrel100%deovejasenceloenlahora72.
Figura 40. Porcentaje de ovejas de cada grupo quemostró celo en las horas posteriores a la retirada de losdispositivos intravaginales(GrupoFGA: tratamientode7díasconesponjasdeFGA+eCG+PGF2αa laretirada,GrupoCIDR:tratamientode7díascondispositivodeprogesterona+eCG+PGF2αalaretirada)
0
5
10
15
20
25
12 16 20 24 28 32 36 40 44 48 52 56 60 64 68 72
Porcentajedeovejasencelo
Momentodesalidaencelo(h)
GrupoFGA
0510152025
12 16 20 24 28 32 36 40 44 48 52 56 60 64 68 72
Porcentajedeovejaencelo
Momentodesalidaencelotraslaretiradadelosdispositivosintravaginales(h)
GrupoCIDR
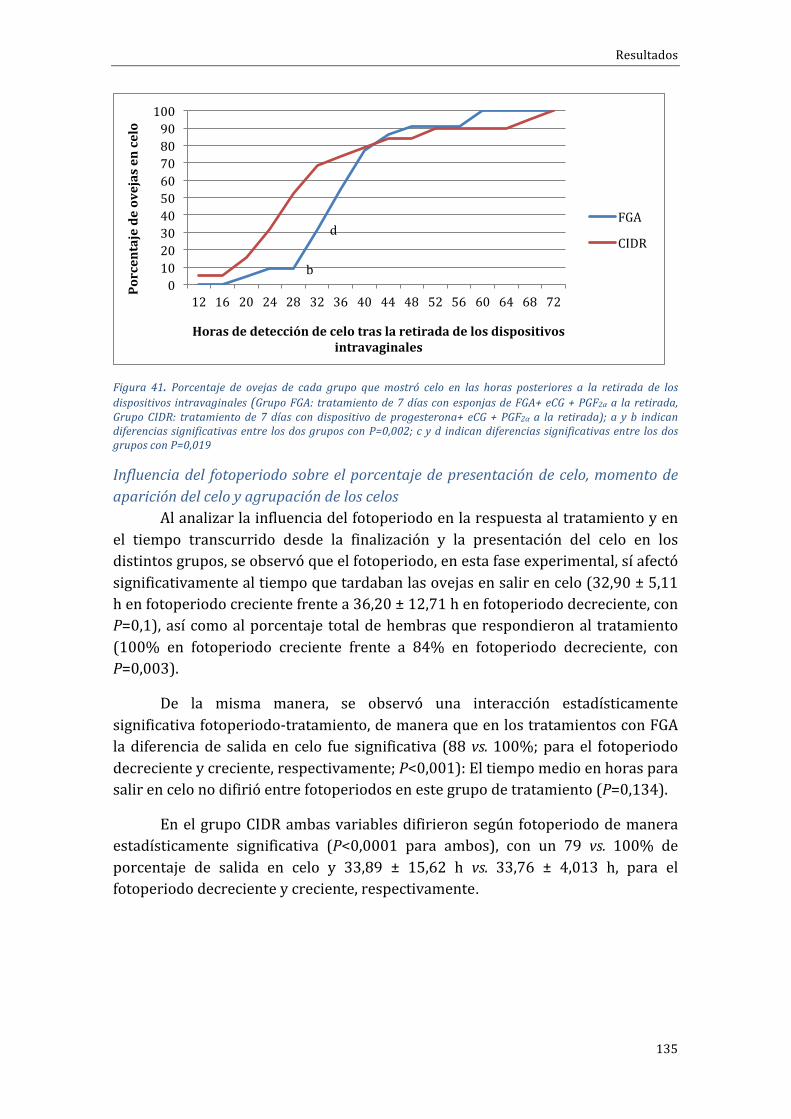
Resultados
135
Figura 41. Porcentaje de ovejas de cada grupo que mostró celo en las horas posteriores a la retirada de losdispositivos intravaginales(GrupoFGA: tratamientode7díasconesponjasdeFGA+eCG+PGF2αa laretirada,GrupoCIDR: tratamientode7días condispositivodeprogesterona+ eCG+PGF2α a la retirada); a y b indicandiferenciassignificativasentre losdosgruposconP=0,002;cyd indicandiferenciassignificativasentre losdosgruposconP=0,019
Influenciadel fotoperiodosobreelporcentajedepresentacióndecelo,momentodeaparicióndelceloyagrupacióndeloscelos
Alanalizarlainfluenciadelfotoperiodoenlarespuestaaltratamientoyenel tiempo transcurrido desde la finalización y la presentación del celo en losdistintosgrupos,seobservóqueelfotoperiodo,enestafaseexperimental,síafectósignificativamentealtiempoquetardabanlasovejasensalirencelo(32,90±5,11henfotoperiodocrecientefrentea36,20±12,71henfotoperiododecreciente,conP=0,1),asícomoalporcentaje totaldehembrasquerespondieronal tratamiento(100% en fotoperiodo creciente frente a 84% en fotoperiodo decreciente, conP=0,003).
De la misma manera, se observó una interacción estadísticamentesignificativafotoperiodo-tratamiento,demaneraqueenlostratamientosconFGAladiferenciade salidaen celo fue significativa (88vs.100%;parael fotoperiododecrecienteycreciente,respectivamente;P<0,001):Eltiempomedioenhorasparasalirencelonodifirióentrefotoperiodosenestegrupodetratamiento(P=0,134).
EnelgrupoCIDRambasvariablesdifirieronsegúnfotoperiododemaneraestadísticamente significativa (P<0,0001 para ambos), con un 79 vs. 100% deporcentaje de salida en celo y 33,89 ± 15,62 h vs. 33,76 ± 4,013 h, para elfotoperiododecrecienteycreciente,respectivamente.
b
d
0102030405060708090100
12 16 20 24 28 32 36 40 44 48 52 56 60 64 68 72
Porcentajedeovejasencelo
Horasdedeteccióndecelotraslaretiradadelosdispositivosintravaginales
FGA
CIDR

DISCUSIÓN


Discusión
139
5 Discusión
5.1 Efecto de la duración del tratamiento desincronización de celos con dispositivos intravaginalesliberadores de progestágenos e influencia de la épocareproductivaydelarazaovina
Enestaprimerafaseexperimentalevaluamoselefectodelareduccióneneltiempodetratamientoconprogestágenos,concretamente,de14a7días,yademásevaluamoslasposiblesdiferenciasdebidasalfotoperiodoyalaraza.Elestudiosehizo en forma de réplicas, una durante el fotoperiodo decreciente (experimento1.1)yotraduranteelfotoperiodocreciente(experimento1.2),analizándosetodoporseparado,yacontinuaciónagrupandolasvariablesqueasílopermitían.
La reducción general del uso de hormonas en producción animal es unatendenciaclaraentodoelmundo,yenespecialenEuropa(Abeciaetal.,2012).Porotro lado, el bienestar animal es cada vez más un objetivo primordial en lasexplotacionesanimales,yanosólodesdeelpuntodevistadelcumplimientodelalegislación2, sino como medio para alcanzar la optimización de la eficienciaproductiva, y por la inquietud y exigencia de los consumidores (Martin et al.,2004).Ligadoaesteámbitodebemoscomentarqueunodelosefectossecundariosmás habituales tras la aplicación de las esponjas vaginales en el ovino, es lainduccióndevaginitis.Enmuchoscasos,alextraerlasesponjasseverificaunaltoporcentaje de éstas con restos hemorrágicos y/o de exudado purulentoindicadoresdeestapatología,quepuedellegaral70%(Swelumetal.,2015;Manesetal.,2016).Además,sabemosquelavaginitisesunprocesoligado,cuantomenos,alaincomodidaddelosanimaleso“disconfort”;(Martin&Kadokawa,2006),yenalgunos casos también se han descrito pérdidas de eficiencia reproductiva en elganadoovinoporestarazón(Manesetal.,2016).Porlotanto,elacortamientodelprotocolo de sincronización en base a dispositivos vaginales, estará ligado abeneficios inmediatos como consecuencia de la reducción de la incidencia devaginitis inducida por el dispositivo per se, aparte de los que pueda haber encuanto a la eficiencia reproductiva, específicamente. Finalmente, es importantecomentarque laduracióndel tratamientodesincronización implicauntrastornoen el manejo de los animales fundamentalmente en sistemas de explotación enextensivo y/o semiextensivo con pastoreo (Delgadillo & Martin, 2015). Lareduccióndelosdíasdetratamientoylaflexibilidadqueestareducciónimplicasetraduceenunamayorfacilidaddemanejo,loque,enproducciónanimal,siemprese traduce en mayor eficiencia productiva. Por lo tanto, éstos son todos
2COMMISSIONREGULATION(EC)No1911/2005of23November2005

Discusión
140
argumentos para considerar especialmente interesante profundizar en elconocimientodelosdenominados“tratamientoscortos”.
Desdeelpuntodevistafisiológico,seríaesperablequeunareducciónenladuración del tratamiento con progestágenos atenuara la caída en los nivelesplasmáticos de progesterona sistémicos, con lo que se evitaría la producción defolículosenvejecidos(Gonzalez-Bulnesetal.,2005),loqueimplicaría,asuvez,unamejoraenlafertilidadtraslostratamientoscortos.Hastaahora,segúnbibliografía,elacortamientoplanteadoenlostratamientosdesincronizacióndecelosbasadosen los dispositivos intravaginales liberadores de progestágenos oscila entratamientosde5, 6, 7 ó9días respectode los tratamientos clásicosde14días(Ungerfeld & Rubianes, 2002; Letelier et al., 2009; Martemucci & D'Alessandro,2011;Ozyurtluetal.,2011).
Laaplicaciónmenosprolongadadelosdispositivosvaginalesdebeincluirlaaplicación de prostaglandinas, ya que un intervalo de aplicación de 6-7 días esmenor que la duración de la vida media de un cuerpo lúteo (Acritopoulou &Haresign, 1980). Los primeros protocolos cortos descritos incluyen la aplicaciónde la prostaglandina simultáneamente a la inserción del dispositivo, aunque,también se puede aplicar a su retirada (Titi et al., 2010; Martemucci &D'Alessandro, 2011), habiendo disparidad de evidencias científicas en labibliografía revisada en cuanto a los resultados obtenidos. Por ello, ésta es lavariacióndetratamientocortoincluidaenelpresentetrabajodondedenominamoscomogrupo “Corto-PG” y “PG-Corto”, según se aplicara laPGF2α al principioo alfinaldelprotocolodesincronización.
Por otro lado, en la especie ovina, sabemos que haymuchos factores queinfluyen de manera determinante en la eficiencia reproductiva de los animales,como la estación en la que se aplique el tratamiento (Martinez-Tinajero et al.,2011), factorquesedebecontrolara lahoradediseñarestudios.El fotoperiodo,componente esencial en la estacionalidad, es un factor determinante en laeficiencia reproductiva de los pequeños rumiantes que ha sido ampliamenteestudiado(Karschetal.,1980;Kennawayetal.,1983;Robinson&Karsch,1987).Así que fue el primer factor a controlar en este trabajo, planteándose losexperimentosen fotoperiodos separados, y sóloenel casodeque los resultadosestadísticos y los criterios metodológicos y fisiológicos lo permitieran, seagregarongruposexperimentalesparaunestudioconjunto.
En el caso de la variable “porcentaje de gestación” y considerando lafertilidadglobaldelosanimalesincluidosenesteestudio,tansóloobservamosunadiferencianumérica(P=0,184)enlasdiferentesestacionesreproductivasyfueenfavor del fotoperiodo decreciente (82,2% vs. 73,7%). Esta “reducidaestacionalidad”yahasidoobservadaporotrosautoresenelcasodelaslatitudesenlasquesehaefectuadoelexperimento(latitud40,70º)oenovejasquehabitan
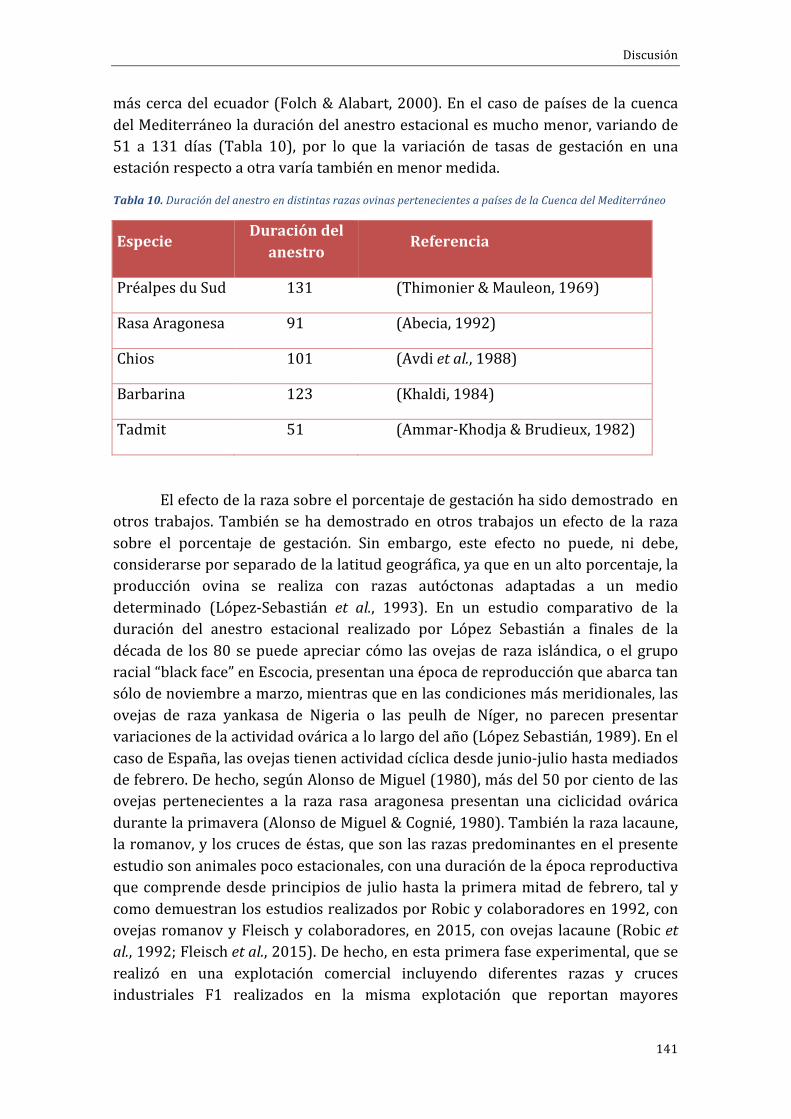
Discusión
141
máscercadelecuador(Folch&Alabart,2000).Enelcasodepaísesde lacuencadelMediterráneoladuracióndelanestroestacionalesmuchomenor,variandode51 a 131 días (Tabla 10), por lo que la variación de tasas de gestación en unaestaciónrespectoaotravaríatambiénenmenormedida.
Tabla10.DuracióndelanestroendistintasrazasovinaspertenecientesapaísesdelaCuencadelMediterráneo
EspecieDuracióndelanestro
Referencia
PréalpesduSud 131 (Thimonier&Mauleon,1969)
RasaAragonesa 91 (Abecia,1992)
Chios 101 (Avdietal.,1988)
Barbarina 123 (Khaldi,1984)
Tadmit 51 (Ammar-Khodja&Brudieux,1982)
Elefectodelarazasobreelporcentajedegestaciónhasidodemostradoenotros trabajos.También sehademostradoenotros trabajosunefectode la razasobre el porcentaje de gestación. Sin embargo, este efecto no puede, ni debe,considerarseporseparadodelalatitudgeográfica,yaqueenunaltoporcentaje,laproducción ovina se realiza con razas autóctonas adaptadas a un mediodeterminado (López-Sebastián et al., 1993). En un estudio comparativo de laduración del anestro estacional realizado por López Sebastián a finales de ladécadade los80sepuedeapreciarcómo lasovejasderaza islándica,oelgruporacial“blackface”enEscocia,presentanunaépocadereproducciónqueabarcatansólodenoviembreamarzo,mientrasqueenlascondicionesmásmeridionales,lasovejas de raza yankasa de Nigeria o las peulh de Níger, no parecen presentarvariacionesdelaactividadováricaalolargodelaño(LópezSebastián,1989).EnelcasodeEspaña,lasovejastienenactividadcíclicadesdejunio-juliohastamediadosdefebrero.Dehecho,segúnAlonsodeMiguel(1980),másdel50porcientodelasovejas pertenecientes a la raza rasa aragonesa presentan una ciclicidad ováricadurantelaprimavera(AlonsodeMiguel&Cognié,1980).Tambiénlarazalacaune,laromanov,yloscrucesdeéstas,quesonlasrazaspredominantesenelpresenteestudiosonanimalespocoestacionales,conunaduracióndelaépocareproductivaquecomprendedesdeprincipiosde juliohasta laprimeramitadde febrero, talycomodemuestranlosestudiosrealizadosporRobicycolaboradoresen1992,conovejasromanovyFleischycolaboradores,en2015,conovejas lacaune(Robicetal.,1992;Fleischetal.,2015).Dehecho,enestaprimerafaseexperimental,queserealizó en una explotación comercial incluyendo diferentes razas y crucesindustriales F1 realizados en la misma explotación que reportan mayores

Discusión
142
resultados productivos y con unas tasas de gestación en general óptimas, tal ycomodescribeOwen(1987)noobservamosdiferenciasracialesenlosporcentajesde gestación (Owen, 1987). En otros trabajos, tampoco se observan diferenciasentre razas como la suffolk, lamerina, la caussenarde, la limousiney algunosdesuscruces(Haresignetal.,1990;Chemineauetal.,1991;Staplesetal.,1992).Enrealidad, otros factores tales como la condición corporal (Forcada et al., 1992;Zhang et al., 2005; Sosa et al., 2010), el nivel higiénico-sanitario (Fragkou et al.,2010), el manejo reproductivo (Anel et al., 2005) o incluso el estrés térmico(Ortavant&Loir,1978;Sawyer,1983)parecensermuchomásdeterminantesenlastasasdegestacióntrasprotocolosdesincronización,quelarazamisma.
Sin embargo, al evaluar la prolificidad, la diferencia entre razas sí fuesignificativa en nuestro trabajo (P=0.050), siendo la raza segureña la demenorprolificidad,encomparaciónconlalacaune,queresultóserlamásprolífica.Estosresultados son semejantes también a los publicados por otros autores que hantrabajadoconestasrazasycrucesraciales(Sierra,1986;Sierra,1989;Perretetal.,1995). Al ser la raza un factor influyente en la prolificidad, el factor se controlódistribuyendodemanerahomogénealasrazasentodoslosgruposexperimentalestal y comoqueda reflejadoenel apartadodematerial ymétodos.Debidoa todoestoes importante,en lamedidade loposible, incluir razasdeterminadasen losestudios científicos, sobre todo cuando éstas son relevantes en sistemasproductivosdedeterminadasregiones(López-Sebastiánetal.,1993),yademás,esmuy importante, asegurar una homogeneidad de razas en los gruposexperimentalesacomparar,talycomohasidoelcasoenelpresenteestudio.
Finalmente,otrofactorqueinfluyedemaneradeterminanteenlaeficienciareproductiva del ovino es la edad, el sistema demanejo ymuy ligada a éste, lacondicióncorporal(Downing&Scaramuzzi,1991;Forcadaetal.,1992;Vinolesetal., 2005;Moeinietal., 2009;Yadietal., 2011).Por esta razón se comprobóquetantoelestadocorporalmedio,comolaedadmediadelosanimalesdentrodelosgrupos experimentales en cada comparativa fuera homogéneo entre gruposexperimentalesencadaréplica.
Una vez controlados todos los posibles factores de confusión pudimoscomprobarelefectoenelporcentajedegestaciónyprolificidaddelacortamientodel protocolo de sincronización con progestágenos intravaginales (tratamientolargo frente a tratamientos cortos), previamente descritos en la literatura(Ungerfeld&Rubianes,1999;Knightsetal.,2001;Letelieretal.,2009).Nuestrosresultados durante el fotoperiodo decreciente (experimento 1.1) no muestrandiferencias significativas en el porcentaje de respuesta al tratamiento desincronización por el hecho de reducir el tiempo de mantenimiento de losdispositivos intravaginalesa7días respectoa14.Elporcentajedegestación fuesimilarparaelgrupoControlodetratamiento“largo”(78,3%)ypara losgrupos

Discusión
143
con tratamiento cortodeprogestágenos independientementedelmomento en elque se administra la prostaglandina grupo PG-Corto: 79,6% y grupo Corto-PG:76,9%P=0.996).
El experimento 1.2 se realizó en fotoperiodo creciente o fase de anestroreproductivo.Lastasasdegestaciónnodifirierontampocoestadísticamenteentreel grupo de tratamiento “largo” (grupo Control; 80%) y los grupos contratamientoscortosPG-Corto:79,2%ygrupoCorto-PG:59,1%P=0,182).Apesarde que las diferencias no alcanzaron significancia estadística, es cierto quenuméricayclínicamentesílaspodemosconsiderarrelevantes,yaquediferenciasde 30 puntos de porcentaje de gestación suponen una diferencia productivasustancial;conseguirunmayornúmerodepartosporovejaesbásicoparamejorarlos resultados económicos, siendo el incremento de los ingresos por venta decorderoscapazdecompensarlosmayorescostes,sobretododealimentación,quesupone la intensificación reproductiva. De hecho, según Pardos (2007) lasexplotacionescapacesdeconseguirmásde1,2partosporhembrapresenteyañotienenmárgenesbrutosporovejasuperioresa25,00€(Pardos&Fantova,2007).Sin embargo, hay otros trabajos que también reportan diferencias numéricas entasas de gestación de 30 puntos tras la aplicación de diferentes protocolos desincronización,sinseréstasestadísticamentesignificativas(60%entratamientoslargos vs.86,7% y 92,3% con PGF2α al inicio y al final respectivamente,P>0,05)(Martemucci&D'Alessandro,2011).Además,ennuestrotrabajoobservamosquela diferencia en el porcentaje de gestación en este protocolo Corto-PG entrefotoperiododecrecienteycrecientesíeraestadísticamentesignificativoindicandounainteracciónsignificativafotoperiodo-tratamientodesincronización,demaneraque durante el fotoperiodo decreciente, el porcentaje de gestación fue del 90%(27/30)mientrasqueenelfotoperiodocrecienteresultóserdel59,1%(13/22),loque sí resultó significativo (P=0,009). Es sorprendente que en el fotoperiodocrecienteoencontraestación,cuandoprecisamentenoseobservancuerposlúteosporelanestroestacionalde losanimales(Hansel&Convey,1983a), fueradondeuntratamientocortoconinclusióndeprostaglandinafuncionarapeor.Unaposibleexplicaciónesquerealmentesísemantuvieraungranporcentajedeanimalesenestro,porlapocaestacionalidadyapreviamentecomentadadelasrazasincluidas(Daza&Blanco,1990),ydadoquelaCCeraóptima,tampocoseveíaagravadoelanestroestacional(Sosaetal.,2010).AsumiendoquesíhubieraCLalprincipiodeltratamiento, está demostrado que altos niveles de P4 durante la fase folicularacarrea efectos negativos sobre el folículo dominante (Gonzalez-Bulnes et al.,2005), lo que podría explicar estas diferencias entre tratamientos. Sin embargo,seríadeesperarestemismoefectotambiénduranteelfotoperiododecreciente,endondepodemoscontardemaneraseguraconCLsenlosovarios,yesunefectoquenohemosobservado,aligualqueenotrostrabajos,quenoobservaronunabajadadefertilidadtraslostratamientoscortosconprostaglandinaalprincipioyalfinal

Discusión
144
(MoradiKoretal.,2012).Porlotanto,esunresultadopendientedeinvestigarmásafondo,atendiendoadetallesfisiológicosydedinámicafolicular.
Los resultados generales obtenidos en nuestro estudio respecto a losporcentajes de gestación, coinciden con lo reportado por Martemucci yD’Alessandro (2011) en la raza italiana altamurana. Estos investigadores noobservaron diferencias significativas al comparar un tratamiento de 14 días conesponjas impregnadas con FGA con un tratamiento corto de esponjas durante 5díasconPGF2αaliniciooalfinaldelmismo(Martemucci&D'Alessandro,2011).Elporcentajedegestaciónobservadono fuediferenteentreelgrupo largo(60%)ycualquiera de los cortos (86,7% y 92,3% con PGF2α al inicio y al finalrespectivamente,P>0,05).Delmismomodo,UngerfeldyRubianes(1999)tampocoobservaron diferencias significativas (P>0,05) en los porcentajes de gestaciónobservadosentresgruposdeovejasalasqueselesadministrótrestratamientosde progestágenos combinados con eCG (Ungerfeld&Rubianes, 1999). A los tresgruposselesinsertaronesponjasintravaginalesimpregnadasenMPAdurante6,9ó 13 días, con administración de eCG a la retirada, obteniendo porcentajes degestaciónde66,7%,75,0%y63,0%,respectivamente.Losresultadosobtenidosenel presente trabajo son igualmente comparables a los observados en cabras porPietroski et al. (2013), que midieron el porcentaje de gestación en tres gruposdistintos de animales tratados con esponjas impregnadas en MAP insertadasintravaginalmente durante 6, 9 ó 12 días, obteniendo exactamente el mismoporcentajedegestaciónenlosgrupostratadosa6y9días(60,0%)ymayorenelgrupo tratado a 12 días (66,7%), pero sin diferencias estadísticamentesignificativas (P>0,05) (Pietroski et al., 2013). Las diferencias específicas en losporcentajesdegestaciónconcretosentre losque reportanestosautoresy lasdenuestrotrabajosepodríanexplicarporelcomponenteracialyladiferenteregióngeográfica.
La prolificidad, otro índice reproductivo esencial en la eficienciareproductivadelovino,evaluadaencadafotoperiodofuetambiénsimilarentrelosgrupos experimentales en ambas estaciones. En el fotoperiodo decreciente en elgrupo de tratamiento largo se obtuvieron 2,51±1,057 corderos nacidos/ovejarespectode los gruposde tratamientos cortos (PG-Corto;2,76±1,052yCorto-PG2,96±1,067 corderos/oveja parida; P=0,929). En el fotoperiodo creciente losresultadosfueron2,41±1,100corderosnacidos/ovejaparaelgrupodetratamientolargo, con respecto a los grupos de tratamientos cortos (PG-Corto; 2,53±1,124 yCorto-PG2,15±1,144corderos/ovejaparida;P=0,904).Enotrostrabajostampocose vio influenciada la prolificidad en función del tipo de tratamientos, ya fuerantratamientos largos o cortos de sincronización, tal y como recogen los autoresMartemucciyD’Alessandro(2011),dondeobtuvierontasasdeprolificidadde1,55corderos/partoconuntratamientode14díasconesponjasimpregnadasenFGA,1,46 corderos/parto y 1,66 corderos/parto con dos tratamiento de 5 días con

Discusión
145
esponjas y PGF2α al final y al inicio del mismo, respectivamente (Martemucci &D'Alessandro, 2011). Estos mismos tipos de tratamiento con esponjasimpregnadasenFGAa14díasrespectodeotroscon5díasconPGF2αalfinalyalinicio los compararon Moradi Kor y colaboradores en 2012 sin observardiferencias significativas entre los porcentajes de gestación (62,5%, 87,5% y93,3%, respectivamente) ni en la prolificidad (1,55, 1,46 y 1,61 corderos/parto,respectivamente) (Moradi Kor et al., 2012). De hecho, como ya comentamospreviamente,larazaesunodelosfactoresmásdeterminantesdelaprolificidadenel ovino (Abdolietal., 2016;McNattyetal., 2016)yno tanto así el protocolodesincronizaciónutilizado.
Sibienesciertoqueunaposiblelimitacióndenuestrotrabajoenestafaseexperimentalalahoradevalorarelporcentajedegestación,pudierasereltamañomuestral utilizado, algo limitado como para detectar estadísticamente unadiferencia en porcentaje de gestación, pero que sí pudiera ser achacable a lostratamientos. Sin embargo, es de reseñar, que numéricamente, el porcentaje deanimalesgestantesfueelmismo,78,3%(47/60),enelcasodeltratamientolargo,respectodeun78,3%(83/106)enelgrupodelostratamientoscortosyque,porotrolado,losporcentajesmediosdegestaciónsonsemejantesalosotrostrabajospreviamentecitados.
Endefinitiva,noobservamosdiferenciasenelporcentajedegestaciónnienla prolificidad según el tratamiento de sincronización hormonal utilizado en unsistema productivo ovino español con razas autóctonas, foráneas y crucesindustriales de las mismas, por lo que podemos concluir que los tratamientoscortos sepuedenconsiderar comoelegiblesenestasexplotacionesganaderasennuestras latitudes, ya que suman las ventajas de la reducción del tratamientohormonal(menostiemporecibiendoprogestágenos)ydeunamejoradebienestaranimal(reduccióndelporcentajedevaginitisasociadasalapresenciadelaesponjaenlavaginadelosanimales),talycomorefierenSwelumycolaboradores(2015),Manesycolaboradores(2016)yMartinyKadokawa(2006).
Una vez comprobado este resultado global de gestación y prolificidad,quisimos profundizar más en la fisiología que sustenta estos resultados, nodescritayaen tantos trabajos.Asípues,pararecabar informaciónmásespecíficaen cuanto al efecto de estos tratamientos cortos sobre la dinámica folicular,funcionalidad hipofisaria y la fase luteal posterior, se planteó la siguiente faseexperimentaldeltrabajo.

Discusión
146
5.2 Efecto de la duración del tratamiento desincronización de celos con dispositivos intravaginalesliberadores de progestágenos sobre la funcionalidadhipofisaria y ovárica en ovejas, en fotoperiododecreciente
Enestafasesecompararonlosmismosmétodosdesincronizacióndecelosqueenlafaseexperimental1peroconunobjetivodiferentequefueelprofundizaren aspectos más fisiológicos referentes a la funcionalidad ovárica, hipofisaria yrespuestaaltratamientodesincronizacióntraslaimplementacióndelosdistintosprotocolos hormonales. Así, nos centramos exclusivamente en la épocareproductiva favorable (fotoperiodo decreciente). Los factores edad y condicióncorporal se controlaron presentando éstos una distribución homogénea entregruposexperimentales.
Losresultadosnosmuestranquesibien,nohubodiferenciassignificativasenlarespuestaaltratamientodesincronizaciónenloqueserefierealporcentajede animales que salieron en celo en las primeras 72h tras la retirada deldispositivo vaginal, observándose los porcentajes del 100% (9/9), vs. 63,6%(7/11) vs.90,0% (10/11) para el grupo Control o largo, frente a los grupos detratamiento cortos (PG-Corto y Corto-PG, respectivamente), realmente, larelevanciaclínicayenproduccióndecasi30puntosentreeltratamientoPG-Cortoylosotrosdos(ControlyCorto-PG)sí loconsideramosimportantes.Dehecho,ladiferencia entre los dos tratamientos cortos mostró una tendenciaestadísticamentesignificativa(P=0,065).Eltiempomediodesalidaencelomedidoenhorassífuesemejanteentrelosgrupos(35,1hvs.43,4hy48htraslaretiradadelosdispositivosvaginales).
En otros estudios publicados, como el de Viñoles y colaboradores (1999)donde se comparaban tratamientos en ovejas de razas tropicales con esponjasimpregnadas en MPA y eCG a la retirada durante 12 y 6 días tampoco seobservarondiferencias significativasa favordeningunode los tratamientos, conporcentajesdegestacióndel67y58%,respectivamente(Vinolesetal.,1999).Sinembargo,losresultadosderespuestaaltratamientofueronengeneralinferioresalos obtenidos en este trabajo, sobre todo, si los comparamos con nuestro grupoControloelgrupoCorto-PGycomparablesa loobservadoennuestrogrupoPG-Corto,aunqueenelpresenteestudionoreflejábamosporcentajedegestaciónsinode salida en celo. Estas diferencias pueden deberse a la raza y las diferentescondicionesdemanejoentrelosdistintossistemas.
Del mismo modo Ungerfeld y Rubianes (2002) establecieron que, entérminosdefertilidad,eltratamientocortode6díasdeesponjasimpregnadasenMPA o FGA es tan efectivo como el tratamiento de 14 días, y sugieren que esta

Discusión
147
flexibilidadpuede resultarventajosaen condicionesde campo, especialmenteensistemasextensivososemiextensivosdeovino(Ungerfeld&Rubianes,2002).Porel contrario, otro estudio realizado en ovejas observó diferencias significativasentre un tratamiento largo de 14 días y uno corto de 7 días con esponjasimpregnadas en FGA. Este experimento se desarrolló al inicio de la estaciónreproductiva,enunalatitudsimilaralanuestra(40°N)yenovejasderazaslocalesturcas,observandoqueun94,6%de lasmismasquehabían sido tratadas conelprotocololargosalieronencelo, frenteaun81,1%delasquehabíanrecibidountratamientocorto (Koyuncu&Altıncekic,2016).Se sabeque la fisiologíaováricapuedevariarentrerazas,perohayotrosmuchosfactoresquesondeterminantes,como la estación en la que se aplique el tratamiento y el tipo de progesteronautilizada,asícomola formadeadministración(Martinez-Tinajeroetal.,2011),elusodeeCGyelmomentodeadministración(Gungoretal.,2007),lapresenciademachosonoenelrebañoenelmomentodelaretiradadelosdispositivos(Moeinietal., 2009), la condición corporal, el sistema demanejo (Yadi etal., 2011) y lalatitud (Martinez-Tinajero et al., 2011). Muchos de estos factores resultan muydiferentesenlosestudiosrespectodeldescritoenestatesisdoctoral,loquelimitalascomparacionesentreunosyotrosestudios.
Enovejasde razaawassihay trabajosque reflejanunapeor respuesta entratamientos cortos frente a los largos. Ustuner y colaboradores obtuvieron en2007diferenciasenrelaciónalostratamientoslargos(12días)ycortos(6días)enhembrastratadasconesponjasimpregnadasenFGA,dondeapreciaronun100%yun 77,1% de respuesta al tratamiento determinada mediante el porcentaje deanimales en celo (P<0,05) (Ustuner et al., 2007). En cambio, Ozyurtlu ycolaboradores no observaron diferencias significativas entre dos grupos dehembras tratadasconesponjas impregnadasenFGAe insertadasdurante7y14días(66,7%y58,3%,respectivamente)enelaño2011,enestamismarazaawassiy sistemas de producción similares (Ozyurtlu et al., 2011). Esta diferencia deresultados entre losdos trabajos en lamisma raza, y en condiciones semejantespodía ser debida a que fue realizada en fotoperiodos distintos, por lo que, en elprimercaso,losautoresaducenqueladiferenciaentregruposfue,probablemente,debida a un retraso en la regresión del CL en algunas ovejas cíclicas, ya que elexperimento se realizó en época reproductiva pero no incluyó PGF2α en eltratamiento.Enelsegundotrabajo,enprincipio,nohabíaovejascíclicas,debidoaque el experimento se realizó en “contraestación”, en el periodo comprendidoentrelasegundamitaddeabrilylaprimerasemanademayo.
Demanera similar,Atamany colaboradores reportan idénticos resultadosen ovejas F1 cruzadas demerino tratadas con esponjas impregnadas en FGA enépoca reproductiva durante 12 y 7 días, determinando como respuesta altratamiento el porcentaje de animales que presentaban celo a la retirada de losdispositivos,conun100%delosanimalesenambosgrupos.Enestetrabajosíseincluía laadministracióndeanálogosdelaprostaglandina,siendoconcretamente

Discusión
148
triaprost trometamina, a la retirada de las esponjas en el tratamiento corto(Atamanetal.,2006).
Otrodatoimportantealahoradevalorarlosprotocolosdesincronizacióneselgradodeagrupacióndelosanimalesquesalenencelo.Lostratamientosquemejoragrupanloscelossuponenunaventajademanejoyeconómicaalunificarenmenostiempo,asuvez,lospartos.Deestamanerasefacilitaelmanejoygarantizauna oferta homogénea de corderos (Parraguez et al., 2000) al permitir laplanificación de inseminación artificial a tiempo fijo (Godfrey et al., 1999) auncuando los resultados de gestación sean semejantes. En nuestro estudio hemospodido comprobar que a las 48h tras la retirada del dispositivo intravaginal lacantidad de ovejas en celo era semejante en los grupos Control y PG-Corto,respectoalgrupoCorto-PG.Sinembargo,enesteexperimento,elrangodetiemposenelquelasovejassalíanencelo(indicadordeagrupación)eramenorenelgrupoCorto-PG(48h)conrespectoalgrupoPG-Corto(62h),mientrasqueelprotocolodesincronización que más agrupaba las salidas en celos fue el largo (16h). OtrosautorescomoMartemucciyD’Alessandro(2011)enuntrabajosimilaralpresente,no encontraron diferencias significativas en el grado de agrupación de celos encabras pertenecientes a la raza autóctona italiana ionica, en un sistema deexplotaciónsemiextensivo,tratadasconFGAdurante11díasyaplicacióndePGF2αel día9 ydos gruposde tratamientos cortos a 5días con aplicacióndePGF2α alprincipiooalfinalconunintervalodetiempodeagrupacióndesalidaencelosenlostresgruposde34,7±6,7h,32,7±7,8hy31,9±8,6h(P<0,05),respectivamente(Martemucci&D'Alessandro,2011).
Por otro lado, en lo referente al grado de agrupación de presentación decelos, Viñoles y colaboradores (2001) observaron, de manera similar a nuestrotrabajo,queelgradoderespuestaaltratamientoenépocareproductivahastalas96horasdesde la retiradade lasesponjas fuesignificativamentemásaltoen lasovejastratadasconprotocoloslargosde12díasrespectoatratamientosde6días(49,0±3,0h y84,8±7,1h;P>0,05, respectivamente) (Vinolesetal., 2001). LosautoresatribuyenestebajogradodesincronizaciónenlostratamientoscortosalapresenciadeunCLfuncionalalfinaldeltratamientoenlamayoríadelasovejas,yaque no incluía el uso de PGF2α, pero este factor no puede aplicarse en nuestrotrabajo.Apesardeello,alas144htraslaretiradadelasesponjas,elnúmerodeovejas que habían mostrado celo era similar en ambos grupos, lo que es muysemejante a lo obtenido en el presente estudio, donde la respuesta final altratamientonofuesignificativaentregrupos.Porotrolado,UngerfeldyRubianes(1999)durantelaépocadeanestro,observaronqueelintervalodetiempodesdela retirada de los dispositivos hasta el inicio del celo fue significativamentemáscortoen lasovejastratadasdurante6díasqueen lasovejastratadasdurante12días, pero en este caso ningún CL estaba interfiriendo con la fase final decrecimientodel folículoovulatoriodebidoa la inclusióndeprostaglandinaen los

Discusión
149
protocolos (Ungerfeld & Rubianes, 1999). Del mismo modo, Sareminejad ycolaboradores en 2014 no encontraron diferencias significativas en época deanestroendosgruposdistintosdeovejas tratadasconMPAdurante14y6días,sin aplicación de PGF2α, y en los que los intervalos de respuesta al tratamientofueronde91,0±0,74y93,33±1,03h,respectivamente(Sareminejadetal.,2014).
Estostrabajos,juntoconnuestrosresultados,demuestranque,enausenciade cuerpo lúteo, ya sea por aplicación de la prostaglandina, ya por realizarlo enhembras en anestro, el acortamiento del tratamiento de sincronización nosupondría diferencias significativas finales en los porcentajes de respuesta alprotocolo de sincronización. No obstante, en nuestro estudio, la capacidad deagrupacióndelcelopermaneciómejorenlostratamientoslargos.Apesardeello,hayquevalorarlasventajasqueaportalagranflexibilidadalahoradesincronizarlos animales con protocolos cortos, situación que se hacemás ventajosa aún ensistemasdeproducciónovinaextensivaysemiextensivaligadaalpastoreo.
Otradelasventajasclarasdelostratamientoscortosrespectodeloslargoseselevitarlainduccióndevaginitisalmantenerlosdispositivosmenostiempoenla vagina de los animales (Manes et al., 2015). Finalmente, debemos considerarcomobeneficiosalareduccióndelaadministracióndehormonas,loquesealineamásconlasproduccionesconsideradas“limpias”,enrespuestaalasdemandasdelasociedad,talycomoproponenensustrabajosMartinycolaboradores(2004)oMartinyKadokawa(2006)(Martinetal.,2004;Martin&Kadokawa,2006).
Aunque la aplicaciónde los tratamientos cortos de esponjas impregnadasenprogestágenosofrece las ventajas citadas, tienenel inconvenientedequehayqueadministrarPGF2αparalisarposiblesCLpresentesenlashembrasconobjetodeevitarunadisminucióndelporcentajedegestación (Vinolesetal.,1999).A lahoradevalorarestostratamientoscortoshayquetenerencuenta,queelpreciodelosmismosseelevadebidoaestainclusiónobligadadelaprostaglandina,yaseaalprincipio (Letelieretal.,2009),yaseaal finaldeprotocolo(Vinolesetal.,1999).Tambiénhayquevalorarelaumentodel trabajoasociadoalaplicar lamisma,deformaqueenunmarcoproductivohayquehacerunbalanceeconómicodeesteefectoyvalorarloteniendoencuentalasventajaspreviamentecomentadas.
En cuanto a la comparativa entre los distintos tratamientos desincronización cortos, en el presente trabajo realizado en ovejas autóctonasespañolas y F1 de cruces industriales, aunque las diferencias fueron sólonuméricas, lasovejasquemejor respondierona la sincronización fueron lasquerecibieron la PGF2α a la retirada de los dispositivos, frente a las que se lesadministróenelmomentodesucolocación.LapresenciadeunCLactivoalteralafuncionalidadylamaduraciónfinaldelosfolículospreovulatoriosylaluteogénesisnormal, debido a que las concentraciones de P4 elevadas durante la fase lutealdisminuyen la secreción de LH, hormona esencial para el crecimiento y

Discusión
150
maduraciónfinalde los folículospreovulatorios(Campbelletal.,1995;Gonzalez-Bulnesetal.,2004).EsporelloquealgunosautoresrecomiendanlaaplicacióndeunadosisdePGF2αosusanálogosenelmomentodelainsercióndelaesponja.Esbien conocida la capacidad de sincronización del estro posterior a laadministración de PGF2α entre los días 5 y 14 del ciclo estral ovino, que induceluteolisis,seguidadeestroyovulación(Acritopoulouetal.,1977;Acritopoulou&Haresign, 1980). Para asegurar una adecuada respuesta a los tratamientos desincronizacióncortosbasadosenprogestágenosserecomienda la inclusiónde lamismaalcomienzodedichoprotocolo(Letelieretal.,2009).Porelcontrario,enotrostrabajosserecomiendalaaplicacióndePGF2αenelmomentodelaretiradade la esponja, durante el fotoperiodo decreciente (Vinoles et al., 1999). VariosautorestambiéncorroboranlanecesidaddeaplicarlaPGF2αenelmomentodelaretirada de la esponja (Greyling et al., 1979). Estos últimos autores obtuvieronporcentajesdesincronizacióndel100%entratamientoscortosde8díasconPGF2αal final del mismo. Fitzgerald y colaboradores (1985) obtuvieron resultadossimilaresdesincronizacióncontratamientosde7díasconPGF2αtambiénalfinal(Fitzgerald et al., 1985). Estos resultados concuerdan con los obtenidos en elpresente estudio en el que, tras un protocolo semejante (Corto-PG), en ovejasautóctonas españolas y F1 de cruces industriales, se observó que el mayorporcentaje de presentación de celo durante la época reproductiva (fotoperiododecreciente) es en el grupo que recibe la PGF2α a la retirada de los dispositivos(90,9% vs. 63.6%). Por lo tanto, parece que dentro de los tratamientos desincronizacióncortos,aplicadosduranteel fotoperiododecreciente, connuestrasrazasyennuestraslatitudes,elprotocolodesincronización“Corto-PG”podríasermásapropiadoqueeltratamiento“PG-Corto”,eigualmenteadecuadorespectodelostratamientoslargos.
Enrelaciónalasdeterminacioneshormonales,lospicosdeliberacióndeLHpreovulatoriosaparecieronenmomentosmuysimilares,sinembargo,estospicosseobservaronmásagrupadosenelgrupoCorto-PGqueenelgrupoPG-Corto.Lospicos de LH más agrupados son muy buscados en los protocolos ligados ainseminaciónartificial, yaquepermitenajustarmásadecuadamenteelmomentodelamismaymejorarlafertilidad(Gonzalez-Bulnesetal.,2004).Deestaforma,alahoradeelegirentrelosdosprotocoloscortos,parecequeeltratamientoCorto-PG tendría ventajas sobre el PG-Corto. Además, este grupo Corto-PG presentanivelesdeLHmuysimilaresalosdelgrupodetratamientolargo(mientrasqueeltratamientoPG-Cortopresentabaniveles inferioresalgrupocontrol),ysonestostratamientos con inserción de dispositivos vaginales durante 14 días yaplenamente demostrados como de una fertilidad adecuada (Vinoles et al., 2001;Evans et al., 2004), por lo que ratifica que este tratamiento, que hemosdenominadoCorto-PG,puedesertanadecuadocomoellargo.

Discusión
151
Encuantoalgradodeagrupacióndelosanimalesquesalenencelodentrode los tratamientos cortos comparados en nuestro trabajo, pudimos comprobarque el rango de tiempo en el que las ovejas salían en celo (indicador deagrupación)eramenoreneltratamientoCorto-PG(48h)respectodeltratamientoPG-Corto(62h).Demanera,queentrelosdostratamientoscortosratificamosotraventajaadicionalenelprotocoloCorto-PGsobreelPG-Corto.
Al evaluar ladinámica folicular, observamosprincipalmente,queel grupode tratamiento Corto-PG daba lugar a un mayor reclutamiento y desarrollofoliculardandocomoresultadomásfolículosmedianosalfinaldelafasefolicular,en comparación con el otro grupo de tratamiento corto (PG-Corto). Esto es otraevidenciaquenoshacepreferireltratamientocortoconlaprostaglandinaalfinal,respectodeltratamientoPG-Corto,yaqueesconocidoquelosfolículosovulatoriosmás jóvenes se correlacionan con una mejor fertilidad (Evans, 2003). Por otrolado,estadiferenciadedinámicafoliculartambiénquedóreflejadaenunamayorliberacióndeE2porpartedeestegrupoCorto-PGrespectodelresto,relaciónyaencontrada enotros trabajospublicados (Campbelletal., 1995).Dehecho, estosmismos autores ratifican que la determinación de niveles de estradiol es unamanera indirecta de estudiar la dinámica folicular (Letelier et al., 2009). En elpresentetrabajo,enloreferentealaliberacióndeestradiol,pudimosobservarunadiferenteproducciónestrogénicaporpartedelosfolículossegúnelmomentodelcicloyelgrupodetratamiento:asíenelprimerdía(momento12h)lasovejasdelgrupo control mostraron una mayor producción de esta hormona ovárica. Sinembargo, a partir ya de las 24h tras la retirada de los dispositivos, fue en estegrupo control donde observamos una producción estrogénica inferiorestadísticamente significativa en losmomentos24h y 60h ymenor, aunque sólonuméricamente, enel restodemomentos.Enel casodelmomento final (60h)elgrupoCorto-PGeselgrupoquepresenta lamayorproduccióndeE2.ResultadossobreliberacióndeE2losobservamosenlostrabajosdeLetelierycolaboradores(2011),dondelafuncionalidaddelosfolículospreovulatoriosseconfirmóporunincrementoenlasconcentracionesdeestradiolenplasmadurantelafasefolicularendosgruposde tratamientodistintos,unodeellos conFGAa14díasyelotrousandotresinyeccionesdePGF2αseparadasporunperiodode10días(Letelieretal.,2011).LasconcentracionesdeE2fueronsimilaresalasobtenidaspreviamentepor otros autores en ovejas de raza manchega (Gonzalez-Bulnes et al., 2005;Gonzalez-Anover et al., 2007) y a las del presente estudio, corroborando la“normalidad”funcionaldenuestrosanimales.Sinembargo,nohemosencontradoestudiosen laespecieovinaquedemuestrennivelesdeestradioldurante la fasefolicular de manera comparada entre diferentes tratamientos de sincronizaciónbasados en la aplicación de progestágenos. Sí hay estudios comparandotratamientosdesincronizaciónbasadosenprostaglandinas(Fierroetal.,2016)ocomparando tratamientosde sincronización respectodel celonatural (Gonzalez-Bulnesetal.,2005).Engeneral,podemosargumentarqueunamayorliberaciónde

Discusión
152
E2 se corresponde con una mejor “salud folicular”, tal y como demuestranGonzalez-Bulnes y colaboradores, (2005) viendo que existe una correlaciónsignificativaentreelniveldeliberacióndeE2ylaposteriorcapacidadluteotróficadelosCL,yademás,estosnivelesdeP4estánpositivamentecorrelacionadosconlacantidadycalidaddeembrionesrecuperados.Asimismo,losnivelesdeE2durantela fase folicular se correlacionan negativamente con el número de oocitos nofertilizados (Gonzalez-Bulnesetal., 2005). Porotro lado, el nivel deE2 liberado,dentro de folículos no quísticos ni persistentes, está correlacionado directa ypositivamenteconlastasasdegestación,yaqueestosnivelessonesencialesparael desarrollo embrionario temprano (Murray et al., 1992; Watson et al., 1994;Nancarrow&Hill, 1995; Fierro etal., 2016). De hecho, Swelum y colaboradores(2015) observaron mejores tasas de fertilidad y prolificidad en el grupoexperimental que liberaba mayores niveles de E2 y por lo tanto de P4 en unestudiocomparativodeprotocolosdesincronizaciónentreesponjasydispositivosCIDR(Swelumetal.,2015).Endefinitiva,parecelógicoesperarunosresultadosdegestaciónbuenos,oinclusomejorestraslostratamientoscortos,ydentrodeellos,traseltratamientoCorto-PG,basadosenestosmayoresnivelesdeliberacióndeE2durantelafasefolicular.Sibien,unavezalcanzadounniveldeestradioladecuado,y que induzca un pico, lo que parece jugar también un papel relevante es elintervaloentreelpicodeE2yeldeLH,observándosequeunretrasoenlarelaciónpicodeE2-picodeLHsepuederelacionarconmenorfertilidad(Veiga-Lopezetal.,2014).EnnuestrotrabajonohemosestudiadoelintervalopicodeE2-picodeLHentregrupos,loquepodríaserinteresanteparafuturosestudios.
Al estudiar la fase luteal y el porcentaje de ovulación no se observarondiferencias estadísticas en cuanto al número medio de CL/oveja, aunque ladistribución del número de cuerpos lúteos sí que varió significativamente entregrupos,conmásanimalescon3CLentrelosgruposcortos(57,1%enCorto-PGy43,5%enPG-Corto)respectodelgrupoControl(11,1%);acambio,seobservaronmenosanimalescon4CLenlosgruposdeprotocoloscortos(0versus22,2%).ElnúmerodeCLreflejaelnúmerodeovulacionesy,enproducciónanimal,lospartosdobles y triples se consideran productivos, especialmente en la producción deaptitudcárnica,apesardelaelevaciónenlastasasdedistociapormalposición.Sinembargo, la aparicióndepartos cuádruples enuna explotación suponeun graveproblema de manejo, ya que la madre no puede mantenerlos y algunos de loscorderosdebenahijarseconotrasovejasocriarseconlactanciaartificial.Porotrolado está demostrada la relación entre el tamañode camada y la dificultadparamamarporpartede los corderosnacidosdepartosmúltiples (Dwyer&Bunger,2012),asícomomayordesgastemetabólicoenlasovejasgestantesdevariosfetos,respectodelasdegestacionessimples(Gonzalez-Garciaetal.,2015)ylaelevacióndelamortalidaddeloscorderossegúnaumentaeltamañodelacamada(Holmoy&Waage,2015).

Discusión
153
Finalmente,laproducciónmediadeP4tampocodifirióentregrupos,loqueratificaría la idoneidad adicional de los tratamientos cortos. En otros trabajosrealizadosdurantelaépocareproductiva,(Greylingetal.,1979),comparandotrestratamientosconesponjasdeMPAdurante8díasconadministracióndetresdosisdistintas de cloprostenol a la retirada, y midiendo concentraciones de P4 enplasma,noobservarondiferenciassignificativasentrelostresgrupos.Ozyurtluyycolaboradores. (2011) tampoco encontraron diferencias significativas en losnivelesdeP4entregruposdeanimalestratadosconesponjasdeFGAa14y7díasdurantelas2y3semanasposterioresalaretiradadelosdispositivos(Ozyurtluetal.,2011).Sinembargo,eltratamientocortomostrónivelesmásaltosdeP4enelmomentoderetirareldispositivo,debido,posiblemente,a lapresenciadeunCL,ya que el tratamiento de sincronización no incluía la administración de PGF2α, adiferenciadelosprotocolosaplicadosennuestrostrabajos.
Porotrolado,AmeryHazzaa(2009),enovejastratadasconFGAyeCGa6y12díasobservaronnivelesbasalesdeP4enlosdías-12,-4(previosalestro)y0(estro) demostrando anestro estacional al comienzo de los protocolos desincronización.Despuésdeldía0lasconcentracionespermanecieronbajas,hastael día 5, consiguiendo que todas las ovejas ovularan durante este periodo. LasconcentracionesdeP4postovulaciónseincrementarongradualmentedesdeeldía5 hasta el día 20 y alcanzaron sumáximo a partir del día 11, pero sin observardiferenciassignificativasentrelosdosgrupos(Amer&Hazzaa,2009),demanerasemejantealoencontradoenelpresentetrabajo.Sinembargo,ennuestroestudio,es importante recalcar, que numéricamente sí observamos una menorconcentración de P4 en el grupo PG-Corto (1,02 ± 1,06 ng/ml) respecto de losniveles observados en el grupo Corto-PG (2,11 ± 2,82 ng/ml) o grupo Control otratamientolargo(2,01±1,82ng/ml).ProbablementelapresenciadeCLdurantela sincronización en las ovejas del grupo Corto-PG dio lugar a niveles deprogesteronaaltos,loqueserelacionaconunamenorsecrecióndeLH(Gonzalez-Bulnesetal.,2005).Estasituaciónhacequelosfolículosdominantespresentesenese momento entren en regresión y desaparezcan, apareciendo a continuaciónfolículosnuevosencrecimiento(losdetamañomediano,observadosenelestudiode la dinámica folicular). Estos folículosmás jóvenes serían los responsables deuna mayor liberación de E2. En general, los resultados de liberación de P4, secorrespondenconfolículosconmenoractividadprevia,reflejadaconlaliberaciónde E2, relación también corroborada previamente en la bibliografía (Gonzalez-Bulnesetal.,2005;Fierroetal.,2016)ademásdeconlosresultadosdeliberacióndeE2delpresentetrabajo.Finalmente,esnotableremarcarquenivelesmenoresdeP4sevencorrelacionadosenotrostrabajosconunamenorfertilidad(Swelumetal.,2015).
De manera que en base a estos resultados y a pesar de la ausencia dediferencias significativas entre tratamientos, tanto en el número medio deCL/oveja como en los niveles medios de P4, podemos esperar semejantes

Discusión
154
resultadosdeeficienciareproductivayproductivaenlasovejasprotocolizadascontratamiento largo o con el tratamiento Corto-PG, que podrían ser usadosindistintamente en granjas, elevando la flexibilidad demanejo y protocolizaciónreproductiva.Porotrolado,recordarqueunamenorduracióndelostratamientoscon dispositivo intravaginal se relaciona, con una menor aparición de vaginitis,hemorragiasyadherencias(Manesetal.,2010),loquesuponeunamayorlimpiezade lavagina, facilitando lavisualizacióndelcérvix, locualesmuyrelevantesi sedecide aplicar inseminación transcervical (Windsor, 1995). Esto seríanargumentos para preferir la utilización del tratamiento Corto-PG, respecto dellargos,unavezdemostradaunaeficaciareproductivasemejante.

Discusión
155
5.3 Efectodeltipodedispositivoprogestativointravaginal(progesterona natural o CIDR versus progestágenosintético o FGA) sobre el porcentaje de gestación enovejas tratadas con un protocolo de sincronización deceloslargo(14días)enfotoperiododecreciente
Enestafaseexperimental3sevalorarondosmétodosdesincronizacióndecelo“largos”usandodosdispositivosdiferentesconprogestativos(progestágenosen esponjas impregnadas en FGA y progesterona en dispositivos tipo CIDR)basadosenelprotocoloclásicode14días,conadministracióndeeCGalaretiradadelosmismos.Elobjetivofuecompararelporcentajedegestaciónylaprolificidad,estimadaporecografíatransabdominaladía40post-monta,durantefotoperiododecreciente.Tambiénsevaloraronotros factorescomoelporcentajedepérdidasde dispositivos y el porcentaje de dispositivos con olor desagradable en elmomento de la retirada. Los factores edad, número de lactaciónmedio /oveja ycondicióncorporalsecontrolaronpresentandoéstosunadistribuciónhomogéneaentregruposexperimentales.
Losporcentajesdegestaciónobtenidosfuerondel61,7%enelgrupoFGAydel 51,6% en el grupo CIDR (P=0,120). La prolificidad estimada también fueligeramentemayor en el grupo FGA (1,62 ± 0,55) que en el grupo CIDR (1,52 ±0,50),conP=0,317.
Puesto que el FGA es un agente progestágeno más potente que laprogesterona natural y, por lo tanto, puede ser usado a dosis más bajas, losresultadosobtenidos,muestranque losdispositivosCIDR,quecontienen300mgde progesterona natural son tan efectivos como método de sincronización delestroenovejascomo20mgdeFGAimpregnadoenesponjas intravaginales,bajolascondicionesdemanejodescritasenelpresentetrabajo,quesonlastípicasdeexplotaciónovinaintensiva(Reynolds,1996).
Nuestros resultadosmuestranquenohubodiferencias significativasenelporcentajedegestacióntalycomoyaobtuvieronafinalesdelosaños80RhodesyNathanielsz, comparando los mismos dispositivos basados en progesteronanatural,durante14días,vs. dispositivosdeMPA,en lugardeFGA.Estosautoresobtuvieronademás,unosporcentajesdegestaciónmuysemejantesalosnuestros,del 56,6%en el grupo tratado con esponjas deMPA y de un 57,7%en el grupotratadoconCIDR.
Porcentajes de gestación un poco más elevados que en el trabajo actual,pero similaresentreellos (P>0,05)apesardeunadiferencianumérica, comoennuestro caso de 10 puntos porcentuales, fueron los obtenidos por Hamra ycolaboradores (1989) al comparar el tratamiento de CIDR impregnados enprogesterona (71%) con el de esponjas impregnadas en cronolona (85%),

Discusión
156
seguidostodosellosdelaadministracióndeeCGalaretiradadelosdispositivos,demanerasemejantealpresentetrabajo(Hamraetal.,1989).AunquelafertilidadmediatraslaaplicacióndelCIDRennuestrotrabajonoesmuyelevada,síestáenconsonancia con otros trabajos publicados previamente, como el de Moeini ycolaboradores (2013) (Moeini et al., 2013). Estos autores compararon distintostratamientos con esponjas y CIDR en ovejas de raza lori durante 13 días enestaciónnoreproductiva,perocondistintasdosisdeeCGenlosdistintosgrupos.Apesardequelasgonadotropinasmejoranlatasadeovulación(Gungoretal.,2007)observaronquelosanimalesa losquese lesadministróunadosismenordeeCG(350UI)presentaronporcentajesdegestaciónmayoresqueaaquellosalosqueseles administraron 500UI tras la retirada de los dispositivos. Los resultados con350UIfueronde54,5%enelgrupotratadoconesponjasdeFGA,frentea45,4%en el grupo tratado con CIDR. En cambio, en los tratamientos que incluían unadosis de 500 UI de gonadotropina se observaron porcentajes de gestación de38,8% en el grupo tratado con esponjas frente al 32,8% observado en el grupotratadoconCIDR.
Resultados similares a los nuestros, obtuvieron Zonturlu y colaboradores(2008)conovejasderazaawassi,unaduracióndelostratamientosde12díasylaadministraciónde300UIdeeCG,conunosporcentajesdegestaciónde52,63%enelgrupotratadoconesponjasyde60,0%enelgrupotratadoconCIDR(Zonturluet al., 2008). El porcentaje de gestación en el grupo tratado con CIDR escomparable igualmente al 63% obtenido por Ungerfeld y Rubianes (2002)(Ungerfeld & Rubianes, 2002). En cambio, los resultados obtenidos por estosmismos autores en los grupos tratados con FGA fueron más altos. En otrosestudiosrevisados, losporcentajesdegestaciónobtenidosentratamientosenlosquesehausadoCIDRhanvariadoentreel44,40%yel55,60%(Fukuietal.,1999;Godfreyetal., 1999;Danieletal., 2001), loquevuelve a concordar connuestrascifrasdeporcentajedegestación.
EstosdosdispositivostambiénhansidocomparadoscondistintasdosisdeeCG(350UI,500UIy600UI)porMoradiKorycolaboradores(2012)ylosgrupostratados con FGA+ 600UI de eCG y CIDR+350UI de eCG obtuvieron idénticosresultadosenloqueaporcentajedegestaciónserefiere(79,78%),siendo,además,significativamentemásaltosqueenelrestodegrupos(MoradiKoretal.,2012).
Swelum y colaboradores (2015), en cambio, obtuvieron porcentajes degestación mayores, y significativamente distintos, en ovejas tratadas con CIDR(77,86%) que en aquellas tratadas con esponjas de FGA (62,41%). También laprolificidad fuemayoren lasovejas tratadasconCIDR frenteaaquellas tratadasconFGA(1,34vs.1,19)(Swelumetal.,2015).
Yadiycolaboradores(2011)obtuvieronporcentajesdepreñezmásaltosenovejasderazaKalkuhitratadasconMPA,queenovejastratadasconCIDR(45%vs.

Discusión
157
35%),encambiolaprolificidadfuemáselevadaenelcasodelosCIDR,yaqueelporcentajedegestacionesgemelaresfrenteagestacionessimplesfuemayorenelgrupo CIDR (14%) que en el grupoMPA (11%). En este trabajo la duración deltratamiento fuede14díaspara laesponja intravaginal impregnadaenMPAyde12díasparaelCIDR, conadministraciónde500UIdeeCGenelmomentode laretirada.Lasdiferenciaspodríanserdebidasalaraza,edad,factoresnutricionales,omanejo reproductivo (Yadietal., 2011).Porotro lado, enel casode laespeciecaprina, Romano (2004) obtuvo resultados exactamente iguales con cabrasnubianasencuantoaporcentajesdegestaciónenlosgruposCIDRyFGA(63%)yun pocomás altos en el grupoMPA (65%), no observando ninguna pérdida dedispositivoenningunodelostresgrupos(Romano,2004).Crosbyycolaboradores(1991) obtuvieron resultados, sólo numéricamente más elevados en ovejastratadasconCIDR(76%)queconprogestágenossintéticos(67%conFGAy66%con MAP) en anestro. En cambio, estos resultados se invirtieron en épocareproductiva,dondeelporcentajedegestaciónobtenidoenanimalestratadosconCIDR(56%)fueinferioralobtenidoenlosotrosdosgrupos(88%enelgrupoFGAy 80% en el grupo MAP) (Crosby et al., 1991). Estudios posteriores en cabrasnadooshantratadasconCIDRdurante18díasreflejanporcentajesdegestacióndel91,3%,muchomayoresalosobtenidosenestetrabajoconesemismodispositivoenovejaslacaune(Shahnehetal.,2008).
La diferencia de fertilidad entre los resultados observados en nuestrotrabajoyloscitadosenotrostrabajospodríanexplicarsesitenemosencuentaquelagranmayoríadelasovejasdelpresenteestudioestabanenlactación,yaque,poruna parte, actividades fisiológicas tales como el movimiento, la lactación o lareproducciónpuedeninducirnivelesbajosdeenergía,yporotra,nivelesbajosdeenergíaprovocanunamenor liberacióndeGnRH, ypor tantodeLHyFSHen lacirculación(Downing&Scaramuzzi,1991).Porotro lado,hayoscilacionesde losvalores concretos de fertilidad según lotes, también observadas en los rebañosnuestros, que, aunque dan lugar a unamedia anual de fertilidadmantenida del75% anual, en determinados lotes de montas, llega a ser menor. Aun así, estacircunstancia no afecta a la comparativa estudiada en el trabajo, ya que en cadaréplica siempre se han incluido animales en grupo CIDR y en grupo FGA, paracontrolar este factor, y por otro lado, y como ha quedado patente, también haynumerosos trabajosqueobtienen cifrasdeporcentajedegestación semejantesalasnuestras.
En cuanto a las otras variables evaluadas, comoporcentaje de pérdidas yporcentaje de dispositivos con olor desagradable, el porcentaje de pérdidas dedispositivosfuemayorenelcasodelasesponjas,dondeun4,46%delosanimaleslas perdieron, frente a un1,90%depérdidas observadas en el casode los CIDR(P=0,332).Enelmomentode laretiradade losdispositivossolamenteun1%delos CIDR presentaron olor desagradable, frente al 76% de esponjas con estacaracterística.

Discusión
158
Omontese y colaboradores (2010) compararon tratamientos con CIDRfrenteaesponjasdeFGAdurante21díasysinadministracióndeeCGalaretiradaenlarazaovinaredsokoto.Enestecasonosevaloróporcentajedegestación,sinoporcentajedeanimalesquesalieronencelo,siendoéstesignificativamenteinferioren los animales tratados con FGA (20%) que en los tratados con CIDR (55,6%)(Omontese et al., 2010). Estos resultados podrían deberse a estros silenciosos,bajosnivelesde actividadovárica (Wheatonetal., 1993), efectode la estaciónocondiciones climatológicas y causas iatrogénicas (Freitas et al., 1997). Sinembargo, los autores del trabajo argumentaban que estos resultados podríanexplicarseporelhechodequeeldispositivohabíaestado21díasinsertadoenlavagina,locualhabríapodidoprovocarunavaginitisyportanto,quelosanimalesnosaliesenencelo.Algunosautores (Alifakiotistetal.,1982),porsuexperienciaprevia con el uso de CIDR en ovejas, indican que las técnicas empleadas parainsertarlasesponjasyfactorestalescomolatexturayconsistenciadelasmismaspodríanteneralgunainfluenciaenlaretencióndelaesponjaenlavaginafrentealCIDR,fabricadoapartirdeunelastómerodesilicona.Talycomoyareseñamosenlarevisiónbibliográfica,adiferenciadeloqueocurreconelCIDR,elcontactoquemantienen lasesponjasvaginalesconelcérvixpredisponeaun incrementode lamicrobiota bacteriana que puede provocar vaginitis y formar adherencias quecausenproblemasreproductivosenelrebaño(Amiridis&Cseh,2012).
Dehecho,y ligadoaestacircunstancia,ennuestrotrabajoobservamosunmenor porcentaje de dispositivos CIDR con olor desagradable (lo que se liga asecreciones anormalesde lamucosavaginal y vaginitis), respectodelporcentajeapreciado en los dispositivos FGA. Estos mismos autores previamente citados,también notaron unamayor pérdida de dispositivos en el caso de las esponjas,comparativamente con losCIDR,hechoque relacionandirectamente a la tasadeirritaciónvaginal.Losporcentajesdepérdidasdescritossonmuchísimomayoresquelosobservadosennuestrotrabajo(40%vs.10%enelestudiocitadoy4%vs.1%enelnuestro).Ladiferenciatanmarcadadelasmediasdepérdidasentreesteestudio y el nuestro, podría deberse al mayor tiempo de permanencia deldispositivo en la vagina (21 días frente a 14). Además los dispositivos de FGAllevan un hilo más largo y de color más llamativo que el del CIDR, que estransparenteymuchomáscorto,deformaquelasovejastienenmayordificultadparaquitarseesteúltimo.
OtrosautorescomoSwelumycolaboradores(2015),tambiéndocumentanqueladescargavaginalmaloliente,consideradacomounarespuestadelamucosavaginal a la presencia de un cuerpo extraño, también fue mucho mayor en elmomentodelaretiradadelasesponjas(98,58%)queeneldelosCIDR(51,91%)(Swelumetal.,2015).

Discusión
159
En el caso del trabajo previamente citado el porcentaje de pérdidas fuemayor en el caso de los CIDR (8,2%) que en el de las esponjas (4%) y tambiénmayor que en nuestro experimento (Rhodes&Nathaniel, 1988). Las razones deestoshechosno laspodemosexplicar enbase a lodescrito en los trabajos, peropodría también depender en parte de la experiencia de aplicación por parte delpersonal encargado delmismo. De hecho, y a diferencia de nuestra experiencia,algunos autores sí describen algunas desventajas en el uso de los dispositivosCIDR, que nosotros no encontramos, como la dificultad para insertarlos en lavaginadelasovejas(Greyling&Brink,1987).
Concluyendo,nuestrosresultadosreflejanporcentajessimilaresrespectoafertilidad y prolificidad, no observando diferencias significativas entre losdispositivoscomparados,porloquelosCIDR,cuandoesténdisponiblesenEuropa,seríanunaalternativa,conventajasenloqueserefierealporcentajedepérdidasyreduccióndevaginitis,loqueequivaleaunamejoraenbienestaranimal.Además,estos dispositivos tienen la posibilidad de ser reutilizados, previa esterilizacióndosoinclusohastatresveces(Coxetal.,2012;Vilarinoetal.,2013).

Discusión
160
5.4 Efectodeltipodedispositivoprogestativointravaginal(progesterona natural o CIDR versus progestágenosintético o FGA) sobre la funcionalidad hipofisaria yovárica en ovejas tratadas con un protocolo desincronización de celos corto (7 días) e influencia de laépocareproductiva
Enlafaseexperimental4sevaloraronposiblesdiferenciasenlarespuestaados dispositivos diferentes con progestativos (progestágenos en esponjasintravaginales con FGA y progesterona en CIDR) basados en el protocolo cortoconsiderado como el de mejor resultado en las fases experimentales anteriores(protocoloCorto-PG).
Habiendo comprobado en la fase experimental 1 que este tratamientoCorto-PG daba lugar a diferencias en el porcentaje de gestación según elfotoperiodo, se llevaron a cabo dos réplicas en distinta estación. Así, elexperimento 4.1 se realizó en fotoperiodo creciente, y el experimento 4.2, enfotoperiododecreciente
En esta parte, los factores de confusión a controlar fueron la condicióncorporal y la edadde los animales, que resultaronhomogéneos entre grupos enambasréplicas.
En época de fotoperiodo creciente se comprobó que no hubo ningunavariación según tratamiento, en lo que respecta al número medio de cuerposlúteos/animal, al tiempo medio en salir en celo ni a la respuesta total altratamiento(estoes,elsalironoencelo),yaqueenlosdosgruposel100%delosanimalesmostraroncelo.Elanálisispormomentosdedeteccióndecelostampocorevelóningunavariaciónentre losgruposencuantoalritmoyagrupaciónde losanimalesenestarespuesta,demostrándoselamismaeficaciaenlasincronizacióndecelosdelosdispositivosCIDRrespectodelastradicionalesesponjas.
Durante el fotoperiodo decreciente no observamos diferencias en elporcentajedeovejasquesalieronencelo,nieneltiempomedioenmostrarlo.Sinembargo,síobservamosunagrupamientomayoren lasalidaenceloenelgrupotratadoconeldispositivoCIDR,demaneraquealas28hy32hhabíamásdeldoblede ovejas en celo en este grupo respecto del grupo FGA. Posteriormente, con eltiempo de observación de celos, esta diferencia se iguala, y no llega a sersignificativa.
En otros trabajos se ha demostrado que el dispositivo CIDR conprogesteronanaturalesigualmenteeficazencomparaciónconlasesponjasdeFGA(Ungerfeld&Rubianes,2002;Moeinietal.,2013;Swelumetal.,2015),resultados

Discusión
161
similares a los del presente estudio, observados en la estación desfavorable, endondeprecisamenteseesperaquelaadicióndeunprogestativoexternoayudedemaneraeficazalosanimalesenanestro,oapuntodesalirdelmismo(Ungerfeld&Rubianes, 1999; Vinoles etal., 2001). De hecho, los resultados de respuesta queobservamos son comparables a los resultados citados enotros trabajos con estetipo de protocolos cortos en estación favorable (Vilarino et al., 2013) odesfavorable (Moradi Kor etal., 2012; Swelum etal., 2015). Por otro lado, en elpresentetrabajo,corroboramosinclusomejorestasasdepresentacióndeceloenlaestacióndesfavorable,respectodelaestaciónreproductiva(79,20%enelgrupoCIDRfrenteal88,00%enesponjasenfotoperiododecreciente,respectodel100%enambosgruposenelfotoperiodocreciente).
Los resultados en la respuesta al tratamiento obtenidos por Yadi et al.(2011) en fotoperiodo creciente no muestran diferencias significativas entre elgrupo tratado con esponjas de FGA y el grupo tratado con CIDR, con unosporcentajesdesalidaencelodelasovejasde45%y35%,respectivamente,aunqueinferiores a los obtenidos en nuestro trabajo en el mismo fotoperiodo (100%)(Yadietal.,2011).
Resultados similares en cuanto a respuesta al tratamiento en fotoperiodocreciente fueron obtenidos por Ungerfeld y Rubianes (2002), donde obtuvieronporcentajesderespuestaaltratamientode91,5%y95,9%enovejastratadasconesponjas de FGA y CIDR, respectivamente. Estos autores también observarontiempos medios de inicio de celo semejantes entre grupos, y por debajo de 40horas,peroligeramentemásaltosquelosnuestros(38,8±1,6y39,9±2,1henlosgrupos FGA y CIDR, frente a los 32,00% ± 6,01 y 33,76% ± 4,01 obtenidos ennuestro trabajo en los mismos grupos). Sin embargo, el tiempo obtenido en elgrupotratadoconesponjasdeFGA(38,8±1,6h)fuemuysimilarporelobservadopornosotrosenelmismogrupoperoenfotoperiododecreciente(38,18±9,46h)(Ungerfeld&Rubianes,2002).
Denuevo,eneltrabajodeMoradiKoretal.(2012),tambiénenfotoperiodocreciente,sedescribenresultadosmuysimilaresalosnuestroseneltiempomediodesalidaencelo,con31,10±1,6henelgrupoCIDRy32,65±1,80henelgrupoFGA,y,aunquelosporcentajesderespuestaaltratamientofueronelevados,síquehubo diferencias significativas entre la respuesta observada en el grupo CIDR(82,45%) y el grupo FGA (92,89%) (Moradi Kor et al., 2012). Por el contrario,Romano(2004),utilizandocabrasnubianasendistintofotoperiodo(decreciente),observódiferenciassignificativaseneltiempomediodesalidaencelo(32,90±9,7horasenlascabrastratadasconesponjasdeFGA,frentea40,2±10,5horasenlascabras tratadasconCIDR).Encuantoa la salidaencelo, los resultados sí fueronidénticos a los obtenidos en nuestro trabajo en fotoperiodo creciente, con unarespuestadel100%delosanimalesenambosgrupos(Romano,2004).

Discusión
162
Estas diferencias en los resultados entre estudios podrían deberse a losdistintos protocolos utilizados (Cam&Kuran, 2003; Gomez-Brunet etal., 2007).Sinembargo,otrosestudiosafirmanqueestadiscrepanciaentredistintosautorespuedeserexplicadaporlafaltadehomogeneidadenlacondicióncorporal,laraza,elsistemademanejooelestadofisiológicoynutricional(Yadietal.,2011;Moeiniet al., 2013). Por ello, es importante ratificar la eficiencia de los diferentesprotocolos y productos hormonales en las razas y en sistemas de producciónconcretos. De hecho, una aportación muy novedosa del presente trabajo es laverificacióndelaeficaciaenrazasutilizadasdemanerahabitualenexplotacionesde ovino de carne, y en las latitudes españolas. Hasta ahora, la mayoría de lostrabajos realizados con ovejas de razas españolas utilizaban animales de razamerina,larazaespañolamásinternacionalmenteconocida(Gardónetal.,2015).
En lo que respecta a la funcionalidad ovárica durante la fase folicular,reflejadaenbasealosanálisisdenivelesdeE2,seobservóunasemejanzaendichafuncionalidad entre ambos tipos de progestativos, excepto en la hora 12, dondeestasdiferenciassísehicieronsignificativas(P<0,0001).LomismoocurrióconlaP4,dondelosnivelesenplasmaobservadosfueronalgomásaltosenelgrupoFGAperoclínicamentenosepuedenconsiderarrelevantes(5,94±1,42ng/mlfrentea4,34±1,96ng/ml,respectivamente).LosresultadosobtenidosennuestrotrabajodifierendelospublicadosporSwelumetal.(2015),dondelosnivelesdeE2apenasvariaron en las horas 0, 24 y 48 posteriores a la retirada de los dispositivosintravaginales, siendo significativamente más altos (P<0,05) en el grupo deanimales a los que se les insertó CIDR que en aquéllos que llevaban esponja deFGA, al igual que los niveles de P4, más altos en el grupo CIDR (Swelum et al.,2015).IgualmenteHamraetal.(1989)yNaderipouretal.(2012)observaronquelos animales tratados con CIDR producían niveles más elevados que aquellostratados con esponjas. Las diferencias entre nuestros protocolos y los de losautores referenciados radicanposiblemente en el tiempodepermanencia de losdispositivosintravaginales,yaquelossuyosfuerontratamientos“largos”(Hamraetal.,1989;Naderipouretal.,2012).
Una vez demostrada semejanza en la eficacia de sincronización (o, segúnalgunostrabajos,superioridad),lautilizacióndelosCIDRrespectodelasesponjaspodríaserdeelecciónporladiferenciaenlastasasdevaginitisinducidas,inclusoenlostratamientoscortos,yaquelasuperficiedecontactoconlamucosavaginales mucho menor, lo que da lugar a una menor irritación, y consiguienteinflamación. La naturaleza de la textura de las esponjas absorbe muchas mássecreciones vaginales, comparado con la siliconano absorbentedelCIDR, loquepermiteelcrecimientoylamultiplicacióndemicroorganismos,loqueprovocaquedesprendanmal olor y además una reacción inflamatoria de la mucosa vaginal,aunquesealeve.Swelumetal.(2015)observaronun9,22%deesponjasadheridasa lamucosa vaginal, causandounmayor porcentaje de lesiones tras su retirada,

Discusión
163
que los tratamientos con CIDR, en los que no se observó ninguna adherencia(Swelumetal.,2015).Dehecho,Manesetal.(2010)yareportaronquelasesponjasintravaginales utilizadas para inducir y sincronizar el estro en ovejas generabanunarespuestainflamatoriaconcambioshistológicosycitológicosenlapareddelavagina(Manesetal.,2010).Estasobservacionesconfirman lahipótesisdequeeluso de estos dispositivos intravaginales producen, no sólo un cambio en lamicrobiotavaginal,sinotambiénunprocesodeinflamaciónsubagudaenlaparedde la vagina (Manesetal., 2014).Asímismo, en este trabajo sedemostróque elflujopresenteenlasovejasdespuésdeuntratamientoconesponjasintravaginalesimpregnadas con progestágenos tiene efectos negativos en la calidad yfuncionalidaddelsemendelmorueco(Manesetal.,2015).
OtradelasventajasdelCIDRessuposiblereutilización(trasesterilizaciónquímicaotratamientoporautoclave)dealmenos,dosveces(Coxetal.,2012),oinclusotres(Vilarinoetal.,2013).Igualmente,Coxetal.(2012)mostraronqueeltratamientocortoconCIDRcombinadoconPGF2αenelmomentode la insercióndel dispositivo permitía la reutilización de los dispositivos sin que afectase a ladinámicafolicularyalintervalodesalidaenceloyovulación.
Enunterceruso,Vilariñoetal.(2013)comprobaronqueCIDRusadosporsegundaoterceravezfuerontanefectivoscomolosnuevosenloqueserefiereadinámica folicular, sincronización del estro y ovulación. Sin embargo, hay quedestacar que en los dispositivos utilizados por tercera vez se observó unadisminuciónenlosnivelesdeP4yundescensoenelporcentajedegestacionesdeaproximadamenteun10%(Vilarinoetal.,2013).
Finalmente, otra de las ventajas de los CIDR las tasas de pérdidas de losdispositivos intravaginales comparando CIDR respecto de esponjas es tambiénmenor(6,3%versus0,8%paraesponjasyCIDR,respectivamente)(Hashemietal.,2006).
Endefinitiva,enelpresentetrabajonoobservamosdiferenciasenlatasaderespuestamedidacomoporcentajedepresentacióndecelo,traslasincronizacióncon losdos tiposdedispositivo intravaginaldeprogestativos comparados (CIDRrespectodeesponjas),aexcepcióndeunamayoragrupacióndelasovejasensalirenceloenelcasodelgrupoCIDRduranteelfotoperiodocreciente(efectopositivodelgrupoCIDR);aexcepcióntambiéndeunamenortasadeliberacióndeE2enlafase folicular y de progesterona en la luteal (efecto que podría ser negativo delgrupoCIDR)yunamayorceleridaddelpicodeLHtraseldeE2(efectopositivodelgrupoCIDR).Porlotanto,secorroboralaposibilidaddeidoneidaddelautilizaciónde ambos dispositivos hormonales en explotaciones ganaderas dentro de unprotocolocortode7dconaplicacióndeprostaglandinaalaretirada.Además,enelcasode losdispositivosCIDR,sesuman lasventajasde lamejoraenelbienestaranimal(reduccióndelporcentajedevaginitisasociadasalapresenciadelaesponja

Discusión
164
en la vagina de los animales), la reducción de pérdidas de los dispositivos porparte de los animales sincronizados y la posibilidad de su reutilización tras suesterilización.

CONCLUSIONES


Conclusiones
167
6 Conclusiones
1. La duración del tratamiento con dispositivos intravaginalesimpregnados con progestágenos sintéticos durante el fotoperiododecreciente puede ser reducida de 14 días utilizados en los tratamientosclásicos hasta 7 días, siempre que se acompañe de la administración deprostaglandina, ya que se obtienen similares porcentajes de gestación yprolificidad.2. El estudio enprofundidadde la funcionalidad ovárica e hipofisariaenfotoperiododecrecientedemostrópatronesendocrinos,decrecimientoyfuncionalidadfolicularsimilaresenel tratamiento largode14díasyeneltratamientocortode7díasconaplicacióndeprostaglandinaa la retiradadelprogestágeno(protocoloCorto-PG)y,enamboscasos,mejoresqueeneltratamiento corto con aplicación de prostaglandina a la inserción delprogestágeno(protocoloPG-Corto).3. El protocolo Corto-PG se relacionó con una menor fertilidad enfotoperiodocreciente.Estamenor fertilidadpodríadebersea lapresenciadecuerposlúteosconbajarespuestaaprostaglandinasduranteelanestroreproductivo.4. EltratamientoControlde14díasconlosdispositivosintravaginalesdeprogesterona(CIDR)yconlasesponjas intravaginalesdeprogestágeno(FGA)danlugaraporcentajesdegestaciónsimilaresduranteelfotoperiododecreciente.5. El tratamiento Corto-PG presenta eficacia similar en cuanto alporcentaje de presentación de celos y funcionalidad ovárica e hipofisaria,tanto con dispositivos intravaginales de progesterona (CIDR), como conesponjas intravaginales de progestágeno (FGA), durante el fotoperiodocreciente.6. Durante el fotoperiodo creciente, el tratamiento Corto-PG condispositivos intravaginales tipo CIDR da lugar a una agrupación de celosmayorperoconunamenorconcentracióndeE2aliniciodelafasefolicularyunamenorconcentracióndeP4durantelafaseluteal,respectoalmismotratamientoconesponjas(FGA),duranteelfotoperiodocreciente.


BIBLIOGRAFÍA


Bibliografía
171
7 Bibliografía
Abdoli R, Zamani P, Mirhoseini SZ, Ghavi Hossein-Zadeh N, Nadri S (2016) Areviewonprolificacygenes in sheep.Reproduction indomesticanimals=Zuchthygiene51,631-637.
AbeciaA,ForcadaF (2011)Bases fisiológicasde la reproducciónen laoveja.En:Manejoreproductivoenganadoovino,Ed.Servet.GrupoAsísBiomedia,S.L.Zaragoza,España.216Pp.
AbeciaJA(1992)Efectodelplanodealimentaciónydelniveldereservassobrelosparámetros reproductivos en ovejas de raza Rasa Aragonesa durante elanoestro estacionario y al inicio de la época de actividad sexual. Tesis dedoctorado,Univ.deZaragoza,309págs.
Abecia JA, Forcada F, Gonzalez-Bulnes A (2011) Pharmaceutical control ofreproduction in sheep and goats. Vet ClinNorthAmFoodAnimPract 27,67-79.
AbeciaJA,ForcadaF,Gonzalez-BulnesA(2012)Hormonalcontrolofreproductioninsmallruminants.Animalreproductionscience130,173-179.
Acritopoulou S, HaresignW (1980) Response of ewes to a single injection of ananalogue of PGF-2 alpha given at different stages of the oestrous cycle.Journalofreproductionandfertility58,219-221.
AcritopoulouS,HaresignW,Foster JP,LammingGE(1977)Plasmaprogesteroneand LH concentrations in ewes after injection of an analogue ofprostaglandinF-2alpha.Journalofreproductionandfertility49,337-340.
AdamsGP (1999) Comparative patterns of follicle development and selection inruminants.JournalofreproductionandfertilitySupplement54,17-32.
Adams GP, Matteri RL, Ginther OJ (1992) Effect of progesterone on ovarianfollicles, emergence of follicular waves and circulating follicle-stimulatinghormoneinheifers.Journalofreproductionandfertility96,627-640.
Alifakiotist T, Michailidis I, Gavrilidis G (1982) Induced breeding in anoestrousmilking ewes on dairy breed: Comparison of norgestometmedroxyprogesterone and fluorogestone in two regimes of PMSG.Theriogenology17,603-610.
AlonsodeMiguelM,CogniéY(1980)Variacionesdelaactividadsexualdelaoveja«RasaAragonesa»duranteelperíododelanoestroestacionario.Efectodelaedad, de las condiciones climáticas y, de la presencia de machos. IXCongresoIntern.deReprod.Anim.eIA,Madrid,16-20dejunio,Vol.IV:387-392.
AmerHA,HazzaaAM(2009)Theeffectofdifferentprogesteroneprotocolsonthereproductive efficiency of ewes during the non-breeding season. .Veterinarskiarhiv79,19-30.
AmiridisGS,CsehS(2012)Assistedreproductivetechnologiesinthereproductivemanagement of small ruminants. Animal reproduction science 130, 152-161.
Ammar-Khodja F, Brudieux R (1982) Seasonal variations in the cyclic lutealovarianactivity in theTadmitewe inAlgeria. Journalof reproductionandfertility65,305-311.

Bibliografía
172
AnelL,KaabiM,AbrougB,AlvarezM,AnelE,BoixoJC,de laFuenteLF,dePazP(2005)Factorsinfluencingthesuccessofvaginalandlaparoscopicartificialinseminationinchurraewes:afieldassay.Theriogenology63,1235-1247.
ArthurGH,PearsonDE (1989)Normal reproductive function.En: SaundersW.B.(ed) Veterinary Reproduction and Obstetrics: 22-24. Balliere Tindal,London.
Ataman MB, Akoz M, Akman O (2006) Induction of synchronized oestrus inakkaramancross-bredewesduringbreedingandanestrusseasons:theuseofshort-termandlong-termprogesteronetreatments.RevueMédVét157,257-260.
AvdiM,VergosV,AlifakiotisT,MichailidisI,DriancourtMA,P.C(1988)Seasonalvariations of oestrous behaviour and ovulation rate in Chios and Semesewes inGreece. Proc. 3rdWorldCongon Sheep andBeef Cattle París, EdINRAPubl.,Pars,19-23dejunio,Vol.2:647-649.
BarilletF,MarieC, JacquinM,LagriffoulG,AstrucJM(2001)TheFrenchLacaunedairysheepbreed:useinFranceandabroadinthelast40years.LivestockProductionScience71,17-29.
BarrettDM,BartlewskiPM,CookSJ,RawlingsNC(2002)UltrasoundandendocrineevaluationoftheovarianresponsetoPGF2alphagivenatdifferentstagesofthelutealphaseinewes.Theriogenology58,1409-1424.
Bartlewski PM, Baby TE, Giffin JL (2011) Reproductive cycles in sheep. Animalreproductionscience124,259-268.
BartlewskiPM,BeardAP,RawlingsNC(1999)Anultrasonographicstudyoflutealfunction inbreedsofsheepwithdifferentovulationrates.Theriogenology52,115-130.
Bartlewski PM, Beard AP, Rawlings NC (2000) An ultrasound-aided study oftemporal relationships between the patterns of LH/FSH secretion,development of ovulatory-sized antral follicles and formation of corporaluteainewes.Theriogenology54,229-245.
Baserga R (1985) The biology of cell reproduction. Harvard University Press,Cambridge,M.A.
BlaschiW,LunardelliPA,MarinhoLS,MaxMC,SantosGM,Silva-SantosKC,Melo-Sterza FA, Baldassarre H, Rigo TR, Seneda MM (2014) Effects ofprogestagen exposure duration on estrus synchronization and conceptionratesofcrossbreedewesundergoingfixedtimeartificialinsemination.JVetSci15,433-437.
BodinL,DiPascualeE,FabreS,BontouxM,MongetP,PersaniL,MulsantP(2007)A novel mutation in the bone morphogenetic proteína 15 gene causingdefectiveproteinsecretionisassociatedwithbothincreasedovulationrateandsterilityinLacaunesheep.Endocrinology148(1),393-400.
BolandMP,GordonI,KelleherDL(1978)Theeffectoftreatment.byprostaglandinanalogue (ICI-80996) or progestagen (SC-9880) on ovulation andfertilizationincyclicewes.JournalofAgriculturalScience91,727-730.
Bowen-Shauver JM, Telleria CM (2003) Luteal regression: a redefinition of theterms.Reproductivebiologyandendocrinology:RB&E1,28.
BradenAWH,LamondR,RadfordHM (1960)Control of the timeof ovulation insheep.AustralianJournalofAgriculturalResearch11(3),389-401.

Bibliografía
173
Buxadé C (1996) Zootecnia. Bases de Producción Animal. Producción ovina, VolVIII.1ªed.Mundi-PrensaLibros,Madrid.
Cahill LP, Mauleón P (1981) A study of the population of primordial and smallfolliclesinthesheep.J.Reprod.Fertil.61:201-206.
CamMA,KuranM(2003)Effectsofasingle injectionofhCGorGnRHagonistonday12postmatingonfetalgrowthandreproductiveperformanceofsheep.Animalreproductionscience80,81-90.
CamberoMuñozP (1999)Cuadernode laexplotacióndeovino (ServicioAgrariodeCajaDuero).
CampbellBK,ScaramuzziRJ,WebbR(1995)Controlofantralfollicledevelopmentand selection in sheep and cattle. Journal of reproduction and fertilitySupplement49,335-350.
Chemineau P, Bodin L, MigaudM, Thiery JC, Malpaux B (2010) Neuroendocrineand genetic control of seasonal reproduction in sheep and goats.Reproductionindomesticanimals=Zuchthygiene45Suppl3,42-49.
Chemineau P, Guillaume D, Migaud M, Thiery JC, Pellicer-Rubio MT, Malpaux B(2008) Seasonality of reproduction in mammals: intimate regulatorymechanismsandpracticalimplications.Reproductionindomesticanimals=Zuchthygiene43Suppl2,40-47.
Chemineau P, Vandaele E, Brice G, Jardon C (1991) Utilisation des implants demelatoninepour l'améliorationdesperformancesde reproductionchez labrebis.Rech.MédVét.,167(3/4):227-239.
Childs GV (2006) Gonadotropes and lactotropes. In: The physiology ofreproduction, Volume 1. J.D., Neill (Ed.). Raven Press, Ltd., New York. pp.1483-1580.
Clarke IJ,PompoloS (2005)Synthesis and secretionofGnRH.Anim.Reprod. Sci.88:29-55.
Contreras-SolisI,VasquezB,DiazT,LetelierC,Lopez-SebastianA,Gonzalez-BulnesA (2009) Efficiency of estrous synchronization in tropical sheep bycombining short-interval cloprostenol-based protocols and "male effect".Theriogenology71,1018-1025.
Cox JF, Allende R, Lara E, Leiva A, Díaz T, Dorado J, Saravia F (2012) Folliculardynamics, interval to ovulation and fertility after AI in short-termprogesterone and PGF2α oestrous synchronization protocol in sheep. .Reproductionindomesticanimals=Zuchthygiene47,946-951.
CrosbyTF,BolandMP,Gordon I (1991)Effect of progestagen treatments on theincidence of oestrous and pregnancy rates in ewes. Animal reproductionscience24,109-118.
Daniel JA, Sterle SW,McFadin-Buff EL, KeislerDH (2001) Breeding ewes out-of-season using melengestrol acetate, one injection of progesterone, or acontrolledinternaldrugreleasingdevice.Theriogenology56,105-110.
DazaA,BlancoPJ(1990)ProduccióndeganadoovinodecarneenEspaña.Razasycruces.Suactualproblemática.MundoGanadero.Madrid.
Daza Andrada A (2002) Mejora de la productividad y planificación deexplotacionesovinas.S.A.AgrícolaEspañola.
DelgadilloJA,GelezH,UngerfeldR,HawkenPA,MartinGB(2009)The'maleeffect'insheepandgoats--revisitingthedogmas.BehavBrainRes200,304-314.

Bibliografía
174
DelgadilloJA,MartinGB(2015)Alternativemethodsforcontrolofreproductioninsmall ruminants: A focus on the needs of grazing industries. AnimalFrontiers5,57-65.
DeNicolo G, Morris ST, Kenyon PR, Morel PC, Parkinson TJ (2008) Melatonin-improved reproductive performance in sheep bred out of season. Animalreproductionscience109,124-133.
Douglas RH, Ginther OJ (1973) Luteolysis following a single injection ofprostaglandinF2alphainsheep.Journalofanimalscience37,990-993.
Downing JA, Scaramuzzi RJ (1991) Nutrient effects on ovulation rate, ovarianfunction and the secretion of gonadotrophic and metabolic hormones insheep.JournalofreproductionandfertilitySupplement43,209-227.
DriancourtMA(1991)Folliculardynamicsinsheepandcattle.Theriogenology.35:55-79.
DriancourtMA,PhiliponP,LocatelliA,JacquesE,WebbR(1988)AredifferencesinFSHconcentrationinvolvedinthecontrolofovulationrateinRomanovandILe-de-Franceewes?J.Reprod.Fertil.83:509-516.
Dutt RH, Casida LE (1948) Alteration of the estrual cycle in sheep by use ofprogesterone and its effect upon subsequent ovulation and fertility.Endocrinology43,208-217.
Dwyer CM, Bunger L (2012) Factors affecting dystocia and offspring vigour indifferentsheepgenotypes.Preventiveveterinarymedicine103,257-264.
Einer-JensenN,HunterR(2005)Counter-currenttransferinreproductivebiology.Reproduction129,9-18.
ElviraL,HernandezF,CuestaP,CanoS,Gonzalez-MartinJV,AstizS(2013)Factorsaffecting the lactation curves of intensively managed sheep based on aclusteringapproach.TheJournalofdairyresearch80,439-447.
Espey LL (1994) Current status of the hypothesis that mammalian ovulation iscomparabletoaninflammatoryreaction.Biologyofreproduction50,233-238.
Evans AC (2003) Characteristics of ovarian follicle development in domesticanimals.Reproductionindomesticanimals=Zuchthygiene38,240-246.
EvansAC,DuffyP,CrosbyTF,HawkenPA,BolandMP,BeardAP(2004)Effectofram exposure at the end of progestagen treatment on estrussynchronisation and fertility during the breeding season in ewes. Animalreproductionscience84,349-358.
Evans AC, Duffy P, Hynes N, Boland MP (2000) Waves of follicle developmentduringtheestrouscycleinsheep.Theriogenology53,699-715.
FAOSTAT (2015) Disponible desde internet:http://www.fao.org/statistics/databases/en/[consulta:agostode2016].
FEAGAS(2010)Razasdeganadoovino.EnCatálogoOficialdeRazasdeGanadodeEspaña (Federación Española Asociaciones Ganado Selecto). Disponibledesde internet: http://new.feagas.com/razas/ovino/segureno/ [consulta:juliode2016].
FieldPA(1991)Relaxinandother lutealsecretorypeptides:cell localizationandfunctionintheovary.In:Ultrastructureoftheovary.G.Familiari,S.MakabeandP.M.Motta(Eds.).KluwerAcademicPress,Boston.Pp.177-198.
FierroS,VinolesC,Olivera-MuzanteJ(2016)Concentrationsofsteroidhormones,estrous,ovarianandreproductiveresponsesinsheepestroussynchronized

Bibliografía
175
withdifferentprostaglandin-basedprotocols.Animalreproductionscience167,74-82.
Fink G (1988) Gonadotrophin secretion and its control. In: The physiology ofreproduction, Volume 2. J.D., Neill (Ed.). Raven Press, Ltd., New York. Pp.1349-1377.
Fitzgerald JA, Ruggles AJ, Stellflug JN, Hansel W (1985) A seven-daysynchronization method for ewes using medroxyprogesterone acetate(MAP)andprostaglandinF2alpha.Journalofanimalscience61,466-469.
FleischA,BollweinH,PiechottaM, JanettF (2015)ReproductiveperformanceofLacaune dairy sheep exposed to artificial long days followed by naturalphotoperiodwithoutandwithadditionalprogestagentreatmentduringthenonbreedingseason.Theriogenology83,320-325.
FlintAPF,SheldrickEL,TheodosisDT,WoodingFBP(1986)Ovarianpeptides:roloflutealoxytocininthecontrolofestrouscyclicityinruminant.J.Anim.Sci.62(Suppl.2):62-71.
Flox JR, Daza A (2000) Mejora de la productividad en ganado ovino. RevistaGanaderíaISSN1695-1123.0:10-17..
FolchJ,AlabartJL(2000)CaracterísticasreproductivasdelaovejaRasaAragonesa.Ovis68,27-36.
Forcada F, Abecia JA, Sierra I (1992) Seasonal changes in oestrus activity andovulation rate in Rasa Aragonesa ewesmaintained at two different bodyconditionlevels.SmallRuminantResearch8,313-324.
Fortune JE (2003) The early stages of follicular development: activation ofprimordial folliclesandgrowthofpreantral follicles.AnimReprodSci.78:135-163.
FragkouAI,MavrogianniSV,FthenakisGC(2010)Diagnosticinvestigationofcasesofdeathsofnewbornlambs.SmallRuminantResearch92(1):41-44.
FreitasVJF,BarilG,MartinGB,SaumandeJ (1997)Physiological limits to furtherimprovement in the efficiency of oestrous synchronization in goats.Reproduction,fertility,anddevelopment9,551-556.
Fuentes FC, Sánchez JM,GonzaloC (2000)Manual de etnología animal: razasderumiantes.1ªed.,Murcia,DiegoMarínLibreroeditor,494págs:259-262.
FukuiY, IshikawaD, IshidaN,OkadaM, ItagakiR,OgisoT(1999)Comparisonoffertility of estrous synchronized ewes with four different intravaginaldevices during the breeding season. Journal of Reproduction andDevelopment45,337-343.
García Álvarez-Coque JM (2004) La Reforma de la PAC y el futuro de la ayudasagrarias.RevistaValencianadeEconomíayHacienda.11:164-183..
GardónJC,EscribanoB,AstizS,RuizS(2015)SynchronizationprotocolsinSpanishMerino Sheep: reduction in time to estrus by the addition of eCG to aprogesterone-basedestrussynchronizationprotocol.AnnAnimSci15,409-418.
Gerlach T, Aurich JE (2000) Regulation of seasonal reproductive activity in thestallion,ramandhamster.Animalreproductionscience58,197-213.
GeschwindI,DeweyR(1968)Dynamicsof luteinisinghormone(LH)secretioninthe cycling ewe: a radioimmunoassay study. Proc Soc Exp BiolMed. 129:451-455.

Bibliografía
176
GibbonsJR,WiltbankMC,GintherOJ(1997)Functionalinterrelationshipsbetweenfollicles greater than 4mm and the follicle-stimulating hormone surge inheifers.Biologyofreproduction57,1066-1073.
Gibbons JR, Wiltbank MC, Ginther OJ (1999) Relationship between folliculardevelopment and the decline in the follicle-stimulating hormone surge inheifers.Biologyofreproduction60,72-77.
GintherOJ,BergfeltDR,KulickLJ,KotK(2000)Selectionofthedominantfollicleincattle: role of two-way functional coupling between follicle-stimulatinghormoneandthefollicles.Biologyofreproduction62,920-927.
Ginther OJ, Kot K, Wiltbank MC (1995) Associations between emergence offollicularwavesandfluctuations inFSHconcentrationsduringtheestrouscycleinewes.Theriogenology43,689-703.
GodfreyRW,CollinsJR,HensleyEL,WheatonJE(1999)Estrussynchronizationandartificialinseminationofhairsheepewesinthetropics.Theriogenology51,985-997.
Gomez-BrunetA,Santiago-MorenoJ,MontoroV,GardeJ,PonsP,Gonzalez-BulnesA,Lopez-SebastianA (2007)Reproductiveperformanceandprogesteronesecretioninestrus-inducedManchegaewestreatedwithhCGatthetimeofAI.SmallRuminantResearch71,117-122.
Gonzalez-Anover P, Encinas T, Veiga-Lopez A, Ammoun I, Contreras I, Ros JM,AriznavarretaC,TresguerresJA,Gonzalez-BulnesA(2007)Effectsofbreedon folliculardynamicsandoestradiol secretionduring the follicularphaseinsheep.Reproductionindomesticanimals=Zuchthygiene42,29-33.
Gonzalez-Bulnes A, García-García RM, Castellanos V, Santiago-Moreno J,Ariznavarreta C, DomínguezV, Lopez-SebastianA, Tresguerres JA, CoceroMJ (2003) Influence of maternal environment on the number oftransferable embryos obtained in response to superovulatory FSHtreatmentsinewes..ReprodNutrDev43,17-28.
Gonzalez-Bulnes A, Souza CJ, Campbell BK, Baird DT (2004) Systemic andintraovarianeffectsofdominantfolliclesonovinefolliculargrowth.Animalreproductionscience84,107-119.
Gonzalez-Bulnes A, Veiga-Lopez A, Garcia P, Garcia-Garcia RM, Ariznavarreta C,Sanchez MA, Tresguerres JA, Cocero MJ, Flores JM (2005) Effects ofprogestagensandprostaglandinanaloguesonovarianfunctionandembryoviabilityinsheep.Theriogenology63,2523-2534.
Gonzalez-GarciaE,TesniereA,CamousS,BocquierF,BarilletF,HassounP(2015)Theeffectsofparity, litter size,physiological state, andmilking frequencyon the metabolic profile of Lacaune dairy ewes. Domestic animalendocrinology50,32-44.
GoodmanAL,HodgenGD(1983)Theovariantriadoftheprimatemenstrualcycle.Recentprogressinhormoneresearch39,1-73.
GoodmanRL(1994)Neuroendocrinecontroloftheovineestrouscycle.En:KnobilE., Neill J.D. (eds) The physiology of reproduction: 659-709. Raven PressLtd.
GordonI(1996)ControlledReproductioninSheepandGoats.(1ªedn),Controlledreproductioninfarmanimals.

Bibliografía
177
Gore-LangtonRE,ArmstrongDT(1994)Follicularsteroidogenesisanditscontrol.En:Thephysiologyofreproduction,Volume1.J.D.,Neill(Ed.).RavenPress,Ltd.,NewYork.Pp.331-385.
Greyling JPC, BrinkWCJ (1987) Synchronization of oestrus in sheep: The use ofcontrolledinternaldrugrelease(CIDR)dispensers.SouthAfricanJournalofAnimalScience17,128-132.
GreylingJPC,WesthuysenJM,VanderandNeikerkCH(1979)Thesynchronizationof oestrus in sheep. 1. Dosage and time of prostaglandin administrationfollowing progestagen pretreatment. . South African Journal of AnimalScience9,185-192.
GungorO,CenesizM,PancarciSM,YildizS,KayaM,KacarC,OzyurtluN,GurbulakK(2007)Effectsofdifferentprogesteronereleasingintravaginaldevicesonestrous synchronization and LH surge in fat-tailed ewes during non-breedingseason..MedycynaWeterynaria63,1316-1319.
Gutierrez-A N (1985) Economic constraints on sheep and goat production indevelopingcountries.InFAOSmallruminantproductioninthedevelopingcountries,Timon,V.M.,Hanrahan,J.P.,eds.(Sofia,Bulgaria,FAO).
HamraAH,McNalleyJW,MarcekJM,CarlsonKM,WheatonJE(1989)Comparisonof progesterone sponges, cronolone sponges and controlled internal drugrelease dispensers on fertility in anestrous ewes. Animal reproductionscience18,219-226.
Hansel W, Convey EM (1983a) Phyiology of the estrous cycle. J. Anim. Sci. 57(Suppl.2):404-424.
HanselW,ConveyEM(1983b)Physiologyof theestrouscycle. Journalofanimalscience57Suppl2,404-424.
HaresignW(1981)Theinfluenceofnutritiononreproductionintheewe1.Effectson ovulation rate, follicle development and luteinizing hormone release.AnimalProduction32,197-202.
Haresign W, Peters AR, Staples LD (1990) The effect of melatonin implants onbreedingactivityandlittersizeincommercialsheepflocksintheUK.AnimProd.,50:111-121.
HashemiM, SafdarianM, Kafi M (2006) Estrous response to synchronization ofestrususingdifferentprogesteronetreatmentsoutsidethenaturalbreedingseasoninewes.SmallRuminantResearch65,279-283.
Hawken PA, Beard AP, Esmaili T, Kadokawa H, Evans AC, Blache D, Martin GB(2007) The introduction of rams induces an increase in pulsatile LHsecretion in cyclic ewes during the breeding season. Theriogenology. 68:56-66.
HendersonDC, Robinson JJ (2007) The reproductive cycle and itsmanipulation.En:Diseasesof sheep.Editedby I.D.Aitken.4th ed.BlackwellPublishingLtd,9600GarsingtonRoad,OxfordOX42DQ,UK.Pp43-53.
Hernandez F, Elvira L, Gonzalez-Martin JV, Gonzalez-Bulnes A, Astiz S (2011)Influence of age at first lambing on reproductive and productiveperformance of Lacaune dairy sheep under an intensive managementsystem.TheJournalofdairyresearch78,160-167.
HolmoyIH,WaageS(2015)TimetrendsandepidemiologicalpatternsofperinatallambmortalityinNorway.ActaveterinariaScandinavica57,65.
HsuehAJW,Adashi EY, JonesPBC,WelshTH (1984)Hormonal regulation of thedifferentiationofculturedovariangranulosecell.EndocrineRev.5:76-127.

Bibliografía
178
Jablonka-ShariffA,Grazul-BilskaAT,RedmerDA,ReynoldsLP(1993)Growthandcellular proliferation of ovine corpora lutea throughout the estrous cycle.Endocrinology133,1871-1879.
JounouR,MolinaE,VillalbaD,Boleda J (2004)Crecimiento en corderosde razaLacaune cárnica: adaptación a las condiciones productivas de la zona deLleida.SEOC.XXIX:59-60.
Juengel JL, Niswender GD (1999) Molecular regulation of luteal progesteronesynthesisindomesticruminants.J.Reprod.Fertil.54(Suppl.):193-205.
Karsch FJ, Bittman EL, Foster DL, Goodman RL, Legan SJ, Robinson JE (1984)Neuroendocrine basis of seasonal reproduction. Recent progress inhormoneresearch40,185-232.
KarschRJ,GoodmanRL,S.L.L(1980)Feed-backbasisofseasonalbreeding:testofanhipothesis.J.Reprod.Fert.,58:521-531.
KennawayDJ,DunstanEA,StaplesLD(1987)Photoperiodiccontroloftheonsetofbreeding activity and fecundity in ewes. Journal of reproduction andfertilitySupplement34,187-199.
KennawayDJ,SanfordLM,GodfreyB,FriesenHG(1983)Patternsofprogesterone,melatonin and prolactin secretion in ewes maintained in four differentphotoperiods.TheJournalofendocrinology97,229-242.
Keyes P, Wiltbank C (1988) Endocrine regulation of the corpus luteum. AnnualReviewsPhysiology.50:465-482.
KhaldiG(1984)Variationssaisonnièresdel'activitéovarienne,ducomportementd'œstrusetdeladuréedeI'anœstruspost-partumdesfemellesovinesdelaraceBarbarine:influencesduniveaualimentaireetdelapresencedumale.TesisdeDoctorado,Univ.desSci.etTech.duLanguedoc,Montpellier,168pgs.
KnickerbockerJJ,WiltbankMC,NiswenderGD(1988)Mechanismsofluteolysisindomesticlivestock.Dom.Anim.Endocrinol.5:91-107.
Knights M, Maze TD, Bridges PJ, Lewis PE, Inskeep EK (2001) Short-termtreatmentwithacontrolledinternaldrugreleasing(CIDR)deviceandFSHto induce fertile estrus and increase prolificacy in anestrous ewes.Theriogenology55,1181-1191.
Koyuncu M, Altıncekic SO (2016) The Effects of Short-Medium and Long-TermApplicationsofFluorogestoneAcetate(FGA)onReproductivePerformanceofKıvırcıkEwesatTheOnsetofTheBreedingSeason.JAgrSci26,360-365.
LahozB, Alabart JL, Folch J, Calvo JH,Martínez-RoyoA, FantovaE (2010)Genesmayoresparaelincrementodelaprolificidad.Albéitar.136:20-21..
Legan SJ, Karsch FJ (1979) Neuroendocrine regulation of the estrous cycle andseasonalbreedingintheewe.Biologyofreproduction20,74-85.
Legan SJ, Karsch FJ (1980) Photoperiodic control of seasonal breeding in ewes:modulation of the negative feedback action of estradiol. Biology ofreproduction23,1061-1068.
LetelierCA,Contreras-Solis I,Garcia-FernandezRA,AriznavarretaC,TresguerresJA, Flores JM, Gonzalez-Bulnes A (2009) Ovarian follicular dynamics andplasmasteroidconcentrationsarenotsignificantlydifferent inewesgivenintravaginal sponges containing either 20 or 40 mg of fluorogestoneacetate.Theriogenology71,676-682.

Bibliografía
179
LetelierCA,Contreras-SolisI,Garcia-FernandezRA,SanchezMA,Garcia-PalenciaP,Sanchez B, Ariznavarreta C, Tresguerres JA, Flores JM, Gonzalez-Bulnes A(2011) Effects of oestrus induction with progestagens or prostaglandinanaloguesonovarianandpituitaryfunctioninsheep.Animalreproductionscience126,61-69.
Leyva V, Buckrell BC, Walton JS (1998) Regulation of follicular activity andovulationinewesbyexogenousprogestagen.Theriogenology50,395-416.
LópezSebastiánA(1989)Estacionalidaddelareproducción.OVISNúm1:59-73..Lopez-Sebastian A, Gonzalez de Bulnes A, Santiago Moreno J, Gomez-Brunet A,
TownsendEC,InskeepEK(1997)PatternsoffolliculardevelopmentduringtheestrouscycleinmonovularMerinodelParisewes.Animalreproductionscience48,279-291.
López-SebastiánA,Santiago-MorenoJ,González-BulnesA,GarcíaLópezM(1993)Aspectos característicos de la fisiología reproductiva de la oveja. Revistacientífica, Vol III. Núm 2. Instituto Nacional de Investigaciones Agrarias.Madrid,España.
LucyMC, Savio JD, Badinga L, De la Sota RL, ThatcherWW (1992) Factors thataffectovarianfolliculardynamicsincattle.J.Anim.Sci.70:3615-3626.
MacriA,PurificatoI,TollisM(2006)AvailabilityofveterinarymedicinalproductsforfoodproducingminoranimalspeciesintheMediterraneanarea.Annalidell'Istitutosuperioredisanita42,422-426.
MAGRAMA (2015) Requisitos y condicionantes de la producción ganadera.Disponible desde internet:http://www.mapama.gob.es/es/ganaderia/temas/requisitos-y-condicionantes-de-la-produccion-ganadera/ganaderia-y-medio-ambiente/[consulta:juliode2016].
MalpauxB,ThieryJC,ChemineauP(1999)Melatoninandtheseasonalcontrolofreproduction.ReprodNutrDev39,355-366.
Malpaux B, Viguie C, Skinner DC, Thiery JC, Chemineau P (1997) Control of thecircannualrhythmofreproductionbymelatoninintheewe.Brainresearchbulletin44,431-438.
Malpaux B, Viguie C, Skinner DC, Thiéry JC, Pelletier J, Chemineau P (1996)Seasonal breeding in sheep: Mechanism of action of melatonin. AnimalReproductionScience42,109-117.
ManesJ,CamperoC,HozborF,AlberioR,UngerfeldR(2015)Vaginalhistologicalchanges after using intravaginal sponges for oestrous synchronization inanoestrous ewes. Reproduction in domestic animals = Zuchthygiene 50,270-274.
Manes J, Fiorentino MA, Kaiser G, Hozbor F, Alberio R, Sanchez E, Paolicchi F(2010) Changes in the vaginal flora after treatment with differentintravaginaldevicesinewes.SmallRuminantResearch94,201-204.
Manes J, Hozbor F, Alberio R, Ungerfeld R (2014) Intravaginal placebo spongeaffect negatively the conception rate in sheep. Small Ruminant Research120,108-111.
Manes J, Rios G, Fiorentino MA, Ungerfeld R (2016) Vaginal mucus from ewestreated with progestogen sponges affects quality of ram spermatozoa.Theriogenology85,856-861.

Bibliografía
180
MartemucciG,D'AlessandroAG(2011)Synchronizationofoestrusandovulationby short time combined FGA, PGF(2alpha), GnRH, eCG treatments fornaturalserviceorAIfixed-time.Animalreproductionscience123,32-39.
MartinGB,KadokawaH(2006)"Clean,greenandethical"animalproduction.Casestudy: reproductive efficiency in small ruminants. J Reprod Dev 52, 145-152.
MartinGB,MiltonJT,DavidsonRH,BancheroHunzickerGE,LindsayDR,BlacheD(2004) Natural methods for increasing reproductive efficiency in smallruminants.Animalreproductionscience82-83,231-245.
Martin GB, Price CA, Thiery JC, Webb R (1988) Interactions between inhibin,oestradiol and progesterone in the control of gonadotrophin secretion intheewe.Journalofreproductionandfertility82,319-328.
Martinez-Tinajero JJ, Ruiz-Herluver I, Montanez-Valdez G, Martinez-Priego G,Velasco-Zebadua ME, Izaguirre F (2011) Reproductive performance inDorperewessynchronizedatestrusduringnon-breedingseasonintropicalconditions.JournalofAnimalandVeterinaryAdvances10,221-223.
McCracken JA,Carlson JC,GlewME,Goding JR,BairdDT,GreenK, SamuelssonB(1972)ProstaglandinF2 identified as a luteolytic hormone in sheep.NatNewBiol238,129-134.
McCrackenJA,GlewME,ScaramuzziRJ(1970)CorpusLuteumRegressionInducedby Prostaglandin F 2α. JOURNAL OF CLINICAL ENDOCRINOLOGY &METABOLISM30(4):544-6.
McNatty KP, Heath DA, Clark Z, Reader KL, Juengel JL, Pitman J (2016) Ovarianfollicular characteristics in sheep with multiple fecundity genes.Reproduction.
McNeillyAS (1984)Changes inFSHand thepulsatile secretionof LHduring thedelay inoestrus inducedbytreatmentofeweswithbovinefollicular fluid.Journalofreproductionandfertility72,165-172.
Menassol JB, Collet A, Chesneau D, Malpaux B, Scaramuzzi RJ (2012) Theinteractionbetweenphotoperiodandnutritionand itseffectsonseasonalrhythmsofreproductionintheewe.Biologyofreproduction86,52.
MenchacaA,VilarinoM,CrispoM,deCastroT,RubianesE(2010)Newapproachesto superovulation and embryo transfer in small ruminants. Reproduction,fertility,anddevelopment22,113-118.
MeuretM(2006)Lespratiquespastoralesentretempscourtdel'alimentationdestroupeaux et temps long des ressources et des milieux. In Actualité etmodernitédupastoralisme(Séance,CRAcadémied'agriculture).
MoeiniMM,Alipour F, SanjabiMR (2013)Efficacy of CIDRor FGA spongeswithhCGtreatmentson theconceptionrateandprolificacy inLoriewesoutofthebreedingseason.IranianJournalofAppliedAnimalScience3,521-525.
Moeini MM, Karami H, Mikaeili E (2009) Effect of selenium and vitamin Esupplementation during the late pregnancy on reproductive indices andmilkproductioninheifers.Animalreproductionscience114,109-114.
Mollard A, Chatelier, V., Codron, J.M., Dupraz, P., Jaccquet, F. (2002) Vers unegestion intégrée agriculture-environnement. Diagnostic, solutions,perspectives. In Agriculture, territoire, environnement dans les politiqueseuropéenes(ATEPE)(Dossierdel'INRA)..

Bibliografía
181
MongetP,MonniauxD(1995)Growthfactorsandthecontroloffolliculogenesis.JReprodFertil49(Suppl.),321-333.
Moradi Kor N, Khanghah KM, Veisi A (2012) Efficiency of short time protocolsbased on combined FGA, PGF2α, GnRH and eCG treatments on oestrussynchronizationandreproductiveperformanceofkermaniewesduringthebreedingseason.IntJBiolMedRes.;3(3):1966-1970.
Murray JF, Downing JA, Evans G, Findlay JK, Scaramuzzi RJ (1992) Epidermalgrowth factor acts directly on the ovary in vivo to inhibit oestradiol-17Pand inhibin secretion and enhance progesterone secretion. . Journal ofEndocrinology137,253-264.
NaderipourH,YadiJ,ShadAG,SirjaniMA(2012)TheeffectsofthreemethodsofsynchronizationonestrusinductionandhormonalprofileinKalkuhiewes:Acomparisonstudy.AfricanJournalofBiotechnology11,530-533.
Naim PC, M.; Gibbons, A. (2009) Timed artificial insemination with ram chilledsemen.ArchivosdeZootecnia,v.58,p.435-440.
NancarrowCD,Hill JL (1995)Oviduct proteins in fertilization and early embryodevelopment.JournalofreproductionandfertilitySupplement49,3-13.
NiswenderGD,JuengelJL,SilvaPJ,RollysonMK,McIntushEW(2000)Mechanismscontrollingthefunctionandlifespanofthecorpusluteum.PhysiolRev80,1-29.
NiswenderGD,SuterDE,SawyerHR(1981)FactorsregulatingreceptorsforLHonovinelutealcells.JournalofreproductionandfertilitySupplement30,183-189.
O'CallaghanD,DonovanA,SunderlandSJ,BolandMP,RocheJF(1994)Effectofthepresenceofmale and female flockmateson reproductive activity in ewes.Journalofreproductionandfertility100,497-503.
O'Mary CC, Pope AL, Casida LE (1950) The use of progesterone in thesynchronizationoftheestrualperiodsinagroupofewesandtheeffectontheirsubsequentlambingrecords.Journalofanimalscience9,499-503.
Omontese BO, Rekwot PI, Makun HJ, Obidi JA, Rwuaan JS, Chiezey NP (2010)Evaluation of EAZI-BreedTM CIDR and FGA-30 intravaginal sponges assynchronizingagentsinprepartumRedSokotodoes.JVetRes3,64-69.
Ortavant R, LoirM (1978) The environment as a factor in reproduction in farmanimals.4thWorldCong.ofAnimProd.,BuenosAires,20-26deabril,Vol.1,423-451.
Owen JB (1987) Group breeding schemes and simplified recording in sheepimprovement.En:Newtechniquesinsheepproduction.FayezI.;MaraiM.;Owen,JB.Ed.Butterworth&Co.(Publishers)Ltd.302pgs:157-162.
Ozyurtlu N, Ay Serhan S, Kucukaslan I, Gungor O, Aslan S (2011) Effect ofsubsequent two short-term, short-term, and long-term progestagentreatmentson fertilityofAwassiewesoutof thebreedingseason.AnkaraUniv.Vet.Fak.Derg.58,105-109.
Padmanabhan VK, F.J.; Lee, J.S. (2002) Hypothalamic, pituitary and gonadalregulationofFSH.Reproduction.59:67-82.
PardosL,FantovaE(2007)Importanciadelcontroldegestióntécnico-económicoen las ganaderías de ovino de carne. Influencia de los diferentes factoresproductivos en los resultados económicos. En: Producción de ovino decarneenmediosemiárido.EditaGobiernodeAragón.Pp.17-29.

Bibliografía
182
Parraguez VH, Blank O, Muñoz C, Latorre E (2000) Inseminación artificial enovinos. . Monografías de Medicina Veterinaria 20, Disponible en:<http://www.monografiasveterinaria.uchile.cl/index.php/MMV/article/view/5019/4903%3E.Fechadeacceso:5017dic.2016.
Peralta JGS, M.T.; García, E.O. (2007) Oestrus synchronization of ewes, usingnorgestomet combined with PGF2α and hCG in the reproductive season.ResearchJournalofAnimalSciences;1:44-48.
PerretG,MercadierM,BodinL(1995)OrganizationofsheepbreedinginFrance–suckling breeds-. III. The Lacaune meat programme. OptionsMéditerranéennes.11:49-59.
PhillipsDJ,deKretserDM(1998)Follistatin:amultifunctionalregulatoryprotein.Frontiersinneuroendocrinology19,287-322.
Picton HM, Tsonis CG, McNeilly AS (1990) FSH causes a time-dependentstimulation of preovulatory follicle growth in the absence of pulsatile LHsecretion in ewes chronically treated with gonadotrophin-releasinghormoneagonist.TheJournalofendocrinology126,297-307.
PietroskiACCA,BrandãoFZ,SouzaJMG,FonsecaJF(2013)Short,mediumorlong-term hormonal treatments for induction of synchronized estrus andovulationinSaanengoatsduringthenonbreedingseason.RevistaBrasileiradeZootecnia42,168-173.
Pineda MH (1989) Sistema reproductor de la hembra. En: EndocrinologíaVeterinaria y Reproducción. L.E. McDonald (Ed.). 4ta. Edición,InteramericanaMcGrowth-HillS.A.MéxicoD.F.,México.Pp.294-344.
Piper PJ, Vane JR,Wyllie JH (1970) Inactivation of prostaglandins by the lungs.Nature225,600-604.
PopeWF,CárdenasH(2004)SensitivityofsheeptoexogenousprostaglandinF2αearlyintheestrouscycle.SmallRuminantResearch55(1),245-248.
PouxX,BeaufoyG,BignalE,B.R(2009)Lessystèmesovinsetcaprinsdel'Unioneuropéenne:implicationspourl'environnement.OptionsMediterraneennesSerieA91,113-123.
Powell MR, Kaps M, Lamberson WR, Keisler DH (1996) Use of melengestrolacetate-based treatments to induce and synchronize estrus in seasonallyanestrousewes.Journalofanimalscience74,2292-2302.
PriceCA,WebbR(1989)OvarianresponsestohCGtreatmentduringtheestrouscycleinheifers.JReprodFertil86,303-308.
Rawlings NC, Cook SJ (1993) LH secretion around the time of the preovulatorygonadotrophinsurgeintheewe.AnimReprodSci30,289-299.
Reynolds JEF (1996) Martindale, the extra pharmacopoeia. (31st edn), RoyalPharmaceuticalSociety,London.RoyalPharmaceuticalSociety.
ReynoldsLP,Grazul-BilskaAT,KillileaSD,RedmerDA(1994)Mitogenicfactorsofcorporalutea.Progressingrowthfactorresearch5,159-175.
ReynoldsLP,RedmerDA(1999)Growthanddevelopmentof thecorpus luteum.JournalofreproductionandfertilitySupplement54,181-191.
RhodesL,NathanielPW(1988)Comparisonofacontrolled internaldrugreleasedevice containing progesterone with intravaginal medrox- yprogesteronespongesforestrussynchronizationinewes..Theriogenology30,831-836.
Robertson DM (1992) Follistatin/activin-binding protein. Trends EndocrinolMetab3,65-68.

Bibliografía
183
RobicZ,LikerB,RupicV(1992)DurationofanestrusinPramenkaandRomanovewesinYugoslavia.Journalofanimalscience70,13-17.
Robinson JE, Karsch FJ (1987) Photoperiodic history and a changing melatoninpattern can determine the neuroendocrine response of the ewe todaylength.Journalofreproductionandfertility80,159-165.
Robinson TJ (1965) Use of Progestagen-Impregnated Sponges InsertedIntravaginally or Subcutaneously for the Control of the Oestrous Cycle intheSheep.Nature206,39-41.
RomanoJE(2004)SynchronizationofestrususingCIDR,FGAorMAPintravaginalpessaries during the breeding season in Nubian goats. Small RuminantResearch55,15-19.
Romero-GonzálezLJ,Muñoz-HurtadoB,Fontaneda-LópezI,López-DíazMC(2008)LenguaAzul:SituacióndelaenfermedadenEspañayEuropa.SEOC.XXXIII:29-39.
RomeuA,MolinaI,TresguerresJA,PlaM,PeinadoJA(1995)Effectofrecombinanthumanluteinizinghormoneversushumanchorionicgonadotrophin:effectson ovulation, embryo quality and transport, steroid balance andimplantationinrabbits.Humanreproduction10,1290-1296.
RosaHJD,BryantMJ(2003)Seasonalityofreproductioninsheep.SmallRuminantResearch48,155-171.
RowlandMT,T.N.(1989)Clinicalpharmacokinetics:conceptsandapplication.LeaandFebiger.
Rubianes E,Menchaca A, Carbajal B (2003) Response of the 1-5 day-aged ovinecorpus luteum toprostaglandinF2alpha.Animal reproduction science78,47-55.
Sánchez-Rodríguez M (2011) Sector ovino lechero. In Máster oficial de laUniversidad de Córdoba en zootecnia y gestión sostenible: ganaderíaecológicaeintegrada.
SantamaríaC,GárrizI,RodríguezA,DíezdeUlzurrunC,OchoaJ(1998)Ovinodecarne:Resultadoseconómicosdediferentessistemasdeproducción.SEOC.XXIII:259-263..
SareminejadP,TabatabaeiS,MamoueiM,MirzadehK,BoujarpourM(2014)TheEffect of Short and Long Term Medroxy Progesterone Acetate (MAP)SpongeTreatmentonReproductivePerformanceduringtheNonBreedingSeasonofArabiaEwes. Iranian Journal ofAppliedAnimal Science4, 747-751.
SawyerGJ(1983)TheinfluenceofradiantheatloadonreproductionintheMerinoewe.LesColloquesdel'INRA,N°20,Ed.INRAPubl.,Paris,225-235..
Scaramuzzi RJ, Adams NR, Baird DT, Campbell BK, Downing JA, Findlay JK,HendersonKM,MartinGB,McNattyKP,McNeillyAS,etal.(1993)Amodelfor follicle selection and the determination of ovulation rate in the ewe.Reproduction,fertility,anddevelopment5,459-478.
SchrickFN,SurfaceRA,PritchardJY,DaileyRA,TownsendEC,InskeepEK(1993)Ovarian structures during the estrous cycle and early pregnancy in ewes.Biologyofreproduction49,1133-1140.
SeeburgPH,MasonAJ,StewartTA,NikolicsK(1987)ThemammalianGnRHgeneanditspivotalroleinreproduction.RecentProg.Horm.Res.43:69-98.

Bibliografía
184
Senger PL (2003) Reproductive cyclicity. Terminology and basic concepts. En:PullmanW.A.(ed)PathwaystoPregnancyandParturition,2aEd:144-163.CurrentConceptions,Inc.
SengerPL(2005)Pathwaystopregnancyandparturation.(2ndrevisededn).Shahneh AZ, Sadeghipanah H, Javaheri H, Emami-mibody MA (2008) Effects of
equine chorionic gonadotropin (eCG) administration and flushing onreproductive performance in Nadooshan goats of Iran. African Journal ofBiotechnology7,3373-3379.
SierraI(1986)LarazasintéticaSalz.MonografíaONE.Ovino.2ªedición.53-60..SierraI(1989)Cruzamientodelaespecieovina.Ovis.4:47-73.Smith MF, McIntush EW, Smith GW (1994) Mechanisms associated with corpus
luteumdevelopment.Journalofanimalscience72,1857-1872.Sosa C, Gonzalez-Bulnes A, Abecia JA, Forcada F, Meikle A (2010) Short-term
undernutrition affects final development of ovulatory follicles in sheepsynchronized for ovulation. Reproduction in domestic animals =Zuchthygiene45,1033-1038.
SouthcottWH,BradenAW,MouleGR(1962)Synchronizationofoestrusinsheepbyorallyactiveprogesteronederivative.AustralianJournalofAgriculturalResearch13,901-906.
SouzaCJ,CampbellBK,BairdDT (1997)Folliculardynamicsandovarian steroidsecretion in sheep during the follicular and early luteal phases of theestrouscycle.Biologyofreproduction56,483-488.
Staples LD, McPhee S, Reeve J, Williams AH (1992) Practical applications forcontrolled release melatonin implants in sheep. En Advances in pinealresearch,Vol.6A,A.Foldes,R.I.Reiter,eds.,JohnLibbey,Londres.
Stellflug JN, Weems YS, Weems CW (1997) Clinical reproductive physiology ofewes. En: Current therapy in large animal theriogenology, Youngquist, RS(Ed)WBSaundersCoPhiladelphia,PennsylvaniaPp594-598.
Suárez IM (2010) Comparación de la efectividad de tres protocolos desincronización de celo en ovejas de raza merina [Tesis]. Córdoba:UniversidaddeCórdoba.FacultaddeVeterinaria.
Swelum AA, Alowaimer AN, Abouheif MA (2015) Use of fluorogestone acetatesponges or controlled internal drug release for estrus synchronization inewes: Effects of hormonal profiles and reproductive performance.Theriogenology84,498-503.
Thatcher WW, Meyer MD, Danet-Desnoyers G (1995) Maternal recognition ofpregnancy.JReprodFertilSuppl.49:15-28.
Thimonier J (1981)Controlof seasonal reproduction in sheepandgoatsby lightandhormones.JournalofreproductionandfertilitySupplement30,33-45.
ThimonierJ,MauleonP(1969)Variationssaisonnièresducomportementd'œstrusetdes activitésovarienneethypophysaire chez lesovins.Ann.Biol.AnimBiochim.Biophys.,9:233-250.
TitiHH,KridliRT,AlnimerMA(2010)Estrussynchronizationinsheepandgoatsusing combinations of GnRH, progestagen and prostaglandin F2alpha.Reproductionindomesticanimals=Zuchthygiene45,594-599.
TondelloLDS,P.C.;Gaudêncio,S.;Pereira,L.;Bertolini,M.;Diniz,A.;Mezzalira,A.(2010)Microbiological and functional evaluation of an alternative device(OB(R))forestroussynchronizationinewes.CiênciaRural;40(2):389-395.

Bibliografía
185
UngerfeldR,RubianesE(1999)Effectivenessofshort-termprogestagenprimingsfor the induction of fertile estrus with eCG in ewes during late seasonalanestrus.AnimalScience68,349-353.
UngerfeldR, Rubianes E (2002) Short termprimingswith different progestogenintravaginal devices (MAP , FGA and CIDR) for eCG-estrous induction inanestrusewes.SmallRumin.Res.46:63-66.
Ustuner B, Gunay U, Nur Z, Ustuner H (2007) Effects of long and short-termprogestagen treatments combinedwith PMSG on oestrus synchronizationandfertilityinAwassiewesduringthebreedingseason.ActaVetBrno76,391-397.
Veiga-Lopez A, Beckett EM, Abi Salloum B, Ye W, Padmanabhan V (2014)Developmental programming: prenatal BPA treatment disrupts timing ofLHsurgeandovarian follicularwavedynamics inadult sheep.Toxicologyandappliedpharmacology279,119-128.
VelleW,HelleO(1979)Experienceswithestrussynchronization insheepoveratwelve-yearperiodusingoralMAPtreatmentfortendays.Journalofanimalscience48,1015-1019.
VilarinoM,RubianesE,MenchacaA(2013)Ovarianresponsesandpregnancyratewithpreviouslyusedintravaginalprogesteronereleasingdevicesforfixed-timeartificialinseminationinsheep.Theriogenology79,206-210.
Vinoles C, Forsberg M, Banchero G, Rubianes E (2001) Effect of long-term andshort-termprogestagentreatmentonfolliculardevelopmentandpregnancyrateincyclicewes.Theriogenology55,993-1004.
VinolesC, ForsbergM,MartinGB,CajarvilleC,Repetto J,MeikleA (2005)Short-term nutritional supplementation of ewes in low body condition affectsfollicledevelopmentduetoanincreaseinglucoseandmetabolichormones.Reproduction129,299-309.
VinolesC,MeikleA,ForsbergM,RubianesE(1999)Theeffectofsubluteallevelsofexogenous progesterone on follicular dynamics and endocrine patternsduringearlylutealphaseoftheewe.Theriogenology51,1351-1361.
Wathes DC, Lamming GE (1995) The oxytocin receptor, luteolysis and themaintenanceofpregnancy.JournalofreproductionandfertilitySupplement49,53-67.
Watson AJ, Watson PH, Arcellana-Panlilio M, Warnes D, Walker SK, Schultz GA,Armstrong DT, Seamark RF (1994) A growth factor phenotype map forovinepreimplantationdevelopment.BiologyReproduction50,725-733.
Webb R, Engeland BG (1982) Identification of the ovulatory follicle in the ewe:associated changes in the follicular size, thecal and granulosa cellluteinizing hormone receptors, antral fluid steroids, and circulatinghormonesduringthepreovulatoryperiod.Endocrinol.110:873-881.
WebbR,GosdenRG,TelferEE,MoorRM(1999)Factorsaffectingfolliculogenesisinruminants.AnimSci68,257-284.
Welch RAS, Andrewes WD, Barnes DR, Bremner K, Harvey TG (1984) CIDRdispensers for oestrus and ovulation control in sheep. Proceedings of the10th International Congress on Animal Reproduction & ArtificialInsemination,vol3,Urbana,IL,USA,pp354-355.
Wheaton JE, Carlson KM, Windels HF, Johnston LJ (1993) CIDR: A newprogesterone-releasingintravaginaldeviceforinductionofestrusandcyclecontrolinsheepandgoats.Animalreproductionscience33(1-4),127-141.

Bibliografía
186
Whitley NC, Jackson DJ (2004) An update on estrus synchronization in goats: aminorspecies.Journalofanimalscience82E-Suppl,E270-276.
WindsorDP(1995)Factors influencing thesuccessof transcervical inseminationinMerinoewes.Theriogenology43,1009-1018.
Yadi J, Moghaddam MF, Khalajzadeh S (2011) Comparison of EstrusSynchronizationbyPGF2α,CIDRandSpongewithPMSGinKalkuhiEwesonEarlyAnoestrousSeason,InternationalConferenceonAsiaAgricultureandAnimal,IPCBEE,vol.13,pp.61-65.
YingSY(1988)Inhibins,activins,andfollistatins:gonadalproteinsmodulatingthesecretionoffollicle-stimulatinghormone-Review.Endocr.Rev.9:267-293.
Zhang S, Blache D, Blackberry MA, Martin GB (2005) Body reserves affect thereproductiveendocrineresponsestoanacutechangeinnutritioninmaturemalesheep.Animalreproductionscience88,257-269.
ZonturluAK,AralF,OzyurtluN,YavuzerU(2008)SynchronizationofestrususingFGAandCIDRintravaginalpessariesduringthetransitionperiodinAwassiewes.JournalofAnimalandVeterinaryAdvances7,1093-1096.





























