Embed Size (px)
DESCRIPTION
Biosintetis asam lemak
Citation preview

B. METABOLIK ANABOLISME
1. BIOSINTESIS ASAM LEMAK
PENDAHULUAN
Seperti proses penguraian dan sintesis lainnya (misal, glikogenolisis dan
glikogenesis), sintesis asam lemak (lipogenesis) dahulu dianggap sebagai reaksi
pembalikan oksidasi di dalam mitokondria. Namun, sekarang jelas bahwa sistem ekstra
mitokondria yang sangat aktif bertanggung jawab untuk sintesis asam lemak yang
lengkap dari asetil-KoA dalam sitosol. Sistem lain untuk perpanjangan rantai asam lemak
juga terdapat diretikulun endoplasmik hati. Di dalam bab ini akan dibahas tentang sintesis
asam lemak palmitat.
Kompetensi khsus, setelah pembahasan sintesis asam palmitat, peserta
pembelajaran mampu menjelaskan tahap-tahap reaksi yang terjadi selama proses sintesis
berlangsung.
LINTASAN UTAMA SINTESIS DE NOVO ASAM PALMITAT.
Sistem sintesis asam palmitat terdapat pada jaringan tubuh termasuk jaringan
hati, ginjal, otak, paru-paru, kelenjar payudara dan adiposa . Sistem ini membutuhkan
kofaktor yang mencakup NADPH, ATP, Mn2+, biotin dan HCO3- (sebagai sumber CO2).
Di dalam sintesis tersebut, asetil-KoA digunakan sebagai substrat dan produk akhirnya
adalah palmitat bebas.
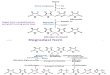
Bikarbonat (HCO3-) digunakan sebagai sumber CO2 pada reaksi pendahuluan
untuk karboksilasi asetil-KoA menjadi malonil KoA dengan adanya ATP dan enzim
asetil-KoA karboksilase. Enzim memindahkan karboksil pada asetil-KoA untuk
membentuk malonil-KoA (gambar 1-1).
Gambar 1-1. Pembentukan malonil-KoA dari asetil-KoA

Ada dua tipe enzim sintase asam lemak, tipe pertama yang terdapat pada
bakteri, tumbuhan dan bentuk organisme lebih rendah lainnya. Masing-masing enzim
pada organisme tersebut saling terpisah, radikal asil terdapat dalam bentuk kombinasi
dengan protein yang disebut protein pembawa radikal asil (ACP-aCye carrier protein).
Tipe kedua, terdapat pada ragi, mamalia dan burung. Pada organisme tersebut, sistem
sintase berupa komplek polipeptida multienzim yang tidak dapat dipisahkan lagi tanpa
kehilangan aktivitasnya, dan ACP adalah bagian dari kompleks tersebut. Komplek
multienzim ini dikode oleh gel tunggal.
Kompleks enzim sintase asam lemak merupakan sebuah dimer (Gambar 1-2).
pada mamalia, setiap dimer identik, dan terdiri atas satu rantai polipeptida yang bisa
ditandai serta mengandung tujuh aktivitas enzim sintase asam lemak dan sebuah ACP
dengan gugus 4’-fosfopantotein-SH. Didekatnya terdapat gugustiol (-SH) dari residu
sistein 3-ketoasil sintase (enzim kondensis).
Gambar 1-2. Kompleks enzim sintase asam lemak

TAHAP-TAHAP REAKSI PADA SINTESIS ASAM PALMITAT
Tahap reaksi diawali oleh kondensasi asetil-KoA dengan gugus sistein-SH yang
dikatalisis oleh asetil transasilase (gambar 1-3).
Gambar 1-3. Tahap reaksi sintesis asam lemak

Malonil –KoA bergabung dengan A’-fosfopantotein ACP yang dikatalisis oleh
malonil transamilase membentuk kompleks enzim asetil (asil)-malonil. Gugus asetil
menyerang metilen pada residu malonil dengan katalis 30 ketoasil sintase yang
membebaskan CO2 dan membentuk kompleks enzim-ketoasil (enzim-asetoasetil). Dalam
reaksi ini gugus sistein-SH yang tadinya ditempati oleh gugus asetil dibebaskan. Tahap
reaksi selanjutnya, adalah reduksi, dehidrasi dan reduksi kembali gugus.
Molekul malonil-KoA baru, yang dibutuhkan untuk perpanjangan rantai,
bergabung dengan gugus-SH pada 4’-fosfopantotein menggantikan residu asiljenuh pada
gugus-SH sistein bebas. Rangkaian reaksi tersebut diulang lebih dari 6 kali sampai
tersusun residu asil 16-karbon (palmitil) yang jenuh. Radikal ini, kemudian dibebaskan
dari enzim oleh aktivitas enzim ketujuh dalam kompleks tersebut yaitu tioesterase
(deasilase), dan membebaskan asam palmitat bebas. Persamaan reaksi keseluruhan
sintesis palmitat dari asetil-KoA dan malonil-KoA adalah.
CH2.CO-S.KoA + 7HOOC.CH2CO ~ S-KoA + 14 NADPH + 14H+
CH3 C(CH2)14 COOH + 7 CO2 + 6H2O +8 KoA-SH + 14NADP+
Asetil-KoA yang digunakan sebagai primer reaksi sintesis asam palmitat,
membentuk atom karbon 15 dan 16 pada asam lemak tersebut. Sedang untuk penambahan
unit C2 pada perpanjangan rantai, berlangsung lewat pembentukan malonil-KoA. Jika
penyiap reaksi adalah propinil-KoA asam lemak rantai panjang yang terbentuk adalah
berantai atom karbon ganjil.
Asetil-KoA yang merupakan molekul pembangun utama asam-asam lemak
dapat dibentuk dari karbohidrat melalui oksidasi di dalam mitokondria. Dalam keadaan
kenyang, enzim ATP-sitrat liase ekstra mitokondria menjadi aktif memecah sitrat menjadi
oksaloasetat dan asetil-KoA. Reaksi pembentukan asetil-KoA ini melibatkan reaksi-
reaksi yang terjadi di dalam mitokondria dan proses translokasi ke luar mitokondria.
Selanjutnya asetil-KoA yang terbentuk digunakan untuk mengawali sintesis asam lemak
palmitat dan asam-asam lemak lainnya (gambar 1-4)

Gambar 1-4. Pembentukan asetil-KoA dari karbohidrat melalui di dalam mitokondria
NADPH yang terlibat dalam sintesis asam palmitat sebagai donor ekuivalen
pereduksi pada tahap reaksi reduksi derivat 3-ketoasil maupun 2,3 asil tak jenuh, sumber
utamanya adalah oksidasi glukosa pada jalur pentosa fosfat. Sumber lain mungkin bukan
utama adalah mencakup reaksi konversi malat menjadi piruvat yang dikatalisis oleh
enzim malat (NADP malat dehidrogenase) dan reaksi isositrat dehidrogenase.
PEMANJANGAN RANTAI ASAM LEMAK
Pemanjangan rantai asam lemak yang disintesis terjadi di dalam retikulum
endoplasma (sistem mikrosom). Di dalam sistem ini untuk memperpanjang rantai asam
lemak terjadi melalui penambahan dua atom karbon. Sumber atom karbon yang
dibutuhkan adalah menggunakan molekul malonil-KoA sebagai donor asetil dan
refuktornya adalah NADPH. Reaksi ini dikatalisis oleh asam lemak elongase mitokondria
(gambar 1-5).
PENGATURAN LIPOGENESIS
Status nutrisi organisme merupakan faktor utama yang mengatur laju
lipogenesis. Pada hewannya yang diberi makan kenyang dan dietnya mengandung
karbohidrat dengan proporsi tinggi, maka lipogenesisnya tinggi, sebaliknya laju
lipogenesisnya akan berkurang pada keadaan asupan kalori yang terbatas, diet tinggi
lemak atau jika terdapat defisiensi insulin. Semua keadaan tersebut berkaitan dengan
peningkatan konsentrasi asam lemak bebas di dalam plasma.

Gambar 1-5. Pemanjangan rantai asam lemak di dalam mikrosom

Sintesis asam lemak rantai panjang juga diatur oleh modifikasi enzim secara
alosterik dan kovalen dalam waktu yang pendek dan oleh perubahan ekpresi gen dalam
jangka waktu panjang. Pengaturan lipogenesis yang paling penting adalah terletak pada
tahap asetil-KoA karboksilase. Sifat enzim ini adalah alosterik dan diaktifkan oleh sitrat.
Pada keadaan kenyang konsentrasi sitrat meningkat, asetil-KoA juga tinggi akibatnya
lipogenesis menjadi cepat akan tetapi, karena asetil-KoA juga dapat menghambat asetil-
KoA karboksilase, maka lipogenesis menjadi berkurang. Lipogenesis juga dapat
dihambat oleh lipolisis atau aliran asam lemak bebas ke dalam jaringan yang
mengakibatkan penghambatan sintesis asam lemak baru.
Hubungan terbalik antara konsentrasi asam lemak bebas dan proporsi enzim
piruvat dehidrogenase yang aktif dan in aktif yang mengatur tersediaanya asetil-KoA,
juga ikut mengatur lipogenesis. Asil-KoA mengakibatkan inhibisi piruvat dehidrogenase
dengan menghambat pengangkutan pertukaran ATP-ADP pada membran interna
mitokondria. Penghambatan ini menyebabkan peningkatan rasio (ATP/ADP)
intramitokondria yang mengakibatkan perubahan bentuk piruvat dehidrogenase aktif
menjadi bentuk in aktif. Demikian pula oksidasi asil-KoA, akibat peningkatan kadar asam
lemak bebas dapat meningkatkan rasio (asetil-KoA)/KoA) dan (NADPH)/(NAD+) dalam
mitokondria yang dapat menghambat piruvat dehidrogenase.
Lipogenesis juga dirangsang oleh insulin melalui beberapa mekanisme. Insulin
meningkatkan pengangkutan glukosa ke dalam sel (misal, jaringan adiposa) sehingga
meningkatkan ketersediaan piruvat untuk sintesis asam lemak maupun gliserol 3-fosfat
untuk esterifikasi asam lemak yang baru terbentuk. Insulin mengkonversi piruvat
dehidrogenase bentuk inaktif menjadi aktif dalam jaringan adiposa. Insulin juga
mengaktifkan asetil-KoA karboksilase yang melibnatkan defosforilasi oleh protein
fosfatse. Hormon ini juga mampu menekan kadar cAMP intraseluler oleh karenanya
menghambat lipolisis di jaringan adiposa sehingga mengurangi asil-KoA rantai panjang
yang merupakan inhibitor lipogenesis.
Dengan mekanisme yang sama, insulin mengantagonis kerja hormon glukagon
dan epinefrin yang menghambat asetil-KoA karboksilase dan oleh karenanya
menghambat lipogenesis. Enzim kompleks sintase dan asetil-Koa dekarboksilase,
beradaptasi dengan kebutuhan fisiologis tubuh lewat melalui peningkatan jumlah totalnya

pada keadaan kenyang dan penurunannya pada keadaan puasa, konsumsi lemak serta
pada penyakit diabetes. Jadi kedua enzim tersebut adalah merupakan enzim adaptif.
RANGKUMAN
1. Lipogenesis dilakukan oleh dua sistem enzim yang terdapat pada silosel sel yaitu :
asetil-KoA karboksilase dan sintase asam lemak.
2. Lintasan lipogenesis, mengkonversi asetil-Koa menjadi asam palmitat yang
memerlukan NADPH, ATP, Mn2+, biotin, asam pantotenat dan HCO3- sebagai
kofaktor.
3. Asetil-KoA karboksilase mengkatalisis konversi asetil-KoA menjadi malonil-
KoA, yang selanjutnya dilanjutkan dengan tujuh aktivitas enzimatik yang
berbeda, akan mengkalisis susunan palmitat dari satu molekul asetil-Koa dan
tujuh molekul malonil-KoA.
4. Pemanjangan rantai asam lemak rantai panjang berlangsung dalam retikulum
endoplasma dan dikatalisis oleh sistem enzim alongase mirkosom.
5. Lipogenesis diatur pada tahap asetil-koA karboksilase oleh pemodifikasi alosterik,
modifikasi kovalen dan induksi serta represi sintesis enzim. Aktivasi enzim
tersebut diatur oleh kadar sitrat dan aktivitasnya dihambat oleh asetil-KoA rantai
panjang. Insulin mengaktifkan asetil-KoA karboksilase dalam waktu pendek
melalui defosforilasi dan dalam waktu pendek melalui induksi sintesis. Glukagon
dan epinefrin mempunyai kerja yang berlawanan dengan insulin.
KEPUSTAKAANKoolman, Jan ; Klaus-Heinrich Röhm. 1994. Atlas Berwarna dan Teks Biokimia. Jakarta : Hipokrates
Lehninger, Albert L. 1994. Dasar-dasar Biokimia Jilid I. Jakarta : Erlangga
Murray, Robert K ; Daryl K.G. ; Peter A.M.; Victor W.R. 2000.Biokimia Harper edisi 25. Jakarta : Buku
Kedokteran EGC
Salati L.M. goodbridge AG: Fahy acid Synthesis in eukaryotes In biochemistry of Lipids.
Smith., Hill RL, Lehman IR., Lefkowitz RIP., White A. 1993. Principle of Biochemistry : Mammalian
Biochemistry, 7th ed. Mc Graw Hill Co. New York

Talwar, GP. 1980. Textbook of Biochemistry and Human Biology Prentice. New Delhi, India.