Embed Size (px)
Citation preview

Eur. J. Biochem. 73, 39-55 (1977)
Assembly of Bacillus subtilis Phage @29 1. Mutants in the Cistrons Coding for the Structural Proteins
Ana CAMACHO, Fernando JIMENEZ, Javier DE LA TORRE, JosC L. CARRASCOSA, Rafael P. MELLADO, Cesar VASQUEZ, Eladio VIRUELA, and Margarita SALAS
Centro de Biologia Molecular, Centro de Investigaciones Biologicas, Consejo Superior de Investigaciones Cientificas and Universidad Autonoma de dadr id
(Received July 2/November 9, 1976)
The effect of mutations in the cistrons coding for the phage structural proteins has been studied by analyzing the phage-related structures accumulated after restrictive infection. Infection with sus mutants in cistron 8, lacking both the major head and the fiber protein, does not produce any phage- related structure, suggesting a single route for the assembly of phage @29; infection with ts mutants in this cistron produces isometric particles. Mutants in cistron 9, coding for the tail protein, TP1, produce DNA-free prolate heads with an internal core ; these particles are abortive and contain the head proteins HPO, HP1 and HP3, the upper collar protein NP2 and the nonstructural proteins p7, p15 and p16. Mutants in cistron 10, coding for the upper collar protein, NP2, produce DNA-free isometric heads also with an internal core ; they contain the head proteins and the nonstructural protein p7, suggesting that this protein forms the internal core. Mutants in cistrons 11 and 12, coding for the lower collar protein, NP3, and the neck appendages, NP1, respectively, give rise to the forma- tion of DNA-containing normal capsids and DNA-free prolate particles, more rounded at the corners than the normal capsids and with an internal core; the DNA-containing 11-particles are formed by the head proteins and the upper collar protein; the DNA-free 11-particles contain, besides these proteins, the nonstructural protein p7 and a small amount of proteins p15 and 16. The DNA- containing 12-particles have all the normal phage structural proteins except the neck appendages, formed by protein NP1; the DNA-free particles are similar to the DNA-free 11-particles.
After restrictive infection mutant sus14(1241) has a delayed lysis phenotype and produces a phage burst higher than normal, after artificial lysis. It produces DNA-containing particles, identical to wild-type phage, which have all the normal phage structural proteins, and DNA-free prolate particles, more rounded at the corners than the final phage particles and with an internal core; the last particles contain the same proteins as the DNA-free 11 or 12-particles. These particles could represent a prohead state, ready for DNA encapsulation. None of the DNA-containing particles have the nonstructural proteins p7, p15 or p16, suggesting that these proteins are released from the proheads upon DNA encapsulation.
Bacteriophage @29 is a small, morphologically complex Bacillus subtilis virus with a double-stranded DNA of molecular weight 11-12 x lo6 [l] (and Talavera, personal communication) closed as a circle by a protein linker [2]. The phage particle consists of a prolate head, a neck region formed by two collars and 12 appendages hanging from the lower collar, and a tail (Fig. 1) [l]. The most likely structure of the head consists of two 5-fold-symmetry end caps based on T= 1 icosahedral symmetry, separated by an equatorial row of five hexamers, formed mainly by protein HP1; protein HP3 forms the fibers irradiating from the
Enzymes. Lysozyme or mucopeptide N-acetylmuramylhydrolase (EC 3.2.1.17); pancreatic ribonuclease (EC 3.1.4.23); pancreatic deoxyribonuclease (EC 3.1.4.5).
phage head [3,4]. Recently a new head protein, named HPO, has been resolved f r o q the major head protein by electrophoresis using gel slabs containing 10 - 20% acrylamide gradients [5]. The neck region contains an upper collar and a lower collar formed by proteins NP2 and NP3, respectively [3,4], and 12 appendages, each one consisting of one subunit of protein NP1 [3,4,6]. The appendage protein, with a molecular weight of 80000 [3] is synthesized as a precursor, P-NP1, of molecular weight 90000 [7,8], which is cleaved before its assembly in the phage particle [5,6].
Up to 23 proteins have been reported to be induced in ultraviolet-irradiated B. subtilis after ~$29 infection [5,9,10]. However, two of these proteins are the precursor and product fragment in the synthesis of the neck appendage protein.

40 Assembly of Phage a29
DNA synthesis
Late control
TPI NP2 NP3
p7 HpIlii- Structural proteins
p15 p16
Lysis
P'7
DNA synthesis
Fig. 1. Genetic, map ~f hrrc,twiophagc, @2Y. The genetic map of Mellado ef a/. [14] has been used. The genetic distance between mutants have been drawn proportional to recombination frequencies. A p before a gene number refers to the identified polypeptide chain of that gene. HPI, HP3, TP1, NP2, NP3 and NP1 are the phage structural proteins
Recently, the mutants from the collections of Tala- Vera et al. [l I], Reilly et al. [I21 and Moreno et al. [13] have been integrated in a common genetic map with a total of 17 genes [14]. Fig. 1 shows the genetic map of phage @29 with the new nomenclature adopted. The protein products of eleven cistrons have been charac- terized. Six cistrons, 1,2, 3, 5, 6 and 17, are involved in phage-specific DNA synthesis [5]. Cistron 4 is involved in the control of late transcription (unpublished results) ; cistrons 8 - 12 code for the phage structural proteins [5, 151. Cistrons 7, 15 and 16 code for late proteins synthesized in high amounts [5, 7, 161. We do not know at present which is the protein product of cistron 13 and whether it codes for an early or late protein. Finally, cistron 14 is involved in the lysis of the infected bacteria since mutants in this cistron, under restrictive conditions, have a delayed lysis phenotype and produce a phage burst higher than normal after artificial lysis [5].
We have started a systematic analysis of bac- teriophage @29 morphogenesis. The phage-related structures accumulated after infection of B. subtilis, under restrictive conditions, with sus or ts mutants in the different cistrons have been analyzed by sucrose gradient sedimentation, electron microscopy and gel electrophoresis. In this paper we describe the results obtained by using mutants in the cistrons coding for the phage structural proteins. A preliminary account of some of these results has already been given [4]. In the accompanying paper, we show the effect on phage morphogenesis of mutations in genes coding for non- structural proteins; we also propose a morphogenetic pathway for the assembly of phage @29.
MATERIALS AND METHODS
Bacteria, Phage and Media
The nonpermissive host, B. subtilis 110NA try- spoA- su- and the permissive bacteria, B. subtilis 168 MO-99 spoA-[met- thr-]+ suf3 were described
The new nomenclature adopted by Mellado et al. [14] for the @29 mutants has been used. The tsmutants were from the collection of Talavera et al. [l 11. Most of the susmutants used were from thecollection of Moreno et al. [13]. Mutant sus12(610) was isolated by Mellado et al. [17] on the B.subtilis strain MO-101-P spoA- thr-[met-]+ S U + ~ . Mutant ts9(l)sus14(1241) was
isolated by Moreno et al. [13], being their mutant HA124; by elimination of the is9 mutation, mutant sus14(1241) was obtained. In all cases, double mutants with susl4( 1241), which produces delayed lysis [5], were used.
Defined medium [I61 and phage buffer [3] were as described.
1131.
Labeling and Purification of Phage @29
B. subtilis 110NA su- was grown in defined medium at 37 "C up to a concentration of 1 x lo8 bacteria/ml. The cells were concentrated 5-fold by centrifugation at room temperature and resuspension in the same medium with 0.5 mM amino acids and infected with @29 at a multiplicity of 20. A 3H-labeled amino acid mixture (50 pCi/ml) was added and the incubation continued until lysis. The phage was

A. Camacho, F. Jimenez, J. de la Torre, J. L. Carrascosa, R. P. Mellado, C. Vasquez, E. Vifiuela, and M. Salas 41
purified as described [3] except that the sucrose gradient centrifugation was omitted for the phage purification. The empty heads, produced in normal lysates, were further purified by centrifugation through 5 ml of a 15-30% (w/v) sucrose gradient in phage buffer for 75 min at 165000 x g at 5 "C.
Labeling and Isolation of Mutant Particles
B. subtilis 110NA su- was grown in defined medium at 42 "C up to a concentration of lo8 bac- teria/ml; the cells were concentrated 5-fold by centrif- ugation at room temperature and resuspension in the same medium with 1 mM amino acids when the cells were labeled with a 14C-labeled protein hydrolysate (5 pCi/ml; 55 Ci/atom C); when the cells were labeled with [35S]methionine (50 pCi/ml) the concentration of methionine was 0.04 mM. To label the DNA, [3H]- thymidine (10 pCi/ml; 19 Ci/mmol) was added in the presence of uridine (200 pg/ml); in this case 2 pCi/ml of 14C-labeled protein hydrolysate were used. In all cases the bacteria were infected with the mutant phages at a multiplicity of 20 and shaken at 42 "C. At different times, aliquots were taken to follow phage development and to assay for acid- insoluble radioactivity. All labeled precursors were obtained from The Radiochemical Center, Amersham.
90 min after infection, a time at which cell lysis had not occurred when infection with a double mutant containing a sus14(1241) mutation was carried out and maximal phage production had already taken place after infection with mutant sus14(1241), the cells were concentrated by centrifugation at 4 "C and subjected to one of the following two lysis and isola- tion procedures.
a) The cell pellet was resuspended in one-tenth the original volume of a buffer containing 20 mM Tris-HCI, pH 7.8, 1 mM EDTA and 25% (w/v) sucrose in the presence of lysozyme (1 mg/ml) and the cells were incubated for 2 h at 0 "C. After this time, more than 90% of the bacteria had been converted to protoplasts ; these were lysed by dilution in phage buffer up to the original volume in the presence of pancreatic DNase I (10 pg/ml) and incubation for 20 rnin at 42 "C. The lysates were centrifuged for 15 min at 10000rev./min in a SS34 rotor of the Sorvall centrifuge to remove bacterial debris. The supernatant, containing most of the phage particles, was centrifuged for 90 min at 133000 x g at 4 "C. The sediment, containing the phage particles, was resus- pended overnight in a small volume of phage buffer and centrifuged in a sucrose gradient as described below.
b) The cell pellet was resuspended in half the original volume of phage buffer in the presence of lysozyme (1 mgiml). After 3-4 rnin at 42 "C, cell
lysis took place and the lysate was immediately centrifuged in a sucrose gradient as described below.
Sucrose Gradient Cen trifigution
Preparative Gradients. Linear 15 - 30% (w/v) sucrose gradients in phage buffer (7.5 ml of each solution) were prepared in 18-ml nitrocellulose tubes. In some cases, when the cell membrane fraction was analyzed,l mlofaCsClsolutionofdensity 1.6g x cm-3 in phage buffer was added at the bottom of the tube. 1 ml of the corresponding lysate or high-speed super- natant was layered on the top of the gradient. Centrifu- gation was carried out at 5 "C and 88000 x g for 3 h or 5.5 h depending on whether DNA-containing or DNA-free particles were to be examined. Fractions of about 0.5 ml were collected and the total radio- activity was determined in an aliquot. The fractions containing the phage particles were pooled and either frozen for further analysis on polyacrylamide gel electrophoresis or dialyzed against phage buffer and stored at 4 "C. Electron microscopy of the particles was carried out both before and after dialysis. The isolation of the particles produced after treatment with 2.5 M NaCl was carried out as described above except that the phage buffer contained 2.5 M NaCl and the centrifugation was for about 22 h.
Analytic Gradients. Linear 15 - 30 "/, or 5 - 20'x (w/v) sucrose gradients in phage buffer (2.2-2.5 ml of each solution) were prepared in 5 ml nitrocellulose tubes. When indicated, 0.5 ml of a CsCl solution of density 1.6 g x cm-3 in phage buffer was added at the bottom of the tube. About 0.2 ml of a solution con- taining the corresponding lysate or high-speed super- natant and, as sedimentation markers, differently labeled phage di29 and/or purified empty heads, were layered on the top of the gradient. The sedimenta- tion coefficient of the different particles produced after infection with mutants was determined in 5 - 20'x sucrose gradients; values of 256 S and 120 S were used for purified phage [I81 and empty heads [3], respectively. When 15 - 30% sucrose gradients were used, centrifugation was carried out at 5 "C for 40 rnin or 75 min at 165000 x g depending on whether DNA- containing or DNA-free particles were being analyzed. When 5 - 20% sucrose gradients were used, centrifuga- tion was carried out at 5 ('C for 30 rnin or 70 rnin at 114000 x g depending on whether DNA-containing or DNA-free particles were analyzed. Fractions of about 0.15 ml were taken, precipitated with 5 o/, trichloroacetic acid in the presence of 100 pg of bovine serum albumin and the acid-insoluble radioactivity was determined as described [3]. For the analysis of the particles resulting from the 2.5 M NaCl treatment, 15 - 30 "/, sucrose gradients were used ; the phage buffer contained 2.5 M NaCl and centrifugation was carried out for 3.5 h at 216000 x g.

42 Assembly of Phage @29
Gel Electrophoresis
Gel electrophoresis was carried out either in slabs or in tubes as indicated in each case. Slab gel electro- phoresis was performed on linear 10 - 20 % acrylamide gradients as described [5]. Electrophoresis was carried out at room temperature for 15 h at a constant current of 20 mA per slab. For a better analysis of the phage structural proteins coelectrophoresis in tubes (15 cm long) was carried out by using linear gradients prepared with 16 -20% acrylamide solu- tions. Electrophoresis was carried out at room temperature at a constant voltage of 100 V for 11.5 h. In all cases the stacking gel was as described [19]. The electrophoresis buffer and the samples for electro- phoresis were prepared as indicated [5]. After electro- phoresis, the gel slabs were dried under vacuum and autoradiography was performed on Kodirex X-ray film [20]. Densitometry was carried out with a chromoscan MK I1 densitometer at 610 -690 nm. The quantitisation of the number of subunits of the non- structural proteins present in phage particles was carried out by densitometry of the autoradiographs, exposed for different time periods to obtain a linear response in the absorbance and weighing the peaks corresponding to the protein to be estimated and that of either protein HP1 or NP2. A number of 85 subunits ofprotein HP1 or 5 subunits of protein NP2 per particle [4] was considered. In the case of mutants in cistron 10, coding for protein NP2, which produce isometric heads, a number of 55 subunits of protein HP1 per particle was considered. When electrophoresis in tubes was carried out, the gels were cut in I-mm slices with a Mickle gel slicer and the radioactivity deter- mined as described [ 161.
Electron Microscopy
Lysis was carried out in situ by an adaptation of the method described by Kellenberger et al. [21]. The cells were grown and infected with double mutants con- taining a sus14(1241) mutation to inhibit cell lysis, as indicated before, At 90 min after infection the infected cells were concentrated 10-fold by centrifugation and resuspension in a buffer containing 20 mM Tris-HC1, pH 7.8, 1 mM EDTA, 25% (w/v) sucrose and lyso- zyme (500 pg/ml) and incubated for 75 min at 0 "C. After this time, 1 pl of a 2% osmium tetroxide solution was added to 0.25 ml of the cell suspension and a drop of this mixture was immediately added to grids with carbon-coated colodion films. After 2 min, the grids were washed twice with phage buffer and finally stained with 2 % uranyl acetate in water. The grids were examined in a Jeol JEM 100-B electron microscope.
For the observation of phage particles with the electron microscope, a drop of the phage suspension
was placed on a grid with a carbon-coated colodion film. After 30 s the grid was dried by touching it with the edge of filter paper. When fractions from a sucrose gradient were analyzed, the grids were washed with phage buffer and dried as before. Finally, the sample was stained by addition of a drop of 2% uranyl acetate in water and dried.
RESULTS
Cistron 14
Infection of B.subti1i.s sup with mutant sus14(1241) gives rise to delayed lysis and produces, after artificial lysis, a burst-size of infective phage higher than that obtained after wild-type phage @29 infection [5] (and results not shown). Even though this mutant contains, besides the mutation which gives rise to the delayed lysis phenotype, two other additional mutations in cistrons 6 and 17, producing, after infection under restrictive conditions, a large fragment of each of these two proteins, it behaves like wild-type phage. Since all the mutants described in this paper contained, besides the corresponding mutation to be analyzed, the sus14( 1241) mutation, to prevent spontaneous lysis, the results obtained after infection of B. subtilis su- with this mutant will be described first. In all the cases, infection with the corresponding single mutant, without the susl4( 1241) mutations, was carried out and the results were the same as those obtained with the double mutant (results not shown) ; this discards the possibility that the two additional mutations in genes 6 and 17, present in mutant sus14(1241), were affecting the results obtained.
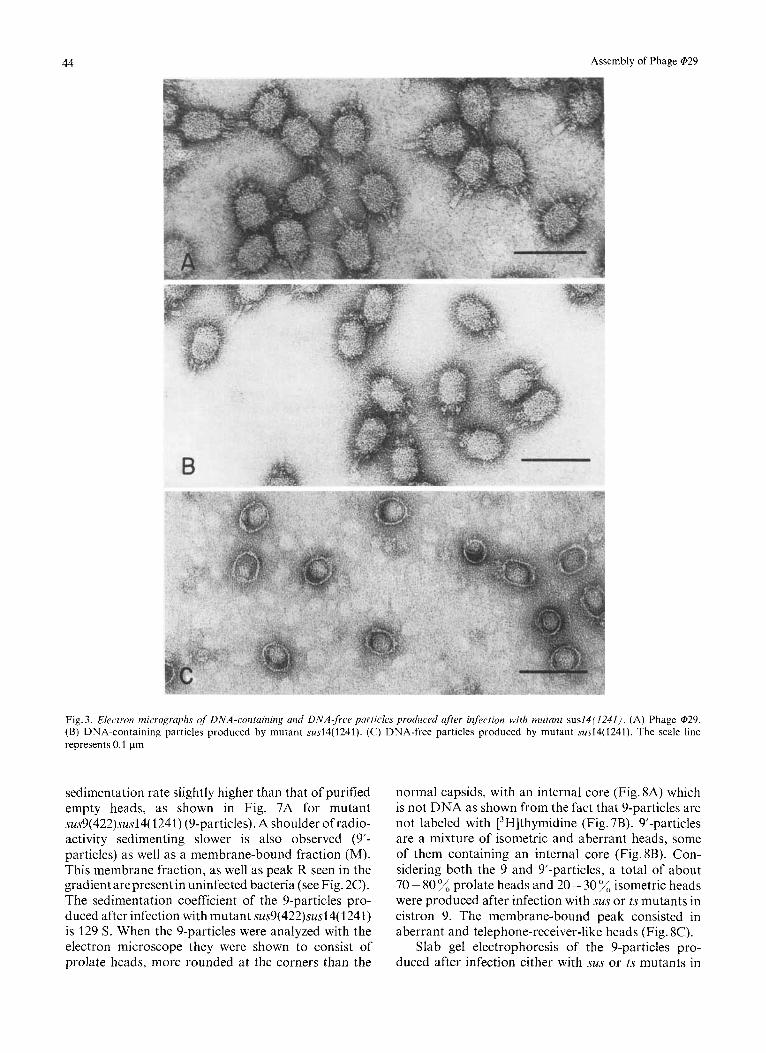
Infection of B.subtilis su- with mutant sus14(1241) produces the formation of DNA-containing infective phage particles, which, in sucrose gradient centrifuga- tion, have the same sedimentation rate as wild-type @29 particles (Fig.2A), and DNA-free heads with a sedimentation rate slightly higher than that of purified wild-type empty heads (Fig. 2B). The proportion of DNA-containing particles is about 40% relative to the total amount of particles produced, as determined from the radioactivity present in the DNA-containing and DNA-free particles (see Fig. 2A). The peak moving with a sedimentation rate slower than that of DNA-free heads is also present in uninfected bacteria (Fig. 2C) and it contains ribosomes (peak R). Fig. 3B shows the electron micrographs of the DNA-containing particles, which are indistinguishable from those produced after infection with wild-type phage (see Fig. 3A) [l, 31. Fig. 3C shows the electron micrographs of DNA-free heads; as can be seen, many of the particles are more rounded at the corners than normal phage and con- tain a core, which could be responsible for the slightly higher sedimentation rate of those particles relative to that of purified empty heads which lose the
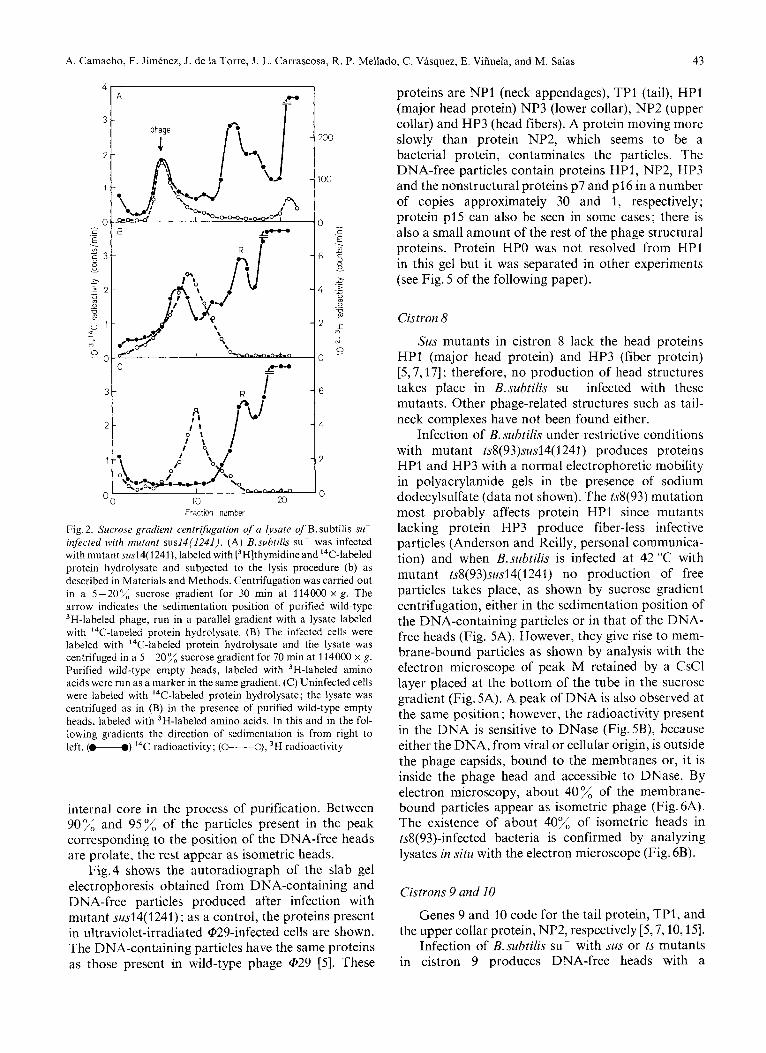
A. Camacho, F. Jimenez, J. de la Torre, J. L. Carrascosa, R. P. Mellado, C. Vasquez, E. Vifiuela, and M. Salas 43
4 - P A
I
- 1
Fraction number
Fig.2. Sucrose gradient centrifugation of a Iysate of B.subtilis su- infected with mutant susI4(1241). (A) B.subtilis su- was infected with mutant susl4( 1241), labeled with [3H]thymidine and I4C-labeled protein hydrolysate and subjected to the lysis procedure (b) as described in Materials and Methods. Centrifugation was carried out in a 5-20% sucrose gradient for 30 min at 114000 x g . The arrow indicates the sedimentation position of purified wild-type 3H-labeled phage, run in a parallel gradient with a lysate labeled with 14C-labeled protein hydrolysate. (B) The infected cells were labeled with 14C-labeled protein hydrolysate and the lysate was centrifuged in a 5 -20% sucrose gradient for 70 min at 114000 x g . Purified wild-type empty heads, labeled with 3H-labeled amino acids were run as a marker in the same gradient. (C) Uninfected cells were labeled with I4C-labeled protein hydrolysate; the lysate was centrifuged as in (B) in the presence of purified wild-type empty heads, labeled with 3H-labeled amino acids. In this and in the fol- lowing gradients the direction of sedimentation is from right to left. (M) 14C radioactivity; (O----O), 3H radioactivity
internal core in the process of purification. Between 90% and 95%, of the particles present in the peak corresponding to the position of the DNA-free heads are prolate, the rest appear as isometric heads.
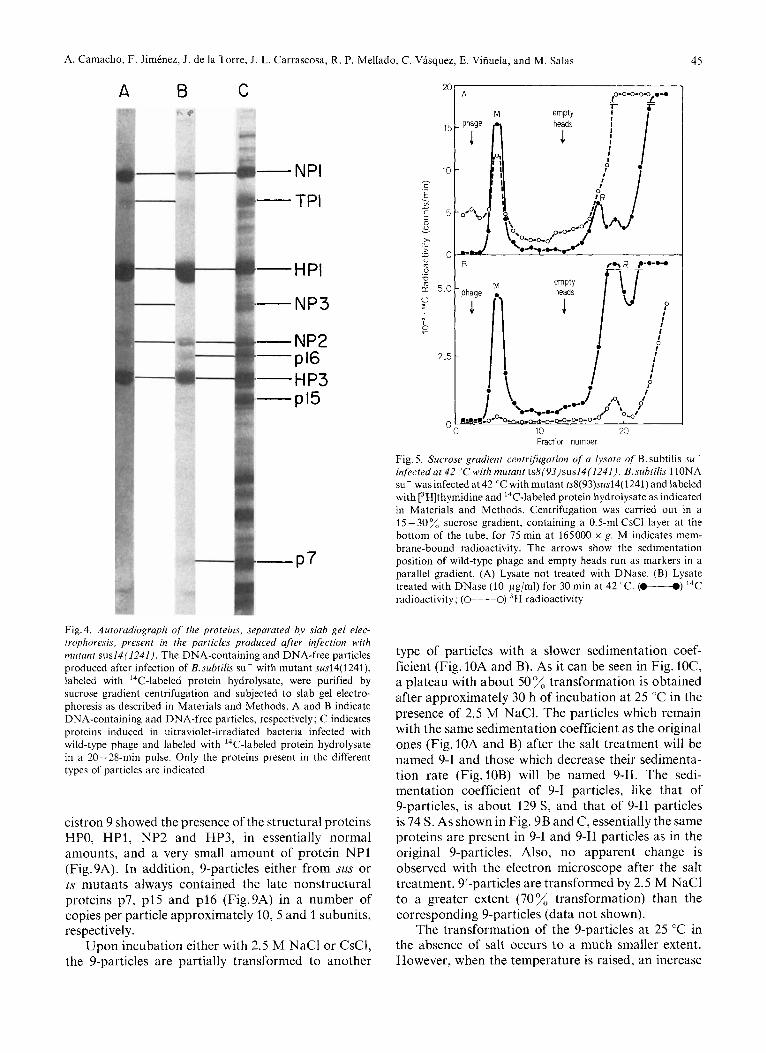
Fig.4 shows the autoradiograph of the slab gel electrophoresis obtained from DNA-containing and DNA-free particles produced after infection with mutant sus14(1241); as a control, the proteins present in ultraviolet-irradiated Qi29-infected cells are shown. The DNA-containing particles have the same proteins as those present in wild-type phage @29 [5]. These
proteins are NP1 (neck appendages), TP1 (tail), HP1 (major head protein) NP3 (lower collar), NP2 (upper collar) and HP3 (head fibers). A protein moving more slowly than protein NP2, which seems to be a bacterial protein, contaminates the particles. The DNA-free particles contain proteins HP1, NP2, HP3 and the nonstructural proteins p7 and p16 in a number of copies approximately 30 and 1, respectively; protein p15 can also be seen in some cases; there is also a small amount of the rest of the phage structural proteins. Protein HPO was not resolved from HP1 in this gel but it was separated in other experiments (see Fig. 5 of the following paper).
Cistron 8
Sus mutants in cistron 8 lack the head proteins HP1 (major head protein) and HP3 (fiber protein) [5,7,17] ; therefore, no production of head structures takes place in B.subtilis sup infected with these mutants. Other phage-related structures such as tail- neck complexes have not been found either.
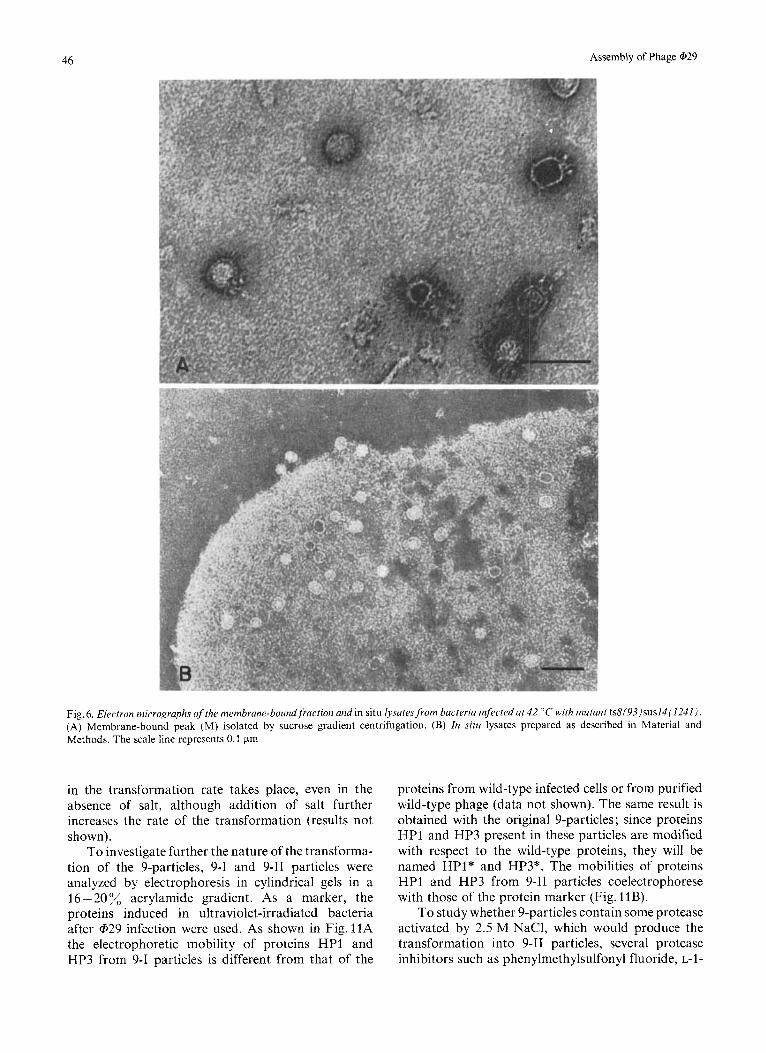
Infection of B. subtilis under restrictive conditions with mutant ts8(93)sus14(1241) produces proteins HP1 and HP3 with a normal electrophoretic mobility in polyacrylamide gels in the presence of sodium dodecylsulfate (data not shown). The ts8(93) mutation most probably affects protein HP1 since mutants lacking protein HP3 produce fiber-less infective particles (Anderson and Reilly, personal communica- tion) and when B.subtilis is infected at 42 "C with mutant ts8(93)sus14( 1241) no production of free particles takes place, as shown by sucrose gradient centrifugation, either in the sedimentation position of the DNA-containing particles or in that of the DNA- free heads (Fig. 5A). However, they give rise to mem- brane-bound particles as shown by analysis with the electron microscope of peak M retained by a CsCl layer placed at the bottom of the tube in the sucrose gradient (Fig. 5A). A peak of DNA is also observed at the same position ; however, the radioactivity present in the DNA is sensitive to DNase (Fig.5B), because either the DNA, from viral or cellular origin, is outside the phage capsids, bound to the membranes or, it is inside the phage head and accessible to DNase. By electron microscopy, about 40% of the membrane- bound particles appear as isometric phage (Fig. 6A). The existence of about 40% of isometric heads in ts8(93)-infected bacteria is confirmed by analyzing lysates in situ with the electron microscope (Fig. 6B).
Cistrons 9 and I0
Genes 9 and 10 code for the tail protein, TPI, and the upper collar protein, NP2, respectively [5,7,10,15].
Infection of B.subtilis su- with sus or ts mutants in cistron 9 produces DNA-free heads with a
44 Assembly of Phage 029
Fig. 3. Electron micrographs of DNA-containing und DNA-free particles produced after infection with mutant sus14(1241). (A) Phage @29. (B) DNA-containing particles produced by mutant .sus14(1241). (C) DNA-free particles produced by mutant sus14(1241). The scale line represents 0.1 pm
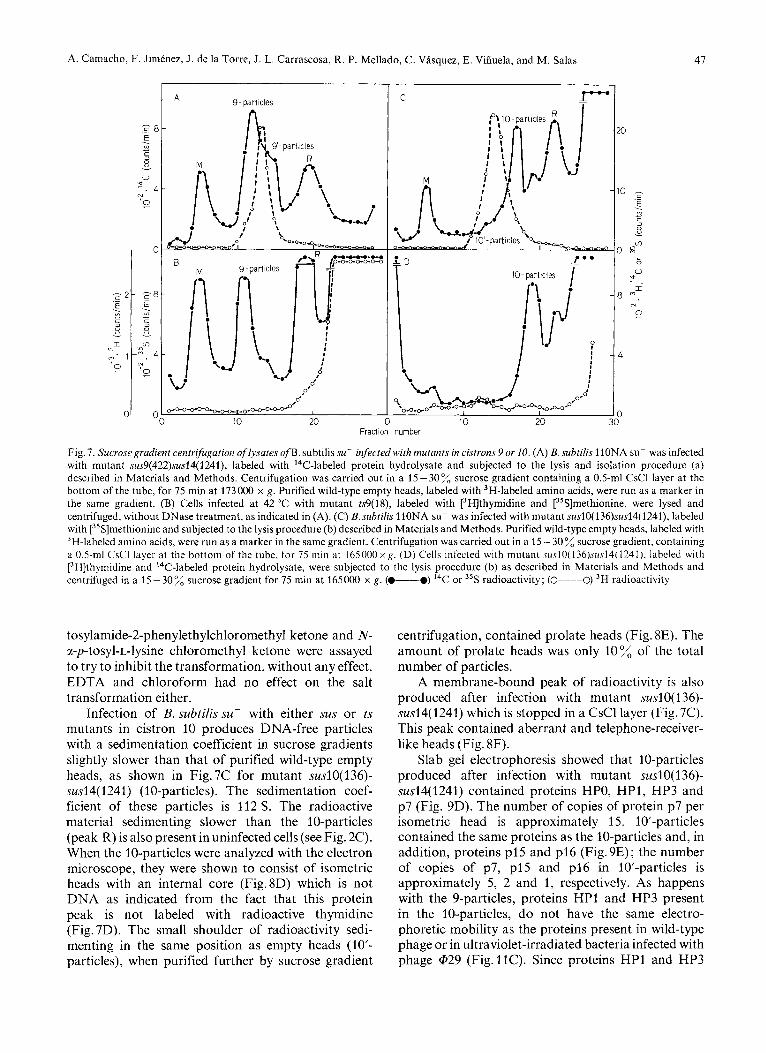
sedimentation rate slightly higher than that of purified empty heads, as shown in Fig. 7A for mutant sus9(422)sus14( 1241) (9-particles). A shoulder of radio- activity sedimenting slower is also observed (9’- particles) as well as a membrane-bound fraction (M). This membrane fraction, as well as peak R seen in the gradient are present in uninfected bacteria (see Fig. 2C). The sedimentation coefficient of the 9-particles pro- duced after infection with mutant sus9(422)sus14(1241) is 129 S. When the 9-particles were analyzed with the electron microscope they were shown to consist of prolate heads, more rounded at the corners than the
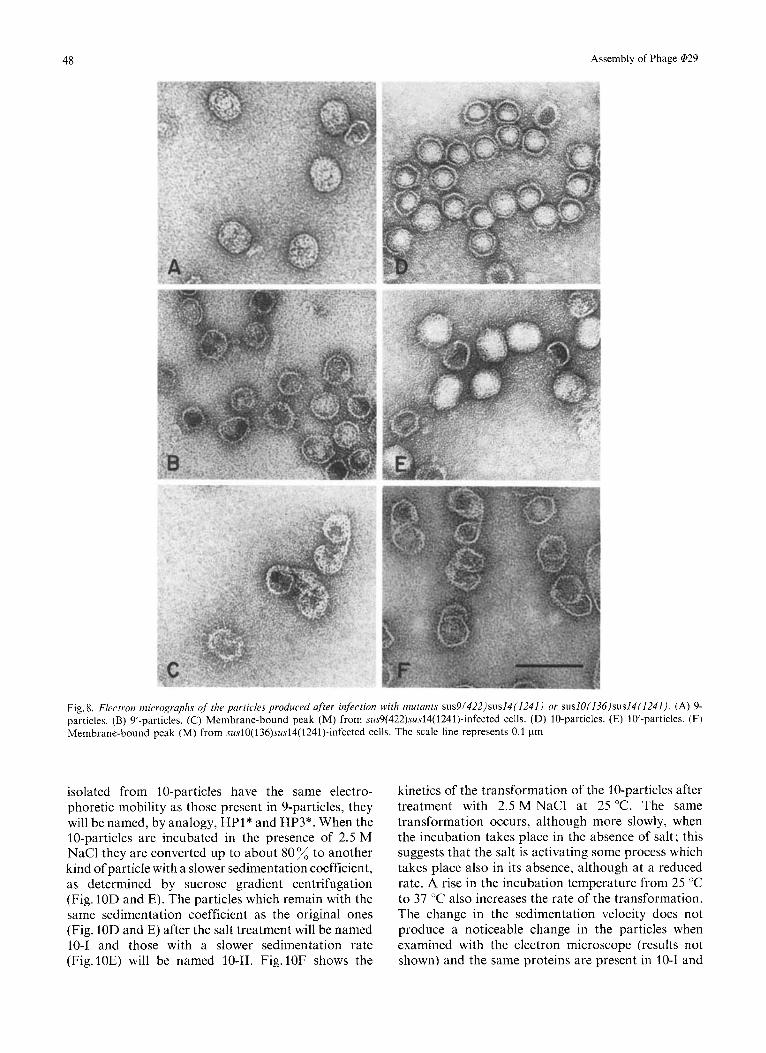
normal capsids, with an internal core (Fig. 8A) which is not DNA as shown from the fact that 9-particles are not labeled with [3H]thymidine (Fig. 7B). 9’-particles are a mixture of isometric and aberrant heads, some of them containing an internal core (Fig.8B). Con- sidering both the 9 and 9’-particles, a total of about 70 - 80 % prolate heads and 20 - 30 isometric heads were produced after infection with sus or ts mutants in cistron 9. The membrane-bound peak consisted in aberrant and telephone-receiver-like heads (Fig. 8C).
Slab gel electrophoresis of the 9-particles pro- duced after infection either with sus or ts mutants in
A. Camacho, F. Jimenez, J. de la Torre, J. L. Carrascosa, R. P. Mellado, C. Vasquez, E. Vihuela, and M. Salas 4s
A B C
- NPI
- TPI
--PI
- NP3
- NP2 -p16 -HP3 --I5
- P 7
Fig.4. Autoradiograph of the proteins, separated by slab gel elec- trophoresis, present in the particles produced after infection with mutant sus14(1241 j . The DNA-containing and DNA-free particles produced after infection of B. subtilis su- with mutant sus14(1241), labeled with ''C-labeled protein hydrolysate, were purified by sucrose gradient centrifugation and subjected to slab gel electro- phoresis as described in Materials and Methods. A and B indicate DNA-containing and DNA-free particles, respectively; C indicates proteins induced in ultraviolet-irradiated bacteria infected with wild-type phage and labeled with ''C-labeled protein hydrolysate in a 20-28-min pulse. Only the proteins present in the different types of particles are indicated
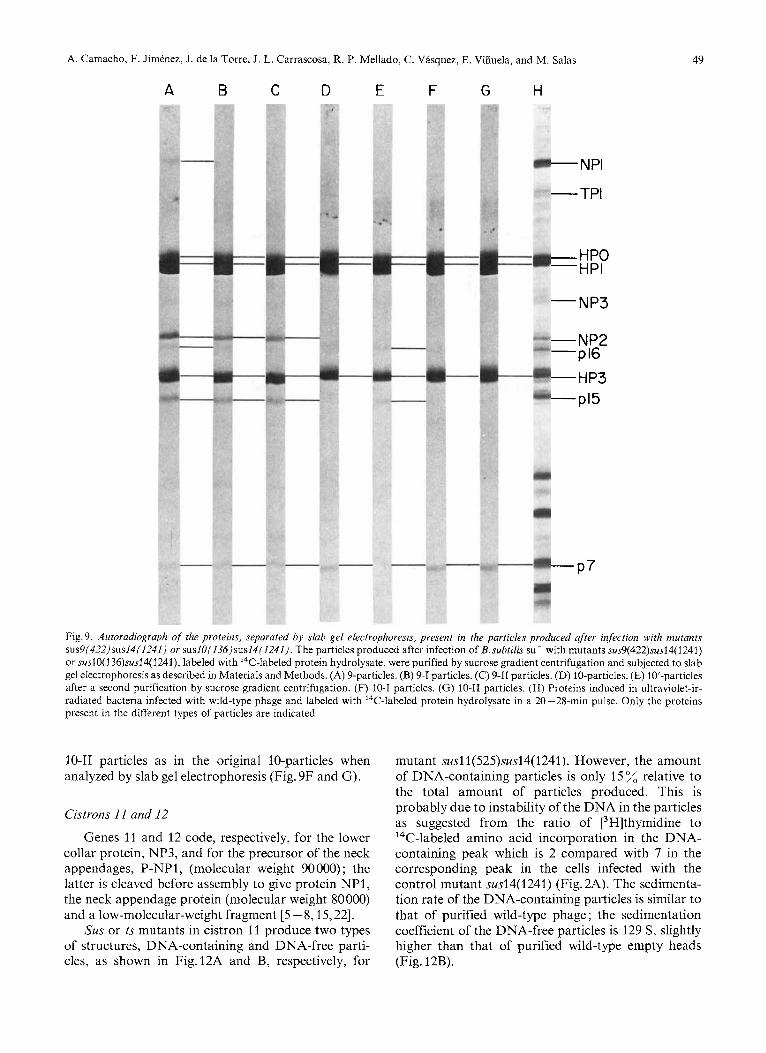
cistron 9 showed the presence of the structural proteins HPO, HP1, NP2 and HP3, in essentially normal amounts, and a very small amount of protein NPI (Fig.9A). In addition, 9-particles either from sus or ts mutants always contained the late nonstructural proteins p7, p15 and p16 (Fig.9A) in a number of copies per particle approximately 10,5 and 1 subunits, respectively.
Upon incubation either with 2.5 M NaCl or CsC1, the 9-particles are partially transformed to another
2
1
t i - c .- E . I v)
C I 3 0 u
>. c > - 1
m U
- .-
0
d 5.1
v I 0
2.
I
Fraction number
Fig. 5. Sucrose gradient centrifugation qf a lysate of B. subtilis su- infected at 42 "C with mutant ts8(93)sus14(I241 j . B.suhtilis 1 lONA su- was infected at 42 "C with mutant ts8(93).sus14( 1241) and labeled with ['Hlthymidine and 14C-labeled protein hydrolysate as indicated in Materials and Methods. Centrifugation was carried out in a 15-30% sucrose gradient, containing a 0.5-ml CsCl layer a t the bottom of the tube, for 75 min at 165000 x g. M indicates mem- brane-bound radioactivity. The arrows show the sedimentation position of wild-type phage and empty heads run as markers in a parallel gradient. (A) Lysate not treated with DNase. (B) Lysate treated with DNase (10 pg/ml) for 30 min at 42 'C. (-) 14C radioactivity; (O----O) 'H radioactivity
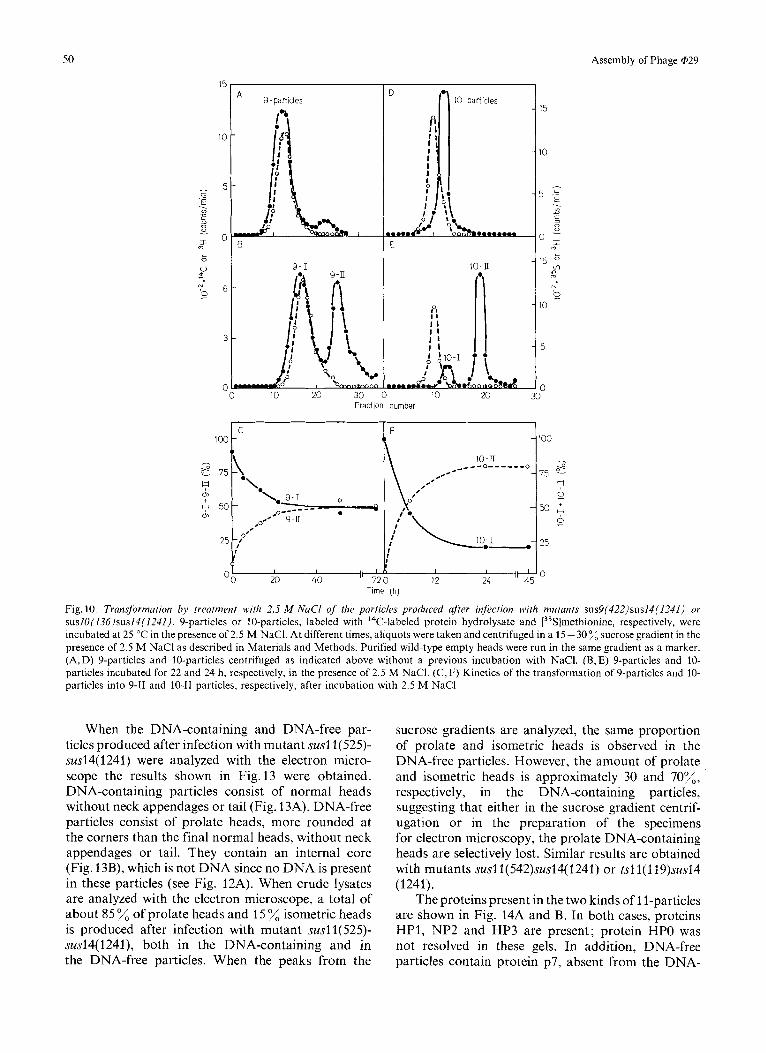
type of particles with a slower sedimentation coef- ficient (Fig. 10A and B). As it can be seen in Fig. lOC, a plateau with about 50 transformation is obtained after approximately 30 h of incubation at 25 OC in the presence of 2.5 M NaC1. The particles which remain with the same sedimentation coefficient as the original ones (Fig. 10A and B) after the salt treatment will be named 9-1 and those which decrease their sedimenta- tion rate (Fig.lOB) will be named 9-11. The sedi- mentation coefficient of 9-1 particles, like that of 9-particles, is about 129 S, and that of 9-11 particles is 74 S. As shown in Fig. 9B and C, essentially the same proteins are present in 9-1 and 9-11 particles as in the original 9-particles. Also, no apparent change is observed with the electron microscope after the salt treatment. 9'-particles are transformed by 2.5 M NaCl to a greater extent (70%) transformation) than the corresponding 9-particles (data not shown).
The transformation of the 9-particles at 25 "C in the absence of salt occurs to a much smaller extent. However, when the temperature is raised, an increase
46 Assembly of Phage @29
Fig. 6 . Electron micrographs of the membrane-boundfraction and in situ lysates from bacteria infected at 42 "C with mutant ts8(93)sus14(1241). (A) Membrane-bound peak (M) isolated by sucrose gradient centrifugation. (B) In siru lysates prepared as described in Material and Methods. The scale line represents 0.1 pm
in the transformation rate takes place, even in the absence of salt, although addition of salt further increases the rate of the transformation (results not shown).
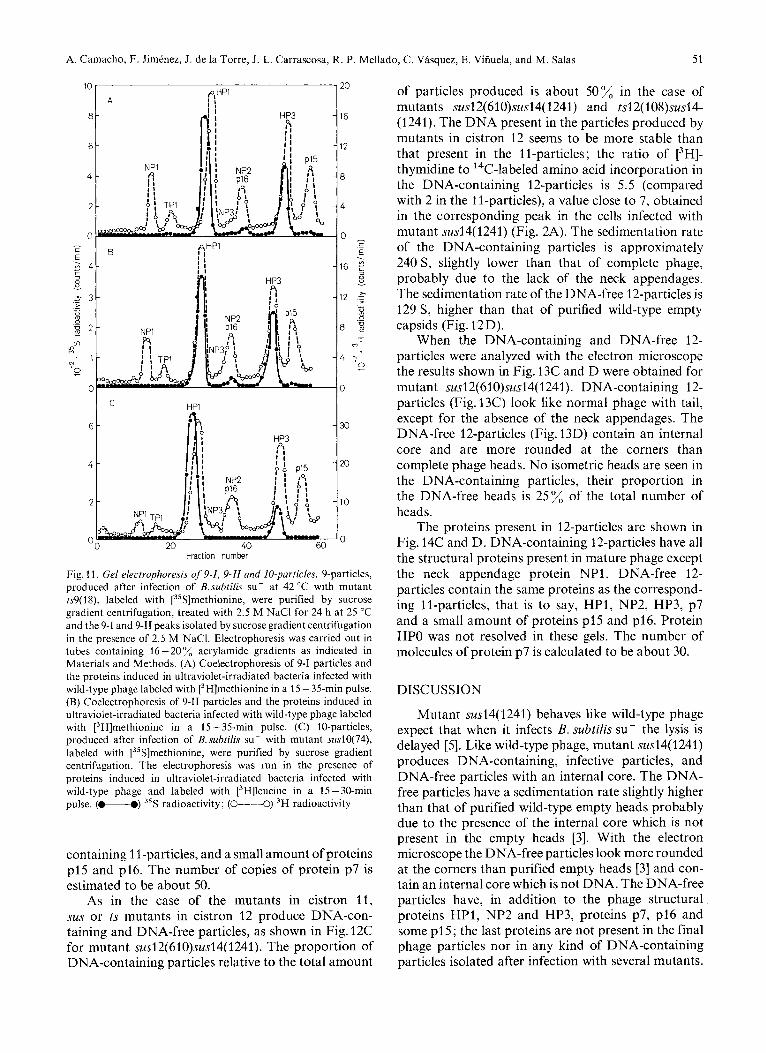
To investigate further the nature of the transforma- tion of the 9-particles, 9-1 and 9-11 particles were analyzed by electrophoresis in cylindrical gels in a 16 - 20 acrylamide gradient. As a marker, the proteins induced in ultraviolet-irradiated bacteria after @29 infection were used. As shown in Fig. 11A the electrophoretic mobility of proteins HP1 and HP3 from 9-1 particles is different from that of the
proteins from wild-type infected cells or from purified wild-type phage (data not shown). The same result is obtained with the original 9-particles ; since proteins HP1 and HP3 present in these particles are modified with respect to the wild-type proteins, they will be named HP1* and HP3*. The mobilities of proteins HP1 and HP3 from 9-11 particles coelectrophorese with those of the protein marker (Fig. 11B).
To study whether 9-particles contain some protease activated by 2.5 M NaCl, which would produce the transformation into 9-11 particles, several protease inhibitors such as phenylmethylsulfonyl fluoride, L-1-
A. Camacho, F. Jimtnez, J. de la Torre, J. L. Carrascosa, R. P. Mellado, C. Vasquez, E. Vifiuela, and M. Salas 41
9 - particles A C
10 20
20
10 .- E . - w c 3
8 - o
3 % L 0
V
I 0 - .
8 -. 0
N
4
? Fraction number
Fig. 7. Sucrose gradient centrifugation of lysates of B. subtilis su- infected with mutunts in cistrons 9 or 10. (A) B. subtilis 110NA su- was infected with mutant sus9(422)sus14(1241), labeled with ''C-labeled protein hydrolysate and subjected to the lysis and isolation procedure (a) described in Materials and Methods. Centrifugation was carried out in a 15-30% sucrose gradient containing a 0.5-ml CsCl layer at the bottom of the tube, for 75 min at 173000 x g. Purified wild-type empty heads, labeled with 3H-labeled amino acids, were run as a marker in the same gradient. (B) Cells infected at 42 "C with mutant ts9(18), labeled with [3H]thymidine and [35S]methionine, were lysed and centrifuged, without DNase treatment, as indicated in (A). (C) B.subtilis 11ONA su- was infected with mutant sus10(136)sus14(1241), labeled with [35S]methionine and subjected to the lysis procedure (b) described in Materials and Methods. Purified wild-type empty heads, labeled with 3H-labeled amino acids, were run as a marker in the same gradient. Centrifugation was carried out in a 15 - 30% sucrose gradient, containing a 0.5-ml CsCl layer at the bottom of the tube, for 75 rnin at 165000xg. (D) Cells infected with mutant su.s10(136)su.s14(1241). labeled with [3H]thymidine and 14C-labeled protein hydrolysate, were subjected to the lysis procedure (b) as described in Materials and Methods and centrifuged in a 15-30% sucrose gradient for 75 min at 165000 x g. (-0) I4C or 35S radioactivity; (0----0) 3H radioactivity
tosylamide-2-phenylethylchloromethyl ketone and N- a-p-tosyl-L-lysine chloromethyl ketone were assayed to try to inhibit the transformation, without any effect. EDTA and chloroform had no effect on the salt transformation either.
Infection of B. subtilis szi- with either sus or ts mutants in cistron 10 produces DNA-free particles with a sedimentation coefficient in sucrose gradients slightly slower than that of purified wild-type empty heads, as shown in Fig.7C for mutant suslO(136)- sus14(1241) (10-particles). The sedimentation coef- ficient of these particles is 112 S. The radioactive material sedimenting slower than the 10-particles (peak R) is also present in uninfected cells (see Fig. 2C). When the 10-particles were analyzed with the electron microscope, they were shown to consist of isometric heads with an internal core (Fig.8D) which is not DNA as indicated from the fact that this protein peak is not labeled with radioactive thymidine (Fig. 7D). The small shoulder of radioactivity sedi- menting in the same position as empty heads (10'- particles), when purified further by sucrose gradient
centrifugation, contained prolate heads (Fig. 8E). The amount of prolate heads was only 10% of the total number of particles.
A membrane-bound peak of radioactivity is also produced after infection with mutant swlO( 136)- sus14(1241) which is stopped in a CsCl layer (Fig. 7C). This peak contained aberrant and telephone-receiver- like heads (Fig. 8F).
Slab gel electrophoresis showed that 10-particles produced after infection with mutant suslO( 136)- sus14(1241) contained proteins HPO, HP1, HP3 and p7 (Fig. 9D). The number of copies of protein p7 per isometric head is approximately 15. 10-particles contained the same proteins as the 10-particles and, in addition, proteins p15 and p16 (Fig. 9E); the number of copies of p7, p15 and p16 in 10'-particles is approximately 5, 2 and 1, respectively. As happens with the 9-particles, proteins HP1 and HP3 present in the 10-particles, do not have the same electro- phoretic mobility as the proteins present in wild-type phage or in ultraviolet-irradiated bacteria infected with phage @29 (Fig.11C). Since proteins HP1 and HP3
48 Assembly of Phage @29
Fig. 8. Electron micrographs of the purticles prodwed after infection w'ith mutants sus9(422)sus14(1241) or sus10(136)sus14(1241). (A) 9- particles. (B) 9'-particles. (C) Membrane-bound peak (M) from sus9(422)sus14(1241)-infected cells. (D) 10-particles. (E) 10-particles. (F) Membrane-bound peak (M) from susl0(l3h)susl4(1241)-infected cells. The scale line represents 0.1 pm
isolated from 10-particles have the same electro- phoretic mobility as those present in 9-particles, they will be named, by analogy, HP1" and HP3*. When the 10-particles are incubated in the presence of 2.5 M NaCl they are converted up to about 80% to another kind of particle with a slower sedimentation coefficient, as determined by sucrose gradient centrifugation (Fig. 10D and E). The particles which remain with the same sedimentation coefficient as the original ones (Fig. 10D and E) after the salt treatment will be named 10-1 and those with a slower sedimentation rate (Fig.lOE) will be named 10-11. Fig.lOF shows the
kinetics of the transformation of the 10-particles after treatment with 2.5 M NaCl at 25 "C. The same transformation occurs, although more slowly, when the incubation takes place in the absence of salt; this suggests that the salt is activating some process which takes place also in its absence, although at a reduced rate. A rise in the incubation temperature from 25 "C to 37 "C also increases the rate of the transformation. The change in the sedimentation velocity does not produce a noticeable change in the particles when examined with the electron microscope (results not shown) and the same proteins are present in 10-1 and
A. Camacho, F. Jimenez, J. de la Torre, J. L. Carrascosa, R. P. Mellado, C. Vasquez, E. Viiiuela, and M. Salas
A B C D E F G H
49
- NPI
- TPI
-HPO -HPI
-NP3
-NP2
-HP3 -p15
-p16
- P 7
Fig. 9. Autoradiograph of the proteins, separated by slab gel electrophoresis, present in the particles produced afer infection with mutants sus9(422)sus14(1241) or sus10(136)sus14(1241). The particles produced after infection of B.subtilis su- with mutants sus9(422)sus14(1241) or suslO( 136)sus14(1241), labeled with 14C-labeled protein hydrolysate, were purified by sucrose gradient centrifugation and subjected to slab gel electrophoresis as described in Materials and Methods. (A) 9-particles. (B) 9-1 particles. (C) 9-11 particles. (D) 10-particles. (E) 10'-particles after a second purification by sucrose gradient centrifugation. (F) 10-1 particles. (G) 10-11 particles. (H) Proteins induced in ultraviolet-ir- radiated bacteria infected with wild-type phage and labeled with ''C-labeled protein hydrolysate in a 20 -28-min pulse. Only the proteins present in the different types of particles are indicated
10-11 particles as in the original 10-particles when analyzed by slab gel electrophoresis (Fig. 9F and G).
Cistrons I I and 12
Genes 11 and 12 code, respectively, for the lower collar protein, NP3, and for the precursor of the neck appendages, P-NP1, (molecular weight 90000) ; the latter is cleaved before assembly to give protein NP1, the neck appendage protein (molecular weight 80000) and a low-molecular-weight fragment [5 - 8,15,22].
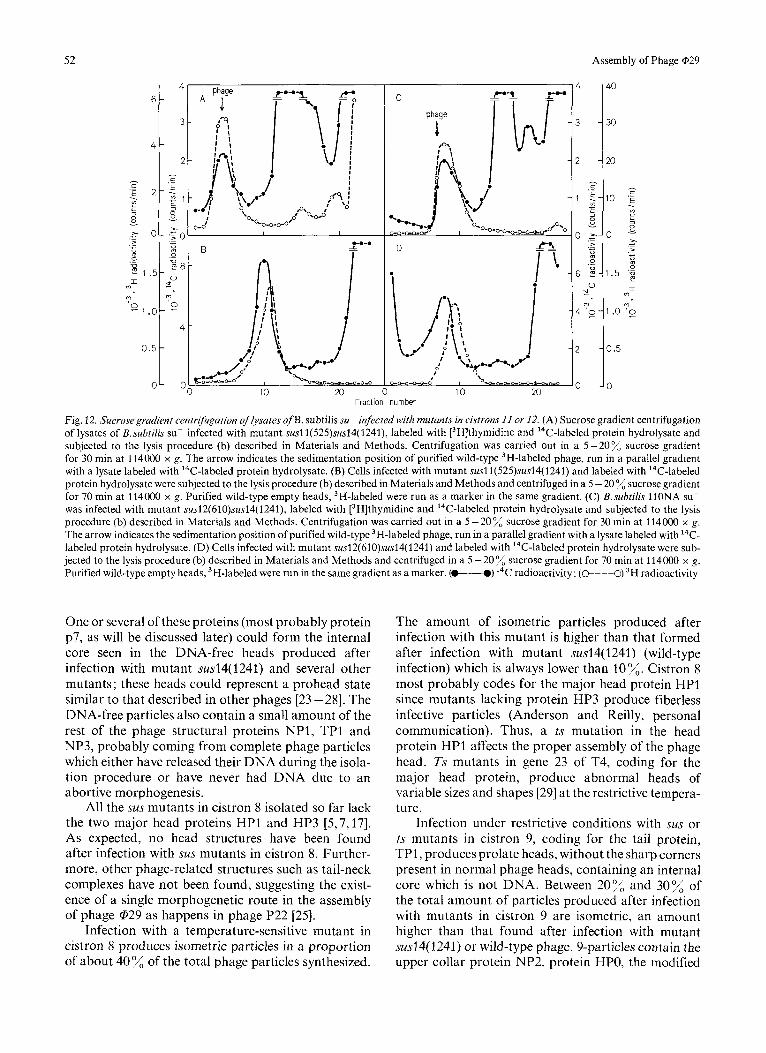
Sus or ts mutants in cistron 11 produce two types of structures, DNA-containing and DNA-free parti- cles, as shown in Fig.12A and B, respectively, for
mutant susl1(525)sus14(1241). However, the amount of DNA-containing particles is only 15 % relative to the total amount of particles produced. This is probably due to instability of the DNA in the particles as suggested from the ratio of rHlthymidine to 14C-labeled amino acid incorporation in the DNA- containing peak which is 2 compared with 7 in the corresponding peak in the cells infected with the control mutant sus14(1241) (Fig. 2A). The sedimenta- tion rate of the DNA-containing particles is similar to that of purified wild-type phage; the sedimentation coefficient of the DNA-free particles is 129 S, slightly higher than that of purified wild-type empty heads (Fig. 12B).
50
100
- 2 7 5 -
? 7 50 m
0,
25
Assembly of Phage @29
C F - 100 0
10-IT - 75 e _-__ -o------o
7 0
50 +
0
; 25 0-0
I I -;
(1 I I 0
*\* ,* \* 9-1 - .o---- --
,o.' 9 - I I
OX'
9-particles
1
15
10
5 .c . Y)
c 3 0
O ,I 15
\D m
N 0
10
5
0 10 20 30
Fractlon number
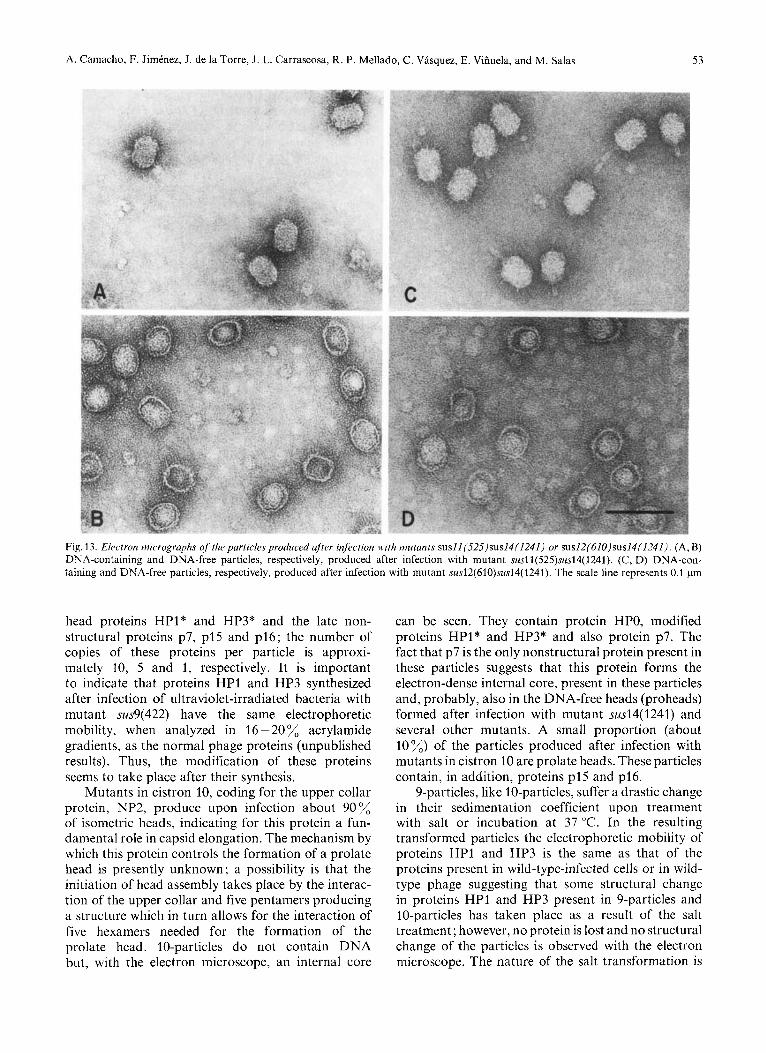
When the DNA-containing and DNA-free par- ticles produced after infection with mutant susl l(525)- susl4( 1241) were analyzed with the electron micro- scope the results shown in Fig.13 were obtained. DNA-containing particles consist of normal heads without neck appendages or tail (Fig. 13A). DNA-free particles consist of prolate heads, more rounded at the corners than the final normal heads, without neck appendages or tail. They contain an internal core (Fig. 13B), which is not DNA since no DNA is present in these particles (see Fig. 12A). When crude lysates are analyzed with the electron microscope, a total of about 85 % of prolate heads and 15 "/, isometric heads is produced after infection with mutant susl l(525)- susl4( 1241), both in the DNA-containing and in the DNA-free particles. When the peaks from the
sucrose gradients are analyzed, the same proportion of prolate and isometric heads is observed in the DNA-free particles. However, the amount of prolate and isometric heads is approximately 30 and 70%, respectively, in the DNA-containing particles, suggesting that either in the sucrose gradient centrif- ugation or in the preparation of the specimens for electron microscopy, the prolate DNA-containing heads are selectively lost. Similar results are obtained with mutants susl1(542)sus14(1241) or tsl 1(119)sus14 (1241).
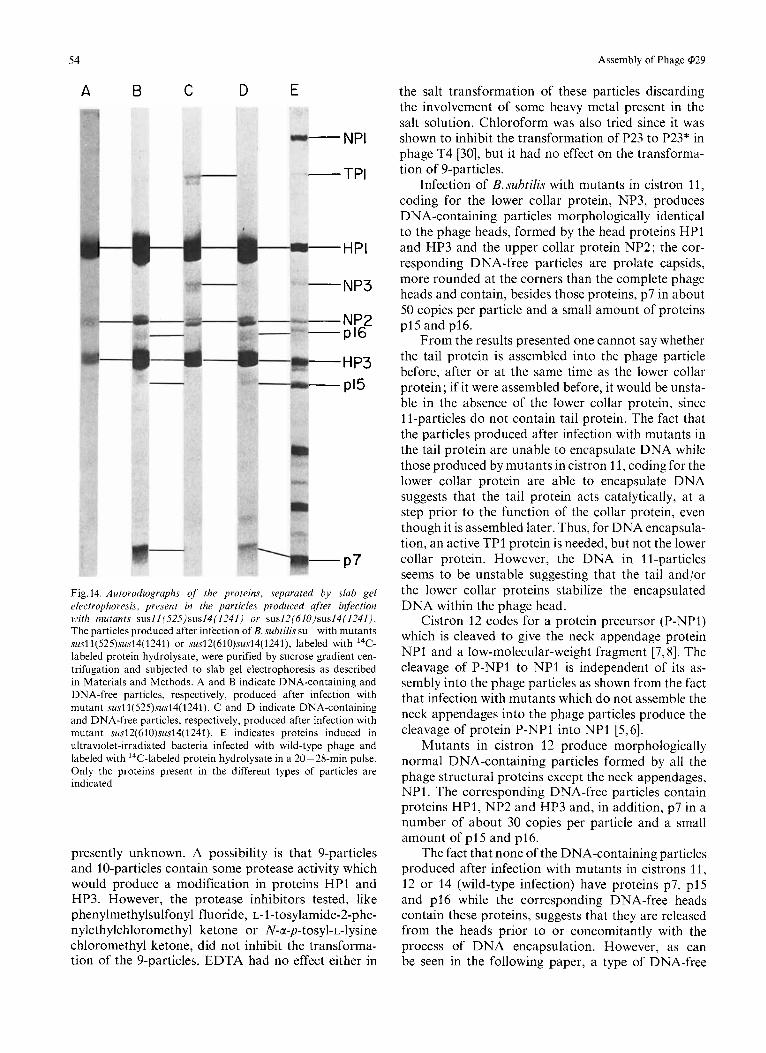
The proteins present in the two kinds of 1 1-particles are shown in Fig. 14A and B. In both cases, proteins HP1, NP2 and HP3 are present; protein HPO was not resolved in these gels. In addition, DNA-free particles contain protein p7, absent from the DNA-
A. Camacho, F. Jimtnez, J. de la Torre, J. L. Carrascosa, R. P. Mellado, C. Vasquez, E. Vifiuela, and M. Salas 51
qHP1 A I !
HPl
Fraction number
Fig. 11. Gel electrophoresis of 9-I, 9-II and I0-particles. 9-particles, produced after infection of B.suhtilis su- at 42 "C with mutant ts9(1 S), labeled with [35S]methionine, were purified by sucrose gradient centrifugation, treated with 2.5 M NaCl for 24 h at 25 "C and the 9-1 and 9-11 peaks isolated by sucrose gradient centrifugation in the presence of 2.5 M NaC1. Electrophoresis was carried out in tubes containing 16 - 20 %, acrylamide gradients as indicated in Materials and Methods. (A) Coelectrophoresis of 9-1 particles and the proteins induced in ultraviolet-irradiated bacteria infected with wild-type phage labeled with [3H]methionine in a 15 - 35-min pulse. (B) Coelectrophoresis of 9-11 particles and the proteins induced in ultraviolet-irradiated bacteria infected with wild-type phage labeled with [3H]methionine in a 15-35-min pulse. (C) 10-particles, produced after infection of B. suhtilis su- with mutant sus10(74), labeled with [35S]methionine, were purified by sucrose gradient centrifugation. The electrophoresis was run in the presence of proteins induced in ultraviolet-irradiated bacteria infected with wild-type phage and labeled with [3H]leucine in a 15-30-min pulse. (-0) 35S radioactivity; (0----0) 3H radioactivity
containing 1 l-particles, and a small amount of proteins p15 and p16. The number of copies of protein p7 is estimated to be about 50.
As in the case of the mutants in cistron 11, sus or ts mutants in cistron 12 produce DNA-con- taining and DNA-free particles, as shown in Fig. 12C for mutant sus12(610)sus14( 1241). The proportion of DNA-containing particles relative to the total amount
of particles produced is about 50% in the case of mutants susl2(61 O)susl4( 1241) and tsl2( 108)susl4- (1241). The DNA present in the particles produced by mutants in cistron 12 seems to be more stable than that present in the ll-particles; the ratio of [3H]- thymidine to 14C-labeled amino acid incorporation in the DNA-containing 12-particles is 5.5 (compared with 2 in the 1 l-particles), a value close to 7, obtained in the corresponding peak in the cells infected with mutant sus14(1241) (Fig. 2A). The sedimentation rate of the DNA-containing particles is approximately 240 S, slightly lower than that of complete phage, probably due to the lack of the neck appendages. The sedimentation rate of the DNA-free 12-particles is 129 S, higher than that of purified wild-type empty capsids (Fig. 12D).
When the DNA-containing and DNA-free 12- particles were analyzed with the electron microscope the results shown in Fig. 13C and D were obtained for mutant sus12(610)sus14(1241). DNA-containing 12- particles (Fig. 13C) look like normal phage with tail, except for the absence of the neck appendages. The DNA-free 12-particles (Fig. 13D) contain an internal core and are more rounded at the corners than complete phage heads. No isometric heads are seen in the DNA-containing particles, their proportion in the DNA-free heads is 25% of the total number of heads.
The proteins present in 12-particles are shown in Fig. 14C and D. DNA-containing 12-particles have all the structural proteins present in mature phage except the neck appendage protein NP1. DNA-free 12- particles contain the same proteins as the correspond- ing ll-particles, that is to say, HPI, NP2, HP3, p7 and a small amount of proteins p15 and p16. Protein HPO was not resolved in these gels. The number of molecules of protein p7 is calculated to be about 30.
DISCUSSION
Mutant susl4( 1241) behaves like wild-type phage expect that when it infects B. subtilis su- the lysis is delayed [5]. Like wild-type phage, mutant sus14(1241) produces DNA-containing, infective particles, and DNA-free particles with an internal core. The DNA- free particles have a sedimentation rate slightly higher than that of purified wild-type empty heads probably due to the presence of the internal core which is not present in the empty heads [3]. With the electron microscope the DNA-free particles look more rounded at the corners than purified empty heads [3 ] and con- tain an internal core which is not DNA. The DNA-free particles have, in addition to the phage structural proteins HP1, NP2 and HP3, proteins p7, p16 and some p15; the last proteins are not present in the final phage particles nor in any kind of DNA-containing particles isolated after infection with several mutants.
52 Assembly of Phage @29
Fraction number
Fig. 12. Sucrosegradient centrijiugation of Iysates o fB . subtilis su- infected with mutunts in cistrons 11 or 12. (A) Sucrose gradient centrifugation of lysates of B. subtilis su- infected with mutant susl1(525)sus14(1241), labeled with 13H]thymidine and 14C-labeled protein hydrolysate and subjected to the lysis procedure (b) described in Materials and Methods. Centrifugation was carried out in a 5-20% sucrose gradient for 30 min at 114000 x g. The arrow indicates the sedimentation position of purified wild-type 3H-labeled phage, run in a parallel gradient with a lysate labeled with 14C-labeled protein hydrolysate. (B) Cells infected with mutant susl1(525)sus14(1241) and labeled with 14C-labeled protein hydrolysate were subjected to the lysis procedure (b) described in Materials and Methods and centrifuged in a 5 - 20 % sucrose gradient for 70 min at 114000 x g. Purified wild-type empty heads, 3H-labeled were run as a marker in the same gradient. (C) B.subtilis 11ONA su- was infected with mutant sus12(610)sus14(1241), labeled with [3H]thymidine and I4C-labeled protein hydrolysate and subjected to the lysis procedure (b) described in Materials and Methods. Centrifugation was carried out in a 5-20% sucrose gradient for 30 min at 114OOO x g. The arrow indicates the sedimentation position of purified wild-type 3H-labeled phage, run in a parallel gradient with a lysate labeled with 14C- labeled protein hydrolysate. (D) Cells infected with mutant sus12(61O)sus14( 1241) and labeled with ''C-labeled protein hydrolysate were sub- jected to the lysis procedure (b) described in Materials and Methods and centrifuged in a 5 -20% sucrose gradient for 70 min at 114000 x g. Purified wild-type empty heads, 3H-labeled were run in the same gradient as a marker. (-) I4C radioactivity; (@---O) 3H radioactivity
One or several of these proteins (most probably protein p7, as will be discussed later) could form the internal core seen in the DNA-free heads produced after infection with mutant susl4( 1241) and several other mutants; these heads could represent a prohead state similar to that described in other phages [23 -281. The DNA-free particles also contain a small amount of the rest of the phage structural proteins NP1, TPI and NP3, probably coming from complete phage particles which either have released their DNA during the isola- tion procedure or have never had DNA due to an abortive morphogenesis.
All the sus mutants in cistron 8 isolated so far lack the two major head proteins HP1 and HP3 [5,7,17]. As expected, no head structures have been found after infection with sus mutants in cistron 8. Further- more, other phage-related structures such as tail-neck complexes have not been found, suggesting the exist- ence of a single morphogenetic route in the assembly of phage @29 as happens in phage P22 [25].
Infection with a temperature-sensitive mutant in cistron 8 produces isometric particles in a proportion of about 40% of the total phage particles synthesized.
The amount of isometric particles produced after infection with this mutant is higher than that formed after infection with mutant susl4( 1241) (wild-type infection) which is always lower than 10%. Cistron 8 most probably codes for the major head protein HP1 since mutants lacking protein HP3 produce fiberless infective particles (Anderson and Reilly, personal communication). Thus, a ts mutation in the head protein HP1 affects the proper assembly of the phage head. Ts mutants in gene 23 of T4, coding for the major head protein, produce abnormal heads of variable sizes and shapes [29] at the restrictive tempera- ture.
Infection under restrictive conditions with sus or ts mutants in cistron 9, coding for the tail protein, TP1, produces prolate heads, without the sharp corners present in normal phage heads, containing an internal core which is not DNA. Between 20% and 30% of the total amount of particles produced after infection with mutants in cistron 9 are isometric, an amount higher than that found after infection with mutant sus14(1241) or wild-type phage. 9-particles contain the upper collar protein NP2, protein HPO, the modified
A. Camacho, F. Jimtnez, J. de la Torre, J. L. Carrascosa, R. P. Mellado, C. Vasquez, E. Vifiuela, and M. Salas 53
Fig. 13. Electron micrographs of the particles produced after infection with mutants susll(525)sus14(1241) or sus12(610)sus14(1241). (A, B) DNA-containing and DNA-free particles, respectively, produced after infection with mutant susl1(525)sus14(1241). (C, D) DNA-con- taining and DNA-free particles, respectively, produced after infection with mutant sus12(610)sus14( 1241). The scale line represents 0.1 pm
head proteins HP1* and HP3* and the late non- structural proteins p7, p15 and p16; the number of copies of these proteins per particle is approxi- mately 10, 5 and 1, respectively. It is important to indicate that proteins HP1 and HP3 synthesized after infection of ultraviolet-irradiated bacteria with mutant sus9(422) have the same electrophoretic mobility, when analyzed in 16 - 20 % acrylamide gradients, as the normal phage proteins (unpublished results). Thus, the modification of these proteins seems to take place after their synthesis.
Mutants in cistron 10, coding for the upper collar protein, NP2, produce upon infection about 90% of isometric heads, indicating for this protein a fun- damental role in capsid elongation. The mechanism by which this protein controls the formation of a prolate head is presently unknown; a possibility is that the initiation of head assembly takes place by the interac- tion of the upper collar and five pentamers producing a structure which in turn allows for the interaction of five hexamers needed for the formation of the prolate head. 10-particles do not contain DNA but, with the electron microscope, an internal core
can be seen. They contain protein HPO, modified proteins HP1* and HP3* and also protein p7. The fact that p7 is the only nonstructural protein present in these particles suggests that this protein forms the electron-dense internal core, present in these particles and, probably, also in the DNA-free heads (proheads) formed after infection with mutant susl4( 1241) and several other mutants. A small proportion (about 10%) of the particles produced after infection with mutants in cistron 10 are prolate heads. These particles contain, in addition, proteins p15 and p16.
9-particles, like 10-particles, suffer a drastic change in their sedimentation coefficient upon treatment with salt or incubation at 37 "C. In the resulting transformed particles the electrophoretic mobility of proteins HP1 and HP3 is the same as that of the proteins present in wild-type-infected cells or in wild- type phage suggesting that some structural change in proteins HP1 and HP3 present in 9-particles and 10-particles has taken place as a result of the salt treatment; however, no protein is lost and no structural change of the particles is observed with the electron microscope. The nature of the salt transformation is
54 Assembly of Phage @29
A B C D E
- NPI
TPI -
--PI
-NP3
-NP2 --I6
-HP3 - pl5
-P?
Fig. 14. Autoradiographs of the proteins, separated by slab gel electrophoresis. present in the particles produced after infection with mutants susll(S2S)sus14(1241) or sus12(61O)sus14(124I). The particles produced after infection of B. subtilis su- with mutants susl1(525)sus14(1241) or sus12(610)sus14(1241), labeled with I4C- labeled protein hydrolysate, were purified by sucrose gradient cen- trifugation and subjected to slab gel electrophoresis as described in Materials and Methods. A and B indicate DNA-containing and DNA-free particles, respectively, produced after infection with mutant susl1(525)sus14(1241). C and D indicate DNA-containing and DNA-free particles, respectively, produced after infection with mutant sus12(61O)sus14(1241). E indicates proteins induced in ultraviolet-irradiated bacteria infected with wild-type phage and labeled with 14C-labeled protein hydrolysate in a 20- 28-min pulse. Only the proteins present in the different types of particles are indicated
presently unknown. A possibility is that 9-particles and 10-particles contain some protease activity which would produce a modification in proteins HP1 and HP3. However, the protease inhibitors tested, like phenylmethylsulfonyl fluoride, L-1-tosylamide-2-phe- nylethylchloromethyl ketone or N-a-p-tosyl-L-lysine chloromethyl ketone, did not inhibit the transforma- tion of the 9-particles. EDTA had no effect either in
the salt transformation of these particles discarding the involvement of some heavy metal present in the salt solution. Chloroform was also tried since it was shown to inhibit the transformation of P23 to P23* in phage T4 [30], but it had no effect on the transforma- tion of 9-particles.
Infection of B.subtilis with mutants in cistron 11, coding for the lower collar protein, NP3, produces DNA-containing particles morphologically identical to the phage heads, formed by the head proteins HPI and HP3 and the upper collar protein NP2; the cor- responding DNA-free particles are prolate capsids, more rounded at the corners than the complete phage heads and contain, besides those proteins, p7 in about 50 copies per particle and a small amount of proteins p15 and p16.
From the results presented one cannot say whether the tail protein is assembled into the phage particle before, after or at the same time as the lower collar protein; if it were assembled before, it would be unsta- ble in the absence of the lower collar protein, since 1 I-particles do not contain tail protein. The fact that the particles produced after infection with mutants in the tail protein are unable to encapsulate DNA while those produced by mutants in cistron 11, coding for the lower collar protein are able to encapsulate DNA suggests that the tail protein acts catalytically, at a step prior to the function of the collar protein, even though it is assembled later. Thus, for DNA encapsula- tion, an active TP1 protein is needed, but not the lower collar protein. However, the DNA in 11-particles seems to be unstable suggesting that the tail and/or the lower collar proteins stabilize the encapsulated DNA within the phage head.
Cistron 12 codes for a protein precursor (P-NPI) which is cleaved to give the neck appendage protein NP1 and a low-molecular-weight fragment [7,8]. The cleavage of P-NP1 to NP1 is independent of its as- sembly into the phage particles as shown from the fact that infection with mutants which do not assemble the neck appendages into the phage particles produce the cleavage of protein P-NP1 into NP1 [5,6].
Mutants in cistron 12 produce morphologically normal DNA-containing particles formed by all the phage structural proteins except the neck appendages, NP1 . The corresponding DNA-free particles contain proteins HP1, NP2 and HP3 and, in addition, p7 in a number of about 30 copies per particle and a small amount of p15 and p16.
The fact that none of the DNA-containing particles produced after infection with mutants in cistrons 11, 12 or 14 (wild-type infection) have proteins p7, p15 and p16 while the corresponding DNA-free heads contain these proteins, suggests that they are released from the heads prior to or concomitantly with the process of DNA encapsulation. However, as can be seen in the following paper, a type of DNA-free

A. Camacho, F. Jimenez, J . de la Torre, J. L. Carrascosa, R . P. Mellado, C. Visquez, E. Viiiuela, and M. Salas 55
heads with an internal core is accumulated after infection with mutants in cistron 16, which contains proteins HPO, HP1, NP2, HP3 and p7 in about 90 copies per particle, but not p15 or p16. This result suggests that the assembly of proteins p15 and p16 takes place on a p7-containing prohead. Since the number of copies of protein p7 in the prolate heads produced after infection with mutants in cistrons 9, 11, 12 and 14 is smaller than that found in the cistron 16 heads one cannot tell at present whether or not protein p7 is released when p15 and p16 are assembled, prior to or concomitant with DNA encapsulation. The heads found after infection with mutants in the above cistrons could represent heads in the process of DNA encapsulation, as will be described in the following paper.
The fact that the only protein missing in 12-particles is NP1 indicates that the neck appendages are the last structural component to be assembled into the phage particles. A similar conclusion has been reached by complementation in vitro [6] (and unpublished results).
The presented results suggest that there is a single morphogenetic route for the assembly of @29 structural proteins, similar to that which happens in phage P22 [25] and unlike the case of the T4 and /z phages which have different morphogenetic pathways for the as- sembly of the head and the tail proteins [31,32]. The role of other nonstructural proteins on the morpho- genesis of the phage particles as well as a pathway for @29 assembly are described in the following paper.
During the course of this research we have shared mutants and information with Drs D. L. Anderson and B. E. Reilly. Our view of @29 assembly results in part from this exchange. This investigation has been aided by Grants from Comisidn Asesora paru el Desarrollo dr Iu Investigacidn Cientifica, Coniisidn Administradora del Descuento Cotnplemmtario ( I . N. P.) and Direccibn General de Sanidad. A.C., F.J., R.P.M., and J.L.C. were Fellows of Fondo Nucional pura lu Forrnucidn de Personal Investigador.
REFERENCES
1. Anderson, D. L., Hickman, I>. D. & Reilly, B. E. (1966) J . Bat,-
2. Ortin, J., Viiiuela, E., Salas, M. & Vasquez, C. (1971) Nat. New teriol. 91, 2081 -2089.
Biol. 234, 275 - 277.
3.
4.
5.
6.
7. 8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19. 20. 21.
22.
23. 24.
25.
26.
27.
28. 29.
30. 31.
32.
Mendez, E., Ramirez, G., Salas, M. & Viiiuela, E. (1971) Virology, 45, 567 - 576.
Vifiuela, E., Carnacho, A, , Jimenez, F., Carrascosa, J. L., Ra- mirez, G. & Salas, M. (1976) Philos. Trans. R. Soc. Lond. Ser. B, Biol. Sci. 276, 29- 35.
Carrascosa, J. L., Camacho, A,, Moreno, F., Jimenez, F., Mel- lado, R. P., Viiiuela, E. & Salas, M. (1976). Eur. J . Bioc,/iem. 66,229 - 241.
Tosi, M. E., Reilly, B. E. &Anderson, D. L. (1975)J. Virol. 16,
Anderson, D. L. & Reilly, B. E. (1974) J . Virol. 13, 211 -221. Carrascosa, J. L., Camacho, A., Viiiuela, E. & Salas, M. (1974)
Hawley, L.A., Reilly, B. E., Hagen, E. H. & Anderson. D. L.
McGuire, J. C., Pene, J. J . & Barrow-Carraway, J. (1974)
Talavera, A,, Jimenez, F., Salas, M. & Viiiuela, E. (1971)
Reilly, B. E., Zeece, V. M. & Anderson, D. L. (1973) J . Virol.
Moreno, F., Camacho, A, , Vifiuela, E. & Salas, M. (1974) Viro-
Mellado, R. P., Moreno, F., Viiiuela, E., Salas, M., Reilly, B. E.
Reilly, B. E., Tosi, M . E. &Anderson, D. L. (1975) J . Virol. 16,
Carrascosa, J . L., Viiiuela, E. & Salas, M. (1973) Virology, 56,
Mellado, R . P.. Vitiucla. h . & Salas. M . (1976) Eur. J . Bio-
Rubio, V., Salas, M., Viiiuela, E., Usobiaga, P., Saiz, J. L. &
Laemmli, U. K. (1970) Nature (Lond.) 227, 680-685. Maizel, J. V., J r (1971) Methods Virol. 5 , 179-246. Kellenberger, E., Eiserling, F. A. & Boy de la Tour, E. (1968)
Camacho, A,, Moreno, F., Carrascosa, J. L., Viiiuela, E. &
Laemmli, U. K. & Favre, M. (1973) J . Mol. B id . 80, 575-599. Bijlenga, R. K. L., Scraba, D. & Kellenberger, E. (1973) Viro-
King, J., Lenk, E. V. & Botstein, D. (1973) J . Mol. Bid. RU,
Hohn, B. & Hohn, T. (1974) Proc. Natl Acad. Sci. U.S.A. 71,
Kaiser, D., Syvanen, M. & Masuda, T. (1975) J . Mol. Bid . Y l ,
Matsuo-Kato, H. & Fujisawa, H. (1975) Virology, 63,105 - 114. Laemmli, U. K., Molbert, E., Showe, M. & Kellenberger, E.
Laemmli, U. K.&Quittner, S. F.(1974) Virology,62,483-499. Wood, W. B., Edgar, R. S., King, J., Lielausis, A. & Hennin-
Weigle, J. (1966) Proc. Nut1 Acad. Sci. U . S . A . 55, 1462- 1466.
1282-1295.
FEBS Lett. 44, 317-321.
(1973)J. Virol. 12, 1349-1159.
J . Virol. 13, 690-698.
Virology, 76. 586- 595.
I I , 156 - 760.
logy, 62, 1 - 16.
& Anderson, D. L. (1976) J . Virol. 19, 495 -500.
1010-1016.
291 - 299.
chem.65,213-223.
Llopis, J. F. (1974) Virology, 57, 112-121.
J . Ultrastruct. Res. 21, 335 - 360.
Salas, M. (1974) Eur. J . Biochem. 47, 199-205.
logy, 56,250 - 267.
697 -731.
2372 - 2376.
175-186.
(1970) J . Mol. Biol. 49, 99-113.
ger, M. (1968) Fed. Proc. 27, 1160-1166.
A. Camacho, F. Jimtnez, J. de la Torre, J . L. Carrascosa, R. P. Mellado, E. Viiiuela, and M. Salas, Centro de Biologia Molecular, Centro de Investigaciones Biologicas, C.S.I.C. and U.A.M., Velazquez 144, Madrid-6, Spain
C . Vasquez, Centro de Virologia Animal, (CONICET-FECIC) Serrano 661, Buenos Aires, Argentina















![Lamda phage[1]](https://img.pdfslide.tips/doc/110x75/58cedaba1a28abd4098b6283/lamda-phage1.jpg)












