Embed Size (px)
Citation preview

AVALIAÇÃO LEUCOCITÁRIA, RELAÇÃO ALBUMINA/GLOBULINA, PROTEÍNA PLASMÁTICA E
FIBRINOGÊNIO DE BOVINOS DA RAÇA NELORE, CONFINADOS E TERMINADOS A PASTO
Ediane Batista da Silva Orientador: Prof. Dr. Dirson Vieira
GOIÂNIA 2006
UNIVERSIDADE FEDERAL DE GOIÁS ESCOLA DE VETERINÁRIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA ANIMAL

EDIANE BATISTA DA SILVA
AVALIAÇÃO LEUCOCITÁRIA, RELAÇÃO ALBUMINA/GLOBULINA, PROTEÍNA PLASMÁTICA E
FIBRINOGÊNIO DE BOVINOS DA RAÇA NELORE, CONFINADOS E TERMINADOS A PASTO
Dissertação apresentada para obtenção do grau de Mestre em Ciência Animal junto à Escola de Veterinária da Universidade Federal de Goiás
Área de concentração: Patologia, Clínica e Cirurgia Animal
Orientador: Prof. Dr. Dirson Vieira - EV/UFG
Comitê de Orientação:
Prof. Dr. Luiz Antônio Franco da Silva - EV/UFG
Prof. Dr. Eugênio Gonçalves de Araújo - EV/UFG
GOIÂNIA 2006

ii
Ao vovô Dete e vovó Vani que sempre
foram fonte de amor, carinho,
dedicação e por serem minha
referência de caráter e humanidade,
com muito amor.
DEDICO

iii
AGRADECIMENTOS
À Universidade Federal de Goiás, à Escola de Veterinária e à Capes
por disponibilizarem suas instalações para a realização desse trabalho e pela
concessão de auxílio financeiro.
Ao professor Dirson Vieira pela orientação e confiança durante esse
período.
Aos professores Olízio Claudino da Silva, Afonso Henrique Miranda,
Paulo Henrique Jorge da Cunha, Adilson Donizeti Damasceno, Eugênio
Gonçalves de Araújo, Valéria de Sá Jaime, Regiani G. Porto, Ana Paula I. Santin,
Ana Paula Junqueira Kipnis e em especial à Professora Maria Clorinda Soares
Fioravanti pelo apoio e constantes esclarecimentos prestados durante as várias
etapas desse projeto.
Ao professor Luiz Antônio Franco da Silva, pelo constante incentivo
durante toda minha vida acadêmica, por depositar confiança em minha
capacidade de execução e principalmente por me ofertar apoio e amizade durante
a nossa longa convivência.
Aos amigos Alline de Paula Reis, Glauciane Ribeiro de castro, Karyne
Oliveira Coelho, Renata Pereira e Bruno Rodrigues Trindade pelo auxílio e divisão
de conhecimentos. Aos colegas de mestrado e doutorado Marcele, Priscila,
Renata, Ana Paula, Luciana, Cleo e Lorena pelo convívio agradável e por dividir
momentos de angústias e alegrias.
Aos amigos e futuros colegas Maria Ivete de Moura, Marco Augusto
Machado e ao médico veterinário Vinícius Rodrigues Sousa pela disponibilidade
em ajudar sempre que possível.
Aos funcionários do Laboratório Clínico Elton e Maria pela paciência e
auxílio inestimável durante a execução desse trabalho.
À empresa frigorífica FRIBOI e seus funcionários, especialmente Éder
e Eliete por auxiliarem na coleta do material desse projeto.
À Marina pelo auxílio incalculável durante as coletas e Sabrina pelo
auxílio na coloração das lâminas e Andréia pelo apoio moral.

iv
À Liliana, minha amiga de todas as horas, pela amizade e atenção
concedida ainda durante a graduação, dias atuais e espero que para toda nossa
vida.
Aos eternos amigos da casa de estudante universitários (CEU II) por
fazerem parte da minha jornada acadêmica e familiar, especialmente a Luciana,
Wagna, Bethânia, Talismã, Raquel, Mara, Ana Maria, Hércules, Marlene,
Eduardo, Verônica e Elcimar.
Aos meus pais Leni e Juvenil e irmãos Marcos, Fábio e Elizângela, por
compreenderem minha ausência e mesmo distante torcerem pelo meu sucesso.
Finalmente, aos meus avós Oldete e Vanilde, tias Ângela, Irene e
Irleide pela dedicação, amor e carinho.

v
SUMÁRIO
1 INTRODUÇÃO ..................................................................................................... 1 2 REVISÃO BIBLIOGRÁFICA ................................................................................ 2
2.1 Resposta leucocitária em bovinos ............................................................. 2 2.3 Mecanismo de ação dos leucócitos ........................................................... 4 2.4 Principais fatores que influenciam o leucograma .................................... 8
2.4.1 Excitação e estresse ............................................................................. 9 2.4.2 Ambiente .............................................................................................. 11 2.4.3 Comportamento dos leucócitos nas doenças respiratórias ........... 13
2.5 Interpretação dos parâmetros leucocitários ........................................... 17 2.5.1 Contagem total de leucócitos ............................................................ 17 2.5.2 Contagem diferencial de leucócitos .................................................. 19
2.6 Constituintes sérico/plasmático ............................................................... 21 2.6.1 Proteína total ....................................................................................... 21 2.6.2 Albumina .............................................................................................. 23 2.6.3 Globulina .............................................................................................. 24 2.6.4 Fibrinogênio ........................................................................................ 25
3 OBJETIVOS ....................................................................................................... 28 3.1 Objetivo geral ............................................................................................. 28 3.2 Objetivos específicos ................................................................................ 28
4 MATERIAL E MÉTODOS .................................................................................. 29 4.1 Local de realização do estudo .................................................................. 29 4.2 Animais ....................................................................................................... 29 4.3 Colheita das amostras ............................................................................... 30 4.4 Exames laboratoriais ................................................................................. 31 4.5 Análise estatística ..................................................................................... 32
5 RESULTADOS E DISCUSSÃO ......................................................................... 33 5.1 Leucograma ............................................................................................... 33 5.2 Proteína total .............................................................................................. 39 5.3 Albumina .................................................................................................... 40 5.5 Fibrinogênio ............................................................................................... 44
CONCLUSÃO ....................................................................................................... 47 REFERÊNCIAS BIBLIOGRÁFICAS ..................................................................... 48 ANEXOS ............................................................................................................... 63

vi
LISTA DE FIGURAS
Figura 1 Mecanismo de rolagem, aderência e quimiotaxia de leucócitos, bem
como a ação das selectinas e integrinas..........................................05
Figura 2 Animais submetidos ao sistema de confinamento (A), as instalações
e o piso de terra (B)..........................................................................15
Figura 3 Animais no curral de espera para abate, em frigorífico - Goiânia,
novembro de 2005............................................................................30
Figura 4 Colheita de sangue na linha de abate - Goiânia, novembro de
2005..................................................................................................31
Figura 5 Média dos valores de leucócitos totais (/μL) de bovinos confinados e
a pasto - Goiânia, julho a novembro de 2005...................................34
Figura 6 Média dos valores de linfócitos (/μL) de bovinos confinados e a
pasto - Goiânia, julho a novembro de 2005......................................35
Figura 7 Média dos valores de neutrófilos segmentados (/μL) de bovinos
confinados e a pasto - Goiânia, julho a novembro de 2005.............36
Figura 8 Média dos valores de neutrófilos segmentados (/μL) de bovinos
confinados e a pasto - Goiânia, julho a novembro de 2005.............36
Figura 9 Média dos valores de eosinófilos (/μL) de bovinos confinados e a
pasto - Goiânia, julho a novembro de 2005.....................................37
Figura 10 Média dos valores de monócitos (/μL) de bovinos confinados e a
pasto - Goiânia, julho a novembro de 2005.....................................38
Figura 11 Média dos valores de proteína total (g/dL) de bovinos confinados e
a pasto - Goiânia, julho a novembro de 2005..................................39
Figura 12 Média dos valores de albumina (g/dL) de bovinos confinados e a
pasto - Goiânia, julho a novembro de 2005.....................................41
Figura 13 Média dos valores da concentração de globulina (g/dL) de bovinos
confinados e a pasto - Goiânia, julho a novembro de 2005.............42
Figura 14 Média da relação Albumina/Globulina (A/G) de bovinos confinados e
a pasto - Goiânia, julho a novembro de 2005...................................43
Figura 15 Média dos valores de fibrinogênio (mg/dL) de bovinos confinados e a
pasto - Goiânia, julho a novembro de 2005..................................... 44

vii
FIGURA 16 Média dos valores da relação Proteína Plasmática Fibrinogênio
(PP:F) de bovinos confinados e a pasto - Goiânia, julho a novembro
de 2005...........................................................................................45

viii
LISTA DE ABREVIATURAS
ACTH Hormônio adrenocorticotrófico
BVDV Vírus da diarréia viral bovina
CD62L Lectina, molécula de adesão de leucócitos
dL Decilitro
g Gramas
H2O2 Peróxido de hidrogênio
IgE Imunoglobulina E
IL Interleucina
LAM-1 Lectina, molécula de adesão de leucócitos
LFA-1 Molécula de adesão celular, inicilmente definidas como anticorpos
monoclonais
L-selectina Molécula de adesão da família selectina encontrada nos linfócitos
Mac-1 Integrina de leucócitos
mL Mililitros
mm3 Milímetro cúbico
MPO Mieloperoxidase
µL Microlitros
μm Micrômetros
ρ Nível de significância
p150, Proteína 150
p95 Proteína 95
pH Potencial hidrogeniônico
PMNs Polimorfonucleares

ix
RESUMO
Bovinos manejados em confinamentos geralmente estão expostos a diferentes situações que podem influenciar negativamente o desempenho dos animais, tais como maior concentração de animais por área, poeira, acúmulo de dejetos e excesso de manipulação. Portanto, é possível que esses fatores criem condições favoráveis à manifestação de diversos problemas sanitários, mesmo subclínicos, que poderão refletir no leucograma e constituintes séricos do organismo do animal. Esse trabalho objetivou avaliar as alterações no leucograma e algumas proteínas séricas de bovinos confinados e manejados à pasto. Utilizou-se 120 amostras sanguíneas, sendo que 60 foram obtidas de bovinos da raça Nelore, macho, confinados e 60 de animais com as mesmas características, porém terminados a pasto. As amostras foram colhidas em frigorífico, no momento do abate, aproveitando a sangria. Os parâmetros estudados foram o leucograma, relação albumina/globulina (A/G) sérica e concentração de fibrinogênio plasmático. Na análise dos dados empregou-se estatística descritiva, obtendo-se as médias, desvio padrão e coeficiente de variação para todos as variáveis avaliadas. As comparações entre médias foram feitas por meio de teste não-paramétrico. A contagem total de leucócitos/μL foi de 7640 ± 2.150 nos bovinos confinados e 7.725 ± 1.843 nos bovinos terminados a pasto. Não houve diferença significativa entre essa variável e a contagem diferencial dos leucócitos. Na avaliação da proteína sérica total em g/dL, houve maior concentração nos bovinos terminados a pasto (6,10 ± 0,53) em comparação com os confinados (5,96 ± 0,49), no entanto essa diferença não foi significativa. Quanto à concentração de globulina nos animais a pasto foi de 3,29 ± 0,76 (ρ<0,05). Esta variável nos bovinos confinados apresentou uma concentração média de 2,99 ± 0,60. No entanto, nessa modalidade de manejo, a concentração sérica de albumina em g/dL foi maior (3,01 ± 0,43) em relação aos bovinos manejados a pasto 2,82 ± 0,45 (ρ<0,05). O índice A/G foi maior para os animais confinados (1,07 ± 8,91) do que os terminados a pasto (0,95 ± 0,38). O fibrinogênio em mg/dL foi mais elevado no animal terminado a pasto (872 ± 610) do que nos submetidos ao confinamento 633 ± 319. Quando se comparou os resultados do fibrinogênio com o do leucograma, a proteína foi mais reativa do que os leucócitos sugerindo que esse componente sangüíneo seja sempre avaliado e em associação com o leucograma. Apesar das adversidades que os bovinos confinados são submetidos, não se identificou alterações sugestivas de reações inflamatórias, tais como elevada contagem de leucócitos e fibrinogênio. A concentração média sérica de albumina mais elevada nos animais confinados, sugerem dieta nutricional mais adequada em comparação com os terminados a pasto. A concentração de globulina dos animais terminados a pasto foram maiores do que os confinados, podendo ser indicativo de maior desafio imunológico sofrido por esse grupo de animais. O nível médio do fibrinogênio plasmático, de bovinos terminados a pasto foi mais elevado do que os animais confinados. Palavras-chave: Confinamento, bovinos, leucócitos, proteínas plasmáticas

x
ABSTRACT Cattle managed in feedlot are generally exposed to different situations that can negatively influence their performance, such as increased concentration of animals, dust, accumulation of dejections and excess manipulation. Therefore, it is possible that those factors generate suitable conditions to the onset of several sanitary problems, even subclinical, that could reflect in the white blood cell count and serum parameters of the animal organism. This work aimed to evaluate the changes in the white blood cell count and some serum proteins of confined or grazing cattle. Of the 120 blood samples harvested, 60 were obtained from confined Nelore male bovines and 60 from animals with the same characteristics but managed extensively. The samples were obtained at the moment of slaughter. The parameters studied were the white blood cell count, serum albumin/globulin ratio and concentration of plasma fibrinogen. In the analysis of the data descriptive statistics was used, and the averages, standard deviation and coefficient of variation calculated for all parameters evaluated. The comparisons between averages were made by non-parametric test. The total counting of white blood cells/ml was 7640 ± 2,150 in confined bovines and 7,725 ± 1,843 in grazing cattle. There was no significant difference between this variable and differential white blood cell count. Regarding the evaluation of the total serum protein in g/dL, grazing bovines (6,10 ± 0,53) had greater concentration than confined ones (5,96 ± 0,49); however, this difference was not significant. The concentration of globulin in the grazing animals was 3,29 ± 0,76 (r<0,05). while confined bovines presented an average concentration of 2,99 ± 0,60. However, in the latter type of management, serum albumin concentration in g/dL was greater (3.01 ± 0,43) when compared to the grazing bovines 2,82 ± 0,45 (r<0,05). Albumin/globulin ratio was greater in confined animals (1,07 ± 8.91) than grazing ones (0,95 ± 0,38). Fibrinogen levels (mg/dL) were greater in grazing animals (872 ± 610) than in confined ones (633 ± 319). When fibrinogen results were compared to the white blood cell count, the protein was more sensitive, suggesting that fibrinogen levels should always be evaluated in association with the white blood cell count. Despite the adversities that confined bovines are subjected, suggestive alterations of inflammatory reactions were not identified. The more elevated serum albumin concentration recorded in the confined animals suggests they were subjected to a more adequate nutritional diet as compared to grazing cattle. Globulin concentration of grazing animals was greater than observed in confined ones, pointing to the fact that the latter were exposed to greater immunological challenge. Plasma fibrinogen levels were greater in grazing bovines when compared to confined animals. Key-words: Bovine, feedlot, leukocytes, plasma proteins

1 INTRODUÇÃO
A sanidade bovina é um tema de grande importância na atualidade.
Questões como a rastreabilidade, alimentos orgânicos e a segurança alimentar
passam a interessar a todos, desde o produtor e principalmente o consumidor.
Existem no Brasil, basicamente dois tipos de manejo de bovinos, destinados à
produção de carne, os bovinos terminados a pasto e os terminados em
confinamento.
De maneira geral, os principais problemas sanitários observados em
confinamentos no Brasil estão relacionados com os próprios animais, com
alimentos e água de bebida e com o ambiente. Durante o manejo, ocorre
situações de estresse causado pelo transporte, vacinação, desverminação,
mudanças na alimentação, hierarquia e aglomeração entre os animais, que cria
condições favoráveis à manifestação de diversos problemas sanitários e dentre
esses, os mais observados são os pulmonares.
Bovinos terminados a pasto, sem assistência médica veterinária
adequada, tem potencial em desenvolver enfermidades como febre aftosa,
brucelose, sofrerem ataques de enteroparasitas, além de parasitas externos como
moscas e carrapatos. Essas enfermidades nem sempre são notadas durante a
terminação dos animais. E se tratando dos parasitas, eles podem causar
depleção do sistema imune do animal, levando-os a apresentarem doenças, que
em condições normais, o organismo do animal controlaria.
Neste contexto, dependendo da agressão que os bovinos confinados e
não confinados são submetidos, o organismo do animal responderá de forma
aguda ou crônica, podendo, o processo inflamatório ser avaliado pela
quantificação dos leucócitos da corrente sangüínea, das proteínas totais,
albumina, globulinas e proteínas de fase aguda, como o fibrinogênio. Portanto, o
estudo dos constituintes sanguíneos fornece dados valiosos, podendo ser
utilizados na avaliação de estado nutricional e das alterações patológicas das
diversas enfermidades.

2
2 REVISÃO BIBLIOGRÁFICA 2.1 Resposta leucocitária em bovinos
As avaliações hematológicas nos bovinos são empregadas para
analisar doenças em um animal ou para avaliar grupos de animais, dentro de um
rebanho e ainda para detectar doenças ocultas e direcionar as condutas clínicas
(WEISS & PERMAN, 1992).
A resposta leucocitária em ruminantes freqüentemente difere das
observadas em outras espécies (TAYLOR, 2000; COLE et al., 1997). KANTEK &
NAVARRO (2005) atribuem a diferença quanto à resposta leucocitária dos
bovinos, ao fato desses animais possuírem um compartimento de reserva
leucocitária na medula óssea, limitado, fazendo com que os neutrófilos imaturos
apareçam rapidamente, nos estados inflamatórios agudos importantes, tais como
na mastite, na peritonite e na pericardite traumática. TAYLOR (2000) acrescenta
que a diferença da resposta leucocitára dificilmente constitui em mudança
hematológica específica para uma doença particular, mas são sugestivos de
considerações diagnósticas, como na septicemia ou endotoxemia.
TENNANT et al. (1975) estudaram a resposta leucocitária de bovinos,
apresentando septicemia e endotoxemia induzida. A leucopenia foi o achado
característico da endotoxemia. Segundo WEISS & PERMAN, (1992) e COLE et
al. (1997) a septicemia bacteriana é freqüentemente caracterizada por
neutropenia na maioria das espécies, entretanto, os ruminantes, especialmente os
bovinos, possuem tendência em desenvolver um quadro de neutrofilia nessas
situações. Esse evento é denominado por JAIN (1993) de neutrofilia de reboque,
pois há desvio de neutrófilos do “pool” marginal para a circulação, acelerando a
liberação de células do compartimento de reserva da medula óssea. Nas
condições inflamatórias menos severas, a neutropenia absoluta pode não ser
evidente. Nesses casos, a contagem de neutrófilos está diminuída, devido ao
efeito de glicocorticóides, sendo a redução da razão neutrófilo:linfócitos a
alteração notada no leucograma (WEISS & PERMAN, 1992; COLE et al., 1997)

3
Já nas doenças inflamatórias agudas dos bovinos, segundo JAIN
(1993), a contagem de leucócitos no estágio inicial, não reflete a severidade do
problema, isso porque, o número de linfócitos excede os neutrófilos nos animais
saudáveis e diminui em situações de estresse, devido à liberação de
corticosteróides endógenos na fase aguda das doenças. Nesses processos, os
neutrófilos maduros que se encontra na medula óssea, entram na circulação e a
resposta neutrofílica ao corticosteróide é marcada pela migração de neutrófilos
para o local da lesão. Para suprir a rápida demanda estabelecida, a medula óssea
lança os neutrófilos maduros, havendo uma depleção dessas células e
conseqüentemente, os neutrófilos imaturos entram na circulação, durante as
primeiras 24 a 48 horas, resultando em desvio de neutrófilos para a esquerda, do
tipo degenerativo. Após esse período, em média três a quatro dias, o desvio para
a esquerda desaparece, pois já há maturação de neutrófilos e também aumento
dos monócitos, marcando um quadro de leucocitose.
Nas inflamações superagudas ou nas septicemias, os leucócitos tóxicos
geralmente acompanham a resposta hematológica nos ruminantes. A basofilia
citoplasmática acomete mais freqüentemente essas espécies porque o citoplasma
eosinofílico é característico de neutrófilo saudável. As vacuolizações
citoplasmáticas, grânulos azurofílicos, corpúsculo de Döhle ou desnaturação
nuclear assemelham-se às mudanças induzidas em outras espécies, sob
condições similares (TAYLOR, 2000).
Nas doenças supurativas crônicas, os neutrófilos circulantes tendem a
apresentar-se maduros, sendo que, nos processos extensivos anteriores ao
estabelecimento da cronicidade, pode-se encontrar contagens elevadas. É
comum verificar contagens de neutrófilos de 20 a 40.000/µL, em lesões
supurativas, associada a microrganismos de baixa virulência (WEISS & PERMAN,
1992). Segundo MATOS & MATOS (1988) e COLE et al. (1997), nos ruminantes
na fase inicial dos processos infecciosos, observa-se um número global de
leucócitos dentro dos limites normais, acompanhado de neutrofilia, sendo que
muitas vezes são acompanhados de neutrófilos jovens. Os autores ressaltam que
nessas espécies, durante um processo infeccioso, a leucocitose não é observada,
devido à influência do estresse sobre a córtex da supra renal, a qual determina a

4
diminuição dos eosinófilos e linfócitos da corrente sanguínea, enquanto que
grande número dos neutrófilos migram para o local da infecção.
Devido à razão neutrófilo:linfócito ser baixa, aproximadamente 0,5,
comparada com outras espécies (3,5 em cães e 1,8 em gatos) e a reserva de
granulócitos maduros na medula óssea dos ruminantes ser limitada, contagens de
leucócitos de 20 a 30 x 103/µL são significantes nos bovinos, comparado com
contagem de 50 a 100 x 103/µL em situações similares em cães (JAIN, 1993).
Devido a vasta variabilidade da resposta leucocitária a doenças nos
bovinos, a avaliação de proteínas de fase aguda oferece melhor parâmetro de
interpretação do que a contagem de leucócitos (SUTHERLAND & WHITNEY,
1995; ECKERSALL & CONNER, 1988). Dentre as proteínas de fase aguda, o
fibrinogênio e a haptoglobina são indicadores mais sensíveis de inflamações
agudas ou crônicas em ruminantes (WEISS & PERMAN, 1992;
VERKKOJULKAISUT, 2000). Geralmente, uma alteração na contagem diferencial
de leucócitos, associada ao número total de leucócitos normal, pode caracterizar
um quadro de resposta inflamatória (JAIN, 1993).
2.3 Mecanismo de ação dos leucócitos
Os leucócitos circulantes podem migrar do vaso para os tecidos em
condições normais ou em circunstâncias patológicas. A migração dos leucócitos
da vasculatura ocorre devido a múltiplos fatores, que são desencadeados pela
ativação seqüencial de proteínas adesivas e seus ligantes nas células endoteliais
e leucocitárias (LAWRENCE & SPRINGER, 1991; KONSTANTOPOULOS &
McINTIRE, 1996). Os monócitos, linfócitos e polimorfonucleares migram
similarmente a esses mecanismos, no entanto, diferem em suas respostas, aos
sinais quimiotáticos e inflamatórios, particularmente quanto à quantidade e
qualidade das moléculas de adesão (DRANSFILD et al., 1992; SPRINGER, 1994;
AGER, 1996; LI et al., 1996). A migração dos polimorfonucleares se inicia com a
sua captura do sangue circulante e posterior “rolagem”, através da parede do
vaso sanguíneo. Esse processo de marginação é um comportamento normal dos

5
polimorfonucleares. Segundo WAGNER & ROTH (2000) apenas após o estímulo
apropriado, ocorre a aderência às células endoteliais e o posicionamento, para
migração do vaso sanguíneo, em direção ao parênquima tecidual (Figura 1).
FIGURA 1 – Mecanismo de rolagem, aderência e quimiotaxia
de leucócitos, bem como a ação das selectinas e integrinas Fonte: COTRAN et al. (2004)
Tanto a captura quanto a rolagem dos polimorfonucleares através do
vaso sanguíneo são devido as ligações reversíveis às selectinas (ALBELDA et al.,
1994; LUSCINSKAS & LAWLER, 1994; CROCKETT-TORABI & FANTONE,
1995).
À análise microscópica intravital da microcirculação vascular no tecido
normal é possível observar, a “fixação e liberação” dos polimorfonucleares à
parede do vaso. A associação tênue dos PMNs com as vênulas pós-capilares, em
tecido não-inflamado, pode ser bloqueado por meio de tratamento com anticorpo
contra selectina leucocitária, como o Mel-14, o LAM-1, o CD62L (SPERTINI et al.,
1991). A expressão construtiva de L-selectina é maior em PMNs recém-liberados
da medula óssea, comparado com PMNs maduros (MATSUBA et al., 1997). A
perda ou soltura de L-selectina da superfície dos PMNs é devida, em parte, à

6
atividade da metaloprotease, que resulta na rápida acumulação de L-selectina
bioativa no sangue (KISHIMOTO et al., 1995).
A adesão de neutrófilos no endotélio vascular é influenciada pela família
das integrinas (ARNAOUT, 1990; SUZUKI et al., 1991). As integrinas são um
grupo de glicoproteína transmembrânica, encontrada nos polimorfonucleares e
outras células hematopoiética que mediam a adesão de célula-célula e célula-
matriz extracelular (HYNES, 1992; LUSCINSKAS & LAWLER, 1994).
A proteína 140 é a segunda classe de proteína de aderência e é
reconhecida pelo anticorpo monoclonal MEL-14. Ela tem sido encontrada em
neutrófilos e linfócitos humanos e de camundongos. A proteína tem envolvimento
inicial de neutrófilos com as células endoteliais, em áreas inflamadas, e também
na aderência de linfócitos às células endoteliais, em veias nódulo-linfáticas. A
capacidade do endotélio em promover a adesão de neutrófilos é aumentada pelas
endotoxinas e citocinas, tais como o fator de necrose tumoral e IL-1 e está inibido
pela presença de outras citocinas, como o fator beta de crescimento (GAMBLE &
VADAS, 1988).
A deficiência de adesão leucocitária nos bovinos, é uma doença
genética autossômica e tem sido associada com a migração reduzida de
neutrófilos, no tecido. Os defeitos da função neutrofílica tais como aderência,
quimiotaxia, fagocitose e citotoxidade implicam em maior susceptibilidade a
infecções recorrentes (KEHRLI et al., 1990; TAJIMA et al., 1993; NAGAHATA et
al., 1993; GERARDI, 1996). Os achados hematológicos na deficiência da
molécula de adesão inclui persistente neutrofilia, hiperplasia granulocítica na
medula e ausência de neutrófilos nas lesões inflamatórias (JAIN, 1993).
A quimiotaxia é a relação entre movimento direcional do leucócito para
o alvo, sobre a influência do gradiente de concentração de substância
quimiotática no ambiente (FRASER, 1997). A ação envolve a participação de
componentes do citoesqueleto como os microfilamentos, microtubos e outras
proteínas de movimento, como a miosina. Além disso a quimiotaxia envolve a
interação de fatores quimiotáticos com específicos receptores de superfície
celular, os quais induzem sinal de transdução e orientam a direção e locomoção
da célula (JAIN, 1993).

7
São várias as substâncias quimiotáticas que podem ser geradas via
endógena ou exógena (HARVATH, 1991) e, incluem componentes do sistema de
complemento, calicreína, produtos bacterianos, vírus, metabólitos do ácido
araquidônico alguns leucócitos e produtos plaquetários, produtos das células
gigantes, fragmentos de IgG e IgM e substâncias liberadas da injúria tecidual. A
deficiência de fatores quimiotáticos ou a presença de substâncias que interferem
na quimiotaxia, podem potencializar as infecções (JAIN, 1993).
Uma vez que os leucócitos chegam ao alvo, sua função é fagocitar o
agente causador da inflamação (KAMPEAN et al., 2004). A fagocitose é um
processo de ingestão da partícula microscópica pelo leucócito, por meio da
extensão de pseudópodes citoplasmáticos em volta do alvo (PAAPE et al., 1979;
JAIN, 1993; BURVENICH et al., 1994; ANDREASEN & ROTH, 2000).
Os neutrófilos ingerem bactérias invasivas ou piogênicas e, as células
mononucleares, realizam a ingestão de microorganismos intracelulares
obrigatórios ou facultativos, como as micobactérias e a Listeria sp. Além disso, a
fagocitose de vírus, certos protozoários e de outras células, tem sido atribuída
também aos neutrófilos. As células imaturas, como bastonetes e metamielócitos,
são menos ativas fagociticamente, portanto, neutrófilos metamielócitos são pouco
funcionais na defesa do organismo (ESPINOSA & PAREDES, 2003).
A fagocitose é influenciada por várias propriedades físicas e químicas,
além das condições ambientais. A fagocitose, tal como a quimiotaxia, é um
processo ativo que requer energia, gerada pela glicólise anaeróbica. A ação
ocorre eficientemente com pH de 6 a 8 e à temperatura de 37º a 40º C. O mais
importante promotor da fagocitose é a opsonina. As opsoninas revestem a
superfície da bactéria e outras partículas estranhas, alterando suas propriedades
de superfície, tornando-as mais propícias à fagocitose (JAIN, 1993; ESPINOSA &
PAREDES, 2003).
As bactérias fagocitadas são destruídas pelos neutrófilos, por diferentes
mecanismos, que são denominados de oxigênio independente ou oxigênio
dependente. Ambos os mecanismos agem em acordo, para a morte e destruição
da bactéria (JAIN, 1993; IHA et al.,1996).
O mecanismo oxigênio independente inclui um ambiente ácido na
fagocitose e a presença de substâncias associadas a grânulos, como as proteínas

8
catiônicas, lactoferrina, lisozima e várias proteases, além das enzimas hidrolíticas
(MURRAY et al., 1989).
O aumento da utilização da glucose por meio da via pentose, aumenta
a atividade respiratória e a ativação da NADPH-oxidase na membrana produz o
metabólito pos-fagocitose ou burst respiratório (JAIN, 1993). Conseqüentemente,
são produzidos muitos metabólitos de oxigênio com atividade microbacteriana,
que inclui o ânion superóxido, H2O2, radicais hidroxil e o oxigênio livre (FARBER
et al., 1990).
O complexo H2O2 (peróxido de hidrogênio) com o MPO
(mieloperoxidase) forma um potente sistema bactericida, capaz de matar várias
bactérias, vírus, micoplasmas, fungos, além de destruir anticorpos cobertos por
células vermelhas. O oxigênio livre e radicais hidroxil matam bactérias por meio
da peroxidação lipídica, da lesão na membrana mitocondrial interna e no ácido
nucléico. Já o hipoclorido, um oxidante reativo, formado como um produto final da
explosão respiratória, é um bactericida e tem sido implicado na lesão tecidual
mediada por neutrófilos (BERNOFSKY et al., 1991).
2.4 Principais fatores que influenciam o leucograma
A resposta hematológica varia de acordo com as condições que os
animais são submetidos e, a quantificação do número de leucócitos, varia
consideravelmente entre os bovinos (COLE et al., 1997). Esta variação pode ser
devido a idade, estado gestacional, raça, atividade muscular e estresse do animal
(JAIN, 1989). FAGLIARI et al. (1998a) encontraram valores mais elevados nos
leucócitos de vaca no momento do parto. No leucograma de bovinos recém-
nascidos, criados a pasto ou confinados, observa-se elevada contagem de
leucócitos. A contagem total diminui em poucos dias de vida e estabiliza aos três
dias de idade (TAYLOR, 2000).
KLINKON & ZADNIK (1999) encontraram elevada contagem de
leucócitos nas vacas, anteriormente ao parto. Nas semanas que antecedem ao
parto e semanas após esse evento, MEGLIA et al. (2005) verificaram um quadro
de neutrofilia, eosinopenia, linfopenia e monocitose.

9
BIRGEL JUNIOR (1991), COSTA (1994), FAGLIARI et al. (1998b),
GONÇALVES et al. (2001) e PAULA NETO (2004), estudaram a influência da
idade na variação leucocitária. Os autores observaram que as contagens dos
leucócitos, linfócitos, segmentados e monócitos decresceram com o avançar da
idade. GONÇALVES et al. (2001) e PAULA NETO (2004) observaram ainda
aumento dos eosinófilos nos bovinos mais velhos.
2.4.1 Excitação e estresse
As mudanças na contagem diferencial de leucócitos pode ser mediada
por meio da liberação de epinefrina endógena, glicocorticóides e ainda, pelas
várias citocinas liberadas durante a inflamação (COLE et al., 1997).
Os exercícios intensos ou a excitação resulta na liberação de
epinefrina, a qual pode levar a mudanças no leucograma, chamado de leucocitose
fisiológica (DELDAR, 1984). A leucocitose fisiológica refere-se a uma contagem
de leucócitos acima do normal, sem nenhuma manifestação de doença
(NORTHINGTON, 2000). As características desta leucocitose são neutrofilia,
linfocitose, contagens variáveis de monócitos e, poucos eosinófilos (COLE et al.,
1997).
Nos animais em repouso, um considerável número de leucócitos
permanecem seqüestrados nos capilares sanguíneos. A liberação de epinefrina
nos animais excitados aumenta a circulação sanguínea e linfática.
Conseqüentemente, os leucócitos seqüestrados circulam para o sangue
periférico, causando leucocitose, acompanhada de neutrofilia e/ou linfocitose
(JAIN, 1993; COLE et al., 1997). TAYLOR (2000) acrescenta que a leucocitose
fisiológica induzida por epinefrina nos bovinos é caracterizada por leucocitose
neutrofílica, com aumento também de linfócitos e monócitos.
A resposta inicial dos bovinos a algum processo de doença é a
liberação de glicocortocóides (WEISS & PERMAN, 1992; COLE et al., 1997). O
efeito do estresse, devido ao corticosteróide exógeno ou à liberação endógena,
resulta em um leucograma típico (TAYLOR, 2000). Os linfócitos, monócitos,
macrófagos e granulócitos exibem receptores para o cortisol e catecolaminas, os

10
quais podem causar mudanças no tráfego celular, proliferação, secreção de
citocinas, produção de anticorpos e atividade citolítica. O efeito sobre a contagem
de leucócitos ocorre em função da espécie e da contagem diferencial dos
leucócitos. Nos bovinos, como os linfócitos são em maior número, ocorre uma
leve leucopenia. Os leucogramas de estresses possuem uma contagem
diferencial característica, que consiste de neutrofilia devida à liberação das
reservas de neutrófilos maduros da medula óssea e, dos neutrófilos marginados
(ANDERSEN, 1970; COLE et al., 1997) sem desvio, linfopenia e eosinopenia
(FRASER, 1997; TAYLOR, 2000). JAIN (1993) acrescenta que os monócitos têm
uma resposta variada, com a aplicação de corticosteróide exógeno. Segundo o
autor, essas células geralmente desaparecem da circulação em doenças
inflamatórias agudas. De fato, o retorno de monócito para o sangue, do ponto de
vista clínico, pode ser considerado um sinal de recuperação ou doença crônica.
JAMES et al. (1982) realizaram um estudo sobre a influência do ACTH,
no comportamento dos linfócitos e dos polimorfonucleares em bovinos e
observaram que, no grupo de bovinos tratados com ACTH, após o terceiro dia de
aplicação, a contagem de leucócitos foi de 16.340/mm3 comparado com os
animais do grupo controle, os quais apresentaram 8.230 leucócitos/mm3. No teste
da função de leucócitos polimorfonucleares, o grupo de animais tratados tiveram
aumento significativo na migração, entretanto, a blastogênese dos linfócitos foi
menor.
Os corticosteróides, direta ou indiretamente, afetam a cinética dos
leucócitos, a fagocitose, as células mediadoras da resposta imune, a imunidade
humoral e a produção de mediadores da inflamação. A linfopenia resultante da
ação do corticosteróide é devida à redistribuição dos linfócitos, da circulação para
os nódulos linfáticos, medula óssea e baço. Quanto à eosinopenia, têm sido
postuladas alterações na aderência e quimiotaxia dessas células (NONNECKE et
al., 1997).
A mudança dos leucócitos, associada a processos inflamatórios, poderá
ser relacionada juntamente com glicocorticóide, o qual promove a diminuição da
contagem de linfócitos durante inflamação aguda (COLE et al., 1997).

11
2.4.2 Ambiente
No sistema de criação de bovinos confinados, a poeira produzida pela
movimentação dos bovinos é um problema ambiental, que pode comprometer a
saúde dos animais, devido a irritação causada no sistema respiratório (MAC
VEAN et al., 1986). WILSON et al. (2002) avaliaram o efeito da poeira, de
tamanho igual ou menor a 2,5 μm, quantidade e diversidade de microorganismos
presentes no ar de confinamentos de bovinos. Ao analisar as bactérias, apenas
microrganismos não patogênicos foram identificados; quanto aos fungos, foram
recuperados em menor número do que as bactérias e nenhum deles patogênico.
FULTON et al. (2002) realizaram uma avaliação do estado sanitário de um
rebanho de bovinos submetidos ao confinamento. Preocupados principalmente,
com a agressão do sistema respiratório dos animais, os autores avaliaram a
sorologia para o vírus da rinotraqueíte infecciosa bovina, diarréia viral bovina 1 e
2, vírus da parainfluenza 3, vírus sincicial respiratório bovino, Mannheimia
haemolytica e Pasteurella multocida, além de swab nasal. A presença de vírus e
bactéria não foi capaz de indicar a existência de doença, no entanto previu a sua
severidade bem como a performance dos animais confinados ou seja, os grupos
de animais com baixa morbidade tiveram níveis mais altos de anticorpos anti
BVDV-1 do que os animais com elevados índices de morbidade. Em estudo
realizado por FAGLIARI (2003), a inoculação intrabronquial de Mannheimia
haemolytica foi feita em bezerros e o autor observou um quadro de neutrofilia e
alto número de neutrófilos e monócitos no fluido broncoalveolar.
RUPPANNER et al. (1978), estudando o manejo de bovinos destinados
a produção de leite e carne, avaliaram o perfil sanguíneo e encontraram elevada
contagem total de leucócitos nos bovinos para abate. De acordo com os autores,
o aumento da contagem leucocitária pode ser atribuída ao manejo excessivo dos
animais, bem como o transporte, principalmente no início do confinamento. Os
autores ressaltaram ainda a importância de se estudar o perfil hematológico de
animais confinados, no sentido de monitorar o estado sanitário desses bovinos,
além de detectar os que não são economicamente viáveis para a produção.
A “performance” e saúde dos bovinos são fortemente influenciadas pelo
clima (MADER, 2003). Durante o verão observa-se elevada contagem de

12
leucócitos (MARAI et al., 1999). MADER (2003), recomenda que algumas
estratégias sejam adotadas nos confinamentos de bovinos, para minimizar o
impacto ambiental na produção desses animais. Segundo o autor deve-se utilizar
barreiras para evitar estresse pelo frio no inverno e, no verão, deve-se
providenciar sombras para minimizar o estresse pelo calor. O efeito do estresse
provocado pelo frio foi estudado por OLSON et al. (1983) nos bovinos. Os autores
não encontraram diferenças hematológicas significativas entre os bezerros
submetidos e os não-submetidos a baixas temperaturas. Entretanto, nos bovinos
submetidos ao frio, houve tendência de maior contagem total de leucócitos, com
neutropenia, comparado aos animais em temperatura normal.
Segundo OHTSUKA et al. (2005), a nutrição adequada de bezerros,
estimula o aumento dos leucócitos durante o período de crescimento.
ANDRESEN (1970) estudou a influência nutricional no hemograma de bovinos, e
observou um quadro de leucocitose e monocitose nas vacas que se alimentavam
com silagem de capim, comparadas com as que se alimentavam com forragem
verde.
Outro parâmetro estudado, em relação à mudança ambiental, é o
transporte dos animais destinados ao abate. O transporte é um evento pouco
familiar para os animais, onde os bovinos são expostos a fatores como calor, frio,
altas ou baixas umidades, além de privação de água, alimento e movimento
(TADICH et al., 2003). Dentre os efeitos adversos produzidos por ocasião do
transporte, destacam-se a possibilidade de morte, perda de peso e mudanças nos
constituintes sanguíneos (KNOWLES, 1999). COLE et al. (1988) submeteram 150
novilhos, ao transporte em caminhões comerciais, durante zero (grupo controle),
12 ou 24 horas. Em todos os grupos, a contagem diferencial dos leucócitos foi
indicativa de resposta ao estresse, observando contraste linear entre a duração
do transporte e a contagem de leucócitos, neutrófilos segmentados, linfócitos e
eosinófilos. SWANSON & MORROW-TESCH (2001) afirmaram que o transporte
de bovinos pode resultar em uma supressão do sistema imunológico, podendo
levar a aumento da susceptibilidade a doenças. Segundo os autores, há
evidencias empíricas que a razão neutrófilo:linfócito aumenta na ocasião que os
animais são manipulados e transportados.

13
No sistema de confinamento de bovinos, manobras como o jejum e
transporte dos animais, para o confinamento ou mesmo no transporte desses
bovinos para o abate, tem-se identificado elevação no número de leucócitos
desses animais (TADICH, et al., 2003) 2.4.3 Comportamento dos leucócitos nas doenças respiratórias
Entre os animais domésticos, os bovinos são mais susceptíveis a
doenças respiratórias devido a algumas deficiências na função pulmonar, tais
como: pequena capacidade nas trocas gasosas, compartimentação anatômica e
pouca ventilação colateral, baixo número de macrófagos residentes e uma fraca
resposta leucocitária à infecção, baixos níveis de lisozima, orientação anatômica
dependente e má perfusão de oxigênio ao nível dos lobos anteroventrais, uma
superfície grande e rugosa nas tonsilas faringeanas, servindo como fonte
microbiana para a “semeadura” do trato respiratório inferior, além da área
endotelial relativamente pequena para a eliminação das toxinas (YATES, 1990).
Nos confinamentos de bovinos de corte, as enfermidades respiratórias
assumem grande importância econômica e sanitária (GRIFFIN, 1997;
LONERAGAN et al., 2001). Atualmente, existem no Brasil aproximadamente
2.006.558 confinamentos. Em Goiás, considerado destaque na produção de
animais de corte, há por volta de 293.250 confinamentos, sendo que 95% destes
possuem piso de terra (ANALPEC, 2005). Devido à movimentação dos animais
nesse ambiente, há formação de grandes quantidades de pó e partículas
suspensas no ar (SWEETEN et al., 1988) (Figura 2), podendo contribuir com a
dispersão de patógenos (MILLER & WOODBURY, 2003). Entre essas partículas,
cujo tamanho varia entre 2,5 a 12,2 µm de diâmetro (MacVEAN et al., 1986;
SWEETEN et al., 1988) encontram-se bactérias, vírus, fungos (WILSON et al.,
2002) que agridem, principalmente o sistema respiratório (MacVEAN et al., 1986;
DUTRA, 2001; WILSON et al., 2002), uma vez que ele comunica-se intensamente
com o mundo exterior (YATES, 1990; GOMES, 2002).
Além desse aspecto, no Brasil os confinamentos de bovinos, são
conduzidos durante a época seca do ano (CARDOSO, 2000). Nesse período,

14
ocorrem também baixas umidades, especialmente no Estado de Goiás, o qual
caracteriza-se por ter clima quente, com até cinco meses de seca (INMET, 2004),
tornando o ambiente altamente prejudicial à respiração dos animais e das
pessoas envolvidas no trabalho do confinamento (DUTRA, 2001).
Vários fatores predisponentes, como as infecções virais, seguidas
pelas infecções bacterianas secundárias, estão envolvidos na patogenia das
doenças respiratórias bovinas (SOETHOUT et al., 2002).
Os agentes causadores da broncopneumonia, ao penetrarem no
sistema respiratório, podem quebrar as barreiras de defesa dos pulmões e
desenvolver doença. A própria divisão das vias aéreas é fator de proteção, pois
além de provocar impacto das partículas agressoras na mucosa brônquica, pela
disposição anatômica das vias condutoras de ar (BREEZE, 1985) propicia
diminuição da velocidade do ar inalado, facilitando a deposição de tais partículas
sobre o muco que recobre as células ciliadas (DIESEL et al., 1991). No momento
da deposição dessas partículas, há o estímulo do mecanismo de limpeza
mucociliar e as partículas são eliminadas pelo movimento dos cílios, que
direcionam o muco à faringe para que seja deglutido (BREEZE, 1985; DIXON,
1992). Conforme a agressão dos agentes patogênicos primários, ou mesmo das
infecções bacterianas secundárias, o sistema mucociliar vai sofrendo alterações
morfofuncionais, culminando com o desaparecimento das células ciliadas
(DIXON, 1992).
Nas porções mais distais das vias aéreas e nos alvéolos, as células
ciliadas diminuem em número ou estão ausentes, de tal maneira que na junção
broncoalveolar e alvéolos, a defesa local é desenvolvida, principalmente, pelos
neutrófilos e macrófagos alveolares (LEID & POTTER, 1985; LIGGITT, 1985;
LOPEZ et al., 1986; DYER et al., 1989).

15
FIGURA 2 – Animais submetidos ao sistema de confinamento
(A), as instalações e o piso de terra (B)
Se o microorganismo conseguir ultrapassar essas defesas do pulmão,
ocorre um processo inflamatório, que provoca algumas modificações no ambiente
pulmonar. A inflamação, por si só estimula a descamação do epitélio
traqueobrônquico (SWEENEY & BEECH, 1991) e a formação do exudato
inflamatório, mediado por fatores quimiotáticos (LIGGITT, 1985; LEID & POTTER,
1985). Os neutrófilos são as células que rapidamente respondem aos agentes
quimiotáxicos, saindo da microcirculação e penetrando no pulmão (COHEN &
ROSSI, 1983; REYNOLDS, 1983; LEID & POTTER, 1985). Devido ao rápido e
volumoso afluxo destas células ao pulmão e à liberação de mediadores químicos
da inflamação, que provocam aumento dos neutrófilos circulantes, levando ao
acúmulo e ativação dos neutrófilos no sítio das lesões, são também fatores
capazes de induzir graves danos ao pulmão (SLOCOMBE et al., 1985; BREIDER
et al., 1988; SLOCOMBE et al., 1990; LAPOINTE et al., 1993) pela liberação de
A
B

16
produtos protéicos, tóxicos ao ambiente pulmonar (LEDWOZYW &
STOLARCZYK, 1991) e também radicais livres, resultante da explosão
respiratória aos neutrófilos (JAIN, 1993).
Alguns estudos em bovinos indicam que microorganismos como o vírus
da Parainfluenza (BRYSON et al., 1983; SLAUSON et al., 1987; BROWN &
ANABA, 1988) o vírus da rinotraqueíte infecciosa bovina (BROWN & ANABA,
1988), o vírus sincicial respiratório bovino (CASTLEMAN et al., 1985) e a
Mannheimia haemolytica (RICHARDS & RENSHAW, 1989; SLOCOMBE et al.,
1984; SLOCOMBE et al., 1990; LEITE, 2002) podem reduzir os mecanismos de
proteção pulmonar. São descritas destruição dos cílios das células epiteliais
cilíndricas ciliadas, com alteração do mecanismo de limpeza mucociliar (BRYSON
et al., 1983; CASTLEMAN et al., 1985; SCOTT, 1994) e redução no número e na
função de macrófagos (SLAUSON et al., 1987; BROWN & ANANABA, 1988) e
neutrófilos (MARKHAN & WILKIE, 1980; RICHARDS & RENSHAW, 1989;
SLOCOMBE et al., 1990).
Dentre os agentes causadores de doenças respiratórias em bovinos, a
pneumonia por bactérias do gênero Pasteurella, Mycoplasma e Haemophilus, M.
haemolytica (CAVERLY et al., 2003; FAGLIARI, 2003) são mais comumente
isoladas.
A M. haemolytica inicia um severo quadro de resposta inflamatória no
pulmão dos ruminantes, caracterizado por um denso infiltrado de neutrófilos
(WITHITELEY et al., 1990; ACKERMANN et al., 1996). Durante a fase de
multiplicação no tecido pulmonar os agentes causadores de pneumonia liberam
substâncias como neuramidase (FRANK & TABATABAI, 1991), protease
(OTULAKOWSKI, 1983) e leucotoxina (MAHESWARAN et al., 1993) que
estimulam os mecanismos de defesa do organismo, especialmente fatores de
coagulação, o sistema de complemento, neutrófilos, monócitos e macrófagos os
quais provocam a liberação de mediadores inflamatórios, como as IL-8 que levam
a diversas alterações sistêmicas (FAGLIARI, 2003). A citocina IL-8, produzida
pelas células endoteliais, células epiteliais, macrófagos ativados, causam a
ativação e quimiotaxia de neutrófilos (BAGGIOLINI et al., 1989).
Estudo realizado por (FAGLIARI, 2003) utilizou a inoculação via
intrabronquial de Mannheimia haemolytica em sete bezerros da raça Holandesa,

17
entre 15 e 30 dias de idade e realizou análise sanguínea e de lavado
broncoalveolar antes e 4, 6, 12 e 24 horas após a inoculação. Os resultados do
leucograma indicaram discretas variações na contagem total de leucócitos. Notou-
se que a partir de quatro horas da inoculação ocorreu diminuição do número de
linfócitos, e das 12 e 24 horas aumento do número de neutrófilos segmentados.
Quanto a avaliação broncoalveolar, houve neutrofilia e aumento do número de
monócitos. Segundo JAIN (1993), essa resposta leucocitária sugere um quadro
de infecção bacteriana. No entanto, o autor ressalta a discreta variação da
resposta inflamatória em ruminantes e o desvio degenerativo para a esquerda nas
primeiras 24 a 48 horas de um processo inflamatório. Nos bovinos a pasto, a pneumonia verminótica causada pelo
Dictyocaulus viviparus assume importância em relação às enfermidades
respiratórias. A doença tem distribuição mundial e ocorre com maior freqüência
durante o verão ou outono (EYSKER, 1994; OGILVIE, 2000; DUTRA, 2005). A
intensidade dos sinais clínicos depende de fatores como estágio da doença e da
carga parasitária, mas usualmente inclui anorexia, emagrecimento, tosse e
taquipnéia; ocorre febre em casos onde já encontra-se estabelecida infecções
bacterianas secundárias (OGILVIE, 2000; ZAJAC et al., 2002).
2.5 Interpretação dos parâmetros leucocitários 2.5.1 Contagem total de leucócitos
O leucograma pode indicar a presença de processos infecciosos,
inflamatórios, toxêmicos, neoplásico e de estresse (PAES et al., 2003). A
contagem total de leucócitos é avaliada quanto ao aumento ou diminuição do
número de células, denominado de leucocitose e leucopenia, respectivamente. A
leucocitose pode ser fisiológica, reativa ou proliferativa, sendo mais comumente
observada do que a leucopenia (JAIN, 1993).
A leucocitose fisiológica pode ser em resposta a epinefrina e em
estresse induzido por corticóide (JAIN, 1993; FRASER, 1997). Segundo VAL

18
BICALHO & CARNEIRO (2005) a leucocitose fisiológica deve ser compreendida,
para que haja discernimento entre esta e a patológica.
A leucocitose reativa ocorre em resposta à doença, podendo ocorrer
com ou sem desvio para a esquerda (JAIN, 1993). A leucocitose por neutrofilia,
geralmente caracteriza infecções bacterianas e problemas relacionados com
extensa necrose tecidual, incluindo queimaduras, traumas, cirurgias extensas e
algumas vezes, neoplasias malignas (FRASER, 1997), enquanto que, a
leucocitose proliferativa, resulta de mudanças neoplasicas no sistema
hematopoético (JAIN, 1993).
Nos bovinos tem-se postulado que a leucopenia está associada à
mastite (MENZIES, 2000; VERKKOJULKAISUT, 2000), hipersensibilidade,
atividade do corticosteróide (JAIN et al., 1989; NONNECKE et al., 1997), doenças
virais e leucose (ANDRESEN, 1970). A leucopenia ocorre precocemente nas
infecções bacterianas dos ruminantes, devido a sua baixa reserva de neutrófilos
na medula óssea e a linfopenia secundária à liberação de glicocorticóides
endógenos. Além desse aspecto, pode ocorrer leucopenia, em resposta à
radiação e drogas radiomiméticas, infecções virais, particularmente as agudas,
não associadas com necrose extensa dos tecidos (FRASER, 1997).
Uma vez que os leucócitos são formados, permanecem durante algum
tempo no espaço extravascular da medula óssea. Depois, através das
fenestrações intercelulares existentes na parede endotelial, eles ganham o
espaço intravascular e a circulação. A produção e a saída para a circulação dos
leucócitos são regulados pelas necessidades do organismo, embora a maior parte
da população leucocitária permaneça no interior da medula. Na circulação, os
leucócitos formam duas populações ou “pools” celulares, de tamanhos
aproximadamente iguais, o “pool” circulante e o marginal, sendo que as células
deste último grupo ficam aderidas à parede dos vasos (KANTEK & NAVARRO,
2005).
Segundo JAIN (1993), a mudança na contagem de leucócitos totais,
pode envolver anormalidades de produção, liberação, distribuição intravascular e
vida média da célula. O autor acrescenta que a circulação de neutrófilos é um
equilíbrio dinâmico com o “pool” marginal e a reserva da medula óssea. Em uma
demanda funcional imediata de neutrófilos, primeiramente há mobilização das

19
células do “pool” marginal e circulante, depois da reserva da medula óssea e
finalmente há aumento da granulopoiese e liberação acelerada, refletindo no
desvio de neutrófilos para a esquerda. O tamanho do “pool” marginal e circulante,
além do tamanho da reserva da medula óssea, bem como a sua capacidade
proliferativa são importantes para a resposta de leucócitos à enfermidades.
2.5.2 Contagem diferencial de leucócitos
A interpretação da contagem diferencial de leucócitos deve ser
empregada na caracterização de processos inflamatórios (RYAN, 1971; COLE et
al., 1997;). As mudanças quantitativas e qualitativas podem ocorrer com vários
leucócitos. É necessário a compreensão da resposta às doenças (ANDRESEN,
1970) e do mecanismo fisiopatológico das anormalidades leucocitárias, para
avaliar as mudanças numéricas e morfológicas dos leucócitos. Nesse aspecto, a
contagem absoluta é preferível do que a contagem relativa, numa interpretação
do leucograma (JAIN, 1993).
Os neutrófilos são as células de primeira linha de defesa nas reações
inflamatórias e contra as infecções. Assim sendo, foram propostos termos, como
desvios, os quais podem ocorrer para a direita e para a esquerda, para descrever
as alterações no sangue, na contagem diferencial dessas células. Esses desvios
são baseados na contagem total de leucócitos, na contagem diferencial de
neutrófilos e no grau de maturação destes (VAL BICALHO & CARNEIRO, 2005).
Além da diferenciação dos neutrófilos, a sua morfologia é muito
importante na determinação da causa de neutrofilia e interpretação do
leucograma (MORRIS & LARGE, 1990). A neutrofilia patológica é observada na
agudização de processos crônicos, anteriormente em equilíbrio (FRASER, 1997),
além de intoxicações metabólicas e não-metabólicas (CUNHA et al., 2004), lesões
com necrose abrangente de órgãos e tecidos como o miocárdio, pâncreas e rins e
nas leucemias mielocíticas (VAL BICALHO & CARNEIRO, 2005). Observa-se
neutrofilia também na fase inicial e de regeneração das hemorragias, quando a
liberação aumentada de eritrócitos jovens pode vir acompanhada de maior
número de neutrófilos (FRASER, 1997).

20
Segundo VAL BICALHO & CARNEIRO (2005), a neutropenia ocorre
basicamente por dois mecanismos. O primeiro é quando há diminuição da
produção de neutrófilos por uma hipoplasia granulocítica da medula óssea, seja
ela de origem infecciosa ou por intoxicação por plantas, como a samambaia, nos
bovinos. O segundo mecanismo é o excesso de consumo dos neutrófilos, em
processos infecciosos graves e demorados. MORRIS & LARGE (1990) citam
ainda como causa de neutropenia as septicemias e/ou endotoxemias bacterianas,
especialmente quando causadas por enfermidades gastrointestinais, metrites ou
mastites, moléstias virais e anafilaxia.
A linfocitose ocorre quando há aumento de linfócitos no sangue e
geralmente é observada em convalescência das doenças infecciosas, quando há
insuficiência adrenocortical, hipertiroidismo (MATOS & MATOS, 1988) e leucemia
linfocítica (MOULTON, 1990). A linfocitose por estimulação imunológica está
associada a inflamações crônicas e é caracterizada pela presença de linfócitos
reativos. Esses linfócitos possuem um citoplasma mais basófilo e um pouco mais
abundante devido ao aumento da síntese protéica; essas células podem ocorrer
em qualquer doença que cause uma imunoestimulação sistêmica, moderada a
acentuada (FRASER, 1997).
MORRIS & LARGE (1990) descreveram que a linfopenia pode ocorrer
em moléstias virais agudas, liberação de endotoxinas, grave infecção bacteriana,
septicemia, imunodeficiência, moléstias riquetsiais e desnutrição. A administração
de antagonistas do ácido fólico e drogas antineoplásicas, também levam à
linfopenia (VAL BICALHO & CARNEIRO, 2005; KANTEK & NAVARRO, 2005).
Embora, a eosinofilia não tenha o mesmo significado clínico das
alterações de neutrófilos e linfócitos, um leucograma que apresenta eosinofilia,
merece atenção (KANTEK & NAVARRO, 2005). A eosinofilia é provocada por
histamina e substâncias aliadas e, pela IgE. Estas indicam alergia e
hipersensibilidade, ou doenças parasitárias, nas quais a intensidade da eosinofilia
é um fator indicativo do contato da quitina do parasita com o hospedeiro
(FRASER, 1997). Segundo MORRIS & LARGE, (1990) raramente ocorre
eosinofilia em bovinos, mas ela pode ser observada, em caso de moléstias que
envolvam a interação entre antígenos, anticorpo, IgE, mastócitos ou basófilos.

21
A eosinopenia ocorre na fase aguda das inflamações, no estresse
emocional, excitação, durante o parto, na presença de endotoxemias, no choque
e estados de anoxia (KANTEK & NAVARRO, 2005) nas situações em que há
excesso de hormônios corticosteróides circulantes, sejam de origem endógena ou
exógena (MATOS & MATOS, 1988; VAL BICALHO & CARNEIRO, 2005).
MORRIS & LARGE, (1990), alertam que um quadro de eosinopenia é difícil de
reconhecer pois o leucograma dos animais clinicamente sadios contem
pouquíssimos eosinófilos.
A monocitose é observada principalmente na fase de recuperação das
inflamações. Outras situações em que há monocitose são a desnutrição,
caquexia, inflamações não específicas, doenças crônicas como a tuberculose,
brucelose e as causadas por protozoários (KANTEK & NAVARRO, 2005) e na
leucemia monocítica (VAL BICALHO & CARNEIRO, 2005; MOULTON, 1990). As
monocitopenias são ocasionalmente observadas, mas não possuem significado
diagnóstico (FRASER, 1997; VAL BICALHO & CARNEIRO, 2005).
Os basófilos raramente são observados no sangue periférico de
bovinos (JAIN, 1989) e as alterações na quantidade dessas células são de difícil
interpretação, geralmente quando ocorre, está relacionada à dermatite alérgica e
reações de hipersensibilidade retardada (MORRIS & LARGE, 1990) e ainda, nas
mesmas condições de alteração do número de eosinófilos (VAL BICALHO &
CARNEIRO, 2005).
2.6 Constituintes séricos/plasmáticos 2.6.1 Proteína total
O fluido no qual as células sanguíneas estão em suspensão é chamado
de plasma (MEYER & HARVEY, 2004). As proteínas plasmáticas são constituídas
por centenas de proteínas, com ampla classificação quanto à função e estrutura.
Baseado na eletroforese, elas podem ser dividas em albumina, α, β e γ globulinas
(KANEKO, 1989).

22
A concentração das proteínas plasmáticas totais são comumente
avaliadas utilizando-se um refratômetro ou bioquimicamente, por meio de
reagentes comerciais utilizando como princípio o método de biureto (KANEKO,
1989; MEYER & HARVEY, 2004).
A maior parte das proteínas plasmáticas são sintetizadas no fígado,
dentre estas, a albumina e a maioria das α e β globulinas. As γ globulinas são
produzidas pelos órgãos linfóides (THOMAS, 2000).
A concentração de proteínas no plasma é maior do que no soro, devido
à presença de fibrinogênio no plasma (MEYER & HARVEY, 2004). A
concentração de proteínas no plasma reflete um equilíbrio entre as concentrações
intra e extra vascular, entretanto, a concentração de proteínas no sangue, não
reflete adequadamente o estoque de proteína total (JAHOOR et al., 1996).
Alguns fatores afetam a concentração de proteína plasmática, tais
como: a idade do animal (JAIN, 1993; CANAVESSI et al.,2000; GONÇALVES et
al., 2001), nutrição (HYVARINEN et al., 1975; JAIN, 1993), sazonalidade
(HYVARINEN et al., 1975), efeito hormonal, balanço de fluidos e em
enfermidades (HARVEY & WEST, 1987; JAIN, 1993; ECKERSALL, 1995;
KLIMIENE et al., 2005).
Segundo MEYER & HARVEY (2004) a hiperproteinemia pode ocorrer
devido a desidratações ou pelo aumento da síntese de globulinas. Já a
hipoproteinemia pode ocorrer pela diminuição da produção de albumina ou
imunoglobulinas, perda de proteína associada com hemorragia, perda de
proteínas em nefropatias ou enteropatias.
As doenças inflamatórias têm um significante efeito nas proteínas
plasmáticas. Durante reparos teciduais, há aumento das proteínas plasmáticas.
Além disso, o processo de inflamação tecidual causa aumento da permeabilidade
vascular, perda de proteínas, a princípio do espaço extra-vascular e
subseqüentemente do espaço vascular (KANEKO, 1989). Durante o estresse e
processos inflamatórios, há aumento da produção de globulina que migra na
fração α e β, como parte à resposta de fase aguda (ECKERSALL, 1995).
Devido ao significado biológico das proteínas, bem como as múltiplas
funções exercida no sistema orgânico, a avaliação dos níveis séricos das

23
proteínas totais e de suas frações, como albumina, α, β e γ-globulinas,
representam um importante auxílio ao diagnóstico clínico (KANEKO, 1989).
2.6.2 Albumina
A albumina é uma proteína sérico-plasmática homogênea, que contém
uma pequena quantidade de carboidrato em sua molécula, podendo ser
glicosilada pela interação monoenzimática com glucose (MEYER & HARVEY,
2004). Dentre as proteínas plasmáticas, a albumina é a mais abundante. Devido
ao grande tamanho da molécula, normalmente ela é retida nos capilares,
entretanto, ela é a primeira proteína a ser perdida durante as injúrias teciduais. A
albumina tem uma função importante na manutenção da função coloidosmótica
sanguínea. Além disso, ela se destaca como proteína transportadora de ácidos
graxos, ácidos biliares, bilirrubina, porfirinas, cetosteróides, diversas drogas, como
as penicilinas, aspirinas, barbitúricos, histaminas e cátions (KANEKO, 1989; JAIN,
1993).
A albumina é sintetizada no fígado e catabolizada em vários tecidos.
Sua síntese é influenciada pelo estado de nutrição, balanço hormonal, condições
gerais do fígado, estresse e a sua própria concentração extravascular. A síntese
da albumina é inversamente proporcional à síntese das proteínas de fase aguda e
é regulada pela interleucina-1 (IL-1) e outras proteínas (JAIN, 1993). Nos animais,
a diminuição da síntese de albumina tem sido relatada como resultante de
processos inflamatórios induzidos pela injeção de turpentina e IL-1 (BALLMER et
al, 1995).
A meia vida da albumina nos bovinos é de 15 a 21 dias. Segundo
MEYER & HARVEY (2004), apesar da concentração de albumina sofrer variação
entre as espécies, geralmente seu valor encontra-se entre 2,5 g/dL a 4,5 g/dL no
plasma ou no soro e esse valor pode ser obtido bioquimicamente, por meio do
verde de bromocresol ou pela eletroforese.
A hipoalbuminemia é melhor avaliada em associação com a razão
albumina:globulina (A/G). Um quadro de hipoalbuminemia, combinada com a
globulina normal a aumentada (razão A/G diminuída), ocorre secundariamente à

24
perda seletiva de albumina, seqüestro dentro do espaço intravascular ou
diminuição da produção. Já a hipoalbuminemia, juntamente com hipoglobulinemia
(razão A/G normal), sugere hiperhidratação, perda sanguínea aguda, lesões
exudativas ou perda de proteínas em enteropatias (KANEKO, 1989; DUNCAN et
al., 1994). Um quadro de hipoalbuminemia auxilia o clínico a limitar o seu
diagnóstico diferencial de uma determinada enfermidade. Segundo WILLARD
(2000), há quatro considerações para se determinar um quadro de
hipoalbuminemia, tais como, diminuição da produção devido a uma insuficiência
hepática, perda de proteína em nefropatias, perda em enteropatias, incluindo
perda de proteínas gástricas, e perdas em lesões cutâneas ou hemorragias.
A hiperalbuminemia geralmente é resultante de hemoconcentração,
devido à elevação do nível da albumina sérica, entretanto, não ocorre aumento da
sua síntese (JAIN, 1993).
2.6.3 Globulina
As globulinas são responsáveis pelo transporte de glicoproteínas,
lipoproteínas, mucoproteínas e cobre (MATOS & MATOS, 1988).
As globulinas podem ser classificadas em α, β e γ-globulinas (KANEKO,
1989; MEYER & HARVEY, 2004).
A concentração de globulina total é obtida, por meio da subtração da
concentração de albumina, da concentração de proteína sérica total,
conseqüentemente, um erro na avaliação da concentração de proteína ou
albumina, pode resultar numa concentração equivocada de globulina (MEYER &
HARVEY, 2004; GRÜNWALDT et al., 2005).
CHORFI et al. (2004) alertaram sobre alguns fatores que podem
influenciar a concentração de globulinas nos animais saudáveis. Dentre eles o
próprio método de avaliação da proteína e o local da coleta do sangue. De acordo
com MEYER & HARVEY (2004) a concentração total de globulina plasmática
pode ser baixa, denominada de hipoglobulinemia em casos de hiperhidratação,
perda de globulinas devido a hemorragias, exudação massiva, perda de proteínas

25
em enteropatias, falhas de transferência ou defeito na síntese de imunoglobulinas
em animais neonatos.
Segundo MEYER & HARVEY, (2004) e THOMAS (2000), são causas
de hiperglobulinemia as desidratações, além do aumento da síntese de proteínas
de fase aguda, as quais ocorrem associadas com resposta inflamatória injúrias
teciduais e/ou corpos estranhos.
A relação albumina/globulina (A/G) é a divisão da concentração de
albumina pela concentração de globulina sérica (RAMÍREZ et al., 2001). O índice
albumina:globulina (A/G) considerado de modo isolado tem pouco valor. Essa
relação adquire importância nas infecções, quando ela se altera, invertendo-se os
valores pelo incremento que ocorre na concentração das imunoglobulinas,
especialmente as γ-globulinas. No soro dos animais, O índice A/G obedece a
valores diversos, sendo que nos bovinos essa relação é menor, comparando com
o soro de humanos e cães (BACILA, 2003). Em estudo bioquímico-clínico de
bovinos da raça Nelore, realizado por BARROS FILHO (1995), verificou que na
faixa etária de 24 a 60 meses, a relação Albumina:Globulina (A:G) foi de 0,81.
RAMÍREZ et al. (2001) caracterizou a relação A/G em vacas leiteiras em três
épocas do ano: seca, entrada da chuva e período chuvoso. Os autores
observaram que a concentração de proteínas plasmáticas totais, assim como a
relação A/G, foi maior na entrada da chuva em relação à seca e ao período
chuvoso.
2.6.4 Fibrinogênio
O fibrinogênio é uma glicoproteína, componente integral da hemostasia,
que fornece substrato para a trombina na formação de fibrina. Em reparos
teciduais, ele promove a matriz para migração de células inflamatórias,
fibroblastos e células endoteliais (THOMAS, 2000). O fibrinogênio liga-se à
superfície celular das integrinas do fagócito migrado e desse modo, provoca uma
cascata de sinais intracelulares que comandam o aumento da degranulação, a
fagocitose e a citotoxidade celular, anticorpo-dependente (SITRIN et al., 1998;
RUBEL et al., 2001).

26
O fibrinogênio comporta-se como uma proteína de fase aguda
(THOMAS, 2000; VEKKOJULKAISUT, 2000; BERRY et al., 2004; MURATA et al.,
2004) e é a proteína mais usada para avaliar a reação de fase aguda nos bovinos
(BENJAMIN, 1978; WISS & PERMAN, 1992; COLE, 1997). A avaliação do
fibrinogênio é utilizada na medicina veterinária, especialmente na bioquímica de
grandes animais, como indicadora da presença de inflamação, infecção
bacteriana ou trauma cirúrgico (MURATA et al., 2004). Isso é possível, devido ao
aumento da síntese de fibrinogênio pelos hepatócitos, que é resultante do
estímulo de IL-1, IL-6 e fator de necrose tumoral, os quais são liberados durante
os eventos inflamatórios e traumáticos (JAIN, 1993). Após 24 a 36 horas da injúria
tecidual, aumenta a concentração plasmática do fibrinogênio, precedido pelo
aumento da gama-globulina. A concentração do fibrinogênio permanece elevada
durante a ação da doença, mas sem exceder a capacidade de produção do fígado
(SUTTON & HOBMAN, 1975). Entretanto, a concentração do fibrinogênio
freqüentemente retorna ao limite normal, vários dias após a destruição tecidual ter
parado, levando à continuada elevação da gama-globulina, o que é característico
de doenças crônicas (PHILLIPS, 1984).
A concentração do fibrinogênio é estimada pela diferença de proteína
plasmática total, por meio de um refratômetro, antes e após a precipitação do
fibrinogênio (WEISS & PERMAN, 1992; FRASER, 1997; COLE et al., 1997;
MEYER & HARVEY, 2004).
A concentração plasmática de fibrinogênio não é afetada pela idade,
sexo, exercício, repetido sangramento e hemorragia (McSHWRRY et al., 1970;
JAIN, 1993), entretanto, o nível da proteína encontra-se elevado em situação de
estresse (GENTRY et al., 1992), em várias condições inflamatórias, infecções
supurativas, processos traumáticos e neoplásicos, em muitas espécies (WEISS &
PERMAN, 1992), parecendo ser um melhor indicador de inflamação para bovinos,
do que as alterações na contagem de leucócitos (McSHERRY et al., 1970;
SUTTON & HOBMAN, 1975; FRASER, 1997).
SUTTON & HOBMAN (1975) avaliaram o sangue de 716 bovinos e
encontraram em 18,6% dessas amostras nível normal de fibrinogênio, mas a
contagem total de leucócitos e/ou contagem de neutrófilos eram anormais. Outros
95% apresentaram elevado nível de fibrinogênio e contagem leucocitária normal.

27
Os autores concluíram portanto, que a dosagem do fibrinogênio é melhor para a
detecção de doenças inflamatórias ou traumáticas, do que a contagem total e
diferencial de leucócitos.
McSHERRY et al. (1970) mediram o nível do fibrinogênio em vacas
doentes e sadias. Segundo os autores, nem todos os animais doentes tiveram
hiperfibrinogemia e, o baixo nível de fibrinogênio detectado em animais doentes, é
sinal de prognóstico ruim, pois pode tratar-se de animais com doenças hepáticas
ou em estado terminal.
BERRY et al. (2004) analisaram a concentração de fibrinogênio em
relação à nutrição, composta por vários níveis energéticos, não encontrando
diferença na concentração de fibrinogênio em animais tratados com diferentes
alimentos energéticos.
Além das causas de aumento e diminuição do fibrinogênio, é importante
observar a relação proteína plasmática:fibrinogênio (PP:F). Este índice é usado
para distinguir hiperfibrinogemia causada por doença, ou associada com
desidratação. Isso porque a hemoconcentração produz um aumento relativo de
todos os componentes das proteínas plasmáticas, incluindo o fibrinogênio
(McSHERRY et al., 1970; SUTTON & HOMAN, 1975; JAIN, 1993), entretanto, sua
síntese elevada, não é acompanhada pela síntese aumentada das outras
proteínas (JAIN, 1993). A relação PP:F é obtida pela subtração da concentração
de fibrinogênio da concentração de proteína plasmática total e dividindo pela
concentração de fibrinogênio. Geralmente, relação PP:F menor que 15:1, indica
aumento relativo do fibrinogênio comparado com as proteínas plasmáticas. A
relação PP:F abaixo de 10:1 indica grande aumento de fibrinogênio (SUTTON &
HOMAN, 1975; BENJAMIN, 1978; JAIN, 1993).

28
3 OBJETIVOS
3.1 Objetivo geral
Avaliar a influência do sistema de criação sobre os valores leucocitários
sanguíneos, protéicos séricos e fibrinogênio plasmáticos em bovinos.
3.2 Objetivos específicos
Avaliar as alterações leucocitárias circulantes, desencadeadas pelos fatores
ambientais no qual esses animais foram submetidos;
Determinar o nível de proteína total, albumina e globulina de bovinos
terminados a pasto e em confinamento;
Avaliar a relação entre albumina/globulina de bovinos terminados em sistemas
extensivos e em confinamento;
Determinar o nível de fibrinogênio plasmático, de bovinos terminados a pasto e
em confinamento;
Determinar a concentração de proteínas plasmáticas de bovinos terminados a
pasto e em confinamento;
Determinar a relação proteína plasmática: fibrinogênio (PP:F);

29
4 MATERIAL E MÉTODOS 4.1 Local de realização do estudo
O trabalho foi desenvolvido no Laboratório de Análises Clínicas do
Hospital Veterinário (LAC/HV) da Escola de Veterinária da Universidade Federal
de Goiás (EV/UFG) e na Empresa Frigorífica FRIBOI, localizada no município de
Goiânia, Estado de Goiás.
O frigorífico foi selecionado, devido ao grande número de animais
abatidos por dia (1.600), por possuir Serviço de Inspeção Federal (SIF) e ainda
por estar localizado próximo ao laboratório no qual as amostras seriam
processadas.
4.2 Animais
Para se realizar esta pesquisa, foram utilizados 120 bovinos, divididos
em dois grupos, sendo um grupo composto por 60 animais, oriundos de
confinamento e outro grupo de 60 animais, terminados a pasto. Os animais de
ambos os grupos eram machos, idade entre 24 a 36 meses, peso médio de 260
kg (17,33 arrobas) e da raça Nelore (Figura 3).
O tamanho da amostra foi determinado em função da variação dos
leucócitos nesta raça.

30
FIGURA 3 – Animais no curral de espera para abate, em frigorífico, Goiânia, novembro de 2005
4.3 Colheita das amostras
Anteriormente ao abate, os animais foram submetidos ao exame “ante-
mortem”, realizado por médicos veterinários, conforme protocolo estabelecido
pelo SIF.
As amostras sanguíneas eram obtidas no momento da sangria dos
bovinos, coletando-se 5 mL de sangue da jugular, em frasco contendo
anticoagulante EDTA (ácido etilediaminotetracético, sal dissódico a 10%, em
solução aquosa – Vacutainer, Bcton, Dickinson Ind. Cirúrgicas Ltda. Brasil), para
a determinação total e diferencial dos leucócitos, além da determinação do
fibrinogênio plasmático. Do mesmo animal era ainda coletado 5 mL de sangue
sem anticoagulante, para determinação das proteínas total e da albumina sérica
(Figura 4).

31
Após a colheita, as amostras eram devidamente acondicionadas em
caixa de isopor, contendo gelo e transportadas para o LAC/HV, onde eram
processadas e analisadas, no mesmo dia da colheita.
Figura 4 – Colheita de sangue na linha de abate, Goiânia,
novembro de 2005
4.4 Exames laboratoriais
Contagem de leucócitos
Após a homogeneização da amostra, em homogeneizador automático,
o sangue era aspirado em pipetas hematimétricas específicas, para a contagem de
leucócitos e diluído em líquido de Türk e posteriormente contados na câmara de
Neubauer. As contagens foram efetuadas de acordo com as recomendações de
JAIN (1993) e MEYER & HARVEY (2004).
Contagem diferencial de leucócitos
As contagens diferenciais dos leucócitos foram feitas em esfregaços
sanguíneos corados pelo corante Giemsa, conforme JAIN (1993).

32
Determinação das proteínas totais séricas, albumina e globulina
As amostras de sangue colhidas dos animais confinados e terminados a
pasto, foram manipuladas de modo que, nas frações de soro separadas,
determinou-se a proteína total, albumina e imediatamente a globulina.
Para a determinação da proteína sérica total, foi utilizado o método de
biureto, utilizando-se reagente comercial (PROTAL, LABORLAB-Produtos para
Laboratórios Ltda., Guarulhos, São Paulo).
A determinação da albumina foi realizada por meio do mesmo reagente
comercial, adotado para a proteína total, mas utilizando-se o método de verde de
bromocresol.
A globulina foi determinada pelo resultante da subtração de albumina,
da proteína total sérica.
Relação entre albumina/globulina
O índice albumina:globulina (A:G) foi calculado dividindo-se o valor da
fração albumina pelo valor total da fração globulina de cada amostra analisada.
Nível de fibrinogênio plasmático
A determinação do fibrinogênio plasmático foi realizada segundo o
método descrito por KANEKO & SMITH (1964).
Nível da proteína plasmática e relação proteína plasmática: fibrinogênio (PP:F)
A determinação da proteína plamática foi realizada segundo JAIN
(1993). A relação PP:F foi obtida de acordo com a seguinte fórmula (JAIN, 1993):
PP:F = [PP] – [F]
[F]
4.5 Análise estatística
A princípio realizou-se a estatística descritiva dos dados, obtendo-se as
médias, desvio padrão e coeficiente de variação para todos os parâmetros
avaliados, para os dois grupos de animais. Após essa avaliação, optou-se por
aplicar o teste não-paramétrico de WILCOXON para a comparação de médias ao
nível de significância de 5% (SMPAIO, 1998). Essas análises estatísticas foram
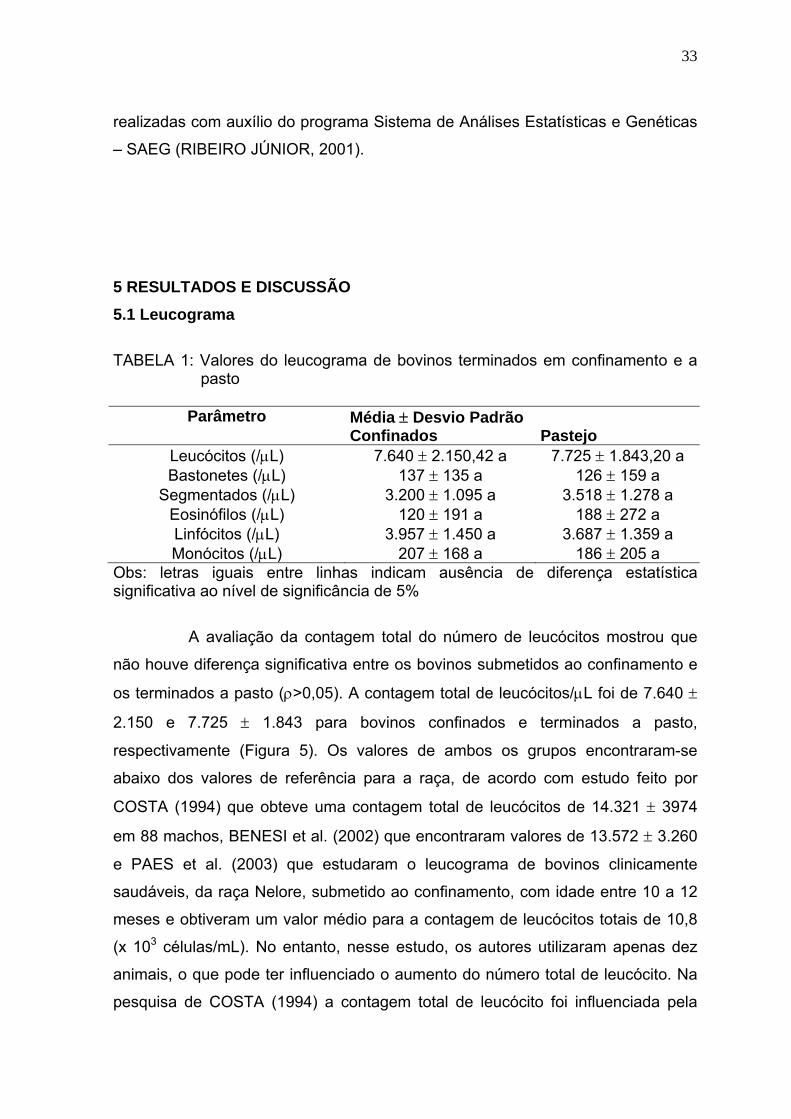
33
realizadas com auxílio do programa Sistema de Análises Estatísticas e Genéticas
– SAEG (RIBEIRO JÚNIOR, 2001).
5 RESULTADOS E DISCUSSÃO 5.1 Leucograma TABELA 1: Valores do leucograma de bovinos terminados em confinamento e a
pasto
Parâmetro Média ± Desvio Padrão Confinados Pastejo
Leucócitos (/μL) 7.640 ± 2.150,42 a 7.725 ± 1.843,20 a Bastonetes (/μL) 137 ± 135 a 126 ± 159 a
Segmentados (/μL) 3.200 ± 1.095 a 3.518 ± 1.278 a Eosinófilos (/μL) 120 ± 191 a 188 ± 272 a Linfócitos (/μL) 3.957 ± 1.450 a 3.687 ± 1.359 a Monócitos (/μL) 207 ± 168 a 186 ± 205 a
Obs: letras iguais entre linhas indicam ausência de diferença estatística significativa ao nível de significância de 5%
A avaliação da contagem total do número de leucócitos mostrou que
não houve diferença significativa entre os bovinos submetidos ao confinamento e
os terminados a pasto (ρ>0,05). A contagem total de leucócitos/μL foi de 7.640 ±
2.150 e 7.725 ± 1.843 para bovinos confinados e terminados a pasto,
respectivamente (Figura 5). Os valores de ambos os grupos encontraram-se
abaixo dos valores de referência para a raça, de acordo com estudo feito por
COSTA (1994) que obteve uma contagem total de leucócitos de 14.321 ± 3974
em 88 machos, BENESI et al. (2002) que encontraram valores de 13.572 ± 3.260
e PAES et al. (2003) que estudaram o leucograma de bovinos clinicamente
saudáveis, da raça Nelore, submetido ao confinamento, com idade entre 10 a 12
meses e obtiveram um valor médio para a contagem de leucócitos totais de 10,8
(x 103 células/mL). No entanto, nesse estudo, os autores utilizaram apenas dez
animais, o que pode ter influenciado o aumento do número total de leucócito. Na
pesquisa de COSTA (1994) a contagem total de leucócito foi influenciada pela

34
elevada contagem de linfócitos. BENESI et al. (2002) atribuíram a elevada
contagem de leucócitos a constante exposição por carrapatos, o que levaria a
uma hemoparasitose enzootica ou natural e contínua premonição dos rebanhos
nas regiões tropicais e subtropicais.
Apesar dos bovinos confinados, sofrerem muitas situações adversas,
tais como, aglomerações dos animais que levam a estresse, infecções por
contágios como a tuberculose, cisticercoses, coccidioses e principalmente
doenças respiratórias como pneumonias (DUTRA, 2001), a contagem total de
leucócitos desses animais estava dentro dos padrões normais. A princípio
acreditava-se que esses grupos poderiam apresentar resposta leucocitária como
aumento ou diminuição dos leucócitos circulantes. RUPPANNER et al. (1978) ao
avaliar o manejo de bovinos destinados à produção de leite e de carne,
encontraram maior contagem total de leucócitos, nos bovinos de abate do que os
bovinos leiteiros, provando que o manejo interfere na quantidade de leucócitos.
No entanto, os autores trabalharam com animais de clima temperado e raças
diferentes da adotada no presente estudo. Acredita-se que os bovinos da raça
Nelore são mais resistentes a adversidades tais como infecção à enfermidades e
infestação por parasitas internos e externos. Portanto a ausência de resposta
leucocitária nesses animais, sobretudo nos animais confinados, pode ser atribuída
ao fator raça.
FIGURA 5 – Média dos valores de leucócitos totais (/μL) de bovinos confinados e a pasto, Goiânia, julho a novembro de 2005
7.5507.6007.6507.7007.750
Confinado Pastejo
Leucócitos totais

35
Exceto pela diferença no tipo de manejo que os animais eram
submetidos, confinados ou a pasto, todas as condições adotadas na pesquisa
foram criteriosamente analisadas com o objetivo de igualar os grupos de animais
e sofrer o mínimo de interferência no leucograma e outros parâmetros estudados.
Portanto, variações leucocitárias descritas por BIRGEL JUNIOR (1991), COSTA
(1994), FAGLIARI et al. (1998b), GONÇALVES et al. (2001) e PAULA NETO
(2004), como idade e raça não ocorreram já que os animais eram pertencentes da
mesma raça e de mesma faixa etária.
Na contagem diferencial dos leucócitos (Tabela 1), não houve diferença
significativa para nenhuma variável estudada (ρ>0,05). Todas as variáveis foram
analisadas pelo seu valor absoluto conforme recomendado por JAIN (1993).
RUPPANER et al. (1978) fizeram uma avaliação do perfil metabólico e celular de
bovinos submetidos ao confinamento, analisando os leucócitos por contagem
diferencial, apesar dos autores não afirmarem tal aspecto; a medodologia adotada
deve ter sido fundamentada nas pesquisas hematológicas daquela ocasião, já
que houveram avanços na hematologia e, atualmente, há consenso quanto à
adoção da contagem diferencial absoluta.
O valor absoluto dos linfócitos/μL foram 3.957 ± 1.449 e 3.687 ± 1.359,
para bovinos confinados e terminados a pasto, respectivamente (Figura 6). Os
valores aqui encontrados estiveram abaixo da referência estabelecida por COSTA
(1994) que foi de 9.940 ± 2.613, de BENESI et al. (2002), que encontraram o valor
de 9.940 ± 3.653 para bovinos Nelore de criação semi-extensiva e de PAES et al.
(2003) que encontraram 7,33 (x103 células/mL) em bovinos da raça Nelore,
submetidos a confinamento. COSTA, 1994 atribuiu o destaque linfocitário da
contagem diferencial celular, à ocorrência de hemoparasitoses e/ou premonição
natural e contínua.
Não foi observado o processo de neutrofilia fisiológica postulado por
MATOS & MATOS (1988), FRASER (1997) e PAES et al. (2003) ainda que os
bovinos do presente estudo antes a colheita, estivessem submetidos a uma
situação de estresse. No entanto, anteriormente à colheita, os animais eram
submetidos ao método de insensibilização, que poderia ter sido atribuído a essa
ausência de neutrofilia.
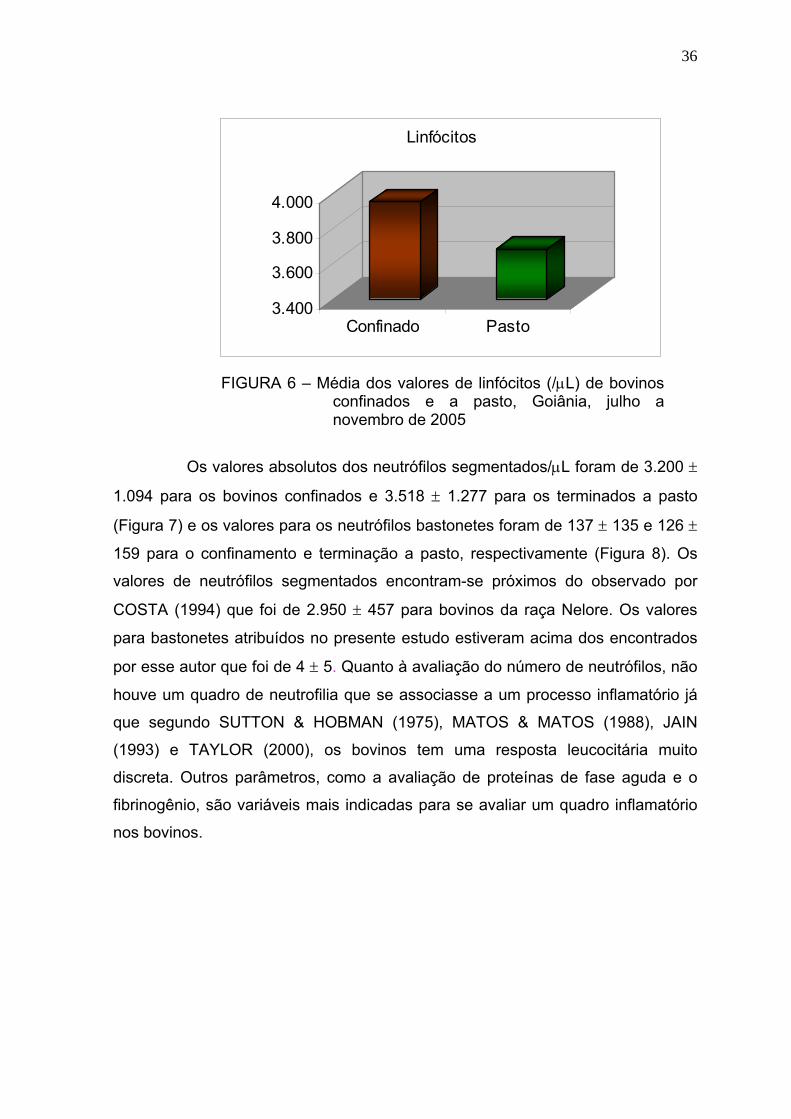
36
3.400
3.600
3.800
4.000
Confinado Pasto
Linfócitos
FIGURA 6 – Média dos valores de linfócitos (/μL) de bovinos
confinados e a pasto, Goiânia, julho a novembro de 2005
Os valores absolutos dos neutrófilos segmentados/μL foram de 3.200 ±
1.094 para os bovinos confinados e 3.518 ± 1.277 para os terminados a pasto
(Figura 7) e os valores para os neutrófilos bastonetes foram de 137 ± 135 e 126 ±
159 para o confinamento e terminação a pasto, respectivamente (Figura 8). Os
valores de neutrófilos segmentados encontram-se próximos do observado por
COSTA (1994) que foi de 2.950 ± 457 para bovinos da raça Nelore. Os valores
para bastonetes atribuídos no presente estudo estiveram acima dos encontrados
por esse autor que foi de 4 ± 5. Quanto à avaliação do número de neutrófilos, não
houve um quadro de neutrofilia que se associasse a um processo inflamatório já
que segundo SUTTON & HOBMAN (1975), MATOS & MATOS (1988), JAIN
(1993) e TAYLOR (2000), os bovinos tem uma resposta leucocitária muito
discreta. Outros parâmetros, como a avaliação de proteínas de fase aguda e o
fibrinogênio, são variáveis mais indicadas para se avaliar um quadro inflamatório
nos bovinos.
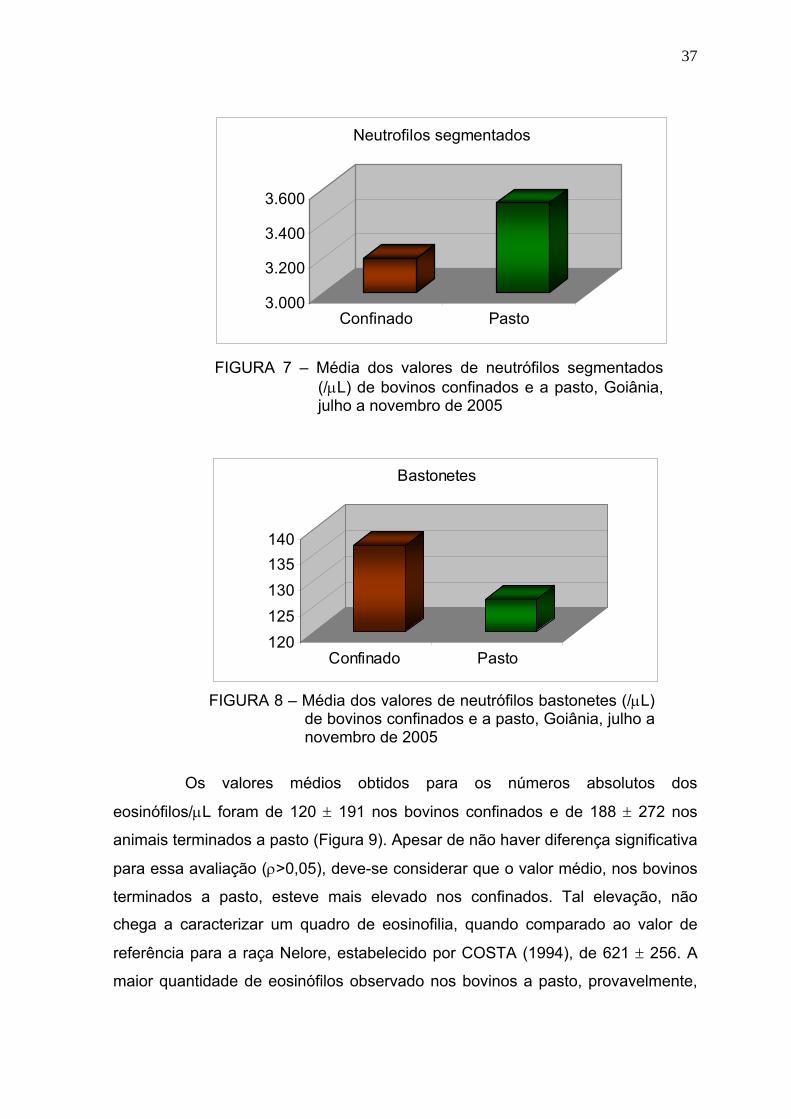
37
3.000
3.200
3.400
3.600
Confinado Pasto
Neutrofilos segmentados
FIGURA 7 – Média dos valores de neutrófilos segmentados
(/μL) de bovinos confinados e a pasto, Goiânia, julho a novembro de 2005
FIGURA 8 – Média dos valores de neutrófilos bastonetes (/μL) de bovinos confinados e a pasto, Goiânia, julho a novembro de 2005
Os valores médios obtidos para os números absolutos dos
eosinófilos/μL foram de 120 ± 191 nos bovinos confinados e de 188 ± 272 nos
animais terminados a pasto (Figura 9). Apesar de não haver diferença significativa
para essa avaliação (ρ>0,05), deve-se considerar que o valor médio, nos bovinos
terminados a pasto, esteve mais elevado nos confinados. Tal elevação, não
chega a caracterizar um quadro de eosinofilia, quando comparado ao valor de
referência para a raça Nelore, estabelecido por COSTA (1994), de 621 ± 256. A
maior quantidade de eosinófilos observado nos bovinos a pasto, provavelmente,
120125130135140
Confinado Pasto
Bastonetes
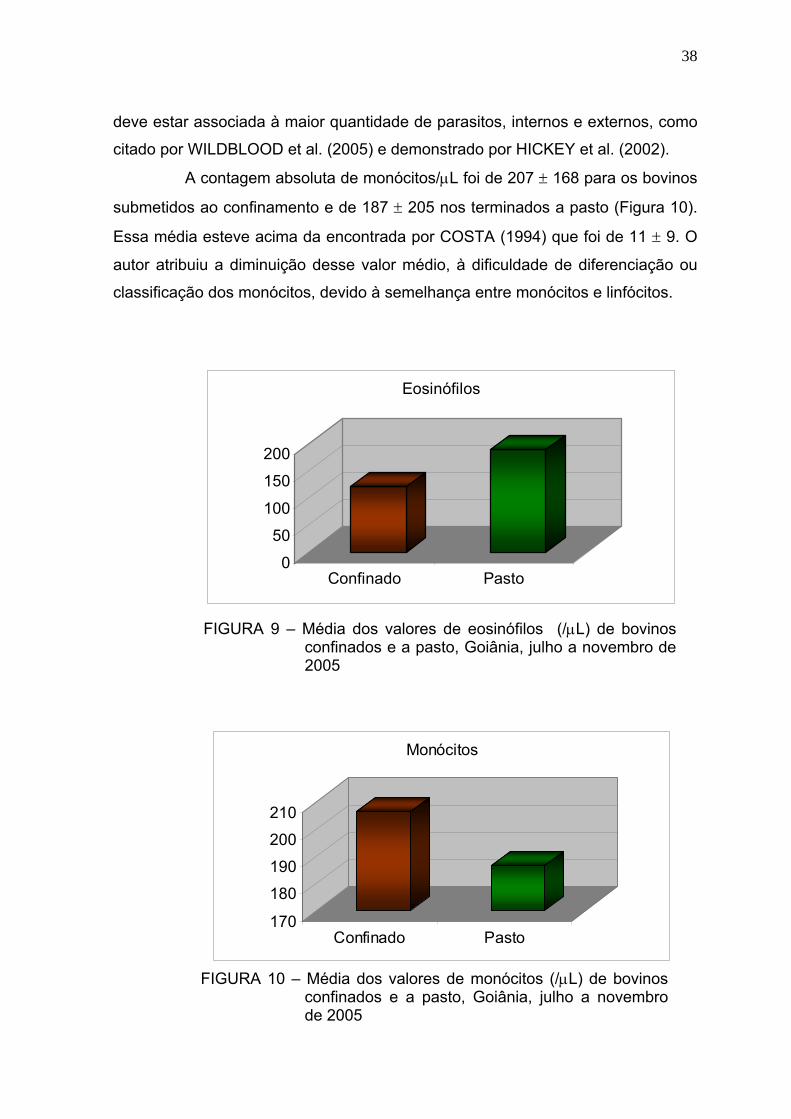
38
deve estar associada à maior quantidade de parasitos, internos e externos, como
citado por WILDBLOOD et al. (2005) e demonstrado por HICKEY et al. (2002).
A contagem absoluta de monócitos/μL foi de 207 ± 168 para os bovinos
submetidos ao confinamento e de 187 ± 205 nos terminados a pasto (Figura 10).
Essa média esteve acima da encontrada por COSTA (1994) que foi de 11 ± 9. O
autor atribuiu a diminuição desse valor médio, à dificuldade de diferenciação ou
classificação dos monócitos, devido à semelhança entre monócitos e linfócitos.
0
50
100
150
200
Confinado Pasto
Eosinófilos
FIGURA 9 – Média dos valores de eosinófilos (/μL) de bovinos
confinados e a pasto, Goiânia, julho a novembro de 2005
FIGURA 10 – Média dos valores de monócitos (/μL) de bovinos
confinados e a pasto, Goiânia, julho a novembro de 2005
170
180
190
200
210
Confinado Pasto
Monócitos
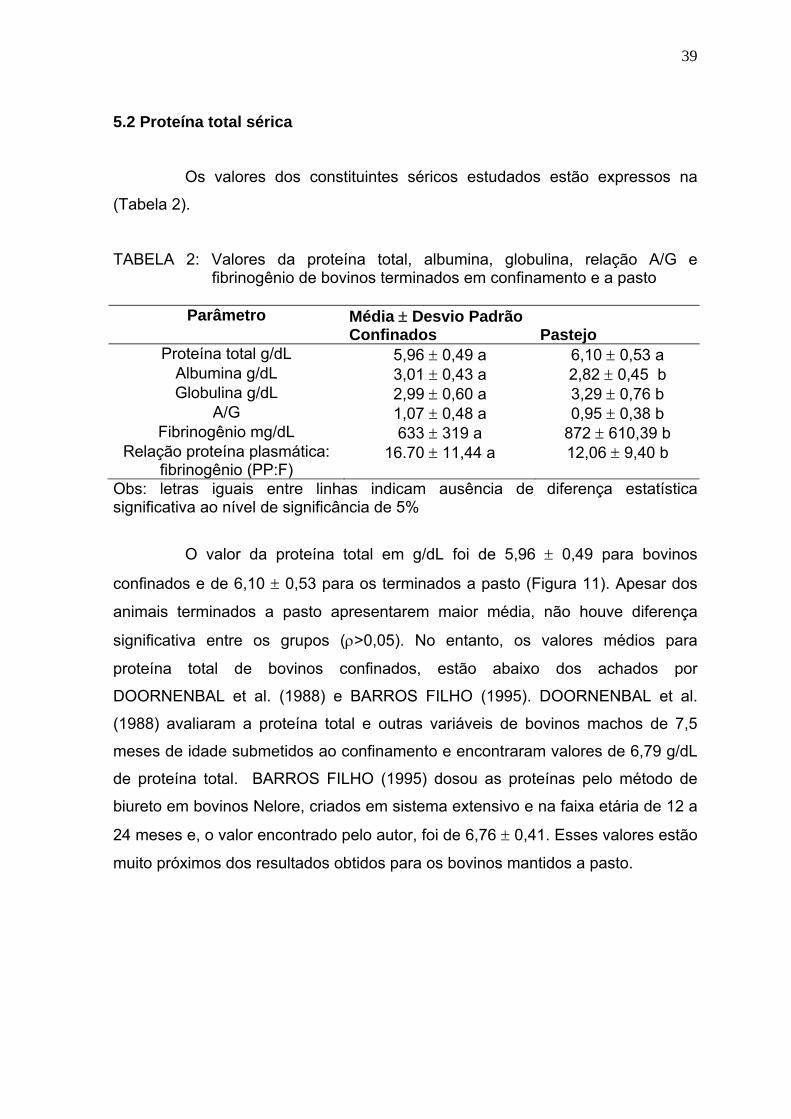
39
5.2 Proteína total sérica
Os valores dos constituintes séricos estudados estão expressos na
(Tabela 2).
TABELA 2: Valores da proteína total, albumina, globulina, relação A/G e fibrinogênio de bovinos terminados em confinamento e a pasto
Parâmetro Média ± Desvio Padrão
Confinados Pastejo Proteína total g/dL 5,96 ± 0,49 a 6,10 ± 0,53 a
Albumina g/dL 3,01 ± 0,43 a 2,82 ± 0,45 b Globulina g/dL 2,99 ± 0,60 a 3,29 ± 0,76 b
A/G 1,07 ± 0,48 a 0,95 ± 0,38 b Fibrinogênio mg/dL 633 ± 319 a 872 ± 610,39 b
Relação proteína plasmática: fibrinogênio (PP:F)
16.70 ± 11,44 a 12,06 ± 9,40 b
Obs: letras iguais entre linhas indicam ausência de diferença estatística significativa ao nível de significância de 5%
O valor da proteína total em g/dL foi de 5,96 ± 0,49 para bovinos
confinados e de 6,10 ± 0,53 para os terminados a pasto (Figura 11). Apesar dos
animais terminados a pasto apresentarem maior média, não houve diferença
significativa entre os grupos (ρ>0,05). No entanto, os valores médios para
proteína total de bovinos confinados, estão abaixo dos achados por
DOORNENBAL et al. (1988) e BARROS FILHO (1995). DOORNENBAL et al.
(1988) avaliaram a proteína total e outras variáveis de bovinos machos de 7,5
meses de idade submetidos ao confinamento e encontraram valores de 6,79 g/dL
de proteína total. BARROS FILHO (1995) dosou as proteínas pelo método de
biureto em bovinos Nelore, criados em sistema extensivo e na faixa etária de 12 a
24 meses e, o valor encontrado pelo autor, foi de 6,76 ± 0,41. Esses valores estão
muito próximos dos resultados obtidos para os bovinos mantidos a pasto.
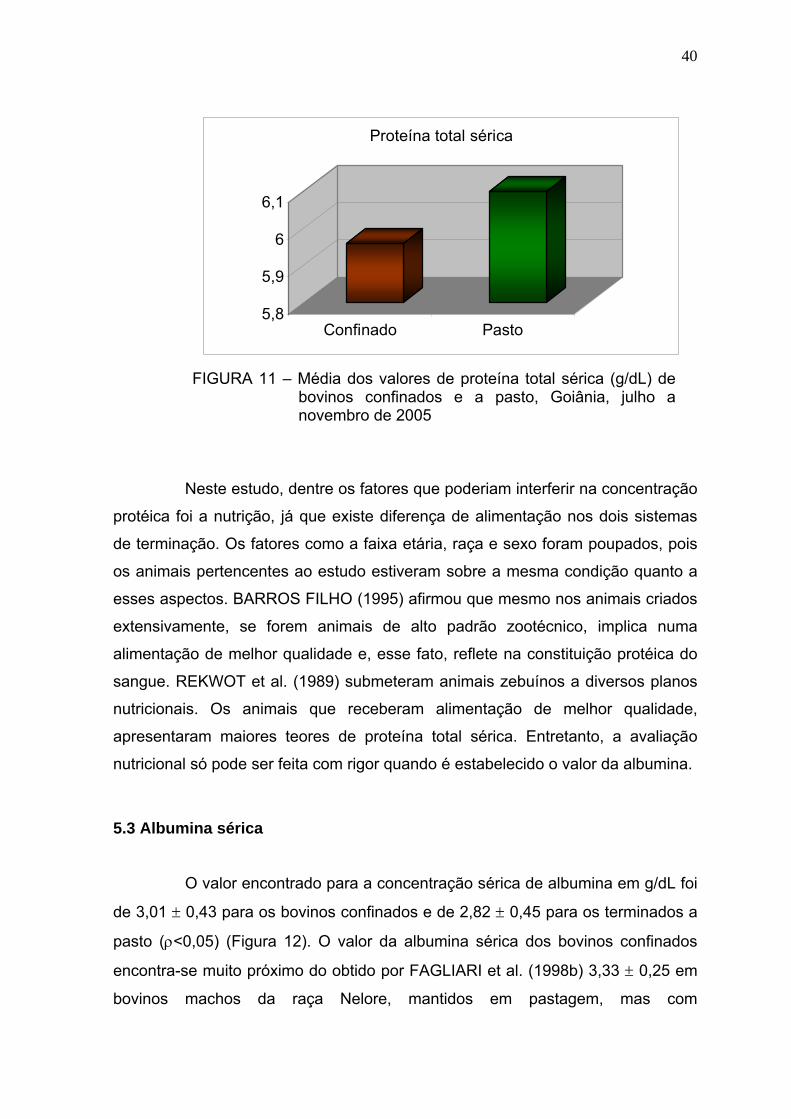
40
5,8
5,9
6
6,1
Confinado Pasto
Proteína total sérica
FIGURA 11 – Média dos valores de proteína total sérica (g/dL) de
bovinos confinados e a pasto, Goiânia, julho a novembro de 2005
Neste estudo, dentre os fatores que poderiam interferir na concentração
protéica foi a nutrição, já que existe diferença de alimentação nos dois sistemas
de terminação. Os fatores como a faixa etária, raça e sexo foram poupados, pois
os animais pertencentes ao estudo estiveram sobre a mesma condição quanto a
esses aspectos. BARROS FILHO (1995) afirmou que mesmo nos animais criados
extensivamente, se forem animais de alto padrão zootécnico, implica numa
alimentação de melhor qualidade e, esse fato, reflete na constituição protéica do
sangue. REKWOT et al. (1989) submeteram animais zebuínos a diversos planos
nutricionais. Os animais que receberam alimentação de melhor qualidade,
apresentaram maiores teores de proteína total sérica. Entretanto, a avaliação
nutricional só pode ser feita com rigor quando é estabelecido o valor da albumina.
5.3 Albumina sérica
O valor encontrado para a concentração sérica de albumina em g/dL foi
de 3,01 ± 0,43 para os bovinos confinados e de 2,82 ± 0,45 para os terminados a
pasto (ρ<0,05) (Figura 12). O valor da albumina sérica dos bovinos confinados
encontra-se muito próximo do obtido por FAGLIARI et al. (1998b) 3,33 ± 0,25 em
bovinos machos da raça Nelore, mantidos em pastagem, mas com

41
suplementação mineral . Quanto aos animais terminados a pasto, os valores da
albumina foram próximos aos encontrados por GONÇALVES et al. (2001) que foi
de 2,92 ± 0,3 para bovinos da raça Guzerá criados a pasto. De acordo com
MEYER & HARVEY (2004) a concentração dessa proteína varia entre as
espécies, mas seu valor sérico ou plasmático pode variar entre 2,5 g/dL a 4,5
g/dL. Os autores acrescentaram ainda que a variabilidade de parâmetros
sanguíneos e bioquímicos podem ser devido a diferenças quanto à raça, estado
fisiológico, composição química do alimento ingerido, temperatura ambiental e
idade do animal. GRUNWALDT et al. (2005) avaliaram alguns parâmetros
bioquímicos como ferramentas para auxiliar no acompanhamento clínico dos
bovinos de corte, além de identificar adversidades na produtividade. Dentre os
parâmetros avaliados, a albumina manteve-se dentro dos padrões normais.
2,7
2,8
2,93
3,1
Confinado Pasto
Albumina sérica
FIGURA 12 – Média dos valores de albumina sérica (g/dL) de
bovinos confinados e a pasto, Goiânia, julho a novembro de 2005
Como os bovinos confinados recebem ração balanceada para atender
suas necessidades metabólicas e assim favorecer o ganho em peso (CARDOSO,
2000), a elevação da concentração de albumina nos animais confinados,
comparados aos criados a pasto, foi influenciada pelo estado de nutrição, como
enfatizado por JAIN (1993) e GARCÉS et al. (2001). GIBES et al. (1999),
recomendam a utilização da avaliação sérica da albumina, rotineiramente, como
uma ferramenta de diagnóstico para detectar estados de mal nutrição e outras
condições adversas.

42
Por outro lado, a concentração de albumina em menor quantidade, nos
bovinos terminados a pasto, pode ter sido influenciada pelo aumento da produção
de fibrinogênio nessa categoria de animal, pois segundo GARCÉS et al. (2001), a
síntese da albumina é inversamente proporcional à síntese das proteínas de fase
aguda.
Os resultados da albumina sérica de animais terminados a pasto foram
menores dos encontrados por BARROS FILHO (1995), em animais criados
extensivamente, que foi de 3,12 ± 0,50. No entanto o autor utilizou a migração
eletroforética para a determinação da albumina e frações da globulina. GREEN et
al. (1982), utilizaram métodos químicos como biureto, para proteína total e verde
de bromocresol para avaliar albumina. Paralelamente comparou com à
eletroforese. A proteína total, a albumina, bem como a razão A/G, foram
semelhantes pelos dois métodos para os bovinos adultos.
5.4 Globulina e relação A/G séricas
A concentração sérica de globulina em g/dL foi mais elevada nos
bovinos terminados a pasto, de 3,29 ± 0,76 (ρ<0,05). Os bovinos confinados
apresentaram uma concentração de 2,99 ± 0,60 (Figura 13). Esse valor esteve
mais próximo dos encontrados por CHORFI et al. (2004), que foi de 2,35 g/dL,
sendo que os autores avaliaram vacas em lactação e, obteveram as amostras
sanguíneas para a análise na veia jugular. Nesse mesmo estudo, o valor da
amostra coletada da veia coccígena, foi menor. Segundo os autores, o local da
coleta das amostras deve ser levado em consideração na interpretação da
globulina sérica. O valor da concentração de globulina, dos bovinos terminados a
pasto, foi próximo ao encontrado por FAGLIARI et al. (1998), que foi de 3,80 ±
0,48, para bovinos Nelore criados também em sistema extensivo.
A concentração de globulina nesse estudo foi maior do que a globulina
nos animais a pasto, que foi 3,29 ± 0,76 e 2,82 ± 0,45, respectivamente.
Concordando com RAMÍREZ et al. (2001), que atribuem o aumento da
concentração de proteínas plasmáticas à resposta imune do organismo do animal
às diversas infecções, além do fato, da concentração de globulina nos animais em

43
condições tropicais, ser proporcionalmente maior do que a concentração de
albumina.
2,8
3
3,2
3,4
Confinado Pasto
Globulina sérica
FIGURA 13 – Média dos valores da concentração de
globulina sérica (g/dL) de bovinos confinados e a pasto, Goiânia, julho a novembro de 2005
A relação A/G apresentou valores mais elevados nos animais
confinados 1,07 ± 0,48 (ρ<0,05). No grupo de animais terminados a pasto foi
obtida uma relação A/G de 0,95 ± 0,38 (Figura 14), próximo ao encontrado por
KAWANO et al. (1997) que foi entre 0,94 e 1,01, apesar dos autores realizarem o
estudo em fêmeas. BARROS FILHO (1995) não encontrou diferença da relação
A/G, inerentes ao sexo dos bovinos.
A razão da elevação do índice A/G foi atribuída ao aumento da
concentração de albumina nos animais confinados. No estudo realizado por
BARROS FILHO (1995), com animais hígidos da raça Nelore, criados em sistema
extensivo, verificou-se que na faixa etária de 24 a 60 meses, a relação (A/G) foi
de 0,81, menores portanto do que a dos animais no mesmo sistema de
terminação da presente pesquisa. Segundo observações feitas por BACILA
(2003), a relação A/G é importante na avaliação de infecções, quando elas se
alteram, invertendo-se os valores pelo incremento que ocorre na concentração
das imunoglobulinas, especialmente as γ-globulinas. WILLARD (2000), acrescenta
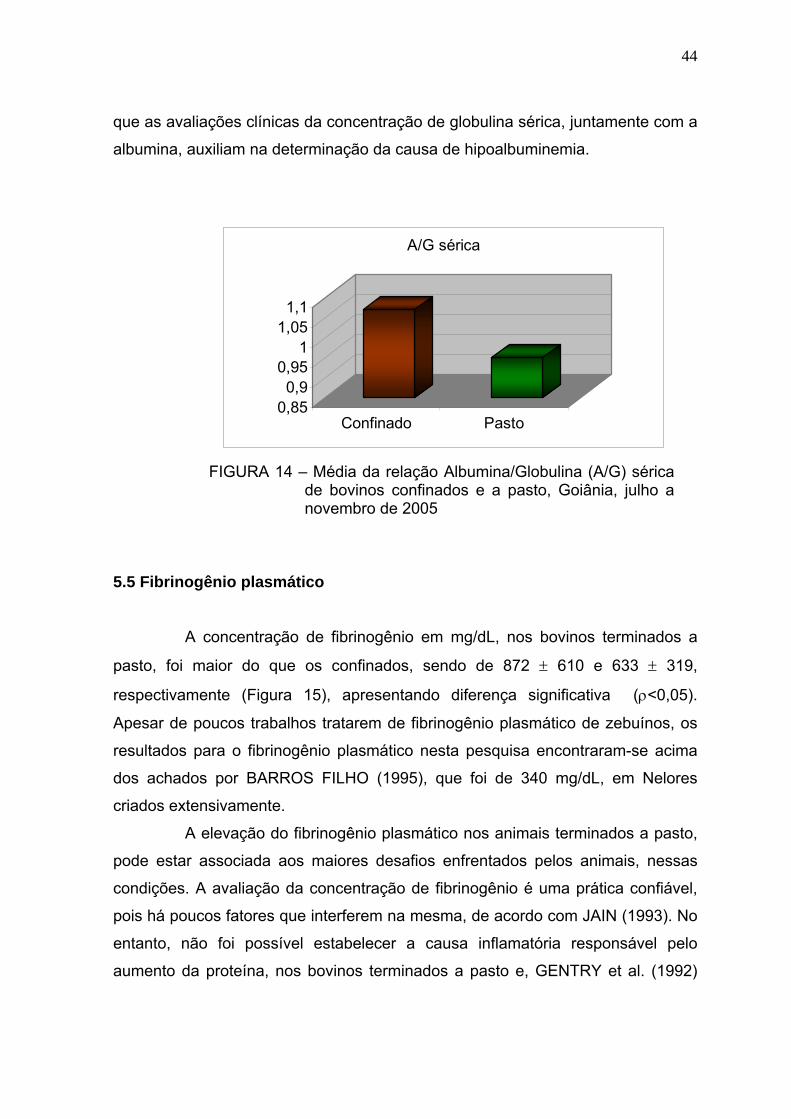
44
que as avaliações clínicas da concentração de globulina sérica, juntamente com a
albumina, auxiliam na determinação da causa de hipoalbuminemia.
0,850,9
0,951
1,051,1
Confinado Pasto
A/G sérica
FIGURA 14 – Média da relação Albumina/Globulina (A/G) sérica
de bovinos confinados e a pasto, Goiânia, julho a novembro de 2005
5.5 Fibrinogênio plasmático
A concentração de fibrinogênio em mg/dL, nos bovinos terminados a
pasto, foi maior do que os confinados, sendo de 872 ± 610 e 633 ± 319,
respectivamente (Figura 15), apresentando diferença significativa (ρ<0,05).
Apesar de poucos trabalhos tratarem de fibrinogênio plasmático de zebuínos, os
resultados para o fibrinogênio plasmático nesta pesquisa encontraram-se acima
dos achados por BARROS FILHO (1995), que foi de 340 mg/dL, em Nelores
criados extensivamente.
A elevação do fibrinogênio plasmático nos animais terminados a pasto,
pode estar associada aos maiores desafios enfrentados pelos animais, nessas
condições. A avaliação da concentração de fibrinogênio é uma prática confiável,
pois há poucos fatores que interferem na mesma, de acordo com JAIN (1993). No
entanto, não foi possível estabelecer a causa inflamatória responsável pelo
aumento da proteína, nos bovinos terminados a pasto e, GENTRY et al. (1992)
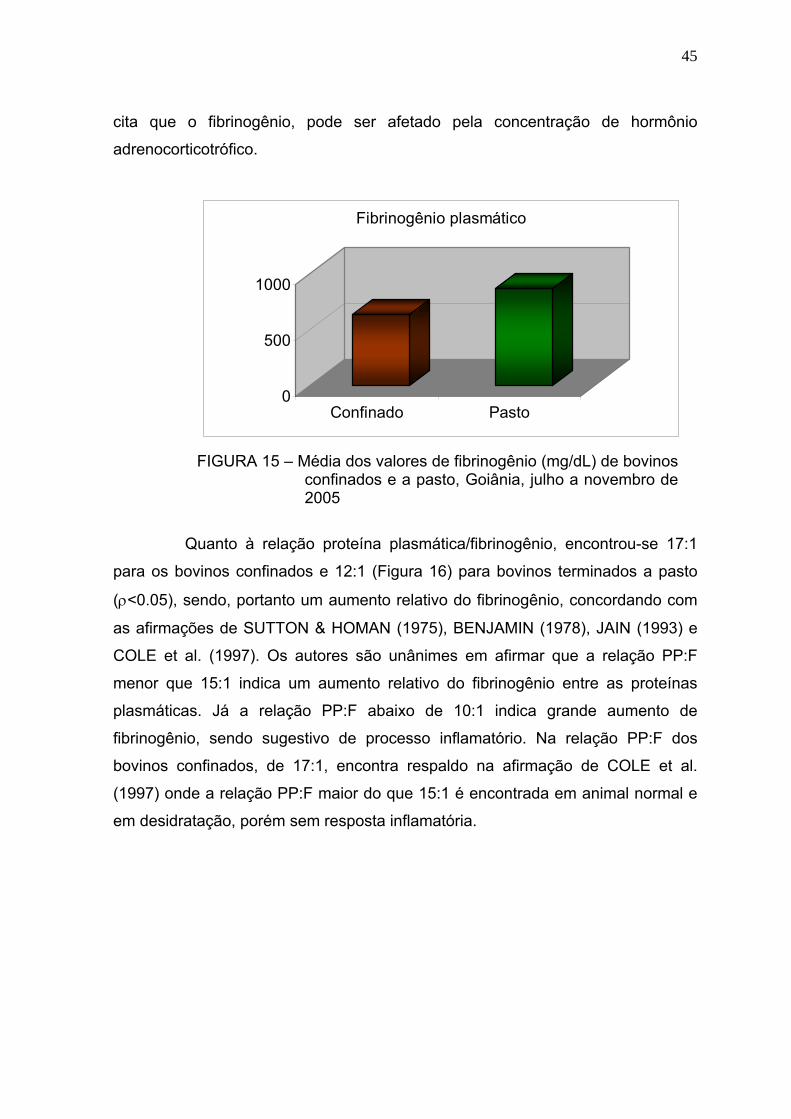
45
cita que o fibrinogênio, pode ser afetado pela concentração de hormônio
adrenocorticotrófico.
0
500
1000
Confinado Pasto
Fibrinogênio plasmático
FIGURA 15 – Média dos valores de fibrinogênio (mg/dL) de bovinos
confinados e a pasto, Goiânia, julho a novembro de 2005
Quanto à relação proteína plasmática/fibrinogênio, encontrou-se 17:1
para os bovinos confinados e 12:1 (Figura 16) para bovinos terminados a pasto
(ρ<0.05), sendo, portanto um aumento relativo do fibrinogênio, concordando com
as afirmações de SUTTON & HOMAN (1975), BENJAMIN (1978), JAIN (1993) e
COLE et al. (1997). Os autores são unânimes em afirmar que a relação PP:F
menor que 15:1 indica um aumento relativo do fibrinogênio entre as proteínas
plasmáticas. Já a relação PP:F abaixo de 10:1 indica grande aumento de
fibrinogênio, sendo sugestivo de processo inflamatório. Na relação PP:F dos
bovinos confinados, de 17:1, encontra respaldo na afirmação de COLE et al.
(1997) onde a relação PP:F maior do que 15:1 é encontrada em animal normal e
em desidratação, porém sem resposta inflamatória.
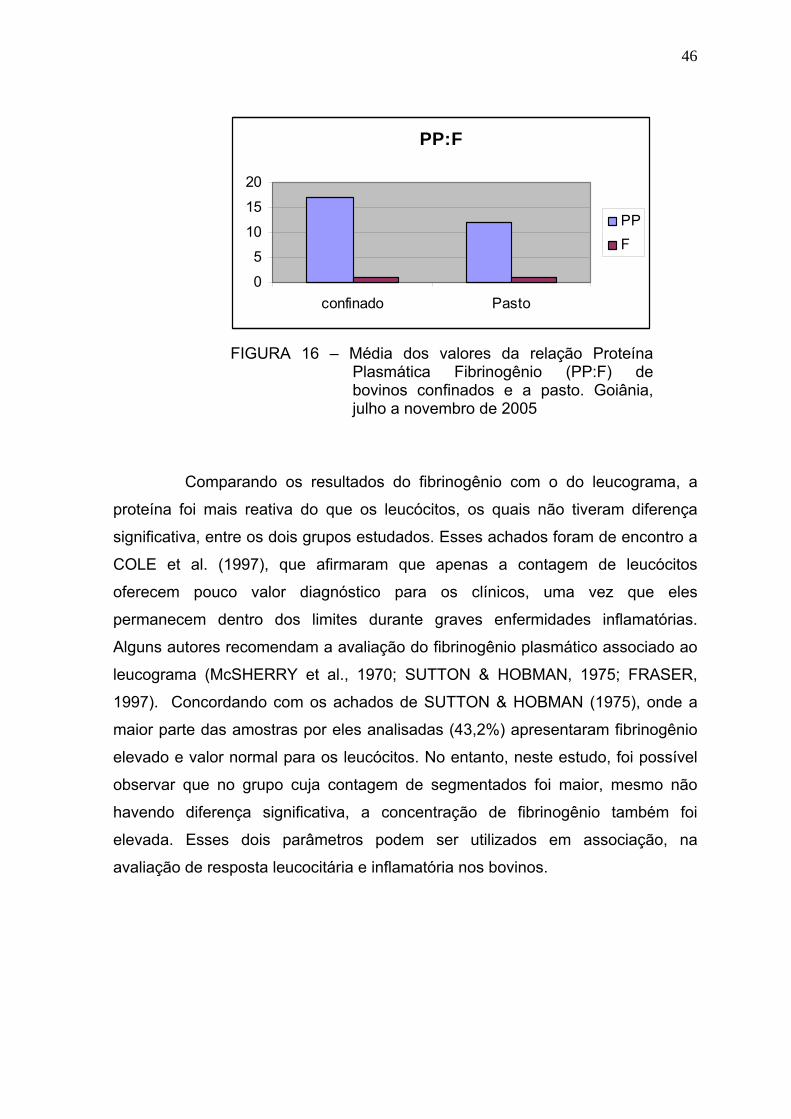
46
PP:F
0
5
10
15
20
confinado Pasto
PPF
FIGURA 16 – Média dos valores da relação Proteína
Plasmática Fibrinogênio (PP:F) de bovinos confinados e a pasto. Goiânia, julho a novembro de 2005
Comparando os resultados do fibrinogênio com o do leucograma, a
proteína foi mais reativa do que os leucócitos, os quais não tiveram diferença
significativa, entre os dois grupos estudados. Esses achados foram de encontro a
COLE et al. (1997), que afirmaram que apenas a contagem de leucócitos
oferecem pouco valor diagnóstico para os clínicos, uma vez que eles
permanecem dentro dos limites durante graves enfermidades inflamatórias.
Alguns autores recomendam a avaliação do fibrinogênio plasmático associado ao
leucograma (McSHERRY et al., 1970; SUTTON & HOBMAN, 1975; FRASER,
1997). Concordando com os achados de SUTTON & HOBMAN (1975), onde a
maior parte das amostras por eles analisadas (43,2%) apresentaram fibrinogênio
elevado e valor normal para os leucócitos. No entanto, neste estudo, foi possível
observar que no grupo cuja contagem de segmentados foi maior, mesmo não
havendo diferença significativa, a concentração de fibrinogênio também foi
elevada. Esses dois parâmetros podem ser utilizados em associação, na
avaliação de resposta leucocitária e inflamatória nos bovinos.

47
6. CONCLUSÃO
A avaliação dos resultados dos leucogramas, albumina, globulina,
relação A/G e fibrinogênio de bovinos sadios confinados e terminados a pasto
possibilitaram as seguintes conclusões:
1. Os manejos de confinamento e a pasto não influenciam na contagem total
e diferencial de leucócitos;
2. A concentração sérica média de albumina dos animais confinados é maior
do que as dos animais terminados a pasto;
3. A concentração de globulina sérica e o índice A/G dos animais terminados
a pasto são maiores do que nos confinados;
4. O nível médio do fibrinogênio plasmático, de bovinos terminados a pasto, é
mais elevado do que nos animais em confinamento.
5. A concentração de proteínas plasmáticas e a relação PP:F é maior nos
animais confinados do que nos animais terminados a pasto.

48
REFERÊNCIAS BIBLIOGRÁFICAS 1. ACKERMANN, M.R.; KEHRLI, M.E.; LAUFER, J.A.; NUSZ, L.T. Alimentary and respiratory-trat lesions in 8 medically fragili Holstein cattle with bovine leukocyte adhesion deficiency (BLAD). Veterinary Pathology, Boston, v.33, p.273-281, 1996. 2. AGER, A. Regulation of lymphocyte migration into lymph by high endothelial venules. Biochemical Society Transactios, London, v.25, p.421-428, 1996.
3. ALBELDA, S.M.; SMITH, C.W.; WARD, P.A. Adhesion molecules and inflammatory injury. The FASEB Journal, Bethesda, v.8, p.504-512, 1994.
4. ANDREASEN, C.B.; ROTH, J.A. Neutrophil functional abnormalities. In: FELDMAN, B.F.; ZINKL, J.G.; JAIN, N.C. Schalm’s Veterinary Hematology. Lippincott, Williams and Williams: Philadelphia, p.356-365, 2000.
5. ANDRESEN, H.A. Evaluation of leukopenia in cattle. Journal American Veterinary Medical Association, Schaumburg, v.156, n.7, p.858-867, 1970.
6. ANUALPEC - ANUÁRIO DA PECUÁRIA BRASILEIRA, FNP Consultoria & Comércio NEHMI, J.M.D.; NEHMI FILHO, V.A.; FERRAZ, J.V. (Coord). São Paulo: Argos, 2005, 485p.
7. ARNAOUT, M.A. Structure and function of the leukocyte adhesion molecules CD11/CD18. Blood, Washington, v.75, n.5, p.1037-1050, 1990.
8. BACILA, M. Bioquímica Veterinária. São Paulo:Robe, 2 ed. 2003, p.583. 9. BAGGIOLINI, M.; WALS, A.; KUNKEL, S.L. Neutrophil-activating peptide-1/interleukin 8, a novel citokine that ativates neutrophil. The Journal of Clinical Investigation, Michigan, v.84, p.1045-1053, 1989.
10. BALLMER, P.E.; IMOBERDORF, R.; ZBÄREN, J. Immunohistochemical demonstration of interleukin-1β induced changes in acute-phase proteins and albumin in rat liver. Acta Histochemica, Löbdegraben, v.97, p. 281-289, 1995.
11. BARROS FILHO, I.R. Contribuição ao estudo da bioquímica clínica em zebuínos da raça Nelore criados no estado de São Paulo: Influência dos fatores etários e do tipo racial. São Paulo, 1995, 132p. Dissertação (Mestrado) Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo.
12. BENESI, F.J.; COSTA, J.N.; BIRGEL, E.H.; D’ANGELINO, J.L.; AYRES, M.C.C.; FILHO, I.R.B. Leucograma padrão de bovines da raça Nelore (Bos indicus) influência de fatores sexuais. Veterinária Notícias, Uberlândia, v.8, n.1, p.59-66, 2002.

49
13. BENJAMIN, M.M. Interpretation of hematologic findings. Outline of Veterinary Clinical Pathology. 3 ed. Ames: Iowa State University, 1978, p.76-107.
14. BERNOFSKY, C. Nucleotide chloramines and neutrophil-mediated cytotoxidade, The FASEB Journal, Bethesda, v.5, p.295-300, 1991.
15. BERRY, B.A.; CONFER, A.W.; KREHBIEL, C.R.; GILL, D.R.; SMITH, R.A.; MONTELONGO, M. Effects of dietary energy and starch concentrations for newly received feedlot calves: II Acute-phase protein response. Journal of Animal Science, Champaign, v.82, p.845-850, 2004.
16. BIRGEL JUNIOR, E.H. O hemograma de bovinos (Bos taurus, Linnaeus, 1758), da raça Jersey, criados no Estado de São Paulo. 1991, 172f. Dissertação (Mestrado em Patologia Bovina) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo.
17. BREEZE, R. Parasitic Bronchitis and Pneumonia. The Veterinary Clinics North America: Food Animal Practice, Philadelfia, v.1, p.277-87, 1985.
18. BREEZE, R. Structure, function, and metabolism in the lung. The Veterinary Clinics North America: Food Animal Practice, Philadelfia, v.1, p.219-35, 1985.
19. BREIDER, M.A.; WALKER, R.D.; HOPKINS, F.M.; SCHULTZ, T.W.; BOWERSOCK, T.L. Pulmonary lesions induced by Pasteurella haemolytica in neutrophil sufficient and neutrophil deficient calves. The Canadian Journal Veterinary Research, Ottawa, v.52, p.205-9, 1988.
20. BROWN, T.T.; ANANABA, G. Effect of respiratory infections caused by bovine herpesvirus-1 or parainfluenza-3 virus on bovine alveolar macrophage functions. American Journal of Veterinary Rearch, Chicago, v.49, p.1447-1451, 1988.
21. BRYSON, D.G.; McNULTY, M.S.; McCRACKEN, R.M.; CUSH, P.F. Ultrastructural features of experimental parainfluenza type 3 virus pneumonia in calves. Journal Comparative Pathology, London, v.93, p.397-414, 1983.
22. BURVENICH, C.; PAAPE, M.J.; HILL, A.W.; GUIDRY, A.J.; MILLER, R.H.; HEYNEMAN, R.; KREMER, W.D.J.; BRAND, A. Role of the neutrophil leukocyte in the local and systemic reactions during experimentally indiced E. coli mastitis in cows immediately after calving. The Veterinary Quarterly, Ultrecht, v.16, p.45-50, 1994.
23. CANAVESSI, A.M.O.; CHIACCHIO, S.B.; SARTORY, R.; CURY, P.R. Valores do perfil eletroforético das proteínas séricas de bovinos da raça Nelore (Bos indicus) criados na região de Botucatu, são Paulo: influência dos fatores etários e sexuais. O Biológico, São Paulo, v.67, p.9-17, 2000.

50
24. CARDOSO, E. G. Confinamento de bovinos, 2000. Disponível em http://cnpgc.embrapa.br/publicacoes/naoseriadas/cursosuplementacao/confinamento. Acesso em 14/06/2004. 25. CASTLEMAN, W.L.; LAY, J.C.; DUBOVI, E.J.; SLAUSON, D.O. Experimental bovine respiratory syncicial virus infection in conventional calves: light microscopic lesions, microbiology, and studies on lavaged lung cells. American Journal of Veterinary Research, Chicago, v.46, p.547-54, 1985.
26. CAVERLY, J.M.; DIAMOND, G.; GALLUO, J.M.; BROGDEN, K.A.; DIXON, R.A.; ACKERMANN, M.R. Coordinated expressions of tracheal antimicrobial peptide and inflammatory-response elements in the lungs. Infection and Immunity, Washington, v.71, n.5, p.2950-2955, 2003.
27. CHORFI, Y.; LANEVSCHI-PIETERSMA, A.; GIRARD, V.; TREMBLAY, A. Evaluation of variation n serum globulin concentrations in dairy cattle. Veterinary Clinical Pathology, Davis, v.33, n.3, p.122-127, 2004.
28. COHEN, A.B.; ROSSI, M. Neutrophils in normal lungs. The American Review of Respiratory Disease, Baltimore, v.127, p.83-9, 1983.
29. COLE, D.C.; ROUSSEL, A.J.; WHITNEY, M.S. Interpreting a bovine CBC:Evaluation the leukon and acute-phase proteins. Veterinary Medicine, London, v.92, n.5, p.470-478, 1997.
30. COLE, N.A.; CAMP, T.H.; ROWE JR, L.D.; STEVENS, D.G.; HUTCHESON, D.P. Effect of transport on feeder calves. American Journal of Veterinary Research, Chicago, v.49, n.2, p.178-1833, 1988.
31. COSTA, J.N. Leucograma de zebuínos (Bos indicus, Linnaeus, 1758) sadios da raça Nelore criados no Estado de São Paulo: Influência dos fatotes etários e sexuais. São Paulo, 1994, 124p. Dissertação (Mestrado) Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo. (1994).
32. COTRAN, R.S.; KUMAR, V.; COLLINS, T. Robbins: Patologia estrutural e functional. 6 ed. Rio de Janeiro: Guanabara Koogan, 2004, 1251p.
33. CROCKETT-TORABI, A.; FANTONE, J.C. The selectins: Insights into selectin-induced intracellular signaling in leukocytes. Immunologic Research, Mississippi, v.14, p.237-251, 1995.
34. CUNHA, P.H.J.; ARAÚJO, G.R.S.; SILVA, L.A.F.; SILVA, O.C.; FRANÇA, T.C.M.; GONZAGA, B.C.F.; ABUD, L.J.; COSTA, G.L. Saturnismo em bovino: relato de caso. In: CONGRESSO PAULISTA DE MEDICINA VETERINÁRIA, 6. 2004, Santos. Anais eletrônicos…[online]. Disponível em http://spmv.org.br/conpavet2004/trabalhos-buiatria093.htm.

51
35. DELDAR, A. Effects of Escherichia coli endotoxin on leukocyte and platelet counts, fibrinogen concentrations and blood clotting in colostrums-fed and colostrums-deficient neonatal calves. American Journal of Veterinary Research, Chicago, v.45, n.4, p.670-677, 1984.
36. DIESEL, D.A.; LEBEL, J.L.; TUCKER, A. Pulmonary particle deposition and airway mucociliary clearance in cold-exposed calves. American Journal Veterinary Research, Chicago, v.52, p.1665-71, 1991.
37. DIXON, P.M. Respiratory mucociliary clearance in the horse in health and disease, and its pharmaceutical modification. Veterinary Record, London, v.131, p.229-35, 1992.
38. DOORNENBAL, H.; TONG, AKW.; MURRAY, N.L. References values of blood parameters in beef cattle of different ages and stages of lactation. Canadian Journal Veterinary Research, Ottawa, v.52, p.99-105, 1988.
39. DRANSFILD, I.; CABANAS, C.; BARRET, J.; HOGG, N. Interation of leukocyte integrins with ligand is necessary but not sufficient for function. The Journal Cell Biology, New York, v.116, p.1527-1535, 1992.
40. DUNCAN, J.R.; PRASSE, K.W.; MAHAFFEY, E.A. Veterinary Laboratory Medicine. 3 ed. Ames IA: Iowa State University Press, 1994, p.112-118.
41. DUTRA, I.S. 2001; Programa sanitário para confinamentos In: ANAIS DO SEMINÁRIO INTERNACIONAL NUTRON SOBRE NUTRIÇÃO DE BOVINOS, 3. Goiânia, 2001.
42. DUTRA, I.S. Pneumonia verminótica no gado de corte. [online] Disponível em http://www.beefpoint.com.br/bn/radarestecnicos/artigo.asp. Acesso em 15/10/2005).
43. DYER, R.M.; ERNEY, S.; SPENCER, P.; BENSON, C.E. Oxidative metabolism of the bovine alveolar macrophage. American Journal Veterinary Research, Chicago, v.50, p.448-54, 1989.
44. ECKERSALL, P.D. Acute phase proteins as markers of inflammatory lesions. Comparative Heamatology International, London, v.5, p.93-97, 1995.
45. ECKERSALL, P.D; CONNER, J.G. Bovine and canine acute phase proteins. Vet Res Commum, v.12, p.169-178, 1988.
46. ESPINOSA, O.R.; PAREDES, P.A. Fagocitosis: mecanismos y consequencias – Primeira parte. Bioquimia, Ciudad del Mexico, v.28, n.4, p.19-30, 2003.
47. EYSKER, M. Dictyocaulosis in cattle. Compendium on Continuing Education for the Practicing Veterinarian, Quebec, v.16, p.669-672, 1994.

52
48. FAGLIARI, J.J. Estudo clínico e laboratorial da pneumonia de bezerros induzida pela inoculação intrabronquial de Mannheimia haemolytica. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, Belo Horizonte. v.55, p. 8-14, 2003.
49. FAGLIARI, J.J.; SANTANA, A.E.; LUCAS, F.A.; CAMPOS FILHO, E.; CURI, P.R. Constituintes sanguíneos de bovinos lactentes, desmamados e adultos das raças Nelore (Bos indicus) e Holandesa (Bos taurus) e de bubalinos (Bos bubalis) da raça Murrah. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, Belo Horizonte. v.50, n.3, p.263-271, 1998b.
50. FAGLIARI, J.J.; SANTANA, A.E.; MARCHIÓ, W.; CAMPOS FILHO, E.; CURI, P.R. Constituíntes sanguíneos de vacas da raça Nelore (Bos indicus) e Holandesa (Bos taurus) e de bubalinos (Bos bubalis) da raça Murrah durante a gestação, no dia do parto e no puerpério. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, Belo Horizonte. v.50, n.3, p.273-282, 1998a.
51. FARBER, J.L.; KYLE, M.E.; COLEMAN, J.B. Mechanisms of cell injury by activated oxygen species. Laboratory Investigation, New York, v.62, p.670-79, 1990.
52. FRANK, G.H.; TABATABAI, L.B. Neuraminidase activity of Pasteurella hemolytica isolates. Infect Immunology, v.32, p.1119-1122, 1991.
53. FRASER, C.M. Manual Merck de Veterinária. 7 ed. São Paulo: ROCA, 1997, 2169p.
54. FULTON, R.W.; COOK, B.J.; STEP, D.L.; CONFER, A.W.; SALIKI, J.T.; MARK, E.P.; BURGE, L.J.; WELSH, R.D.; BLOOD, K.S. Evaluation of health status of calves and impact on feedlot performance: assessment of a retained ownership program for postweaning calves. The Canadian Journal of Veterinary Research, Ottawa, v.66, p.173-180, 2002.
55. GAMBLE, J.R.; VADAS, M.A. Endothelial adhesiveness for blood neutrophils in inhibited by transforming growth factor-beta. Science, Washington, v.242, p.97-99, 1988.
56. GARCÉS, M.L.V.; RIVERO, H.F.; RIOS, G.R.; QUIÑONES, R.N.; GARCIA, L.C.; BLANCO, E.S. Observaciones hematoquímicas em um rebanho de vacas lecheras com freqüentes estados de anemia. Ver. Prod. Anim, v.13, n.1, p.83-88, 2001.
57. GENTRY, P.A; LIPTRAP, R.M.; TREMBLAY, R.R.; LICHEN, L.; ROSS, M.L. Adrenocorticotrophic hormone fails to alter plasma fibrinogen and fibronectin values in calves but does so in rabbits. Veterinary Research Communications, Netherlands, v.16, n.4, p.253-264, 1992.

53
58. GERARDI, A.S. Bovine leukocyte adhesion deficiency: a review of a modern disease and its implications. Research Veterinary Science, Saskatoon, v.6, p.183-186, 1996.
59. GIBES, J.; CULL, W.; HENDERSON, W.; DALEY, J.; HUR, K.; KHURI, S.F. Preoperative serum albumin level as a predictor of operative mortality and morbidity. Archives of Surgery, Chicago, v.134, p.36-42, 1999. 60. GOMES, M.J.M. Ambiente e pulmão. Jornal de Pneumologia, São Paulo, v.28, n.5, p.261-269, 2002.
61. GONÇALVES, R.C.; PAES, P.R.O.; ALMEIDA, C.T.; FONTEQUE, J.H.; LOPES, R.S.; KUCHEMBUCK, M.R.G.; CROCCI, A.J. Influência da idade e do sexo sobre o hemograma, proteínas séricas totais, albumina e globulina de bovinos sadios da raça Guzerá (Bos indicus). Veterinárias Notícias, Uberlândia, v.7, n.1, p.61-68, 2001.
62. GREEN, S.A.; JEMKINS, S.J.; CLARK, P.A. A comparison of chemical and eletrophoretic methods of serum protein determination in clinically domestic animals of various age. Cornell Veterinarian, New York, v.72, n.4, p.416-426, 1982.
63. GRIFFIN, D. Economic impact associated with respiratory disease in beef cattle. The Veterinary Clinics North American Food Animal Practices, Philadelphia, v.13, p.367-377, 1997.
64. GRÜNWALDT, E.G.; GUEVARA, J.C.; ESTÉVEZ, O.R.; VICENTE, A.; ROUSSELLE, H.; ALCUTEN, N.; AGUERREGARAY, D.; STASI, C.R. Biochemical and heamatological measurements in beef cattle in Mendonza Plain Rangelands (Argentina). Tropical Animal Helth and Production, Netherlands, v.37, p.527-540, 2005.
65. HARVATH, L. Neutrophil chemiotatic factors. Experientia Supplementarum, v.59, p.35-52, 1991.
66. HARVEY, J.W.; WEST, C.L. Predinisone-induced increases in serum alpha-2-globulin and haptoglobin concentrations in dog. Veterinary Pathology, Boston, v.24, p.90-92, 1987.
67. HICKEY, M.C.; EARLEY, B.; FRENCH, P.; KAENE, M.G. Evaluation of parasite control programmes in beef cattle, Beef Production, Dunsany, v.43, p.1-20, 2002.
68. HYNES, R.O. Integrins: versatility, modulation, and signaling in cell adhesion. Cell, Massachusetts, v.69, p.11-25, 1992.
69. HYVARINEN, H.; HELLE, T.; VÄYRYNEN; VÄYRYNEN, P. Seasonal and nutritional effects on serum proteins and urea concentration in the reinder (Rangifer tarandus tarandus L.) British Journal Nutrition, v.33, p.63-72, 1975.

54
70. IHA, S.; ORITA, K.; KANNO, T.; UTSUMI, T.; SATO, E.F.; INOUE, M.; UTSUMI, K. Oxygen-dependent inhibition of neutrophil respiratory burst by nitric oxide. Free Radic Research, Oxford, v.25, n.6, p.489-98, 1996.
71. INMET (Instituto Nacional de Metereologia) [online]. Disponível em http://www.inmet.gov.br. Acesso em 24/06/2004.
72. JAHOOR, F.; BHATTIPROLU, S.; DEL ROSARIO, M. Chronic protein deficiency diffentially affects the kinectis of plasma protein in young pigs. J Nutr, v.121, p.1489-1495, 1996.
73. JAIN NC, VEGAD JL, SHRIVASTAVA AB, JAIN NK, GARG UK, KOLTE GN.Haematological changes in buffalo calves inoculated with Escherichia coli endotoxin and corticosteroids Research Veterinary Science, London, v.47, n.3, p.305-8, 1989.
74. JAIN, N.C. Essentials of Veterinary Hematology. Philadelphia: Lea & Febiger, 1993, 417p.
75. JAMES, A.R.; KAEBERLE, M.L.; HSU, W.H. Effects of ACTH administration on bovine polymorphonuclaear leukocyte function and lymphocyte blastogenese. Journal of Veterinary Research, v.43, p.412-418, 1982.
76. KAMPEAN, A.H.; TOLLERSRUD, T.; LARSEN, S.; ROTH, J.A.; FRANK, D.E.; LUND, A. Repeatability of flow cytometric and classical measurement of phagositosis and respiratory burst in bovine polymorphonuclear leukocytes. Veterinary Immunology and Immunopathology, Amhers, v.97, p.105-114, 2004.
77. KANEKO, J.J. Clinical biochemistry of domestic animal. 4 ed., San Diego: Academic Press, 1989, 932p.
78. KANEKO, J.J.; SMITH, R. The estimation of plasma fibrinogen and its clinical significance in the dog. California Veterinarian, Sacramento, v.21, n.4, p.21-24, 1964.
79. KANTEK, C.E.; NAVARRO, K.G. Manual de Hematologia Veterinária. 2 ed. São Paulo: Varela, 2005, 206p.
80. KAWANO, H.; TSUNO, K.; NOMURA, Y.; KATSURA, N.; ARIKAWA, A.; TSUJI, A.; ONIMARU, T. Values of serum components in Japanese black beef steers at farms with high productivity and low frequencies of disease and death in Miyazaki prefecture. Journal Veterinary Medicine Science, Tokyo, v.59, p.873-877, 1997.
81. KEHRLI, J.R.; SCHMALSTIEY, M.E.; ANDERSON, F.C.; 0VANDERMAATEN, D.C.; HUGHES, M.J.; ACKERMANN, B.J.; WILHELMSEN, M.R.; BRAWN, C.L.; STEVENS, G.B.; WHETSTONE, M.G. Molecular definition

55
of the bovine granulocytopathy syndrome: Identification of deficiency of the Mac-1 (CD11b/CD18) glycoprotein. American Journal of Veterinary Research, Chicago, v. 51, p.1826-1836, 1990.
82. KISHIMOTO, T.K.; KAHN, J.; MIGAKI, G.; MAINOLFI, E.; SHIRLEY, F.; INGRAHAM, S.F.; ROTHLEIN, R. Regulation of L-selectin expression by membrane proximal proteolysis. Agents Actions Suppl, v.47, p.121-134, 1995.
83. KLIMIENE, I.; SPAKAUSKAS, V.; MATUSEVIEIUS, A. Correlation on different biochemical parameters in blood sera of healthy and sick cows. Veterinary Research Comunications, Netherlands. v.29, p.95-102, 2005.
84. KLINKON, M.; ZADNIK, T. Dynamics of red and white blood picture in dairy cows during the periparturient period. Comparative Hematology International, London, v.9, p.156-161, 1999.
85. KNOWLES, T.G. A review of the road transport of cattle, Veterinary Record, London, v.144, p.197-201, 1999.
86. KONSTANTOPOULOS, K.; McINTIRE, L.V. Effects of fluid dynamics on vascular cell adhesion. The Journal of Clinical Investigation, Michigan, v.98, p.2661-2665, 1996.
87. LAPOINTE, J.M., LAVOIE, J.P., VRINS, A.A. Effects of triamcinolone acetonide on pulmonary function and bronchoalveolar lavage cytologic features in horses with chronic obstructive pulmonary disease. American Journal of Veterinary Research, Chicago, v.54, p.1310-6, 1993.
88. LAWRENCE, M.B.; SPRINGER, T.A. Leukocytes roll on a selectin at physiologic flow rates: distrinction and parequesit for adhesion through integrins. Cell, Massachusets, v.65, p.859-873, 1991.
89. LEDWOZYW, A.; STOLARCZYK, H. The involvement of polymorphonuclear leukocytes in the pathogenesis of bronchopneumonia in calves I. Activated granulocyte induced lipid peroxidation in red blood cells. Acta Veterinaria Hungarica, Budapest, v.39, p.175-85, 1991.
90. LEID, R.W.; POTTER, K.A. Inflammation and mediators of lung injury The Veterinary Clinics North America: Food Animal Practice, Philadelfia, v.1, p.377-400, 1985.
91. LEITE, F.; O’BRIEN, S.; SYLTE, M.J.; PAGE, T.; ATAPATTU, D.; CZUPRYNSKI, C.J. Inflammatory cytokines enhance the interaction of Mannheimia haemolytica leukotoxin with bovine peripheral blood neutrophils in vivo. Infection and Immunity, Washington, v.70, p.4336-4343, 2002.
92. LI, X.; ABDI, K.; RAWN, J.; MACKAY, C.R.; MENTZER, S.J. LFA-1 and L-selectin regulation recirculating lymphocyte tethering and rolling on lung

56
endothelium. American Journal of Respiratory Cell and Molecular Biology, New York, v.14, p.398-406, 1996.
93. LIGGITT, H.D. Defense mechanisms in the bovine lung. The Veterinary Clinics North America: Food Animal Practice, Philadelfia, v.1, p.347-66, 1985.
94. LONERAGAN, G.H.; DARGATZ, D.A.; MORLEY, P.S.; SMITH, M.A. Trends in mortality ratios among cattle in U.S. feedlots. Journal Animal Medical Association, Schaumburg, v.219, p.1122-1127, 2001.
95. LOPEZ, A.; MAXIE, G.; RUHNKE, L.; SAVAN, M.; THOMSON, R.G. Cellular inflamatory response in the lungs of calves exposed to bovine viral diarrhea virus, Mycoplasma bovis, and Pasteurella haemolytica. American Journal Veterinary Research, Chicago, v.47, p.1283-6, 1986.
96. LUSCINSKAS, F.W.; LAWLER, J. Integrins as dynamic regulators of vascular function. The FASEB Journal, Bethesda, v.8, p.929-938, 1994.
97. MAC VEAN, D.W.; FRANZEN, D.K.; KEEFE, T.J.; BENNETT, B.W. Airborne particle concentration and meteorologic condictions associated with pneumonia incidence in feedlot cattle. American Journal Veterinary Research, Chicago, v.47, p.2676-2682, 1986.
98. MADER, T.L. Envirommental stress in confined beef cattle. Journal of Animal Science, Champaign, v.81, p.110-119, 2003.
99. MAHESWARAN, S.K.; WEISS, D.J.; KANNAN, M.S. Neutrophil-mediated injury to bovine pulmonary endothelial cells: enhancement by Pasteurella hemolytica leukotoxin. Infect Immunology, v.61, p.2618-2625, 1993.
100. MARAI, I.F.; HABEEB, A.A.; FARGHALY, H.M. Productive, physiological and biochemical changes in imported and locally born Friesian and Holstein lactating cows under hot summer conditions of Egypt. Tropical Animal Health Production, Netherlands, v.31, n.4, p.233-243, 1999.
101. MARKHAN, R.J.F., WILKIE, B.N. Interaction between Pasteurella haemolytica and bovine alveolar macrophages: cytotoxic effect on macrophages and impaired phagocytosis. American Journal Veterinary Research, Chicago, v.41, p.18-22, 1980.
102. MATOS , M.S.; MATOS, P.F. Laboratório Clínico Médico Veterinário. 2 ed. São Paulo: Atheneu, 1988, 238p.
103. MATSUBA, K.T.; VAN EEDEN, S.F.; BICKNELL, S.G.; WALKER, B.A.M. HAYASHI, S.; HOGG, J.C. Apoptosis in circulating PMN: increased susceptibility in L-selectin-deficient PMN. American Journal of Physiology, Maryland, v.272, p.852-2858, 1997.

57
104. McSHERRY, B.; HORNEY, F.D.; deGROOT, J.J. Plasma fibrinogen levels in normal and sick cows. Can. J. Com. Med. V.34, p.191-197, 1970.
105. MEGLIA, G.E.; JOHANNISSON, A.; AGENÄS, S.; HOLTENIUS, K.; PERSSON, W. K. Effects of feeding intensity during the dry period on leukocyte and lymphocyte sub-populations, neutrophil function and health in periparturient dairy cows. The Veterinary Journal, Switzerland, v.169, p.376-384, 2005.
106. MENZIES F.D; MCBRIDE, S.H; MCDOWELL, S.W.; MCCOY, M.A.; MCCONNELL, W.; BELL, C. Clinical and laboratory findings in cases of toxic mastitis in cows in Northern Ireland. Veterinary Record, London, 2000 Jul 29;147(5):123-8.
107. MEYER, D.J.; HARVEY, J.W. Veterinary laboratory medicine: interpretation & diagnosis. 2 ed. Philadelphia:Sauders, 2004, 351p.
108. MILLER, D.N.; WOODBURY, B.L. Simple protocols to determine dust potentials from cattle feedlot soil and surface samples. Journal Enviromental Quality, Madison, v.32, p.1634-1640, 2003.
109. MORRIS, D.D.; LARGE, S.M. Alterações no leucograma. In: SMITH, B.P. Tratado de Medicina Veterinária de Grandes Animais. São Paulo: Manole, p.437-446, 1990.
110. MOULTON, J.E. Tumors in domestic animals, 3ed. Los Angeles: University of California, 1990, 672p.
111. MURATA, H.; SHIMADA, N.; YOSHIOKA, M. Current research on acute phase proteins in veterinary diagnosis: an overview. The Veterinary Journal, v.168, p.28-40, 2004.
112. MURRAY, H.W.; SZURO-SUDOL, A.; WELLNER, D.; OCA, M.J.; GRANGER, A.M.; LIBBY, D.M.; ROTHERMEL, C.D.; RUBIN, B.Y. Role of tryptophan degradation in respiratory burst-independent antimicrobial activity of gamma interferon-stimulated human macrophages. Infect Immun. v.57, n.3, p.845-849, 1989.
113. NAGAHATA, H.; NOCHI, H.; TAMOTO, K.; TANIYAMA, H.; NODA, H.; MORITA, M.; KANAMAKI, M; KOCIBA, G.J. Bovine leukocyte adhesion deficiency in Holstein cattle. Canadian Journal of Veterinary Research, Ottawa, v.57, p. 255-261, 1993.
114. NONNECKE, B.J.; BURTON, J.L.; KEHRLI, M.E. Associations between function and composition of blood mononuclear leukocyte populations from Holstein Bulls treated with dexamatasone. Journal of Dairy Science, Lancaster, v.80, p.2403-2410, 1997.

58
115. NORTHINGTON, W. Hematology: leukocytes and their responses. 25/10/2000.[online].Disponível:http://www.campus.murraystate.edu/academic/faculty/Wade.Northington/leukocyt.htm. Acesso em 27/12/2005.
116. OGILVIE, T.H. Medicina interna de grandes animais. Porto Alegre: Artes Médicas Sul, 2000, 568p.
117. OHTSUKA, H.; FUKUNAGA, N.; HATSUGAYA, A.; HAYASHI, T.; HARA, H.; KOYA, M.; ABE, R.; KAWAMURA, S. Effect of nutritional condictions on changes in leukocyte populations in Japanese black calves. Journal Veterinary Medical Science, Oxford, v.67, p.183-185, 2005.
118. OLSON, D.P.; SOUTH, P.J.; HENDRIX, K. Hematologic values in hypothermic and rewarmed young calves. American Journal of Veterinary Research, Chicago, v.44, n.4, p.572-576, 1983.
119. OTULAKOWSKI, G.L. Proteolysis of sialolycoprotein by Pasteurella hemolytica cytotoxic culture supernate. Infect Immunology, v.42, p.64-70, 1983.
120. PAAPE, M.J.; WERGIN, W.P.; GUIDRY, A.J.; PEARSON, R.E. Leukocytes – second line of defense against invading mastitis pathogens. Journal of Dairy Science, Lancaster, v.62, p.135-153, 1979.
121. PAES, P.R.O.; ROSA, E.P.; BARIONI, G.; FONSCECA, L.A.; ARRIGONI, M.B.; MARTINS, C.L.; TAKAHIRA, R.K.; GONÇALVES, R.C. Leucograma de bovinos Nelore, Brangus e mestiços Nelore x Angus com idade entre 10 a 12 meses, submetidos ao mesmo regime de manejo. In: CONGRESSO LATINO AMERICANO, 11, CONGRESSO BRASILEIRO, 5 e CONGRESSO NORDESTINO DE BUIATRIA, 3., Salvador. Resumos… Salvador: Associação Brasileira de Buiatria, p.37, 2003.
122. PAULA NETO, J.B. Hemogramas de bovinos (Bos taurus) sadios da raça Curraleiro de diferentes idades, machos e fêmeas, gestantes e não gestantes. Goiânia, 2004, 65p. Dissertação (Mestrado) Escola de Veterinária, Universidade Federal de Goiás.
123. PHILLIPS, W.A. The effects of assembly and transit stressors on plasma fibrinogen concentration of beef calves. Can J Comp Med, v.48, p.35-41, 1984.
124. RAMÍREZ, M.N.; SALAZAR, F.C.; LEONARDI, F.; BRAVO, H. Relación albumina:globulina plasmáticas en tres épocas Del año en vacas de la raza Carora Del estado Lara-Venezuela. CONGRESSO NACIONAL DE BUIATRIA, Vera Cruz, 16-18 agosto, 2001.
125. REKWOT, P.I.; OYEDIPE, E.O.; AKEREJOLA, O.O.; VOH, A.A.; DAWUDA, P.M. Serum biochemistry of Zebu bulls and their Friesian crosses fed two planes of protein. British Veterinary Journal, London, v.145, n.1, p.85-88, 1989.

59
126. REYNOLDS, H.Y. Lung inflamation: role of endogenous chemotactic factors in attracting polymorphonuclear granulocytes. The American Review of Respiratory Disease, Baltimore, v.127, p.816-24, 1983.
127. RIBEIRO JÚNIOR. Análises estatísticas no SAEG. Viçosa – UFV, 2001. 301p.
128. RICHARDS, A.B.; RENSHAW, H.W. Functional and metabolic activity of bovine pulmonary lavage cells phagocytically stimulated with pathogenic isolates of Pasteurella haemolytica. American Journal Veterinary Research, Chicago, v.50, p.329-34, 1989.
129. RUBEL, C.; FERNANDEZ, G.C.; DRAN, G.; BOMPADRE, M.B.; ISTURIZ, M.A.; PALERMO, M.S. Fibrinogen promotes neutrophil activation and delays apoptosis. Journal of Immunology, Bethesda, v.66, p.2002-2010, 2001.
130. RUPPANNER, R.; NORMAN, B.B.; ADAMS, C.J.; ADDIS, D.J.; LOFGREEN, G.P.; CLARK, J.G.; DUNBAR, J.R. Metabolic and cellular profile testing in calves under feedlot condictions: blood cellular components – Reference values and changes over time in feedlot. American Journal Research, v.39, n.5, p.851-854, 1978.
131. RYAN, G.M. Blood values in cows: leucocytes . Research in Veterinary Science, Saskatoon, v.12, p.576-578, 1971.
132. SAMPAIO, I.B.M. Estatística aplicada à experimentação animal. Belo Horizonte: Fundação de Ensino e Pesquisa em Medicina Veterinária e Zootecnia, 1998. 221p.
133. SCOTT, P.R. Field study of undifferentiated respiratory disease in housed beef calves. The Veterinary Record, London, v.134, p.325-7, 1994.
134. SITRIN, R.G.; PAN, P.M.; SRIKANTH, S.; TODD, R.F. Fibrinogen activates NF-kappa B transcription factorsin mononucleariphagocytes. Journal of Immunology, Bethesda, v.161, p.1462-1470, 1998.
135. SLAUSON, D.O.; LAY, J.C.; CASTLEMAN, W.L.; NEILSEN, N.R. Alveolar macrophage phagocytic kinetics following pulmonary parainfluenza-3 virus infection. Journal of Leukocyte Biology, Bethesda, v.41, p.412-20, 1987.
136. SLOCOMBE, R.; MALARK, J.; INGESOLL, R.; DERKSEN, F.J.; ROBINSON, N.E. Importance of neutrophils in the pathogenesis of acute pneumonic pasteurelosis in calves. American Journal Veterinary Research, Chicago, v.46, p.2253-8, 1985.
137. SLOCOMBE, R.; MULKS, M.; KILLINGSWORTH, C.R.; DERKSEN, F.J.; ROBINSON, N.E. Effect of Pasteurella haemolytica — derived endotoxin on pulmonary structure and function in calves. American Journal Veterinary Research, Chicago, v.51, p.433-8, 1990.

60
138. SLOCOMBE, R.F.; DERKSEN, F.J.; ROBINSON, N.E.; GUPTA, A.; NEWMAN, J.P. Interations of cold stress and Pasteurela haemolytica in the phatogenesis of pneumonic pasteurelosis in calves: method of introduction and hematologic and pathologic changes. American Journal Veterinary Research, Chicago, v.45, p.1757-1763, 1984.
139. SOETHOUT, E.C.; MÜLLER, K.E.; RUTTEN, V. P.M.G. Neutrophil migration in the lung, general and bovine-specific aspects. Veterinary Immunology and Immunopathology, Amhers, v.87, p.277-285, 2002.
140. SPERTINI, O.; LUSCINSKAS, F.W.; KANSAS, G.S.; MUNRO, J.M.; GRIFFIN, J.D.; GIMBRONE, M.A.; TEDDER, T.F. Leukocyte adhesion molecule-1(LAM-1, L-selectin) interacts with an inducible endothelial cell ligand to support leukocyte adhesion. Journal Immunology, v.147, p.2565-2573, 1991.
141. SPRINGER, T.A. Traffic signals for lymphocyte recirculation and leukocyte emigration: The multistep paradigm. Cell, Massachusets, v.76, p.301-314, 1994.
142. SUTHERLAND, R.J.; WHITNEY, M.S. Acute phase proteins in bovine inflammatory diseases. Texas Vet, Austin, v.57, n.6, p.27-29, 1995.
143. SUTTON, R.H.; HOBMAN, B. The value of plasma fibrinogen estimations in cattle: a comparison with total leukocyte and neutrophil counts. New Zeland Veterinary Journal, Palmeston, v.23, n.3, p.21-27, 1975.
144. SUZUKI, M.; ASAKO, H.; KUBES, P. Neutrophil-derived oxidants promote leukocyte adhence in postcapillary venules. Micro Vasc Res, v.42, p.125-31, 1991.
145. SWANSON, J.C.; MORROW-TESCH, J. Cattle transport: historical, research, and future perspectives, Journal of Animal Science, Champaign, E. Suppl. p.102-109, 2001.
146. SWEENEY, C.R.; BEECH, J. Bronchoalveolar lavage In: BEECH, J. Equine respiratory disorders. Pennsylvania: Lea & Febiger, 1991. Cap. 4, p.55-61.
147. SWEETEN, J.M.; PARNELL, C.B.; ETHEREDJE, R.S.; OSBORNE, D. Dust emissions in cattle feedlots. Veterinary Clinics North Animal Food Animal Practices, Philadelphia, v.4, p.557-578, 1988.
148. TADICH, N.; GALLO, C.; ECHEVERRIA, R.; SCHAIK, V.G. Efecto del ayuno durante dos tiempos de confinamiento y de transporte terrestre sobre algumas variables sanguíneas indicadoras de estrés en novillos. Archivos del Medicina Veteterínaria, Valdivia, v.35, n.2, 2003.
149. TAJIMA, M.; IRIE, M.; KIRISAWA, R.; HAGIWARA, K.; KUROSAWA, T.; TAKAHASHI, K. The detection of a mutation of CD18 gene in bovine leukocyte

61
adhesion deficiency (BLAD). The Journal of Veterinary Medical Scince, Tokyio, v.55, p.145-146, 1993.
150. TAYLOR, J.A. Leukocyte responses in ruminants. In: FELDMAN, B.F.; ZINKL, J.G.; JAIN, N.C. Schalm’s Veterinary Hematology, 5 ed. Philadelphia: Wilkins, p.891-898, 2000.
151. TENNANT, B.; HARROLD, D.; REINA-GUERRA, M. Hematology of the neonatal calf. II. Response associated with acute enteric infections, gram-negative septicemia, and experimental endotoxemia. Cornell Veteterinary, London, v.65, n.4, p.457-75, 1975.
152. THOMAS, J.S. Overview of plasma proteins. In: FELDMAN, B.F.; ZINKL, J.G.; JAIN, N.C. Schalm’s Veterinary Hematology, 5 ed. Philadelphia: Wilkins, p.891-898, 2000.
153. VAL BICALHO, A.P.C.; CARNEIRO, R.A. Os leucócitos, Belo Horizonte: Departamento de Medicina Veterinária da Universidade Federal de Minas Gerais, 2005, [Apostila de Patologia Clínica]. Disponível em http://www.vet.ufmg.br:8080/clinica/clinica/documentos_/00000001 Acesso em 21/12/2005.
154. VEKKOJULKAISUT, H.Y. Hirvonen’s Thesis on acute phase response in dairy cattle, University of Helsinki, Pyörälä: Satu, 2000, 79p.
155. WAGNER, J.G.; ROTH, R.A. Neutrophil migration mechanisms, with an emphasis on the pulmonary vasculature. Pharmacological, Bethesda, v.52, p.349-374, 2000.
156. WEISS, D.J.; PERMAN, V.P. Assessment of the hematopoietic system in ruminants. Veterinary Clinics of North America: Food Animal Pratice, Philadelphia, v.8, n.2, p.411-429, 1992.
157. WILDBLOOD, L.A.; KERR, K.; CLARK, D.A.S.; CAMERON, A.; TURNER, D.G.; JONES, D.G. Production of eosinophil chemoattractant activity by ovine gastrointestinal nematodes. Veterinary Immunology and Immunopathology, Amhers, v.107, p.57-65, 2005.
158. WILLARD, M.D. Hypoalbuminemia. In: FELDMAN, B.F.; ZINKL, J.G.; JAIN, N.C. Schalm’s Veterinary Hematology, 5 ed. Philadelphia: Wilkins, p.891-898, 2000.
159. WILSON, S.C.; MORROW-TESCH, J.; STRAUS, D.C.; COOLEY, J.D.; WONG, W.C.; MITLÖHNER, F.M.; McGLONE, J.J. Airbone microbial flora in a cattle feedlot. Applied and Environental Microbiology, Washington, v.68, n.7, p.3238-3242, 2002.
160. WITHITELEY, L.O.; MAHESWARAN, S.K.; WEISS, D.J.; AMES, T.R. Immunohistochemical localization of Pasteurella haemolytica A1-derivaded

62
endotoxin, leukotoxin, and capsular polysaccharide in experimental bovine pasteurella pneumonia. Veterinary Pathology, Boston, v.27, p.150-161, 1990.
161. YATES, W.D.G. Sistema respiratório. In: THOMPSON, R.G. Patologia Veterinária Especial. São Paulo:Manole, 1990, p.79-142.
162. ZAJAC, A.M. Parasitic bronchitis and pneumonia. In: SMITH, B.P. Large animal internal medicine: diseases of horses, cattle, sheep and goats. 3ed. Missouri: Mosby, 2002, p.577-579.

63
ANEXOS
ANEXO 1 – Valores do leucograma dos bovinos confinados e terminados a pasto
Quadro 1: Contagem total e diferencial de leucócitos de bovinos submetidos ao confinamento
Amostra
Contagem Total de
Leucócitos Neutrófilos Batonetes
Neutrófilos Segmentados Eosinófilos Linfócitos Monócitos
1 8.700 147 4.146 147 3.915 261 2 11.900 119 3.927 595 6.902 357 3 7.950 0 4.373 80 3.499 0 4 6.250 188 2.750 0 3.313 0 5 11.550 0 3.003 231 8.201 0 6 9.800 490 4.116 0 5.096 98 7 4.150 0 830 83 3.238 0 8 2.900 29 1.247 0 1.566 58 9 5.650 396 2.260 0 2.995 0 10 6.400 384 1.856 0 4.032 128 11 5.950 60 2.559 0 3.154 119 12 5.100 102 2.040 51 2.703 204 13 5.350 107 2.408 214 2.247 375 14 6.000 60 2.700 120 2.880 240 15 6.050 61 2.965 182 2.420 424 16 5.350 54 2.836 54 2.033 375 17 7.800 156 3.978 0 3.510 156 18 8.250 83 4.208 165 3.300 495 19 6.700 134 3.752 67 2.613 134 20 8.850 354 3.806 89 4.248 354 21 7.400 222 3.182 0 3.848 148 22 8.850 89 3.363 89 5.133 177 23 9.350 0 3.460 0 5.704 94 24 11.000 110 4.730 220 5.720 330 25 9.350 187 4.582 0 4.301 187 26 4.200 168 1.008 0 2.856 42 27 4.750 0 1.568 48 2.993 95 28 6.400 448 2.368 0 3.328 192 29 7.800 390 4.056 0 3.042 234 30 5.750 345 2.818 0 2.473 58 31 10.750 323 3.978 215 5.483 323 32 7.100 142 2.840 71 3.408 639 33 4.650 372 1.581 0 2.651 47 34 6.900 69 3.795 0 2.760 138 35 9.550 191 4.584 0 4.393 287 36 4.600 46 2.116 0 2.346 92 37 7.450 298 4.396 0 2.608 149 38 4.150 42 2.117 42 1.743 208 39 7.050 212 3.102 0 3.525 212 40 11.600 0 5.568 0 5.800 232 41 10.350 0 4.140 207 5.693 311 42 9.850 99 1.970 0 7.486 296 43 6.450 65 2.774 0 3.354 258 44 10.900 218 4.578 109 5.232 763
45 8.800 88 2.992 0 5.544 176 46 7.900 237 2.923 79 4.266 395 47 8.700 87 1.914 0 6.003 696 48 8.500 85 3.400 255 4.590 170 49 8.300 0 4.731 1.162 2.075 332 50 7.250 0 2.103 73 4.930 145 51 6.050 61 2.057 303 3.570 61 52 6.700 67 2.546 201 3.752 134 53 9.900 0 4.752 99 4.851 198 54 10.450 0 4.807 418 5.016 209 55 6.650 133 3.458 266 2.727 67 56 8.750 0 4.988 350 3.325 88 57 8.900 0 4.183 0 4.628 89 58 9.000 90 3.240 360 4.950 360 59 5.750 0 2.703 230 2.815 0 60 9.950 398 2.786 398 6.369 0
Quadro 2: Contagem total e diferencial de leucócitos de bovinos terminados à pasto
Amostra
Contagem Total de Leucócito
Neutrófilos Batonetes
Neutrófilos Segmentados Eosinófilos Linfócitos Monócitos
1 10.050 503 6.533 0 2.814 2012 9.050 91 3.711 362 4.797 913 4.100 0 1.681 82 2.214 1234 4.650 93 2.883 279 1.303 935 9.250 93 1.758 93 7.308 06 8.150 82 2.771 0 4.727 5717 10.450 418 5.330 0 4.494 2068 12.800 640 5.632 128 6.400 09 8.850 89 5.222 89 3.275 177
10 8.600 172 3.698 172 4.386 8611 7.150 429 3.718 143 2.860 012 7.150 143 2.717 0 4.076 21513 8.600 0 3.784 688 4.042 8614 8.650 87 2.422 1.298 3.720 26015 8.250 0 4.868 165 2.888 33016 8.340 83 4.170 83 3.670 16717 6.050 182 3.691 121 2.057 018 6.200 0 2.976 0 2.914 24819 9.550 191 5.062 0 4.298 020 6.400 0 4.416 128 1.856 021 8.350 0 5.428 84 2.505 33422 9.050 91 3.620 0 4.887 45323 9.800 98 3.724 196 5.292 39224 6.800 68 3.740 68 2.788 13625 8.850 354 5.222 0 3.186 8926 8.400 84 4.620 0 3.528 16827 5.250 0 3.728 0 1.365 15828 7.050 141 5.006 0 1.763 14129 8.700 0 4.872 0 3.741 8730 8.800 0 6.072 0 2.552 17631 5.950 119 3.035 179 2.380 238
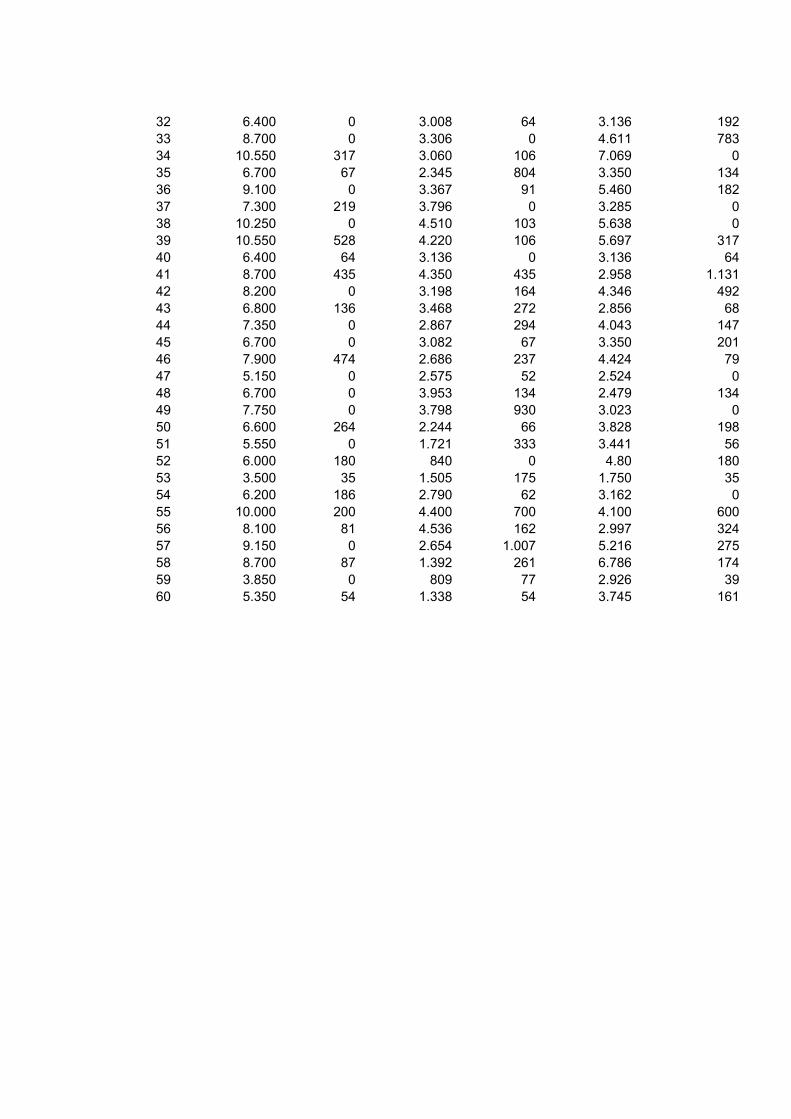
32 6.400 0 3.008 64 3.136 19233 8.700 0 3.306 0 4.611 78334 10.550 317 3.060 106 7.069 035 6.700 67 2.345 804 3.350 13436 9.100 0 3.367 91 5.460 18237 7.300 219 3.796 0 3.285 038 10.250 0 4.510 103 5.638 039 10.550 528 4.220 106 5.697 31740 6.400 64 3.136 0 3.136 6441 8.700 435 4.350 435 2.958 1.13142 8.200 0 3.198 164 4.346 49243 6.800 136 3.468 272 2.856 6844 7.350 0 2.867 294 4.043 14745 6.700 0 3.082 67 3.350 20146 7.900 474 2.686 237 4.424 7947 5.150 0 2.575 52 2.524 048 6.700 0 3.953 134 2.479 13449 7.750 0 3.798 930 3.023 050 6.600 264 2.244 66 3.828 19851 5.550 0 1.721 333 3.441 5652 6.000 180 840 0 4.80 18053 3.500 35 1.505 175 1.750 3554 6.200 186 2.790 62 3.162 055 10.000 200 4.400 700 4.100 60056 8.100 81 4.536 162 2.997 32457 9.150 0 2.654 1.007 5.216 27558 8.700 87 1.392 261 6.786 17459 3.850 0 809 77 2.926 3960 5.350 54 1.338 54 3.745 161
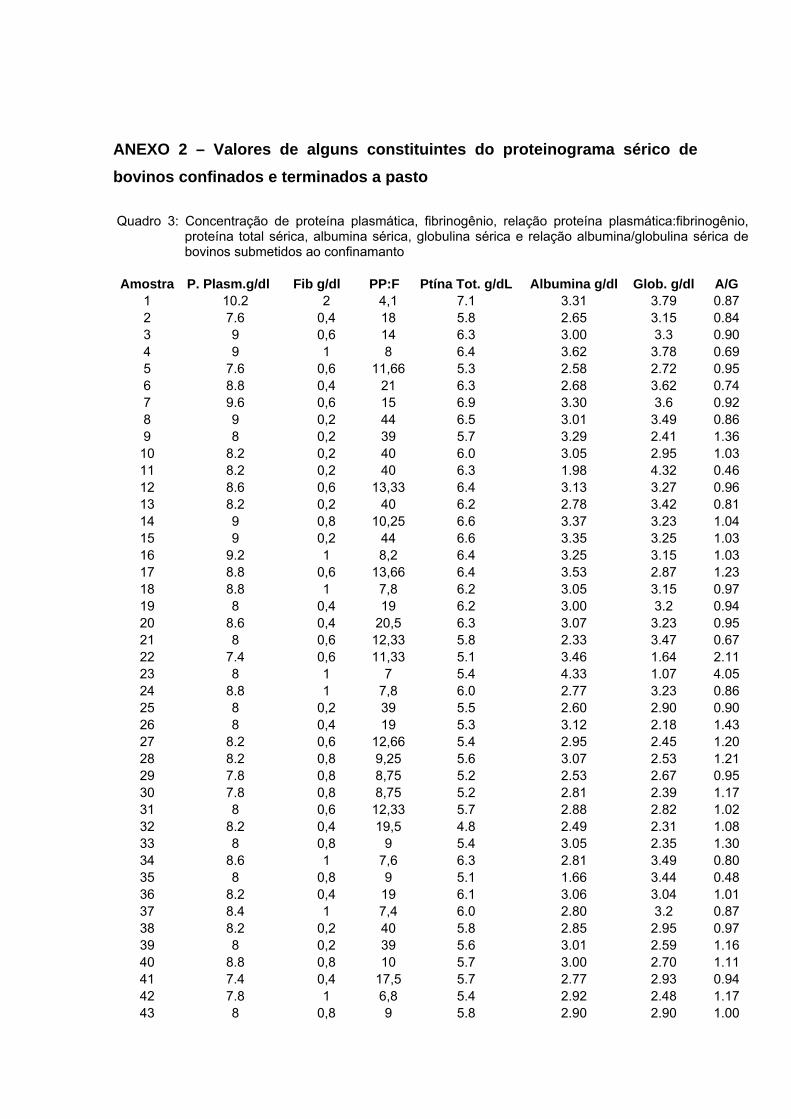
ANEXO 2 – Valores de alguns constituintes do proteinograma sérico de bovinos confinados e terminados a pasto Quadro 3: Concentração de proteína plasmática, fibrinogênio, relação proteína plasmática:fibrinogênio,
proteína total sérica, albumina sérica, globulina sérica e relação albumina/globulina sérica de bovinos submetidos ao confinamanto
Amostra P. Plasm.g/dl Fib g/dl PP:F Ptína Tot. g/dL Albumina g/dl Glob. g/dl A/G
1 10.2 2 4,1 7.1 3.31 3.79 0.87 2 7.6 0,4 18 5.8 2.65 3.15 0.84 3 9 0,6 14 6.3 3.00 3.3 0.90 4 9 1 8 6.4 3.62 3.78 0.69 5 7.6 0,6 11,66 5.3 2.58 2.72 0.95 6 8.8 0,4 21 6.3 2.68 3.62 0.74 7 9.6 0,6 15 6.9 3.30 3.6 0.92 8 9 0,2 44 6.5 3.01 3.49 0.86 9 8 0,2 39 5.7 3.29 2.41 1.36 10 8.2 0,2 40 6.0 3.05 2.95 1.03 11 8.2 0,2 40 6.3 1.98 4.32 0.46 12 8.6 0,6 13,33 6.4 3.13 3.27 0.96 13 8.2 0,2 40 6.2 2.78 3.42 0.81 14 9 0,8 10,25 6.6 3.37 3.23 1.04 15 9 0,2 44 6.6 3.35 3.25 1.03 16 9.2 1 8,2 6.4 3.25 3.15 1.03 17 8.8 0,6 13,66 6.4 3.53 2.87 1.23 18 8.8 1 7,8 6.2 3.05 3.15 0.97 19 8 0,4 19 6.2 3.00 3.2 0.94 20 8.6 0,4 20,5 6.3 3.07 3.23 0.95 21 8 0,6 12,33 5.8 2.33 3.47 0.67 22 7.4 0,6 11,33 5.1 3.46 1.64 2.11 23 8 1 7 5.4 4.33 1.07 4.05 24 8.8 1 7,8 6.0 2.77 3.23 0.86 25 8 0,2 39 5.5 2.60 2.90 0.90 26 8 0,4 19 5.3 3.12 2.18 1.43 27 8.2 0,6 12,66 5.4 2.95 2.45 1.20 28 8.2 0,8 9,25 5.6 3.07 2.53 1.21 29 7.8 0,8 8,75 5.2 2.53 2.67 0.95 30 7.8 0,8 8,75 5.2 2.81 2.39 1.17 31 8 0,6 12,33 5.7 2.88 2.82 1.02 32 8.2 0,4 19,5 4.8 2.49 2.31 1.08 33 8 0,8 9 5.4 3.05 2.35 1.30 34 8.6 1 7,6 6.3 2.81 3.49 0.80 35 8 0,8 9 5.1 1.66 3.44 0.48 36 8.2 0,4 19 6.1 3.06 3.04 1.01 37 8.4 1 7,4 6.0 2.80 3.2 0.87 38 8.2 0,2 40 5.8 2.85 2.95 0.97 39 8 0,2 39 5.6 3.01 2.59 1.16 40 8.8 0,8 10 5.7 3.00 2.70 1.11 41 7.4 0,4 17,5 5.7 2.77 2.93 0.94 42 7.8 1 6,8 5.4 2.92 2.48 1.17 43 8 0,8 9 5.8 2.90 2.90 1.00
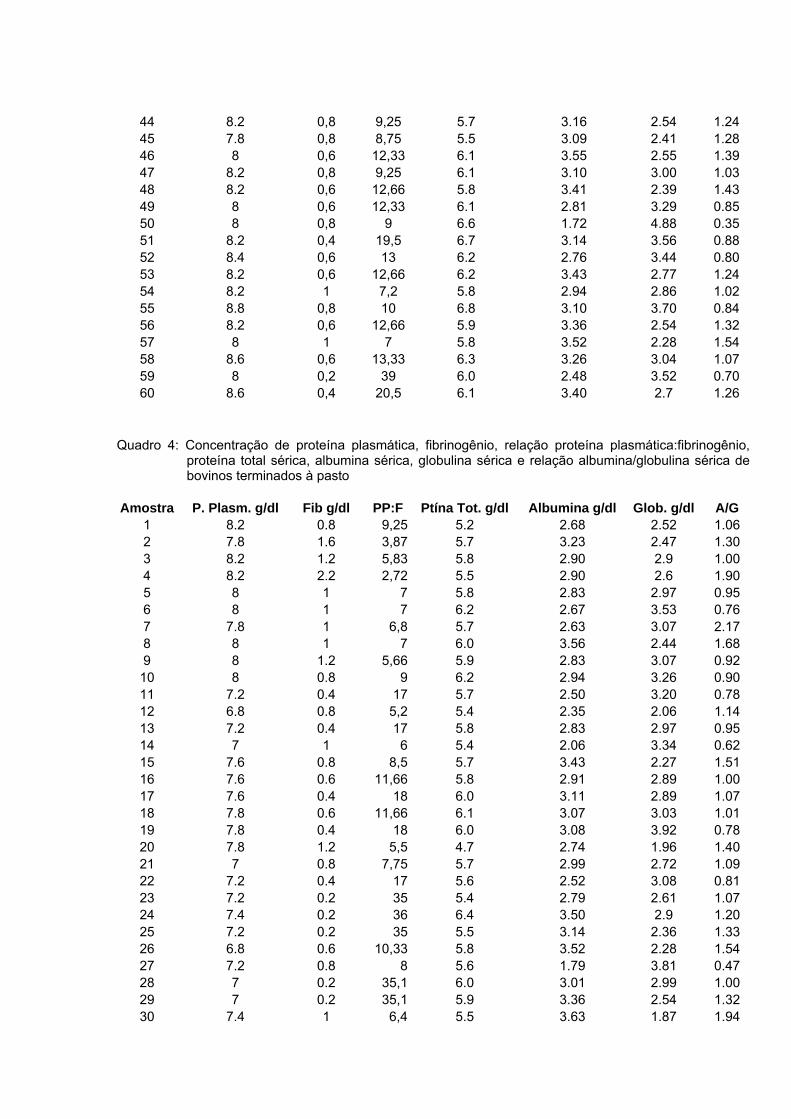
44 8.2 0,8 9,25 5.7 3.16 2.54 1.24 45 7.8 0,8 8,75 5.5 3.09 2.41 1.28 46 8 0,6 12,33 6.1 3.55 2.55 1.39 47 8.2 0,8 9,25 6.1 3.10 3.00 1.03 48 8.2 0,6 12,66 5.8 3.41 2.39 1.43 49 8 0,6 12,33 6.1 2.81 3.29 0.85 50 8 0,8 9 6.6 1.72 4.88 0.35 51 8.2 0,4 19,5 6.7 3.14 3.56 0.88 52 8.4 0,6 13 6.2 2.76 3.44 0.80 53 8.2 0,6 12,66 6.2 3.43 2.77 1.24 54 8.2 1 7,2 5.8 2.94 2.86 1.02 55 8.8 0,8 10 6.8 3.10 3.70 0.84 56 8.2 0,6 12,66 5.9 3.36 2.54 1.32 57 8 1 7 5.8 3.52 2.28 1.54 58 8.6 0,6 13,33 6.3 3.26 3.04 1.07 59 8 0,2 39 6.0 2.48 3.52 0.70 60 8.6 0,4 20,5 6.1 3.40 2.7 1.26
Quadro 4: Concentração de proteína plasmática, fibrinogênio, relação proteína plasmática:fibrinogênio,
proteína total sérica, albumina sérica, globulina sérica e relação albumina/globulina sérica de bovinos terminados à pasto
Amostra P. Plasm. g/dl Fib g/dl PP:F Ptína Tot. g/dl Albumina g/dl Glob. g/dl A/G
1 8.2 0.8 9,25 5.2 2.68 2.52 1.06 2 7.8 1.6 3,87 5.7 3.23 2.47 1.30 3 8.2 1.2 5,83 5.8 2.90 2.9 1.00 4 8.2 2.2 2,72 5.5 2.90 2.6 1.90 5 8 1 7 5.8 2.83 2.97 0.95 6 8 1 7 6.2 2.67 3.53 0.76 7 7.8 1 6,8 5.7 2.63 3.07 2.17 8 8 1 7 6.0 3.56 2.44 1.68 9 8 1.2 5,66 5.9 2.83 3.07 0.92 10 8 0.8 9 6.2 2.94 3.26 0.90 11 7.2 0.4 17 5.7 2.50 3.20 0.78 12 6.8 0.8 5,2 5.4 2.35 2.06 1.14 13 7.2 0.4 17 5.8 2.83 2.97 0.95 14 7 1 6 5.4 2.06 3.34 0.62 15 7.6 0.8 8,5 5.7 3.43 2.27 1.51 16 7.6 0.6 11,66 5.8 2.91 2.89 1.00 17 7.6 0.4 18 6.0 3.11 2.89 1.07 18 7.8 0.6 11,66 6.1 3.07 3.03 1.01 19 7.8 0.4 18 6.0 3.08 3.92 0.78 20 7.8 1.2 5,5 4.7 2.74 1.96 1.40 21 7 0.8 7,75 5.7 2.99 2.72 1.09 22 7.2 0.4 17 5.6 2.52 3.08 0.81 23 7.2 0.2 35 5.4 2.79 2.61 1.07 24 7.4 0.2 36 6.4 3.50 2.9 1.20 25 7.2 0.2 35 5.5 3.14 2.36 1.33 26 6.8 0.6 10,33 5.8 3.52 2.28 1.54 27 7.2 0.8 8 5.6 1.79 3.81 0.47 28 7 0.2 35,1 6.0 3.01 2.99 1.00 29 7 0.2 35,1 5.9 3.36 2.54 1.32 30 7.4 1 6,4 5.5 3.63 1.87 1.94
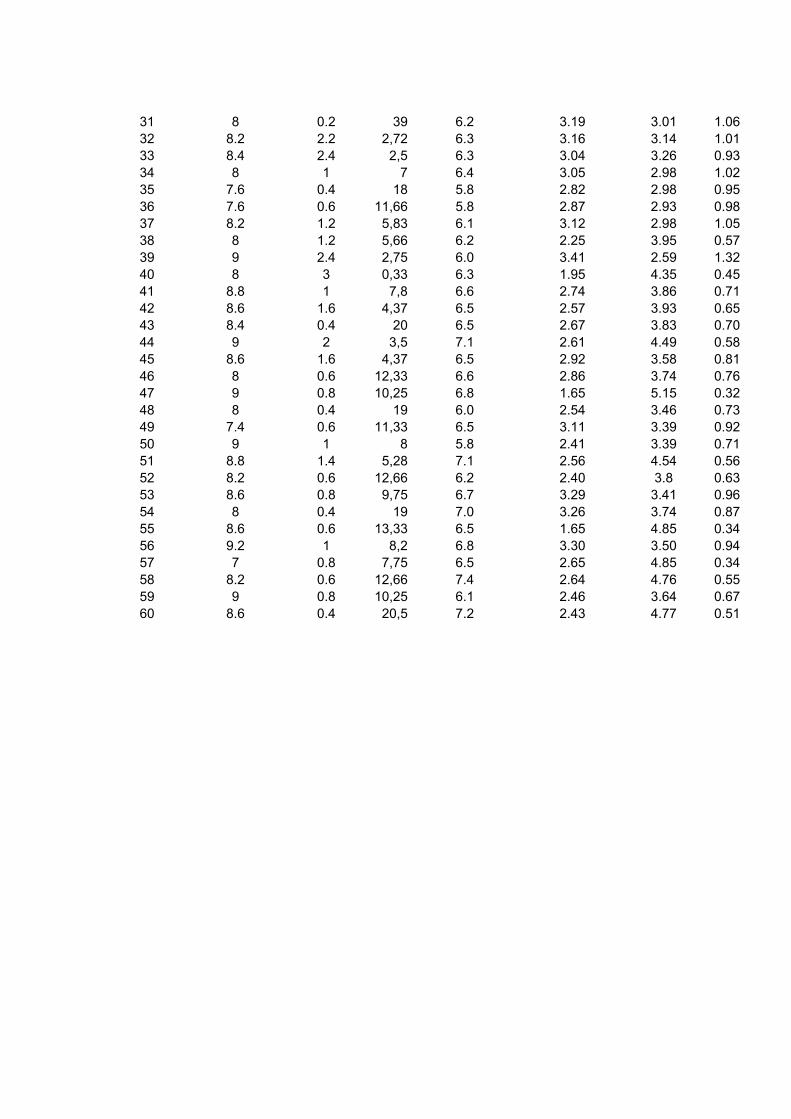
31 8 0.2 39 6.2 3.19 3.01 1.06 32 8.2 2.2 2,72 6.3 3.16 3.14 1.01 33 8.4 2.4 2,5 6.3 3.04 3.26 0.93 34 8 1 7 6.4 3.05 2.98 1.02 35 7.6 0.4 18 5.8 2.82 2.98 0.95 36 7.6 0.6 11,66 5.8 2.87 2.93 0.98 37 8.2 1.2 5,83 6.1 3.12 2.98 1.05 38 8 1.2 5,66 6.2 2.25 3.95 0.57 39 9 2.4 2,75 6.0 3.41 2.59 1.32 40 8 3 0,33 6.3 1.95 4.35 0.45 41 8.8 1 7,8 6.6 2.74 3.86 0.71 42 8.6 1.6 4,37 6.5 2.57 3.93 0.65 43 8.4 0.4 20 6.5 2.67 3.83 0.70 44 9 2 3,5 7.1 2.61 4.49 0.58 45 8.6 1.6 4,37 6.5 2.92 3.58 0.81 46 8 0.6 12,33 6.6 2.86 3.74 0.76 47 9 0.8 10,25 6.8 1.65 5.15 0.32 48 8 0.4 19 6.0 2.54 3.46 0.73 49 7.4 0.6 11,33 6.5 3.11 3.39 0.92 50 9 1 8 5.8 2.41 3.39 0.71 51 8.8 1.4 5,28 7.1 2.56 4.54 0.56 52 8.2 0.6 12,66 6.2 2.40 3.8 0.63 53 8.6 0.8 9,75 6.7 3.29 3.41 0.96 54 8 0.4 19 7.0 3.26 3.74 0.87 55 8.6 0.6 13,33 6.5 1.65 4.85 0.34 56 9.2 1 8,2 6.8 3.30 3.50 0.94 57 7 0.8 7,75 6.5 2.65 4.85 0.34 58 8.2 0.6 12,66 7.4 2.64 4.76 0.55 59 9 0.8 10,25 6.1 2.46 3.64 0.67 60 8.6 0.4 20,5 7.2 2.43 4.77 0.51

ANEXO 3 – Análise de variância não paramétrica para o sistema de terminação de bovinos
Teste de Wilcoxon
Tabela 1: Análise de variância não paramétrica do número de leucócitos em função do sistema de terminação de bovinos – Goiânia, 2005
Sistema de terminação Médias No. de amostras Confinamento 7.640 60 Pasto 7.725 60 Valor do Teste = 0,23624 Significância =0,40663
Tabela 2: Análise de variância não paramétrica do número de bastonete em função do sistema de terminação de bovinos – Goiânia, 2005
Sistema de terminação Médias No. de amostras Confinamento 136,77 60 Pasto 126,30 60 Valor do Teste = 0,97641 Significância = 0,16444
Tabela 3: Análise de variância não paramétrica do número de neutrófilo segmentado em função do sistema de terminação de bovinos – Goiânia, 2005
Sistema de terminação Médias No. de amostras Confinamento 3.200 60 Pasto 3.518 60 Valor do Teste = 1,35940 Significância = 0,08702
Tabela 4: Análise de variância não paramétrica do número de eosinófilo em função do sistema de terminação de bovinos – Goiânia, 2005
Sistema de terminação Médias No. de amostras Confinamento 120,37 60 Pasto 188,37 60 Valor do Teste = 1,60684 Significância = 0,05405

Tabela 5: Análise de variância não paramétrica do número de linfócitos em função do sistema de terminação de bovinos – Goiânia, 2005
Sistema de terminação Médias No. de amostras Confinamento 3.957 60 Pasto 3.687 60 Valor do Teste = 1,08647 Significância = 0,13864
Tabela 6: Análise de variância não paramétrica do número de monócitos em função do sistema de terminação de bovinos – Goiânia, 2005
Sistema de terminação Médias No. de amostras Confinamento 206,83 60 Pasto 186,53 60 Valor do Teste = 1,32007 Significância = 0,09341
Tabela 7: Análise de variância não paramétrica da quantidade de proteína total sérica em função do sistema de terminação de bovinos – Goiânia, 2005
Sistema de terminação Médias No. de amostras Confinamento 5,96 60 Pasto 6,10 60 Valor do Teste = 1,23660 Significância = 0,10812
Tabela 8: Análise de variância não paramétrica da quantidade de albumina sérica em função do sistema de terminação de bovinos – Goiânia, 2005
Sistema de terminação Médias No. de amostras Confinamento 3,01 60 Pasto 2,82 60 Valor do Teste = 2,16527 Significância = 0,01518

Tabela 9: Análise de variância não paramétrica da quantidade de globulina sérica em função do sistema de terminação de bovinos – Goiânia, 2005
Sistema de terminação Médias No. de amostras Confinamento 2,99 60 Pasto 3,29 60 Valor do Teste = 2,0539 Significância = 0,02007
Tabela 10: Análise de variância não paramétrica da relação Albumina/Globulina (A/G) em função do sistema de terminação de bovinos – Goiânia, 2005
Sistema de terminação Médias No. de amostras Confinamento 1,07 60 Pasto 0,95 60 Valor do Teste = 1,79012 Significância = 0,03672
Tabela 11: Análise de variância não paramétrica da concentração de fibrinogênio plasmático em função do sistema de terminação de bovinos – Goiânia, 2005
Sistema de terminação Médias No. de amostras Confinamento 633 60 Pasto 872 60 Valor do Teste = 2,44111 Significância = 0,00732
Tabela 12: Análise de variância não paramétrica da relação proteína plasmática: fibrinogênio em função do sistema de terminação de bovinos – Goiânia, 2005
Sistema de terminação Médias No. de amostras Confinamento 16.69867 60 Pasto 12.06850 60 Valor do Teste = 3.29747 Significância = 0,00049