Embed Size (px)
Citation preview

Biochimie - Curs 12 - 2019
1
Structura acizilor nucleici
În orice organism acizii nucleici constituie o sursă care codează informaţia
biologică. Forma şi activităţile diverselor celule sunt în mare măsură determinate de
instrucţiunile genetice conţinute de ADN (sau ARN în unele virusuri). În conformitate cu
dogma centrală a biologiei moleculare, secvenţa de baze nucleotidice din ADN codează
secvenţa aminoacizilor din proteine. Multe proteine din celulă sunt enzime care participă
în procesele metabolice. Alte proteine au rol participa în menţinerea şi transmiterea
informaţiei genetice.
Există două tipuri de acizi nucleici, ADN-ul și ARN-ul, care sunt purtători ai
informaţiei genetice şi determină ca această informaţie să fie disponibilă pentru celulă.
Structurile acestor molecule trebuie să fie în concordanţă cu următoarele aspecte:
1. Informaţia genetică trebuie păstrată într-o formă stabilă pentru o perioadă
îndelungată.
2. Informaţia genetică trebuie sa fie decodată înainte de a fi utilizată. Transcripţia
este procesul în care secvenţa nucleotidelor din ADN este copiată sub forma
ARN-ului mesager în aşa manieră încât să determine sinteza proteinelor (proces
de translaţie care se desfășoară în ribozomi).
3. Informaţia conţinută pe ADN sau ARN trebuie să fie accesibilă atât proteinelor
cât şi acizilor nucleici. Aceşti agenţi pot recunoaşte (se pot lega de) acizii nucleici
astfel încât să poată determina schimbări ale funcţiilor acestor molecule.
4. Progenitorii unui organism trebuie să fie echipaţi cu acelaşi set de instrucţiuni ca
ale părintelui. Astfel, ADN-ul este replicat (copiat) astfel încât celulele nou
formate să primească aceeaşi informaţie genetică.
În general, este acceptat faptul că pentru exercitarea funcţiei lor acizii nucleici
necesită şi unele componente celulare.
Structura elicoidală a ADN-ului
Perioada de după anii 1900 până la cel de-al doilea Razboi Mondial a fost
considerată “vârsta de aur” a geneticii. Cu toate acestea cercetătorii nu au reuşit să
confirme faptul că ADN-ul şi nu proteinele constituie materialul ereditar. Oricum,
această perioadă s-a distins printr-o serie de descoperiri genetice care au permis stabilirea
unor corelaţii între genetică şi evoluţie.
În 1869, F. Miescher a izolat ADN-ul din leucocite. Acesta a colectat “puroiul”
(care conţine o cantitate apreciabilă de leucocite) din bandajele folosite la un spital, după
care a îndepărtat acest lichid de pe bandaje utilizând o soluţie salină. După adăugarea
unei soluţii slab alcaline peste celulele albe a constat faptul că nucleele precipită din
soluţie. Meischer a observat faptul că aceasta substanţă din nucleu, numită nucleină, are
un raport constant fosfor:azot (P:N).
ADN-ul s-a dovedit a fi unul din principalii componenţi ai nucleului (Mendel şi
Darwin au publicat descoperirea lor în acelaşi timp). Deoarece s-a izolat din nucleu,
initial compusul (ADN-ul) a fost numit nucleină. Mai târziu a fost denumit acid nucleic şi
în ultima instanţă acid deoxiribonucleic (ADN).
Biochimie - Curs 12 - 2019
2
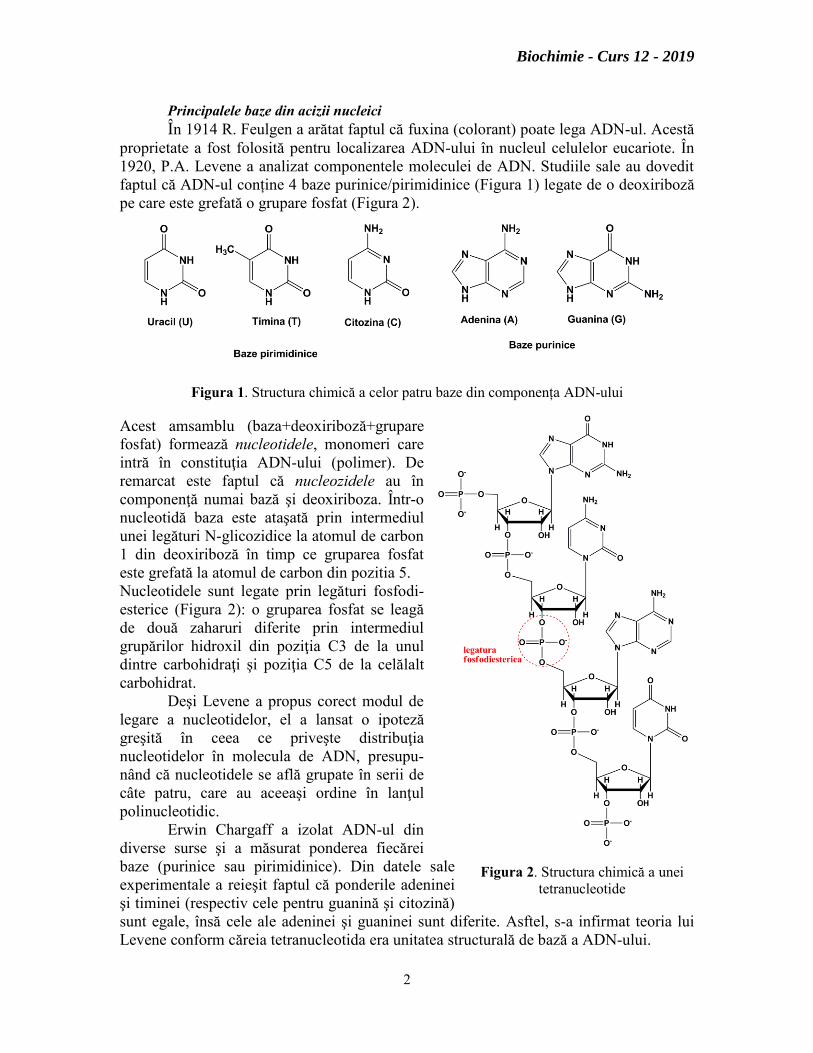
Principalele baze din acizii nucleici
În 1914 R. Feulgen a arătat faptul că fuxina (colorant) poate lega ADN-ul. Acestă
proprietate a fost folosită pentru localizarea ADN-ului în nucleul celulelor eucariote. În
1920, P.A. Levene a analizat componentele moleculei de ADN. Studiile sale au dovedit
faptul că ADN-ul conține 4 baze purinice/pirimidinice (Figura 1) legate de o deoxiriboză
pe care este grefată o grupare fosfat (Figura 2).
Figura 1. Structura chimică a celor patru baze din componența ADN-ului
Acest amsamblu (baza+deoxiriboză+grupare
fosfat) formează nucleotidele, monomeri care
intră în constituţia ADN-ului (polimer). De
remarcat este faptul că nucleozidele au în
componenţă numai bază şi deoxiriboza. Într-o
nucleotidă baza este ataşată prin intermediul
unei legături N-glicozidice la atomul de carbon
1 din deoxiriboză în timp ce gruparea fosfat
este grefată la atomul de carbon din pozitia 5.
Nucleotidele sunt legate prin legături fosfodi-
esterice (Figura 2): o gruparea fosfat se leagă
de două zaharuri diferite prin intermediul
grupărilor hidroxil din poziţia C3 de la unul
dintre carbohidraţi şi poziţia C5 de la celălalt
carbohidrat.
Deşi Levene a propus corect modul de
legare a nucleotidelor, el a lansat o ipoteză
greşită în ceea ce priveşte distribuţia
nucleotidelor în molecula de ADN, presupu-
nând că nucleotidele se află grupate în serii de
câte patru, care au aceeaşi ordine în lanţul
polinucleotidic.
Erwin Chargaff a izolat ADN-ul din
diverse surse şi a măsurat ponderea fiecărei
baze (purinice sau pirimidinice). Din datele sale
experimentale a reieşit faptul că ponderile adeninei
şi timinei (respectiv cele pentru guanină şi citozină)
sunt egale, însă cele ale adeninei şi guaninei sunt diferite. Asftel, s-a infirmat teoria lui
Levene conform căreia tetranucleotida era unitatea structurală de bază a ADN-ului.
Figura 2. Structura chimică a unei
tetranucleotide

Biochimie - Curs 12 - 2019
3
Figura 3. Interacțiuni între bazele
nucleotidice din lanțurile
ADN-ului dublu-catenar
După 1900 Garrod a propus o legătură între gene şi metabolismul existent la
naştere. Una din întrebările care au survenit în acea perioadă a fost următoarea: ce este o
genă? Răspunsul a venit după 20 de ani atunci când F. Griffith a studiat diferenţa dintre
două tipuri de bacterii (una care cauzează pneumonia-S şi alta care nu cauzează
pneumonia-R). Tipul S de bacterie era încapsulat, pe când tipul R nu. Griffith a
demonstrat (1928) faptul că un tip de bacterie nepatogenic poate deveni patogenic prin
intermediul unui “factor de transformare”. Cercetătorul a injectat diferite tipuri de bacterii
în șoarece şi a constatat faptul că tipul S induce moartea, în timp ce tipul R nu afectează
organismul. Mai mult, folosind tipul S al bacteriei care este distrus de temperatură a
observat faptul că organismul a supravieţuit. Combinarea ultimului tip de bacterie
(distrusă prin încălzire) cu tipul R (nepatogen) a indus apariţia pneumoniei, respectiv a
decesului. În anul 1944, O. Avery, C. MacLeod și M. McCarty au concluzionat faptul că
“factorul de transformare” este de fapt ADN-ul. Dovada că ADN-ul este materialul
ereditar a fost adusă de M. DelBruck şi S. Luria. Aceştia au analizat bacteriofagele, un tip
de virus (conţinând un inveliş proteic care încapsulează ADN-ul) care atacă bacteriile
E.Coli (din intestinul gros).
În 1952, A. D. Hershey şi M. Chase au demonstrat prin marcare cu radioizotopi a
bacteriofagelor (32P pentru ADN şi 35S pentru proteină) faptul că ADN-ul (care s-a regăsit
în celula infectată) este “purtătorul fizic” al informaţiei genetice.
Structura ADN-ului
ADN-ul este un polimer care are două lanţuri de deoxinucleotide, molecule care
sunt legate prin legături fosfodiesterice. Cea mai cunoscută formă a ADN-ului este aceea
a B-ADN-ului, care are trăsăturile structurale a ADN-ului descris de J. Watson şi F.
Crick, împreună cu R. Franklin şi colaboratorii acesteia:
1. Cele două lanţuri antiparalele sunt rotite spre dreapta în
jurul unei axe comune rezultând o structură elicoidală al
cărei diametru este de aproximativ 20 Å.
2. Planele formate de bazele nucleotidice, care formează
perechi de legături de hidrogen, sunt aproximativ
perpendiculare pe axa centrală a helixului. În B-ADN,
bazele ocupă ”miezul” helixului iar resturile de
fosfozahar se regăsesc în afara acestuia, formând
adâncituri majore sau minore. Doar porţiuni mici din
perechile de baze sunt expuse spre exterior.
3. Fiecare pereche de baze are aproximativ aceeaşi
înclinaţie, ceea ce conferă o simetrie moleculei de
ADN. Perechile de baze A-T şi G-C (Figura 3) pot fi
interschimbabile fără să afecteze poziţia resturilor de
zahar din afara helixului. Guanina formează 3 legături
de hidrogen cu citozina, iar adenina 2 legături de
hidrogen cu timina. Alte perechi de baze perturbă
semnificativ dispunerea de tip bielicoidal.
4. B-ADN helixul are 10 perechi de baze (eng. “base
pairs” = bp) pentru o rotaţie completă şi pasul de 34 Å.
Pentru a demonstra trăsăturile structurale ale ADN-ului Watson şi Crick s-au
bazat pe informaţiile provenite din modul de difracţie al razelor X de către această

Biochimie - Curs 12 - 2019
4
moleculă. În practică un fascicul de raze X este direcţionat spre un cristalul unei
substanţe. După bombardarea cristalului unele raze sunt împraştiate sau reflectate în
momentul în care întâlnesc atomi. Razele X împrăştiate pot interfera între ele şi produc
spoturi de diferite intensităţi, care pot fi înregistrate sub forma unei hărţi. Difractograma
(harta rezultată) este asemenea unei semnături caracteristice fiecărei molecule. R.
Franklin şi W. Maurice au obţinut difractogramele specifice ADN-ului. Din aceste
fotografii reieşea faptul că molecula de ADN este simetrică. Distribuţia sub forma literei
“X”, din fotografie, este o dovadă a structurii elicoidale a ADN-ului. Într-o difractogramă
de raze X cu cât spoturile sunt mai apropiate, cu atât distanţa este mai mare. Astfel
“barele orizontale” corespund de fapt cu pasului helixului. Distanţa verticală dintre bare,
34 Å, este o măsură a înălţimii pasului. Distanţa de la mijlocul difractogramei la partea de
sus a acesteia, 3,4 Å, este echivalentă cu distanţa dintre două baze suprapuse. Dat fiind
faptul că înălţimea pasului era de 34 Å iar distanţa dintre baze 3,4 Å cercetătorii au dedus
numărul de baze pe pas - 10 nucleotide. Pasul helixului poate fi calculat din unghiul pe
care “X-ul” îl face cu axa orizontală. Astfel, dacă unghiul este mai mare pasul este mai
mic, iar ADN-ul mai compact. Din difractograma obținută s-a dedus faptul că ADN-ul
prezintă o modalitate de împachetare dublu-elicoidală cu grupările fosfat orientate spre
exterior, iar bazele în interior.
Watson şi Crick erau în concurenţă cu Pauling în rezolvarea structurii tridimen-
sionale a ADN-ului. Ultimul, după ce a rezolvat structura -helixului din proteine,
încerca să deslușească şi structura ADN-ului. Aproape în acelaşi timp Pauling a trimis o
publicaţie despre structura ADN-ului. Watson şi Crick au verificat structura lui Pauling
cu ajutorul unui model cu bile şi bețe, structura care s-a dovedit a fi un triplu-helix.
Distribuţia grupărilor fosfat era în centrul helixului, iar bazele în exteriorul acestuia.
Această orientare a grupărilor fosfat în interiorul helixului era practic imposibilă datorită
repulsiilor electrostatice dintre sarcinile negative, lucru care ar fi îngreunat împachetarea
moleculei.
Figura 4. Cele mai cunoscute variante de împachetare ale ADN-ului dublu catenar (A, B şi Z)
Strcutura dublu-elicoidală poate avea câteva împachetări distincte în funcție de
compoziţia solventului şi secvenţa de baze. Varianta de ADN propusă de Watson și Crick
era aceea B-ADN-ului (Figura 4). Mai târziu au fost propuse și alte variante structurale
ale ADN-ului: A-ADNul şi Z-ADN-ul.

Biochimie - Curs 12 - 2019
5
5S-rARNul Phe-tARNul
(118 nucleotide) (77 nucleotide)
Figura 5. Tipuri de structuri ale ARN-ului
Figura 6. Conformerii sin și anti ai adenozinei
diferă prin rotația în jurul legăturii N-glicozidice
Perechile de baze ale A-ADN-ului sunt inclinate în raport cu axa helixului
În condiții de deshidratare B-ADN-ul suferă schimbări conformaționale
reversibile și este transformat în A-ADN o macromoleculă a cărei structură este mai
aplatizată și mai largă decât aceea a B-ADN-ului. A-ADNul are 11,6 bp pe rotație și
pasul de 34 Å. Diferenta majoră dintre B-ADN și A-ADN este aceea că perechile de baze
sunt inclinate cu un unghi de 20º în raport cu axa helixului. Mai mult, A-ADN-ul are
adâncituri majore mai profunde comparativ cu B-ADN-ul.
Z-ADN-ul formează un helix orientat spre stânga
La 25 de ani de la descoperirea B-ADN-ului, studiul structurii deoxi(CGCGCG)
de către A. Wang și A. Rich a evidenţiat o altă modalitate de împachetare a ADN-ului.
Lanțurile acestui tip de ADN s-au dovedit a fi orientare spre stânga (Z-ADN). Acest tip
de ADN are 12 bp pe rotaţie şi pasul de 44 Å, adâncitura minoră mai profundă, iar
adâncitura majoră se distinge mai puţin. Difracţia fibrelor şi studiile RMN au arătat faptul
că polinucleotidele complementare cu purinele şi pirimidinele afectate (de exemplu poli
d(GC)·poli d(GC) sau poli d(AC)·poli d(GC)) adoptă conformaţia Z la o concentraţie
mare de sare. Stabilizarea Z-ADN-ului de către săruri se explică prin faptul că repulsiile
dintre gruparile fosfat vecine sunt mai atenuate în prezenţa sărurilor.
ARN-ul formează un A-helix
ARN-ul dublu-helix este materialul genetic al unor virusuri, dar este sintetizat numai sub
forma unui singur lanţ. Acest lanţ poate
forma punţi de hidrogen intramoleculare
formând un lanţ dublu, respectiv bucle
(Figura 5).
Segmente scurte din lanţul ARN-ului sunt
implicate în multiplicarea unor gene.
În mod uzual ARN-ul formează con-
formații de tip A-ADN şi are 11 bp pe
rotaţie şi pasul de 30,9 Å, iar perechile de
baze sunt înclinate cu aproximativ 16,7º
în raport cu axa elicoidului.
Flexibilitatea conformaţională a
ADN-ului este limitată
Conformaţia unei unităţi de nucle-
otidă indică prezenţa a 6 unghiuri de tor-
siune pentru partea de fosfo-zahar şi un
unghi de torsiune atribuit legăturii glicozidice. Rotaţia în jurul legăturii glicozidice este

Biochimie - Curs 12 - 2019
6
impiedicată. Rezidurile purinice posedă două conformaţii, sin şi anti, ultima dintre
acestea fiind mai stabilă (Figura 6). În marea majoritate a acizilor nucleici, toate bazele
adoptă conformaţia anti. Numai în cazul Z-ADN-ului apar resturi de purină şi pirimidină
a căror conformaţie alternează (anti și sin).
Conformaţiile plic ale ribozei sunt esenţiale în acizii nucleici, determinând
orientarea relativă a substituienţilor fosfat la fiecare rest de riboză. În B-ADN
conformaţia este C2’-endo, pe când în A-ADN conformaţia este C3’-endo. În schimb în
nucleotidele purinice din Z-ADN au conformaţia de tip 3’-endo iar nucleotidele
pirimidice au conformaţia 2’-endo.
Proprietăţile ADN-ului în soluţie
Moleculele de ADN de diferite dimensiuni pot fi studiate prin diverse metode
fizico-chimice. Dimensiunile genomului bacteriofagilor și virușilor variază de la câteva
mii de baze (kilobaze) la câteva sute de kilobaze. Genomul bacterian poate varia de la 0,5
Mb la 10 Mb. Genomul eucariotelor este divers și variază de la aproximativ 10 Mb în
unele ciuperci la mai mult de 100000 Mb în anumite plante.
Proprietăţile acido-bazice
Grupările fosfat din legăturile diesterice ale ADN-ului, care se repetă periodic în
fiecare nucleotidă, au valori scăzute ale pKa-ului şi din acest motiv sunt ionizate la valori
ale pH-ului mai mari de valoarea 4, fapt care conferă ADN-ului un caracter acid.
Grupările fosfat sunt orientate spre exteriorul dublu-helixului şi pot fi interacţiona cu
moleculele de apă, ioni divalenţi (Ca2+, Mg2+) sau amine policationice (spermidina şi
spermina-sunt asociate cu moleculele de ADN virale sau bacteriene). Stabilitatea
legăturilor de hidrogen din perechile de baze din ADN depinde de gradul de ionizare al
gruparilor amino (pH 4-11).
Vâscozitatea
Rigiditatea şi lungimea apreciabilă a lanţului conferă ADN-ului o vâscozitate
apreciabilă. Măsurătorile de vâscozitate sunt folosite pentru a urmări gradul de denaturare
al ADN-ului şi implicit gradul de împachetare al duplexului.
Coeficientul de sedimentare
Coeficientul de sedimentare şi masa moleculară a tipurilor de ADN pot fi deter-
minate prin ultracentrifugare. Masa moleculară a ADN-ului poate fi determinată prin
compararea vitezei de sedimentare într-un gradient de densitate de zaharoză cu o probă
de ADN cu dimensiuni şi coeficient de sedimentare determinate în prealabil.
Sedimentarea la echilibru în gradient de CsCl este folosită pentru determinarea
densităţii de plutire a ADN-ului. Moleculele de ADN se concentrează într-o banda stabilă
la nivelul care densitatea de plutire este egală cu densitatea CsCl din acea zonă. ADN-ul
monocatenar are o densitate mai mare decât ADN-ul dublucatenar, care la rândul său are
în general o densitate mai mare decât proteinele. ARN-ul poate fi diferenţiat de ADN
(mono sau bicatenar) prin faptul că primul are o densitate mai mare. Densitatea de plutire
a ADN-ului poate furniza informaţii referitoare la ponderea perechilor G-C şi A-T din
moleculă. Mai mult, ADN-urile virale intacte, omogene, prezintă benzi înguste, în timp
ce fragmetele heterogene de ADN din celulele eucariotelor apar sub forma unor benzi
(grupuri de benzi) mai largi.

Biochimie - Curs 12 - 2019
7
Figura 7. Denaturarea (desfacerea)
ADN-ului dublu-catenar în mono-catenar
Denaturarea și renaturarea
În cazul în care o soluţie care conţine
duplex-ADN-ul este încălzită peste o anumită
temperatură, structura nativă a acestuia este alterată
rezultând separat cele două lanţuri complementare
care au conformaţii aleatorii (Figura 7). Astfel
procesul de denaturare este însoţit de schimbări
calitative în cazul proprietăţilor fizice ale ADN-ului
(scăderea vâscozităţii şi creşterea absorbanţei în
UV în cazul formei denaturate). Denaturarea ADN-
ului este un fenomen cooperativ în care alterările
dintr-o parte a moleculei destabilizează celelalte
legături (dintre cele două lanţuri rămase). Această
denaturare are loc într-un interval îngust de
temperatură. Mijlocul acestui interval poartă
numele de temperatură de topire, Tm (m = eng.
melting-topire). Stabilitatea duplex-ADN-ului
(ilustrată de Tm) depinde de o serie de factori:
natura solventului, tăria ionică, conţinutul de perechi G-C (care conţin o legătură de
hidrogen suplimentară în comparaţie cu perechea A-T) sau de pH.
În 1960, J. Marmur a arătat faptul că ADN-ul denaturat poate fi renaturat (să
revină la starea iniţială) în condiţiile în care temperatura este menţinută cu 25 mai jos
decât Tm.
Perechile de baze
Împerecherea bazelor este asemeni unui “relipiri” a celor două lanţuri din acizii
nucleici. În cazul modelului proous de Watson și Crick aceste perechi de baze se
formează între oligonucleotide complementare. Există însă şi situaţii diferite. De
exemplu, perechea de baze A-T poate avea atomul de azot din poziția 7 (perechea
Hoogsteen) drept acceptor şi nu cel din pozitia 1 care corespunde situaţiei clasice
(modelului Watson-Crick). Oricum, măsurătorile experimentale au arătat faptul că numai
perechile de baze Watson-Crick au o stabilitate mai mare față de celelalte perechi. Există
şi situaţii diferite în care segmentele ce se împachetează dublu-elicoidal, din multe ARN-
uri, conţin perechi de baze de tipul G-U care au rol în stabilizarea structurii terţiare a
acestora.
Figura 8. Tipuri de perechi de baze

Biochimie - Curs 12 - 2019
8
Interacţiunile de van der Waals
Purinele şi pirimidinele din interiorul structurii dublu elicoidale au tendinţa de a
se orienta într-o manieră paralelă prin intermediul interacţiunilor de tip van der Waals
(interacţiuni electronice de tip -) sau hidrofobe. Aceste interacţiuni contribuie în mod
esenţial la stabilizarea structurii ADN-ului.
Interacţiunile ionice
Interacţiunile electrostatice care au loc între grupările fosfat trebuie considerate,
alături de legăturile de hidrogen şi interacţiunile hidrofobe din acizii nucleici, factori care
contribuie la stabilizarea structurii acizilor nucleici. De exemplu, Tm a duplex-ADNului
creşte dacă concentraţia ionilor de Na+ este mărită. Acest fapt se datorează intercalării
acestor ioni între grupările fosfat. Analog ionii divalenţi, Mg2+, Mn2+ şi Co2+ se leagă
specific de grupările fosfat şi astfel constituie agenţi de protecţie a acizilor nucleici. Ionii
de Mg2+ joacă de asemenea un rol esenţial în stabilizarea structurilor complexe adoptate
de diverse molecule de ARN.
Reacția de polimerizare în lanț (PCR-The polymerase chain reaction)
Geneticienii au înțeles faptul că sunt necesare mai multe componente pentru a
realiza replicarea ADN-ului. În acest scop este nevoie de enzimă denumită ADN
polimeraza și de deoxinucleotide care constituie “cărămizile” de bază ale ADN-ului.
Reacția de polimerizare în lanț (PCR) este o reacție enzimatică de amplificare
mediată de primeri (secvențe scurte de ADN) specifici secvențelor de ADN genomic sau
clonat. Metoda PCR a fost inventată de Karry Mullis în 1983 și implică utilizarea unor
primeri (a căror lungime este de obicei 20-25 pb) și a unei ADN polimeraze termostabile,
cele mai utilizate fiind Taq, Pfu sau Pwo polimeraza. ADN-ul matriță conține secvența
țintă, care poate avea de la zeci la zeci de mii de nucleotide lungime. Tehnica PCR
standard realizează amplificarea unei singure secvențe de ADN care are o lungime de cel
puțin 5 kb (5000 de baze). Long PCR este tehnica prin care se pot amplifica fragmente
mai lungi de ADN (de până la 40 kb). A treia variantă de PCR, multiplex, este utilizată
pentru amplificarea unor secvențe multiple care au o lungime sub 5 kb. Taq polimeraza
catalizează reacția într-un sistem tampon, în care există un exces de perechi de primeri și
cele patru fosfat-deoxinucleotide (dNTP), generându-se astfel milioane de copii ale
secvenței țintă. PCR poate fi utilizat și pentru amplificarea secvenței de ARN, care
trebuie convertită inițial în ADN de enzima revers transcriptaza. Spre deosebire de ADN
trebuie luată în considerare instabilitatea și susceptibilitatea ARN-ului la degradare.
ADN-ul utilizat ca matriță pentru PCR poate proveni din diferite surse: sânge
liofilizat, salivă, țesut parafinat, păr. De asemenea reacția PCR poate fi utilizată pentru
amplificarea ADN-ului din materiale degradate: mumii sau fosile.
Replicarea ADNului in vitro - reactia PCR
Reacția PCR este constituită din serii de trei pași esențiali care definesc un ciclu
PCR: denaturarea ADN-ului matriță dublu catenar, alinierea perechilor de primeri la
matrițele de ADN monocatenar și extinderea enzimatică a primerilor, prin care se produc
copii care servesc drept matrițe în ciclurile ulterioare. ADN-ul matriță este suspendat într-

Biochimie - Curs 12 - 2019
9
un amestec alcătuit din apă distilată sterilizată (fără ARN), soluție tampon (ce conține
clorura de magneziu), polimeraza (Taq, Pfu sau Pwo polimeraza respectiv un amestec de
polimeraze) și cele patru dNTP. De asemenea există o pereche de primeri a căror
secvențe sunt complementare cu cele ale ADN-ului care flanchează regiunea țintă.
Pentru construirea primerilor se iau în considerare următoarele criterii:
- trebuie să aibă un conținut de baze în jur de 50% GC;
- lungimea trebuie sa fie între 15-30 pb;
- cei doi primeri trebuie să nu formeze duplexuri între ei;
- trebuie evitați primerii care conțin bucle;
- capătul 3’ trebuie să prezinte o complementaritate perfectă cu a ADN-ului țintă;
- temperatura de aliniere a primerilor sa fie între 50-64 ºC;
- temperatura la care jumătate din molecule sunt monocatenare și cealaltă jumă-
tate bicatenare este temperatura de topire (Tm). Temperatura de topire poate fi
aproximată după formula:
Tm = 2 ∙ (număr de baze A/T) + 4 ∙ (număr de baze C/G)
Temperatura de aliniere a primerilor este de obicei aleasă cu 5 ºC mai scăzută
decât temperatura de topire a ADN-ului. Din acest motiv este foarte importantă alegerea
temperaturii de topire a celor doi primeri.
Figura 9. Etapele reacției PCR

Biochimie - Curs 12 - 2019
10
Amestecul de reacție este mai întâi încălzit la temperatura de 94 ºC pentru
denaturarea (separarea) celor două catene de ADN (Figura 9) și apoi răcit la o
temperatură optimă care facilitează alinierea (alipirea acestora la secvența de ADN)
primerilor.
Primerii sunt orientați unul în amonte și unul în aval față de regiunea ce urmează
a fi amplificată, ambii cu capătul 3’ spre interiorul secvenței țintă. Poziția relativă a
regiunilor complementare ale acestor primeri determină lungimea secvenței de ADN care
va fi copiată.
În timpul extinderii primerilor ADN polimeraza adaugă progresiv dNTP-urile
complementar cu secvența matriță, la capătul 3’ al fiecărui primer, generându-se asftel o
nouă copie. Astfel prin cicluri repetate de răcire și încălzire se poate forma o cantitate
semnificativă de ADN a cărui secvență este identică cu aceea a ADN-ului matriță.
Rezultatul, după 20 de cicluri de amplificare, este producerea a 220 (peste un milion) copii
ale ADN-ului matriță.
Inițial exista un impediment legat de stabilitatea polimerazei care era utilizată la
replicarea ADN-ului. Enzima respectivă se degrada în momentul încălzirii amestecului la
temperatură optimă neceară pentru separara lanțurilor ADN-ului dublu-catenar. Din
fericire, biologii au extras diverse polimeraze din bacterii termofile, care traiau în medii
la temperaturi extreme. Prin această strategie s-au izolat polimeraze rezistente la
temperaturi mari. Taq polimeraza este o polimerază termostabilă care a fost extrasă din
bacteria Termus acvaticus (bacterie care trăiește în izvoarele calde). Unul dintre
dezavantajele folosirii Taq polimerazei este acela al fidelității scăzute (o eroare de
copiere la aproximativ 9000 de nucleotide), fapt datorat lipsei activității exonucleazice. În
schimb Pfu polimeraza, o polimerază termostabilă (enzima care are în componență
tungsten) numită după specia hipertermofilă anaerobă Pyrococcus furiosus (acest
organism se dezvoltă la temperaturi de 100 ºC), posedă și activitate de verificare a
corectitudinii polimerizării realizând astfel o amplificare cu o acuratețe ridicată a
secvenței ADN dorite. Pwo polimeraza a fost izolată din archeobacteria hipertermofilă
Pyrococcus woesei.
Cele aproximativ 30 de cicluri sunt realizate într-un aparat optimizat să oscileze
temperatura pe diferite intervale de timp, numit thermal cycler. Numărul de cicluri PCR
trebuie optimizat în functie de numărul de copii țintă dorit. Pornindu-se de la o singură
copie, cea mai eficientă reacție de PCR atinge în câteva ore un platou după 40 de cicluri
de amplificare. Acest proces este denumit PCR deoarece în decursul fiecărui ciclu
cantitatea de ADN se dublează. Mărimea fragmentului de ADN copiat rezultat în urma
PCR este controlată prin intermediul unei tehnici de separare numită electroforeză în
geluri de agaroză.
Aplicații ale tehnicii PCR
Unul din exemplele elocvente ale aplicabilității PCR-ului are rezonanță istorică.
Această tehnică a permis demascarea unui impostor care pretindea a fi membru al
familiei imperiale ruse. În anul anul 1918, Nicolae Romanov II, ultimul țar al Rusiei,
împreună cu familia au fost asasinați în timpul revoluției bolșevice. Ei au fost înhumați
într-un loc nemarcat. În 1993, oasele lor au fost supuse testului de identificare a ADN-
ului. Acest lucru a fost posibil prin comparerea secvențelor de ADN ale soției țarului
Romanov cu acelea ale prințului Philip de Edinburg, soțul regiei Elizabeta II. Interesant,
la scurt timp după asasinarea familiei regale, au circulat zvonuri referitoare la

Biochimie - Curs 12 - 2019
11
supraviețuirea unei fiice a țarului, Anastasia, în urma asasinatului. O persoană a convins o
parte din nobilimea rusă din Berlin de apartenența sa la familia regală. Mai târziu, această
persoană a emigrat în SUA unde a decedat în 1984. Prin intermediul unei probe de țesut
care era depozitată într-un spital unde aceasta a fost supusă unei intervenții, ADN-ului
acesteia a putut fi amplificat. În urma investigațiilor s-a demonstrat faptul că împărăteasa
Alexandra și printul Filip nu sunt rude cu această persoană.
Mai mult, aplicațiile Reacției PCR sunt diverse:
- în criminalistică;
- manipularea genetică;
- detecția HIV (ADN-ul este izolat din celulele roșii și amplificat cu primeri
corespunzători secvențelor HIV);
- diagnosticare prenatală a bolilor genetice.
Balizele moleculare (BM)
Tehnologia balizelor moleculare a fost introdusă începând cu anul 1996 de către
Tyagi şi Kramer. În această tehnică o probă fluorescentă (etichetată la unul din capete cu
un fluorofor și la altul cu un stingător) a fost utilizată pentru a dovedi prezenţa unor
secvenţe complementare atribuite unor acizi nucleici din soluţie. O baliză moleculară este
o oligonucleotidă sub formă de ac conţinând o secvenţă ţintă specifică flancată de două
secvenţe complementare care pot hibridiza pentru a forma o tijă. Majoritatea probelor au
25-40 nucleotide; tija poate conţine 5-10 pb. Un fluorofor şi un stingător adecvat sunt
plasaţi la capetele 3’ şi 5’ ale tijei şi semnalul fluorescenţei este mediat de transferul de
energie prin rezonanţă. În absenţa secvenţei complementare, fluorescenţa fluoroforului
de pe tija balizei (BM) este stinsă de către stingătorul adiacent. Legarea unei BM la
secvenţa ţintă induce o disociere a tijei, fapt care determină o creştere a distanţei dintre
fluorofor şi stingător, conducând la o creştere a intensităţii fluorescenţei.
O BM se poate lega reversibil sau să disocieze de secvenţa ţintă, şi acestă
reversibilitate poate afecta sensibilitatea si selectivitatea detecţiei. Performanţa tehnicii
este dictată de o serie de factori (temperatură, pH sau conţinutul secvenţei tijei şi buclei).
La o temperatură scăzută, o BM este caracterizată prin stabilitate ridicată a intra-
duplexului chiar în prezenţa secvenţei ţintă. Din acest motiv fluorescenţa este scăzută
datorită stingerii intramoleculare a BM care nu hibridizează cu secvenţa ţintă (Figura 10).
Figura 10. Tehnica hibridizării ADN-ului
La o temperatură optimă, BM va fi instabilă şi va hibridiza cu secvenţa de interes,
rezultând un semnal puternic pentru fluorescenţă. În final, la o temperatură mai mare,

Biochimie - Curs 12 - 2019
12
porţiunile de pe tija BM şi complexul BM-ADN ţintă sunt instabile dat fiind faptul că
BM rămâne într-o conformaţie liniară.
BM pot fi folosite în numeroase aplicaţii: detecţia agenţilor patogeni şi la
stabilirea genotipului alelelor.



























