Embed Size (px)
Citation preview

5/12/2018 artigo premio n bel 2004 - ubiquitina proteassoma - slidepdf.com
http://slidepdf.com/reader/full/artigo-premio-nobel-2004-ubiquitina-proteassoma 1
QUÍMICA NOVA NA ESCOLA N° 20, NOVEMBRO 2004Prêmio Nobel de Química 2004
Recebido em 28/10/04, aceito em 3/11/04
ATUALIDADES EM QUÍMICA
Nas últimas décadas, é notável,mas não surpreendente, afreqüente premiação com o
Nobel de Química ou de Medicina dedescobertas de estruturas e proces-sos moleculares de crucial impactona Biologia e saúde humana. Afinal,as demandas sociais por informaçãonas áreas da saúde, meio ambiente,tecnologia “verde”, computação e se-gurança continuam acrescer vertiginosa-mente e a Química seenraíza rapidamentenas bases de váriasáreas do conhecimen-to. Ela passa a cons-tituir-se cada vez maisem linguagem privile-giada para discussão
e compreensão dosfenômenos da vida. OPrêmio Nobel de Química de 2004 foiconferido a três pesquisadores (AaronCiechanover, Avram Hershko e IrwinRose) que conjuntamente desvenda-ram o processo de degradação deproteínas sinalizado por cadeias depoliubiquitina. Dois deles são israe-
Marilene Demasi e Etelvino J.H. Bechara
O Prêmio Nobel de Química de 2004 foi outorgado a três pesquisadores que desvendaram como ocorre oprocesso de degradação celular de proteínas mediado por ubiquitina. Este artigo relata como os estudosdesse processo avançaram, desde as descobertas iniciais no final da década de 70 até os dias de hoje.
Prêmio Nobel, ubiquitina, proteólise, bioquímica celular
lenses, sendo esta a primeira vez queum Prêmio Nobel em uma área deciências é outorgado a pesquisa-dores de Israel.
Neste artigo procuramos recupe-rar os passos percorridos por Ciecha-nover, Hershko e Rose que culmina-ram com o prêmio da Academia RealSueca de Ciências. Primeiramente,salientamos que nada mais justo do
que esse reconhe-cimento, uma vezque esses pesqui-sadores contribuí-ram para a desco-berta e elucidaçãode um processoenvolvido numaampla rede de re-gulação bioquí-
mica das células.Essa descoberta
revelou uma dinâmica intracelular deregulação fina, inimaginável atéentão, que inclui desde o controle dequalidade das sínteses protéicas atéa regulação do ciclo e da respostacelular a diversos estímulos fisioló-gicos e patológicos. As proteínas têm
vida média bastante variável, de 30segundos até muitos dias. As proteí-nas degradadas pelo processo sina-lizado pela proteína ubiquitina têmvida média compreendida entre se-gundos e algumas horas. Entre essasproteínas estão incluídas aquelas quepossuem defeitos conseqüentes deerros de síntese e, muito importante,aquelas que participam da sinaliza-ção e regulação celular e que devem,num dado momento, ser imediata-mente degradadas para que a célulase divida, interrompa seu cresci-mento, “morra” ou simplesmente ex-ecute adequadamente suas funções.Portanto, “decisões” celulares preci-sas, tais como parada de crescimentoe morte celular (apoptose), são regu-ladas por esse processo, além da res-
posta imunológica e antiinflamatória.Naturalmente, espera-se que um pro-cesso tão abrangente como o desco-berto por esses pesquisadores tenhareflexos em inúmeras doenças huma-nas e aponte possibilidades paraminimizar os seus efeitos ou mesmorevertê-las.
Degradação protéica
A primeira referência à degrada-ção intracelular de proteínas foi feita
A seção “Atualidades em Química” procura apresentar assuntos que mostrem como a Química é uma ciência viva,
seja com relação a novas descobertas, seja no que diz respeito à sempre necessária revisão de conceitos.
“Decisões” celularesprecisas, tais como parada
de crescimento e mortecelular (apoptose), além da
resposta imunológica eantiinflamatória, são
reguladas pelo processode degradação de
proteínas mediado por
ubiquitina

5/12/2018 artigo premio n bel 2004 - ubiquitina proteassoma - slidepdf.com
http://slidepdf.com/reader/full/artigo-premio-nobel-2004-ubiquitina-proteassoma 2
16
QUÍMICA NOVA NA ESCOLA N° 20, NOVEMBRO 2004Prêmio Nobel de Química 2004
por Schoenheimer em 1942, que,através de marcação radioisotópica,demonstrou que proteínas são cons-tantemente sintetizadas e desmon-tadas. A partir de então, várias pro-teases envolvidas na degradaçãoprotéica não-dependente de energiaforam descritas. Uma das primeirasa ser amplamente estudada foi atripsina, na luz intestinal, responsávelpela catálise da hidrólise de proteínas
ingeridas na dieta alimentar (proteó-lise extracelular).O processo intracelular de degra-
dação protéica dependente de ener-gia, captada na forma de ATP, foi des-crito pela primeira vez por Sympsonem 1953 e retomado por Hershko em1971. Este pesquisador, durante umestágio de pós-doutoramento nolaboratório de Gordon Tomkins, naUniversidade da Califórnia, em SãoFrancisco, publicou um trabalho reali-zado com cultura de células hepáti-cas, mostrando que a degradaçãointracelular da enzima tirosina amino-transferase era um processo depen-dente de ATP. Na época, o fato de quea degradação de proteínas pudesseconsumir energia era paradoxal, umavez que a proteólise é uma reação dehidrólise e, portanto, um processoexotérmico. Hershko, no entanto, in-sistiu nessa investigação exatamentepela aparente contradição. Suas con-clusões nesse trabalho, que foi consi-
Proteólise
Proteólise é a reação de hidró-lise das ligações peptídicas forma-das entre os aminoácidos de umaproteína. Dependendo da protea-se, os produtos de hidrólise pro-téica catalisada por proteases po-
dem ser tanto pequenos peptídeosquanto aminoácidos isolados.Uma técnica bastante utilizadapara a medida de proteólise é amarcação isotópica de proteínas.Consiste na utilização, dentre umasérie de compostos radioativos, doisótopo radiativo 125I, quando a pro-teína é marcada in vitro, ou de mis-tura dos aminoácidos cisteína/ metionina marcados com outroisótopo radiativo, o 35S, quando a
marcação é feita em proteínasintracelulares. Essa mistura é adi-cionada ao meio de cultura celulare as células utilizam esses aminoá-cidos marcados para a síntese pro-téica; portanto, as proteínas intra-celulares ficam rotuladas comradioisótopos. As proteínas podemser precipitadas com alguns áci-dos fortes, como, por exemplo, oácido tricloroacético ou perclórico.No entanto, nem todos os produtosde hidrólise são precipitados poresses ácidos, somente peptídeoscom cadeia acima de 12 aminoáci-dos. A avaliação do grau de pro-teólise se faz tomando-se o con-teúdo protéico, tanto de proteínasmarcadas com 125I quanto com 35S;precipitando-se as proteínas comácido tricloroacético e, após centri-fugação para separar o precipitadoprotéico da fração solúvel, faz-se
a contagem radioativa na fraçãosolúvel, uma vez que os produtosde hidrólise protéica (pequenospeptídeos ou aminoácidos) nãosão precipitados com ácido triclo-roacético e, portanto, são recupe-rados na fração solúvel. Essa téc-nica permite a avaliação do graude proteólise em lisado celular, emcélulas intactas ou mesmo in vitro,incubando-se proteínas marcadascom proteases purificadas.
Ubiquitina
A ubiquitina é uma proteína de ape-nas 76 aminoácidos. Foi isolada pelaprimeira vez de timo bovino, em 1972.Em seguida, constatou-se que essaproteína era encontrada em todos ostecidos e organismos eucarióticos, po-
rém nada era conhecido sobre suafunção intracelular. Na mesma época,foi descrita sua ligação covalente aoutra proteína, uma histona (proteínanuclear), desconhecendo-se, no en-tanto, a razão disto. Sua função intra-celular foi quase que totalmente eluci-dada no final da mesma década porCiechanover, Hershko e Rose.
Figura 1: Estrutura tridimensional da ubiquitina. As setas mostradas indicam estru-turas secundárias do dobramento protéico denominadas de folha β e a espiral nocentro da estrutura refere-se a outro tipo de dobramento denominado de α hélice.
derado um marco para investigaçõesposteriores, convergiam para a possi-bilidade da existência de um pro-cesso proteolítico mais complexo doque aqueles conhecidos até então.Concluiu afirmando que: “Estadependência por ATP para a degra-dação protéica intracelular parece serincompatível com um mecanismoproteolítico simples e sugere que oprocesso seja de múltiplas etapas...
Pode ser que o ATP esteja diretamen-te envolvido na modificação da proteí-na antes de sua degradação ou naformação de um metabólito interme-diário”.
Um importante avanço nos estu-dos da proteólise ATP-dependente foio modelo experimental publicado porEtlinger e Goldberg, em 1977, utili-zando lisado de reticulócitos de coe-lho. Lisado é o material citoplasmáticoobtido experimentalmente pelo rompi-mento mecânico de células e poste-rior centrifugação para se obter afração solúvel do citoplasma celular.Esse sistema levantou a possibilidadede se poder distinguir a atividadeproteolítica lisossomal da não-lisos-somal. O lisossoma é o comparti-mento intracelular responsável peladegradação de inúmeras macromolé-culas, incluindo proteínas extracelu-lares, que são trazidas para o interiordas células pelo processo de endo-citose, além de proteínas de membra-

5/12/2018 artigo premio n bel 2004 - ubiquitina proteassoma - slidepdf.com
http://slidepdf.com/reader/full/artigo-premio-nobel-2004-ubiquitina-proteassoma 3
QUÍMICA NOVA NA ESCOLA N° 20, NOVEMBRO 2004Prêmio Nobel de Química 2004
nas e proteínas intracelulares de vidamédia longa. Um exemplo de proteínacom vida longa é a hemoglobina:aproximadamente 110 dias. No mo-delo de estudo proteolítico com lisado
de eritrócitos foi possível a diferen-ciação entre atividade proteolíticanão-lisossomal e lisossomal, uma vezque as enzimas lisossomais têm ativi-dade proteolítica em pH ácido, sendo
que a atividade ATP-dependente,
então descrita, possuía atividade óti-ma em pH ligeiramente alcalino,nominalmente 7,8.
Foi a partir de então, entre o finalda década de 70 e início da de 80,que Ciechanover, Hershko e Rose de-senvolveram uma série de estudosque culminaram com a elucidação domecanismo de proteólise dependentede ATP e da ubiquitina. Os encontrosque mantiveram e o trabalho realizadoem conjunto foram possíveis graçasa uma série de estágios sabáticos de
Figura 2: Seqüência de eventos desde a marcação da proteína-alvo com ubiquitina (Ub)até sua degradação pelo proteassoma. (1) A Ub liga-se covalentemente à enzima E1denominada de ativadora de Ub. Essa etapa é dependente de ATP. (2) A Ub é transferidapara a enzima E2, conjugadora de Ub, que, uma vez complexada à Ub, interage com aenzima E3, denominada de ligase. É nessa etapa (3) que a Ub é transferida para o subs-
trato ou proteína-alvo. As proteínas-alvo são “selecionadas” (reconhecidas) pela enzimaE3 por possuírem “sinais” em sua estrutura. (4) A etapa anterior é repetida até a formaçãode uma cadeia de poliubiquitina na proteína-alvo (5). A cadeia assim formada será reco-nhecida por proteínas integrantes da unidade regulatória do proteassoma. (6) A proteína-alvo será desdobrada por proteínas especializadas das unidades regulatórias (essa etapatambém é ATP-dependente) e dirigida para a câmera catalítica (proteassoma 20S) parahidrólise. Os fragmentos de degradação são liberados no lado oposto do proteassoma.
Degradação protéica sinalizada
por ubiquitina
Este processo consiste na “mar-cação” de proteínas intracelularespara a degradação por intermé-dio de adições sucessivas de ubi-quitina à proteína-alvo, criando-se
dessa maneira uma cadeia depoliubiquitina na proteína quesofrerá degradação. Envolve aparticipação de três classes deenzimas ubiquitinadoras denomi-nadas de E1, E2 e E3. A cadeiade poliubiquitina e, portanto, aproteína alvo, será reconhecidapor unidades regulatórias do pro-teassoma, que é a protease intra-celular responsável pela degrada-ção de proteínas rotuladas com
cadeias de poliubiquitina. Umavez havendo o reconhecimentoda cadeia de poliubiquitina poressas unidades do proteassoma,a proteína alvo será desdobrada,para ser hidrolisada gerando ca-deias polipeptídicas de 3-21 ami-noácidos. Esses pequenos peptí-deos são liberados da câmeraproteolítica e hidrolisados poruma série de peptidases intrace-lulares, resultando unidades de
aminoácidos que podem entãoser reutilizados pela célula para asíntese protéica. A cadeia depoliubiquitina é liberada da proteí-na e as unidades de ubiquitinasão recuperadas também porprocesso enzimático. Enzimasdenominadas desubiquitinadorasestão envolvidas nesse processo.

5/12/2018 artigo premio n bel 2004 - ubiquitina proteassoma - slidepdf.com
http://slidepdf.com/reader/full/artigo-premio-nobel-2004-ubiquitina-proteassoma 4
18
QUÍMICA NOVA NA ESCOLA N° 20, NOVEMBRO 2004Prêmio Nobel de Química 2004
Ciechanover e Hershko no laboratóriode Rose, na Pensilvânia (EUA).
A descoberta
Partindo do princípio de que ofenômeno que estavam observandodependia de uma proteína (protease)que efetuava a hidrólise do substrato-alvo na presença de ATP, e que essaprotease era parte integrante do con-teúdo protéico do lisado de reticuló-citos, esses pesquisadores aplicaramo lisado a uma coluna cromatográficade Sepharose® DEAE com o objetivode separar (isolar) essa protease des-conhecida. Duas das frações obtidasnesse processo, quando eram mistu-radas, reconstituíam a proteólise ATP-dependente. No entanto, quando
testadas separadamente não apre-sentavam atividade. Continuando apurificação das proteínas contidas emcada uma dessasfrações, eles isola-ram uma proteínatermoestável demassa molecular9000 u. Essa proteí-na foi identificadacomo um compo-nente ativo para queocorresse proteólise ATP-dependente.Quando essa proteí-na era adicionada àoutra fração ocorria proteólise, porémnunca na sua ausência. Denomina-ram essa proteína de APF-1 ( Active
Principle of Fraction 1). Ela foi identi-ficada em 1980 como sendo a ubiqui-tina que já havia sido descrita anterior-mente, porém desconhecia-se suaatividade. Rapidamente, verificou-seque a molécula dessa proteína tinha76 resíduos de aminoácidos e eraaltamente conservada nas células
eucarióticas, sendo essencialmenteidêntica em organismos tão distintoscomo uma levedura ou uma vaca.
Continuando o fracionamento portécnica cromatográfica, conseguiramidentificar uma fração que continha asenzimas tiólicas E1 e E2 e a enzimaE3 e, numa outra fração, identificaramuma proteína de alta massa molecu-lar, também essencial para a atividadeque haviam descrito. Muito provavel-mente essa proteína era a proteaseque buscavam. No entanto, foi so-
mente 10 anos mais tarde que ela foiidentificada por outros investigadorescomo sendo o proteassoma, na ver-dade um complexo protéico de altamassa molecular capaz de reconhe-cer e catalisar a hidrólise de proteínasmarcadas com cauda de ubiquitina.Os experimentos descritos acima e asconclusões a que chegaram estãodescritos em publicações de 1978 e1979 (Ciechanover et al., 1978;Hershko et al., 1979).
Ciechanover, Hershko e Rosecontinuaram buscando os elementosdo processo de proteólise ATP-de-pendente, desviando-se daí emdiante da protease e concentrando-se nos demais componentes do pro-cesso.
Uma vez tendo purificado a ubiqui-tina (Ub), chamada de APF-1 até
aquele momento,eles a marcaram comiodo radioativo (125I) e
demonstraram ine-quivocamente queinúmeras proteínasno lisado de reticuló-citos estavam ligadascovalentemente àUb. Logo em segui-da, numa outra sériede experimentos,fizeram marcação de
três proteínas diferentes com 125I aoinvés de marcar a Ub. Utilizandoessas proteínas como substratos da
O proteassoma
O proteasoma foi descrito em meados da década de 80 por vários gruposde pesquisa que investigavam diferentes temas bioquímicos. Esse complexoprotéico (constituído por várias cadeias peptídicas) recebeu diferentesdenominações antes de ser definitivamente chamado de proteassoma( proteasome no inglês: protea de protease e some de soma, partícula), devidoà sua alta massa molecular. O proteassoma tem massa molecular de 2500ku. Possui uma unidade central catalítica denominada de 20S, flanqueadanas duas extremidades por unidades regulatórias que reconhecem a cadeiade poliubiquitina na proteína-alvo, desdobram a proteína para hidrólise edirecionam a proteína para a câmera catalítica, denominada de proteassoma20S. Acredita-se que 80% das proteínas de origem intracelular sejamdegradadas pelo proteassoma. As proteínas alvo de degradação peloproteassoma são aquelas de vida média ultra-curta e curta: de algunssegundos até algumas horas, desde a síntese até hidrólise.
Figura 3: Micrografia eletrônica do proteassoma. Ao centro, a unidade catalítica 20S.Essa unidade é formada por subunidades denominadas de α e β. Cada uma delaspossui sete cadeias peptídicas, diferentes entre si, formando heptâmeros que serepetem dos dois lados. A atividade hidrolítica está nas cadeias β. A estrutura doproteassoma é simétrica: o substrato pode se ligar a qualquer uma das duasextremidades das unidades regulatórias.
No final da década de 70,Ciechanover, Hershko e
Rose identificaram umaproteína de alta massamolecular, essencial para oprocesso de degradação
protéica. Ela só foiidentificada como o
proteassoma dez anos mais tarde, por outros
pesquisadores
5/12/2018 artigo premio n bel 2004 - ubiquitina proteassoma - slidepdf.com
http://slidepdf.com/reader/full/artigo-premio-nobel-2004-ubiquitina-proteassoma 5
QUÍMICA NOVA NA ESCOLA N° 20, NOVEMBRO 2004Prêmio Nobel de Química 2004
proteólise, eles demonstraram quemúltiplas unidades de Ub podiamconjugar-se com a proteína-alvo. Es-sa seqüência de trabalhos (Ciecha-nover et al., 1980; Hershko et al.,1980) incluiu ainda uma série dedetalhes sobre como é a ligação deUb às proteínas-alvo e, também, ante-
cipou a existência de uma enzima queremove da proteína-alvo a cauda depoliubiquitina.
Entre 1981 e 1983, tendo já sidocriada a hipótese sobre o sistema demarcação com Ub da proteólise ATP-dependente, eles se dedicaram à elu-cidação completa do processo. Os
trabalhos dessa época culminaramcom a purificação e caracterizaçãodas três enzimas ubiquitinadoras (E1-E3) e elucidaram o mecanismo com-pleto de marcação por Ub.
Os trabalhos desse período sãoainda textos-referência para a descri-ção do sistema. Eles utilizaram proto-
colos experimentais bastante elegan-tes àquela época. Purificaram a enzi-ma E1 por cromatografia de afinidadeligando a Ub covalentemente a umacoluna de Sepharose®. O lisado dereticulócitos era aplicado à coluna,sendo que a enzima E1 ficava retidana coluna por ligação à Ub; dessamaneira conseguiram purificar e de-monstrar o tipo de ligação covalenteentre Ub e E1. Utilizando-se de meto-dologia semelhante conseguiram
purificar as duas outras enzimas: E2e E3. Neste caso, usaram como estra-tégia eluições diferenciadas - lem-brando que a eluição consiste naetapa cromatográfica que usa um sol-vente ou solução para retirar o mate-rial retido na coluna. Dessa maneiraisolaram as três enzimas e demons-traram que E2 liga-se a E1 na colunadiferentemente de E3, que era eluídada coluna isoladamente. Todos essespassos estão descritos em publica-
ções feitas por eles em 1981 e 1983(Ciechanover et al., 1981; Hershko et
al., 1983).
Implicações fisiológicas e patológicas
O processo de degradação deproteínas sinalizado por ubiquitinaparticipa de inúmeros processos in-tracelulares muito importantes para amanutenção da homeostase ou dadefesa celular. Alguns exemplos sãoenumerados a seguir.
Sistema imunológico
Alguns dos peptídeos geradosdentro da célula pelo sistema Ub-pro-teassoma são utilizados pela célulacomo antígenos de superfície celulare são reconhecidos pelos linfócitos T.Se esses peptídeos não forem pro-dutos de uma proteína humana, oslinfócitos T reconhecem o peptídeo nasuperfície celular como um antígeno,um componente estranho, e
O método cromatográfico
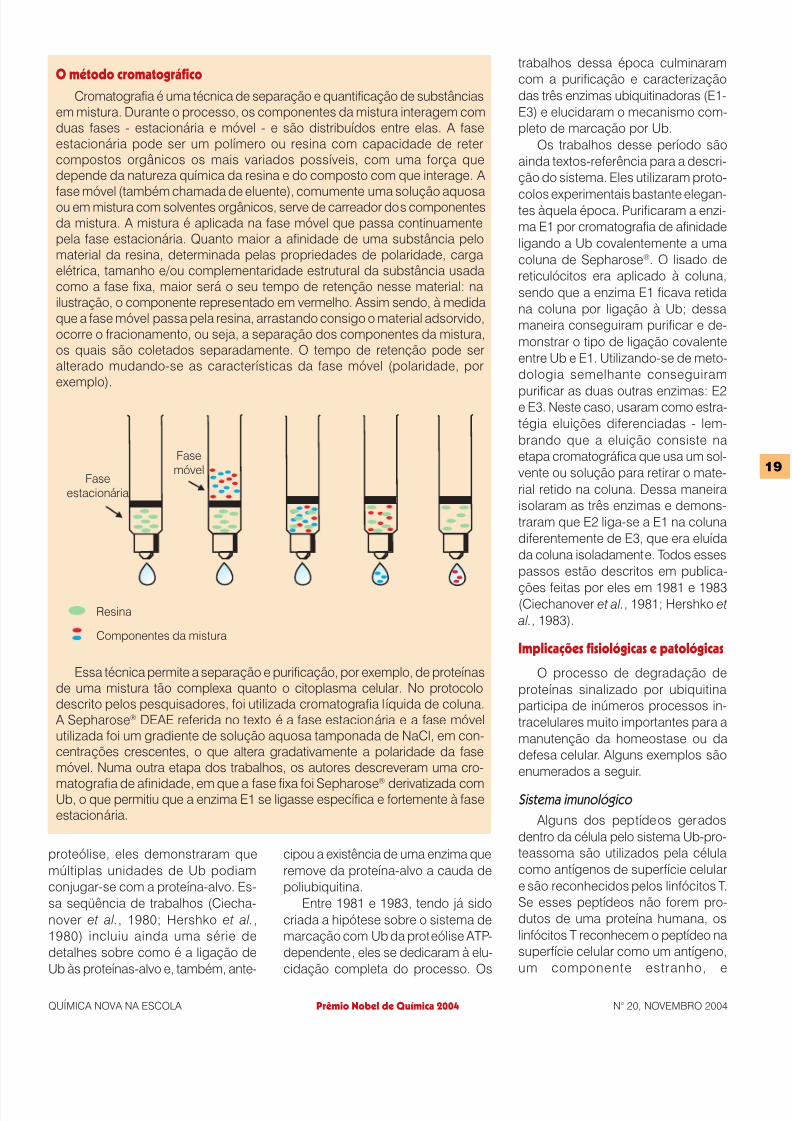
Cromatografia é uma técnica de separação e quantificação de substânciasem mistura. Durante o processo, os componentes da mistura interagem comduas fases - estacionária e móvel - e são distribuídos entre elas. A faseestacionária pode ser um polímero ou resina com capacidade de retercompostos orgânicos os mais variados possíveis, com uma força quedepende da natureza química da resina e do composto com que interage. A
fase móvel (também chamada de eluente), comumente uma solução aquosaou em mistura com solventes orgânicos, serve de carreador dos componentesda mistura. A mistura é aplicada na fase móvel que passa continuamentepela fase estacionária. Quanto maior a afinidade de uma substância pelomaterial da resina, determinada pelas propriedades de polaridade, cargaelétrica, tamanho e/ou complementaridade estrutural da substância usadacomo a fase fixa, maior será o seu tempo de retenção nesse material: nailustração, o componente representado em vermelho. Assim sendo, à medidaque a fase móvel passa pela resina, arrastando consigo o material adsorvido,ocorre o fracionamento, ou seja, a separação dos componentes da mistura,os quais são coletados separadamente. O tempo de retenção pode seralterado mudando-se as características da fase móvel (polaridade, porexemplo).
Resina
Componentes da mistura
Fase
estacionária
Fase
móvel
Essa técnica permite a separação e purificação, por exemplo, de proteí nasde uma mistura tão complexa quanto o citoplasma celular. No protocolodescrito pelos pesquisadores, foi utilizada cromatografia lí quida de coluna. A Sepharose® DEAE referida no texto é a fase estacionária e a fase móvelutilizada foi um gradiente de solução aquosa tamponada de NaCl, em con-centrações crescentes, o que altera gradativamente a polaridade da fasemóvel. Numa outra etapa dos trabalhos, os autores descreveram uma cro-
matografia de afinidade, em que a fase fixa foi Sepharose® derivatizada comUb, o que permitiu que a enzima E1 se ligasse especí fica e fortemente à faseestacionária.

5/12/2018 artigo premio n bel 2004 - ubiquitina proteassoma - slidepdf.com
http://slidepdf.com/reader/full/artigo-premio-nobel-2004-ubiquitina-proteassoma 6
20
QUÍMICA NOVA NA ESCOLA N° 20, NOVEMBRO 2004Prêmio Nobel de Química 2004
Abstract: Nobel Prize in Chemistry 2004: ATP-Dependent Proteolysis of Ubiquitin-Marked Proteins - The Nobel Prize in Chemistry 2004 was awarded to three scientists that unveiled how the process
of ubiquitin-mediated cellular degradation of proteins occurs. This article reports how the studies of this process advanced, from the initial discoveries in the end of the seventies up to nowadays.
Keywords: Nobel Prize, ubiquitin, proteolysis, celular biochemistry
Os laureados
Aaron Ciechanover Aaron Ciechanover Aaron Ciechanover Aaron Ciechanover Aaron Ciechanover ----- Nascido em1947 na cidade de Haifa, Israel, foialuno de doutorado (1981) de Avram Hershko, na área de Medi-cina, no Instituto de Tecnologia deIsrael (Technion), em Haifa. Atual-mente é professor de Bioquí micado Instituto de Pesquisas em Ciên-cias Médicas Famí lia Rappaport doTechnion. Em 2000, já recebera,juntamente com Hershko, o Prêmio
Albert Lasker para Pesquisa Mé-dica Básica; em 2003, foi-lhe outor-gado o Prêmio Israel pela desco-berta do “sistema ubiquitina”.
Avram Hershko Avram Hershko Avram Hershko Avram Hershko Avram Hershko ----- Nascido em 1937 em Karcag, na Hungria, emigroupara Israel em 1950 e hoje é cidadão israelense. Doutorou-se em 1969 pelaUniversidade Hebraica, em Jerusalém, sendo professor no mesmo centrode pesquisa onde atua Ciechanover. Em 1994, já recebera o Prêmio Israel e,em 2001, o Prêmio Wolf de Medicina.
Ir Ir Ir Ir Ir win Rosewin Rosewin Rosewin Rosewin Rose ----- Nascido em 1926 em Nova Iorque (EUA),
doutorou-se em 1952 pela Universidade de Chicago.Foi professor na Faculdade de Medicina da Universi-dade de Yale, de 1953 a 1963, quando se mudou parao Centro Fox Chase para Câncer, na Filadélfia, do qualse aposentou em 1995. Atualmente é pesquisador emé-rito no Departamento de Fisiologia e Biofí sica da Facul-
dade de Medicina da Universidade da Califórnia em Irvine, Califórnia (EUA).
Referências bibliográficas
CIECHANOVER, A.; HELLER, H.;
ELIAS, S.; HAAS, A.L. e HERSHKO, A.
ATP-dependent conjugation of reticulo-cyte proteins with the polypeptide required
for protein degradation. Proc. Natl. Acad.
Sci., v. 77, p. 1365-1368, 1980.
CIECHANOVER, A.; HELLER, H.; KATZ-
ETZION, R. e HERSHKO, A. Activation of
the heat-stable polypeptide of the ATP-
dependent proteolytic system. Proc. Natl.
Acad. Sci., v. 78, p. 761-765, 1981.
CIECHANOVER, A.; HOD, Y. e
HERSHKO, A. A heat-stable polypeptide
component of an ATP-dependent pro-
teolytic system from reticulocytes.
Biochem. Biophys. Res. Commun., v. 81,
p. 1100-1105, 1978.HERSHKO, A.; CIECHANOVER, A.;
HELLER, H.; HAAS, A.L. e ROSE, I.A. Pro-
posed role of ATP in protein breakdown:
Conjugation of proteins with multiple
chains of the polypeptide of ATP-depen-
dent proteolysis. P roc. Natl. Acad. Sci., v.
77, p. 1783-1786, 1980.HERSHKO, A.; CIECHANOVER, A. e
ROSE, I.A. Resolution of the ATP-depen-
desencadeiam o processo que levaráà destruição daquela célula. Esse me-
canismoé muito importante em casos
de infecção viral. Uma célula infec-
tada com um ví rus terá como produ-
tos de hidrólise peptí deos originários
de proteí nas virais que serão reconhe-
cidos pelos linfócitos T como estra-
nhos ao organismo e, portanto, acélula será destruí da, evitando a con-
tinuidade da replicação viral.
Resposta inflamatória
As células possuem proteí nas de-
nominadas fatores de transcrição.
Essas proteí nas se ligam ao DNA e
induzem a transcrição de determi-
nados genes. Um desses fatores de
transcrição, denominado NFkB, loca-
liza-se no citoplasma associado a
uma proteí na que inibe sua atividadejunto ao DNA. Quando a célula éinfectada por uma bactéria, esse inibi-
dor sofre uma alteração estrutural, éubiquitinado e degradado pelo pro-
teassoma. Dessa maneira a proteí na
NFkB fica livre e desloca-se para o
núcleo, desencadeando a transcrição
de inúmeros genes relacionados àresposta inflamatória que se constitui
em um processo vital de defesa.
Terapêutica anti-tumoral
Inibidores do proteassoma ou daubiquitinação protéica estão prestesa serem introduzidos na terapêuticaanti-tumoral. Essa estratégia se ba-seia no fato de que a inibição irrever-sí vel do sistema Ub-proteassoma levaa célula à morte celular programada
dent proteolytic system from reticulo-
cytes: A component that interacts with
ATP. Proc. Natl. Acad. Sci., v. 76, p. 3107-
3110, 1979.
HERSHKO, A.; HELLER, H.; ELIAS, S.e CIECHANOVER, A. Components of
ubiquitin-protein ligase system. J. Biol.
Chem., v. 258, p. 8206-8214, 1983.
Para saber mais
Na Internet
S í tio da Fundaçã o Nobel: www.
nobel.se.
Hershko (esquerda) e Ciechanover, no Technion.
- apoptose -, muito desejável em setratando de células tumorais, pois im-pede que elas proliferem.
Marilene Demasi ([email protected]), gra-
duada em Ciências Farmacêuticas pela Universi-
dade de São Paulo (USP), mestre em Psicobiologiapela Universidade Federal de São Paulo e doutora
em Ciências Biológicas (Bioquí mica) pela USP,é pes-
quisadora do Instituto Butantã, em São Paulo - SP.
Etelvino J.H. Bechara ([email protected]), gradua-
do em Quí mica e doutor em Ciências (Bioquí mica)pela USP, édocente do Departamento de Bioquí mica
do Instituto de Quí mica da USP, em São Paulo - SP
(http://www.iq.usp.br/wwwdocentes/ebechara). Fo
presidente da Sociedade Brasileira de Quí mica nobiênio 1988-1990.





























