Embed Size (px)
Citation preview

Kazutoshi Murakami,1,2 Jun Eguchi,1 Kazuyuki Hida,1 Atsuko Nakatsuka,1
Akihiro Katayama,1 Miwa Sakurai,3 Haruki Choshi,1 Masumi Furutani,4
Daisuke Ogawa,3 Kohji Takei,5 Fumio Otsuka,2 and Jun Wada1
Antiobesity Action of ACAM byModulating the Dynamics of CellAdhesion and Actin Polymerizationin AdipocytesDiabetes 2016;65:1255–1267 | DOI: 10.2337/db15-1304
Coxsackie virus and adenovirus receptor-like membraneprotein (CLMP) was identified as the tight junction–associated transmembrane protein of epithelial cellswith homophilic binding activities. CLMP is also rec-ognized as adipocyte adhesion molecule (ACAM), andit is upregulated in mature adipocytes in rodentsand humans with obesity. Here, we present that aP2promoter–driven ACAM transgenic mice are protectedfrom obesity and diabetes with the prominent reductionof adipose tissue mass and smaller size of adipocytes.ACAM is abundantly expressed on plasma membrane ofmature adipocytes and associated with formation ofphalloidin-positive polymerized form of cortical actin(F-actin). By electron microscopy, the structure of zonulaadherens with an intercellular space of ∼10–20 nm wasobserved with strict parallelism of the adjoining cellmembranes over distances of 1–20 mm, where ACAMand g-actin are abundantly expressed. The formationof zonula adherens may increase the mechanicalstrength, inhibit the adipocyte hypertrophy, and improvethe insulin sensitivity.
Quite a few adhesion molecules have been identified bymolecular genetics as well as gene expression profile studiesin adipocytes derived from experimental models and humanstudies. For instance, only cadherins were reported to beexpressed in premature adipocytes. In the cell lines, such as
C3H10T1/2 and 3T3-L1 cells, N-cadherin and cadherin-11are expressed, and they are prominently suppressed bythe induction of adipocyte differentiation and down-regulated to very low levels after the full differentia-tion (1). Transgenic (Tg) expression of dominant-negativeN-cadherin decreased bone formation, delayed acquisitionof peak bone mass, and increased body fat (2). Althoughthe information of adhesion molecules in adipocyte biol-ogy is limited, we identified adipocyte adhesion molecule(ACAM) from the visceral adipose tissues of OLETF (OtsukaLong-Evans Tokushima fatty) rats by PCR-based cDNA sup-pressive subtraction methods (3). Mouse ACAM was inde-pendently identified as adipocyte-specific protein 5 (ASP5)from 3T3-L1 cells by using signal sequence trap by aretrovirus-mediated expression screening method (4). HumanACAM had been identified as coxsackie virus and adenovirusreceptor-like membrane protein (CLMP) by bioinformaticsapproaches, and Raschperger et al. (5) demonstrated thatCLMP is a component of the tight junction of epithelial cellsand colocalized with zonula occludens-1. ACAM/CLMP be-longs to CTX (cortical thymocyte marker in Xenopus), andthey are characterized by extracellular variable (V-type) andconstant (C2-type) immunoglobulin domains, which areinvolved in the homophilic adhesion and aggregation ofthe cells. Although we reported the expression of ACAMincreased in mature adipocytes in genetically obese db/dband diet-induced obesity mice, and also in adipose tissues
1Department of Nephrology, Rheumatology, Endocrinology and Metabolism,Okayama University Graduate School of Medicine, Dentistry and PharmaceuticalSciences, Okayama, Japan2Department of General Medicine, Okayama University Graduate School of Med-icine, Dentistry and Pharmaceutical Sciences, Okayama, Japan3Department of Diabetic Nephropathy, Okayama University Graduate School ofMedicine, Dentistry and Pharmaceutical Sciences, Okayama, Japan4Central Research Laboratory, Okayama University Graduate School of Medicine,Dentistry and Pharmaceutical Sciences, Okayama, Japan5Department of Biochemistry, Okayama University Graduate School of Medicine,Dentistry and Pharmaceutical Sciences, Okayama, Japan
Corresponding author: Jun Wada, [email protected].
Received 16 September 2015 and accepted 22 February 2016.
This article contains Supplementary Data online at http://diabetes.diabetesjournals.org/lookup/suppl/doi:10.2337/db15-1304/-/DC1.
© 2016 by the American Diabetes Association. Readers may use this article aslong as the work is properly cited, the use is educational and not for profit, andthe work is not altered.
Diabetes Volume 65, May 2016 1255
OBESITYSTUDIES

in the subjects with obesity, the functional role of ACAMin mature adipocytes and in obesity remained totally un-known. Here, we generated aP2 promoter–driven ACAMTg mice, and they are protected from obesity and diabeteswith reduced accumulation of white and brown adiposetissues. In ACAM Tg mice fed high-fat, high-sucrose(HFHS), chow, ACAM is abundantly expressed on plasmamembranes of mature adipocytes, where the zonulaadherens–like structure is formed. The adhesion processand formation of zonula adherens are associated with theformation of actin polymerization on the surface of themature adipocytes. The formation of zonula adherensmay increase the mechanical strength, inhibit the adipocytehypertrophy, alter the signaling events, and improve theinsulin sensitivities. Our finding provides the new thera-peutic modalities targeting the processes of cell adhesionand actin polymerization of adipocytes in the treatment ofobesity and diabetes.
RESEARCH DESIGN AND METHODS
Generation of ACAM Tg Miceb-Globin intron and human growth hormone poly-A sig-nal were ligated into BamHI-EcoRI and EcoRI-XhoI sites ofpcDNA3.1Zeo vector (Invitrogen), respectively (Supplemen-tary Fig. 1A). Coding region of mouse ACAM cDNA wasinserted into EcoRI site by blunt end ligation after filling-in reaction. BamHI and XhoI fragment was subjected toblunt end ligation into SmaI site of pBluescript SKII(+)(Stratagene), in which mouse aP2 promotor was insertedinto EcoRV-PstI site by blunt end ligation. The insert wasexcised with HindIII and NotI, and transgene was generated.Microinjected C57BL/6JJcl one-cell stage zygotes were ovi-duct transferred and permitted to develop to term. Three Tgfounders were obtained, and Southern blot analysis wasperformed using EcoRI and SmaI fragment of the transgene.Genotyping of Tg mice was performed by PCR using primers59-GACATTGAATGGCTGCTCACCG-39, 59-GCTCTGCACATACTGTACAGTC-39, 59-GTTGGAACGCTGGGAACTCACACTGAGATC-39, and 59-GGTTCAGAACCTCTCACTTCCGGTCCTATG-39.
AnimalsMale C57BL/6JJcl mice were housed in cages and main-tained on a 12-h light-dark cycle. For the animal experi-ments with mice, standard chow (NMF; Oriental Yeast) andHFHS (D12331; Research Diet) diets were used and the micewere killed at 30 weeks of age. Oxygen consumption wasmeasured using an O2/CO2 metabolism–measuring system(MK-5000; Muromachi). All animal experiments were ap-proved by the Animal Care and Use Committee of the De-partment of Animal Resources, Advanced Science ResearchCenter, Okayama University. Liver triglyceride was measuredby the Folch method (Skylight Biotech, Tokyo, Japan).
Cell Culture and Adipocyte DifferentiationMouse 3T3-L1 fibroblasts (American Type Culture Collection)were cultured in vitro and differentiated into mature adipo-cytes. 3T3-L1 cells were grown in DMEM (Sigma-Aldrich)
supplemented with 10% calf serum (Hyclone), 100 units/mLpenicillin, and 0.1 mg/mL streptomycin (Sigma-Aldrich).Confluent cells were induced to differentiate by the addi-tion of 0.5 mmol/L 1-methyl-3-isobutylxanthine (IBMX),0.25 mmol/L dexamethasone (DEX), and 10 mg/mL insu-lin (Sigma-Aldrich). After 48 h, induction medium wasremoved and cells were cultured with DMEM supple-mented with 10% FBS (Invitrogen) and 10 mg/mL insulinfor 14 days. For determination of the major inducer forACAM expression, the confluent preadipocytes weretreated with DEX/IBMX/insulin, DEX, IBMX, insulin,DEX/IBMX, IBMX/insulin, DEX/insulin, FBS, pioglitazone,DEX/IBMX/insulin/pioglitazone, 8-bromoadenosine–cAMP(Calbiochem), or forskolin (Sigma-Aldrich) at the con-centrations indicated. In separate experiments, confluentpreadipocytes were preincubated with H89 (Sigma-Aldrich),SB203580 (Calbiochem), PD98059 (Calbiochem), andLY294002 (Sigma-Aldrich) for 30 min, respectively, in priorto induction of differentiation.
Poly A+ RNA Analysis of Various TissuesFor quantitative real-time PCR analysis in various tissues inmice, cDNA synthesized from 2 mg total RNA was analyzedin a Sequence Detector (model 7900; Applied Biosystems)with specific primers and SYBR Green PCR Master (AppliedBiosystems). The relative abundance of mRNAs was stan-dardized with 36B4 mRNA as the invariant control. ThemRNA expression was determined by SYBR Green. Gene-specific primers are indicated in Supplementary Table 1.
RNA Isolation and Northern Blot Analysesof 3T3-L1 CellsTotal RNA was isolated from 3T3-L1 cells by using RNeasyLipid Tissue Mini kit (Qiagen). For Northern blot analysis,10 mg total RNA was subjected to 2.2 mol/L formaldehyde1% agarose gel electrophoresis and capillary transferred tothe Hybond XL nylon membranes (GE Healthcare Life Sci-ences). The membranes were hybridized with [a-32P]dCTP-radiolabeled mouse ACAM, mouse C/EBPb, mouse C/EBPd,mouse C/EBPa, mouse peroxisome proliferator–activatedreceptor g, mouse LPL (lipoprotein lipase), and mouse 18Sribosomal RNA (American Type Culture Collection) cDNAsat 68°C in ExpressHyb Hybridization Solution (Clontech)for 1 h. Filters were washed in high-stringency conditions,i.e., four times in 13 sodium chloride–sodium citrate/0.1%SDS at 20°C, followed by two times at 50°C in 0.13 sodiumchloride–sodium citrate/0.1% SDS.
AntibodiesSheep polyclonal anti-ASAM (adipocyte-specific adhesionmolecule) (ACAM) (R&D Systems), rat monoclonal anti-F4/80 (Cl:A3-1) (AbD Serotec), rabbit polyclonal anti–nonmusclemyosin heavy-chain II-A (Covance), and mouse monoclo-nal anti–g-actin (Sigma-Aldrich) antibodies were used forimmunoblotting, immunofluorescence, and immunogoldstudies. For secondary antibodies, rabbit anti-sheep IgG,horseradish peroxidase–conjugated (Millipore), Alexa Fluor594 donkey anti-sheep IgG (H+L), Alexa Fluor 488 goat
1256 ACAM in Obesity Diabetes Volume 65, May 2016

anti-rat IgG (H+L), and Alexa Fluor 488 chicken anti-rabbitIgG (H+L) (Invitrogen) were used.
High-Performance Liquid Chromatography–TandemMass Spectrometry and Matrix-Assisted LaserDesorption/Ionization–Top of Flight MassSpectrometryRecombinant full-length mouse ACAM tagged withcalmodulin-binding peptide (CBP) and streptavidin-binding peptide (SBP) (Adeno-pCTAP-mACAM) (InterPlayMammalian TAP System; Stratagene) were prepared byadenovirus expression kit (Takara). Adeno-pCTAP-mACAMwas introduced to 3T3-L1 cells. Soluble proteins werepurified by CBP and SBP binding resin (InterPlay MammalianTAP System; Stratagene) and subjected to SDS-PAGE andCoomassie blue staining. Visible bands were excised andin-gel digested with trypsin and analyzed with liquidchromatography–tandem mass spectrometry (LC-MS/MS)and matrix-assisted laser desorption/ionization–top offlight mass spectrometry (MALDI-TOF/MS).
Cell Fractionation of Cultured 3T3-L1 Cells3T3-L1 cells at 24 h after the induction were fractioned.Cells were washed in PBS and resuspended in suspensionbuffer (20 mmol/L Tris-HCl, pH 7.4; 1 mmol/L EDTA; and255 mmol/L sucrose) containing protease inhibitor cock-tail (Sigma-Aldrich) and homogenized with a precooledmotor-driven Potter-Elvehjem grinder with 20 strokesat 1,400 rpm. Nuclei were pelleted by centrifugation at1,000g for 10 min. The postnuclear supernatant wascentrifuged at 16,000g for 20 min. The pellet containingmitochondria, peroxisomes, and plasma membrane frac-tions was resuspended in 5 mL resuspension buffer andlayered onto 5 mL sucrose cushion (1.12 mol/L sucrose,1 mmol/L EDTA, and 20 mmol/L Tris-HCl, pH 7.4) andcentrifuged at 101,000g for 25 min. The mitochondriaand peroxisomes were collected as a pellet and resus-pended in suspension buffer. The plasma membrane wascollected at the interface, resuspended in 10 mL suspen-sion buffer, centrifuged at 16,000g for 15 min, and resus-pended in suspension buffer. The postnuclear supernatantwas centrifuged at 48,000g for 20 min, and the pellet con-taining high-density microsomes (HDMs) was obtained. Thesupernatant was centrifuged at 212,000g for 70 min, sepa-rating the pellet containing low-density microsomes (LDMs)and the supernatant containing the cytosol. Pellets fromfractions containing HDMs and LDMs were resuspendedin suspension buffer.
Immunoelectron MicroscopyAdipose tissues and 3T3-L1 cells were fixed with 4% para-formaldehyde and 0.05% glutaraldehyde in 0.01 mol/Lphosphate buffer, pH 7.4, for 30 min and then scrapedfrom the dish, and a pellet was prepared with a microfuge.The tissues and cell pellets were then dehydrated in agraded series of ethanol, and they were embedded andpolymerized in LR-White (Polysciences, Inc.) at 50°C for48 h. Approximately 60-nm-thin sections were picked up
with a 300 mesh nickel grid. The sections were pretreatedwith 20 mmol/L Tris-buffered saline, pH 8.2, containing0.25% BSA. They were incubated with anti-ACAM anti-body and then with anti-sheep IgG conjugated with 10 nmcolloidal gold particles for 4 h at room temperature. Theywere washed with Tris-buffered saline and stained withlead citrate and uranyl acetate and examined by an elec-tron microscope at accelerating voltage of 75 kv. Trans-mission and scanning electron microscopy was performedas previously described (6).
Adenoviral Vectors and InfectionThe adenoviral vector encoding a constitutively activeform of C/EBPb (liver-enriched activator protein [LAP])was generously provided by Professor Hiroshi Sakaue(Tokushima University, Tokushima, Japan). For adenovi-ral infection, 3T3-L1 preadipocytes were cultured to 90%confluency and infected at a multiplicity of infection of 10plaque-forming units/cell 48 h before induction of differ-entiation (7). The cells were collected at indicated timepoints and were subjected to Northern blot analysis. As acontrol, the adenoviral vector encoding LacZ was used.
Plasmid Constructs and Luciferase Reporter AssayThe ACAM 1,869–base pair (21,741 to 128) luciferaseplasmid was generated by ligating into the cloning siteof the promoterless luciferase reporter plasmid pGL3-Basic (Promega). Various 59-deletion constructs 21,341,2584, 2464, 2235, 297, 279, and 257 were made.The mutant constructs of promoter sequence using 279deletion construct—wild type (WT) (272 to 259), 59-AGCTAACCCCAAAC-39; mutant (MT)1, 59-AGCTAACAATGGCC-39; and mutant 2 (MT2), 59-AGCTAAACCCGAAC-39—were generated by PCR. 3T3-L1 preadipocyteswere transfected using a Neon electroporation transfec-tion system (Invitrogen). Transfections were performedusing 10 mg pBIND (Renilla luciferase)-SV40 (internalcontrol) along with 10 mg pGL3-Basic plasmids containingthe ACAM promoter or with pGL3-Basic plasmid. Forty-eight hours after transfection, cells were incubated with0.5 mmol/L IBMX for 6 h, and luciferase reporter assayswere performed using the Dual Luciferase Reporter AssaySystem (Promega). Transfection efficiencies were nor-malized to the Renilla luciferase activity. For the cyto-megalovirus promoter–driven transient overexpressionof Kruppel-like factor (KLF)4, GATA-binding protein 1 andGATA-binding protein 6, EX-Mm19461-M02 (pReceiver-M02), EX-Mm02659-M02 (pEZ-M02), and EX-Mm02664-M02 (pEZ-M02) (OmicsLink Expression Clone, Genecopia)were used, respectively. For negative control, EX-NEG-M02(pReceiver-M02CT) was used.
Statistical AnalysisData are expressed as the mean 6 SE, and the multiplecomparisons were performed by a one-way ANOVA withBonferroni and Tukey corrections. A value of P , 0.05 wasregarded as statistically significant. The data were analyzedusing the IBM SPSS Statistics software program.
diabetes.diabetesjournals.org Murakami and Associates 1257

RESULTS
ACAM Tg Mice Are Protected From ObesityThree independent aP2-driven ACAM Tg lines of C57BL/6JJcl mice were established (Supplementary Fig. 1A). Thecopy numbers of transgenes differed in L11 (high-), L22(intermediate-), and L4 (low-expression) lines, and theprotein expression of ACAM in both epididymal and sub-dermal fat tissues corresponded to the copy numbersof transgene (Supplementary Fig. 1B). On HFHS chow,mRNA expression of ACAM was significantly augmentedin epididymal, subdermal, and brown adipose tissues andnot in liver and skeletal muscle (Supplementary Fig. 1C).The body weight gains on HFHS chow were prominentlyreduced in ACAM Tg mice in parallel with the expressionof ACAM (Supplementary Fig. 1D–F). Throughout the fol-lowing experiments, intermediate-expression (L22) linewas used, since the mice derived from high-expression(L11) line were completely protected from obesity andthe reduction of body weight may dominantly influencethe phenotype such as improvement of glucose metabo-lism. In ACAM Tg mice, the body and fat pad weight wassignificantly reduced on HFHS chow compared with WTmice (Fig. 1A and B), and the adipocyte size was also sig-nificantly reduced (Fig. 1C–E). In ACAM Tg mice, the im-munoreactivity of ACAM was enhanced on the cell surfaceof adipocytes in both immunofluorescence and immuno-peroxidase studies (Fig. 1F). The staining of ACAM mergedwith F4/80, and the adipose tissue macrophages abun-dantly expressed ACAM (Fig. 1G). Both glucose toleranceand insulin sensitivity were significantly improved inACAM Tg mice on HFHS chow (Fig. 2A–C), and serumsmall-sized LDL cholesterol was significantly reduced com-pared with WT mice (Fig. 2D). In contrast, the lipid drop-lets in liver tissue, liver weight, and triglyceride contentwere not altered in ACAM Tg mice on HFHS chow (Fig.2E–G). The data suggested that ACAM Tg mice are pro-tected from obesity and insulin resistance.
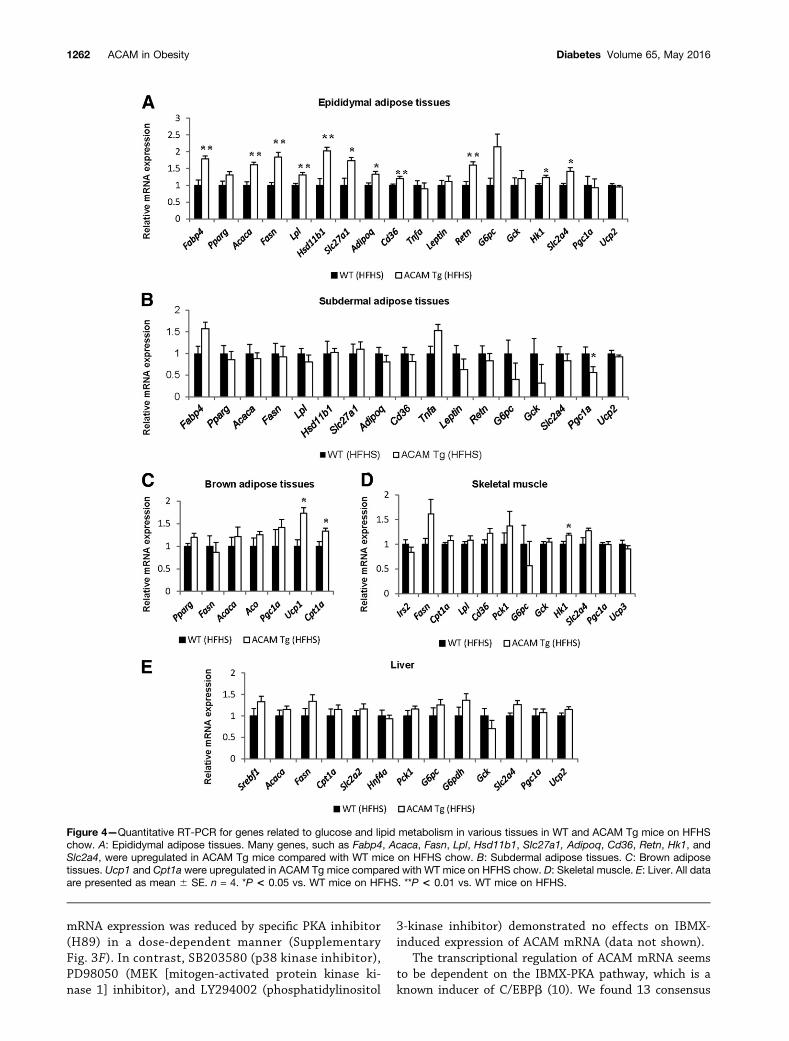
Although the locomotor activity, food intake, andrespiratory quotient were not altered in ACAM Tg miceon HFHS chow, the weight of brown adipose tissues wassignificantly reduced in ACAM Tg mice on HFHS chowcompared with WT mice (Fig. 3A–D). The light and elec-tron microscopy demonstrated that lipid droplets wereprominently reduced in ACAM Tg mice on HFHS chowin brown adipose tissues (Fig. 3D). The oxygen consump-tion rate was significantly elevated in both dark and lightperiods in ACAM Tg mice on HFHS chow (Fig. 3E–G).Quantitative RT-PCR demonstrated that lipid metabo-lism–related genes (Fabp4, Acaca, Fasn, Lpl, Hsd11b1, andSlc27a1), inflammation-related genes (Adipoq, Cd36,and Retn), and glucose metabolism–related genes (Hk1and Slc2a4) significantly increased in ACAM Tg mice com-pared with WT mice (Fig. 4A). In brown adipose tissues,the gene expression of Ucp1 and Cpt1a significantly in-creased in ACAM Tg mice compared with WT (Fig. 4C). Incontrast, most of the genes expressed in subdermal adipose
tissues, skeletal muscle, and liver were not altered in ACAMTg mice on HFHS chow (Fig. 4B, D, and E). Taken together,the overexpression of ACAM in adipocytes directly alteredthe biological functions of adipose tissues in the status ofobesity and it increased brown fat activity. We also checkedconversion of white to bright/beige adipocyte in subdermalWATs; however, we did not observe such browning in thetissues (Supplementary Fig. 1G).
ACAM Is Differentially Expressed in 3T3-L1 AdipocytesTo explore the function of ACAM in adipocytes, we nextinvestigated the mRNA expression during the 3T3-L1adipocyte differentiation. ACAM mRNA revealed thefirst peak at 6 h after induction, i.e., the early stage ofdifferentiation, and was downregulated within 48 h, whichcoincided with the induction of Cebpb and Cebpd (Fig. 5Aand Supplementary Fig. 2A). At 14 days, ACAM mRNAdemonstrated the second peak after the appearance ofadipocyte maturation markers, such as Cebpa, Pparg, andLpl (Fig. 5B and Supplementary Fig. 2B). By the fraction-ation of the 3T3-L1 cells, ACAM localized mainly in theplasma membrane fraction, to a lesser extent in LDM andHDM fractions (Fig. 5C and Supplementary Fig. 2C). West-ern blot analyses demonstrated the upregulation of ACAMafter the induction of differentiation (Supplementary Fig.2D). Adeno-pCTAP-mACAM was introduced to 3T3-L1cells, and soluble proteins were purified by CBP and SBPbinding resin and finally subjected to SDS-PAGE and Coo-massie blue staining (Fig. 5D). Visible bands were excisedand in-gel digested with trypsin and analyzed with LC-MS/MS and MALDI-TOF/MS, by which myosin II-A and g-actinwere identified, respectively. We next confirmed the com-plex formation of ACAM and g-actin by the immunopre-cipitation using 3T3-L1 cells (Fig. 5E). Before the inductionand at the early stage of differentiation, ACAM expressedmainly on the cell processes of 3T3-L1 cells revealed byimmunofluorescence and immunogold studies (Fig. 5F–H).Before the induction, ACAM did not colocalized with myo-sin II-A; however, it merged with myosin II-A concentratedon the cell surface as well as perinuclear cytosol when3T3-L1 cells fully differentiated to the mature adipocytes(Fig. 5G).
We then searched the critical inducers for the upregu-lation of ACAM mRNA expression. The induction withIBMX or its combination with other inducers upregulatedACAM mRNA levels; the effects were almost comparablewith those from DEX/IBMX/insulin stimulation (Supple-mentary Fig. 3A). IBMX induced ACAM mRNA expressionin a dose- and time-dependent manner, and it peaked at6 h in the presence of 500 nmol/L IBMX (SupplementaryFig. 3B and C). IBMX is known to inhibit phosphodiesterases,stimulate adenyl cyclase activity, and increase the intra-cellular cAMP accumulation (8). Both 8-bromoadenosine–cAMP and forskolin, a cAMP-elevating agent, upregulatedACAM mRNA levels in a dose-dependent manner (Supple-mentary Fig. 3D and E). It is also known that cAMPstimulates protein kinase A (PKA) activity (9) and ACAM
1258 ACAM in Obesity Diabetes Volume 65, May 2016
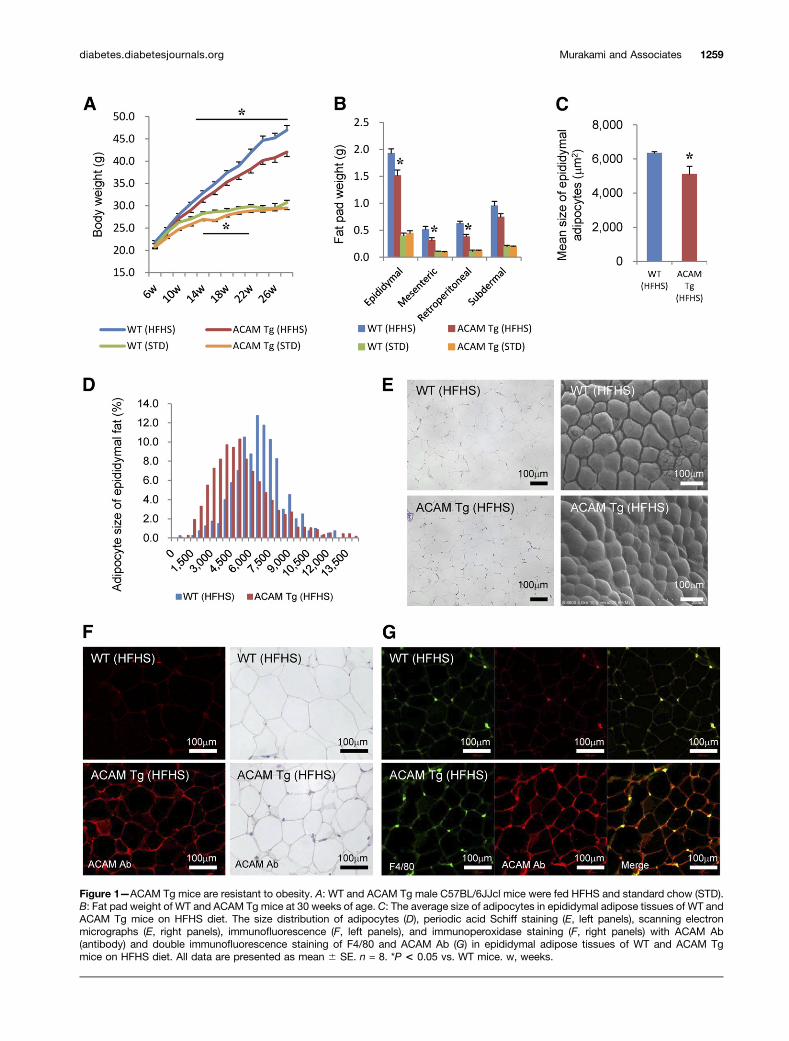
Figure 1—ACAM Tg mice are resistant to obesity. A: WT and ACAM Tg male C57BL/6JJcl mice were fed HFHS and standard chow (STD).B: Fat pad weight of WT and ACAM Tg mice at 30 weeks of age. C: The average size of adipocytes in epididymal adipose tissues of WT andACAM Tg mice on HFHS diet. The size distribution of adipocytes (D), periodic acid Schiff staining (E, left panels), scanning electronmicrographs (E, right panels), immunofluorescence (F, left panels), and immunoperoxidase staining (F, right panels) with ACAM Ab(antibody) and double immunofluorescence staining of F4/80 and ACAM Ab (G) in epididymal adipose tissues of WT and ACAM Tgmice on HFHS diet. All data are presented as mean 6 SE. n = 8. *P < 0.05 vs. WT mice. w, weeks.
diabetes.diabetesjournals.org Murakami and Associates 1259
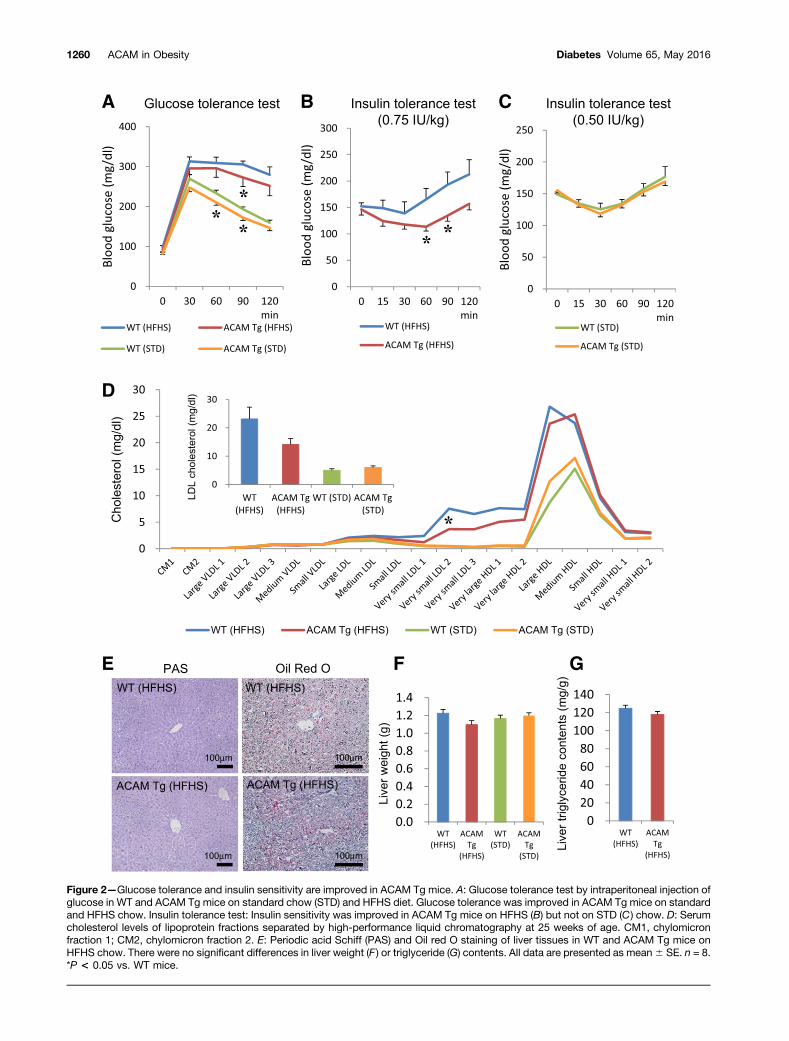
Figure 2—Glucose tolerance and insulin sensitivity are improved in ACAM Tg mice. A: Glucose tolerance test by intraperitoneal injection ofglucose in WT and ACAM Tg mice on standard chow (STD) and HFHS diet. Glucose tolerance was improved in ACAM Tg mice on standardand HFHS chow. Insulin tolerance test: Insulin sensitivity was improved in ACAM Tg mice on HFHS (B) but not on STD (C ) chow. D: Serumcholesterol levels of lipoprotein fractions separated by high-performance liquid chromatography at 25 weeks of age. CM1, chylomicronfraction 1; CM2, chylomicron fraction 2. E: Periodic acid Schiff (PAS) and Oil red O staining of liver tissues in WT and ACAM Tg mice onHFHS chow. There were no significant differences in liver weight (F ) or triglyceride (G) contents. All data are presented as mean6 SE. n = 8.*P < 0.05 vs. WT mice.
1260 ACAM in Obesity Diabetes Volume 65, May 2016
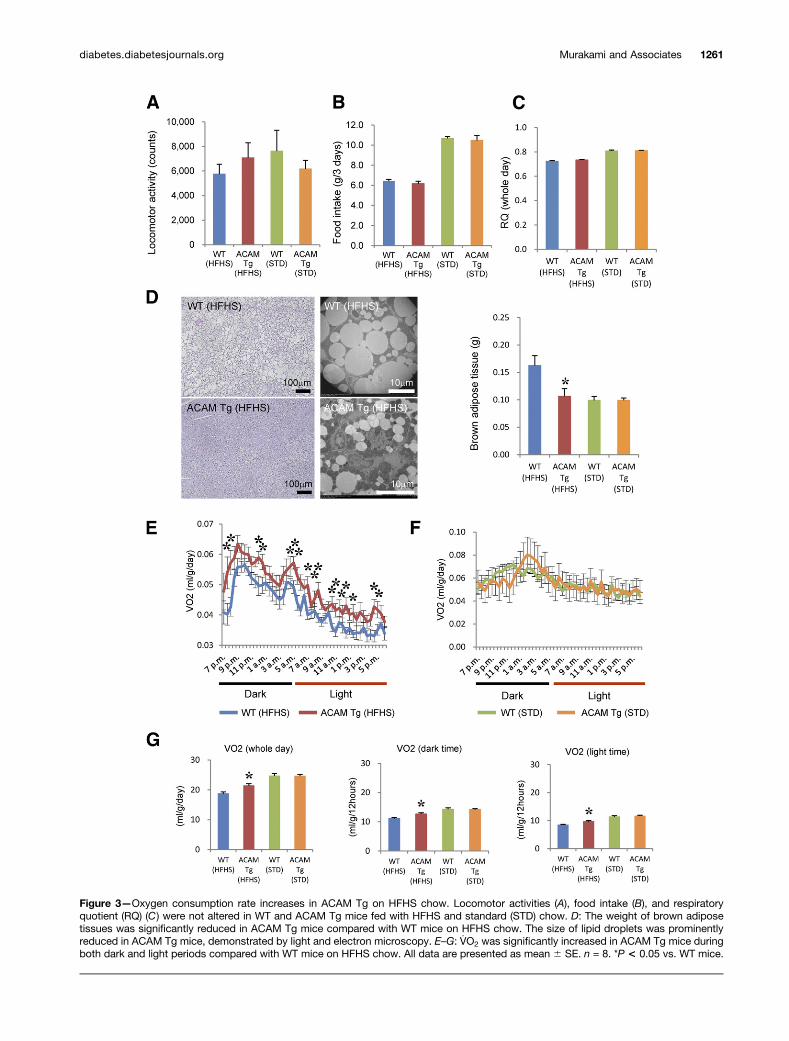
Figure 3—Oxygen consumption rate increases in ACAM Tg on HFHS chow. Locomotor activities (A), food intake (B), and respiratoryquotient (RQ) (C) were not altered in WT and ACAM Tg mice fed with HFHS and standard (STD) chow. D: The weight of brown adiposetissues was significantly reduced in ACAM Tg mice compared with WT mice on HFHS chow. The size of lipid droplets was prominentlyreduced in ACAM Tg mice, demonstrated by light and electron microscopy. E–G: V̇O2 was significantly increased in ACAM Tg mice duringboth dark and light periods compared with WT mice on HFHS chow. All data are presented as mean 6 SE. n = 8. *P < 0.05 vs. WT mice.
diabetes.diabetesjournals.org Murakami and Associates 1261
mRNA expression was reduced by specific PKA inhibitor(H89) in a dose-dependent manner (SupplementaryFig. 3F). In contrast, SB203580 (p38 kinase inhibitor),PD98050 (MEK [mitogen-activated protein kinase ki-nase 1] inhibitor), and LY294002 (phosphatidylinositol
3-kinase inhibitor) demonstrated no effects on IBMX-induced expression of ACAM mRNA (data not shown).
The transcriptional regulation of ACAM mRNA seemsto be dependent on the IBMX-PKA pathway, which is aknown inducer of C/EBPb (10). We found 13 consensus
Figure 4—Quantitative RT-PCR for genes related to glucose and lipid metabolism in various tissues in WT and ACAM Tg mice on HFHSchow. A: Epididymal adipose tissues. Many genes, such as Fabp4, Acaca, Fasn, Lpl, Hsd11b1, Slc27a1, Adipoq, Cd36, Retn, Hk1, andSlc2a4, were upregulated in ACAM Tg mice compared with WT mice on HFHS chow. B: Subdermal adipose tissues. C: Brown adiposetissues. Ucp1 and Cpt1a were upregulated in ACAM Tg mice compared with WT mice on HFHS chow. D: Skeletal muscle. E: Liver. All dataare presented as mean 6 SE. n = 4. *P < 0.05 vs. WT mice on HFHS. **P < 0.01 vs. WT mice on HFHS.
1262 ACAM in Obesity Diabetes Volume 65, May 2016
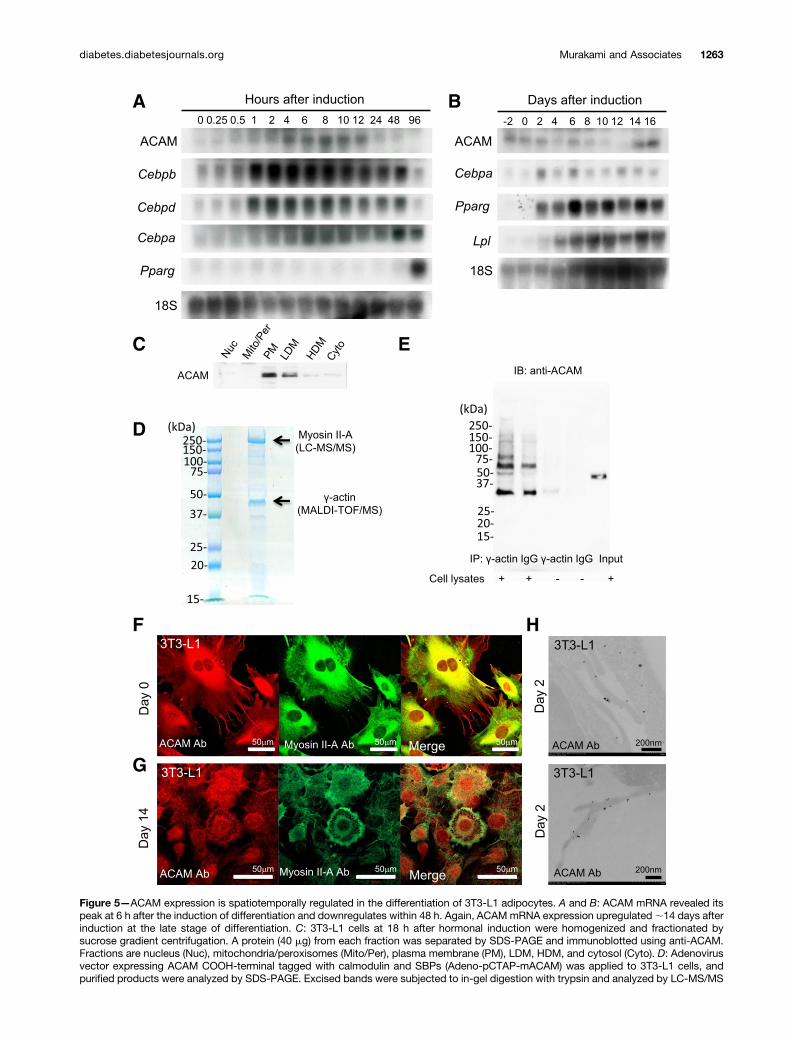
Figure 5—ACAM expression is spatiotemporally regulated in the differentiation of 3T3-L1 adipocytes. A and B: ACAM mRNA revealed itspeak at 6 h after the induction of differentiation and downregulates within 48 h. Again, ACAMmRNA expression upregulated;14 days afterinduction at the late stage of differentiation. C: 3T3-L1 cells at 18 h after hormonal induction were homogenized and fractionated bysucrose gradient centrifugation. A protein (40 mg) from each fraction was separated by SDS-PAGE and immunoblotted using anti-ACAM.Fractions are nucleus (Nuc), mitochondria/peroxisomes (Mito/Per), plasma membrane (PM), LDM, HDM, and cytosol (Cyto). D: Adenovirusvector expressing ACAM COOH-terminal tagged with calmodulin and SBPs (Adeno-pCTAP-mACAM) was applied to 3T3-L1 cells, andpurified products were analyzed by SDS-PAGE. Excised bands were subjected to in-gel digestion with trypsin and analyzed by LC-MS/MS
diabetes.diabetesjournals.org Murakami and Associates 1263

C/EBPb binding sites by analyzing the21,741–base pairACAM promoter region (Supplementary Fig. 4A) andobserved approximately eightfold induction of luciferaseactivities in 2584, 2464, 2235, 297, and 279 ACAMpromoter constructs by the treatments of IBMX or ad-enoviral vector encoding active-form C/EBPb (LAP)(Supplementary Fig. 4B and C). A possible C/EBPb bindingconsensus site between272 and259, AGCTAACCCCAAAC,was mutated by using 279 ACAM promoter construct toproduce 279 (MT1) and 279 (MT2) ACAM promoterconstructs (Supplementary Fig. 4A), which resulted in aloss of transactivation induced by IBMX and LAP. KLF4has been reported to induce adipocyte differentiation byactivating C/EBPb. KLF4-expressing plasmids in the com-bination with GATA1- and/or GATA6-expressing plasmidssynergistically enhanced luciferase activity using279 ACAMpromoter construct (Supplementary Fig. 4D).
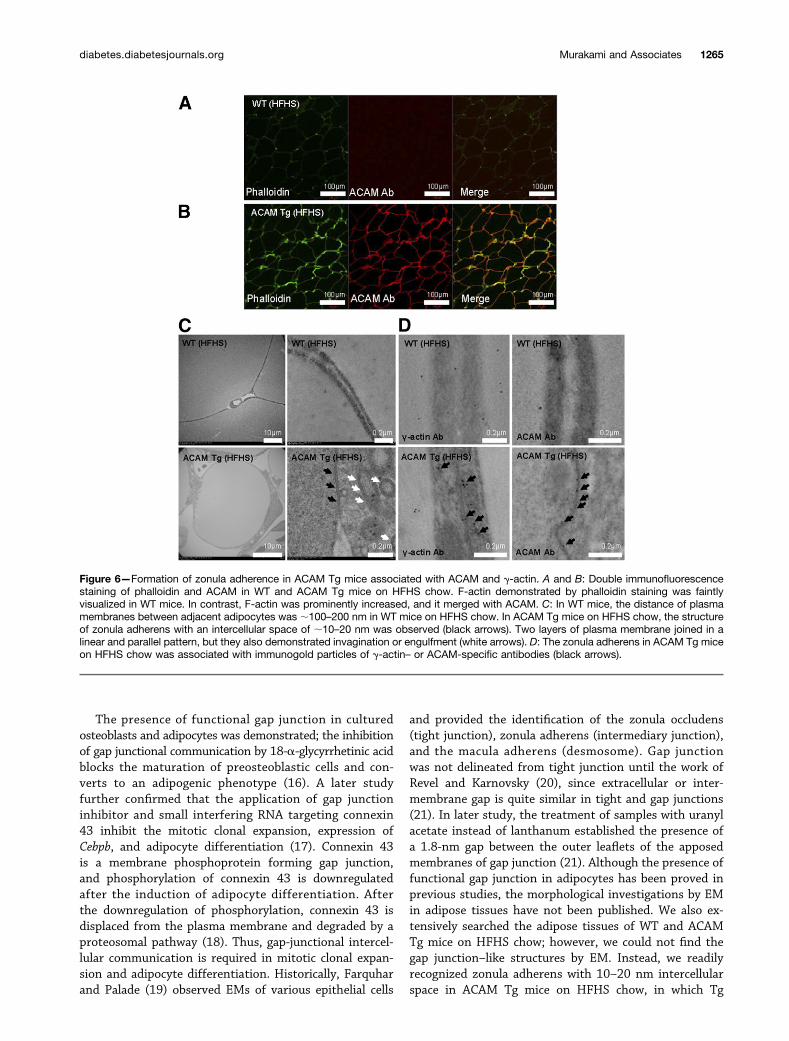
ACAM Promotes the Polymerization of Actin andForms Zonula Adherens in AdipocytesWe investigated the expression of ACAM and polymerizedform of actin (F-actin) by phalloidin staining. In WT miceon HFHS chow, both F-actin and ACAM were faintlystained surrounding the adipocytes in epididymal adiposetissues, while they colocalized and accentuated in a patchyfashion on the adipocytes (Fig. 6A and B). In WT mice, thedistance of plasma membranes between adjacent adipo-cytes was ;100–200 nm in WT mice on HFHS chow (Fig.6C). In ACAM Tg mice on HFHS chow, we observed thezone with an intercellular space of ;10–20 nm, the struc-ture of zonula adherens (Fig. 6C and SupplementaryFigs. 5A and 7, black arrows). The zonula adherens wascontinuous and characterized by strict parallelism of theadjoining cell membranes over distances of 1–20 mm(Supplementary Figs. 6 and 7). The intercellular spacewas occupied by homogeneous and amorphous materialof low density and conspicuous bands of dense materiallocated in the subjacent cytoplasmic matrix. In the zonulaadherens, two layers of plasma membrane joined in alinear and parallel pattern, but they also demonstratedinvagination or engulfment (Fig. 6C and SupplementaryFigs. 5A and 6 , white arrows). The zonula adherens wasassociated with immunogold particles when the sectionswere stained with g-actin– or ACAM-specific antibodies(Fig. 6D and Supplementary Fig. 5B, arrows). In brownadipose tissues in ACAM Tg mice on HFHS chow, similarstructures of zonula adherens were observed and immuno-gold particles associated with ACAM were demonstratedalong the zonula adherens (Supplementary Fig. 8). The
ACAM-mediated homophilic adhesion of adipocytes andformation of zonula adherens promoted the actin polymer-ization. The formation of zonula adherens inhibited the ad-ipocyte hypertrophy by actin polymerization, which resultedin the improvement of obesity and insulin resistance.
DISCUSSION
In 3T3-L1 adipocytes, IBMX is a major inducer of ACAMmRNA expression, and it is a known inducer of C/EBPb.We identified C/EBPb binding sites in the promoter re-gions of the ACAM gene, and luciferase-reporter geneassay demonstrated that the promoter activities were en-hanced by C/EBPb expressed by LAP. KLF4 functions asan immediate early regulator of adipogenesis to induceC/EBPb (11), and KLF4, GATA1, and GATA6 have beenreported to enhance the transcriptional activity of ACAMgene in Sertoli cells (12). In the current study, we dem-onstrated that the promoter activities in luciferase assaywere further enhanced by the expression of KLF4, GATA1,and GATA6 and they are the critical coactivator of thetranscription of the ACAM gene. Immunofluorescenceand immunogold studies demonstrated the presence ofACAM at the cell processes of the 3T3-L1 cells, and wespeculate that ACAM may serve the heterophilic bindingby interacting with certain extracellular matrix (ECM)glycoproteins and also with cytoskeletal componentssuch as g-actin and myosin II-A complex.
The mature adipocytes are surrounded by ECM glycopro-teins such as collagens type I, IV, V, and VI; fibronectin; andthrombospondin, and the ECM environment regulates theadipogenesis and adipocyte function (13). The remodeling ofECM surrounding the adipocytes takes place and the basallamina are thickened in omental adipose tissues of the ratsfed with a high-fat diet (14). The administration of matrixmetalloproteinase inhibitor tolylsam into the mice fed with ahigh-fat diet resulted in lower body weight and lower sub-cutaneous and gonadal adipose tissue mass (15). Thus, theperiadipocyte ECM remodeling is tightly related to the adi-pogenesis, tissue inflammation, fibrosis, insulin sensitivity,and cardiovascular diseases (13). Since periadipocyte ECMplays an important role in adipocyte biology and the closecell-cell contact of the adipocytes has not been visualized inEM (electron microscopy) observation, the role of ACAMin mature adipocytes remains elusive. ACAM/CLMP isexpressed in tight junction of epithelial cells and involvedin the development and maintenance of epithelial cells.Recently, we have developed ACAM knockout mice, andACAM2/2 mice demonstrate the elongation of small intes-tine, dilatation of bronchi, and formation of huge renal cysts.
and MALDI-TOF/MS, and myosin II-A and g-actin were identified. E: Immunoprecipitation (IP) study. Cell lysates of 3T3-L1 cells wereimmunoprecipitated with anti–g-actin and blotted with anti-ACAM antibodies. IB, immunoblot. F: Before the induction of adipocytedifferentiation (day 0), ACAM localized on the cell processes and did not merge with myosin II-A. G: In mature 3T3-L1 adipocytes at14 days after the induction (day 14), ACAM merged with myosin II-A, and they colocalized on the cell surface. H: The immunogold studiesusing ACAM antibody demonstrated that they were preferentially expressed on the cell process of 3T3-L1 cells at 2 days after the induction(day 2).
1264 ACAM in Obesity Diabetes Volume 65, May 2016
The presence of functional gap junction in culturedosteoblasts and adipocytes was demonstrated; the inhibitionof gap junctional communication by 18-a-glycyrrhetinic acidblocks the maturation of preosteoblastic cells and con-verts to an adipogenic phenotype (16). A later studyfurther confirmed that the application of gap junctioninhibitor and small interfering RNA targeting connexin43 inhibit the mitotic clonal expansion, expression ofCebpb, and adipocyte differentiation (17). Connexin 43is a membrane phosphoprotein forming gap junction,and phosphorylation of connexin 43 is downregulatedafter the induction of adipocyte differentiation. Afterthe downregulation of phosphorylation, connexin 43 isdisplaced from the plasma membrane and degraded by aproteosomal pathway (18). Thus, gap-junctional intercel-lular communication is required in mitotic clonal expan-sion and adipocyte differentiation. Historically, Farquharand Palade (19) observed EMs of various epithelial cells
and provided the identification of the zonula occludens(tight junction), zonula adherens (intermediary junction),and the macula adherens (desmosome). Gap junctionwas not delineated from tight junction until the work ofRevel and Karnovsky (20), since extracellular or inter-membrane gap is quite similar in tight and gap junctions(21). In later study, the treatment of samples with uranylacetate instead of lanthanum established the presence ofa 1.8-nm gap between the outer leaflets of the apposedmembranes of gap junction (21). Although the presence offunctional gap junction in adipocytes has been proved inprevious studies, the morphological investigations by EMin adipose tissues have not been published. We also ex-tensively searched the adipose tissues of WT and ACAMTg mice on HFHS chow; however, we could not find thegap junction–like structures by EM. Instead, we readilyrecognized zonula adherens with 10–20 nm intercellularspace in ACAM Tg mice on HFHS chow, in which Tg
Figure 6—Formation of zonula adherence in ACAM Tg mice associated with ACAM and g-actin. A and B: Double immunofluorescencestaining of phalloidin and ACAM in WT and ACAM Tg mice on HFHS chow. F-actin demonstrated by phalloidin staining was faintlyvisualized in WT mice. In contrast, F-actin was prominently increased, and it merged with ACAM. C: In WT mice, the distance of plasmamembranes between adjacent adipocytes was ;100–200 nm in WT mice on HFHS chow. In ACAM Tg mice on HFHS chow, the structureof zonula adherens with an intercellular space of ;10–20 nm was observed (black arrows). Two layers of plasma membrane joined in alinear and parallel pattern, but they also demonstrated invagination or engulfment (white arrows). D: The zonula adherens in ACAM Tg miceon HFHS chow was associated with immunogold particles of g-actin– or ACAM-specific antibodies (black arrows).
diabetes.diabetesjournals.org Murakami and Associates 1265

overexpression of ACAM facilitated the formation ofzonula adherens. Originally, Farquhar and Palade (19) de-fined zonula adherens by the presence of an intercellularspace (;20 nm) occupied by homogeneous, apparentlyamorphous material of low density; by strict parallelism ofthe adjoining cell membranes over distances of 0.2–0.5 mmand by conspicuous bands of dense material located in thesubjacent cytoplasmic matrix (19), the observed structuresof ACAM Tg mice on HFHS chow exactly corresponded withthe definition of zonula adherens.
To further characterize the molecular organization ofzonula adherens in adipocytes, we performed TAP purifi-cation and immunoprecipitation studies and demonstratedthat ACAM interacts with myosin II-A and g-actin in 3T3-L1 cells. We further confirmed that ACAM and g-actincolocalized with zonula adherens observed in adipose tis-sues of ACAM Tg mice by immunoelectron microscopy.Coxsackie and adenovirus receptor (CAR) is a homolog ofACAM with 35% identity, and it also interacts with andbinds to actin (22). CAR is strongly expressed in the de-veloping nervous system, it uniformly expresses on all neu-ral cells at an initial stage, and it downregulates and thenrestricts to axonal and dendritic surface at more advancedstages (23). The membrane proximal Ig domain of CARbinds to a fibronectin fragment and is involved in theheterophilic interactions, while two extracellular Ig do-mains are involved in the homophilic interactions of CAR(23). Similarly, ACAM is localized on the cell processes ofpreadipocytes, downregulates after differentiation, and re-distributes on the plasma membrane of 3T3-L1 cells. Inmature adipocytes in ACAM Tg mice, ACAM colocalizedwith polymerized actin concentrated on the area of zonulaadherens. Recently, the actin cytoskeleton dynamics of po-lymerization and depolymerization cycles have been re-ported to drive the adipocyte differentiation. Adipogenicstimuli downregulate RhoA–Rho-associated protein kinasesignaling, induce the disruption of actin stress fibers, andresult in the conversion to the monomeric globular-actin.The binding globular-actin to MKL1 (megakaryoblastic leu-kemia 1) inhibits the nuclear translocation of MKL1, thereduction of MKL1 in nuclei activates the transcriptionalactivity of Pparg gene, and it results in adipocyte differ-entiation (24). In fully differentiated adipocytes, corticalF-actin is again formed and it regulates the insulin-simulatedtranslocation of GLUT4 from intracellular pool to theplasma membrane. The overexpression of ACAM in Tgmice prominently facilitates the formation of corticalF-actin on the HFHS chow. One can speculate that theenhanced formation of cortical F-actin may increase themechanical strength per unit area, and it inhibits the adi-pocyte hypertrophy in the status of obesity induced byHFHS chow. In addition, the formation of cortical F-actinenhances the translocation of GLUT4, and it may improvethe insulin sensitivity in ACAM Tg mice. We have alsogenerated ACAM knockout mice; however, they were smallfor age and died around 14 weeks, and we were unable toanalyze the size of adipocytes.
In 3T3-L1 preadipocytes, the expression of ACAM isinduced by the cAMP-PKA-C/EBPb pathway, and it con-centrates on the cellular processes. ACAM expressiondeclines and again appears on cell surface of the fullydifferentiated 3T3-L1 adipocytes. ACAM Tg mice areprotected from obesity and insulin resistance. The cur-rent investigation also provides evidence that the ma-ture adipocytes develop the zonula adherens with themolecular organization of ACAM and cortical F-actin inACAM Tg mice on HFHS chow. The promotion andmaintenance of cortical F-actin by targeting ACAM con-stitute a candidate for the new therapeutic modalities inthe treatment of obesity and diabetes (SupplementaryFig. 9).
Funding. This work was supported by Japan Society for the Promotion ofScience Grant-in-Aid for Scientific Research (25126716, 26293218, 26461361,26461362, and 40620753). K.M. is supported by the Okayama MedicalFoundation and is a recipient of the Biological Study Award for Encouragement(Ryobi Teien Memory Foundation).Duality of Interest. J.W. receives speaker honoraria from Astellas,Boehringer Ingelheim, Novartis, Novo Nordisk, and Tanabe Mitsubishi andreceives grant support from Bayer, Daiichi Sankyo, Kyowa Hakko Kirin, MerckSharp & Dohme, Novo Nordisk, Otsuka, Torii, Pfizer, Takeda, Taisho Toyama, andTanabe Mitsubishi. No other potential conflicts of interest relevant to this articlewere reported.Author Contributions. K.M., J.E., K.H., and J.W. participated in thedesign of the whole study. K.M., J.E., K.H., A.N., and J.W. participated in thegeneration of ACAM Tg mice. Cell culture studies were performed by K.M., J.E.,A.N., A.K., M.S., and H.C. Electron microscopy was performed by M.F. and J.W.Western blot analyses were performed by D.O., K.T., and F.O. K.M., K.T., F.O.,and J.W. conceived of the study, participated in coordination, performed thestatistical analyses, and helped to draft the manuscript. All authors read andapproved the final manuscript. J.W. is the guarantor of this work and, as such,had full access to all the data in the study and takes responsibility for the integrityof the data and the accuracy of the data analysis.
References1. Shin CS, Lecanda F, Sheikh S, Weitzmann L, Cheng SL, Civitelli R. Relativeabundance of different cadherins defines differentiation of mesenchymal pre-cursors into osteogenic, myogenic, or adipogenic pathways. J Cell Biochem2000;78:566–5772. Castro CH, Shin CS, Stains JP, et al. Targeted expression of a dominant-negative N-cadherin in vivo delays peak bone mass and increases adipogenesis.J Cell Sci 2004;117:2853–28643. Eguchi J, Wada J, Hida K, et al. Identification of adipocyte adhesion mol-ecule (ACAM), a novel CTX gene family, implicated in adipocyte maturation anddevelopment of obesity. Biochem J 2005;387:343–3534. Tsuruga H, Kumagai H, Kojima T, Kitamura T. Identification of novelmembrane and secreted proteins upregulated during adipocyte differentiation.Biochem Biophys Res Commun 2000;272:293–2975. Raschperger E, Engstrom U, Pettersson RF, Fuxe J. CLMP, a novel memberof the CTX family and a new component of epithelial tight junctions. J Biol Chem2004;279:796–8046. Makino H, Yamasaki Y, Haramoto T, et al. Ultrastructural changes of ex-tracellular matrices in diabetic nephropathy revealed by high resolution scanningand immunoelectron microscopy. Lab Invest 1993;68:45–557. Sakaue H, Ogawa W, Matsumoto M, et al. Posttranscriptional control ofadipocyte differentiation through activation of phosphoinositide 3-kinase. J BiolChem 1998;273:28945–28952
1266 ACAM in Obesity Diabetes Volume 65, May 2016

8. Parsons WJ, Ramkumar V, Stiles GL. Isobutylmethylxanthine stimulatesadenylate cyclase by blocking the inhibitory regulatory protein, Gi. Mol Pharmacol1988;34:37–419. Chernogubova E, Cannon B, Bengtsson T. Norepinephrine increases glucosetransport in brown adipocytes via beta3-adrenoceptors through a cAMP, PKA,and PI3-kinase-dependent pathway stimulating conventional and novel PKCs.Endocrinology 2004;145:269–28010. Cao Z, Umek RM, McKnight SL. Regulated expression of three C/EBP iso-forms during adipose conversion of 3T3-L1 cells. Genes Dev 1991;5:1538–155211. Birsoy K, Chen Z, Friedman J. Transcriptional regulation of adipogenesis byKLF4. Cell Metab 2008;7:339–34712. Sze KL, Lee WM, Lui WY. Expression of CLMP, a novel tight junction protein,is mediated via the interaction of GATA with the Kruppel family proteins, KLF4and Sp1, in mouse TM4 Sertoli cells. J Cell Physiol 2008;214:334–34413. Chun TH. Peri-adipocyte ECM remodeling in obesity and adipose tissuefibrosis. Adipocyte 2012;1:89–9514. Aslan H, Altunkaynak BZ, Altunkaynak ME, Vuraler O, Kaplan S, Unal B.Effect of a high fat diet on quantitative features of adipocytes in the omentum:an experimental, stereological and ultrastructural study. Obes Surg 2006;16:1526–153415. Van Hul M, Lupu F, Dresselaers T, Buyse J, Lijnen HR. Matrix metalloproteinaseinhibition affects adipose tissue mass in obese mice. Clin Exp Pharmacol Physiol2012;39:544–550
16. Schiller PC, D’Ippolito G, Brambilla R, Roos BA, Howard GA. Inhibition ofgap-junctional communication induces the trans-differentiation of osteoblasts toan adipocytic phenotype in vitro. J Biol Chem 2001;276:14133–1413817. Yanagiya T, Tanabe A, Hotta K. Gap-junctional communication is requiredfor mitotic clonal expansion during adipogenesis. Obesity (Silver Spring) 2007;15:572–58218. Yeganeh A, Stelmack GL, Fandrich RR, Halayko AJ, Kardami E, Zahradka P.Connexin 43 phosphorylation and degradation are required for adipogenesis.Biochim Biophys Acta 2012;1823:1731–174419. Farquhar MG, Palade GE. Junctional complexes in various epithelia. J CellBiol 1963;17:375–41220. Revel JP, Karnovsky MJ. Hexagonal array of subunits in intercellularjunctions of the mouse heart and liver. J Cell Biol 1967;33:C7–C1221. Grosely R, Sorgen PL. A history of gap junction structure: hexagonal arraysto atomic resolution. Cell Commun Adhes 2013;20:11–2022. Huang KC, Yasruel Z, Guérin C, Holland PC, Nalbantoglu J. Interaction of theCoxsackie and adenovirus receptor (CAR) with the cytoskeleton: binding to actin.FEBS Lett 2007;581:2702–270823. Patzke C, Max KE, Behlke J, et al. The coxsackievirus-adenovirus receptorreveals complex homophilic and heterophilic interactions on neural cells. J Neurosci2010;30:2897–291024. Nobusue H, Onishi N, Shimizu T, et al. Regulation of MKL1 via actin cy-toskeleton dynamics drives adipocyte differentiation. Nat Commun 2014;5:3368
diabetes.diabetesjournals.org Murakami and Associates 1267





























