Embed Size (px)
Citation preview

Toda correspondencia relativa a la ASOCIACIÓN ESPAÑOLA DE LIMNOLOGÍA y a la revista LIMNETICA, incluida lapetición de altas, bajas, intercambios, suscripciones y ejemplares atrasados debe dirigirse a la Secretaria de la AsociaciónEspañola de Limnología, C/ Los Ángeles, 33, 46920-Mislata (Valencia).
Los manuscritos de trabajos científicos para su publicación en LIMNETICA deben ser enviados a Joan Armengol,Departament d'Ecologia. Facultat de Biologia. Universitat de Barcelona. Av. Diagonal, 645. 08028-Barcelona.
Limnetica está indexada en las siguientes bases de datos:Aquatic Sciences and Fisheries Abstracts (ASFA); Zoological Record of BIOSIS® database; Freshwater BiologicalAssociation (FBA); NISSC’s FISHLIT database; Sistema de Información en Línea para Revistas Científicas de América Latina,Caribe, España y Portugal (LATINDEX); Library of Natural Sciences of Russian Academy of Science (LNS); Indice Españolde Ciencia y Tecnología (ICYT).
Presidencia: SERGI SABATER. GironaVicepresidencia: JULIA TOJA. SevillaSecretaría: JUAN MIGUEL SORIA. ValenciaTesorería: EUGENIO RICO. Madrid
ASOCIACIÓN ESPAÑOLADE
LIMNOLOGÍA
LIMNETICALIMNETICA es una revista internacional publicada por la Asociación Española de Limnología.
EditorJOAN ARMENGOL
Editores adjuntosISABEL MUÑOZ
FRANCESC SABATER
Editores invitadosIRMA VILA
ALICIA FERNÁNDEZ CIRELLI
Comité editorialJ. Alba. Granada A. Palau. LleidaM.J. Boavida. Lisboa, Portugal C. Pedrós-Alió. BarcelonaX. Casamitjana. Girona D. Planas. Montreal, CanadáJ. Catalán. Barcelona N. Prat. BarcelonaM.A.S. Graça. Coimbra, Portugal A. Quesada. MadridC. Granado. Sevilla K. Simek. Ceske Budejovice, Rep. ChecaR.D. Gulati. Nieuwersluis, Holanda T. Tundisi. Sao Carlos, BrasilR. Margalef. Barcelona E. Vicente. Valencia
H.L. Golterman. Francia
Secretaria de RedacciónELENA GUERRA



Volumen 22. Número 1-2. 2003
LIMNETICARevista de la
Asociación Española de Limnología
Volumen especialII Jornadas Internacionales sobre Eutrofización
de lagos y embalses
EditoresIrma Vila (Universidad de Chile)
Alicia Fernández Cirelli (Universidad de Buenos Aires)Joan Armengol (Universitat de Barcelona)
Publicacions

© Asociación Española de Limnología
Depósito Legal:V-2404-1986
ISSN: 0213-8409
Autoedición: Servei Gràfic NJR, SL
Impresión: Gráficas Rey
Impreso en España
Printed in Spain

Este volumen contiene una serie de artículos pre-sentados en las II Jornadas Internacionales sobreEutrofización de Lagos y Embalses realizado enBarcelona del 2 al 4 de Junio de 2001. EstasJornadas fueron organizadas conjuntamente porla Universidad de Barcelona, Aigües Ter-Llobregat y por el Programa Iberoamericano deCiencia y Tecnología para el Desarrollo(CYTED) como parte de las actividades desarro-lladas dentro del Subprograma XVII Aprove-chamiento y Gestión de Recursos Hídricos en elmarco de la Red Temática XVIIB Eutrofizaciónde de lagos y embalses de CYTED.
Las Jornadas se organizaron de acuerdo conlos criterios que rigen el Programa CYTED-XVII, el intercambio de información y presenta-ción de experiencias entre especialistas de dife-rentes países Iberoamericanos en el tema deaprovechamiento y gestión de recursos hídricos.En estas Jornadas se abordó la problemática dela eutrofización desde tres puntos de vista com-plementarios: 1) Los estudios ecológicos y lasexperiencias de gestión de lagos y embalseseutróficos con el objetivo de mejorar su estadotrófico, 2) La problemática de la calidad delagua desde el punto de vista del abastecimiento,con énfasis en los sistemas de gestión, de vigi-lancia y control del agua que llega a las plantaspotabilizadoras, y 3) La incidencia de laDirectiva Marco del Agua de la UE en los lagosy embalses de los países europeos participantesen el programa CYTED.
Con la asistencia de cerca de 100 participan-tes de de diferentes países Iberoamericanos,Argentina, Brasil, Chile, Colombia, España,Portugal, Uruguay y Venezuela, y la participa-ción de las empresas de abastecimiento de aguapotable de España, Aigües Ter-Llobregat,Aguas de Barcelona, Canal de Isabel II deMadrid, EMASESA de Sevilla, Consorcio deAgua de Bilbao, y Brasil, Companhia deSaneamento Básico do Estado de Sao Paulo, lasJornadas se celebraron de acuerdo con los obje-tivos previamente indicados.
A lo largo de las jornadas se presentaron18 comunicaciones orales en sesiones plenariasen las que se abordaron, entre otros temas, elestudio ecológico de lagos y embalses, ejemplosde gestión y adecuación ambiental para reducirla eutrofización, el efecto de la realización deplanes de saneamiento en cuencas hidrográficassobre el estado trófico de los sistemas lacustres,el efecto de la actividad humana en la evolucióntrófica de los lagos, los ciclos del fósforo y delnitrógeno como factores clave en la eutrofiza-ción, los sistemas de control y seguimiento delos embalses a diferentes escalas temporales, laactividad de los organismos planctónicos desdeel bacterioplancton al zooplancton o el efecto dela luz como factor clave en la actividad del fito-plancton de los embalses. La participación acti-va de algunas de las diferentes empresas deabastecimiento de agua potable a grandes ciuda-des españolas dio lugar a una seria de ponenciassobre modelos de gestión ecosistémica de losembalses asociado a la problemática de pota-bilización del agua. A lo largo de las Jornadasse realizaron mesas redondas sobre eutrofiza-ción, criterios de gestión de los embalses deabastecimiento de agua potable, la DirectivaMarco del Agua de la UE y mejoras técnicas enla potabilización del agua.
Para la realización de las II JornadasInternacionales sobre Eutrofización de Lagos yembalses ha sido fundamental la colaboraciónde diferentes organismos públicos y privadosentre los que destacan de forma especial elPrograma CYTED-XVII, Aigües Ter-Llobregat,la Universidad de Barcelona y el ParqueCientífico de Barcelona.
El desarrollo de algunas de las ponencias ple-narias a dado lugar a los trabajos que se presentanen este volumen de Limnetica. Para su publica-ción se han seguido los criterios se revisión cientí-fica habituales de esta revista con el objetivo deque se sigan los criterios de calidad de la revista.
Este volumen es también un modesto, perosincero homenaje a uno de los participantes en
Exordio

las Jornadas, el Dr. Vicente Clavero de laUniversidad de Málaga. El Dr. Clavero fue invi-tado para presentar una síntesis de los trabajosrealizados durante muchos años sobre el ciclodel fósforo en sistemas acuáticos, en especial enel río Palmones y el embalse de CharcoRedondo. Lamentablemente, su trágica muerteno solo nos ha privado de su trabajo publicadosino que ha truncado una trayectoria científicaque, más que una promesa, era ya una realidad ycon una gran proyección de futuro. Sus compa-ñeros del Departamento de Ecología y losmiembros de su equipo han querido tambiénrendirle homenaje enviando un artículo comoun recuerdo a su memoria. Igualmente, el Dr.Xavier Niell, su director de Tesis, codirector demuchos de sus proyectos y publicaciones, y unbuen amigo, ha escrito la necrológica con la queiniciamos este volumen.
En la medida que estos trabajos publicadosson el resultado de estudios concretos y aplica-ciones a lagos y embalses Iberoamericanos y
cubren un amplio espectro de enfoques y solu-ciones a algunos problemas planteados cumplenuno de los objetivos más importantes delPrograma CYTED-XVII. Por este motivo pen-samos que puede ser una herramienta de trabajode interés para muchos científicos y técnicosque en estos momentos están trabajando en elárea de los recursos hídricos. En igual medida,este volumen supone el resultado de un trabajocomún, de intercambio de ideas y experienciasentre especialitas de diferentes países que estántrabajando con objetivos similares. Por estemotivo los editores de este volumen queremosagradecer desde estas páginas toda la ayudarecibida empezando por los participantes en lasJornadas y por haber elegido este marco y estarevista para la presentación de sus resultados.
Los editores de este volumen Irma Vila
Alicia Fernández CirelliJoan Armengol

EN MEMORIA DE
Vicente Clavero
El agua que corre libreEs el agua que se bebeEl hombre que sueña vive.
C. MarquezEn sueños en el aire. (J. Peña El Lebrijano)
Vicente, nació en 1961, el día de San Elías. Nos dejó el día 14 de Agosto del año 2002.Era licenciado y doctor en Biología por “su” Universidad de Málaga. Me refiero a “su”Universidad porque además de dedicarse a ella, desde alumno, hasta que fue Profesor, actuócomo un elemento dinamizador.Vicente y yo trabajamos siempre juntos durante su investigación sobre EcosistemasAcuáticos. Su capacidad y dedicación le permitieron embestir trabajos de modo versátilsobre micrométodos, su tesina versa sobre una puesta a punto de un microsoxhlet, o biensobre procesos a gran escala, como los balances de nutrientes que preparaba sobre la cuencade “nuestro” río Palmones, en el que realizó su Tesis, sobre su tema de investigación preferi-do, el intercambio de fosfato entre el agua y el sedimento.Empezó a trabajar en el Departamento de Ecología de la Universidad de Málaga en 1985 ydesde entonces hasta el año 2002 confirió alegría y compañerismo a todos. No tenía preven-ciones a la hora de enfocar problemas nuevos, era un entusiasta de la reproducción de proce-sos a gran escala en modelos experimentales en el laboratorio. Desde 1994 era Profesor deEcología de “nuestra” Universidad de la que fue Profesor Titular en 1999. Sus cualidadescomo docente fueron, y son aún, muy estimadas, comunicaba, muy directamente, con losalumnos de “su” Málaga, hablaba igual que ellos y trabajaba arduamente las clases, yo ledecía que las sudaba.Personalmente recuerdo qué gran entusiasmo derrochó explicando una asignatura que él hizopopular: Bioestadística y recuerdo la atracción que ejercía en asignaturas como ImpactoAmbiental y Biogeoquímica de las cuales fue un pionero.Participó activamente en la ejecución de 10 proyectos de investigación y publicó más de40 trabajos de investigación. Su actividad era madrugadora, bromeaba con él diciéndole queabría la Facultad. Su humanidad era formidable, quería a su Virgen de “La Zamarrilla” y le caracterizaba serdel F. C. Barcelona que tanto le hizo sufrir. Era apasionado, le gustaba Curro Romero y ElCamarón. Los que le conocieron comprenden el significado de lo que he escrito, sin estascosas, Vicente no era Vicente.En la flor de su juventud, lleno de fuerza, terminó de reírse, pero su sonrisa queda por los rin-cones de todos los sitios donde le recordamos. Estaremos contigo siempre Vicente.
F. Xavier Niell, Amigo de Vicente y Profesor de “su” Universidad.


Medidas de gestión y adecuación ambiental de embalses frente a la eutrofia
Antoni Palau Ibars
Departamento de Medio Ambiente y Desarrollo Sostenible. Endesa Servicios S.L. Carretera de Tarragona,Km 89,300. 25001-Magraners (Lleida)
RESUMEN
La eutrofización es una alteración prácticamente inherente a la construcción de embalses y a su explotación. Ciertamente,existen diferentes tipos de embalses y no en todos la eutrofia se manifiesta con la misma intensidad. Por la misma razón, nosiempre son aplicables con igual éxito las distintas medidas de atenuación del grado de eutrofia que se conocen.En la ponencia se muestra una revisión cualitativa, pero suficientemente exhaustiva, de las medidas de predicción, prevencióny corrección de la eutrofia, asociables a embalses, en sentido amplio.En cualquier propuesta de control de perturbaciones provocadas por actuaciones humanas en sistemas naturales (como es elcaso de la construcción de embalses), el punto de partida es la predicción; es decir, el reconocimiento previo a la actuación, decual será la respuesta del medio a la perturbación en función de las características de este. Conocida la respuesta más probable,si esta se evalúa como crítica a los efectos de los objetivos de la obra hidráulica proyectada y/o a los costes y funcionalidad(eficiencia) de su explotación, se pueden adoptar una serie de medidas preventivas con el fin de minimizar en lo posible, laeutrofia y/o sus síntomas.En la fase de explotación de un embalse, se reducen los grados de libertad en cuanto a posibles actuaciones de control y ate-nuación de la eutrofia y adquiere una importancia decisiva los usos a los cuales se dedica el embalse (abastecimiento urbano,riegos, hidroelectricidad, etc.) a la hora de escoger las opciones de actuación.Finalmente, la eutrofia es una alteración que no sólo afecta a los embalses, sino también a los tramos de río aguas abajo deellos de modo que es necesario contemplar también medidas de control en este tipo de ambientes.
Palabras clave: Eutrofización, embalse, medidas de control y gestión.
ABSTRACT
Eutrophication is a rather inherent ecological disturbance of dam building and reservoir management. Certainly, there are dif-ferent types of dams and not all of them show the same degree of eutrophication. For the same reason, not all the knowneutrophy control measures can be applied with the same success. In this paper a qualitative but rather exhaustive revision ispresented, about eutrophy’s prediction, prevention, management and correction measures that can be related to dams in ageneral sense. As with any proposal for control of human disturbances in natural systems (such as the case of dam building),the starting point is prediction; that is, the acknowledgement before action is taken, about what will be the response of theenvironment to that perturbation according to its’ characteristics. Once the most likely response in terms of potential eutrophi-cation is known, and if this response is evaluated as a critique to the effects of the projected hydraulic construction objectives,and/or to the costs and functionality (efficiency) of it’s operation, then a number of preventive measures can be adopted tominimize, as much as possible, eutrophy and/or its’ symptoms. During a dam’s operation phase, the degrees of freedom for pos-sible eutrophy control and mitigation actions are reduced with respect to other phases, and the uses to which the dam is orien-ted (urban water provision, irrigation, hydroelectricity generation, etc.) become critical when is time to select eutrophy’s con-trol options. Finally, eutrophication is a change that not only affects reservoirs, but also sections of the rivers downstream thedam, so it is also necessary to keep in mind the control measures for this type of environment.
Keywords: Eutrophication, reservoir, control and management measures.
1
Limnetica 22(1-2): 1-13 (2003)© Asociación Española de Limnología, Madrid. Spain. ISSN: 0213-8409

INTRODUCCIÓN
La eutrofización denominada “cultural” es per-cibida desde hace años como una alteración pre-ocupante de la calidad de las aguas continenta-les (Margalef, 1976; Lee et al., 1980) y deamplia distribución en los ecosistemas acuáti-cos. En algunos lugares, no obstante, la eutrofi-zación no es vivida como una perturbación delos ecosistemas acuáticos sino como una formade cultivo acuático, fuente de alimento, que hayque potenciar (Ryding & Rast, 1993). Por otrolado, la eutrofia como proceso y sus consecuen-cias, bien pudo jugar un papel clave hace unos2.000 millones de años, en la transformación dela atmósfera de la tierra hacia las condicionesoxidantes actuales.
En el caso de los embalses es una alteraciónprácticamente inherente a su construcción y a suexplotación, especialmente en zonas semiáridas,bien soleadas, con un régimen hidrológico mar-cadamente estacional y redes hidrográficas den-samente ocupadas por población o usos agrope-cuarios. La mayor susceptibilidad de losembalses a la eutrofia, con relación a los lagos,se explica por la carga de materia orgánica quelos primeros deben procesar de golpe en su inicioy por la alta relación entre las superficies decuenca y de lámina de agua, que favorece el man-tenimiento de unas mayores aportaciones relati-vas de nutrientes por unidad de superficie.
Este artículo no es ni pretende ser una revi-sión bibliográfica sobre las medidas de gestióny adecuación ambiental de los embalses frente ala eutrofia. Se trata de una recopilación cualita-tiva de propuestas fundamentadas en actuacio-nes, técnicas y procedimientos, que se sabe, enalgunos casos por experiencia propia, puedendar buenos resultados en el control de la eutro-fia de los embalses, tanto desde un punto devista predictivo, como preventivo o corrector.
Son en general propuestas bien conocidasdesde hace años, recopiladas de diferentes fuen-tes, que se consideran absolutamente vigentesen la actualidad e incluso, algunas de ellas,insuficientemente documentadas, contrastadas ysancionadas por la práctica.
Un par de aspectos importantes son lossiguientes. En primer lugar hay tener en cuentaque existe una amplia tipología de embalses ypor tanto de condiciones y condicionantes parala manifestación de la eutrofia. Por esta razón,no todas las propuestas de control de la eutro-fia son aplicables a todos los embalses y, en elcaso de las que presentan un más amplioespectro de aplicación, no necesariamente hande alcanzar la misma eficiencia en cualquierembalse donde se apliquen.
En segundo lugar, si bien la eutrofización esun concepto originariamente acuñado para eco-sistemas leníticos, afecta también y de formamuy significativa a los tramos de río reguladospor embalses eutróficos, de modo que las actua-ciones de prevención y corrección deben contem-plar también esa parte del ecosistema fluvial.
EL CONTROL DE LA EUTROFIA EN EMBALSES
Las actuaciones de control de la eutrofia enembalses deben tratarse como cualquier otroaspecto técnico constructivo relevante y comotal deben empezar a considerarse en la propiaconcepción del proyecto de construcción de lapresa que formará el embalse, o en su defecto,en el estudio de impacto ambiental que le acom-paña. A este nivel, la primera cuestión a plante-arse es la vocación trófica que tendrá el futuroembalse (Fig. 1), de acuerdo con la informacióndisponible sobre el lugar donde se pretendeconstruir y las características básicas del tipo deobra hidráulica que se proyecta.
Esta fase de predicción del grado de eutrofiaque tendrá el embalse, puede tener un margende error apreciable, pero debe permitir en cual-quier caso precisar la necesidad o no de proyec-tar y planificar medidas de atenuación de laeutrofia. En esta fase es donde tiene mayor sen-tido la detección de las causas y/o los orígenesde la posible situación de eutrofia, al objeto deevaluar las opciones de actuación sobre ellos.
El diseño y la consideración a nivel de pro-yecto (o de su correspondiente estudio de
2 A. Palau

impacto ambiental), de las posibles medidas deatenuación de la eutrofia, constituye la fasede prevención, que podrá incluir tanto obras einfraestructuras, como actuaciones de gestiónya programadas, encaminadas a evitar, en lamedida de lo posible, la eutrofización de lamasa de agua embalsada.
Finalmente, si a pesar de todo se llega (oexiste) un grado de eutrofia inaceptable, biensea desde un punto de vista ecológico, socioeco-nómico o de explotación de la obra hidráulica,no quedan más opciones que la adopción demedidas correctoras que, se aplicarán funda-mentalmente sobre los síntomas de eutrofia, enforma de tratamientos y/o gestión del volumende agua embalsada.
A lo largo de estas tres fases (predicción,prevención y corrección) desde la concepciónde un proyecto de embalse hasta la consecu-ción de un nivel aceptable de eutrofización, sevan perdiendo grados de libertad a nivel de
posibilidades de actuación, en el sentido quelas opciones de aplicación de medidas (obras,actuaciones, etc.) se van reduciendo o encare-ciendo, desde la fase de proyecto hasta la deexplotación del embalse; es decir la faltade previsión en las consideraciones inicia-les del proyecto de embalse, redunda en posi-bles limitaciones del uso del agua o en unoscostes de control de la eutrofia crecientes, enla fase de explotación, lo que en definitivapuede interpretarse como una pérdida de laef iciencia o rendimiento del proyecto deembalse y el uso del agua, en la medida que larelación coste/beneficio aumenta.
Predicción de la vocación trófica
La eutrofia no se manifiesta por igual en cual-quier ecosistema acuático, siendo los más afecta-dos los de tipo lenítico (lagos y embalses) y aúndentro de estos, una misma aportación de fósforo
Control de la eutrofización en embalses 3
Figura 1. Esquema general del enfoque que recibe el control de la eutrofia en el presente artículo. General diagram of the eutro-phication control approach in this paper.

puede tener efectos muy distintos según algunascaracterísticas del ecosistema acuático receptor.
Toda predicción supone deducir la respuestade un proceso o sistema concretos, frente a unasdeterminadas condiciones y por tanto en el casode la eutrofización de embalses, hay que partirdel conocimiento del tipo de embalse que se pro-yecta y de las características del medio donde sepretende construir. La tabla 1 muestra la informa-ción básica necesaria para la predicción.
A partir de la localización geográfica dellugar donde se proyecta el embalse y de su ante-proyecto constructivo, no resulta demasiadocomplicado obtener información sobre losaspectos referidos en la tabla 1. De ellos puedededucirse, o al menos estimarse, el origen y lacarga de nutrientes y/o de materia orgánica querecibirá el embalse, la dureza del agua y su con-tenido habitual en oxígeno disuelto o bien eltiempo medio de residencia del agua embalsaday su circulación por el vaso de embalse, asícomo el tipo de ciclo térmico anual del embalsey su estabilidad térmica, relacionándolo todoello con la disponibilidad y el comportamientode nutrientes. También puede obtenerse unaserie de índices referidos a las aptitudes morfo-lógicas del futuro vaso de embalse (Wetzel,1981) de cara a la eutrofización que, aún nosiendo definitivos, complementan el análisis de
la vocación trófica de un embalse. Conocidosestos aspectos, la predicción de la eutrofia, en lamayoría de casos, pasa por la cuantificación delas cargas de fósforo que recibirá el embalse y lamodelización de sus consecuencias mediantealguno de los modelos, de diferente compleji-dad y precisión, que existen para la determina-ción de la vocación trófica del futuro embalse(Ryding & Rast, 1993).
Para el reconocimiento de los posibles efec-tos del estado trófico del embalse en el ríoaguas abajo, interesa conocer la profundidadde la toma (o tomas) de agua desde el embalse,así como las características de la incorporaciónde esas aguas al río. Resulta también esencialsaber el régimen de caudales de mantenimientoy de explotación del embalse, así como lascaracterísticas morfológicas, hidráulicas y lim-nológicas del cauce y las riberas que afectarána su capacidad de procesado de nutrientes y,en definitiva, a la intensidad de la manifesta-ción de la posible eutrofia.
La estimación de las cargas de nutrientes ymateria orgánica que recibe el cauce principaly los tributarios río abajo, permite reconocerposibles efectos sinérgicos o acumulativos, enespecial en embalses que bien sea por capacidadde regulación o de derivación de aguas, reducendrásticamente los caudales circulantes río abajo.
4 A. Palau
Tabla 1. Información básica en la predicción de la vocación trófica de un futuro embalse y de sus consecuencias río abajo. Basic informationfor the prediction of possible trophic states of a future reservoir and its’ consequences downstream.
FUENTES DE INFORMACIÓN ASPECTOS BÁSICOS A CONSIDERAR
Localización geográfica Climatologíadel futuro embalse Hidrología
HidrogeologíaCaracterísticas físico-químicas de las aguasUsos y estado de conservación de la cuencaUsos del futuro vaso de embalseMorfología del cauce y riberas en el tramo reguladoUsos y estado de conservación de la ribera en el tramo regulado
Características del anteproyecto Morfología del futuro vaso de embalsede embalse Usos y manejo del agua embalsada
Profundidad y características de las tomas de agua en la presaCaudales de mantenimiento en el tramo regulado

Prevención de la eutrofia
El principal resultado de la predicción sobre lavocación trófica de un futuro embalse, es elreconocimiento de una parte de los posiblesefectos ambientales del embalse proyectadosobre el ecosistema fluvial y la identificación delas limitaciones que la eutrofia puede suponerpara los usos y objetivos de explotación delembalse. De todo ello, se podrán deducir lasnecesidades (y posibilidades) de actuación en lareducción de las cargas de nutrientes o en la ate-nuación de los síntomas de la posible eutrofiza-ción, que haya que incorporar al proyecto cons-tructivo, con carácter preventivo. La Tabla 2recoge las consideraciones más habituales en la
prevención de la eutrofia, susceptibles de tener-se en cuenta en nuevos embalses y algunas deellas también como medidas de corrección delgrado de eutrofia en embalses ya existentes.
Las actuaciones preventivas, para ser efica-ces, deben quedar recogidas en el propio pro-yecto constructivo o en el preceptivo estudio deimpacto ambiental que le acompaña. Es impor-tante destacar que todas estas actuaciones, adiferencia de las indicadas en la fase de predic-ción, suponen ya unos costes económicosimportantes y unos compromisos (decisiones)bastante irreversibles.
Con relación al diseño de las obras, equipos einfraestructuras (tabla 2) una primera considera-ción debe ser siempre el análisis de la reducción
Control de la eutrofización en embalses 5
Tabla 2. Niveles de actuación y posibles aspectos a considerar en cada uno de ellos, con relación a la prevención de la eutrofización en futurosembalses y la reducción de sus efectos negativos en el río aguas abajo. Levels of intervention and possible issues to be incorporated to eachone of them, in relation to eutrophication prevention the eutrophication of a new reservoir and to reduce its negative consequences downstre-am the dam.
NIVEL DE ACTUACIÓN POSIBLES ASPECTOS A CONSIDERAR
Proyecto técnico o estudio de impacto • Depuración de aguas residuales en la cuenca tributariaambiental. Aspectos referidos a obras, • Restauración hidrológico-forestal de la cuencaequipos e infraestructuras • Desviación (“by-pass”) de tributarios
• Características y ubicación (localización en profundidad) de las tomas de agua
• Dimensionado del vaso de embalse• Construcción de presas de cola
Proyecto técnico o estudio de impacto • Reducción (extracción) de la carga de materia orgánica oxidableambiental. Aspectos referidos al tratamiento • Aprovechamiento de recursos inundables (suelos, vegetación,previo del futuro vaso de embalse madera, áridos)
Proyecto técnico o estudio de impacto • Usos y tiempos de residencia del aguaambiental. Aspectos referidos a los usos • Ordenación (limitación) de usos y actividades en el embalsedel embalse
Proyecto técnico o estudio de impacto • Época del año para el inicio del llenado del embalseambiental. Planificación del primer llenado • Curva (velocidad) de llenado de embalsedel embalse • Plan de extracción selectiva de agua a diferentes profundidades
según la época del año
Proyecto técnico o estudio de impacto • Vaciados periódicos totales o parciales del embalseambiental. Gestión del tramo de río regulado • Régimen de caudales de mantenimiento
• Restauración y/o consolidación de la vegetación de ribera• Capacidad de aireación del cauce• Características y ubicación (localización en profundidad)
de las tomas de agua

en origen del exceso de nutrientes que vaya arecibir el embalse. Así, en función de la impor-tancia socioeconómica del embalse, sería admisi-ble plantearse actuaciones de saneamiento deaguas residuales directas y difusas, así como larestauración hidrológico-forestal a nivel de lacuenca tributaria. Una opción hasta cierto puntoalternativa puede ser la construcción de una presade cola, que además de poder contribuir a laretención (y extracción de nutrientes), a menudopuede convertirse en una infraestructura social yeconómicamente dinamizadora de la zona, si sediseña con capacidad para acoger actividadeslúdicas y recreativas. Por otro lado, no es infre-cuente que estas zonas represadas en las colas deembalses, acaben convirtiéndose en zonas húme-das con cierto valor de conservación. En elmismo sentido, si las condiciones del entorno delembalse y sus dimensiones lo permiten, puedeplantearse la desviación parcial (o temporal) deltributario responsable de la eutrofización, haciazonas húmedas de alrededor o río abajo, si bienhay que tener en cuenta que con ello se modificala tasa de renovación del agua embalsada, laestructura de la comunidad algal y las relacionesagua-sedimentos (Moss et al., 1989).
El dimensionado del vaso de embalse sueleadmitir pocas modificaciones, dado que estáacotado por las características topográficas dela cerrada donde se proyecta la presa y por losobjetivos del embalse, que corren el riesgo deperder sentido si se altera significativamente elvolumen embalsable. En cambio, la ubicación yel diseño de las tomas de agua, desde las super-ficiales hasta los desagües de fondo, si admitecambios que no sólo no tienen porque penalizarel aprovechamiento del embalse, sino que pue-den mejorarlo al aumentar las garantías deexplotación; así, en la utilización de uno u otrotipo de válvula o compuerta en una toma, o tam-bién en su profundidad, puede estar la diferenciaen suministrar río abajo unas aguas con poco onada de oxígeno disuelto durante la estratifica-ción estival o bien prácticamente saturadas al100% durante cualquier época del año.
Sobre el tratamiento previo del futuro vaso deembalse, son bien conocidas las propuestas de
retirada de la vegetación como medida dereducción de la carga orgánica oxidable bajo elagua. La eficacia de esta medida no siempre haquedado demostrada, dado que una fracciónimportante de la materia orgánica oxidable seencuentra en los horizontes superiores del suelo,componente este mucho más difícil de retirar, apesar de ser, ecológica y económicamente, bas-tante más valioso que la cubierta vegetal. Encualquier caso, cualquier aprovechamiento quepueda hacerse de los recursos situados en lazona inundable por el embalse (madera, suelos,áridos, etc.) es, por principio, recomendable.
En cuanto a la prevención aplicada a los usosdel agua y del embalse, dado que como ya se haindicado el dimensionado de su volumen no esfácilmente modificable y la aportación de los tri-butarios tampoco (salvo si son ríos regulados),resulta que las posibilidades de gestión de lostiempos de residencia medios del total de aguaembalsada, aspecto realmente clave en la preven-ción y la contención de la eutrofia (Straskraba,1999), son limitados. Sin embargo, si se disponede un sistema de tomas de agua a distintas pro-fundidades suficientemente versátil (Cassidy,1989), puede llevarse a cabo una actuación eficazen el control de la producción primaria y portanto de la manifestación de la eutrofia, medianteuna gestión selectiva de la tasa de renovación delagua a diferentes profundidades y épocas del año(Toja, 1982; Palau, 1991).
Las propuestas de ordenación de usos y acti-vidades son una opción con signo preventivo,pero de interés también en casos de embalses enexplotación sujetos a aprovechamientos diver-sos, tanto de tipo hidráulico (riegos, producciónhidroeléctrica, abastecimientos) como de tiporecreativo (navegación, pesca, etc.). En algunoscasos, los resultados en materia de control yprevención de la eutrofia pueden ser apreciablesa partir de medidas tan simples y aparentementetan indirectas como la gestión de la pescadeportiva, por ejemplo controlando los efectosde la resuspensión de nutrientes (bioturbidez)por un exceso de ejemplares de gran tamaño dedeterminadas especies de peces de hábitos ben-tónicos detritívoros. En el mismo sentido, la
6 A. Palau

ordenación de otros usos frecuentes en embal-ses, como el baño y la navegación recreativa,además de lógicamente el control de vertidosdirectos o difusos al embalse mediante una pla-nificación de usos y actividades en sus orillas ymárgenes (Wilson & O’Sullivan, 1993) tambiénpueden contribuir significativamente en la pre-vención y mitigación de la eutrofia.
Pero independientemente de los resultadosque se consigan con medidas preventivas deltipo de las indicadas, no hay que olvidar que laeutrofia es, en la práctica, una condición inhe-rente a cualquier nuevo embalse, de modo quesiempre habrá que contar, con un proceso deeutrofia inicial más o menos intenso, asociadoal primer llenado del embalse.
Una forma de incidir en una reducción de laintensidad y la duración de este proceso demaduración de un nuevo embalse es medianteuna planificación del primer llenado del embal-se, que debe tener en consideración al menos losaspectos relacionados al respecto en la tabla 2.Si bien no siempre es posible aplicar una pro-gramación de este tipo, en general es mejor pro-ceder al primer llenado con aguas frías (másdensas y oxigenadas) e ir llenando el embalseprogresivamente en varios años, con unos tiem-pos de residencia del agua lo más bajos posibleal inicio, gestionados mediante vaciados parcia-les (o totales, según el volumen de embalse y laaportación), preferentemente con extracciónselectiva del agua a diferentes profundidadessegún la época del año. Los costes añadidos quepuede suponer la aplicación de esta medida, seven compensados en la mejor calidad del aguaembalsada, lo que reduce la afectación ambien-tal del embalse en el río y no limita ni condicio-na su explotación por tanto tiempo como el quese requiere sin una gestión del primer llenado.
Río abajo, las medidas preventivas frente a laeutrofización de un embalse tienen como objeti-vo común el favorecer la mezcla y la aireacióndel agua, junto con el mantener (o restablecer)el control trófico que la vegetación de riberarealiza sobre el cauce. Así, se trata de implantarun régimen de caudales de mantenimiento quecontemple, no sólo un caudal mínimo o “ecoló-
gico” sino una distribución temporal del caudalacorde con el régimen hidrológico natural,incluyendo crecidas controladas (caudales gene-radores o renovadores) y otras consideraciones(Palau, 1994). Estos caudales de mantenimientodeben permitir una adecuada aireación delcauce y una limitada proporción de ambienteslimnófilos, más propensos a permitir las forma-ciones de macrófitos y con ello a reconcentraren un tramo habitualmente demasiado corto, lamayor parte del reciclado de nutrientes proce-dentes del embalse. Cabe indicar que en algunoscasos sería planteable una situación totalmentecontraria a la descrita; es decir dedicada a favo-recer el estancamiento del agua en las inmedia-ciones del pie de presa para que la producciónprimaria del cauce retenga la mayor cantidadposible de nutrientes, evitando su dispersiónaguas abajo.
Por otro lado, la restauración y/o consolida-ción de la vegetación de ribera, en la medidaque ejerce un control sobre la temperatura delagua y la disponibilidad de luz y nutrientes paralos productores primarios acuáticos, es tambiénde interés si la opción escogida es la del proce-sado dosificado de los nutrientes, río abajo.
Tal y como ya se ha comentado, la profundi-dad de procedencia del agua embalsada y suforma de incorporación al cauce (en salto concaída libre, en conducción a presión), es la clavede la calidad del agua río abajo y por tanto sontambién aspectos a tener en consideración. En elcaso de aprovechamientos hidroeléctricos,donde la conducción forzada (a presión) delagua hasta las turbinas es obligada, hay laopción de instalar turbinas con un diseño quefavorece la aireación del agua y/o equipos deaireación propiamente (ASCE/EPRI, 1989).Con la primera opción, se consiguen resultadosfrancamente significativos en cuanto a recupe-ración del contenido de oxígeno disuelto.
Corrección de la eutrofia
Si la eficacia de las medidas preventivas se fun-damentaba en la precisión de las prediccionessobre la vocación trófica y su reflejo en fases
Control de la eutrofización en embalses 7

tempranas del proyecto constructivo del nuevoembalse, del éxito de la prevención dependerá lanecesidad o no de llevar a cabo medidas correc-toras durante la explotación que, en cualquiercaso pueden evaluarse como indispensablespara embalses ya eutróficos.
La corrección de la eutrofia en embalsespuede abordarse desde dos perspectivas nonecesariamente excluyentes, pero sí de muy dis-tinto alcance y ámbito de aplicación. Tal y comose recoge en la figura 1, una opción es la gestióny la otra el tratamiento.
Gestión
La gestión se refiere a actuaciones de manejodel agua embalsada y/o de las comunidadesnaturales acuáticas. Como muestra la tabla 3,
estas actuaciones son en buena medida, comu-nes a las de tipo preventivo expuestas en la tabla2, pero en este caso no se aplican para que noaparezcan los síntomas de la eutrofia sino paraatenuar su manifestación o para que esta tengalugar dentro de unos límites admisibles.
Como ya se ha indicado en el apartado sobremedidas preventivas, la reducción de los apor-tes de nutrientes en origen, bien sea median-te actuaciones directas (depuración de aguasresiduales) o indirectas (restauración hidroló-gico-forestal de la cuenca y control de aportesdifusos a partir de prácticas agropecuariasinadecuadas), debe ser siempre la opción departida a considerar.
La conservación, o si es el caso la restaura-ción hidrológico-forestal de la cuenca, ademásde evitar problemas de erosión y aterramiento
8 A. Palau
Tabla 3. Principales opciones para la corrección de la eutrofia en embalses y/o la atenuación de sus síntomas. Main options for eutrophycorrection in reservoirs and/or for mitigation of its’ symptoms.
TIPO DE NIVELES DE POSIBLES ASPECTOS A CONSIDERARACTUACIÓN APLICACIÓN O SOBRE LOS QUE ACTUAR
Gestión Cuenca tributaria • Depuración de aguas residuales• Desviación (“by-pass”) de tributarios• Control de la contaminación difusa• Restauración hidrológico-forestal
Gestión Agua embalsada • Curva-guía de explotación del nivel de embalse y control de tiempo de residencia medio global y selectivo
• Programación de vaciados periódicos• Instalación de equipos de aireación hipolimnética o de mezcla
vertical de la columna de agua
Comunidades Acuáticas • Modificación de la estructura tróficadel embalse • Manejo de zonas de cola de embalses como “filtros verdes”
Agua y comunidades • Aireación de las aguas salientes del embalserío abajo • Aumento de la capacidad de aireación del cauce
• Implantación de un régimen de caudales de mantenimiento
Tratamiento Agua embalsada • Floculantes y precipitantes• Oxidantes
Sedimentos del embalse • Secado y aireación mecánica• Extracción• Oxidación
Comunidades acuáticas • Extracción mediante medios mecánicosdel embalse • Alguicidas y desinfectantes
• Aplicación de productos químico-biológicos

de embalses (que a su vez son un agravante de laeutrofización), puede contribuir eficazmente ala retención de nutrientes y a la conservación delos acumulados en el suelo.
El volumen de agua embalsada a lo largo delaño, es un factor determinante en el estado tró-fico de los embalses (Pérez-Martinez et al.,1989) por lo que cabe plantearse la posibilidadde establecer una curva-guía de explotacióndel nivel de embalse, coordinada con unaextracción selectiva del agua a distintas pro-fundidades, según la época del año, de modoque se limite al máximo el desarrollo algal y ladesoxigenación hipolimnética (Vicente et al.,2000). Esta curva-guía podría incluir vaciadosperiódicos del embalse (por ejemplo decena-les), que además de mejorar la aireación delsedimento, permitirían actuaciones mecánicassobre él, si fueran necesarias. El vaciado deembalses es, por otro lado, una forma de super-visar el estado de conservación de la presa ysus órganos de regulación, lo cual es obligato-rio en Francia cada 10 años.
Desde hace años existen equipos y experien-cia práctica en la inyección controlada de aire,oxígeno o agua en el hipolimnion de lagos yembalses (Jaeger, 1989; ASCE/EPRI, 1989).Estas técnicas, si bien permiten contener uno delos principales problemas de la eutrofización (eldéficit de oxígeno), son bastante costosas, com-plejas (Ashley & Hall, 1989) y pueden llegar atener un efecto contrario al buscado en funciónde la escala de tiempo considerada para evaluarlos resultados, por ejemplo si se resuspendensedimentos (y materia orgánica) y se rompe laestratificación térmica de la columna de agua,alimentando la zona fótica con más nutrientes.
Por el contrario, en sistemas poco profundosy bajo determinadas condiciones y objetivos, ladesestratificación (mezcla vertical forzada) dela columna de agua, puede ser una buenaopción para evitar o reducir la manifestacióndel déficit de oxígeno disuelto de fondo, aso-ciable a la eutrofización.
A nivel de comunidades acuáticas, se han rea-lizado experiencias sobre modificación de laestructura trófica para limitar el desarrollo algal,
fundamentalmente mediante la eliminación(Faafeng & Brabrand, 1989; Reinersten et al.,1997) o la introducción de especies de peces fil-tradores (Proulx et al., 1993; Bertolo et al., 2000)o comedores de macrófitos. En todos los casos,los resultados conducen a una significativa corre-lación positiva entre la biomasa de peces filtrado-res y la de fitoplancton, sin duda debida al cam-bio de presión de predación sobre el zooplanctony su mayor eficiencia en el control del fitoplanc-ton. En zonas como la península ibérica, querepresentan un “nicho ecológico” vacío para estetipo de especies por la falta de lagos, la introduc-ción de especies de peces filtradores para el con-trol de la eutrofia, como en algunas ocasiones seha insinuado, podría conducir a resultados espec-tacularmente contrarios a los esperados.
En cualquier caso, la retirada de materiaorgánica (vegetación acuática, peces) está claroque es una forma de reducir la carga denutrientes que moviliza el ecosistema, de modoque debe considerarse como una opción decontrol de la eutrofia. En este sentido, la ges-tión (retirada) de la materia orgánica producidaen zonas de cola de los embalses que funcionencomo humedales (alta productividad) puedeinscribirse como una medida de control de laeutrofia. Paradójicamente, suele ocurrir queeste tipo de ambientes artif iciales, dada ladiversidad de especies que albergan, en espe-cial de aves, son objeto de algún tipo de protec-ción que limita las posibilidades de uso como“filtro verde” para el embalse.
Aún dentro de las medidas de gestión, en elrío aguas abajo, las actuaciones de correcciónde la eutrofia se centran en lo ya indicado parasu prevención; es decir en maximizar la capaci-dad de oxigenación de las aguas, tanto a la sali-da del embalse como a lo largo del río, conser-vando como aspecto clave, un régimen decaudales de mantenimiento adecuado y un buenestado de la vegetación de ribera.
Tratamiento
En cuanto a la aplicación de tratamientoscorrectivos de la eutrofia, la tabla 3 recoge las
Control de la eutrofización en embalses 9

principales opciones existentes. Cabe indicarque se trata de actuaciones en su mayoríainaplicables a grandes embalses o en generalmasas de agua importantes, tanto por su costeeconómico, como por su eficacia y posibilida-des de control. Son medidas por tanto adecua-das para pequeñas masas de agua, de usos muyespecíficos y fácilmente controlables a todonivel, como es el caso de balsas de riego, depó-sitos de abastecimiento de agua, lagunas paraactividades recreativas, etc.
A nivel del agua, existe la opción de aplicartratamientos con productos químicos floculan-tes, precipitantes y oxidantes, que reduzcan lacarga orgánica en la columna de agua a favor dela formación de sedimento más o menos mine-ralizado, que después se podrá retirar o tratar.
Dentro de los tratamientos del sedimento,está su oxidación mediante productos químicos,su secado y aireación mecánica para su minera-lización, todo ello combinado o no con su retira-da (Sebetich & Ferriero, 1997). Algunas expe-riencias de secado y aireación mecánica oretirada de sedimentos, realizadas sobre lagunaseutróficas de cierta extensión, pero poco pro-fundas, se apuntan como soluciones eficaces enla reducción de la carga de sedimentos disponi-bles (Phillips & Jackson, 1989).
En cuanto a tratamientos sobre comunidadesde organismos acuáticos, está la aplicación dealguicidas y productos desinfectantes, conlas limitaciones inherentes al manejo de estetipo de sustancias, en ecosistemas acuáticosabiertos. Está también la ya citada retiradamecánica de organismos acuáticos (vegetación,peces), como una opción de reducción de lacarga de materia orgánica (y nutrientes) proce-sable por el embalse.
También se han realizado algunas experien-cias de corrección de la eutrofia en pequeñasmasas de agua, basadas en aplicar compuestosquímico-biológicos, de acción múltiple, forma-dos por coprecipitantes, floculantes y microor-ganismos nitrif icantes con sus capacidadesenzimáticas aumentadas mediante ingenieríagenética. Se trata probablemente del campopor el que avanzará (o debería avanzar) el tra-
tamiento de la eutrofia en pequeñas masas deagua, dado que se fundamente en la potencia-ción de reacciones y procesos naturales delpropio ecosistema acuático.
CONCLUSIONES
La corrección definitiva del proceso de eutrofi-zación pasa forzosamente por la eliminación, oal menos la atenuación de la fuente causante dela alteración: el aporte excesivo de nutrientes.El problema es que esta opción no es siempreposible y en cualquier caso suele ser de difícilejecución, no sólo por el carácter difuso quepueden tener parte de las aportaciones, sino por-que la retirada de nutrientes requiere tratamien-tos poco extendidos en los planes de saneamien-to de cuencas. De esta forma, el control de laeutrofia tiende a centrarse habitualmente en elcontrol de sus efectos o síntomas, más que en laerradicación de sus causas.
En el caso de embalses además, se trata deecosistemas “condenados” a padecer un ciertogrado de eutrofia al menos en su primer llenado,como consecuencia de la mineralización bajo elagua, de la materia orgánica presente en suvaso. Sólo por esta razón, y al margen del inte-rés social y económico que tiene, resulta obliga-da la consideración de la eutrofia como unaspecto relevante más del proyecto técnico decualquier nuevo embalse.
Para embalses nuevos que se juzguen inapela-blemente necesarios, a partir del reconocimien-to del lugar donde se proyectan y de las caracte-rísticas técnicas de su presa y su cubeta,recogidas a nivel de anteproyecto, es posiblemediante modelos sencillos, establecer la voca-ción trófica de la futura masa de agua artificial(Fig. 2). En función de los resultados de estapredicción, se debería tratar la eutrofizaciónpotencial como un condicionante económica-mente relevante para embalses con determina-dos usos y objetivos, recogiendo si es el caso,dentro del propio proyecto técnico definitivoy/o en el preceptivo estudio de efectos ambien-tales que lo acompaña, aquellas medidas pre-
10 A. Palau

ventivas de la eutrofia que se consideren másadecuadas y que como mínimo podrán centrarseen el tratamiento previo del futuro vaso inunda-ble del embalse, la programación del primer lle-nado y de los usos futuros del embalse, el dise-ño de elementos estructurales y las actuacionesy medidas de gestión necesarias para el tramode río que quedará regulado.
Si a pesar de todo la eutrofia se manifiestapor encima de lo deseable, o se está delante deun embalse eutrófico ya en explotación, puedenaplicarse medidas correctoras a distintos niveles(aguas, sedimentos, organismos), recogidas enforma de actuaciones de gestión, bastante coin-
cidentes con las posibles actuaciones preventi-vas, o bien como tratamientos, en ese caso fun-damentalmente adecuados para pequeños volú-menes de agua.
La disponibilidad de tomas de agua a diferen-tes niveles de embalse y la gestión selectiva delos tiempos de residencia del agua, se presentancomo medidas eficaces en el control de la eutro-fia, con una capacidad a la vez preventiva ycorrectora.
Desde la fase de proyecto a la de explota-ción de un embalse, las opciones de control dela eutrofia recogidas de forma genérica en lafigura 2, tienden a ser cada vez menos eficien-
Control de la eutrofización en embalses 11
Figura 2. Principales medidas de control y gestión de la eutrofia a partir de la predicción del estado trófico de un anteproyecto deembalse, en la prevención a nivel de proyecto y en la corrección en un embalse en explotación (EsIA: Estudio de ImpactoAmbiental). Main measures for eutrophication control and management, from the trophic state prediction in the reservoir’s preli-minary draft phase, the incorporation of preventive measures on the definitive draft phase, and the corrective actions when thereservoir is operative.

tes, bien sea por su mayor coste o la crecientedisminución de la capacidad de autorecupera-ción del embalse potencialmente eutróf ico.Puede ocurrir perfectamente que no se puedancontrolar del todo los efectos de la eutrofia, apesar de las medidas tomadas y de las inver-siones realizadas, dado que otros factores nocontrolables (meteorología, aportación hidro-lógica, etc.), poco previsibles (situacionesaccidentales, etc.) o poco controlables (verti-dos ilegales, etc.), pueden modificar de formasignificativa el sentido de los resultados, almenos transitoriamente.
Por último, la eutrofización de embalses nohay que interpretarla como una perturbaciónlocalizada sólo aguas arriba de las presas, sinotambién con consecuencias importantes ríoabajo, donde por las propias características delecosistema acuático lótico, el control de laeutrofia admite menos opciones, pero las dispo-nibles son más simples y eficaces, todas ellascentradas básicamente en fomentar la aireacióny evitar o promover (según los objetivos) zonasde acumulación (alto tiempo de residencia) denutrientes y materia orgánica.
BIBLIOGRAFÍA
ASCE/EPRI. 1989. Civil Engineering Guidelines forPlanning and Designing HydroelectricDevelopments. American Society of CivilEngineers (ASCE) y Electric Power ResearchInstitute (EPRI). ASCE/EPRI Guides 1989, Vol.1. N.Y. USA.
ASHLEY, K. I. & K. J. HALL. 1989. Factors influen-cing oxygen transfer in hypolimnetic aerationsystems. Verh. Internat. Verein. Limnol., 24:179-183.
BERTOLO, A., F. LESCHER-MOUTOUÉ & G.LACROIX. 2000. Interaction effects of depthand planktivorous fish on plankton biomass.Verh. Internat. Verein. Limnol., 27: 1747-1751.
CASSIDY, R. A. 1989. Water temperature, dissolvedoxygen and turbidity control in reservoir relea-ses. In: Alternatives in regulated river manage-ment. J.A. Gore & G.E. Petts (eds.): 28-62.CRC Press. Boca Raton, Florida (USA).
FAAFENG, B. A. & A. BRABRAND. 1989.Biomanipulation of a small, urban lake-removalfish exclude bluegreen blooms. Verh. Internat.Verein. Limnol., 24: 597-602.
JAEGER, D. 1989. TIBEAN- a new hypolimneticaeration plant. Verh. Internat. Verein. Limnol.,24: 184-187.
LEE, G. F., W. RAST, R. A. JONES & J. L. ORTIZ.1980. Progresos recientes en la estimación dela respuesta de los lagos y embalses a las apor-taciones de nutrientes. Centro de Estudios yExperimentación del Centro de EstudiosHidrográficos (MOPU). Publicación nº 137.Madrid
MARGALEF, R. 1976. Biología de los embalses.Investigación y Ciencia, 1: 50-62.
MOSS, B., J. STANSFIELD & K. IRVINE. 1989.Problems in the restoration of a hypereutrophiclake by diversion of a nutrient-rich inflow. Verh.Internat. Verein. Limnol., 24: 568-572.
PALAU, A. 1991. Evolution of annual chlorophyllconcentration in a high mountain pump-storagereservoir. Verh. Internat. Verein. Limnol., 24:1401-1404.
PALAU, A. 1994. Los mal llamados caudales “ecoló-gicos”. Bases para una propuesta de cálculo.OP (Obras Públicas), 28 (2): 84-95.
PÉREZ-MARTÍNEZ, C., R. MORALES-BAQUERO& P. SÁNCHEZ-CASTILLO. 1989. The effectof the volume decreasing on the trophic statusin four reservoirs from Southern Spain. Verh.Internat. Verein. Limnol., 24: 1382-1385.
PHILLIPS, G. & R. JACKSON. 1989. The control ofeutrophication in very shallow lakes, theNorfolk Broads. Verh. Internat. Verein. Limnol.,24: 573-575.
PROULX, M., F. R. PICK, A. MAZUMDER & D. R.S. LEAN. 1993. Effects of plancktivorous fishon phytoplancton biomass and communitystructure. Verh. Internat. Verein. Limnol., 25:331-334.
RAVERA, O. 1989. The effects of hypolimneticoxygenation in the shallow and eutrophicLake Comabbio (Northern Italy) studied byenclosure. Verh. Internat. Verein. Limnol., 24:188-194.
REINERSTEN, H., J. I. KOKSVIK & A. HAUG.1997. Effects of fish elimination on the phyto-plankton and zooplankton in a small eutro-phic lake. Verh. Internat. Verein. Limnol., 26:593-598.
12 A. Palau

RYDING, S. O. & W. RAST. 1993. Le contrôle del’eutrophisation des lacs et des reservoirs. Ed.Masson. UNESCO. París.
SEBETICH, M. J. & N. FERRIERO. 1997. Lake res-toration by sediment dredging. Verh. Internat.Verein. Limnol., 26: 776-781.
STRASKRABA, M. 1999. Retention Time as a KeyVariable of Reservoir Limnology. In:Theoretical Reservoir Ecology and itsApplications. J.G. Tundisi & M. Straskraba(eds.): 385-410. International Institute ofEcology. Brazilian Academy of Sciences andBackhuys Publishers.
TOJA, J. 1982. Control de la eutrofia en embalsespor utilización selectiva de agua a distintas pro-
fundidades. Obras Públicas (Abril-Mayo): 223-231.
VICENTE, E., A. CAMACHO, M. D. SENDRA, D.SANCHIS, J. M. SORIA, M. J. DASÍ & M. R.MIRACLE. 2000. Limnological management ofthe Amadorio Reservoir (Spain) during anextremely dry summer. Verh. Internat. Verein.Limnol., 27: 2298-2302.
WETZEL, R. G. 1981. Limnología. Ed. Omega.Barcelona.
WILSON, H. M. & P. E. O’SULLIVAN. 1993. Thecontrol of eutrophication of small shallowlakes in the United Kingdom: The legal fra-mework. Verh. Internat. Verein. Limnol., 25:461-464.
Control de la eutrofización en embalses 13


15
Fitoplancton de cinco embalses de Venezuela con diferentes estadostróficos
Ernesto J. González, Mario Ortaz, Carlos Peñaherrera, Enrique Montes, María L. Matos y Janin Mendoza
Universidad Central de Venezuela, Instituto de Biología Experimental, Apartado 47106 – Los Chaguaramos, Caracas, 1041 – Venezuela. Fax: 58-212-7535897. E-mail: [email protected]
RESUMEN
Se estudiaron las variaciones de abundancia, biomasa y producción primaria del fitoplancton en cinco embalses de la zonanorte de Venezuela empleados para el suministro de agua potable, y que poseen diferentes estados tróficos: Pao-Cachinche,Quebrada Seca, La Mariposa, La Pereza y Lagartijo. Los muestreos se realizaron en diferentes momentos entre septiembrede 1997 y agosto de 1999. El fitoplancton se recolectó con una botella de captación del tipo van Dorn (3 l), y se fijó con solu-ción de lugol hasta la posterior cuantificación de abundancia por sedimentación en cámaras de Utermöhl y conteo con laayuda de un microscopio invertido. La biomasa se estimó mediante la extracción de clorofila-a con etanol y la producciónprimaria se estimó mediante la evolución de la concentración del oxígeno disuelto en botellas claras y oscuras e incubación“in situ” por dos horas. En Pao-Cahinche (hipereutrófico) se identificaron 59 especies del fitoplancton, Cyanobacteria fue elgrupo dominante, la abundancia varió entre 25,9 x 109 y 410,2 x 109 org. m-2, la biomasa entre 32,4 y 186,6 mg m-2, mientrasque la producción primaria bruta (PPB) promedio fue de 3229 y 2482 mg C m-2 d-1 en las estaciones E1 y E2, respectiva-mente. En La Mariposa (hipereutrófico) se identificaron 50 especies, las Cryptophyta dominaron, la abundancia varió entre6,2 x 109 y 35,5 x 109 org. m-2, el máximo valor de biomasa fue de 161,8 mg m-2, y la PPB varió entre 968 y 2633 mg C m-2
d-1. En Lagartijo (oligotrófico) se identificaron 67 especies, siendo las algas verdes las más diversas y las dominantes; laabundancia fue relativamente baja en este ecosistema, con valores entre 4,7 x 109 y 27,2 x 109 org. m-2; la biomasa varióentre 0,01 y 50,0 mg m-2, mientras que los valores de PPB variaron entre 376 y 2088 mg C m-2 d-1. En La Pereza (eutrófico)se identificaron 48 especies, Cryptophyta dominó, la abundancia varió entre 26,7 x 109 y 102,9 x 109 org. m-2, la biomasavarió entre 5,5 mg m-2 y 199,2 mg m-2 y la PPB entre 265 y 5619 mg C m-2 d-1. En Quebrada Seca (hipereutrófico) se identi-ficaron 48 especies, y Cyanobacteria dominó el fitoplancton; la abundancia varió entre 26,9 x 109 y 69,3 x 109 org. m-2, labiomasa entre 53,7 y 355,4 mg m-2 y la PPB entre 1763 y 3494 mg C m-2 d-1. La mayoría de los embalses evaluados emplea-dos para el suministro de agua potable se encuentran eutrofizados, y en ellos predominan las Cyanobacteria y lasCryptophyta, con elevados valores de abundancia, biomasa y producción primaria, como consecuencia de la entrada excesivade nutrientes desde sus cuencas de drenaje altamente erosionadas e intervenidas y a través de las aguas servidas sin trata-miento previo.
Palabras claves: Eutrofización, fitoplancton, abundancia, biomasa, producción primaria, Venezuela.
ABSTRACT
Seasonal variations of phytoplankton abundance, biomass, and primary production were studied in five drinking water reser-voirs located in the northern region of Venezuela, and that show different trophic states: Pao-Cachinche, Quebrada Seca, LaMariposa, La Pereza y Lagartijo. Samples were taken at different moments from September 1997 to August 1999.Phytoplankton was collected with a van Dorn type bottle (3 l) and fixed with lugol solution until later abundance quantifica-tion by sedimentation with Utermöhl chambers, and counting with inverted microscope. Biomass was estimated by chlo-rophyll-a extraction with ethanol, and primary production was estimated by dissolved oxygen evolution in light and dark bot-tles and “in situ” incubation for two hours. A total of 59 phytoplankton species were identified in Pao-Cachinche(hypertrophic); Cyanobacteria was the dominant group, abundance ranged between 25.9 x 109 and 410.2 x 109 cells m-2,biomass ranged between 32.4 and 186.6 mg m-2, while mean gross primary production (GPP) were 3229 and 2482 mg C m-2
d-1 in stations E1 and E2, respectively. In La Mariposa (hypertrophic), a total of 50 species were identified; Cryptophytadominated, abundance ranged between 6.2 x 109 and 35.5 x 109 cells m-2, the highest biomass value was 161.8 mg m-2, andGPP ranged between 968 and 2633 mg C m-2 d-1. A total of 67 species were identified in Lagartijo (oligotrophic), beinggreen algae the dominant and more diverse group; abundance was relatively low in this ecosystem, and ranged between
Limnetica 22(1-2): 15-35 (2003)© Asociación Española de Limnología, Madrid. Spain. ISSN: 0213-8409

INTRODUCCIÓN
Los problemas asociados con la eutrofizaciónson generalmente de experiencia reciente en lospaíses tropicales (Harper, 1992). La construc-ción de embalses nuevos para el suministro deagua potable, aunado al aumento de asentamien-tos humanos en sus cuencas de drenaje, hanresultado en un amplio espectro de problemas,con el consecuente incremento acelerado en laentrada de nutrientes.
Las masas de agua eutróficas se caracterizanpor tener una productividad elevada y altosvalores de biomasa en todos los niveles tróficos(Ryding & Rast, 1992). En estos cuerpos deagua proliferan las cianobacterias, y el hipolim-nion presenta condiciones de anoxia durante losperíodos de estratificación térmica, y a menudoposeen menos especies de plantas y animalesque los sistemas oligotróficos.
En el caso de los cuerpos de agua empleadospara el suministro de agua potable, los altosniveles de nutrientes producen el crecimientoexcesivo de algas, las cuales pueden conferirolores desagradables al agua (Edmondson,1969), además de bloquear los filtros en lasestaciones de tratamiento y bombeo, lo cualencarece y retarda el tratamiento del agua(Ryding & Rast, 1992).
En Venezuela existen numerosos embalses enlos que se desconocen completamente suscaracterísticas limnológicas (González, 2000),por lo que se hace necesario conocer el funcio-namiento de estos ecosistemas, para así lograr
un aprovechamiento racional de ellos (Infante etal., 1992, Infante et al., 1995).
Los embalses seleccionados en este estudio(Pao-Cachinche, La Mariposa, Lagartijo, LaPereza y Quebrada Seca), se emplean para elsuministro de agua potable para densas regionesurbanas de la región centro – norte de Venezuela.En la mayoría de ellos se presentan grandes den-sidades de algas, especialmente de cianobacte-rias, que causan los problemas antes descritos.
En vista de lo anterior, y dado el interés de lascompañías hidrológicas Hidroven, Hidrocentroe Hidrocapital en conocer las característicaslimnológicas de los cinco embalses citados, seplanteó el objetivo de estudiar las variaciones deabundancia, biomasa y producción primariade esta comunidad en cada uno de los embalses.
MATERIALES Y MÉTODOS
En cada embalse se estimó la producción pri-maria del fitoplancton mediante el método de laevolución de oxígeno en botellas claras y oscu-ras, e incubación “in situ” por dos horas(Wetzel & Likens, 1991). Las muestras setomaron con una botella de captación del tipovan Dorn (3 l) hasta tres veces la profundidaddel disco de Secchi.
Las muestras de fitoplancton se recolectaroncon la botella de captación del tipo van Dorn(3 l) a las mismas profundidades a las cuales seestimó la producción primaria. Para la determi-nación de abundancia, las muestras se fijaron
16 González et al.
4.7 x 109 and 27.2 x 109 cells m-2; biomass ranged between 0,01 and 50,0 mg m-2, while GPP varied from 376 to 2088 mg Cm-2 d-1. In La Pereza (eutrophic), a total of 48 species were identified; Cryptophyta was the dominant group, abundancevaried between 26.7 x 109 and 102.9 x 109 cells m-2, biomass ranged between 5.5 mg m-2 and 199.2 mg m-2 and GPP rangedbetween 265 and 5619 mg C m-2 d-1. A total of 48 phytoplankton species were identified in Quebrada Seca (hypertrophic);Cyanobacteria were the dominant group, abundance ranged between 26.9 x 109 and 69,3 x 109 cells m-2, biomass rangedbetween 53.7 and 355.4 mg m-2, and GPP ranged between 1763 and 3494 mg C m-2 d-1. Most of these reservoirs that areemployed for drinking water supply are eutrophic and Cyanobacteria and Cryptophyta were the dominant phytoplanktongroups with elevated abundance values, biomass, and primary production as a result of the high nutrient input from theirhighly erosionated basins and from waste water without previous treatment.
Keywords: Eutrophication, phytoplankton, abundance, biomass, primary production, Venezuela

con solución de lugol en envases de 100 ml,mientras que para la determinación de biomasa,las muestras se almacenaron en envases de 2 l yse mantuvieron refrigeradas y en oscuridad hastael momento de su análisis en el laboratorio.
Una vez en el laboratorio, la biomasa se esti-mó como concentración de clorofila-a por elmétodo de extracción de pigmentos con etanol(Nusch & Palme, 1975).
La abundancia del fitoplancton se estimó porsedimentación en cámaras de Utermöhl y con-teo en un microscopio invertido de luz (Wetzel& Likens, 1991). Su identificación se realizócon la ayuda de claves taxonómicas.
ÁREAS DE ESTUDIO
Los estudios se realizaron en los embalses Pao-Cachinche, La Mariposa, Lagartijo, La Pereza yQuebrada Seca, cuyas localizaciones enVenezuela y sus principales características mor-fométricas se resumen en la Tabla 1.
Embalse Pao-Cachinche
Está ubicado en la región centro norte deVenezuela, en el límite entre los estadosCarabobo y Cojedes, cerca de la ciudad deValencia, aguas debajo de la confluencia de losríos Paito y Chirgua, aproximadamente hacia el
centro de la cuenca del río Pao. En él confluyenlos principales cursos que drenan de la cuencaalta: ríos Chirgua, Paito, Pirapira, Paya y SanPedro. Se encuentra a una cota de 353 m.s.n.m.(Ginez & Olivo, 1984). Sus aguas se utilizanpara agricultura de subsistencia y uso pecuariointensivo y extensivo, además de suministraragua potable al área metropolitana de Valencia yotras poblaciones de los estados Carabobo,Cojedes y Aragua. El río Paito transporta aguasresiduales y domésticas, mientras que el ríoChirgua transporta las aguas provenientes degranjas avícolas y porcinas. El embalse estárodeado por más de 50 cochineras y polleras, yrecibe una alta carga de nutrientes (especial-mente P), principalmente del río Paito.
Se realizaron muestreos mensuales desdeseptiembre de 1997 hasta febrero de 1999. Seseleccionaron 2 estaciones, ubicadas en el brazooeste y en el brazo este del embalse, los cualesse encuentran separados por un canal estrecho.En el brazo oeste se encuentran el dique, el ali-viadero y la torre-toma, y se ubicó la estacióndenominada E1. La estación ubicada en el brazoeste se denominó E2.
Embalse La Mariposa
El embalse está ubicado en el Distrito Capital,en la ciudad de Caracas. La mayor parte delagua que recibe el embalse proviene del río El
Fitoplancton de cinco embalses venezolanos 17
Tabla 1. Características morfométricas de los embalses estudiados. Morphometric features of the studied reservoirs.
Pao-Cachinche La Mariposa Lagartijo La Pereza QuebradaSeca
Ubicación geográfica 9º 53’ N 10º 24’ N 10º 11’ N 10º 27’ N 10º 13’ N68º 08’ W 66º 33’ W 66º 43’ W 66º 46’ W 66º 43’ W
Area del 16 100 000 540 000 4 550 000 562 500 950 000embalse (m2)Volumen del 170 000 000 7 000 000 80 000 000 8 000 000 7 250 000embalse (m3)Profundidad media (m) 10.6 13.0 17.5 14.0 7.6Tiempo de 281 40 243 12 17renovación (días)

Valle al cual le vierten una considerable canti-dad de agua bombeada desde el embalseLagartijo mediante el Sistema Tuy. El embalsetambién recibe aguas de una pequeña quebradaconocida como Quebrada Los Indios ubicada enla zona oriental del mismo. El embalse estásituado a 980 m.s.n.m. y su cubeta es alargadacon dos brazos principales al sur y al estecorrespondientes a las desembocaduras del ríoEl Valle y la Quebrada Los Indios, respectiva-mente. El embalse se encuentra intervenidoantrópicamente y su cuenca está altamente ero-sionada (González & Ortaz, 1998).
Se realizaron muestreos mensuales de sep-tiembre de 1998 a agosto de 1999. Las muestrasse tomaron en la estación ubicada frente aldique y frente a la torre-toma.
Embalse Lagartijo
Se encuentra ubicado en el estado Miranda. Suprincipal tributario es el río Lagartijo, queaporta aproximadamente 3000 l/s (promedioanual) de agua relativamente limpia ya queeste río nace dentro del Parque NacionalGuatopo, de modo que buena parte de su reco-rrido ocurre dentro de una zona protegida. Elembalse además recibe de manera artif icialagua bombeada desde el río Tuy, la cual antesde entrar al embalse es sometida a un trata-miento de sedimentación y cloración. SegúnInfante et al. (1992) el agua bombeada desdeel río Tuy entra por un lugar que se encuentracerca del punto de toma de agua que se envíapara Caracas y posiblemente por esta razón, suefecto se deja sentir localmente, sin afectarsignificativamente las condiciones del agua enel resto del embalse.
El embalse Lagartijo es de baja altitud (188m.s.n.m.) y muy dendrítico con pendientes pro-nunciadas en las montañas adyacentes de modoque la zona litoral, al igual que en el embalse LaMariposa, está poco desarrollada.
Los muestreos se realizaron de septiembre de1998 a julio de 1999 en una estación cercana auno de los diques del embalse, lejana de la zonade influencia del punto afectado por el río Tuy.
Embalse La Pereza
Este embalse se encuentra ubicado en el estadoMiranda a una cota de 1062 m.s.n.m. (Castillo etal., 1973). Su principal función es el suministrode agua potable a las poblaciones aledañas al áreametropolitana de Caracas. Este embalse es el máspequeño de los cinco estudiados, y recibe agua dela quebrada La Pereza y, principalmente, de laproveniente de otros embalses del sistema Tuy.
Sus alrededores están muy intervenidos, conuna alta erosión en las laderas contiguas. Asímismo, se pudo observar la descarga al embalsede aguas de desecho de origen doméstico eindustrial, y fue evidente la presencia de cochi-neras en áreas aledañas así como de ganadovacuno. Todos los productos de desecho de lasfuentes citadas son vertidas al embalses o arras-tradas a él por escorrentía.
Los muestreos se realizaron en noviembre de1998, febrero, abril, mayo y junio de 1999. Lasmuestras se tomaron en una localidad ubicadafrente al dique.
Embalse Quebrada Seca
Este embalse se encuentra ubicado cerca delembalse Lagartijo en el estado Miranda, a unacota de 167 m.s.n.m. (Castillo et al., 1973). Estárodeado de construcciones no planificadas ydepósitos de basura en sus alrededores, lo que lohace muy vulnerable a cualquier tipo de activi-dad contaminante.
Los muestreos se realizaron durante losmeses de octubre y diciembre de 1998, y enmarzo, mayo y junio de 1999. Las muestras setomaron en una localidad frente al dique.
Los embalses La Mariposa, Lagartijo, La Perezay Quebrada Seca se encuentran interconectadosmediante el sistema Tuy, un sistema de bombeo deagua entre los embalses de la cuenca del río Tuy.
RESULTADOS Y DISCUSIÓN
En la Tabla 2 se presentan algunas característi-cas físicas y químicas de los embalses estudia-
18 González et al.
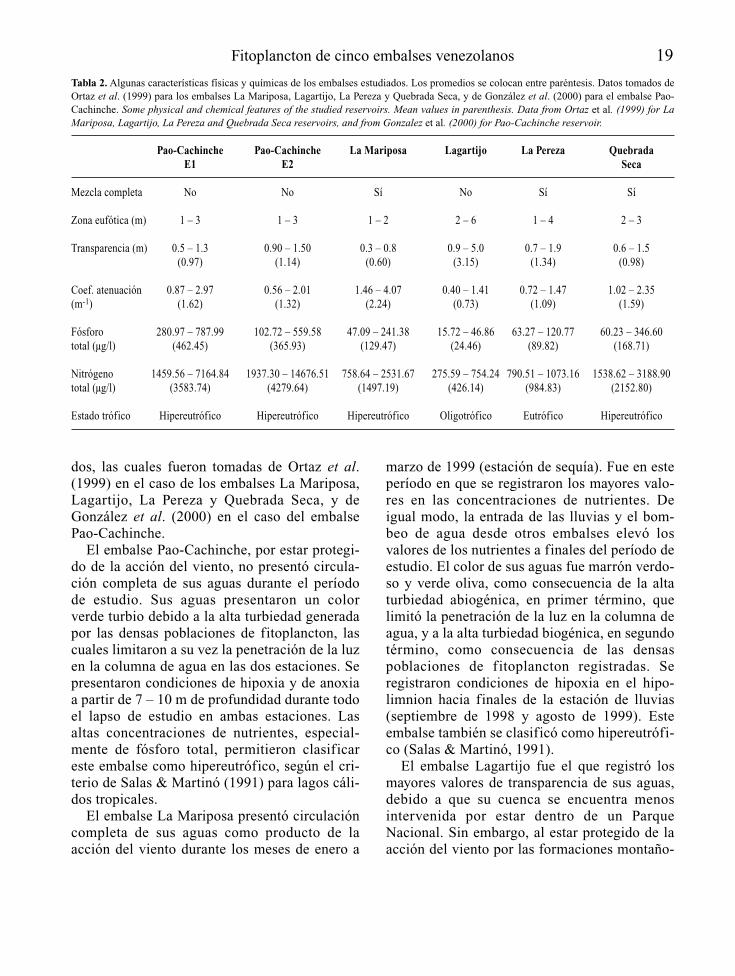
dos, las cuales fueron tomadas de Ortaz et al.(1999) en el caso de los embalses La Mariposa,Lagartijo, La Pereza y Quebrada Seca, y deGonzález et al. (2000) en el caso del embalsePao-Cachinche.
El embalse Pao-Cachinche, por estar protegi-do de la acción del viento, no presentó circula-ción completa de sus aguas durante el períodode estudio. Sus aguas presentaron un colorverde turbio debido a la alta turbiedad generadapor las densas poblaciones de fitoplancton, lascuales limitaron a su vez la penetración de la luzen la columna de agua en las dos estaciones. Sepresentaron condiciones de hipoxia y de anoxiaa partir de 7 – 10 m de profundidad durante todoel lapso de estudio en ambas estaciones. Lasaltas concentraciones de nutrientes, especial-mente de fósforo total, permitieron clasificareste embalse como hipereutrófico, según el cri-terio de Salas & Martinó (1991) para lagos cáli-dos tropicales.
El embalse La Mariposa presentó circulacióncompleta de sus aguas como producto de laacción del viento durante los meses de enero a
marzo de 1999 (estación de sequía). Fue en esteperíodo en que se registraron los mayores valo-res en las concentraciones de nutrientes. Deigual modo, la entrada de las lluvias y el bom-beo de agua desde otros embalses elevó losvalores de los nutrientes a finales del período deestudio. El color de sus aguas fue marrón verdo-so y verde oliva, como consecuencia de la altaturbiedad abiogénica, en primer término, quelimitó la penetración de la luz en la columna deagua, y a la alta turbiedad biogénica, en segundotérmino, como consecuencia de las densaspoblaciones de fitoplancton registradas. Seregistraron condiciones de hipoxia en el hipo-limnion hacia finales de la estación de lluvias(septiembre de 1998 y agosto de 1999). Esteembalse también se clasificó como hipereutrófi-co (Salas & Martinó, 1991).
El embalse Lagartijo fue el que registró losmayores valores de transparencia de sus aguas,debido a que su cuenca se encuentra menosintervenida por estar dentro de un ParqueNacional. Sin embargo, al estar protegido de laacción del viento por las formaciones montaño-
Fitoplancton de cinco embalses venezolanos 19
Tabla 2. Algunas características físicas y químicas de los embalses estudiados. Los promedios se colocan entre paréntesis. Datos tomados deOrtaz et al. (1999) para los embalses La Mariposa, Lagartijo, La Pereza y Quebrada Seca, y de González et al. (2000) para el embalse Pao-Cachinche. Some physical and chemical features of the studied reservoirs. Mean values in parenthesis. Data from Ortaz et al. (1999) for LaMariposa, Lagartijo, La Pereza and Quebrada Seca reservoirs, and from Gonzalez et al. (2000) for Pao-Cachinche reservoir.
Pao-Cachinche Pao-Cachinche La Mariposa Lagartijo La Pereza Quebrada E1 E2 Seca
Mezcla completa No No Sí No Sí Sí
Zona eufótica (m) 1 – 3 1 – 3 1 – 2 2 – 6 1 – 4 2 – 3
Transparencia (m) 0.5 – 1.3 0.90 – 1.50 0.3 – 0.8 0.9 – 5.0 0.7 – 1.9 0.6 – 1.5(0.97) (1.14) (0.60) (3.15) (1.34) (0.98)
Coef. atenuación 0.87 – 2.97 0.56 – 2.01 1.46 – 4.07 0.40 – 1.41 0.72 – 1.47 1.02 – 2.35(m-1) (1.62) (1.32) (2.24) (0.73) (1.09) (1.59)
Fósforo 280.97 – 787.99 102.72 – 559.58 47.09 – 241.38 15.72 – 46.86 63.27 – 120.77 60.23 – 346.60total (µg/l) (462.45) (365.93) (129.47) (24.46) (89.82) (168.71)
Nitrógeno 1459.56 – 7164.84 1937.30 – 14676.51 758.64 – 2531.67 275.59 – 754.24 790.51 – 1073.16 1538.62 – 3188.90total (µg/l) (3583.74) (4279.64) (1497.19) (426.14) (984.83) (2152.80)
Estado trófico Hipereutrófico Hipereutrófico Hipereutrófico Oligotrófico Eutrófico Hipereutrófico

sas aledañas, no presentó circulación completade sus aguas durante el período de estudio,lo cual produjo, a su vez, el desarrollo de con-diciones de hipoxia y de anoxia a partir de los6 – 7 m de profundidad. Se registraron bajasconcentraciones de nutrientes, y se clasificócomo oligotrófico (Salas & Martinó, 1991).
El embalse La Pereza presentó grandes varia-ciones en su nivel de agua durante el período deestudio, lo cual es común en este sistema debidoal manejo al que es sometido mediante laextracción y bombeo de agua desde y hacia elsistema Tuy, y a su tamaño pequeño. Esto fue
registrado por González et al. (1997) en un estu-dio previo, cuando el embalse disminuyó sunivel en diez metros y luego volvió a incremen-tarse otros diez metros en meses sucesivos(octubre a noviembre de 1997). Este hecho, másque las funciones de fuerza, dirigen la dinámicade este ecosistema, permitiendo así la mezclacompleta de sus aguas. Se presentó, al igual queen La Mariposa, una alta turbiedad abiogénica,en primer término, y una alta turbiedad biogéni-ca, en segundo término. Las altas concentracio-nes de fósforo total permitieron clasificarlocomo eutrófico (Salas & Martinó, 1991).
20 González et al.
Tabla 3. Lista de especies del fitoplancton encontradas en el embalse Pao-Cachinche. List of phytoplankton species collected in Pao-Cachinche reservoir.
Cyanobacteria1. Anabaena aphanizomenoides2. Anabaena spiroides3. Aphanocapsa sp.4. Aphanothece nidulans5. Chroococcus limneticus6. Chroococcus pallidus7. Coelosphaerium kuetzingianum8. Coelosphaerium minutissimun9. Coelosphaerium negelianum10. Cylindrospermopsis raciborskii11. Dactylococcopsis acicularis12. Gomphosphaeria aponina13. Lyngbya limnetica14. Limnothrix sp.15. Merismopedia elegans16. Merismopedia tenuissima17. Microcystis aeruginosa18. Microcystis flos-aquae19. Microcystis holsatica20. Oscillatoria limosa21. Pseudoanabaena mucicola22. Rhaphidiopsis curvata23. Spirulina sp.24. Synechococcus bigranulatus25. Synechocystis aquatilis
Chlorophyta26. Botryococcus braunii27. Chlorella vulgaris28. Closterium macilentum29. Closterium sp.30. Cosmarium sp.31. Crucigenia apiculata32. Crucigenia rectangularis
33. Dictyosphaerium pulchellum34. Elakatothrix gelatinosa35. Kirchneriella lunaris36. Monoraphidium braunii37. Monoraphidium tortile38. Nephrocytium limneticum39. Oocystis lacustris40. Phacotus lenticularis41. Pleurotaenium sp.42. Schroederia setigera43. Sphaeocystis schroeteri44. Staurastrum leptocladum45. Volvox sp.
Bacillariophyta46. Aulacoseira granulata47. Cyclotella sp.48. Denticula sp.49. Gomphonema sp.50. Navicula sp.51. Rhizosolenia sp.52. Ropalodia sp.53. Synedra ulna
Euglenophyta54. Euglena variabilis55. Trachelomonas superba56. Trachelomonas volvocina
Pyrrophyta57. Peridinium sp.
Cryptophyta58. Chroomonas sp.59. Cryptomonas erosa
Fitoplancton de cinco embalses venezolanos 21
El embalse Quebrada Seca presentó una zonaeufótica restringida a unos 2 – 3 m, producto dela alta turbiedad biogénica (densas poblacionesde fitoplancton). Este embalse presentó circula-ción completa de sus aguas entre enero y marzode 1999, lo cual seguramente también fue afec-tado por la influencia del bombeo de agua desdey hacia el sistema Tuy. Se presentaron condicio-nes de anoxia a partir de 5 – 7 m. También seregistraron altas concentraciones de fósforototal, lo cual permitió clasificarlo como hipe-reutrófico (Salas & Martinó, 1991).
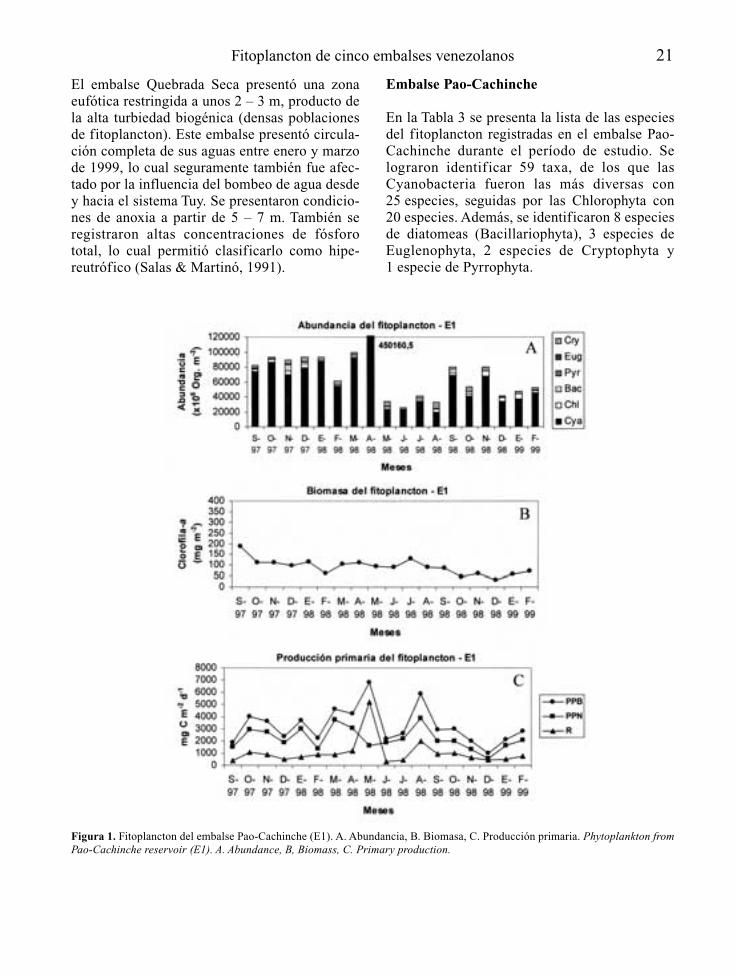
Embalse Pao-Cachinche
En la Tabla 3 se presenta la lista de las especiesdel fitoplancton registradas en el embalse Pao-Cachinche durante el período de estudio. Selograron identificar 59 taxa, de los que lasCyanobacteria fueron las más diversas con25 especies, seguidas por las Chlorophyta con20 especies. Además, se identificaron 8 especiesde diatomeas (Bacillariophyta), 3 especies deEuglenophyta, 2 especies de Cryptophyta y1 especie de Pyrrophyta.
Figura 1. Fitoplancton del embalse Pao-Cachinche (E1). A. Abundancia, B. Biomasa, C. Producción primaria. Phytoplankton fromPao-Cachinche reservoir (E1). A. Abundance, B, Biomass, C. Primary production.
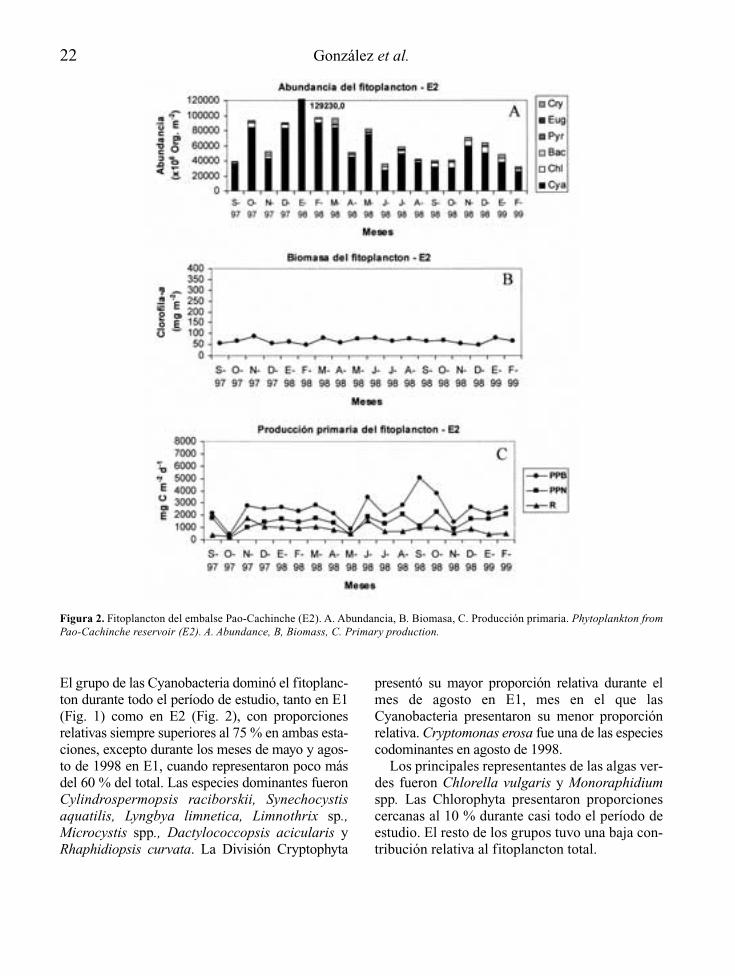
El grupo de las Cyanobacteria dominó el fitoplanc-ton durante todo el período de estudio, tanto en E1(Fig. 1) como en E2 (Fig. 2), con proporcionesrelativas siempre superiores al 75 % en ambas esta-ciones, excepto durante los meses de mayo y agos-to de 1998 en E1, cuando representaron poco másdel 60 % del total. Las especies dominantes fueronCylindrospermopsis raciborskii, Synechocystisaquatilis, Lyngbya limnetica, Limnothrix sp.,Microcystis spp., Dactylococcopsis acicularis yRhaphidiopsis curvata. La División Cryptophyta
presentó su mayor proporción relativa durante elmes de agosto en E1, mes en el que lasCyanobacteria presentaron su menor proporciónrelativa. Cryptomonas erosa fue una de las especiescodominantes en agosto de 1998.
Los principales representantes de las algas ver-des fueron Chlorella vulgaris y Monoraphidiumspp. Las Chlorophyta presentaron proporcionescercanas al 10 % durante casi todo el período deestudio. El resto de los grupos tuvo una baja con-tribución relativa al fitoplancton total.
22 González et al.
Figura 2. Fitoplancton del embalse Pao-Cachinche (E2). A. Abundancia, B. Biomasa, C. Producción primaria. Phytoplankton fromPao-Cachinche reservoir (E2). A. Abundance, B, Biomass, C. Primary production.

La abundancia del fitoplancton se estimó en lasdos estaciones limnéticas, y en E1 la mayorabundancia se registró durante el mes de abrilde 1998, con valores de 450 x 109 org./m2 (Fig.1A). Este valor extremo se debió a un grandesarrollo o floraciones (“bloom”) de las espe-cies Synechocystis aquatilis y Microcystis spp.Por el contrario, la menor abundancia del fito-plancton se registró en junio, con una abundan-cia de 25.9 x 109 org./m2, posiblemente comoconsecuencia de una menor disponibilidad denutrientes (principalmente ortofosfatos). Tam-bién pudo haber influido el hecho de que lasaguas de esta estación fueron tratadas conalguicidas y otros químicos (sulfato de cobre,sulfato de aluminio e hipoclorito de calcio)durante el mes de mayo de 1998, a fin de con-trolar las densas poblaciones de algas. La abun-dancia promedio del fitoplancton fue de 86,1 x109 org./m2 en E1. Este valor puede considerar-se bastante elevado.
La biomasa del fitoplancton en E1 varió entre32.42 mg/m2 (diciembre de 1998) y 186.63 mg/m2
(septiembre de 1997), con un valor promedio de92.13 mg/m2 (Fig. 1B). Estos valores se puedenconsiderar elevados. En términos volumétricos, elvalor promedio fue de 42.40 µg/l.
Se encontró una correlación estadísticamentesignificativa entre la abundancia del fitoplanc-ton y el coeficiente de atenuación de la luz enE1 (r= 0.5081; p<0.05), lo que confirmó que lasalgas fueron responsables en gran medida de lapobre penetración de la luz hacia los estratosmás profundos en E1. La transparencia del aguaestuvo significativamente correlacionada enforma negativa con la abundancia del fitoplanc-ton (González et al., 2000), reforzando la afir-mación anterior (r= -0.5623; p<0.05). De igualforma, la biomasa del f itoplancton, medidacomo concentración de clorofila-a, tambiénestuvo correlacionada de manera significativacon el coeficiente de atenuación de la luz(r = 0.5748; p<0.05).
Por su parte, en E2 se registraron valores deabundancia del fitoplancton con menores fluc-tuaciones que en E1 (Fig. 2A). La abundanciade las algas varió entre 31.6 x 109 org./m2
(febrero de 1999) y 129.2 x 109 org./m2 (enerode 1998), con una media de 64.6 x 109
org./m2. Este valor puede considerarse muyelevado, a pesar de ser menor que el determi-nado para E1.
La variación de la biomasa del fitoplancton enE2 también fue elevada, con valores entre 45.74mg/m2 (febrero de 1998) y 85.40 mg/m2 (noviem-bre de 1997), y un promedio de 65.35 mg/m2
(Fig. 2B). Este promedio también fue menor queel determinado para E1. La concentración de clo-rofila-a promedio, expresada en unidades volu-métricas, fue de 25.08 µg/l.
Los mayores valores y las mayores fluctua-ciones de abundancia y de biomasa del fito-plancton registrados en la estación ubicada allado de la torre – toma, en relación con losestimados en la estación ubicada en el brazoeste del embalse, pudieran ser el resultado dela presencia de un aliviadero en E1. El fito-plancton en esta región pareció responder rápi-damente a la entrada de nutrientes desde lostributarios (especialmente desde el río Paito),pero en los meses en que se dio el flujo de sali-da del agua por el aliviadero se pudo acelerarla pérdida de nutrientes y de algas a una mayortasa que en E2, en la que no había aliviadero yen la que las aguas parecieron estancarse unpoco como consecuencia del estrecho quesepara ambos brazos del embalse, el cual impi-de un flujo libre de agua entre ambas regionesde este cuerpo de agua.
Las figuras 1C y 2C muestran las variacionesde la producción primaria del fitoplancton enE1 y E2, respectivamente. E1 presentó mayoresfluctuaciones que E2, probablemente debido ala extracción de agua desde la torre-toma. Eneste embalse, sólo en E1 las aguas fueron trata-das con alguicidas (CuSO4 y AlSO4) y, segura-mente, esto también afectó los procesos del fito-plancton. La respiración representó, enpromedio, el 32% y el 34% de la producciónprimaria bruta (PPB) en E1 y E2, respectiva-mente. Los valores promedios de la PPB fueron3229 mg C m-2 d-1 y 2482 mg C m-2 d-1 para E1y E2, respectivamente, que pueden considerarsecomo valores elevados.
Fitoplancton de cinco embalses venezolanos 23

Embalse La Mariposa
En la Tabla 4 se presenta la lista de especies delfitoplancton presentes en el embalse LaMariposa durante el período de estudio. Selograron identificar 50 especies. Las divisionesmás diversas fueron las Chlorophyta (algas ver-des) y las Cyanobacteria, cada una con 18 espe-cies, seguidas por las Bacillariophyta con 8 es-pecies. La especie dominante durante casi todoel período de estudio fue Cryptomonas erosa(Cryptophyta). Otras especies abundantes fueronChroomonas sp. (Cryptophyta), Mallomonas sp.(Chrysophyta), Chlorella vulgaris y Schroederiasetigera (Chlorophyta) y Merismopedia elegans
(Cyanobacteria). Aulacoseira granulata (Ba-cillariophyta) fue abundante durante los mesesde septiembre y febrero.
La figura 3 presenta las variaciones de abun-dancia, biomasa y de producción primaria delfitoplancton durante el período de estudio.
La abundancia del fitoplancton, presentó elmismo patrón de variación que la biomasa, conpicos de 35.5 x 109 org./m2 y 34.8 x 109 org./m2
en septiembre y febrero, respectivamente. Elgrupo dominante fue el de las Cryptophytadurante casi todo el período de estudio, con pro-porciones relativas que a veces superaron el80 %. Las Chlorophyta dominaron en enero,durante la circulación completa de las aguas, y
24 González et al.
Tabla 4. Lista de las especies del fitoplancton encontradas en el embalse La Mariposa. List of phytoplankton species collected in La Mariposareservoir.
Cyanobacteria1. Anabaena sp.2. Aphanocapsa sp.3. Coelosphaerium kuetzingianum4. Cylindrospermopsis raciborskii5. Chroococcus limneticus6. Chroococcus minutus7. Dactylococcopsis acicularis8. Gomphosphaeria aponina9. Lyngbya limnetica10. Microcystis aeruginosa11. Microcystis delicatissima12. Merismopedia glauca13. Merismopedia punctata14. Oscillatoria sp.15. Phormidium sp.16. Spirulina sp.17. Synechococcus sp.18. Synechocystis aquatilis
Chlorophyta19. Actinastrum hantzschii20. Asterococcus sp.21. Closterium ehrenbergii22. Coelastrum microporum23. Cosmarium sp.24. Crucigenia tetrapedia25. Chlorella vulgaris26. Dictyosphaerium pulchellum27. Eudorina elegans28. Micractinium pusillum29. Monoraphidium minutum
30. Monoraphidium tortile31. Oocystis sp.32. Pediastrum simplex33. Scenedesmus acuminatum34. Schroederia setigera35. Sphaerocystis schroeteri36. Staurastrum sp.
Chrysophyta37. Mallomonas sp.
Bacillariophyta38. Aulacoseira granulata39. Cyclotella meneghiniana40. Denticula sp.41. Gomphonema sp.42. Navicula sp.43. Nitzschia sp.44. Rhizosolenia sp.45. Synedra sp.
Euglenophyta46. Euglena sp.47. Trachelomonas volvocina
Cryptophyta48. Cryptomonas erosa49. Chroomonas sp.
Pyrrophyta50. Peridinium sp.
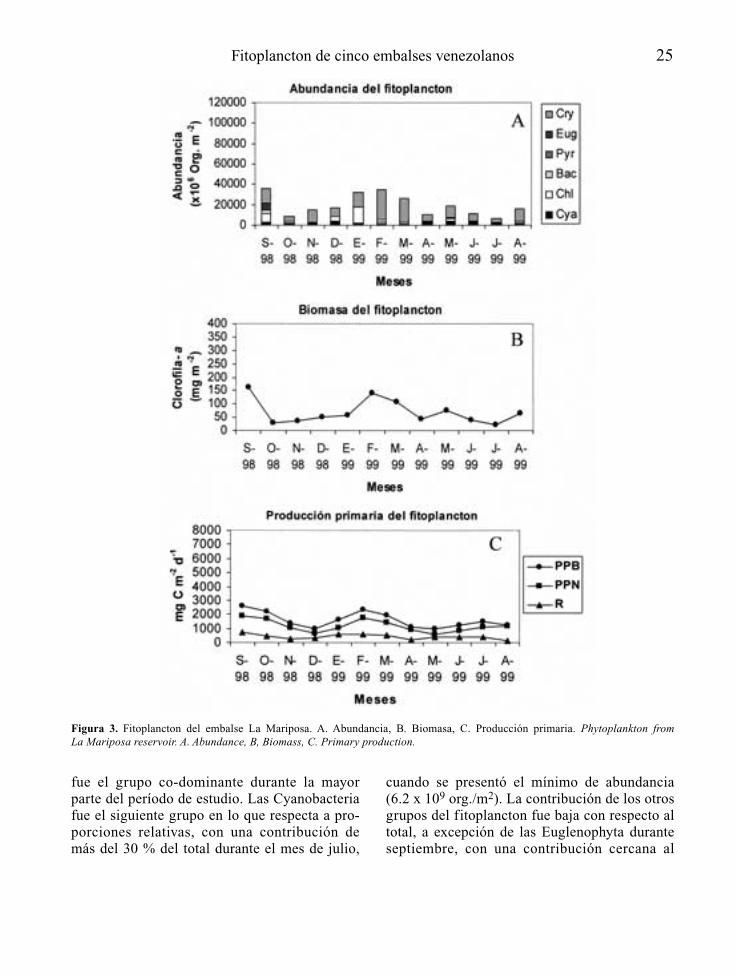
fue el grupo co-dominante durante la mayorparte del período de estudio. Las Cyanobacteriafue el siguiente grupo en lo que respecta a pro-porciones relativas, con una contribución demás del 30 % del total durante el mes de julio,
cuando se presentó el mínimo de abundancia(6.2 x 109 org./m2). La contribución de los otrosgrupos del fitoplancton fue baja con respecto altotal, a excepción de las Euglenophyta duranteseptiembre, con una contribución cercana al
Fitoplancton de cinco embalses venezolanos 25
Figura 3. Fitoplancton del embalse La Mariposa. A. Abundancia, B. Biomasa, C. Producción primaria. Phytoplankton fromLa Mariposa reservoir. A. Abundance, B, Biomass, C. Primary production.

20 % del total, y de las Bacillariophyta duranteseptiembre y febrero, meses en que su contribu-ción fue cercana al 10 %.
El máximo valor de biomasa (como clorofila-a) fue de 161.77 mg/m2, y se registró durante elmes de septiembre de 1998; otro pico de magni-tud similar se registró en febrero de 1999, des-pués de la mezcla completa de las aguas delembalse (valor de 140.16 mg/m2). El valor pro-medio fue de 68.74 mg/m2 (media por unidad de
volumen = 41.92 µg/l). El patrón de la variaciónde la biomasa coincidió con los patrones de varia-ción de la conductividad, nitrógeno total, fósforototal y los nitratos y ortofosfatos (Ortaz et al.,1999). El fitoplancton, por lo tanto, respondiódirectamente a la mayor disponibilidad denutrientes en el agua, los cuales fueron resuspen-didos durante la época de circulación de las aguas(enero a marzo de 1999) y cuando se bombeóagua desde otros embalses (septiembre de 1998).
26 González et al.
Tabla 5. Lista de especies del fitoplancton encontradas en el embalse Lagartijo. List of phytoplankton species colleted in Lagartijo reservoir.
Cyanobacteria1. Anabaena aphanizomenoides2. Aphanothece sp.3. Cylindrospermopsis raciborskii4. Chroococcus limneticus5. Chroococcus minimus6. Chroococcus pallidus7. Dactylococcopsis acicularis8. Gomphosphaeria sp.9. Lyngbya limnetica10. Microcystis holsatica11. Oscillatoria annae12. Spirulina laxissima13. Synechococcus leopoliensis14. Synechococcus linearis15. Synechocystis aquatilis
Chlorophyta16. Actinotaenium minutissimum17. Botryococcus braunii18. Closterium dianae19. Coelastrum microporum20. Coelastrum reticulatum21. Crucigenia fenestra22. Crucigenia quadrata23. Crucigeniella rectangularis24. Chlamydomonas sp.25. Chlorella saccarophila26. Chlorella vulgaris27. Dictyosphaerium pulchellum28. Elakatothrix gelatinosa29. Eudorina sp.30. Gyoerffyana humicola31. Kirchneriella contorta32. Monoraphidium minutum33. Monoraphidium tortile34. Nephrochlamys sp.35. Nephrocytium aghardianum36. Oocystis eremosphaeria37. Oocystis lacustris
38. Oocystis parva39. Phacotus sp.40. Scenedesmus arcuata41. Scenedesmus ecornis42. Schroederia setigera43. Sphaerocystis schroeteri44. Spirotaenia minuta45. Staurastrum tetracerum46. Tetraedron incus47. Tetraedron sp.48. Tetrastrun triangulare49. Volvox sp.
Chrysophyta50. Mallomonas sp.
Bacillariophyta51. Actinella sp.52. Achnanthes sp.53. Aulacoseira granulata54. Cyclotella meneghiniana55. Cymbella sp.56. Denticula sp.57. Gomphonema gracile58. Navicula sp.59. Nitzschia sp.60. Rhizosolenia sp.61. Stephanodiscus hantzschii62. Synedra ulna
Euglenophyta63. Euglena sp.64. Trachelomonas volvocina
Cryptophyta65. Cryptomonas erosa66. Chroomonas nordstedtii
Pyrrophyta67. Peridinium sp.

González & Ortaz (1998), realizando experi-mentos de fertilización con N y P en microcos-mos en este embalse, demostraron que el enri-quecimiento con estos nutrientes, a pesar de lasaltas concentraciones presentes en las aguasnaturales, condujeron a un aumento de la bio-masa del fitoplancton dentro de los microcos-mos. Esto indica que el f itoplancton puedeseguir aumentando sus densidades si continúael proceso de eutrofización del embalse, yaque la entrada de nutrientes al embalse conti-núa siendo elevada.
La PPB del fitoplancton, presentó la mismatendencia que la abundancia y la biomasadel fitoplancton, con picos de 2633.29 y2366.01 mg C m-2 d-1 durante los meses de sep-tiembre y febrero, respectivamente. El primerpico pudo deberse a la entrada de nutrientes porescorrentía en plena estación de lluvias del año1998, mientras que el pico de febrero pudo serconsecuencia de una mayor disponibilidad denutrientes en la columna de agua como conse-cuencia de la circulación de las aguas. Un picomenor de producción primaria bruta (PPB) de1516.66 mg C m-2 d-1 registrado en julio, pudodeberse al ligero aumento en la conductividad(aumento de la concentración de electrólitos) y alaumento en las concentraciones de nitratos, amo-nio y ortofosfatos (Ortaz et al., 1999). El menorvalor de la PPB fue de 968.21 mg C m-2 d-1, y seregistró en mayo de 1999.
La PPB promedio fue de 1601.61 mg C m-2 d-1,valor que puede considerarse alto, y la respiraciónrepresentó el 26 % de este valor.
Embalse Lagartijo
En la Tabla 5 se presenta la lista de especies delfitoplancton presentes en el embalse Lagartijoidentificadas durante el período de estudio. Selograron identificar 67 especies, siendo las másdiversas las Chlorophyta (algas verdes) con34 especies.
La figura 4 muestra las variaciones de abun-dancia, biomasa y de producción primaria delfitoplancton. La abundancia del fitoplanctontambién fue relativamente baja en este ecosis-
tema, con valores entre 4.6 x 109 org./m2
(mayo) y 27.2 x 109 org./m2 (diciembre). Elpico de abundancia de diciembre se debió alnotable aumento de densidad de la especieSphaerocystis schroeteri (alga verde). Otrospicos secundarios de abundancia se registraronen octubre y en julio, debido al desarrollo delas poblaciones de Gomphonema gracile(Bacillariophyta o diatomea) Lyngbya limneti-ca (Cyanobacteria) y Cryptomonas erosa(Cryptophyta), respectivamente.
Las algas verdes dominaron el fitoplanctondurante casi todo el lapso de estudio, tal comose aprecia en la figura 4A. Las Chlorophyta lle-garon a representar más del 80 % del total delfitoplancton presente en diciembre, cuando sedesarrolló la especie colonial Sphaerocystisschroeteri, tal como se explicó anteriormente.En octubre y noviembre, las Bacillariophytadominaron ligeramente sobre las Chlorophyta,debido al desarrollo de las poblacionesde Gomphonema gracile. En febrero hubo unacontribución importante de la diatomeaStephanodiscus hantzschii, que generó un ligeroaumento en la proporción de las diatomeas acasi el 25 % del total. Las Cyanobacteria contri-buyeron con menos del 10 % del total de sep-tiembre a diciembre, con un ligero aumento enenero a más del 25 % debido al desarrollo de laspoblaciones de Synechococcus spp., para dismi-nuir a menos del 10 % en febrero nuevamente.A partir de marzo, la contribución de este grupoaumentó gradualmente, hasta contribuir conmás del 80 % del total del fitoplancton en junio,debido al incremento en las densidades deLyngbya limnetica y Cylindrospermopsis raci-borskii. En julio, Lyngbya limnetica fue la espe-cie dominante del fitoplancton. Esta alta abun-dancia de las Cyanobacteria en el período delluvias también fue registrado por Infante et al.,(1992) y por González et al. (1997).
La mayor contribución de las Cryptophyta seregistró en julio de 1999, con cerca de un 25 %del total, cuando Cryptomonas erosa fue laespecie co-dominante. Durante el resto del perí-odo, su contribución al fitoplancton del embalseLagartijo fue baja, aunque en septiembre ésta
Fitoplancton de cinco embalses venezolanos 27
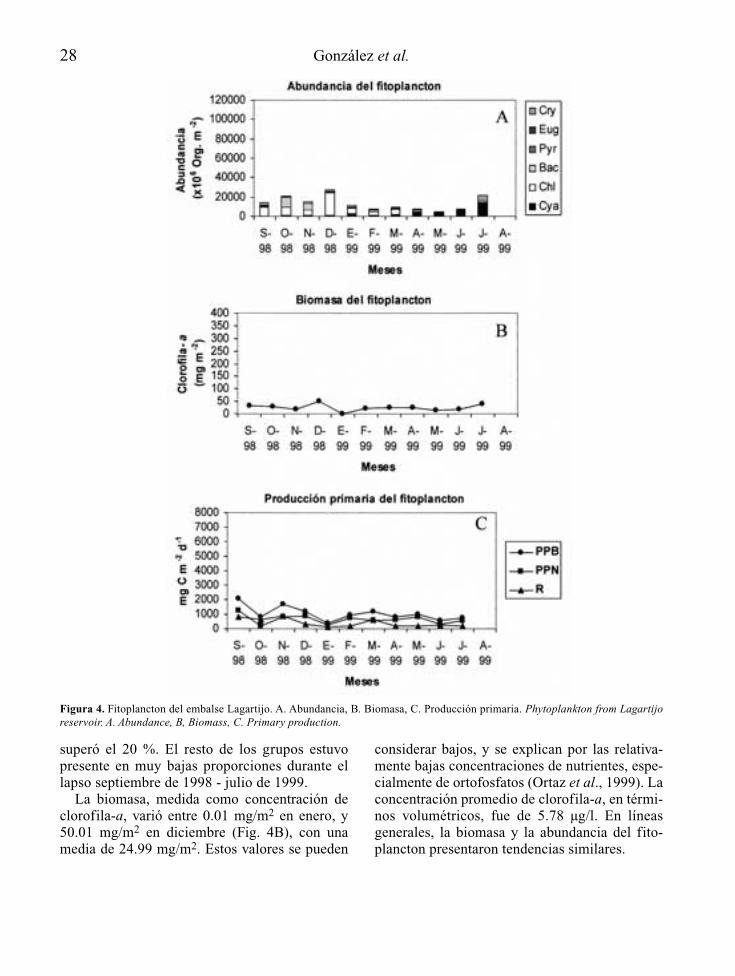
superó el 20 %. El resto de los grupos estuvopresente en muy bajas proporciones durante ellapso septiembre de 1998 - julio de 1999.
La biomasa, medida como concentración declorofila-a, varió entre 0.01 mg/m2 en enero, y50.01 mg/m2 en diciembre (Fig. 4B), con unamedia de 24.99 mg/m2. Estos valores se pueden
considerar bajos, y se explican por las relativa-mente bajas concentraciones de nutrientes, espe-cialmente de ortofosfatos (Ortaz et al., 1999). Laconcentración promedio de clorofila-a, en térmi-nos volumétricos, fue de 5.78 µg/l. En líneasgenerales, la biomasa y la abundancia del fito-plancton presentaron tendencias similares.
28 González et al.
Figura 4. Fitoplancton del embalse Lagartijo. A. Abundancia, B. Biomasa, C. Producción primaria. Phytoplankton from Lagartijoreservoir. A. Abundance, B, Biomass, C. Primary production.

La producción primaria bruta (PPB) presentógrandes variaciones durante el período de estudio(Fig. 4C), con valores entre 376.05 mg C m-2 d-1
en enero y 2087.80 mg C m-2 d-1 en septiembre.Estos valores se pueden considerar de bajos amoderados. El menor valor de producción prima-ria coincidió con el menor valor de biomasa,cuando dominaron las especies de pequeño tama-ño, como Cryptomonas erosa y Chlorella spp.(unicelulares) y Monoraphidium spp. (filamento-sas). Las diatomeas presentaron bajas densidadesdurante este mes. El aumento de la PPB en juliocoincidió con el incremento de la abundancia y dela biomasa durante este mes. En promedio, laPPB fue de 1031 mg C m-2 d-1, y la respiracióndel fitoplancton representó el 38 % de la PPB.Debido a su condición oligotrófica, este embal-se fue el que presentó los menores valores de
abundancia, biomasa y producción primaria delos cinco embalses estudiados.
Embalse La Pereza
En la Tabla 6 se presenta la lista de las especiesidentificadas en el embalse La Pereza. Se lograronidentificar 48 especies, siendo las más diversas lasChlorophyta, con 21 especies. Cryptomonas erosa(Cryptophyta) y las diatomeas Cyclotella mene-ghiniana, Aulacoseira granulata, Synedra sp. yNavicula sp. dominaron el fitoplancton durantecasi todo el período de estudio. Las CyanobacteriaSynechococcus sp. y Synechocystis aquatilis, y elalga verde Chlorella vulgaris dominaron el fito-plancton durante el mes de junio.
La abundancia, biomasa y producción prima-ria del fitoplancton se presentan en la Fig. 5. Las
Fitoplancton de cinco embalses venezolanos 29
Tabla 6. Lista de las especies del fitoplancton encontradas en el embalse La Pereza. List of phytoplankton species colleted in La Perezareservoir.
Cyanobacteria1. Cylindrospermopsis raciborskii2. Chroococcus sp.3. Dactylococcopsis acicularis4. Gomphosphaeria sp.5. Lyngbya limnetica6. Marssoniella sp.7. Merismopedia sp.8. Microcystis holsatica9. Oscillatoria sp.10. Phormidium sp.11. Rhaphidiopsis sp.12. Spirulina sp.13. Synechococcus sp.14. Synechocystis aquatilis
Chlorophyta15. Actinastrum hantzschii16. Asterococcus sp.17. Closterium sp.18. Coelastrum sp.19. Cosmarium sp.20. Chlamydomonas sp.21. Chlorella vulgaris22. Chodatella sp.23. Crucigenia sp.24. Dictyosphaerium sp.25. Elakatohrix gelatinosa26. Eudorina sp.27. Monoraphidium sp.
28. Nephrocytium sp.29. Oocystis sp.30. Pandorina sp.31. Scenedesmus ecornis32. Schroederia setigera33. Sphaerocystis schroeteri34. Staurastrum sp.35. Tetraedron sp.
Bacillariophyta36. Aulacoseira granulata37. Cyclotella meneghiniana38. Eunotia sp.39. Gomphonema sp.40. Navicula sp.41. Nitzschia sp.42. Stephanodiscus hantzschii43. Synedra sp.
Euglenophyta44. Euglena sp.45. Trachelomonas sp.
Cryptophyta46. Cryptomonas erosa47. Chroomonas sp.
Pyrrophyta48. Peridinium sp.
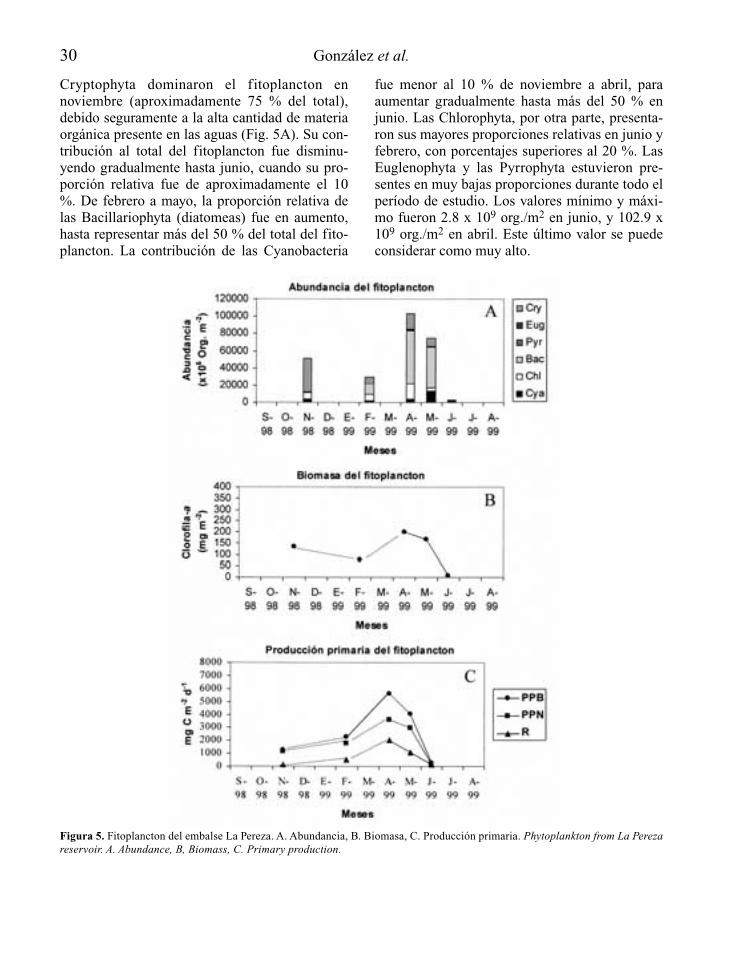
Cryptophyta dominaron el fitoplancton ennoviembre (aproximadamente 75 % del total),debido seguramente a la alta cantidad de materiaorgánica presente en las aguas (Fig. 5A). Su con-tribución al total del fitoplancton fue disminu-yendo gradualmente hasta junio, cuando su pro-porción relativa fue de aproximadamente el 10%. De febrero a mayo, la proporción relativa delas Bacillariophyta (diatomeas) fue en aumento,hasta representar más del 50 % del total del fito-plancton. La contribución de las Cyanobacteria
fue menor al 10 % de noviembre a abril, paraaumentar gradualmente hasta más del 50 % enjunio. Las Chlorophyta, por otra parte, presenta-ron sus mayores proporciones relativas en junio yfebrero, con porcentajes superiores al 20 %. LasEuglenophyta y las Pyrrophyta estuvieron pre-sentes en muy bajas proporciones durante todo elperíodo de estudio. Los valores mínimo y máxi-mo fueron 2.8 x 109 org./m2 en junio, y 102.9 x109 org./m2 en abril. Este último valor se puedeconsiderar como muy alto.
30 González et al.
Figura 5. Fitoplancton del embalse La Pereza. A. Abundancia, B. Biomasa, C. Producción primaria. Phytoplankton from La Perezareservoir. A. Abundance, B, Biomass, C. Primary production.

La biomasa, estimada como concentración declorofila-a, varió entre 5.48 mg/m2 en junio, y199.21 mg/m2 en abril (Fig. 5B), con una mediade 117.10 mg/m2. En términos de unidades devolumen, el promedio fue de 44.37 µg/l. Las con-diciones de hipoxia en toda la columna de aguaen junio, limitaron el crecimiento del fitoplanc-ton, que presentó una notable disminución de subiomasa y abundancia de mayo a junio. El patrónde variación de la abundancia del fitoplanctoncoincidió con el de la variación de biomasa.
Los bajos valores de biomasa en junio sedebieron a la dominancia de especies de pequeñotamaño, como Synechococcus sp., Synechocystisaquatilis y Chlorella vulgaris. El pico de bioma-sa en abril y los altos valores de mayo se debie-
ron a una alta tasa de desarrollo de las diatomeasCyclotella meneghiniana, Aulacoseira granulata,Synedra sp. y Navicula sp. y de la especie flage-lada Cryptomonas erosa, como consecuencia dela alta disponibilidad de nutrientes en el cuerpode agua. La alta abundancia de diatomeas le con-firieron al agua un color pardo amarillento enestos meses. La alta abundancia del fitoplanctonen los meses de abril y mayo, fueron responsa-bles de los menores valores de transparencia delagua y de los mayores valores del coeficiente deatenuación de la luz registrados en las aguas delembalse (Ortaz et al., 1999).
La producción primaria bruta del fitoplanctonsiguió la misma tendencia de la abundancia y dela biomasa (Fig. 5C). El pico de producción se
Fitoplancton de cinco embalses venezolanos 31
Tabla 7. Lista de especies del fitoplancton encontradas en el embalse Quebrada Seca. List of phytoplankton species collected in QuebradaSeca reservoir.
Cyanobacteria1. Anabaena sp.2. Aphanocapsa sp.3. Aphanothece sp.4. Cylindrospermopsis raciborskii5. Chroococcus sp.6. Dactylococcopsis acicularis7. Gomphosphaeria sp.8. Lyngbya limnetica9. Merismopedia elegans10. Microcystis aeruginosa11. Oscillatoria sp.12. Rhaphidiopsis sp.13. Spirulina sp.14. Synechococcus sp.15. Synechocystis aquatilis
Chlorophyta16. Actinastrum hantzschii17. Asterococcus sp.18. Closterium sp.19. Coelastrum sp.20. Cosmarium sp.21. Crucigenia sp.22. Chlamydomonas sp.23. Chlorella vulgaris24. Dictyosphaerium pulchellum25. Elakatothrix gelatinosa26. Eudorina sp.27. Monoraphidium sp.28. Nephrocytium sp.
29. Oocystis sp.30. Pandorina sp.31. Phacotus sp.32. Pediastrum sp.33. Scenedesmus sp.34. Schroederia setigera35. Sphaerocystis sp.36. Tetraedron sp.
Chrysophyta37. Mallomonas sp.
Bacillariophyta38. Aulacoseira granulata39. Cyclotella sp.40. Navicula sp.41. Nitzschia sp.42. Rhizosolenia sp.43. Synedra sp.
Euglenophyta44. Euglena sp.45. Trachelomonas sp.
Cryptophyta46. Cryptomonas erosa47. Chroomonas sp.
Pyrrophyta48. Peridinium sp.

registró en abril, con un valor de 5618.67 mg Cm-2 d-1, valor comparable a uno de los registradosen uno de los ecosistemas más productivos yeutrofizados del país, como lo es el Lago deValencia (Infante, 1997). Esta alta PPB fue la res-ponsable de la sobresaturación de oxígeno regis-trada durante el mes de abril (Ortaz et al.,1999).En mayo también se registró un alto valor dePPB, equivalente a 4055.85 mg C m-2 d-1, res-ponsable de la sobresaturación de oxígeno super-ficial en este mes. En cambio, durante junio,debido a la limitación causada por los bajos valo-res de oxígeno disuelto en el agua, la produccióndisminuyó a 264.49 mg C m-2 d-1. La PPB pro-medio fue de 2690.68 mg C m-2 d-1, mientras quela respiración promedio fue de 753.94 mg C m-2
d-1, lo cual representó el 29 % de la PPB.
Embalse Quebrada Seca
La Tabla 7 presenta la lista de especies del fito-plancton encontradas en el período de estudio. Entotal, se encontraron 48 especies, siendo lasChlorophyta las más diversas con 21 especies. LaCyanobacteria Cylindrospermopsis raciborskiidominó durante casi todo el período de estudio.
La figura 6A muestra que las Cyanobacteriadominaron el fitoplancton de octubre a marzo,con proporciones superiores al 60 % del total.Cylindrospermopsis raciborskii, Oscillatoria sp.,y Anabaena sp. estuvieron presentes en altas den-sidades en este período. Las Cyanobacteria co-dominaron con las algas verdes en junio, conuna proporción de 50 %; las CyanobacteriaDactylococcopsis acicularis y Synechocystisaquatilis, y las Chlorophyta Chlorella vulgaris ySchroederia setigera fueron las especies predo-minantes en este mes. En mayo dominaron lasBacillariophyta, debido al desarrollo de las pobla-ciones de Aulacoseira granulata. Las Chlo-rophyta co-dominaron también en este mes, debi-do a las altas abundancias de Phacotus sp. yNephrocytium sp. Las Cryptophyta presentaronsus mayores proporciones relativas en octubre,superando el 10 % del total. En el resto del perío-do de estudio, este grupo presentó bajas contribu-ciones relativas al total del fitoplancton presente,
al igual que los demás grupos de esta comunidad.La abundancia del fitoplancton varió entre
26.9 x 109 org./m2 y 69.3 x 109 org./m2 en octu-bre y diciembre, respectivamente, con un pro-medio de 42.5 x 109 org./m2.
La biomasa del f itoplancton presentó sumáximo valor durante marzo, con una concen-tración de clorofila-a de 355.35 mg/m2 (Fig.6B). La mayor disponibilidad de ortofosfatos enla zona eufótica hizo posible este valor extremo(Ortaz et al., 1999). Los menores valores seregistraron en mayo (53.73 mg/m2) y diciembre(59.20 mg/m2), con un promedio de 134.74mg/m2 (en términos volumétricos, el promediofue de 62.71 µg/l).
La abundancia del fitoplancton presentó unatendencia inversa a la de la biomasa del fitoplanc-ton. El pico de abundancia de diciembre se debióa las altas densidades de Cylindrospermopsisraciborskii y gran abundancia de especies depequeño tamaño (Cryptomonas erosa, Chlorellavulgaris) o de filamentos delgados (Oscillatoriasp., Rhaphidiopsis sp.), las cuales poseen menorcontenido de pigmentos fotosintéticos que las quepredominaron en marzo, como lo fueron los gran-des filamentos de Anabaena sp. Otro pico deabundancia se presentó en mayo, cuando la diato-mea Aulacoseira granulata fue dominante. Estadiatomea tiene en el citoplasma vacuolas de grantamaño, por lo que el contenido de clorofila-a eslimitado, lo cual explica los mínimos valores debiomasa encontrados.
La producción primaria bruta mostró la mismatendencia que la biomasa del fitoplancton, convalores entre 1762.50 y 3493.99 mg C m-2 d-1 enmayo y marzo, respectivamente (Fig. 6C), conuna media de 2466.92 mg C m-2 d-1. Estos valo-res se pueden considerar altos. La respiraciónrepresentó en promedio el 21 % de la PPB.
En la figura 7 se presentan, a modo de resu-men, las proporciones promedios de los diferen-tes grupos del fitoplancton en cada uno de losembalses estudiados. Se puede notar que lasCyanobacteria dominaron en aquellos embalsescon alto grado de eutrofización, en los que la tur-biedad se debió en primer término a la materiaorgánica (fitoplancton), como lo fueron Pao-
32 González et al.
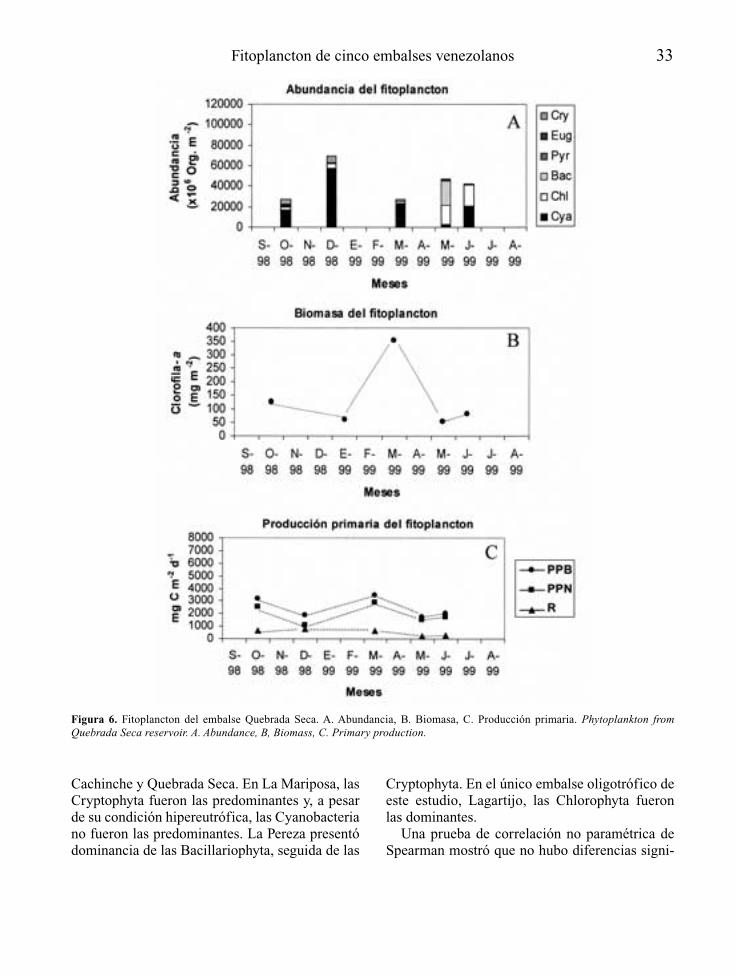
Cachinche y Quebrada Seca. En La Mariposa, lasCryptophyta fueron las predominantes y, a pesarde su condición hipereutrófica, las Cyanobacteriano fueron las predominantes. La Pereza presentódominancia de las Bacillariophyta, seguida de las
Cryptophyta. En el único embalse oligotrófico deeste estudio, Lagartijo, las Chlorophyta fueronlas dominantes.
Una prueba de correlación no paramétrica deSpearman mostró que no hubo diferencias signi-
Fitoplancton de cinco embalses venezolanos 33
Figura 6. Fitoplancton del embalse Quebrada Seca. A. Abundancia, B. Biomasa, C. Producción primaria. Phytoplankton fromQuebrada Seca reservoir. A. Abundance, B, Biomass, C. Primary production.

ficativas (p<0.05) entre el orden jerárquico de losgrupos del fitoplancton de E1 y E2 del embalsePao-Cachinche (rs = 0.9429), ya que pertenecen aun mismo embalse y con una condición hipereu-trófica similar, y entre los embalses Lagartijo yQuebrada Seca (rs = 0.9429), los cuales sólodifieren en el orden jerárquico de dos grupos:Cyanobacteria dominó en Quebrada Seca, segui-da de Chlorophyta, mientras que en Lagartijodominó Chlorophyta, seguida de Cyanobacteria.
CONSIDERACIONES FINALES
Además de las funciones de fuerza naturales(viento, temperatura y precipitación), el manejode los cuerpos de agua estudiados afecta nota-blemente la dinámica de todos los parámetrosdeterminados en todos los embalses evaluados,lo que lo convierte en una función de fuerzabásica también (Tundisi & Matsumura-Tundisi,1990). Las variaciones de volumen de los embal-ses, especialmente en La Pereza, Quebrada Secay La Mariposa, pudo contribuir enormementecon la resuspensión de los elementos nutritivosdesde el fondo de la columna de agua, así comotambién a su reposición en la zona eufóticadonde pudieron ser asimilados por el fitoplanc-ton. Adicionalmente, las grandes fluctuacionesen el nivel de agua aumentan las posibilidadesde erosión de la línea de costa de los embalses,
lo cual contribuiría a aumentar la carga de sedi-mentos inorgánicos y al pobre desarrollo decomunidades litorales.
En el caso del embalse Pao-Cachinche, las fluc-tuaciones en la abundancia, biomasa y producciónprimaria del fitoplancton se debieron a la entradade nutrientes, al autosombreo de las algas y al tra-tamiento con los químicos CuSO4 y AlSO4.
La mayoría de los embalses evaluados en esteestudio, y que son empleados para el suministrode agua potable, se encuentran eutrofizados, yen ellos predominan las Cyanobacteria y lasCryptophyta, con elevados valores de abundan-cia, biomasa y producción primaria, como con-secuencia de la entrada excesiva de nutrientesdesde sus cuencas de drenaje altamente erosio-nadas e intervenidas y a través de las aguas ser-vidas sin tratamiento previo. Aún en el embalseoligotrófico, durante el inicio de la temporadade lluvias, las Cyanobacteria aumentaron susproporciones relativas, lo cual puede ser un pro-blema al momento de la utilización de las aguas,ya que todos estos son embalses que se empleanpara el suministro de agua potable.
Los embalses La Mariposa, La Pereza yQuebrada Seca pudieran presentar mayores gra-dos de eutrofización, de no ser por el bajo tiempode residencia de sus aguas (Straskraba, 1999).
De los cuatro embalses eutrofizados, sólo enel embalse Pao-Cachinche se tiene previsto unplan para su mitigación (desestratificación), aser aplicado próximamente. En todos los casos,el control de la entrada de nutrientes (especial-mente fósforo) a los embalses, parece ser la víapara mitigar la eutrofización y controlar efecti-vamente el crecimiento de las algas indeseables(colonias y filamentos de Cyanobacteria, espe-cialmente), de acuerdo a los sugerido porGonzález & Ortaz (1998), Ortaz et al. (1999) yGonzález et al. (2000).
AGRADECIMIENTOS
A las empresas hidrológicas Hidroven, Hidro-centro e Hidrocapital, y a la empresa Hidro-impacto, por financiar los proyectos de investiga-
34 González et al.
Figura 7. Proporciones promedios de los grupos del fito-plancton en los cinco embalses estudiados. Mean proportionsof the phytoplankton groups in the five studied reservoirs.

ción. A Pasquale Molinaro, Pablo Borjas, AldoCróquer, Sandra Cano y Sandra Keri, por su co-laboración en la ejecución del proyecto. Alpersonal de vigilancia y de operaciones deHidrocapital y de Hidrocentro en los embalses LaMariposa, La Pereza y Pao-Cachinche, por suapoyo en los trabajos de campo.
BIBLIOGRAFÍA
CASTILLO, C., J. GÓMEZ & C. MONTES. 1973.Embalses de Venezuela. Serie El Agua Nº 1.Caracas.
EDMONDSON, W. T. 1969. Eutrophication in NorthAmerica. Nat. Acad. Sci.: 124-149.
GINEZ, A. & M. L. OLIVO. 1984. Inventario de losembalses con información básica para la activi-dad piscícola. I. Sinopsis de los embalses admi-nistrados por el MARNR. Ministerio del Ambientey de los Recursos Naturales Renovables. SerieInformes Técnicos DGSPOA/IT/183. Caracas,142 pp.
GONZÁLEZ, E. J., M. ORTAZ, C. PEÑA-HERRERA & E. MONTES. 1997. Estudio con-trol de algas en los embalses La Pereza, QuebradaSeca, La Mariposa, Lagartijo y Camataguamediante técnicas de biomanipulación. Informetécnico presentado a las compañías HidroimpactoC.A. e Hidrocapital.
GONZÁLEZ, E. J. & M. ORTAZ. 1998. Efectos delenriquecimiento con N y P sobre la comunidad delfitoplancton en microcosmos de un embalse tropi-cal (La Mariposa, Venezuela). Rev. Biol. Trop., 46:27-34.
GONZÁLEZ, E. J., M. ORTAZ & C.PEÑAHERRERA. 2000. Caracterización limno-lógica del embalse Pao-Cachinche (EstadosCarabobo y Cojedes). Informe técnico presentadoa las compañías Hidroven e Hidrocentro.
GONZÁLEZ, E. J. 2000. Caracterización limnológi-ca del embalse El Pueblito (Estado Guárico,Venezuela). Trabajo de ascenso a la categoría deProfesor Agregado. Universidad Central deVenezuela. Caracas, 40 pp.
HARPER, D. 1992. Eutrophication of freshwaters.Principles, problems and restoration. Chapman &Hall. London, 327 pp.
INFANTE, A. 1997. Primary production of phyto-plankton in lake Valencia (Venezuela). Int. Revueges. Hydrobiol., 82: 469-477.
INFANTE, A., O. INFANTE & T. VEGAS. 1992.Caracterización limnológica de los embalsesCamatagua, Guanapito y Lagartijo, Venezuela.Proyecto Multinacional de Medio Ambiente yRecursos Naturales. Organización de los EstadosAmericanos y Universidad Central de Venezuela,Caracas.
INFANTE, A., O. INFANTE. & E. GONZÁLEZ.1995. Caracterización limnológica de los embalsesEl Cují y El Andino, Venezuela. ProyectoMultinacional de Medio Ambiente y RecursosNaturales. Organización de los Estados Americanosy Universidad Central de Venezuela, Caracas.
NUSCH, E. A. & G. PALME. 1975. BiologischeMethoden für der Praxis der Gewässer-untersuchung, Bestimmung des Chlorophyll-a undPhaeopigment-gehaltes in Oberflachenwässer.GWF-Wässer/Abwässer, 116: 562-565.
ORTAZ, M., E. J. GONZÁLEZ, J. MANDUCA, C.PEÑAHERRERA & E. MONTES. 1999.Características limnológicas de los embalses LaPereza, Lagartijo, La Mariposa y Quebrada Secay preservación del zooplancton como controladorde microalgas, mediante técnicas de biomanipula-ción. Informe técnico presentado a las compañíasHidroimpacto C.A. e Hidrocapital.
RYDING, S. O. & W. RAST. 1992. El control de laeutroficación en lagos y pantanos. EdicionesPirámide S.A. Unesco. Madrid, 385 pp.
SALAS, H. & P. MARTINÓ. 1991. A simplifiedphosphorus trophic model state for warm-watertropical lakes. Wat. Res., 25: 341-350.
STRASKRABA, M. 1999. Retention time as a keyvariable of reservoir limnology. En: Theoreticalreservoir ecology and its applications. J. G.Tundisi & M. Straskraba (eds.): 385-410.International Institute of Ecology, BrazilianAcademy of Sciences & Blackhuys Publishers.São Carlos, Rio de Janeiro & Leiden.
TUNDISI, J. G. & T. MATSUMURA-TUNDISI. 1990.Limnology and eutrophication of Barra Bonitareservoir, S. Paulo State, Southern Brazil. Arch.Hydrobiol. Beih. Ergebn. Limnol., 33: 661-676.
WETZEL, R. G. & G. E. LIKENS. 1991. Limnologicalanalyses. 2nd edition. Springer-Verlag, New York,391 p.
Fitoplancton de cinco embalses venezolanos 35


INTRODUCTION
The change in nitrogen and phosphorus loa-dings to lakes led to changes in the biogeoche-mical cycles of these nutrients in lakes.However, the cycling of N is expected to bemore affected by external loadings due to theopen characteristic of the N cycle.
That increases in nutrient loadings to lakesimplied the augmentation of in-lake nutrientconcentrations, and the subsequent enhance-ment of lake biological standing stocks, is now
widely recognized (Sakamoto, 1966; Dillonand Rigler, 1974; Schindler et al., 1978;Hanson and Leggett, 1982; Hanson and Peters,1984; Bird and Kalff, 1984; Yan, 1986;Quiros, 1990; Brown et al., 2000; amongmany others). Increased levels of living anddead organic matter in eutrophic and hypertro-phic lakes when compared with oligotrophiclakes permit us to predict an augmentation ofreductive characteristics of the lake environ-ment with the increase in lake trophic status(Stumm and Morgan, 1996).
The relationship between nitrate and ammonia concentrations in the pelagic zone of lakes
Rolando Quirós
Area de Sistemas de Producción Acuática, Facultad de Agronomía, Universidad de Buenos Aires. Av. SanMartín 4453, (1417) Buenos Aires, Argentina. E-mail: [email protected]
ABSTRACT
Both nitrate (NO3-N) and ammonia (NH4-N) concentrations are in general highly variable during a lake’s seasonal cycle.During the growing seasonal peak, such concentrations can be very low due mainly to its’ sequestration by phytoplankton.With the aim to study NO3-N and NH4-N mean concentration variation along the increase of the trophic state of lakes, thisstudy simultaneously analyses the information for more than 400 lakes worldwide. Both forms of nitrogen are directly rela-ted to the trophic state of a lake when lakes are compared with total phosphorous concentration (TP) ranging across fivemagnitude levels. However, in lakes a pronounced non-linearity dominates the relationships among the different forms ofnitrogen and phosphorous. Our results suggest that the empirical basis for both the limiting resource and inorganic nitrogenhypotheses are related. The NO3-N: NH4-N ratio is directly related to the TN: TP ratio for lakes in general and shallow onesin particular.
Key words: lakes, nutrients, ammonia, nitrate, lake comparisons, trophic state
RESUMEN
Las concentraciones de nitrato (NO3-N) y de amonio (NH4-N) generalmente son altamente variables durante el ciclo esta-cional de un lago. Durante el pico de la estación de crecimiento tales concentraciones pueden llegar a ser muy bajas, debi-do principalmente a su captación por parte del fitoplancton. Con el objeto de estudiar la variación de las concentracionesmedias de NO3-N y de NH4-N con el aumento del estado trófico de los lagos, fue analizada información simultánea paramás de 400 lagos distribuidos mundialmente. Ambas formas de nitrógeno están directamente relacionadas con el estadotrófico de un lago, cuando son comparados lagos con concentraciones de fósforo total (TP) que abarcan cinco órdenes demagnitud. Sin embargo, una pronunciada no-linealidad domina las relaciones entre las diversas formas de nitrógeno y fós-foro en lagos. Nuestros resultados sugieren que la base empírica de las hipótesis del nutriente limitante y del nitrógenoinorgánico están relacionadas. La relación NO3-N:NH4-N esta directamente vinculada con la relación TN:TP para loslagos en general y para los poco profundos en particular.
Palabras clave: lagos, nitrógeno disuelto, amonio, nitratos, estado trófico
37
Limnetica 22(1-2): 37-50 (2003)© Asociación Española de Limnología, Madrid. Spain. ISSN: 0213-8409

Both nitrate (NO3-N) and ammonia (NH4-N)concentrations are highly variable during lakeseasonal cycles. For deep stratified lakes nitrateis higher during mixing events and usuallydecreses in late summer and fall. For the tropho-genic zone of shallow lakes, both concentrationswould be lower during periods of water columnstability and they will increase during verticalmixing events. NH4-N is generated by hetero-trophic bacteria as the primary nitrogenous endproduct of decomposition of organic matter, andis readily assimilated by plants in the trophoge-nic zone (see Wetzel, 2001). NH4-N concentra-tions are usually low in oxygenated waters ofoligo- to mesotrophic deep lakes because of uti-lization by plants in the photic zone and nitrifi-cation to N oxidized forms. At relatively lowdissolved oxygen, nitrification of ammonia cea-ses, the absorptive capacity of the sediments isreduced, and a marked increase of the release ofNH4-N from the sediments then occurs. As aresult, the NH4-N concentration would increasefor the water column of shallow lakes and ofdeep lakes during mixing processes. NO3-N isthe common form of inorganic nitrogen enteringlakes from the drainage basin; in relatively aero-bic waters nitrification prevails (see Wetzel,2001). When NO3-N from external sources rea-ches lakes it is taken by autotrophs and bacteria,transformed in organic matter that on decay andfood web transmission ultimately goes to theNH4-N pool. Therefore, NH4-N accumulatesrespect to NO3-N in relatively standing waterslike those of lakes and reservoirs. In eutrophicand hypertrophic lakes, a large lake concentra-tion of organic matter implies a large potentialpool of ammonia. In more reductive media deni-trif ication and nitrate reduction to ammoniawould prevail (Stumm and Morgan, 1996). Theremoval of N from lakes is usually dominatedby NO3-N denitrification concomitantly withthe oxidation of organic matter, but in highlyproductive surface waters, high pH would favorN release to the atmosphere as NH3.
Therefore, and due mainly to in-lake Ncycling, it has to be expected that the ratio ofNO3-N to NH4-N (NO3-N:NH4-N) in lakes
would be less variable in relation to natural andpollutional sources of both forms of nitrogenthat in direct connection to total lake biomassand other lake characteristics like morphometry,water alkalinity and pH, and dissolved oxygenregime. For lake steady states, the supply ofnutrients to lake surface waters includes bothexternal inputs and internal inputs such as relea-ses from sediments, transport from deep-waterlayers, and, in the case of nitrogen, from N2fixation. Following P and N loadings, the prin-cipal destiny of both nutrients usually is incor-poration into lake biota followed by sedimenta-tion. The principal sinks for N usually are thesame as for P, the uptake by the biota followedby sedimentation. However, a lake could controlTN concentration in its waters by denitrifica-tion, NH4-N releases from sediments and N2fixation. All these mechanisms would contribu-te to determine both the TN:TP and the ratioNO3-N:NH4-N for lake surface waters.
A large number of theories have been propo-sed to explain the success of pelagic cyanobac-teria in lakes. These hypotheses embrace manyspatial and temporal scales (see Hyenstrand etal. (1998) for a review), and the most generaltheories are environmentally connected and it isprobable that their empirical bases would berelated (Reynolds and Petersen, 2000). One ofmost recognized theories, the low TN:TP hypo-thesis (Schindler, 1977; Smith, 1982; 1983;Bulgakov and Levich, 1999), predicts that cya-nobacteria will dominate lakes with a lowTN:TP ratio. However, lakes with low TN:TPratios usually also have high TP concentrations(Downing and McCauley, 1992; Quiros, 2002).With basis in their physiological requirements,it has been proposed that non-N2-fixing cyano-bacteria would be disfavored in nitrate rich(McQueen and Lean, 1987) oxidizing environ-ments and favored in ammonia rich (Blomqvistet al., 1994) more reductive environments. TheTN:TP hypothesis has been seriously challen-ged by Reynolds (1998) argumentation aboutselective process in the phytoplankton, and theinorganic N hypothesis had to be modified toinclude iron effects (Hyenstrand et al.,1999).
38 R. Quirós

In this paper, the relationship between the empiri-cal bases of the limiting resource theory (Tilmanet al., 1982; Smith, 1983) and the inorganic nitro-gen hypothesis (Blomqvist et al., 1994) is studiedin among lake comparisons. More specifically,the relationship between the TN:TP and the NO3-N:NH4-N ratios is explored.
MATERIALS AND METHODS
With the purpose of exploring trophic stateeffects on nitrate and ammonia concentrationsfor the pelagic zone of lakes a variety of publi-shed and unpublished data was used, comprisingmeasurements of total phosphorus (TP), totalnitrogen (TN), total organic nitrogen (TON),dissolved inorganic N, and mean depth for 425world lakes and ponds ranging from ultraoligo-trophic to hypertrophic.
Depending on available information, the data-base includes data for: a) Northwest CanadianTerritories (Pienitz et al., 1997a; 1997b), Quebec(D`Arcy and Carignan, 1997), north-temperatelakes in Wisconsin (LTER data, Bowser et al.,1999), and northeastern USA lakes (EMAP data,Larsen et al., 1991); b) world lake data forEurope (ILEC, 1994), Asia and Oceania (ILEC,1993), and the Americas (ILEC, 1995); c)Mississippi alluvial valley lakes (Wylie andJones, 1986); d) Southamerican reservoirs(Tundisi et al., 1991) and Brazilian lakes(Mitamura et al., 1997); d) Andean-Patagonianlakes (Pedrozo et al., 1993; Diaz andPedrozo,1996); e) fish ponds in Thailand, Kenya,and Honduras (PD/A CRSP data); f) water qua-lity data for USA lakes and reservoirs (Nebraska(STORET, 2001) and USGS (USGS, 2001); g)others lakes and reservoirs distributed throughtemperate and tropical regions (Quiros files).Morphometric and nutrient data were used aspublished and each water body is representedonce in data. Most nutrient data are mean annualvalues but for some lakes, available data for justone year were included.
Several data subsets were used for nutrientanalyses. The total world lake and pond set
(total lake data, TOTAL) comprised nutrientdata for 425 lakes, reservoirs, and ponds distri-buted in both temperate and tropical regions,independently of their trophic conditions, mor-phometry, and water characteristics. A subset ofthe TOTAL (n = 425), is comprised by lakesand reservoirs with available morphometricdata (ZLAKES, n = 191). For the latter, twolake subsets for deep (DEEP, mean depth >10m, n = 42) and shallow (SHALLOW, meandepth < 10m, n = 149) lakes were also contem-plated. To compare lake depth effects betweendeep and shallow lakes, a SHALLOW65 lakesubset (n = 80) was defined as a part of theSHALLOW subset but only including lakeswith TP concentrations ranging the same rangeof TP concentrations than deep lakes (< 65mg·m-3). In order to stabilize the variance forcorrelation and regression analysis, all thevariables were log-transformed. Curvilineartrends in data were studied using robust locallyweighted regression and smoothing graphictechniques (LOWESS). The Number CruncherStatistical System (NCSS 2000) (Hintze, 1998)was used. Throughout this paper, the TP con-centration will be considered as the main indi-cator of lake trophic state.
RESULTS
Lake data
Total lake data represents better eutrophic andhypertrophic lakes, and TP concentrations rangedover five orders of magnitude (Table 1). However,TP ranges were narrow when deep and shallowlakes were considered separately (Table 1).
As a result of lake trophic state augmentation,all lake nutrient forms increase their concentra-tions, as expected. Mean NO3-N and NH4-Nconcentrations in surface waters increase 7 andmore than 20 times, respectively, from oligotro-phic and mesotrophic to hypertrophic lakes(Table 1). Correlation analysis showed thatlakes with the highest total nutrient levels ten-ded to have higher NO3-N and NH4-N concen-
Nitrate and ammonia in lakes 39
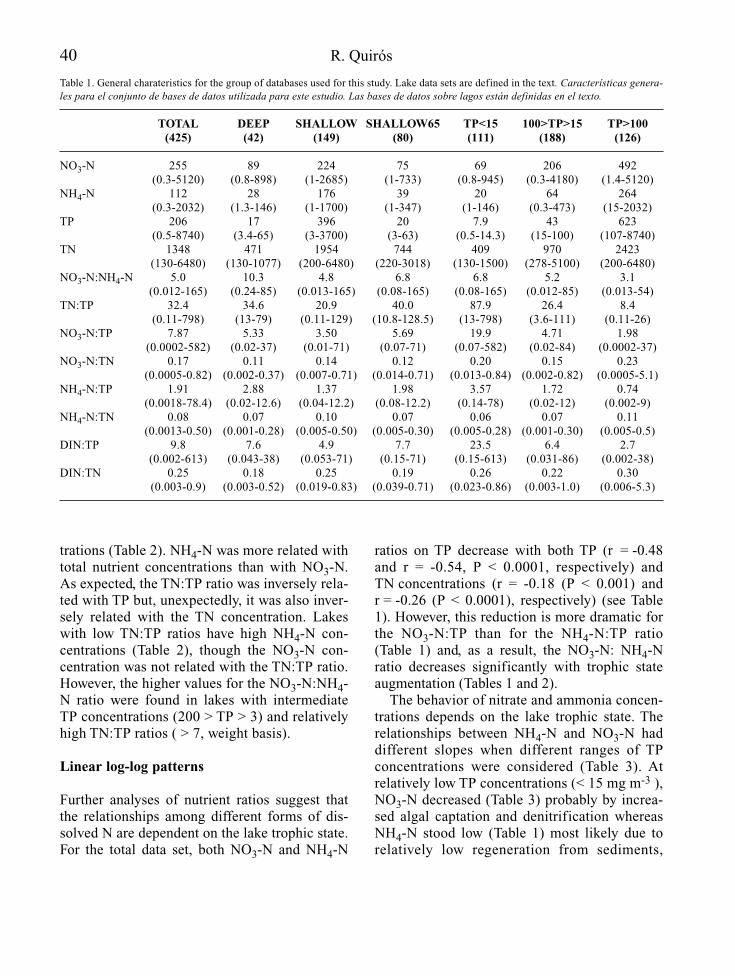
trations (Table 2). NH4-N was more related withtotal nutrient concentrations than with NO3-N.As expected, the TN:TP ratio was inversely rela-ted with TP but, unexpectedly, it was also inver-sely related with the TN concentration. Lakeswith low TN:TP ratios have high NH4-N con-centrations (Table 2), though the NO3-N con-centration was not related with the TN:TP ratio.However, the higher values for the NO3-N:NH4-N ratio were found in lakes with intermediateTP concentrations (200 > TP > 3) and relativelyhigh TN:TP ratios ( > 7, weight basis).
Linear log-log patterns
Further analyses of nutrient ratios suggest thatthe relationships among different forms of dis-solved N are dependent on the lake trophic state.For the total data set, both NO3-N and NH4-N
ratios on TP decrease with both TP (r = -0.48and r = -0.54, P < 0.0001, respectively) andTN concentrations (r = -0.18 (P < 0.001) andr = -0.26 (P < 0.0001), respectively) (see Table1). However, this reduction is more dramatic forthe NO3-N:TP than for the NH4-N:TP ratio(Table 1) and, as a result, the NO3-N: NH4-Nratio decreases significantly with trophic stateaugmentation (Tables 1 and 2).
The behavior of nitrate and ammonia concen-trations depends on the lake trophic state. Therelationships between NH4-N and NO3-N haddifferent slopes when different ranges of TPconcentrations were considered (Table 3). Atrelatively low TP concentrations (< 15 mg m-3 ),NO3-N decreased (Table 3) probably by increa-sed algal captation and denitrification whereasNH4-N stood low (Table 1) most likely due torelatively low regeneration from sediments,
40 R. Quirós
Table 1. General charateristics for the group of databases used for this study. Lake data sets are defined in the text. Características genera-les para el conjunto de bases de datos utilizada para este estudio. Las bases de datos sobre lagos están definidas en el texto.
TOTAL DEEP SHALLOW SHALLOW65 TP<15 100>TP>15 TP>100(425) (42) (149) (80) (111) (188) (126)
NO3-N 255 89 224 75 69 206 492(0.3-5120) (0.8-898) (1-2685) (1-733) (0.8-945) (0.3-4180) (1.4-5120)
NH4-N 112 28 176 39 20 64 264(0.3-2032) (1.3-146) (1-1700) (1-347) (1-146) (0.3-473) (15-2032)
TP 206 17 396 20 7.9 43 623(0.5-8740) (3.4-65) (3-3700) (3-63) (0.5-14.3) (15-100) (107-8740)
TN 1348 471 1954 744 409 970 2423(130-6480) (130-1077) (200-6480) (220-3018) (130-1500) (278-5100) (200-6480)
NO3-N:NH4-N 5.0 10.3 4.8 6.8 6.8 5.2 3.1(0.012-165) (0.24-85) (0.013-165) (0.08-165) (0.08-165) (0.012-85) (0.013-54)
TN:TP 32.4 34.6 20.9 40.0 87.9 26.4 8.4(0.11-798) (13-79) (0.11-129) (10.8-128.5) (13-798) (3.6-111) (0.11-26)
NO3-N:TP 7.87 5.33 3.50 5.69 19.9 4.71 1.98(0.0002-582) (0.02-37) (0.01-71) (0.07-71) (0.07-582) (0.02-84) (0.0002-37)
NO3-N:TN 0.17 0.11 0.14 0.12 0.20 0.15 0.23(0.0005-0.82) (0.002-0.37) (0.007-0.71) (0.014-0.71) (0.013-0.84) (0.002-0.82) (0.0005-5.1)
NH4-N:TP 1.91 2.88 1.37 1.98 3.57 1.72 0.74(0.0018-78.4) (0.02-12.6) (0.04-12.2) (0.08-12.2) (0.14-78) (0.02-12) (0.002-9)
NH4-N:TN 0.08 0.07 0.10 0.07 0.06 0.07 0.11(0.0013-0.50) (0.001-0.28) (0.005-0.50) (0.005-0.30) (0.005-0.28) (0.001-0.30) (0.005-0.5)
DIN:TP 9.8 7.6 4.9 7.7 23.5 6.4 2.7(0.002-613) (0.043-38) (0.053-71) (0.15-71) (0.15-613) (0.031-86) (0.002-38)
DIN:TN 0.25 0.18 0.25 0.19 0.26 0.22 0.30(0.003-0.9) (0.003-0.52) (0.019-0.83) (0.039-0.71) (0.023-0.86) (0.003-1.0) (0.006-5.3)
algal uptake, and higher nitrification to moreoxidized N forms. At intermediate TP (100 > TP> 10-15), both NO3-N and NH4-N concentra-tions were directly related with TP (Table 3). Anincreased NH4-N regeneration from sedimentsand a relative acceleration of nitrification can besupposed for this range of TP concentrations.N2- fixation usually is an important N compen-sation mechanism at this intermediate TP levels(Levine and Schindler, 1999). For high TP con-centrations (> 100 mg m-3 ), NO3-N did notincrease or its increse was negligible. For thisrange of TP, NH4-N was directly related with TPbut with a lower slope for the NH4-N regressionon TP (Table 3). Nitrification appears to behighly reduced when compared with NH4-Nregeneration from sediments. The latter fact isultimately related to higher organic matter con-
centration and living biomass for high TP lakes.In conclusion, this is also an expected resultbecause eutrophic and hypertrophic lakes aremore reductive environments when comparedwith oligotrophic and mesotrophic lakes(Stumm and Morgan, 1996).
Our described pattern for NO3-N and NH4-Nvariability in lakes presented some differenceswhen deep and shallow lakes have been analy-zed separately (Table 4). Shallow lakes presen-ted higher level of nutrients when comparedwith deep lakes. Most of those deviations maybe due to differences in lake trophic state(Table 1, compare DEEP and SHALLOW data).However, some differences between shallowand deep lakes were apparent when lakes withequivalent TP variability had been compared(Table 1, compare DEEP and SHALLOW65data). Both NH4-N and TN concentrations werehigher for SHALLOW65 lakes, as well as theNO3-N:NH4-N ratio was lower for SHA-LLOW65 than for DEEP lakes. High data varia-bility precludes us to conclude about significantdifferences for means except those for TN andTON (t-test, P < 0.01 and 0.02, respectively).When lakes were compared using multipleregression models including a “dummy” varia-ble for lake depth, TN, TON, and TN:TP ratioswere higher for shallow than for deep lakes(P < 0.05). On the other hand, NO3-N:NH4-Nratios were lower for SHALLOW65 lakes(P < 0.05) when TP concentration was heldconstant. The huge increase of NO3-N at inter-
Nitrate and ammonia in lakes 41
Table 2. Matrix correlation coefficients among nutrients, andsome ratios for whole lake data set (TOTAL). Variables as theyhave been defined in the text. Matriz de coeficientes de correla-ción entre los nutrientes y algunos cocientes para todo el conjuntode la base de datos (TOTAL). Las variables según han sido defini-das en el texto.
TN TN:TP NO3-N NH4-N NO3:NH4
TP 0.75 -0.88 0.47 0.72 -0.20TN -0.35 0.57 0.72 -0.05 (a)TN:TP -0.19 (a) -0.50 0.29NO3 0.63 0.57NH4 -0.27
N = 425. (a); t-test, P > 0.00001
Table 3. Slope values for the log-log regression models between nutrient and total phosphorus (TP) for the whole data set (TOTAL) and forthree data subsets as defined by its’ variation range of variation. Standard error for slopes are indicated. Valores de las pendientes para losmodelos de regresión log-log entre nutrientes y el fósforo total (TP) para todo el conjunto de datos (TOTAL) y para tres subconjuntos defi-nidos por su rango de variación. Se indican los errores estándar de las pendientes.
n NH4-N NO3-N NO3-N:NH4-N TN NH4-N:TP NO3-N:TP TN:TP
TP < 15 111 0.225 (0.151) (a) -0.710 (0.228) -0.947 (0.200) 0.102 (0.073) (a) -0.775 (151) -1.710 (0.223) -0.904 (0.073)100>TP>15 188 0.705 (0.131) 1.024 (0.184) 0.307 (0.186) (a) 0.479 (0.063) -0.295 (0.131) 0.024 (0.184) (a) -0.521 (0.064)TP>100 126 0.486 (0.079) -0.047 (0.118) (a) -0.628 (0.106) 0.204 (0.057) -0.514 (0.078) -1.047 (0.118) -0.787 (0.056)TOTAL 425 0.620 (0.029) 0.490 (0.045) -0.171 (0.041) 0.387 (0.018) -0.380 (0.029) -0.510 (0.045) -0.620 (0.018)
(a), P > 0.05
mediate TP concentrations appears to bemore related with deep than with shallow lakes(Table 4). More than half of lakes included inthe DEEP subset (55 %) were lakes withTP < 15 mg.m-3. On the other hand, the dissol-ved N increases for the SHALLOW65 subset atlow and intermediate TP levels was related withNH4-N (Table 4). That TN:TP ratios are higherfor shallow than for deep lakes when TP is heldconstant has been shown before for a larger setof world lakes (Quirós, 2002).
Non-linear log-log patterns
As it has been shown above, NO3-N and NH4-Nconcentrations were both positively log-logcorrelated with TP (Table 2, Fig. 1). However,those results also suggest that the relationshipsbetween the dissolved forms of N and the laketrophic state are more complex than just log-loglinear relationships. A close inspection of figure1 shows that NO3-N did not increase when TPwas very high (> 100 mg m-3) or even decreaseswhen it was low (< 8-10 mg m-3) (Table 3). Incontrast, NO3-N showed a sharp increase whenTP was between 15 and 100 mg m-3. This range
of TP is in close coincidence with the TP rangewhere lake TN increases sharply with TP (Fig. 2).For this latter TP range an acceleration of lakenitrification processes fueled by higher organicmatter contents and sustained by enough dissol-ved oxygen, can be suspected. Moreover, a noti-ceable change of curvature was displayed for TPbetween 20 and 30 mg m-3 (Fig. 1). From this TPrange upward NO3-N diverged from NH4-N up toTP near 100 mg m-3. It has to be stressed herethat a TP concentration of 25 mg m-3 has beenrepeatedly mentioned in literature as a lower limitfor Microcystis plankton dominance (seeReynolds et al., 2000). The NO3-N:TP ratiodeclined for all the range of TP concentrationswith exception of TP ranging between 15 and100 mg m-3 (Table 3, Fig. 3a).
Contrasting with the behavior of NO3-N,NH4-N did not present huge changes of slopeunder TP augmentation. It increased at a lowrate for TP below 8-10 mg m-3, its rate of incre-ase was maximum for TP between 15 and 100mg m-3, and increased at a declining rate for TPhigher than 100 mg m-3 (Table 2, Fig. 1). Thisfact may indicate that NH4-N returns from sedi-ments increased with lake trophic state (Tables1 and 2) but also that those returns, per unit of
42 R. Quirós
Figure 2. Relationship between total nitrogen (TN) and totalphosphorus (TP). The relationship after a LOWESS smoo-thing is indicated. Relación entre el nitrógeno total (TN) y elfósforo total (TP). Se indica la relación con suavizadoLOWESS.
Figure 1. Relationships between nitrate (NO3-N), ammonia(NH4-N), and total phosphorus (TP), for all lakes and ponds(TOTAL data, n = 425). LOWESS smoothing relationshipsand the TP line are indicated. Relaciones entre nitrato (NO3-N) , amonio (NH4-N) y fósforo total (TP), para todos los lagosy lagunas (TOTAL data, n=425). Se indican las relacionessegún un suavizado LOWESS y la linea TP.
TP, declines from oligotrophic to hypertrophiclakes (Fig. 2). One stricking result is that NH4-N:TP ratio decline was a minimum for lakeswith TP between 10 and 100 mg m-3 (Fig. 3a).For this latter TP range, NH4-N returns fromsediments must be higher than losses producedby nitrif ication to NO3-N. Anyway, NH4-Naccumulates continuously in the water columnduring lake eutrophication.
The proportions of NO3-N and NH4-N in totalnitrogen both change with lake trophic state.NO3-N and NH4-N accounted for 30-35 % and2 % of the N in the water, respectively, for oligo-trophic lakes, and about 10% and 20%, respecti-vely, for high TP lakes (Fig. 3b). The dissolvedinorganic N had a minimum at low TP concentra-tions (TP = 5-6 mg m-3) (Fig. 1), but for TP ofabout 20 mg m-3 the proportion of NO3-N in TNwas minimum (Fig. 3b). For total lake data, NO3-N:TN was not log-log related to TP. However, ithad its greater values for low TP lakes and decre-ased abruptly up to TP near 20 mg m-3. From thisTP concentration upwards it decreased steadily(Fig. 3b). On the other hand, NH4-N:TN increa-sed gently from its smaller values for low TPlakes to its higher values for high TP lakes. Othersurprising result was that the LOWESS line slopefor the relationship between NH4-N:TN and TPdeclined considerably for TP concentrations bet-ween 20 and 50-60 mg m-3. A close inspection ofFigure 3b reinforce that NO3-N was the mostimportant form of dissolved inorganic N for oli-gotrophic lakes whereas NH4-N was critical forhypertrophic lakes with very high TP concentra-tions (> 700-800 mg m-3).
The behaviors of NH4-N and TN concentra-tions under TP augmentation were similar (Fig.3a, and compare Figures 1 and 2). The changeof slopes for the TN:TP ratio was close to thechanges displayed for the NH4-N:TP ratio whendifferent ranges of TP concentrations were con-sidered (Table 3).
Relationship between TP-TN and NO3-N:NH4-N ratios
Lakes with high TP concentrations and lowTN:TP ratios also tend to have low NO3-N:NH4-N ratios (Table 2). For total lake data, bothTN:TP and NO3-N:NH4-N ratios were inverselyrelated to TP (r = -0.88 (P < 10-6) and r = -0.20(P < 10-4)). Both ratios were significantly corre-lated between them (r = 0.29, P < 10-6).
For the low range of TP (< 10-15 mg m-3), theslope for the TP-TN relationship was the smallestwhen compared with upper ranges of TP (Fig. 2,
Nitrate and ammonia in lakes 43
Figure 3. a) Relationships between NO3-N:TP, NH4-N:TP, andTN:TP ratios and total phosphorus concentration for (TP) forthe whole data set; b) Relationships between NO3-N:TN, andNH4-N:TN, and total phosphorus concentration (TP) for thewhole data set; the constant total nitrogen (TN) lines andthe LOWESS smoothing relationships are indicated. a) Relaciones entre los cocientes NO3-N:TP, NH4-N:TP, yTN:TP y el concentración de fósforo total (TP) para todo elconjunto de datos; b) Relaciones entre NO3-N:TN, y NH4-N:TN, y la concentración de fósforo total (TP) para todo elconjunto de datos; se indican las líneas de la relación cons-tante para el nitrógeno total (TN) y para las relaciones consuavizados LOWESS.
a
b
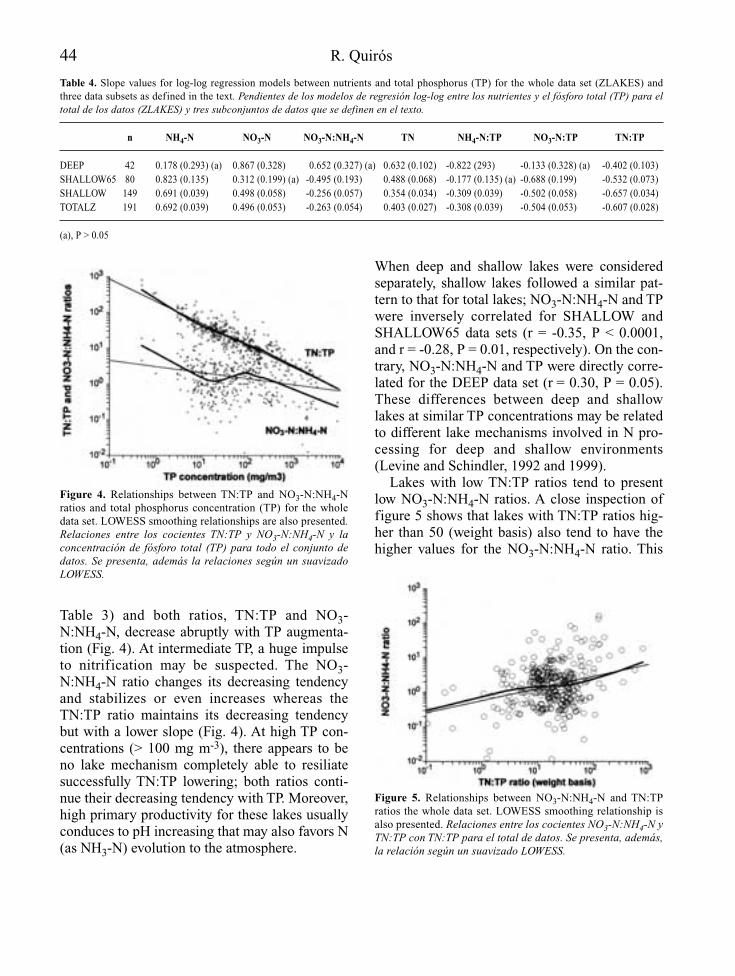
Table 3) and both ratios, TN:TP and NO3-N:NH4-N, decrease abruptly with TP augmenta-tion (Fig. 4). At intermediate TP, a huge impulseto nitrification may be suspected. The NO3-N:NH4-N ratio changes its decreasing tendencyand stabilizes or even increases whereas theTN:TP ratio maintains its decreasing tendencybut with a lower slope (Fig. 4). At high TP con-centrations (> 100 mg m-3), there appears to beno lake mechanism completely able to resiliatesuccessfully TN:TP lowering; both ratios conti-nue their decreasing tendency with TP. Moreover,high primary productivity for these lakes usuallyconduces to pH increasing that may also favors N(as NH3-N) evolution to the atmosphere.
When deep and shallow lakes were consideredseparately, shallow lakes followed a similar pat-tern to that for total lakes; NO3-N:NH4-N and TPwere inversely correlated for SHALLOW andSHALLOW65 data sets (r = -0.35, P < 0.0001,and r = -0.28, P = 0.01, respectively). On the con-trary, NO3-N:NH4-N and TP were directly corre-lated for the DEEP data set (r = 0.30, P = 0.05).These differences between deep and shallowlakes at similar TP concentrations may be relatedto different lake mechanisms involved in N pro-cessing for deep and shallow environments(Levine and Schindler, 1992 and 1999).
Lakes with low TN:TP ratios tend to presentlow NO3-N:NH4-N ratios. A close inspection offigure 5 shows that lakes with TN:TP ratios hig-her than 50 (weight basis) also tend to have thehigher values for the NO3-N:NH4-N ratio. This
44 R. Quirós
Figure 4. Relationships between TN:TP and NO3-N:NH4-Nratios and total phosphorus concentration (TP) for the wholedata set. LOWESS smoothing relationships are also presented.Relaciones entre los cocientes TN:TP y NO3-N:NH4-N y laconcentración de fósforo total (TP) para todo el conjunto dedatos. Se presenta, además la relaciones según un suavizadoLOWESS.
Figure 5. Relationships between NO3-N:NH4-N and TN:TPratios the whole data set. LOWESS smoothing relationship isalso presented. Relaciones entre los cocientes NO3-N:NH4-N yTN:TP con TN:TP para el total de datos. Se presenta, además,la relación según un suavizado LOWESS.
Table 4. Slope values for log-log regression models between nutrients and total phosphorus (TP) for the whole data set (ZLAKES) andthree data subsets as defined in the text. Pendientes de los modelos de regresión log-log entre los nutrientes y el fósforo total (TP) para eltotal de los datos (ZLAKES) y tres subconjuntos de datos que se definen en el texto.
n NH4-N NO3-N NO3-N:NH4-N TN NH4-N:TP NO3-N:TP TN:TP
DEEP 42 0.178 (0.293) (a) 0.867 (0.328) 0.652 (0.327) (a) 0.632 (0.102) -0.822 (293) -0.133 (0.328) (a) -0.402 (0.103)SHALLOW65 80 0.823 (0.135) 0.312 (0.199) (a) -0.495 (0.193) 0.488 (0.068) -0.177 (0.135) (a) -0.688 (0.199) -0.532 (0.073)SHALLOW 149 0.691 (0.039) 0.498 (0.058) -0.256 (0.057) 0.354 (0.034) -0.309 (0.039) -0.502 (0.058) -0.657 (0.034)TOTALZ 191 0.692 (0.039) 0.496 (0.053) -0.263 (0.054) 0.403 (0.027) -0.308 (0.039) -0.504 (0.053) -0.607 (0.028)
(a), P > 0.05

ratio have a tendency to be stabilized for TN:TPratios between 50 and 10 and decreases forTN:TP ratios lesser than 8-7. A change of curva-ture for the fitting line is noticeable when TN:TPequals 25 (Fig. 5). However, when the relation-ships between NO3-N:NH4-N and TN:TP havebeen analyzed for deep and shallow lakes separa-tely there were not noticeable differences in bet-ween. NO3-N:NH4-N and TN:TP were directlyrelated for both deep and shallow lakes though itwas not significant for deep lakes (r = 0.27,P = 0.14, and r = 0.44, P < 0.01, for DEEP andSHALLOW65 data sets, respectively).
DISCUSSION AND CONCLUSIONS
Our results strongly support that the concentra-tion of NH4-N and the proportion of NH4-N intotal nitrogen in lake surface waters rise steadilywith lake trophic state for lakes and shallowlakes. The analysis of our data for deep lakes didnot show that NH4-N increase for deep lakeswith TP concentrations lower than 65 mg m-3.However, our results also support that NH4-Nwill also increase for deep eutrophic and hyper-trophic lakes with very high TP concentrations.The behavior of NO3-N under lake eutrophica-tion is relatively more complex than NH4-Nbehavior. The concentrations of NO3-N in lakesurface waters increase clearly with lake trophicstate only for lakes with TP concentrations bet-ween 10 and 100 mg m-3, as well as for deep andshallow lakes, but it did not show a clear intensi-fication for shallow lakes with TP concentrationslower than 65 mg m-3. As well as for total nitro-gen, when the total trophic continuum is consi-dered, both forms of inorganic N decline per unitof TP, but that declination is reduced for TP con-centrations between 10-15 and 100 mg m-3.
The NO3-N:NH4-N ratio decreases with TP forlakes and shallow lakes, but it increased for deeplakes with TP concentrations lower than65 mg m-3 in our total lake data set. NH4-N wasgreater than NO3-N for TP concentrations from600-700 mg m-3 upwards (Fig. 1). When onlyshallow lakes were considered, it was higher than
NO3-N for TP concentrations from 70-90 mg m-3
upwards. These results are consistent with theview that shallow lakes compensate high TP loa-dings better than deep lakes, mainly throughNH4-N processing. Moreover, total nitrogen, aswell as total organic nitrogen, is higher for sha-llow than for deep lakes at equivalent concentra-tions of total phosphorus (Quiros, 2002; thispaper). However, both NO3-N:TP and NH4-N:TPratios decrease with lake trophic state for lakesand shallow lakes, but neither NO3-N:TP norNH4-N:TP decreased for deep and shallow lakes,respectively, when lakes have had TP concentra-tions lower than 65 mg m-3. This latter result sug-gests that NO3-N is a major form in N dynamicsfor surface waters of deep lakes whereas NH4-Nis prominent for shallow lakes, at low and inter-mediate TP concentrations (< 70-100 mg m-3).
Some similarities are apparent between thebehaviors of inorganic N dissolved forms displa-yed for low and very high TP lakes. The NO3-N:TP and NH4-N:TP ratios both decrease withTP for oligotrophic and hypertrophic lakes. Theproportion of NO3-N in total nitrogen decreaseswith TP for low and very high TP concentrations,whereas the proportion of NH4-N increases forboth oligotrophic and hypertrophic lakes. Anincrease of the reductive characteristics of theenvironment with the lake trophic state (Stummand Morgan, 1996) may explain both NH4-Nintensification and NO3-N reduction. It is to beexpected that both deep and shallow lakes willpresent a similar pattern for NO3-N reductionand NH4-N rise for high TP concentrations.However, a change for in-lake operating mecha-nisms is apparent for TP concentrations between8-15 and 100 mg m-3. This range of TP is coinci-dental with non-linear nitrogen and phytoplank-ton biomass increases showed for several datasets distributed worldwide. The lower part of thisrange of TP is where occur most of the changesin species composition for lake communities(e.g.: Colby et al., 1972; Leach et al., 1977;Quiros, 1998; Reynolds et al., 2000).
Our results support that there are not relati-vely simple log-log models to explain relations-hips between lake nutrient concentrations. Both
Nitrate and ammonia in lakes 45

NO3-N and NH4-N concentrations presentedcomplex non-linear log-log patterns when haverelated with TP. The multiple changes of slopefor log-log regressions may reflect the complexinterplay of internal mechanisms and theinfluences of external forces. The relationshipsin lakes are still more complex because all theanalyzed forms of N and the biomass for lakecommunities are really a potentially decreasingfunction of lake TP (Quiros, 2000). Curvilinearrelationships among total phosphorus, totalnitrogen, and phytoplankton biomass have beenshown for many lake data sets (Straskraba,1976; Prairie et al., 1989), but we still have arudimentary understanding of the factors thatdetermine TP-TN and TP-Chl sigmoid patterns.McCauley et al. (1989) called for an elucidationof the underlying biological mechanisms thatproduce the nonlinear response with TP.Furthermore, Watson et al. (1992) suggestedthat non-linearity reflects an underlying syste-matic variation in the biomass of functionalalgal groups. Mazumder (1994) showed that sig-moid patterns appear to be related to the transi-tion from stratified to mixed lakes along the TPgradient, and Quiros (1998) have related thosepatterns to the transition from lakes with balan-ced fish populations to planktivore dominatedlakes. Our earlier results (Quiros, 2002) suggestthat previous explanations for nonlinear patternsmight be included in a systemic view for Nvariability under P loading and its differencesbetween deep and shallow lakes. It has been pro-posed that underlying mechanisms for observednonlinear patterns are hidden behind differencesin the N biogeochemical cycling for lakes withdifferent trophic states and that an impulse tolake nitrification process may be suspected forintermediate lake trophic states (Quiros, 2002).
The physical-chemical characteristics of lakewater are influenced by external and in-lake pro-cesses. The complexity of these processes oftenlimits us to discriminate between internal andexternal causes for some in-lake process. As isusual in studying open systems with complexbehaviors, a mix of both internal and externalinfluences on in-lake processes can be identified.
It has been proposed two sources of change forthe TN:TP ratio change in lakes. The first, mostlyexternal (Downing and McCauley, 1992) hypo-thesis, proposes that the lake TN:TP ratio decrea-ses under nutrient enrichment is due related pri-marily with the N:P ratio for the nutrient loads. Asecond mostly internal theory (Schindler, 1977)considers that lakes compensate high P loadingsthroughout the most open N cycling. However,our results suggest that internal and externalinfluences may be not equally distributedthroughout the trophic continuum. The quantitiesof NO3-N and NH4-N from external sources arehighly diverse and variable because of markeddifferences in the inputs from surface andgroundwater sources, particularly as affected byhuman activities (see Wetzel, 2001). NH4-N isoxidized rapidly to nitrate in aerobic waters ofrivers and streams. However, in lakes, NO3-N israpidly reduced into organic compounds withinorganisms and liberated as ammonia during themetabolism of these organisms. External influen-ces would be most important at low and veryhigh TP concentrations but in-lake N processingis highly active in determining nutrient relation-ships for lakes at TP concentrations between 10-15 and 100 mg m-3. The more holistic external-internal view (Quiros, 2002) suggests that in-lakeprocesses always work against external changesbut they are particularly successful in opposingexternal influences only at intermediate lake tro-phic states. Experimental results support part ofour contentions; the external sources may domi-nate the N inputs to lake waters for high TN:TPlakes whereas waters of low TN:TP lakes couldreceive more than half of their N from internallake sources such as NH4-N from sediments orN2 fixation (Levine and Schindler, 1992).
The TN:TP and NO3-N:NH4-N ratios aredependent on lake trophic state but also on lakemorphometry. The NO3-N:NH4-N ratio is con-sistently very low for eutrophic and hypertro-phic lakes. However, it appears to be relativelylow also for ultraoligotrophic lakes (Fig. 4).Maximum values for NO3:NH4 were displayedat TP concentrations between 3 and 30 mg m-3.Anyway, our results suggest that the em-
46 R. Quirós

pirical basis for both the limiting resources(Tilman et al., 1982; Smith, 1982; Smith andBennett,1999) and the inorganic nitrogen hypo-theses (Blomqvist et al., 1994) are related. TheNO3-N:NH4-N ratio is directly related to theTN:TP ratio for lakes and shallow lakes. A simi-lar relationship was shown for deep lakes but itis something that needs to be investigated fur-ther. As was repeatedly suggested in literature(see Hyenstrand et al. (1998) for a review),NO3-N and NH4-N would not be equivalentforms of dissolved inorganic N for differentspecies composing the phytoplankton commu-nity. A high organic matter content in a high tro-phic state lake implies a more reductive envi-ronment where ammonia accumulates respect tonitrate (Fig. 3b). Therefore, it is to be expected,that different proportions of NO3-N and NH4-Nin total nitrogen or some other factors (e.g.some elements other than N or P) related withthe continuum that goes from high oxidizing tomore reductive environments throughout thetrophic continuum, may be reflected in phyto-plankton structure.
Our results suggest that leading mechanismsresponsible of N cycling in lakes would be dif-ferent for different lake trophic states and bet-ween deep and shallow lakes. The central role ofP in lake ecology was shown more than 30 yearsago (Schindler, 1977; Schindler et al., 1971),and most lakes distributed worldwide werephosphorus (P) restricted before cultural eutro-phication, independent of its original trophicstate. However, the role of N has been discussedfor many years in literature. The effect of inter-nal compensating processes is to increase ratherthan lower, the TN:TP ratio (Fig. 2). Internalmechanisms that can compensate low TN:TPnutrient loadings include: N2 fixation, sedimen-tation of N and P at a ratio different from theexternal supply ratio, differential P and N relea-ses from sediments, and denitrification. At thelake system level, these mechanisms would ope-rate differently for deep than for shallow lakes.It is not our intention here to hypothesize abouthidden mechanisms behind described patterns.We believe that an adequate mix of well-known
N processing mechanisms may explain quiteadequately our described behavior for inorganicdissolved forms. The high variability in our dataprecludes us to conclude that some of those dif-ferences are significantly.
Our obtained relationships will be only validfor stagnant surface waters. Therefore, for theNO3-N:NH4-N ratio, a variable dependence onthe water renewal time may be suspected.Flowing waters usually have higher dissolvedoxygen contents that modify N processingand conduct to higher NO3-N:NH4-N ratios forrunning waters when lakes and rivers werecompared at constant TP concentration (Quirós,unpublished results).
ACKNOWLEDGMENTS
R.Quirós acknowledges research support fromthe Consejo Nacional de InvestigacionesCientíficas y Tecnológicas (CONICET). Wethank Hugo T. von Bernard for technical assis-tance. This work was supported by AgenciaNacional de Promoción Científica y Técnicagrant PICT 04698.
REFERENCES
BIRD, D. F., & J. KALFF. 1984. Empirical relations-hips between bacterial abundance and chlorophyllconcentration in fresh and marine waters. Can. J.Fish. Aquat. Sci., 41: 1015-1023.
BLOMQVIST, P., A. PETTERSON & P. HYENS-TRAND. 1994. Ammonium-nitrogen: A key regu-latory factor causing dominance of non-nitrogen-fixing cyanobacteria in aquatic systems. Arch.Hydrobiol., 132: 141-164.
BOWSER, C. J., D.E. ARMSTRONG, J. J. MAGNU-SON & T. K. KRATZ. 1999. Nutrient Data ofNorth Temperate Lakes Primary Studies—TroutLake Area. LTER Program, USA. http://limno-sun.limnology.wisc.edu/catalog/html#chemical
BROWN, C. D., M. V. HOYER, R. W. BACHMANN,AND D. E. CANFIELD Jr. 2000. Nutrient-chlo-rophyll relationships: an evaluation of empiricalnutrient-chlorophyll models using Florida and
Nitrate and ammonia in lakes 47

north-temperate lake data. Can. J. Fish. Aquat.Sci., 57: 1574-1583.
BULGAKOV, N. G. & A. P. LEVICH. 1999. Thenitrogen to phosphorus ratio as a factor regulatingphytoplankton community structure. Arch.Hydrobiol., 146: 3-22.
COLBY, P. J., G. R. SPANGLER, D. A. HURLEY &A. M. MCCOMBIE. 1972. Effects of eutrophica-tion on salmonid communities in oligotrophiclakes. J. Fish. Res. Board Can., 29: 975-983.
D´ARCY, P., AND R. CARIGNAN. 1997. Influenceof catchment topography on water chemistry insoutheastern Quebec Shield lakes. Can. J. Fish.Aquat. Sci., 54: 2215-2227.
DIAZ, M. & F. PEDROZO. 1996. Nutrient limita-tions in Andean-Patagonian lakes at latitude40’41o S. Arch. Hydrobiol., 138: 123-143.
DILLON, P. J., & F. H. RIGLER. 1974. The phos-phorus-chlorophyll relationship in lakes. Limnol.Oceanogr., 19: 767-773.
DOWNING, J. A. & E. MCCAULEY. 1992. Thenitrogen:phosphorus relationship in lakes. Limnol.Oceanogr., 37: 936-945.
EMAP. Surface Waters Northeast Lakes 1991-94data. EPA, USA. http://www.epa.gov/emap/-html/datal/surfwatr/data/nelakes
HANSON, J. M., & W .C. LEGGETT. 1982.Empirical prediction of fish biomass and yield.Can. J. Fish. Aquat. Sci., 39: 257-263.
HANSON, J. M., & R. H. PETERS. 1984. Empiricalprediction of crustacean zooplankton biomass andprofundal macrobenthos biomass in lakes. Can. J.Fish. Aquat. Sci., 41: 439-445.
HINTZE, J. L., 1998. Number cruncher statisticalsystem (NCSS). Version 2000. Graphics. Dr. JerryL. Hintze. Kaysville, Utah, USA.
HYENSTRAND, P., P. BLOMQVIST & A. PETTER-SON. 1998. Factors determining cyanobacteria suc-cess in aquatic systems - a literature review. Arch.Hydrobiol. Spec. Issues Advanc. Limnol., 51: 41-62.
HYENSTRAND, P., E. RYDIN & D. M. GUNNER-HERD. 1999. Response of pelagic cyanobacteriato iron additions – enclosure experiments fromLake Erken. Journal of Plankton Research, 22:1113-1126.
ILEC, 1993. Data Book of World Lake Environments.1. Asia and Oceania. International LakeEnvironment Committee. United NationsEnvironment Programme. Kusatsu, Shiga, Japan.
ILEC, 1994. Data Book of World Lake Environments.2. Africa and Europe. International Lake
Environment Committee. United NationsEnvironment Programme. Kusatsu, Shiga, Japan.
ILEC, 1995. Data Book of World Lake Environments.3. The Americas. International Lake EnvironmentCommittee. United Nations EnvironmentProgramme. Kusatsu, Shiga, Japan.
LARSEN, D. P., D. L. STEVENS, A. R. SELLE & S.G. PAULSEN. 1991. Environmental monitoringand assessment program, EMAP-surface waters: anortheast lakes pilot. Lake and ReservoirManagement, 7: 1-11.
LEACH, J. H., M.G. JOHNSON, J. R. M. KELSO, J.HARTMANN, J. NUMANN & B. ENTZ. 1977.Responses of percid fishes and their habitats toeutrophication. J. Fish. Res. Board. Can., 34:1964-1971.
LEVINE, S. N. & D. W. SCHINDLER. 1992.Modification of the N:P ratio in lakes by in situprocesses. Limnol. Oceanogr., 37: 917-935.
LEVINE, S. N. & D. W. SCHINDLER. 1999.Influences of nitrogen to phosphorus supply ratiosand physicochemical conditions on cyanobacteriaand phytoplankton species composition in theExperimental Lakes Area, Canada. Can. J. Fish.Aquat. Sci., 56: 451-466.
MAZUMDER, A., 1994: Phosphorus - chlorophyllrelationships under contrasting herbivory andthermal stratification: predictions and patterns.Can. J. Fish. Aquat. Sci., 51: 390-400.
MCCAULEY, E., J. F. DOWNING & S. WATSON.1989. Sigmoid relationships between nutrientsand chlorophyll among lakes. Can. J. Fish. Aquat.Sci., 46: 1171-1175.
MCQUEEN, D. J. & D. R. S. LEAN. 1987. Influenceof water temperature and nitrogen to phosphorusratios on the dominance of blue-green algae inLake St. George, Ontario. Can. J. Fish. Aquat.Sci., 44: 598-604.
MITAMURA, O., Y. SAIJO, K. HINO & F.A.R.BARBOSA. 1997. The significance of regenera-ted nitrogen for phytoplankton productivity inlakes Dom Helvecio, Jacare and Carioca. In:Limnological Studies on the Rio Doce ValleysLakes, Brazil. J. S. Tundisi and Y. Saijo (eds.):141-153. Brazilian Academy of Sciences, Brazil.
PD/A CRSP. Pond Dynamics/Aquaculture CRSPdata base, Oregon State University, Corvallis,Oregon, USA.
PEDROZO, F., S. CHILLRUD, P. TEMPORETTI &M. DIAZ. 1993. Chemical composition andnutrient limitation in rivers and lakes of Northern
48 R. Quirós

Patagonian Andes (39.5o-42o S; 71o W) (Rep.Argentina). Verh. Internat. Verein. Limnol., 25:207-214.
PRAIRIE, Y. T., C. M. DUARTE & J. KALFF.1989. Unifying nutrient - chlorophyll relations-hips in lakes. Can. J. Fish. Aquat. Sci., 46: 1176-1182.
QUIROS, R., 1990b: Factors related to variance ofresiduals in chlorophyll - total phosphorus regres-sions in lakes and reservoirs of Argentina.Hydrobiologia, 200/201: 343-355.
QUIROS, R. 1998. Trophic cascade effects in a tem-perate - subtropical water body continuous. Verh.Internat. Verein. Limnol., 26: 2315-2319.
QUIROS, R. 2002. The nitrogen to phosphorus ratiofor lakes: A cause or a consequence of aquatic bio-logy? In: El agua en Iberoamérica: De la limnolo-gía a la gestión en Sudamérica. A. Fernández y G.Chalar (eds.): 11-26. Programa Iberoamericano deCiencia y Tecnología para el Desarrollo (CYTED),Buenos Aires, Argentina.
PIENITZ, R., J. P. SMOL & D. R. S. LEAN. 1996a.Physical and chemical limnology of 59lakes located between the southern Yukon andthe Tuktoyaktuk Peninsula, NorthwestTerritories (Canada). Can. J. Fish. Aquat. Sci.,54: 330-346.
PIENITZ, R., J. P. SMOL & D. R. S. LEAN. 1996b.Physical and chemical limnology of 24 lakes locatedbetween Yellowknife and Contwoyto Lake,Northwest Territories (Canada). Can. J. Fish. Aquat.Sci., 54: 347-358.
REYNOLDS, C. S. 1998. What factors influence thespecies composition of phytoplankton in lakes ofdifferent trophic status? In: Phytoplankton andTrophic Gradients. Hydrobiologia, 369/370: 11-26.
REYNOLDS, C. S. 1999. Non-determinism to probabi-lity, or N:P in the community ecology of phytoplank-ton. Arch. Hydrobiol., 146: 23-65.
REYNOLDS, C. S., S. N. REYNOLDS, I. F.MUNAWAR & M. MUNAWAR. 2000. The regu-lation of phytoplankton dynamics in the world`slargest lakes. Aquatic Ecosystem Health andManagement, 3: 1-21.
REYNOLDS, C. S. & A. C. PETERSEN. 2000. Thedistribution of planktonic Cyanobacteria in Irishlakes in relation to their trophic states.Hydrobiologia, 424: 91-99.
SAKAMOTO, M. 1966. Primary production by thephytoplankton community in some Japanese lakes
and its dependence on lake depth. Arch.Hydrobiol., 62: 1-28.
SCHINDLER, D. W. 1977. Evolution of phosphoruslimitation in lakes. Science, 195: 260-262.
SCHINDLER, D. W., F. A. ARMSTRONG, S. K.HOLMGREN & G. J. BRUNSKILL. 1971.Eutrophication of Lake 227, Experimental LakesArea, northwestern Ontario, by addition of phos-phate and nitrate. J. Fish. Res. Bd. Can., 28: 1763-1782.
SCHINDLER, D. W., E. J. FEE, & T.RUSZCZYNSKI. 1978. Phosphorus input and itsconsequences for phytoplankton standing cropand production in the Experimental Lakes Areaand in similar lakes. J. Fish. Res. Board Can., 35:190-196.
SMITH,V. H. 1982. The nitrogen and phosphorusdependence of algal biomass in lakes: An empiri-cal and theoretical analysis. Limnol. Ocenogr., 27:1101-1112.
SMITH,V.H. 1983. Low nitrogen to phosphorusratios favor dominance by blue-green algae in lakephytoplankton. Science, 221: 669-671.
SMITH, V. H. & S. J. BENNETT. 1999.Nitrogen:phosphorus supply ratio and phytoplank-ton community structure in lakes. Arch.Hydrobiol., 146: 37-53.
STORET, 2001. STORET Water Quality Data forNebraska Lakes, USA. http://nrcnt2.nrc.state.ne.us/cgi-win/storet.exe/station_search
STRASKRABA, M., 1976. Empirical and analyticalmodels of eutrophication . In: Proceedings of anInternational Symposium on eutrophication andrehabilitation of surface waters “EUTROSYM76”. : 352-371. Karl-Marx-Stadt, GDR, Vol. III.
STUMM, W. & J. J. MORGAN. 1996. AquaticChemistry: Chemical Equilibria and Rates inNatural Waters. 3rd. ed. J. Wiley & Sons, Inc. NewYork, USA.
TILMAN, D., S. S. KILHAM & P. KILHAM. 1982.Phytoplankton community ecology: the role oflimiting nutrients. Annu. Rev. Ecol. Syst., 13: 349-372.
TUNDISI, J. G., T. MATSUMURA TUNDISI, M. C.CALIJURI, AND E. M. L. NOVO. 1991.Comparative limnology of five reservoirs in themiddle Tiete River, S.Paulo State. Verh. Internat.Verein. Limnol., 24: 1489-1496.
USGS, 2001. Water Quality Samples for USA. USGeological Service, USA. http://water.usgs.gov/-nwis/qwdata
Nitrate and ammonia in lakes 49

WATSON, S., E. MCCAULEY & J. A. DOWNING.1992: Sigmoid relationships between phosphorus,algal biomass, and algal community structure.Can. J. Fish. Aquat. Sci., 49: 2605-2610.
WETZEL, R.G. 2001. Limnology. Lake and RiverEcosystems. 3rd. Ed. Academic Press, San Diego,CA, USA.
WYLIE, G. D. & J. R. JONES. 1986. Limnology of awetland complex in the Mississippi alluvial valleyof southeast Missouri. Arch. Hydrobiol./Suppl.,74: 288-314.
YAN, N.D. 1986. Empirical prediction of crustaceanzooplankton biomass in nutrient-poor CanadianShield lakes. Can. J. Fish. Aquat. Sci., 43: 788-796.
50 R. Quirós

INTRODUCCIÓN
Los lagos se originan principalmente por la obs-trucción del drenaje superficial debido particu-larmente a procesos morfogenéticos, conforman-do una estructura temporal, en la historiaerosional de los sistemas geográficos (Bellair &
Pomero, 1977; Mosetti, 1977). Ellos recibenaportes sólidos y líquidos de su cuenca de drena-je, mediante escurrimiento lineal, laminar y sub-terráneo, razón por la cual las características dela calidad del agua y de las comunidades bioló-gicas allí presentes, reflejan los efectos acumula-dos de todos los aportes de agua y materiales
Caracterización y tendencias tróficas de cinco lagos costeros de Chile Central
Oscar Parra, Claudio Valdovinos, Roberto Urrutia, Marcos Cisternas, Evelyn Habit y María Mardones
Unidad de Sistemas Acuáticos, Centro de Ciencias Ambientales EULA-Chile, Universidad de Concepción.Casilla 160-C, Concepción, Chile. E-mail: [email protected]
RESUMEN
El presente trabajo corresponde a una caracterización geográfica y limnológica de un sistema de 5 lagos costeros, localiza-dos en la fachada occidental de la Cordillera de Nahuelbuta (VIII Región Chile), denominados “Lagos Nahuelbutanos”(Laguna Grande de San Pedro, Laguna Chica de San Pedro, Laguna Quiñenco, Lago Lanalhue y Lago Lleulleu). Estos exhi-ben diversos niveles de intervención humana, debido principalmente al uso turístico y recreativo, a la infuencia urbana y alintenso uso forestal de sus cuencas hidrográficas. Los estudios realizados comprenden la geomorfología y origen de loslagos, usos históricos y actuales del suelo, caracterización y evolución de la cubierta vegetal, morfometría y batimetría,características sedimentológicas y paleolimnológicas, calidad del agua y biota acuática (fitoplancton, bentos y fauna ícti-ca), así como algunos aspectos asociado al metabolismo de la columna de agua. Se realizó un análisis limnológico yambiental comparativo en relación con la condición trófica actual de estos sistemas. Los resultados indican diferencias enlos niveles de trofía, siendo el Lago LleuLleu el más oligotrófico y Laguna Grande de San Pedro el mas eutrófico. En gene-ral el grado de trofía y de calidad del agua de estos cuerpos de agua se relacionan con indicadores biológicos y con el nivelde intervención a que han sido sometidas sus cuencas de drenaje.
Palabras clave: cuencas hidrográficas, uso del suelo, paleolimnología, calidad del agua, biota, eutrofización, intervenciónantrópica.
ABSTRACT
The present work correspond to a geographical and limnological characterization dealing with a coastal five-lake systemlocated at the eastern piedmont of the Nahuelbuta mountain range (VIII Region, Chile) named “Lagos Nabuelbutanos”(Laguna Grande de San Pedro, Laguna Chica de San Pedro, Laguna Quillenco, Lago Lanalhue y Lago Lleulleu). The lakesshow several levels of human intervention, mainly due to tourist and recreational uses, urban influence, and forestal inten-sive use in their hydrographic basins. The performed studies comprise the geomorphology and origin of the lakes, historicaland current soil use, characterization and evolution of the plant coverage, morphometry and bathymetry, sedimentologicaland paleolimnological characteristics, water quality and aquatic biota (i.e., phytoplankton, benthos, and ichthyic fauna, aswell as some topics associated with the water column metabolism. A comparative environmental and limnological analysiswas performed, in relation to the current trophic conditions of these systems. The results show differences in trophic state,being Lago Lleulleu the most oligotrophic, and Laguna Grande de San Pedro the most eutrophic. In general, the trophicdegree and water quality of these water bodies are related with biological indicators and with the level of intervention towhich their drainage basins have been subjected.
Keywords: basins, soil usage, paleolimnology, water quality, biota, eutrophication, anthropic intervention.
51
Limnetica 22(1-2): 51-83 (2003)© Asociación Española de Limnología, Madrid. Spain. ISSN: 0213-8409

procedentes del entorno. Diversos autores desta-can la estrecha relación existente entre el estadotrófico de un sistema limnético y las condicionesgeográficas y particularmente geomorfológicasdel lago y de su cuenca de drenaje (Ryding &Rast, 1992). Aspectos tales como la extensión, laprofundidad de un lago y el aporte de materialparticulado, son relevantes en la determinacióndel ciclo de vida del sistema limnético; éstos serelacionan estrechamente con los procesos mor-fogenéticos de la cuenca lacustre.
La diversidad climática y geológica de Chilecontinental conforma una realidad territoriallatitudinal, de tal magnitud, que permite ladiferenciación de una gran variedad de ecosis-temas terrestres y acuáticos, algunos de ellosambientalmente relevantes. Entre estos ecosis-temas acuáticos continentales relevantes seobservan de norte a sur (Fig. 1), el sistema delagos del altiplano, el sistema de lagunas hiper-salinas de la segunda región, el sistema delagos nord-patagónicos o araucanos, el sistemade lagos de la isla de Chiloé, el sistema delagos magallánicos o patagónicos y el sistemade lagos costeros de la región centro sur deno-minados como “Lagos Nahuelbutanos” (Parraet al., 1999) por estar insertos en un dominiogeográfico común; la vertiente litoral centro-norte de la Cordillera de Nahuelbuta.
Particularmente, en la Región del Biobío, ladisponibilidad de cuerpos de aguas lénticos esbastante escasa comparada con la magnitud delas aguas corrientes. En esta Región los sistemasde lagos más importantes se encuentran en la pre-cordillera de los Andes, sobre los 800 m.s.n.m, enlas zonas de nacimiento del sistema fluvial delrío Biobío. Estos son el lago Laja (“Laguna dela Laja”) y las lagunas Icalma y Galletué. En elsector costero de la Región, se encuentra el siste-ma de lagos anteriormente nombrado, los quecorresponden a lagos costeros de aguas dulces,localizados en las estribaciones occidentales dela Cordillera de Nahuelbuta. El sistema de lagos“Nahuelbutanos” esta constituído por 6 cuerposlénticos localizados en la vertiente sur occiden-tal de la Cordillera de Nahuelbuta (parte de laCordillera de la Costa) que se alinean de norte a
sur entre los sistemas fluviales de los ríosBiobío e Imperial. Entre ellos destacan: LagunaChica de San Pedro, Laguna Grande de SanPedro, Laguna La Posada, Laguna Quiñenco,Lago Lanalhue y Lago Lleu-Lleu.
A diferencia de los lagos que se encuentranen la parte alta y la precordillera andina de laRegión, cuyas aguas tienen bajos contenidosde nutrientes, debido a una mínima interven-ción de sus cuencas de drenaje y ningunainfluencia de asentamientos urbanos, los lagosnahuelbutanos exhiben diversos niveles deintervención humana de norte a sur. Así, loslagos nahuelbutanos septentrionales se encuen-tran rodeados en parte por centros urbanos,desarrollándose una importante actividad turís-tica y recreativa en ellos. Además, la mayorparte de sus cuencas hidrográficas han sido yestán sometidas a una intensa actividad fores-tal. Por otra parte, los dos lagos nahuelbutanosmeridionales (Lanalhue y Lleulleu) correspon-den a áreas de desarrollo indígena, lo cualconstituye un componente cultural, actualmen-te relevante en Chile para la toma de decisio-nes respecto al uso de estos recursos acuáticos.
Desde esta perspectiva, los procesos queafectan la calidad y usos de estos recursos,como la eutrofización, requieren una especialatención de la comunidad científica. Es necesa-rio generar el conocimiento de base para decidiracciones de protección ambiental que permitansu control y su uso sustentable. La eutrofizaciónrepresenta el proceso de envejecimiento naturalde los lagos, como resultante de la acumulacióngradual de nutrientes, un incremento de la pro-ductividad biológica y la depositación paulatinade sedimentos provenientes de su cuenca de dre-naje. En condiciones naturales el proceso deeutrofización es lento, y las tasas de cambioocurren normalmente a escala temporal de mile-nios. Sin embargo, por causas antrópicas rela-cionadas con el mal uso del suelo, el incrementode la erosión y por la descarga de aguas servidasdomésticas, se ve acelerado a escala temporal dedécadas o menos (Vollenweider, 1968).
La evaluación cuantitativa del estado tróficoy el grado de contaminación de los sistemas
52 Parra et al.

lacustres, es de gran trascendencia en gestiónambiental territorial, ya que permite determinarrestricciones de uso de estos recursos (e.g.abastecimiento de agua para consumo humano,baño), y orientar medidas de recuperación ymitigación cuando corresponda (e.g. colectoresde aguas lluvia, forestación). Los sistemas tra-dicionales de evaluación del estado trófico desistemas lacustres, se basan fundamentalmenteen el contenido de fósforo y nitrógeno, y losclasifican en oligo-, meso- y eutróficos. Sinembargo, en las últimas décadas, esta aproxi-mación ha sido motivo de discusión en la litera-
tura científica (Tundisi et al., 1997), sugiriendola utilización de un mayor número de indicado-res ambientales, no solo del cuerpo de agua ensí, sino también de las relaciones con su cuencade drenaje y con una escala temporal másamplia, que permita explicar en el tiempo lasacciones y los procesos que determinaron lascondiciones actuales.
Nuestro grupo de trabajo ha estudiado loscuerpos de aguas anteriormente nombrados através de sus distintos componentes, obteniendoinformación básica de las características limno-lógicas de cada uno de ellos. Esto ha permitido
Caracterización de cinco lagos costeros chilenos 53
Figura 1. Localización latitudinal de los principales sistemas lacustres de Chile. Latitudinal location for themain lake systems in Chile.

realizar un análisis comparativo de los cincocuerpos de agua y de sus respectivas cuencashidrográficas, explicando en parte el estado tró-fico actual, sus causas, sus efectos, sus indica-dores y sus tendencias.
Considerando el valor como patrimonio natu-ral, el uso actual y potencial que representa elsistema de lagos nahuelbutanos para la segundaárea mas poblada e industrializada del país, seha decidido realizar la presente contribución,que tiene como objetivo integrar esta informa-ción y aquella todavía no publicada, realizandoun análisis ambiental comparativo de las carac-terísticas limnológicas de los cinco cuerposacuáticos, incluidas sus cuencas de drenaje,
poniendo especial énfasis en la condición trófi-ca y estado de eutrofización y estimar su ten-dencia, para posteriormente identificar accionesde conservación y protección ambiental.
ÁREA DE ESTUDIO
Los sistemas acuáticos estudiados (Fig. 2), tie-nen la característica de ser cuerpos de aguacercanos a la zona litoral marina. Estos poseensimilar base geológica y climatológica, dondelas diferencias radican fundamentalmente en eluso de suelo, la intensidad de la actividadforestal silvícola, las actividades turísticas y laocupación urbana de sus respectivas cuencashidrográficas (Tabla 1).
MATERIALES Y MÉTODOS
Los estudios realizados comprendieron: geo-morfología y el origen de los lagos (Mardones& Reuther, 1999), erosión, sedimentación, usoshistóricos y actuales del suelo de las cuencaslacustres y la caracterización y evolución de lacubierta vegetal (Cisternas et al., 1997, 1999a,1999b, 2000, 2001), morfometría y batimetríade los cuerpos de agua, características sedimen-tológicas y paleolimnología (Urrutia et al.,2000a, 2000b), calidad del agua y biota acuáti-ca (fitoplancton, bentos y fauna íctica) en cuan-to a biodiversidad y como organismos indi-cadores, y algunos aspectos asociados almetabolismo de la columna de agua (Parra,1989; Parra et al., 1976, 1989, 1999; Scasso &Campos, 1998, 1999; Valdovinos & Figueroa,2000; Valdovinos et al., en prensa).
Geología, Hidrografía, Clima y Geomorfología
La información geológica, hidrográfica y delclima se obtuvo de los trabajos de Ferraris(1981), Ferraris & Bonilla (1981), Fuenzalida(1971), Cier-Serplac (1976), Devynck (1970),Endlicher & Mackel (1985), Veil (1961), Katz
54 Parra et al.
Figura 2. Localización de las lagunas Grande de San Pedro,Chica de San Pedro y Quiñenco, y de los lagos Lanalhue yLleu-lleu. Geographic location of the lagoons Grande de SanPedro, Chica de San Pedro y Quiñenco, and the Lanalhue yLleu-lleu lakes.

(1970). El levantamiento geomorfológico deta-llado se efectuó mediante la fotointerpretaciónde los vuelos (1994-1995) y de un vuelo parti-cular (1996) solicitado por los investigadores.El sistema morfogenético fue clasificado basán-dose en criterios morfológicos, morfométricos,sedimentológicos, litológicos y de uso delsuelo, aspectos que fueron relevados, analizadosy evaluados directamente en terreno (Mardones& Reuther, 1999).
Uso histórico del uso del suelo
Se realizó un detallado análisis de fotointerpre-tación para una de las cuencas más representati-vas. Se utilizaron fotografías aéreas de la cuen-ca de Laguna Chica (vuelo Trimetrogon 1943,vuelo Hycon 1955, vuelo OEA 1961, vueloSAF, 1981, vuelo SAF 1994). La información setraspasó, corrigiendo digitalmente las diferentesescalas, a la cartografía base (1:10 000). Selograron seis diferentes cartas de uso mostrandosu evolución de los últimos 50 años Esta carto-
grafía fue ingresada al SIG Arc/Info, separandolos usos en diferentes coberturas. A través delSIG se obtuvo el Cambio Total entre periodos,es decir, se identificó, por traslape digital, lasáreas que sufrieron cambios entre los años estu-diados, independientemente de los tipos de uso(Cisternas et al., 2001).
Cambios en las tasas de erosión y sedimentación
Se determinaron las tasas de erosión-sedimenta-ción de Laguna Chica, considerándola comorepresentativa de los 5 lagos en estudio. Estainformación fue posteriormente comparada conlas modificaciones de uso del suelo. Mediantegeocronología isotópica (210Pb y 137Cs) y paleo-palinología (Cisternas et al., 1997, Urrutia et al.,2000, Cisternas et al., 2001), se determinaron lastasas de sedimentación de un core obtenido en laparte más profunda del lago. Considerando deter-minadas premisas se infirieron las tasas históri-cas y actuales de erosión en la cuenca .
Caracterización de cinco lagos costeros chilenos 55
Tabla 1. Características generales de las cuencas hidrográficas y grado de intervención humana. General characteristics of the hydrogra-phic basins and degree of human intervention.
PARAMETROS Y L. CHICA L. GRANDE L. QUIÑENCO L. LANALHUE L. LLEU-LLEUUSOS DEL SUELO
Area del lago (km2) 0.82 1.55 0.29 31.9 39.8Area cuenca (km2) 4.5 12.7 3.0 327.43 538.82Rel. Área cuenca / área lago 5.5 8.2 10.3 10.2 16.8Rel. Área cuenca / vol. lago 523.3 992.2 3333.3 777.5 717.3Desarrollo línea de costa 1.8 2.1 1.5 2.9 4.2% Bosque y matorral nativo 27.13% 5.68% 17% 25.2% 14.42%% Plantaciones 48.86% 52.35% 70.3% 52.42% 40.21%% Agricultura 3.35% 1.02% 1.6 % 22.35% 45.32%% Urbano 4.9% 4.1% 0% ¿ 0%%Prot.ecológica 0% 0% 0% 0% 0%Evacuación aguas servidas no antes no antes noActividad turística y intensa regular ninguna 0.11% 0.05%recreativa (camping y playas)Invasión de macrófitas Sí Sí No No NoGrado trófico Mesotrófico Eutrófico Distrófico Eutrófico OligotróficoMacrófitas acuáticas Abundantes Muy abundantes Escasas Regular EscasasEstratificación de verano Sí No No Sí Sí

Comparación entre las condicionesprehispánicas y actuales
Se tomaron cores en la parte más profunda delas Lagunas Grande y Chica de San Pedro y encada caso se obtuvieron los centímetros superfi-ciales (representando a las condiciones actuales)y los más profundos (representando a las condi-ciones prehispánicas). Los sedimentos inferio-res fueron datados con 14C, para corroborar sucondición prehispánica. Se analizaron las diato-meas, para determinar calidad del agua, y supolen, para reconocer la evolución de la vegeta-ción en la cuenca. (Cisternas et al., 1999a, b;Urrutia et al., 2000, Cisternas et al., 2001).
Morfometría y batimetría
El levantamiento batimétrico se realizó utilizandoun ecosonda Lowrance X-16 con 192 kHz defrecuencia y un transductor de 8° (LowranceElectronics, INC.) con una sensibilidad de ±0,1m. El posicionamiento de los transectos se realizóutilizando las cartas del IGM, escala 1:50 000 ypuntos notables en las orillas de los lagos. En elcaso de Laguna Chica de San Pedro, LagunaGrande de San Pedro y el Lago Lanalhue seprocedió a una actualización de los mapasbatimétricos realizados por Dellarossa & Parra(1985) y Scasso (1996). La determinación de losprincipales parámetros morfométricos se realizósiguiendo a Hutchinson (1957).
Sedimentología
La determinación de MOT se realizó de acuerdoa la metodología propuesta por Mills (1978), elanálisis granulométrico y textural siguiendoWentworth (1922).
Calidad del agua
Con el objeto de considerar la variabilidad tem-poral de las características físicas y químicas delos lagos, fueron muestreados estacionalmentedurante los períodos de verano (enero 1997),otoño (mayo 1997), invierno (agosto 1997) y pri-
mavera (diciembre 1997). Las muestras fueronobtenidas a las distintas profundidades con unabotella Rüttner de 2 L de capacidad y transporta-das al laboratorio en frascos plásticos de 5 L a ca.4°C, y analizadas dentro de las primeras 6 horasde obtenidas. Las muestras fueron analizadas deacuerdo a las metodologías estándar indicadaspor la American Public Health Association(APHA). Las determinaciones realizadas in situfueron las siguientes: temperatura (termómetrode mercurio), pH (pH-metro Shott), conductivi-dad (conductivímetro Hanna), transparencia(disco Secchi Hydro-Bios) y turbidez (turbidíme-tro Milton-Roy Co.). Las determinaciones reali-zadas en laboratorio fueron las siguientes: oxíge-no disuelto (método de Winckler), nitrógeno total(método de Kjendall), fósforo total (métodomolibdato-tungstato), alcalinidad (titulación conHCl) y seston total (gravimetría).
Fitoplancton
En cada lago se establecieron dos o tres estacio-nes de muestreo, una en el sector de entrada del(o los) afluentes principales, otra en la zona demayor profundidad y que generalmente corres-pondió al centro del lago y finalmente una ter-cera, en las proximidades del efluente principaldel lago. Estos se efectuaron en verano (enero1997), otoño (mayo 1997), invierno (agosto1997) y primavera (diciembre 1997). Se utiliza-ron una red de 55 mm de trama y la botellamuestreadora Rüttner. Se trabajó con un micros-copio invertido Karl Zeiss y para el recuento seutilizó el procedimiento descrito en Utermöhl(1957). Mayor información sobre la metodolo-gía relacionada con la comunidad fitoplanctóni-ca referirse a Parra et al., (1999). La determina-ción taxonómica se basó fundamentalmente enlos trabajos de Parra et al., 1976, 1980,1981,1982, 1983 y Rivera et al., 1983.
Metabolismo bentónico
Empleando un core de gravedad (48 mm dediámetro interior; 0.51 de longitud), en enero de1997 se obtuvieron muestras no perturbadas de
56 Parra et al.

sedimento en los sectores más profundos de loslagos. Las tasas de decaimiento de oxígenofueron determinadas incubando los cores enoscuridad a temperatura constante (18±0.5°C)siguiendo a (Newrkla & Gunatilaka, 1982). Parael estudio de la dinámica de la temperatura ycontenido de oxígeno sobre el fondo de loslagos, en enero, mayo, agosto y diciembre de1997, se obtuvieron muestras de agua con unabotella Rüttner a ca. 30 cm del fondo. Latemperatura fue medida con un termómetro dealcohol (Hydro-Bios), y el oxígeno disueltodeterminado con el método de Winckler (verStrickland & Parsons, 1979).
Zoobentos
Los muestreos se realizaron en los meses deagosto-septiembre de 1997. Se tomaron 8 répli-cas para la parte litoral y 4 réplicas para la zonaprofundal. En la zona litoral se muestreómanualmente empleando cuadrantes de 0.1 m2.Las muestras de la parte profundal se obtuvie-ron mediante una draga Petit-ponar de 0.02 m2
de mascada. Todas las muestras se tamizaroncon una trama de 500 µm, se fijaron con forma-lina al 10%. La macrofauna se separó e identifi-có una lupa binocular. Para el análisis espacialde las comunidades bentónicas, se analizó unamatriz de densidad (individuos por m2 por espe-cie y estación de muestreo) empleando un análi-sis de conglomerados y escaleo no-métricomultidimensional (NMDS). Las estaciones(muestras) se compararon y agruparon, paradefinir áreas con similar composición biológica(análisis normal o tipo-q).
Fauna íctica
El análisis de las comunidades de peces se reali-zó sobre la base de la información aportada porCorfo (1995) para los lagos Lleulleu yLanalhue, y Scasso (1996) y Scasso & Campos(1998 y 1999) para lagunas Grande y Chica. Noexisten antecedentes sobre la ictiofauna del lagoQuiñenco. El estudio de la fauna íctica deLleulleu y Lanalhue se basó en dos muestreos
realizados en invierno y verano de 1995. Enambos casos se utilizó un único arte de pesca,consistente en redes de trasmallo, por lo que nose incluyó en este estudio el muestreo de la zonalitoral de los lagos. El estudio de lagunasGrande y Chica incluyó 5 muestreos en losmeses de noviembre de 1993, enero, marzo,junio y septiembre de 1994. Las comunidadespelágicas se estudiaron mediante métodoshidroacústicos (ecosonda Lowrance X16) y cap-turas con redes monofilamento de 40 m de largoy 2.5 m de ancho y 32, 35, 45 y 55 mm de entre-nudo. La producción se estimó mediante losmodelos de Downing et al., (1990) y Downing& Plante (1993) y el máximo rendimiento soste-nido mediante el método de Graham (Ricker,1981). La comparación de estos parámetrosentre ambos sistemas se efectuó utilizandoANOVA. La comunidad litoral se muestreómediante redes de arrastre en las mismas épocasde año. Para el estudio de edades y crecimientode Odontesthes bonariensis se utilizó el métodode von Bertalanffy (1938).
RESULTADOS
Geología
Los sistemas lacustres se distribuyen de Norte aSur en la ladera occidental de la Cordillera deNahuelbuta, unidad morfoestructural de laCordillera de la Costa de Chile Centro-Sur(Región del Biobío). Sus cuencas hidrográficasse abren y drenan hacia las llanuras costeras enel sector septentrional y hacia las plataformasmarinas y llanuras litorales en el Sur.
Los afloramientos rocosos varían en natu-raleza y en edad de E a W; mientras que la ver-tiente cordillerana donde se emplazan las cuen-cas lacustres está conformada por rocascristalinas y metamórficas, la franja más cerca-na al litoral, se estructura en rocas sedimenta-rias clásticas y sedimentos de origen marino,fluvial y eólico.
Desde el margen septentrional del lagoLanalhue hasta el río Biobío, aflora el Basa-
Caracterización de cinco lagos costeros chilenos 57

mento metamórfico Serie Oriental, constituidopor metagrauvacas, f ilitas, rocas córneas ygneises, de edad Paleozoico superior. Al Sur dellago Lanalhue y en toda la hoya del lagoLlelleu, las rocas estructurantes del relievecorresponden a la Serie Occidental delBasamento metamórfico. Esta serie comprendeprincipalmente micaesquistos y metacherts,muy deformados; profundamente meteorizadospor los diferentes agentes climáticos, los quehan generado suelos arcillosos de color rojizo-castaño, interestratif icados con depósitos deladera (Endlicher & Mäckel, 1985). Las rocascristalinas intrusivas, reconocidas principal-mente en el margen nororiental de la cuenca dellago Lanalhue, corresponden al Batolito de laCosta, unidad informal compuesta de granitos,tonalitas, granodioritas y dioritas, con zonas demigmatitas y gneises. Este emerge en la partecentral de la Cordillera de Nahuelbuta, intruyeal Basamento metamórfico y ha sido datado delPaleozoico superior (Ferraris, 1981).
En el margen N del área de estudio, en lazona de contacto entre la Cordillera costera ylas llanuras litorales afloran rocas sedimentariascontinentales eocénicas atribuidas a laFormación Cosmito (Veyl, 1961), conformadaprincipalmente por lutitas y arcosas. Estos aflo-ramientos se distribuyen como una franja dis-continua de dirección NE-SO, alternando conlas rocas metamórficas Paleozoicas, al S deLaguna Grande de San Pedro. Hacia el extremoSur de dicha área estas características litológi-cas se mantienen, ya que se adosan al cordónmontañoso terrazas esculpidas en rocas sedi-mentarias de edad Terciaria; las formacionesmás antiguas observadas son de edad Eocénicay las más recientes, corresponden a rocas de laFormación Tubul, de edad Plioceno superior. Enambos casos, predominan rocas del tipo lutitas yareniscas (Ferraris & Bonilla, 1981).
Las llanuras situadas al Oeste de las cuencaslacustres están constituidas por sedimentosmarinos, fluviales y eólicos de edad Pleisto-cénica-Holocénica.
Desde el punto de vista tectónico, el rasgomás destacado es la foliación de la Serie meta-
mórfica Occidental, la que está asociada a unproceso de metamorfismo-deformación y falla-mientos en bloque, de edad Postmiocénica;deformaciones que se relacionan con el alza-miento de la Cordillera de Nahuelbuta . Lasfracturas de gran ángulo (mayor de 45º), lamayoría con manteo cercano a la vertical, pare-cen corresponder a movimientos de tipo normal,presentando rumbos generalmente ENE, NW yNNE. Dichas fracturas, son fundamentales paracomprender tanto el trazado geométrico de lared hidrográfica, como la morfología y la bati-metría de las cuencas lacustres. Esta influenciaes clara en la Laguna Grande de San Pedro,cuyo sistema léntico ocupa un valle de línea defalla, de dirección NNE-SSO, que discurre casiparalelo a la línea costera. También tiene inci-dencia en el flujo de agua subterránea que ali-menta los sistemas lacustres.
Clima
Este sistema de lagos está localizado en la zonamedia y húmeda del área costera dentro delárea de influencia Mediterránea (Fuenzalida,1971). Según Cier-Serplac (1976) las precipita-ciones incrementan de Oeste a Este debido alefecto climático de barrera que ejerce laCordillera de Nahuelbuta. Desde el norte hacialos lagos mas al sur del sistema, las precipita-ciones varían entre 1.250 mm a cerca de 2500mm. Las lluvias se concentran principalmenteen invierno (Mayo-Octubre). La temperaturamedia anual es 12 a 13 °C. Desde Mayo aAgosto la dirección de viento dominante varíaentre W y N y durante el resto del añovaría entre S y SW. Esta alternancia es debida auna influencia anticiclónica durante el verano,y durante la influencia ciclónica con la aproxi-mación del frente polar que es el que generaalteraciones atmosféricas en el invierno.
Geomorfología
Los cinco lagos se emplazan en valles de laCordillera de Nahuelbuta cuyo drenaje local hasido bloqueado. Existen importantes diferencias
58 Parra et al.

morfométricas y morfogenéticas entre los treslagos situados en el margen N del área de estu-dio (Lagunas Chica y Grande de San Pedro yLaguna Quiñenco) y aquellos situados en lazona meridional (lagos Lanalhue y Lleulleu).
Los sistemas lacustres septentrionalesLas cabeceras de las cuencas lacustres se empla-zan en los cordones montañosos de erosiónhídrica, de la naciente Cordillera de Nahuelbuta,estructurados en roca metamórfica Paleozoica;cuyos interfluvios presentan orientación N yONO y altitudes > 400m. Adosada a la cordillerase aprecia una terraza de erosión marina de 65 a100m de altitud, 450m de ancho dispuesta comouna franja continua de dirección N-S y pendiente<5º, conformada tanto por roca sedimentariaEocénica, como por filitas y esquistos Paleo-zoicos. El contacto con la cordillera es general-mente abrupto y lineal, lo que permitiría suponerque se trata de un escarpe adaptado a líneas defalla. Tanto los cordones como las terrazas estándisectados por pequeños y estrechos valles defuertes pendientes longitudinales.
Al Oeste y ONO de estas unidades morfoló-gicas se sitúa la llanura litoral de San Pedro-Coronel, cuya altitud media es de aproximada-mente 6 m.s.n.m. Conforma una franjacontinua de 1-4 km de ancho, de dirección N-S,modelada por cordones litorales y dunas. Por elnúmero y grado de conservación de cordoneslitorales observados, se estima que la llanura sehabría modelado entre el Tardiglacial y elReciente, al menos en seis estadios de retroce-so del mar. Este tipo de modelado se presentaparticularmente, en el frente de los lagosQuiñenco y San Pedro Grande. En el margenNorte de las dos Lagunas de San Pedro, el ríoBiobío construye una terraza de acumulaciónconstituida por sedimentos arenosos basálticos,de edad Pleistocénica-Holocénica, parcialmen-te modelada por dunas.
Los sistemas lacustres meridionalesEn el margen Sur del área de estudio, los lagosLanalhue y Lleulleu emplazan sus cabeceras enel eje mismo de la Cordillera de Nahuelbuta. La
cordillera está modelada por cordones y restosde superficies de erosión de origen continental,estructurados en rocas metamórficas (filitas yesquistos) de edad Paleozoica, los que se elevanprogresivamente hacia el E de 500 a 800m.s.n.m., a través de bloques quebrados por unsistema de fallas ONO-ESE y ENE-OSO.
Siguiendo las mismas direcciones estructura-les citadas anteriormente, los ejes de los lagosse han emplazado en antiguos valles adaptadosa la estructura, tal como se aprecia por su mor-fología y batimetría. Del mismo modo, lascuencas hidrográficas de ambos sistemas, pre-sentan trazados angulosos que evidencian lainfluencia estructural.
El contacto con la llanura litoral se produce através de niveles decrecientes de terrazas con-formadas por areniscas y lutitas Terciarias,cuyas altitudes varían entre 25 y poco más de200 m.s.n.m. La morfología plana, su disposi-ción en franjas paralelas a la línea costera, elafloramiento de rocas sedimentarias marinas deedad Terciaria testigos de antiguas fasesde transgresión marina, permiten asumir queestos niveles de terraza adosados a la Cordillerade Nahuelbuta, son de abrasión marina. Losniveles de terraza mejor desarrollados son aque-llos situados más próximos a la costa, suponién-dose por esta razón que son también los másjóvenes. Ellos se distribuyen irregularmenteentre los 25-30m, 50m y 70-80m.s.n.m. El restode las terrazas, están representadas por retazosmuy fragmentados, exceptuando el nivel de 200 m.s.n.m. que presenta mayor continuidad.
El frente del lago Lleulleu está modelado porla terraza marina de 25-30m, sobre la cual sobre-yacen dunas longitudinales, las que no estánrelacionadas con la obstrucción del drenaje,puesto que sobreyacen a las terrazas marinas, aun nivel superior al espejo de agua. La terraza de50m de altitud, está ampliamente representadaen el frente del lago Lanalhue; sobre ella no seobservan modelados de origen eólico. Esta pla-nicie de origen marino-eólico inclinada hacia elOeste, tiene 6 km de ancho en el sector Norte yse estrecha progresivamente hacia el Sur, hastadesaparecer en el sector de Quidico, donde las
Caracterización de cinco lagos costeros chilenos 59

estribaciones cordilleranas caen directamente alocéano formando acantilados rocosos. Se reco-nocen aquí una secuencia de cordones dunariosde diferentes edades y de altitudes que varíanentre 16 y 47m. En el lago Lanalhue las dunasmás próximas al cuerpo de agua, quedan distan-tes unos 4 km del frente del lago, de modo quesus riberas están modeladas en areniscas de edadTerciaria. En el frente del lago Lleulleu, la llanu-ra litoral tiene cerca de 2.6 km de ancho y lasdunas se aproximan al frente mismo del lago,observándose allí que los sedimentos eólicossobreyacen una formación de rocas sedimenta-rias de edad probablemente Eocénica. Estemodelado no tiene mayor influencia en la forma-ción del lago, pero lo nutren de sedimentos porla deflación eólica cuando la cobertura vegetales débil. Los lagos están drenados por los ríosLleulleu y Puyehue-Paicaví.
El lago Lanalhue se sitúa a 12 m.s.n.m.,inunda un estrecho valle de la Cordillera deNahuelbuta de orientación SE-NO. Su morfolo-gía y batimetría tienen estrecha relación con losaccidentes tectónicos que estructuran la cuen-ca. El lago se divide en tres secciones: la sec-ción interior es la menos profunda (4m) y defondo plano; se dispone en dirección Norte-Sury tiene 3 km de longitud y 1.5 km de ancho.Está parcialmente seccionada por el delta de losríos Elicura y Calebu y se comunica con elresto del lago, a través de un estrecho (Pta. ElSapo). La parte central tiene 8 km de longitud y2-3 kms de ancho; se dispone en dirección SE-NO y su fondo tiene la forma de un estrechocanal, de laderas irregulares, cuya profundidadaumenta en dirección a la costa, de 14 a 22m.b.n.m. El trazado de las riberas se presentaaccidentado por penínsulas y ensenadas.Finalmente, el sector frontal es el más estrechoy profundo: tiene la forma de una T volcadahacia el Oeste, 3 km de largo, entre 0.5 y 0.8km de ancho y 24 m.b.n.m. de profundidad.Hacia el Este, una península estructurada porrocas sedimentarias de la terraza de abrasiónmarina localizada en el sector de El Banquete,genera un umbral que dif iculta la conexiónentre las dos últimas secciones.
El lago Lleulleu tiene una forma lobulada,angulosa, cuyo trazado repite las direccionesestructurales observadas en la cordillera: NNE-SSO y O-E. Se ha formado por la obstrucciónde dos valles principales de dirección E-O: losvalles de Huillinco y Mahuilque y un eje dedirección NNE-SSO, casi paralelo a la líneacostera, situado en la falla que separa laCordillera de Nahuelbuta de las terrazas litora-les. El lago tiene una altitud de 20 m.s.n.m.,0.5 a 3 km de ancho y el eje mayor, de direcciónNNE-SSO se extiende por aproximadamente17 km de longitud. En cuanto a su batimetría, laparte frontal del lago que se inserta en las terra-zas litorales es la de menor profundidad (10 m),mientras que en los lóbulos internos se ha esti-mado en más de 40m. La compartimentaliza-ción batimétrica del sistema, al igual que en elLanalhue se relaciona con líneas de falla queaccidentan el borde cordillerano.
Origen de los lagos
Los sistemas lacustres de Lanalhue y Lleulleu,emplazados al sur del área de estudio tienen unorigen distinto a aquellos situados en las cercaní-as de Concepción (las Lagunas Chica y Grandede San Pedro y laguna Quiñenco). La formaciónde los primeros, ha sido interpretada a la luz delos modelos neotectónicos para la plataformade Arauco. Estos autores demuestran que a partirdel Pleistoceno, la plataforma de Arauco habríafuncionado como una zona de deformación co-sísmica, tectónicamente independiente de laCordillera de Nahuelbuta. Movimientos de emer-sión de la plataforma estarían asociados a la ocu-rrencia de grandes terremotos, con recurrencia deal menos uno por siglo. Se estiman movimientosde solevantamiento de las terrazas costeras de0.25m/siglo en la parte frontal de los lagos. Estopermite sugerir que dichos lagos corresponden avalles costeros, cuyo drenaje habría sido obstrui-do en el Pleistoceno reciente, por las formacionesrocosas de la plataforma de Arauco debido a lasperiódicas crisis sísmicas.
Los lagos litorales Lagunas Chica y Grandede San Pedro y Laguna Quiñenco, se emplazan
60 Parra et al.

en antiguos valles de la Cordillera de la Costacuyo drenaje ha sido bloqueado por dunas y cor-dones litorales. Tres factores se combinan paraexplicar su formación: el importante aporte dearenas basálticas a la costa provocado por des-cargas lacustres en el alto valle del Laja(Mardones & Jaque, 1996), las transgresiones yregresiones marinas de origen glacioeustático yla neotectónica co-sísmica Cuaternaria, que eneste sector ha actuado con menor intensidad ydistintamente en los bloques y fosas que estruc-turan la llanura de Concepción y Coronel.
En relación a Las lagunas de San Pedro, ajuzgar por la batimetría y la potencia de losdepósitos de arena en su frente, es posible queéstas se hayan formado y destruido subsecuen-temente desde fines del Riss al Actual, debido alas variaciones glacioeustáticas del nivel marinoen el Pleistoceno superior. De hecho, la últimatransgresión marina ocurrió entre el 8000 BP yel 6400 BP; ésta aumentó en 5m el nivel actualdel mar el que habría inundado al menos par-cialmente éstos valles, formando lagunas.A partir de este momento, viene una fase deregresión marina posiblemente asociada a sole-vantamientos co-sísmicos, lo que permite la for-mación sucesiva de crestas de playa y cordonesdunarios, que finalmente construyen los siste-mas lacustres actuales, desconectados del mar.En cuanto a la laguna Quiñenco, ésta ha sidooriginada en un periodo previo a la llegada de laúltima formación de arenas negras, ya que elcordón litoral que la cierra está construido porarenas cuarcíferas. Si se considera la propuestamorfogenética de Mardones & Jaque (1996), suformación sería previa al periodo Tardiglaciar(>15 000 BP) y relacionada estrictamente con latransgresión flandriana y los solevantamientoscosteros posteriores a este evento.
Hidrografía
Las hoyas hidrográficas de estos cinco lagosestán conformadas por pequeñas cuencas coste-ras exorreicas, que drenan la vertiente occiden-tal de la Cordillera de Nahuelbuta y tienen comonivel de base los sistemas lénticos. La jerarquía
de estas cuencas va desde drenes de orden 6 enlas cuencas más grandes observadas en el LagoLleulleu (ríos Huillinco y Mahuilque) hastaorden 1 en las cuencas más pequeñas. En laslagunas de San Pedro las cuencas de mayorjerarquía son de orden 3. La red de drenaje secaracteriza por distintos patrones y densidadesde canales, debido al control geológico y estruc-tural del área. Sobre el Basamento cristalino, elpatrón de drenaje es dendrítico con densidadmedia. En las terrazas de abrasión marina, encambio, se observa un patrón de drenaje parale-lo, con orientación principal de NNE a SSW oSSE-NNW. Según Ferraris y Bonilla (1981),esto se debe a una adaptación de los cursos flu-viales a los lineamientos estructurales. Aquí ladensidad de drenaje es gruesa, debido a la altapermeabilidad de estas rocas, lo que ademáspermite el desarrollo de valles profundos.
Vegetación y uso del suelo
La vegetación original del área de localizacióndel sistema lacustre se inserta en la transiciónentre las regiones meso e hidromórfica (DiCastri, 1968). El bosque esclerófilo se contactacon el del roble-laurel-lingue (Nothofagus spp. -Crptocarya alba, Persea lingue), que se desa-rrolla más al Sur. En las partes altas de las cuen-cas todavía existen especies de Nothofagus, N.oblicua (“Roble”) y N. procera (“Raulí”). Estebosque al ser talado origina matorrales escleró-filos, los cuales a su vez al ser explotados, seconvierten en comunidades arbustivas bajas.Estos hábitats son rápidamente colonizados pormalezas introducidas, como la “retamilla”(Telline monspessulana), las que forman comu-nidades asociadas a renovales del antiguo bos-que esclerófilo. Sobre esta situación vegetacio-nal y edáfica se ha desarrollado el proceso dereforestación con especies exóticas, Pinusradiata y Eucalyptus spp., el primero en muchomayor porcentaje (cerca del 80 % de la actualcubierta vegetacional). En las quebradas masprofundas de las cuencas del sistema se obser-van restos de vegetación nativa como especiesde “Olivillo” (Aextoxicum punctatum), “Peumo”
Caracterización de cinco lagos costeros chilenos 61
(Cryptocarya alba), “Boldo” (Peumus boldus),“Avellano” (Gevuina avellana), “Litre” (Lithreacaustica), “Maqui” (Aristotelia chilensis) yCanelo (Drymis winteri). En los últimos años lavegetación presenta una tendencia notoriamentedegradativa hacia comunidades del tipo mato-rral y pastizal, degradación que se traduce enerosión de los suelos (Barrientos, 1990).
Los usos del suelo de los cinco lagos son: a)bosque y matorral nativo, generalmente unaforma degradada de bosque, debido a la tem-prana intervención humana (e.g. corta y que-mas), b) plantaciones forestales, áreas ocupadaspor plantaciones forestales principalmente P.radiata y Eucalyptus globulos, c) áreas defores-tadas con escasa o nula cobertura vegetal gene-radas por diversas intervenciones humanas(corta, quemas, incendios), con suelos expues-
tos, d) áreas con sectores de humedales ye) áreas con cobertura urbana y residencial. LaTabla 1 presenta un cuadro comparativo de losusos del suelo de las respectivas cuencas dedrenaje de los lagos estudiados.
Por otra parte es necesario indicar que la acti-vidad forestal en la Región, basada mayormenteen plantaciones de especies exótica, es el carác-ter dominante en el paisaje, ocupando cerca del40 % de todo el territorio regional y más del50% del área de estudio. Al mismo tiemporepresenta la principal actividad económica encuanto a usos de recursos naturales.
Morfometría y batimetría
La morfología de los lagos estudiados presentóun amplio rango de variación tanto en forma
62 Parra et al.
Figura 3. Batimetría de los cuerpos de agua. Bathymetry of the water bodies.

como tamaño (Fig. 3). Es así que el tamaño delos lagos fluctuó entre 39.8 km2 (Lleulleu) y los3.0 km2 (Quiñenco) de superficie, con profun-didades que fluctuaron entre los 46.5 m(Lleulleu) y 6.1 m (Quiñenco). Por su parte laforma fluctuó entre contornos muy regulares, conbajo desarrollo de costa, como el caso de lagunaQuiñenco (1.5, Laguna Chica (1.8) y LagunaGrande (2.1) de San Pedro, hasta formas ycontornos muy irregulares, representados poraltos valores de desarrollo de línea de costa de loslagos Lleulleu (4.2) y Lanalhue (2.9).
La poca profundidad de los lagos estudiadoses consistente con la uniformidad geológica delárea y la escasa altura del relieve donde seencuentran ubicados (Mardones & Reuther,1999). Sólo el Lago Lleulleu presentó una pro-fundidad máxima superior a los 30 m. Estosresultados indican que todos los lagos, conexcepción del Lleulleu, presentan el 100% de suárea sobre la cota que determina la zona deaguas someras que según Campos et al., (1992)está determinada por la isóbata de los 30 m. Ellago Lleulleu presentó una zona de aguas some-ras que corresponden aproximadamente al 50%de su área total. La batimetría de estos cuerposde agua esta estrechamente asociado al origende cubetas que contienen las masas de agua. Los
lagos Quiñenco, Chica y Grande de San Pedropresentan una sola gran cubeta de fondo plano,que ocupa la mayor extensión en superficie, confuertes pendientes en las riberas asociadas a lacordillera de Nahuelbuta y una suave batimetríahacia el extremo opuesto. En cambio la batime-tría de los lagos Lanalhue y Lleulleu esta deter-minado por el origen tectónico de estos cuerposde agua. Es posible encontrar una completacaracterización morfométrica de los lagos enUrrutia et al., (2000), (Tabla 2).
Caracterización sedimentológica
Los sedimentos de los cinco lagos estudiados secaracterizaron por presentar texturas fangosas(limos y arcillas) muy homogéneas, con ausen-cia de fracciones gruesas (arenas y gravas). Entodos los casos se trata de fangos sin caracterís-ticas reductoras, de colores “Grises mediana-mente oscuros” (N4) con tonalidades “Griscafé” (5YR4/1) y “Gris oliva” (5Y4/1), deacuerdo a la Carta de Colores de la GeologicalSociety of América (1975).
Considerando el tamaño medio de las partícu-las de sedimento, el fango de todos los lagos seclasificó texturalmente (Wentworth, 1922)como “limos finos”. Por otra parte y de acuerdo
Caracterización de cinco lagos costeros chilenos 63
Tabla 2. Características morfométricas de los lagos. Lakes’ morphometric characteristics.
PARAMETROS L. CHICA L. GRANDE L. QUIÑENCO L. LANALHUE L. LLEU-LLEU
Latitud (S) 36º 51’ 36º 51’ 36º 59’ 37º 55’ 38º 09’Longitud (W) 73º 05’ 73º 06’ 73º 07’ 73º 19’ 73º 19’Altura (m.s.n.m.) 5.0 4.0 5.0 12.0 20.0Profundidad máx. (m) 18.0 13.5 6.1 26.0 46.5Profundidad media (m) 10.3 8.3 3.0 13.1 23.5Largo máx. (km) 1.9 2.7 1.1 9.6 13.2Ancho máx. (km) 0.87 1.4 0.36 4.3 3.7Perímetro (km) 5.7 9.4 2.94 58.6 93.3Area del lago (km2) 0.82 1.55 0.29 31.9 39.8Area cuenca (km2) 4.5 12.7 3.0 325.0 670.0Volumen (km3) 0.0086 0.0128 0.0009 0.418 0.934Desarrollo línea de costa 1.8 2.1 1.5 2.9 4.2Rel. Prof. media / Prof. máx. 0.57 0.61 0.49 0.50 0.50Rel. Área cuenca / área lago 5.5 8.2 10.3 10.2 16.8Rel. Área cuenca / vol. lago 523.3 992.2 3333.3 777.5 717.3Prof. criptodepresión (m) 13.0 9.5 1.1 14.0 26.9
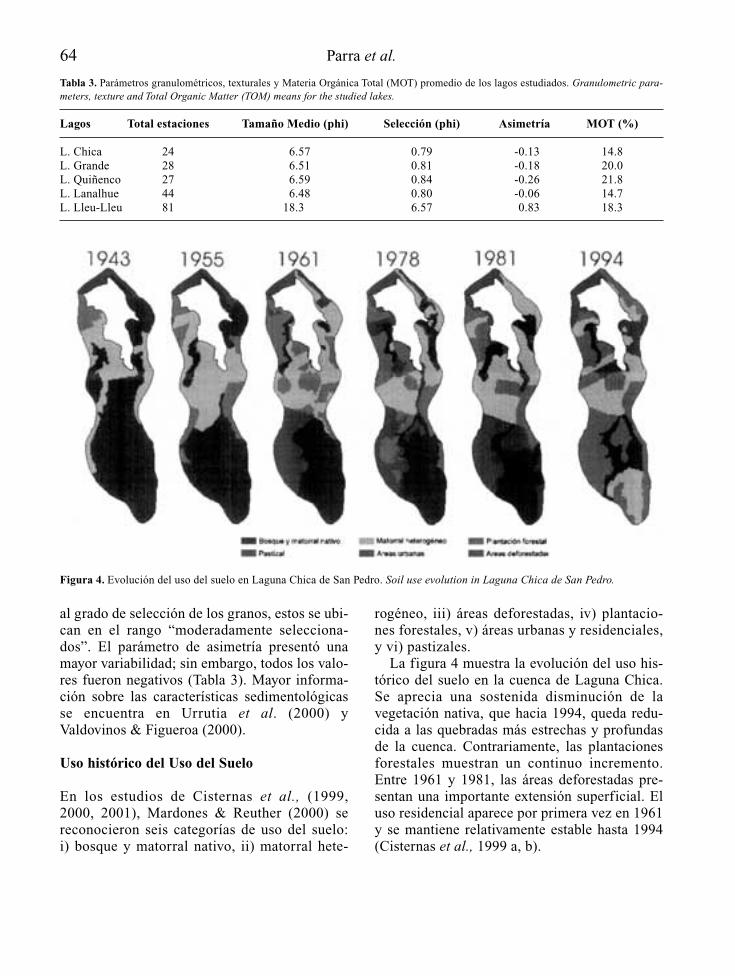
al grado de selección de los granos, estos se ubi-can en el rango “moderadamente selecciona-dos”. El parámetro de asimetría presentó unamayor variabilidad; sin embargo, todos los valo-res fueron negativos (Tabla 3). Mayor informa-ción sobre las características sedimentológicasse encuentra en Urrutia et al. (2000) yValdovinos & Figueroa (2000).
Uso histórico del Uso del Suelo
En los estudios de Cisternas et al., (1999,2000, 2001), Mardones & Reuther (2000) sereconocieron seis categorías de uso del suelo:i) bosque y matorral nativo, ii) matorral hete-
rogéneo, iii) áreas deforestadas, iv) plantacio-nes forestales, v) áreas urbanas y residenciales,y vi) pastizales.
La figura 4 muestra la evolución del uso his-tórico del suelo en la cuenca de Laguna Chica.Se aprecia una sostenida disminución de lavegetación nativa, que hacia 1994, queda redu-cida a las quebradas más estrechas y profundasde la cuenca. Contrariamente, las plantacionesforestales muestran un continuo incremento.Entre 1961 y 1981, las áreas deforestadas pre-sentan una importante extensión superficial. Eluso residencial aparece por primera vez en 1961y se mantiene relativamente estable hasta 1994(Cisternas et al., 1999 a, b).
64 Parra et al.
Tabla 3. Parámetros granulométricos, texturales y Materia Orgánica Total (MOT) promedio de los lagos estudiados. Granulometric para-meters, texture and Total Organic Matter (TOM) means for the studied lakes.
Lagos Total estaciones Tamaño Medio (phi) Selección (phi) Asimetría MOT (%)
L. Chica 24 6.57 0.79 -0.13 14.8L. Grande 28 6.51 0.81 -0.18 20.0L. Quiñenco 27 6.59 0.84 -0.26 21.8L. Lanalhue 44 6.48 0.80 -0.06 14.7L. Lleu-Lleu 81 18.3 6.57 0.83 18.3
Figura 4. Evolución del uso del suelo en Laguna Chica de San Pedro. Soil use evolution in Laguna Chica de San Pedro.

Respecto al Cambio Total, la figura 5 muestra lasáreas que modificaron su uso entre los periodosestudiados. Se aprecia un incremento de las modifi-caciones con el tiempo, del mismo modo, se reco-noce que el cambio se fue trasladando hacia el sur,a los sectores de altas pendientes (Cisternas et al.,1999 a, b). La Figura 6 muestra el total de modifi-caciones ocurridas entre 1943 y 1994. Se observaque la casi la totalidad de la superficie de la cuencafue transformada en un periodo de 50 años.
Considerando estas tendencias y la evoluciónhistórica de la vertiente occidental de la cordi-llera de Nahuelbuta, es posible inferir que lascinco cuencas lacustres sufrieron un proceso dereemplazo de la vegetación nativa de un modosimilar al reconocido para Laguna Chica. Deeste modo, Cisternas et al., (1999 b) proponenun modelo de reemplazo de vegetación nativapor plantaciones forestales para toda la cordi-llera de Nahuelbuta (Fig. 7). El Periodo I,representado por los años cuarenta, muestrauna cobertura de bosque y matorral nativodegradado, minoritariamente en los sectoresplanos y mayoritariamente en las áreas de pen-dientes. Con el avance de las plantaciones(Periodo II), principalmente ubicadas en luga-res llanos, los remanentes de nativo están limi-tados sólo a los sectores altos (probablemente
por las dificultades de explotación). A diferen-cia de la rápida transición en los dos primerosperiodos, el Periodo III presenta áreas defores-tadas, que se mantienen por largo tiempo en lossectores más frágiles de las cuencas (PeriodosIII-IV). Finalmente, el propio proceso de fores-tación hará desaparecer las áreas deforestadas(Periodo V), quedando las cuencas completa-mente cubiertas con las plantaciones exóticas.
Cambios en las tasas de erosión y sedimentación
En la figura 8 es posible observar los resultadosdel análisis cronológico para Laguna Chica.Tanto el 137Cs como la concentración de polen deP. radiata comprueban el modelo cronológicopropuesto (Cisternas et al., 2001). De acuerdocon el modelo cronológico utilizado (CRS) latasa neta de acumulación de sedimentos deLaguna Chica ha variado en un orden de magni-tud, desde 50 g m-2 a-1 hacia 1883 a 600 g m-2 a-1
en 1968. Los cambios en las tasas de sedimenta-ción muestran tres pulsos durante el s. XX. Elprimero comienza a fines del s. XIX y alcanza sumáximo al final de los años cuarenta (580 g m-2
a-1). Otro pulso comienza a inicios de los añoscincuenta (240 g m-2 a-1) y alcanza su máximo a
Caracterización de cinco lagos costeros chilenos 65
Figura 5. Evolución del Cambio Total durante los intervalos temporales estudiados (A= 1943-1955; B= 1955-1961; C= 1961-1978; D= 1978-1981; E= 1981-1994). Total change evolution during the studied temporal intervals (A= 1943-1955; B= 1955-1961; C= 1961-1978; D= 1978-1981; E= 1981-1994).

finales de los sesenta (600 g m-2 a-1). El eventomás reciente comienza alrededor de 1978 (260 gm-2 a-1) y continua en los noventa, cuando elmáximo fue 520 g m-2 a-1 (Cisternas et al., 2001).En Chile, como en el resto del hemisferio sur, nose detectaron aumentos de las concentraciones de137Cs ambiental por efecto del accidente deChernobyl ocurrido en Ucrania en 1986 (SurfaceAir Sampling Program; Larsen et al., 1995). Enun monitoreo realizado en Chile entre los años1966-1994 (Piñones & Tomicic, 1995), no sedetectaron los efectos de dicho accidente nuclear.Del mismo modo, análisis realizados en sedimen-tos de la Antártica (Appleby et al., 1995) no sereconocieron indicios de fallout radioactivogenerado por dicho accidente. Lo anterior, sedebe a que la inyección de 137Cs en Chernobyl nofue de una magnitud suficiente para incorporarsea la estratosfera. De este modo, la contaminaciónno fue transportada a escala planetaria, afectandoprincipalmente al norte de Europa (Larsen et al.,1995). Considerando lo anterior, el peak detecta-do en la columna sedimentaria de Laguna Chicade San Pedro, debe responder más bien al máxi-mo de emisión estratosférica de 1963.
La erosión en la cuenca del lago fue calcula-da como S*AL/AW, donde S es la tasa de sedi-mentación calculada para el core, AL es el áreadel fondo del lago, y AW el área de la cuencahídrica (Cisternas et al., 2001). Se obtuvieronlas tasas de erosión para los 10 primeros centí-metros del núcleo (1942-1996). El registro delluvia para el área (Figura 9 A) muestra que lasprecipitaciones no han tenido una tendencia aaumentar o disminuir durante los últimos50 años. De este modo, es posible inferir que lasvariaciones en la erosión de la cuenca han esta-do controladas principalmente por el uso desuelo. Este mismo periodo temporal es cubiertopor la información de uso del suelo obtenidamás arriba. La tasa de erosión, a escala linear detiempo (Figura 9 B), presenta los mismos tresmáximos que las tasas de sedimentación; sinembargo la escala linear muestra que fueronbreves. Los primeros dos peaks, alrededor de1950 y 1970, duraron unos pocos años; el tercermáximo en los años noventa, dura 7 años. Los
tres máximos se aproximan a 1 tm ha-1 a-1.Valores bajo la media (0.5 tm ha-1a-1) predominanpor periodos más largos (Cisternas et al., 2001).
66 Parra et al.
Figura 6. Total de modificaciones ocurridas entre 1943 y1994. Total changes occured between 1943 and 1994.
Figura 7. Modelo conceptual del proceso de cambio de usodel suelo para la cordillera de Nahuelbuta (Cisternas et al.,1999 b). Conceptual model of the soil use change process forthe Nahuelbuta range (Cisternas et al., 1999 b)
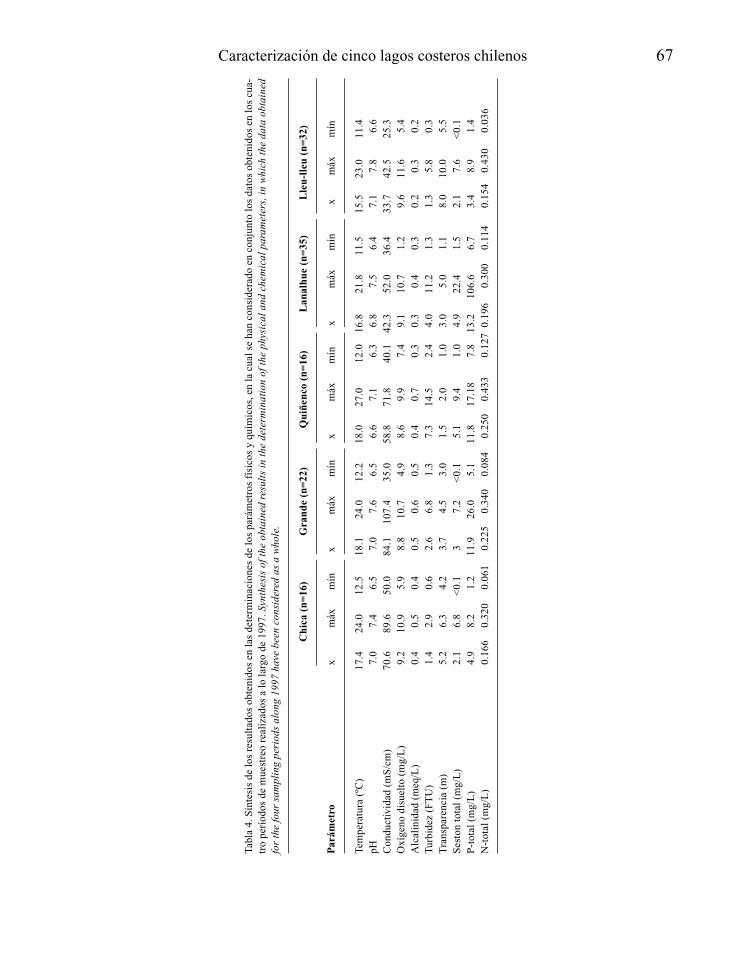
Caracterización de cinco lagos costeros chilenos 67
Tabl
a 4.
Sín
tesi
s de
los
resu
ltad
os o
bten
idos
en
las
dete
rmin
acio
nes
de lo
s pa
rám
etro
s fí
sico
s y
quím
icos
, en
la c
ual s
e ha
n co
nsid
erad
o en
con
junt
o lo
s da
tos
obte
nido
s en
los
cua-
tro
perí
odos
de
mue
stre
o re
aliz
ados
a l
o la
rgo
de 1
997.
Syn
thes
is o
f th
e ob
tain
ed r
esul
ts i
n th
e de
term
inat
ion
of t
he p
hysi
cal
and
chem
ical
par
amet
ers,
in
whi
ch t
he d
ata
obta
ined
for
the
four
sam
plin
g pe
riod
s al
ong
1997
hav
e be
en c
onsi
dere
d as
a w
hole
.
Ch
ica
(n=
16)
Gra
nd
e (n
=22
)Q
uiñ
enco
(n
=16
)L
anal
hu
e (n
=35
)L
leu
-lle
u (
n=
32)
Par
ámet
rox
máx
mín
xm
áxm
ínx
máx
mín
xm
áxm
ínx
máx
mín
Tem
pera
tura
(ºC
)17
.424
.012
.518
.124
.012
.218
.027
.012
.016
.821
.811
.515
.523
.011
.4pH
7.0
7.4
6.5
7.0
7.6
6.5
6.6
7.1
6.3
6.8
7.5
6.4
7.1
7.8
6.6
Con
duct
ivid
ad (
mS
/cm
)70
.689
.650
.084
.110
7.4
35.0
58.8
71.8
40.1
42.3
52.0
36.4
33.7
42.5
25.3
Oxí
geno
dis
uelt
o (m
g/L
)9.
210
.95.
98.
810
.74.
98.
69.
97.
49.
110
.71.
29.
611
.65.
4A
lcal
inid
ad (
meq
/L)
0.4
0.5
0.4
0.5
0.6
0.5
0.4
0.7
0.3
0.3
0.4
0.3
0.2
0.3
0.2
Turb
idez
(F
TU
)1.
42.
90.
62.
66.
81.
37.
314
.52.
44.
011
.21.
31.
35.
80.
3T
rans
pare
ncia
(m
)5.
26.
34.
23.
74.
53.
01.
52.
01.
03.
05.
01.
18.
010
.05.
5S
esto
n to
tal
(mg/
L)
2.1
6.8
<0.
13
7.2
<0.
15.
19.
41.
04.
922
.41.
52.
17.
6<
0.1
P-t
otal
(m
g/L
)4.
98.
21.
211
.926
.05.
111
.817
.18
7.8
13.2
106.
66.
73.
48.
91.
4N
-tot
al (
mg/
L)
0.16
60.
320
0.06
10.
225
0.34
00.
084
0.25
00.
433
0.12
70.
196
0.30
00.
114
0.15
40.
430
0.03
6
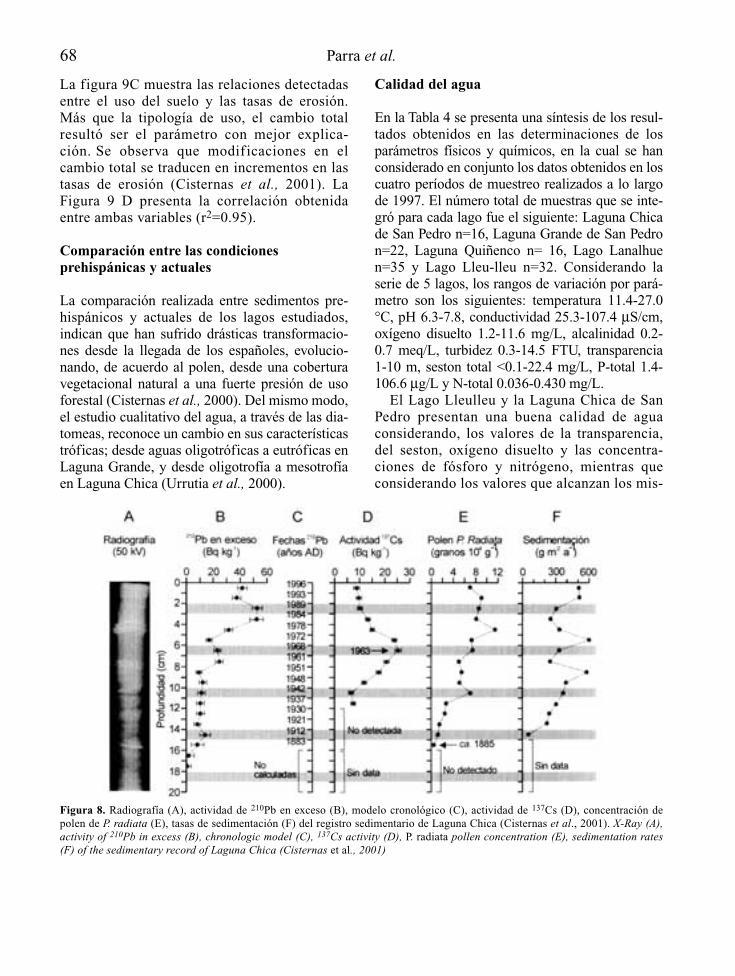
La figura 9C muestra las relaciones detectadasentre el uso del suelo y las tasas de erosión.Más que la tipología de uso, el cambio totalresultó ser el parámetro con mejor explica-ción. Se observa que modif icaciones en elcambio total se traducen en incrementos en lastasas de erosión (Cisternas et al., 2001). LaFigura 9 D presenta la correlación obtenidaentre ambas variables (r2=0.95).
Comparación entre las condicionesprehispánicas y actuales
La comparación realizada entre sedimentos pre-hispánicos y actuales de los lagos estudiados,indican que han sufrido drásticas transformacio-nes desde la llegada de los españoles, evolucio-nando, de acuerdo al polen, desde una coberturavegetacional natural a una fuerte presión de usoforestal (Cisternas et al., 2000). Del mismo modo,el estudio cualitativo del agua, a través de las dia-tomeas, reconoce un cambio en sus característicastróficas; desde aguas oligotróficas a eutróficas enLaguna Grande, y desde oligotrofía a mesotrofíaen Laguna Chica (Urrutia et al., 2000).
Calidad del agua
En la Tabla 4 se presenta una síntesis de los resul-tados obtenidos en las determinaciones de losparámetros físicos y químicos, en la cual se hanconsiderado en conjunto los datos obtenidos en loscuatro períodos de muestreo realizados a lo largode 1997. El número total de muestras que se inte-gró para cada lago fue el siguiente: Laguna Chicade San Pedro n=16, Laguna Grande de San Pedron=22, Laguna Quiñenco n= 16, Lago Lanalhuen=35 y Lago Lleu-lleu n=32. Considerando laserie de 5 lagos, los rangos de variación por pará-metro son los siguientes: temperatura 11.4-27.0°C, pH 6.3-7.8, conductividad 25.3-107.4 µS/cm,oxígeno disuelto 1.2-11.6 mg/L, alcalinidad 0.2-0.7 meq/L, turbidez 0.3-14.5 FTU, transparencia1-10 m, seston total <0.1-22.4 mg/L, P-total 1.4-106.6 µg/L y N-total 0.036-0.430 mg/L.
El Lago Lleulleu y la Laguna Chica de SanPedro presentan una buena calidad de aguaconsiderando, los valores de la transparencia,del seston, oxígeno disuelto y las concentra-ciones de fósforo y nitrógeno, mientras queconsiderando los valores que alcanzan los mis-
68 Parra et al.
Figura 8. Radiografía (A), actividad de 210Pb en exceso (B), modelo cronológico (C), actividad de 137Cs (D), concentración depolen de P. radiata (E), tasas de sedimentación (F) del registro sedimentario de Laguna Chica (Cisternas et al., 2001). X-Ray (A),activity of 210Pb in excess (B), chronologic model (C), 137Cs activity (D), P. radiata pollen concentration (E), sedimentation rates(F) of the sedimentary record of Laguna Chica (Cisternas et al., 2001)
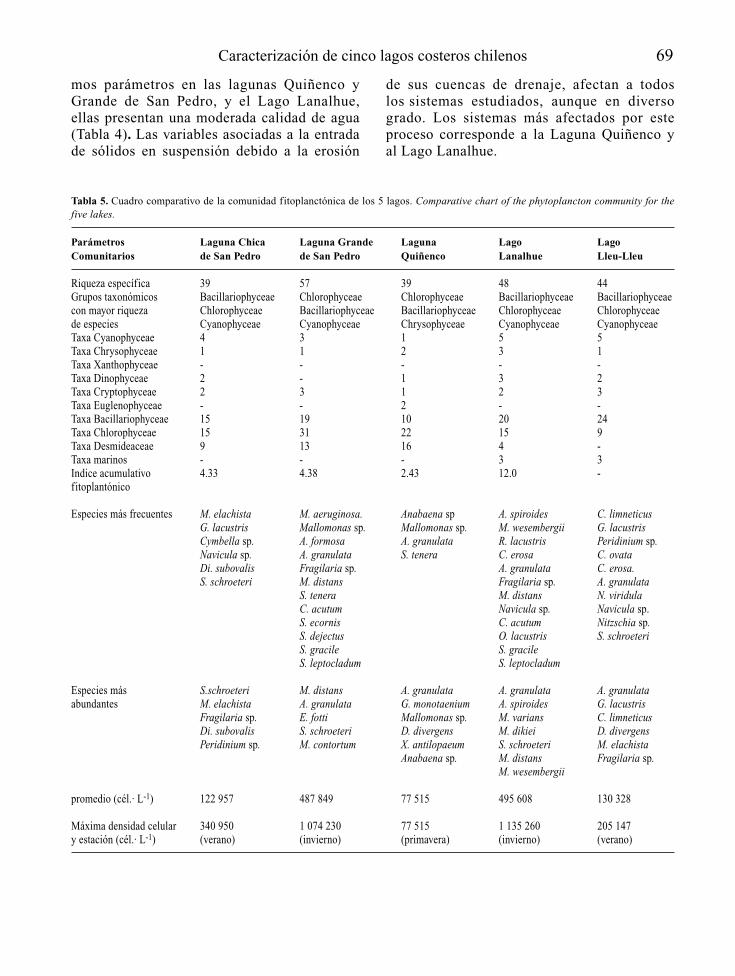
Caracterización de cinco lagos costeros chilenos 69
Tabla 5. Cuadro comparativo de la comunidad fitoplanctónica de los 5 lagos. Comparative chart of the phytoplancton community for thefive lakes.
Parámetros Laguna Chica Laguna Grande Laguna Lago Lago Comunitarios de San Pedro de San Pedro Quiñenco Lanalhue Lleu-Lleu
Riqueza específica 39 57 39 48 44Grupos taxonómicos Bacillariophyceae Chlorophyceae Chlorophyceae Bacillariophyceae Bacillariophyceaecon mayor riqueza Chlorophyceae Bacillariophyceae Bacillariophyceae Chlorophyceae Chlorophyceaede especies Cyanophyceae Cyanophyceae Chrysophyceae Cyanophyceae CyanophyceaeTaxa Cyanophyceae 4 3 1 5 5Taxa Chrysophyceae 1 1 2 3 1Taxa Xanthophyceae - - - - -Taxa Dinophyceae 2 - 1 3 2Taxa Cryptophyceae 2 3 1 2 3Taxa Euglenophyceae - - 2 - -Taxa Bacillariophyceae 15 19 10 20 24Taxa Chlorophyceae 15 31 22 15 9Taxa Desmideaceae 9 13 16 4 -Taxa marinos - - - 3 3Indice acumulativo 4.33 4.38 2.43 12.0 -fitoplantónico
Especies más frecuentes M. elachista M. aeruginosa. Anabaena sp A. spiroides C. limneticusG. lacustris Mallomonas sp. Mallomonas sp. M. wesembergii G. lacustrisCymbella sp. A. formosa A. granulata R. lacustris Peridinium sp.Navicula sp. A. granulata S. tenera C. erosa C. ovataDi. subovalis Fragilaria sp. A. granulata C. erosa.S. schroeteri M. distans Fragilaria sp. A. granulata
S. tenera M. distans N. viridulaC. acutum Navicula sp. Navicula sp.S. ecornis C. acutum Nitzschia sp.S. dejectus O. lacustris S. schroeteriS. gracile S. gracileS. leptocladum S. leptocladum
Especies más S.schroeteri M. distans A. granulata A. granulata A. granulata abundantes M. elachista A. granulata G. monotaenium A. spiroides G. lacustris
Fragilaria sp. E. fotti Mallomonas sp. M. varians C. limneticusDi. subovalis S. schroeteri D. divergens M. dikiei D. divergensPeridinium sp. M. contortum X. antilopaeum S. schroeteri M. elachista
Anabaena sp. M. distans Fragilaria sp.M. wesembergii
promedio (cél.· L-1) 122 957 487 849 77 515 495 608 130 328
Máxima densidad celular 340 950 1 074 230 77 515 1 135 260 205 147y estación (cél.· L-1) (verano) (invierno) (primavera) (invierno) (verano)
mos parámetros en las lagunas Quiñenco yGrande de San Pedro, y el Lago Lanalhue,ellas presentan una moderada calidad de agua(Tabla 4). Las variables asociadas a la entradade sólidos en suspensión debido a la erosión
de sus cuencas de drenaje, afectan a todoslos sistemas estudiados, aunque en diversogrado. Los sistemas más afectados por esteproceso corresponde a la Laguna Quiñenco yal Lago Lanalhue.
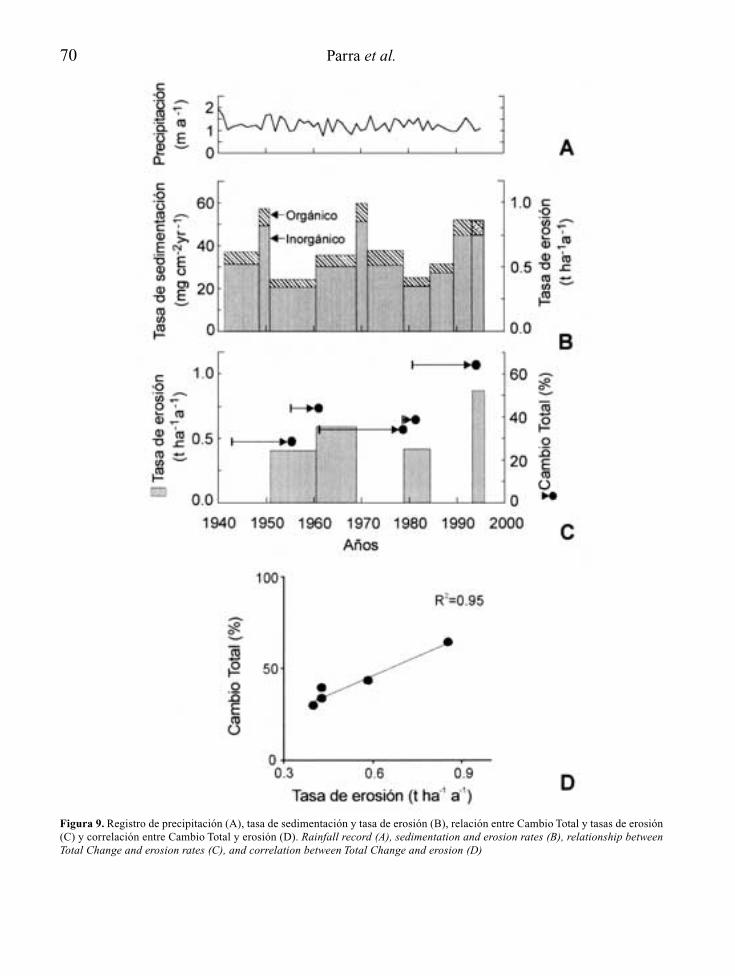
70 Parra et al.
Figura 9. Registro de precipitación (A), tasa de sedimentación y tasa de erosión (B), relación entre Cambio Total y tasas de erosión(C) y correlación entre Cambio Total y erosión (D). Rainfall record (A), sedimentation and erosion rates (B), relationship betweenTotal Change and erosion rates (C), and correlation between Total Change and erosion (D)
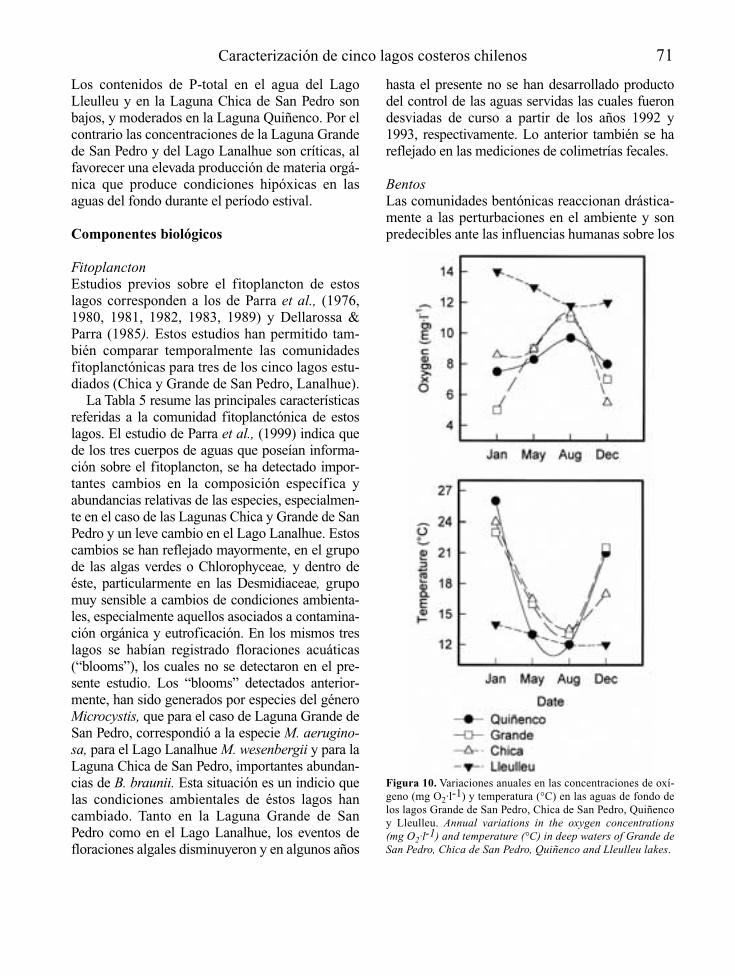
Los contenidos de P-total en el agua del LagoLleulleu y en la Laguna Chica de San Pedro sonbajos, y moderados en la Laguna Quiñenco. Por elcontrario las concentraciones de la Laguna Grandede San Pedro y del Lago Lanalhue son críticas, alfavorecer una elevada producción de materia orgá-nica que produce condiciones hipóxicas en lasaguas del fondo durante el período estival.
Componentes biológicos
FitoplanctonEstudios previos sobre el fitoplancton de estoslagos corresponden a los de Parra et al., (1976,1980, 1981, 1982, 1983, 1989) y Dellarossa &Parra (1985). Estos estudios han permitido tam-bién comparar temporalmente las comunidadesfitoplanctónicas para tres de los cinco lagos estu-diados (Chica y Grande de San Pedro, Lanalhue).
La Tabla 5 resume las principales característicasreferidas a la comunidad fitoplanctónica de estoslagos. El estudio de Parra et al., (1999) indica quede los tres cuerpos de aguas que poseían informa-ción sobre el fitoplancton, se ha detectado impor-tantes cambios en la composición específica yabundancias relativas de las especies, especialmen-te en el caso de las Lagunas Chica y Grande de SanPedro y un leve cambio en el Lago Lanalhue. Estoscambios se han reflejado mayormente, en el grupode las algas verdes o Chlorophyceae, y dentro deéste, particularmente en las Desmidiaceae, grupomuy sensible a cambios de condiciones ambienta-les, especialmente aquellos asociados a contamina-ción orgánica y eutroficación. En los mismos treslagos se habían registrado floraciones acuáticas(“blooms”), los cuales no se detectaron en el pre-sente estudio. Los “blooms” detectados anterior-mente, han sido generados por especies del géneroMicrocystis, que para el caso de Laguna Grande deSan Pedro, correspondió a la especie M. aerugino-sa, para el Lago Lanalhue M. wesenbergii y para laLaguna Chica de San Pedro, importantes abundan-cias de B. braunii. Esta situación es un indicio quelas condiciones ambientales de éstos lagos hancambiado. Tanto en la Laguna Grande de SanPedro como en el Lago Lanalhue, los eventos defloraciones algales disminuyeron y en algunos años
hasta el presente no se han desarrollado productodel control de las aguas servidas las cuales fuerondesviadas de curso a partir de los años 1992 y1993, respectivamente. Lo anterior también se hareflejado en las mediciones de colimetrías fecales.
BentosLas comunidades bentónicas reaccionan drástica-mente a las perturbaciones en el ambiente y sonpredecibles ante las influencias humanas sobre los
Caracterización de cinco lagos costeros chilenos 71
Figura 10. Variaciones anuales en las concentraciones de oxí-geno (mg O2·l-1) y temperatura (°C) en las aguas de fondo delos lagos Grande de San Pedro, Chica de San Pedro, Quiñencoy Lleulleu. Annual variations in the oxygen concentrations(mg O2·l-1) and temperature (°C) in deep waters of Grande deSan Pedro, Chica de San Pedro, Quiñenco and Lleulleu lakes.

ecosistemas acuáticos, como es el caso de laeutroficación cultural (Rosenberg & Resh, 1993).En el presente estudio, se analizó el efecto de latrofía de los sistemas lénticos, sobre la estructurade las comunidades de macroinvertebrados bentó-nicos. Los resultados muestran que el estado tró-fico de un lago tiene un importante efecto sobrelas comunidades bentónicas. En la figura 12 sepresenta un cuadro que resume la relación exis-tente entre las características bióticas de los lagosy su nivel de trofía (niveles de materia orgánica delos sedimentos, y niveles de Seston orgánico, P-total y N-total). Nótese que los 5 lagos están orde-nados en el espacio bidimensional de acuerdo a sumacrofauna, coincidiendo con el incremento delnivel de trofía de los lagos siguiendo la secuencia:L. Lleulleu, L. Chica de San Pedro, L. Quiñenco,L. Grande de San Pedro y L. Lanalhue.
En la figura 13 se señalan las sucesiones deespecies (o formas) responsables de la ordenaciónde las 23 estaciones de muestreo. La línea diagonaldivide el NMDS en dos campos: el derecho incluyea las estaciones de fondos blandos (círculos blan-cos) y el izquierdo a las de fondos duros (círculosnegros). Cada campo posee grupos característicosde taxa que están ordenados secuencialmente deacuerdo al nivel trófico de los lagos. En fondosduros de lagos pobres en nutrientes dominanMeriadialaris sp., Aegla sp. y Oligochaeta indet.3;mientras que en lagos ricos en nutrientes dominanDugesia anceps, Arrenuridae indet., Micropsectrasp. y Chironomus sp. Por otra parte, en fondos
blandos de lagos pobres en nutrientes dominanDiplodon chilensis y Pisidium chilense, mientrasque en los más ricos en nutrientes, Oribatuloideaindet. y Chironomidae indet.3.
Los resultados del análisis de regresión múlti-ple entre las variables abióticas y bióticas (Tabla7), indican que la agrupación de las estaciones demuestreo basándose en su macrofauna, es expli-cada fundamentalmente (en orden jerárquico) porel contenido de materia orgánica de los sedimen-tos, Seston orgánico, Seston total, Nitrógenototal, Fósforo total y Seston inorgánico.
Los resultados sugieren que: a) los factoresabióticos de la columna de agua y sedimentos,afectan la estructura comunitaria de los macroin-vertebrados bentónicos; b) existe una sucesión deespecies a lo largo del gradiente de trofía de loslagos, tanto en fondos duros litorales como enfondo blandos sublitorales. En fondos duros delagos pobres en nutrientes dominan Meriadialarissp., Aegla sp. y Oligochaeta indet.3; mientras queen lagos ricos en nutrientes dominan Dugesiaanceps, Arrenuridae indet., Micropsectra sp. yChironomus sp. Por otra parte, en fondos blandosde lagos pobres en nutrientes dominan Diplodonchilensis y Pisidium chilense, mientras que en losmás ricos en nutrientes, Oribatuloidea indet. yChironomidae indet.3.
Las tasas de decaimiento de oxígeno disueltoen columnas de sedimento no perturbadas, hansido empleadas como una medida integradora dela actividad metabólica de las comunidades ben-
72 Parra et al.
Tabla 6. Caracterización de los sedimentos superficiales (macroinvertebrados, granulometría, potencial redox (Eh) y carbono orgánico) ycontenido de nutrientes de las aguas de fondo (P-total y N-total) de los lagos Quiñenco, Grande de San Pedro, Chica de San Pedro yLeulleu. Superficial sediments characterization (mcroinvertebrates, granulometry, redox potential,(Eh) and organic carbon), and nutrientcontent of benthic waters (Total-P and Total-N) for lakes Quiñenco, Grande de San Pedro, Chica de San Pedro, and Leulleu.
Quiñenco Grande Chica Lleulleu
Core 1 Core 2 Core 1 Core 2 Core 1 Core 2 Core 1 Core 2
Macroinvertebrados (>250 µm) No No No No No No No NoGranulometría (Phi medio) 6.5 6.5 6.5 6.5 6.5 6.5 6.5 6.5Potencial redox1 mV) -250 -270 -350 -310 +45 -10 +200 +170Carbono orgánico (%) 25.3 28.5 21.4 22.5 15.3 19.0 14.0 17.1Total-P (µg l-1) 35.6 35.0 52.0 52.4 16.4 16.1 4.76 4.70Total-N (mg l-1) 0.86 0.84 0.68 0.60 0.64 0.65 0.07 0.07
1bajo los primeros 50 mm de la columna de sedimento.
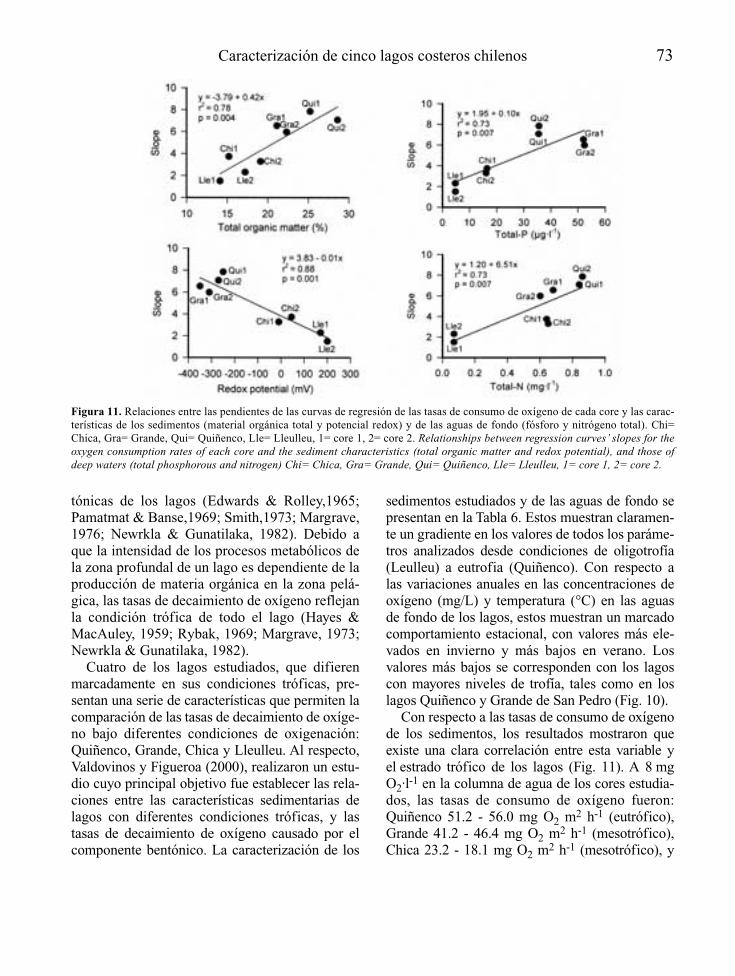
tónicas de los lagos (Edwards & Rolley,1965;Pamatmat & Banse,1969; Smith,1973; Margrave,1976; Newrkla & Gunatilaka, 1982). Debido aque la intensidad de los procesos metabólicos dela zona profundal de un lago es dependiente de laproducción de materia orgánica en la zona pelá-gica, las tasas de decaimiento de oxígeno reflejanla condición trófica de todo el lago (Hayes &MacAuley, 1959; Rybak, 1969; Margrave, 1973;Newrkla & Gunatilaka, 1982).
Cuatro de los lagos estudiados, que difierenmarcadamente en sus condiciones tróficas, pre-sentan una serie de características que permiten lacomparación de las tasas de decaimiento de oxíge-no bajo diferentes condiciones de oxigenación:Quiñenco, Grande, Chica y Lleulleu. Al respecto,Valdovinos y Figueroa (2000), realizaron un estu-dio cuyo principal objetivo fue establecer las rela-ciones entre las características sedimentarias delagos con diferentes condiciones tróficas, y lastasas de decaimiento de oxígeno causado por elcomponente bentónico. La caracterización de los
sedimentos estudiados y de las aguas de fondo sepresentan en la Tabla 6. Estos muestran claramen-te un gradiente en los valores de todos los paráme-tros analizados desde condiciones de oligotrofía(Leulleu) a eutrofia (Quiñenco). Con respecto alas variaciones anuales en las concentraciones deoxígeno (mg/L) y temperatura (°C) en las aguasde fondo de los lagos, estos muestran un marcadocomportamiento estacional, con valores más ele-vados en invierno y más bajos en verano. Losvalores más bajos se corresponden con los lagoscon mayores niveles de trofía, tales como en loslagos Quiñenco y Grande de San Pedro (Fig. 10).
Con respecto a las tasas de consumo de oxígenode los sedimentos, los resultados mostraron queexiste una clara correlación entre esta variable yel estrado trófico de los lagos (Fig. 11). A 8 mgO2·l-1 en la columna de agua de los cores estudia-dos, las tasas de consumo de oxígeno fueron:Quiñenco 51.2 - 56.0 mg O2 m2 h-1 (eutrófico),Grande 41.2 - 46.4 mg O2 m2 h-1 (mesotrófico),Chica 23.2 - 18.1 mg O2 m2 h-1 (mesotrófico), y
Caracterización de cinco lagos costeros chilenos 73
Figura 11. Relaciones entre las pendientes de las curvas de regresión de las tasas de consumo de oxígeno de cada core y las carac-terísticas de los sedimentos (material orgánica total y potencial redox) y de las aguas de fondo (fósforo y nitrógeno total). Chi=Chica, Gra= Grande, Qui= Quiñenco, Lle= Lleulleu, 1= core 1, 2= core 2. Relationships between regression curves’ slopes for theoxygen consumption rates of each core and the sediment characteristics (total organic matter and redox potential), and those ofdeep waters (total phosphorous and nitrogen) Chi= Chica, Gra= Grande, Qui= Quiñenco, Lle= Lleulleu, 1= core 1, 2= core 2.
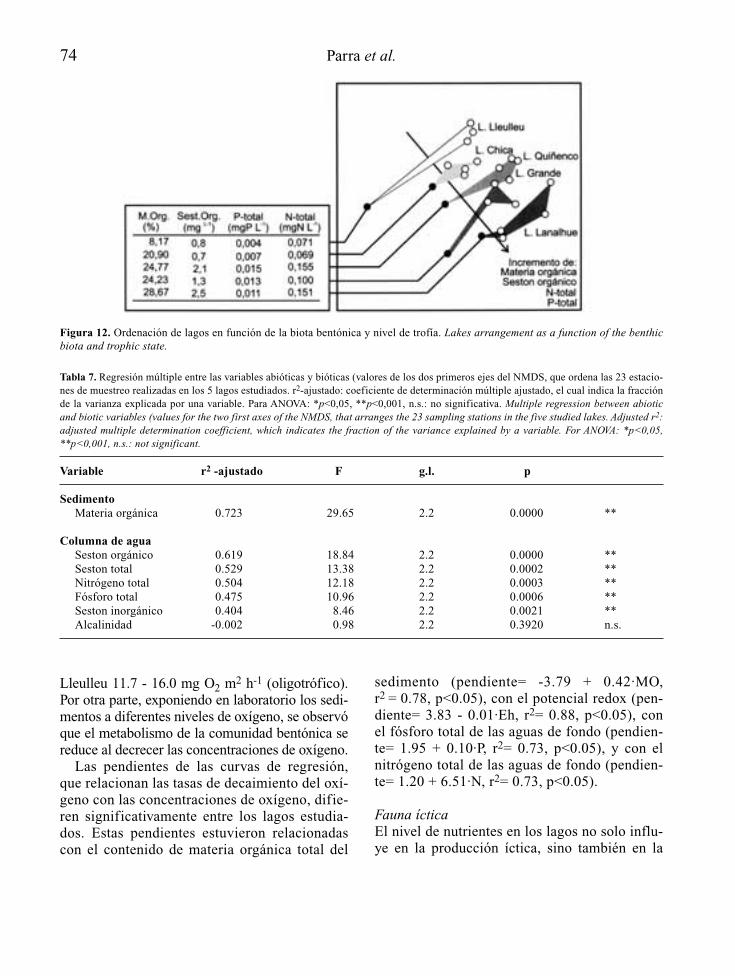
Lleulleu 11.7 - 16.0 mg O2 m2 h-1 (oligotrófico).Por otra parte, exponiendo en laboratorio los sedi-mentos a diferentes niveles de oxígeno, se observóque el metabolismo de la comunidad bentónica sereduce al decrecer las concentraciones de oxígeno.
Las pendientes de las curvas de regresión,que relacionan las tasas de decaimiento del oxí-geno con las concentraciones de oxígeno, difie-ren significativamente entre los lagos estudia-dos. Estas pendientes estuvieron relacionadascon el contenido de materia orgánica total del
sedimento (pendiente= -3.79 + 0.42·MO,r2 = 0.78, p<0.05), con el potencial redox (pen-diente= 3.83 - 0.01·Eh, r2= 0.88, p<0.05), conel fósforo total de las aguas de fondo (pendien-te= 1.95 + 0.10·P, r2= 0.73, p<0.05), y con elnitrógeno total de las aguas de fondo (pendien-te= 1.20 + 6.51·N, r2= 0.73, p<0.05).
Fauna ícticaEl nivel de nutrientes en los lagos no solo influ-ye en la producción íctica, sino también en la
74 Parra et al.
Figura 12. Ordenación de lagos en función de la biota bentónica y nivel de trofía. Lakes arrangement as a function of the benthicbiota and trophic state.
Tabla 7. Regresión múltiple entre las variables abióticas y bióticas (valores de los dos primeros ejes del NMDS, que ordena las 23 estacio-nes de muestreo realizadas en los 5 lagos estudiados. r2-ajustado: coeficiente de determinación múltiple ajustado, el cual indica la fracciónde la varianza explicada por una variable. Para ANOVA: *p<0,05, **p<0,001, n.s.: no significativa. Multiple regression between abioticand biotic variables (values for the two first axes of the NMDS, that arranges the 23 sampling stations in the five studied lakes. Adjusted r2:adjusted multiple determination coefficient, which indicates the fraction of the variance explained by a variable. For ANOVA: *p<0,05,**p<0,001, n.s.: not significant.
Variable r2 -ajustado F g.l. p
SedimentoMateria orgánica 0.723 29.65 2.2 0.0000 **
Columna de aguaSeston orgánico 0.619 18.84 2.2 0.0000 **Seston total 0.529 13.38 2.2 0.0002 **Nitrógeno total 0.504 12.18 2.2 0.0003 **Fósforo total 0.475 10.96 2.2 0.0006 **Seston inorgánico 0.404 8.46 2.2 0.0021 **Alcalinidad -0.002 0.98 2.2 0.3920 n.s.

composición específica de la comunidad (Colbyet al., 1972). En los sistemas estudiados lariqueza de especies varía de 3 en Lleulleu a12 especies en Laguna Grande (Tabla 8), refle-jando una tendencia al incremento en la riquezay diversidad de especies en este ambiente demayor trofía (Scasso, 1996). En los cuatrolagos, la relación de especies nativas a introdu-cidas es cercana a 1:1, a excepción del lagoLleulleu (Corfo, 1995), donde, debido al uso deun único arte de pesca, no ha sido registradala presencia de especies en la zona litoral. En la
zona pelágica, Odontesthes bonariensis corres-ponde a la especie más abundante de los cuatrosistemas, seguida por Oncorhynchus mykiss enLaguna Chica y Cyprinus carpio en Lanalhue yLaguna Grande. La alta abundancia de O. bona-riensis y C. carpio es común en sistemas eutró-ficos, donde son especies exitosas gracias a sushábitos alimentarios oportunistas (Prochelle &Campos, 1985; Vila & Soto, 1984).
Por su parte, las especies de salmónidos res-ponden inicialmente a la eutrofización con unaumento en su tasa de crecimiento corporal, peroluego disminuyen su reproducción, siendo final-mente reemplazadas por otras especies (Colby etal., 1972). Ello se refleja en la ausencia de S.trutta en los lagos Lanalhue, Grande y Chica, entanto que habita en Lleulleu, al igual que en otrostres lagos oligotróficos de la región (Laja, Icalmay Galletué; Campos et al., 1993, Scasso &Campos, 1998). La segunda especie salmonídeapresente en estos sistemas, O. mykiss, pareceadaptarse mejor que S. trutta a mayores nivelesde eutroficación, ya que se encuentra presente enlas lagunas Chica y Grande. Sin embargo, suaporte porcentual a la biomasa de la comunidadpelágica disminuye de un 26% en Laguna Chicaa un 19% en Laguna Grande (Scasso, 1996). Sumejor adaptación a niveles intermedios de eutro-ficación quedan demostrados además en la
Caracterización de cinco lagos costeros chilenos 75
Tabla 8. Especies de peces registradas en los 4 sistemas estudiados. Fish species registered for the 4 studied systems.
Especies Lleulleu Chica Lanalhue Grande
Nativas Percilia irwini � �Galaxias maculatus � � �Cheirodon galusdae � �Nematogenis inermis � �Percichthys trucha � �Geotria australis �
Introducidas Oncorhynchus mykiss � �Salmo trutta �Odontesthes bonariensis � � � �Cyprinus carpio � � � �Gambusia holbrooki � �Cichlasoma facetum � �Carassius carassius �
Total Nativas / introducidas 0/3 4/5 2/2 6/6Total 3 9 3 12
Figura 13. Especies indicadoras del bentos de la condicióntrófica de los lagos para ambientes litorales y profundos.Benthos trophic condition index species of the lakes, for litto-ral and deep environements

mayor abundancia, biomasa y producción quealcanza en laguna Chica respecto de lagunaGrande (Scasso & Campos, 1998), (Tabla 9). Deigual forma, O. bonariensis también refleja unamayor adaptación a ambientes mesotróficos,alcanzando mayor biomasa y producción enLaguna Chica y obteniendo un largo asintótico yedades significativamente mayores en LagunaChica (longitud total máxima observada 53 cmy VIII años) que en Laguna Grande (43 cm yV años) (Scasso & Campos, 1999). Sin embargo,la mayor tasa de crecimiento de O. bonariensis seproduce en Laguna Grande, lo que se relacionaríacon una mayor disponibilidad de alimento(Scasso & Campos, 1999).
A pesar del mayor nivel de trófía de LagunaGrande, los valores de densidad íctica (1171peces ha-1) y biomasa íctica (563±177 kg ha-1)no presentan diferencias significativas con losobtenidos en Laguna Chica (896 peces ha-1 y591±86 kg ha-1). Sin embargo, ambos valoresson significativamente mayores a los del LagoIcalma, oligotrófico (184 peces ha-1 y 164±24kg ha-1; Scasso, 1996). De igual forma, la pro-ducción íctica y el máximo rendimiento sosteni-do no son significativamente diferentes enambos sistemas, aún cuando, Laguna Grandepermite la extracción de mayores cuotas de cap-tura por especie (Tabla 9).
DISCUSIÓN Y CONCLUSIONES
Los ecosistemas acuáticos continentales interactú-an con el sistema terrestre de diferentes maneras,así, la mantención de los procesos ecológicos sonmuy dependientes de esta relación (Tundisi et al.,1997), las que también dependen de las caracterís-ticas geomorfológicas que han dado origen a lascuencas lacustres. Por lo tanto, es importante ana-lizar estas interacciones en un contexto regional,histórico y a una escala geológica y actual. Laevolución física y biológica de un lago es el resul-tado de un proceso dinámico el cual a su vez esdependiente de un sistema geomórfico (Tundisi etal., 1997). Así, las relaciones entre superficieterrestre, la hidrología de pendientes, la superficiedel cuerpo de agua y el desarrollo de la línea decosta, entre otras, son fundamentales para enten-der la magnitud y carácter de las entradas de mate-rial alóctono a los lagos y en la formación de lospatrones estacionales de estratificación física, quí-mica y biológica. Pero también es relevante tenerpresente la acción humana en una escala temporalapropiada a dicha acción, la que puede incidirfuertemente en el establecimiento de nuevospatrones geomorfológicos y también en las inter-acciones entre los sistemas terrestres y acuáticos.
El sistema estudiado ofrece una oportunidadpara comprender las características ecológicas
76 Parra et al.
Tabla 9. Producción íctica, máximo rendimiento sostenido y cuotas de captura por especie en Laguna Grande y Chica (Basado en Scasso1996). Ichthyc production
Especie
Laguna Chica de San Pedro Laguna Grande de San Pedro
Producción Máximo Cuota de Producción Máximo Cuota íctica Rendimiento captura íctica Rendimiento de captura
(kg ha-1 año-1) Sostenido (peces/día) (kg ha-1 año-1) Sostenido (peces/día)(kg ha-1 año-1) (kg ha-1 año-1)
O. mykiss 94 33.7 32 63 21.7 50O.bonariensis 269 123.5 36 159 114.3 96P. trucha - - - 33 12.5 45C.carpio - - - 65 21.5 s/r
TOTAL 384 157 68 364 170 191
s/r: sin restricción

presentes considerando que estas son las conse-cuencias de las interacciones en el tiempo que sehan dado con los ecosistemas terrestres. Esto esposible por: a) la información limnológica basedisponible y generada, b) el conocimiento que seha logrado últimamente sobre los sistemas terres-tres y c) por la evaluación de la magnitud de lasactividades humanas efectuadas en ambos siste-mas. De lo anterior resulta importante definir unmarco de referencia respecto a los puntos indica-dos lo cual facilitará el entendimiento de algunasrelaciones causa-efecto postuladas.
1. El sistema de lagos nahuelbutanos compartenen gran medida importantes característicasnaturales básicas relacionadas con la geolo-gía, climatología, geomorfología, hidrografía,cobertura vegetal pretérita y actual.
2. Al mismo tiempo, producto de las relacionesque emergen entre el sistema geomórfico y elcuerpo de agua, se generan particularidadesde cada ecosistema acuático, determinadaspor relaciones cuantitativas como por ejem-plo, las relaciones entre las superficies de lasáreas de drenaje y la de los cuerpos de aguas,el desarrollo de la línea de costa, tiempo derenovación de las aguas, que a su vez deter-minan algunas de las características limnoló-gicas (estratificación térmica, concentraciónde nutrientes) y el grado de impacto de algu-nas acciones humanas (contaminación orgá-nica, concentración de coliformes, modifica-ción de sustratos de fondo).
3. Las principales acciones humanas efectuadasen el sistema son:• Remoción del bosque nativo en las áreas de
drenaje y su reemplazo por actividad agrí-cola y/o en mayor parte por plantacionesforestales (Pinus radiata y Eucalyptus spp.).
• Establecimiento de asentamientos huma-nos, actividades turísticas y recreativas, yconstrucción de caminos civiles y foresta-les afectando la configuración de pendien-tes cerca de los lagos.
• Introducción de especies de peces (Cyprinuscarpio, Carassiuss carassius, Odontesthesbonariensis, Oncorhynchus mykis).
• Introducción de especies de malezas acuá-ticas (e.g. Egeria densa ).
A lo anterior se debe agregar que ninguno delos sistemas acuáticos y sus respectivas cuencasse encuentran sometidos a un régimen de protec-ción especial y prácticamente todas sus super-ficies son de carácter productivo y de desarro-llo indígena (áreas de los Lagos Lanalhue yLleulleu), lo que explica el alto nivel de interven-ción ha que ha sido sometido el sistema.
Como se indicó en los resultados, durantegran parte del presente siglo, el área de empla-zamiento del sistema de lagos nahuelbutanos,ha sufrido una de las más importante modifica-ciones medio ambientales generadas desde lallegada de los colonizadores españoles; el reem-plazo del bosque nativo por plantaciones fores-tales de P. radiata y Eucalyptus spp. Este proce-so es el factor mas importante que ha producidoel paulatino incremento de la exportación desedimentos y nutrientes (Cisternas et al., 1997,1999, 2000). Esta situación también ha sido des-crita para otras áreas de la Región (Oyarzun1993, 1997). Por las características de la activi-dad forestal, los periodos de cosechas son nega-tivos, especialmente si las plantaciones se loca-lizan en pendientes altas. Según los mismosautores, mas que las plantaciones forestales porsi mismas, aparentemente es negativa la locali-zación adoptada, y más aún la aplicación de latala rasa como método de cosecha. La historia“ambiental” propuesta para laguna Chica de SanPedro y aplicable a todos las cuencas de drenajedel sistema de lagos nahuelbutanos consiste en:i) fase de degradación antrópica del bosquenativo, caracterizado principalmente por tala yquemas, para realizar siembras; ii) fase de pre-paración del suelo para las primeras plantacio-nes forestales y finalmente, iii) fase de activoproceso de crecimiento urbano desarrollado conmayor intensidad en las cuencas cercanas a laconurbación de Concepción.
Todo lo anterior indica que durante los últi-mos 50 años las cuencas hidrográficas localiza-das en el sector norte de la Cordillera deNahuelbuta han sido modificadas entre un 50 a
Caracterización de cinco lagos costeros chilenos 77

un 90% de sus superficies, lo que en el transcur-so del tiempo ha significado modificación delos gradientes de pendientes, incremento y alte-raciones en los patrones de sedimentación yaportes de nutrientes a los cuerpos acuáticos,situación que se mantiene hasta hoy día.
La circulación de materia orgánica de origenterrestre en los lagos produce una considerableproducción de nutrientes inorgánicos y pasa aconstituir la base de varias cadenas tróficas a par-tir del detritus. En el área de estudio se han esti-mado aportes de sedimentos entre 0.10 a 1.16 tmha-1 por año-1 (Cisternas et al., 2000). Así una delas consecuencias primaria del corte de los bos-ques son los cambios en las tasas de sedimenta-ción. Después de la remoción de la cubierta vege-tal, los suelos pueden permanecer descubiertospor meses (5-6 meses hasta un año). Todo loanterior explicaría los cambios en la composiciónquímica de los sedimentos indicados por Urrutiaet al. (2000) y Valdovinos y Figueroa (2000). Porotra parte las modificaciones en la coberturavegetal produce también alteración en la pen-diente modificando los patrones de escorrentía yde erodabilidad afectando también la entrada desedimentos y nutrientes afectando las caracterís-ticas de los sustratos litorales para invertebrados.
Otro efecto de las plantaciones tanto de Pinuscomo de Eucalyptus es la homogenización deldetritus vegetal, muy diferente al producido porcoberturas vegetales nativas, lo que producecambios en la composición iónica de las aguas(Tundisi et al., 1997).
La influencia humana ha sido mayor en loslagos Grande y Chica de San Pedro y en el LagoLanalhue por las influencia de áreas urbanas deConcepción y Contulmo respectivamente. Loanterior ha significado por algún tiempo la entra-da de aguas lluvias y aguas servidas, la modifica-ción de sus pendientes por la construcción decaminos y accesos públicos y una fuerte altera-ción de la línea de costa por infraestructura turís-tica y recreativa. Es justamente en los dos prime-ros lagos donde se han introducido malezasacuáticas dominadas por Egeria densa, que hastahoy cubren toda la zona litoral de los lagos. Otrosefectos fueron sobre la calidad del agua, particu-
larmente en aquellos parámetros relacionadoscon la calidad microbiológica, la transparencia,las concentraciones de nutrientes, la presencia defloraciones algales, y la presencia de indicadoresalgales y bentónicos de incremento de la trofía.
Si se comparan los resultados de la clasifica-ción de la calidad de agua de los lagos, con varia-bles de respuesta biológica a los procesos deeutroficación, tales como indicadores fito-planctónicos (Parra et al., 1999), bentónicos(Valdovinos et al., 2000) y fauna íctica (Scaso yCampos, 1998; 1999), se observan claras diferen-cias, las cuales pueden ser atribuidas al estadotrófico de estos cuerpos de aguas. En los lagosque presentan una menor concentración denutrientes y mejor calidad de agua, e.g. LagoLleulleu y la Laguna Chica de San Pedro, laabundancia fitoplanctónica es de moderada abaja (<150 000 cél./L). Esta condición es con-gruente con algunas características de las comu-nidades bióticas que son indicadores de buenacalidad de agua, típicas de lagos oligo- a me-sotróficos, tales como la elevada relaciónBacillariophyceae / Chlorophyceae (>8). Conrespecto a las comunidades de macroinvertebra-dos bentónicos, se observa una gran abundanciade bivalvos suspensívoros tales como Diplodonchilensis (Bivalvia: Hyriidae), lo cual de igualforma sugiere condiciones de buena calidadde agua. Lo mismo ocurre con la presencia deSalmo trutta en el lago Lleulleu, especies salmo-nídea de altos requerimientos de calidad de agua.
Por el contrario, en los lagos que presentanuna mayor trofía y calidad de agua inferior, talescomo Laguna Grande de San Pedro y el LagoLanalhue, la abundancia fitoplanctónica es ele-vada (>400 000 cél./L) y la comunidad poseeindicadores de moderada a mala calidad de agua,típicos de lagos meso- eutróficos tales como labaja proporción Bacillariophyceae / Chloro-phyceae (<8), y la presencia de Microcystisaeruginosa y M. wesembergii y desarrollo defloraciones algales. En relación a las comunida-des de macroinvertebrados bentónicos, se obser-va la presencia de bioindicadores de regular cali-dad de agua tales como isópodos del géneroAsellus y ácaros Oribatuloidea que también son
78 Parra et al.

bioindicadores de moderada a mala calidad deagua. En los peces, son abundantes especiesgeneralistas y tolerantes tales como Cyprinuscarpio y Carassius carassius.
El análisis comparativo de los parámetrosasociados a la comunidad fitoplanctónica de loscinco lagos, demostró que cada uno de ellospresenta una comunidad particular, con diferen-cias notables entre ellos y por lo tanto, no existeun patrón que los asimile. En ninguno de loslagos estudiados en periodo del año 1997 sedetectaron floraciones algales o abundanciasrelevantes de especies indicadoras de contami-nación orgánica. Se postula, sobre la base de lacomposición específica, especies más frecuen-tes y más abundantes, así como la densidad totaldel fitoplancton, la siguiente clasificación trófi-ca: oligotrofía para el lago Lleulleu, una meso-trofía a una ligera eutrofía para la laguna Chicade San Pedro, eutrofía para Laguna Grande yLanalhue y una condición trófica particular, quecorrespondería a una distrofía producto delmaterial particulado orgánico y probablementeuna importante concentración de material húmi-co, para laguna Quiñenco.
Laguna Quiñenco posee una condición demoderada calidad de agua, sin embargo, a dife-rencia de las Laguna Grande de San Pedro y elLago Lleulleu, posee una baja abundancia fito-planctónica (<80 000 cél./L), a pesar de poseersignificativas concentraciones de nutrientes. Estose debería a que el factor limitante de la comuni-dad fitoplanctónica sería la baja penetración de laluz, asociada fundamentalmente a la elevadacarga de seston y al color, debido a la presenciade ácidos húmicos, por lo cual corresponde a untípico lago distrófico (Wetzel, 1983). Por el con-trario, la comunidad de macroinvertebrados ben-tónicos está compuesta por taxa indicadores deregular a buena calidad de agua.
Lo anterior también se ajusta a los resulta-dos obtenidos sobre la tasa de consumo deoxígeno de los sedimentos (Valdovinos &Figueroa, 2000). Este parámetro se utilizacomo una medida integradora de la actividadmetabólica de las comunidades bentónicas.Considerando que la intensidad de los procesos
metabólicos de la zona profundal de un lago esdependiente de la producción de materia orgá-nica en la zona pelágica, las tasas de consumode oxígeno de los sedimentos reflejan la condi-ción tróf ica de todo el lago (Newrkla yGunatilaka, 1982). A mayor tasa de consumode oxígeno, menor es su calidad de agua. Loslagos con bajos contenidos de nutrientes ybuena calidad de agua, coinciden con los queposeen las tasas de consumo de oxígeno másbajas (Lleulleu y Chica de San Pedro). Por elcontrario, el lago con una calidad inferior,posee una elevada tasa de consumo de oxígeno(Grande de San Pedro). Laguna Quiñencoposee una condición intermedia. No se cuentacon datos de consumo de oxígeno del LagoLanalhue que puedan ser discutidos.
Los lagos pueden ser clasificados tambiéncomo oligotróficos, mesotróficos o eutróficos,basados en sus concentraciones de nutrientes oen las tasas de producción de materia orgánica(Valdovinos & Figueroa, 2000). En los lagosestudiados, asumiendo que las entradas alócto-nas de materia orgánica no tienen tanta relevan-cia como la producida en el interior de los lagos,la producción de la zona pelágica se traduce enuna significativa sedimentación de materialalóctono. De esta forma, los nutrientes asocia-dos a la materia orgánica, son removidos tempo-ralmente de la columna de agua. Estos nutrien-tes estarán nuevamente disponibles para laproducción primaria, como resultado de la acti-vidad metabólica de la comunidad bentónica.Debido a que la densidad y composición deespecies de la comunidad bentónica profundales fuertemente dependiente de la cantidad demateria orgánica producida en la zona pelágica,su actividad metabólica está ligada a la produc-tividad del lago (Hayes & MacAuley, 1959;Rybak, 1969; Margrave, 1973; Newrkla &Gunatilaka, 1982; Riise & Roos, 1997).
En el estudio de Valdovinos y Figueroa (2000),las tasas de consumo de oxígeno más elevadasfueron medidas en los sedimentos de Quiñencoseguidas por Grande. Comparativamente, bajosvalores fueron medidos en los sedimentos deChica y Lleulleu. Los resultados también mostra-
Caracterización de cinco lagos costeros chilenos 79

ron que exponiendo en laboratorio los sedimen-tos a diferentes niveles de oxígeno, se observóque el metabolismo de la comunidad bentónicase reduce al decrecer las concentraciones de oxí-geno. Eso es explicable debido a que la falta deoxígeno favorece la descomposición anaeróbicade la materia orgánica e incrementa la cantidadde metabolitos, incrementando el déficit de oxí-geno en los sedimentos (Rybak, 1969; Dechev etal., 1977; Newrkla & Gunatilaka, 1982; Riise &Roos 1997). La falta de oxígeno suficiente actúadisminuyendo las tasas de consumo bentónico deoxígeno, como fue demostrado en los cuatrolagos. Las altas pendientes en las curvas de regre-sión en Quiñenco y Grande.
Los métodos empleados en este estudio com-parativo de las tasas de consumo de oxígeno sonsimplificados y no necesariamente duplican lascondiciones ocurridas en condiciones naturales,sin embargo, a pesar de estas dificultades, estasherramientas serían útiles para la evaluacióncuantitativa del compartimento bentónico de unlago, en relación a su nivel trófico. Los resulta-dos obtenidos sugieren que los factores abióticosde la columna de agua y sedimentos, afectan laestructura comunitaria de los macroinvertebradosbentónicos. Las comunidades varían siguiendoun continuo a lo largo de un gradiente de niveltrófico, a lo largo del cual se observa una clarasucesión de especies. Los datos son consistentescon la hipótesis que señala que un incremento delnivel trófico de un lago, debido a un aumento demacronutrientes como P y N, favorece la produc-tividad fitoplanctónica generando un incrementodel contenido de materia orgánica de los sedi-mentos, lo cual afecta la estructura comunitariade los macroinvertebrados bentónicos.
En cuanto a la ictiofauna, la dispar informa-ción disponible hace difícil el reconocimientode patrones ictiofaunísticos debidos a los dife-rentes niveles de trofía. A pesar de ello, lainformación existente muestra algunas tenden-cias, tales como el aumento en el número deespecies en el sistema más eutrófico, corres-pondiente a laguna Grande. Según Scasso &Campos (1999) este hecho se debe a la mayoradición y mejor adaptación de especies exóticas
como C. carpio y C. carassius a las aguaseutróficas. Además, es posible que la mayorheterogeneidad ambiental de la zona litoral deLaguna Grande, dada por la abundancia dehidrófitas vasculares, favorezca la presencia deun mayor número de especies. A pesar de estamayor riqueza específica, la información exis-tente muestra la tendencia de Laguna Grande(eutrófica) a presentar una menor productivi-dad que Laguna Chica (mesotrófica).
Esto resulta consistente con la tendencia dela productividad íctica de sistemas leníticos aaumentar con el nivel de trofía sólo hasta cier-to límite, ya que los estados avanzados deeutrof icación presentan efectos negativossobre las poblaciones de peces (Campos,1993). Al respecto, Scasso (1996) indica que lamenor productividad de Laguna Grande puedeser explicada en parte por el déficit de oxígenodurante primavera y verano, lo cual implicaque los peces sólo utilicen los primeros9 metros de la columna de agua en dichas épo-cas. Al evitar las zonas profundas de baja oxi-genación las poblaciones de peces ven reduci-do el espacio disponible (Scasso, 1996; Scasso& Campos, 1999), lo que estaría frenando laproductividad en este sistema.
De todo lo expuesto queda de manifiesto queel grado de trofía y de calidad del agua de estesistema lacustre se relacionan con indicadoresbiológicos expresados en las diversas comuni-dades existentes y estos a su vez con el nivel deintervención de las cuencas hidrográficas.Queda todavía por avanzar en el conocimientode mayor detalle sobre estas relaciones, ya queellas permitirían predecir con mayor precisiónla incidencia de acciones humanas sobre la inte-gridad de estos ecosistemas.
AGRADECIMIENTOS
Se agradece a Yely Ambiado, Mauricio Aguayo,Ricardo Faúndes por su valiosa ayuda para lapreparación del material gráfico. Esta investiga-ción fue financiada por el Proyecto FON-DECYT 196-0600.
80 Parra et al.

BIBLIOGRAFÍA
AMERICAN PUBLIC HEALTH ASSOCIATION(APHA). 1980. Standard methods for the exami-nation of water and waste-water, New York.
APPLEBY, P.G., V. J. JONES & J. C. ELLIS-EVANS. 1995. Readiometric dating of lake sedi-ments from Signy Island. Journal ofPaleolimnology, 13 :179-191.
BARRIENTOS, C. 1990. Estructura dinámica de lavegetación en la hoya hidrográfica de la LagunaChica de San Pedro. Tesina de Diplomado enAnálisis y Gestión del Ambiente. Centro EULA-Chile, Universidad de Concepción, 23 pp.
BELLAIR , P. & CH. POMEROL. 1977. Elementosde Géologie. A. Collin.
BERTALANFFY, L. 1938. A quantitative theory oforganic growth inquiries on growth laws. Humanbiology, 10(2): 181-213.
CAMPOS, H., W. STEFFEN, G. AGÜERO, O.PÁRRA. & L. ZÚÑIGA, L. 1992. Limnology oflake Ranco (Chile). Limnológica, 22 (4): 337-353.
CAMPOS, H., V. H. RUIZ, J. F. GAVILÁN & F.ALAY. 1993. Peces del Río Biobío. SeriePublicaciones de Divulgación EULA, 5
CAMPOS, H. 1993. Procesos de eutroficación enlagos del sur de Chile. Estimación de los efectosde la acuicultura intensiva. SeminarioInternacional Acuicultura y Medio AmbienteFundación Chile.
CIER-SERPLAC. 1976. Atlas Regional, VIII Regióndel Biobío. Chile.
CISTERNAS, M., A. ARANEDA, O. RETAMAL &R. URRUTIA. 1997. Variaciones Históricas en lastasas de erosión-sedimentación de un cuerpolacustre antropizado: Utilización de geocronolo-gía radioisotópica. Revista de Geografía NorteGrande, 24: 151-156.
CISTERNAS, M., P. DEBELS, P. MARTÍNEZ & R.SANHUEZA. 1999a. Cambios Históricos en elUso del Suelo de una Pequeña Cuenca Lacustre deNahuelbuta. Revista Geográfica de Chile TerraAustralis, 44: 141-153.
CISTERNAS, M., P. MARTÍNEZ, C. OYARZÚN. &P. DEBELS. 1999b. Caracterización del procesode reemplazo de vegetación nativa por plantacio-nes forestales en una cuenca lacustre de laCordillera de Nahuelbuta, VIII Región, Chile.Revista Chilena de Historia Natural, 72: 661-676.
CISTERNAS, M., L. TORRES, R. URRUTIA, A..ARANEDA & O. PARRA.. 2000 Comparación
ambiental mediante registros sedimentarios entrelas condiciones prehispánicas y actuales de un sis-tema lacustre. Revista Chilena de HistoriaNatural, 73: 151-162.
CISTERNAS, M., A. ARANEDA, P. MARTINEZ &S. PEREZ. 2001. Effects of historical land use onsediment yield from a lacustrine watershed in cen-tral Chile. Earth Surface Processes andLandforms, 26(1): 63-76.
COLBY, P., G. R. SPANGLER, D. A. HURLEY & A.M. MCCOMBIE. 1972. Effects of eutrophicationon salmonid communities in oligotrophic lakes. J.Fish. Res. Bd. Can., 29: 975-983.
CORFO. 1995. Potencialidad de las aguas interioresen la zona sur de la Provincia de Arauco (comunasde Cañete a Tirúa): Lagos Lanalhue y Lleulleu.Fondos Estudios e Investigaciones CORFO.
DECHEV, G., S. YORDANOV & E. MATVEEVA.1977. Oxygen consumption and oxygen debt in bot-tom sediments. Arch. Hydrobiol. Suppl. 52: 63-71.
DELLAROSSA, V. & O. PARRA. 1985. EstadoAmbiental de la laguna Grande de San Pedro y dellago Lanalhue. Red Mínima de Control de Lagos.Convenio Dirección General de Aguas yUniversidad de Concepción, Chile. Informe Final,209 pp.
DEVYNCK, J. L. 1970. Contribución al estudio dela circulación atmosférica en Chile y clima de laregión del Biobío. Universidad de Concepción,Depto. Geofísica, 174 pp.
DI CASTRI, F. 1968. Esbozo ecológico de Chile. InBiologie de L’Amerique australe. Deboutteville. C& E. Rapoport (Eds.). Centre National de laRecherche Scientifique, 34 pp.
DOWNING, J. A., C. PLANTE & S. LALONDE.1990 Fish production correlated with primary pro-ductivity, not the morphoedaphic index. Can. J.Fish. Aquat. Sci., 47: 1929-1936.
DOWNING, J. A. & C. PLANTE. 1993 Productionof fish populations in lakes. Can. J. Fish. Aquat.Sci., 50: 110-120.
EDWARDS, R. W. & H. L. J. ROLLEY. 1965.Oxygen consumption of river muds. J. Ecol., 53:1-19.
ENDLICHER, W. & W. MACHEL. 1985. NatouralResources, land use and degradation in the coastalzone of Arauco and Nahuelbuta Range.Geojournal, 11 (1):43-64.
FERRARIS, F. 1981. Hoja Los Angeles-Angol,Región del Biobío, 1: 250.000. Mapa GeológicoPreliminar N° 5, 10 pp, Sernageomin, Santiago.
Caracterización de cinco lagos costeros chilenos 81

FERRARIS, F. & R. BONILLA. 1981. Hoja Arauco-Lebu, Región del Biobío, 1: 250.000. MapaGeológico Preliminar N° 6, 26pp, Sernageomin,Santiago.
FUENZALIDA, H. 1971. Climatología de Chile.Publicación Int. Fac. de Ciencias Físicas yMatemáticas, Universidad de Chile, 73 pp.
HARGRAVE, B. T. 1973. Coupling carbon flowthrough some pelagic and benthic communities. J.Fish. Res. Bd Canada, 30: 1317-1326.
HARGRAVE, B. T. 1976. Metabolism at the benthicboundary. In: The benthic boundary layer. I. N.McCave (ed.). Plenum Press. 232 pp.
HAYES, F. . & M. A. McAULEY, M.A. 1959. Lakewater and sediment V. Oxygen consumed in waterover sediment cores. Limnol. Oceanogr., 4: 291-298.
HUTCHINSON, G. E. 1957. A treatise on Limnology,Vol. 1. John Wiley, New York.
KATZ, H. R . 1970. Randpazifische Bruchtektonikam Beipiel Chiles und Neuseelands. GeologischeRundschau, 59: 898-926.
LARSEN, R. J., C. G. SANDERSON & J. KADA.1995. EML Surface Air Sampling Program. DataReport. ONU, 248 pp.
LEWIS, D.W. 1984. Practical Sedimentology.Hutchinson Ross Publishing company. Strowdsburg,Pennsylvania. 229 pp.
MARDONES, M. & E. JAQUE. 1996. Geomor-fología del valle del río Laja. I Taller Interna-cional de geoecología de Montaña y desarrolloSustentable de los Andes del Sur. The UnitedNations University: 271-286.
MARDONES, M. & C. D. REUTHER. 1999.Geomorphological aspects of the drainage patternarround lake Lanalhue and lake Lleulleu in theactice convergent margin setting of South-CentralChile. Mitteilungen Geologische undPaläontologisches Institut und Museum,Universität Hamburg, 83: 75-88.
MILLS, A. 1978. A comparison of methods of deter-mining carbon in marine sediments from deNational Status and Trends Program. NOAATechnical Memorandum NOS OMA 59: 112 pp.
MOSSETTI, F. 1977. Le acqua. Colección II NostroUniverso. UTET.
NEWRKLA, P. & A. GUNATILAKA. 1982. Benthiccommunity metabolism of three Austrian pre-alpi-ne lakes of different trophic conditions and itsoxygen dependency. Hydrobiologia, 92: 531-536.
NYGAARD, G. 1949. Hydrobiological studies insome ponds and lakes. II. The quotient hypothesis
and some new or little known phytoplankton orga-nisms. Biol. Skr., 7: 1-293.
OYARZUN, C. E. 1993. Evaluación del modeloU.S.L.E. para predecir pérdidas de suelo en áreasforestadas de la cuenca del río Bio-Bio. Bosque,14(1): 45-54.
OYARZUN, C. E. 1995. Land use, hydrological pro-perties, and soil erodibilities in the Bio-Bio riverbasin, Central Chile. Mountain Research andDevelopment, 5(4): 331-338.
PAMATMAT, M. M. & K. BANSE. 1969. Oxygenconsumption by the seabed - II. In situ measure-ments to a depth of 180 m. Limnol. Ocenogr., 14:250-259.
PARRA O. 1989. La eutroficación de la LagunaGrande de San Pedro: Un caso de estudio.Ambiente y Desarrollo, 5(1): 117-136.
PARRA, O., V. DELLAROSSA & E. UGARTE.1976. Estudio limnológico de las lagunas “Chicade San Pedro”, La Posada” y “Lo Méndez” I.Análisis cuali y cuantitativo del plancton invernal.Bol. Soc. Biol. Concepción, 50: 73-86.
PARRA, O., E. UGARTE, E. BALABANOFF, S.MORA, M. LIEBERMANN & A. ARON. 1980.Remarks on a bloom of Microcystis aeruginosaKuetzing. Nova Hedwigia, 33: 971-1004.
PARRA, O., E. UGARTE & V. DELAROSSA. 1981.Periodicidad estacional y asociaciones en el fito-plancton de tres cuerpos lénticos de la Región deConcepción, Chile. Gayana Botánica, 36 :1-35.
PARRA, O., M. GONZALEZ, V. DELLAROSSA, P.RIVERA. & M. ORELLANA. 1982-1983.Manual taxonómico del fitoplancton de aguascontinentales de Chile. Vol. I, 70 pp; Vol. II, 82pp.; Vol. III, 99 pp. Vol. V 353 pp.
PARRA, O., C. JARA & L. GUZMÁN. 1989. Laslagunas intraurbanas de Concepción: Estadoactual y perspectivas de recuperación y uso. Actasdel III Encuentro Nacional del Medio Ambiente,301-313.
PARRA, O., H. CAMPOS, W. STEFFENS, G.AGÜERO, S. BASUALTO, D. AVILES & M.VIGHI. 1993. Estudios limnológicos de los lagosIcalma y Galletué: Lagos de origen del río Biobío(Chile Central). Monografía Científicas EULA, Ed.Universidad de Concepción, Chile. 12: 161-188.
PARRA, O., C. VALDOVINOS & A. FIGUEROA.1998. Caracterización física y química de 5 lagosde Chile Central. Gayana, (in press).
PARRA O., S. BASUALTO, R. URRUTIA & C.VALDOVINOS. 1999. Estudio comparativo de la
82 Parra et al.

diversidad fitoplanctónica de cinco lagos de dife-rentes niveles tróficos. Gayana. 56(2) :25-40.
PIÑONES, O. & I. TOMICIC. 1995. Estudio de losniveles radiológicos-ambientales en Chile duranteel periodo 1966-1994. Nucleotécnica, 29:67-82.
PROCHELLE, O. & H. CAMPOS. 1985. The biologyof the introduced carp Cyprinus carpio L., in theriver Cayumapu, Valdivia, Chile. Studies on Neotro-pical Fauna and Environment., 20(29): 65-82.
RICKER, W.E. 1981. Computation and interpreta-tion of biological statistics of fish populations.Bull. Fish. Res. Bd. Canada:191-382.
RIISE, J. C. & N. ROOS. 1997. Benthic metabolismand the effects of bioturbation in a fertilised poly-culture fish pond in Northeast Thailand.Aquaculture, 150(1-2): 45-62.
RIVERA, P., O. PARRA, M. GONZÁLEZ, V. DEA-LLAROSSA & M. ORELLANA. 1983. ManualTaxonómico del Fitoplancton de AguasContinentales. Editorial Universidad deConcepción, Vol. IV, Bacillariophyceae. 97 pp.
ROSENBERG, D. M. & V. H. RESH. 1993.Freshwater biomonitoring and benthic macroin-vertebrates. Chapmann & Hall.
RYBAK, J. I. 1969. Bottom sediments of lakes ofvarious trophic type. Ecol. Pol., 17: 611-662.
RYDING, S. & W. RAST. 1992. El control y de laeutrofización en lagos y pantanos. UNESCO, Ed.Pirámide, Madrid.
SCASSO, F. 1996. Productividad íctica en lagos dediferente estado trófico: recomendaciones de con-servación para pesca deportiva. Tesis de GradoCentro EULA-Chile Universidad de Concepción.
SCASSO, F. & H. CAMPOS. 1998. Oncorhynchusmykiss (Pisces, Salmonidae) populations in lakesof different trophic levels of the Biobío riverbasin, Chile. Verh. Internat. Verein. Limnol., 26:2320-2323.
SCASSO, F. & H. CAMPOS. 1999. Comparison oftwo populations of silverside (Odontesthes bona-riensis) in Eutrophic lakes of Central Chile.Journal of Freshwater Ecology, 14(1): 61-70.
SMITH, K. L. 1973. Respiration of a sublittoralcommunity. Ecology, 54: 1065-1075.
STRICKLAND, J. D. H. & PARSONS, T. R. 1978. Apractical handbook of seawater analysis. Bull.Fish. Res. Bd Canada, 167.
TUNDISI, J. G., Y. SAIJO & T. SUNAGA. 1997.Ecological effects of Human Activities in theMiddle Rio Doce Lakes. In: Limnological Studieson the Rio Doce Valley Lakes, Brazil. J. G. Tundisi& Y. Saigo (eds.).
URRUTIA, R., K. SABBE, F. CRUCES, K. POZO, J.BECERRA, A. ARANEDA, W. VYVERMAN &O. PARRA. Paleolimnological studies of LagunaChica of San Pedro (VIII Region): Diatom, hydro-carbons and fatty acid records. Revista Chilena deHistoria Natural, 73:593-604. 2000.
URRUTIA, R., M. CISTERNAS, A. ARANEDA, O.RETAMAL, O. PARRA & M. MARDONES.2000. Caracterización morfométrica y sedimento-lógica de cinco lagos costeros de la VIII Región,Chile. Revista Geográfica de Chile TerraAustralis, 45:7-24.
UTERMOHL, H. 1957. Zur Vervollkommung derquantitativen Phytoplankton Methodik. Mitt. Int.Vereinigung Theor. Angew. Limnol., 15: 158-163.
VALDOVINOS, C. & A. R. FIGUERO. 2000.Benthic community metabolism and trophic con-ditions of four South American lakes.Hydrobiologia, 429: 151-156.
VALDOVINOS. C., O. PARRA, R. FIGUEROA, R.URRUTIA & M. CISTERNAS. 2000.Clasificación de la Calidad de las Aguas de CincoSistemas Lacustres de Chile Central sometidos adistintos niveles de intervención Humana (envia-da).
VEYL, C., 1961. Contribución al conocimiento de laGeología regional de la provincia de Concepción.Inst. de Química. Geología. Universidad deConcepción. Publicación interna.
VILA, I. & D. SOTO. 1984. Odontesthes bonarien-sis “pejerrey argentino”, una especie para culti-vo extensivo. FAO Documento técnico, 4: 224 -228.
VOLLENWEIDER, R. A. 1968. Scientifical funda-mentals of the eutrophication of lakes and flowingwaters, with particular reference to nitrogen andphosphorus as factors in eutrophication. Rep.Organis. Econ. Coop. And Dev., DAS/CSI/68.27.Paris.
WENTWORTH, C. K. 1922. A scale of grade andclass terms for clastic sediments. J. Geol., 30:377-392.
Caracterización de cinco lagos costeros chilenos 83


INTRODUCCIÓN
Los embalses constituyen una discontinuidad enla dinámica fluvial. Por este motivo, el agua alsalir de un embalse presenta características físi-cas, químicas y biológicas diferentes a las queposeía aguas arriba del mismo. En particular se hamodificado un parámetro de gran importancia enla dinámica de las comunidades fluviales y en lacalidad del agua en los embalses: la temperatura.
La gran inercia térmica que presenta un embal-se provoca que aguas abajo el río altere subs-tancialmente su comportamiento térmico. Asípor ejemplo, en verano el agua que sale delembalse, situado en una región de clima medi-terráneo, puede presentar una temperatura nota-blemente inferior a la que tiene el río aguasarriba del mismo. Por tal motivo, aguas abajode la presa el río incrementa la temperatura a lolargo de su circulación. Este incremento tiende
85
Incidencia de los embalses en el régimen térmico del río. El caso del tramo final del río Ebro
Rafael Val1. Daniel Niñerola1. Joan Armengol2. José Dolz1
1Departament d’Enginyeria Hidràulica, Marítima i Ambiental, E.T.S.E.C.C.P. Universidad Politécnica deCatalunya. C/J. Girona, 1-3, D1.08034-Barcelona2Departament d’Ecologia. Facultat de Biologia. Universidad de Barcelona. Avd. Diagonal, 645. 08028-Barcelona
RESUMEN
El agua del río mantiene un intercambio de calor con la atmósfera y con el fondo; este proceso puede lograr su equilibrio alo largo de un tramo de río con características geológicas y climáticas similares. Su comportamiento puede ser modificadopor efectos artificiales; tales como el uso del agua del río para refrigeración de centrales termoeléctricas o por la existenciade embalses.En el caso de regiones con clima mediterráneo, donde se encuentran los embalses aquí estudiados, los efectos de las altera-ciones en el régimen térmico producido por éstos, provocan una tendencia a la constancia térmica estacional, un aumentode la uniformidad diaria, una elevación de las temperaturas invernales y la disminución de las temperaturas estivales. Latemperatura del agua es un factor importante en la ecología del río, ya que condiciona en gran medida la vida de las comu-nidades fluviales. Aguas abajo de las termoeléctricas o presas, la temperatura del agua evoluciona de tal forma que tiende alograr el equilibrio con el medio ambiente.
Palabras clave: balance térmico, radiación solar, embalse, clima mediterráneo.
ABSTRACT
The river water maintains a heat exchange with the atmosphere and with the bottom; this process can reach its equilibriumalong a stretch of the river with similar geologic and climatic characteristics. Its behavior can be modified by artificialeffects, such as the use of the river water for the cooling of thermal power stations or by the presence of reservoirs.In the case of regions with Mediterranean climate, where the studied reservoirs are located, the effects of changes in thenatural thermal regime caused by them produce a trend towards a constant seasonal thermal condition, an increase in dailyuniformity, an increase in winter temperatures, and a decrease in summer temperatures. Water temperature is an importantfactor in the river’s ecology, because it greatly conditions the life of fluvial communities. Downstream from power stationsor dams, the water temperature evolves in such a way that tends to achieve equilibrium with the environment.
Keywords: heat budget, solar radiation, long wave radiation, reservoir, Mediterranean climate.
Limnetica 22(1-2): 85-92 (2003)© Asociación Española de Limnología, Madrid. Spain. ISSN: 0213-8409

a restablecer el equilibrio térmico entre el río ysu entorno (atmósfera y cauce) alterado por elembalse.
Este fenómeno se presenta en el río Ebro,tomando especial relieve la gran influencia que elsistema de embalses existente aguas arriba de laCentral Nuclear de Ascó (sistema Mequinenza –Ribarroja – Flix) tiene en la temperatura del río.Esta influencia, más acusada en unas estacionesque en otras, condiciona fuertemente el equili-brio térmico del río. A lo largo de la presentecomunicación se presentaran datos y reflexionesen este sentido.
DESCRIPCIÓN DE LA ZONA EN ESTUDIO
La Central Nuclear de Ascó se asienta en la mar-gen derecha del río Ebro, a la altura de la po-
blación que le da nombre, cinco kilómetrosaguas abajo de la central hidroeléctrica de Flix.
El río tiene un importante núcleo de regulaciónexactamente en la zona aguas arriba de la C. N.Ascó; así, si bien la central de Flix cuenta con unareducida capacidad de embalse (del orden de11 hm3), inmediatamente aguas arriba se encuen-tra la central de Ribarroja, con una capacidad deembalse de 210 hm3 y en su cola se encuentra elembalse de Mequinenza, con 1534 hm3 de capa-cidad y verdadero centro de regulación del tramobajo del río Ebro (figura 1). La media anual de laaportación total (Ebro más Segre y Cinca) enRibarroja es de 18018 hm3.
Teniendo en cuenta los volúmenes de embal-se y las aportaciones naturales, observamos queel tiempo de residencia medio del agua seráprácticamente despreciable en Flix y muchomayor en Mequinenza.
86 Val et al.
Figura 1. Localización de la zona en estudio. Aportación natural de los ríos y capacidad de almacenamiento de los embalses, (1) RíoEbro (11491 hm3/año), (2) Río Segre (2915 hm3/año), (3) Río Cinca (3438 hm3/año), (4) Embalse de Mequinenza (1534 hm3),(5) Embalse de Ribarroja (210 hm3) (6) Embalse de Flix (11 hm3). La longitud del río estudiado(7) tiene 5.3 km. Location of thestudy area. Annual flow of the rivers and reservoirs’ volume capacity: (1) Ebro River (11 491 hm3/y), (2) Segre River (2915 hm3/y),(3) Cinca River (3438 hm3/y), (4) Mequinenza reservoir (1534 hm3), (5) Ribarroja reservoir (210 hm3), (6) Flix reservoir (11 hm3).The length of the studied river (7) is 5.3 km.
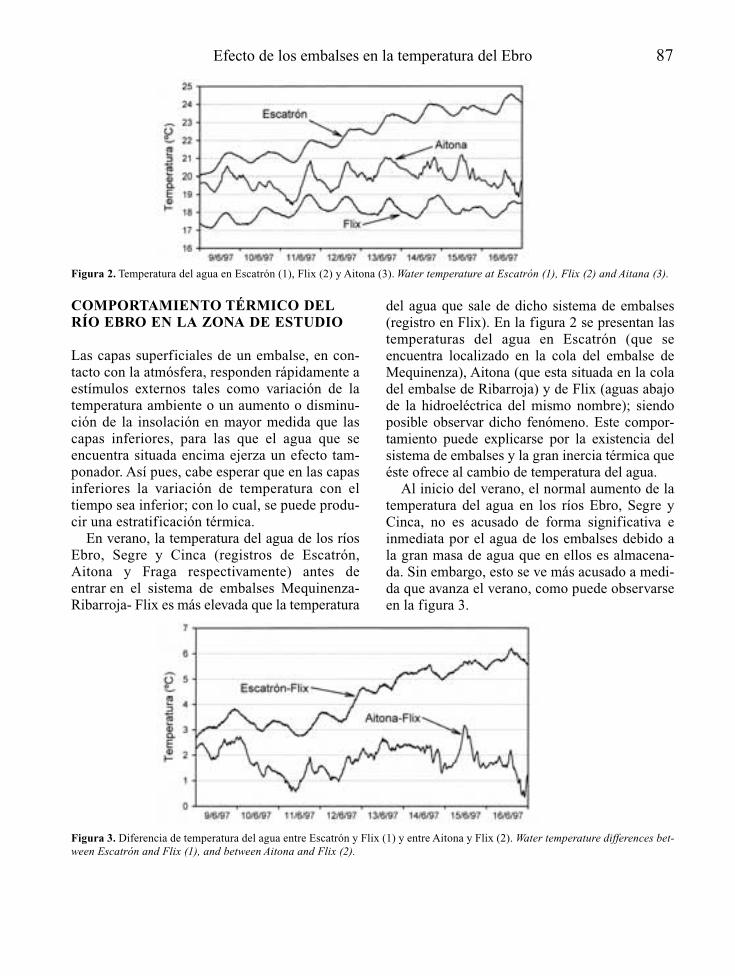
COMPORTAMIENTO TÉRMICO DELRÍO EBRO EN LA ZONA DE ESTUDIO
Las capas superficiales de un embalse, en con-tacto con la atmósfera, responden rápidamente aestímulos externos tales como variación de latemperatura ambiente o un aumento o disminu-ción de la insolación en mayor medida que lascapas inferiores, para las que el agua que seencuentra situada encima ejerza un efecto tam-ponador. Así pues, cabe esperar que en las capasinferiores la variación de temperatura con eltiempo sea inferior; con lo cual, se puede produ-cir una estratificación térmica.
En verano, la temperatura del agua de los ríosEbro, Segre y Cinca (registros de Escatrón,Aitona y Fraga respectivamente) antes deentrar en el sistema de embalses Mequinenza-Ribarroja- Flix es más elevada que la temperatura
del agua que sale de dicho sistema de embalses(registro en Flix). En la figura 2 se presentan lastemperaturas del agua en Escatrón (que seencuentra localizado en la cola del embalse deMequinenza), Aitona (que esta situada en la coladel embalse de Ribarroja) y de Flix (aguas abajode la hidroeléctrica del mismo nombre); siendoposible observar dicho fenómeno. Este compor-tamiento puede explicarse por la existencia delsistema de embalses y la gran inercia térmica queéste ofrece al cambio de temperatura del agua.
Al inicio del verano, el normal aumento de latemperatura del agua en los ríos Ebro, Segre yCinca, no es acusado de forma significativa einmediata por el agua de los embalses debido ala gran masa de agua que en ellos es almacena-da. Sin embargo, esto se ve más acusado a medi-da que avanza el verano, como puede observarseen la figura 3.
Efecto de los embalses en la temperatura del Ebro 87
Figura 2. Temperatura del agua en Escatrón (1), Flix (2) y Aitona (3). Water temperature at Escatrón (1), Flix (2) and Aitana (3).
Figura 3. Diferencia de temperatura del agua entre Escatrón y Flix (1) y entre Aitona y Flix (2). Water temperature differences bet-ween Escatrón and Flix (1), and between Aitona and Flix (2).
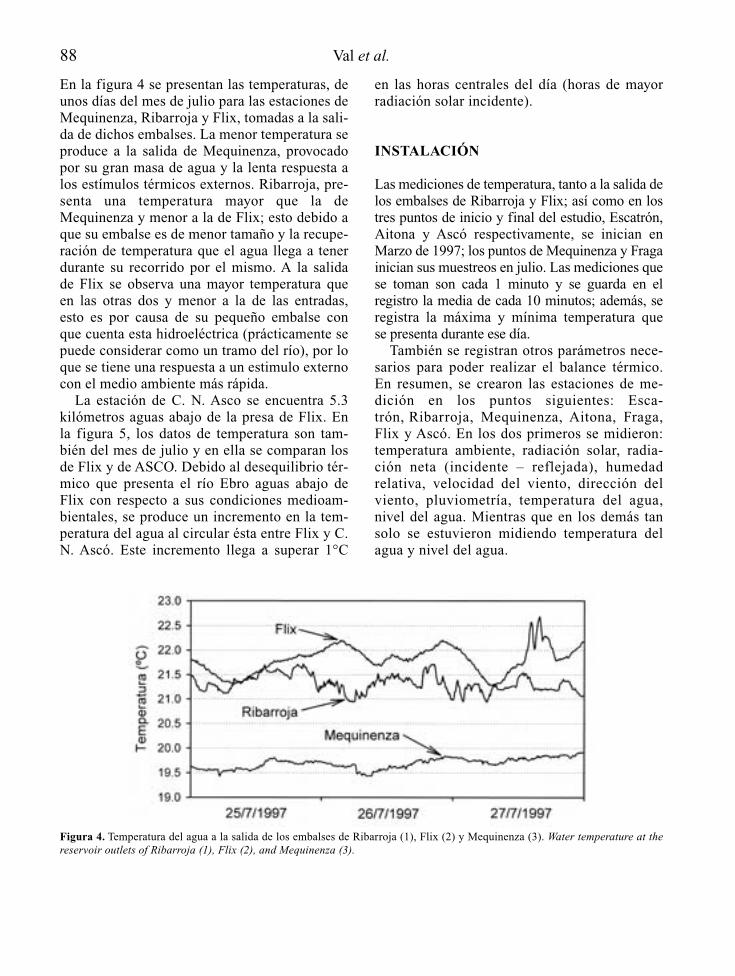
En la figura 4 se presentan las temperaturas, deunos días del mes de julio para las estaciones deMequinenza, Ribarroja y Flix, tomadas a la sali-da de dichos embalses. La menor temperatura seproduce a la salida de Mequinenza, provocadopor su gran masa de agua y la lenta respuesta alos estímulos térmicos externos. Ribarroja, pre-senta una temperatura mayor que la deMequinenza y menor a la de Flix; esto debido aque su embalse es de menor tamaño y la recupe-ración de temperatura que el agua llega a tenerdurante su recorrido por el mismo. A la salidade Flix se observa una mayor temperatura queen las otras dos y menor a la de las entradas,esto es por causa de su pequeño embalse conque cuenta esta hidroeléctrica (prácticamente sepuede considerar como un tramo del río), por loque se tiene una respuesta a un estimulo externocon el medio ambiente más rápida.
La estación de C. N. Asco se encuentra 5.3kilómetros aguas abajo de la presa de Flix. Enla figura 5, los datos de temperatura son tam-bién del mes de julio y en ella se comparan losde Flix y de ASCO. Debido al desequilibrio tér-mico que presenta el río Ebro aguas abajo deFlix con respecto a sus condiciones medioam-bientales, se produce un incremento en la tem-peratura del agua al circular ésta entre Flix y C.N. Ascó. Este incremento llega a superar 1°C
en las horas centrales del día (horas de mayorradiación solar incidente).
INSTALACIÓN
Las mediciones de temperatura, tanto a la salida delos embalses de Ribarroja y Flix; así como en lostres puntos de inicio y final del estudio, Escatrón,Aitona y Ascó respectivamente, se inician enMarzo de 1997; los puntos de Mequinenza y Fragainician sus muestreos en julio. Las mediciones quese toman son cada 1 minuto y se guarda en elregistro la media de cada 10 minutos; además, seregistra la máxima y mínima temperatura quese presenta durante ese día.
También se registran otros parámetros nece-sarios para poder realizar el balance térmico.En resumen, se crearon las estaciones de me-dición en los puntos siguientes: Esca-trón, Ribarroja, Mequinenza, Aitona, Fraga,Flix y Ascó. En los dos primeros se midieron:temperatura ambiente, radiación solar, radia-ción neta (incidente – reflejada), humedadrelativa, velocidad del viento, dirección delviento, pluviometría, temperatura del agua,nivel del agua. Mientras que en los demás tansolo se estuvieron midiendo temperatura delagua y nivel del agua.
88 Val et al.
Figura 4. Temperatura del agua a la salida de los embalses de Ribarroja (1), Flix (2) y Mequinenza (3). Water temperature at thereservoir outlets of Ribarroja (1), Flix (2), and Mequinenza (3).
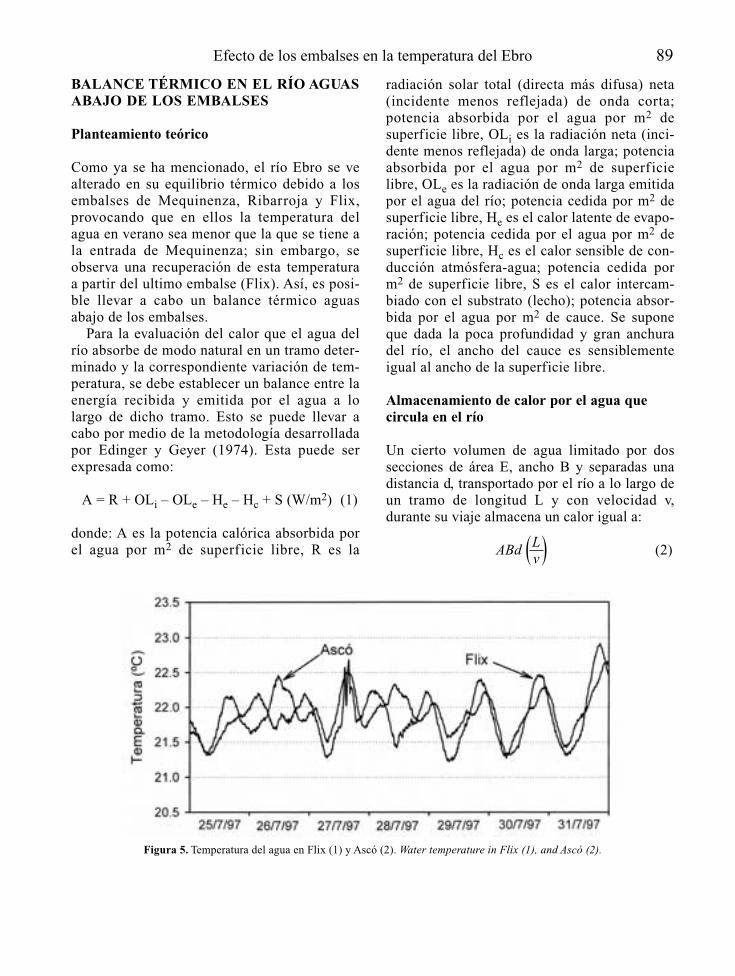
BALANCE TÉRMICO EN EL RÍO AGUASABAJO DE LOS EMBALSES
Planteamiento teórico
Como ya se ha mencionado, el río Ebro se vealterado en su equilibrio térmico debido a losembalses de Mequinenza, Ribarroja y Flix,provocando que en ellos la temperatura delagua en verano sea menor que la que se tiene ala entrada de Mequinenza; sin embargo, seobserva una recuperación de esta temperaturaa partir del ultimo embalse (Flix). Así, es posi-ble llevar a cabo un balance térmico aguasabajo de los embalses.
Para la evaluación del calor que el agua delrío absorbe de modo natural en un tramo deter-minado y la correspondiente variación de tem-peratura, se debe establecer un balance entre laenergía recibida y emitida por el agua a lolargo de dicho tramo. Esto se puede llevar acabo por medio de la metodología desarrolladapor Edinger y Geyer (1974). Esta puede serexpresada como:
A = R + OLi – OLe – He – Hc + S (W/m2) (1)
donde: A es la potencia calórica absorbida porel agua por m2 de superficie libre, R es la
radiación solar total (directa más difusa) neta(incidente menos reflejada) de onda corta;potencia absorbida por el agua por m2 desuperficie libre, OLi es la radiación neta (inci-dente menos reflejada) de onda larga; potenciaabsorbida por el agua por m2 de superficielibre, OLe es la radiación de onda larga emitidapor el agua del río; potencia cedida por m2 desuperficie libre, He es el calor latente de evapo-ración; potencia cedida por el agua por m2 desuperficie libre, Hc es el calor sensible de con-ducción atmósfera-agua; potencia cedida porm2 de superficie libre, S es el calor intercam-biado con el substrato (lecho); potencia absor-bida por el agua por m2 de cauce. Se suponeque dada la poca profundidad y gran anchuradel río, el ancho del cauce es sensiblementeigual al ancho de la superficie libre.
Almacenamiento de calor por el agua quecircula en el río
Un cierto volumen de agua limitado por dossecciones de área E, ancho B y separadas unadistancia d, transportado por el río a lo largo deun tramo de longitud L y con velocidad v,durante su viaje almacena un calor igual a:
ABd ( ) (2)Lv
Efecto de los embalses en la temperatura del Ebro 89
Figura 5. Temperatura del agua en Flix (1) y Ascó (2). Water temperature in Flix (1), and Ascó (2).

deberá verificarse la igualdad:
ABd ( ) = ∆TdEρCe (3)
de donde:
A = = (4)
donde ∆T es el incremento de temperatura delagua en su viaje a lo largo del tramo (ºC), Q es elcaudal circulante (m3/s), Ce es el calor específicodel agua (1Kcal/ºCKg), ρ es la densidad del agua(1000 kg/m3), L es la longitud del tramo estudia-do (m), B es el ancho eficaz del río (m) es elancho asociado al agua en movimiento y no inclu-ye el agua inmovilizada junto a las márgenes.
Radiación solar total neta de onda corta
El valor de la radiación solar total neta de ondacorta sobre la superficie del río, diferencia entrela radiación incidente y la reflejada. Esta ha sidomedida directamente.
Radiación neta de onda larga absorbida
La radiación atmosférica incidente de ondalarga se calcula mediante la ley de Stefan-Boltzmann. Tomando en cuenta los valores delos parámetros físicos involucrados, esto seexpresa como:
OLi = 5.14 10-13 (TA)6 (W/m2 ) (5)
donde TA es la temperatura del aire (K), medidadirectamente.
Radiación de onda larga emitida
La radiación de onda larga emitida a la atmósfe-ra por el río puede asimismo evaluarse con laley de Stefan-Boltzmann:
OLe = 5.49 10-8 ( Ts )4 (W/m2) (6)
donde Ts es la temperatura de la superficie delagua (K), medida directamente.
Pérdida de calor por evaporación
Tomando en cuenta la expresión obtenida porSeco y Garmendia (1973)
He = 1.49 ( es – ed ) U0.76 ( W/m2) (7)
donde es es la tensión de vapor de saturación delaire a la temperatura del agua (mm Hg), ed es latensión de vapor del aire (mm Hg), U es la velo-cidad del viento (m/s), medida directamente.
Pérdida de calor por conducción
El calor sensible intercambiado por conducciónentre el agua y el aire se obtiene mediante larelación de Bowen, entre el calor cedido porconducción y el calor latente de evaporación
Hc = 0.6917 ( TA – Ts ) U0.76 (W/m2) (8)
Calor intercambiado con el lecho
El agua también intercambia calor con el lecho.La importancia de este supuesto intercambio decalor ha sido puntualizada por Fristchen y VanBavel (1963). Este intercambio consiste enalmacenar el calor en las horas de fuerte radia-ción solar y cederlo al agua durante la noche. Elflujo de calor entre el agua y el lecho afecta alos estratos que se encuentran situados por enci-ma de una cierta profundidad.
Si consideramos el aporte neto diario delsubstrato, al ser las temperaturas del río sensi-blemente cíclicas a lo largo del día, cabe esperarque tienda anularse. Los aportes horarios, noobstante, no son despreciables, y si se realiza unbalance energético con esta frecuencia de mues-treo, deben ser tenidos en cuenta.
Ajuste de parámetros en el balance energético
La ecuación de almacenamiento de calor, des-crita anteriormente, esta dada como
A = (9)∆TρQCe
LB
∆TρQCeLB
∆TρEvCeLB
Lv
90 Val et al.

Agrupando los términos que son conocidos, setiene que
A = (10)
Así, es posible que la ecuación de almacena-miento de calor este en función del ancho efec-tivo del río, B; de tal forma se tiene que
A = (11)
Por otra parte, se sabe que el balance de ener-gía esta dado por
A = R + OLi – OLe – He – Hc + S (W/m2) (12)
de la cual, casi todos los valores son conocidosa excepción del calor intercambiado con ellecho, S; de ahí que
A = C+ S (W/m2) (13)
donde
C = R + OLi – OLe – He – Hc (W/m2) (14)
Así, dadas un conjunto de medidas con fre-cuencias horarias, se puede establecer la ecua-ción del balance energético, teniendo así unconjunto de expresiones del tipo
= Ci + Si i = 1 ... 24 (15)
donde: Ki es el valor conocido para cada hora, Bes el ancho eficaz del río Ci es el parámetroconocido.
Si es el calor intercambiado con el lecho, 1...24representa el conjunto de medidas que cubren undía completo, en el que, como se ha comentado,la suma de todos los Si debe ser nula
i=1Σ24
Si = 0 (16)
Realizando el balance energético a lo largo deun día completo, debe verificarse
i=1Σ24
= i=1Σ24
Ci (17)
por lo que
B = i=1Σ24
Ki / 1Σ24
Ci (18)
permite ajustar el ancho eficaz del río, conindependencia de Si. Este procedimiento esválido para series de 24, 48, etc. y en generalun número de horas tal que comprenda un nú-mero entero de días.
Conocido el valor de B, se tiene
Si = * Ci (19)
RESULTADOS
Con anterioridad al estudio que actualmente seesta realizando, se llevo a cabo un primer balan-
KiB
KiB
KiB
KB
∆TρQCeL
Efecto de los embalses en la temperatura del Ebro 91
Figura 6. Balance energético a lo largo de un día. Daily energy budget.

ce energético en un tramo de 5.3 km. entre Flixy ASCO con un conjunto de datos correspon-dientes al mes de agosto de 1990.
Al aplicar la ecuación de balance de acuerdocon lo expuesto anteriormente, fue posibletener unos primeros resultados; de loscuales se obtiene que el ancho eficaz del ríovale B=120 m. Teniendo presente que de losplanos topográficos (1:5 000) se valoraba en140m; sin embargo, en esta medida se tomabaen cuenta la zona de agua inmovilizada junto alas márgenes.
En la figura 6, se muestra el resultado delbalance energético de uno de los días para esteperiodo. En él, se refleja la acumulación decalor de agua (A), la radiación de onda cortaneta recibida (R), el aporte del calor del lecho(S), el balance de radiación de onda larga recibi-da y emitida (OLi – OLe) y el balance entre elcalor latente de evaporación y el calor sensiblede conducción (He – Hc).
En la gráfica se puede observar que el valorde la radiación de onda corta es quien juegauno de los papeles importantes en el balanceglobal junto con los términos de acumulaciónde calor por el agua y de incremento decalor con el lecho. Los balances de onda largay de evaporación-conducción prácticamente notienen incidencia.
La aportación neta diaria del lecho es prácti-camente nula, como ya se había comentado enel planteamiento teórico; por lo que el caloracumulado por el agua es casi igual al recibidopor la radiación de onda corta. Sin embargo, ellecho juega un papel regulador, de modo queabsorbe calor cuando el agua tiende a calentarse(horas centrales del día) y cederlo cuando tiendea enfriarse (durante la noche).
CONCLUSIONES
A continuación se presentan las conclusionesmás importantes de este estudio:
1. Como se ha visto, en verano y en climamediterráneo, los datos reportan un enfria-
miento del agua por efecto de los embalses;lo que condiciona la vida en las comunida-des fluviales.
2. El agua que circula aguas abajo del embalse,tiende a recuperar la temperatura de equili-brio térmico de acuerdo con las condicionesambientales.
3. Aplicando la ecuación del balance energéticoa un tramo de 5.3 km. situado inmediatamen-te aguas abajo del sistema de embalsesMequinenza-Ribarroja-Flix en el río Ebro(España), para la época de verano, fue posibleobservar que las variables más importantesson la radiación solar total neta de onda corta,almacenamiento de calor y el calor intercam-biado con el lecho.
AGRADECIMIENTOS
El presente estudio se realiza dentro de los pro-yectos de investigación REN 2001-2185-CO2-O1 y O2 financiados por el Programa deRecursos Hídricos del Plan Nacional deInvestigación y Desarrollo. Agradecemos alInstituto de Cooperación Iberoamericano labeca que concedida a Rafael Val Segura parapoder llevar a cabo el doctorado en laUniversidad Politécnica de Cataluña, Barcelona.
REFERENCIAS
EDINGER, J. E.& D. K. BRADY & J. C. GEYER.1974. Heat Exchange and Transport in theEnvironment, Rep. 14, Electric Power ResearchInstitute, Palo Alto, California.
FRITSCHEN, L. J. & C. H. M. VAN BAVEL, 1963:Evaporation from Shallow Water and RelatedMicrometeorological Parameters. Journal ofApplied Meteorology, 2(3): 407–411.
DOLZ, J. y J. PUERTAS. 1990. Estudio delComportamiento del Río Ebro en Verano.Universidad Politécnica de Cataluña, Barcelona,España. 124 pp
SECO, J & J. GARMENDIA. (1973). Evaporaciónen España. Instituto Nacional de Meteorología,Publicación A-52, Madrid, España. 30 pp
92 Val et al.

INTRODUCCIÓN
Clásicamente ha sido demostrado que la compo-sición de las asociaciones fitoplanctónicasdepende en gran medida del clima lumínico yfísico químico. Durante los últimos años se hacomprobado además, que las interacciones devariables hidráulicas tales como períodos y pro-fundidad de mezcla y estratificación junto conel tiempo de retención de los sistemas acuáticosserían las variables forzantes de mayor impor-
tancia en la dinámica de estas asociaciones(Ford, 1990; Reynolds 1997, 1999 y 2000;Armengol et al., 1999; Straskraba, 1999).
Es el caso de los sistemas límnicos chilenos,los cuales se desarrollan en una franja continen-tal que bordea al Océano Pacífico, comprendidaentre dos o tres meridianos como promedio(66º30’ W-76º 30’W), esta influencia marítimaconfiere un clima templado y con variacionestérmicas poco marcadas estacionalmente.(Romero, 1985). Con la excepción de los lagos
93
Respuesta de la estructura fitoplanctónica a las perturbacionesantrópicas de un lago templado
Irma Vila y Rodrigo Pardo
Departamento de Ciencias Ecológicas. Facultad de Ciencias. Universidad de Chile. Casilla 653 [email protected]
RESUMEN
El Embalse Rapel (34º S; 71ºW) fue generado en 1968, y la comunidad fitoplanctónica señala los efectos de procesos de eutro-ficación y posterior contaminación por la recepción de agroquímicos y aguas claras de relave de minería del cobre.La distribución de las diversas asociaciones fitoplanctónicas es homogénea a lo largo del embalse debido a las característicashidrodinámicas del sistema con tiempos de retención entre 12 días y tres meses en las áreas diferentes del embalse y vientosdiarios que mantienen una mezcla turbulenta de la columna de agua.Las microalgas se asocian en tres grupos correspondientes a los períodos de diferente calidad química del agua y las asocia-ciones aquí descritas señalan que taxa como Microcystis aeruginosa, Oscillatoria sp. y Aulacoseira granulata aunque toleran-tes a eutrofía son rapidamente afectadas por cobre. La respuesta al aumento de la concentración de cobre señala una disminu-ción en riqueza de especies y un incremento de Dictyiosphaerium pulchellum, Coelastrum microporum y Pediastrum simplextolerantes al metal.
Palabras claves: Microalgas, fitoplancton, eutrofización, cobre.
ABSTRACT
The Rapel reservoir (34º S; 71ºW) was impounded in 1968 and the phytoplankton community shows the effects of eutrophica-tion processes and later contamination due to the affluence of agrochemicals and copper mining clear water tailings.Phytoplankton diverse associations distribution along the reservoir is homogenous because of the system’s hydrodynamic cha-racteristics with retention times between 12days and three months at the different areas of the reservoir and daily winds thatmaintain a turbulent mix of the water column.Micro algae associate in three groups that correspond to periods of different chemical water quality and the associations heredescribed show that taxa such as Microcystis aeruginosa, Oscillatoria sp., and Aulacoseira granulata although tolerant toeutrophy are rapidly affected by copper. The response to the increase of the copper concentration correspons with a decreasein species richness and with a higher abundance of Dictyiosphaerium pulchellum, Coelastrum microporum and Pediastrumsimplex as copper tolerant species.
Keywords: Micro algae, phytoplankton, eutrophication, copper.
Limnetica 22(1-2): 93-102 (2003)© Asociación Española de Limnología, Madrid. Spain. ISSN: 0213-8409

altoandinos, los lagos ubicados entre los 17º S alos 54ºS se caracterizan por ser monomícticostemplados y presentar termoclinas poco establesdurante el período de primavera y verano debidoa la presencia diaria de vientos SW (Campos,1984). La mayoría de estos lagos, a semejanzade los sistemas neozelandeses se caracterizanpor presentar rangos térmicos reducidos, epi-limnion profundo y períodos de estratificaciónbreves (Viner, 1985 y 1987; Geller, 1992). Estascaracterísticas se acentúan en los embalses porla regulación en la extracción del agua queinfluiría en el tiempo de residencia hidráulica.
El Embalse Rapel (34º S; 71ºW) fue generadoen 1968, y a ocho años de ser embalsado presentóun proceso de eutroficación acelerado debido a larecepción de agroquímicos y desarrollo rápido delturismo en sus orillas. Las asociaciones de micro-algas en este período mostraron la sucesión clásicadescrita para embalses templados (Diatomeas-Clorófitas- Cianófitas) (Hutchinson, 1957;Margalef, 1983; Vila et al., 1987) y con dominan-cia significativa de Aulacoseira granulata(Ehremberg) Simonsen durante todo el año. Estaespecie es muy abundante además, en los lagos delpaís y de acuerdo con Reynolds (1997) es exitosa
en sistemas turbulentos por recuperarse desde lossedimentos. Al incremento de nutrientes por lasactividades de turismo y agrícolas de la cuenca sesumó en 1987 el ingreso de aguas claras de relavesde la minería por el cual se incorporó cantidadessignificativas de sulfatos (1500 mg L-1) asociadosprincipalmente a molibdeno y cobre. Desde laincorporación de sulfatos la dinámica fitoplanctó-nica se ha visto modificada significativamente alperder la sucesión estacional, disminuir su bioma-sa y las especies ser reemplazadas por aquellastolerantes al efecto alguicida del sulfato de cobre(Vila et al., 2000). Sin embargo, durante 1998 sedisminuyó la afluencia de sulfatos a 250 mg L-1 ylas microalgas señalan nuevos cambios en las aso-ciaciones estacionales.
La distribución de las diversas asociaciones fito-planctónicas ha sido homogénea a lo largo delembalse y ésto posiblemente sería originado por lascaracterísticas hidrodinámicas del sistema que tienetiempos de retención entre 12 días y tres meses enlas áreas diferentes del embalse, con vientos diariosque mantienen una mezcla turbulenta de la colum-na de agua (Contreras et al., 1994).
Este trabajo intenta asociar la estructura de lasasociaciones fitoplanctónicas como respuesta a
94 Vila y Pardo
Figura 1. Ubicación geográfica y cubetas del Embalse Rapel. Cubeta 1: Estación fluvial; Cubeta 2 Estación de transición –lacus-tre y Cubeta 3 Estación represa. Geographic location and basins of the Rapel reservoir. Basin 1: Fluvial station; Basin 2:Transitional station; Basin 3: Dam station.
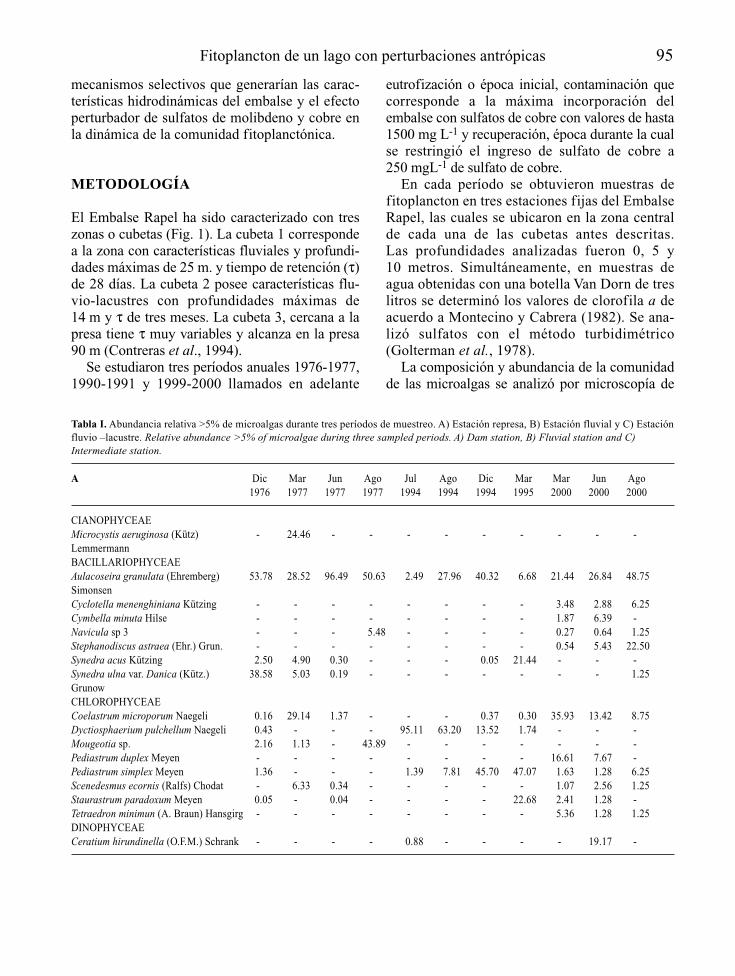
mecanismos selectivos que generarían las carac-terísticas hidrodinámicas del embalse y el efectoperturbador de sulfatos de molibdeno y cobre enla dinámica de la comunidad fitoplanctónica.
METODOLOGÍA
El Embalse Rapel ha sido caracterizado con treszonas o cubetas (Fig. 1). La cubeta 1 correspondea la zona con características fluviales y profundi-dades máximas de 25 m. y tiempo de retención (τ)de 28 días. La cubeta 2 posee características flu-vio-lacustres con profundidades máximas de14 m y τ de tres meses. La cubeta 3, cercana a lapresa tiene τ muy variables y alcanza en la presa90 m (Contreras et al., 1994).
Se estudiaron tres períodos anuales 1976-1977,1990-1991 y 1999-2000 llamados en adelante
eutrofización o época inicial, contaminación quecorresponde a la máxima incorporación delembalse con sulfatos de cobre con valores de hasta1500 mg L-1 y recuperación, época durante la cualse restringió el ingreso de sulfato de cobre a250 mgL-1 de sulfato de cobre.
En cada período se obtuvieron muestras defitoplancton en tres estaciones fijas del EmbalseRapel, las cuales se ubicaron en la zona centralde cada una de las cubetas antes descritas.Las profundidades analizadas fueron 0, 5 y10 metros. Simultáneamente, en muestras deagua obtenidas con una botella Van Dorn de treslitros se determinó los valores de clorofila a deacuerdo a Montecino y Cabrera (1982). Se ana-lizó sulfatos con el método turbidimétrico(Golterman et al., 1978).
La composición y abundancia de la comunidadde las microalgas se analizó por microscopía de
Fitoplancton de un lago con perturbaciones antrópicas 95
Tabla I. Abundancia relativa >5% de microalgas durante tres períodos de muestreo. A) Estación represa, B) Estación fluvial y C) Estaciónfluvio –lacustre. Relative abundance >5% of microalgae during three sampled periods. A) Dam station, B) Fluvial station and C)Intermediate station.
A Dic Mar Jun Ago Jul Ago Dic Mar Mar Jun Ago 1976 1977 1977 1977 1994 1994 1994 1995 2000 2000 2000
CIANOPHYCEAEMicrocystis aeruginosa (Kütz) - 24.46 - - - - - - - - -LemmermannBACILLARIOPHYCEAEAulacoseira granulata (Ehremberg) 53.78 28.52 96.49 50.63 2.49 27.96 40.32 6.68 21.44 26.84 48.75SimonsenCyclotella menenghiniana Kützing - - - - - - - - 3.48 2.88 6.25Cymbella minuta Hilse - - - - - - - - 1.87 6.39 -Navicula sp 3 - - - 5.48 - - - - 0.27 0.64 1.25Stephanodiscus astraea (Ehr.) Grun. - - - - - - - - 0.54 5.43 22.50Synedra acus Kützing 2.50 4.90 0.30 - - - 0.05 21.44 - - -Synedra ulna var. Danica (Kütz.) 38.58 5.03 0.19 - - - - - - - 1.25GrunowCHLOROPHYCEAECoelastrum microporum Naegeli 0.16 29.14 1.37 - - - 0.37 0.30 35.93 13.42 8.75Dyctiosphaerium pulchellum Naegeli 0.43 - - - 95.11 63.20 13.52 1.74 - - -Mougeotia sp. 2.16 1.13 - 43.89 - - - - - - -Pediastrum duplex Meyen - - - - - - - - 16.61 7.67 -Pediastrum simplex Meyen 1.36 - - - 1.39 7.81 45.70 47.07 1.63 1.28 6.25Scenedesmus ecornis (Ralfs) Chodat - 6.33 0.34 - - - - - 1.07 2.56 1.25Staurastrum paradoxum Meyen 0.05 - 0.04 - - - - 22.68 2.41 1.28 -Tetraedron minimun (A. Braun) Hansgirg - - - - - - - - 5.36 1.28 1.25DINOPHYCEAECeratium hirundinella (O.F.M.) Schrank - - - - 0.88 - - - - 19.17 -
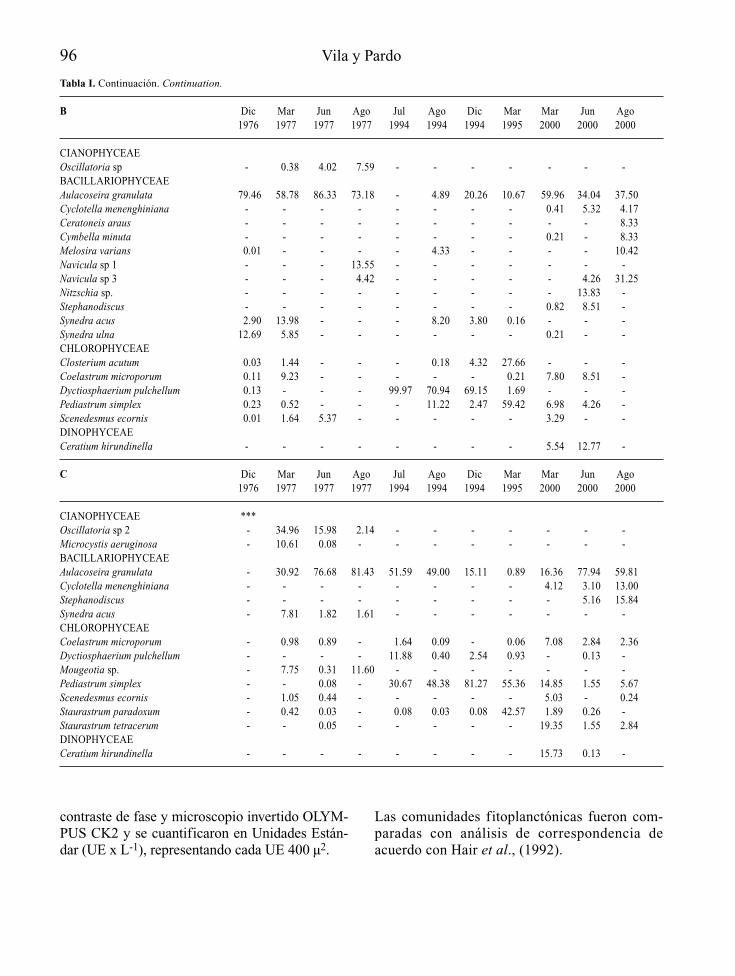
96 Vila y Pardo
Tabla I. Continuación. Continuation.
B Dic Mar Jun Ago Jul Ago Dic Mar Mar Jun Ago 1976 1977 1977 1977 1994 1994 1994 1995 2000 2000 2000
CIANOPHYCEAEOscillatoria sp - 0.38 4.02 7.59 - - - - - - -BACILLARIOPHYCEAEAulacoseira granulata 79.46 58.78 86.33 73.18 - 4.89 20.26 10.67 59.96 34.04 37.50Cyclotella menenghiniana - - - - - - - - 0.41 5.32 4.17Ceratoneis araus - - - - - - - - - - 8.33Cymbella minuta - - - - - - - - 0.21 - 8.33Melosira varians 0.01 - - - - 4.33 - - - - 10.42Navicula sp 1 - - - 13.55 - - - - - - -Navicula sp 3 - - - 4.42 - - - - - 4.26 31.25Nitzschia sp. - - - - - - - - - 13.83 -Stephanodiscus - - - - - - - - 0.82 8.51 -Synedra acus 2.90 13.98 - - - 8.20 3.80 0.16 - - -Synedra ulna 12.69 5.85 - - - - - - 0.21 - -CHLOROPHYCEAEClosterium acutum 0.03 1.44 - - - 0.18 4.32 27.66 - - -Coelastrum microporum 0.11 9.23 - - - - - 0.21 7.80 8.51 -Dyctiosphaerium pulchellum 0.13 - - - 99.97 70.94 69.15 1.69 - - -Pediastrum simplex 0.23 0.52 - - - 11.22 2.47 59.42 6.98 4.26 -Scenedesmus ecornis 0.01 1.64 5.37 - - - - - 3.29 - -DINOPHYCEAECeratium hirundinella - - - - - - - - 5.54 12.77 -
C Dic Mar Jun Ago Jul Ago Dic Mar Mar Jun Ago 1976 1977 1977 1977 1994 1994 1994 1995 2000 2000 2000
CIANOPHYCEAE ***Oscillatoria sp 2 - 34.96 15.98 2.14 - - - - - - -Microcystis aeruginosa - 10.61 0.08 - - - - - - - -BACILLARIOPHYCEAEAulacoseira granulata - 30.92 76.68 81.43 51.59 49.00 15.11 0.89 16.36 77.94 59.81Cyclotella menenghiniana - - - - - - - - 4.12 3.10 13.00Stephanodiscus - - - - - - - - - 5.16 15.84Synedra acus - 7.81 1.82 1.61 - - - - - - -CHLOROPHYCEAECoelastrum microporum - 0.98 0.89 - 1.64 0.09 - 0.06 7.08 2.84 2.36Dyctiosphaerium pulchellum - - - - 11.88 0.40 2.54 0.93 - 0.13 -Mougeotia sp. - 7.75 0.31 11.60 - - - - - - -Pediastrum simplex - - 0.08 - 30.67 48.38 81.27 55.36 14.85 1.55 5.67Scenedesmus ecornis - 1.05 0.44 - - - - - 5.03 - 0.24Staurastrum paradoxum - 0.42 0.03 - 0.08 0.03 0.08 42.57 1.89 0.26 -Staurastrum tetracerum - - 0.05 - - - - - 19.35 1.55 2.84DINOPHYCEAECeratium hirundinella - - - - - - - - 15.73 0.13 -
contraste de fase y microscopio invertido OLYM-PUS CK2 y se cuantificaron en Unidades Están-dar (UE x L-1), representando cada UE 400 µ2.
Las comunidades fitoplanctónicas fueron com-paradas con análisis de correspondencia deacuerdo con Hair et al., (1992).
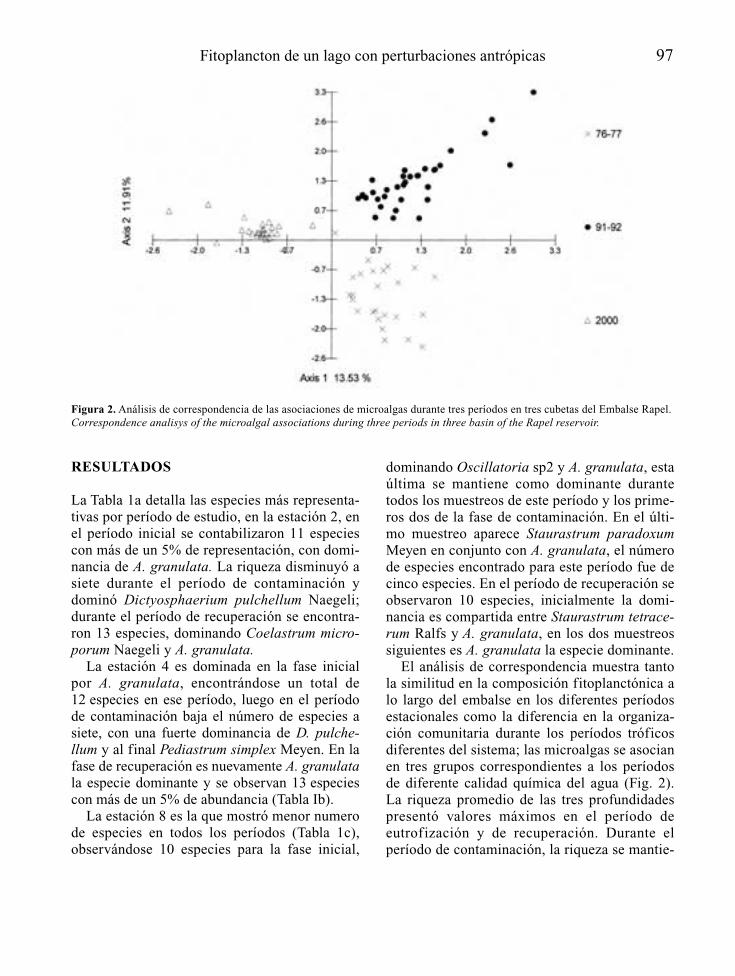
RESULTADOS
La Tabla 1a detalla las especies más representa-tivas por período de estudio, en la estación 2, enel período inicial se contabilizaron 11 especiescon más de un 5% de representación, con domi-nancia de A. granulata. La riqueza disminuyó asiete durante el período de contaminación ydominó Dictyosphaerium pulchellum Naegeli;durante el período de recuperación se encontra-ron 13 especies, dominando Coelastrum micro-porum Naegeli y A. granulata.
La estación 4 es dominada en la fase inicialpor A. granulata, encontrándose un total de12 especies en ese período, luego en el períodode contaminación baja el número de especies asiete, con una fuerte dominancia de D. pulche-llum y al final Pediastrum simplex Meyen. En lafase de recuperación es nuevamente A. granulatala especie dominante y se observan 13 especiescon más de un 5% de abundancia (Tabla Ib).
La estación 8 es la que mostró menor numerode especies en todos los períodos (Tabla 1c),observándose 10 especies para la fase inicial,
dominando Oscillatoria sp2 y A. granulata, estaúltima se mantiene como dominante durantetodos los muestreos de este período y los prime-ros dos de la fase de contaminación. En el últi-mo muestreo aparece Staurastrum paradoxumMeyen en conjunto con A. granulata, el númerode especies encontrado para este período fue decinco especies. En el período de recuperación seobservaron 10 especies, inicialmente la domi-nancia es compartida entre Staurastrum tetrace-rum Ralfs y A. granulata, en los dos muestreossiguientes es A. granulata la especie dominante.
El análisis de correspondencia muestra tantola similitud en la composición fitoplanctónica alo largo del embalse en los diferentes períodosestacionales como la diferencia en la organiza-ción comunitaria durante los períodos tróficosdiferentes del sistema; las microalgas se asocianen tres grupos correspondientes a los períodosde diferente calidad química del agua (Fig. 2).La riqueza promedio de las tres profundidadespresentó valores máximos en el período deeutrofización y de recuperación. Durante elperíodo de contaminación, la riqueza se mantie-
Fitoplancton de un lago con perturbaciones antrópicas 97
Figura 2. Análisis de correspondencia de las asociaciones de microalgas durante tres períodos en tres cubetas del Embalse Rapel.Correspondence analisys of the microalgal associations during three periods in three basin of the Rapel reservoir.
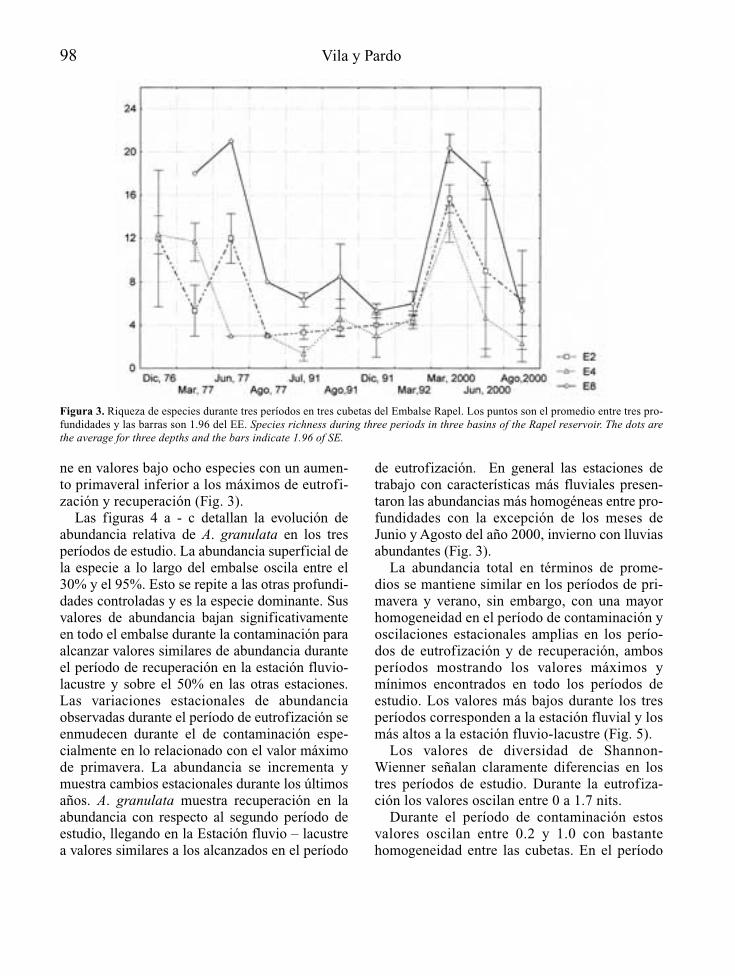
ne en valores bajo ocho especies con un aumen-to primaveral inferior a los máximos de eutrofi-zación y recuperación (Fig. 3).
Las figuras 4 a - c detallan la evolución deabundancia relativa de A. granulata en los tresperíodos de estudio. La abundancia superficial dela especie a lo largo del embalse oscila entre el30% y el 95%. Esto se repite a las otras profundi-dades controladas y es la especie dominante. Susvalores de abundancia bajan significativamenteen todo el embalse durante la contaminación paraalcanzar valores similares de abundancia duranteel período de recuperación en la estación fluvio-lacustre y sobre el 50% en las otras estaciones.Las variaciones estacionales de abundanciaobservadas durante el período de eutrofización seenmudecen durante el de contaminación espe-cialmente en lo relacionado con el valor máximode primavera. La abundancia se incrementa ymuestra cambios estacionales durante los últimosaños. A. granulata muestra recuperación en laabundancia con respecto al segundo período deestudio, llegando en la Estación fluvio – lacustrea valores similares a los alcanzados en el período
de eutrofización. En general las estaciones detrabajo con características más fluviales presen-taron las abundancias más homogéneas entre pro-fundidades con la excepción de los meses deJunio y Agosto del año 2000, invierno con lluviasabundantes (Fig. 3).
La abundancia total en términos de prome-dios se mantiene similar en los períodos de pri-mavera y verano, sin embargo, con una mayorhomogeneidad en el período de contaminación yoscilaciones estacionales amplias en los perío-dos de eutrofización y de recuperación, ambosperíodos mostrando los valores máximos ymínimos encontrados en todo los períodos deestudio. Los valores más bajos durante los tresperíodos corresponden a la estación fluvial y losmás altos a la estación fluvio-lacustre (Fig. 5).
Los valores de diversidad de Shannon-Wienner señalan claramente diferencias en lostres períodos de estudio. Durante la eutrofiza-ción los valores oscilan entre 0 a 1.7 nits.
Durante el período de contaminación estosvalores oscilan entre 0.2 y 1.0 con bastantehomogeneidad entre las cubetas. En el período
98 Vila y Pardo
Figura 3. Riqueza de especies durante tres períodos en tres cubetas del Embalse Rapel. Los puntos son el promedio entre tres pro-fundidades y las barras son 1.96 del EE. Species richness during three periods in three basins of the Rapel reservoir. The dots arethe average for three depths and the bars indicate 1.96 of SE.
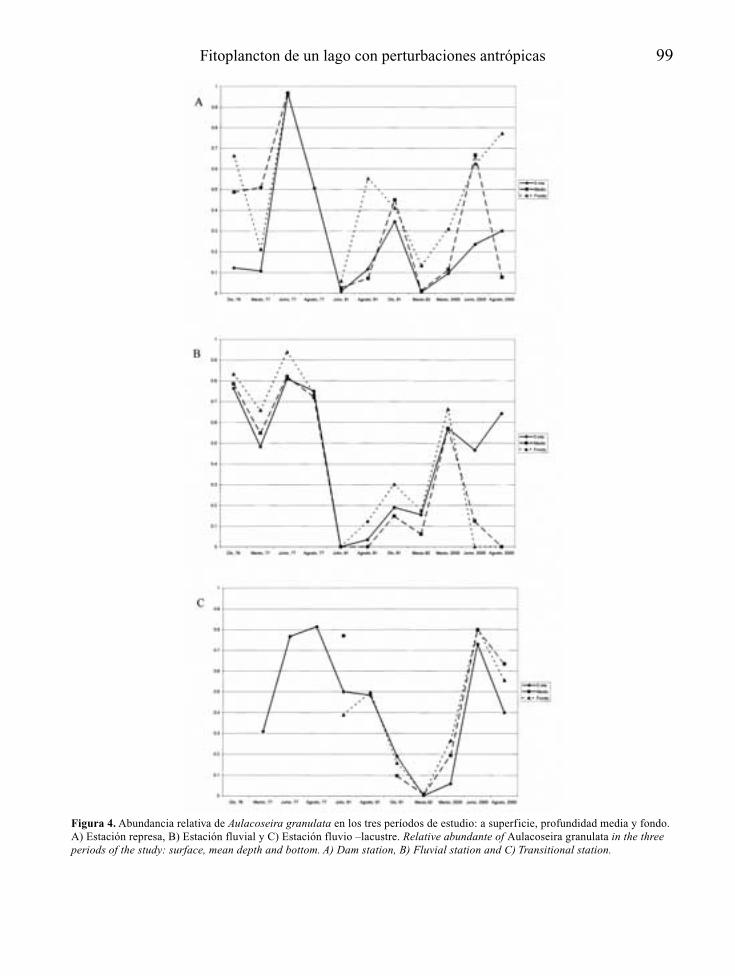
Fitoplancton de un lago con perturbaciones antrópicas 99
Figura 4. Abundancia relativa de Aulacoseira granulata en los tres períodos de estudio: a superficie, profundidad media y fondo.A) Estación represa, B) Estación fluvial y C) Estación fluvio –lacustre. Relative abundante of Aulacoseira granulata in the threeperiods of the study: surface, mean depth and bottom. A) Dam station, B) Fluvial station and C) Transitional station.
de recuperación la diversidad oscila entre 1 y1.8 (Fig. 6). Los valores más bajos de diversidadse estiman para la estación fluvial.
DISCUSIÓN
Ha sido ampliamente documentada la variabili-dad de la composición del fitoplancton en losembalses, especialmente como respuesta a lamorfología de los mismos (Kimmel et al., 1990;Reynolds, 1999). En el caso de los sistemas chile-nos, a la morfología se suman las variables carac-terísticas de clima oceánico. La mezcla turbulentacasi permanente del agua de los lagos monomícti-cos templados de Chile, ha favorecido la presen-cia especies como A. granulata, la cual mantieneo incrementa sus poblaciones por resuspensióndesde los sedimentos (Reynolds et al., 1985).Bajo condiciones de sucesión normal de losembalses templados, la composición fitoplanctó-nica ha sido predecible tanto en su composicióncomo su respuesta a los cambios estacionales. Enel período inicial de este embalse, la sucesiónestacional de las microalgas estaba conformadapor especies representativas de la mayoría de losordenes comunes en estos sistemas. Entre éstas,Chlorophyceae y Cyanophyceae (Margalef, 1983;
Reynolds, 2000), a pesar de la dominancia de A.granulata. La disminución marcada de esta espe-cie durante el período de contaminación se expli-caría principalmente por el incremento de materiaorgánica y sulfatos los cuales estarían producien-do extensas zonas anóxicas cercanas a los sedi-mentos en las cuales se solubilizarían metalescomo Cu+2, afectando rápidamente a los inóculosde A. granulata. Trabajos realizados con molus-cos bentónicos muestran el efecto contaminantede los sulfatos de cobre provenientes de los sedi-mentos (Valdovinos et al., 1998). En general semenciona que existe información insuficienteacerca de los mecanismos de respuesta a la toxici-dad de metales pesados como el cobre y que serequiere de más investigación en el tema y espe-cialmente en aspectos relacionados con lasmicroalgas tanto en sistemas naturales comoexperimentales. Al respecto, IPCS (1998) resumeinformación que señala que a concentracionesentre ≥ 10 y ≥ 20 µg L-1 de cobre, la clorofila a, yla fijación de nitrógeno y carbono disminuyensignificativamente. Filipek et al. (1987) discutenampliamente y demuestran que las concentracio-nes de cobre disuelto reflejan la acidez del agua.A pH neutro generalmente la concentración decobre disuelto está bajo los límites de detecciónen la columna de agua, pero en los sedimentos,
100 Vila y Pardo
Figura 5. Abundancia total promedio de microalgas entre tres profundidades, a escala logarítmica y barras de 1.96 del EE), en tresperíodos. Average total abundance of microalga between three depths, in logarithmic scale and 1.96 SE bars, in three periods.
que son el depósito y reservorio de metales ocu-rren cambios diarios y/o estacionales que modifi-can las interacciones sedimento-agua. Así, aún apH neutro Winner y Owen (1991) han demostra-do que a valores entre 20 µg L-1 y 40 µg L-1
de cobre se reduce la riqueza de especies demicroalgas. Tratamientos con 5 µg L-1 de cobredurante 2 a 5 semanas, en Cazenoria, Lake NewYork, señalaron alteraciones de corto plazo en lasucesión estacional de las microalgas, similar-mente a lo ocurrido en el embalse Rapel (Effleret al., 1980; Vila et al., 1997).
Por otra parte, ha sido frecuentemente reportadoque la solubilización de Cu+2, afectaría en toda lacolumna de agua especialmente a las Cyanobac-terias descritas como altamente sensibles a estemetal (Boyd, 1972; Wurtsbaugh & Horne, 1982).
La perturbación producida por ralaves demetales como cobre, disminuye significativa-mente la riqueza de especies en este embalse,sin embargo la presencia de microalgas toleran-tes en mayor abundancia apoya la resilienciadel sistema, el cual muestra puntos de compen-sación en relación con los productores prima-rios, pero es de importancia evaluar el efectoque los cambios en las asociaciones de lasmicroalgas y la perturbación producirían en latrama trófica del sistema.
BIBLIOGRAFÍA
ARMENGOL, J., J. C. GARCÍA, M. COMERMA,M. ROMERO, J. DOLZ, M. ROURA, B. H. HAN,A. VIDAL, & K. SIMEK. 1999. LongitudinalProcesses in Canyon Type Reservoirs: The case ofSau (N.E. Spain). In: Theoretical ReservoirEcology and its applications. J. G Tundisi & M.Straskraba (eds.): 313-345. International Instituteof Ecology. Brazilian Academy of Sciences andBackhuys Publishers.
BOYD, C. 1972. Water quality in warmwater fishponds. Auburn University, AgriculturalExperiment Station, R. Dennis Rouse,Director/Auburn, Alabama.
CAMPOS, H. 1984. Limnological study ofAraucanian lakes (Chile). Verh. Internat. Verein.Limnol., 22: 1319-1327.
CONTRERAS, M., H. VILLAGRÁN Y C. SALA-ZAR. 1994. Características hidrodinámicas delembalse Rapel. Medio Ambiente, 12(1): 41-49.
EFFLER S. W., S. LITTEN, S. D. FIELD, T. TONG-NORK, F. HALE, M. MEYER & M. QUIRK1980. Whole lake response to low level coppersulphate treatment. Water Res., 14: 1489-1499.
FILIPEK L. H., D. K. NORDSTROM & W. H. FIC-KLIN. 1987. Interaction of acid mine drainagewith waters and sediments of West Squaw Creekin the west Shasta mining district, California.Environ. Sci. Technol., 21:388-396
Fitoplancton de un lago con perturbaciones antrópicas 101
Figura 6. Valores de diversidad de Shannon-Wienner de microalgas en tres cubetas, durante tres períodos. Las barras indican 1.96de EE. Shannon-Wienner diversity values of microalgae in three basins during three periods. The bars show 1.96 of SE.

FORD, D. E. 1990. Reservoir transport processes. In:Reservoir Limnology. Ecological perspectives. K.W. Thorton, B. L. Kimmel & F. E. Payne(eds.):15-41. John Wiley & Sons Inc. N. Y.
GELLER, W. 1992. The temperature stratificationand related characteristics of Chilean lakes inmidsummer. Aquatic Sciences, 54: 1-16.
GOLTERMAN, H. L., R. S. CLYMO & M. A.OHNSTAD. 1978. Methods for physical and che-mical analisis of freshwaters. 2ond ed. Blackwell,Oxford.
HAIR, J., R. ANDERSON, R. TATHAM & W.BLACK. 1992. Multivariate Analysis withReadings. Mac Millan Publishing Co. N. Y.
HUTCHINSON, G. E. 1957. A Treatise onLimnology. Vol. I: Geogrpahy, Physics andChemistry. N. Y.
IPCS (International Programme on ChemicalSafety). 1998. Environmental Health Criteria2000. Copper. World Health Organization,Geneva.
KIMMEL, B. L., T. L. OWEN & L. J. PAULSON.1990. Reservoir Primary Production. In:Reservoir Limnology. Ecological Perspectives. K.W. Thorton, B. L. Kimmel & F. E. Payne (eds.):133-194. John Wiley & Sons Inc. N. Y.
MARGALEF, R. 1983. Limnología. Ed. Omega.Barcelona.
MONTECINO, V. & S. CABRERA. 1982.Phytoplankton activity and standing crop in animpoundment of Central Chile. J. Plankton Res. 4:943-950.
REYNOLDS, C., V. MONTECINO, M. E. GRAF &S. CABRERA. 1985. Short-term dynamics of aMelosira population in the plankton of animpundment in Central Chile. J. Plankton Res. 8:715-740.
REYNOLDS, C. S. 1989. Physical determinants ofphytoplankton succession. In: Plankton Ecology.U. Sommer (ed.): 9-56. Springer. Berlin.
REYNOLDS, C. S. 1997. Vegetation processes in thepelagic: A model for ecosystem theory. Excellencein Ecology, Book 9. Ecology Institute, Nordbunte,Oldendorf/Luhe, Germany.
REYNOLDS, C. S. 1999. Phytoplankton assembla-ges in Reservoirs. In: Theoretical ReservoirEcology and its applications. J. G Tundisi & M.Straskraba (eds.): 439-456. International Institute
of Ecology. Brazilian Academy of Sciences andBackhuys Publishers.
REYNOLDS, C. S. 2000. Phytoplankton designer orhow to predict compositional responses to trophicstate change. Hydrobiologia, 424: 123-132.
ROMERO, H. 1985. Geografía de Chile. Tomo XI:Geografía de los climas. Instituto GeográficoMilitar. Santiago, Chile. 243 pp.
STRASKRABA, M. 1999. Retention Time as a KeyVariable in Reservoir Limnology. In: TheoreticalReservoir Ecology and its applications. J. GTundisi & M. Straskraba (eds.): 385-410.International Institute of Ecology. BrazilianAcademy of Sciences and Backhuys Publishers.
VALDOVINOS, C., R. FIGUEROA, H. CID, O.PARRA, E. ARAYA, S. PRIVITERA Y V.OLMOS. 1998. Transplante de organismos bentó-nicos entre sistemas lénticos: ¿refleja la biodispo-nibilidad de metales traza en el ambiente? Bol.Soc. Chile. Quim., 43: 467-475.
VILA, I., I. BARENDS & V. MONTECINO. 1987.Composición, abundancia y distribución espacialy temporal del fitoplancton del embalse Rapel(1976-1977). Revista Chilena de Historia Natural,60: 37-55.
VILA, I., M. CONTRERAS & J. PIZARRO. 1997.Eutrophication and phytoplankton selective res-ponses in a temperate reservoir. Verh. Internat.Verein. Limnol., 26: 798-802.
VILA, I., M. CONTRERAS, V. MONTECINO, J.PIZARRO & D. ADAMS. 2000. Rapel: A 30 yearstemperate reservoir. Eutrophication or contamina-tion? Arch. Hydrobiol. Spec. Issues Advanc.Limnol., 55: 31-44.
VINER, A. B. 1985. Thermal stability and phytoplanktondistribution. In: Perspectives in Southern Hemispherelimnology. R. B. Davies & R.D. Wamsley (eds.). Dev.Hydrobiol., 28, Dordrecht, Junk.
VINER, A.B. (ed.). 1987. Inland waters of NewZealand. DSIR Bull. 241, Wellington.
WINNER, R.W. & T. L. OWEN 1991. Seasonalvariability in the sensitivity of freshwater phyto-plankton communities to a chronic copper stress.Aquat. Toxicol., 19: 73-88.
WURTSBAUGH, W.A., & A. J. Horne. 1982. Effectsof copper on nitrogen fixation and growth of blue-green algae in natural plankton associations. Can.J. Fish. Aquat. Sci., 39: 1636-1641.
102 Vila y Pardo

INTRODUCCIÓN
El embalse de Salto Grande constituye una obrade represamiento del Río Uruguay de carácterbinacional Uruguayo-Argentino. Fue creado parageneración de energía hidroeléctrica entrando enoperación en 1979. Es utilizado además paraabastecimiento de agua potable, navegación,riego y recreación. Según Salas y Martino(1990), de acuerdo a su temperatura media anualmayor a 15ºC y su temperatura mínima superior a10ºC, el sistema podría considerarse como cálidotropical. En base a las características físico-quí-micas, el embalse está formado por dos subsiste-mas definidos por el cauce principal y los brazos
laterales (Berón, 1990; Chalar et al. 1993; Condeet al, 1996) y presenta características de ambien-te meso-eutrófico. La comunidad fitoplanctónicaestá dominada por diatomeas del géneroMelosira (Aulacoseira spp), en casi todo el siste-ma y durante gran parte del año, siendo acompa-ñadas por cyanobacterias Microcystis aeruginosaen los brazos Gualeguaycito y Mandisoví (mar-gen argentino), al final del verano y otoño(Quirós y Luchini, 1983).
Este estudio pretende aportar informaciónnueva sobre la composición y dinámica de lacomunidad fitoplanctónica y analizar su distri-bución espacial y temporal, en relación a losparámetros abióticos.
103
Abundancia y diversidad del fitoplancton en el Embalse de SaltoGrande (Argentina – Uruguay). Ciclo estacional y distribución espacial
Lizet De León1 y Guillermo Chalar2
Sección Limnología, Facultad de Ciencias, Universidad de la República. Iguá 4225 CP: 11400. Montevideo– Uruguay. 1- e-mail: [email protected]; 2- e-mail: [email protected]
RESUMEN
El embalse de Salto Grande es un ecosistema artificial creado para la producción hidroeléctrica. Este sistema tiene un tiempode residencia bajo, 12.3 días, su régimen térmico es polimíctico y la morfología es dendrítica. En este embalse se ha estudiadola composición, abundancia y diversidad del fitoplancton en relación con el ciclo estacional y los parámetros físicos y quími-cos. La abundancia anual ha variado de 16 a 1963 ind ml-1. Los máximos valores se registraron en verano en las estacionessituadas en brazos con dominancia de Aulacoseira y Microcystis. Los valores mínimos se encontraron en el canal principal ydurante el invierno donde dominaban los fitoflagelados nanoplanctónicos. La diversidad anual mostró un patrón inverso conun rango de variación entre 0.6 y 4.4 bits ind-1. En este trabajo se discute una sucesión estacional de estrategas C a R y S, comosugiere Reynolds 1988.
Palabras clave: fitoplancton, diversidad, sucesión de estrategas, embalses.
ABSTRACT
Salto Grande reservoir is an artificial ecosystem created for hydroelectric power generation. This system has low retentiontime, 12.3 days, polymictic regime, and dendritric morphology. The composition, abundance and diversity of phytoplankton inrelation with the seasonal cycle and physicochemical parameters were investigated in this reservoir. Annual abundance variedfrom 16 to 1963 ind ml-1. The maximum values were registered in summer, in the stations located in the branches withAulacoseira and Microcystis dominance. The minimum ones corresponded to the main channel in winter were nanoplanktonicphytoflagellates dominated. The annual diversity showed an inverse trend ranging between 0.6 and 4.4 bits ind-1. In this paper,a succession from C to R and S strategy is discussed, as suggested by Reynolds (1988).
Keywords: phytoplankton, diversity, strategist succession, reservoirs.
Limnetica 22(1-2): 103-113 (2003)© Asociación Española de Limnología, Madrid. Spain. ISSN: 0213-8409

ÁREA DE ESTUDIO
Las características morfométricas del Embalsede Salto Grande se detallan en la tabla 1. El RíoUruguay es el principal tributario. El mismonace aproximadamente a 60 km del OcéanoAtlántico en la Serra do Mar (Santa Catarina -Brasil) y recorre 1800 km en dirección Oeste ySur, hasta su desembocadura en el Río de laPlata. Su lecho es principalmente de roca basál-tica, siendo el margen uruguayo más elevadoque el argentino. El régimen hidrológico del ríopresenta sus mayores caudales durante el invier-no y los menores en verano. El caudal medio en
72 años de registro fue de 4643 m3 s-1 con míni-mos y máximos de 92 y 36100 m3 s-1 respecti-vamente (CTM, 1982).
La cuenca del Río Uruguay abarca 297 199 km2
de extensión en territorios de Brasil, Argentina yUruguay, mientras que la cuenca del embalse es de224 000 km2 . Las precipitaciones anuales en lacuenca tributaria varían entre 1000 y 2000 mm.En la cuenca del embalse se registra un pro-medio anual de 1260 mm. Las temperaturasmedias anuales son de 19 ºC y los vientos predo-minantes son del NE y SO con promedios anualesde 7 y 14 km h-1 respectivamente.
MATERIAL Y MÉTODOS
Se seleccionaron cuatro estaciones de muestreoen la cabecera del embalse, dos en el curso prin-cipal denominadas Represa (R) y La Paloma (A),una en el brazo Gualeguaycito (G) en el margenargentino, al Este, y otra en el brazo Itapebí (I) enel margen uruguayo, al Oeste (Fig. 1). Las mues-tras se tomaron con una frecuencia bimensualentre mayo y setiembre (invierno) y mensualentre setiembre y abril (primavera a otoño).
En cada estación se midió la temperatura y setomaron muestras para la determinación de sóli-dos suspendidos totales, fósforo total, fosfato,nitratos y silicatos, según la metodología descritaen Conde et al. (1997). Se tomaron muestras parael análisis cualitativo de fitoplancton mediantearrastre superficial con una red de plancton de 33µm y para el análisis cuantitativo y de clorofila acon bomba de succión a las profundidadescorrespondientes al 100 %, 50 %, 1 % de luzincidente en la superficie de la columna de aguay a 1 m, 5 m y 10 m por debajo del límite del1 %. Esta última profundidad no fue consideradapara la clorofila a. Las muestras cualitativas sefijaron in situ con formalina neutralizada al 4%de concentración final. Para la clasificación delas Divisiones se siguió el criterio de Silva (1972,en Parra y Bicudo, 1995). Los organismos flage-lados nanoplanctónicos (< 30 µm) no identifica-dos se agruparon bajo la denominación de“Fitoflagelados”. Para el análisis cuantitativo, las
104 De León y Chalar
Tabla 1. Características morfométricas del Embalse Salto Grande.Morphometric features of the Salto Grande Reservoir.
Superficie 780 km2
Volumen 5109 x 106 m3
Profundidad media 6.4 mProfundidad máxima 35.0 mAncho medio 5.4 kmAncho máximo 9.0 kmLongitud 100 kmDesarrollo línea de costa 11.9Perímetro 1190 kmTiempo de residencia promedio 11.3 días
Figura 1. Esquema mostrando la ubicación del Embalse deSalto Grande en la región y las estaciones de muestreo en elembalse. R- Represa, A- La Paloma, G- Gualeguaycito e I-Itapebí. Diagram showing Salto Grande Reservoir location inthe region, and the sampling stations in the reservoir. R-Dam, A- La Paloma, G- Gualeguaycito e I- Itapebí.
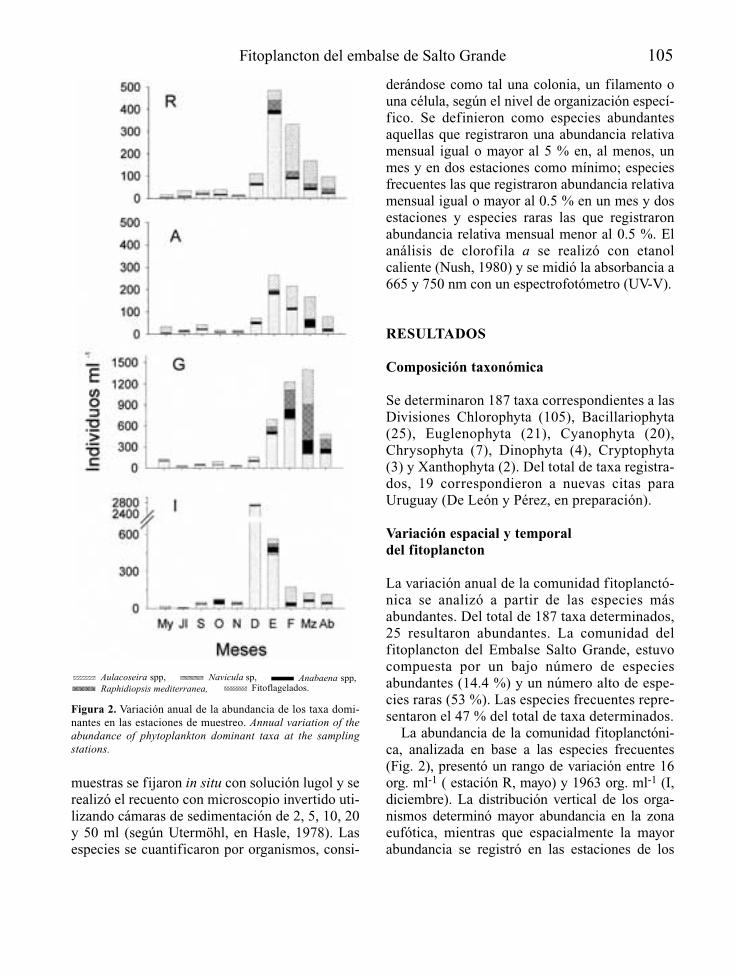
muestras se fijaron in situ con solución lugol y serealizó el recuento con microscopio invertido uti-lizando cámaras de sedimentación de 2, 5, 10, 20y 50 ml (según Utermöhl, en Hasle, 1978). Lasespecies se cuantificaron por organismos, consi-
derándose como tal una colonia, un filamento ouna célula, según el nivel de organización especí-fico. Se definieron como especies abundantesaquellas que registraron una abundancia relativamensual igual o mayor al 5 % en, al menos, unmes y en dos estaciones como mínimo; especiesfrecuentes las que registraron abundancia relativamensual igual o mayor al 0.5 % en un mes y dosestaciones y especies raras las que registraronabundancia relativa mensual menor al 0.5 %. Elanálisis de clorofila a se realizó con etanolcaliente (Nush, 1980) y se midió la absorbancia a665 y 750 nm con un espectrofotómetro (UV-V).
RESULTADOS
Composición taxonómica
Se determinaron 187 taxa correspondientes a lasDivisiones Chlorophyta (105), Bacillariophyta(25), Euglenophyta (21), Cyanophyta (20),Chrysophyta (7), Dinophyta (4), Cryptophyta(3) y Xanthophyta (2). Del total de taxa registra-dos, 19 correspondieron a nuevas citas paraUruguay (De León y Pérez, en preparación).
Variación espacial y temporaldel fitoplancton
La variación anual de la comunidad fitoplanctó-nica se analizó a partir de las especies másabundantes. Del total de 187 taxa determinados,25 resultaron abundantes. La comunidad delfitoplancton del Embalse Salto Grande, estuvocompuesta por un bajo número de especiesabundantes (14.4 %) y un número alto de espe-cies raras (53 %). Las especies frecuentes repre-sentaron el 47 % del total de taxa determinados.
La abundancia de la comunidad fitoplanctóni-ca, analizada en base a las especies frecuentes(Fig. 2), presentó un rango de variación entre 16org. ml-1 ( estación R, mayo) y 1963 org. ml-1 (I,diciembre). La distribución vertical de los orga-nismos determinó mayor abundancia en la zonaeufótica, mientras que espacialmente la mayorabundancia se registró en las estaciones de los
Fitoplancton del embalse de Salto Grande 105
Figura 2. Variación anual de la abundancia de los taxa domi-nantes en las estaciones de muestreo. Annual variation of theabundance of phytoplankton dominant taxa at the samplingstations.
Aulacoseira spp, Navicula sp, Anabaena spp,Raphidiopsis mediterranea, Fitoflagelados.
brazos y temporalmente en verano (entre diciem-bre y abril). En las estaciones R y A (cauce princi-pal), las abundancias promedio anuales fueronsimilares entre sí y menores que en G e I (brazos).
La comunidad fitoplanctónica registró sumenor abundancia (< 100 ind ml-1), entre mayoy noviembre en las cuatro estaciones de mues-treo. En diciembre se incrementó la densidad deorganismos principalmente en I (ca. 2000 org.ml-1), mientras que en enero, el incremento seregistró en todas las estaciones. Estas diferen-cias se debieron principalmente al crecimientode Bacillariophyta en I (en diciembre) y deCyanophyta en G (febrero, marzo y abril),seguidas de Bacillariophyta y flagelados (Fig.2). Dentro de estos grupos las especies másabundantes fueron: Aulacoseira sp y A. ambiguaen I y Aulacoseira sp, A. ambigua, Anabaenaplanctonica y Raphidiopsis mediterranea en G.
Biomasa del fitoplancton
La biomasa fitoplanctónica, medida como la con-centración de clorofila a , registró un valor pro-medio anual de 2.01 µg l-1 para las 4 estaciones, 5profundidades y 9 meses analizados. Los meno-res valores se registraron en las estaciones delcauce principal (R y A), con un mínimo de0.15 µg l-1 en mayo (Fig. 3). La mayor concentra-ción de clorofila a se registró en diciembre, conun máximo de 17.2 µg l-1 (promedio para la
columna de agua) en I. Dicho aumento se encon-tró asociado a un incremento de la abundancia deAulacoseira spp. En G la mayor concentración seregistró en marzo y abril (7.0 µg l-1 y 5.7 µg l-1,respectivamente) y estuvo asociada al aumentode las poblaciones de Cyanophyta (Anabaenaplanctonica, Raphidiopsis mediterranea).
Diversidad del fitoplancton
Para el análisis de diversidad se utilizaron losdatos de abundancia de las especies más fre-cuentes. Se excluyeron los fitoflagelados portratarse de un grupo multiespecífico.
Los valores de diversidad oscilaron entre 0.6y 4.4 bits ind-1 con un promedio de 2.68 paratodo el sistema. Los valores más altos corres-pondieron generalmente a los meses fríos (mayoa octubre) mientras que la menor diversidad fueregistrada en los meses cálidos (diciembre,febrero y marzo) (Fig. 4).
En las estaciones R y A, el índice de diversi-dad fue mayor al comienzo del período de estu-dio, disminuyendo progresivamente. G e I pre-sentaron la misma tendencia en el tiempo, perocon mayores oscilaciones. En diciembre seregistró una disminución de la diversidad entodas las estaciones y en enero un incremento.
Parámetros ambientales
La temperatura mínima del agua se registró enjulio (11.8 ºC), aumentando hasta alcanzar el
106 De León y Chalar
Figura 4. Variación anual de la diversidad fitoplanctónica enlas estaciones de muestreo. Annual variation of phytoplanktondiversity at the sampling stations.
Figura 3. Variación anual de la clorofila a en superficie, enlas estaciones de muestreo. Annual variation of surface chlo-rophyll a at the sampling stations.

máximo en enero (26.5 ºC). No se registrarondiferencias verticales que permitieran definiruna estratificación térmica. El caudal de ingreso(Qi) y el caudal de salida (Qs) fueron altosdurante el invierno y menores en verano (Fig.5). Durante los meses de verano se registró unaumento del nivel del agua del embalse de apro-ximadamente 1 m. Consecuentemente con lasvariaciones de los aportes y salidas de agua y elmanejo del nivel de la cota, el tiempo de resi-dencia del embalse fue mayor en verano con unmáximo de 24 días en diciembre y menor eninvierno con un mínimo de 4 días en julio(media anual = 14 días). La temperatura delagua se correlacionó positivamente con el tiem-po de residencia (p<0.01) (Fig. 6A). La precipi-tación local medida en la estación meteorológi-ca cercana al área de estudio presentó uncomportamiento similar al tiempo de residencia,mostrando que la magnitud de dichos aportes nofueron significativos comparados con los apor-tes provenientes de la cuenca media y alta apor-tados por el cauce principal.
La concentración de sólidos suspendidos,fosfatos y fósforo total presentaron los valoresmáximos en invierno y se correlacionaron posi-tivamente entre sí (p<0.01) y negativamente conel tiempo de residencia (p<0.01) (Fig. 6B). Laconcentración de nitratos y silicatos (Fig. 6C),presentó un comportamiento diferente de los
parámetros anteriores, con máximos en losmeses de verano, relacionándose con el aumen-to de la precipitación local.
DISCUSIÓN
Composición y ciclo estacional
La comunidad fitoplanctóncia del embalse estu-vo dominada por Bacillariophyta, Cyanophyta yfitoflagelados nanoplanctónicos. Junto condichos grupos se determinaron una importantevariedad de especies de Chlorophyta y Eugle-nophyta y pocas especies de Chrysophyta,Cryptophyta y Dinophyta. Esta composición fuesimilar a la encontrada por Quirós y Lucchini(1983), en el mismo sistema y por Bonilla (1997)y Pérez et al. (1999), en los embalses sobre elRío Negro (Uruguay). La variedad de especies deChlorophyta, Bacillariophyta y Euglenophyta,coincidió con los resultados encontrados porO’Farrell e Izaguirre (1994), referidos a la com-posición del fitoplancton del Río Uruguay.Espacialmente, se registraron diferencias entre lacomposición taxonómica del cauce principal ylos brazos, principalmente debido a la dominan-cia de diatomeas en las primeras y la mayor den-sidad de cianobacterias en las últimas. Entre lasespecies de Aulacoseira, fue muy frecuente yabundante la especie A. ambigua y su forma spi-ralis. Esta especie no fue encontrada en otros ríosde la región, constituyendo un taxa característicodel Río Uruguay (O’Farrell, 1994).
Según estudios anteriores (O’Farrell, 1994;O’Farrell e Izaguirre, 1994 y O’Farrell et al.,1996), realizados en el Río Uruguay y en quinceimportantes ríos de la cuenca del Río de la Plataincluyendo diversos tramos del Río Paraná yafluentes, la composición de la comunidad fito-planctónica del Embalse de Salto Grande puededefinirse como eupotamoplanctónica.
De acuerdo con Reynolds (1988), la comuni-dad fitoplanctónica se estructura, básicamente,en relación a la disponibilidad de nutrientes yluz. En ambientes con disponibilidad denutrientes, predominan organismos oportunis-
Fitoplancton del embalse de Salto Grande 107
Figura 5. Variación anual de los parámetros físicos y quími-cos (media de las cuatro estaciones). A- temperatura y tiempode residencia. B- Sólidos suspendidos totales (SST), fosfatos(PO4) y fósforo total (PT). C- Precipitación local, nitratos(NO2-
3) y silicatos (SiO3-2). Annual variation of the physical
and chemical parameters (mean of the four sampling stations).A- temperature and retention time. B- Total suspended solids(SST), phosphates (PO4) and total phosphorus (PT). C- Localrainfall, nitrates (NO2-
3) and silicates (SiO3-2).
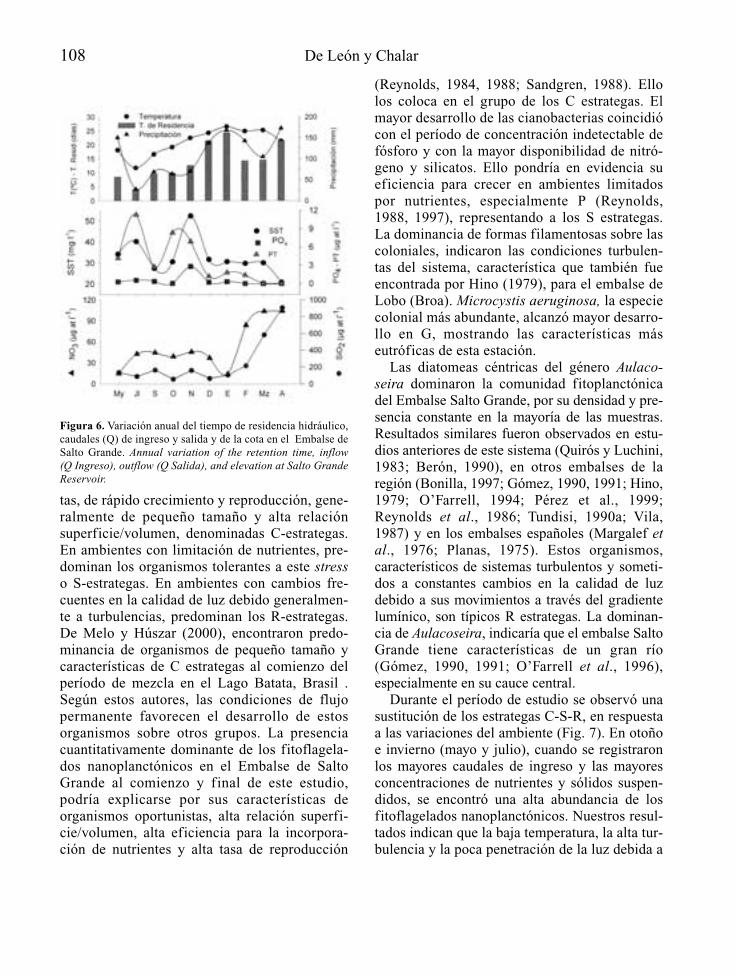
tas, de rápido crecimiento y reproducción, gene-ralmente de pequeño tamaño y alta relaciónsuperficie/volumen, denominadas C-estrategas.En ambientes con limitación de nutrientes, pre-dominan los organismos tolerantes a este stresso S-estrategas. En ambientes con cambios fre-cuentes en la calidad de luz debido generalmen-te a turbulencias, predominan los R-estrategas.De Melo y Húszar (2000), encontraron predo-minancia de organismos de pequeño tamaño ycaracterísticas de C estrategas al comienzo delperíodo de mezcla en el Lago Batata, Brasil .Según estos autores, las condiciones de flujopermanente favorecen el desarrollo de estosorganismos sobre otros grupos. La presenciacuantitativamente dominante de los fitoflagela-dos nanoplanctónicos en el Embalse de SaltoGrande al comienzo y final de este estudio,podría explicarse por sus características deorganismos oportunistas, alta relación superfi-cie/volumen, alta eficiencia para la incorpora-ción de nutrientes y alta tasa de reproducción
(Reynolds, 1984, 1988; Sandgren, 1988). Ellolos coloca en el grupo de los C estrategas. Elmayor desarrollo de las cianobacterias coincidiócon el período de concentración indetectable defósforo y con la mayor disponibilidad de nitró-geno y silicatos. Ello pondría en evidencia sueficiencia para crecer en ambientes limitadospor nutrientes, especialmente P (Reynolds,1988, 1997), representando a los S estrategas.La dominancia de formas filamentosas sobre lascoloniales, indicaron las condiciones turbulen-tas del sistema, característica que también fueencontrada por Hino (1979), para el embalse deLobo (Broa). Microcystis aeruginosa, la especiecolonial más abundante, alcanzó mayor desarro-llo en G, mostrando las características máseutróficas de esta estación.
Las diatomeas céntricas del género Aulaco-seira dominaron la comunidad fitoplanctónicadel Embalse Salto Grande, por su densidad y pre-sencia constante en la mayoría de las muestras.Resultados similares fueron observados en estu-dios anteriores de este sistema (Quirós y Luchini,1983; Berón, 1990), en otros embalses de laregión (Bonilla, 1997; Gómez, 1990, 1991; Hino,1979; O’Farrell, 1994; Pérez et al., 1999;Reynolds et al., 1986; Tundisi, 1990a; Vila,1987) y en los embalses españoles (Margalef etal., 1976; Planas, 1975). Estos organismos,característicos de sistemas turbulentos y someti-dos a constantes cambios en la calidad de luzdebido a sus movimientos a través del gradientelumínico, son típicos R estrategas. La dominan-cia de Aulacoseira, indicaría que el embalse SaltoGrande tiene características de un gran río(Gómez, 1990, 1991; O’Farrell et al., 1996),especialmente en su cauce central.
Durante el período de estudio se observó unasustitución de los estrategas C-S-R, en respuestaa las variaciones del ambiente (Fig. 7). En otoñoe invierno (mayo y julio), cuando se registraronlos mayores caudales de ingreso y las mayoresconcentraciones de nutrientes y sólidos suspen-didos, se encontró una alta abundancia de losfitoflagelados nanoplanctónicos. Nuestros resul-tados indican que la baja temperatura, la alta tur-bulencia y la poca penetración de la luz debida a
108 De León y Chalar
Figura 6. Variación anual del tiempo de residencia hidráulico,caudales (Q) de ingreso y salida y de la cota en el Embalse deSalto Grande. Annual variation of the retention time, inflow(Q Ingreso), outflow (Q Salida), and elevation at Salto GrandeReservoir.

la alta turbidez inorgánica, habrían limitado lapresencia de otras especies (Gliwitz, 1999;Reynolds, 1984, 1997; Sommer, 1993), favore-ciendo a las más eficientes en la incorporaciónde nutrientes, con alta tasa de crecimiento (estra-tegas C) y con mayor eficiencia fotosintéticapara desarrollarse en un ambiente turbio y turbu-lento (estrategas R). A finales de invierno y enprimavera (setiembre a diciembre) la disponibili-dad de nutrientes y el incremento de la radiaciónsolar y la temperatura favorecieron el creci-miento de las Chlorophyta de pequeño tamaño(C-estrategas). En noviembre, coincidiendo conuna alta concentración de SiO2 y el máximo dePO4 y SST, se registró un aumento de la abun-dancia de las Bacillariophyta (R-estrategas). Elmáximo poblacional alcanzado en diciembre, sereflejó en el incremento de la concentración declorofila a y en la disminución de los nutrientesal final de la primavera y verano (diciembre amarzo). Desde febrero hasta abril, la concentra-ción de P fue indetectable, determinando la pre-sencia de las Cyanophyta, tolerantes al stress denutrientes o S-estrategas. A partir de febrero dis-
minuye la abundancia de las Bacillariophyta yaumenta la población de fitoflagelados (C-estra-tegas), indicando el comienzo de un nuevo cicloa partir de otoño. Los resultados indican una pre-sencia de estrategas C en otoño e invierno, susti-tuidas por R en primavera y verano y éstas por Sal final del verano y otoño.
Abundancia, biomasa y diversidad
La abundancia del fitoplancton del embalse pre-sentó dos períodos claramente diferenciados: 1-abril a noviembre (invierno) y 2- diciembre amarzo (verano). Durante el primer período, laabundancia promedio fue inferior a 100 ind ml-1,similar a la registrada para lagos oligotróficos(Izaguirre et al., 1990), de régimen hidrológicovariable (García, 1980) y sistemas lóticos(García y Anselmi, 1989; Zalocar y Vallejos,1982) de la misma región. En cambio, la abun-dancia registrada a partir de diciembre, superiora 1000 ind ml-1, estuvo en el rango de losambientes lénticos (García y Anselmi, 1989)mesotróficos a eutróficos (Calijuri y Tundisi,
Fitoplancton del embalse de Salto Grande 109
Figura 7. Sucesión estacional de los estrategas fitoplanctónicos C, S y R y su relación con los factores ambientales en el EmbalseSalto Grande. Seasonal succession of phytoplanktonic C, S and R strategists, and its relationship with environmental factors at theSalto Grande Reservoir.

1990; García, 1977; Izaguirre y Vinocur, 1994a;Izaguirre et al., 1990) o de grandes ríos(O’Farrell y Izaguirre, 1994; Schiaffino, 1981;Zalocar y Vallejos, 1982).
La biomasa fitoplanctónica medida comoconcentración de clorofila a fue baja. Según laescala utilizada por Tundisi et al. (1988), enbase al índice de Carlson para 23 embalses delEstado de Sao Paulo, el sistema se clasificaríacomo ultraoligotrófico a oligotrófico (concen-tración promedio menor a 2.5 µg l-1). Según elíndice utilizado por Salas y Martino (1990) parasistemas cálidos tropicales, Salto Grande seríaoligotrófico (62 % de probabilidad (Chalar etal., 1993). El patrón de variación temporal de labiomasa fitoplanctónica fue igual al registradopor Montecino y Cabrera (1982), para el embal-se Rapel, un sistema eutrófico monomícticotemplado y por Bonetto et al. (1972) paraembalses de la región subtropical templada.
La diversidad del fitoplancton de SaltoGrande fue relativamente baja (2.98 bits ind-1
promedio, 0.5 – 4.1 bits ind-1), en relación a losvalores encontrados por O’Farrell y Izaguirre(1994), para el Río Uruguay aguas arriba (4.1-5.8 bits ind-1) y abajo del embalse (3.2-4.3 bitsind-1). Estos valores indican la alteración de lascondiciones ambientales del río al pasar de unrégimen fluvial a un embalse (Tundisi 1983,1990b), pero no pueden considerarse per se indi-cadoras de determinado estado trófico. Izaguirreet al. (1990), determinaron un rango de 0.67 a4.51 bits ind-1 en 20 cuerpos de agua ultra-oligoa eutróficos, de la región andino patagónica deArgentina. Sin embargo, no pudieron establecerdiferencias de nivel trófico en base al valor dediversidad. Lobo y Kobayasi (1990), encontra-ron mayor diversidad en ambientes con mayoríndice de saprobiedad, cuando esperaban lo con-trario. Los mejores indicadores de cambiosen las características tróficas de ríos y arroyosserían las variaciones en la composición y abun-dancia del fitoplancton, más que los índices dediversidad (Stoermer y Smol, 1999).
En Salto Grande, los valores más altos dediversidad se registraron en invierno, cuando ladensidad fitoplanctónica fue baja (< 100 ind ml-1)
y el sistema presentó características lóticas. Losmenores valores se registraron en el verano, en losbrazos cuando se observaron las mayores densida-des de organismos (> 1000 ind ml-1) y el sistemapresentó características más lénticas. Las comuni-dades sometidas a disturbios frecuentes de media-na intensidad presentan diversidad alta (Holzman,1993; Pádisak, 1993; Reynolds, 1993; Sommer,1993; Sommer et al., 1993). Esto indicaría que elrégimen lótico del embalse constituye un distur-bio frecuente para la comunidad, probable-mente debido a los pulsos provocados por losaportes de nutrientes, materiales particulados yla turbulencia generada en el sistema. Izaguirrey Vinocur (1994b) encontraron valores de diver-sidad entre 4.0 y 4.8 en ambientes lénticos de laregión con baja densidad de organismos (690 a16500 ind ml-1), coincidiendo con Branco ySenna (1996), que encontraron correlación nega-tiva entre densidad y diversidad. Según Margalef(1997), la disminución de la diversidad indica unincremento de la actividad metabólica de losorganismos, de modo que las variaciones dediversidad están asociadas a las característicasproductivas de la comunidad. La marcada estacio-nalidad de la abundancia y biomasa del fitoplanc-ton y de las características ambientales estudia-das, serían los factores determinantes de lasdiferencias temporales en la diversidad encontra-das en este sistema.
Estado Trófico
En la caracterízación trófica de los cuerpos deagua se utilizan diversos indicadores entre loscuales se encuentran la composición, abundan-cia, biomasa y diversidad fitoplanctónica. Lacomposición del Embalse Salto Grande estuvogeneralmente dominada por diatomeas deambientes eutróficos, coincidiendo con losresultados de Gómez (1991) en el Embalse delRio III. También fue similar a la composición delos embalses eutróficos analizados por Bonettoet al. (1976) y a las asociaciones eutróficas ymesotróficas determinadas por O’Farrell eIzaguirre (1994b). Según Duarte et al. (1992),los lagos dominados por diatomeas tendrían
110 De León y Chalar

características mesotróficas, mientras que losdominados por cianobacterias serían eutróficos(Lampert y Sommer, 1997; Margalef, 1983;Paerl, 1988; Pizzolón 1996; Scheffer et al.,1997). En Salto Grande, las cianobacterias fue-ron más abundantes en los brazos, especialmen-te en G. Berón (1990) y Quirós y Lucchini(1983), mencionaron la presencia de floracionesde M aeruginosa, prácticamente uniespecífica,entre abril y diciembre de 1980 en G. La densi-dad de M. aeruginosa registrada en este estudiofue alta pero no alcanzó a los valores reportadospor estos autores.
AGRADECIMIENTOS
Este trabajo fue financiado por el Convenioentre la Comisión Técnica Mixta de Salto Grandey la Facultad de Ciencias, Universidad de laRepública Oriental del Uruguay. Los autoresdesean agradecer a los compañeros de la Sec.Limnología por su colaboración en los muestreosy análisis químicos, a la Lic. Ma. del CarmenPérez por su asistencia en la identificación taxo-nómica del fitoplancton, al Lic. AlejandroOtaegui, Dto. de Ecología de la CTM, por suapoyo a esta investigación y al Dr. Oscar Parra,Centro EULA de la Universidad de Concepción-Chile, por sus sugerencias.
BIBLIOGRAFÍA
BERON, L. 1990. Features of the limnological beha-vior of Salto Grande’s reservoir (Argentina-Uruguay). Ecological Modelling, 52: 87-102.
BONILLA, S. 1997. Composición y abundancia fito-planctónica de tres embalses en cadena sobre el RíoNegro, Uruguay. Iheringia, ser. Bot., 49:47-61.
BONETO, A., D. DI PERSIA, R. MAGLIANESI yM. CORIGLIANO. 1976. Caracteres limnológi-cos de algunos lagos eutróficos de embalse de laregión central de Argentina. Ecosur, 3(5): 47-120.
BRANCO, C. y P. SENNA. 1996. Phytoplanktoncomposition, community structure and seasonalchanges in a tropical reservoir (Paranoa Reservoir,Brazil). Algological Studies, 81: 69-84.
CALIJURI, M. C. y J. G. TUNDISI. 1990.Limnologia comparada das Rs do Lobo (BROA) eBarra Bonita. Estado de Sâo Paulo. Mecanismosde funcionamento e bases para o gerenciamento.Rev. Brasil. Biol., 50 (4): 893-913.
CHALAR, G., L. DE LEON, R. DE LEON, D.FABIAN y J. GORGA. 1993. Evaluación de laeutrofización del Embalse de Salto Grande. Análisisde las relaciones entre los parámetros físico-quími-cos y biológicos. Informe Final de la primer etapa.CTM-SG/Universidad de la República–Facultad deCiencias-Sección Limnología.
CONDE, D., W. PINTOS, J. GORGA, R. DE LEON,G. CHALAR y R. SOMMARUGA. 1997. Themain factors inducing chemical spatial heteroge-neity in the Salto Grande, a reservoir on theUruguay River. Arch. Hydrobiol. Suppl. 113.Large Rivers, 10 (1-4): 571-578.
DE MELO, S. y V. HUSZAR. 2000. Phytoplanktonin an amazonian flood-plain lake (Lago Batata,Brazil): diel variation and species strategies. J.Plankton. Res., 22(1): 63-76.
DUARTE, C. M., S. AGUSTÍ y D. CANFIELD.1992. Patterns in phytoplankton community struc-ture in Florida Lakes. Limnol. & Oceanogr.,37(1): 155-161.
GARCÍA, M. O. 1980. Fitoplancton de una lagunadel valle aluvial del Paraná medio (“LosMatadores”, Santa Fe, Argentina). I. Estructura ydistribución en relación a factores ambientales.Ecología, Argentina, 4: 127-140.
GARCÍA, M. O. 1977. Ciclo anual del fitoplanctonen el Embalse San Roque (Córdoba, Argentina).Rev. Asoc. Cienc. Nat. Litoral, 8: 1-12.
GARCÍA, M. O. y M. I. ANSELI. 1989. Fitoplanctony variables ambientales en la cuenca del RíoSaladillo (Sta. Fe, Argentina). Rev. Brasil. Biol.,49(4): 957-967.
GLIWICZ, Z. M. 1999. Predictability of seasonal anddiel evens in tropical ans temperate lakes and reser-voirs. En: Theoretical Reservoir ecology and itsApplications. J. G. Tundisi & M. Straskaba (eds.):99-124. International Institute of Ecology, BrazilianAcademy of Sciences and Bachuys Publishers.
GOMEZ, N. 1990. Variaciones morfológicas deAulacoseira granulata (Ehrenberg.) Simonsen(Bacillariophyceae) en el embalse Río III,Córdoba, Argentina. Biota, Chile, 6: 45-51.
GOMEZ, N. 1991. Poblaciones de diatomeas planc-tónicas en una R subtropical: Embalse Río III,Argentina. Annls. Limnol., 27(1): 3-14.
Fitoplancton del embalse de Salto Grande 111

HASLE, F., 1978. Counting phytoplankton. En :Phytoplankton Manual. Monographs inOceanographic Methodology. Sournia, A. (ed.).UNESCO, 6: 88 - 96.
HINO, K. 1979. Analise qualitativa e quantitativa domicrofitoplancton da R do Lobo (BROA) SâoCarlos, Sâo Paulo. Tese de Maestrado. Univ.Federal de Sâo Carlos-SP-Brasil.
HOLZMAN, R. 1993. Seasonal fluctuations in thediversity and compositional stability of phyto-plankton communities in small lakes in upperBavaria. Hydrobiol., 249: 101-109.
IZAGUIRRE, I. y A. VINOCUR. 1994a. Typology ofshallow lakes of the Salado River Basin(Argentina), based on phytoplankton communii-ties. Hydrobiol., 277:49-62
IZAGUIRRE, I. y A. VINOCUR. 1994b. Algalassemblages from shallow lakes of the SaladoRiver Basin (Argentina). Hydrobiol., 289:57-64
IZAGUIRRE, I., del GIORGIO, I. O’FARRELL y G.TELL. 1990. Clasificación de 20 cuerpos de aguaandino-patagónicos (Argenitna) en base a laestructura del fitoplancton estival. Cryptogamie,Algol., 11: 31-46.
LAMPERT, W. y U. SOMMER. 1997.Limnoecology. Oxford University Press.
LOBO, E. y H. KOBAYASI. 1990. Shannon’sDiversity Index applied to some diatoms assem-blages in the Sakawa River system (KanagawaPref., Japan) and its use as an indicator of waterquality. Jpn. J. Phycol. (Sôrvi), 38: 229-243.
MARGALEF, R. 1997. Our Biosphere. En:Excellence in Ecology nº 10. O. Kinne (Ed.)..Ecology Institute – Germany.
MARGALEF, R., D. PLANAS, J. ARMENGOL, A.VIDAL, N. PRAT, A. GUISET, J. TOJA y M.ESTRADA. 1976. Limnología de los embalsesespañoles. Ministerio de Obras Públicas. Madrid.
MARGALEF, R. 1983. Limnología. Omega.MONTECINO, V. y S. CABRERA. 1982.
Phytoplankton activity and standing crop in animpoundment of central Chile. J. Plankton Res.,4(4).
NUSCH, E.A. 1980. Comparison of differentmethods for chlorophyll and phaeopigmentsdetermination. Arch. Hydrobiol. Beih. Ergebn.Limnol., 17:14-36.
O’FARRELL, I. 1994. Comparative analysis of thephytoplankton of fifteen lowland fluvial systemsof the River Plate Basin (Argentina). Hydrobiol.,289: 109-117.
O’FARRELL, I., I. IZAGUIRRE y A. VINOCUR.1996. Phytoplankton ecology of the Lower ParanáRiver (Argentina). Arch. Hydrobiol. Suppl.,115/Large Rivers, 11 (1): 75-89.
O’FARRELL, I y I. IZAGUIRRE. 1994.Phytoplankton ecology and limnology of a RiverUruguay Lower Basin (Argentina). Arch.Hydrobiol./Suppl., 99 (Monographische Beiträge)(1/2): 155-179.
PAERL, H. 1988. Growth and reproductive strategiesof freshwater blue-green algae (cianobacteria).En: Growth and reproductive strategies of fres-hwater phytoplankton. Sandgren C. (ed.).Cambridge University Press.
PARRA, O. y C. BICUDO. 1995. Introducción a labiología y sistemática de las algas de aguas conti-nentales- Univ. De Concepción.
PÉREZ, M. C.; S. BONILLA y G. MARTÍNEZ.1997. Phytoplankton community of a polymicticreservoir, La Plata River Basin, Uruguay. Rev.Brasil. Biol., 59(4): 353-541.
PIZZOLON, L. 1996. Importancia de las cianobacte-rias como factor potencial de toxicidad en lasaguas continentales. Interciencia, 21: 239-245.
PLANAS, D. 1975. Distribution and productivity ofthe phytoplankton in Spanish reservoirs. Verh.Internat. Verein. Limnol., 19: 1860-1870.
QUIRÓS, R y L. LUCHINI. 1983. Característicaslimnológicas del embalse de Salto Grande III:Fitoplancton y su relación con parámetrosambientales. Rev. Asoc. Cienc. Nat. Litoral, 13:19-66.
REYNOLDS, C. S., 1997. Vegetations processes inthe pelagic: a model for eccosystem theory. En:Excellence in Ecology nº 9. Kinne, O. (Ed.).Ecology Institute, Germany.
REYNOLDS, C. S., 1993. Scales of disturbance andtheir importance in plankton ecology. Hydrobiol.,249: 157-171.
REYNOLDS, C. S., 1988. Functional morphologyand adaptative strategies of freshwater phyto-plankton. En: Growth and reproductive strategiesof freshwater phytoplankton. Sandgren C. (Ed.).Cambridge University Press.
REYNOLDS, C. S. 1984. The ecology of freshwaterphytoplankton. Cambridge University Press.
REYNOLDS, C.; V. MONTECINO; M. GRAF y S.CABRERA. 1986. Short-term dynamics of aMelosira populations in the plankton of animpoundment in Central Chile. J. Plankton Res.,8(4): 715-740.
112 De León y Chalar

SANDGREN C. (Ed.). 1988. Ecology and reproduc-tive strategies of freshwater phytoplankton.Cambridge Univ. Press.
SALAS, H. J. y P. MARTINO. 1990. Metodologíassimplificadas para la evaluación de la eutrofiza-ción en lagos cálidos tropicales. CEPIS/HPE/OPS.
SCHEFFER, M., S. RINALDI, A. GRAGNANI, L.MUR Y E. VANNES. 1997. On the dominance offilamentous cyanobacteria in shallow turbid lakes.Ecology, 78: 272-282.
SCHIAFFINO, M. 1981. Campaña limnológica“Keratella I” en el Río Paraná medio. Rev. Asoc.Cienc. Nat. Litoral, 12: 140-147.
SOMMER, U. 1993. Disturbance-diversity relationshipsin two lakes of similar nutrient chemistry but contras-ting disturbance regimes. Hydrobiol., 249: 59-65.
SOMMER, U., J. PADISAK, C. REYNOLDS y P.JUHASZ-NAGY. 1993. Hutchinson’s heritage: thediversity – disturbance relationship in phytoplank-ton. Hydrobiol., 249-1-7.
STOERMER, E. y J. SMOL. 1999. The DiatomsApplications for the Environmental and EarthSciences. Cambridge Univ. Press.
TUNDISI, J. G. 1990a. Distribuiçâo espacial,sequência temporal e ciclo sazonal do fitoplânc-
ton em Reservatorios: Fatores limitantes e contro-ladores. Rev. Brasil Biol., 50(4): 937-955.
TUNDISI, J. G. 1990b. Limnology and eutrophica-tion of Barra Bonita Reservoir, S. Paulo State,Southern Brazil. Arch. Hydrobiol. Beih. Ergebn.Limnol., 33: 661-676.
TUNDISI, J. G. 1983. A review of basic ecologicalprocesses interactin with production and standing-stock of phytoplankton in lakes and reservoirs inBrazil. Hydrobiol., 100: 223-243.
TUNDISI, J. G., T. MATSUMURA-TUNDISI, R.HENRY, O. ROCHA y K. HINO. 1988.Comparaçâo do estado trófico de 23 reservató-rios do Estado de Sao Paulo: Eutrof izaçâo emanejo. En: Limnologia e Manejo de Reserva-tórios. V. 1, T. 1. J. G. Tundisi (ed.). Universidadde Sao Paulo.
VILA, I., I. BARENS y V. MONTECINO. 1987.Abundancia y distribución temporal del fitoplanc-ton en el Embalse Rapel, Chile Central. Rev.Chilena de Hist. Nat., 60: 37-55.
ZALOCAR, Y. y E. VALLEJOS. 1982. Fitoplanctondel Río Alto Paraná. Variación estacional y distri-bución en relación a factores ambientales. Ecosur,9(17): 1-28.
Fitoplancton del embalse de Salto Grande 113