Embed Size (px)
Citation preview

Biotecnologie ambientaliaa 2012-2013

Programma di Biotecnologie ambientaliModulo II - 3 crediti (dr. P. Morandini)
• Piante coltivate e piante selvatiche: la sindrome da domesticazione e i relativi caratteri principali (non-shattering e dormienza ridotta) e le implicazioni per l’invasività e la persistenza nell’ambiente.
• Rischi e benefici ambientali delle piante transgeniche in paragone a quelle convenzionali.
• Convenzione di Rio, Protocollo di Cartagena e normativa sulle piante create con tramite ingeneria genetica (scopi, effetti).
• Piante per una maggiore sostenibilità ambientale (plastiche biodegradabili), per il risanamento di siti contaminati (approcci, specie in uso, tipi di contaminanti rimossi).
• Interazione simbiotiche pianta-microrganismo: fissazione dell’azoto (batteri azoto fissatori e funghi vescicolo arbuscolari.
• Interazione pianta-microrganismo: le risposte di difesa delle piante e generazione di specie resistenti.

Che cosa è la domesticazione?
• Definizione di domesticazione: “Processo tramite il quale le piante o gli animali selvatici vengono adattati all’uomo e all’ambiente che egli fornisce”
• Ben di più che il semplice allevare in cattività, domare o coltivare
• Processo di selezione che porta a caratteristiche morfologiche, fisiologiche, genetiche e “comportamentali” ereditabili
• In molti casi esistono progenitori selvatici (o loro discendenti)
• I principali caratteri sotto selezione sono stati identificati (“sindrome da domesticazione)
• Lasso di tempo richiesto: (1.000 – 10.000 anni)
Perché si parla di “sindrome”? Sindrome (gr. Syn = con + dromos = circuito, strada. Stessa radice del verbo gr. dramai, agire, manifestare, da cui anche dramma). Aggregazione di segni, sintomi e/o altre manifestazioni considerate parte di un'entità morbosa.

Accumulo di mutazioni dannose per la pianta, ma favorevoli all’uomo (oltre a quelle dannose tramite linkage)
Seme trattenuto a maturità della spiga (frutto)
Riduzione del contenuto di tossine (patogeni…)
Dimensioni, forma e colore del frutto/seme/parte commestibile
Aumento nel numero di semi/fertilità
Nanismo/accestimento (in generale “plant architecture”)
Dormienza del seme (infestanza, permanenza..)
Richieste nutrizionali (fertilizzanti…)
Adattamento della fioritura alle condizioni locali
La sindrome da domesticazione

Dimensioni del frutto/parte commestibile: due caratteri evidenti che distinguono una pianta domesticata dalla sua controparte selvatica

Pomodoro selvatico / cultivar moderna
Per migliaia di anni l’uomo ha selezionato tra le varianti offerte spontaneamente da madre natura
Il risultato di questo lento processo (accumulo di “mutazioni”) è stata la creazione di piante coltivate a partire da quelle selvatiche

• Sindrome da domesticazione
Qu’est ce qu’une plante domestiquée?
• Rendement prioritaireBanane, arancie, uvetta scomparsa dei semi
Sterilità propagazione vegetativa
Diapositiva di Benoît Pujol

•Fully domesticated plants are characterized by a similar set of traits that confer adaptation to the human environment
•The two most important traits of the syndrome are loss of seed dispersal and reduced seed dormancy
•The specific details for each trait and the affected genes depend on the crop
•Fully domesticated plants have most of the traits of the syndrome but generally not all of them; within a crop, differences in the degree of domestication can be observed
Esempi di piante con sindrome “incompleta”:
Girasole
Colza
...
The domestication syndrome: the essentials

La selezione fatta dall’uomo che effetti ha sulla pianta?
Avena selvatica
Avena coltivata

Un bel carattere
Spighetta in una pianta matura:i semi cadono a terra entro pochi giorni.
Spighetta in una pianta matura:i semi rimangono per anni attaccati alla spighetta. Il loro distacco richiede un trattamento vigoroso (trebbiatura).

Orzo selvatico (murino) - coltivato

Teosinte – Mais
Semi di teosinte
Semi di mais
i semi si staccano facilmente
Per staccare i semi occorre un’azione vigorosa

I mutanti ci mettono in evidenza cambiamenti nel DNA che hanno un effetto sul fenotipo.
Ci permettono di scoprire geni importanti per i vari processi
Ci dicono che esistono dei geni ma non dove si trovino o quali siano.
Come possiamo identificarli?
Possiamo utilizzare queste conoscenze (quali siano i geni e dove siano) per sviluppare nuove varietà?
Per entrambe le cose si sfruttano i marcatori molecolari
- permettono di mappare / identificare i geni responsabili dei caratteri- sono usati per selezionare più velocemente le migliori piante nei programmi di miglioramento genetico: è il cosiddetto MAB (Marker Assisted Breeding) o MAS (Marker Assisted Selection), la selezione assistita da marcatori.

F2
P2
F1
P1 x
Grande popolazione con migliaia di piante
Non dormienteDormiente
Utilità dei marcatori molecolari: identificare i geni
genotipizzazione (determinare gli alleli del marcatore presenti nei diversi individui
Marcatore molecolare polimorfico tra i due genitori
Fenotipizzazione
Si combina l’informazione sul genotipo con quella sul fenotipo:
la maggior parte degli individui con il carattere delle dormienza mostrano uno solo dei due alleli del marcatore il marcatore è vicino al gene.
Ci sono solo due individui che deviano, è avvenuto un crossing over tra il marcatore e l’allele della domienza
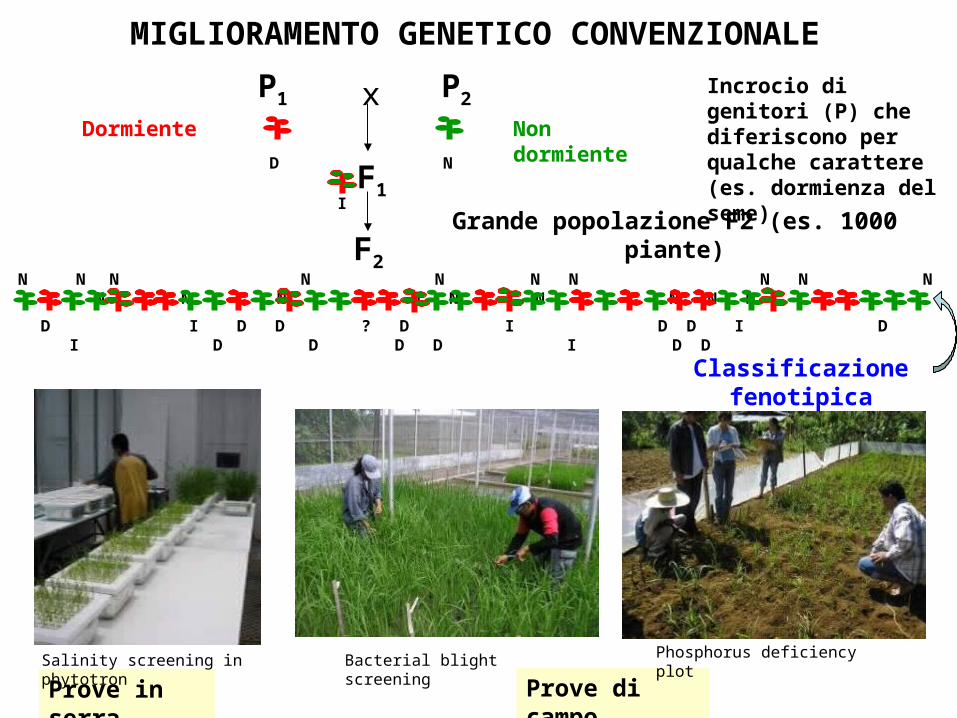
F2
P2
F1
P1 x
Grande popolazione F2 (es. 1000 piante)
Classificazione fenotipica
Prove di campoProve in serra
MIGLIORAMENTO GENETICO CONVENZIONALE
Salinity screening in phytotron Bacterial blight screening Phosphorus deficiency plot
Non dormienteDormiente
Incrocio di genitori (P) che diferiscono per qualche carattere (es. dormienza del seme)D N
N N N N N N N N N N N N N N N N N N N
D I D D ? D I D D I D I D D D D I D D
I

F2
P2
F1
P1 x
Grande popolazione con migliaia di piante
Non dormienteDormiente
si sfrutta il linkage disequilibrium (associazione allelica) tra il gene desiderato ed un marcatore molecolare prossimo ad esso
Utilità dei marcatori molecolari: MAS
In questo modo è possibile prevedere quali individui possiedano alleli di interesse
genotipizzazione
Marcatore molecolare polimorfico tra i due genitori

I marcatori devono essere polimorfici nei genitori
1 2 3 4 5 6 7 8RM296
P1 P2
Polymorphic!
1 2 3 4 5 6 7 8
RM84
P1 P2
Not polymorphic

…TCGCCTGAGAGAGAGAGATCCC…
…TCGCCTGAGAGATCCCCGTCCA…
…AAGATTGGATCCCCATATCCA…
…AAGATTAGATCCCCATATCCA…
Marcatore A (indel) Marcatore B (SNP)Gene X
P1
P2
allele X1 (P1)
allele X2 (P2)
allele A1 (P1)
allele A2 (P2)
allele B1 (P1)
allele B2 (P2)
Fie
sta
Dis
cove
ryG
ala
Flo
rin
a
Nov
a E
asyg
ro
TN
10-8
Du
rell
o d
i For
lì
Pri
ma
Mod
ial G
ala
Fu
ji
L’allele X1 del gene X e gli alleli (A1 e B1) dei due marcatori molecolari (A e B) che derivano dallo stesso genitore (P1) tendono a segregare insieme alla meiosi. Analogamente per gli alleli dell’altro genitore (P2).
Marcatore molecolare del tipo SSR con amplificati variabili (230-257 bp) tra diverse cultivar di melo. Immagine di L. Gianfranceschi.

P1 x P2
F1
Screening fenotipico
F2
F3
F4
F5
F6
F7
F8 – F12
Phenotypic screening
Plants space-planted in rows for individual plant selection
Families grown in progeny rows for selection.
Preliminary yield trials. Select single plants.
Further yield trials
Multi-location testing, licensing, seed increase and cultivar release
P1 x P2
F1
F2
F3
MAS
MAS per individuare i candidati da sottoporre ad analisi fenotipiche
F4Families grown in progeny rows for selection.
Pedigree selection based on local needs
F6
F7
F5
F8 – F12Multi-location testing, licensing, seed increase and cultivar release
Only desirable F3 lines planted in field
La MAS permette di concentrare l’indagine su poche linee

Immagine dell’IRRI (Los Banos, Filippine)
Swarna: sensibile alla sommersione
varietà Swarna SUB1 resistente alla sommersione
Swarna SUB-1 è come Swarna ma contiene il gene SUB1 che conferisce tolleranza alla sommersione. SUB1 deriva da un’altra varietà; la sua presenza nei vari stadi del backcross è stata rivelata tramite MAS
Un esempio di MAS

14-day-old seedlings were submerged for 14 days and photographed 14 d after de-submergence
The Sub1 region donor line IR49830 (an FR13A derivative) was introduced into the submergence-intolerant indica variety Swarna by backcrossing (BC) with MAS
P2 P1

Genotipo grafico di Swarna-Sub1
Linea BC3F2: contiene circa 2,9 MB di DNA del donatore
Una linea di questo tipo è detta NIL (nearly isogenic line) perchè contiene solo un frammento di ungenitore, il resto proviene dall’altro (introgressione)

Il gene qSH1 di riso
• Uno dei geni che conferiscono un fenotipo non-shattering
• Carattere quantitativo (diversi genei posson concorrere al fenotipo
• Presente in diverse (ma non tutte) le varietà.
• Mappato al fondo del cromosoma 1 di riso
• Clonato nel 2006

Konishi et al., (2006) Science 312:1392-1396
Chromosomal locations of QTLs for seed-shattering degree, based on an F2 population
Nipponbare Kasalath
QTL (quantitative trait locus) analysis between a shattering-type indica cultivar, Kasalath, and a nonshattering type japonica cultivar, Nipponbare.
Five QTLs were detected on five chromosomes; all contributed to shattering reduction loss of seed shattering occured independently in japonica and indica.
Most rice cultivars have somehow lost the seed-shattering habit use natural variations in seed shattering among cultivars.
Generally, indica cultivars exhibit relatively strong seed shattering, whereas some japonica cultivars do not exhibit it at all.

marcatorePosizione
(in cM)
0,6
7,0
12,2
19,9
25,4
43,2
52,1
58,1
64,9
74,1
103,1
103,7
116,5
118,9
123,5
124,8
127.3
129,3
134,2
140,5
142,4
157,6
167,2
169,5
176,3
181,8
C602
C1679
C30
C470
G359
G317
C1211
R1712
S1501
R596
G302
R2374
S2717
R1012
S1870
C137
R1416
C1282
R758
G370
R2125
R2414
R503
S10581
R117
C112
Centromero
SP1
LOG
Gn1a
qSH1
qSD1
Mappa di associazione del cromosoma 1 di riso. In rosso sono evidenziati alcuni geni, tra cui qSH1.
Nipponbare alleles
Kasalath alleles

Increase in value = loss of shattering =non shattering
= Nipponbare, ma con un pezzo di cromosoma (che contiene qSH1)che deriva da Kasalath
Breaking tensile strength upon detachment of seeds from the pedicels by bending and pulling
The QTL with the largest effect, termed QTL of seed shattering in chromosome 1 (qSH1), explained 68.6% of the total phenotypic variation in the population. We therefore made a near-isogenic line (NIL) that contained a short chromosomal segment from Kasalath at the qSH1 region in a Nipponbare genetic background.

position of abscission layer formation Scanning electron microscope (SEM) photos of
pedicel junctions after detachment of seeds.
Nipponbare
Kasalath
NIL (qSH1)
= Nipponbare ma shattering
abscission layer
The NIL exhibited the formation of a complete abscission layer between pedicel and spikelet at the base of the rice seed
A seed-shattering phenotype stronger than either Kasalath or Nipponbare

C434 was the nearest marker to qSH1 upon the rough mapping. MarkersC283 and R3265 were used to select recombinants near the qSH1 locus.
B and A are homozygous for Kasalath and Nipponbare, respectively
Hanno cercato eventi di ricombinazione tra i due marcatori in 10.388 linee segreganti

Ci sono alcune differenze nella zona trascritta
Il gene di riso è simile a geni caratterizzati in Arabidopsis

No distinct open reading frame (ORF) in the SNP region. Located 12 kb away from the SNP, there is one ORF for a rice ortholog of the Arabidopsis REPLUMLESS (RPL) gene
Come si può spiegare il dato?
Il gene di Arabidopsis
a BEL1-type homeobox) and is involved in the formation of a dehiscence zone (or abscission layer) alongside the valve in the Arabidopsis fruit (silique).

In situ analysis of qSH1 expression.qSH1 è espresso nella zona dove si formerà la zona di abscissione
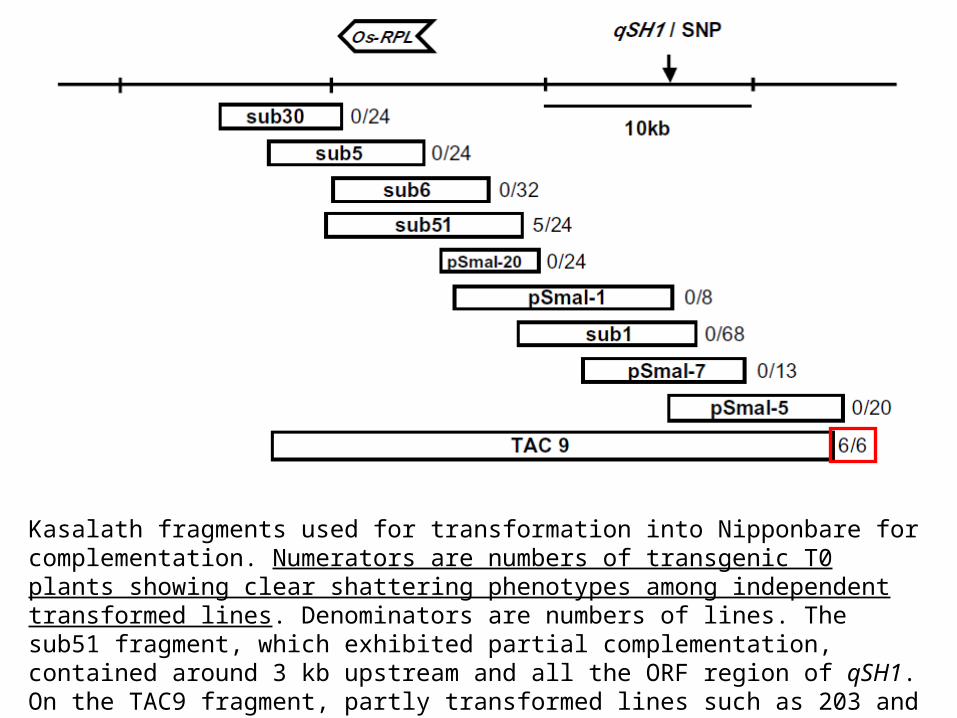
Kasalath fragments used for transformation into Nipponbare for complementation. Numerators are numbers of transgenic T0 plants showing clear shattering phenotypes among independent transformed lines. Denominators are numbers of lines. The sub51 fragment, which exhibited partial complementation, contained around 3 kb upstream and all the ORF region of qSH1. On the TAC9 fragment, partly transformed lines such as 203 and 204 in Fig. 2 were excluded.

Complementazione di Nipponbare con un frammento genomico di Kasalath da 26 kbp (linee 1, 201, 203 e 204): la complementazione è evidente solo nella linea 1 e 201 (è come la NIL)
Dall’analisi risulta che 1 e 201 contengono il DNA intatto, mentre 203 e 204 risultano troncati (dots and crosses indicate DNA markers used to confirm the transformed and nontransformed parts).
these lines lost the ORF region upon transformation
nonshattering of T0 plants
Only transgenic lines that contained the Kasalath fragment with both the ORF and the SNP exhibited complete seed shattering,

Non-shattering degrees of T1 selfing progeny in Line 1 (see two slides above) which was complemented
transg
ene
lost b
y seg
regatio
n
V, vector control N, Nipponbare

Temperate japonica cultivars of Chinese origin
Selection for the qSH1 allele was not as intense nor as expansive as selection for the sh4 allele, as qSH1 is not fixed, even within the temperate japonica subpopulation, let alone the whole of O. sativa
Cultivars in temperate japonica
Controls: Kasalath & Nipponbare
La presenza di “G” a qSH1 correla bene con un fenotipo shattering, mentre la presenza di “T” correla bene con non-shattering specialmente nelle cultivar di origine cinese.Non è però 100% ci sono altri geni che influenzano il fenotipo

Statistical analysis of the association of seed shattering with genotype
Andando ad esaminare polimorfismi fiancheggianti qSH1 si trova che uno è associato con lo shattering/non-shattering proprio perchè è associato a qSH1
Tutte le evidenze prese insieme permettono di concludere che:
- this RPL ortholog is the qSH1 gene
- the identified SNP affected only the spatial mRNA expression pattern of qSH1 at the abscission layer
the SNP was highly associated with the degree of seed shattering among temperate japonica rice cultivars (a subgroup of japonica) and implied that this SNP had been a target of artificial selection for non-shattering habit during rice domestication

Nipponbare: ...TCATGAAATGT...
Kasalath: ... TCATGCAATGT...
Pro
mo
tor
eR
egio
ne
tras
crit
ta
Idealmente basta il cambiamento di una sola base sull’intero genoma (400 milioni di basi) per stravolgere totalmente la biologia riproduttiva di una specie

Sulla base dei vari polimorfismi (tra cui qSH1) è possibile cercare di ricostruire la storia della domesticazone del riso
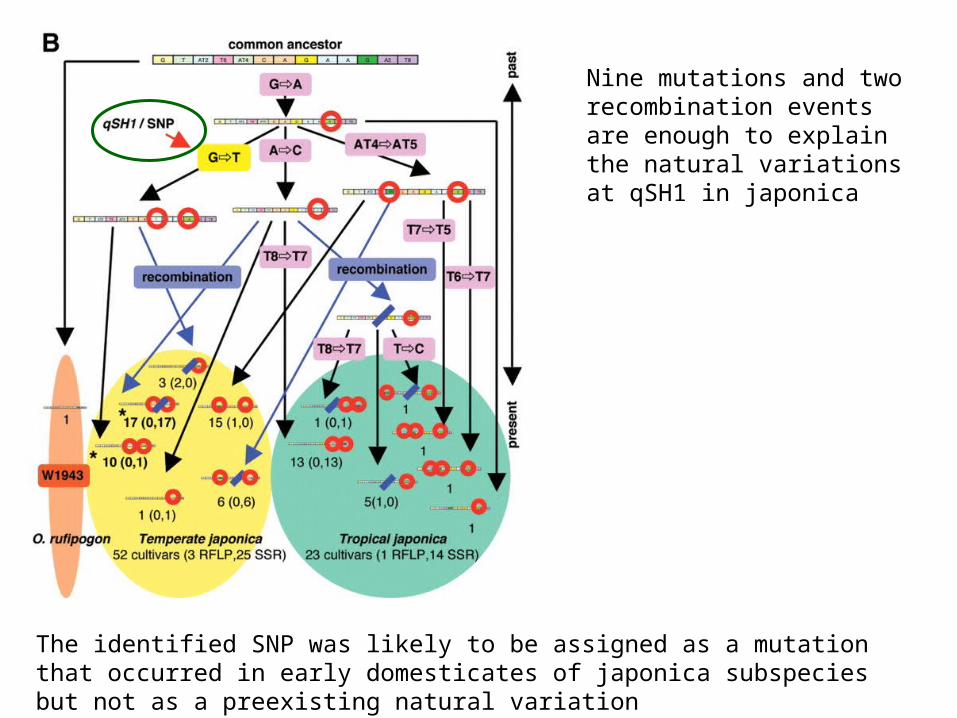
The identified SNP was likely to be assigned as a mutation that occurred in early domesticates of japonica subspecies but not as a preexisting natural variation
Nine mutations and two recombination events are enough to explain the natural variations at qSH1 in japonica

Rice domestication by reducing shattering
Genetic analysis of an F2 population derived between O. sativa ssp. indica and the wild annual species O. nivara identified three quantitative trait loci (QTL) -sh3, sh4, and sh8- responsible for the reduction of grain shattering in cultivated rice.
Gene sh4
Li et al (2006)

The remaining accessions of the wild species with confirmed shattering differed invariably from the cultivars by one mutation, d, which was a nucleotide substitution of G for T or an amino acid substitution of asparagine for lysine in O. sativa.

Subcellular localization of sh4. Rice roots transformed with Ubi::sh4-GFP
epidermal cells
DAPI sh4-GFP Merge
La proteina SH4 è una proteina nucleare

Expression of sh4
RNA from leaves (L) and the flower/grain and pedicel junction 3 days before flower opening (–3), the day of flower opening (1), and every 3 days thereafter during seed development.
Real-time RT-PCR estimate of relative expression of sh4
Force required to pull flowers or grains away from pedicels on:
Days starting from flower opening
Force required to pull away grains of transformants

First, qSH1 was selected during the domestication of japonica rice, but was subsequently selected against and eliminated from most of the japonica cultivars after sh4 was introduced from cultivars of different origins, such as indica. The combination of qSH1 and sh4 could have made threshing too difficult and laborious at the time. Second, qSH1 was derived in the japonica cultivars that already had sh4 to further reduce shattering when the requirement for stronger threshing force was no longer a problem.
The sh4 mutation preceded and spread more widely in landraces than the qsh1 mutation. Because the qsh1 mutation, but not the sh4 mutation, caused abscission-layer loss, it is very likely that the archeological data on the short rachillae indicate that the qsh1 mutation had already been selected by 7000 bc.
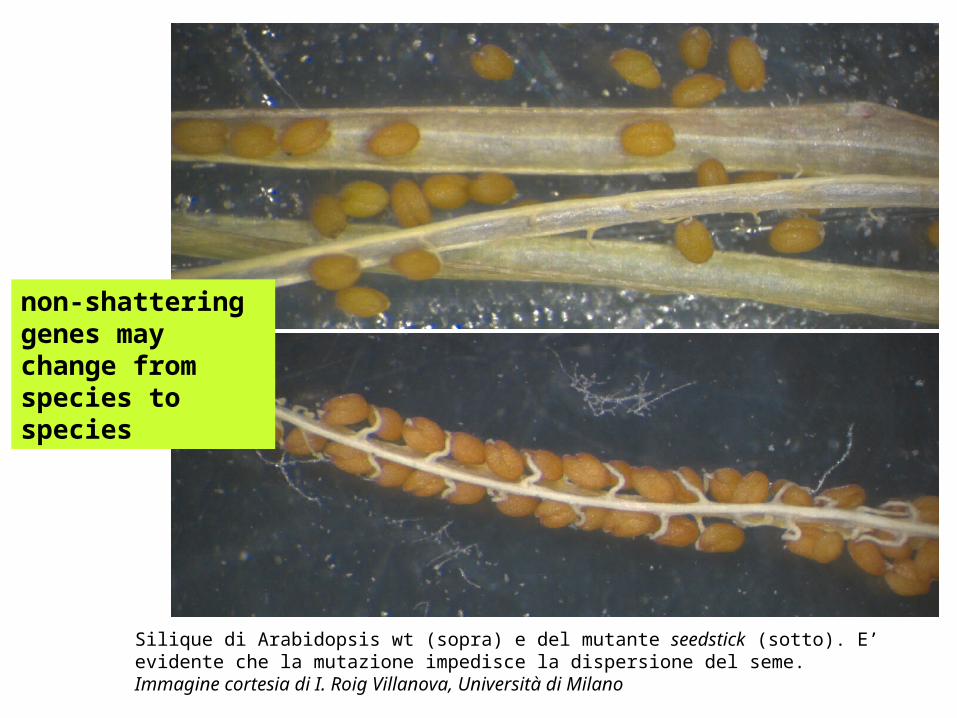
Silique di Arabidopsis wt (sopra) e del mutante seedstick (sotto). E’ evidente che la mutazione impedisce la dispersione del seme. Immagine cortesia di I. Roig Villanova, Università di Milano
non-shattering genes may change from species to species

La coltivazione del riso: il problema del crodo
Il riso crodo è molto simile al riso coltivato (stessa specie).
Difficile da distinguere quando è allo stadio di germinello.
A maturità, il riso crodo è più alto.
Infesta 40-75 % dell’area coltivata a riso in Europa.

riso crodo (red rice)

Riso crodoRiso coltivato
* Un grosso problema nelle zone a semina diretta* Riso crodo e coltivato sensibili agli stessi erbicidi

Semi dispersi prima del raccolto
Dormienti per 2-5 a.
11 piante m-2 40% perdita di resa1
2 3
23
1
Binasco
(Mi)Foto prese a Binasco (Ott. 2011) nei tre campi contrassegnati sulla mappa
Livello di infestazione con riso crodo:
Bassa Media Alta

Riso crodo
Riso coltivato
Esempio di rischi: riso crodo e riso HT
Cecilia
In breve: i pericoli per l’ambiente sono essenzialmente di tipo agronomico (il danno avviene nei campi, non nelle zone selvatiche)
FOOD SECURITY !!!
HT rice (transgenic or not) gives excellent control of red rice for 2-3 yrs, BUTthen resistance gene is introgressed into weedy rice technology is lost

Campo n.3 nel 2012 (ad alta infestazione di crodo nel 2011)
Neanche più una pianta di crodo… Motivo? varietà Clearfield!(tollerante agli imidazolinoni, che inibiscono l’acetolattato sintasi)

Svetta sopra la coltura, ma è la stessa specie.
Probabilmente è una forma de-domesticata che disperde i semi e cresce più alta
Girasole coltivato
girasole selvatico
Un caso analogo al riso crodo
Foto prese a sud di Siena (Aug. 2011)

BibliografiaShattering
Konishi et al. (2006) An SNP Caused Loss of Seed Shattering During Rice Domestication. Science 312, 1392-1396.
Li et al (2006) Rice Domestication by Reducing Shattering. Science 311, 1936-1939.
Crodo
Gressel e Valverde (2009) A strategy to provide long-term control of weedy rice while mitigating herbicide resistance transgene flow, and its potential use for other crops with related weeds. Pest Manag Sci 65:723–731





























