Embed Size (px)
Citation preview

Placenta CI987),8,545-553
Biochemical, Ultrastructural and Histochemical Studies of Cat Placentae Deficient in Activity of Lysosomal a-Mannosidase
JOSEPH ALROY”?“, CHRISTOPHER D. WARRENb, SRINIVASA S. RAGHAVAN”, PETER F. DANIEL’, KENNETH L. SCHUNKd & EDWIN H. KOLODNY”
u Department of Pathology, Tufts University School of Medicine and Veterinary Medicine, Boston, MA 02111, USA
b Laboratory for Carbohydrate Research, Departments of Biological Chemistry and Medicine, Harvard Medical School and Massachusetts General Hospital, Boston, MA 02114, USA
1 E. K. Shriver Center for Mental Retardation Inc., Department of Biochemistry, Waltham, MA 02254, USA
J Department of Medicine, Tufts University School of Veterinary Medicine, Grafton, MA 01536, USA
e To whom correspondence should be addressed
Paper accepted 20.2.1987
INTRODUCTION
a-Mannosidosis is an autosomal recessive disorder of glycoprotein catabolism (Beaudet, 1983) caused by deficient activity of lysosomal a-mannosidase. It occurs in humans (Kjellman et al, 1969), cattle (Jolly and Thompson, 1978) and cats (Vandevelde et al, 1982). This enzyme removes terminal a-linked mannosyl residues from N-linked oligosaccharides (Warner and O’Brien, 1983). The deficiency results in massive accumulation in tissues and excretion in urine of oligosaccharides with terminal a-mannosyl residues, due to incomplete catabolism of glycoprotein saccharide chains. In all three species the disorder is characterized by progressive neurological impairment as well as visceral, ocular and skeletal involvement. An early diagnosis of this disease is essential for early therapeutic intervention in human patients (Rattazi, 1982) and for experimental therapy in affected animals.
In this study we compared biochemical and morphological features of placentae obtained at birth from normal, presumptive heterozygous and affected kittens. The features examined in- cluded activity of lysosomal ol-mannosidase, the amount and nature of oligosaccharides stored due to incomplete degradation, ultrastructure and patterns of lectin staining. We now report that by using enzyme assay and oligosaccharide determination in placenta, it is possible to dis- tinguish between normal, presumptive heterozygous and affected kittens. Morphological features and lectin histochemistry cannot distinguish between normal and presumptive hetero-
545

546 Placenta (,987), Vol. N
zygotes, while placentae from affected animals exhibit a distinctive pattern by these tech- niques.
MATERIALS AND METHODS
Twenty placentae were obtained at birth from five litters of cats numbers 34 to 38,40 to 42,56 to 59, 61 to 64 and 65 to 68. From each placenta, specimens were taken for determination of lysosomal a-mannosidase activity and for oligosaccharide analysis and were frozen until used. In addition, samples were fixed in IO per cent buffered formalin for light microscopy and lectin histochemistry. Corresponding samples were fixed in 2 per cent (v/v) Trumps solution for electron microscopy.
For determination of enzyme activity, a IO per cent (w/v) homogenate of placental tissue in water was prepared. Protein was determined according to the method of Lowry et al (1951). The reaction mixture contained 5 to ~opg of enzyme protein from placental tissue homo- genate, 5 mM 4-methylumbelliferyl-cl-D-mannopyranoside as substrate, and o. I M sodium acetate pH 4.0 in a total volume of IOO ~1. After incubation for 6 min at 37’C, the reaction was terminated by the addition of a 0.25 M glycine_KOH buffer pH 10.3. The amount of 4-methylumbelliferone released was quantified on an Aminco-Bowman spectrofluorimeter (Kolodny and Mumford, 1976). Venous blood was obtained from the cats. Leucocytes were isolated and a sonicated homogenate in water was prepared (Kolodny and Mumford, 1976). a-Mannosidase activity was assayed as described above using either 20 to 40 pg of protein from leucocytes, or 5 ~1 of plasma.
For oligosaccharide determination, placentae (I g of tissue) were thawed, dried, weighed and scissored, and then homogenized with water (5 ml/g) at 0°C (Polytron PTIO ST, Brinkman, Westbury, NY). The homogenate was centrifuged at 49 ooog for 45 min at 4°C and the super- natant lyophilized. The residue left after freeze-drying was treated with ethanol/water (I:I (v/v); 0.5 ml), mixed vigorously, and allowed to stand for 15 min after which the liquid was transferred into clean tubes and recentrifuged. The supernatant was evaporated to dryness (NJ, and the residue redissolved in water (I ml). One millilitre of tissue extract was applied to a column of Bio-Gel P-2 (1.2 x 14cm) and eluted with water. The first fraction (6 ml) was deionized by passage through coupled columns (0.5 x 2.5 cm) of AG5ow-X8 (H+) and AGI- X8 (HCO,-) ion-exchange resins (Bio-Rad, Richmond, CA). The solution was evaporated (N,) to I ml and oligosaccharides reduced with NaBH, (20 mg) overnight at room temperature (Matsuura et al, 1981). Isocratic, normal-phase high pressure liquid chromatography (HPLC) was performed on a 5 pm Amino Spherisorb column (Warren et al, 1983). For gradient reversed-phase HPLC, a portion of the sample solution (50 out of 200 ~1) was dried, treated with an internal standard of sucrose (25 ng) and perbenzoylated (Daniel et al, 1981). Processing of the sample to remove excess reagents, and HPLC on a C-8 column, were done as described previously (Warren et al, 1986).
Formalin-fixed paraffin-embedded placenta sections were cut at 5 pm, deparaffinized and hydrated. From each placenta, one section was stained with haematoxylin and eosin. For lectin study, placenta sections were incubated with 2 per cent (v/v) H,O, to block endogenous per- oxidase activity. They were then covered for IO min with filtered mouse liver powder solution at a concentration of ~oopg/ml in phosphate-buffered saline (PBS), the excess solution was shaken off and the slide around the tissue was blotted. The sections were then incubated with one of ten different lectins (Vector Laboratories, Burlingame, CA) for 30min, washed three times in PBS followed by incubation with avidin-biotin-peroxidase complex (ABC) (Vector

Alro,~~ et al: Placentae o/‘Feline a-Mannosidosts
Tuble I. Lectins used for identifying carbohydrate residues in cat placenta
.547
Lectin origin
Concen- tration Major sugar Binding
Common name Acronym (g/ml) specilicatiow inhibitor
Arachrs hypogeu Peanut PNA 20 Gal-/&( I -+ 3)-GalNAc Lactose Canavalia enstformt~ Jack bean Con-A 10 a-D-Glc, sc-D-Man a-D-Methyl-Man Doltchos btjorus Horse gram DBA IO cc-o-GalNac cc-o-GalNAc GJycine mu Soybean SBA IO a-o-GalNAc, a-D-Gal cc-o-GalNAc Griffonia srmpliclfolia Bandeiraea GS-I 50 c&o-Gal Lactose Lens rulrnans Common lentil LCA IO cc-o-Glc, cc-o-Man cc-o-Methyl-Man Ricmus communt~ (lastor bean RCA-I 50 P-o-Gal Lactose Trrtmm ml~am Wheat germ WGA 50 (/&(I +.+)-r)-GlcNAc),NeuNAc NeuNAc
Succinyl-WGA S-WGA IO (,4( I ++I>-GlcNAc), p-D-GIcNAc C,‘le.s europarus Gorse UEA-I IO a-I.-Fucose r-L-Fucose
‘s Gal = galactose; GulNAr = h;acetylgalactosamine; GlcNAc = .V-acetylglucosamine; NeuNAc = .V-acetyl- ncuraminic acid (sialic acid); Man = Mannonc.
Laboratories) for 30 min, and washed again three times with PBS. The ‘visualant’ peroxidase was activated by incubation for 8 to ~omin with PBS solution containing diaminobenzidine (DAB) and H,O,. The sections were washed in tap water for 5 min, counterstained with methyl green, dehydrated, and coverslipped with permount. Table I lists the lectins used in this study, their acronyms, the concentration used, their major sugar specificities, and the cor- responding sugars used to inhibit their tissue binding. Incubation of lectins with 0.2 M solu- tions of their corresponding sugar hapten before application of the lectin solution to the tissue section served as a control for binding specificity. Incubation of ABC or lectin solution alone served as a negative control for non-specific binding.
For electron microscopy, 5 mm3 tissue blocks fixed in z per cent (v/v) Trump’s solution were postfixed with I per cent (v/v) osmium tetroxide in 0.1 M cacodylate buffer, pH7.4, dehydrated through graded ethanol solutions, and embedded in Epon 812. For tissue orien- tation r-pm thick Epon-embedded placentae were stained with toluidine blue. Sections 50 to 7onm thin were cut on an LKB 8801 ultramicrotome, stained with uranyl acetate and lead citrate, and photographed with a Philips EM-300 electron microscope.
RESULTS
x-Mannosidase activity in placentae 37, 56,57 and 58 varied from 417 to 88~ units, with 589 % 214 (mean f standard deviation) which was less than IO per cent of control (6286 * 1155), while the enzyme activity of presumptive heterozygotes had a mean of 2581 f 751, which is less than 50 per cent of control (see Table 2). The genetic status of these cats was also con- firmed by the level of cr-mannosidase activity in leucocytes, and the values for controls, carriers and affected cats in leucocytes were 1141 Ifr IO, 286 f 48 and 8.0 f 3.6, respectively. The corresponding values in plasma for control, carriers and affected cats were 4405 f 949, 1336 f 199 and 55 f 6, respectively.
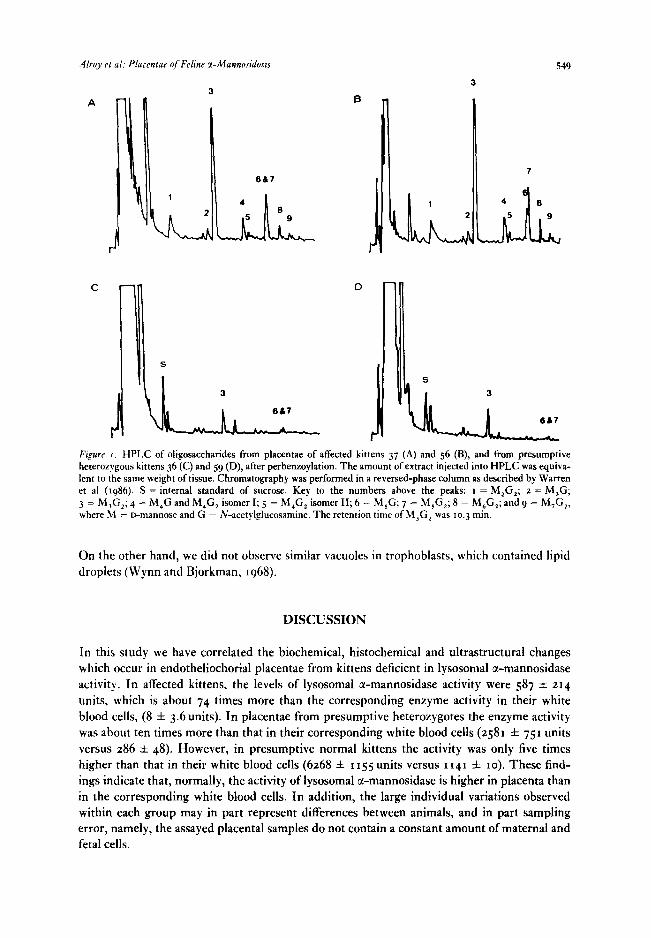
Oligosaccharides from I I placentae were analysed by isocratic HPLC. Placentae 37, 56, 57 and 58 showed high levels of oligosaccharides with a pattern typical of tx-mannosidosis, having a series of peaks with retention times showing a logarithmic relationship. The total amount of oligosaccharides accumulated in these placentae was 446 f 265 pg/g tissue. Placentae 34, 35,
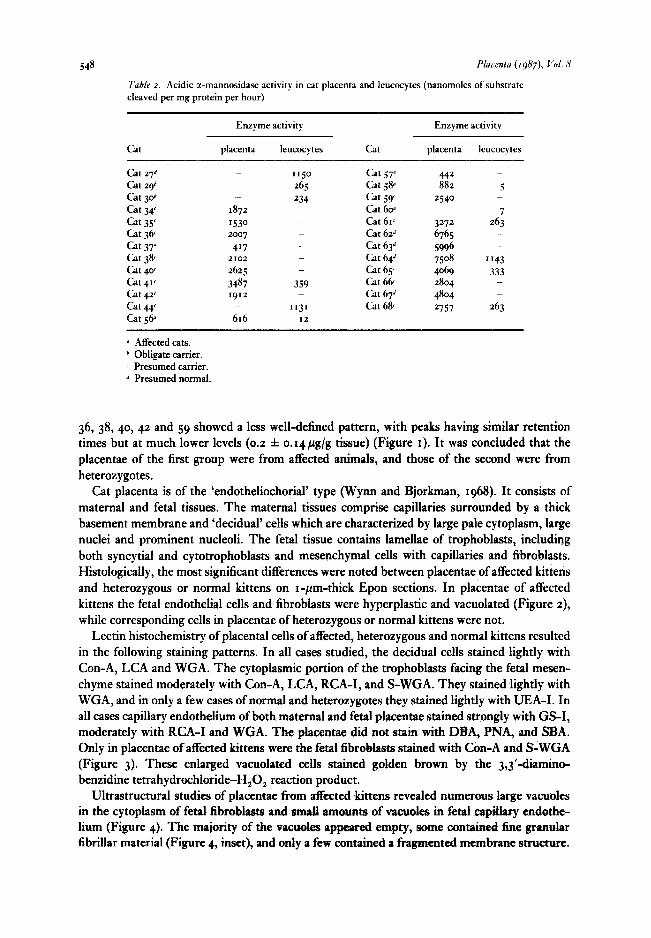
548 Placenra (1987), Vol. N
Table 2. Acidic a-mannosidase activity in cat placenta and leucocytes (nanomoles of substrate cleaved per mg protein per hour)
Cat
Enzyme activity
placenta leucocytes Cat
Enzyme activity
placenta leucocytes
cat 276 Cat ~9~ cat 306 cat 34 Cat 35 Cat 3& a 37 Cat 3% cat 40’ cat 4“ Cat 42’ cat++‘ cat 56”
1872 1530
2IO2
2625 3487 I912
616
II50
265 234
_. _. _ _
359
“3’ I2
Cat 57” Cat 5@ Cat 59 Cat 6oa Cat 61< Cat 626 Cat 636 Cat 646 Cat 65’ Cat 6& Cat 67d Cat 6%
442 882 5
2540 7
3272 263 6765 _ 5996 _
7508 “43 4069 333 2804 4804 2757 263
L1 Affected cats. b Obligate carrier. c Presumed carrier. d Presumed normal.
36, 38,40, 42 and 59 showed a less well-defined pattern, with peaks having similar retention times but at much lower levels (0.2 f o.t4,ug/g tissue) (Figure I). It was concluded that the placentae of the first group were from affected animals, and those of the second were from heterozygotes.
Cat placenta is of the ‘endotheliochorial’ type (Wynn and Bjorkman, 1968). It consists of maternal and fetal tissues. The maternal tissues comprise capillaries surrounded by a thick basement membrane and ‘decidual’ cells which are characterized by large pale cytoplasm, large nuclei and prominent nucleoli. The fetal tissue contains lamellae of trophoblasts, including both syncytial and cytotrophoblasts and mesenchymal cells with capillaries and fibroblasts. Histologically, the most significant differences were noted between placentae of affected kittens and heterozygous or normal kittens on t-pm-thick Epon sections. In placentae of affected kittens the fetal endothelial cells and fibroblasts were hyperplastic and vacuolated (Figure 2),
while corresponding cells in placentae of heterozygous or normal kittens were not. Lectin histochemistry of placental cells of affected, heterozygous and normal kittens resulted
in the following staining patterns. In all cases studied, the decidual cells stained lightly with Con-A, LCA and WGA. The cytoplasmic portion of the trophoblasts facing the fetal mesen- thyme stained moderately with Con-A, LCA, RCA-I, and S-WGA. They stained lightly with WGA, and in only a few cases of normal and heterozygotes they stained lightly with UEA-I. In all cases capillary endothelium of both maternal and fetal placentae stained strongly with GS-I, moderately with RCA-I and WGA. The placentae did~not stain with DEA, PNA, and SBA. Only in placentae of affected kittens were the fetal fibroblasts stained with Con-A and S-WGA (Figure 3). These enlarged vacuolated cells stained golden brown by the 3,3’diamino- benzidine tetrahydrochloride-H,0, reaction product.
Ultrastructural studies of placentae from affected kittens revealed numerous large vacuoles in the cytoplasm of fetal fibroblasts and small amounts of vacuoles in fetal capillary endothe- lium (Figure 4). The majority of the vacuoles appeared empty, some contained fine granular fibrillar material (Figure 4, inset), and only a few contained a fragmented membrane structure.
.4lr11y PI ul: Placentae qf Feline a-Mannoszdosrs 549
667
B
t
D
I Figure I. HPLC of oligosaccharides from placentae of affected kittens 37 (A) and 56 (B), and from presumptive heterozygous kittens 36 (C) and 59 (D), after perbenzoylation. The amount of extract injected into HPLC was equiva- lent to the same weight of tissue. Chromatography was performed in a reversed-phase column as described by Warren et al (1986). S = internal standard of sucrose. Key to the numbers above the peaks: I = M,G,; a = M,G; 3 = M,G,; 4 = M,G and M,G, isomer I; 5 = M,G, isomer II; 6 = M,G; 7 = M,G,; 8 = M,G,; and 9 = M,G,, where J&I = D-ItIannOSe and G = N-acetylglucosamine. The retention time of M,G, was to.3 mm.
On the other hand, we did not observe similar vacuoles in trophoblasts, which contained lipid droplets (Wynn and Bjorkman, 1968).
DISCUSSION
In this study we have correlated the biochemical, histochemical and ultrastructural changes which occur in endotheliochorial placentae from kittens deficient in lysosomal a-mannosidase activity. In affected kittens, the levels of lysosomal a-mannosidase activity were 587 f 214 units, which is about 74 times more than the corresponding enzyme activity in their white blood cells, (8 f 3.6 units). In placentae from presumptive heterozygotes the enzyme activity was about ten times more than that in their corresponding white blood cells (2581 f 751 units versus 286 f 48). However, in presumptive normal kittens the activity was only five times higher than that in their white blood cells (6268 f I 155 units versus I 141 f IO). These find- ings indicate that, normally, the activity of lysosomal a-mannosidase is higher in placenta than in the corresponding white blood cells. In addition, the large individual variations observed within each group may in part represent differences between animals, and in part sampling error, namely, the assayed placental samples do not contain a constant amount of maternal and fetal cells.
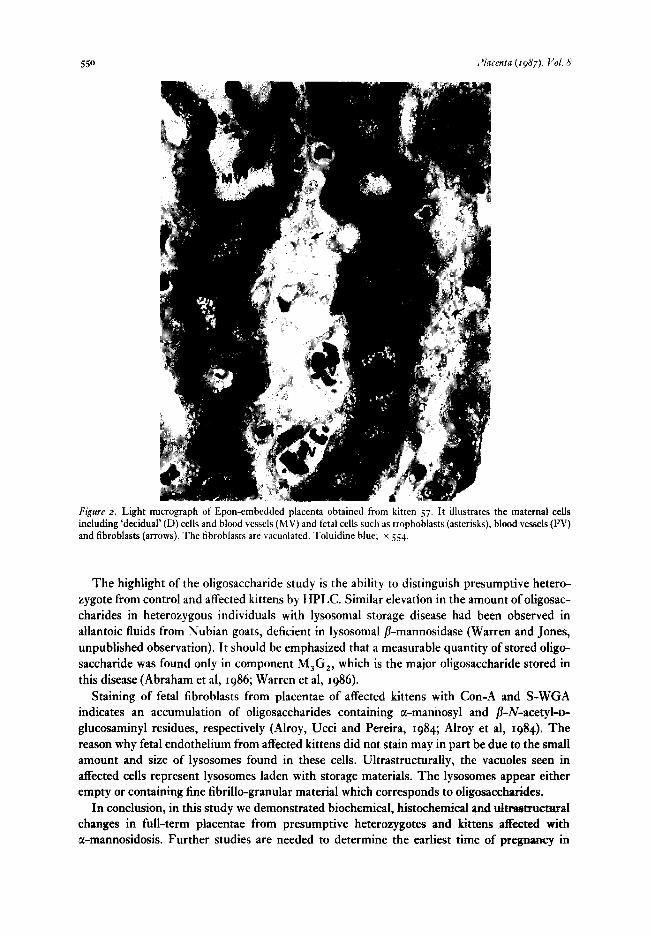
Figure 2. Light micrograph of Epon-embedded placenta obtained from kitten 57. It illustrates the maternal cells including ‘decidual’ (D) cells and blood vessels (MV) and fetal cells such as trophoblasts (asterisks), blood vessels (FV) and fibroblasts (arrows). The fibroblasts are vacuolated. Toluidine blue; x 554.
The highlight of the oligosaccharide study is the ability to distinguish presumptive hetero- zygote from control and affected kittens by HPLC. Similar elevation in the amount of oligosac- charides in heterozygous individuals with lysosomal storage disease had been observed in allantoic fluids from Nubian goats, deficient in lysosomal /?-mannosidase (Warren and Jones, unpublished observation). It should be emphasized that a measurable quantity of stored oligo- saccharide was found only in component M,G,, which is the major oligosaccharide stored in this disease (Abraham et al, 1986; Warren et al, 1986).
Staining of fetal fibroblasts from placentae of affected kittens with Con-A and S-WGA indicates an accumulation of oligosaccharides containing cc-mannosyl and P-N-acetyl-D- glucosaminyl residues, respectively (Alroy, Ucci and Pereira, 1984; Alroy et al, 1984). The reason why fetal endothelium from affected kittens did not stain may in part be due to the small amount and size of lysosomes found in these cells. Ultrastructurally, the vacuoles seen in affected cells represent lysosomes laden with storage materials. The lysosomes appear either empty or containing fine fibrillo-granular material which corresponds to oligosaccharides.
In conclusion, in this study we demonstrated biochemical, histochemical and ultrastructural changes in full-term placentae from presumptive heterozygotes and kittens affected with a-mannosidosis. Further studies are needed to determine the earliest time of pregnancy in

Alro.~~ et ~1: Placentae of Felrne a-Mannoridosis 55’
Figure 3. Light micrograph of paraffin-embedded placenta obtained from kitten 37. Only the trophoblasts and fetal fibroblasts (arrows) stained with S-WGA. Methyl green; x 554.
which one may be able to distinguish between these conditions, by using methodology similar to chorionic villi biopsies (Miller, 1986).
SUMMARY
Lysosomal a-mannosidase activity, oligosaccharide profiles, light and electron microscopy and lectin histochemistry studies were performed on full-term placentae obtained from five litters of cats. They resulted from breeding related cats who are obligate heterozygotes for lysosomal a-mannosidase deficiency. c+Mannosidase activity in placentae from affected kittens was less than IO per cent of control, while in placentae from presumptive heterozygotes the activity was less than 50 per cent of control. High-pressure liquid chromatographic analysis of oligosac- charides revealed massive accumulation of undegraded oligosaccharides in placentae of affected kittens. A small elevation was found in placentae from presumptive heterozygous kittens, and none was detected in placentae of normal kittens. Light and electron microscopic examinations revealed vacuolization of fetal endothelial and mesenchymal cells only in placentae of affected kittens. Succinylated wheat germ agglutinin and concanavalin A stained the fetal fibroblasts only in placentae of affected kittens.

Placenta (19#7), Vol. H
Figure 4. Electron micrograph of placenta obtained from kitten 56. It illustrates trophoblasts (Tr), fetal vessels (FV) and fetal fibroblasts (Mes). Both the endothelial cells and fibroblasts are vacuolated. x 6895. Inset. Close-up of fetal endothelial cell illustrating enlarged vacuole containing ‘stored’ fibrillar and granular material. x 42 980.
ACKNOWLEDGEMENTS
This study was supported by grant NSz1765 from the National Institute of Neurological and Communicative Dis- orders and Stroke. The authors are thankful to Mrs Vibha Goyal for her technical assistance and to MS Cindy Welch for assistance with the manuscript.
REFERENCES
Abraham, D., Daniel, P. F., Dell, A. et al (1986) Structural analysis of the major urinary oligosaccharides in feline a-mannosidosis. BiochcmicalJoumal, 233,899-904.
Alroy, J., Ucci, A. A. & Pereira, M. E. A. (1984) Lectins: histochemical probes for specific carbohydrate residues. In Diagrrosric Zmmunohisrochemisrry (Ed.) DeLellis, R. A., Volume z, pp. 67-88. New York: Masson.
AIroy, J., Orgad, U., Ucci, A. A. & Pereira, M. E. A. (1984) Identification of glycoprotein storage diseases by lec- tins: a new diagnostic method. Journal of Histochemistry and Cytochemistry, 32, 1280-1284.
Beaudet, A. L. (1983) Disorders of glycoprotein degradation: mannosidosis, fucosidosis, sialidosis and aspartylgly- cosaminuria. In The Metabolic Basis of Znherired Discuses (Ed.) Stanbury, J. B., Wyngaarden, J. B., Frederickson, D. A. S. et al, 5th Edition, pp. 78842. New York: McGraw-Hill.
Daniel, P. F., DeFeudisj D. F., Lott, 1. T. br McCluer, R. H. (I&) Qpantitative microanalysis of oligosaccharides by high performance liquid chromatography. Carbohydrate Research, 97, 161-180.
jolly, R. D. & ThoanPson, K. G. (1978) The Pathology of bovine tmumosidosis. Verninary Pa&&gy, 1g,141--152. Kjellman, B., Gamstotp, I., Brun, A. et al (1969) Mannosidosis: a clinical and histopattmlogial study. 3oumal of
Pediatrics, 75,366-373. Kohxlny, E. H. & Mumford, R. A. (1976) H uman leukocyte acid hydrolases: characterization of eleven lysosomal
enzymes and study of reaction conditions for their automated analysis. Ctinica C&r&r Acts, 70,247-257. Lowry, 0. H., Rosonbrough, N. J., Fart, A. L. L Rat&II, R. J.( 1951) Protein measurement with the folk phenol
reagent. 3ournal of Biological Chemistry, 193,265-275. Matauuq F., Nunez, H. A., Gsabowski, G. A. & Sweeley, C. C. (rq81) Smmtural studies of’urinaryoligosacchar-
idea from patiUm3 w&h nuum&&&. Arc&w of &oCti?y ad Bio&sks, 2q,337-352. Slkr, K. (1986) ChorionbioPsie-ke nave mogliehkeit der prenatalen &goose.. &r~ck&rt, 73,125-1a8. Rnttazl, M. C. (1~8~) Enzyme thesapy in 1ysocomPt storage dii: current approaches. In If&ran Ccttetirs, Part B:
Medical Aspects (Ed.) Bonne-Tamir, B. pp. 573-583. New York Alan Liss.

A@ et rrl: Placentae of Feline a-Mannosidosis 5.53
Vandevelde, M., Fankauser, R., Bischsel, P. et al (1982) Hereditary neurovisccral mannosidosis associated with ” r-mannosidase deficiency in a family of Persian cats. Acta Neuropathologica, 58,6448.
Warner, G. T. & O’Brien, J. S. (1983) Genetic defects in glycoprotcin metabolism. .4nnual Reviem of‘Cenetcrs, 17,
395-44’. Warren, C. D., Sadeh, S., Daniel, P. F. et al (1983) Induced mannosidosis-excrction of oligosaccharides b)
locoweed-intoxication in sheep. FEBS Letters, 163,99-103. Warren, C. D., AIroy, J., Bugge, B. et al (1986) Oligosaccharides from placenta: early diagnosis of feline mannosi-
dosis. FEB.7 Letters, 1952477252. Wynn, R. M. & Bjorkman, N. (1968) Ultrastructure of feline placental membrane. .4merican 3ournal of Obstetrri-s
und G,ynrcology, IOZ, 34-43.








![Diseases related to pregnancy - med.nu.ac.th · PDF file• abruptio placentae • rupture of spiral artery ... Microsoft PowerPoint - เอกสาร4....ppt [Compatibility Mode]](https://img.pdfslide.tips/doc/110x75/5ab108cb7f8b9abc2f8c2699/diseases-related-to-pregnancy-mednuacth-abruptio-placentae-rupture-of.jpg)




















