Embed Size (px)
Citation preview

NUM 11. MAYO-AGOSTO 2011
Los RNAs pequeños no codificantes comomoduladores de la expresión génica
pág. 10
Identificación de la proteína eIF-5a dependientede poliaminas en Trichomonas vaginalis
pág. 21
Aspectos genéticos de algunasarritmias hereditarias.
pág. 7

PLANTA ACADÉMICA
Dra. Esther Orozco O.Rectora de la UACM
y Fundadora del Posgrado
Dra. Elizbeth ÁlvarezDra. Elisa Azuara
Dra. Minerva CamachoDr. César López Camarillo
Dr. Jesús FandiñoDra. Mavil López Casamichana
Dr. Humberto NicoliniDr. José de Jesús Olivares
Dra. Martha YocupicioDr. Mauricio Castañón
Dra. Selene ZárateDra. Itzel Quintas
RESPONSABLE DE LA EDICIÓN DE ESTE NÚMERO
Dra. Mavil López Casamichana
RESPONSABLE DE GENÓMICAS HOY
Dr. César López Camarillo
Posgrado en Ciencias GenómicasUniversidad Autónoma de la Ciudad de México
PLANTEL DEL VALLE
Avenida San Lorenzo 290, Colonia Del ValleDelegación Benito Juárez, C.P. 03100, Ciudad de México
5488 6661 ext. 15313http://www.uacm.edu.mx/genomicas
Publicación cuatrimestral, 2500 ejemplares.
Contenido

1
Nuestros Investigadores pág. 2
Publicaciones científicas recientes del PCG-UACM pág. 3
Noticias del PGC pág. 4
Anuncios pág. 5
De nuestros colaboradores pág. 7
Los RNAs pequeños no codificantes como moduladores de la expresión génica pág. 10
La Proteómica como herramienta para el estudio de las células T. pág. 16
Identificación de la proteína eIF-5a dependiente de poliaminas en Trichomonas vaginalis pág. 21
Noticias del mundo de la Ciencia pág. 25
Nuevos proyectos del PGC-UACM con financiamiento pág. 27
Graduados pág. 28
Participación del PCG-UACM en congresos nacionales e internacionales pág. 29
CienciArte: Requiem en Sol Mayor pág. 31
Desde el portaobjetos pág. 32
Contenido
La academia del PCG se conduele ante el reciente fallecimiento del Dr. Leobardo Mendoza Alcántar, profesor de la Escuela Su-perior de Medicina del Instituto Politécnico Nacional, y se une solidariamente a la congoja y resignación que embarga a sus familiares y seres queridos.
El Dr. Mendoza, quien fuera nuestro colega desde el año 2004 hasta el 2007, implementó el área de Microscopía Confocal del PCG donde se desempeñó exitosamente y, sin lugar a dudas,
contribuyó al avance de numerosos proyectos de investigación científica desarrollados tanto por pro-fesores y estudiantes de la UACM, como de otras instituciones educativas.
Ante su dolorosa pérdida física, no sólo quedan para la posteridad, su desempeño como académico, su trayectoria científica, su ejemplo de vida y su calidad como ser humano; también, desde esta trinchera, subsistirá perennemente su valiosa contribución al crecimiento del, en aquel entonces, naciente Posgrado en Ciencias Genómicas.

2
Nuestros
Dra. Mavil López CasamichanaProfesora InvestIgadora del
Posgrado en CIenCIas genómICas
Oriunda de la Ciudad de La Habana, Cuba, obtuvo el título profesional de Licenciada en Bioquímica por la Facultad de Ciencias Biológicas de la Univer-sidad de la Habana. Arribó a México para cursar la Maestría en Ciencias con especialidad en Bio-química en el Centro de Investigación y de Estu-dios Avanzados del Instituto Politécnico Nacional y, posteriormente, se incorporó a la Universidad Autónoma de la Ciudad de México donde egresó con mención honorífica del Doctorado en Ciencias Genómicas en el año 2008.
Dentro de la planta académica del PCG, la Dra. Ló-pez Casamichana se desempeña como profesora de los cursos de posgrado “Metabolismo Celular e Introducción a la Proteómica”, “Antropología Molecular” y “Genética Forense”. Además, se encuentra a cargo del Laboratorio de Análisis y Diagnóstico de ADN del PCG, donde desarrolla las líneas de investigación en Genética Forense y Diag-nóstico Molecular. Desde el año 2010 es miembro del Sistema Nacional de Investigadores.
Actualmente, dirige varios proyectos de investiga-ción científica que vinculan la formación integral de estudiantes de maestría y doctorado del PCG con la generación de nuevos conocimientos, entre ellos se encuentran:
Polimorfismos genéticos en el establecimiento de la pigmentación humana y su utilidad en el área forense.
Análisis molecular de marcadores polimórficos de utilidad forense en una población de occisos des-conocidos de la Ciudad de México.
Estudio del fenotipo bioenergético celular de neo-plasias intraepiteliales cervicales de bajo y alto gra-do.
INvestIgadores

3
Carvajal-Gamez BI, Arroyo R, Camacho-Nuez M, Lira R, Martínez-Benitez M, Alvarez-Sán-chez ME. Putrescine is required for the expres-sion of eif-5a in Trichomonas vaginalis. Mol Bio-chem Parasitol 180(1):8-16. 2011.
Alcaraz-Estrada, Sofia Lizeth; Yocupicio-Mon-roy Martha and del Angel Rosa María. Insights into dengue virus genome replication. Future Vi-rology. 5: 575-592. 2011.
C. López-Camarillo, E. Azuara-Liceaga, A. Zamorano, O. Hernández de la Cruz, I. López Rosas, Esther Orozco, L. Marchat. Genom-ics and proteomics approaches to understand virulence of Entamoeba histolytica. In Science against microbial pathogens: communicating
current research and technological advances. A. Méndez-Vilas Ed. Spain. 2011
Carlos Pérez-Plasencia, Patricia Piña-Sanchez, Fe-lipe Vaca-Paniagua, Veronica Fragoso-Ontiveros, Nadia Jacobo-Herrera, Cesar Lopez-Camarillo and Omar Vargas-Hernández. Molecular events towards Wnt pathway activation in Cervical Cancer: changing the balance on NKD/DVL sig-
nals. In Human Papillomavirus /Book 2. ISBN 979-953-307-684-2. In Tech Publishers. 2011.
Urraca N, Camarena B, Aguilar A, Fresán A, Apiquian R, Orozco L, Carnevale A, Nicolini H. Association study of DRD3 gene in schizophrenia in mexican sib-pairs. Psychiatry Res. Jul 5, 2011.
Fernandez-Sanchez V, Pelayo R, Flores-Guzman P, Flores-Figueroa E, Villanueva-Toledo J, Garrido E, Ruiz-Sanchez E, Alvarez-Sanchez E, Mayani H. In vitro effects of stromal cells expressing differ-ent levels of Jagged-1 and Delta-1 on the growth of primitive and intermediate CD34+ cell subsets
from human cord blood. Sep 11 (Epub ahead of print) (in press)
Absalom Zamorano-Carrillo, Beatriz Zamo-ra-López, Claudia-Guadalupe Benítez-Cardoza, Rosas-Trigueros Jorge, César
López-Camarillo and Laurence A Marchat. Transcriptional Networks: Dynamic Information Fluxes. In Bioinformatics: Genome bioinformatics and computational biology. ISBN 978-1-62100-913-9. Nova Science Publishers. USA. 2011.
PUBLICACIONES CIENTÍFICASrecientes del PCG
La pubLicación de resuLtados de Los proyectos de investigación en revistas científicas internacionaLes con arbitraje estricto, constituye un importante indicador de La caLidad e impacto de Las investiga-ciones reaLizadas en eL pcg-uacm.
Imag
en: P
aul &
Lin
dam
arie
Am
bros
e

4
NOTICIAS DEL PCG
investigaciones deL pcg generan Las primeras patentes registradas de La uacm
abordado, cuando se presenta la tricomonosis pue-den encontrarse consecuencias como desarrollo de cáncer de próstata e infertilidad, principalmente.
La Dra. María Elizbeth Álvarez Sánchez, profesora investigadora y actual coordinadora del Posgrado en Ciencias Genómicas de nuestra institución jun-to con su grupo de trabajo propuso posibles bio-marcadores para la tricomonosis en el hombre. Sus proyectos científicos han contado con el apoyo de la UACM, del Instituto de Ciencia y Tecnología del DF y del CONACYT.
En entrevista a los medios de prensa, el pasado 27 de septiembre, la Dra. Álvarez-Sánchez señaló:“El proyecto está enfocado al análisis proteómi-co de este parásito, y una línea es precisamente
encontrar moléculas que puedan ser utilizadas como biomarcadores de la trico-monosis en el hombre, este estudio se abordó por ser el hombre portador de esta infección y por lo tanto ser potencial transmisor de este patógeno.
Como resultado del esfuerzo encausado para en-contrar biomarcadores para el diagnóstico de la tricomonosis, enfermedad de transmisión sexual de alta incidencia mundial causada por el pará-sito Trichomonas vaginalis, la UACM registró sus dos primeras patentes nacionales ante el Instituto Mexicano de la Propiedad Intelectual (IMPI). La tri-comonosis, es una infección que afecta el tracto urogenital del humano. Las mujeres son las que presentan mayor sintomatología y las que pueden presentar complicaciones tales como ruptura pre-matura de membranas, abortos, predisposición a la adquisición del cáncer cérvico-uterino, enferme-dad pélvica crónica y es un factor predisponente en la adquisición del virus de inmunodeficiencia hu-mana VIH. En el caso del hombre, la mayoría son asintomáticos por lo que su estudio ha sido poco

5
Se solicitan estudiantes para realizar su tesis de licenciatura y posgrado en el estudio de la expresión génica y proteómica de T. vaginalis.Se desarrollaran proyectos en mecanismos de regulación de la expresión génica de T. vaginalis mediado por poliaminas así como proyec-tos en expresión proteómica de la tricomonosis en hombres. Requisitos: Estar inscrito en una institución de educación superior. Contar al menos con el 75% de los créditos de la licenciatura y con promedio general no menor a 8.0. Informes: Dra.Elizbeth Alvarez Sánchez.Tel: 5488-6661 ext. 15306.Email: [email protected]
Solicito dos estudiantes de licenciatura para desarrollar proyectos en Genómica y Proteómica del Cáncer de Mama.Se desarrollarán proyectos de investigación enfocados al análisis funcional de microRNAs y análisis proteómico de biopsias de carcinomas mamarios. Requisitos: Estar inscrito en una institución de educación superior. Contar al menos con el 75% de los créditos de la licencia-tura con promedio general mayor a 8.0.Informes: Dr. César López-Camarillo. Tel: 5488-6661 ext. 15307 y 15312.Email: [email protected]
Solicito estudiantes para realizar su tesis de licenciatura, servicio social y/o en proyectos relacionados con el parásito Enta-moeba histolytica.Requisitos. Estar inscrito en una institución de educación superior. Contar con al menos el 75% de los créditos de la licenciatura.Informes: Dra. Elisa Azuara, Posgrado en Ciencias. Tel: 5488-6661 ext. 15312.Email: [email protected]
La 4° edición del ya tradicional Diplomado en Investigación Ge-nómica, organizado por el Posgrado en Ciencias Genómicas de la Universidad Autónoma de la Ciudad de México y el Instituto de Ciencia y Tecnología del Distrito Federal, cerró exitosamente su programa mensual de sesiones el pasado 25 de julio de 2011.
Un total de 49 especialistas nacionales y 10 internacionales se pre-sentaron en el Auditorio del Plantel del Valle para dar a conocer los más recientes avances de sus investigaciones científicas en Ge-nómica, Transcriptómica, Proteómica, Fenómica, Farmacogenómi-ca, Oncogenómica, Genética Forense y Antropología Molecular, Metabolómica e Infectómica. Tal y como se ha concebido, este tipo actividades tiene como propósito despertar el interés de los asistentes hacia la investigación genómica y sus avances científicos
ANUNCIOS
se soLicitan estudiantes de Licenciatura para reaLizar tesis, servicio sociaL y/o posgrado en proyectos de investigación genómica y proteómica en eL pcg-uacm.
concLuye exitosamente eL
4o. dipLomado en investigación genómica
y tecnológicos; además de proporcionar conocimiento actualiza-do de la mano de expertos, tales como herramientas científicas y metodológicas para el diseño e impulso de estrategias enfocadas a solventar problemas de impacto de nuestra sociedad. Al evento asistieron 262 participantes y se recibieron 114 trabajos, de los cuales se certificaron 83 con altos estándares de calidad.
Por su elevado nivel científico, originalidad, profunda labor de investigación, capacidad de síntesis y revisión bibliográfica actua-lizada, 4 tesinas certificadas recibieron un reconocimiento como las “Mejores Tesinas” en la ceremonia de clausura. Una versión corta de estas tesinas será publicada en la Gaceta Genómicas hoy. Las tesinas premiadas fueron las siguientes:

6
1° Lugar -“Los RNAs pequeños no codificantes como moduladores la expresión génica”Autores: Frías Soria Christian Lizette y Cervantes Villagrana Rodolfo
2° Lugar -“La proteómica como herramienta para el estudio de las células T” Autora: López Torres Donají.
3° Lugar -“Genotipificación del virus de la hepatitis B (VHB) y su resistencia a fármacos antivirales” Autores: Briones Peña Saida Jessica, Fuentes Romero Luis León y Rodríguez Díaz Roberto Alejandro.
4° Lugar -“Implicación de los procesos de acetilación y desacetilación de histonas en el desarrollo del cáncer” Autores: Guerrero Parra Hilda Adriana y Solís Paredes Juan Mario.

7
Dra. Angélica Rueda y Sánchez de la VegaDepartamento de BioquímicaCentro de Investigación y deEstudios Avanzados del IPN
55-57473800 Ext. [email protected]
http://www.biochem.cinvestav.mx
Palabras clave: Arritmias hereditarias, canales ióni-cos, receptor de rianodina, corazón.
INTRODUCCIÓN
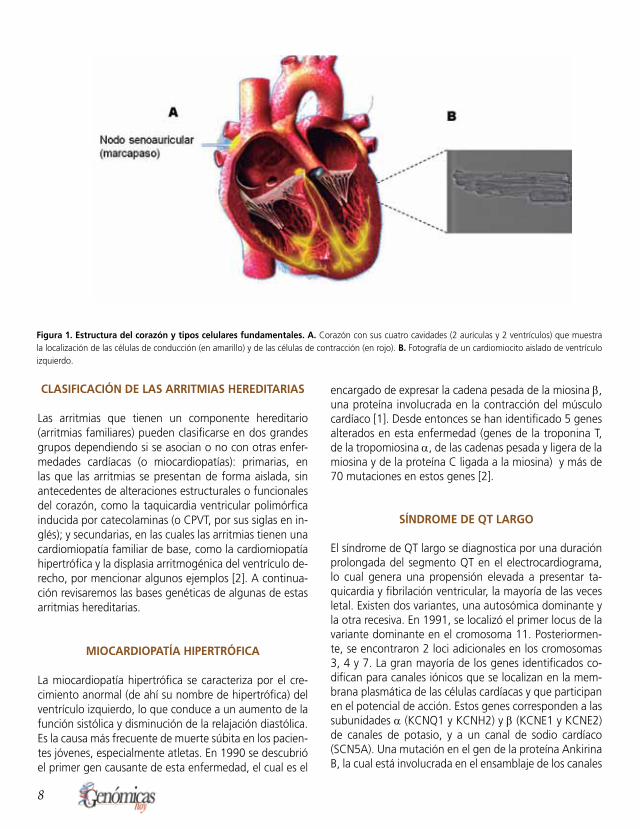
Las arritmias son variaciones anormales de la frecuencia y del ritmo cardíaco que se originan por alteraciones del ci-clo, ya sea a nivel de las fibras de conducción del corazón que son las encargadas de propagar el impulso eléctrico que antecede a la contracción de aurículas y ventrículos; o bien, a nivel de las células contráctiles, o cardiomioci-tos, que inician eventos automáticos y espontáneos, no coordinados con el impulso eléctrico (Fig.1). Dependien-do de si la arritmia induce una disminución o un aumento en la frecuencia cardíaca, ésta se clasifica como bradi-cardia o taquicardia, respectivamente. Adicionalmente, existen arritmias letales que generan un patrón irregular del ritmo cardíaco y que pueden producir fibrilación y muerte súbita. Fue en 1990 que se identificó el primer gen causante de una cardiomiopatía [1]. Desde entonces, muchos centros de investigación han dedicado sus esfuerzos a la identificación de los genes causantes de las arritmias hereditarias, principalmente aquellas monogenéticas, ya sean dominantes o recesivas. Estas enfermedades son
DE NUESTROS COLABORADORES:Aspectos genéticos de algunasarritmias hereditarias
Las arritmias son una de Las mayores causas de mortaLidad en La pobLación, especiaLmente en aqueLLos pacientes con enfermedad cardiaca previa. Los avances actuaLes en Las técnicas de bioLogía moLecuLar y genética humana apLicadas a La medicina genómica nos están LLevando a La identificación de Las aLte-raciones genéticas responsabLes de aLgunas de Las arritmias cardiacas hereditarias. a pesar de que La cardioLogía tardó en incorporar estos enfoques experimentaLes, Los avances actuaLes en La cardioLogía moLecuLar han sido espectacuLares. en esta úLtima década se han descubierto y reportado numerosas mutaciones puntuaLes en genes causantes de enfermedades cardiacas famiLiares. actuaLmente, La tera-pia cardiovascuLar incLuye medicamentos diseñados mediante ingeniería genética y muchos proyectos con terapias génicas están en marcha en diversos Laboratorios deL mundo. en Los próximos años pre-senciaremos numerosos descubrimientos de Los mecanismos moLecuLares que participan en La generación de Las arritmias hereditarias. sin duda aLguna, Las personas con afecciones cardiacas hereditarias se beneficiarán en muy corto pLazo de todos estos avances en La medicina genómica, aL tener a su aLcance mejores opciones terapéuticas y preventivas.
poco frecuentes en la población, pero al ser un solo gen el responsable del desarrollo de la patología, se facilita el estudio detallado de las alteraciones moleculares que resultan de la expresión del gen alterado, permitiendo incluso desarrollar animales transgénicos que albergan uno o los dos alelos del gen mutado
8
encargado de expresar la cadena pesada de la miosina β, una proteína involucrada en la contracción del músculo cardíaco [1]. Desde entonces se han identificado 5 genes alterados en esta enfermedad (genes de la troponina T, de la tropomiosina α, de las cadenas pesada y ligera de la miosina y de la proteína C ligada a la miosina) y más de 70 mutaciones en estos genes [2].
SÍNDROME DE QT LARGO
El síndrome de QT largo se diagnostica por una duración prolongada del segmento QT en el electrocardiograma, lo cual genera una propensión elevada a presentar ta-quicardia y fibrilación ventricular, la mayoría de las veces letal. Existen dos variantes, una autosómica dominante y la otra recesiva. En 1991, se localizó el primer locus de la variante dominante en el cromosoma 11. Posteriormen-te, se encontraron 2 loci adicionales en los cromosomas 3, 4 y 7. La gran mayoría de los genes identificados co-difican para canales iónicos que se localizan en la mem-brana plasmática de las células cardíacas y que participan en el potencial de acción. Estos genes corresponden a las subunidades α (KCNQ1 y KCNH2) y β (KCNE1 y KCNE2) de canales de potasio, y a un canal de sodio cardíaco (SCN5A). Una mutación en el gen de la proteína Ankirina B, la cual está involucrada en el ensamblaje de los canales
CLASIFICACIÓN DE LAS ARRITMIAS HEREDITARIAS
Las arritmias que tienen un componente hereditario (arritmias familiares) pueden clasificarse en dos grandes grupos dependiendo si se asocian o no con otras enfer-medades cardíacas (o miocardiopatías): primarias, en las que las arritmias se presentan de forma aislada, sin antecedentes de alteraciones estructurales o funcionales del corazón, como la taquicardia ventricular polimórfica inducida por catecolaminas (o CPVT, por sus siglas en in-glés); y secundarias, en las cuales las arritmias tienen una cardiomiopatía familiar de base, como la cardiomiopatía hipertrófica y la displasia arritmogénica del ventrículo de-recho, por mencionar algunos ejemplos [2]. A continua-ción revisaremos las bases genéticas de algunas de estas arritmias hereditarias.
MIOCARDIOPATÍA HIPERTRÓFICA
La miocardiopatía hipertrófica se caracteriza por el cre-cimiento anormal (de ahí su nombre de hipertrófica) del ventrículo izquierdo, lo que conduce a un aumento de la función sistólica y disminución de la relajación diastólica. Es la causa más frecuente de muerte súbita en los pacien-tes jóvenes, especialmente atletas. En 1990 se descubrió el primer gen causante de esta enfermedad, el cual es el
Figura 1. Estructura del corazón y tipos celulares fundamentales. A. Corazón con sus cuatro cavidades (2 aurículas y 2 ventrículos) que muestra la localización de las células de conducción (en amarillo) y de las células de contracción (en rojo). B. Fotografía de un cardiomiocito aislado de ventrículo izquierdo.

9
iónicos a la membrana plasmática también está asociada al tipo 4 de síndrome de QT largo [3].
CPVT
La taquicardia ventricular polimórfica inducida por cate-colaminas o CPVT (por sus siglas en inglés) es una arrit-mia hereditaria que se caracteriza por la aparición de una taquicardia ventricular inducida por estrés o ejercicio. En la mayoría de los casos, esta taquicardia produce la fi-brilación ventricular y conduce a la muerte súbita de las personas que la presentan, sin muestra de alteraciones estructurales de su corazón. Esta arritmia hereditaria se presenta en dos formas: dominante y recesiva. La forma dominante de esta arritmia se encontró asociada a mu-taciones en el gen que expresa al canal de calcio (Ca2+) intracelular, comúnmente conocido como receptor de rianodina (RyR); mientras que la forma recesiva está aso-ciada a mutaciones en el gen de la calsecuestrina (CSQ), proteína que se encuentra en el lumen del retículo sarco/endoplámico (RS), que tiene una gran capacidad para unir Ca2+ y que participa junto con triadina y junctina en modular la respuesta del RyR al Ca2+ luminal [4].
Las primeras mutaciones puntuales del RyR asociadas a la presencia de CPVT (entre ellas la R4497C-RyR) se iden-tificaron en 4 familias diferentes que presentaban una taquicardia ventricular hereditaria inducida por estrés o ejercicio [5]. Desde entonces se han identificado 155 mutaciones para el gen de RyR y 12 para el gen de CSQ que originan variantes de estas proteínas, las cuales están asociadas a la presencia de CPVT [4].
Debido a la importancia que tiene el determinar las bases genéticas y moleculares de las arritmias, se han generado ratones transgénicos que expresan al RyR con alguna de las mutaciones puntuales encontradas en CPVT. Éste es el caso del ratón knock in para la variante R4497C-RyR [6], el cual recapitula muchas de las características fenotípicas de los pacientes con CPVT. Los cardiomiocitos ventricu-lares del ratón heterocigoto presentan alteraciones en su manejo del Ca2+ intracelular debidas a un aumento en la sensibilidad del RyR al Ca2+ del citoplasma, lo cual genera eventos arritmogénicos, como contracciones de los car-diomiocitos fuera del ritmo de estimulación [6]. Ésto ha permitido dilucidar las bases moleculares de una arritmia hereditaria lo cual permitirá, en un futuro, diseñar he-rramientas farmacológicas para el tratamiento de estas afecciones cardÍacas.
AGRADECIMIENTOS
Al Fondo SEP-Conacyt Ciencia Básica (proyecto No. 80960) y al Instituto de Ciencia y Tecnología del Distrito Federal (ICyTDF, proyecto No.331).
REFERENCIAS
[1] Geisterfer-Lowrance AA, Kass S, Tanigawa G, Vosberg HP, McKenna W, Seidman CE, Seidman JG. A molecular basis for familial hypertrophic cardiomyopathy: a beta cardiac myosin heavy chain gene missense mutation. Cell 1990, 62:999-1006.[2] Campuzano O, Beltrán-Alvarez P, Iglesias A, Scornik F, Pérez G, Brugada R. Genetics and cardiac channelopathies. Genet Med 2010, 12:260-7.[3] Inherited arrhythmias database. http://www.fsm.it/cardmoc/[4] Cerrone M, Napolitano C, Priori SG. Catecholaminergic po-lymorphic ventricular tachycardia: A paradigm to unders-tand mechanisms of arrhythmias associated to impaired Ca(2+) regulation. Heart Rhythm 2009, (11):1652-9. Review[5] Priori SG, Napolitano C, Tiso N, Memmi M, Vignati G, Bloise R, Sorrentino V, Danieli GA. Mutations in the cardiac ryano-dine receptor gene (hRyR2) underlie catecholaminergic polymorphic ventricular tachycardia. Circulation 2001, 103:196-200.[6] Fernández-Velasco M, Rueda A, Rizzi N, Benitah JP, Colombi B, Napolitano C, Priori SG, Richard S, Gómez AM. Increased Ca2+ sensitivity of the ryanodine receptor mutant Ry-R2R4496C underlies catecholaminergic polymorphic ven-tricular tachycardia. Circ Res 2009, 104:201-9.
GLOSARIO
Cardiomiopatía: término que refiere las enfermedades del músculo cardíaco, lo que incluye anormalidades en la estructura o la función del corazón. Existen tres tipos principales de cardio-miopatía: congestiva dilatada, hipertrófica y restrictiva.
Transgénico: organismo cuyo material genético ha sido modi-ficado para que deje de expresar un gen específico (knock-out), para que exprese una variante de algún gen o para que exprese un gen que normalmente no contiene (knock-in).

10
M. en C. Christian Lizette Frías Soria([email protected])
M. en C. Rodolfo Daniel Cervantes Villagrana([email protected])
Centro de investigación y de estudios avanzados del IPN, Departamento de Farmacobiología, Sede Sur.
Mejor tesina del
en Investigación Genómica
4° diplomado
El estudio de la regulación de la expresión de genes ad-quiere relevancia día a día gracias a los nuevos enfoques y herramientas de la biología molecular que han permiti-do conocer nuevas moléculas y mecanismos que partici-pan en la expresión génica. Tradicionalmente, el dogma central de la biología molecular postula que el DNA ge-nómico se expresa mediante su transcripción a mRNA el cual es traducido a una proteína. De esta manera, tanto el DNA como el RNA que no llevará a la traducción de una proteína (con excepción del RNA ribosomal o rRNA, y RNA de transferencia o tRNA), eran catalogados como material genético inservible o basura. En la actualidad, se considera que la totalidad del material genético es fun-cional, tal es el caso de algunos segmentos del DNA que son transcritos a un precursor del mRNA y sin ser necesa-riamente traducidos, son capaces de regular la expresión de otros genes e inclusive, de cumplir funciones determi-nantes en la vida celular. Estos segmentos transcritos del DNA que no codifican para una proteína son llamados RNAs no codificantes (non-coding RNAs, ncRNAs) y des-empeñan un papel importante en la modulación de la síntesis de proteínas [1].
LOS RNAs PEQUEÑOS NO CODIFICANTES COMOMODULADORES DE LA EXPRESIÓN GÉNICA
A diferencia de las moléculas largas de mRNA, los ncR-NAs tienen una longitud menor a 300 nucleótidos y se les ha acuñado los términos de RNA pequeños (small RNA), RNA no mensajero (non-messenger RNA, nmRNA), RNA no mensajero pequeño (small non-messenger RNA, snmRNA), RNA pequeño no codificante (tiny non-coding RNA, tncRNA), RNA modulador pequeño (small modu-latory RNA, smRNA) o RNA regulador pequeño (small regulatory RNA, srRNA) [1]. Las funciones de los RNAs pequeños van desde la formación de heterocromatina, hasta la desestabilización del mRNA y control traduccio-nal [2]. Los primeros detalles del silenciamiento de genes mediado por el RNA fueron observados a principios de los 90´s en varios estudios realizados en plantas y ani-males.
En 1993, Ambros y colaboradores descubrieron el RNA pequeño lin-4 que regula el desarrollo larval de Cae-norhabditis elegans y fue reconocido como el miembro fundador de una clase de pequeños RNAs llamados mi-croRNAs (miRNAs); posteriormente fue identificado en algas verdes, virus, animales y otros organismos com-plejos. Mientras tanto, otros tipos de RNAs pequeños

11
fueron descubiertos en animales, plantas y hongos, entre estos se encuentran los pequeños RNAs de interferencia endógenos (endogenous small interfering RNAs, endo-siRNAs), los RNAs asociados a Piwi (piRNAs) [3,4] y los RNAs pequeños nucleolares (small nucleolar RNAs, snoR-NAs), ver Tabla 1. Al igual que los miRNAs son capaces de regular la expresión de genes a través del reconocimiento de secuencias específicas en los mRNAs.
LOS MICRORNAs (miRNAs)
Los miRNAs son RNAs no codificantes que tienen una longitud aproximada de 21 nucleótidos y forman pares de bases con sitios específicos en la región 3’ no traduci-da (untranslated región, 3’-UTR) de mRNAs, impidiendo la traducción y guiando a la degradación del mRNA por un mecanismo similar al RNA de interferencia (interfering RNA, iRNA) [5], ver Figura 1. Se estima que por encima del 30% de los genes humanos están regulados por miR-
Figura 1. Biosíntesis y mecanismo de acción de los miRNAs. Los miRNAs son producidos por dos cortes consecutivos por endonucleasas en el núcleo y en el citoplasma. La biosíntesis de los miRNAs inicia con la transcripción de un miRNA primario (pri-miRNA) de varias kilobases de longitud con una estructura de tallo burbuja por la RNA-Polimerasa II (pol II). Los pri-miRNAs son poliadenilados y posteriormente fragmentados en el núcleo por la enzima Drosha dando lugar a un miRNA precursor (pre-mRNA). El pre-miRNA es transportado al citoplasma por la exportina 5 y Ran-GTP (un cofactor unido a GTP), donde es procesado por la enzima Dicer resultando un producto maduro de doble cadena con 19 24 nucleótidos de longitud. La maduración del miRNA culmina con la formación de un complejo de proteínas llamado complejo silenciador inducido por RNA (RISC) para llevar a cabo la represión de la traducción del mRNA, la cadena que es incorporada al RISC es el miRNA maduro, mientras que la cadena opuesta es liberada y degradada. Los miRNAs maduro se unen a la secuencia complementaria en el mRNA, impidiendo su traducción y favoreciendo el almacenamiento o degradación del mRNA en los cuerpos P. Adaptada de Lee et al., 2004; Chu y Rana, 2009; Kim y Siomi, 2009.
NAs [6] y los descubrimientos en los últimos años apoyan el papel de los miRNAs en la regulación de procesos vita-les como la proliferación celular [7], desarrollo [8], dife-renciación [9], metabolismo celular [10] y apoptosis [11]. Los niveles de miRNA precisan un riguroso control para mantener las funciones celulares y las alteraciones en la regulación de miRNAs usualmente han sido asociadas a patologías como el cáncer [2].
LOS RNAs ASOCIADOS A PIWI (piRNAs)
Las proteínas Piwi son elementos clave en las vías de silenciamiento del mRNA y se especializan en modular la unión de pequeños RNAs a los mRNAs. Las proteínas Piwi se asocian a pequeños RNAs endógenos de 24 a 30 nucleótidos, llamados RNAs asociados a Piwi (piRNAs) [12]. Los piRNAs fueron descubiertos al estudiar los RNAs pequeños en el desarrollo de Drosophila melanogaster, son procesados a partir de una cadena sen

12
Figura 2. La biogénesis de los piRNAs. La síntesis de los piRNAs se lleva a cabo a través de dos fases, la vía de procesamiento primario y el ciclo ping pong. La vía de procesamiento primario involucra la trans-cripción de un gen, generando una cadena antisenti-do para un mRNA blanco. Posteriormente en el ciclo ping pong se produce un proceso de amplificación, donde el remanente del mRNA silenciado, induce la maduración de más piRNAs. La proteína Piwi se une a la cadena antisentido y corta los transcritos blanco para generar el extremo 5´ en la cadena sentido, esta última se asocia con la proteína Ago3 la cual corta y metila el extremo 3´ de la misma cadena. El complejo de Ago/cadena sentido reconoce y corta la cadena antisentido precursora del piRNA para permitir la unión a la proteína Piwi, de esta manera se cierra el ciclo de amplificación. Adaptada de Chu y Rana, 2009; Ghildiyal y Zamore, 2009.
cilla de RNA precursora que es transcrita de elementos repetitivos intergénicos, transposones o grandes clusters de piRNAs [2]. En mamíferos se reveló que existen cien-tos de especies individuales de miRNA, mientras que las secuencias individuales de piRNA constan de cientos de miles y podrían ser millones. La principal función de los piRNAs es silenciar elementos transponibles, este papel está altamente conservado a través de las distintas espe-cies animales [13]. De esta manera los RNAs asociados a Piwi constituyen un sistema de defensa que protege a células germinales y somáticas contra la expresión de transposones.
Estudios bioquímicos y genéticos efectuados en moscas y ratones han llevado a proponer dos vías en la biogé-nesis de piRNAs: la vía de procesamiento primario y el ciclo ping pong (Figura 2). Inicialmente la vía primaria provee un pool de piRNAs dirigidos a múltiples transpo-sones, posteriormente en el ciclo ping pong se forman más piRNAs (proceso de amplificación) que se dirigen a transposones activos [13]. Adicionalmente, los piRNAs están involucrados en el silenciamiento transcripcional y las proteínas Piwi promueven modificaciones en histonas de la eucromatina, para incrementar su expresión, ade-más de mejorar la transcripción de piRNA en heterocro-matina de regiones subteloméricas de cromosomas [14]. Este hallazgo sugiere que los piRNAs pueden tener un control epigenético a través de múltiples mecanismos, ya sea silenciando o activando genes blanco a nivel trans-cripcional o post-transcripcional.
LOS PEQUEÑOS RNAs DE INTERFERENCIA ENDÓGE-NOS (ENDO-siRNAs)
La secuenciación de RNAs pequeños en el tejido somático de D. melanogaster, cultivos celulares y ovarios, permitió la identificación de endo siRNAs. Estos RNAs pequeños constan de aproximadamente 21 nucleótidos y se deri-van de transcritos de transposones, pares de transcritos sentido-antisentido o de largas estructuras tallo-burbuja [2]. Actualmente es claro que tanto plantas y animales producen una amplia gama de endo-siRNAs y juegan un papel importante en el silenciamiento de retrotransposo-nes y gametogénesis [15]. Los endo-siRNAs producidos por gusanos y plantas dependen de la acción de RNA polimerasas dependientes de RNA (RdRPs), las cuales uti-lizan el mRNA blanco como plantilla para la síntesis de novo de endo-siRNAs [16].
LOS RNAs PEQUEÑOS NUCLEOLARES (snoRNA)
La biosíntesis de ribosomas en células eucariotas es com-pleja, involucra numerosos eventos para generar rRNAs maduros y el subsecuente ensamblado de rRNAs con de-cenas de proteínas ribosomales. Los snoRNAs son funda-mentales en la maduración ribosomal, por ser requeridos en los procesos de corte para generar rRNAs individuales, y por su capacidad de guiar las modificaciones necesarias en sitios específicos de los rRNAs como son la metilación

13
Figura 3. Identificación de nuevos RNAs pequeños. Herramientas de las ciencias genó-micas utilizadas para la identificación, selección y caracterización de las funciones biológicas de los RNAs pequeños no codificantes. Adaptada de Soppa et al., 2009.
y la pseudouridilación [17,18,19]. Para ellos, los snoRNAs presentan motivos de secuencias consenso cortas que re-conocen los sitios donde se realizaran las modificaciones en un rRNA y un simple snoRNA puede dirigir una o varias modificaciones en los rRNAs [19]. Además, participan en la regulación de la expresión de proteínas y guían modifi-caciones en otros snoRNAs.
Los snoRNA se originan principalmente de intrones du-rante el proceso de splicing del pre-mRNA, la mayoría de los transcritos que alojan snoRNA intrónicos codifican para proteínas involucradas en la biosíntesis o función del ribosoma. El número de especies de snoRNAs es aproxi-madamente 200 en vertebrados [19], pero cabe destacar que existen snoRNAs que carecen de un blanco en rRNAs y snoRNAs, la mayoría de estos snoRNAs huérfanos tie-nen una expresión general y algunos tienen una expre-sión tejido-específica [20]. Se han identificado muchos snoRNAs huérfanos en todas las especies, lo que sugiere que es probable que tengan funciones adicionales a las mencionadas [21].
LOS RNAs PEQUEÑOS Y LA GENÓMICA
A través del análisis del transcriptoma y proteoma ha sido posible identificar la generación y la función biológica de los distintos RNAs pequeños no codificantes tanto en or-ganismos primitivos [22], como en el humano [23,24] (ver la Fig. 3). El entendimiento de la biogénesis de los RNAs
pequeños no codificantes ha permi-tido el desarrollo de diversas estra-tegias para activar vías de iRNA con fines terapéuticos. Se han diseñado RNAs recombinantes inhibitorios para imitar a los miRNAs primarios o miRNAs precursores, la expresión miRNAs artificiales se logra a través de plásmidos o empleando vectores virales. Esta tecnología ha sido em-
pleada en el tratamiento de infecciones respiratorias vi-rales en modelos animales y el iRNA se ha empleado para inhibir la infección producida por el virus del ébola en un modelo de primates [25].
Estudios recientes sugieren que los RNAs pequeños no codificantes, particularmente los miRNAs, contribuyen en la patogénesis y progresión de síndromes mielodisplá-sicos (MDS). Se identificaron especies de miRNAs que ac-túan como reguladores importantes del transcriptoma y se encuentran alterados en una gran cantidad de pacien-tes con MDS. Por otro lado, en pacientes con MDS de bajo grado está enriquecido con piRNAs, lo cual protege potencialmente al DNA de mutaciones, contrario a lo que se observa en pacientes con MDS de alto grado [26]. Debido al vínculo con algunas condiciones patológicas y en particular con el desarrollo y progresión del cáncer, los miRNAs y otros ncRNAs pueden convertirse en biomar-cadores útiles con fines diagnósticos. Diversos estudios han identificado a través de biopsias, asociaciones entre miRNAs y pronóstico de enfermedades, supervivencia y mortalidad en cáncer de colon, cáncer pancreático, glioblastoma [27], leucemia [28], cáncer de mama [29] y daño al miocardio. Además, los miRNAs plasmáticos y otros ncRNAs representan novedosos biomarcadores específicos para la detección temprana de diversas pa-tologías, debido al fácil acceso a la muestra, así como el uso de técnicas eficientes y cada vez más sensibles que las ciencias genómicas ofrecen [27].

14
CONCLUSIONES Y PERSPECTIVAS
Los ncRNA en animales juegan un papel crucial en el mantenimiento de la función celular. La habilidad de pre-decir el destino de los ncRNAs en base a su secuencia y a sus características estructurales es esencial para su uso efectivo como herramienta experimental y para su potencial uso terapéutico. Si bien, se ha logrado diluci-dar algunos aspectos de la biogénesis y función de los ncRNA, todavía quedan muchos detalles por aclararse. Las respuestas a estas interrogantes podrían ayudar a comprender las bases moleculares de varias patologías en las cuales la expresión de genes se encuentra alterada, por ejemplo el cáncer. El desarrollo terapéutico de siRNAs dirigidos a genes relacionados con diversas enfermeda-des ya está en proceso, así como estudios para inhibir miRNAs específicos ligados a patología. Sin embargo, es importante el desarrollo de nuevas herramientas bioin-formáticas que permitan distinguir entre RNAs pequeños no codificantes funcionales y no funcionales para conti-nuar con el desarrollo de herramientas terapéuticas.
GLOSARIO
Cuerpos P: son regiones citoplasmáticas con una eleva-da tasa de secuestro y degradación por deadenilación del mRNA. Algunos mRNA controlados por miRNAs pueden ser liberados de los cuerpos P en respuesta a condiciones de estrés, con la capacidad de reanudar su traducción.
Transposones: son “parásitos genómicos” que ame-nazan la integridad del genoma huésped. Estos pueden moverse hacia nuevos sitios por inserción o transposición y en consecuencia dividir genes y alterar el genoma.
Síndromes mielodisplásicos: un grupo de trastornos heterogéneos de células madre hematopoyéticas que frecuentemente genera leucemia mieloide aguda.
REFERENCIAS
[1] Ying SY, Miller JD, Lin SL: Non-coding RNAs–de-velopment of man-made vector-based intronic mi-croRNAs (miRNAs). In MicroRNAs, from basic science to disease biology. Edited by Krishnarao Appasani. USA:
Tabla 1. Tipos de RNAs pequeños no codificantes, origen y función. Adaptada de Bachellerie et al., 2002; Chu y Rana, 2009; Ghildiyal y Zamore, 2009; Kim y Siomi, 2009

15
Cambridge University Press, 2008: 22-41.[2] Kim VN, Han J, Siomi M: Biogenesis of small RNAs in animals. Nat Rev Mol Cell Biol 2009, 10: 126-139.[3] Bartel DP: MicroRNAs: genomics, biogenesis, me-chanism, and function. Cell 2004 116: 281-297.[4] Bartel DP: MicroRNAs: Target Recognition and Re-gulatory Functions. Cell 2009, 136: 215-233.[5] Royo H, Cavaillé J: Non-coding RNAs in imprinted gene clusters. Biol Cell 2008, 100: 149-166.[6] Rajewsky N, Socci ND: Computational identifica-tion of microRNA targets. Dev Biol 2004, 267: 529-535.[7] Cheng, AM, Byrom MW, Shelton J, Ford LP: Antisen-se inhibition of human miRNAs and indications for an involvement of miRNA in cell growth and apop-tosis: Nucleic Acid Res 2005, 33: 1290-1297.[8] Karp X, Ambros, V: Developmental biology. En-countering microRNAs in cell fate signaling. Science 2005, 310: 1288-1289.[9] Chen CZ, Li L, Lodish HF, Bartel DP: MicroRNAs mo-dulate hematopoietic lineage differentiation. Scien-ce 2004, 303: 83-85.[10] Poy MN, Eliasson L, Krutzfeldt J, Kuwajima S, Ma X, MacDonald P, Pfeffer S, Tuschl T, Rajewsky N, Rorsman P, Stoffel M: A pancreatic-islet specific miRNA regula-tes insulin secretion. Nature 2004, 432: 226-230.[11] Xu, P, Guo M, Hay BA: MicroRNAs and the regu-lation of cell death. Trends Genet 2004, 20: 617-624.[12] Chu CY, Rana TM: Structure and Gene-Silencing Mechanisms of Small. In Noncoding RNAs Non-Protein Coding RNAs. Edited by Walter NG, Woodson SA, Ba-tey RT. Germany: Springer Series in Biophysics 13, 2009: 335-356.[13] Siomi MC, Sato K, Pezic D, Aravin AA: PIWI-interac-ting small RNAs: the vanguard of genome defence. Nat Cell Biol 2011, 12: 246-258.[14] Yin H, Lin H: An epigenetic activation role of Piwi and a Piwi-associated piRNA in Drosophila melano-gaster. Nature 2007, 450: 304-308.[15] Ohnishi Y, Totoki Y, Toyoda A, Watanabe T, Yama-moto Y, Tokunaga K, Sakaki Y, Sasaki H, Hohjoh H: Small RNA class transition from siRNA/piRNA to miRNA during pre-implantation mouse development. Nu-cleic Acids Res 2010, 38: 5141–5151.[16] Ghildiyal M, Zamore PD: Small silencing RNAs: an expanding universe. Nature Rev Gen 2009, 10: 94-108.[17] Gardner PP, Bateman A, Poole AM: SnoPatrol: how many snoRNA genes are there? J Biol 2010, 9: 1-4[18] Taft RJ, Glazov EA, Lassmann T, Hayashizaki Y, Car-
ninci P, Mattick JS: Small RNAs derived from snoR-NAs. RNA 2009, 15: 1233-1240.[19] Bachellerie JP, Cavaillé J, Hüttenhofer A: The ex-panding snoRNA world. Biochimie 2002, 84: 775-790.[20] Rogelj B: Brain-Specific Small Nucleolar RNAs. J Mol Neurosci 2006, 28: 103-109.[21] Rodor J, Letelier I, Holuigue L, Echeverria M: Nu-cleolar RNPs: from genes to functional snoRNAs. Biochem Soc Trans 2010, 38: 672-676. [22] Straub J, Brenneis M, Jellen-Ritter A, Heyer R, Soppa J, Marchfelder A: Small RNAs in haloarchaea: iden-tification, differential expression and biological function. RNA Biol 2009, 6: 281-92. [23] Hansen KF, Sakamoto K, Obrietan K: MicroRNAs: a potential interface between the circadian clock and human health. Genome Med 2011, 3: 10.[24] Takizawa T, Gemma A, Ui-Tei K, Aizawa Y, Sadovsky Y, Robinson JM, Seike M, Miyake K: Basic and clinical studies on functional RNA molecules for advanced medical technologies. J Nippon Med Sch 2010, 77: 71-79.[25] Davidson BL, McCray PB: Current prospects for RNA interference-based therapies. Nat Rev Genet 2011, 12: 329-340.[26] Beck D, Ayers S, Wen J, Brandl MB, Pham TD, Webb P, Chang CC, Zhou X: Integrative analysis of next generation sequencing for small non-coding RNAs and transcriptional regulation in Myelodysplastic Syndromes. BMC Med Genomics 2011, 4:19.[27] Galasso M, Sana ME, Volinia S: Non-coding RNAs: a key to future personalized molecular therapy? Genome Med 2010, 2: 12.[28] Tanaka M, Oikawa K, Takanashi M, Kudo M, Oh-yashiki J, Ohyashiki K, Kuroda M: Down-regulation of miR-92 in human plasma is a novel marker for acu-te leukemia patients. PLoS ONE 2009, 4: e5532.[29] Iorio MV, Ferracin M, Liu CG, Veronese A, Spizzo R, Sabbioni S,Magri E, Pedriali M, FabbriM, Campiglio M, Menard S, Palazzo JP, Rosenberg A, Musiani P, Volinia S, Nenci I, Calin GA, Querzoli P, Negrini M, Croce CM: MicroRNAgene expression deregulation in human breast cancer. Cancer Res 2005, 65: 7065–7070.[30] Soppa J, Straub J, Brenneis M, Jellen-Ritter, Heyer R, Fischer S, Granzow M, Voss B, Hess WR, Tjaden B, Mar-chfelder A: Small RNAs of the halophilic archaeon Halofera volcanii. Biochem Soc Trans 2009, 37: 133-136.

16
López-Torres Donají. Laboratorio de Biología Celular y Citometría de Flujo.Universidad Autónoma Metropolitana-Iztapalapa. [email protected]
Palabras clave: linfocitos T, proteómica, patologías, señalización, activación.
Mejor tesina del 4° Díplomado en Investigación Genómica
La ProteómicaLa Proteómicacomo herramienta parael estudio de las células T.
Introducción
Nuestro sistema inmunológico nos protege contra agen-tes infecciosos (virus, bacterias, toxinas, etc.), sin embargo las alteraciones en su funcionamiento están involucradas en diversas patologías, como la autoinmunidad, inmu-nodeficiencia, hipersensibilidad y cáncer. Las técnicas y metodología actuales, permiten dilucidar cada vez mejor, el funcionamiento y regulación de las células del sistema inmunológico y detectar sus alteraciones en patologías, así como la generación de moléculas terapéuticas.
La investigación proteómica permite vislumbrar nuevas aplicaciones biomédicas y farmacéuticas [1]. En los últi-mos diez años la inmunología ha adoptado a la proteó-mica, originando a la inmunoproteómica [2]. La identi-ficación de las proteínas que intervienen en las diversas etapas, permite comprender las bases moleculares y la naturaleza de una enfermedad; y las proteínas identifica-das pueden utilizarse como biomarcadores de diagnósti-co o pronóstico de la enfermedad [1].
Las principales células linfoides que participan en la in-ducción, expresión y regulación de la respuesta inmuno-lógica adquirida, incluyen a los linfocitos T y B, y células presentadoras de antígenos (APCs). En general, hay dos tipos de subpoblaciones de linfocitos: los linfocitos T (cé-
lulas T) y los linfocitos B (células B) [3]. La función de las células B es producir anticuerpos y secretarlos, mientras que los linfocitos T cumplen funciones diversas, implica-das en interacciones con otras células.
Todas las células T tienen la molécula TCR (T cell recep-tor), que es el receptor para antígeno en estas células; la molécula CD3, que es un complejo tetramolecular asociado al TCR y relacionado con la transducción de se-ñales. Algunas células T expresan, el correceptor CD4 o CD8, y otras ninguna de las dos, aunque siguen siendo TCR+, CD3+. Las células TCD4+ y TCD8+ constituyen la mayoría de las células T en el organismo [3]. Los linfoci-tos TCD8+ destruyen células infectadas por virus o algún otro patógeno intracelular. Las células TCD4+ ayudan a otras células del sistema inmunológico a responder con-tra las infecciones de origen extracelular [4].
Proteoma, proteómica y análisis proteómico.
Se denomina proteoma al conjunto completo de proteí-nas que componen una célula en condiciones determina-das [5]. Para el estudio del proteoma se han desarrolla-do técnicas de análisis de proteínas a gran escala y alta velocidad [6]; la proteómica es la ciencia que estudia la

17
Figura 1. Diagrama de flujo del análisis proteómico (basado en Pando-Robles, 2009) [1]. Para llevarla a cabo, las proteínas se separan por isoelectroenfoque en gel de poliacrilamida. A continuación el gel se equilibra con un detergente y las proteínas se separan nuevamente en una segunda dirección, perpendicular a la primera, con base a su masa molecular. Esto permite separar en un único gel miles de proteínas, detectadas con reactivos de coloración, por fluorescencia o radiomarcaje y posteriormente, cuantificadas. El procedimiento de identificación más habitual de las proteínas es por digestión con proteasas y análisis de los fragmentos peptídicos resultantes por espectrometría de masas [14].
La proteómica en la activación en las Células T.
La proteómica ha permitido identificar un gran núme-ro de proteínas y determinar la secuencia de activación, posterior a la presentación del antígeno en células T; por ejemplo, se ha demostrado que la fosforilación de pro-teínas adaptadoras, como Shc y Grb2, está involucrada en el acoplamiento del TCR a la vía de señalización Ras. También se demostró la existencia de dos poblaciones de TCR en la superficie celular de los linfocitos T no ac-tivados: uno relacionado con el citoesqueleto y el otro no. Los niveles de fosforilación y formas ubiquitinadas, en cada una de las dos poblaciones de TCR, muestra un papel específico/separado en eventos mediados por re-ceptor [7]. Esta asociación entre el TCR y el citoesqueleto ha dado lugar a una oleada de interés sobre la actina del citoesqueleto asociada a la activación de Src, ZAP70 y
identidad, estructura, función bioquímica y celular de to-das las proteínas de un organismo, órgano o célula y sus variaciones según el tiempo, espacio y estado fisiológico [5]. La descripción del proteoma humano permite resol-ver cuestiones no resueltas por la información contenida en el genoma. Nuestro genoma contiene unos 25 000 genes, comunes en nuestras 10 trillones de células. Sin embargo, podemos clasificar más de 200 tipos celulares altamente especializados [6]. La técnica más utilizada para el análisis proteómico, es la electroforesis bidimen-sional en gel de poliacrilamida (2D-PAGE), ver Figura 1.

18
Figura 2. Activación de célula TCD4+ mediada por TCR (modificado de Moreno y cols., 2008). La susceptibilidad a la artritis reumatoide, involucra a proteínas que participan en la activación o inhibición de funciones celulares, especialmente en linfocitos T CD4+ que inducen inflamación sinovial crónica. La fosfatasa de tirosina PTPN22, regula negativamente la activación de linfocitos T. Las principales proteínas desfosforiladas por PTPN22 en linfocitos T son las cinasas de tirosina (PTK) Lck y ZAP70. Ambas PTK tienen dos sitios principales de fosforilación, uno que inhibe su función y otro que la activa. PTPN22 desfosforila las formas activas: Lck 505P y ZAP70 493P, su efecto es la inhibición de la función de las PTKs y, consecuentemente, de la activación de linfocitos T a través del TCR; debido a esto, los cambios estructurales en PTPN22 podrían relacionarse con susceptibilidad a artritis reumatoide, lupus y diabetes mellitus I [15].
Estudio proteómico de células T en patologías.
Otra ruta importante estudiada por análisis proteómico, es la regulación de las cascadas de transducción de seña-les a través de oxidación/reducción de aniones tiolato en los residuos de cisteína. Hay varios ejemplos que implican la oxidación de tioles aberrantes en las células T en artri-tis reumatoide. Muchos factores de transcripción (NFkB, STAT, AP-1, y el elemento de unión de respuesta al AMPc (CREB)) también están sujetos a la regulación redox/tiol. Por lo que la oxidación/reducción es importante en la ac-tivación de las células, la proliferación o muerte celular programada [8].
CD3β, en la exocitosis de gránulos de CTL, y en la contri-bución a los déficits en la señalización de células T duran-te el envejecimiento [7] [8].
La activación mediante el TCR produce cambios drásti-cos en el proteoma de una célula T: la entrada en ciclo celular, producción de citocinas, y diferenciación de una célula T efectora o de memoria [7], ver Figura 2. La pro-teómica es excepcionalmente adecuada para identificar modificaciones post-traduccionales utilizando marcado-res globales de modificación [9].

19
La proteómica ha permitido conocer alteraciones en las células T en enfermedades o infecciones. En células T hu-manas infectadas con el virus inmunodeficiencia humana 1 (VIH-1), se observó cambios en la abundancia relativa de 93 proteínas, principalmente aquellas involucradas en el metabolismo, señalización apoptótica, y proteínas de transporte intracelular [2]. Este análisis permite identificar proteínas implicadas en las interacciones virus-huésped, que pueden convertirse en el futuro blancos de fármacos.
Por otra parte, el asma y la alergia han surgido como pro-blemas importantes de salud pública. Ahora se sabe que la fase aguda de la inflamación pulmonar, se caracteriza por el incremento en la actividad proteolítica local; dado por metaloproteíncinasas de matriz activadas (MMPs), importante en la progresión de la disnea y asma. La ex-presión local de citocinas IL-4, IL-5, IL-13; y quimiocinas CCL7, CCL11, CCL17, dirigen la respuesta inmunológica a los alérgenos. Muchos miembros de la familia de MMP son sobre reguladas en pulmón de individuos asmáticos, coordinada por las citocinas [10].
Conclusiones y perspectivas.
Las bases modernas de la inmunología básica permiten resolver en gran medida el problema de la identificación de histocompatilidad entre donadores y receptores para el trasplante de órganos; modular el sistema inmunoló-gico en el tratamiento de enfermedades auto-inmunoló-gicas, crónicas infecciosas, neoplásicas; y recientemente otras enfermedades crónico-degenerativas. La FDA ha aprobado once anticuerpos monoclonales solos o con-jugados con toxinas o radionucleótidos, para uso tera-péutico. Sus usos van desde para prevenir el rechazo de órganos trasplantados, como el daclizumab, dirigido hacia la subunidad del receptor de IL-2Rα de linfocitos activados; en enfermedades como la artritis reumatoide y la enfermedad de Crhon; en pacientes con recaída de linfoma no Hodkin-CD20, en donde el infliximab inhibe la unión del TNF-α a su receptor, o el rituximab dirigido contra CD20 [11].
La sangre representa, probablemente la fuente más acce-sible para la búsqueda de biomarcadores de diagnóstico y pronóstico. El número de proteínas en la sangre huma-na es superior a las 106 moléculas diferentes, que repre-sentaría el producto de 25 000 a 30 000 genes. El poten-cial de la sangre en contener biomarcadores de utilidad clínica es grande [12]. Los linfocitos pueden representar sensores moleculares para la evaluación de la modifica-
ción de la expresión génica en condiciones fisiológicas y patológicas. Proporcionan un modelo único para los estudios integrados de la expresión génica en humanos, teniendo en cuenta que: (i) son extremadamente sensi-bles a modificaciones del medio, (ii) la variación de estos estímulos determina el patrón de las modificaciones de proteínas en linfocitos, y (iii) están fácilmente disponibles en gran número en la sangre periférica, sin necesidad de una biopsia invasiva y/o de riesgo [13].
La proteómica provee de demasiada información, a la espera de ser explotada; por lo que se ha propues-to la aceptación de la inmunoinformática, la rama de la bioinformática asociada con la inmunología, para sacar el máximo provecho de la información que se genera y traducir los datos en conocimiento biológico [8].
GLOSARIO
Antígeno: Es una sustancia propia o extraña, que es re-conocida por las células del sistema inmunológico como dañina y estimula la formación de anticuerpos contra ella.
Biomarcadores: Son elementos biológicos analizados a nivel molecular y/o celular, con base a los cuales se de-sarrollan medidas sensibles para ser identificados como indicadores del estado o progresión de una patología o tratamiento.
Citocinas: Son péptidos reguladores, de gran importan-cia en el sistema inmunológico. Permiten la comunica-ción intercelular, activación de receptores de membrana, secreción de anticuerpos, proliferación, diferenciación y migración celular.
Factor de transcripción: Es un elemento que partici-pa en la regulación de la transcripción del ADN. Pueden reconocer y unirse a secuencias específicas de ADN o uniéndose a otros factores.
FDA: Agencia de Alimentos y Medicamentos de Nor-teamérica, responsable de la regulación de alimentos, suplementos alimenticios, medicamentos, cosméticos, aparatos médicos, productos biológicos y derivados san-guíneos.
Genoma: Es la totalidad de la información genética que posee un organismo en particular.

20
Radionucleótidos: Son elementos químicos con confi-guración inestable que experimentan una desintegración radiactiva que se manifiesta en la emisión de radiación en forma de partículas alfa o beta y rayos X o gama.
REFERENCIAS
[1] Pando-Robles RV, Lanz-Mendoza H: La importan-cia de la proteómica en la salud pública. Sal Púb de Méx 2009, 51 (Suppl 3): 386-394.[2] Ringrose JH, Jeeninga RE, Berkhout B, Speijer D: Proteomic Studies Reveal Coordinated Changes in T-Cell Expression Patterns upon Infection with Hu-man Immunodeficiency Virus Type 1. J of Virol 2008, 82: 4320-4330.[3] Rojas-Espinosa O: Inmunidad adquirida: el siste-ma linfoide. In Inmunología (de memoria). 3th edition. Madrid, España. Médica Panamericana; 2006: 72-82.[4] Parham P: Inmunidad mediada por células T. In Inmunología/Immunology. 2nd edition. Madrid, España. Médica Panamericana; 2006: 157-160.[5] López-Muñoz F, Álamo C: Contribución de las nuevas técnicas genómicas y proteómicas al estu-dio de la patogenia de las emfermedades mentales y al desarrollo de la psicofarmacología. In Historia de la psicofarmacología. Volume 3. Madrid, España. Médica Panamericana; 2007:1720-1726.[6] Prieto-Valtueña JM, Yuste-Ara JR: Inmunidad me-diada por células. Serología y diagnóstico inmuno-biológico. In La clínica y el laboratorio. 21st edition. Bar-celona, España. Masson-Elsevier; 2010: 425-427.[7] Caplan S, Baniyash M: Normal T cells express two T cell antigen receptor populations, one of which is linked to the cytoskeleton via zeta chain and dis-plays a unique activation-dependent phosphoryla-tion pattern. J Biol Chem 1996, 271: 705–712.[8] Grant MM, Scheel-Toellner D, Griffiths HR: Contri-butions to our understanding of T cell physiology through unveiling the T cell proteome. Clin & Exp Immunol 2007, 149:9-15.[9] Zheng H, Hu P, Quinn D, Wang Y: Phosphotyro-sine proteomic study of interferon alpha signaling pathway using a combination of immunoprecipita-tion and immobilized metal affinity chromatogra-phy. Mol Cell Proteomics 2005, 4: 721-730.[10] Kheradmend F, Corry D: Discovery of novel mar-kers in allergic lung infalmmation through proteo-mic-based technologies. Expert Rev Proteomics 2008, 5: 9-12.[11] Valdespino-Gómez VM: Elementos necesarios
para el ejercicio moderno de las ciencias médicas. Cir y cirujanos 2003, 71: 80.[12] López-Farré A, Macaya-Miguel C: Aportaciones de la investigación en el área cardiovascular en Espa-ña. In Libro de la salud cardiovascular del Hospital Clínico San Carlos y la Fundación BBVA. Madrid, España. Funda-ción BBVA; 2007: 676-678.[13] Borro M, Gentile G, Stigliano A, Misiti S, Toscano V, Simmaco M: Proteomic analysis of peripheral T lym-phocytes, suitable circulating biosensors of strictly related diseases. Clin and Exp Immunol 2007, 150: 494–501.[14] Pallás V, Escobar C, Rodríguez-Palenzuela P, Marcos JF: Proteómica: Conceptos y metologías. In Herra-mientas biotecnológicas en fitopatología. Madrid, España: Mundi-Prensa Libros, s. a; 2008: 75-88.[15] Moreno J, Vázquez-Ortiz G, López-Blanco JA, Ló-pez-Romero R, Medina F: Hacia un tratamiento no empírico de la artritis reumatoide basado en su pa-togenia molecular. Reumatol Clín 2008, 4: 19-31.

21
PROYECTOS DEL PCGen desarrollo
en esta sección se resumen aLgunos de Los trabajos de tesis experimentaL que desarroLLan actuaLmente Los es-tudiantes deL pcg.
Identificación de la proteína eIF-5a dependientede poliaminas en Trichomonas vaginalis
M. en C. Bertha Isabel Carvajal GámezEstudiante de Doctorado en Ciencias Genómicas
Trichomonas vaginalis es un parásito protozoario cla-sificado taxonómicamente en el grupo de los excava-tas [1] (Fig. 1). Este microorganismo oportunista es el agente patógeno responsable de la tricomonosis, infección de transmisión sexual (ITS), que ocupa el pri-
mer lugar a nivel mundial en lo que a infecciones de transmisión sexual se refiere. La Organización Mundial de la Salud (OMS) reporta una incidencia de tricomo-nosis de más de 250 millones de casos al año (reporte de la OMS año 2010). En la República Mexicana, hasta la semana 14 del presente año, se han alcanzado al-rededor de 29, 011 casos. Esta infección se encuentra distribuida principalmente en mujeres de la zona cen-tro y sur del país. En los estados de Veracruz, Estado de México, Puebla y el Distrito Federal existe una alta incidencia de la tricomonosis, a diferencia de la zona norte del país donde existe una baja incidencia de la infección [2]. La tricomonosis afecta el tracto urogeni-tal de los humanos [3] y ha sido asociada a la gonorrea y vaginitis bacteriana [4, 5]. Esta enfermedad ocasiona graves daños en la salud tanto de mujeres como de hombres, pues constituye un factor de riesgo para la transmisión del VIH (virus de inmunodeficiencia adqui-rida) [6, 7, 8] así como también se le ha relacionado con la predisposición a adquirir cáncer cervical [9] a de-sarrollar de enfermedad pélvica crónica [10], infertili-dad [11] y susceptibilidad a infecciones concomitantes causadas por otros agentes patógenos como Neisseria gonorrea y Clamidia trachomatis [12].
La infección por T. vaginalis puede manifestarse como aguda o crónica, según los signos y síntomas. En la forma crónica se puede presentar dolor durante las re-laciones sexuales y escozor en los genitales externos. En la fase aguda suele aparecer, además de los
Figura 1. Microscopía electrónica de barrido de Trichomonas va-ginalis, mostrando la forma ovoide y ameboide de este parásito en contacto con una célula de la línea celular DU-147 de cáncer de próstata.
Palabras claves: Trichomonas vaginalis, tricomono-sis, poliaminas, factor de inicio de la traducción 5A, deoxihipusina sintasa, deoxihipusina hidroxilasa.

22
síntomas anteriores, inflamación en la vulva, y ardor en la cavidad baja y durante la micción. Un signo clí-nico propio de esta infección es el denominado ‘cérvix de fresa’, observado sólo en el 2% de las pacientes [4]. Uno de las características más delimitadas de esta infección es la gran cantidad de descargas vaginales con olor fétido, debido a la presencia de putrescina y cadaverina, las cuales pertenecen al grupo de las po-liaminas.
El descubrimiento de las poliaminas se remonta al año 1678, cuando Antonie van Leeuwenhoek aisló algunos cristales provenientes de semen humano. Sin embargo, no fue hasta 1924 que la fórmula empíri-ca de los cristales fue descrita. En los siguientes años, posteriores a la síntesis del producto, los nombres que se dieron a dos de las poliaminas fueron basados en el descubrimiento original “espermidina y espermina”. Por otra parte, la putrescina (1,4-diaminobutano) fue aislada de la bacteria Vibrio cholerae y su nombre de-riva de una gran cantidad de carne en putrefacción y la cadaverina fue nombrada así porque presenta un olor similar a los organismos en proceso de descom-posición. Hasta hoy las poliaminas están consideradas como reguladores críticos del crecimiento celular, di-ferenciación y muerte celular [13]. Las poliaminas son cationes pequeños que se caracterizan por tener una carga neta positiva que se distribuye a lo largo de su estructura [13] (Fig. 2). Su mecanismo de acción es capaz de regular dos propiedades de virulencia en T. vaginalis: la adhesión y la citotoxicidad, específica-mente a través de la modulación de la expresión de al menos una cisteín proteinasa de 65 kDa (CP65) [14, 15, 16]. Este parásito carece de la vía metabólicas de síntesis de poliaminas, por lo cual utiliza un sistema denominado antiporte acoplado a la secreción de dos moléculas de putrescina por cada molécula de esper-mina internalizada [17, 18]. Las poliaminas son com-
puestos que influyen en la transcripción, la traducción y que distintivamente participan en una modificación post-traduccional específica y única denominada hi-pusinación, que es llevada a cabo sobre la proteína implicada en el inicio de la traducción denominada eIF-5A [19, 20]. Estudios recientes sugieren que eIF-5A es capaz de unirse al extremo 3´UTR de los RNA mensajeros (RNAm), específicamente a la secuencia denominada elemento de respuesta a eIF-5A (ERE); lo cual podría indicar su participación en el mecanismo de regulación de la expresión genética a nivel post-transcripcional [21]. El factor eIF-5A de humano es una proteína de 140 aminoácidos la cual se encuentra muy conservada evolutivamente y por tanto está pre-sente en arqueabacterias hasta en mamíferos, pero no en eubacterias [20, 22]. eIF-5A es conocido como un factor dependiente de las poliaminas el cual se carac-teriza por tener 12 aminoácidos conservados (Ser-Thr-Ser-Lys-Thr-Gly-Hpu-His-Gly-His-Ala-Lys) rodeando un residuo de lisina. Este arreglo de aminoácidos no se ha identificado en otras proteínas lo cual sugiere su ori-gen temprano en la evolución de los eucariontes [20, 22]. eIF-5A es activado mediante la hipusinación, una modificación post-traduccional que ocurre específica-mente en el residuo de lisina [20, 22]. El aminoácido añadido conocido como hipusina (Ne-[4-amino-2-hi-droxibutil]lisina) fue denominado así en base a su es-tructura que consiste de dos hidroxiputrescinas y una lisina [20, 22]. Esta modificación post-traduccional se lleva a cabo en dos etapas: en la primera etapa la en-zima deoxihipusinasintasa (DHS) transfiere un carbono aminobutil de la espermidina al residuo específico de lisina de la proteína eIF-5A precursora. Esta reacción es dependiente de NAD+ y da paso a la siguiente reac-ción enzimática catalizada por la homoespermidina sintasa (DOOH)que hidroxila el carbono aminobutil para generar la hipusina. De este modo, eIF5A precur-
Figura 2. Estructura y for-mula química de las polia-minas más abundantes en las secreciones vaginales y prostáticas.

23
sora es transformada a la forma activa o proteína ma-dura (Fig. 3) [20, 21, 22]. Recientemente, se identificó y clono los genes que codifican para el factor eIF-5A de T. vaginalis. Este parásito contiene dos genes tveif-5a1 y tveif-5a2 que comparten un 98% de identidad; ambos genes se encontraron en dos locus diferentes dentro del genoma del parásito, poseen un tamaño de 504 pb y su contenido de G+C es del 49.2%. Así mismo tveif-5a1 y tveif-5a2 contienen secuencias de regulación tipo Inr en su región promotora en las po-siciones -23 y -28 río arriba del ATG respectivamente, los cuales podrían modular su expresión a nivel trans-cripcional. Las proteínas TveIF-5A1 y TveIF-5A2 codifi-cadas por estos dos genes comparten una identidad del 99%, presentando únicamente dos cambios de aminoácidos. Ambas secuencias presentan el domi-nio de 12 aminoácidos incluyendo el residuo de lisina
necesarios para la hipusinación [23]. TveIF-5A tiene dos dominios, el N-terminal que se caracteriza por ser básico e hidrofílico y se encuentra situado en un loop estructural dentro del cual existe un residuo de lisina expuesto y, el C-terminal que es ácido y contie-ne una alfa hélice que le permite unirse al RNAm y a otras proteínas (Fig. 4). Esta estructura se encuentra altamente conservada en los diferentes organismos analizados ya que mediante predicción de estructuras en 3D utilizando las estructuras terciarias obtenidas de cristales de proteínas relacionadas provenientes de Homo sapiens, Leishmania y Methanococcus, se ha demostrado que poseen una alta homología no
Figura 3. Modificación post-traduccional de eIF-5A por hipusi-nación, para la generación de eIF-5A madura o activa. La hipusi-nación es llevada a cabo mediante dos pasos enzimáticos por las enzimas deoxihipusina sintasa (DHS) y deoxihipusina hidroxilasa (DOOH).
Figura 4. Modelo de la estructura tridimensional de eIF-5A de T. vaginalis. En morado se muestran las alfa hélices y en azul las hojas beta plegadas. El loop expuesto en la región conservada donde se lleva a cabo la hipusinación, muestra el arreglo de 12 aminoácidos incluyendo al residuo de lisina marcado con un asterisco.
solo en secuencia sino también en su plegamiento es-tructural en 3 dimensiones. TveIF-5A se localiza en el citoplasma de los trofozoítos de T. vaginalis [20, 22, 23]. Interesantemente, en parásitos crecidos en au-sencia de poliaminas, la expresión del gen tveif-5A se encontró disminuida en comparación con los niveles de expresión observados en las células crecidas en el medio normal, condición considerada como control. Además, los niveles de transcrito de tveif-5a fueron recuperados con la adición de putrescina exógena, demostrándose que las poliaminas regulan la expre-sión del gen tveif-5a. Aunado a esto, en ausencia de putrescina la cantidad de TveIF-5A madura disminu-ye en comparación con el control y es recuperada al adicionar putrescina exógena lo que sugiere que putrescina es necesaria para la expresión y modifi-cación post-traduccional de TveIF-5A. En base a los antecedentes, conocemos que las poliaminas influyen en la citotoxicidad de T. vaginalis y que estos catio-nes podrían estar regulando diferentes funciones ce-lulares, incluyendo la modificación post-traduccional de la proteína TveIF-5A. Nuestros hallazgos nos han permitido proponer la existencia de un mecanismo de regulación de la citotoxicidad en T. vaginalis mediado por poliaminas y TveIF-5A [24]. Este proceso podría ser similar al descrito a nivel post-transcripcional para el gen cox-2, el cual también es regulado por la pro-teína dependiente de poliaminas eIF-5A. El grupo de investigación de la Doctora Elizbeth Álvarez Sánchez continúa llevando a cabo estudios genómicos y pro-teómicos que coadyuvarán en la obtención de nuevos datos de interés para la comprensión de la biología molecular de este agente patógeno.

24
GLOSARIO
Tricomonosis: Infección de transmisión sexual que afecta el aparato urogenital del hombre y de la mujer ocasionada por el parásito protozoario Trichomonas vaginalis.Putrescina: también conocida como butano-1,4-diamina, es una diamina que se crea al podrirse la carne dándole su olor característico.
Cadaverina: conocida también como 1,5-diaminopentano, es una diamina que se obtiene por la descomposición del aminoácido lisina. Su nombre es debido a su olor característi-co de materia orgánica muerta.
Diamina: es una sustancia orgánica en cuya molécula hay dos grupos –NH2 unidos a uno o dos carbonos de radicales de hidrocarburos.
Poliaminas: son moléculas de naturaleza policatiónica pre-sentes tanto en plantas, animales y microorganismos que contienen por lo menos dos grupos –NH2 unidos a uno o dos carbonos.Espermina y espermidina: son parte del metabolismo de las poliaminas envuelta en el crecimiento celular y presente en gran cantidad en el semen del hombre.
Transcripción: es el proceso de la expresión génica, me-diante la cual se transfiere la información contenida en la secuencia de DNA hacia la secuencia de prorteínas utilizando el RNAm como intermediario.
Hipusinación: modificación post-traduccional que se lleva a cabo específicamente en la proteína eIF-5A precursora.
REFERENCIAS
[1] Parfrey LW, Barbero E, Lasser E, Dunthorn M, Bhatta-charya D, Patterson DJ, Katz LA: Evaluating Support for the Current Classification of Eukaryotic Diversity. Plos Genetics 2006, 2: 2062-2073.[2] Instituto de Diagnóstico y Referencia Epidemiológicos[InDRE].[3] Alderete JF, Crouch ML y Arroyo R: Mecanismos de patogénesis de la trichomonosis: La enfermedad de transmisión sexual no viral número uno causada por Trichomonas vaginalis. La frontera: genética molecular de la enfermedad. Capitulo 6. Tomo I. Editado por Luna-Arias J.P Y Orozco-Orozco E. Editorial IPN México; 2004: 167-200.[4] Schwebke JR, Burgués D: Trichomoniasis. Clin Microbiol 2004, 17:794-803.[5] Swygard H, Sena A, Hobbs M, Cohen M: Trichomonia-sis: clinical manifestations, diagnosis and management. Sex Transm Infect 2004, 81: 91–95. [6] Sorvillo F, Smith L, Kerndt P, Ash L: Trichomonas vagi-nalis, HIV, and African Americans. Emerg Infect Dis 2001,
7:927–932.[7] Kapiga SH, Lyamuya EF, Lwihula GK, Hunter DJ: The in-cidence of HIV infection among women using family planning methods in Dar es Salaam, Tanzania. AIDS 1998,12:75–84.[8] Laga M, Manoka A, Kivuvu M: Non-ulcerative sexually transmitted diseases as risk factors for HIV-1 transmis-sion in women: results from a cohort study. AIDS 1993, 7:95–102.[9] Viikki M, Pukkala E, Nieminen P, Hakama M: Gynaeco-logical infections as risk determinants of subsequent cervical neoplasia. Acta Oncol 2000, 39:71–75.[10] Schwebke JR, Burgess D: Trichomoniasis. Clin Micro-biol Rev 2004, 17:794–803.[11] Cotch MF, Pastorek IIJG, Nugent RP, Hillier SL, Gibbs RS, Martin DH: Trichomonas vaginalis associated with low-birth weight and preterm delivery. The vaginal infec-tions and Prematurity Study Group. SexTransm Dis 1997, 24:353–360.[12] Anorlu RI, Fagbenro-Beyioku AF, Fagorala T: Prevalence of Trichomonas vaginalis in patients with vaginal dis-charge in Lagos Nigeria. Nigeria Post Med 2001, 8:183-186.[13] Tabor CW y Tabor H: Polyamines in Microorganisms. Microbiol Rev 1985, 49:81-99.[14] Hernández-Gutiérrez R, Avila-González L, Ortega-López J, Cruz-Talonia F, Gómez-Gutierrez G, Arroyo R: Trichomo-nas vaginalis: characterization of a 39-kDa cysteine proteinase found in patient vaginal secretions. Exp Pa-rasitol 2004, 4:125-135. [15] Alvarez-Sánchez ME, Solano-González E, Yañez-Gómez C, Arroyo R: Negative iron regulation of the CP65 cys-teine proteinase cytotoxicity in Trichomonas vaginalis. Microbes Infect 2007, 9:1597-605. [16] Alvarez-Sanchez ME, Avila-Gonzalez L, Becerril García C, Fattel Facenda LV, Ortega-Lopez J, Arroyo R: A novel cysteineproteinase [CP65] of Trichomonas vaginalis in-volved in citotoxicity. Microb Pathogenesis 2000, 28:193-202.[17] Yarlet N, Martínez MP, Golberg DL, Kramer DL, Porter CW: Dependence of Trichomonas vaginalis upon pol-yamine backconversion. Microbiol 2000, 146: 2715-2722.[18] Yarlet N, Bacchi CJ: Parasite polyamine metabolism: tar-gets for chemotherapy. Biochem Soc Trans 1994, 22: 875-879.[19] Liu L, Guo X, Rao JN, Bernard TS, Jie J, Casero RA, Jian-Ying W: Polyamine-modulated c-Myc expression in nor-mal intestinal epithelial cellsregulates p21Cip1 trans-cription through a proximal promoter region. Biochem J 2006, 398: 257-267.[20] Park MH: The Post-Translational Synthesis of a Po-lyamine-Derived Amino Acid, Hypusine, in the Eukar-yotic Translation Initiation Factor 5A [eIF5A]. J. Biochem 2006, 139:161–169.

25
NOTICIASdel mundo de la ciencia
Los tumores suprarrenaLes conducen a Los investi-gadores a La hipertensión reLacionada a mutacio-nes genéticas
Más de mil millones de humanos en todo el mundo sufren hipertensión severa, lo cual los hace propen-sos a un mayor riesgo de sufrir ataques cardíacos o derrame cerebral. Los científicos saben que algunas personas están genéticamente predispuestas a mani-festar una presión arterial alta, pero tanto la comple-jidad de la condición como la del genoma humano han hecho la identificación de los factores de riesgo extremadamente difícil.
Investigadores del Instituto Médico Howard Hughes (HHMI) han utilizado la próxima generación de her-ramientas de secuenciación del ADN para identificar una mutación en un gen que subyace a una de las formas más comunes de hipertensión severa.
Richard P. Lifton del HHMI y sus colegas informaron a la revista Science que las mutaciones en el gen que codifica para el canal de potasio KCNJ5 impulsan el desarrollo de tumores productores de aldosterona en la glándula suprarrenal. La hormona aldosterona normalmente sólo se produce por la glándula suprar-renal en respuesta a la deshidratación o a los altos niveles de potasio en la sangre. Los niveles excesivos de aldosterona producen hipertensión.
“Estos tumores son clínicamente importantes, pues se encuentran entre el cinco y el diez por ciento de los pacientes con hipertensión grave. También pensamos que podrían ser biológicamente interesantes porque, a diferencia de muchos otros tipos de cáncer, rara vez se convierten en invasivos o dan lugar a metástasis distantes”, dijo Lifton, también profesor y catedrático en la Escuela de Medicina de la Universidad de Yale.
Lifton, quien trabajó con un equipo de investiga-dores de las Universidades Yale y Uppsala, del Cole-gio Médico de New York y del Hospital Henry Ford en Detroit, señaló que su análisis de los tumores fue un problema particularmente conveniente gracias a las nuevas tecnologías que permiten secuenciar efici-entemente todos los genes del genoma.
“debido aL aLto impacto de estos tumores en La pobLación hipertensa, estos resuLtados aL-canzan una connotación cLínica sustanciaL.”- richard p. Lifton
Mediante una estrategia de secuenciación del ADN llamada secuenciación completa del exoma, que se desarrolló en el laboratorio por el primer autor del artículo, Murim Choi, se secuenciaron 23.000 genes humanos presentes en los tumores de cuatro pacien-tes. Los investigadores compararon las secuencias de cada paciente con las del ADN de sus propias célu-las sanguíneas, con el objeto de buscar mutaciones que se hayan producido en los tumores de manera somática, es decir, después del desarrollo normal del individuo. Ellos encontraron muy pocas mutaciones somáticas que pudieran alterar proteínas, sólo alre-dedor del dos por tumor. Sorprendentemente, un gen que codifica para un canal de potasio llamado KCNJ5 se halló doblemente mutado. Cuando estu-diaron otros adenomas productores de aldosterona encontraron que cualquiera de las dos mutaciones en KCNJ5 estuvo presente en ocho de los 22 tumores es-tudiados. “Resulta excepcionalmente poco probable

26
que estos hallazgos se deban a un hecho fortuito y es-tablecen que estas mutaciones son la causa de estos tumores”, expresó Lifton…
Los canales de potasio son poros celulares que nor-malmente sólo le permiten a los iones potasio difundir dentro y fuera de la célula. El análisis estructural su-giere que las mutaciones del gen KCNJ5, observadas en los adenomas, introducen cambios en la proteína que podrían disminuir su preferencia por los iones de potasio. El análisis electrofisiológico mostró que cuando el canal mutante se expresaba en células en cultivo, permitió el paso iones de sodio a través de la membrana.
“En la década de los 90’s, en el laboratorio de Rod MacKinnon se demostró que la base para la especi-ficidad del canal es un filtro de selectividad definido por aminoácidos específicos que permiten a los iones potasio pasar a través del canal y no así a los de sodio, apuntó Lifton, refiriéndose a su compañero investiga-dor del HHMI Rod MacKinnon, de la Universidad de Rockefeller, quien recibiera el Premio Nobel de Quími-ca en 2003 por trabajar en la estructura y función del canal. “Una de las dos mutaciones encontradas enlos tumores estudiados altera precisamente uno de los aminoácidos que MacKinnon había definido”.
Posteriormente, el equipo demostró que la entrada de sodio causada por KCNJ5 mutado en las células tumorales, pone en marcha una cadena de eventos que resulta en un aumento de los niveles de calcio intracelular, lo cual induce tanto la producción de al-dosterona como la proliferación celular. “A nuestro entender, éste es el primer informe de una mutación en los canales iónicos con un papel en la neoplasia”, dijo Lifton. Si bien las nuevas mutaciones identifica-das se producen somáticamente, se planteó la inter-rogante de qué ocurriría si una mutación así estuviera presente en cada célula del cuerpo. Esto llevó a Lifton a revisar un estudio realizado en su laboratorio, que fue publicado en 2008. En éste, su equipo describió la condición hereditaria de una familia, con una enfer-medad en la que el padre y sus dos hijas tenían hiper-tensión severa desde la infancia, debido a la secreción no controlada de aldosterona y al crecimiento masivo de todas las células de las glándulas suprarrenales que normalmente producen aldosterona. En aquella oca-sión, los investigadores no pudieron identificar al gen responsable de la condición.
Ahora, armados con un gen candidato, ellos re-ex-aminaron las muestras de ADN de esa familia y en-contraron que el padre tenía una mutación similar en KCNJ5 que había transmitido a sus dos hijas afecta-das. Este descubrimiento, también reportado en la revista Science, vincula las mutaciones que alteran las regiones críticas de la proteína KCNJ5, ya sean con-génitas o adquiridas somáticamente, con la excesiva producción de aldosterona en tumores suprarrenales que desencadenan una presión arterial alta.
El análisis realizado por este equipo no habría sido posible sin la poderosa tecnología de secuenciación de ADN actual. En lugar de aplicar este proceso a to-das las muestras de ADN tumorales en su estudio, el grupo de Lifton redujo su búsqueda, utilizando un método llamado secuenciación completa del exoma. Conceptualmente, el método es sencillo. Sólo el uno por ciento del ADN de una célula contiene los genes que codifican proteínas, pero estos genes (“exoma”) son sitios que contienen alrededor del 85 por ciento de las mutaciones que causan enfermedades en el humano. La Metodología de secuenciación completa del exoma permite a los investigadores hacer caso omiso a secuencias extrañas y centrarse en las poco más de 30 millones de pares de bases que confor-man ese uno por ciento. “La secuenciación completa del exoma está permitiendo a los genetistas estudiar a la vez enfermedades complejas y poco comunes”, afirmó Choi, refiriéndose a condiciones como la hip-ertensión o la diabetes que emergen de una constel-ación de factores.
Choi aprendió las técnicas computacionales necesar-ias para llevar a cabo el estudio. “Cuando comencé mi proyecto en 2007, necesitaba desarrollar habili-dades para analizar grandes conjuntos de datos, así que compré un libro para aprender PERL”, dijo, re-firiéndose a un lenguaje de programación utilizado en el análisis bioinformático. “Hice un programa de trabajo durante el día y leía el libro por la noche. Lo terminé en dos semanas”. Claramente, esas fueron dos semanas bien invertidas. En 2009, Choi fue el primer autor del estudio “Exoma completo” realiza-do en el laboratorio de Lifton que fue publicado en Proceedings of the National Academy of Sciences. Ya para el documento actual en Sciences, Choi analizó todos los datos genómicos.
“Debido al alto impacto de estos tumores en la po-

27
blación hipertensa, estos resultados alcanzan una connotación clínica sustancial”, comentó Lifton. Él dice que los hallazgos de su equipo revelan la biología sorprendentemente sencilla de estos tumores suprar-renales, y aumenta la posibilidad de desarrollar una prueba de detección para identificar a los pacientes con estas neoplasias, mediante la búsqueda de una de estas dos mutaciones en células o ADN de los tu-mores que circulen en la sangre. “Una simple prueba de detección de estos tumores tendría un impacto transcendental en el tratamiento de esta enfermedad, ya que esta forma de hipertensión se puede curar me-diante la eliminación del tumor “, señaló...
Fuente de la informaciónNoticias del Instituto Howard-Hughes.
Traducido y reproducido con autorización del HHMIhttp://www.hhmi.org/news/lifton20110211.html
NUEVOS PROYECTOS DEL PCG-UACMCON FINANCIAMIENTO.
Proyecto aprobado:
Diseño de nanopartículas superparamagnéticas dirigidas contra células tumorales de cáncer de mama Her2+.
Área: Medicina y Ciencias de la salud. Modalidad: Joven investigador.
Responsable técnico: Dr. Jesús Fandiño ArmasPCG - UACM
El pasado 28 de julio de 2011 se publicaron los resultados correspondientes a los proyectos
de investigación aprobados en apego a las bases establecidas en la Convocatoria de Investi-
gación Científica Básica 2010 del Conacyt. Esta convocatoria se emite con el objeto de apo-
yar propuestas de investigación científica básica que generen conocimiento de frontera y
contribuyan a mejorar la calidad de la educación superior y a la formación de científicos y
académicos.

28
GRADUADOS
desde eL año 2003 a La fecha, se han graduado en eL pcg-uacm
40 estudiantes de maestría y 6 de doctorado en ciencias genómicas.
MAESTRÍA EN CIENCIAS
GENERACIÓN 2009
DOCTORADO EN CIENCIAS
GENERACIÓN 2003
Dr. en C. Álvaro Díaz Badillo.Tesis: Genética de poblaciones del complejo Culex pipiens (diptera: culicidae) en la Ciudad de México. Fecha de titulación: Agosto, 2011.Directoras de Tesis: Dra. Minerva Camacho Nuez y Dra. María de Lourdes Muñoz Moreno.
M. en C. José Alí Flores.Tesis: Silenciamiento del gen rad50 en líneas celulares de cáncer de mama y análisis de su efecto en la respuesta al tratamiento con las combinaciones cis-platino/paclitaxel y cis-pla-tino/doxorubicina. Fecha de titulación: Julio,
2011. Director de tesis: Dr. César López Camarillo. UACM.
M. en C. Alma María de la Luz Villalobos Osnaya.Tesis: Caracterización de un posible inhibidor de TveIF-5A de Trichomonas vaginalis. Fecha de titulación: Julio, 2011.Directora de tesis: Dra. María Elizbeth Álvarez
Sánchez. UACM.
M. en C. José Luis Villalpando Aguilar.Tesis: Caracterización de una metaloproteinasa de Trichomonas vaginalis expresada en presen-cia de Zinc. Fecha de titulación: Julio, 2011. Directora de tesis: Dra. María Elizbeth Álvarez Sánchez. UACM.
M. en C. Cinthia Berenice García Luna.Tesis: Cambios en la expresión del transcrito regulado por cocaína y anfetaminas en el hi-potálamo lateral, en el núcleo paraventricular y en el núcleo arcuato del hipotálamo de ratas macho en dos modelos de anorexia. Fecha de
titulación: Junio, 2011. Director de tesis: Dr. Humberto Nicolini Sánchez. UACM.
M. en C. Marcos Agustín Muñoz Lino.Tesis: Búsqueda e identificación de proteínas que se expresan diferencialmente en tumores de cáncer de mama triple negativo. Fecha de titulación: Julio, 2011. Director de tesis: Dr. César López Camarillo.
UACM.

29
Yocupicio Monroy M. Escalera Cueto M., Zárate S., del Angel, R. M. Effect of human miRNAs on dengue virus infection. 30th Annual Meeting American society for Virology, Julio 16-20 2011, Minneapolis Minnesota.
Ramírez, R. Zárate S., del Angel, R. M., Yocupicio Monroy M. RIG-I hampers dengue virus infection through IFN-β dependent and independent mechanisms. 30th Annual Meeting American society for Virology, Julio 16-20 2011, Minneapolis Minnesota.
A partir del mes de mayo del 2011, el Dr. Humberto Nicolini Sánchez, profesor-Investigador del PCG-UACM pasó a ser miembro del Comité Editorial de la revista ita-liana Clinical Neuropsychiatry Journal.
DISTINCIONES
TRABAJOS PRESENTADOS
PARTICIPACIÓN DEL PCG-UACM EN CONGRESOS NACIO-NALES E INTERNACIONALES
eL trabajo reaLizado en eL pcg-uacm ha sido divuLgado en diversos congre-sos nacionaLes e internacionaLes. en estos eventos se exponen Los resuLtados más reLevantes de Las investigaciones que reaLizan Los profesores-investigado-res y estudiantes de maestría y doctorado.

30
PLANTA ACADÉMICADra. Minerva Camacho, Dr. César López, Dra. Martha Yocupicio, Dra. Elizbeth Álvarez, Dr. Humberto Nicolini, Dra. Elisa Azuara,
Dr. José de Jesús Olivares, Dr. Mauricio Castañón,Dra. Selene Zárate,Dr. José FandiñoDra. Sara Frías, INP/UACM
UACMUACMUACMUACMUACMUACM
UACMUACMUACMUACM
• Genómica humana• Genómica de agentes infecciosos en humanos• Genómica de agentes infecciosos de importancia veterinaria
LÍNEAS DE INVESTIGACIÓN El Programa de Maestría en Ciencias Genómicas pertenece al Pro-grama Nacional de Posgrados de Calidad (PNPC) en la vertiente de Programa de Fomento a la Calidad del Posgrado (PFCP) del CONACyT, por lo que los estudiantes aceptados podrán solicitar una beca para realizar estudios de Maestría.
INFORMESCatalina Sánchez,
Posgrado en Ciencias Genómicas, UACMSan Lorenzo # 290 Col. Del Valle, México, D.F.
Tel. 5488-6661 ext. [email protected]
www.uacm.edu.mx/genomicas
REQUISITOS• Maestría en área afín
• Tiempo completo• Comprensión de inglés científico
DOCUMENTOS• Solicitud de admisión
• 2 Curriculum vitae con copia de comprobantes• Original y 2 copias del certificado de
estudios de licenciatura• Original y 2 copias del acta de examen de maestría
• 2 Cartas de recomendación con copia• Original y 2 copias del acta de nacimiento
• 1 Fotografía tamaño infantil
RECEPCIÓN DE DOCUMENTOSFecha abierta
ADMISIÓN• Entrevista
• Presentación de la tesis de maestría• Calendario de inscripción abierto
Maestría DoctoradoREQUISITOS
• Licenciatura afín con promedio mínimo de 8.00• Tiempo completo
• Comprensión de inglés científico
DOCUMENTOS• Solicitud de admisión
• 2 Curriculum vitae con copia de comprobantes• 2 Copias del certificado total de estudios de licenciatura
• 2 Copias del título de licenciatura • 2 Cartas de recomendación con copia
• 2 Copias del acta de nacimiento• 2 Fotografías tamaño infantil
RECEPCIÓN DE DOCUMENTOS14 de marzo al 29 de abril de 2011
PROCESO DE ADMISIÓN4 al 13 de mayo de 2011
CURSO PROPEDÉUTICO16 de mayo al 1 de julio de 2011
RESULTADOS FINALES28 y 29 de julio de 2011
FECHA DE INICIO DE CURSOS
1 de agosto de 2011
La Universidad Autónoma de la Ciudad de México,a través del Posgrado de Ciencias Genómicas, invita a inscribirse en los programas de
CienciasMaestría y Doctorado
en
Genómicas

31
CIenCIArteRequiem en Sol Mayor
aré el acto insolente de explicar tu hazaña Soles negros pisaban tu entraña y días siderales guardaste en pestañas Tu hermoso diseño de rostro solarlucía el reflejo del gato, de ojos brillar
Quién hizo de tí un rayo tan fuerteque el cuerpo duró las vidas felinasy el corazón trasciende tu muerte. Me imagino que fuiste depositando tus ojos, tu mente, tus manos y frente, la voz incesante, en seres y amores Dejaste mirarte silencio potente Ganaste a la muerte su horrible presencia, te diste a sí misma la vida consciente, de la mujer que bebe su cáliz constante, pero devuelve a los días los años infantes Vi tus manos crear al instante, vi tus pasos pisar muy distante, Sollange Archer Motivaste la furia de vivir delirante, moviste de sitio el blanco a su flecha, jugaste tremenda cambiando tu cuerpo, haciéndolo leve, dejaste saliera a vagar con el alma, bromeaste con dios y su dados siniestros, soñaste con él y tu amor despertó su poder otra vez Hago hoy como si te hablara ayer, que vi tus exvotos llorar de mujer Las horas, minutos, segundos hiciste en pedazos de objetos rasgados de tiempo, metidos en cajas que son un juguete, un cofre de inversos tesoros, dolencias tejidas en trozos matéricos, esculturas del viaje en el cuerpo, análogos de tu tristeza silente, volcada en seres de amor estridente Te escribo fragmentos de vuelta a tu amistad generosa, a tu decente humanidad, a tu amor sorjuanesco siempre constante y, a la complicidad del poder vital de nuestro antiguo arte. Descansa, ahora sí, de tu cuerpo mínimo, Sol-archer, y avanza tu espíritu a una mejor noche, dejándonos tu luz de nuestra parte…
EFS, abril 2011

32
desde el
Imágenes del MicroUniverso
Portaobjetos:
Alteraciones en la proteína sorcina podrían estar asociadas a la presencia de hipertensión arterial esencial. La sorcina regula al receptor de rianodina cuya función es fundamental en el control del tono vascular y la presión arterial. La imagen muestra la distribución tanto del receptor de rianodina (A, rojo) como de sorcina (B, verde) en cardiomiocitos de ventrículo izquierdo de ratón. La fluorescencia se adquirió en canales separados que fueron superpues-tos (C). Se consideraron positivas para la colocalización aquellas imágenes en las cuales la sobreposición de la señal correspondiente a la sorcina y el receptor de rianodina fue mayor al 50% (D).
Créditos: Micrografía cortesía de la Dra. Angélica Rueda y Sánchez de la Vega obtenida en Microscopio Confocal Zeiss LSM510 META.

UNIVERSIDAD AUTÓNOMA DE LA CIUDAD DE MÉXICO
Esther Orozco Orozco
RECTORA
Minerva Camacho Nuez
Coordinadora Académica
Nora Isabel Huerta Guajardo
Coordinación de Difusión Cultural
y Extensión Universitaria
Jesús Fandiño
Encargado del Colegio de Ciencia y Tecnología
Genómicas hoyBoletín cuatrimestral del Posgrado en Ciencias Genómicas UACM
fué impresa en julio de 2011 en el taller de impresión de laUniversidad Autónoma de la Ciudad de Méxicocon un tiraje de dos mil quinientos ejemplares

Genómicas hoy es una publicación delPosgrado en Ciencias Genómicas de la UACM
Diseño: Alfredo Padilla














![CAMARILLO Arquitectos Ampe 2.0o [Reparado]](https://img.pdfslide.tips/doc/110x75/577c78191a28abe0548eb635/camarillo-arquitectos-ampe-20o-reparado.jpg)