Embed Size (px)
Citation preview
ANÁLISIS DE LOS DATOS DEL PROGRAMA SACRE EN LA
COMUNIDAD AUTÓNOMA DEL PAÍS VASCO
2007 2007

Documento: ANÁLISIS DE LOS DATOS DEL PROGRAMA SACRE EN LA COMUNIDAD AUTÓNOMA DEL PAÍS VASCO
Fecha de edición: 2007
Autor: SOCIEDAD ESPAÑOLA DE ORNITOLOGÍA (SEO/BIRDLIFE)
Elaboración del informe
Virginia Escandell
Juan Carlos del Moral
Coordinación: Virginia Escandell, José Antonio Gainzarain y José María Fernández
Revisión / Dirección: IKT S.A.
Propietario: Gobierno Vasco. Departamento de Medio Ambiente y Ordenación del Territorio. Dirección de Biodiversidad y Participación Ambiental

ÍNDICE
1. INTRODUCCIÓN 1
2. OBJETIVOS 2
3. METODOLOGÍA DE CENSO 3
3.1. Instrucciones 3
3.2. Material entregado a cada colaborador 5
3.3. Cursos de perfeccionamiento y estandarización de censadores 6
4. RESULTADOS 7
4.1. Cobertura cuadrículas 7
4.2. Hábitat representados 9
4.2. Área de estudio 8
4.3. Evolución de las poblaciones 10
5. EQUIPO DE TRABAJO 13
6. AGRADECIMIENTOS 14
ANEXOS
I. Material entregado a cada colaborador
II. Cobertura en el País vasco en el año 2007
III. Porcentaje de cambio anual obtenido para cada especie y tendencia general
IV. Gráficas de evolución obtenidas para cada especie entre 1998-2007
V. Índice de cambio anual para distintos grupos de especies
VI. Gráficas de evolución de los tres grupos considerados
VII. Análisis estadístico de cada especie generado por el programa TRIM


1. INTRODUCCIÓN
El presente informe corresponde al “Análisis de los datos del programa SACRE en
la CAPV (1998-2007)”, siguiendo las indicaciones establecidas en el contrato adoptado
entre IKT S.A. y SEO/BirdLife.
En el año 1996, SEO/BirdLife puso en marcha el programa de Seguimiento de Aves
Comunes en España (programa SACRE). En concreto, en el ámbito de la CAPV, el trabajo
de campo comenzó a realizarse en 1997.
Con el objetivo de obtener un indicador de biodiversidad de la CAPV, los datos de
esta comunidad pueden ser extraídos de la base de datos general para obtener índices
mensurables, objetivos, renovables y con significación desde el punto de vista de la
evolución de la biodiversidad en este territorio, ya que el tamaño muestral y el número de
años que se ha realizado el censo lo permite.
1

2. OBJETIVOS
El principal objetivo es conocer la tendencia (positiva o negativa) de todas las especies
de aves comunes y dispersas, difícilmente contables y para las que no existe información
sobre su evolución poblacional. Quedarán incluidas todas aquellas especies que se detecten
de una manera significativa con la metodología establecida. Es decir, la mayoría de
paseriformes y otros grupos afines, quedando excluidos grupos como garzas, anátidas,
grandes rapaces y otros taxones para las que es necesario un seguimiento más específico.
Con los muestreos realizados para el Programa SACRE, además de lo anterior, se
obtiene:
· Mejor conocimiento de la biología de las poblaciones de aves comunes y, en particular,
de los factores responsables de su evolución.
· Información sobre las preferencias de hábitat de cada especie.
· Abundancias relativas de cada especie por tipo de hábitat y por comarcas en la
comunidad.
· Identificación de las zonas donde se producen tendencias decrecientes. Esto permitirá
centrar los análisis y las posteriores labores de conservación en aquellos puntos que se
consideren prioritarios.
· Identificación de los tipos de hábitat que estén sufriendo mayores transformaciones, y
en los que la conservación de sus especies de aves pueda estar más amenazada.
· Elaborar una red de ornitólogos con una cualificación adecuada, que permita realizar
éste y otros trabajos relacionados con aves y que asegure una cobertura adecuada y una
calidad de información óptima para toda la comunidad.
2

3. METODOLOGÍA DE CENSO Y ANÁLISIS DE DATOS
La metodología utilizada para la realización de los muestreos es la siguiente:
• La unidad de muestreo es la cuadrícula UTM de 10x10 km, seleccionada por orden de
prioridad, y de forma estratificada en todo el territorio.
• La estratificación para la selección de las cuadrículas está realizada en función de los
tipos de vegetación, de los pisos bioclimáticos de la clasificación de Allué Andrade y de
la división provincial, de forma que se asegura una cobertura proporcional tanto de
ambientes como geográficamente.
• En cada cuadrícula se realizan 20 estaciones de escucha y se registran todas las aves
vistas u oídas en cada una de ellas, en dos categorías de distancia (dentro y fuera de un
radio de 25 m). Estas estaciones se repiten cada año exactamente en los mismos puntos.
• Se realizan dos visitas por temporada a cada cuadrícula, una entre el 15 de abril y el 15
de mayo centrada en el periodo de máxima actividad de reproductores sedentarios y
presaharianos, y otra entre el 15 de mayo y el 15 de junio centrada en el periodo de
máxima actividad de reproductores transaharianos.
• Las fechas de censo son ajustadas en cada zona en función de sus condiciones
climáticas. Así, los censos se reparten en fechas tempranas o tardías dentro de cada
periodo dependiendo de si se localizan en costa, áreas bajas y calurosas o en zonas de
alta montaña.
• En cada estación se describe el hábitat y se anotan anualmente los cambios observados,
de forma que se pueda relacionar la evolución de cada especie con las transformaciones
de hábitat que se produzcan.
3

• Las estaciones son distribuidas homogéneamente por toda la cuadrícula de forma que
aseguren una cobertura geográfica homogénea.
• Los puntos de muestreo son repartidos en los distintos ambientes de la cuadrícula de
forma proporcional a la extensión de cada uno.
• Para el análisis de la evolución de las poblaciones, se utiliza el programa estadístico
TRIM (TRends & Indices for Monitoring data), desarrollado por Statistics Netherlands.
Este programa ha sido diseñado especialmente para el análisis de la evolución de
poblaciones animales con series de datos de varios años, que permite considerar valores
intermedios “perdidos” que son recalculados en función de los datos de años anteriores
y posteriores y de la calidad de la población en otras estaciones en esa temporada.
TRIM genera un índice anual de abundancia para cada especie considerada, que se basa
en la regresión loglineal de Poisson, pero con la posibilidad de corregir dicha regresión
con correlaciones seriales a través de estimas de los parámetros del modelo, dado que
las series temporales de un año a otro no son totalmente independientes. Dichos índices
anuales se recalculan cada año en función de la nueva información incorporada, y el
programa permite analizar la evolución de las especies en relación con variables del
medio. Además, TRIM es la herramienta estadística cuyo uso recomienda el EBCC
(European Bird Census Council) para este tipo de análisis y es la que se aplica en el
Programa Pan-Europeo de seguimiento de aves reproductoras de BirdLife International,
cuyo diseño ha sido encomendado a la RSPB, con participación de otras sociedades
representadas en el EBCC (SEO/BirdLife entre ellas).
• Para obtener la evolución por grupos de especies (urbanas, de cultivos y forestales), se
ha realizado la media ponderada de los índices de las especies consideradas en cada
caso; se generaron dando peso a cada especie según la abundancia de cada especie. De
esta forma, influye en el índice tanto una especie escasa como una abundante, y éstas no
enmascaran la tendencia de las escasas.
4

3.1. Material entregado a cada colaborador
Para que el trabajo de campo sea fácilmente registrado y que permita el volcado de
la información a la base de datos central de una forma rápida e inequívoca se proporciona el
siguiente material (incluido en el Anexo I):
· Instrucciones. Incluyen todos los pasos que debe seguir el censador para realizar
correctamente el trabajo, tanto los planteamientos previos de selección y colocación de
estaciones en cada cuadrícula, como los puntos a tener en cuenta para realizar los
censos correctamente.
· Fichas de campo. Han sido realizadas para que los registros obtenidos se anoten de
forma rápida y ordenada y permitan perder el menor tiempo posible en el campo. En
ellas se registran los contactos de las aves en el campo.
· Fichas resumen. Útiles para pasar a limpio la información sobre aves obtenida en el
campo. Han sido diseñadas para facilitar la informatización rápida de los datos y el
archivado de seguridad de los mismos.
· Fichas de vegetación. Necesarias para indicar el ambiente presente en cada punto de
muestreo y los cambios registrados en cada temporada.
· Mapa de cuadrícula. Imprescindible para diseñar adecuadamente el censo dentro de la
unidad mediante el reconocimiento de sus límites, orografía, infraestructuras, etc.
· Mapa de hábitat. Muy necesario para la identificación de los ambientes que hay en la
zona y para la correcta colocación de las estaciones de escucha.
· Tabla de estaciones recomendadas. En función del cálculo de la superficie de cada
ambiente dentro de una cuadrícula, se ha realizado un cálculo del número de estaciones
que hay que realizar en cada tipo de hábitat dentro de la unidad. Este listado, en
principio, es el ideal y el censador situará o no el número de estaciones recomendadas
en cada ambiente en función de la accesibilidad a las distintas partes de la cuadrícula.
· Base de datos personal. Permite el volcado de los datos por cada colaborador. Este
aspecto, además de facilitar la inclusión de errores por parte de terceros, facilita que el
propio censador observe de año en año la evolución de cada especie en su cuadrícula así
como la evolución general de la misma.
5

· Guía interactiva de entrenamiento para los programas de seguimiento de aves
comunes en España de SEO/BirdLife. Se facilita este CD a todos los participantes con
el objetivo de que puedan mejorar la identificación de las aves por el canto y obtener
datos de calidad.
3.2. Cursos de perfeccionamiento y estandarización de censadores
Se realizan cursos todos los años para homogeneizar la capacidad de detección de
especies por parte de los participantes, así como el sistema de trabajo (uniformizar
descripciones de hábitat, estandarizar sistemas de establecimiento de bandas alrededor del
observador, establecer de forma común qué especies tener en cuenta en función de su
actividad, etc.).
El objetivo de estos cursos es que todo participante tenga bien claro su modo de
censar y que conozca los errores que se suelen cometer, tanto durante los muestreos, como
al cumplimentar las fichas. De esta forma se homogeneizan los datos entre todos los
participantes. Otro de los objetivos del curso es ampliar el conocimiento mediante jornadas
de campo donde se procede al entrenamiento en la identificación de las especies a través de
los cantos, que es el método de identificación básico para la participación en el programa.
El incremento de los conocimientos de los participantes y la aclaración de dudas o
eliminación de errores en la identificación, hace que la calidad del programa se incremente
año a año. Esto no afectará a los análisis realizados, pues cada colaborador está clasificado
en función de sus conocimientos y el porcentaje de especies que identifica, y los análisis se
realizan en función de estos conocimientos. Así, cuando se disponga de datos suficientes,
las especies más comunes, al ser detectadas por un mayor número de participantes de forma
fiable, tendrán un tamaño muestral diferente a las especies que son más difíciles de
identificar, para las que sólo se tendrá en cuenta las cuadrículas realizadas con personas con
nivel alto en la identificación. Estos cursos se han realizado en los últimos cinco años en el
País Vasco, esta temporada el curso tuvo lugar el 14 de abril.
6

4. RESULTADOS DE LOS CENSOS ENTRE 1998-2007
4.1. Cobertura de cuadrículas
Como se indica en la metodología, la unidad de muestreo es la cuadrícula UTM de
10 kilómetros de lado. Existen cuadrículas con tres grados de prioridad (Figura 1), para
poder realizar siempre muestreos homogéneamente repartidos en función de la superficie
de hábitat (Figura 2). Durante los primeros años se intentaron cubrir las cuadrículas de
prioridad 1, una vez cubiertas y a medida que aumenta el número de colaboradores, se
intenta cubrir la totalidad de la comunidad muestreando a continuación las zonas de
prioridad 2 y por último, las de prioridad 3. La cobertura en el año 2007 ha sido de 26
cuadrículas (Anexo II), repartidas en las distintas provincias y con distintos grados de
prioridad (Figura 3). No se han tenido en cuenta los datos de dos cuadrículas (WN2050 y
WN7090) porque no se disponía de los datos en el momento de hacer el informe.
La provincia con mayor cobertura de cuadrículas ha sido Álava con 16, seguida de
Vizcaya con 6 y Guipúzcoa con 4 (Tabla 1).
Figura 1. Cuadrículas seleccionadas en el País Vasco para ser muestreadas con prioridad.
7
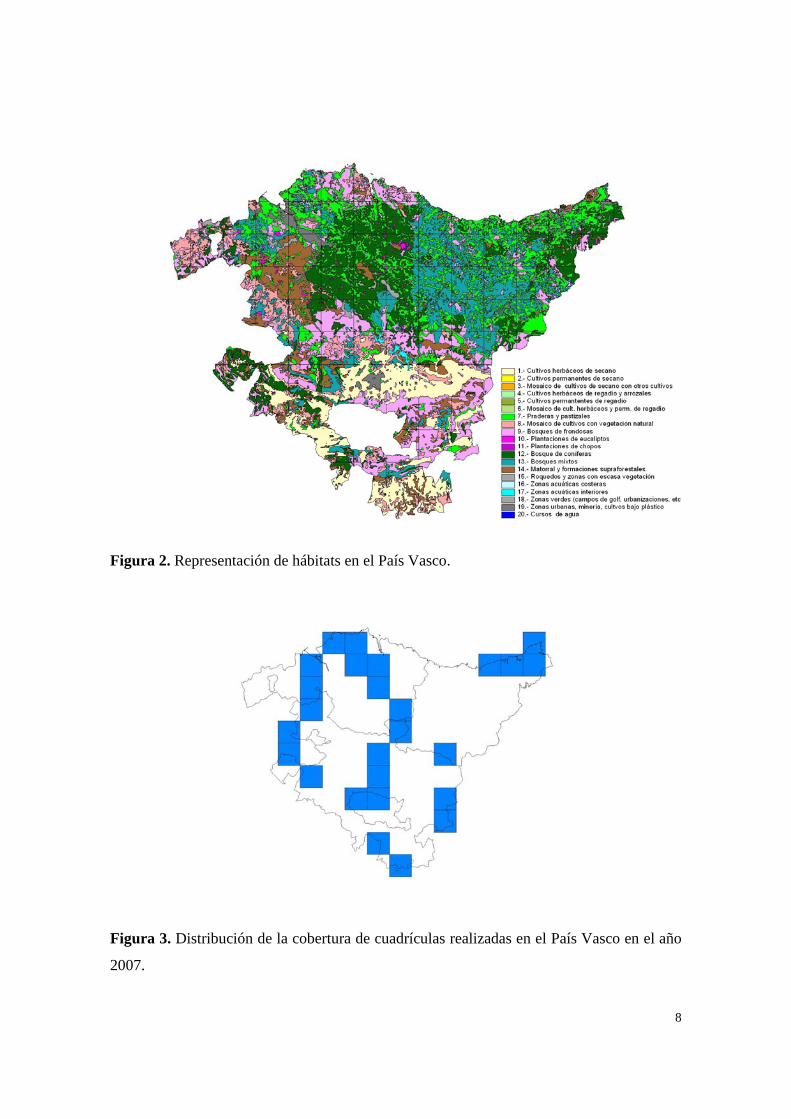
Figura 2. Representación de hábitats en el País Vasco.
Figura 3. Distribución de la cobertura de cuadrículas realizadas en el País Vasco en el año
2007.
8
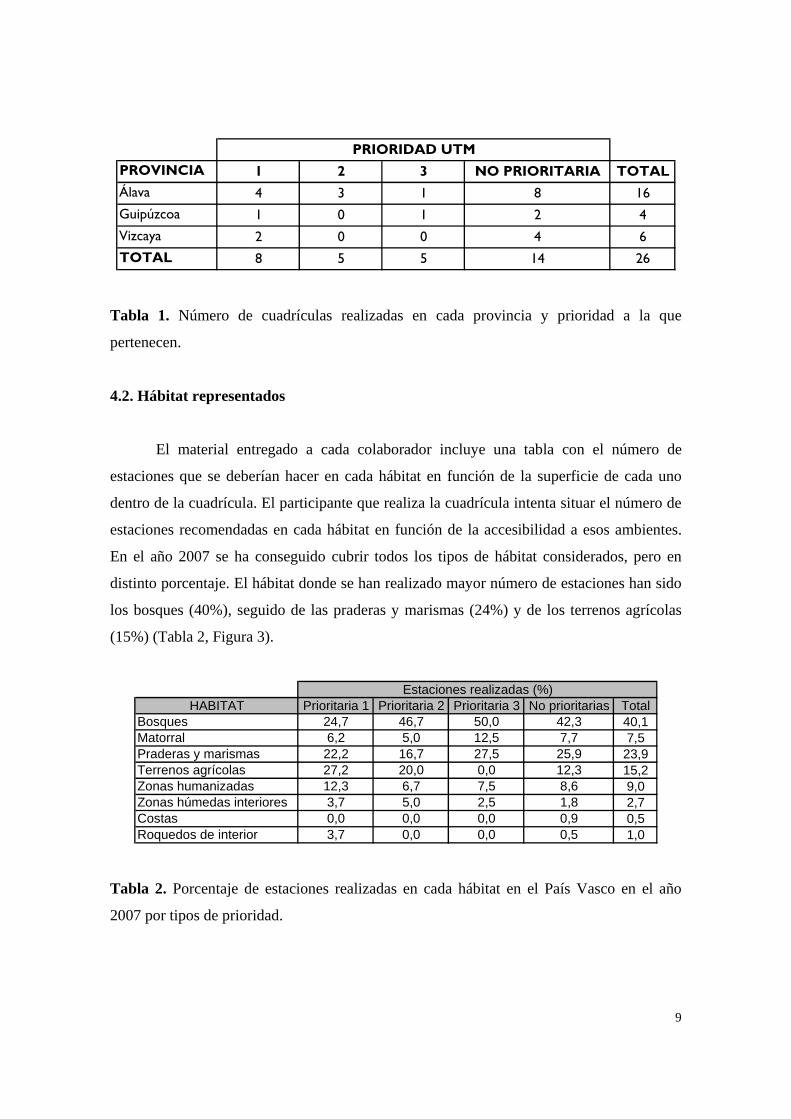
PRIORIDAD UTM PROVINCIA 1 2 3 NO PRIORITARIA TOTAL Álava 4 3 1 8 16 Guipúzcoa 1 0 1 2 4 Vizcaya 2 0 0 4 6 TOTAL 8 5 5 14 26
Tabla 1. Número de cuadrículas realizadas en cada provincia y prioridad a la que
pertenecen.
4.2. Hábitat representados
El material entregado a cada colaborador incluye una tabla con el número de
estaciones que se deberían hacer en cada hábitat en función de la superficie de cada uno
dentro de la cuadrícula. El participante que realiza la cuadrícula intenta situar el número de
estaciones recomendadas en cada hábitat en función de la accesibilidad a esos ambientes.
En el año 2007 se ha conseguido cubrir todos los tipos de hábitat considerados, pero en
distinto porcentaje. El hábitat donde se han realizado mayor número de estaciones han sido
los bosques (40%), seguido de las praderas y marismas (24%) y de los terrenos agrícolas
(15%) (Tabla 2, Figura 3).
Estaciones realizadas (%) HABITAT Prioritaria 1 Prioritaria 2 Prioritaria 3 No prioritarias Total
Bosques 24,7 46,7 50,0 42,3 40,1 Matorral 6,2 5,0 12,5 7,7 7,5 Praderas y marismas 22,2 16,7 27,5 25,9 23,9 Terrenos agrícolas 27,2 20,0 0,0 12,3 15,2 Zonas humanizadas 12,3 6,7 7,5 8,6 9,0 Zonas húmedas interiores 3,7 5,0 2,5 1,8 2,7 Costas 0,0 0,0 0,0 0,9 0,5 Roquedos de interior 3,7 0,0 0,0 0,5 1,0
Tabla 2. Porcentaje de estaciones realizadas en cada hábitat en el País Vasco en el año
2007 por tipos de prioridad.
9
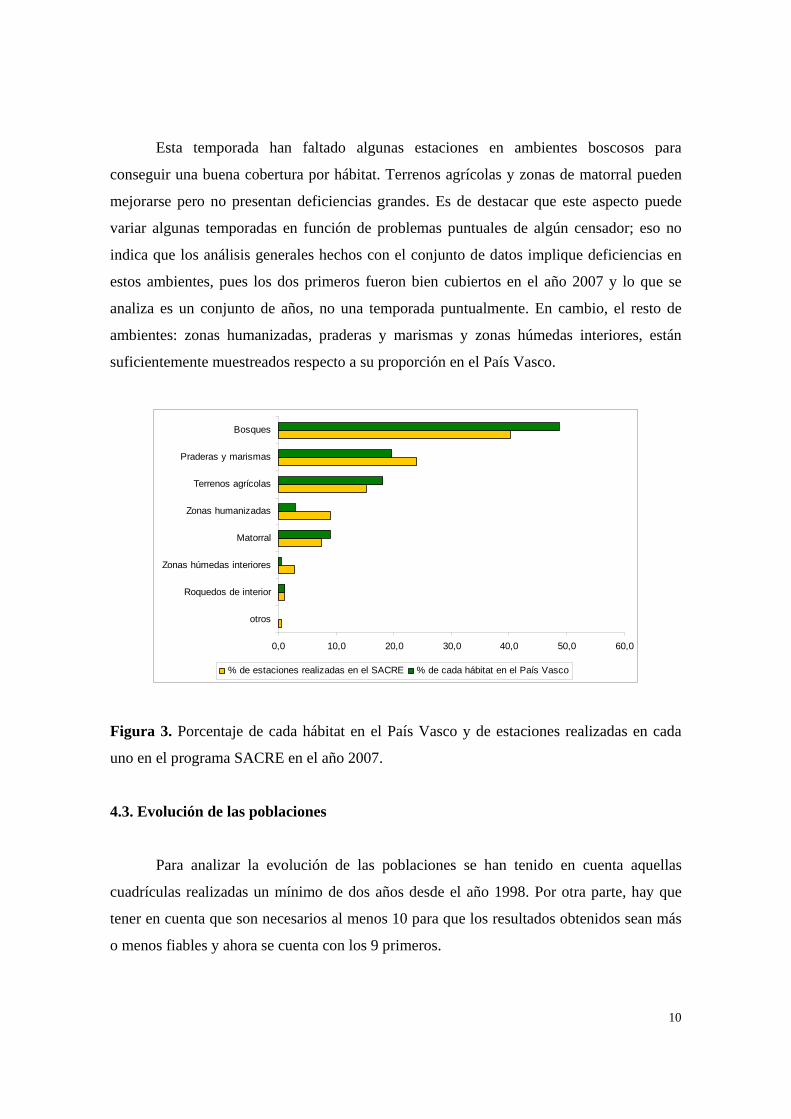
Esta temporada han faltado algunas estaciones en ambientes boscosos para
conseguir una buena cobertura por hábitat. Terrenos agrícolas y zonas de matorral pueden
mejorarse pero no presentan deficiencias grandes. Es de destacar que este aspecto puede
variar algunas temporadas en función de problemas puntuales de algún censador; eso no
indica que los análisis generales hechos con el conjunto de datos implique deficiencias en
estos ambientes, pues los dos primeros fueron bien cubiertos en el año 2007 y lo que se
analiza es un conjunto de años, no una temporada puntualmente. En cambio, el resto de
ambientes: zonas humanizadas, praderas y marismas y zonas húmedas interiores, están
suficientemente muestreados respecto a su proporción en el País Vasco.
i
i
i i l
0,0 10,0 20,0 30,0 40,0 50,0 60,0
otros
Roquedos de interior
Zonas húmedas nteriores
Matorral
Zonas humanizadas
Terrenos agrícolas
Praderas y mar smas
Bosques
% de estac ones real zadas en el SACRE % de cada hábitat en e País Vasco
Figura 3. Porcentaje de cada hábitat en el País Vasco y de estaciones realizadas en cada
uno en el programa SACRE en el año 2007.
4.3. Evolución de las poblaciones
Para analizar la evolución de las poblaciones se han tenido en cuenta aquellas
cuadrículas realizadas un mínimo de dos años desde el año 1998. Por otra parte, hay que
tener en cuenta que son necesarios al menos 10 para que los resultados obtenidos sean más
o menos fiables y ahora se cuenta con los 9 primeros.
10

En el anexo III se indica la tendencia obtenida para cada especie y en el anexo IV
las gráficas de evolución de cada una de ellas.
Los resultados indican un fuerte declive para cinco especies: buitrón (Cisticola
juncidis), grajilla (Corvus monedula), escribano cerillo (Emberiza citrinella), Lavandera
blanca (Motacilla alba) y gorrión molinero (Passer montanus). También se ha detectado
declive, pero moderado en carricero común (Acrocephalus scirpaceus), alondra común
(Alauda arvensis), Pardillo común (Carduelos cannabina), ruiseñor bastardo (Cettia cetti),
cuervo (Corvus corax), codorniz común (Coturnix coturnix), triguero (Emberiza calandra),
cogujada común (Galerita cristata), alcaudón dorsirrojo (Lanius collurio), papamoscas gris
(Muscicapa striata), gorrión común (Passer domesticus), tórtola europea (Streptopelia
turtur), curruca mosquitera (Sylvia borin) y zorzal común (Turdus philomelos). Aunque
también aparece el mosquitero común (Phylloscopus collybita) con moderado declive, no
debe considerarse como tal. Este resultado puede deberse por la consideración del
mosquitero ibérico (Phylloscopus ibericus) como nueva especie a partir de los últimos años.
Antes todos los registros de estaban referidos a P. collybita y a partir de la separación de
esta especie en dos, los registros de collybita fueron de repente inferiores y los de ibericus
se incrementaron. Por ello, también esta especie presenta un fuerte incremento.
Se ha detectado un fuerte incremento en milano negro (Milvus migrans) y colirrojo
tizón (Phoenicurus ochruros) y un incremento moderado en agateador común (Certhia
brachydactyla), paloma torcaz (Columba palumbus), cuco común (Cuculus canorus), avión
común (Delichon urbicum), pinzón vulgar (Fringilla coelebs), herrerillo común (Parus
caereleus), tórtola turca (Streptopelia decaocto) y mirlo común (Turdus merula).
Presentan una tendencia estable: perdiz común (Alectoris rufa), vencejo común
(Apus apus), jilguero (Carduelos carduelis), verderón común (Carduelis chloris),
golondrina común (Hirundo rustica), ruiseñor común (Luscinia megarhynchos), carbonero
común (Parus major), verdecillo (Serinus serinus), curruca capirotada (Sylvia atricapilla) y
chochín común (Troglodytes troglodytes).
11

Las demás especies muestran una tendencia incierta, por lo que en algunos casos,
aunque se han incluido los valores de tendencia en la tabla, no se muestra su gráfica.
En el anexo V se incluye la tendencia obtenida por grupos de especies en medio
urbano, de cultivos y forestales. En el anexo VI se muestran las gráficas de evolución de
cada grupo. Tanto las especies de medios urbanos como las de cultivos presentan un
porcentaje de cambio anual negativo. En cambio, en las especies de medios forestales este
cambio es positivo.
En el anexo VII se incluyen todos los análisis estadísticos realizados para cada
especie. En ellas se puede apreciar el tamaño muestral de contactos de cada especie (tantos
positivos como negativos), sus resultados en la recta de regresión y su grado de
significación.
12

5. EQUIPO DE TRABAJO
- Coordinación nacional: Virginia Escandell
- Coordinación regional en el País Vasco: José Antonio Gainzarain.
- Participantes: Alejandro Onrubia Baticón, Brian Webster, David Alday Irure, David
Cantalejo González, David Henderson, Ernesto Reyes Lara, Fernando Sánchez
Aranaz, Gorka Belamendia Cotorruelo, Iñaki Martínez, J. Miguel Devesa Pérez,
Javier Anderez Unquera, Javier García Sáez, Javier Uriarte Jairo, Jon Hidalgo
Múgica, Jorge Echegaray Fernández, José Ángel Isasi Zurbanobeaskoetxea, Jose
Antonio Gainzarain Diaz, José Antonio Herrero Fernández, José Mª Salazar Alonso,
José Manuel Cabrita Duarte, José María Fernández García, Josema Verdugo del
Val, Juan González De Suso Mata, Juan Mª Domínguez Robledo, Juan Puche
Fernandez, Julen Zuberogoitia, Leandro Arroyo Delgado, Luis Lobo Urrutia, Mario
Castaños Ortega, Merche Larrea Santa-Olalla, Miguel Ángel López De Armentia
Castillo, Nuria Busto González, Óscar Prada Campaña, Pedro Cruzado Díaz,
Ramón Elosegui Borinaga, Ramón Martín Martín, Ricardo Ibáñez García, Sergio de
Juan Zuloaga, Unai Fuente Gómez, Unai Oskoz Aretxaga, Victoria Badiola
Careaga, Xabier Iturrate, Xavier Erdozia Martínez.
13

6. AGRADECIMIENTOS
Agradecemos al Departamento de Medio Ambiente y Ordenación del Territorio del
Gobierno Vasco, haber confiado en SEO/BirdLife para utilizar los datos obtenidos
mediante el programa de seguimiento de aves comunes reproductoras (programa SACRE),
en la obtención de un indicador de biodiversidad de la CAPV, especialmente a José Mª
Fernández por facilitar que esto sea posible. También agradecemos a Pedro Silos la
creación de la base de datos que permite coordinar y gestionar informáticamente todos los
datos obtenidos.
14

Anexo I Material entregado a cada colaborador.

Anexo II COBERTURA EN EL PAÍS VASCO EN EL AÑO 2007. Cuadrículas realizadas, provincia y prioridad a la que pertenecen (N.P.: no prioritaria).
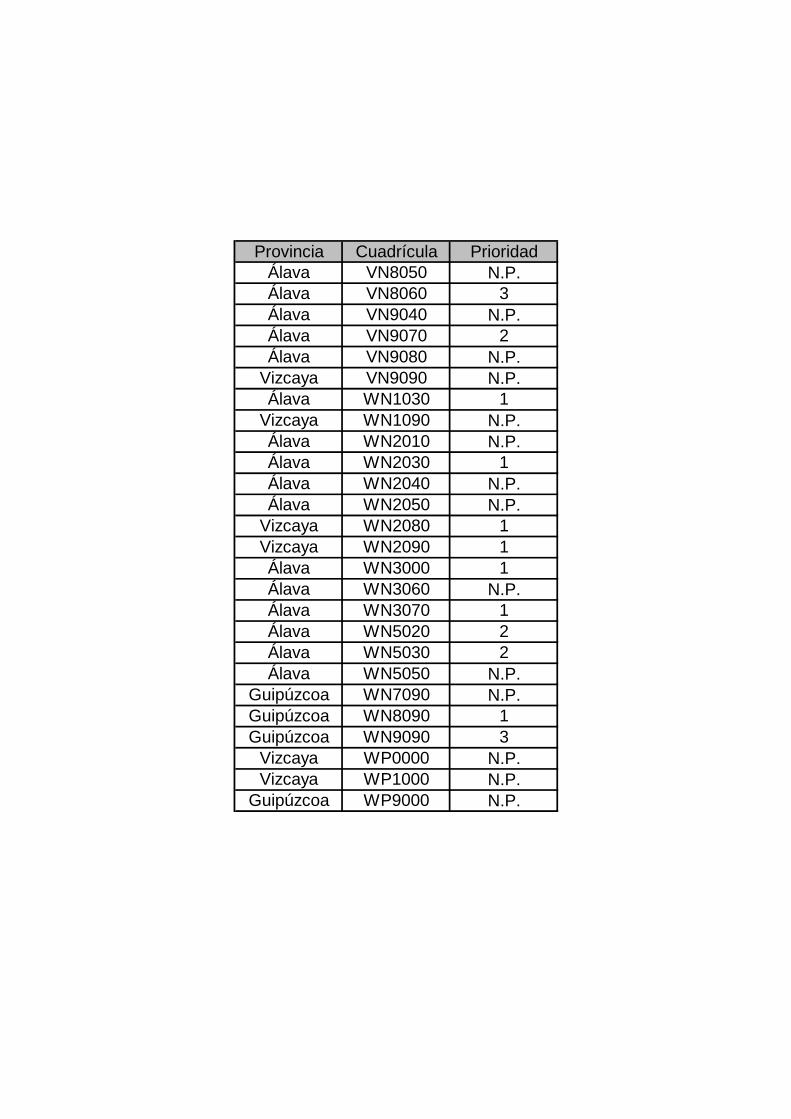
Provincia Cuadrícula Prioridad Álava VN8050 N.P. Álava VN8060 3 Álava VN9040 N.P. Álava VN9070 2 Álava VN9080 N.P.
Vizcaya VN9090 N.P. Álava WN1030 1
Vizcaya WN1090 N.P. Álava WN2010 N.P. Álava WN2030 1 Álava WN2040 N.P. Álava WN2050 N.P.
Vizcaya WN2080 1 Vizcaya WN2090 1 Álava WN3000 1 Álava WN3060 N.P. Álava WN3070 1 Álava WN5020 2 Álava WN5030 2 Álava WN5050 N.P.
Guipúzcoa WN7090 N.P. Guipúzcoa WN8090 1 Guipúzcoa WN9090 3
Vizcaya WP0000 N.P. Vizcaya WP1000 N.P.
Guipúzcoa WP9000 N.P.

Anexo III Índice de cambio obtenido para cada especie y tendencia general. (%: porcentaje de
cambio anual, máx y mín: valores máximo y mínimo del intervalo de confianza al 95%. Significación de las tendencias (Test de Wald): * p<0,05 ** p<0,01 ***p<0,001). Se
presentan en verde las especies con tendencia positiva, en rojo las especies con tendencia negativa, en azul las que tienen una tendencia estable y en gris aquellas cuya
tendencia obtenida es incierta.
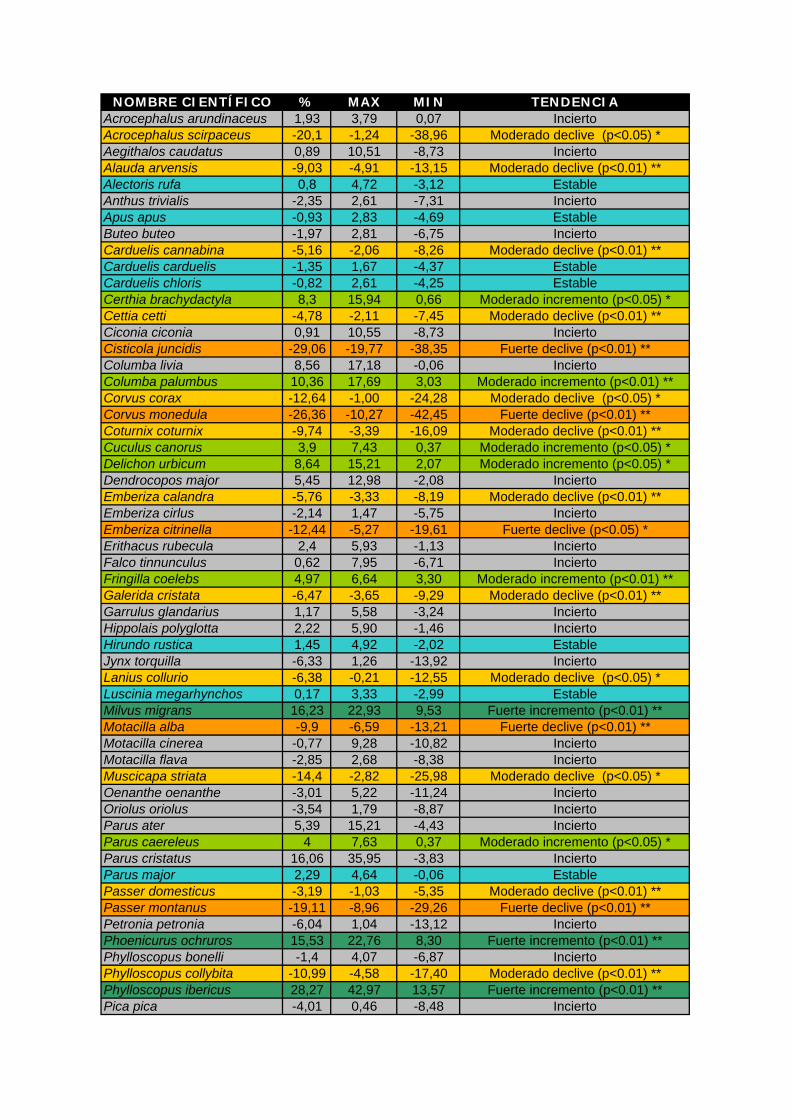
NOMBRE CIENTÍFICO % MAX MIN TENDENCIA Acrocephalus arundinaceus 1,93 3,79 0,07 Incierto Acrocephalus scirpaceus -20,1 -1,24 -38,96 Moderado declive (p<0.05) * Aegithalos caudatus 0,89 10,51 -8,73 Incierto Alauda arvensis -9,03 -4,91 -13,15 Moderado declive (p<0.01) ** Alectoris rufa 0,8 4,72 -3,12 Estable Anthus trivialis -2,35 2,61 -7,31 Incierto Apus apus -0,93 2,83 -4,69 Estable Buteo buteo -1,97 2,81 -6,75 Incierto Carduelis cannabina -5,16 -2,06 -8,26 Moderado declive (p<0.01) ** Carduelis carduelis -1,35 1,67 -4,37 Estable Carduelis chloris -0,82 2,61 -4,25 Estable Certhia brachydactyla 8,3 15,94 0,66 Moderado incremento (p<0.05) * Cettia cetti -4,78 -2,11 -7,45 Moderado declive (p<0.01) ** Ciconia ciconia 0,91 10,55 -8,73 Incierto Cisticola juncidis -29,06 -19,77 -38,35 Fuerte declive (p<0.01) ** Columba livia 8,56 17,18 -0,06 Incierto Columba palumbus 10,36 17,69 3,03 Moderado incremento (p<0.01) ** Corvus corax -12,64 -1,00 -24,28 Moderado declive (p<0.05) * Corvus monedula -26,36 -10,27 -42,45 Fuerte declive (p<0.01) ** Coturnix coturnix -9,74 -3,39 -16,09 Moderado declive (p<0.01) ** Cuculus canorus 3,9 7,43 0,37 Moderado incremento (p<0.05) * Delichon urbicum 8,64 15,21 2,07 Moderado incremento (p<0.05) * Dendrocopos major 5,45 12,98 -2,08 Incierto Emberiza calandra -5,76 -3,33 -8,19 Moderado declive (p<0.01) ** Emberiza cirlus -2,14 1,47 -5,75 Incierto Emberiza citrinella -12,44 -5,27 -19,61 Fuerte declive (p<0.05) * Erithacus rubecula 2,4 5,93 -1,13 Incierto Falco tinnunculus 0,62 7,95 -6,71 Incierto Fringilla coelebs 4,97 6,64 3,30 Moderado incremento (p<0.01) ** Galerida cristata -6,47 -3,65 -9,29 Moderado declive (p<0.01) ** Garrulus glandarius 1,17 5,58 -3,24 Incierto Hippolais polyglotta 2,22 5,90 -1,46 Incierto Hirundo rustica 1,45 4,92 -2,02 Estable Jynx torquilla -6,33 1,26 -13,92 Incierto Lanius collurio -6,38 -0,21 -12,55 Moderado declive (p<0.05) * Luscinia megarhynchos 0,17 3,33 -2,99 Estable Milvus migrans 16,23 22,93 9,53 Fuerte incremento (p<0.01) ** Motacilla alba -9,9 -6,59 -13,21 Fuerte declive (p<0.01) ** Motacilla cinerea -0,77 9,28 -10,82 Incierto Motacilla flava -2,85 2,68 -8,38 Incierto Muscicapa striata -14,4 -2,82 -25,98 Moderado declive (p<0.05) * Oenanthe oenanthe -3,01 5,22 -11,24 Incierto Oriolus oriolus -3,54 1,79 -8,87 Incierto Parus ater 5,39 15,21 -4,43 Incierto Parus caereleus 4 7,63 0,37 Moderado incremento (p<0.05) * Parus cristatus 16,06 35,95 -3,83 Incierto Parus major 2,29 4,64 -0,06 Estable Passer domesticus -3,19 -1,03 -5,35 Moderado declive (p<0.01) ** Passer montanus -19,11 -8,96 -29,26 Fuerte declive (p<0.01) ** Petronia petronia -6,04 1,04 -13,12 Incierto Phoenicurus ochruros 15,53 22,76 8,30 Fuerte incremento (p<0.01) ** Phylloscopus bonelli -1,4 4,07 -6,87 Incierto Phylloscopus collybita -10,99 -4,58 -17,40 Moderado declive (p<0.01) ** Phylloscopus ibericus 28,27 42,97 13,57 Fuerte incremento (p<0.01) ** Pica pica -4,01 0,46 -8,48 Incierto
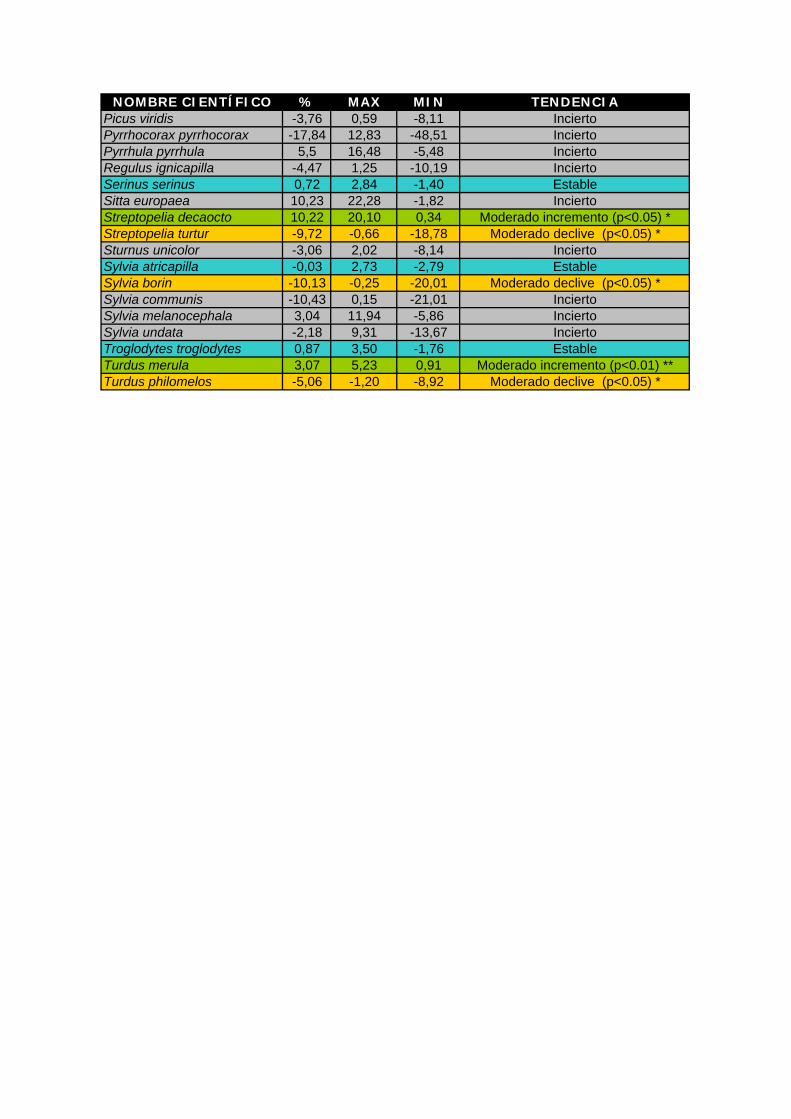
NOMBRE CIENTÍFICO % MAX MIN TENDENCIA Picus viridis -3,76 0,59 -8,11 Incierto Pyrrhocorax pyrrhocorax -17,84 12,83 -48,51 Incierto Pyrrhula pyrrhula 5,5 16,48 -5,48 Incierto Regulus ignicapilla -4,47 1,25 -10,19 Incierto Serinus serinus 0,72 2,84 -1,40 Estable Sitta europaea 10,23 22,28 -1,82 Incierto Streptopelia decaocto 10,22 20,10 0,34 Moderado incremento (p<0.05) * Streptopelia turtur -9,72 -0,66 -18,78 Moderado declive (p<0.05) * Sturnus unicolor -3,06 2,02 -8,14 Incierto Sylvia atricapilla -0,03 2,73 -2,79 Estable Sylvia borin -10,13 -0,25 -20,01 Moderado declive (p<0.05) * Sylvia communis -10,43 0,15 -21,01 Incierto Sylvia melanocephala 3,04 11,94 -5,86 Incierto Sylvia undata -2,18 9,31 -13,67 Incierto Troglodytes troglodytes 0,87 3,50 -1,76 Estable Turdus merula 3,07 5,23 0,91 Moderado incremento (p<0.01) ** Turdus philomelos -5,06 -1,20 -8,92 Moderado declive (p<0.05) *

Anexo IV Gráficas de evolución obtenidas para cada especie entre los años 1998-2007 ordenadas alfabéticamente. Se presentan en verde las especies con tendencia positiva, en rojo las especies con tendencia negativa, en azul las que tienen una tendencia estable y en gris
aquellas cuya tendencia obtenida es incierta.
Acrocephalus arundinaceus
-42
13,5
53,3
-25
-25
86,9
-27,2-36,1
54,6
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Acrocephalus scirpaceus
-74,8 -53,1 -90,6-53,3
-81,1
-29,7
-7,10,0
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
Porc
enta
je d
e ca
mbi
o (1
998=
0)
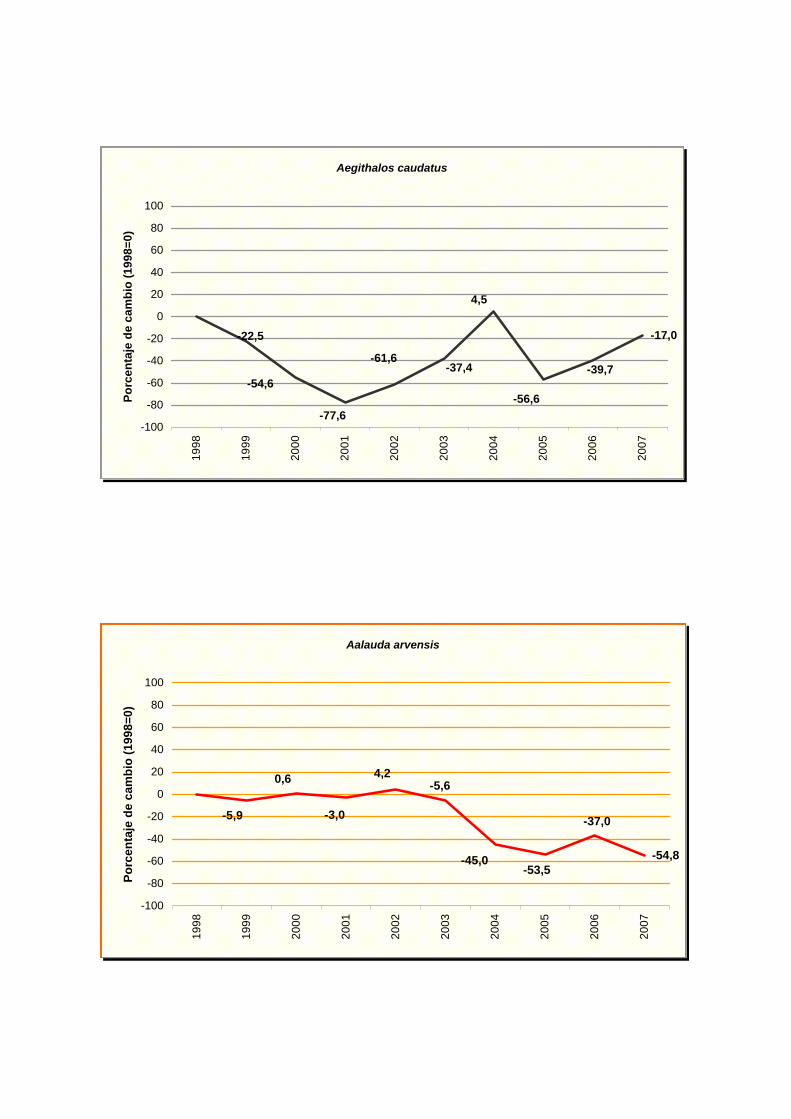
Aegithalos caudatus
-17,0-22,5
-54,6
-61,6
4,5
-37,4 -39,7
-77,6 -56,6
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Aalauda arvensis
-54,8
-5,9
0,6 4,2
-45,0
-5,6
-37,0-3,0
-53,5
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
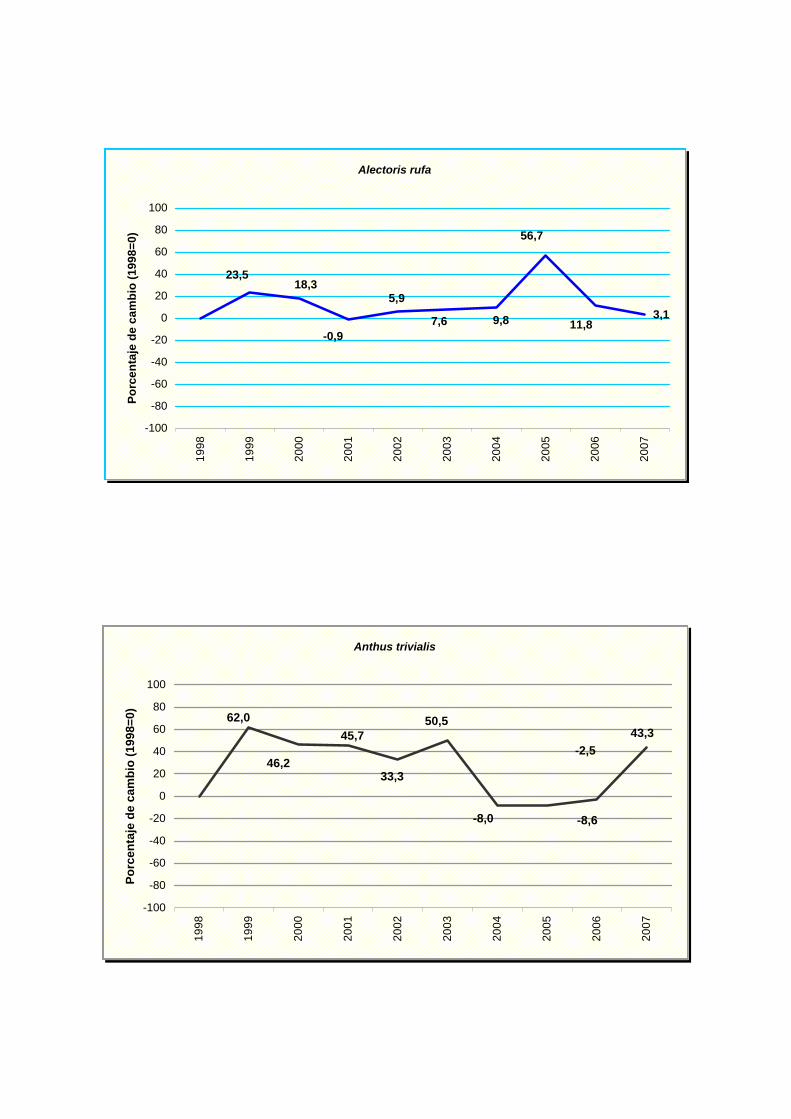
Alectoris rufa
3,1
56,7
-0,9 11,87,6 9,8
5,9 18,3
23,5
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Anthus trivialis
43,3
-8,6
45,7 -2,5
50,5
-8,0
33,3 46,2
62,0
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
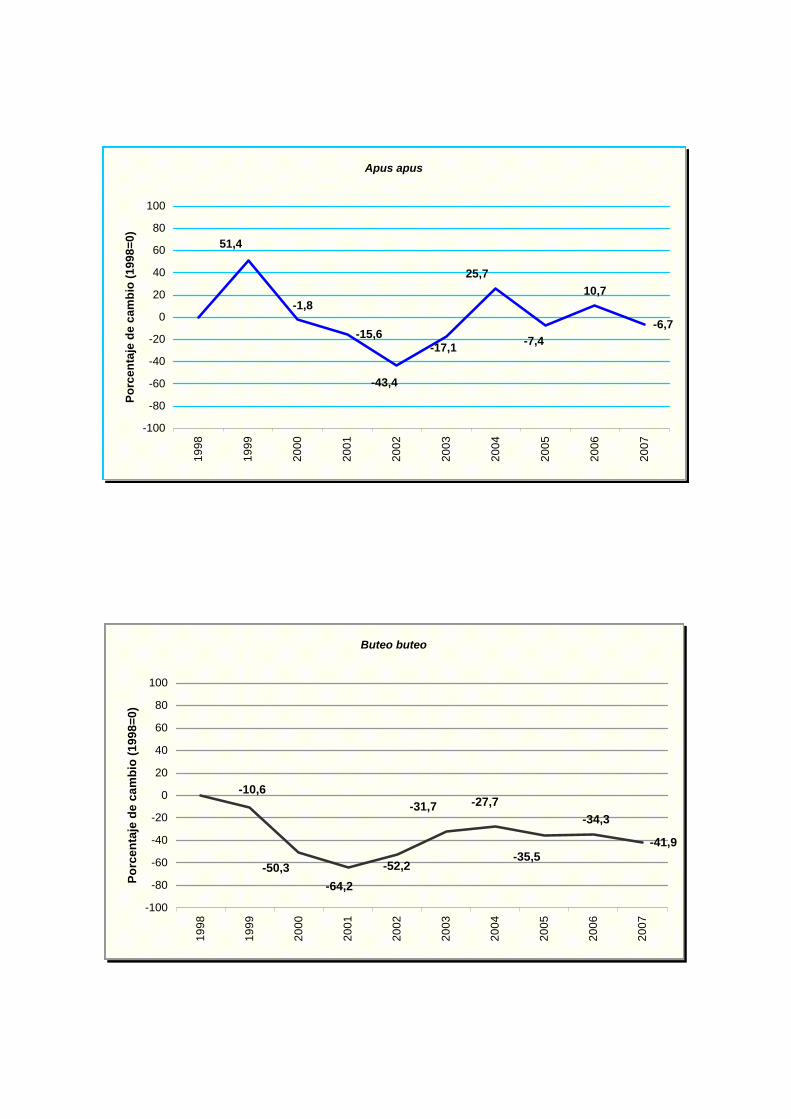
Apus apus
-15,6 -6,7
-7,4
10,7
-17,1
25,7
-43,4
-1,8
51,4
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Buteo buteo
-41,9 -35,5
-64,2
-34,3 -31,7 -27,7
-52,2-50,3
-10,6
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
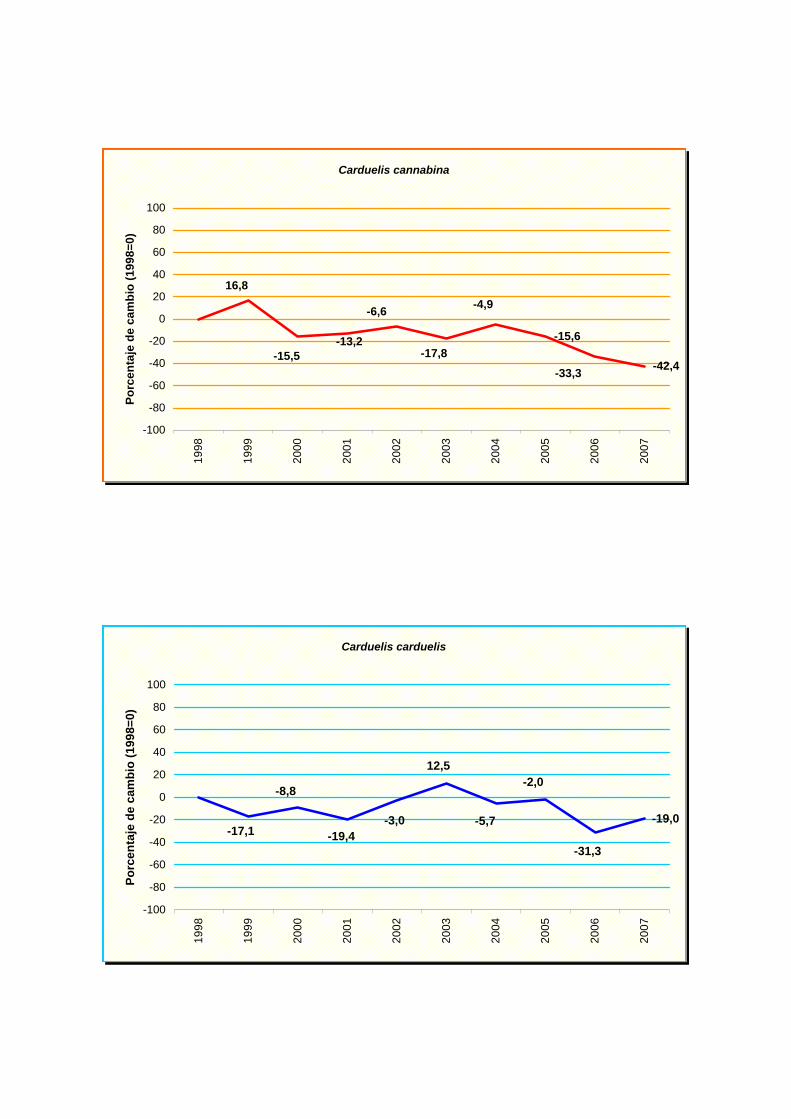
Carduelis cannabina
-15,6
-42,4
-13,2
-33,3 -17,8
-4,9-6,6
-15,5
16,8
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Carduelis carduelis
-19,0 -19,4
-2,0
-17,1
-8,8
-3,0 -5,7
12,5
-31,3
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
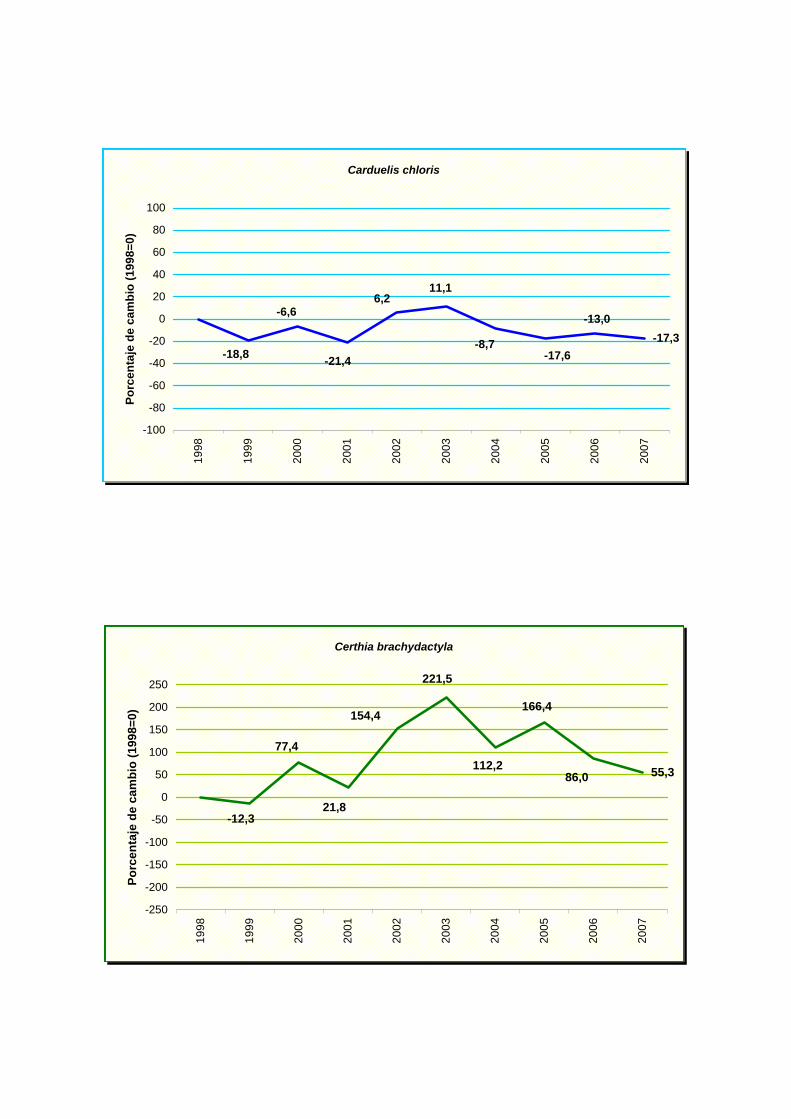
Carduelis chloris
-17,3
-21,4 -17,6-18,8
-6,6 6,2
-8,7
11,1
-13,0
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Certhia brachydactyla
55,3
166,4
21,8-12,3
77,4
154,4
112,2
221,5
86,0
-250
-200
-150
-100
-50
0
50
100
150
200
250
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
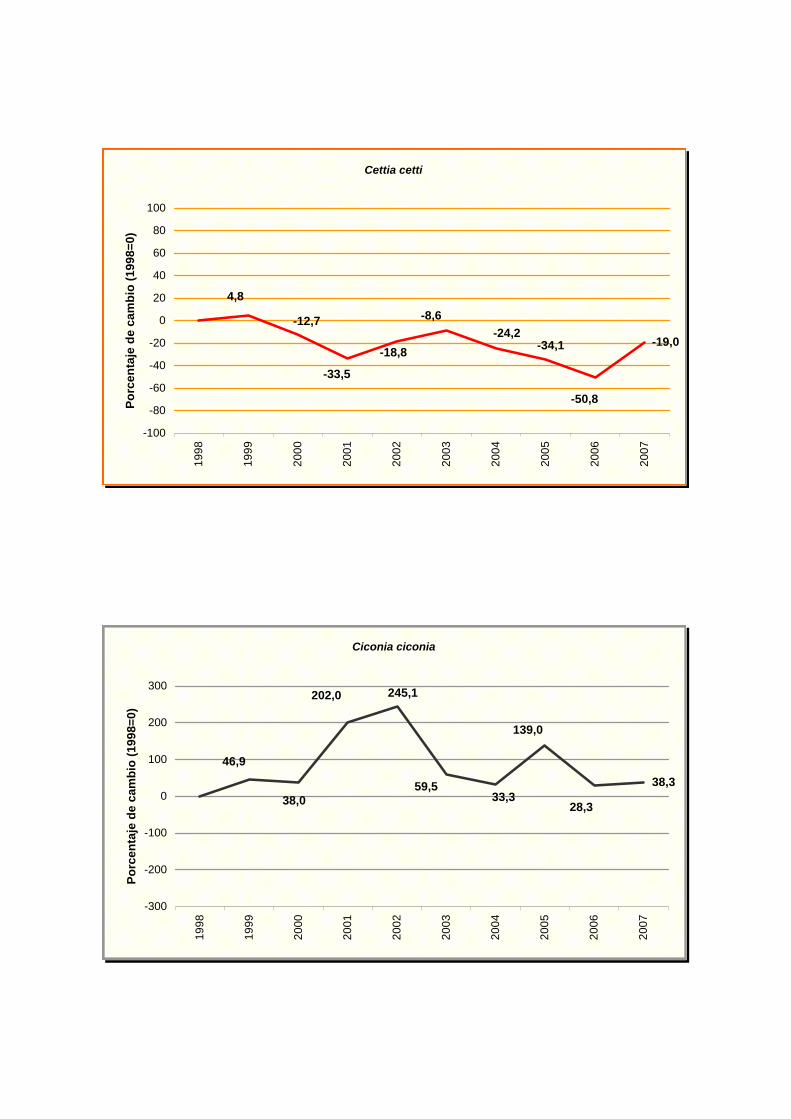
Cettia cetti
-19,0-34,1
-33,5
4,8
-12,7
-18,8 -24,2
-8,6
-50,8
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Ciconia ciconia
38,3
139,0
202,0
46,9
38,0
245,1
33,359,5
28,3
-300
-200
-100
0
100
200
300
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
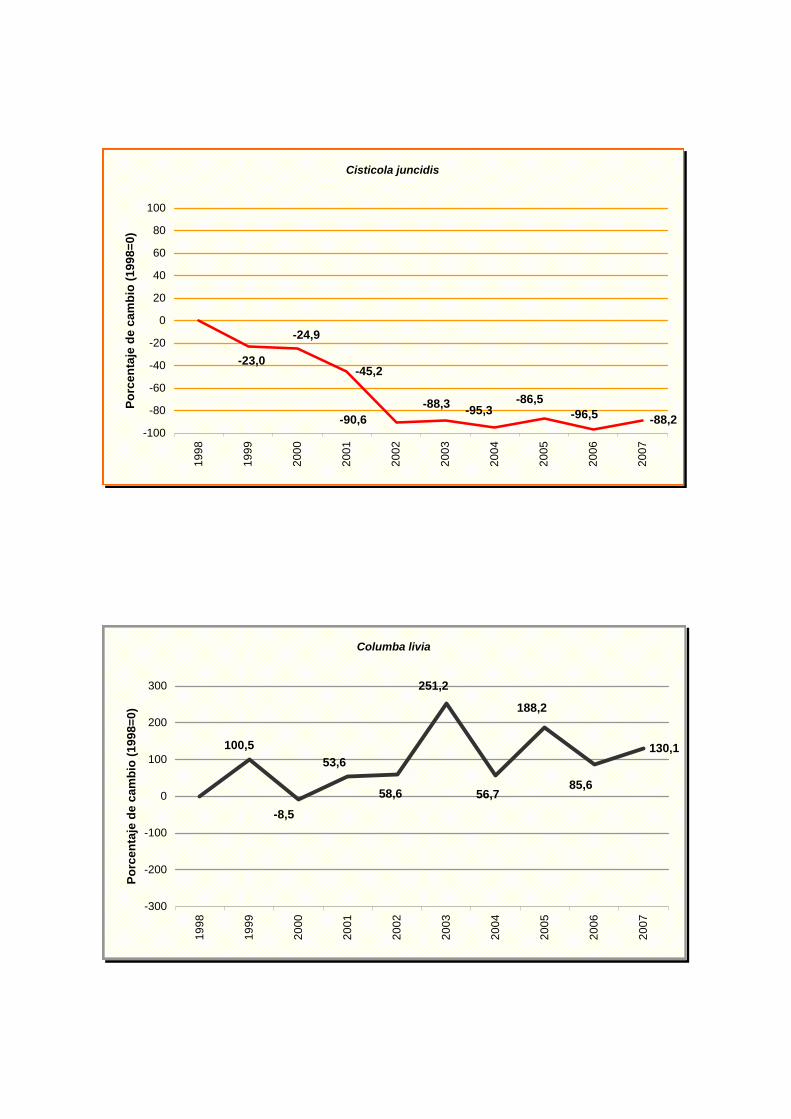
Cisticola juncidis
-45,2
-88,2 -86,5
-23,0
-24,9
-90,6 -95,3-88,3
-96,5 -100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Columba livia
130,1100,5
-8,5
58,6 56,7
251,2
85,6
53,6
188,2
-300
-200
-100
0
100
200
300
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
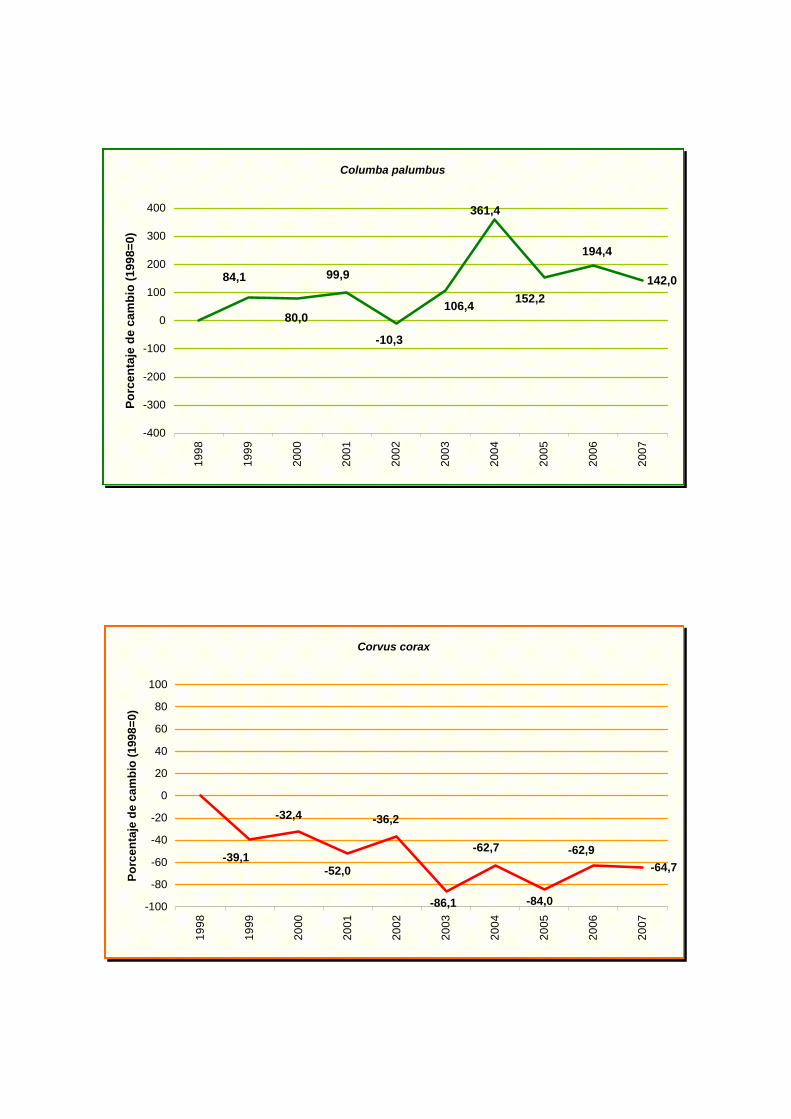
Columba palumbus
142,099,9
152,2
84,1
80,0
-10,3
361,4
106,4
194,4
-400
-300
-200
-100
0
100
200
300
400 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Corvus corax
-64,7
-84,0
-52,0 -39,1
-32,4 -36,2
-62,7
-86,1
-62,9
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
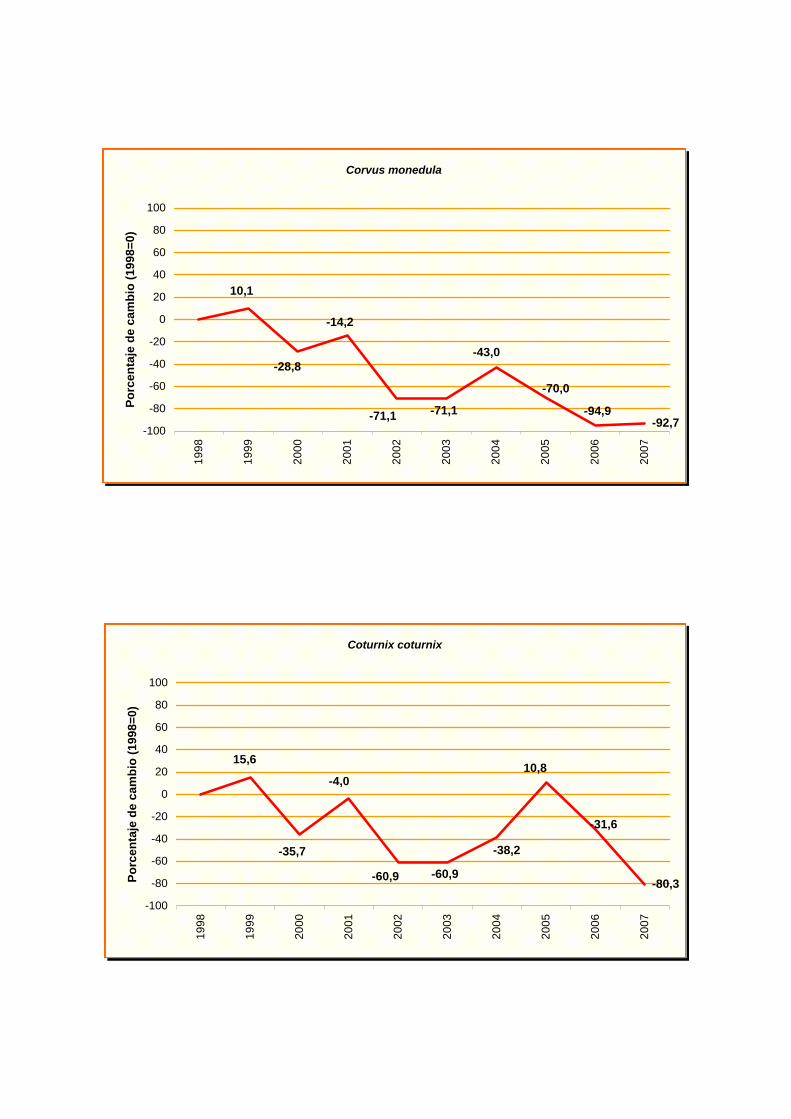
Corvus monedula
-92,7
-70,0
-14,2
-94,9-71,1
-43,0
-71,1
-28,8
10,1
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Coturnix coturnix
-80,3
10,8 -4,0
15,6
-35,7
-60,9
-38,2
-60,9
-31,6
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
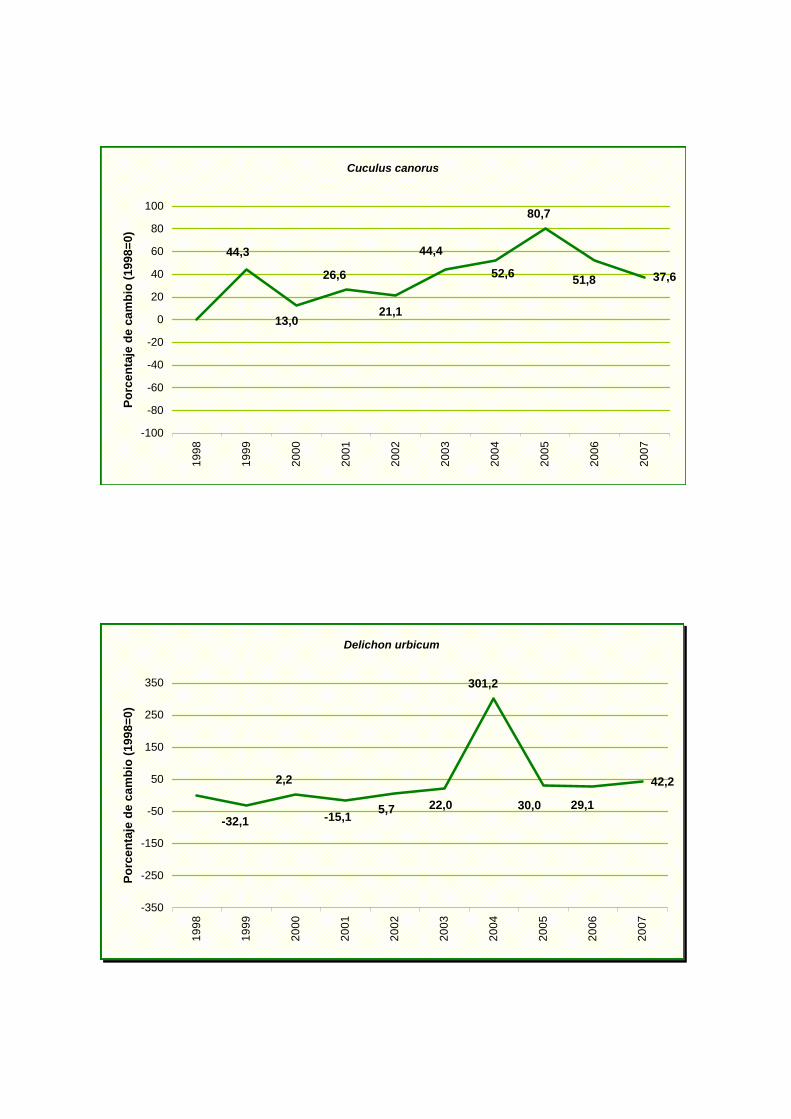
Cuculus canorus
37,651,8
44,4
52,6
21,113,0
44,3
26,6
80,7
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Delichon urbicum
42,2
30,0-15,1-32,1
2,2
5,7
301,2
22,0 29,1
-350
-250
-150
-50
50
150
250
350
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
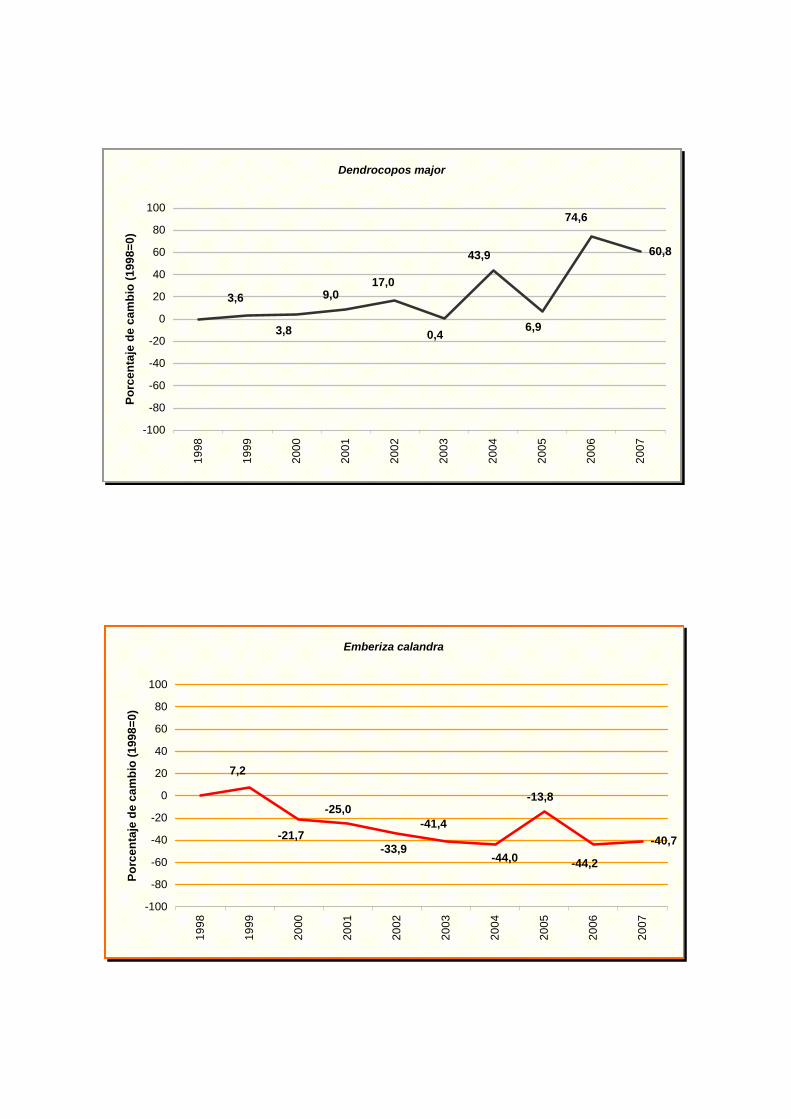
Dendrocopos major
60,8
74,6
0,4
43,9
17,0
3,8
3,6 9,0
6,9
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Emberiza calandra
-40,7
-44,2
-41,4
-44,0-33,9
-21,7
7,2
-25,0 -13,8
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
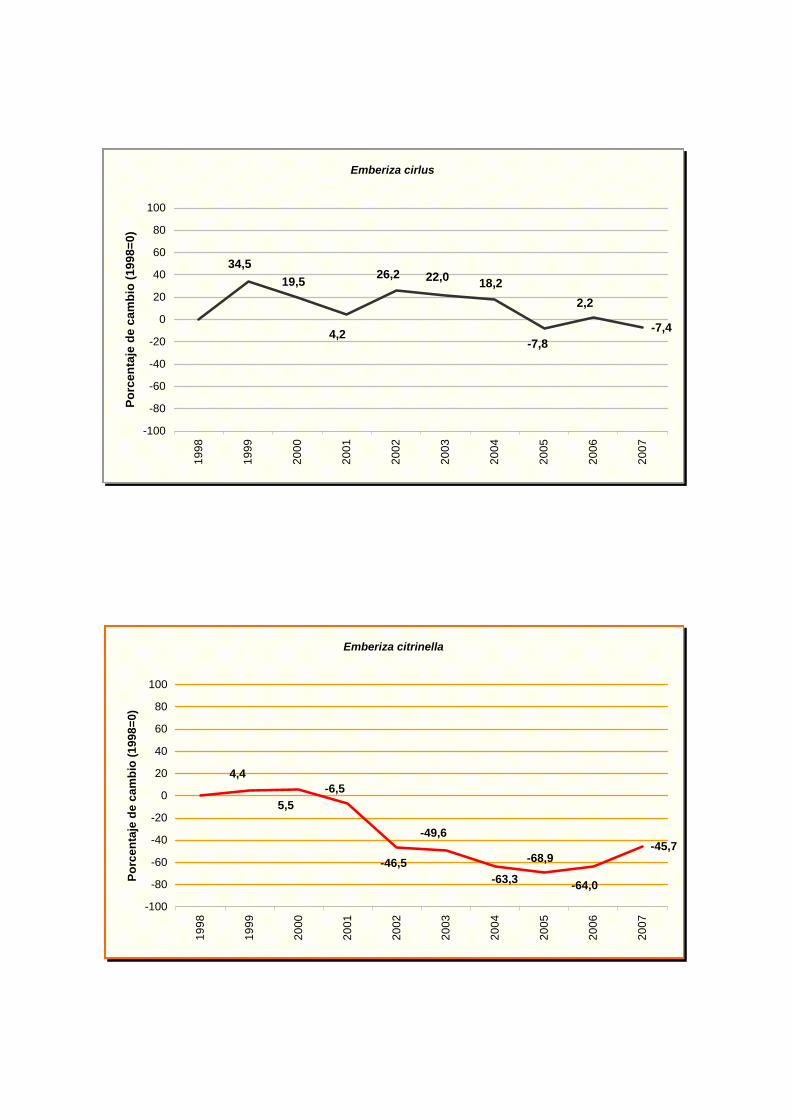
Emberiza cirlus
-7,4 -7,8
4,2
34,5 19,5 26,2
18,222,0
2,2
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Emberiza citrinella
-45,7
-64,0
-49,6
-63,3 -46,5
5,5
4,4 -6,5
-68,9
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
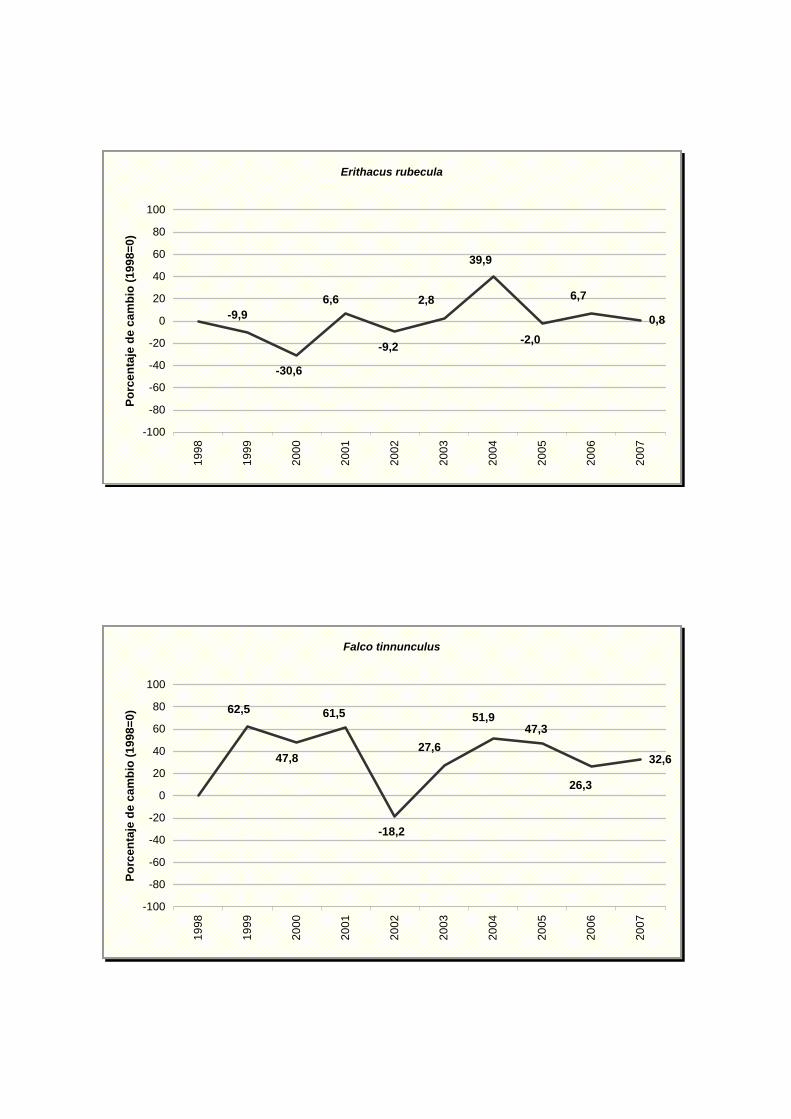
Erithacus rubecula
0,8
6,72,8
39,9
-9,2
-30,6
-9,9 6,6
-2,0
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Falco tinnunculus
32,6
26,3
27,6
51,9
-18,2
47,8
62,5 61,5 47,3
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
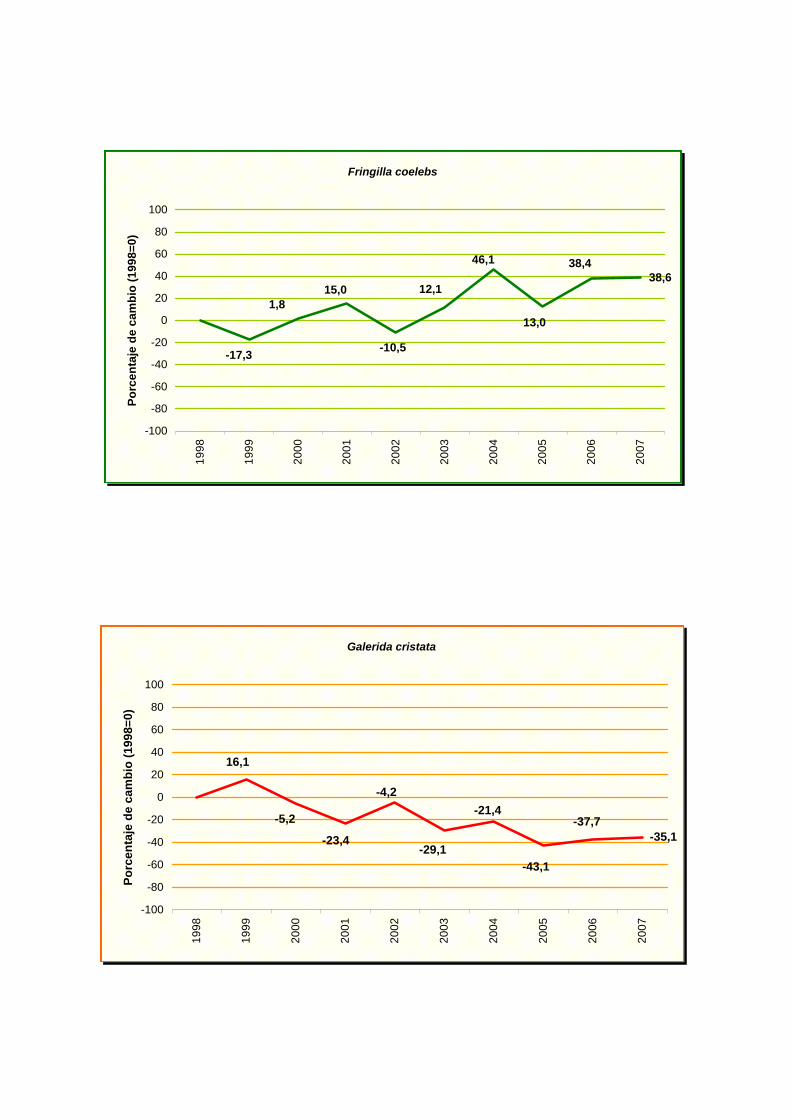
Fringilla coelebs
38,6 38,4
12,1
46,1
-10,5
1,8
-17,3
15,0
13,0
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Galerida cristata
-35,1
-43,1
-23,4
16,1
-5,2
-4,2 -21,4
-29,1
-37,7
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
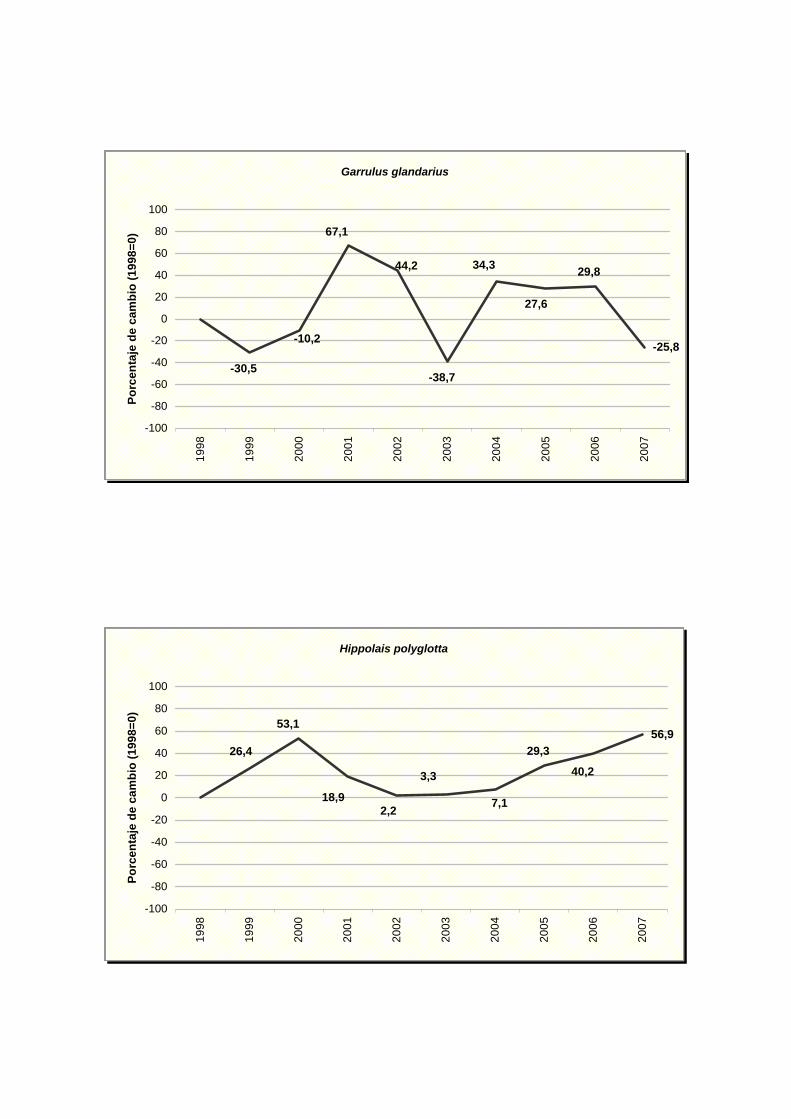
Garrulus glandarius
-25,8
27,6
67,1
-30,5
-10,2
44,2 34,3
-38,7
29,8
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Hippolais polyglotta
56,9
40,23,3
7,12,2
53,1
26,4
18,9
29,3
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
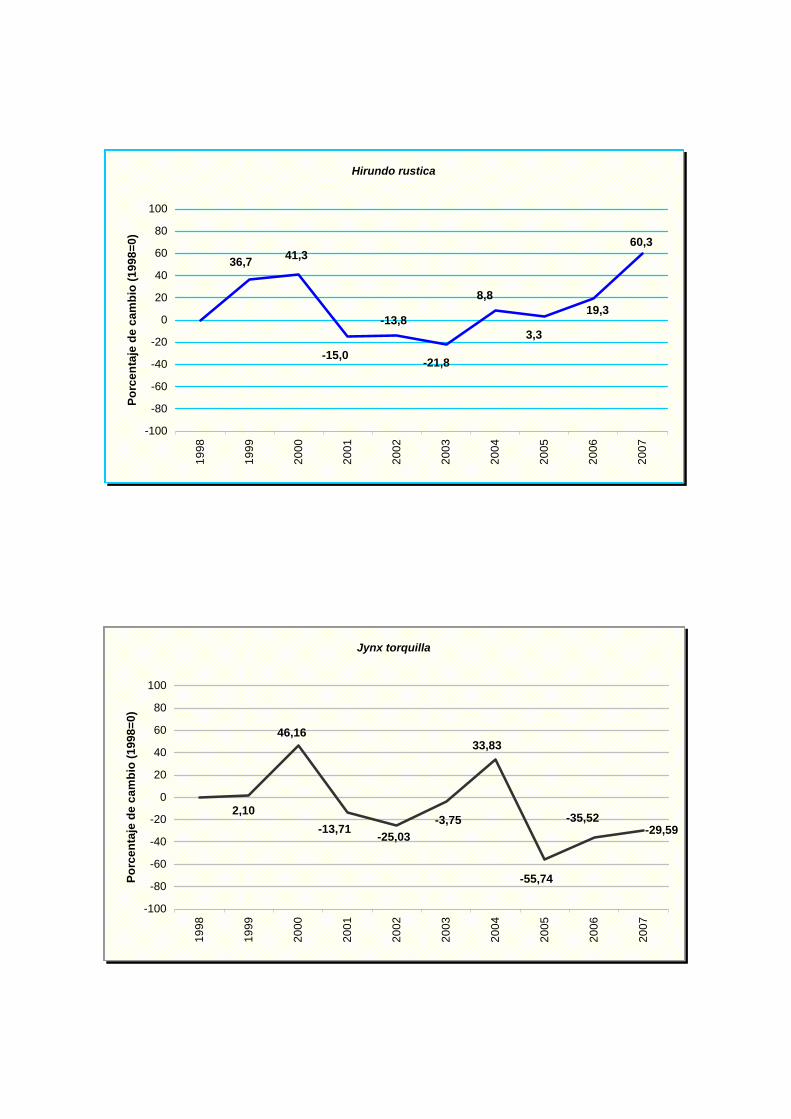
Hirundo rustica
60,3
19,3
-21,8
8,8
-13,8
41,336,7
-15,0
3,3
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Jynx torquilla
-29,59
-55,74
-13,71 2,10
46,16
-25,03
33,83
-3,75 -35,52
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
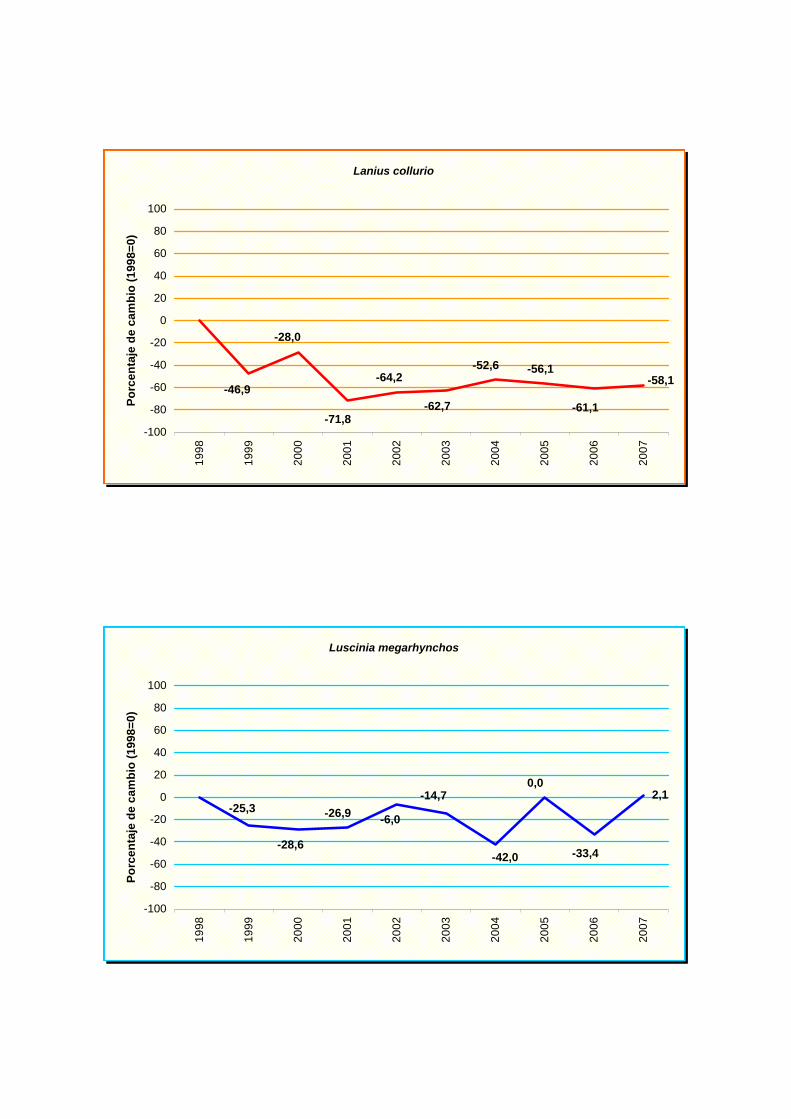
Lanius collurio
-58,1
-61,1-62,7
-52,6 -64,2
-28,0
-46,9
-71,8
-56,1
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Luscinia megarhynchos
2,1
-33,4
-14,7
-42,0
-6,0
-28,6
-25,3 -26,9
0,0
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
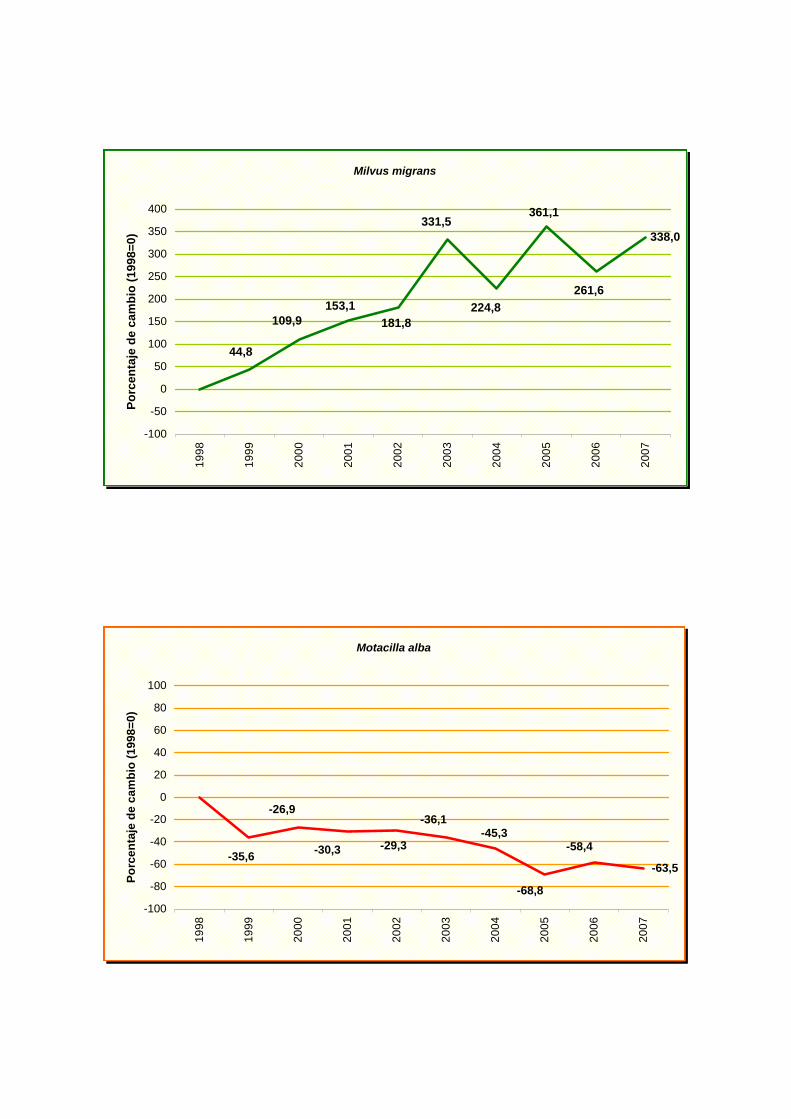
Milvus migrans
338,0
261,6
331,5
224,8 181,8109,9
44,8
153,1
361,1
-100
-50
0
50
100
150
200
250
300
350
400 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Motacilla alba
-63,5
-68,8
-30,3-35,6
-26,9
-29,3 -45,3
-36,1
-58,4
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
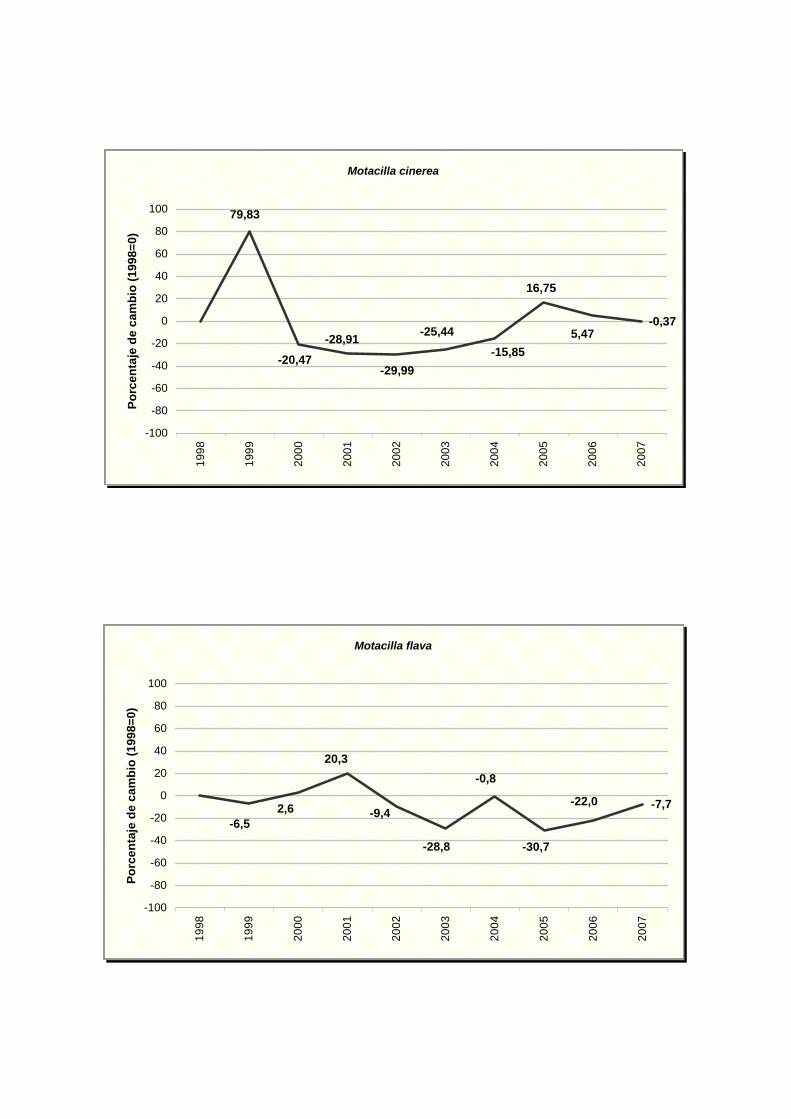
Motacilla cinerea
-0,37 5,47-25,44
-15,85 -29,99
-20,47
79,83
-28,91
16,75
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Motacilla flava
-7,7-22,0
-28,8
-0,8
-9,42,6 -6,5
20,3
-30,7
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
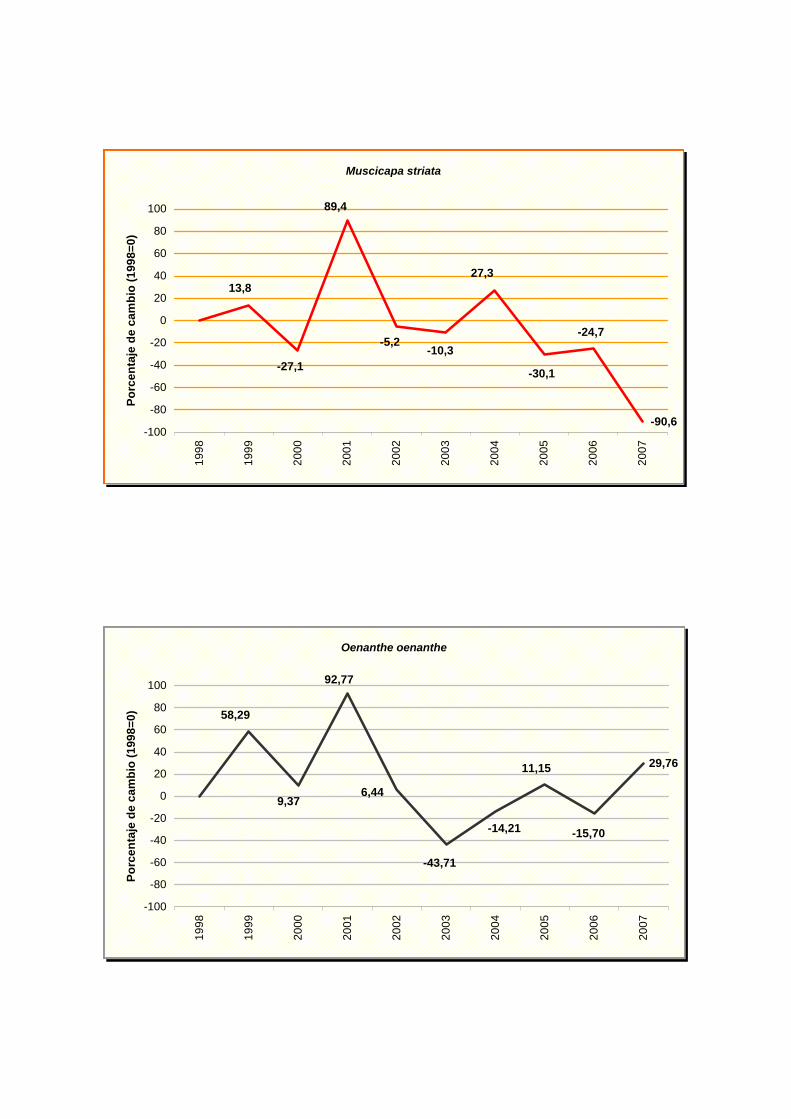
Muscicapa striata
-90,6
-24,7 -10,3
27,3
-5,2
-27,1
13,8
89,4
-30,1
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Oenanthe oenanthe
29,76
-15,70
-43,71
-14,21
6,449,37
58,29
92,77
11,15
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
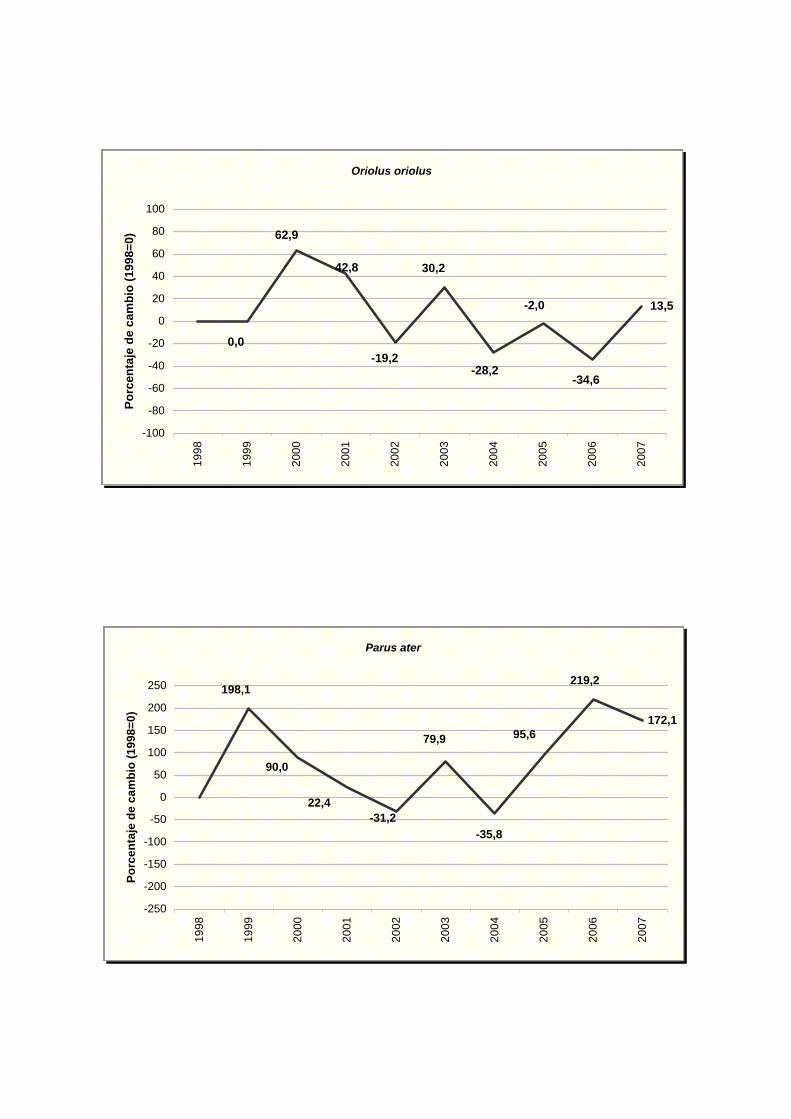
Oriolus oriolus
13,5-2,0
42,8
0,0
62,9
-19,2 -28,2
30,2
-34,6
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Parus ater
172,1 95,6
22,4
198,1
90,0
-31,2 -35,8
79,9
219,2
-250
-200
-150
-100
-50
0
50
100
150
200
250
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
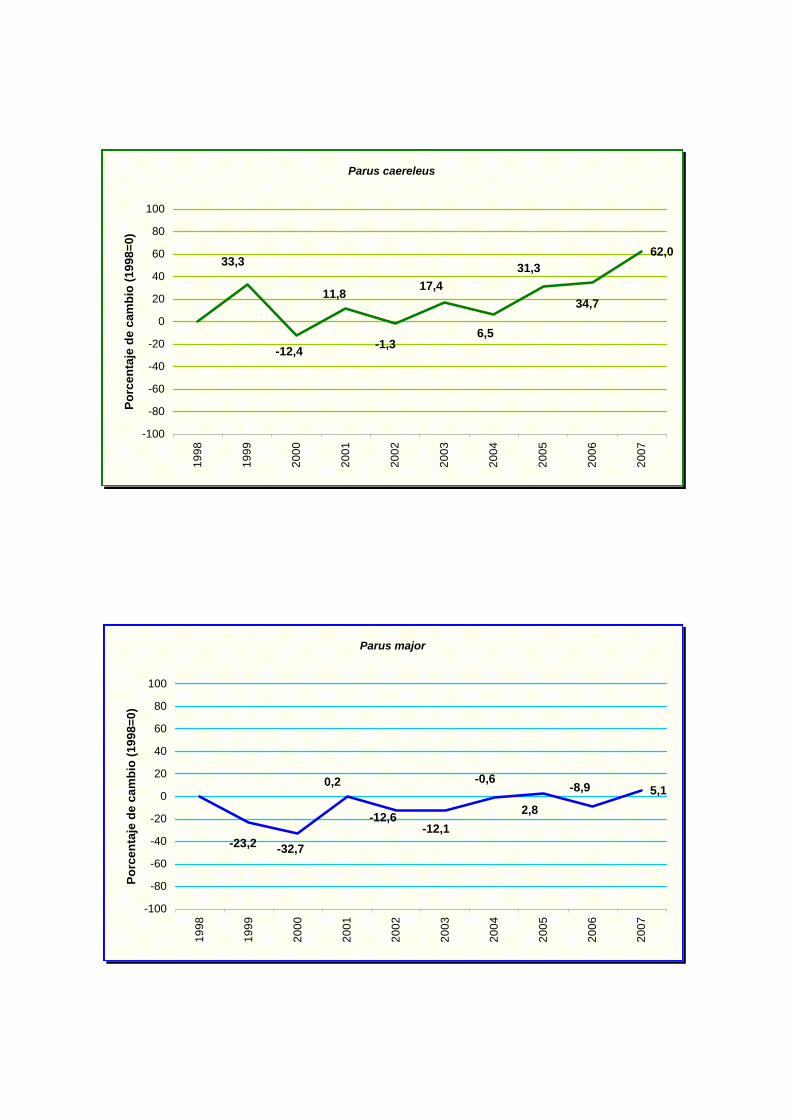
Parus caereleus
62,0 31,3
11,8
33,3
-12,4 -1,3 6,5
17,4 34,7
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Parus major
5,1 2,8
0,2
-23,2 -32,7
-12,6
-0,6
-12,1
-8,9
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
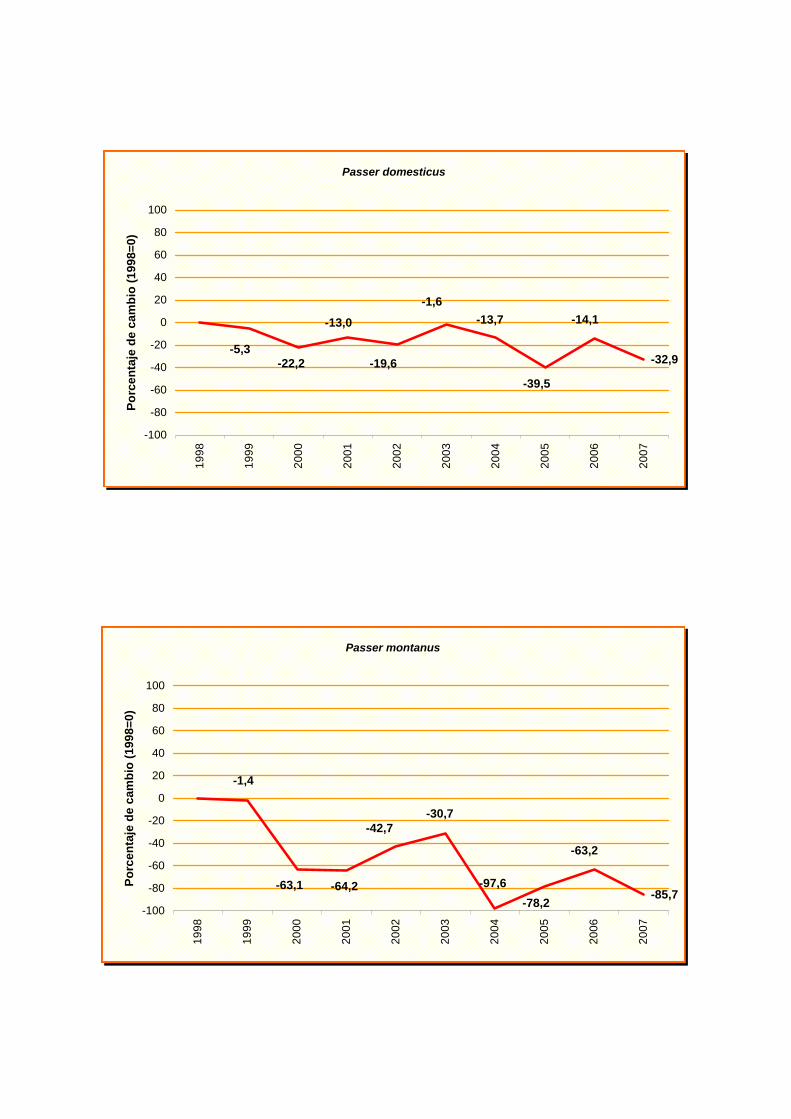
Passer domesticus
-32,9
-39,5
-13,0
-5,3 -22,2 -19,6
-13,7 -1,6
-14,1
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Passer montanus
-85,7-78,2
-64,2
-1,4
-63,1
-42,7
-97,6
-30,7
-63,2
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
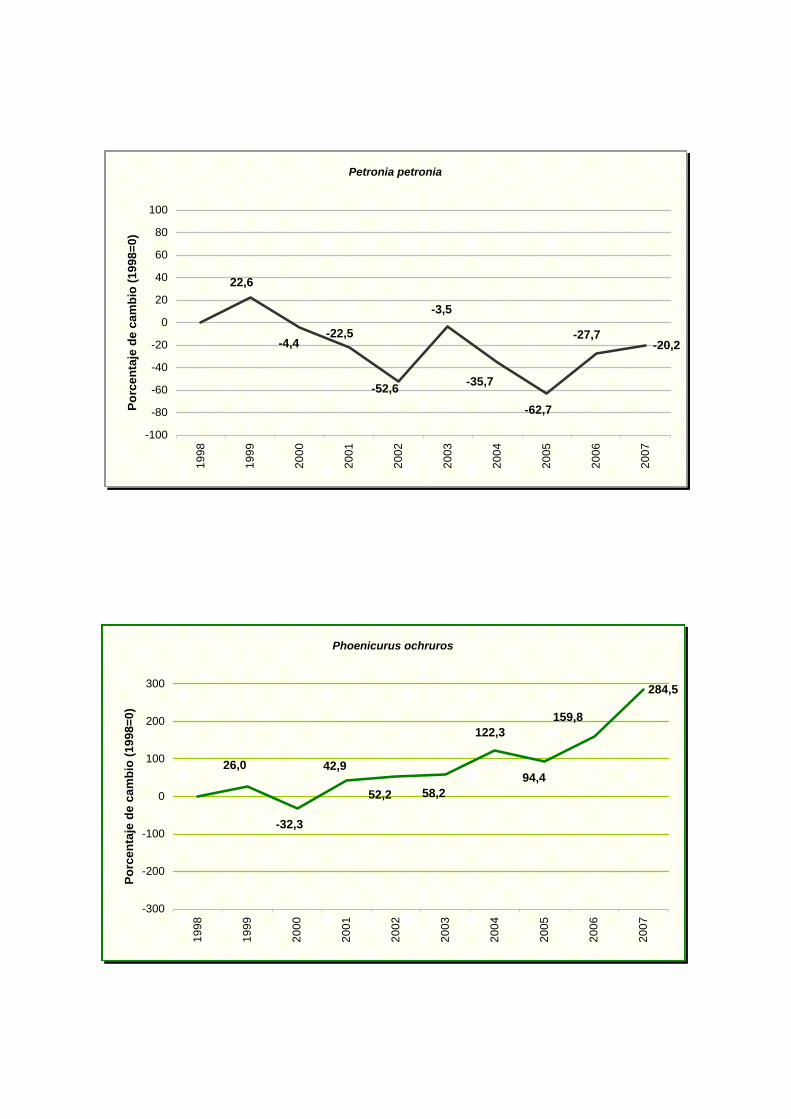
Petronia petronia
-20,2
-62,7
-22,5
22,6
-4,4
-52,6 -35,7
-3,5
-27,7
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Phoenicurus ochruros
284,5
94,442,926,0
-32,3
52,2
122,3
58,2
159,8
-300
-200
-100
0
100
200
300
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
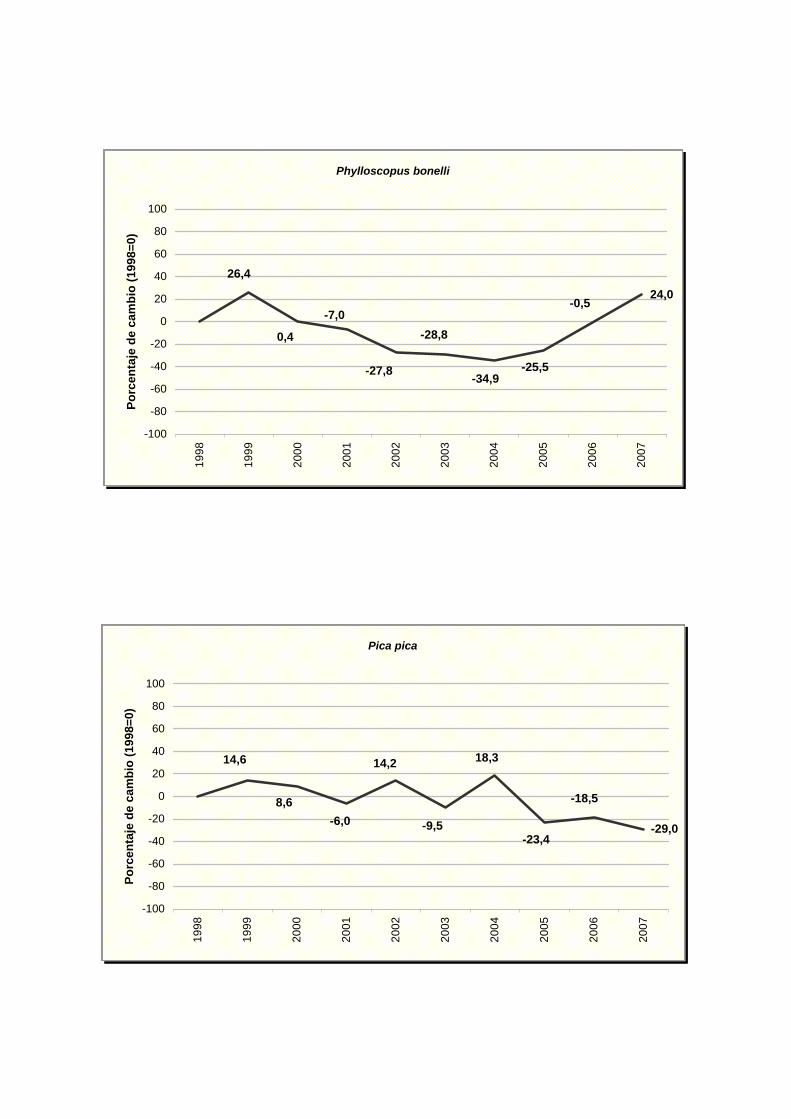
Phylloscopus bonelli
24,0-0,5
-28,8
-34,9-27,8
0,4
26,4
-7,0
-25,5
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Pica pica
-29,0-23,4
-6,0
14,6
8,6
14,2 18,3
-9,5
-18,5
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
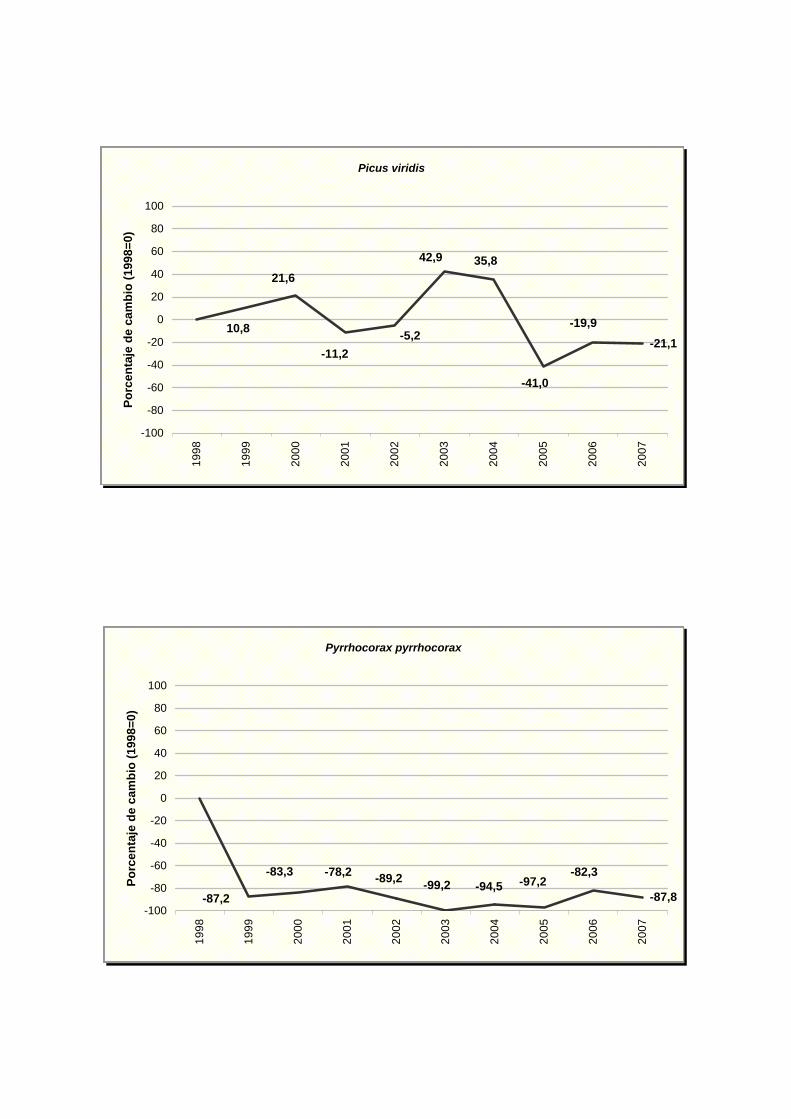
Picus viridis
-21,1 -19,9
42,9 35,8
-5,2
21,6
10,8
-11,2
-41,0
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Pyrrhocorax pyrrhocorax
-87,8
-82,3 -99,2 -94,5-89,2-83,3
-87,2
-78,2 -97,2
-100
-80
-60
-40
-20
0
20
40
60
80
100
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
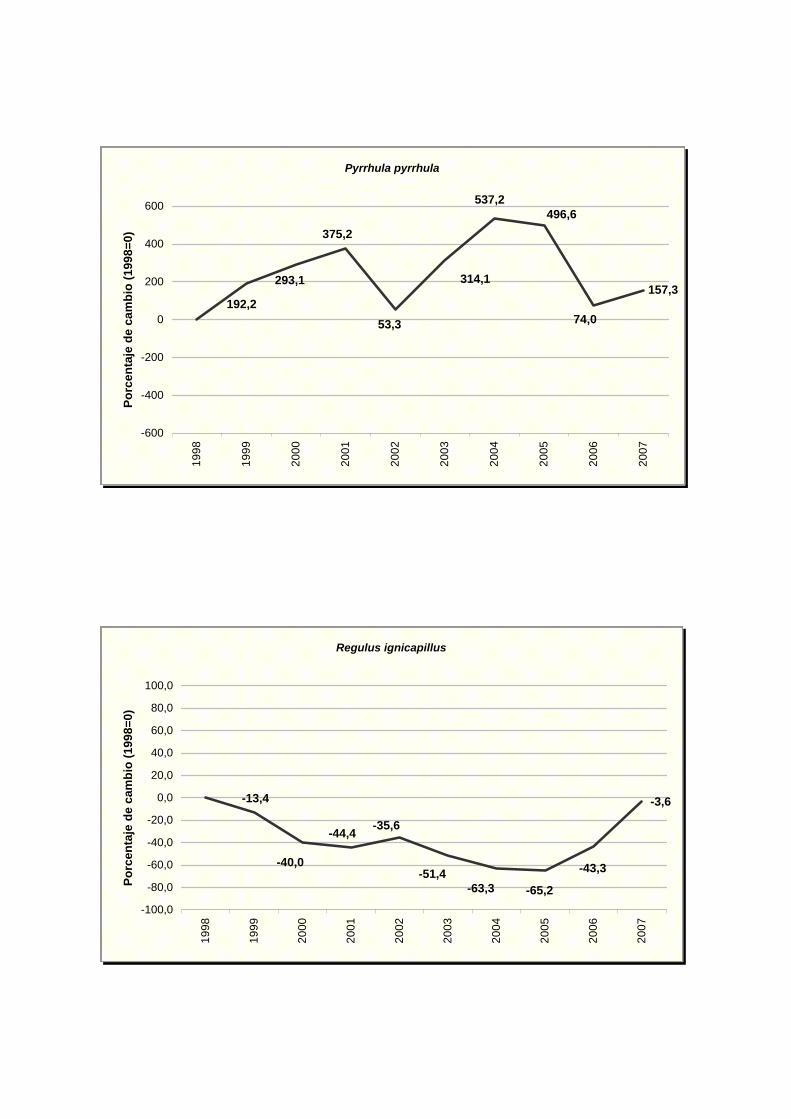
Pyrrhula pyrrhula
157,3
496,6 375,2
192,2
293,1
53,3
537,2
314,1
74,0
-600
-400
-200
0
200
400
600 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Regulus ignicapillus
-3,6
-43,3-51,4 -63,3
-35,6
-40,0
-13,4
-44,4
-65,2 -100,0
-80,0
-60,0
-40,0
-20,0
0,0
20,0
40,0
60,0
80,0
100,0
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
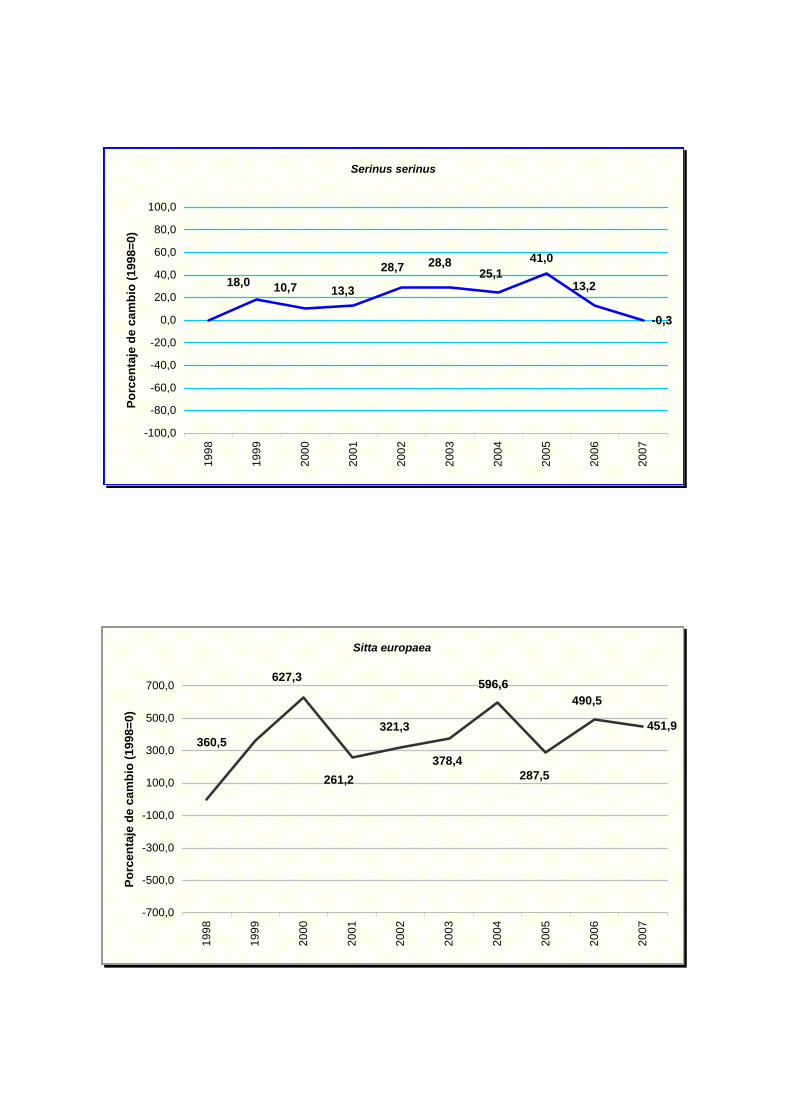
Serinus serinus
-0,3
13,2
28,8 25,128,7
10,718,0 13,3
41,0
-100,0
-80,0
-60,0
-40,0
-20,0
0,0
20,0
40,0
60,0
80,0
100,0 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Sitta europaea
451,9
490,5
378,4
596,6
321,3
627,3
360,5
261,2 287,5
-700,0
-500,0
-300,0
-100,0
100,0
300,0
500,0
700,0
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
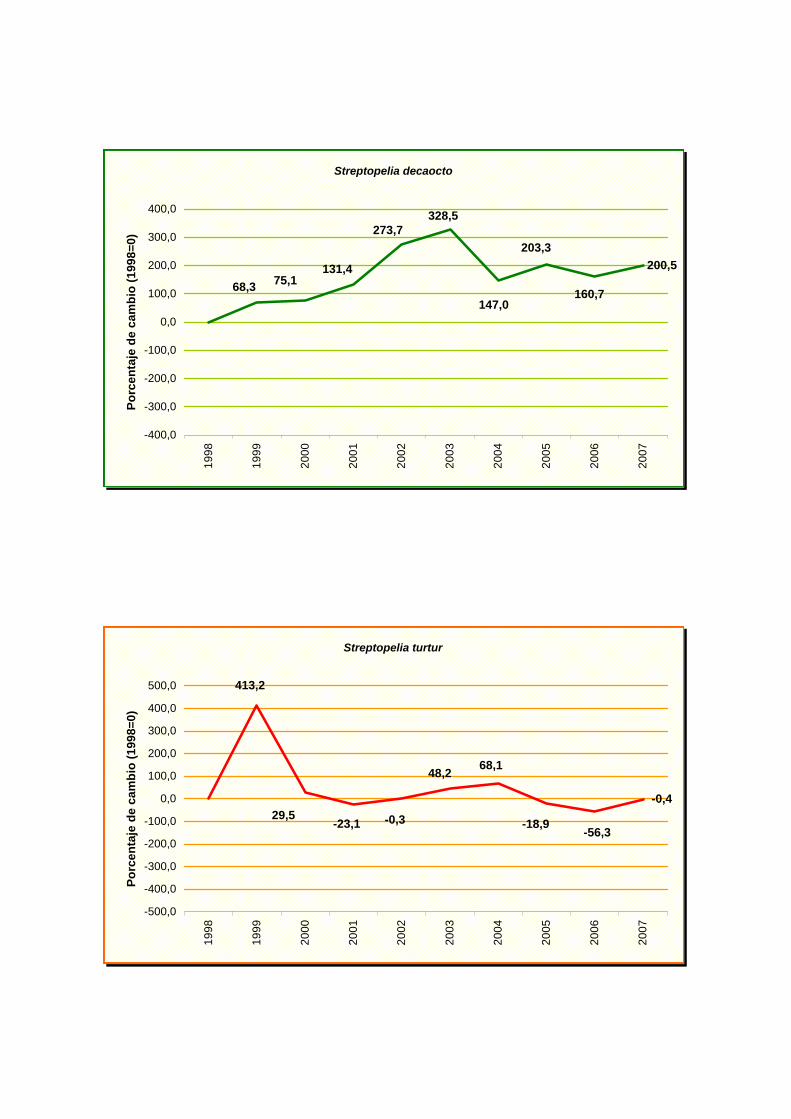
Streptopelia decaocto
200,5
160,7
328,5
147,0
273,7
75,168,3 131,4
203,3
-400,0
-300,0
-200,0
-100,0
0,0
100,0
200,0
300,0
400,0 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Streptopelia turtur
-0,4
-56,3
48,2 68,1
-0,329,5
413,2
-23,1 -18,9
-500,0
-400,0
-300,0
-200,0
-100,0
0,0
100,0
200,0
300,0
400,0
500,0
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
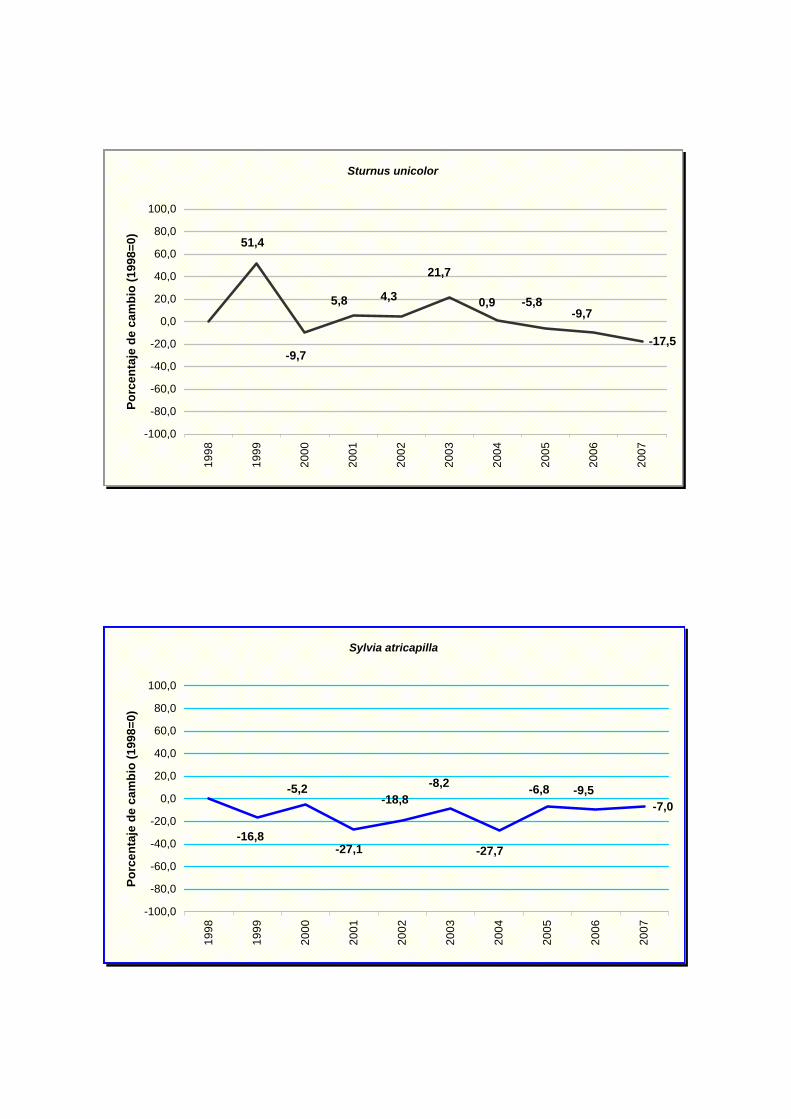
Sturnus unicolor
-17,5
-9,7
21,7
0,94,3
-9,7
51,4
5,8 -5,8
-100,0
-80,0
-60,0
-40,0
-20,0
0,0
20,0
40,0
60,0
80,0
100,0 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Sylvia atricapilla
-7,0 -9,5-8,2
-27,7
-18,8-5,2
-16,8 -27,1
-6,8
-100,0
-80,0
-60,0
-40,0
-20,0
0,0
20,0
40,0
60,0
80,0
100,0
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
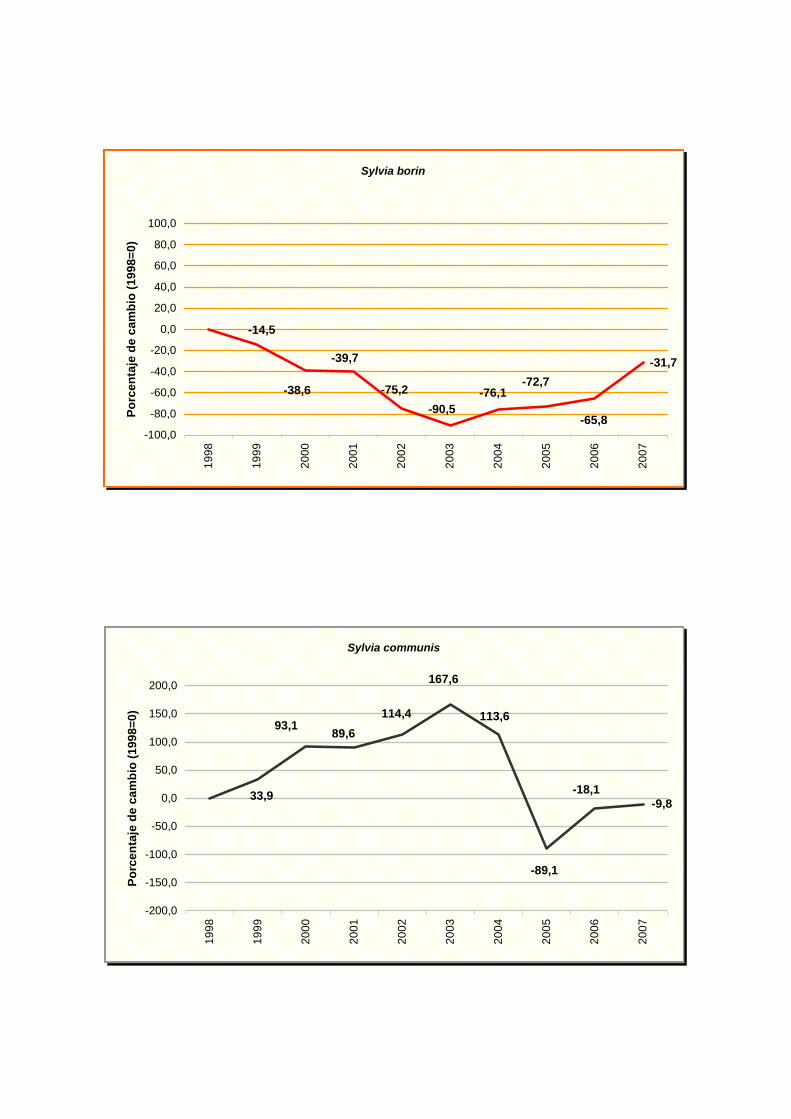
Sylvia borin
-31,7
-65,8-90,5
-76,1-75,2-38,6
-14,5
-39,7
-72,7
-100,0
-80,0
-60,0
-40,0
-20,0
0,0
20,0
40,0
60,0
80,0
100,0 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Sylvia communis
-9,8
-89,1
89,6
33,9
93,1 114,4 113,6
167,6
-18,1
-200,0
-150,0
-100,0
-50,0
0,0
50,0
100,0
150,0
200,0
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
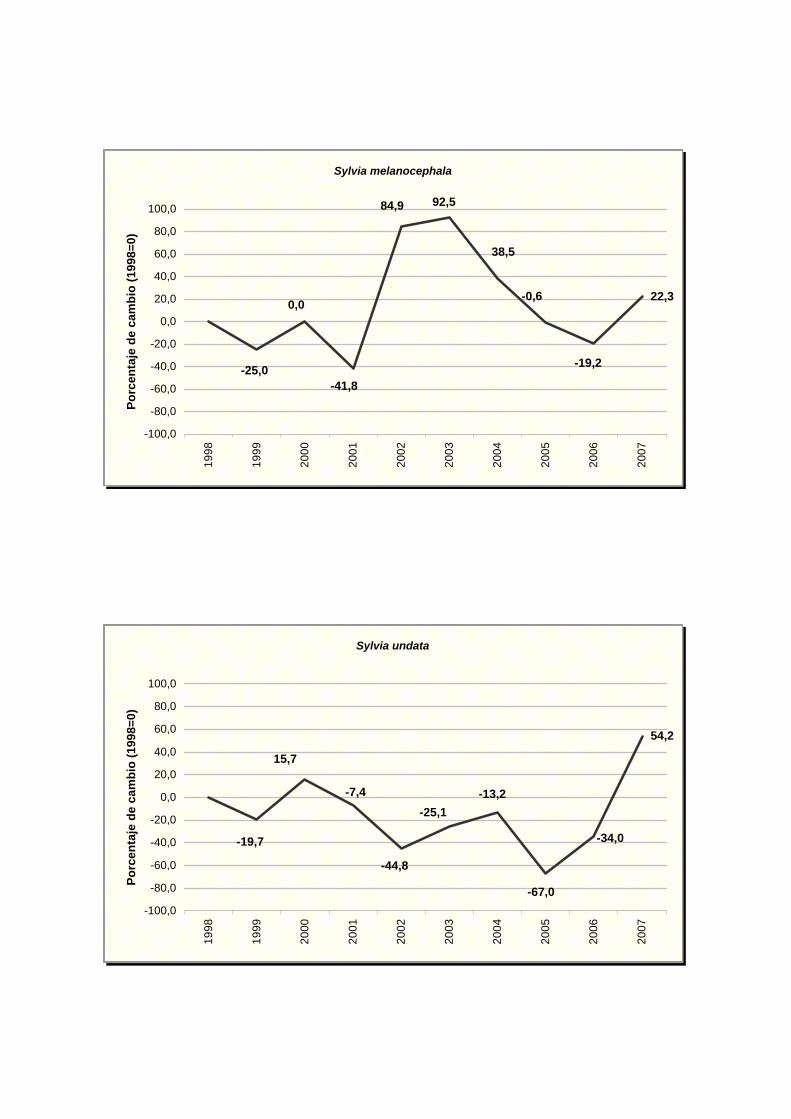
Sylvia melanocephala
22,3
-19,2
92,5
38,5
84,9
0,0
-25,0 -41,8
-0,6
-100,0
-80,0
-60,0
-40,0
-20,0
0,0
20,0
40,0
60,0
80,0
100,0 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Sylvia undata
54,2
-34,0
-25,1 -13,2
-44,8
15,7
-19,7
-7,4
-67,0 -100,0
-80,0
-60,0
-40,0
-20,0
0,0
20,0
40,0
60,0
80,0
100,0
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
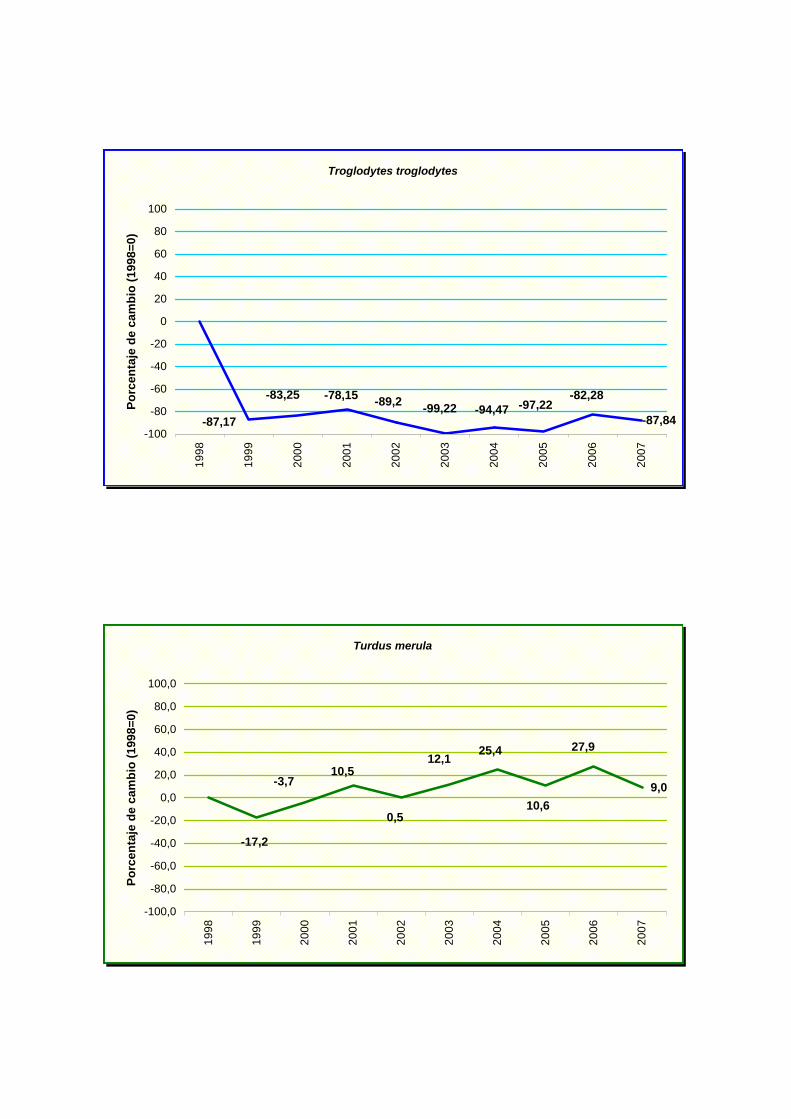
Troglodytes troglodytes
-87,84
-82,28 -99,22 -94,47-89,2-83,25
-87,17
-78,15 -97,22
-100
-80
-60
-40
-20
0
20
40
60
80
100 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
Turdus merula
9,0
27,912,1 25,4
0,5
-3,7
-17,2
10,5
10,6
-100,0
-80,0
-60,0
-40,0
-20,0
0,0
20,0
40,0
60,0
80,0
100,0
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)
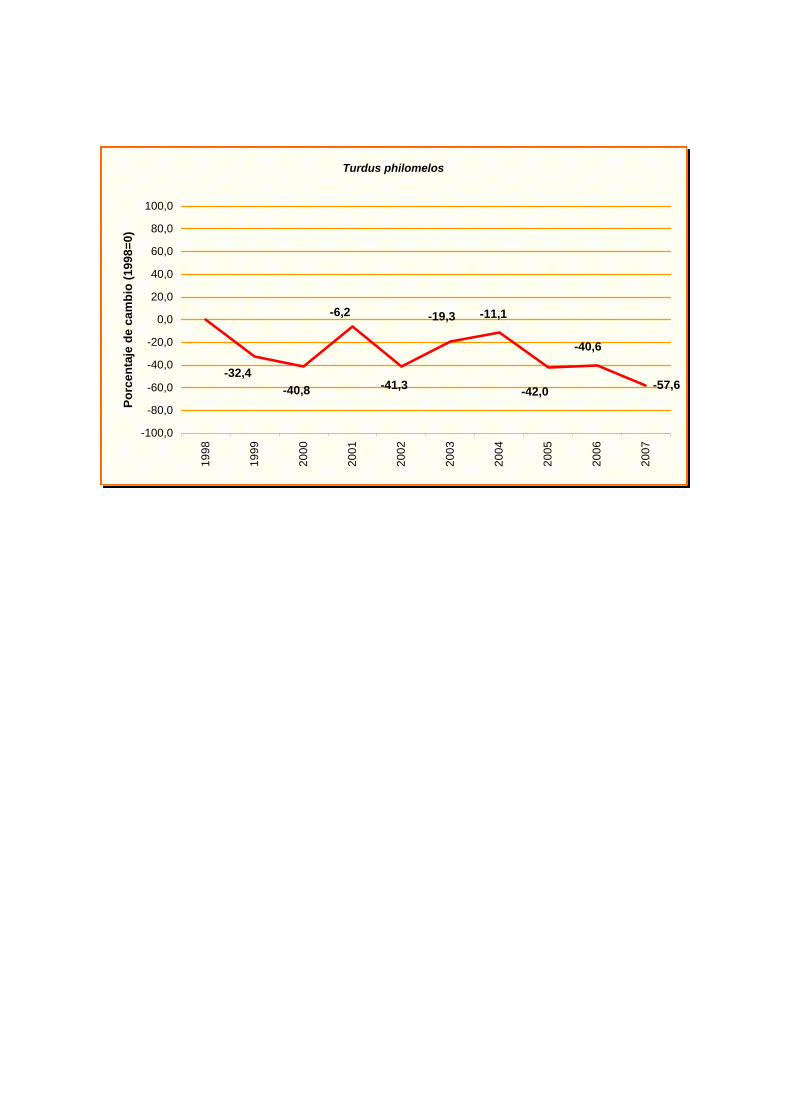
Turdus philomelos
-57,6-42,0
-6,2
-32,4 -40,8 -41,3
-11,1-19,3
-40,6
-100,0
-80,0
-60,0
-40,0
-20,0
0,0
20,0
40,0
60,0
80,0
100,0 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
Porc
enta
je d
e ca
mbi
o (1
998=
0)

Anexo V Índice de cambio anual obtenido para distintos grupos de especies: urbanas, de cultivos
y forestales. Se presentan en verde los grupos con tendencia positiva y en rojo con tendencia negativa.

TENDENCIAS OBTENIDAS POR GRUPOS DE ESPECIES
GRUPO % CULTIVOS EN GENERAL -3,1 FORESTALES 2,4 URBANAS -1,1

Anexo VI Gráficas de evolución de los tres grupos considerados (urbanas, de cultivos y forestales) entre 1998-2007. Se presentan en verde los grupos con tendencia positiva y en rojo con
tendencia negativa.
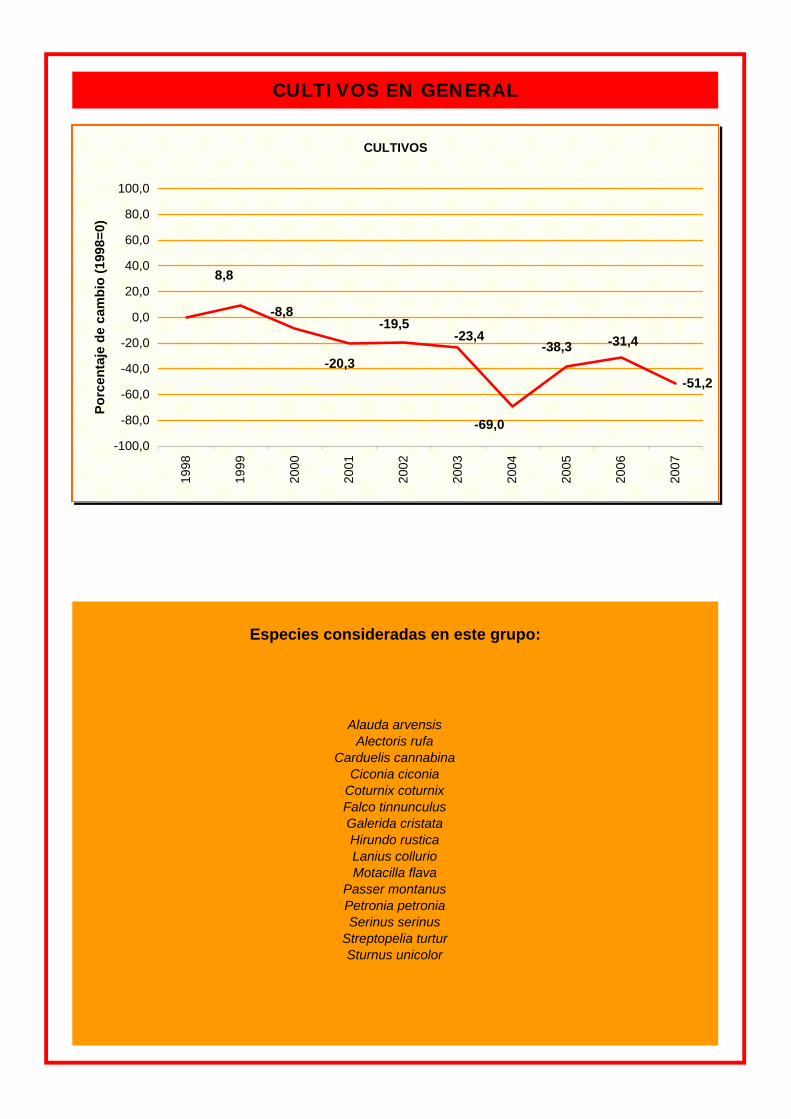
Especies consideradas en este grupo:
Coturnix coturnix
CULTIVOS EN GENERAL
Alauda arvensis Alectoris rufa
Carduelis cannabina Ciconia ciconia
Falco tinnunculus Galerida cristata Hirundo rustica Lanius collurio
Streptopelia turtur Sturnus unicolor
Motacilla flava Passer montanus Petronia petronia Serinus serinus
CULTIVOS
-51,2
-38,3 -20,3
-31,4-23,4
-69,0
-19,5-8,8
8,8
-100,0
-80,0
-60,0
-40,0
-20,0
0,0
20,0
40,0
60,0
80,0
100,0 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
(Po
rcen
taje
de
cam
bio
1998
=0)
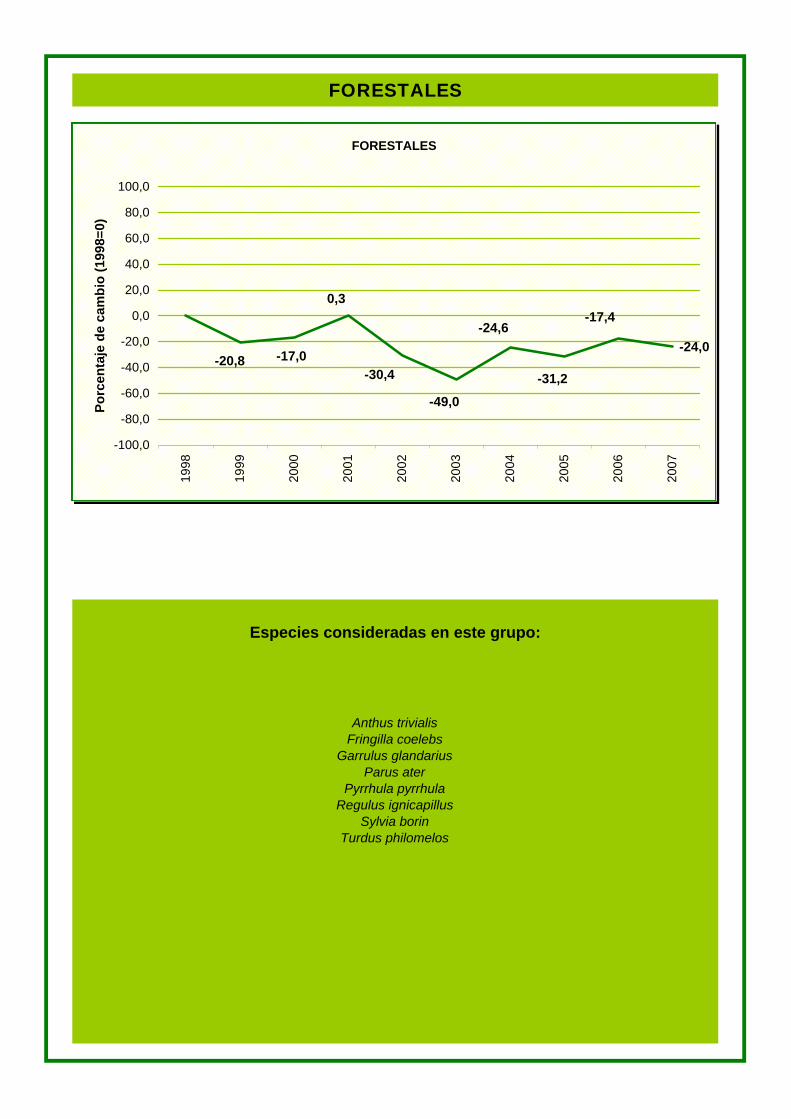
Especies consideradas en este grupo:
FORESTALES
Anthus trivialis Fringilla coelebs
Garrulus glandarius Parus ater
Pyrrhula pyrrhula Regulus ignicapillus
Sylvia borin Turdus philomelos
FORESTALES
-24,0
-31,2
0,3
-20,8 -17,0 -30,4
-24,6
-49,0
-17,4
-100,0
-80,0
-60,0
-40,0
-20,0
0,0
20,0
40,0
60,0
80,0
100,0 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
(Po
rcen
taje
de
cam
bio
1998
=0)
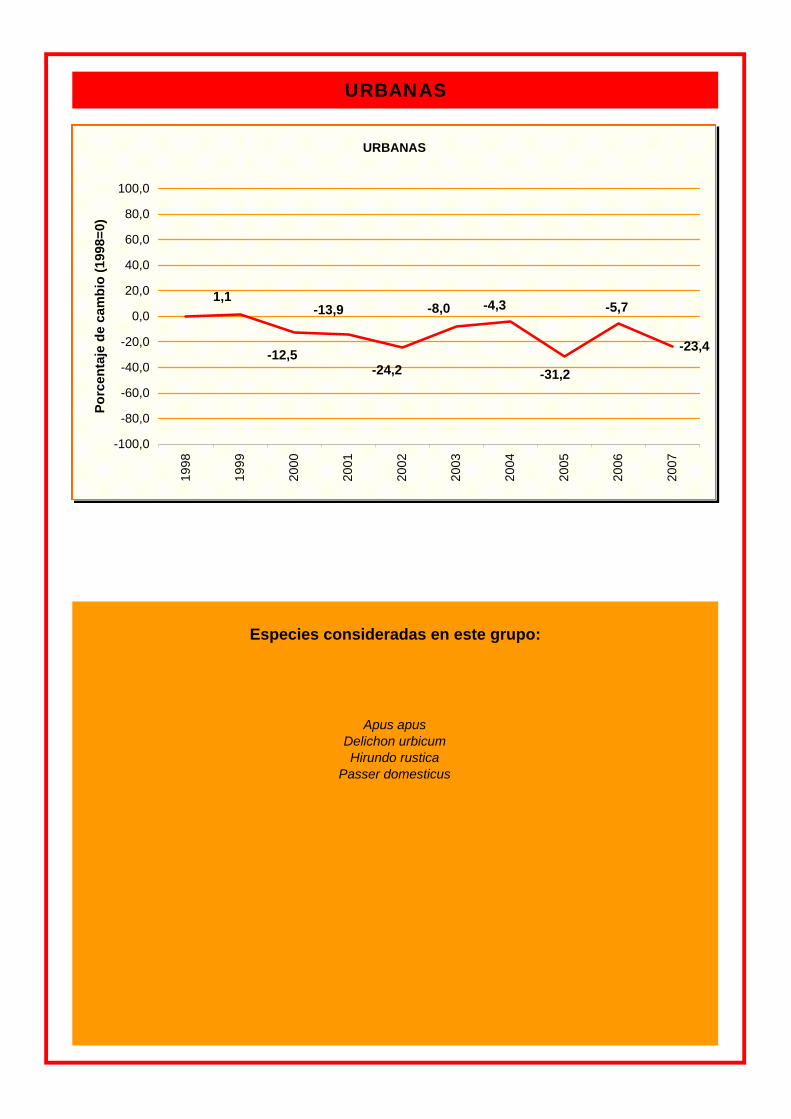
Especies consideradas en este grupo:
URBANAS
Hirundo rustica Passer domesticus
Apus apus Delichon urbicum
URBANAS
-23,4
-31,2
-13,9 -5,7-8,0 -4,3
-24,2 -12,5
1,1
-100,0
-80,0
-60,0
-40,0
-20,0
0,0
20,0
40,0
60,0
80,0
100,0 19
98
1999
2000
2001
2002
2003
2004
2005
2006
2007
(Po
rcen
taje
de
cam
bio
1998
=0)

Anexo VII Análisis estadístico de cada especie generado por el programa TRIM.





























