Embed Size (px)
Citation preview

Addition of the Antioxidant Probucol to Angiotensin IIType I Receptor Antagonist Arrests ProgressiveMesangioproliferative Glomerulonephritis in the Rat
Shuji Kondo,* Maki Shimizu,* Maki Urushihara,* Koichiro Tsuchiya,†
Masanori Yoshizumi,† Toshiaki Tamaki,† Akira Nishiyama,‡ Hiroshi Kawachi,§
Fujio Shimizu,§ Mark T. Quinn,� David J. Lambeth,¶ and Shoji Kagami*Departments of *Pediatrics and †Pharmacology, The Institute of Health Bioscience, The University of TokushimaGraduate School, Tokushima, Japan; ‡Department of Pharmacology, Kagawa Medical University, Kagawa, Japan;§Department of Cell Biology, Institute of Nephrology, Niigata University School of Medicine, Niigata, Japan;�Department of Veterinary Molecular Biology, Montana State University, Bozeman, Montana; and ¶Department ofPathology and Laboratory Medicine, Emory University, Atlanta, Georgia
Angiotensin II (Ang II) and reactive oxidative species (ROS) that are produced by NADPH oxidase have been implicated inthe progression of glomerulonephritis (GN). This study examined the effect of simultaneously interrupting Ang II and ROSwith an Ang II receptor blocker (ARB), candesartan, and a free radical scavenger, probucol, in a model of progressivemesangioproliferative GN induced by the injection of anti–Thy-1 antibody into uninephrectomized rats. Nephritic rats weredivided into four groups and given daily oral doses of the following: Vehicle, 1% probucol diet, 70 mg/L candesartan indrinking water, and probucol plus candesartan. These treatments lasted until day 56. Vehicle-treated nephritic rats developedprogressively elevated proteinuria and glomerulosclerosis. Candesartan kept proteinuria significantly lower than vehicle orprobucol. The addition of probucol to candesartan normalized urinary protein excretion. Increases in BP in nephritic rats werelowered by these treatments, except with probucol. It is interesting that both glomerular cell number and glomerulosclerosiswere significantly decreased by candesartan and normalized by the addition of probucol. Immunohistochemical studies forTGF-�1, collagen type I, and fibronectin revealed that the combined treatment abolished glomerular fibrotic findingscompared with candesartan. In addition, glomerular expression of NADPH oxidase components and superoxide productionsuggested that the combined treatment completely eliminated NADPH oxidase–associated ROS production. In conclusion, ourstudy provides the first evidence that the antioxidant probucol, when added to an Ang II receptor blockade, fully arrestsproteinuria and disease progression in GN. Furthermore, the data suggest that NADPH oxidase–associated ROS productionmay play a pivotal role in the progression of GN. The combination of probucol and candesartan may represent a novel routeof therapy for patients with progressive GN.
J Am Soc Nephrol 17: 783–794, 2006. doi: 10.1681/ASN.2005050519
T he marked accumulation of extracellular matrix (ECM),including collagens and fibronectin, in glomeruli is ahallmark in the development of human and experimen-
tal progressive glomerulonephritis (GN) (1–4). TGF-�1, a primefibrogenic factor, stimulates the deposition of ECM by mesan-gial cells and thereby may induce glomerulosclerosis (2,5).
Recently, the interventions with angiotensin II (Ang II) typeI receptor blocker (ARB) and/or angiotensin-converting en-zyme inhibitor (ACEI) have shown their protective potentialagainst human nephropathies, including IgA nephropathy anddiabetic nephropathy, and progressive experimental models
such as mesangioproliferative GN and nephrectomized rats(6–10). These clinical and experimental in vivo studies havestrongly suggested that the renoprotective effect of these anti-hypertensive agents results from not simply the inhibition of itshemodynamic action but also blockade of its hemodynamic-independent actions. In fact, many studies, including our pre-vious in vitro studies, have disclosed the mechanisms and therole of Ang II–induced ECM deposition through mesangialcells (11–13). Ang II stimulates ECM synthesis through theinduction of TGF-�1 expression in cultured rat glomerularmesangial cells (11) and affects the plasminogen activator/plasminogen activator inhibitor-1 system to result in acceler-ated deposition of ECM (12), which constitutes evidence thatAng II can act as a potent fibrotic molecule independent of itseffects on BP. Although the current therapy by inhibition ofAng II retards renal fibrosis, it is not enough to prevent com-pletely the progression of GN. This suggests that other fibro-genic factors such as PDGF-BB, TNF-�, and IL-1� might also be
Received May 20, 2005. Accepted December 29, 2005.
Published online ahead of print. Publication date available at www.jasn.org.
Address correspondence to: Dr. Shoji Kagami, Department of Pediatrics, TheInstitute of Health Bioscience, The University of Tokushima Graduate School,Kuramoto-cho-3-chome, Tokushima 770-8503, Japan. Phone: �81-88-633-7135;Fax: �81-886-31-8697; E-mail: [email protected]
Copyright © 2006 by the American Society of Nephrology ISSN: 1046-6673/1703-0783

involved in the progression of GN (3,14). For maximizing thetherapeutic potential of antifibrotic treatment, additional ther-apy that is aimed at suppression of these fibrogenic factors andtheir related signaling pathways may be required.
Recently, reactive oxygen species (ROS) that are produced byNADPH oxidase has been implicated in the development andprogression of GN (15–17). The administration of various nat-ural or synthetic antioxidants has been shown to be beneficialin the prevention and attenuation of renal scarring in numerousanimal models of renal disease (18). These compounds includevitamins, N-acetylcysteine, �-lipoic acid, lipid-lowering agents,and many others. Probucol is a lipid-soluble, cholesterol-low-ering drug with potent antioxidant properties that has beenshown to scavenge lipid peroxidation (19). This agent seems toplay a role in eliminating ROS produced by NADPH oxidase(20).
It is interesting that Ang II is considered a representativestimulator of NADPH oxidase activity that leads to the en-hanced production of superoxide (O2
·�) in several cell lines,including mesangial cells and vascular smooth muscle cells.These studies have also demonstrated that NADPH oxidasesare composed of gp91phox-homolog and produce ROS includ-ing O2
·� not only in phagocytes but also in nonphagocytic cells(21–25). In addition, these studies indicated that NADPH oxi-dase plays an important role as a cell-signaling molecule inaddition to being a classical mediator of tissue injury (21,22).Whereas gp91phox, which has been called Nox2 (for NADPHoxidase), is generally expressed in inflammatory cells such asleukocytes, the Nox family is also composed of various mem-bers from Nox1 to Nox5, which are expressed in noninflamma-tory cells, including mesangial cells and podocytes. In particu-lar, Nox1, Nox2, and Nox4 have been investigated in AngII–induced ROS production in renal and cardiovascular dis-eases (21,22,25). Besides, the levels of these Nox proteins areassociated with the activity and O2
·� production (21,22). Otherfibrogenic factors, such as PDGF-BB, TNF-�, and IL-1�, all ofwhich are involved in the progression of GN, also stimulateROS production in mesangial cells, although the detailed mech-anism of their production remain unknown (15,23,24). There-fore, we hypothesized that the blockade of both the action ofAng II and ROS production associated with NADPH oxidaseactivation might be a novel strategy for preventing the progres-sion of glomerulosclerosis. We examined whether simulta-neously interrupting Ang II and ROS with the ARB candesartanand probucol could reduce proteinuria and disease progressionin a rat model of irreversible mesangioproliferative GN.
Materials and MethodsAntibodies and Reagents
Previously characterized mouse mAb that specifically recognizedNox2 (clone 54.1) and rabbit polyclonal antibody against Nox4 wereused in this study (26,27). Horseradish peroxidase–conjugated goatanti-mouse IgG (H�L; Bio-Rad, Hercules, CA) and horse anti-rabbitIgG (H�L; Cell Signaling Technology Inc., Beverly, MA) were used forWestern blot analysis. For immunohistochemistry, previously charac-terized rabbit polyclonal anti–TGF-�1 antibody was used, and rabbitantibodies against collagen type I (COL I) and fibronectin (FN) wereobtained from Chemicon International Inc. (Temecula, CA) (8).
Experimental DesignAll experimental procedures were performed according to the guide-
lines for the care and use of laboratory animals established by NationalInstitutes of Health and the Institute for Animal Experimentation, theUniversity of Tokushima Graduate School. A progressive model ofmesangioproliferative GN was induced as described previously, withsome modifications (8). Briefly, rats were uninephrectomized and 1 wklater received an intravenous injection of 1.0 ml of PBS that contained2 mg of the nephritogenic anti–Thy-1 mAb 1-22-3. Nephritic rats weredivided into four groups (n � 6) and given daily oral doses as follows:(1) Vehicle, (2) 1% probucol diet, (3) 70 mg/L candesartan in drinkingwater, or (4) probucol plus candesartan. The dose concentration ofprobucol in the rat diet and candesartan in drinking water were se-lected on the basis of previous reports (28–31). The therapeutic regimenthat was used in our study was pharmacologically valid and efficientaccording to the manufacturer’s information for probucol. For achiev-ing a therapeutically adequate concentration of probucol, this reagentwas given 7 d before the injection of mAb 1-22-3. The dose of cande-sartan (70 mg/L) that was used in this experiment was high enough (10mg/kg per d) to inhibit Ang II receptor binding to kidney tissue (28,29).Candesartan was started on the same day as disease induction (day 0).Six rats in each group were killed at 7 d and 56 d after the injection ofmAb 1-22-3. As controls, six rats received injections of PBS 1 wk afterunilateral nephrectomy and were killed on days 7 and 56.
Urinary Protein Excretion, Blood Parameters, and BPUrine was periodically collected from the rats in a metabolic cage
during the course of GN. The amount of urinary protein excretion wasmeasured by the Bradford method (Bio-Rad, Oakland, CA). Theamount of food consumed and water intake were measured and cal-culated on day 56. At the end of the study period, serum concentrationsof blood urea nitrogen, creatinine, total cholesterol, and nonesterifiedfatty acid were measured using the reagents that were manufacturedby WAKO Chemical Industries (Osaka, Japan). Serum total protein wasmeasured by a BCA protein assay (Pierce, Rockford, IL). The serumlevel of triglyceride was assessed with a Lipidos Kit (TOYOBO, Osaka,Japan). Systolic BP (SBP) was measured noninvasively by the tail-cuffmethod (32). Concentrations of serum probucol were measured byHPLC at the laboratories of Otsuka Pharmaceutical Co. Ltd. (To-kushima, Japan).
Histology and ImmunohistochemistryThe right kidney of each rat was removed immediately, fixed in 10%
buffered formalin and embedded in paraffin, and 4-�m sections werestained with periodic acid-Schiff reagent. Glomerular cell number wasassessed on the basis of total glomerular cell count per glomerularcross-section. It was counted in 30 glomeruli per kidney in a periodicacid-Schiff–stained section, and the mean number of cells per glomer-ulus was calculated. A pathologist who was blinded to other findingssemiquantitatively analyzed the glomerulosclerosis score. The percent-age of each glomerulus that was occupied by mesangial matrix wasestimated and assigned a code as follows: 0 � absent, 0.5 � 1 to 5%; 1 �
5 to 25%; 2 � 25 to 50%; 3 � 50 to 75%; or 4 � �75%.Frozen sections (3 �m) were incubated with anti–COL I antibodies,
anti-FN antibodies, and anti–TGF-�1 Ab for 1 h and then with FITC-conjugated secondary antibodies. To evaluate the level of glomerularstaining with each antibody, we performed semiquantitative analysisas follows: 0 � diffuse, very weak, or absent mesangial staining; 1� �
1 to 25% of focally increased mesangial staining; 2� � 25 to 50% ofglomerular tuft demonstrating strong mesangial staining; 3� � 50 to75% of glomerular tuft demonstrating strong mesangial staining; and
784 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 783–794, 2006

4� � �75% of glomerular tuft stained strongly. For each kidneysection, 30 glomeruli were selected at random and evaluated by thesame blinded pathologist; the mean value per section was calculated(33).
Formalin-fixed tissue sections (3 �m) were deparaffinized with xy-lene and rehydrated with a graded series of ethanol. Endogenousperoxidase was blocked with hydrogen peroxide, and the samples wereheated at 121°C for 15 min in 0.01 mol/L citrate buffer (pH 6.0). Thesections were incubated with anti–ED-1 mAb (Serotec, Kinglington,UK) diluted in PBS that contained 1% BSA at 4°C for 24 h. After beingwashed with PBS, the sections were incubated with biotinylated sec-ondary antibody, avidin-biotin-peroxidase complex (ABC Elite; VectorLaboratories, Burlingame, CA), and then 3,3�-diaminobenzidine(Dojindo, Kumamoto, Japan). Each section was counterstained withMayer’s hematoxylin (Wako, Tokyo, Japan), dehydrated, and cover-slipped. The number of ED-1–positive cells was counted and calculatedin 30 full-size glomeruli (34).
Western BlottingWestern blotting was performed as described previously (35). Total
protein was extracted from glomeruli that were isolated by the sievingmethod (11). Protein samples (30 �g) were separated by 12.5% SDS-PAGE and transferred to nitrocellulose membranes. The membraneswere probed with mouse monoclonal anti-Nox2 antibodies, rabbitpolyclonal anti-Nox4 antibodies, and mouse monoclonal anti–�-actinantibodies (Sigma Chemical Co., St. Louis, MO) as a standard and thenincubated with an horseradish peroxidase–conjugated secondary anti-bodies. Immunoreactive proteins were detected with an enhancedchemiluminescence detection system (Amersham Corp., ArlingtonHeights, IL). Densitometric analysis was performed using an LKB
UltroScan XL apparatus (Pharmacia LKB Biotechnology, Uppsala, Swe-den).
In Situ Superoxide ProductionAs described elsewhere, in situ production of O2
·� was determinedusing 30-�m sections of frozen tissue that were incubated withdihydroethidium (DHE; 10 �mol/L) in PBS for 30 min at 37°C in ahumidified chamber that was protected from light. DHE is oxidizedupon reacting with O2
·� to ethidium bromide, which binds to DNAin the nucleus and fluoresces red. To detect ethidium bromide, weused a 543-nm He-Ne laser combined with a 560-nm long-pass filter.Glomerular O2
·� content was estimated by analyzing the meanfluorescence intensity in glomeruli using National Institutes ofHealth imaging software (27). To identify O2
·�-producing cells inglomeruli, we performed immunofluorescent staining combinedwith DHE assay (red) and FITC labeling (green) using anti–ED-1mAb and anti–Thy-1 mAb as markers of macrophages and mesan-gial cells, respectively.
Statistical AnalysesThe values are expressed as mean � SEM. The differences were
evaluated with the Stat Mate III software package (ATMS Co., Ltd.,Tokyo, Japan). For urinary protein excretion, blood parameters, and BP,statistical significance was evaluated using a t test (8,33). The data forhistology and immunohistochemistry were analyzed using the non-parametric Kruskal-Wallis test for multiple comparisons (8). The sta-tistical significance level was defined as P � 0.05.
Table 1. Total food intake, total water intake, body weight, and the level of serum probucol in rats that had GNon day 56 and had been treated with probucol and/or ARB (candesartan)a
Control GN GN � Probucol GN � ARB GN � Probucol � ARB
Total food intake (g/d) 22.7 � 0.8 21.2 � 1.0 23.8 � 1.4 23.3 � 2.9 22.8 � 1.9Total water intake (ml/d) 37.8 � 2.8 41.7 � 4.0 39.2 � 3.7 35.8 � 3.0 41.5 � 3.0Body weight (g) 449 � 6 442 � 12 429 � 14 420 � 22 433 � 12Serum probucol (�g/ml) ND ND 11.2 � 3.8 ND 4.5 � 0.9aThe values are expressed as mean � SEM (n � 6). ARB, angiotensin II receptor blocker; GN, glomerulonephritis; ND, not
detected, indicating the level of serum probucol is below the measurement limit.
Table 2. Time course of urinary protein excretion (mg/d) in rats that had GN on day 56 and had been treatedwith probucol and/or ARB (candesartan)a
Day 0 Day 7 Day 14 Day 28 Day 56
Control 20 � 1.1 23 � 2.4 18 � 2.6 23 � 2.4 16 � 1.7GN 20 � 2.1 74 � 12.9b 78 � 17.1b 170 � 64.5c 273 � 100.6c
GN � probucol 19 � 2.1 90 � 5.6b 88 � 20.6b 117 � 17.3b 140 � 33.6b
GN � ARB 20 � 3.5 88 � 6.3b 37 � 2.5b,d,e 33 � 5.3f 23 � 5.0d,f
GN � probucol � ARB 20 � 2.8 56 � 8.0b,f,g 33 � 1.7b,d,e 22 � 2.5d,f 10 � 2.6d,f,h
aThe values are expressed as mean � SEM (n � 6).bP � 0.01 versus control.cP � 0.05 versus control.dP � 0.05 versus GN.eP � 0.05 versus GN � probucol.fP � 0.01 versus GN � probucol.
J Am Soc Nephrol 17: 783–794, 2006 ARB � Probucol Arrests Progressive Mesangioproliferative GN 785

ResultsEffect of Probucol and/or ARB on Urinary ProteinExcretion, BP, and Blood Parameters
As shown in Table 1, total food intake, total water intake, andbody weight were comparable among all groups. The serumlevel of probucol was 11.2 � 3.8 �g/ml in probucol-treatednephritic rats, 4.5 � 0.9 �g/ml in probucol plus ARB-treatednephritic rats, and below the measurement limit in rats that
were not treated with probucol, indicating that the serum levelof probucol was significantly elevated by a 1% probucol diet.
There was no difference in 24-h urinary protein excretionamong all groups before the induction of GN. As shown inTable 2, proteinuria was significantly increased in all nephriticrats on day 7 after disease induction. In vehicle-treated ne-phritic rats, proteinuria gradually increased until the end of theexperiment. Treatment with ARB or probucol plus ARB re-duced proteinuria compared with vehicle treatment from 2 wkafter the induction of nephritis. Conversely, treatment withprobucol did not significantly reduce proteinuria. Treatmentwith probucol plus ARB significantly prevented proteinuriacompared with treatment with probucol or ARB on days 7 and 56.
SBP of vehicle-treated nephritic rats on day 56 was elevated,although not significantly. Treatment with ARB or probucolplus ARB significantly reduced SBP compared with vehicletreatment. Conversely, treatment with probucol did not lowerSBP (Figure 1).
Blood parameters on day 56 are shown in Table 3. Remark-able hypoproteinemia was not detected in any of the groups. Asignificant elevation of serum total protein was observed inprobucol plus ARB-treated nephritic rats. The level of bloodurea nitrogen in vehicle-treated nephritic rats was elevated,whereas the level in probucol plus ARB-treated nephritic ratswas significantly lowered to the level in the control. The serumlevel of creatinine in probucol plus ARB-treated nephritic ratswas slightly lower than that in vehicle-treated nephritic rats,although the difference was not statistically significant. Hyper-lipidemia was observed in vehicle-treated nephritic rats, asshown in Table 3. A significant improvement of hyperlipidemiawas not observed in rats with probucol or ARB treatment.Combination treatment with probucol and ARB significantlydecreased the levels of triglyceride and nonesterified fatty acid,respectively.
Figure 1. Systolic BP in rats that had glomerulonephritis (GN)on day 56 and had been treated with probucol and/or angio-tensin II receptor blocker (ARB; candesartan). �, nephrecto-mized control rats; f, vehicle-treated GN rats; o, probucol-treated GN rats; u, ARB-treated GN rats; z, probucol � ARB–treated GN rats. Data are mean � SEM. *P � 0.01 versus control;†P � 0.01 versus GN; ‡P � 0.01 versus GN�probucol.
Table 3. Serum levels of total protein, BUN, creatinine, T-CHO, TG, and NEFA in rats that had GN on day 56 andhad been treated with probucol and/or ARB (candesartan)a
Control GN GN �probucol GN � ARB GN � probucol
� ARB
Total protein (g/dl) 6.3 � 0.1 6.1 � 0.2 6.0 � 0.2 6.0 � 0.2 6.6 � 0.1b,c
BUN (mg/dl) 21 � 1.6 32 � 3.3d 26 � 2.3 23 � 2.4 22 � 1.2e
Creatinine (mg/dl) 0.88 � 0.04 1.16 � 0.15 0.97 � 0.04 0.89 � 0.03 0.87 � 0.02f
T-CHO (mg/dl) 71.0 � 3.3 171.3 � 42.0d 124.5 � 18.6d 98.1 � 3.5 82.7 � 3.5TG (mEq/L) 147.2 � 34.5 281.3 � 47.7d 283.6 � 64.5 216.9 � 39.2 145.8 � 20.2e
NEFA (�g/ml) 0.35 � 0.01 0.63 � 0.08g 0.63 � 0.04g 0.57 � 0.08 0.35 � 0.03f,h,i
aThe values are expressed as mean � SEM (n � 6). BUN, blood urea nitrogen; T-CHO, total cholesterol; TG, triglyceride;NEFA, nonesterified fatty acid.
bP � 0.05 versus GN � probucol.cP � 0.05 versus GN � ARB.dP � 0.05 versus control.eP � 0.05 versus GN.fP � 0.01 versus GN � probucol.gP � 0.01 versus control.hP � 0.01 versus GN.iP � 0.01 versus GN � ARB.
786 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 783–794, 2006

Figure 2. Light micrographs showing the effects of probucol and/or ARB (candesartan) on glomerular structural changes in ratswith GN on day 56. (A) Control. (B) Vehicle-treated GN. (C) Probucol-treated GN. (D) ARB-treated GN. (E) Probucol �ARB–treated GN. Periodic acid-Schiff–stained sections. Magnification, �200.
Figure 3. The glomerular cell number (A and D), the glomerulosclerosis score (B and E), and the number of ED-1–positivemacrophages (C and F) in rats that had GN on days 7 (A through C) and 56 (D through F) and had been treated with probucoland/or ARB (candesartan). �, nephrectomized control rats; f, vehicle-treated GN rats; o, probucol-treated GN rats; u,ARB-treated GN rats; z, probucol � ARB–treated GN rats. Data are mean � SEM. *P � 0.01 versus control; †P � 0.01 versus GN;‡P � 0.01 versus GN�ARB.
J Am Soc Nephrol 17: 783–794, 2006 ARB � Probucol Arrests Progressive Mesangioproliferative GN 787

Renal HistologyThe effects of treatment with probucol and/or ARB on renal
histology were examined on days 7 and 56 after disease induc-tion. None of the treatments significantly affected morphologicchanges such as mesangial cell proliferation accompanied bymild mesangial matrix accumulation and segmental mesangio-lytic lesions seen on day 7 in vehicle-treated nephritic rats (datanot shown). Representative light micrographs of glomerularlesions on day 56 after each treatment are shown in Figure 2.The most striking glomerular changes that were observed invehicle-treated nephritic rats were severe mesangial prolifera-tion and marked ECM accumulation, which were accompanied
by adhesion to Bowman’s capsule, glomerulosclerosis, and tu-bulointerstitial fibrosis (Figure 2B). Treatment with ARB im-proved these pathologic findings (Figure 2D). Furthermore,glomerular injury could not be detected in probucol plus ARB-treated nephritic rats (Figure 2E).
To examine the effects of each treatment on nephritic glomer-uli, we performed histologic and histochemical analyses ondays 7 and 56 of the study period (Figure 3). On day 7, theglomerular cell number and the glomerulosclerosis score weresignificantly increased to similar levels in all nephritic rats thatreceived each treatment (Figure 3, A and B). Conversely, theincreased number of ED-1–positive macrophages seen in day 7
Figure 4. Representative immunohistochemical micrographs showing the effects of probucol and/or ARB (candesartan) on theexpression of TGF-�1, collagen type I (COL I), and fibronectin (FN) in rats with GN on day 56. Magnification, �200.
Figure 5. Semiquantitative assessment of the expression of TGF-�1, COL I, and FN in rats that had GN on days 7 (A) and 56 (B)and had been treated with probucol and/or ARB. �, nephrectomized control rats; f, vehicle-treated GN rats; o, probucol-treatedGN rats; u, ARB-treated GN rats; z, probucol � ARB–treated GN rats. Data are mean � SEM. *P � 0.01 versus control; #P � 0.05versus control; †P � 0.01 versus GN; ‡P � 0.01 versus GN�ARB; §P � 0.05 versus GN�ARB.
788 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 783–794, 2006

vehicle-treated nephritic rats was significantly reduced in ratsthat were given probucol and/or ARB (Figure 3C). On day 56,a striking increase in both glomerular cell number and theglomerulosclerosis score was observed in vehicle-treated ne-phritic rats (Figure 3, D and E). Treatment with ARB but notprobucol led to a considerable reduction in both parameters.Furthermore, a remarkable reduction of these parameters to thecontrol level was achieved in probucol plus ARB-treated ne-phritic rats. There were no significant changes in the number ofED-1–positive macrophages in any of the groups (Figure 3F).
Renal Expression of TGF-�1 and ECM ProteinsRepresentative immunostaining micrographs on day 56 after
disease induction showed that a marked increases in the ex-pression of TGF-�1, COL I, and FN were observed in themesangial area in �90% of the glomeruli from vehicle-treatednephritic rats (Figure 4). Treatment with ARB significantlyreduced glomerular immunofluorescence staining for TGF-�1,COL I, and FN, whereas treatment with probucol did not. Theadministration of probucol plus ARB improved staining to thecontrol level. Glomerular staining scores for TGF-�1, COL I,and FN are shown in Figure 5. A semiquantitative analysis on day
7 revealed that the glomerular expression of COL I and FN but notTGF-�1 was upregulated in all nephritic rats regardless of treat-ment (Figure 5A). Conversely, on day 56, the levels of immuno-fluorescence for these factors indicated that combined treatmentwith probucol plus ARB normalized immunofluorescence inten-sity for the expression of TGF-�1 and ECM proteins to the controllevel, in parallel with histologic improvement (Figure 5B).
Glomerular Expression of NADPH Oxidase Componentsand In Situ Production of Superoxide
To examine whether probucol and/or ARB affected the lev-els of Nox2 and Nox4 protein, the major component of NADPHoxidase, we performed Western blotting on days 7 and 56 afterdisease induction (Figure 6). On day 7, a significant increase inNox2 protein was observed in only vehicle-treated nephriticrats, whereas Nox4 protein was significantly increased in allnephritic rats regardless of treatment (Figure 6, A through C).The increased level of Nox2 protein in vehicle-treated nephriticrats seemed to be slightly suppressed by probucol and/or ARBadministration even though this effect was NS (Figure 6, A andB). On day 56, the levels of Nox2 and Nox4 protein in vehicle-treated nephritic rats were upregulated 4.6- and 1.6-fold, re-
Figure 6. Expression of Nox2 and Nox4 in rats that had GN on days 7 (A through C) and 56 (D through F) and had been treatedwith probucol and/or ARB (candesartan). Total protein was extracted from glomeruli that were isolated by the sieving method.The samples were adjusted to a standard content (30 �g) and analyzed by Western blotting using the mouse monoclonal anti-Nox2antibody and rabbit polyclonal anti-Nox4 antibody. (A and D) Representative Western blotting is shown. (B, C, E, and F) Resultsof densitometric analysis. The results represent the mean � SEM. �, nephrectomized control rats; f, vehicle-treated GN rats; o,probucol-treated GN rats; u, ARB-treated GN rats; z, probucol � ARB–treated GN rats. *P � 0.01 versus control; †P � 0.05 versusGN; ‡P � 0.01 versus GN.
J Am Soc Nephrol 17: 783–794, 2006 ARB � Probucol Arrests Progressive Mesangioproliferative GN 789

spectively (Figure 6, D through F). This increased level of Nox2protein was significantly decreased in probucol-treated ne-phritic rats and was attenuated by approximately 50% in ARB-or probucol plus ARB-treated nephritic rats (Figure 6, D and E).In addition, ARB or probucol plus ARB treatment tended todecrease the level of Nox4 in nephritic rats, although thesechanges were not statistically significant (Figure 6, D and F).
For evaluation of the effect of probucol and/or ARB onglomerular O2
·� content, in situ superoxide production of days7 and 56 was detected by DHE (Figure 7). A significant increasein glomerular O2
·� production was observed in only vehicle-treated nephritic rats on day 7 (Figure 7F). Treatment of ne-phritic rats with probucol or ARB seemed to decrease its pro-duction, although it was numerically NS. Treatment withprobucol plus ARB significantly attenuated glomerular O2
·�
production to near the control level. On day 56, O2·� produc-
tion was strikingly enhanced in vehicle-treated nephritic rats,and this was significantly reduced to the control level in pro-bucol- or ARB-treated nephritic rats (Figure 7, B through D andG). Of note, combined treatment completely eliminated theglomerular O2
·� content from day 56 nephritic glomeruli (Fig-ure 7, E and G).
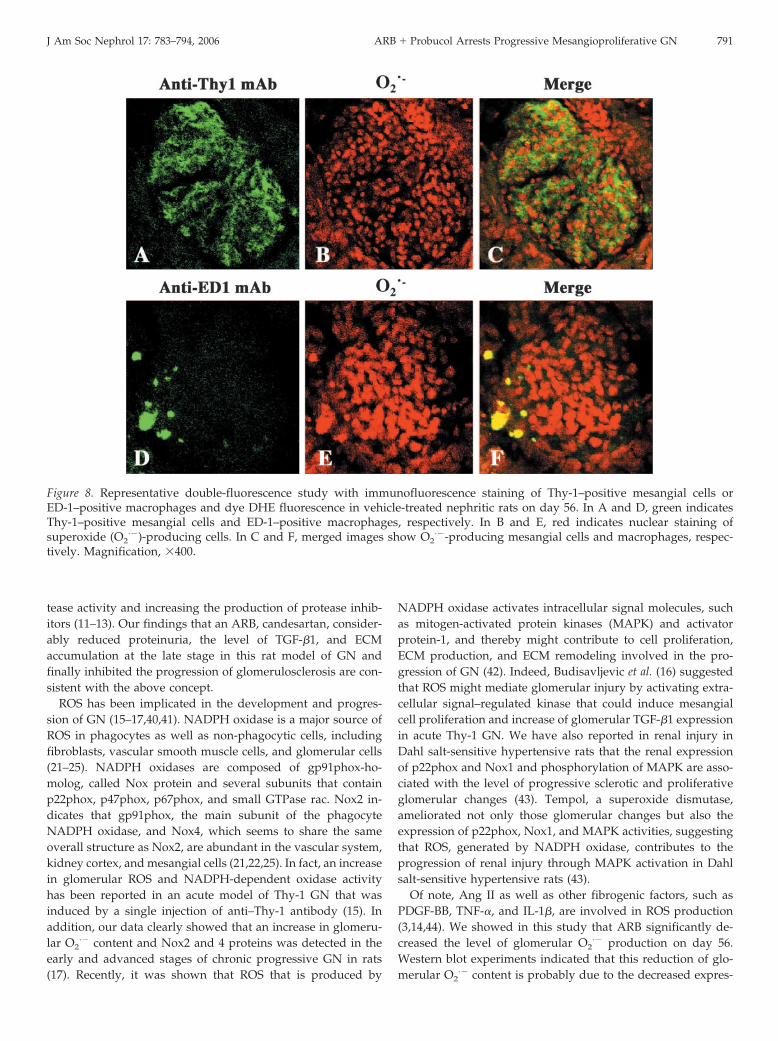
Next, double-immunofluorescent staining of mesangial cellsor macrophages combined with the detection of O2
·� by DHEon frozen sections was performed to identify the O2
·�-produc-ing cells in day 56 nephritic glomeruli. Figure 8, A through C,shows that immunostaining of Thy-1–positive mesangial cellswas co-localized with most of the nuclear staining of O2
·�-
producing cells. Staining of O2·� was also detected on a few
ED-1–positive macrophages that had infiltrated nephritic glo-meruli (Figure 8, D through F). These results suggest thatmesangial cell O2
·� production, possibly via NADPH oxidaseactivation, is involved mainly in the level of glomerular injury,including morphologic parameters in this rat model of GN.
DiscussionIn this study, we demonstrated that the combined adminis-
tration of probucol and ARB fully arrests proteinuria and theprogression of severe mesangioproliferative GN in the rat. Itseems likely that the beneficial effect of such combined treat-ment is due to the synergistic action of Ang II inhibition with areceptor antagonist and the elimination of ROS with a radicalscavenger.
At present, blocking of Ang II action is one of the most usefulstrategies for treating chronic progressive mesangioprolifera-tive GN, including IgA nephropathy. ARB and/or ACEI limitsproteinuria and reduces the decline in GFR and the risk forESRD more effectively than other antihypertensive treatments(6,36–38). Several in vivo animal experiments have indicatedthat the effect of ARB and/or ACEI in preventing the progres-sion to end-stage renal failure is due to the ability of ARB andACEI to modulate the effects of Ang II on the production ofTGF-�1 and ECM as well as on glomerular hypertension(8,10,39). In vitro studies have demonstrated that Ang II stim-ulates TGF-�1 expression, which promotes the synthesis ofECM proteins and inhibits ECM turnover by decreasing pro-
Figure 7. In situ detection of superoxide (O2·�) in glomeruli of rats that had GN and had been treated with probucol and/or ARB
(candesartan). Representative fluorescence micrographs of glomeruli that were stained with the O2·�-sensitive dye dihydro-
ethidium (DHE; red fluorescence) were from nephrectomized control rats (A), vehicle-treated GN rats (B), probucol-treated GNrats (C), ARB-treated GN rats (D), and probucol plus ARB-treated GN rats (E) on day 56 after disease induction. Semiquantitativeassessment of mean fluorescence intensity for glomerular O2
·� content detected on days 7 (F) and 56 (G) in all groups that receivedeach treatment. �, nephrectomized control rats; f, vehicle-treated GN rats; o, probucol-treated GN rats; u, ARB-treated GN rats;z, probucol � ARB–treated GN rats. Data are mean � SEM. *P � 0.01 versus control; †P � 0.01 versus GN; ‡P � 0.01 versusGN�probucol or ARB; §P � 0.05 versus control.
790 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 783–794, 2006
tease activity and increasing the production of protease inhib-itors (11–13). Our findings that an ARB, candesartan, consider-ably reduced proteinuria, the level of TGF-�1, and ECMaccumulation at the late stage in this rat model of GN andfinally inhibited the progression of glomerulosclerosis are con-sistent with the above concept.
ROS has been implicated in the development and progres-sion of GN (15–17,40,41). NADPH oxidase is a major source ofROS in phagocytes as well as non-phagocytic cells, includingfibroblasts, vascular smooth muscle cells, and glomerular cells(21–25). NADPH oxidases are composed of gp91phox-ho-molog, called Nox protein and several subunits that containp22phox, p47phox, p67phox, and small GTPase rac. Nox2 in-dicates that gp91phox, the main subunit of the phagocyteNADPH oxidase, and Nox4, which seems to share the sameoverall structure as Nox2, are abundant in the vascular system,kidney cortex, and mesangial cells (21,22,25). In fact, an increasein glomerular ROS and NADPH-dependent oxidase activityhas been reported in an acute model of Thy-1 GN that wasinduced by a single injection of anti–Thy-1 antibody (15). Inaddition, our data clearly showed that an increase in glomeru-lar O2
·� content and Nox2 and 4 proteins was detected in theearly and advanced stages of chronic progressive GN in rats(17). Recently, it was shown that ROS that is produced by
NADPH oxidase activates intracellular signal molecules, suchas mitogen-activated protein kinases (MAPK) and activatorprotein-1, and thereby might contribute to cell proliferation,ECM production, and ECM remodeling involved in the pro-gression of GN (42). Indeed, Budisavljevic et al. (16) suggestedthat ROS might mediate glomerular injury by activating extra-cellular signal–regulated kinase that could induce mesangialcell proliferation and increase of glomerular TGF-�1 expressionin acute Thy-1 GN. We have also reported in renal injury inDahl salt-sensitive hypertensive rats that the renal expressionof p22phox and Nox1 and phosphorylation of MAPK are asso-ciated with the level of progressive sclerotic and proliferativeglomerular changes (43). Tempol, a superoxide dismutase,ameliorated not only those glomerular changes but also theexpression of p22phox, Nox1, and MAPK activities, suggestingthat ROS, generated by NADPH oxidase, contributes to theprogression of renal injury through MAPK activation in Dahlsalt-sensitive hypertensive rats (43).
Of note, Ang II as well as other fibrogenic factors, such asPDGF-BB, TNF-�, and IL-1�, are involved in ROS production(3,14,44). We showed in this study that ARB significantly de-creased the level of glomerular O2
·� production on day 56.Western blot experiments indicated that this reduction of glo-merular O2
·� content is probably due to the decreased expres-
Figure 8. Representative double-fluorescence study with immunofluorescence staining of Thy-1–positive mesangial cells orED-1–positive macrophages and dye DHE fluorescence in vehicle-treated nephritic rats on day 56. In A and D, green indicatesThy-1–positive mesangial cells and ED-1–positive macrophages, respectively. In B and E, red indicates nuclear staining ofsuperoxide (O2
·�)-producing cells. In C and F, merged images show O2·�-producing mesangial cells and macrophages, respec-
tively. Magnification, �400.
J Am Soc Nephrol 17: 783–794, 2006 ARB � Probucol Arrests Progressive Mesangioproliferative GN 791

sion of glomerular Nox2 protein by ARB, because several re-ports based on cell culture studies have demonstrated that AngII stimulated the protein and mRNA expression of Nox2(45,46). Therefore, the reduction of glomerular O2
·� content inGN by ARB might be linked to some of the renoprotectiveeffects that are induced by Ang II blockade.
Probucol, a cholesterol-lowering drug with potent antioxi-dant properties, reduced glomerular O2
·� production in GN toalmost the normal level, although a significant amelioration ofproteinuria, glomerular pathology, and expression of TGF-�1and ECM was not seen in probucol-treated nephritic rats. Pro-bucol seems to act mainly as an antioxidant in nephritic ratsbecause the level of serum cholesterol was not significantlyreduced. The level of Nox2 protein was slightly decreased onday 7 and significantly downregulated on day 56 by probucol.This suppressive effect on Nox2 might lead to significant O2
·�
reduction in long-term probucol-treated nephritic rats via coor-dination with the action as a radical scavenger.
The combination of probucol and ARB not only completelyeliminated O2
·� production but also normalized urinary pro-tein excretion and the expression of TGF-�1 and ECM seen inGN, resulting in full prevention of the progression of GN.Long-term combination therapy with ARB and probucol mayconfer a strong, synergistic antioxidative action on nephriticglomeruli by inhibiting Nox2 protein expression associatedwith NADPH oxidase activity and by scavenging glomerularO2
·� content and thereby enhance the renoprotective effect ofARB against the progressive nature of the disease in this ratmodel of GN. It is interesting that HMG-CoA reductase inhib-itors or statins, which are cholesterol-lowering agents, alsoshow strong antioxidant ability involved in the inhibition ofRac1 geranylgeranylation and enhance the beneficial effects ofRAS inhibition on several types of organ damage (47,48). Nie etal. (49) reported in a model of CCl4-induced liver fibrosis thatpitavastatin enhances the antifibrogenesis effects of candesar-tan. Pitavastatin significantly enhanced the suppressive effectsof ARB on TGF-�1 expression and fibrogenesis, whereas italone did not affect liver fibrogenesis or TGF-�1 expression.Regarding the effect of statins on glomerular injury, Zoja et al.(39) reported that therapy with either ACEI and ARB or ceriv-astatin had limited effects, but combination therapy with theseagents further reduced glomerulosclerosis and urinary proteinexcretion in a rat model of Heymann nephritis. The expressionof TGF-�1 was significantly decreased by triple combined ther-apy, whereas ACEI, ARB, and cerivastatin did not significantlyaffect TGF-�1 expression in GN. Although neither study men-tioned substantial changes in O2
·� production in damagedtissues by statins, several studies have demonstrated that st-atins reduced oxidative stress in damaged tissues (47,48).Therefore, the combination of an antioxidant and Ang II block-ade might be a useful strategy for preventing the progression ofchronic GN. Further investigations, including in vitro studies,are needed to explore the molecular mechanisms that are re-sponsible for the inhibitory effects of combined therapy on theprogression of GN by especially focusing on the signal path-ways that lead to the suppression of the expression of TGF-�1.
Probucol is a well-established pharmacologic agent that has
been shown to reduce restenosis after coronary balloon angio-plasty by improving vascular remodeling in humans (50). Fur-thermore, Haas et al. (51) reported that probucol also hadfavorable renal effects in membranous nephropathy in a clinicalstudy. Therefore, further studies on rat models of progressiveGN and patients with chronic GN, such as IgA nephropathyand diabetic nephropathy, will be necessary to address whethercombined therapy with probucol and ARB could have clinicalsignificance.
ConclusionOur study provides important evidence that the antioxidant
probucol when added to ARB fully arrests proteinuria and theprogression of GN. The data suggest that the beneficial effectsof this combined therapy might be mediated through the elim-ination of ROS that is produced mainly by NADPH oxidase inGN. The combination of probucol and ARB, both of which arefrequently used in daily clinical practice, may represent a novelroute of therapy for patients with progressive GN.
AcknowledgmentsThis work was supported by grants from the Japanese Ministry of
Welfare (16591035).Parts of this work were published in abstract form at the 37th Annual
Meeting of the American Society of Nephrology; October 28 to Novem-ber 1, 2004; St. Louis, MO.
We are grateful to Dr. Masuko Ushio-Fukai and Dr. Tohru Fukai(Emory University) and Dr. Katsuyuki Matsui (Teikyo University) forhelpful discussions. We also thank Dr. Masanori Takamatsu and Ms.Naomi Okamoto for excellent technical assistance. Probucol and can-desartan were kindly provided by Otsuka Pharmaceutical Co. Ltd. andTakeda Chemical Industries (Osaka, Japan), respectively.
References1. Klahr S, Schreiner G, Ichikawa I: The progression of renal
disease. N Engl J Med 318: 1657–1666, 19982. Border WA, Noble NA: Transforming growth factor beta in
tissue fibrosis. N Engl J Med 331: 1286–1292, 19943. Abboud HE: Growth factors in glomerulonephritis. Kidney
Int 43: 252–267, 19934. Floege J, Johnson RJ, Gordon K, Iida H, Pritzl P, Yoshimura
A, Campbell C, Alpers CE, Couser WG: Increased synthe-sis of extracellular matrix in mesangial proliferative ne-phritis. Kidney Int 40: 477–488, 1991
5. Border WA, Noble N: Maximizing hemodynamic-indepen-dent effects of angiotensin II antagonists in fibrotic dis-eases. Semin Nephrol 21: 563–572, 2001
6. Russo D, Minutolo R, Pisani A, Esposito R, Signoriello G,Andreucci M, Balletta MM: Coadministration of losartanand enalapril exerts additive antiproteinuric effect in IgAnephropathy. Am J Kidney Dis 38: 18–25, 2001
7. Ruggenenti P, Fassi A, Ilieva AP, Bruno S, Iliev IP, Bruse-gan V, Rubis N, Gherardi G, Arnoldi F, Ganeva M, Ene-Iordache B, Gaspari F, Perna A, Bossi A, Trevisan R,Dodesini AR, Remuzzi G; Bergamo Nephrologic DiabetesComplications Trial (BENEDICT) Investigators: Prevent-ing microalbuminuria in type 2 diabetes. N Engl J Med 351:1941–1951, 2004
8. Nakamura T, Obata J, Kimura H, Ohno S, Yoshida Y,
792 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 783–794, 2006

Kawachi H, Shimizu F: Blocking angiotensin II amelioratesproteinuria and glomerular lesions in progressive mesangio-proliferative glomerulonephritis. Kidney Int 55: 877–889, 1999
9. Mackenzie HS, Ots M, Ziai F, Lee KW, Kato S, Brenner BM:Angiotensin receptor antagonists in experimental modelsof chronic renal failure. Kidney Int Suppl 63: S140–S143,1997
10. Peters H, Border WA, Noble NA: Targeting TGF-beta over-expression in renal disease: Maximizing the antifibroticaction of angiotensin II blockade. Kidney Int 54: 1570–1580,1998
11. Kagami S, Border WA, Miller DE, Noble NA: AngiotensinII stimulates extracellular matrix protein synthesis throughinduction of transforming growth factor-beta expression inrat glomerular mesangial cells. J Clin Invest 93: 2431–2437,1994
12. Kagami S, Kuhara T, Okada K, Kuroda Y, Border WA,Noble NA: Dual effects of angiotensin II on the plasmino-gen/plasmin system in rat mesangial cells. Kidney Int 51:664–671, 1997
13. Singh R, Alavi N, Singh AK, Leehey DJ: Role of angiotensinII in glucose-induced inhibition of mesangial matrix deg-radation. Diabetes 48: 2066–2073, 1999
14. Striker LJ, Peten EP, Elliot SJ, Doi T, Striker GE: Mesangialcell turnover: Effect of heparin and peptide growth factors.Lab Invest 64: 446–456, 1991
15. Gaertner SA, Janssen U, Ostendorf T, Koch KM, Floege J,Gwinner W: Glomerular oxidative and antioxidative sys-tems in experimental mesangioproliferative glomerulone-phritis. J Am Soc Nephrol 13: 2930–2937, 2002
16. Budisavljevic MN, Hodge LA, Barber K, Fulmer JR,Durazo-Arvizu RA, Self SE, Kuhlmann M, Raymond JR,Greene EL: Oxidative stress in the pathogenesis of exper-imental mesangial proliferative glomerulonephritis. Am JPhysiol Renal Physiol 285: F1138–F1148, 2003
17. Kondo S, Kagami S, Shimizu M, Kitamura A, Quinn MT,Kawachi H, Shimizu F, Kuroda Y: Role of gp91phox-con-taining NADPH oxidase in progressive model of rat me-sangioproliferative glomerulonephritis (GN) [Abstract].J Am Soc Nephrol 14: SA-PO542, 2003
18. Tylicki L, Rutkowski B, Horl WH: Antioxidant: A possiblerole in kidney protection. Kidney Blood Press Res 26: 303–314, 2003
19. Kerjaschki D, Neale TJ: Molecular mechanisms of glomer-ular injury in rat experimental membranous nephropathy(Heymann nephritis). J Am Soc Nephrol 7: 2518–2526, 1996
20. Kuzuya M, Kuzuya F: Probucol as an antioxidant andantiatherogenic drug. Free Radic Biol Med 14: 67–77, 1993
21. Griendling KK, Sorescu D, Ushio-Fukai M: NAD(P)H ox-idase: Role in cardiovascular biology and disease. Circ Res74: 1141–1148, 2000
22. Lassegue B, Clempus RE: Vascular NAD(P)H oxidases:Specific features, expression, and regulation. Am J PhysiolRegul Integr Comp Physiol 285: R277–R297, 2003
23. Radeke HH, Meier B, Topley N, Floge J, Habermehl GG,Resch K: Interleukin 1-alpha and tumor necrosis factor-alpha induce oxygen radical production in mesangial cells.Kidney Int 37: 767–776, 1990
24. Jones SA, Hancock JT, Jones OT, Neubauer A, Topley N:The expression of NADPH oxidase components in humanglomerular mesangial cells: Detection of protein and
mRNA for p47phox, p67phox, and p22phox. J Am SocNephrol 5: 1483–1491, 1995
25. Gorin Y, Ricono JM, Kim NH, Bhandari B, Choudhury GG,Abboud HE: Nox4 mediates angiotensin II-induced activa-tion of Akt/protein kinase B in mesangial cells. Am JPhysiol Renal Physiol 285: F219–F229, 2003
26. Burritt JB, Quinn MT, Jutila MA, Bond CW, Jesaitis AJ:Topological mapping of neutrophil cytochrome b epitopeswith phage-display libraries. J Biol Chem 270: 16974–16980,1995
27. Szocs K, Lassegue B, Sorescu D, Hilenski LL, Valppu L,Couse TL, Wilcox JN, Quinn MT, Lambeth JD, GriendlingKK: Upregulation of Nox-based NAD(P)H oxidases in re-stenosis after carotid injury. Arterioscler Thromb Vasc Biol22: 21–27, 2002
28. Taal MW, Zandi-Nejad K, Weening B, Shahsafaei A, KatoSM, Lee KW, Ziai F, Jiang TM, Brenner BM, Mackenzie HS:Proinflammatory gene expression and macrophage recruit-ment in the rat remnant kidney. Kidney Int 58: 1664–1676,2000
29. Fabiani ME, Dinh DT, Nassis L, Casley DJ, Johnston CI: Invivo inhibition of angiotensin receptors in the rat kidneyby candesartan cilexetil: A comparison with losartan. Am JHypertens 13: 1005–1013, 2000
30. Modi K, Morrissey J, Shah SV, Schreiner GF, Klahr S:Effects of probucol on renal function in rats with bilateralureteral obstruction. Kidney Int 38: 843–850, 1990
31. Kita T, Nagano Y, Yokode M, Ishii K, Kume N, Ooshima A,Yoshida H, Kawai C: Probucol prevents the progression ofatherosclerosis in Watanabe heritable hyperlipidemic rab-bit, an animal model for familial hypercholesterolemia.Proc Natl Acad Sci U S A 84: 5928–5931, 1987
32. Nishiyama A, Yao L, Nagai Y, Miyata K, Yoshizumi M,Kagami S, Kondo S, Kiyomoto H, Shokoji T, Kimura S,Kohno M, Abe Y: Possible contributions of reactive oxygenspecies and mitogen-activated protein kinase to renal in-jury in aldosterone/salt-induced hypertensive rats. Hyper-tension 43: 841–848, 2004
33. Kagami S, Urushihara M, Kondo S, Hayashi T, Yamano H,Loster K, Vossmeyer D, Reutter W, Kuroda Y: Effects ofanti-alpha1 integrin subunit antibody on anti-Thy-1 glo-merulonephritis. Lab Invest 82: 1219–1227, 2002
34. Kondo S, Kagami S, Kido H, Strutz F, Muller GA, KurodaY: Role of mast cell tryptase in renal interstitial fibrosis.J Am Soc Nephrol 12: 1668–1676, 2001
35. Kondo S, Kagami S, Urushihara M, Kitamura A, ShimizuM, Strutz F, Muller GA, Kuroda Y: Transforming growthfactor-beta1 stimulates collagen matrix remodelingthrough increased adhesive and contractive potential byhuman renal fibroblasts. Biochim Biophys Acta Mol Cell Res1693: 91–100, 2004
36. Nakao N, Yoshimura A, Morita H, Takada M, Kayano T,Ideura T: Combination treatment of angiotensin-II receptorblocker and angiotensin-converting-enzyme inhibitor innon-diabetic renal disease (COOPERATE): A randomisedcontrolled trial. Lancet 361: 117–124, 2003
37. Taal MW, Brenner BM: Renoprotective benefits of RASinhibition: From ACEI to angiotensin II antagonists. KidneyInt 57: 1803–1817, 2000
38. Dillon JJ: Angiotensin-converting enzyme inhibitors andangiotensin receptor blockers for IgA nephropathy. SeminNephrol 24: 218–224, 2004
J Am Soc Nephrol 17: 783–794, 2006 ARB � Probucol Arrests Progressive Mesangioproliferative GN 793

39. Zoja C, Corna D, Camozzi D, Cattaneo D, Rottoli D, Batani C,Zanchi C, Abbate M, Remuzzi G: How to fully protect thekidney in a severe model of progressive nephropathy: Amultidrug approach. J Am Soc Nephrol 13: 2898–2908, 2002
40. Neale TJ, Ullich R, Ojha P, Poczewski H, Verhoeven AJ,Kerjaschki D: Reactive oxygen species and neutrophil re-spiratory burst cytochrome b558 are produced by kidneyglomerular cells in passive Heymann nephritis. Proc NatlAcad Sci U S A 90: 3645–3649, 1993
41. Shah SV: The role of reactive oxygen metabolites in glo-merular disease. Annu Rev Physiol 57: 245–262, 1995
42. Viedt C, Soto U, Krieger-Brauer HI, Fei J, Elsing C, KublerW, Kreuzer J: Differential activation of mitogen-activatedprotein kinases in smooth muscle cells by angiotensin II.Involvement of p22phox and reactive oxygen species. Ar-terioscler Thromb Vasc Biol 20: 940–948, 2000
43. Nishiyama A, Yoshizumi M, Hitomi H, Kagami S, KondoS, Miyatake A, Fukunaga M, Tamaki T, Kiyomoto H,Kohno M, Shokoji T, Kimura S, Abe Y: The SOD mimetictempol ameliorates glomerular injury and reduces mito-gen-activated protein kinase activity in Dahl salt-sensitiverats. J Am Soc Nephrol 15: 306–315, 2004
44. Jaimes EA, Galceran JM, Raij L: Angiotensin II inducessuperoxide anion production by mesangial cells. Kidney Int54: 775–784, 1998
45. Rueckschloss U, Quinn MT, Holtz J, Morawietz H: Dose-dependent regulation of NAD(P)H oxidase expression byangiotensin II in human endothelial cells: Protective effectof angiotensin II type 1 receptor blockade in patients withcoronary artery disease. Arterioscler Thromb Vasc Biol 22:1845–1851, 2002
46. Touyz RM, Chen X, Tabet F, Yao G, He G, Quinn MT,Pagano PJ, Schiffin EL: Expression of a functionally ac-tive gp91phox-containing neutrophil-type NADPH oxi-dase in smooth muscle cells from human resistance ar-teries. Regulation by angiotensin II. Circ Res 90: 1205–1213, 2002
47. Takemoto M, Node K, Nakagami H, Liao Y, Grimm M,Takemoto Y, Kitakaze M, Liao JK: Statins as antioxidanttherapy for preventing cardiac myocyte hypertrophy.J Clin Invest 108: 1429–1437, 2001
48. Maack C, Kartes T, Kilter HM, Schafers HJ, Nickenig G,Bohm M, Laufs U: Oxygen free radical release in humanfailing myocardium is associated with increased activity ofrac1-GTPase and represents a target for statin treatment.Circulation 108: 1567–1574, 2003
49. Nie L, Imamura M, Itoh H, Ueno H: Pitavastatin enhancesthe anti-fibrogenesis effects of candesartan, an angiotensinII receptor blocker, on CCl4-induced liver fibrosis in rats. JUOEH 26: 165–177, 2004
50. Cote G, Tardif JC, Lesperance J, Lambert J, Bourassa M,Bonan R, Gosselin G, Joyal M, Tanguay JF, Nattel S, GalloR, Crepeau J; for the Multivitamins and Probucol StudyGroup: Effect of probucol on vascular remodeling aftercoronary angioplasty. Circulation 99: 30–35, 1999
51. Haas M, Mayer G, Wirsberger G, Holzer H, Ratschek M,Neyer U, Neuweiler J, Kramar R, Schneider B, Breiteneder-Geleff S, Regele HM, Horl WH, Kerjaschki D: Antioxidanttreatment of therapy-resistant idiopathic membranous ne-phropathy with probucol: A pilot study. Wien Klin Wochen-schr 114: 143–147, 2002
794 Journal of the American Society of Nephrology J Am Soc Nephrol 17: 783–794, 2006











![[2] Whitening Bleaching and Antioxidant](https://img.pdfslide.tips/doc/110x75/563db985550346aa9a9e1e68/2-whitening-bleaching-and-antioxidant.jpg)