Embed Size (px)
Citation preview

Università degli Studi di Padova
FACOLTÀ DI INGEGNERIACorso di Laurea in Ingegneria dell'Informazione
ANALISI DI ALCUNI MODELLI
NEURONALI
Relatore:Chiar.mo Prof.Mauro Bisiacco
Laureanda:Carlotta Tecchio
Anno Accademico 2012/2013

2
Ai miei genitori, unica vera fonte di ispirazione.

Indice
1 Introduzione 5
2 Elementi di biologia e �siologia 7
2.1 Neuroni . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 72.2 Segnali neuronali e potenziale d'azione . . . . . . . . . . . . . . . . . . . 82.3 Le sinapsi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 92.4 La membrana cellulare . . . . . . . . . . . . . . . . . . . . . . . . . . . . 102.5 Correnti ioniche e canali ionici . . . . . . . . . . . . . . . . . . . . . . . . 112.6 Origine del potenziale di riposo (L'equazione di Nernst) . . . . . . . . . . 12
3 Modelli neuronali 15
3.1 Modello di Hodgkin-Huxley dell'assone gigante del calamaro . . . . . . . 153.2 Tecnica di blocco del voltaggio . . . . . . . . . . . . . . . . . . . . . . . . 163.3 Derivazione delle equazioni di Hodgkin-Huxley . . . . . . . . . . . . . . . 183.4 Simulazioni . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
4 Modelli ridotti 27
4.1 Modello di FitzHugh-Nagumo . . . . . . . . . . . . . . . . . . . . . . . . 274.2 Simulazioni . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
5 Conclusioni 33
3

4 INDICE

Capitolo 1
Introduzione
Nel presente lavoro di tesi si è studiato un modello matematico sviluppato a partire dailavori pionieristici di A.L. Hodgkin e A.F. Huxley sulla �siologia del neurone. Scopo diquesta tesi di laurea è lo studio dei comportamenti dinamici di modelli di un singoloneurone, analizzando la risposta del singolo neurone sottoposto a continue stimolazioni.Viene studiata successivamente una variante del modello di Hodgkin-Huxley, ovvero ilmodello sempli�cato di FitzHugh-Nagumo (che è un modello paradigmatico per i sistemieccitabili). Il comportamento di tali modelli verrà in�ne simulato tramite il pacchettoSimulink del software Matlab con l'obiettivo di osservare che le curve dei gra�ci ottenutedalle simulazioni coincidano con quelle ottenute dai due scienziati americani.
5

6 CAPITOLO 1. INTRODUZIONE

Capitolo 2
Elementi di biologia e �siologia
In questa sezione si andranno ad esaminare alcune caratteristiche generali dei neuroni ele loro interazioni con altri tipi di cellule. Dopo una breve descrizione delle caratteristichemorfologiche e funzionali del neurone, ci si so�ermerà ad analizzare i potenziali di azioneche sono impulsi di tensione tipici generati durante la dinamica neuronale e sono anchei responsabili dello scambio di informazione fra i neuroni. In ultimo verrà data unadescrizione dettagliata della membrana cellulare prestando una particolare attenzionealle sue proprietà elettriche.
2.1 Neuroni
I neuroni (si veda la �gura 2.1) costituiscono le unità fondamentali del sistema nervo-so, grazie alle loro peculiari proprietà �siologiche e chimiche sono in grado di ricevere,integrare e trasmettere segnali elettrici de�niti impulsi nervosi che associati a correntiioniche attraversano la membrana del neurone. I neuroni sono in numero molto elevato,il cervello umano ne è costituito da circa 100 miliardi, ed hanno diverse forme e dimen-sioni. Pur nella loro varietà possiamo quasi sempre distinguere tre regioni principali: idendriti, l'assone e il corpo cellulare o soma (si veda la �gura 2.2).
Il soma ha una struttura compatta che risulta circa sferica (70µm di diametro) edè l'unità deputata all'elaborazione dell'informazione. I dendriti sono estensioni relati-vamente corte del soma; essi si rami�cano subito e quindi circondano il corpo cellularecon un reticolo che fornisce un'enorme area mediante la quale il neurone può prenderecontatto e ricevere segnali da altri neuroni. Gli assoni sono lunghe protuberanze che siproiettano dal soma lontano dal corpo cellulare e la loro funzione è quella di portare leinformazioni generate dal soma ai dendriti di un altro neurone. Se si dovesse esprimerein termini elettronici questa interazione, i dendriti potrebbero essere visti come ′′input′′ ,gli assoni come ′′output′′ e il soma come la CPU. Attraverso la �gura 2.2 è possibile farsi
7

8 CAPITOLO 2. ELEMENTI DI BIOLOGIA E FISIOLOGIA
Figura 2.1: Struttura di un neurone
Figura 2.2: Forme e dimensioni dei neuroni
un'idea del numero di connessioni attraverso cui una rete neuronale espleta le propriefunzioni.
2.2 Segnali neuronali e potenziale d'azione
La di�erenza di carica elettrica che sussiste sui due versanti della membrana plasmatica,cioè fra interno ed esterno della cellula neuronale, viene de�nita potenziale di membrana;

2.3. LE SINAPSI 9
la cui variazione temporale e spaziale è indicata dal termine segnale neuronale. In unneurone non stimolato, cioè a riposo, il potenziale di membrana assume un valore dicirca -70 mV, chiamato appunto potenziale di riposo. I potenziali di azione sono rapidevariazioni nel potenziale di membrana che passa dal normale valore negativo verso unopositivo, e termina con una variazione che ripristina il potenziale negativo; essi hannouna forma pressochè stereotipata e in �gura 2.3 ne è riportato un esempio.
Si notino le caratteristiche principali:
• L'impulso ha una durata di circa 1-2 ms ed una ampiezza di circa 100-120 mV
• Nella fase di discesa l'impulso prima di ritornare al valore di riposo passa attraversouna fase di iperpolarizzazione, della durata di circa 10 ms
• Il potenziale di azione viaggia lungo l'assone ed è trasmesso inalterato agli al-tri neuroni, e costituisce l'unità elementare associata alla trasmissione dei segnalineuronali
Figura 2.3: Forma tipica di un impulso
2.3 Le sinapsi
Sotto forma di impulsi elettrici l'informazione viene trasmessa da un neurone all'altroa livello delle sinapsi. In tale contesto il neurone ′′trasmittente′′ prende il nome dipresinaptico e quello ′′ricevente′′ di postsinaptico.
Si distinguono essenzialmente due tipi di sinapsi: la sinapsi chimica e la sinapsi elet-trica. La sinapsi chimica, della quale lo schema è riportato nella �gura 2.4, è la più

10 CAPITOLO 2. ELEMENTI DI BIOLOGIA E FISIOLOGIA
Figura 2.4: Esempio di sinapsi chimica
comune nel cervello dei vertebrati, e si basa sul meccanismo descritto in seguito. Ilpotenziale di azione generato dal neurone presinaptico, giunto all'estremità dell'assonedepolarizza la membrana cellulare causando il rilascio all'interno della fessura sinapti-ca di vescicole contenenti neurotrasmettitori; tale neurotrasmettitore, raggiunto l'altroneurone, è rivelato da speciali molecole che provocano l'apertura di canali attraverso cuiuna corrente ionica �uisce causando una variazione del potenziale di membrana. Quindiin una sinapsi chimica si ha prima una trasformazione di un segnale elettrico in unochimico e poi la successiva trasformazione di un segnale chimico in uno elettrico. Lasinapsi elettrica invece realizza una accoppiamento elettrico tra due neuroni adiacenti,permettendo un �usso di corrente elettrica diretto tra essi.
2.4 La membrana cellulare
La membrana cellulare del neurone è un sottile rivestimento, con spessore di 5 nm, chedelimita la cellula in tutti gli organismi viventi, la separa dall'ambiente esterno e ne regolagli scambi con questo. La membrana separa delle cariche, svolge quindi passivamenteuna azione di capacità. Essa è composta da molecole di lipidi e proteine, tali proteineprendono il nome di canali proteici, canali acquosi o canali ionici. I canali proteici sonoappunto molecole proteiche che attraversano tutto lo spessore della membrana cellulareentrando in contatto sia con l'interno della cellula che con il liquido extracellulare. Gliioni inorganici (sodio, potassio, calcio e cloro) costituiscono le correnti ioniche alla basedell'attività elettrica neuronale, essi si muovono attraverso la membrana in modo attivo

2.5. CORRENTI IONICHE E CANALI IONICI 11
legandosi a particolari molecole dette molecole di trasporto. I canali proteici, come si puòvedere nella �gura 2.5, sono fatti di:
• un poro centrale pieno d'acqua;
• una regione del poro che agisce da �ltro di selettività regolando il transito degliioni in base alle dimensioni ed alle caratteristiche chimico-�siche;
• un sistema di porte che si aprono e si chiudono in modo stocastico, di norma lostato di chiusura predomina quando il potenziale di membrana è al valore di riposo.
Figura 2.5: Struttura della membrana cellulare e del canale ionico
2.5 Correnti ioniche e canali ionici
I canali per gli ioni consistono di strutture molecolari cilindriche che attraversano ilduplice strato lipidico della membrana formando un ′′poro′′ che può essere attraversatoda ioni. Per lo più, i canali ionici si comportano come se fossero corredati di un ′′cancello′′
in grado di consentire o impedire il passaggio dello ione speci�co, modi�cando a secondadella condizione, la propria forma. Quando il canale ionico si trova in uno stato diapertura viene detto attivato, mentre quando si trova in uno stato di chiusura vienedetto disattivato. In particolare, i canali voltaggio-dipendenti si aprono o si chiudono inrisposta a cambiamenti nel potenziale elettrico della membrana plasmatica. Riguardoalla selettività della speci�cità ionica i canali si possono distinguere in cationici e anionicia seconda che risultino permeabili agli ioni positivi o negativi. I complicati meccanismialla base della mobilità di ogni specie ionica possono essere empiricamente tenuti in contoin termini di una permeabilità p speci�ca della membrana ad una data specie ionica. Lade�nizione di p si basa sulla seguente relazione:
J = −p∆[C] (2.1)

12 CAPITOLO 2. ELEMENTI DI BIOLOGIA E FISIOLOGIA
dove J è il �usso molare e ∆[C] rappresenta la di�erenza di concentrazione ionica aidue lati della membrana; p ha le dimensioni di una velocità ed è solitamente misuratain cm/s. La permeabilità dipende solo dal tipo e dal numero di canali ionici presentisulla membrana. I canali anionici sono principalmente permeabili al cloro, che è digran lunga il maggior permeante anionico nelle soluzioni biologiche. Limitandoci alleproprietà passive della membrana di solito viene riportata la conduttanza che è unamisura dell'abilità della membrana di trasportare corrente elettrica ed è misurata inSiemens. La conduttanza di una membrana non dipende solo dalla sua permeabilità,perché la corrente è trasportata dagli ioni, e quindi è necessario tener conto anche dellaconcentrazione ionica all'interno ed all'esterno della cellula. In de�nitiva la corrente cheattraversa un canale ionico dipenderà:
• dalla di�erenza di potenziale applicata alla membrana;
• dalla conduttanza del canale;
• dal moto degli ioni dentro la membrana che è descrivibile in termini e�caci da unmoto di�usivo che tende a produrre un �usso dalla zona a maggior concentrazionea quella a minor concentrazione secondo la legge enunciata da Fick :
Jdiff = −Dd[C]
dx(2.2)
dove Jdiff rappresenta il �usso dovuto alla di�usione, D è il coe�ciente di di�usionee [C] è la concentrazione ionica.
2.6 Origine del potenziale di riposo (L'equazione di
Nernst)
Il potenziale di riposo di una cellula è determinato dall'asimmetrica distribuzione degliioni nei due ambienti intra ed extra-cellulare e della di�erente conduttanza alle diversespecie ioniche. Un esempio che coglie l'essenza dell'origine del potenziale di riposo dellacellula neuronale è presentato in �gura 2.6. Gli ioni K+, Na+, Cl+, etc., si muovonoverso zone di minor concentrazione essendo soggetti al campo elettrico generato dal-l'insieme degli altri ioni. Poiché la cellula ha verso gli ioni un comportamento di tiposelettivo, gli ioni non si di�ondono allo stesso modo dentro e fuori la cellula (ad esempiola cellula è molto permeabile allo ione potassio piuttosto che allo ione sodio). Lo ioneK+ viene trasportato all'interno della cellula mentre lo ione Na+ viene espulso a spesedell'organismo. La cellula viene quindi a possedere un potenziale negativo all'internorispetto all'esterno.

2.6. ORIGINE DEL POTENZIALE DI RIPOSO (L'EQUAZIONE DI NERNST) 13
Figura 2.6: Modello sempli�cato per le distribuzioni ioniche
Il potenziale di equilibrio di ogni specie ionica è legato alle concentrazioni intracellularied extracellulari attraverso la cosiddetta equazione di Nernst :
Eione =kT
qln
[n]e[n]i
(2.3)
dove: Eione è il potenziale di equilibrio dello ione; [n]e, [n]i sono rispettivamente leconcentrazioni extracellulari ed intracellulari; k ' 1.38·10−23 è la costante di Boltzmann;T è la temperatura assoluta in gradi Kelvin; q è la carica elettrica (in Coulomb) dellaspecie ionica. L'origine di questa equazione deriva dal seguente ragionamento. Dallameccanica statistica di Boltzmann per sistemi in equilibrio termico si ha che la probabilitàp(U) che una molecola si trovi in uno stato di energia U risulta: p(U) ∝ exp(−U/kT ).Considerando ora gli ioni con carica positiva q in un campo elettrico statico, la loroenergia nel punto x risulta U(x) = qV (x), dove V (x) è il potenziale nel punto x. Ladensità di probabilità sarà proporzionale alla densità ionica [n(x)] quindi:
p(U(x1))
p(U(x2))=
[n(x1)]
[n(x2)]= exp(−q(V (x1)− V (x2))/kT ) (2.4)
e dunque posto ∆V = (V (x1)− V (x2)) la di�erenza di potenziale fra esterno ed internodella cellula si trova l'equazione di Nernst.

14 CAPITOLO 2. ELEMENTI DI BIOLOGIA E FISIOLOGIA

Capitolo 3
Modelli neuronali
In questo capitolo verranno discussi due modelli che descrivono la dinamica del potenzialedi membrana del singolo neurone. La prima parte tratterà la presentazione del notomodello matematico di Hodgkin-Huxley, questi due �siologi inglesi si dedicarono allostudio delle proprietà elettriche delle membrane assoniche, grazie anche alla scelta degliassoni giganti del calamaro come materiale di studio. Hodgkin e Huxley contribuironoin maniera fondamentale alle conoscenze relative ai potenziali di azione che sono statidescritti in precedenza, tanto che il loro lavoro li portò a vincere un premio Nobel perla medicina nel 1963. La parte �nale verterà sulla trattazione dei modelli ridotti, inparticolare saranno trattati i modelli bidimensionali e tra questi il modello di Fitzhugh-Nagumo. L'importanza dei modelli ridotti bidimensionali risiede nel fatto che la bassadimensionalità rende più agevole l'analisi e consente di cogliere gli aspetti essenziali delladinamica di modelli più complicati.
3.1 Modello di Hodgkin-Huxley dell'assone gigante del
calamaro
In questa sezione verrà descritta la tecnica del blocco del voltaggio o voltage-clamp svi-luppata alla �ne degli anni quaranta ed ampiamente sfruttata in quegli anni per studiarele caratteristiche elettro�siologiche della membrana dell'assone gigante del calamaro, cheha permesso a Hodgkin e Huxley di caratterizzare le conduttanze all'origine del poten-ziale di azione. Come si vedrà il modello di Hodgkin-Huxley è costituito da un sistema diquattro equazioni di�erenziali che descrivono la dinamica del potenziale di membrana edelle tre correnti ioniche fondamentali: la corrente del sodio (INa), la corrente del potas-sio (IK) e la corrente di perdita (IL) dovuta principalmente al cloro ma riassume l'e�ettoanche di altre correnti ioniche minori. Verrà evidenziato inoltre come il meccanismoalla base di queste correnti ioniche risieda nel fatto che le conduttanze del sodio e delpotassio sono dipendenti dalla di�erenza del potenziale di membrana; e come dal valore
15
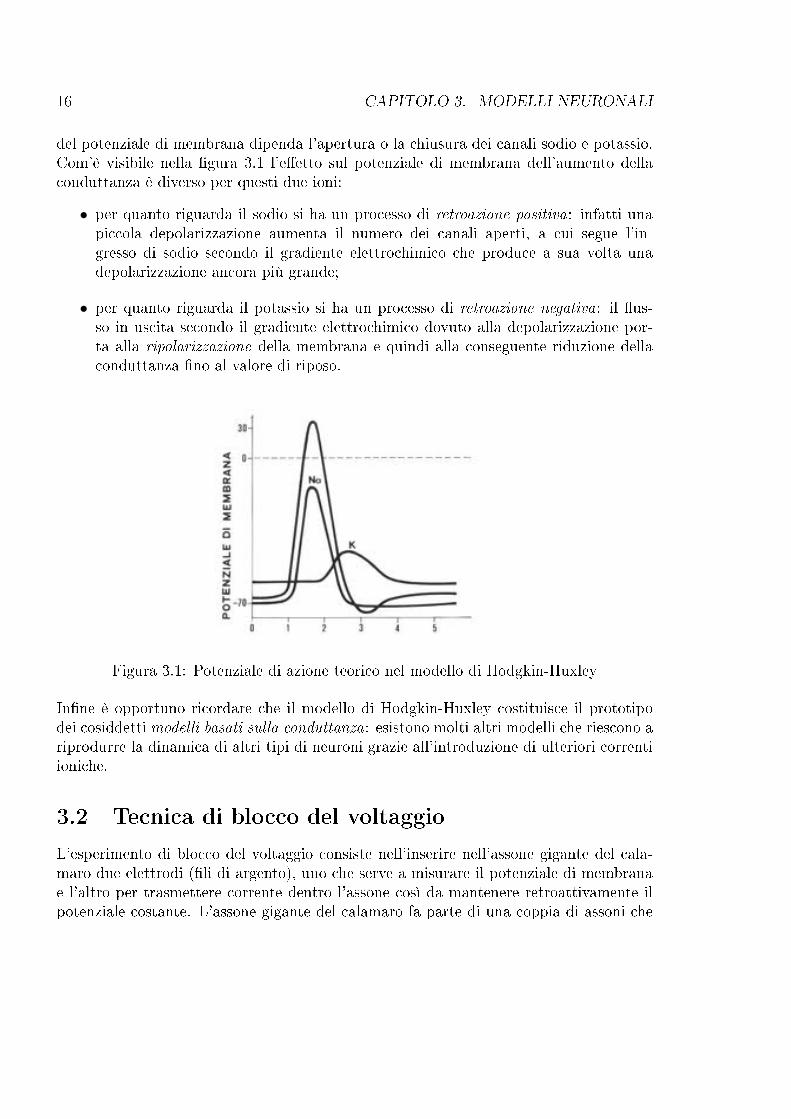
16 CAPITOLO 3. MODELLI NEURONALI
del potenziale di membrana dipenda l'apertura o la chiusura dei canali sodio e potassio.Com'è visibile nella �gura 3.1 l'e�etto sul potenziale di membrana dell'aumento dellaconduttanza è diverso per questi due ioni:
• per quanto riguarda il sodio si ha un processo di retroazione positiva: infatti unapiccola depolarizzazione aumenta il numero dei canali aperti, a cui segue l'in-gresso di sodio secondo il gradiente elettrochimico che produce a sua volta unadepolarizzazione ancora più grande;
• per quanto riguarda il potassio si ha un processo di retroazione negativa: il �us-so in uscita secondo il gradiente elettrochimico dovuto alla depolarizzazione por-ta alla ripolarizzazione della membrana e quindi alla conseguente riduzione dellaconduttanza �no al valore di riposo.
Figura 3.1: Potenziale di azione teorico nel modello di Hodgkin-Huxley
In�ne è opportuno ricordare che il modello di Hodgkin-Huxley costituisce il prototipodei cosiddetti modelli basati sulla conduttanza: esistono molti altri modelli che riescono ariprodurre la dinamica di altri tipi di neuroni grazie all'introduzione di ulteriori correntiioniche.
3.2 Tecnica di blocco del voltaggio
L'esperimento di blocco del voltaggio consiste nell'inserire nell'assone gigante del cala-maro due elettrodi (�li di argento), uno che serve a misurare il potenziale di membranae l'altro per trasmettere corrente dentro l'assone così da mantenere retroattivamente ilpotenziale costante. L'assone gigante del calamaro fa parte di una coppia di assoni che

3.2. TECNICA DI BLOCCO DEL VOLTAGGIO 17
si estendono per tutta la lunghezza del suo mantello nel nervo stellato (si veda la �gura3.2) e che sono deputati al controllo dei muscoli che provocano il movimento del calama-ro; l'assone ha il vantaggio di presentare un diametro molto grande (circa un mm) e ciòconsente di inserire facilmente dentro ad esso un elettrodo assiale. In sostanza la tecnica
Figura 3.2: Assone gigante del calamaro
di blocco del voltaggio consente di registrare il �usso di corrente attraverso la cellulaneuronale in una situazione sperimentale in cui la di�erenza di potenziale applicata allamembrana è mantenuta costante nel tempo. L'apparato per e�ettuare esperimenti con
Figura 3.3: Illustrazione dell'apparato per e�ettuare esperimenti con la tecnica del bloccodel voltaggio
la tecnica del blocco del voltaggio è illustrato in �gura 3.3; l'assone è perfuso con acquadi mare e due sottili �li di argento vengono inseriti all'interno di esso da una estremità.Un elettrodo fornisce la misura del potenziale di membrana (Vm), ossia la di�erenza dipotenziale fra l'interno dell'assone e l'acqua del mare. Inoltre esso è anche connessoall'ingesso invertente (-) di un ampli�catore detto ampli�catore di voltage clamp. L'in-gresso non invertente (+) è a sua volta connesso ad un generatore di tensione variabile

18 CAPITOLO 3. MODELLI NEURONALI
che fornisce il potenziale di comando. In presenza di una di�erenza di potenziale tra gliingressi, l'ampli�catore eroga una corrente in uscita la quale scorre attraverso la mem-brana cellulare e l'acqua del mare. La corrente è misurata mediante la caduta di tensioneattraverso una resistenza in serie; e consta di quattro componenti:
• corrente capacitiva (IC), impulso di corrente di breve durata;
• corrente di dispersione (ID), costante in uscita per tutta la durata del gradino ditensione;
• corrente transitoria entrante (IT ), dovuta all'ingresso del sodio che si attiva moltovelocemente;
• corrente ritardata (IR), dovuta all'uscita del potassio, che si attiva molto piùlentamente di quella del sodio.
La presenza della (IC) risulta del tutto prevedibile in quanto è semplicemente dovutaall'adeguamento del potenziale di membrana alla tensione forzante (-9mV) così comesuccederebbe per qualsiasi capacità ai cui capi vi fosse una di�erenza di potenziale diversada quella del generatore di tensione su cui è chiusa. La corrente (ID) è quella che ci siaspetterebbe se le conduttanze della membrana a riposo non fossero alterate dal gradinodi tensione, ed ha una dipendenza lineare dallo spostamento del voltaggio dal valore diriposo. Costituisce invece un fatto nuovo la componente (IT ) in quanto evidenzia il fattoche un aumento di permeabilità comporta un conseguente �usso di ioni che entra nellacellula. Gli e�etti positivi del voltage clamp in de�nitiva sono:
• si possono misurare le dipendenze temporali delle varie correnti a Vm costante;
• inserendo gli elettrodi si ha anche uno space-clamp, nel senso che tutta la lunghezzadell'assone ha lo stesso Vm.
3.3 Derivazione delle equazioni di Hodgkin-Huxley
Le equazioni di Hodgkin-Huxley si basano sull'analogia di un neurone con un circuitoelettrico composto da resistenze e condensatori. Bisogna però considerare il fatto chele forze che agiscono sui vari ioni non sono solo di tipo elettrico, ma anche di tipoosmotico; ossia il moto medio attraverso la membrana sarà nella direzione della minoreconcentrazione. Con considerazioni di tipo termodinamico, si dimostra che esiste, perogni ione i, un potenziale Vi di equilibrio, al quale cioè le forze elettriche sono compensateda quelle osmotiche. Applicando ora la legge di Ohm, la corrente Ii dello ione i, soddisfa
Ii = gi(V − Vi) (3.1)

3.3. DERIVAZIONE DELLE EQUAZIONI DI HODGKIN-HUXLEY 19
dove gi rappresenta la conduttanza, e si è adottata la convenzione che V è il potenzialeall'interno meno il potenziale all'esterno, mentre Ii è espresso nel senso del moto dicariche positive dall'interno verso l'esterno. Poiché la membrana funge da conduttore, siapplica anche la legge di Faraday, ossia
Q = CV (3.2)
dove Q è la carica separata dal condensatore e C la sua capacità. Queste idee vengono oraapplicate all'assone del calamaro, considerato come un cilindro di raggio a. Si procededividendo il cilindro in tante sezioni lunghe ∆x, in cui potenziale e correnti vengonosupposti costanti. Sia q(x,t) la densità di carica al punto x al tempo t. Ossia la caricatotale nelle celle centrate in x è q(x, t)∆x. Se C è la capacità della membrana per unitàdi area, la legge di Faraday dà
q(x, t)∆x = 2πa∆xCv(x, t) (3.3)
Sia poi Ii(x, t) la densità di corrente dello ione i attraverso la membrana. Come dettosopra vale
Ii(x, t) = gi(v(x, t)− Vi) (3.4)
dove gi è una conduttanza per unità di area. La densità di corrente da x a x + ∆x,indicata con i+(x, t), soddisfa
r∆xi+(x, t) = v(x, t)− v(x+ ∆x, t) (3.5)
dove r è la resistenza per unità di lunghezza. Integrando sull'area della sezione, si hache la corrente totale da x a x+ ∆x è data da
I+(x, t) =πa2
r∆x(v(x, t)− v(x+ ∆x, t)) (3.6)
Usando (3.4) e (3.6) si ha che la derivata della carica totale nelle celle in x soddisfa
d
dtq(x, t)∆x = −
∑i
gi(v(x, t)− Vi)2πa∆x− I+(x, t) + I+(x−∆x, t) =
−2πa∆x∑i
gi(v(x, t)− Vi) +a
2r(v(x+ ∆x, t)− 2v(x, t) + v(x−∆x, t)
(∆x)2).
Usando (3.3) e dividendo per 2πa∆x si ha
Cδ
δtv(x, t) = −
∑i
gi(v(x, t)− Vi) +a
2r(v(x+ ∆x, t)− 2v(x, t) + v(x−∆x, t)
(∆x)2) (3.7)

20 CAPITOLO 3. MODELLI NEURONALI
Facendo tendere ∆x a 0 si ottiene
δ
δtv(x, t) = − 1
C
∑i
gi(v(x, t)− Vi) +a
2rc
δ2
δx2v(x, t). (3.8)
Sulla base dei vari esperimenti in voltage-clamp, Hodgkin e Huxley hanno ipotizzato chegNa e gK si potessero descrivere nel modo seguente:
gNa = gNam3h (3.9)
gk = gkn4
dove gNa e gk rappresentano le conduttanze massime e m,h,n rappresentano la porzionedi molecole (di tipo m, h, e n) in posizione ON. Le espressioni (3.9) verrebbero alloraa signi�care che perché un canale del potassio sia aperto devono esserci 4 molecole n inposizione ON; un canale del sodio è aperto invece quando siano in posizione ON 3 mole-cole m e 1 molecola h. Va notato però che meccanismi come quelli descritti sembravanoplausibili a Hodgkin-Huxley ma mancava completamente una conferma sperimentale. m,n e h soddisfano poi le equazioni di�erenziali
dn
dt= δn(v)(1− n)− βn(v)n
dm
dt= δm(v)(1−m)− βm(v)m (3.10)
dh
dt= δh(v)(1− h)− βh(v)h
Il signi�cato di (3.10) si comprende meglio riscrivendole come
dl
dt= − 1
τl(v)(l − l∞(v)) (3.11)
dove l∞(v) = δl(V )δl(V )+βl(V )
per [l = m,n, h] rappresenta il livello a cui l tenderebbe se
V fosse costante e τl(V ) = 1δl(V )+βl(V )
è il tempo medio in cui avviene una transizione.
Mettendo insieme (3.8) e (3.11) si ottiene il sistema completo:
δVδt
= −gNam3h(V − VNa)− gkn4(V − Vk)− gl(V − Vl) + ηVxxdmdt
= − 1τm(V )
(m−m∞(V ))dndt
= − 1τn(V )
(n− n∞(V ))dhdt
= − 1τh(V )
(h− h∞(V ))
(3.12)
dove gl è la conduttanza rispetto ad altri ioni che si suppone indipendente da V . Lacomplessità di questo modello è tale da non consentirne una risoluzione analitica. Fi-nora il suo comportamento è stato studiato grazie a numerose simulazioni numericheconfrontate con i dati sperimentali; alcune simulazioni vengono riportate nel prossimoparagrafo.

3.4. SIMULAZIONI 21
3.4 Simulazioni
Al �ne di simulare le equazioni della (3.12), alle costanti gNa, gk, gl, VNa, Vk, Vl si sosti-tuiscono i valori presenti nella tabella riportata in �gura 3.4, V , m, n, h sono le variabilidi stato, ηVxx rappresenta l'impulso di corrente elettrico esterno applicato tramite l'e-lettrodo ed è funzione del tempo, successivamente se ne impone l'andamento temporaleusando un ingresso arbitrariamente scelto, ed in�ne verranno analizzati i gra�ci ottenutilavorando con Simulink.
Figura 3.4: Funzioni δl(V ) e βl(v) (l = [m,n, h]) nel modello di Hodgkin-Huxley
Figura 3.5: Valore dei parametri del modello di Hodgkin-Huxley
δVδt
= −120m3h(V − 50)− 36n4(V − (−77))− 0.3(V − (−54.4)) + ηVxxdmdt
= − 1τm(V )
(m−m∞(V ))dndt
= − 1τn(V )
(n− n∞(V ))dhdt
= − 1τh(V )
(h− h∞(V ))
(3.13)
Tramite Simulink si è costruito il seguente schema a blocchi per analizzare il modellodi Hodgkin-Huxley precedentemente descritto. Volendo fare ulteriore chiarezza, al �ne disempli�care la comprensione dello schema, viene riscritta la prima equazione della (3.12)mettendo bene in evidenza la corrente totale composta da: corrente sodio, correntepotassio e corrente di perdita del canale.
Iion = gNam3h(V − ENa) + gkn
4(V − Ek) + gL(V − EL) (3.14)

22 CAPITOLO 3. MODELLI NEURONALI
In �gura 3.9 è presentata una tipica soluzione delle equazioni di Hodgkin-Huxley. Altempo t = 0 il potenziale V (0) è spostato dal suo valore di equilibrio (ca. −60mV ) a−20mV , la curva V (t) ha la tipica forma del potenziale di azione caratterizzata da unaiperpolarizzazione (V (t) arriva a −70mV per t ' 4ms) e un lento ritorno all'equilibrio.In �gura 3.12 sono riportate le variabili m, n ed h dopo la perturbazione. E' inoltreevidente come le simulazioni di seguito riportate riproducano abbastanza fedelmente igra�ci ottenuti dagli stessi Hodgkin e Huxley.Hodgkin e Huxley hanno ottenuto anche ottimi confronti con gli esperimenti (�gura 3.11).Dal punto di vista qualitativo le equazioni (3.12) hanno le seguenti caratteristiche:
• Soglia: se lo spostamento iniziale di V è piccolo, si ha un veloce rilassamento allostato di equilibrio; se lo spostamento supera un valore di soglia, allora si ha ungrande cambiamento delle variabili del sistema.
• Risposta binaria (0-1): quando la soglia viene superata, il comportamento succes-sivo è quasi uguale qualunque sia il valore esatto di V (0), nella classica forma dettapotenziale di azione.
• Periodo refrattario: se il potenziale viene nuovamente elevato subito dopo la �-ne di un potenziale di azione non si ha una nuova eccitazione ma solo un nuovorilassamento all'equilibrio.
I tre fenomeni sopracitati sono visibili nelle simulazioni in �gura 3.9, in particolare:
• In 3.9(a) il potenziale V (0) è posto a −40mV , spostamento di V piccolo e quindi sinota un veloce rilassamento allo stato di equilibrio, noto come fenomeno di Soglia.
• In 3.9(b) il potenziale V (0) è posto a 30mV , dopo che il potenziale di azione èstato elevato e si nota un rilassamento all'equilibrio senza passare attraverso unanuova eccitazione, il cosiddetto Periodo refrattario.
• In 3.9(c) il potenziale V (0) è posto a −80mv,la curva V (t) ha la tipica forma delpotenziale di azione, tale caratteristica è infatti la Risposta binaria (0-1).

3.4. SIMULAZIONI 23
Figura 3.6: Schema a blocchi del modello di Hodgkin-Huxley

24 CAPITOLO 3. MODELLI NEURONALI
Figura 3.7: m ed h rappresentano le variabili di gating dei canali del sodio
Figura 3.8: n rappresenta la variabile di gating dei canali del potassio

3.4. SIMULAZIONI 25
Figura 3.9: Il potenziale V dopo una perturbazione dallo stato di riposo
Figura 3.10: Andamento dei modelli m(t), n(t) e h(t)

26 CAPITOLO 3. MODELLI NEURONALI
Figura 3.11: Alcuni dei confronti e�ettuati da Hodgkin-Huxley fra teoria ed esperimenti

Capitolo 4
Modelli ridotti
Per il modello di Hodgkin-Huxley la non linearità e l'alta dimensionalità rendono dif-�coltosa l'analisi e la visualizzazione della dinamica nello spazio delle fasi. Si ricorreperciò ai cosiddetti modelli ridotti bidimensionali grazie ai quali è possibile ricostruirele caratteristiche essenziali del modello completo tramite un'analisi standard nel pianodelle fasi: uno di questi modelli è appunto il modello di FitzHugh-Nagumo.
4.1 Modello di FitzHugh-Nagumo
Come visto nel precedente capitolo, le equazioni di Hodgkin e Huxley sono su�ciente-mente accurate per la propagazione dell'impulso negli assoni giganti del calamaro, macertamente non per tutti i tipi di neurone in tutte le specie; rimane quindi la necessità dicapire quali sono le caratteristiche delle equazioni che rendono possibile il meccanismodel potenziale di azione. Per questi motivi Fitzhugh nel 1961 ha formulato un sistemapiù semplice di equazioni con le stesse caratteristiche, noto come sistema di Fitzhugh-Nagumo. Prima di tutto, si sfrutta l'idea delle due scale temporali, supponendo che Vvari su una scala veloce, mentre un'altra variabile (w che sostituisce n e h) abbia unadinamica lenta. L'equazione per V abbia le caratteristiche simili alla �gura 4.1: ossiaper w intermedio dV
dtabbia 3 punti di equilibrio, mentre per w elevato vi sia solo un equi-
librio. Un sistema di questo tipo è stato ottenuto da Fitzhugh modi�cando l'equazionedi Van der Pol, un'equazione nota per descrivere le oscillazioni dei circuiti elettrici:
x+ µ(x2 − 1)x+ x = 0 (4.1)
ponendo y = −x3
3+ x− x
µ, si ottiene da (4.1)
{x = µ(x− x3
3)− µy
µy = x(4.2)
27

28 CAPITOLO 4. MODELLI RIDOTTI
Figura 4.1: Quando n è basso si ha un solo equilibrio con V alto; quando n è alto si haun solo equilibrio con V basso; per n intermedio si hanno tre equilibri
Ponendo ora τ = µt e ε = 1µ2
si ha da (4.2)
{dxdt
= x− x3
3− y
dydτ
= εx(4.3)
Se in (4.1) µ� 1 sarà ε� 1. L'equazione di Fitzhugh-Nagumo è una lieve modi�ca di(4.3) ottenuta sostituendo a x− x3
3un'altra cubica e aggiungendo, per maggior �essibilità,

4.1. MODELLO DI FITZHUGH-NAGUMO 29
un altro termine in dydτ. Precisamente{
dVdt
= V (V − a)(1− V )− w ≡ f(V )− wdwdτ
= ε(V − γw)(4.4)
con 0 < a < 1/2 e 0 ≤ γ ≤ 1/3. E' immediato veri�care che (0,0) è l'unico punto di
equilibrio di (4.4). Lo Jacobiano calcolata in (0,0) è J0 =
(−a −1ε −εγ
). Si ha tr(J)<0,
det J0>0, per cui (0,0) è asintoticamente stabile. Dalla �gura 4.2 si vede che dato unpunto iniziale (V0, 0), se 0 < V0 < a si avrà un rilassamento verso l'origine.
Figura 4.2: Il piano delle fasi del sistema descritto dalla (4.4)
Se invece a < V0 < 1, V e W dovranno crescere �no a raggiungere W = f(V ).Per studiare meglio il comportamento delle soluzioni in quest'ultimo caso, si decide discegliere ε = 0. Il sistema (4.4) non è singolare in ε = 0, segue che in ogni intervallodi tempo �nito [0,t] la soluzione (Vε(t),Wε(t)) di (4.4) con dato iniziale Vε(0) = V0,Wε(0) = W0, converge per ε −→ 0+ alla soluzione di (4.4) con ε = 0 ossia W0(t) = W0,V0(t) = z(t) dove z(t) risolve {
dzdτ
= f(V )−W0
z(0) = V0(4.5)
Questo risultato non dice però molto sul comportamento asintotico delle soluzioni di(4.4). Bisogna quindi cambiare la scala temporale ponendo t = ετ e si ottiene

30 CAPITOLO 4. MODELLI RIDOTTI
Figura 4.3: Viene mostrata con la linea tratteggiata la soluzione non di�erenziabile limiteper ε → 0 per V0 > a. Essa è costituita da due segmenti orizzontali ai valori w = 0 ew = wm e dai due tratti dell'isoclina w = f(V )
{εdVdt
= f(V )−WdWdt
= V − γW (4.6)
Gli zeri di dVdt
sono tutti i punti in cui W = f(V ). Si possono scrivere tali punti comeV = φ(W ) de�nendo tre funzioni:
• φ1(W ) de�nita su (Wm,+∞) e a valori in (−∞, Vm);
• φ2(W ) de�nita su (Wm,WM) e a valori in (Vm, VM);
• φ3(W ) de�nita su (−∞,WM) e a valori in (VM ,+∞);
tali che f(φi(W )) = W , i=1,2,3. Sull'intervallo (Wm,WM) in cui tutte le funzioni sonode�nite si ha φ1(W ) < φ2(W ) < φ3(W ). Dalla �gura 4.3 è visibile che φ1(W ) e φ3(W )sono equilibri asintoticamente stabili per il sistema (4.5), mentre φ2(W ) è un equilibrioinstabile. Scegliendo la funzione φ1(W ) con V0 < a, il sistema degenere:
dW
dt= φ1(W )− γ(W ) (4.7)
ha 0 come equilibrio globalmente asintotico; infatti f(φ(W )) = W > γ(W ) per W < 0 ef(φ(W )) = W < γ(W ) per W < 0. Se invece V0 > a il sistema degenere ottenuto è:
dW
dt= φ3(W )− γ(W ) (4.8)

4.2. SIMULAZIONI 31
ricordando che φ3 è de�nita solo sull'intervallo (−∞,WM). Inoltre in ogni punto ditale insieme φ3(W ) > W ; quindi W (t) ha sempre derivate positive, anzi non inferiori aφ3(WM) −WM > 0. Si è quindi ottenuto un risultato preciso di soglia. Se V0 < a siha un ritorno immediato a 0; se V0 > a si ha il ciclo del potenziale di azione. Per ogniε > 0 �ssato, esisteranno pochi dati iniziali per cui si ha una risposta intermedia, questoa causa della dipendenza continua delle soluzioni di (4.4) da ε.
4.2 Simulazioni
Figura 4.4: Schema a blocchi del modello di FitzHugh-Nagumo
Nello schema riportato in �gura (4.4): e rappresenta ε, eg rappresenta εγ ed f(u) =V (V − a)(1− V ). Le equazioni della (4.6) vengono simulate nei due seguenti casi:
• Caso a: V (0) < aε = 0V (0) = −20W (0) = 0
• Caso b: V (0) > aε = 0.1

32 CAPITOLO 4. MODELLI RIDOTTI
V (0) = 40W (0) = 1
Figura 4.5: Andamento di V
Figura 4.6: Andamento di W

Capitolo 5
Conclusioni
In questo elaborato è stato illustrato il modello ideato da Hodgkin e Huxley per spiegarela genesi dei potenziali di azione attraverso esperimenti e�ettuati sull'assone gigante delcalamaro. Tale situazione viene descritta con quattro equazioni di�erenziali non lineari,le soluzioni di queste equazioni, non sono facilmente esprimibili in forma analitica, per cuisi è ricorso all'utilizzo del simulatore Simulink. Obiettivo di codesta tesi è stato quello didimostrare come simulazioni coincidano quasi perfettamente con quelle e�ettuate dagliscienziati negli anni 50 quando ancora non si disponeva dell'ausilio di certi strumentitecnologici, ma soltanto attraverso l'utilizzo di dati sperimentali si è riuscito a capire cheil potenziale di membrana assume la tipica forma del potenziale di azione.
33

34 CAPITOLO 5. CONCLUSIONI

Bibliogra�a
[1] S.E. Luria, S.Y. Gould, S.Singer (1989), Una visione della vita, introduzione allabiologia, Zanichelli, Bologna.
[2] William K. Purves, David Sadava, Gordon H. Orians, H.Craig Heller (2001), Biologia,Zanichelli, Bologna.
[3] A.Torcini, Modelli semplici di neuroni, CNR, Firenze.
[4] Corrado Mascia, Eugenio Montefusco (2011), Modelli analitici per le applicazioni,Università La Sapienza di Roma.
35





























