Embed Size (px)
DESCRIPTION
Biotehnologii in industria alimentara
Citation preview

BIOTEHNOLOGII IN INDUSTRIA
ALIMENTARA
SUPORT CURSANUL III TPPA +CEPA

BIOTEHNOLOGII CLASICE SI MODERNE
Progresele considerabile înregistrate în ultimul secol în domeniul biotehnologiei , având la baza o serie de descoperiri ştinţiifice, dintre care amintim – descoperirea ADN-ului, în 1953 şi stabilirea tehnicilor de manipulare genetică în 1973, au pus bazele unei biotehnologii moderne cu considerabile efecte economice şi sociale .
Realizările din ultimele decenii în domeniul ingineriei genetice, au determinat pe oamenii de ştiinţă să anticipeze că secolul XXI va fi dominat de această activitate, aşa cum secolul XX a fost dominat de descoperirile fizicii: tehnica informatică, fisiunea nucleară, explorarea cosmosului .
Cele mai cunoscute descoperiri ale ingineriei genetice din secolul XX sunt: interferonul (o proteină sintetizată de celula umană ca răspuns la atacul unor virusuri şi care astăzi se foloseşte pentru tratarea cancerului hepatic), insulina produsă de bacterii, hormoni de creştere umani sau diferite vaccinuri .
Cele mai multe cercetări, cu cele mai multe rezultate au fost realizate în domeniul medicinei şi industriei farmaceutice: producţia de hormoni, de antigene, de enzime, de reactivi necesari diagnosticării.
In domeniul industriei, biotehnologiile asigură valorificarea deşeurilor industriale pentru obţinerea de materiale plastice, combustibili sau substanţe chimice .
Microorganismele sunt utilizate să transforme biomasa necomestibilă în hrană şi energie A apărut aşadar o ramură a ingineriei genetice, biotehnologia modernă, care foloseşste ca maşini şi aparate de producţie microorganismele sau diferiţi constituenţi celulari ai microorganismelor .
Dezvoltarea biotehnologiilor va determina schimbări fundamentale în domeniul agroalimentar. Astfel prin tehnologia manipularii materialului genetic, ADN recombinat, se anticipează o cale de realizare de recolte mai bogate prin transferul genelor fixatoare de azot la plantele de interes major din punct de vedere al alimentaţtiei .
Au fost realizate astfel materii prime vegetale cu conţinut nutritiv mai ridicat, mai rezistente la boli şi atacul dăunătorilor, secetă, frig.
Este posibil, spun specialiştii, ca peste 100 de ani, configuraţia plantelor de cultură şi configuraţia animalelor să fie foarte diferite faţă de cele din zilele noastre.
Industria alimentară este poate cel mai vechi utilizator al biotehnologiilor, fermentaţia ca proces biotehnologic fiind cunoscută de milenii. In antichitate, drojdiile erau folosite pentru fabricarea pâinii, vinului , berii. In urmă cu 5000 de ani sumerienii fabricau 20 de tipuri de bere.
Biotehnologiile moderne au un domeniu vast de exploatare în industria modernă.A crescut producţia mondială de enzime exogene, utilizate în aproape toate ramurile industriale, sinteza de aminoacizi esenţiali cu ajutorul bacteriilor, obţinerea de alcool folosit ca substituent al petrolului sau producerea de biomasaă prin valorificarea pe calea fermentaţiei a unor plante cu creştere rapidă.
Beneficiile biotehnologiilor moderne au impus dezvoltarea lor foarte rapidă la nivel mondial .1.Creşterea şi diversificarea producţiei vegetale Primele generaţii de produse alimentare modificate genetic (OMG sau GMO) au fost bine primite de
fermieri în SUA, prin creşterea producţiei agricole. Spre exemplu, studiile au arătat că, o specie de porumb modificată genetic, rezistent la acţiunea dăunătoare a insectelor, a manifestat o rezistenţă crescută şi la la acţiunea altor microorganisme: fungi, mucegaiuri care atacau porumbul nemodificat.
De asemenea, nivelul de micotoxine produse de diferite specii de mucegaiuri sunt mult mai mici la porumbul-GMO.
2.Beneficii nutritive şi igienice O mare varietate de produse biotehnologice din clasa uleiurilor sau grăsimilor alimentare sunt deja pe
piaţă.Cu ajutorul biotehnologiilor vegetale a fost redus conţinutul de acizi graşi saturaţi din câteva sortimente de uleiuri vegetale .
Prin biotehnologie a fost rezolvată o problemă de igienă alimentară, atunci când uleiurile vegetale erau hidrogenate pentru creştera stabilităţii la tratament termic sau pentru obţinerea margarinei.
Cercetarile biotehnologice au vizat de asemenea creşterea conţinutului proteic al cartofului. Aceşti cartofi transgenici conţin un număr important de aminoacizi, care în mod normal nu se găsesc în cartof. Pentru orz au fot create specii bogate în vitaminaA sau cu un conţinut mai bogat de fier.
2

A fost creată o roşie modificată genetic care conţine de trei ori mai mult antioxidant-licopen.Consumul de licopen este asociat cu scăderea riscului producerii cancerului şi scăderea nivelului colesterolului din sânge .
Din punct de vedere al igienei alimentare, biotehnologia încearcă să resolve şi următoarele probleme: Identificarea proteinelor alergenice din lapte, soia, alune şi eliminarea lor din producţiile
vegetale viitoare Descreşterea coţinutului natural de substanţe toxice, prezente în produsele alimentare
3. Creşterea calităţii produselor Biotehnologiile sunt de asemenea utilizate pentru modificarea caracteristicilor materiilor prime pentru
a fi mai atractive pentru consumator şi mai uşor de procesat. Cercetarile efectuate au determinat: creşterea conservabilităţii fructelor şi vegetalelor proaspete ardeii, morcovii şi salata sunt mai crocante obţinerea de soiuri cu sâmburi puţini pentru pepeni şi struguri creşterea sezonalităţii soiurilor de tomate, căpşuni creşterea aromei roşiilor, ardeilor, perelor crearea de specii de ceai şi cafea fără cofeină
Cercetătorii japonezi au identificat enzima care produce substanţa care declanşează lăcrimarea atunci când tăiem ceapa. Este numai o problemă de timp până când va fi creată o specie de ceapă fără această enzimă.
Foarte multe cercetări au fost efectuate pentru schimbarea raportului apă-amidon din diferite leguminoase. Astfel, un cartof cu un conţinut ridicat de amidon, va fi mai sănătos pentru om, deoarece el va absorbi mai puţin ulei în timpul prăjirii. De asemenea, obţinerea amidonului din cartof se va face cu un consum energetic mult mai redus. Cu aceleaşi scopuri, este în cercetare şi o specie de tomate pentru obţinerea pastei de tomate. Creşterea cu 1.2% a substanţei uscate din cartofi, ar aduce o economie energetică de 35 milioane de $ procesatorilor americani .
De beneficiile biotehnologiilor beneficiază şi industria de lactate. Specialiştii din Noua Zeelandă utilizează în present biotehnologiile animale pentru creşterea conţinutului de cazeină din lapte cu 13% , pentru obţinerea unei cantităţi mărite de brânzeturi.
ORGANISMELE MODIFICATE GENETIC
1. Organisme modificate genetic Având în vedere reacţia consumatorilor privind introducerea pe piaţă a unor plante, dar şi la adresa
alimentelor şi furajelor modificate genetic, este necesară cunoaşterea noţiunii de OMG , aşa cum este cunoscută pe plan naţional şi internaţional .
In Germania organismele modificate genetic(OMG) sunt organisme al căror material genetic a fost modificat într-un mod care nu există în natură în condiţii naturale sau de recobinare naturală. OMG trebuie să fie o unitate capabilă de autoreplicare sau transmitere a materialului genetic .
In SUA, OMG se referă la plante şi animale care conţin gene transferate de la alte specii, pentru a obţine anumite caracter , precum rezistenţa la anumite pesticide şi ierbicide.
In România, conform ordonanţei de guvern OG nr.49/2000, OMG este un organism care conţine o combinaţie nouă de material genetic, obţinut prin tehnicile biotehnologiilor moderne care îi conferă noi caracteristici.
2. Ingineria geneticăIngineria genetică poate fi definită ca un ansamblu de metode şi tehnici care permit introducerea în
patrimonial genetic al unei celule a uneia sau a mai multor gene noi, numite de interes, fie modificarea unor gene prezente deja în celulă.
Genele trasferate sunt denumite transgene. Ingineria genetică mai este denumită şi modificare genetică, transformare genetică sau transgeneză, iar produsele produsele obţinute poartă numele de organisme modificate genetic sau organisme transgenice.
3

Pentru modificarea genetică a plantelor este nevoie de : gene de interes metoda de integrare a transgenelor în nucleul celulei care va fi originea noii plante selectarea plantelor la care transgena se exprimă la un nivel ridicat , adecvat
scopului(rezistenţă la ierbicid, culoare, aromă)Comparativ cu metoda clasică de ameliorare, transformarea prin inginerie genetică are cel puţin două
avantaje: permite introducerea unui singur caracter la o varietate gena transferată poate proveni din orice sursă, ceea ce extinde în mod nelimitat posibilităţile
de exploatare Cercetările de inginerie genetică necesită laboratore scumpe, echipamente, reactivi speciali şi
specialişti vizionari. In aceste condiţii este evident că cercetările de inginerie genetică s-au dezvoltat prioritar în ţările cu mai multe resurse financiare şi umane.
3.Posibilele reacţii adverse ale organismelor modificate genetic OMG sunt subiectul unor dezbateri aprinse, pe de o parte din motive etice, iar pe de altă parte
din motive legate de riscul utilizării şi consumării lor. Ca urmare a presiunii consumatorilor, consiliul Comunităţii Europene a emis directive care se referă la obţinerea, utilizarea, eliberarea deliberată în mediu şi comercializarea OMG-urilor şi a alimentelor obţinute din materii prime modificate genetic.
In directive nr.18/2001/CEE sunt menţionate potenţialele efecte adverse ale eliberării OMG-urilor în mediu:
apariţia unor boli umane prin efecte toxice şi alergice apariţia unor boli la plante şi animale prin efecte toxice şi alergice efecte negative asupra dinamicii şi diversităţii genetice a populaţiilor de specii din
mediu diminuarea rezistenţei la patogeni compromiterarea tratamentelor profilactice sau terapeutice vegetale, veterinare sau
umane, prin transferul de gene care conferă rezistenţă la antibioticele utilizate în medicina umană şi veterinară
efecte asupra ciclului biogeochimic(ciclul compuşilor în natură, în special ciclul C şi N)
Această directivă stabileşte că introducerea alimentelor obţinute din OMG-uri să se realizeze prin metoda pas cu pas. Etichetarea, care este obligatorie pentru toate produsele care conţin OMG , asigură consumatorii că pot alege un produs modificat genetic sau unul tradiţionale.
4

ENZIME UTILIZATE IN PROCESELE BIOTEHNOLOGICE DIN INDUSTRIA ALIMENTARA
1. GENERALITĂŢI ASUPRA ENZIMELOR
Enzimele definite drept componente de natură proteică, produse de celulele vii care catalizează reacţii de sinteză şi degradare din organismele animale, vegetale şi microorganisme, prezintă numeroase implicaţii şi aplicaţii în industria alimentară. Fiind componente ale materiilor prime vegetale şi animale utilizate în industria alimentară, a căror activitate nu încetează odată cu recoltarea, depozitarea, conservarea şi prelucrarea tehnologică a acestora, enzimele pot manifesta acţiuni favorabile, dorite, care conduc la îmbunătăţirea calităţilor naturale constituţionale, gustative etc. sau nefavorabile şi nedorite, determinînd degradarea şi pierderea valorilor nutritive. Dacă în acest sens se are în vedere activitatea enzimelor proprii materiilor prime vegetale şi animale, enzimele ca atare sub formă de preparate enzimatice, obţinute din diferite surse bogate în enzime, îşi găsesc multiple aplicaţii ca adaosuri, fiind folosite încă din cele mai vechi timpuri ca de exemplu cheagul la prepararea brînzeturilor, „Koji" (kabi-taki = „floare de mucegai", preparat enzimatic obţinut prin cultivarea lui Aspergillus oryzae pe un decoct de orez sau alte cereale), la prepararea unor produse tradiţionale-fermentate, larg răspîndite în Asia de Sud-Est.
In ultimele decenii, preparatele enzimatice şi-au găsit numeroase utilizări în diferite sectoare ale industriei alimentare. Această creştere spectaculoasă a gradului de utilizare a preparatelor enzimatice în industria alimentară (dar şi nealimentară) este explicată prin eficienţa şi precizia, versatilitatea şi economicitatea cu care acţionează aceste preparate enzimatice.
Se postulează existenţa în materia vie a circa 10 000 de enzime diferite, dintre care circa 2 000 au fost izolate, în stare mai puţin sau mai mult purificată şi a căror intervenţie şi acţiune în diferitele procese metabolice este mai mult sau mai puţin cunoscută. Catalizînd reacţiile biochimice din organismele vegetale şi animale, enzimele condiţionează desfăşurarea, coordonarea şi autoreglarea acestor reacţii prin care se realizează procesele metabolice (anabolice şi catabolice) ale creşterii, dezvoltării, reproducerii şi tuturor activităţilor celulare.
In industria alimentară, prelucrătoare de materii prime vegetale şi animale, practic nu există procese tehnologice în care să nu fie implicate enzimele endogene, proprii materiilor biologice folosite sau ale microorganismelor utilizate în prelucrarea acestora; în plus, în multe cazuri, în prezent, se recurge la suplimentarea acestor activităţi enzimatice cu enzime exogene, cu preparate enzimatice obţinute din diverse surse bogate în enzime, ţesuturi vegetale, animale şi mai ales în ultimii ani din culturile diverselor microorganisme.
Indiferent că se au în vedere enzimele endogene, proprii materiilor prime sau cele adăugate acestora, ele se caracterizează prin următoarele proprietăţi generale:— sunt cei mai eficienţi catalizatori cunoscuţi astăzi; în concentraţii extrem de mici ( 10 -6 M sau chiar şi 10-9 M) determină realizarea reacţiilor cu viteze extrem de mari;— reacţiile catalizate enzimatic se produc în condiţii compatibile cu viaţa, la temperatură şi presiune obişnuită, în mediu slab acid, neutru sau slab alcalin ;
— manifestă specificitate de acţiune (determină producerea unui anumit tip de reacţii, de exemplu de oxidorcducere, de hidroliză, de sinteză etc.) şi de substrat (recunoaşte numai un anumit reactant, sau un grup limitat de reactanţi);— asigură coordonarea, reglarea şi controlul proceselor biochimice la care participă şi care stau la baza metabolismului celular.

1.1. Constituţia enzimelor
Determinantă pentru funcţia catalitică a enzimelor este configuraţia lor, respectiv structura şi organizarea spaţială a moleculei proteice care manifestă activitatea enzimatică.
Din punct de vedere structural, enzimele se împart în două categorii:— enzime de natură exclusiv proteică, constituite în întregime din proteine (de exemplu: pepsina, tripsina, papaina etc.) ;— enzime de natură heteroproteică formate dintr-o parte proteică (apo-enzimă) şi una neproteică
denumită cofactor enzimatic. Cele două părţi separate sînt inactive catalitic; împreună formează complexul molecular apo-enzimă:cofactor, respectiv holoenzima care manifestă activitate catalitică.În structura holoenzimei, cofactorul imprimă specificitate de acţiune, respectiv tipul şi viteza de
reacţie catalitică, în timp ce apoenzima imprimă specificitatea de substrat, deci determină substanţa asupra căreia acţionează enzima.
Apoenzima, fiind de natură proteică, va manifesta proprietăţile generale ale proteinelor: este termolabilă şi nedializabilă; stabileşte legătura enzimei cu substratul; manifestă grade diferite de afinitate pentru cofactor; este susceptibilă de modificări conformaţionale în anumite limite.Cofactorul enzimatic reprezintă componente neproteice, de natură chimică foarte diferită, care
sunt indispensabile pentru manifestările activităţii catalitice a numeroase enzime.După natura chimică şi modul lor de legare la apoenzimă, cofactorii se clasifică: coenzimă; grupări prostetice; ioni metalici.Coenzimele sunt compuşi organici, derivaţi din vitamine, care se ataşează temporar la apoenzimă
şi care sunt uşor disociabili de acestea. Ele trec uşor de la o apoenzimă la alta, putând să participe la transformarea altor molecule de substrat după terminarea unei anumite reacţii. Din ei fac parte: NAD+, NADP+, FMN, FAD, ATP, CTP, acidul lipoic etc.
Grupările prostetice sunt substanţe organice fixate pe apoenzimă, care disociază greu, deoarece sunt legate prin legături covalente. Aceste grupări sunt: TPP, piridoxalfosfatul, hemul. Gruparea prostetică imprimă mecanismul unor procese enzimatice: transportul de electroni, de grupări –NH2, acetic etc.
Ionii metalici sunt indispensabili pentru exercitarea funcţiei catalitice a unor enzime, participând în calitate de cofactori sau de componente structurale ale acestora. Ele se mai numesc metal-enzime, iar printre ionii care îndeplinesc rol de cofactor se pot menţiona: Mg2+, Mn2+, Cu2+, Zn2+, Fe2+, Fe3+, Mo2+. Unele enzime pot conţine chiar doi ioni metalici.
Exercitarea proprietăţilor catalitice ale enzimelor se realizează prin intermediul situsurilor catalitice — centrilor sau zonelor active — care reprezintă ansamblul grupărilor chimice din structura enzimei ce participă efectiv în manifestarea activităţii enzimatice (de exemplu, anumite resturi din aminoacizii constituenţi ai lanţurilor polipeptidice şi cofactorii enzimatici, care vin în contact direct cu substratul).
În funcţie de organizarea lor structurală se disting: enzime monomerice, constituite dintr-un lanţ polipeptidic care nu poate fi disociat în subunităţi fără pierderea activităţii lor catalitice şi enzime oligomerice care sunt agregate moleculare constituite din doi sau mai mulţi protomeri, respectiv lanţuri polipeptidice similare sau diferite, care prin disociere sau din contră prin asociere devin catalitic-active. Pentru o serie de enzime s-au stabilit existenta unor forme moleculare multiple sau izoenzime, care catalizează realizarea aceleiaşi reacţii biochimice, dar care diferă între ele prin configuraţii spaţiale diferite (compoziţie în aminoacizi constituenţi), prin proprietăţile lor fizico-chimice (pH sau temperatură optimă, cinetică de reacţie, comportare faţă de inhibitori sau activatori etc.). imunologice etc.
Se cunosc, de asemenea, aşa-numitele sisteme sau complexe multienzimatice constituite din enzime intim legate prin interacţiuni necovalente, care participă la realizarea unor secvenţe de reacţii biochimice consecutive şi înlănţuite, în care produsul de reacţie al primei enzime devine substrat pentru cea de a doua enzimă, al cărui produs de reacţie devine substrat pentru a treia enzimă etc., pînă la
6

realizarea întregului şir de reacţii prin care se metabolizează în celule un component biochimic. Astfel de sisteme sau complexe multi-enzimatice sunt, în general, localizate în celule la nivelul diferitelor formaţiuni sau organite subcelulare.
1.2. Cinetica reacţiilor şi factorii care influenţează activitatea enzimelorEnzimele catalizează producerea reacţiilor chimice, termodinamic posibile, fără schimbarea
echilibrului de reacţie, prin scăderea energiei de reacţie a reactanţilor şi prin creşterea vitezei de reacţie a reactanţilor (in vivo ele condiţionează desfăşurarea, coordonarea şi autoreglarea reacţiilor biochimice ale materiei vii, prin care se realizează procesele metabolice ale creşterii, dezvoltării, reproducerii şi tuturor activităţilor celulare).
Într-o reacţie catalizată enzimatic, enzimă (E) formează cu substratul (S) un complex intermediar de tranziţie (ES) foarte reactiv şi instabil, care se transformă rapid, conducînd la eliberarea produsului sau produşilor de reacţie (P) şi a enzimei ce poate relua reacţia, după schema generală:
E + S ES EP P + E
O serie de enzime catalizează reacţii în care pot interacţiona două sau chiar mai multe substraturi. Aşa, de exemplu, sunt transferazele care catalizează transferul unei grupări chimice funcţionale de pe un substrat (donor) pe alt substrat (acceptor), realizîndu-se reacţii de simplă sau dublă deplasare.
Mecanismul catalizei enzimatice, sub raportul interacţiunii dintre enzimă si substrat, al formării complexului reactiv enzimă — substrat şi transformarea acestuia în produşi de reacţie, se bazează pe diferite interpretări, explicîndu-se prin ipotezele „broască-cheie", potrivirii induse, catalizei covalente şi catalizei generale prin acizi şi baze.
Viteza reacţiilor catalizate enzimatic este dependentă de: concentraţia enzimei şi a substratului, afinitatea enzimei faţă de substrat, temperatură, pH, efectori (activatori sau inhibitori), potenţial redox, radiaţii etc.
7

In anumite limite, viteza reacţiilor catalizate enzimatic este direct proporţională cu concentraţia enzimei, pentru ca, la concentraţii crescute ale enzimei, să devină aproape constantă. Creşterea concentraţiei substratului determină iniţial o creştere rapidă a vitezei de reacţie, pentru ca ulterior viteza de reacţie să crească lent, iar la concentraţii mari de substrat să devină maximă, căpătând o valoare con-stantă (fig. 1). Expresia matematică care defineşte relaţia dintre viteza de reacţie catalizată enzimatic şi concentraţia substratului este dată de ecuaţia Michaelis-Menten : vmax*[S]
V = ----------------- KM+[S]
în care: v- este viteza de reacţie la un moment dat; vmax — viteza maximă de reacţie, corespunzătoare concentraţiei mari de substrat, cînd
enzima este saturată cu substrat ; [S] —concentraţia substratului ; kM — constanta Michaelis-Menten, moli/1.
Din punct de vedere grafic, ecuaţia Michaelis-Menten reprezintă o hiperbolă, indicînd o tranziţie de la o fază în care viteza de reacţie este dependentă de concentraţia substratului (fază în care reacţia este de ordinul 1) la o fază independentă de concentraţia substratului (fază în care reacţia este de ordinul zero), în care viteza de reacţie devine maximă, enzima fiind saturată cu substrat. La o viteză egală cu 1/2 din viteza maximă corespunde o valoare determinată a concentraţiei substratului denumită kM sau constanta Michaelis-Menten, care se exprimă în moli/1 şi care indică afinitatea enzimei faţă de substrat; cu cît kM are o valoare mai mică, cu atît afinitatea enzimei pentru substrat este mai mare şi invers, cu cît kM are o valoare mai mare cu atît afinitatea enzimei pentru substrat va fi mai mică.
În anumite limite (pînă la circa 60 ... 80°C), viteza reacţiilor enzimatice creşte o dată cu temperatura (temperatura la care viteza de reacţie este maximă corespunde temperaturii optime de acţiune a enzimei). La temperaturi mai mari decît temperatura optimă, viteza de reacţie scade rapid, avînd loc un proces de denaturare termică a enzimei (fig. 2). Temperatura optimă a enzimelor este influenţată şi condiţionată de structura chimică a enzimei, a substratului, şi de concentraţia acestora în sistemul de reacţie. In general, activitatea enzimelor scade la temperaturi peste 50 ... 60°C; totuşi există enzime care îşi păstrează încă în mare măsură activitatea şi la temperaturi de peste 60°C, ca de exemplu: papaina, o protează vegetală activă la 80°C; peroxidaza din hrean care rămîne activă şi după o încălzire la 100°C.
Activitatea enzimelor este dependentă de pH-ul mediului în care îşi exercită acţiunea. pH-ul la care activitatea este maximă este denumit pH optim al enzimei, iar pH-ul la care enzima îşi menţine activitatea în cele mai bune condiţiuni se numeşte pH optim de stabilitate. Cele două pH-uri optime ale
8

unei enzime, de activitate şi de stabilitate, pot avea valori apropiate, pot să coincidă sau pot să fie diferite (fig. 3).
Activitatea enzimelor este, de asemenea, influenţată de prezenţa unor efectori (diferite substanţe chimice) în mediul de exercitare a activităţii lor; activatorii influenţează pozitiv, iar inhibitorii negativ activitatea enzimelor. In funcţie de natura şi modul de acţiune al inhibitorilor, inhibiţia enzimatică este de mai multe tipuri: ireversibilă, reversibilă (competitivă şi necompetitivă), şi alosterică.
1.3. Clasificarea enzimelorRecenta clasificare şi nomenclatură a enzimelor se bazează pe principiile şi regulile stabilite şi
publicate în anul 1964, revizuite şi republicate în 1973 de Comisia de Enzime a Uniunii Internaţionale de Biochimie (I.U.B.) şi a Uniunii Internaţionale de Chimie Pură şi Aplicată (I.U.P.A.C.). In acest sens, enzimele au fost clasificate în 6 clase (numeroase subclase şi sub-subclase) şi anume:— oxidoreductaze — catalizează reacţiile de oxidoreducere prin transfer de hidrogen sau electroni, sau prin combinarea unui substrat cu oxigenul molecular ;— transferaze — catalizează transferul diferitelor grupări chimice de la un substrat donator la un alt substrat acceptor;— hidrolaze— catalizează scindarea hidrolitică a diferitelor substraturi, prin adiţia apei la nivelul diferitelor grupări chimice;— liaze — catalizează adiţia sau îndepărtarea unor grupări chimice din substraturi, prin mecanisme diferite faţă de hidroliză;— izomeraze—catalizează rearanjări intramoleculare ;— ligaze sau sintetaze — catalizează sinteza unor noi legături prin unirea a doi compuşi într-unul singur, folosind ca sursă energetică nucleozidtrifosfaţii.
Clasele de enzime se împart în subclase şi sub-subclase, în funcţie de o serie de detalii privind grupările supuse transformării şi natura cofactorilor implicaţi în reacţia catalizată enzimatic.Pentru nomenclatura enzimelor se folosesc nume uzuale sau tradiţionale, care sînt de obicei formate din numele substratului asupra căruia acţionează enzima, urmat de terminaţia -ază şi nume sistemice (recomandate de Comisia de Enzime) formate din numele substratului sau substraturilor şi tipul de reacţie catalizat, urmat de terminaţia ază, însoţite de un cod (alcătuit din patru cifre care reprezintă clasa, subclasa, sub-subclasa şi numărul de ordine), precedat de EC (Enzyme Commission). Cîteva exemple, privind numele unor enzime importante pentru industria alimentară:
E.C. 3.2.1.2 α-1-4-Glucan maltohidrolaza ( β-amilaza)E.C. 3.1.1.3. Glicerol ester hidrolaza (lipaza)
9

1.4. Unităţi de măsură ale activităţii enzimelorIn principiu, determinarea activităţii enzimelor se efectuează prin : măsurarea gradului de
transformare a substratului, măsurarea concentraţiei produsului de reacţie sau măsurarea cineticii de reacţie, urmărite într-un anumit interval de timp prin metode fizice sau chimice adecvate. Activitatea enzimelor se exprimă cantitativ în unităţile propuse de Comisia de Enzime (CE) şi anume: — Unitatea de activitate enzimatică (U) reprezintă cantitatea de enzima care catalizează transformarea a l mol substrat/min în condiţii standard (25°C, pH şi concentraţie de substrat optime). Această unitate de măsură recomandată de CE în 1961 se foloseşte încă în prezent; CE recomandă renunţarea sau abandonarea progresivă a folosirii unităţii U şi trecerea la Kat.— Katalul (Kat) reprezintă cantitatea de enzimă care catalizează transferarea a l mol substrat/s în condiţii standard. (Prin definiţie această unitate de măsură se apropie mai mult de dimensiunile constantelor de viteză folosite în cinetica chimică respectiv mol/s.) Se foloseşte şi multiplul kilokatal (K Kat) şi respectiv submultiplii milikatul (mKat), microkatul (Kat), nanokatul (nKat) şi picokatul (pKat).— Activitatea specifică — reprezintă numărul de unităţi enzimatice/mg proteină (respectiv Kat/kg proteină şi Kat/kg proteină).— Activitatea enzimatică molară (număr de transfer = turnover number) — reprezintă numărul de molecule de substrat transformate de către o moleculă de enzimă în timp de l min sau l s (Kat/mol enzimă).Cu toate aceste recomandări ale Comisiei de Enzime, în lucrări mai vechi sau chiar şi în prezent se folosesc şi alte moduri, arbitrare, de exprimare a activităţii enzimelor, care de obicei sînt definite de cei ce le utilizează.
2. PREPARATE ENZIMATICE ŞI ENZIME IMOBILIZATE
Preparatele enzimatice şi de enzime imobilizate folosite în realizarea diferitelor procese biotehnologice din industria alimentară sunt considerate ca adjuvanţi de transformare. In anul 1982, Comitetul mixt FAO/OMS de experţi în aditivi alimentari a stabilit o serie de „norme generale pentru preparatele enzimatice utilizate în prepararea alimentelor". Conform acestui comitet, preparatele enzimatice, folosite ca aditivi în industria alimentară, sunt obţinute din materii prime de origine animală, vegetală sau microbiană, fiind constituite din celule întregi, din părţi de celule sau extracte complet lipsite de celule. Pot conţine una sau mai multe componente enzimatice active, suporturi, solvenţi, agenţi de conservare, antioxidanţi şi alte substanţe necesare şi conforme unei bune practici de fabricare. Ele se pot prezenta sub formă lichidă, semilichidă, uscate sau imobilizate, avînd o culoare care poate să varieze de la qvasi incolor la brun închis.
In ceea ce priveşte materiile prime din care sînt obţinute preparatele enzimatice, normele Comitetului mixt FAO/OMS prevăd ca:— ţesuturile de origine animală să răspundă normelor veterinare aplicate cărnii şi manipularea lor să satisfacă exigenţele unei bune practici igienice;— materialele de origine vegetală, folosite ca surse de enzime sau ca ingrediente în prepararea mediilor de cultură pentru microorganismele producătoare de enzime, trebuie să nu elibereze nici un reziduu nociv pentru sănătate, în condiţii normale de utilizare;— preparatele enzimatice de origine microbiană trebuie produse prin folosirea controlată fără penetrare de microorganisme susceptibile de a conduce la apariţia de substanţe toxice sau alte produse nedorite.
În cazul preparatelor de enzime imobilizate, în care insolubilizarea enzimei se realizează prin procedee fizice şi/sau chimice, suportul şi în special agentul de imobilizare folosit trebuie să fie inert sau admis de a fi utilizat în produsele alimentare, iar orice eliberare de enzimă de pe suport, şi mai ales eliberarea de agent de imobilizare, trebuie să rămînă în limitele acceptabile care vor fi precizate pentru fiecare preparat enzimatic imobilizat.
Aditivii (inclusiv adjuvanţii de transformare) şi ingredientele care intervin în producerea preparatelor enzimatice trebuie să fie substanţe acceptate pentru a fi utilizate în produsele alimentare ca: apa, substanţe insolubile care pot fi îndepărtate o dată ce s-a produs procesul de transformare.
Limitele şi metodele de determinare a componentelor contaminante ale preparatelor enzimatice folosite în industria alimentară sînt arătate în tabelul 2.
10

Limite şi metode de determinare pentru componentele contaminante,recomandate de Comitetul mixt FAO/OMS (Etude FAO: Alimentations et Nutrion, 19/1982)
Componente contaminante ale
preparatelor enzimatice
Limite admisibile Metodele de determinare recomandate sînt publicate în:
ArsenPlumb Metale grele
Maximum 3 mg/kg Maximum 10 mg/kg Maximum 40 mg/kg (exprimate ca plumb)
Me"thodes Generales (Directives Generales pour l'usage des normes IECFA d'identite et de purete)
Număr total de germeni viabili
Coliformi
Maximum 5. 1O4 /g
Maximum 30/g
Microbiologic — Directives generales pour le denombrement de microorganisme Directives generales pour le denombrement des coliformes Methode par le comptage de colonies obtenues à 30°C. ISO. Norme internaţionale Ref. nr. ISO 4833, Premier Edition (1978-02-01)
Escherichia coli
Salmonclla
Absent pe o probă de 25 g
Absent pe o probă de 25 g
FDA — Bacteriogical Analvtical Manual - Fifth Edition ( 1978) Cap. X. Enteropathogenic Escherichia coli ; Cap. XII. Isolaion and identification of Salmonella
Activitatea antibiotică de origine bacteriană
Absentă Etude FAO: Alimentation et Nutrition, 19/1982; Appendice A
2.1. Preparate enzimaticePreparatele enzimatice folosite în industria alimentară trebuie produse în condiţii similare unei
bune practici de fabricare a produselor alimentare, iar prin utilizarea lor să nu se ajungă la o creştere a numărului total de germeni (NTG) şi la creşterea conţinutului în săruri peste limitele admise pentru un anumit produs alimentar luat în considerare.
Pentru obţinerea preparatelor enzimatice sunt folosite de obicei surse bogate în enzimele dorite, care sunt ieftine, uşor accesibile şi care se prelucrează uşor. Aceste surse pot fi de origine vegetală, animală sau microbiană. In plante, concentraţii mari de enzime se pot întâlni în seminţe, cereale germinate sau negerminate, rădăcini, sevă şi latexuri, frunze şi în unele cazuri chiar în coajă.La animale, concentraţii mari de enzime se găsesc în special în ficat, pancreas, mucoasa stomacală sau intestinală, inimă, rinichi, creier etc. In scopuri practice sunt folosite mai ales pancreasul, mucoasa stomacală şi intestinală.
O sursă foarte importantă de enzime, pentru producerea preparatelor enzimatice, o constituie diferitele microorganisme ca: bacteriile, drojdiile şi microciupercile (mucegaiurile). Faţă de sursele de enzime de origine vegetală sau animală, culturile diferitelor microorganisme prezintă o serie de avantaje care explică în mare măsură tendinţa manifestată în ultimele 2—3 decenii de a fi folosite din ce în ce mai mult pentru obţinerea de preparate enzimatice. Microorganismele pot fi obţinute în cantităţi mari, prin înmulţire în instalaţii speciale, pe medii de cultură ieftine (de obicei subproduse ale industriei alimentare ca: tărîţe de grîu, extract de porumb obţinut prin concentrarea apelor de înmuiere de la fabricarea amidonului, melasă, şroturi de soia şi de floarea-soarelui etc.).
Ciclul de creştere şi dezvoltare al microorganismelor este foarte scurt faţă de cel al plantelor şi animalelor, iar obţinerea microorganismelor în cantităţi mari nu necesită angajarea de terenuri cultivabile, cum este cazul la materiile prime vegetale. In plus, microorganismele prezintă şi avantajul că producţia lor de enzime poate fi mult mărită prin selectarea şi utilizarea de tulpini şi mutante înalt productive precum şi prin stabilirea condiţiilor fizice şi chimice optime (medii de cultură şi condiţii optime de cultivare) pentru producerea de enzime.
In cazul utilizării microorganismelor ca surse de enzime pentru industria alimentară, selectarea acestora se va face luîndu-se în considerare o serie de criterii cum sînt următoarele: să nu manifeste
11

putere patogenă şi să nu producă toxine (endo-, exotoxine sau micotoxine), să nu manifeste activitate antibiotică sau potenţial alergen şi să producă cu precădere şi în cantităţi mari enzima sau complexul enzimatic dorit, pe medii de cultură ieftine şi în condiţii avantajoase.
Indiferent de sursa de materii prime, prelucrarea lor pentru obţinerea de preparate enzimatice de uz alimentar este în linii generale aceeaşi (fig. 4).
Enzimele obţinute ca preparate brute sau parţial purificate, sub formă lichidă, semilichidă sau uscată sunt utilizate în industria alimentară ca atare, fiind adăugate şi acţionînd în mediile pe care urmează să le transforme ca enzime „libere", respectiv solubilizate în medii apoase şi fără a mai putea fi ulterior recuperate. Activitatea lor, după ce au realizat transformările dorite, este de obicei oprită prin diferite tratamente, mai ales pe cale termică, prin acidulare sau alcalinizare. In unele cazuri ele mai rămîn active şi în produsul finit.
2.2. Preparate enzimatice imobilizate Principiul general al procedeelor de imobilizare a enzimelor constă în legarea sau fixarea unei
enzime sau a unui sistem multienzimatic de suportul insolubil în apă, în condiţiile păstrării proprietăţilor catalitice, respectiv specificitatea de acţiune şi posibilitatea de a acţiona la pH şi temperatură asemănătoare enzimelor libere (neimobilizate).
Suporturile sau matricile utilizate în imobilizarea enzimelor pot fi de natură anorganică: silicea coloidală, caolinita, particule sau perle de sticlă cu grad de porozitate controlat, oxizi metalici (alumina, oxid de zirconiu, oxid de titan, cărbune etc.), şi de natură organică: celuloza şi derivaţii acesteia, agaroză, amidon, dextran, colagen, polimeri obţinuţi prin polimerizarea unor monomeri de tipul acrilamidă, acid metacrilic ş.a., răşini formaldehidice etc.
In funcţie de natura legăturilor care se stabilesc între enzime şi suportul de imobilizare, procedeul sau metodele de imobilizare pot fi fizice sau chimice.
12

Procedeele fizice se bazează pe imobilizarea enzimelor prin intermediul legăturilor fizice ca de exemplu, interacţiuni electrostatice, formarea de legaturi ionice, formarea de legături de hidrogen, interacţiuni proteină-proteină etc., diferenţiindu-se în acest sens (fig. 5.):— adsorbţia pe suporturi insolubile în apă (cărbune, clei, răşini schimbătoare de ioni, celuloză, sticlă etc.) ;— includerea în structuri macromoleculare (această incluziune se realizează prin polimerizarea unor materiale ca poliacrilamidă în silicagel, amidon, în prezenţa moleculelor de enzimă, astfel încît se formează o matrice de polimer în care sînt incluse moleculele de enzimă şi în care atît substratul cît şi produsul poate să difuzeze) ;— microîncapsulare în membrane semipermeabile ;— imobilizarea în celule de ultrafiltrare.
Procedeele chimice de imobilizare a enzimelor se referă la cele care conduc la formarea de legături covalente sau parţial covalente, între grupările funcţionale, care însă nu sînt esenţiale pentru manifestarea activităţii catalitice (nu sînt implicate în situsul catalitic al enzimei), şi un suport activat chimic, insolubil în apă. In cadrul acestor procedee se pot evidenţia (fig. 6):— legarea covalentă a enzimelor de suporturi insolubile, care posedă grupări reactive sau care pot fi activate prin diferite reacţii chimice;— copolimerizarea enzimelor cu un monomer reactiv şi legarea încrucişată (cross-linking) sau reticulară intra- şi intermoleculară a enzimelor legate de un suport cu un reactiv multifuncţional. La imobilizarea enzimelor trebuie să se aibă în vedere următoarele: conformaţia spaţială a moleculelor de enzimă în sistemul imobilizat este diferită de cea a mediului natural din care s-a extras enzimă. Pe de altă parte, structura tridimensională a moleculei de enzimă este distorsionată de legăturile sale cu suportul; micromediul moleculei de enzimă imobilizat este diferit de cel al enzimei aflate în soluţie, afectîndu-se viteza de difuzie a substratului şi a produşilor de reacţie. Inhibiţia de substrat şi de produs poate, de asemenea juca un rol important; procesul de transport al substanţelor este un proces pasiv, în comparaţie cu situaţia enzimei din celula vie, iar aceasta va avea desigur influenţă şi asupra vitezei de reacţie.
Imobilizarea enzimelor pe un suport anorganic sau organic provoacă schimbări în comportamentul acestora şi în cinetica reacţiilor: se măreşte stabilitatea enzimei (stabilitatea enzimei atît în stare statică cît şi în stare dinamică fiind influenţată de procedeul de imobilizare, măsura acestei stabilităţi fiind 1/2 din durata de viaţă a enzimei); se schimbă afinitatea enzimei faţă de substrat, aceasta fiind influenţată de durata lor de contact care, la rîndul ei, este determinată de viteza fluxului sau viteza de agitare a substratului în contact cu sistemul suport-enzimă, de viteza de difuzie a substratului la enzimă imobilizată; se modifică caracterul catalizei prin trecere de la cataliza omogenă la cea eterogenă.
13

La folosirea enzimelor imobilizate poate avea loc o variaţie a pH-ului optim iar randamentul în produsul de transformare este micşorat. De exemplu, prin folosirea amiloglucozidazei în soluţie, plecînd de la o suspensie de amidon cu 33% s.u., se ajunge la un randament de transformare în glucoza de 95,5— 96%, în timp ce la folosirea amiloglucozidazei imobilizate, randamentul este de 92—93%.
In orice caz, preparatele de enzime imobilizate, faţă de cele libere sau solubile, prezintă o serie de avantaje printre care se amintesc următoarele:
— refolosirea repetată a enzimei, cu aceeaşi cantitate de enzimă putându-se transforma cantităţi mai mari de substrat;— se poate lucra în sistem semicontinuu sau continuu, cu automatizarea procesului, ceea ce asigură un control precis al parametrilor de lucru ;— are loc o creştere a vitezei de lucru, prin controlul riguros al vitezei fluxului de substrat şi al concentraţiei acestuia;— se poate stopa reacţia enzimatică la momentul dorit şi se evită trecerea enzimei în produsul transformat ;— costurile globale de producţie sunt mai mici în comparaţie cu procedeele de folosire .a enzimelor libere ;— se pot utiliza şi enzime care nu sunt trecute pe lista GRAS (Generally Recognised as Safe).
Factorii critici ce trebuie luaţi în considerare la folosirea enzimelor imobilizate sînt următorii:— eficienţa economică a imobilizării determinată de costul enzimei, suportului şi metoda de imobilizare;— activitatea enzimei imobilizată care este în funcţie de tehnica de imobilizare, caracteristicile materialului de suport, viteza de difuzie a substratului la enzimă şi a produsului de transformare.— caracteristicile substratului ce trebuie transformat ;— stabilitatea enzimei care trebuie menţinută un timp cât mai îndelungat ;— contaminarea microbiologică a sistemului enzimă-suport în timpul utilizării reactorului respectiv.
Cu toate avantajele pe care le prezintă enzimele imobilizate, pînă în prezent, industria alimentară utilizează la nivel industrial numai glucozizomeraza imobilizată pentru izomerizarea glucozei în fructoză şi lactaza imobilizată pentru hidroliza lactozei din lapte sau zer. Sunt efectuate însă o serie de cercetări la nivel de laborator şi chiar la nivel pilot, care permit să se întrevadă în viitor extinderea folosirii prepa-ratelor de enzime imobilizate şi în alte sectoare ale industriei alimentare; în acest sens, sunt însă necesare cercetări care să conducă, probabil, la modificarea sau adoptarea unor tehnologii „clasice" de obţinere a diferitelor produse alimentare, la utilizarea preparatelor de enzime imobilizate sau la stabilirea de noi tipuri de bioreactoare şi eventual noi moduri de folosire a enzimelor imobilizate în medii eterogene, consistente şi vîscoase de felul celor prelucrate într-o serie de sectoare ale industriei alimentare.
2.3 Enzime de fermentaţie
Cea mai importantă problemă care trebuie rezolvată la obţinerea de preparate enzimatice cu ajutorul microorganismelor este gasirea unui microorganism care să producă în cantitate mare enzima sau sistemul enzimatic dorit. In acest caz un interes practic îl reprezintă microorganismele heterotrofe.
Pentru ca un organism să poată ataca un substrat şi să-l metabolizeze, el trebuie să posede enzimele necesare care să catabolizeze reacţiile de metabolism în condiţiile de mediu proprii pentru dezvoltare.
Totalitatea enzimelor pe care un organism le posedă în permanenţă sau pe care poate să le elaboreze la nevoie formează echipamentul enzimatic potenţial al microorganismului. Echipamentul enzimatic este determinat la rândul lui de genetica microorganismului. Zestrea ereditară constituită din gene care sintetizeză enzimele microorganismelor variază de la o specie la alta.
Majoritatea enzimelor care alcătuiesc echipamentul enzimatic sunt enzime intracelulare. Aceasta înseamnă că enzimele sintetizate de gene, după formare, rămân în celulă, nefiind secretate în mediul înconjurător în care microorganismele se dezvoltă. Activitatea lor se desfăşoară în interiorul celulei. Unele microorganisme cum sunt drojdiile, bacteriile lactice, nu conţin decât enzime intracelulare. La astfel de microorganisme eliberarea enzimelor în mediul înconjurător are loc după moartea celulelor, în
14

urma proceselor de autoliză. Din acestă cauză, astfel de microorganisme nu metabolizează decât substraturi nutritive solubile şi permeabile prin membranele lor celulare.
Alte microorganisme, aşa cum sunt bacteriile din genul Baccillus sau fungii din genul Aspergillus, elaborează pe lângă un mare număr de enzime intracelulare şi enzime extracelulare, pe care le secretă în mediul înconjurător. Enzimele extracelulare sunt, în general hidrolaze şi sunt secretate de microorganisme cu scopul de a degrada substraturile insolubile şi solubile cu molecule mari la compuşi solubili uşor asimilabili.
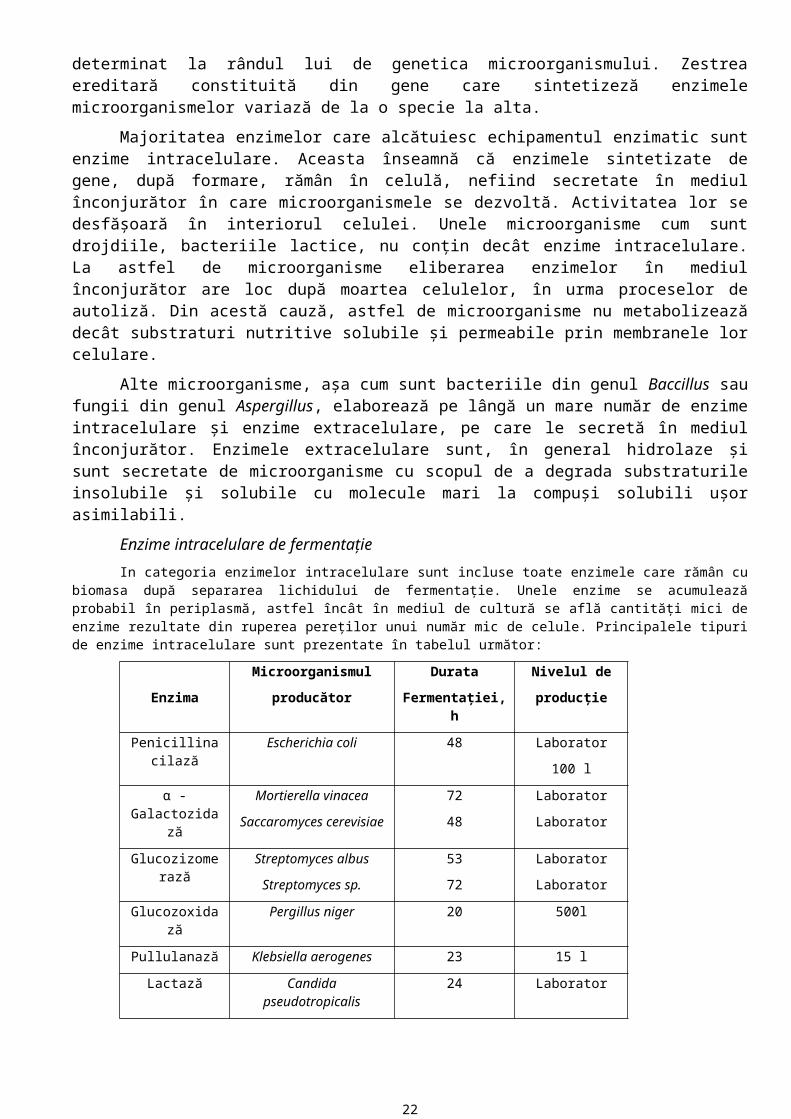
Enzime intracelulare de fermentaţie
In categoria enzimelor intracelulare sunt incluse toate enzimele care rămân cu biomasa după separarea lichidului de fermentaţie. Unele enzime se acumulează probabil în periplasmă, astfel încât în mediul de cultură se află cantităţi mici de enzime rezultate din ruperea pereţilor unui număr mic de celule. Principalele tipuri de enzime intracelulare sunt prezentate în tabelul următor:
Enzima
Microorganismul
producător
Durata
Fermentaţiei, h
Nivelul de
producţie
Penicillinacilază Escherichia coli 48 Laborator
100 l
α - Galactozidază Mortierella vinacea
Saccaromyces cerevisiae
72
48
Laborator
Laborator
Glucozizomerază Streptomyces albus
Streptomyces sp.
53
72
Laborator
Laborator
Glucozoxidază Pergillus niger 20 500l
Pullulanază Klebsiella aerogenes 23 15 l
Lactază Candida pseudotropicalis 24 Laborator
Majoritatea acestor enzime acţionează asupra unor substraturi cu masă moleculară mică. Aceasta se explică prin faptul că substraturile mici pot pătrunde în celule şi funcţionează astfel ca inductori ai enzimelor care le degradează. Spre deosebire de acestea, enzimele extracelulare acţionează asupra substraturilor cu masă moleculară mare.
Enzimele intracelulare se împart în două mari categorii: unele au o poziţie centrală în metabolismul organismului în creştere şi se produc în cantităţi mari in biomasă, altele au un rol secundar, minor şi se produc în cantităţi mici. Pentru utilizarea acestora din urmă în scopuri industriale este necesară imobilizarea lor, astfel încât să fie economică şi justificată folosirea lor.
In ceea ce priveşte activitatea enzimatică a acestor preparate, de cele mai multe ori constituie un secret de serviciu, deoarece poate face oricând obiectul unui brevet de invenţie. Pentru ca o enzimă să fie economică, ea trebuie să aibă o activitate enzimatică minimă( 100 UE/ml). De aceea majoritatea enzimelor intracelulare la care nu se atinge acestă valoare se imobilizează. Alteori, aceste enzime se utilizează pentru atacul unor impurităţi care incomodează procesul principal, dar fără atacul componentului principal. De exemplu, hidroliza enzimatică completă a amidonului presupune şi hidroliza legăturilor α – 1,6 glucozidice, aflate în număr mic în structura amilopectinei.
Pullulanazaeste folosită în acest caz pentru hidroliza legaturilor α – 1,6 facilitând în acest fel acţiunea amiloglucozidazei şi β- amilazei.
Glucozoxidaza este folosită pentru îndepărtarea urmelor de glucoză din praful de ouă folosit la obţinerea maionezei şi în acest caz nu trebuie să aibă o activitate enzimatică mare.
Producerea enzimelor intracelulare ridică probleme specifice. Bioprocesul trebuie astfel condus încât pe parcursul biosintezei să se evite spargerea celulelor. Un alt aspect se referă la stabilitatea enzimelor; unele enzime sunt stabile în mediul intracelular şi devin instabile în afara celulei.
15

Unul dintre dezavantajele procedeelor de obţinere prin biosinteză a enzimelor intracelulare îl constituie necesitatea eliberării acestora din celule, prin distrugerea membranei celulare, extracţie şi ulterior separarea extractului de resturile celulare. In plus enzimele sunt impurificate cu toate proteinele intracelulare, chiar dacă enzime dorită se află în properţie mare în totalul proteinelor extrase din celule.
Un avantaj tehnologic al enzimelor intracelulare îl constituie faptul că extracţia se poate efectua cu un volum mic de soluţie tampon, ceea ce conduce la obţinerea unui extract brut cu conţinut mare de enzimă şi nu mai este necesară concentrarea ulterioară a acestuia.
Enzime extracelulare de fermentaţie
Cele mai importante enzime microbiene extracelulare sunt, în general hidrolazele( proteinaze, amilaze, celulaze) care actionează asupra substraturilor cu masă moleculară mare. Pentru microorganisme, este normal, din punct de vedere fziologic, să producă nutrienţi din polimeri biologici disponibili în mediul înconjurător celular.
Pentru a ataca mai uşor substraturile, microorganismele îşi produc singure enzimele necesare hidrolizei acestora şi uneori această producţie de enzimă este optimă chiar la tulpinile sălbatice.
Enzimele hidrolitice produse prin fermentaţie şi utilizate în cantităţi mari în industrie sunt proteazele, amilazele, celulazele. Acestea sunt comercializate şi utilizate în industria alimentară, textilă, detergenţilor.
Pentru că productia unei enzime extracelulare utilizează în cea mai mare parte resursele diponibile ale celulei, biosinteza şi secreţia unei astfel de enzime sunt supuse unui complex de factori regulatori. In general, producţia acestor enzime este indusă de niveluri reduse ale polimerilor, iar uneori polimerii înşişi funcţionează ca inductori. Alteori, compuşii care nu sunt substraturi pentru enzime, dar sunt înrudiţi ca structură cu acestea, pot funcţiona ca inductori. De exemplu, producţia de celulaze este indusă atât de lactoză, cât şi de celobioză. Producţia acestor enzime este de asemenea supusă represiei prin cataboliţi. Atâta timp cât nutrienţii cu masă moleculară mică sunt diponibili, celulele cresc fără să producă hidrolaze şi abia la epuizarea nutrienţilor începe biosinteza enzimelor hidrolitice extracelulare. Astfel, inducţia şi represia biosintezei enzimelor sunt fenomene complexe ce au loc în concordanţă cu condiţiile de mediu în care se găsesc celulele.
Trebuie reţinut faptul că factorii care determină înmulţire şi dezvoltarea celulei vor determina şi declanşarea biosintezei enzimelor. Un rol important în formarea enzimei îl are existenţa în mediul de cultură a substratului care induce formarea enzimei specifice degradării respectivului substrat. El constituie inductorul operonului enzimei.
Primul studiu sistematic efectuat în această direcţie a fost efectuat de Karström, care împarte enzimele în două grupe: enzimele constitutive şi enzimele adaptative.
Enzimele constitutive se sintetizează în celule în mod permanent. Concentraţia lor este însă influenţată de prezenţa sau absenţa în mediul de cultură a substraturilor pe care ele le metabilizează. Concentraţia aceastor enzime poate să varieze şi sub influenţa altor factori: sursa de azot, sursa de carbon, factorii de creştere, sărurile minerale, temperatura, pH-ul.
In schimb, sinteza enzimelor adaptative este declanşată numai de prezenţa substraturilor specifice în mediul de cultură sau atunci când este nevoie de prezenţa lor în procesele metabolice.
Pe baza unor studii îndelungate şi de profunzime cu privire la biosinteza enzimelor induse de substrat s-a ajuns la următoarele concluzii:
- Elaborarea de către un organism a unei enzime induse are loc numai în prezenţa inductorului; ca inductor funcţioneză, de obicei, substratul care trebuie metabolizat; funcţia de inductor poate să o îndeplinescă şi substanţele înrudite structural cu acesta, dar care nu poate îndeplini funcţia de substrat;
- Biosinteza are loc pornind întotdeauna de la substanţe cu structură simplă;
- Procesul de biosinteză decurge cu consum de energie şi din acest motiv, pe lângă substanţele necesare sintezei(aminoacizi, vitamine, diferiţi cationi) este nevoie în mediu şi de o sursă de carbon, de obicei un glucid;
16

- Sinteza enzimelor induse are loc, de regulă, în timpul înmulţirii microorganismului; ea poate sa aibă loc în faza staţionară, dacă sunt prezente inductorul şi substraturile de sinteză;
- Procesul de biosinteză al enzimelor este inhibat de toate substanţele care inhibă biosinteza substanţelor proteice, de exemplu cloramfenicolul;
- Biosinteza enzimelor induse este inhibată ori de câte ori în mediul de cultură este prezent un substrat uşor metabolizabil de către enzimele constitutive ale microorgamismului;
- Biosinteza unei enzime induse este inhibată selectiv şi de metaboliţii rezultaţi din reacţiile de transformare ale inductorului, datorită activităţii acestuia; fenomenul se numeşte represie;
2.4 Enzime microbiene cu aplicaţii industriale. Domenii de utilizare
Principalele aplicaţii ale enzimelor utilizate pe scară largă în ultimele 2 decenii sunt prezentate în tabelul următor(Frost şi Moss,1985):
Denumirea
enzimei
Industriile
utilizatoare
Tipul de
fermentaţie
I Felul enzimei Forma de condiţionare
1 2 3 4 5
1.Enzime proteolitice
Enzime proteolitice alcaline bacteriene
Detergenţi
Procese alimentare
Pielărie
Submers Extracelulare(Ex)
Lichid(L)
Solid(S)
Enzime proteolitice bacteriene neutre
Industria alimentară Submers Extracelulare(Ex) Lichid(L)
Enzime proteolitice fungice
Panificatie
Industria brânzeturilor
Submers
Submers în culturi de suprafată
Extracelulare(Ex) Lichid(L)
Solid(S)
2.Enzime amilolitice
α –amilaze bacteriene Amidon, bere
Detergenţi, textile
Submers Extracelulare(Ex) Lichid(L)
Solid(S)
α –amilaze fungice Amidon
Panificaţie
Submers Extracelulare(Ex) Lichid(L)
Solid(S)
Amiloglucozidaze Amidon
Bere
Submers Extracelulare(Ex) Lichid(L)
Solid(S)
Pullulanaze Amidon Submers în culturi de suprafaţă
Extracelulare(Ex)
Intracelulare(In)
Lichid(L)
Solid(S)
3. Alte glucozidaze
Lactază(β-galactozidază)
Industria laptelui Submers in culturi de suprafaţă
Extracelulare(Ex)
Intracelulare(In)
Lichid(L)
Solid(S)
Invertază Industria textilă Submers Intracelulare(In) Lichid(L)
Solid(S)
Rafinază(α -galactozidază)
Rafinarea zahărului Submers Intracelulare(In) Imobilizată(Im)
Celulaze Industria alimentară(sucuri)
Submers Extracelular(Ex) Lichid(L)
Solid(S)
Β-1,3(4)-Glucanaze Industria berii Submers Extracelular(Ex) Lichid(L)
17

Lipaze Industria alimentară
Teste de diagnostic
Submers Extracelular(Ex) Solid(S)
Pectinaze Industria sucurilor şi băuturilor alcoolice
Submers in culturi de suprafaţă
Extracelular(Ex) Lichid(L)
Solid(S)
4. Alte enzime
Glucozizomeraza Amidon Submers Intracelulare(In) Imobilizată(Im)
Glucozoxidaza Industria sucurilor şi băuturilor alcoolice
Teste de diagnostic
Submers Intracelulare(In) Lichid(L)
Solid(S)
Catalaza Industria sucurilor Submers Intracelulare(In) Lichid(L)
Solid(S)
Penicilinacilaza Antibiotice Submers Intracelulare(In) Imobilizată(Im)
O analiză a pieţei comerciale arată că enzimele obţinute prin procese fermentative microbiene reprezintă aproximativ 80% din totalul producţiei de enzime produse astăzi în lume. Dintre celelelte enzime, obţinute prin extracţie, necesitătile sunt acoperite de următoarele preparate: cheag de viţel, β-amilază din orz, proteinază pancreatică, etc.
Dintre enzimele de fermentatie cea mai mare cantitate o reprezintă proteinazele alcaline bacteriene utilizate în industria detergenţilor.
3. OXIDOREDUCTAZE IMPORTANTE ÎN BIOTEHNOLOGIILE ALIMENTARE
Oxidoreductazele sunt enzimele participante în procesele de oxidare biologică, care catalizează reacţiile de oxidoreducere printr-o serie de mecanisme: transfer de hidrogen (transhidrogenaze sau dehidrogenaze), transfer de electroni (transelectronaze), combinarea directă a unui substrat cu oxigenul molecular (oxidare) etc. Ele acţionează asupra unei perechi de substraturi, A şi B, dintre care cel redus se va oxida, iar cel oxidat se va reduce, după schema generală:
Ared + Boxid Aoxid + Bred
In funcţie de grupările chimice de substanţele donatoare de hidrogen asupra cărora acţionează, sau de substanţele care încorporează oxigenul etc., se împart în 17 subclase, în cadrul cărora se diferenţiază diferitele sub-subclase de oxidoreductaze. Aceste enzime îşi exercită acţiunea lor catalizatoare prin intermediul unor coenzime (nicotin-adenindinucleotidice NAD+, NADP+; flavinice ca "flavinmononucleotidul FMN sau flavinadenindinucleotidul FAD, heminice, etc.), care se comportă ca acceptori intermediari între substratul donor, iniţial, şi cel acceptor, final .
Numeroase oxidoreductaze intervin în procesele metabolice, atît anaerobe cât şi aerobe, ca, de exemplu, glicoliza şi diferitele fermentaţii, ciclul acizilor tricarboxilici, catena de respiraţie celulară etc., care se desfăşoară în diversele materii prime alimentare şi în procesele fermentative de obţinere a unor produse alimentare. Unele oxidoreductaze proprii materiilor prime, în special vegetale, folosite în industria alimentară sunt implicate în procese degradative ca, de exemplu: în îmbrunarea enzimatică (polifenoloxidaze, peroxidaze etc.), în procesele de albire sau decolorare (lipooxigenaza), în degradarea acidului ascorbic (acid ascorbic oxidaza), deteriorarea oxidativă a unor produse (peroxidaze, catalaze) etc. De asemenea, unele preparate enzimatice cu activitate oxidoreductazică sunt folosite în diferite scopuri în industria alimentară ca, de exemplu- glucozoxidaza pentru protejarea unor alimente împotriva deteriorărilor oxidative şi îmbrunarea Maillard, sau catalaza pentru eliminarea H2O2 reziduale în „pasteurizarea" la rece a laptelui.3.1. Oxidoreductaze NAD+ sau NADP+ dependente
18

Aceste enzime catalizează procesele reversibile de oxidoreducere caracterizate prin transfer de hidrogen de pe un substrat donor pe altul acceptor şi au caracter anaerob, deoarece acceptorul de hidrogen este altul decît oxigenul. Sunt denumite adesea dehidrogenaze, transhidrogenaze şi dehidraze. Coenzimele lor, NAD+ sau NADP+, care se comportă ca acceptori sau donori intermediari de hidrogen între substraturile participante în reacţie, se detaşează uşor de apoenzimă, puţind să-şi exercite acest rol pentru mai multe enzime. Oxidoreductazele NAD+ dependente intervin în numeroase procese catabolice oxidative ale alcoolilor, aldehidelor, aminoacizilor, iar cele NADP+ dependente catalizează mai ales procese anabolice reductive. Ele sînt foarte răspîndite în diverse organe şi ţesuturi animale şi vegetale, precum şi în diferite microorganisme. Dehidrogenazele prezintă deci importanţă şi în numeroase procese fermentative şi în procesul de respiraţie care se desfăşoară în diverse materii prime sau în procesele fermentative de obţinere a unor produse alimentare.
Alcool dehidrogenaza (alcool: NAD+—oxidoreductaza). Catalizează transformarea acetaldehidei, rezultată din decarboxilarea acidului piruvic, în alcool etilic, după reacţia:
Glucide Acid piruvic Aldehidă acetică Alcool etilic
Reacţia prezintă importanţă deosebită în procesul de fermentaţie alcoolică; în condiţii aerobe, enzima poate să catalizeze reacţia de oxidare a alcoolului etilic în aldehidă acetică. Enzima are masa moleculară 150000 şi conţine Zn2+ în structura ei.
Lactat dehidrogenaza (L-lactat: NAD-oxidoreductaza). Catalizează (în condiţii anaerobe) reducerea acidului piruvic cu formare de acid lactic în prezenţă de NADH + H+ conform reacţiei:
Acidul lactic constituie etapa finală a glicolizei şi a fermentaţiei lactice. Lactat dehidrogenaza izolată din inima şi muşchii mamiferelor sau din unele bacterii ca, de exemplu, Bacillus subtilis, conduce la formarea de acid L-lactic, deosebindu-se de lactat dehidrogenază obţinută din Lactobacillus plantarum care duce la formarea de acid D-lactic. In general, microorganismele care realizează fermentaţia lactică produc acid lactic racemic, deoarece acidul L-lactic rezultat este parţial izomerizat la acid D-lactic, prin intervenţia unei izomeraze.
Malicdehidrogenaza (L-malat: NAD+ -oxidoreductaza). Catalizează reacţia de oxidare a acidului malic în acid oxalilacetic, care mai departe este transformat în acid lactic.
Malatoxidaza. Denumită şi enzimă malică (malat: NAD—oxidoreductaza), malatoxidaza catalizează reacţia de transformare a acidului malic în acid piruvic, acesta fiind apoi transformat în acid lactic.
19

Aceste două enzime intervin în fermentaţia malolactică a vinurilor. Alături de aceste două oxidoreductaze, în fermentaţia lactică a vinurilor intervine şi enzimă malolactică (L-malat: NAD-carboxiliaza) care catalizează reacţia:
Acetaldehiddehidrogenaza (Acetaldehida: NAD-oxidoreductaza). Catalizează oxidarea aldehidei acetice în acid acetic, mai ales în procesul de oţetire a vinurilor.In condiţii aerobe, alcoolul etilic din vin, sub influenţa alcooldehidrogenazei, este transformat în aldehidă acetică iar aceasta sub influenţa acetaldehiddehidrogenazei se oxidează în acid acetic.
3.2. Oxidoreductaze FAD sau FMN dependenteAceste enzime cunoscute şi sub denumirea de flavinenzime au drept coenzime FAD-ul sau
FMN-ul care sînt derivaţi ai riboflavinei (vitamina B2). Unele flavinenzime conţin în molecula lor şi ioni metalici ( MO, Fe), care le conferă uneori şi funcţia de a transporta electroni. In general coenzimele flavinenzimelor sînt strîns asociate moleculelor de apoenzimă (deo-sebindu-se din acest punct de vedere de NAD+ şi NADP+ care se pot detaşa de apoenzimă). Unele flavinenzime acţionează ca dehidrogenaze anaerobe, iar altele au caracter aerob, putând transfera hidrogenul direct de la substratul donor către oxigenul molecular cu formare de apă oxigenată. In cadrul acestor din urmă enzime se încadrează glucozoxidaza, care îşi găseşte multiple utilizări în industria alimentară.
Glucozoxidaza (Notatina, -D-glucozo-O2-transhidrogenaza, -D-gluco-oxigen-oxidoreductaza, EC 1.1.3.4). Este o flavoenzimă care conţine 2 moli de flavinadenindinucleotid (FAD) ca grupare prostetică/mol enzimă.
Se obţine din Aspergillus niger (S.U.A.), Penicillium amagasakiense (Japonia) şi Penicillium vitale (Rusia). Glucozoxidaza din Aspergillus este considerată enzimă intracelulară implicată în mecanismul metabolismului energetic In practică, tulpinile de Aspergillus niger sînt cultivate submers şi miceliul respectiv este separat de mediul de cultură; miceliul, care conţine enzimă, este dezintegrat şi glucozoxidaza trece în soluţie care este centrifugată pentru separarea pereţilor celulari. După purificări repetate, enzima este precipitată cu acetonă sau alcool etilic, precipitatul fiind separat prin filtrare sau centrifugare, uscat, apoi extras cu soluţie tampon. Enzimă în soluţie este standardizată şi stabilizată, putând fi păstrată cîţiva ani la temperatura de refrigerare.Glocozoxidaza din Penicillium este purificată direct din mediul de cultură, fiind considerată o enzimă extracelulară, fapt care este în neconcordantă cu funcţia sa în procesele bioenergetice ale celulei. De fapt, miceliul de Penicillium se autolizează rapid, în comparaţie cu Aspergillus, şi enzima este trecută în mediul de biosinteză. Prin urmare, glucozoxidaza din Penicillium este tot o enzimă intracelulară. Activitatea acestor enzime rămîne neschimbată în intervalul de temperatură 30...60°C.
Enzimă posedă o specificitate de substrat foarte ridicată, glucoza fiind oxidată de 160 ori mai repede decît -glucoza, dar aceasta nu are o semnificaţie practică deoarece mutarotaza (aldozmutarotaza, EC 5.1.3.3.) restabileşte permanent echilibrul între şi -D-glucoză.
3.3. Transelectronaze
20

Sunt enzime care catalizează reacţiile de oxidoreducere prin transfer de electroni de pe un donor către un acceptor. In funcţie de natura acceptorului, transelectronazele pot fi: anaerobe, cînd acceptorul este o altă substanţă diferită faţă de oxigenul molecular, şi aerobe, cînd acceptorul este oxigenul. Transelectronazele anaerobe sînt cunoscute şi sub denumirea de citocromi, care sînt localizaţi la nivelul mitocondriilor, unde participă ca transportori de electroni în catena de respiraţie celulară.
3.4. Oxidaze
Sunt oxidoreductaze care catalizează reacţiile unor substraturi cu oxigen molecular sau al peroxizilor, fiind caracterizate prin transfer de hidrogen de pe un donator pe un acceptor care este oxigenul molecular. Ca produs de reacţie se formează H2O2.
În funcţie de mecanismele de acţiune oxidazele pot fi:— oxigenaze: dioxigenaze şi monooxigenaze;—oxidaze transportoare de electroni: citocromoxidaza, cupruoxidazele (tirozinaze, polifenoloxidazele, cateholoxidaza, ascorbatoxidaza etc.);— hidroxiperoxidaze (peroxidaze şi catalaze).
Lipoxigenaza este o dioxigenază (lipoxidaza linoleat: oxigenoxidoreductaza, EC 1.13.1.13), fiind cunoscută şi sub denumirea de carotenoxidază. Catalizează oxidarea acizilor graşi polinesaturaţi care conţin legăturile cis, cis- 1,4 pentadienice cu ajutorul O2. Acizii graşi polinesaturaţi oxidaţi sunt acidul linoleic (9, 12 octadecadienoic), linolenic (9, 12, 15 octadecatrienoic) şi arahidonic (5,8, 11, 14 eicosatetraenoic). Aceşti acizi graşi sînt denumiţi esenţiali. Sînt oxidate şi gliceridele şi metilesterii acizilor graşi menţionaţi. Se formează peroxizii respectivi .
Lipoxigenaza se găseşte în mazăre, fasole, arahide, cartofi, ridichi, dar activitate lipoxigenazică extrem de mare o au boabele de soia. Enzima este activă si la temperaturi scăzute şi prin acţiunea ei se formează mirosuri nedorite în produsele vegetale respective.
Boabele de soia conţin două tipuri de lipoxigenază, una activată de Ca2+ iar cealaltă inhibată de Ca2+, vîrful de activitate fiind suma efectelor pozitive şi negative, funcţie de modificările concentraţiei de calciu. Lipoxigenaza din soia nu necesită activare cu metale sau grupare prostetică. Antioxidanţii obişnuit folosiţi sunt inhibitori slabi ai lipoxigenazei iar acidul ascorbic şi tocoferolii accelerează reacţiile de oxidare.Lipoxigenaza din soia se utilizează pentru albirea făinii de grîu prin distrugerea carotenilor.
Polifenoloxidazele (denumite cresolaze, catecholoxidaze, cateholaze, tirozinaza, fenolaza; denumire sistemică O-difenol: oxigenoxidoreductaza, EC 1.10.3.1) sunt enzime care transportă electronii direct pe oxigen. Catalizează hidroxilarea monofenolilor cu formare de O-difenoli, care sunt transformaţi în O-chinone. Sunt larg răspândite în plante, găsindu-se în cantitate mai mare în special în ciuperci, tuberculi de cartofi, frunze de ceai şi tutun, boabe de cafea şi în diferite fructe ca mere, piersici, caise, banane etc.
Polifenoloxidazele din plante nu prezintă o specificitate deosebită, puţind să acţioneze asupra unor mari varietăţi de compuşi monofenolici şi O-difenolici. In schimb, polifenoloxidazele din ţesuturile mamiferelor au o specificitate mult mai restrînsă, acţionează doar asupra tirozinei şi dihidroxifeni-Lalaninei.
Îmbrunarea materialelor vegetale (legume, fructe proaspete tăiate) în contact cu oxigenul din aer este datorată polimerizării sau polimerizării oxidative a chinonelor formate din mono- şi difenoli, asupra cărora au acţionat polifenoloxidazele. Prin folosirea unor tratamente termice ca, de exemplu: în cazul blanşării legumelor şi fructelor, are loc inactivarea polifenoloxidazelor. De asemenea, concentraţii relativ mici de clorură de sodiu, de agenţi de complexare a cuprului, de acid ascorbic, S02 pot preveni sau minimaliza procesele de îmbrunare enzimatică a legumelor şi fructelor cauzate de polifenoloxidaze.
Peroxidazele sînt enzime care catalizează reacţii de oxidoreducere după reacţia generală:ROOH + AH2 → H2 + ROH + Aîn care: ROOH poate fi peroxidul de hidrogen (H202) sau un alt peroxid organic.Aceste enzime sînt larg distribuite în microorganisme (bacterii, drojdii), în plante ca, de exemplu,
napi, gulii, cartofi, smochine şi diferite alte legume şi fructe, în ţesuturi şi lichide biologice animale ca, de exemplu, ficat, lapte, salivă, în leucocite etc.
21
Din punct de vedere structural pot fi grupate în: peroxidaze feriprotoporfirice (cu grupare prostetică feriprotoporfirina III) prezente în plantele superioare (legume şi fructe), în animale (ca, de exemplu, triptofan-pirolaza, tiroid-iodin-peroxidaza) şi în microorganisme (citocrom c-peroxidaza din drojdii); verdo-peroxidaze (cu grupare prostetică porfirinică diferită de feri-porfirina III) de culoare verzuie, cum este lactoperoxidaza din lapte şi flavo-protein-peroxidaza (cu FAD ca grupare prostetică) de felul celor găsite în diferiţi streptococi dar şi în unele ţesuturi animale.
În general, peroxidazele prezintă o mare stabilitate termică. De exemplu, pentru unele peroxidaze s-a constatat păstrarea a circa 50% din activitatea iniţială după un tratament termic de 32 min la 85°C, 12 min la 100°C, 2,5 min la 116°C şi 0,4 min la 145°C. Datorită termostabilităţii lor, determinarea activităţii peroxidazice din diferite legume şi fructe este folosită ca un indice al eficienţei tratamentelor termice aplicate în procesele de blanşare, sterilizare etc. Determinarea activităţii lactoperoxidazei este folosită la urmărirea eficienţei pasteurizării în industria laptelui.
Diferitele peroxidaze intervin în degradarea fructelor şi legumelor, fiind implicate în modificarea mirosului şi gustului (mai ales la mazăre, fasole etc.) în procesele de îmbrunare enzimatică.
Sub formă de preparate enzimatice, în special peroxidaza obţinută din hrean, sînt folosite, alături de glucozoxidază, în determinarea specifică a glucozei din diferite materiale biologice.
4. HIDROLAZE IMPORTANTE ÎN INDUSTRIA ALIMENTARĂ
Hidrolazele sunt enzime care catalizează scindarea hidrolitică a substraturilor, determinând desfacerea legăturilor dintre atomii de carbon şi alţi atomi prin introducerea elementelor apei. Principalele legături scindate la hidrolaze sunt legăturile: ester glicozidazică şi peptidică.
4.1. Lipaze şi esterazeEnzimele lipolitice sînt importante în metabolismul şi degradarea lipidelor. Importanţa lor
fiziologică constă în aceea că hidrolizează grăsimile alimentare cu formare de di-, monogliceride şi acizi graşi, produşi care sînt transportaţi, oxidaţi şi sintetizaţi în gliceride şi fosfolipide proprii organismului. Importanţa lor tehnologică constă în faptul că realizează hidroliza controlată a lipidelor, în vederea formării de arome în cadrul alimentelor (de exemplu,, în cazul brânzeturilor) sau în sisteme pregătite special pentru obţinerea de aromatizanţi. Enzimele lipolitice sunt implicate însă şi în degradarea unor produse alimentare cu conţinut ridicat în lipide (lapte şi produse lactate, carne, peşte, produse vegetale bogate în lipide, cum ar fi arahidele, seminţe de floarea soarelui, soia, germeni de grîu şi porumb etc.).
Lipazele se diferenţiază de esteraze prin afinitatea lor mai mare pentru acizii graşi cu lanţ lung din structura gliceridelor. Pe de altă parte, pe plan structural, lipazele prezintă o porţiune hidrofilă şi alta lipofilă, fapt ce le permite plasarea lor la interfaţa ulei/apă, acţiunea lor fiind favorizată de calitatea emulsiilor de tipul ulei/apă. Sărurile de calciu şi proteinele sunt activatori (efectori) ai lipazelor. In cazul lipazelor din sucul pancreatic, rol de activator îl au şi sărurile acizilor biliari.
Enzimele lipolitice de tipul lipazelor şi fosfolipazelor sunt prezente ca enzime endogene în diferite ţesuturi animale, lapte, produse vegetale.
Laptele conţine o lipaza (cea mai importantă) care are pH optim la 8,5—9 şi alte 2—3 lipaze care includ şi o lipaza acidă. Proteinele din lapte au acţiune inhibitoare faţă de lipaze. Clorura de calciu (CaCl2) inhibă semnificativ lipazele din lapte. Lipazele din lapte hidrolizează rapid trigliceridele simple, viteza de hidroliză micşorîndu-se o data cu creşterea lanţului de acid gras din structura trigliceridei. Lipazele din lapte posedă si activitate de sinteză:glicerol + acid gras → gliceride + apă
Sinteza gliceridelor este influenţată de concentraţia enzimei, concentraţia glicerolului, temperatura şi durata incubării.
Fructele şi legumele, dar în special seminţele de grîu, ovăz, secară, soia etc., conţin lipaze care provoacă deteriorări ale acestora în condiţii nefavorabile de umiditate relativă mare a aerului, conţinut mare de umiditate al cerealelor, temperatură ridicată. Datorită creşterii conţinutului de acizi graşi liberi scad proprietăţile de panificare a făinurilor de grîu. Germenii de grîu, secară, orz au o activitate lipolitică mai mare decît endospermul, deci sunt mai bogaţi în lipaze. Scutelum şi stratul aleuronic, de asemenea, conţin mai multe lipaze decît endospermul.
22

Lipazele produse de microorganisme sînt produse de drojdii (Candida, Torulopsis) , mucegaiuri (Rhizopus, Penicillimmi, Aspergillus, Geotrichum, Mucor), bacterii (Pseudomonas, Achromobacter, Staphylococcus). Lipazele din microorganisme diferă între ele prin pH şi temperatura optimă de acti-vitate, durata/temperatura de inactivitare, respectiv stabilitatea termică.
Lipazele produse de microorganisme sînt considerate ca lipaze adevărate care hidrolizează grăsimile şi uleiurile naturale ca şi gliceridele sintetice. Concentraţii scăzute de săruri de Ca, Na, K, Mg activează lipazele, dar metalele grele sînt inhibitori puternici.
Avînd în vedere acţiunea negativă a lipazelor asupra caracteristicilor senzoriale ale produselor alimentare, este necesar să se ia următoarele măsuri:— să se reducă conţinutul în apă liberă al produsul alimentar (acolo unde este posibil), pentru a micşora activitatea apei (aw), deci pentru micşorarea vitezei reacţiilor enzimatice;— să se păstreze produsele alimentare la temperaturi cât mai scăzute (de refrigerare sau congelare) sau să se inactiveze lipazele prin tratament termic ;— să se reducă timpul de depozitare al materiilor prime ;— să se elimine factorii care concură la dezvoltarea microflorei lipolitice.
4.2. Enzime oligozidazice
Dintre oligozidaze, mai importante pentru industria alimentară sunt -glucozidaza (EC 3.2.1.21), -D-galactozidaza (EC 3.2.1.22), -D-galactozidaza (EC 3.2.2.23), -D-fructofuranozidaza (EC 3.2.1.26).
-Glucozidaza (EC 3.2.1.21), denumită şi emulsină, scindează legătura -glucozidică din -glucozii răspîndiţi în plante cum ar fi amigdalină, prunazină, naringină, dar şi din celobioză şi gentiobioză. Această enzimă este produsa şi de unele microorganisme: Alcaligenes faecalis, Botryodiplodia theobromae, Phoma strasseri, Septoria licopersici, Aspergillus niger, Myrothecium verucaris, Trichoderma viride, Saccharomyces lactis etc.
-D-Galactozidaza (EC 3.2.1.32) este prodsuă de B. stearothermophilus, Penicillium duponti, Absidia griseola, Mortierella vinaceae, Aspergillus awamori. Substratul natural al acestei enzime este rafinoza, trizaharid format din galactoză, glucoza şi fructoză. Enzima desface rafinoza în galactoză şi zaharoză şi are aplicaţii în industria zahărului şi industria derivatelor proteice vegetale.
23

-D-Galactozidaza (EC 3.2.2.23), cunoscută şi sub numele de lactază hidrolizeză lactoza la glucoza şi galactoză (fig. 13). Este găsită în unele plante, microorganisme şi în mucoasa intestinală a mamiferelor. Este obţinută din tulpini de mucegaiuri (Aspergillus niger, Aspergillus oryzae, Aspergillus foetidis), din drojdii (Kluyveromyces fragilis, Kluyveromices lactis, Candida pseu-dotropicalis, Torulopsis lactis) şi bacterii (Lactobacillus bulgaricus, Streptococcus thermophilus, Escherichia coli).
Preparatele lactazice din drojdii acţionează bine la pH ≈6 şi la temperatura de 35 ... 40°C, deci la pH-ul substratului natural (lapte degresat, lapte concentrat), cele de origine fungică au activitate maximă la pH ≤ 5 şi la temperatura de 50 ... 60 "C, fiind recomandate pentru hidroliza lactozei din zer. Din Aspcrgillus oryzae şi Aspergillus niger se obţin lactaze acidoactive şi acidostabile, folosite la hidroliza lactozei în mediu acid. Lactaza de origine bacteriană (E. coli) acţionează bine la pH ~ 7,3 şi la 46 °C. Ionii de sodiu şi potasiu sînt activatori ai lactazei din E. coli şi din drojdii.
Preparatele enzimatice de lactază se utilizează pentru obţinerea de produse lactate destinate şi adulţilor care prezintă intoleranţă la lactoză (deficientă congenitală, primară sau secundară de lactază), pentru obţinerea siropului'de lactoză ce are multiple aplicaţii în industria alimentară, pentru împiedicarea apariţiei unor defecte în produsele lactate concentrate şi congelate. Se utilizează şi în panificaţie, în cazul utilizării zerului pulbere la fabricarea pîinii.
4.3. AmilazeAmidonul, cel mai important homoglucid de rezervă din regnul vegetal, se găseşte în diferite
materii prime folosite în industria alimentară, de obicei sub formă de granule, în cele două componente principale, amiloza şi amilopectina, care sînt dispuse în straturi concentrice, fiind însoţite de mici cantităţi de alte substanţe organice (protide, lipide) şi substanţe anorganice (H3PO4, Si03, H2O). Amiloza care reprezintă 20—25% din amidon, este alcătuită din unităţi de glucoza legate între ele prin legături α l-4 glucozidice, gradul de polimerizare fiind de 200—6000 (în medie 103). Amilopectina care reprezintă 75—80% din amidon este formată din lanţuri liniare de glucoza legate α l-4 glucozidic, lanţuri care se grefează unele pe altele prin legături α l-6 glucozidice. Numărul legăturilor α l -6 glucozidice reprezintă 5—6% din ansamblul de legături din amilopectină, gradul de polimerizare al acesteia fiind de 105—109
resturi de glucoza. Sub formă de granule, amidonul este rezistent la acţiunea enzimelor amilolitice. In general,
enzimele amilolitice acţionează numai în condiţiile în care granulele de amidon sunt cel puţin lezate (de exemplu în cazul amidonului din cereale supuse măcinării) şi mai ales cînd amidonul a fost gelificat prin încălzire în prezenţă de apă, realizîndu-se pe această cale o hidratare, o umflare şi deci o spargere a granulelor de amidon. Sub această formă amidonul devine uşor atacabil de enzimele amilolitice care, acţionînd asupra amilozei şi amilo-pectinei, produc desfacerea hidrolitică a legăturilor α l-4 glucozidice, determinând depolimerizarea acestora şi formarea de dextrine de diferite mărimi, maltoză, glucoza.
Enzimele implicate în degradarea amidonului pot fi endogene, care se găsesc în cantitate mai mare în cereale încolţite, importante în cazul orzului ce este transformat în malţ, şi exogene, care în cea mai mare parte sunt de natură bacteriană sau fungică. Amilazele sunt bine caracterizate şi diferenţiate pe baza tipului de legătură hidrolizată, modul lor de atac (endo sau exo), natura substratului limită şi produşii de degradare. Enzimele care atacă amidonul sînt glucozilhidrolaze şi aparţin următoarelor grupe:a) Enzime care hidrolizează legăturile l-4 glucozidice
.-Amilaza, cunoscută şi sub denumirea de amilază dextrinogenică, diastaza, ptialină, glucogenază, -l,4-glucon 4-glucanohidroxilază (EC 3.2.1.1.), este o metalproteină care conţine calciu în proporţie de l atom gram/moleculă, strîns legat de enzimă. Prin îndepărtarea calciului, enzimă devine inactivă şi instabilă la căldură. Calciul nu participă direct la formarea complexului enzimă-substrat, dar menţine molecula de enzimă în configuraţia optimă pentru un maxim de activitate şi stabilitate, (în practica de hidroliza a amidonului se adaugă calciu sub formă de săruri). Enzimă are activitate la pH cuprins între 4,5 şi 7,0, pH-ul optim depinzînd de originea enzimei. Stabilitatea la căldură este de asemenea diferită în funcţie de originea enzimei. α-Amilaza hidrolizează mai mult sau mai puţin la întîmplare legăturile l-4 glucozidice din interiorul moleculelor de amiloză şi amilopectină, conducînd la dextrine de diferite mărimi, respectiv la lichefierea amidonului şi la o hidroliza prelungită şi la maltoză şi maltotrioze.
24

-Amilazele pot fi de origine animală (amilază produsă de glanda salivară şi pancreas), de origine vegetală (cereale încolţite în special malţ) cu importanţă în industria berii, spirtului şi panificaţiei şi de origine microbiană (bacterii, mucegaiuri şi mai puţin drojdii), cele de origine bacteriană fiind mai stabile la căldură decît cele de origine fungică.
-Amilaza, denumită şi amilază zaharigenică, α l-4 glucano-maltohidrolaza (EC 3.3.1.2) hidrolizează legăturile α l-4 glucozidice ale amidonului de la capetele nereducătoare ale lanţurilor poliglucidice (este exoenzimă). Amiloza cu număr impar de unităţi de glucoza este transformată în maltoză în proporţie de 90% şi în maltotrioze. La concentraţii mari ale enzimei şi la hidroliză prelungită, maltotrioza este degradată lent la glucoza şi maltoză. Amilopectina este degradată şi ea în maltoză dar hidroliză se opreşte la legăturile α l—6 glucozidice care nu pot fi hidrolizate şi nici by-passate. Se for-mează în acest fel 60—70% maltoză, restul fiind dextrine limită. -amilaza are pH-ul optim între 5 şi 6 iar temperatura optimă ~55 ... 63 °C. Pentru activitatea enzimatică sînt necesare grupările —SH.Surse importante de -amilază sunt orzul, grâul, secara, cartofii dulci şi soia. Recent s-a dovedit că este produsă şi de unele microorganisme (Bacillus, Streptomyces ), avînd pH-ul optim la 5—6 şi temperatura optimă la 60 °C.b) Enzime care hidrolizează legăturile α l-6 glucozidice. Sunt cunoscute şi sub denumirea de enzime de debranşare (deramificare) ce acţionează asupra arnilopectinei din amidon, în cadrul acestora sînt mai bine cunoscute următoarele enzime:— pullulanaza (EC 3.2.1.41), care hidrolizează poliglucidul pullulan, conducînd la eliberarea de maltotrioza. Acţionează, de asemenea, şi asupra legăturilor α l -6 glucozidice din amilopectină şi în mai mică măsură asupra celor din structura glicogenului. Este produsă de numeroase microorganisme, dar se extrage, în principal, din Klebsiella aerogenes.— izoamilaza (EC 3.2.1.68) poate deramifica atît amilopectină cit şi glicogenul. Acţionează foarte slab asupra pullulanului. Este produsă de specii dePseudomonas şi Citophaga. Aceste două enzime de deramificare pot completa 100% acţiunea de dextrinizare şi zaharificare a amidonului produsă de α şi β amilaze. Ca şi -amilaza, enzimele de deramificare nu acţionează asupra amidonului negelificat.c) Enzime care hidrolizează atît legăturile l-4cît şi l-6 glucozizidice. În cadrul acestora o deosebită importanţă o prezintă: glucoamilaza ce hidrolizează mai repede legăturile α l-4 dar şi cele α l-6 de la capetele nereducătoare ale lanţurilor poliglucidice din amidon, conducînd la glucoza. Randamentul de transformare a amidonului în glucoza este afectat de capacitatea enzimei de a polimeriza glucoza la maltoză şi maltotrioza, reacţie care este dependentă de concentraţia ridicată a substratului. Preparatele comerciale de amiloglucozidaza sînt de origine fungică (Aspergillus, Rhizopus), fiind impurificate cu lipaze, tanaze, celulaze, hemicelulaze, proteaze.
4.4. Enzime pectolitice :Enzimele care acţionează asupra substanţelor pectice pot fi clasificate în două grupe: enzime
saponifiante sau pectinesteraze şi enzime depolimerizante sau pectinglicozidaze.Pectinesterazele (pectinmetilesteraza — pectindemetoxilaza, pectinpectilhidrolaze, EC 3.1.1.11),
notate PME sau PE, hidrolizează esterul metilic al acidului galacturonic.Pectinesterazele se găsesc în diferite produse vegetale (tomate, ceapă, pepeni, mere, pere etc.) şi
au un optim de pH aproape de neutralitate. De exemplu, pectinesterazele din tomate şi citrice au pH-ul optim la 7,5, cele din mere la 6,6 şi cele din struguri la 5,6. Sînt relativ stabile la căldură.
Pectinglicozidazele sunt enzime depolimerizante. Preparatele enzimatice pectolitice sînt utilizate în mare măsură pentru ameliorarea extracţiei sucului celular din fructe şi legume (prin degradarea pereţilor celulari), pentru macerarea fructelor şi legumelor în vederea obţinerii de nectaruri (prin degradarea substanţelor pectice ale lamelelor mijlocii care asigură legătura dintre celule, fără a se distruge pereţii celulelor parenchimatoase) şi pentru limpezirea (clarificarea) diferitelor categorii de sucuri.
In cazul preparatelor enzimatice pectolitice folosite pentru macerare, este necesar ca acestea să posede şi activitate celulazică, hemicelulazică, proteazică şi amilazică.
4.5. Peptidhidrolaze (proteinaze şi peptidaze)Peptidhidrolazele — proteinazele şi peptidazele — cunoscute şi sub denumirea generală de
enzime proteolitice catalizează scindarea hidrolitică a legăturilor peptidice, din moleculele proteidelor,
25
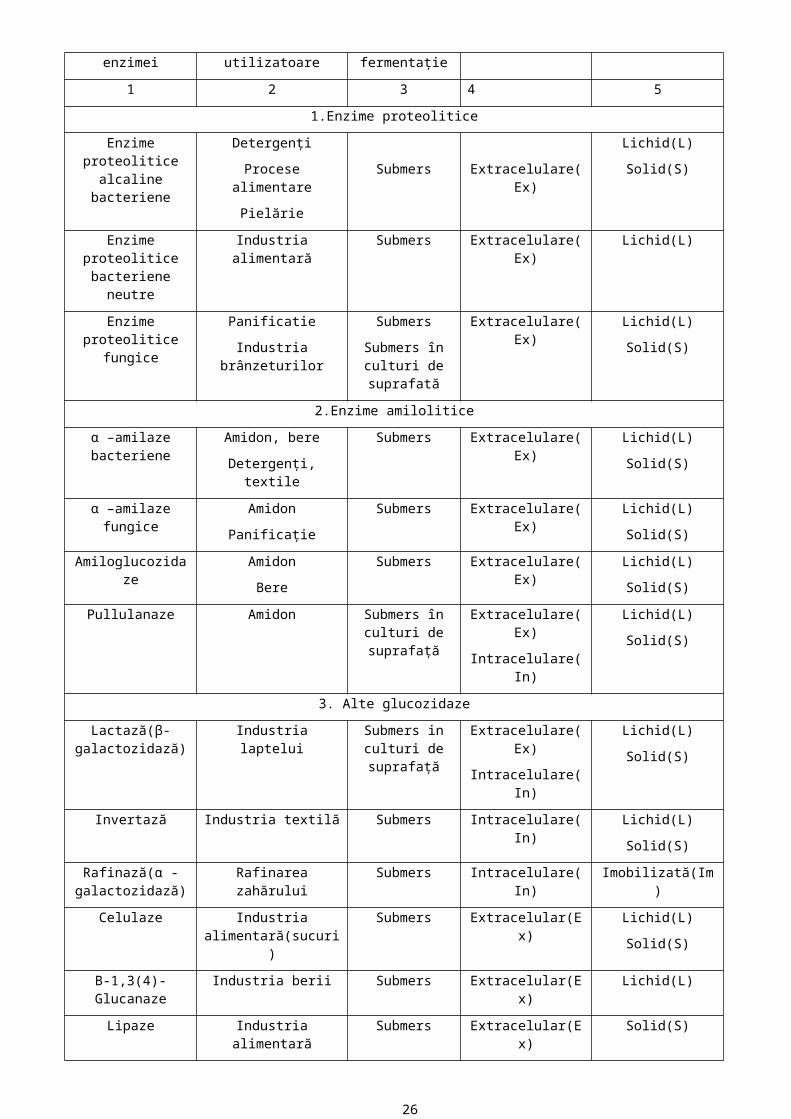
polipeptidelor şi oligopeptidelor, conducînd la eliberarea de peptide de diferite mase moleculare, sau chiar la aminoacizi, după următoarea reacţie generală:
În funcţie de regiunea din molecula proteidelor sau polipeptidelor la nivelul căreia acţionează, acestea se subdivid în endo- şi exopeptidhidrolaze ; endo-peptidhidrolazele, denumite şi proteinaze sau peptidilpeptidhidrolaze, scindează hidrolitic legăturile peptidice din interiorul catenelor polipeptidice ale proteidelor şi proteinelor cu masa moleculară mare, producând catene polipeptidice de masă moleculară mai mică, iar exo-peptidhidrolazele, peptidazele sau exopeptidazele catalizează desfacerea hidrolitică a legăturilor peptidice de la extremităţile catenelor polipeptidice sau oligopeptidice, conducînd, în general, la eliberarea de aminoacizi. Exo-peptidazele pot acţiona atât asupra produşilor de hidroliză a endo-peptidhidrolazelor cât şi asupra unor proteine nehidrolizate. La rîndul lor, exopeptidazele, se subdivid în aminopeptidaze, care desfac hidrolitic aminoacizii de la capetele N terminale ale catenelor peptidice (corespunzătoare resturilor de aminoacizi cu gruparea —NH2 liberă) şi carboxipeptidaze ce scindează hidrolitic aminoacizii de la capetele C terminale (corespunzătoare resturilor de aminoacizi cu gruparea —COOH liberă).
În cadrul exo-peptidazelor se grupează şi dipeptidazele, care desfac legăturile peptidice din dipeptide, precum şi dipeptidil-peptidhidrolazele, care scindează dipeptide de la capătul N terminal al lanţului polipeptidic şi peptidil-dipeptidhidrolazele ce eliberează dipeptide din capătul C terminal.
In cazul endo-peptidhidrolazelor, luîndu-se în considerare unele grupări funcţionale din centrii activi ai enzimelor, comportarea faţă de unii inhibitori sau activatori şi efectul pH-ului, se pot distinge:— proteinaze serinice, care au în centrul activ un rest de serină şi care sunt inhibate de unii compuşi organofosforici; acestui grup îi aparţin: tripsina, chimotripsina, elastaza, subtilizina (din Bacillus sp.), proteinaza alcalină din Aspergillus sp. etc.;— proteinaze tiolice sau SH-dependente, care au în centrul activ grupări —SH libere, corespunzătoare resturilor de cisteină şi care pot fi inactivate prin blocarea acestora cu metale grele, cu agenţi alchilanţi sau oxidanţi; din cadrul acestora fac parte: papaina, ficina şi bromelina;— proteinaze carboxilice (acide) al căror centru activ cuprinde grupări carboxilice ionizate şi care sunt active în domeniu de pH acid; din acest grup fac parte proteinazele care coagulează laptele, chimozina, pepsina, proteinazele produse de Mucor sp., precum şi proteinazele acide produse de Aspergillus sp. ;— metal-proteinaze, care sunt activate de ioni metalici legaţi de centrul activ (ca, de exemplu, Ca 2+, Zn2+, Mg2+, Fe2+); din grupul acestora fac parte: colagenazele, proteinaza neutră produsă de Bacillus subtilis, sau Aspergillus oryzae etc.Dintre peptidhidrolazele vegetale :
Papaina. Sub denumirea de papaină se înţelege preparatul brut ce se colectează din fructele de Caryca papaia, cu puţin înainte de coacerea acestora. Latexul colectat în contact cu aerul se coagulează, după care se usucă sub vid, se macină şi se cerne. Preparatul brut de papaia conţine de fapt trei fracţiuni: papaină propriu-zisă (10%), chimio-papaină (45%) şi lizozim (20%), ultima fracţiune fiind denumită şi papain-peptidază. Aceste fracţiuni diferă între ele prin masa moleculară şi punctul izoelectric.
Papaina propriu-zisă este o proteinază tiolică constituită dintr-un lanţ polipeptidic format din 212 resturi aminoacizi. Papaina are o activitate specifică largă hidrolizînd atît peptidele mici cît şi proteinele. Este foarte activă în hidroliza amidelor şi esterilor. Are pH-ul optim în jur de 6. Papaina are o stabilitate bună.
Chimopapaina. A fost obţinută în stare cristalizată din latexul de papaya şi este formată din chimopapaina A şi B. Are acţiune asemănătoare cu papaină, însă este mai puţin eficace asupra hemoglobinei şi este mai stabilă decît papaina. Chimopapaina îşi pierde 50% din activitatea sa dacă este menţinută 75 min la 75°C (pH = 7,2), faţă de 56 min pentru papaină.Preparatul brut de papaină atacă relativ bine fibrele musculare şi pe cele elastice dar mai puţin pe cele de colagen.
Bromelina. Este o enzimă proteolitica cu acţiune energică care se găseşte în sucul de ananas. Ca şi papaină şi ficină, bromelina este o sulfhidril-protează, însă are structura unei glicoproteine. Optimul de
26
activitate este la pH = 6,3 şi la 55°C. Are o acţiune bună asupra fibrelor de colagen, dar o acţiune slabă asupra celor elastice. Acţiunea asupra fibrelor musculare este foarte redusă.
Peptidhidrolazele de origine animală mai bine cunoscute şi mai des folosite sînt: chimozina, pepsina şi tripsina, cărora li se adaugă chimotripsina, elastaza şi carboxipeptidazele, aminopeptidazele şi dipeptidazele, produse de diferite glande digestive. Cu excepţia ultimelor două tipuri de exo-peptidaze, celelalte peptidhidrolaze sunt produse de glandele digestive sub formă de pro-enzime (preenzime sau zimogene) inactive, care devin active numai în urma scindării hidrolitice a unor porţiuni din structura lor, ce blochează activitatea lor enzimatică. Conversia proenzimelor în enzime active se realizează pe cale autocatalitică, care conduc la enzimă activă şi fragmente polipeptidice inactive enzimatic.
Chimozina. Cunoscută şi sub denumirea de cheag, rennină, presură sau labferment, chimozina constituie un preparat enzimatic cu acţiune coagulantă (asupra laptelui) şi proteolitica, asemănător pepsinei, obţinut din stomacuri de viţel, miel sau ied, folosit în special la prepararea brînzeturilor. Chimozina rezultă din precursorul inactiv — prochimozina — produsă de mucoasa stomacului glandular (al patrulea stomac al viţeilor) prin hidroliza parţială, proces influenţat de pH şi concentraţia în săruri.
Chimozina este o pretează serinică, cu specificitate de acţiune similară pepsinei, acţionînd mai ales la nivelul legăturilor peptidice în care sunt implicate resturi de triptofan, fenilalanina, tirozină, metionină şi leucină; manifestă activitate coagulantă şi proteolitică.
Pepsina. Endo-peptizadă produsă sub formă de pepsinogen de glandele stomacale ale mamiferelor inclusiv ale omului şi care se găseşte şi în sucul gastric al păsărilor, reptilelor şi peştilor, pepsina scindează hidrolitic legăturile peptidice din interiorul catenelor polipeptidice ale proteinelor, acţionînd de preferinţă la nivelul legăturilor peptidice în care sunt implicaţi aminoacizi aromatici (fenilalanină, tirozină sau triptofan) şi dicarboxilici (acizii glutamic şi aspartic). Transformarea pepsinogenului în pepsina catalitic activă se realizează la pH mai mic decit 5, în prezenţa ionilor H+ sau a unor mici cantităţi de pepsina, activarea fiind însoţită de eliberarea unui polipeptid alcătuit din 29 de resturi de aminoacizi, care acţionează ca inhibitor al pepsinei, şi cinci peptide mici. Pepsina este constituită dintr-un singur lanţ polipeptidic cu un rest izoleucinic la capătul N terminal şi un rest de valină la capătul C terminal. pH-ul optim de acţiune este 1,5—2,5. Este folosită la precipitarea cazeinei, obţinerea de peptone, în unele ţări pentru limpezirea berii la rece, ca adjuvant digestiv şi este inclusă în reţeta de preparare a unor băuturi tonice şi a chewing-gum-ului.
Tripsina. Este tot o endo-peptidază, care desface hidrolitic legăturile peptidice în interiorul catenelor polipeptidice, de preferinţă cele constituite între gruparea carboxil a unui aminoacid diaminic (arginină, lizină) şi gruparea aminică a altui rest de aminoacid. Este produsă de pancreas sub formă de tripsinogen, forma inactivă, care se transformă în tripsină activă în intestinul subţire sub acţiunea enterochinazei (o enzimă intestinală) sau pe cale autocatalitică, în prezenţa unor mici cantităţi de tripsină. Tripsina este constituită din 223 resturi de aminoacizi a căror secvenţiabilitate este cunoscută. Acţionează la pH neutru, slab alcalin, pH-ul optim fiind 8,5.
În ultimele decenii, datorită dezvoltării impetuoase a proceselor şi instalaţiilor de cultivare a microorganismelor la nivel industrial, pentru realizarea de diferite produse (antibiotice, vitamine, preparate enzimaticc etc.) pe cale biosintetică, s-a înregistrat un interes deosebit şi pentru cercetarea şi obţinerea de preparate peptidhidrolazice din microorganisme, în special din bacterii şi fungi. Interesul pentru astfel de preparate proteolitice a crescut şi mai mult după anul 1965, cînd s-au folosit astfel de preparate pentru obţinerea de detergenţi cu enzime, de tip Biotex, care conţin proteaze produse de Bacillus licheniformis.Multe din microorganismele folosite la producerea de preparate proteolitice produc aceste enzime extracelular, secretându-le în lichidele de cultură, ceea ce uşurează foarte mult condiţiile de prelucrare industrială pentru obţinerea de preparate enzimatice. Datorită acestor fapte, există în prezent tendinţa ca o serie de preparate proteolitice de natură vegetală sau animală să fie înlocuite din ce în ce mai mult cu preparate proteolitice microbiene, care sunt în general mai ieftine şi de o mai mare diversitate.
În prezent sînt cunoscute numeroase tulpini de microorganisme care secretă în mediile lor de cultură cantităţi apreciabile de enzime proteolitice; dintre acestea, însă, sunt folosite pentru obţinerea de preparate proteazice numai tulpinile nepatogene şi care nu produc toxine. De mai mulţi ani, în diferitele sectoare ale industriei alimentare, se folosesc proteaze obţinute mai ales prin cultivarea unor microorganisme ca: Bacillus subtilis, Bacillus licheniformis, Bacillus megaterium, Aspergillus niger, Aspergillus oryzae, Streptomyces sp., Rhizopus sp., Byssoclamys fulva, Endothia parasiiica, Mucor niiehei, Mucor pusillus etc.
27

Proteinazele microbiene nu sunt, în general, foarte diferite de cele vegetale sau animale; aparţin şi ele serin - sulfhidril- şi carboxil-proteinazelor, precum şi metalo-proteinazelor.
MICROORGANISME FOLOSITE IN PROCESELE BIOTEHNOLOGICE
Viaţa Terrei nu ar fi posibilă fără existenţa microorganismelor, pentru că acestea stabilesc echilibrul între substanţele organice şi cele anorganice, între materia vie şi materia nevie. Industria alimentară, în mare parte, este de neconceput fără existenţa microorganismelor care sînt folosite pentru conservare, obţinerea de produse alimentare datorită acţiunii lor fermentative (produse lactate acide, unt, brînzeturi, bere, vin, pâine, alimente fermentate din cereale şi leguminoase etc.). In general, produsele rezultate în urma acţiunii microorganismelor pot fi clasificate în celule şi biomasă, metaboliţi primari, metaboliţi secundari şi enzime.
Celule şi biomasă. Numeroase produse alimentare sunt consumate împreună cu celulele microorganismelor folosite pentru fermentare (produse lactate fermentate, produse de carne şi peşte fermentate, produse tradiţionale fermentate de origine vegetală, pâine) sau se consumă numai mediul fermentat, celulele fiind îndepărtate, cum este cazul băuturilor alcoolice de tipul berii, cidrului, vinului etc. In cazul biomasei, aceasta poate fi utilizată ca aditiv de fermentare (drojdie de panificaţie) sau aditiv de îmbogăţire a produselor alimentare cu proteine, respectiv ca furaje pentru păsări, animale, peşti. O serie de microorganisme sunt folosite sub forma de inocul de celule ca bioinsecticide, micoerbicide, vaccinuri.
Metaboliţii primari. Sunt molecule mici, vitale pentru celulele vii în vederea producerii de macromolecule sau pentru convertire în coenzime (nucleotide, vitamine). Metaboliţii primari de importanţă pentru industria alimentară sunt alcoolul etilic, poliolii (manitolul, sorbitolul, xilitolul), conservanţi (acidul acetic, acidul lactic, acidul citric, acidul propionic), antioxidanţi şi sechestranţi (acidul ascorbic, acidul citric etc.), potenţiatori de aromă (glutamatul monosodic, 5'—IMP, 5'—GMP), acidulanţi (acidul acetic, acidul citric, acidul malic, acidul gluconic, acidul lactic, acidul tartric, acidul fumărie etc.), aminoacizi (L-lizina, L-triptofanul, L-fenilalanină etc.), vitamine (riboflavina, ciancobalamine, vitamina C, vitamine D), gaze (CO2).
Metaboliţii secundari. Sunt produşi de un spectru larg de microorganisme şi pot fi folosiţi ca aditivi de stimulare (giberelinele) în producţia de malţ, de conservare (nizina) sau ca: coccidiostate, pesticide, bioinsecticide, inocul pentru seminţe, vaccinuri, antihelmintici etc., în producţia vegetală sau animală pentru a mări producţia de alimente necesare omului . De remarcat că, printre metaboliţii secundari se numără şi micotoxiele, în special aflatoxinele cu acţiune dăunătoare omului şi animalelor.
Producţia de metaboliţi secundari este afectată de mecanismele determinate genetic ca: derepresia (inducţia), reglarea catabolică, reglarea feed-back la care trebuie adăugată reglarea de by-pass.
Reacţiile de producere a celulelor, biomasei şi a metaboliţilor pot fi paralele, perfect consecutive (sau decuplate) sau parţial cuplate In funcţie de tipul de reacţie se conduce şi fermentaţia respectivă. Fermentaţiile pot fi conduse discontinuu sau continuu, impunîndu-se optimizarea lor. În această direcţie, în cazul fermentaţiei discontinue, optimizarea trebuie să rezolve două situaţii:
când substratul este inhibitor pentru producţia de metabolit (sau biomasă) când produsul este inhibitor (biomasă sau metabolitul).
Prima situaţie se întîlneşte, de exemplu, la obţinerea biomasei de drojdie de panificaţie, unde represiunea catabolică de către glucide antrenează o producţie de alcool etilic care trebuie evitată, cele două soluţii posibile fiind evitarea excesului de substrat, lucrîndu-se cu cultura in „pat" şi, respectiv, plasarea unui detector de alcool în efluentul de gaz (CO2), care permite să se cunoască aportul de substrat ce trebuie furnizat.
A doua situaţie se întîlneşte, de exemplu, la producţia de culturi lactice, a căror dezvoltare este însoţită de formarea de acid lactic (bacterii homofermentative) sau alcool etilic/acid lactic (bacterii heterofermentative). Pentru a obţine celule de bacterii în cantitate de 60 g/1, este necesară o dializă a culturii, tehnică ce se poate aplica şi la fabricarea drojdiei de panificaţie, în care caz celulele de drojdie se separă prin centrifugare iar „laptele" de drojdie este reciclat. In acest fel se obţin 130 g drojdie/1.
28

Sistemele de fermentaţie continuă se pot clasifica după natura biocatalizatorului (imobilizat sau nu, sistem eterogen sau nu, celule incluse, adsorbite, grefate prin covalenţă), gradul de reciclare al celulelor, gradul de amestecare a fluidelor reacţionabile în reactorul continuu .
Producţia de enzime. Microorganismele produc o serie de enzime folositoare pentru industria alimentară şi alte industrii, constituind deci surse de enzime. Producţia de enzime va depinde de tipul de microorganism folosit şi de condiţiile de dezvoltare: compoziţia chimică a mediului de cultură, pH, temperatură, adaosul de substanţe de inducere sau scăderea cantităţii de substanţe represoare în mediu.
Enzimele produse de microorganisme sunt utilizate în industria alimentară pentru bioconversia unor substanţe în altele, dar şi în alte scopuri (îmbunătăţirea calităţii produselor alimentare, creşterea capacităţii de conservare, creşterea digestibilităţii prin hidroliza enzimatică, îndepărtarea compuşilor nedoriţi din materiile prime sau produse alimentare, îmbunătăţirea tehnologiilor de prelucrare a produselor alimentare în sensul simplificării, scurtării procesului tehnologic şi micşorarea costurilor energetice etc
Microorganismele intervin însă şi în sens negativ: datorită capacităţii lor patogene manifestată prin însuşirea de a pătrunde în organismul viu, se înmulţesc în acest organism, secretă toxine şi anihilază sistemele de apărare ale organismului viu. Nu trebuie neglijată nici capacitatea unor micro-organisme de a elibera toxine (exotoxine) în mediul în care se dezvoltă, inclusiv în produse alimentare - prin al căror consum se produc toxiinfecţii alimentare.
1. BACTERIILE
Dintre bacterii, cele mai importante, care se utilizează în industria alimentară, aparţin următoarelor genuri:
1.1. Genul Streptococcus
Gilliland împarte streptococii în patru grupe: Pyogenic (Str. aglactiae, Str. pyogenes), Viridans (Str. salivarum, Str. bovis, Str. thermophilus), Enterococous (Str. faecalis, Str. faecium, Str. durans), Lactic (Str. lactis, Str. cremons, Str. diacetilactis).
Alţi cercetători consideră că grupul streptococilor lactici cuprinde doar specia Str. lactis cu 3 subspecii: Str. lactis subspecia lactis, Str. lactis, subspecia cremoris, numit uzual Str. cremoris şi Str. lactis, subspecia diacetilactis (Str. diacetilactis).
Streptococii lactici fermentează lactoza, zaharoza, maltoza la acid lactic, dezvoltându-se bine pe un substrat proteic nutritiv care trebuie să conţină peptide, aminoacizi şi vitamine din grupul B. Toate speciile sunt homo-fermentative. Streptococii lactici se grupează în diplo sau sub formă de lanţuri, mai lungi la dezvoltarea în bulion şi mai scurte la dezvoltarea în lapte. Habitatul obişnuit îl constituie laptele şi produsele lactate.
Str. lactis şi Str. diacetilactis se izolează din laptele crud, iar Str. cremoris din produsele lactate acide. Str. thermophilus, inclus în grupa streptococilor Viridans de către Gilliland, este totuşi cooptat în grupa streptococilor lactici, deşi nu se încadrează din punct de vedere serologic. Acesta este termofil (temperatura optimă de dezvoltare 37 ... 45°C), fapt care pledează pentru originea sa intestinală. Se poate dezvolta pînă la 50°C, dar nu şi la 20°C. Este sensibil la o concentraţie de 2% NaCl în mediu, fermentează un număr redus de zaharuri (zaharoza, lactoza, glucoza). Nu fermentează maltoza. Pentru dezvoltare are nevoie de vitamine din grupul B dar nu şi de purine şi pirimidine.
Metabolismul glucidelor cu producere de acid lactic ca produs final principal. Streptococii lactici fermentează lactoza prin transformare mai întîi în glucoza şi galactoză-6-P. Streptococii lactici deşi sunt homofermentativi, în condiţiile în care glucidele din substrat sunt limitate, formează produşi finali de heterofermentaţie.
Activitatea proteolitică. Activitatea proteolitică a streptococilor este apreciată ca fiind mai slabă decît a lactobacililor, fapt constatat prin determinarea azotului neproteic în laptele degresat inoculat cu Str. lactis. Se consideră că diferitele tulpini de streptococi degradează foarte puţin (sau chiar de loc) cazeina, deşi anumiţi streptococi (Str. cremoris) produc atît peptidaze extra-celulare cât şi intracelulare.
Enterococii. Grupul enterococilor cuprinde speciile Streptococus faecalis, Streptocccus faecium şi Streptococcus durans. Se dezvoltă la 10°C dar şi la 45°C. Str. durans şi Str. faecium se găsesc în
29

brînzeturile fabricate din lapte crud, ultimul posedând şi o bună activitate acidifiantă. Prin proteoliza cazeinei stimulează dezvoltarea bacteriilor din genul Leuconostoc. Str. faecium şi Str. durans se utilizează la fabricarea brînzei Cheddar în S.U.A. în scopul accelerării maturării.
1.2. Genul LeuconostocAcest gen cuprinde bacterii cu formă sferică, lenticulară, grupate în perechi sau lanţuri, imobile,
asporogene, gram negative, facultativ anaerobe, catalazo-negative, heterofermentative. Au nevoie pentru dezvoltare de vitamine (acid nicotinic, acid pantotenic, tiamină, biotină) şi zaharuri fermentescibile. Specii de leuconostoc formează polimeri glucidici (dextran). Nu sînt proteolitici, nu reduc azotaţii. Se dezvoltă bine la temperaturi cuprinse între 20 şi 30°C.
Genul Leuconostoc cuprinde speciile: Leuconostoc lactis, Leuconostoc paramezenteroides şi Leuconostoc mezenteroides, acesta din urmă cuprinzînd subspeciile Leuconostoc citrovonim (Leuconostoc cremoris) şi Leconostoc dextranicum. Intre specii există diferenţe în ceea ce priveşte zaharurile fermentate, datorită prezenţei sau absenţei unor enzime implicate în metabolismul gluci-dic. Leuconostoc citrovonim fermentează lactoza, galactoza şi glucoza.
Cu excepţia lui Leuconostoc lactis, cele mai multe specii se dezvoltă greu în laptele fără adaos de substanţe stimulatoare Intr-un mediu nutritiv complex, mulţi leuconostoci se dezvoltă bine la o incubare de 10—12 ore, la 30°C. Leuconostoc citrovorum constituind o excepţie, deoarece multe tulpini necesită 2—3 zile de incubare la 30°C. Adăugarea de cisteină în mediu îi intensifică dezvoltarea. Toţi leuconostocii au nevoie pentru creştere de valină şi glutamat, mulţi au nevoie de alanină, metionina fiind stimulatorie pentru alte multe specii.
Metabolismul glucidelor. Fermentarea glucidelor de către leuconostoci se face pe calea fosfocetohexozelor, dintr-un mol de hexoză, rezultînd cîte un mol de lactat, alcool etilic şi CO2.
Metabolismul citratului. Metabolismul citratului la Leuconostoc este acelaşi ca şi la Str. diacetilactis, cu menţiunea că leuconostocii produc acetoină şi diacetil la pH scăzut. Calea de metabolizare a citratului este arătată în figura urmatore. Acetoina se poate forma pe două căi: prin decarboxilarea acetolactatului şi din diacetil prin intermediul diacetilreductazei. Această ultimă cale este redusă sau lipseşte la Str. diacetilactis, din cauza capacităţii limitate a acestuia de a produce acetil-CoA, unul din precursorii diacetilului.
Leuconostocii se utilizează la fermentarea unor produse vegetale (varză, castraveţi măsline), intervenind în cadrul microflorei spontane sau ca culturi starter, la fermentarea malo-lactică a vinurilor, pentru producerea de dextran etc. O utilizare mai mare a leuconostocilor are loc in industria laptelui pentru fabricarea unor produse lactate fermentate (lapte acru, smântână fermentată pentru consum sau destinată fabricării untului, brînzeturilor).
In acest caz se folosesc culturi lactice care pe lîngă streptococi acidifianţi (St. lactis, Str. cremoris) conţin şi bacterii producătoare de aromă şi anume specii de Leuconostoc, inclusiv Str. diacetilactis care este deopotrivă aromatizant şi acidifiant.
Bacteriile aromatizante îndeplinesc două funcţii majore: formează compuşi de aromă (diacetil, acetoina, acetat etc.) şi produc CO2 care determină apariţia aşa-numitelor „ochiuri" în unele tipuri de brînzeturi (Edam, Gouda, Tilsit etc.). Ambele funcţii sunt consecinţa metabolizării citratului însă sunt specii de Leuconostoc care produc CO2 şi din lactoză. Leuconostocii ca şi lactobacilii, pot produce uneori defecte la brînzeturi: crăparea pastei la brînza Cheddar (defect numit „Blit openness"), apariţia timpurie de gaze la brînza Gouda. Str. diacetilactis poate produce defectul de „plutire a coagulului" la brânza cottage. Toate aceste defecte se datoresc formării de CO2. Pentru a preveni defectul de „plutire a coagulului", brânza cottage se fabrică cu o cultură care nu conţine bacterii aromatizante, adăugîndu-se apoi, în brînza obţinută, o cultură de Leuconostoc citrovorum cultivată pe lapte cu adaos de citrat, sau o cultură concentrată de Leuconostoc citrovorum.
1.3. Genul Pediococcus Aparţine familiei Streptococaceae şi cuprinde bacterii sub formă de coci perechi sau tetrade, ca rezultat al diviziunii alternative pe cele două planuri perpendiculare. Sunt imobile, asporogene. Metabolismul lor este predominant fermentativ, homolactic. Se produce acid lactic racemic (DL) din glucoza, fructoză şi manoză, sorbitolul şi amidonul nefiind fermentate. Nu lichefiază gelatina şi nu reduc azotaţii la azotiţi. Sunt anaerobi — microaerofili şi au necesităţi nutritive complexe. Multe specii sunt catalazo-negative.
30

P. acidilacti este utilizat pentru produsele de carne fermentate la temperaturi mai ridicate, deoarece are dezvoltare bună la 40 ... 52°C, producînd rapid acid lactic şi deci scade efectiv pH-ul, produsul obţinut avînd gust acrişor. Atunci cînd se utilizează la fermentarea unor produse ce se menţin la temperaturi mai scăzute (15 ... 27°C), producţia de acid lactic este mai lentă.
Avînd în vedere că pediococii produc acid lactic, antibiotice şi/sau bacteriocine, ei exercită şi o acţiune inhibitoare faţă de microorganismele patogene şi cele de alterare.
1.4. Genul Lactobacillus Aparţine familiei Lactobacilaceae conform manualului lui Bergey. Această familie cuprinde
bacterii sub forma de bastonaşe de lungimi şi grosimi variabile precum şi cocobacili scurţi, aşezaţi obişnuit în lanţuri în faza de înmulţire logaritmică. Sunt asporogene, imobile, gram pozitive, ana-erobe sau facultativ anaerobe. În general sunt catalazo-negative, citocrom oxidazonegative, nu reduc azotaţii, nu lichefiază gelatina. Au activitate proteolitică şi lipolitică redusă. Glucidele cele mai bine fermentate sînt lactoza, maltoza, zaharoza (mai ales în faza de dezvoltare), apoi hexozele (glucoza fructoza, galactoza). Pentru dezvoltare necesită substanţe minerale şi toate vitaminele din grupul B. Se dezvoltă bine în medii cu pH = 5,5 — 5,8 dar şi la pH < 5. Se pot dezvolta în limite largi de temperatură (5 ... 53°C), dar temperatura optimă este cuprinsă între 30 şi 45°C. În funcţie de temperatura optimă de dezvoltare, lactobacilii pot fi termofili (L. lactis, L. helveticius, L. acidophilus), temperatura optimă fiind 37...45°C şi mezofili (L. casei, L. plantarum), temperatura optimă fiind 28...30°C. Ambele grupe de lactobacili sînt homofermentative.
Unii autori au clasificat streptobacteriile în acidorezistente şi neacidorezistente. Lactobacilii neacidorezistenţi produc compuşi aromatici (diacetil, acetonă), ceea ce înseamnă metabolizarea diversă a acidului piruvic. Produc de două ori mai mult diacetil şi acetoină decît cei acidorezistenţi. Se prezintă sub formă de bastonaşe cu lungime variabilă. Rar formează lanţuri. Se pot dezvolta la 2...15°C. Nu se dezvoltă la pH ≤ 5,6. Se utilizează la fermentaţia salamurilor crude cu pH = 5,9—6,1, produsele obţinute avînd durata mare de maturare, iar gustul final este dulceag. Streptobacteriile acidorezistente au forma de bastonaşe care formează lanţuri. Se dezvoltă bine la pH < 5 şi din această cauză se utilizează la fabricarea salamurilor crude cu maturare scurtă.
Metabolismul lactozei. Lactobacilii hidrolizează lactoza cu ajutorul -galactozidazei şi fosfo--galactozidazei . Dacă Lactobacillus casei foloseşte pentru hidroliză lactozei fosfo--galactozidaza, majoritatea lactobacililor (Lactobacillus bulgaricus, lactis, helveticus) folosesc mai mult -galacto-zidaza. Această enzimă, în cazul lui Lactobacillus bulgaricus, are optimul de pH l= 7 şi este activată de Mg, Mn, Fe. In cele mai multe cazuri, lactobacilii preferă glucoza ca sursă de energie în comparaţie cu lactoza sau galactoza. Unii lactobacili eliberează din celulă galactoză obţinută din lactoza, ceea ce conduce la acumularea galactozei în mediul de cultură. Pentru a fi fermentată galactoză este transformată în glucozo-6-P iar aceasta este metabolizată în continuare pe cale homo- sau heterofermentativă.
Activitatea proteolitică a lactobacililor. Deşi lactobacilii nu sînt consideraţi „proteolitici", ei îşi pot obţine azotul necesar dezvoltării din proteinele laptelui. Se consideră că activitatea endopeptidazică a lactobacililor este asociată cu peretele celular iar activitatea exopeptidazica este localizată intracelular. Enzimele de suprafaţă produc hidroliză parţială a moleculelor mari de proteine din care se formează peptide suficient de mici pentru a putea fi transportate în interiorul celulei unde exopeptidazele continuă degradarea lor pînă la aminoacizii necesari creşterii. Activitatea proteolitică a lactobacililor este optimă la 45...50°C şi la pH = 5,2—5,8. Enzimele sînt complet inactivate prin încălzirea culturii la 70°C, timp de l min.
In industria laptelui se utilizează Lactobacillus bulgaricus şi Lactobacillus lactis, singuri sau în amestec, la fabricarea iaurtului, chefirului, brînzei Emmenthal, brînzeturilor italiene. Lactobacillus helveticus este şi el implicat în unele din aceste produse. Lactobacillus acidophilus este folosit la fabricarea laptelui acidofil, laptelui acidofil nefermentat şi a altor produse acidofile. Lactobacillus casei se utilizează pentru obţinerea produsului japonez denumit yakult.
Pentru industria cărnii interesează L. sake, L. curvatus dar în special L plantantm care se diferenţiază de celelalte specii ale genului Lactobactllus (29 specii), prin faptul că nu produce CO2 la fermentarea glucozei dar produce CO2 din gluconat.
Alte utilizări. Lactobacilii interesează şi în fermentarea măslinelor, în fermentaţia produselor vegetale (varză, castraveţi), în producţia spirtului si a drojdiei presate în vederea acidificării plămezilor, la fabricarea unor produse de cereale (braga, cvas), la fabricarea acidului lactic prin fermentare.
31

1.5. Genurile Micrococcus şi StaphylococcusAparţin familiei Micrococcaceae care cuprinde bacterii sub formă de coci cu Ø = 0,3—0,5 ., cu
diviziune în mai multe planuri, formînd grămezi sau pachete. Pot fi mobile sau imobile, asporogene, gram-pozitive, cu metabolism respirator sau fermentativ, catalazo-pozitive, aerobe sau facultativ anaerobe. Se pot dezvolta în medii conţinînd 15% NaCl. Produc acizi din glucoza fără a produce gaze. Pentru industria cărnii interesează anumite specii de micrococi şi stafilococi care sînt folosite pentru capacitatea lor de a reduce azotaţii şi azotiţii (contribuie la formarea culorii), pentru activitatea lor catalazică, de acidificare, proteolitică şi lipolitică. Dintre speciile de micrococi, mai des utilizate sînt Micrococcus aurantiacus şi Micrococcus varians.
În industria laptelui, micrococii formează partea principală a populaţiei nelactice din lapte şi brînzeturi, tulpini aparţinînd genului Micrococcus utilizîndu-se pentru maturarea laptelui destinat brînzeturilor. Micrococcus freudenreichii, izolat din brinza Cheddar fabricată din lapte crud, a fost ulterior folosit în scopul accelerării formării aromei brînzei fabricate din laptele pasteurizat, datorită activităţii proteolitice şi lipolitice. In scopul accelerării maturării unor brinzeturi cu pastă presată s-a încercat adăugarea în lapte a proteazelor produse de micrococi (s-a folosit Rulactina, o metalo-protează cu activitate strict endopeptidazică, produsă de o tulpină de Micrococcus caseolyticus). Combinaţiile de micrococi şi stafilococi sunt eficace pentru activitatea azotat-reductazică şi catalazică. Staphylococcus carnosus acţionează mai bine decît micrococii în formarea culorii cărnii, reducînd azotaţii la azotiţi si respectiv azotiţii la oxid de azot, chiar în condiţii de aciditate ridicată a substratului. Aroma produselor la care este utilizată cultura starter de Staphylococcus carnosus este superioară.
Combinaţia L. pentosus + Staphylococcus carnosus se comportă mai bine decît combinaţia L. plantarum + Staphylococcus carnosus, deoarece L. pentosus produce mai rapid şi mai intens acid lactic în comparaţie cu L. plantarum.
1.6. Genul PropionibacteriumLa fabricarea unor tipuri de brînzeturi (Emmenthal, Swiss, Gruyere) caracterizate prin prezenţa
„ochiurilor" în pastă şi printr-o anumită aromă se folosesc bacterii din genul Propionibacterium, alături de bacterii lactice. Acestea sunt folosite şi pentru producţia de vitamină B12, a acidului propionic, a unor băuturi lactate.Sunt gram-pozitive, nesporulate sub formă de bastonaşe imobile, cu un capăt rotunjit şi celălalt ascuţit şi colorat mai slab. Celulele unor culturi pot fi sub formă de coci, elongate, bifurcate sau chiar ramificate. De obicei, celulele sînt singure, perechi, sau se aranjează în configuraţii V, Y, lanţuri scurte sau îngrămădiri.
2 . DROJDIILE
Drojdiile, ciuperci unicelulare care se înmulţesc prin înmugurire, mai rar prin sciziparitate şi care formează ascospori (sunt şi drojdii care nu fac spori, acestea denumindu-se drojdii false-torule şi micoderme) sunt agenţi tipici ai fermentaţiei alcoolice. Se prezintă sub formă de celule rotunde sau ovoide (Saccharomyces cerevisiae), eliptice (Saccharomyces ellipsoideus), foarte alungite (Saccharomyces pasteurianus), de forma unei lămîi (Saccharomyces apculatus), de forma unei sticle (Saccharomyces ludwigii) sau sub formă de cilindru (Pichia). La unele specii, ca rezultat al procesului de înmugurire, celulele de înmugurire rămân ataşate de celula mamă şi, înmugurind la rândul lor, formează un pseudomiceliu, asemănător în structură cu cea a amilopectinei, componentă a amidonului. Desigur, forma drojdiilor este în funcţie de vârsta şi condiţiile de cultură. Pe medii solide, drojdiile formează colonii caracteristice speciei.
Dintre drojdii interesează (în sens util) cele aparţinînd familiei Saccharomycetaceae, genul Saccharomyces care cuprinde drojdii alcooligene folosite în industria berii, vinului şi pîinii, genul Kluyveromyces care fermentează lactoza (levuri ale laptelui), genul Debaryomices care se utilizează în industria cărnii. Mai interesează drojdiile aparţinînd familiei Criptococcaceae (genurile Candida, Torulopsis) care se folosesc ca agenţi de fermentaţie şi ca producători de biomasă.
Cea mai largă utilizare a drojdiilor este cea de a produce fermentaţie alcoolică. Acestea sînt de fapt drojdiile „adevărate" care aparţin genului Saccharomyces (Meyen) Rees şi care se caracterizează prin
32

aceea că nu formează miceliu tipic, produc l-4 spori, iar puterea de fermentare depăşeşte puterea de respiraţie.Sunt adaptate la condiţii de anaerobioză şi, prin urmare, nu formează voaluri la suprafaţa lichidelor. Acestea sunt de fapt drojdii cultivate, izolate şi selecţionate prin culturi în scopuri utile-industriale şi se deosebesc de cele care şi-au păstrat caracterele primitive, fiind cunoscute sub denumirea de drojdii sălbatice.
Drojdiile, după modul lor de comportare în timpul fermentaţiei, pot fi grupate în drojdii de fermentaţie superioară şi drojdii de fermentaţie inferioară. Cele de fermentaţie superioară se ridică în cantitate mare la suprafaţă în spuma care acoperă lichidul de fermentare, au optimul de temperatură la 28 ... 30°C şi fermentează numai la 1/3 din rafinoză. Drojdiile de fermentaţie inferioară nu se ridică la suprafaţă decît în cantitate mică, au optimul de temperatură la 8 ... 12°C şi fermentează complet rafinoză. Zaharurile fermentate de drojdii sunt triozele (aldehida glicerică şi dioxiacetona), aldohexozele (D-glucoza, D-manoza şi D-galactoza), cetohexozele (D-fructoza). Di- şi trizaharidele sunt fermentate în măsura în care drojdiile respective conţin enzimele necesare transformării acestora în zaharuri simple. Poliglucidele nu sunt transformate de drojdii. Excepţie fac Saccharomyces fragilis care fermentează inulina şi Endomycopsis fbuligerea care fermentează amidonul.
Fermentaţia alcoolică produsă de drojdii este influenţată de concentraţia zaharului fermentescibil în must sau plămadă, temperatură, conţinutul în alcool şi felul drojdiei. Concentraţia favorabilă de zahăr fermentescibil în must sau plămadă este de 10—15%. Fermentaţia alcoolică se desfăşoară bine la pH = 4—4,5, în mediu alcalin sensul fermentaţiei fiind schimbat. Viteza maximă a fermentaţiei este la 30°C, dar în practică aceasta se realizează la temperaturi cuprinse între 4 şi 28°C.
Alcoolul, pe măsura acumulării în mediu, devine toxic pentru drojdii. Există rase de drojdii care se dezvoltă la 16—18% alcool, însă în cele mai multe cazuri fermentaţia se opreşte la 12—14% alcool. O dată cu creşterea temperaturii de fermentare se măreşte toxicitatea alcoolului. Fermentaţia alcoolică este un proces anaerob. Dacă drojdiile se cultivă în aerobioză pe substraturi adecvate, ele se înmulţesc foarte mult şi deci produc biomasă.
Fermentaţia alcoolică nu este o fermentaţie absolut pură. Chiar în cazul unei fermentaţii alcoolice normale, în afară de alcoolul etilic iau naştere glicerina (produs secundar predominant care se formează din ~ 8% din totalul zaharurilor existente in mediu), acid lactic, acid acetic, substanţe acetoinice (acetil metil carbinol = acetoină şi diacetil), acid malic, acid succinic, acid propionic, acid citramalic, acid dimetilgliceric, alcooli superiori: izobutilic, izoamilic, amilic, care provin din aminoacizi rezultaţi la degradarea substanţelor proteice din plămezi şi musturi. Aceşti alcooli superiori se formează prin reacţii de dezaminare şi decarboxilare ale aminoacizilor leucină, izoleucină şi valină.
De remarcat că în funcţie de modul de dirijare al fermentaţiei alcoolice, (pH, substanţe adăugate), fermentaţia alcoolică poate fi deviată substanţial de la cursul „normal". De exemplu, în prezenţă de acid sulfuros sau bisulfit se produce multă glicerina. Acelaşi lucru se întîmplă dacă pH-ul este alcalin. Fermentaţia alcoolică se aplică în mod deosebit la fabricarea alcoolului din produse amidonoase (cartofi, porumb, secară), produse care conţin zaharoză (sfeclă, melasă), lactoza (zer), glucoza (hidrolizate celulozice, leşii sulfitice). Fermentaţia alcoolică stă la baza fabricării vinului şi berii.
Drojdiile sunt folosite pentru obţinerea de biomasă (inclusiv drojdia de panificaţie), pentru producere de glicerina, grăsimi, băuturi fermentate lactate şi pe bază de cereale şi leguminoase, cît şi pentru producţia unor enzime (lactază, invertază).
Dintre drojdii, în industria cărnii se foloseşte Debaromyces hansenii care este tolerantă la clorură de sodiu, nu reduce azotatul şi necesită oxigen atmosferic pentru multiplicare. Se dezvoltă bine în straturile periferice ale salamului neafumat sau puţin afumat. Are capacitatea de a consuma oxigenul din pastă şi de a distruge peroxizii formaţi de unele bacterii lactice.
In cazul brînzeturilor, drojdiile se dezvoltă rapid la suprafaţa brînzeturilor şi în interiorul brînzeturilor cu pasta moale sau cu mucegai în pastă, în timpul presării, zvântării şi maturării, avînd următoarele acţiuni pozitive:— consumă lactatul şi în acest fel contribuie la neutralizarea pastei, îmbunătăţind prin aceasta consistenţa şi favorizînd implantarea bacteriilor;— produc factori de creştere pentru dezvoltarea bacteriilor ;— produc o „acoperire" la anumite tipuri de brînzeturi (Roquefort);— produc proteoliză şi lipoliză şi prin urmare contribuie la consistenţa şi aroma brînzei ;— produc compuşi volatili de aromă.
33

Dintre drojdii, Candida kansei a fost cultivată în asociaţie cu lactobacili şi Str. thermophilus, favorizînd prin oxidarea lactatului scăderea potenţialului de oxidoreducere şi producerea de factori de creştere, dezvoltarea culturilor starter în care este asociată. Drojdia Candida lipolitică este uneori folosită la fabricarea brînzeturilor cu mucegai în pastă, datorită faptului că prin lipazele conţinute realizează o hidroliză a grăsimilor din brînză, fapt ce favorizează utilizarea acestora de către Penicillium.
3. MUCEGAIURILE
Mucegaiurile sînt cuprinse în patru clase mari din care, pentru industria alimentară, interesează Phycmycetes. Din clasa Phycomycetes, interesează (în sens pozitiv), genurile Mucor şi Rhizopus, aparţinînd familiei Mucoraceae. Aceste mucegaiuri care se întîlnesc pe diferite produse vegetale şi alimentare, sub formă de colonii pufoase, au o acţiune fcrmentativă netă. In afară de faptul că unele specii de Mucor şi Rhizopus se folosesc la obţinerea unor produse alimentare fermentate, mucegaiurile aparţinînd acestor genuri sunt surse importante de enzime, în principal, amilolitice şi proteolitice.
Din clasa Ascomycetes genurile Aspergillus şi Penicillium. Dacă mucegaiurile din genul Aspergillus se utilizează mai puţin la fabricarea unor produse alimentare (de exemplu şuncă S.U.A. şi Spania), obţinerea de produse fermentate tradiţionale din Orientul îndepărtat, aceste mucegaiuri au o largă utilizare în obţinerea de preparate enzimate amilazice, proteazice, pectolitice. Mucegaiurile din genul Penicillium sunt mult utilizate în industria laptelui (brînzeturilor), cărnii (salamurilor crude), dar şi ca surse de enzime amilolitice, proteolitice şi pectolitice.
Genul Mucor cuprinde specii cu aparatul vegetativ dintr-o singură celulă puternic ramificată, cu numeroase nuclee. Majoritatea speciilor sunt saprofite, preferând însă medii uşor acide, bogate în zahăr. Se înmulţesc pe două căi: asexuat şi sexuat. In primul caz sporii se formează în sporangi rotunzi sau piriformi lipsiţi de apofize, aflaţi la extremitatea unor prelungiri simple sau ramificate numite sporangiofori. Genul Rhizopus se deosebeşte de Mucor prin faptul că sporangioforii simpli — neramificaţi, pornesc mai mulţi din acelaşi loc iar sporangii au apofize.
Genul Aspergillus cuprinde specii pluricelulare care produc pe suprafaţa substraturilor pâsle colorate. Speciile acestui gen au conidiofori unicelulari cu extremitatea aproape sferică, pe suprafaţa cărora se află prelungiri — sterigme ce produc şiraguri de conidii.
Genul Penicillium se deosebeşte de Aspergillus prin conidiofori pluricelulari ramificaţi în formă de penson.
În industria cărnii, culturile starter de spori de mucegai se utilizează la maturarea unor tipuri de şuncă şi a unor tipuri de salamuri crude fabricate în România, Italia, Ungaria, Elveţia, Spania, Franţa, Bulgaria, Austria, Belgia, R.F.G. (şi mai puţin în S.U.A., Israel, Iugoslavia, Polonia) In cazul salamurilor crude afumate sau neafumate, mucegaiurile de acoperire contribuie la reglarea eliminării apei din produs şi a schimbului de gaze, la formarea aromei şi la aspectul comercial al produsului. Aroma este, în general, mai evidentă la salamurile cu diametru mai mic. Produsele pot fi livrate cu mucegaiul de acoperire intact sau după „periere". Culoarea miceliului rămas este dependentă de varietatea mucegaiului folosit, în Italia se preferă o culoare alb-ivorie, în Ungaria o culoare gri, în România o culoare alb-mat. Prin utilizarea culturilor starter de spori de mucegai, se suprimă apariţia pe salamuri a mucegaiurilor toxicogene , precum şi a mucegaiurilor de „pătare" (care produc spoturi de culoare verde sau neagră), în special a mucegaiului Penicillium stoloniferum .
3.1. Penicillium nalgiovensisTulpina selecţionată de Penicillium nalgiovensis este cea mai frecvent utilizată pentru
însămînţarea de suprafaţă a salamurilor crude, deoarece acoperă în mod uniform si rapid produsul cu miceliu de culoare albă din care se dezvoltă condiofori purtători de conidii (spori). Acoperirea cu miceliu a produsului are loc după 3—4 zile de la însămînţare cu spori, iar corpii de fructificaţie cu condii apar după 5—6 zile de la însămînţare. Temperatura optimă de dezvoltare a acestui mucegai este de 22 ... 23°C.
3.2. Penicillium expansumPenicillium expansum se dezvoltă bine la 22°C, umiditatea relativă pentru sporulare fiind de 82%, pentru germinare 88%, iar pentru dezvoltare 92—95%. După însămînţarea batoanelor cu spori, produsul depozitat în încăperi cu ~ 90% umiditate se acoperă cu un strat compact şi pufos de miceliu, după
34

aproximativ 8 zile; maturizarea deplină a mucegaiului (produs gata pentru periere) are loc după 30 zile de la însămînţare. Penicillium expansum nu produce micotoxine, dacă substratul conţine proteine cu sulf (cazul pastei salamurilor crude).In industria laptelui culturile fungice sunt necesare atât în fabricarea brînzeturilor cu mucegai în pastă cât şi cu mucegai la suprafaţă. Pentru brînzeturile cu mucegai în pastă (brînza Roquefort, Gorgonzola, Stilton, Gammelost etc.) se utilizează tulpini de Penicillium roqueforti. Pentru brînza de tip Camembert, cu mucegai la suprafaţă, se folosesc două specii de Penicillium: P. camemberti şi P. caseicolum. Aceste mucegaiuri au un rol fundamental în formarea aromei, consistenţei şi aspectului brînzeturilor respective, prin procesele de proteoliză şi lipoliză pe care le realizează.
3.3. Penicillium roqueforti ThomAcest mucegai creşte în colonii cu aspect pufos, catifelat, azonate, în general subţiri, cu
conidiofori scurţi şi abundenţi, cu marginea cu aspect arhnoid, stelat sau ca o reţea. Conidiile sunt lungi, de culoare galben-verzui închis, adesea verzui, până aproape de negru pe spatele coloniei. Tolerează cantităţi mari de NaCl (5—8%) în mediu acid şi se poate dezvolta într-o atmosferă conţinînd doar 5% oxigen. Se dezvoltă bine la temperaturi de 20 ... 25°C (dar nu şi la temperaturi > 35°C), pe medii cu umiditate ridicată. Dezvoltarea abundentă a miceliului are loc la pH = 4,5—7,5. Mucegaiul poate folosi sulfatul de amoniu ca singura sursă de azot. Nu utilizează aminoacizii leucină, cisteină şi metionină. Produce enzime proteolitice şi lipolitice, substratul avînd o deosebită influenţă asupra activităţii enzimatice. Prin cultivarea pe medii îmbogăţite în grăsime se obţine o cultură cu puternică activitate proteolitică şi lipolitică, acizii graşi eliberaţi fiind transformaţi în compuşi carbonilici şi în special metilcetonele rezultate prin -oxidarea acizilor graşi. Datorită echipamentului enzimatic proteolitic, conţinutul în aminoacizi liberi din brînzeturi creşte simţitor.
P. roqueforti se dezvoltă în canalele şi golurile practicate în pasta brînzeturilor moi, producînd spori de culoare verde-închis şi astfel conferă brînzei respective aspect marmorat.
Cercetările efectuate la Universitatea din Wisconsin, asupra toxinogenezei lui Penicillium roqueforti, au pus în evidenţă faptul că acest mucegai produce PR-toxina, nocivă pentru şobolani la care induce modificări histologice la nivelul ficatului, rinichiului precum şi hemoragii, fenomene tremorgene, avorturi şi retenţia placentei la mamifere.
Pe lingă PR-toxină, în miceliul de Penicilliiim roqueforti, s-au pus în evidenţă şi doi alcaloizi asemănători cu cei produşi de către ergot: roquefortina şi izofumigaclavina. Aceşti doi alcaloizi au fost puşi în evidenţă şi în brînzeturile fabricate cu Penicillium roqueforti, în proporţie de 0,66—6,8 mg/kg produs. Ambele substanţe posedă acţiune neurotoxică. Roquefortina are DL50 = 15—20 mg/kilocorp la şoareci (administrare prin injectare intraperitoneală).
3.4. Penicillium camemberti şi Penicillium caseicolum
Aceste două specii de mucegai sunt strâns înrudite şi sunt utilizate în producţia brînzeturilor de tip Camembert, incluzînd Camembert, Brie, Neufchâtel, Coulommier, Garre de l'Est, Olivet.
Tulpinile au fost izolate de pe brînzeturi sau din camerele (pivniţele) de maturare a brînzeturilor.Deşi strâns înrudite, cele două specii P. camemberti Thom şi P. caseicolum Bainer se deosebesc între ele prin caracteristicile fiziologice şi biochimice.
Penicillium camemberti Thom denumit şi Penicillium album se utilizează pentru obţinerea de brînză tip Camembert cu textură mai untoasă, mai parfumată, dar de culoare gri-alburie.
Penicillium caseicolum Bainer, denumit şi Penicillium candidum, se utilizează pentru obţinerea de brînză tip Camembert cu o textură mai compactă, aromă mai delicată şi o culoare alb-imaculată.
CULTURI STARTER DE MICROORGANISME
DEFINITIE Culturile starter sunt definite ca acele culturi care se obţin plecând de la cultură pură stoc şi care
prin trecere prin culturi intermediare devin apte de a fi folosite pentru producţia unor alimente
35
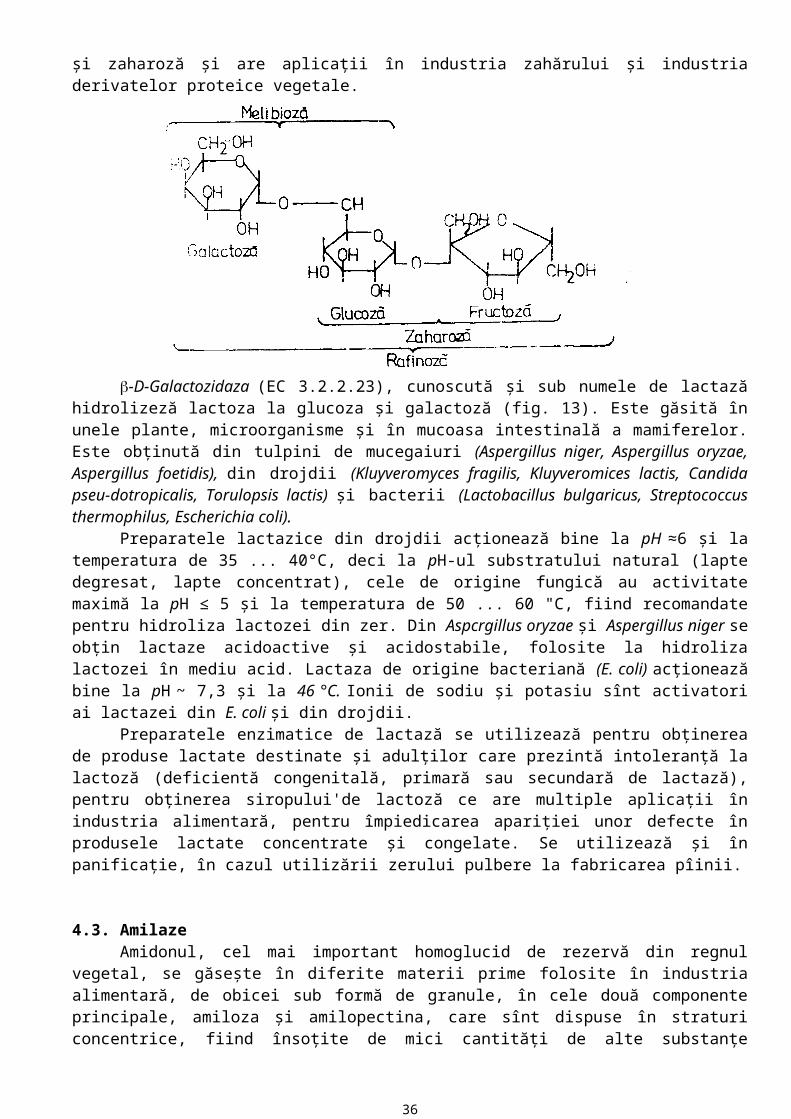
fermentate.Culturile starter pot fi formate dintru-un singur microorganism sau din mai multe microorganisme.Culturile starter sunt utilizate în vederea: dirijării unor procese biochimice prin care se asigură produsului un anumit grad de inocuitate(inclusive capacitatea de conservare) asigurării unor însuşiri senzoriale asigurării unor însuşiri nutritiveLa folosirea culturilor starter în industria alimentară trebuie să se aibă în vedere următoarele : să aibă o anumită concentraţie de microorganisme pe unitatea de produs şi un număr cât mai redus de germeni nedoriţi. produşii de metabolism primari şi secundari să nu prezinte pericol pentru sănătatea oamenilor. să nu conţină şi să nu producă antibiotice care se utilizează în scop terapeutic la om. să aibă anumite activităţi specifice:producerea alcoolului, acidului lactic, reducerea azotului. microorganismele existente să fie declarate cu numele ştiinţific întreg speciile(suşele) noi, care se introduce în producţie, trebuie să fie înregistrate la Ministerul Sănătăţii şi să fie depozitate în colecţii cu nomenclator; înainte de introducerea în producţie să fie testate din punct de vedere al inocuităţii, în conformitate cu legislaţia în vigoare. speciile care pe baza noilor cunoştinţe ştiinţifice au fost recunoscute ca având potenţial patogen sau toxicogen, trebuie să fie supuse unui control riguros pentru fiecare suşă, realizându-se studii de de toxicitate pe termen lung, de carcinogenitate şi mutagenitate.Toate cele menţionate trebuie să constituie ca o obligativitate absolută , deoarece:
culturile starter se pot consuma odată cu produsul alimentar, în stare vie cum este cazul produselor lactate acide , brânzeturilor şi salamurilor crude , unele sortimente de bere, unele produse vegetale fermentate(varză murată, castraveţi muraţi , măsline);
culturile starter se pot consuma după distrugerea lor, rămânâmd în produsul alimentar atât ele cât şi produşii lor de metabolism , aşa cum este cazul brânzei proaspete de vaci şi a iaurturilor care au fost pasteurizate, produsele de panificaţie;
produsele de metabolism se consumă odată cu produsele alimentare , însă microorganismele sunt eliminate în cea mai mare parte, aşa cum este cazul berii , vinului, alcoolului alimentar , oţetului;
1. CULTURI STARTER DE BACTERII LACTICE Culturile starter de bacterii lactice sunt folosite în diferite domenii: industria laptelui, industria
cărnii, a produselor vegetale murate, a vinului,a panificaţiei,a sucurilor de fructe şi legume fermentate.
Folosirea culturilor starter de bacterii lactice asigură produselor alimentare în care se introduce un anumit grad de inocuitate , deoarece bacteriile lactice produc:
acizi organici , în special acid lactic, acid acetic, alcool, CO2
substanţe bacteriocine eliberate în mediul de cultură peroxizi ( H 2O2)
In plus, bacteriile lactice intră în competiţie cu bacteriile patogene şi cu cele de alterare în ceea ce priveşte consumul de substanţe nutritive, iar datorită acidifierii mediului, consecinţă a acumulării acizilor organici, bacteriile patogene şi de alterare sunt inhibate în dezvoltarea lor(stafilococi,salmonele, etc.). Datorită acidităţii se inhibă şi dezvoltarea microflorei cu activitate proteolitică şi decarboxilazică, deci se formează cantităţi mai mici de amine biogene, iar în cazul cărnurilor sărate mai puţine nitrozamine.
1.1CULTURI STARTER UTILIZATE IN INDUSTRIA LAPTELUI
La obţinerea culturilor starter trebuie să se aibă în vedere , în mod deosebit următoarele :- mediul de cultură - tratamentul termic aplicat laptelui
36

- condiţiile de incubare- interacţiunile dintre speciile din cultura starter- eventualele infectari cu bacteriofagi- instabilitatea culturilor starter de bacterii lactice
1.1.1 LAPTELE MEDIU DE CULTURA PENTRU CULTURI STARTERIn ceea ce priveşte mediul de cultură –laptele, se menţionează faptul că,acesta nu este un mediu ideal pentru dezvoltarea bacteriilor lactice din două motive:
a) Laptele crud nu are un conţinut optim de substanţe nutritive azotate, vitamine şi alţi factori de creştere, indispensabili bacteriilor lactice .
Există diferenţe genetice între specii şi suşe în ceea ce priveşte utilizarea lactozei şi galactozei. Astfel, culturile starter de iaurt (Lactobacillus bulgaricus şi Str.thermophilus) produc mai rapid acid lactic, dacă lactoza a fost în prealabil hidrolizată cu β-galactozidază. Din lapte, Lb.bulgaricus foloseşte numai glucoza. De asemenea, azotul neproteic din lapte nu este satisfăcător cantitativ pentru bacteriile lactice care trebuie să posede activitate proteolitică pentru a hidroliza proteinele în scopul eliberării aminoacizilor necesari pentru nutriţia azotată. In această direcţie există diferenţe între specii şi tulpini. Tulpinile lente utilizează azotul neproteic, dar se dezvoltă greu.Tulpinile rapide, utilizează peptidele hidrolizate de ele, deoarece posedă echipament proteolitic puternic. In fapt, diferenţele dintre tulpinile lente şi rapide, constau în absenţa şi, respective prezenţa unor plasmide care conţin gene ce codifică sinteza de peptihidroloze (proteinaze şi peptidase).Tulpinile lente au pierdut aceste plasmide când celulele s-au divizat în cursul dezvoltării, iar cele rapide le-au păstrat.Această situaţie poate apărea şi în cazul utilizării lactozei. Tipul de proteină din lapte, are influenţă, de asemenea, asupra dezvoltării bacteriilor lactice. Astfel, Lb.bulgaricus utilizează preferenţial β-cazeina ca sursă de azot, în timp ce leuconostocii prefera azotul dintru-un extract de drojdie. Tipul de aminoacizi folosiţi este diferit în funcţie de specie. Streptococii folosesc valina, glicina, histidina, izoleucina, în timp ce leuconostocii preferă valina şi glutamatul. In general, laptele conţine 20% din totalul aminoacizilor şi peptidelor necesare dezvoltării optime a leuconostocilor şi streptococilor. Adaosul de substanţe stimulatoare sub formă de extract de drojdie, extract din pancreas, sirop de porumb, suc de tomate, are drept scop îmbogăţirea mediului de cultură, laptele, cu factori stimulatori: aminoacizi, vitamine, substanţe minerale sau enzime proteolitice necesare hidrolizei iniţiale a proteinelor din lapte. In cazul bacteriilor lactice de aromatizare este necesară prezenţa în mediul de cultură a citratului şi a ionilor de Mg2+ şi Mn2+.
b) Laptele crud conţine o serie de inhibitori naturali, cum ar fi lactotransferina, care exercită actiune bacteriostatică asupra tulpinilor care necesită prezenţa fierului în mediul de reacţie, lactoperoxidaza care manifestă actiune bacteriostatică faţa de strptococi, lizozimul care are acţiune bactericidă faţa de toate bateriile lactice.
1.1.2 TIPURI DE CULTURI STARTER UTILIZATE IN INDUSTRIA LAPTELUI
Culturile starter utilizate în industria laptelui pot fi clasificate în: mezofile şi termofile.Culturile starter mezofile pot fi singulare, multiple şi mixte.Culturile starter singulare (single strain starter) conţin numai Str.lactis subspecia lactis ,
respective Str.lactis subspecia cremoris, ambii homofermentativi, care produc acid lactic (L+) în proporţie de 0.8%. Folosirea acestor culturi starter singulare a apărut ca o necesitate de a evita formarea ochiurilor de fermentare la unele brânzeturi.
Culturile starter singulare prezintă următoarele avantaje: Se poate utiliza continuu aceeaşi clutură cu activitate relativ constantă şi previzibilă Nu este necesară alternarea culturilor, eliminându-se riscul deprecierii acestora de bacteriofagi Se folosesc cantităti mici de inocul pentru obţinerea de cultură primară şi secundară Influenţele sezoniere ale compoziţiei laptelui sunt reduse Se creează condiţiile unei producţii standardizate de produse lactate (consistenţa calităţii
produsului)
37

Cultura poate fi controlată şi supravegheată din punct de vedere al caracteristicilor sale Culturile starter mezofile prezintă, următoarele dezavantaje:
Pot fi depreciate de tulpinile producătoare de bacteriocine Pot suferi pierderi de plasmide, deci pot pierde una sau mai multe funcţii metabolice
Culturile starter multiple mezofile sunt acele culturi care se bazează pe folosirea a 5-6 tulpini selecţionate, neînrudite pe plan fagic şi cultivate separat până la stadiul de cultură primară sau până la stadiul de cultură starter de producţie, când se amestecă între ele. In aceste condiţii, tulpinile nu se dezvoltă împreună decât cel mult 10 generaţii, ceea ce face ca nici o tulpină să nu devină predominantă.Culturile starter mezofile mixte sunt formate, de regulă, din două tipuri de bacterii lactice:
Bacterii lactice acidifiante: Str.lactis sau Str. Cremoris Bacterii lactice producătoare de aromă: Str.diacetilactis sau specii de leuconostoci.
Pentru a avea o aromă corespunzătore trebuie să se aibă în vedere următoarele:- temperatura optima pentru leuconostoci este de 24-27oC , cu o durată a unei generaţii de 3.2 ore la 26 oC şi 3.5 ore la 22 oC. Str. cremoris are durata unei generaţii de 1.5 ore la 22 oC, ceea ce înseamnă că există posibilitatea ca streptococii să domine leuconostocii în culturile starter mixte, de unde se impune condiţia să existe un anumit raport între streptococci/leuconostoci.- după producerea diacetilului, acesta se poate transforma în acetoină, respectiv 2,3 butilenglicol cu pierderea semnificativă a aromeiCulturi starter termofile pot fi: acidulante: Lb.acidophilus acidulante –aromatizante, care la rândul lor pot fi constituite din una sau mai multe specii de lactobacili şi dintr-o specie de streptococci. Din această categorie se întâlnesc:-cultură starter termofilă pentru iaurt : Lb. bulgaricus şi Str.thermiphilus-cultură starter termofilă pentru brînzeturi: Lb. bulgaricus +Lb.lactis+Lb.helveticus+ Str.thermophilus.
Utilizarea culturilor starter temofile este benefică deoarece: produc acid lactic, deci scade pH-ul laptelui şi determină coagularea acestuia (iaurt); la brânzeturi acidifierea favorizează eliminarea zerului din coagul au activitate proteolitică şi contribuie la îmbunătăţirea proprietăţilor reologice şi la aroma produselor fermentate; produc aminoacizi prin proteoliză care stimulează dezvoltarea bacteriilor lactice produc compuşi de aromă , aldehidă acetică şi urme de acetoină produc substanţe cu caracter filant care influenţează vâscozitatea produsului(Str.termophilus în cultura pentru iaurt) produc bacteriocine produc apă oxigenată (Lb.bulgaricus)
2. CULTURI STARTER CONCENTRATE DE MICROORGANISME
Culturile starter concentrate sunt definite ca acele culturi dezvoltate în condiţii controlate, concentrate într-un volum mic şi conservate prin congelare sau uscare în vederea depozitării sau transportului.
Culturile concentrate pot fi: bacterii, drojdii, spori de mucegai. In ceea ce priveşte culturile starter concentrate de bacterii, cele mai des utilizate sunt culturile de bacterii lactice.Culturile concentrate de bacterii pot fi utilizate pentru:
obţinerea culturilor starter de producţie obţinerea produselor fermentate prin utilizarea directă în lapte , compoziţie cârnaţi, etc.
2.1 OBTINEREA CULTURILOR STARTER CONCENTRATETehnologia de obţinere a culturilor starter concentrate cuprinde urmatoarele operaţii:
1. inocularea mediului de cultură cu microorganismul din cultura stoc 2. incubare pentru multiplicare3. concentrarea mediului împreună cu celulele sau separarea celulelor, de regulă prin centrifugare şi resuspendare într-un lichid adecvat 4. conservare prin congelare şi uscare 5. depozitare în stare congelată sau uscată
38

In tehnologia de obţinere a culturilor starter concentrate trebuie ales un mediu de cultură care să asigure toate substanţele pentru dezvoltare, realizându-se un control riguros al temperaturii şi menţinerea unui nivel optim de pH.Conservarea prin congelare a concentratului de microorganisme se face în următoarele moduri : sub formă lichidă, în care caz , concentratul de bacterii se suspendă într-un antigel solubil în apă, care se utilizează în proporţie de 40-50% faţa de concentrat. Congelarea se realizează la -40oC(culturi subrăcite). Acest gen de conservare prezintă următoarele avantaje : -se împiedică acţiunea dăunătoare a gheţii asupra celulelor de bacterii -manupularea concentratului este mai uşoară -concentratul se poate încălzii până la temperatura de utilizare congelarea în azot lichid (-196 oC), în care caz concentratul se amestecă cu un concentrat crioprotector: 10% glicerol, 7.5% lactoză (în cazul bacteriilor lactice ) conservarea prin reducerea conţinutului de apă, prin liofilizare, în care caz concentratul de bacterii se amestecă cu un suport adecvat (lapte praf degresat,lactoproteine pulbere, lactoză, zaharoză) după care se liofilizează. La liofilizare temperatura produsului nu trebuie să depăşească 40-45oC. După liofilizare se face ambalarea sub vid sau în atmosferă controlată de gaz inert. Depozitarea produselor liofilizate trebuie să se facă la temperaturi scăzute (- 18oC).Culturile starter concentrate trebuie să conţină 2.5*1010 şi 5.5*1010 celule viabile active /g sau ml produs.
2.2 CULTURI STARTER CONCENTRATE PENTRU INDUSTRIA LAPTELUI Pentru industria laptelui se folosesc, în special culturi concentrate de bacterii lactice şi propionice.
Culturile concentrate de bacterii lactice se folosesc pentru: brânza Cheddar, brânzeturi de tip italian, elveţian brânza de vaci, smântână, lapte bătut, iaurt
Culturile de bacterii concentrate propionice se adaugă direct în laptele destinat fabricării brânzeturilor (nu se mai folosesc culturi intermediare).
2.3 CULTURI STARTER CONCENTRATE PENTRU INDUSTRIA CARNII
Culturile starter concentrate pentru industria cărnii pot fi culturi singulare sau în amestec. Ele pot fi formate din bacterii lactice (Lb.Plantarum, Lb.sake, Lb.alimentarius), pediococi (P.acidilacti , P.pentosaceus), stafilococi(S.carnosus, S.xilosus), micrococi(M.varians), etc.
Culturile starter de micrococi sau stafilococi sunt recomandate la maturarea lentă a salamurilor crude, unde se realizează o scădere lentă a pH-lui la o valoare de 5.6-6.1, conferind produselor un gust acrişor iar realizarea consistenţei se realizează în timp.
Culturile mixte de micrococi-stafilococi-lactobacili se utilizează pentru maturarea rapidă a salamurilor crude, în care caz realizarea consistenţei merge paralel cu acidifierea.
Culturile starter concentrate utilizate în industria cărnii se gasesc sub formă lichidă sau liofilizate.Adaosul în compoziţie trebuie să asigure o concentraţie de cel puţin 106-107 microorganisme/g pastă. Modul de folosire al culturilor depinde de modul lor de conservare. Culturile conservate în antigel sunt folosite ca atare, cele liofilizate se suspendă în apă pentru o distribuţie mai uniformă în compoziţie.
Culturile starter concentrate de bacterii lactice pentru industria cărnii nu se amestecă cu sare ,condimente, acid ascorbic, acizi alimentari deoarece se inactivează rapid.
Culturile starter concentrate folosite în industria cărnii trebuie să îndeplinescă urmatoarele condiţii:
să fie tolerante la NaCl (concentraţie 2.5-3%) să se dezvolte bine în prezenţa de 80-100 mg NaNO3 /kg compoziţie să producă numai acid lactic(specii homofermentive) să fie puţin lipolitice şi proteolitice să nu producă gusturi şi mirosuri neplăcute din cauza produşilor secundari de reacţie să fie nepatogene să fie inactivate la 57-60oC
Folosirea culturilor starter concentrate de bacterii în industria cărnii prezintă urmatoarele avantaje:
39

- micşorează durata de maturare a produselor din carne- se îmbunătăţesc proprietăţile senzoriale(gust, miros, consistenţă)- se asigură un grad înalt de inocuitate produsului alimentar (salamurile şi cârnaţii cruzi)
prin controlul dezvoltării microorganismelor patogene şi de alterare (Staphilococcus aureus , Salmonella, Cl.botulinum)
2.4 CULTURI STARTER CONCENTRATE DIN SPORI DE MUCEGAI
Tehnologia de obţinere include următoarele operaţii: prepararea mediului de cultură, sterilizarea acestuia şi repartizarea în eprubete (agar înclinat) însămânţarea mediului din eprubete cu spori de mucegai din cultura dorită termostatarea pentru germinarea sporilor cu organe purtătoare de spori şi sporularea preluarea sporilor cu 2 ml ser fiziologic 0.8% şi însămânţarea mediilor de cultură Czapek, cu 2%
agar, aflate în vase Roux recoltarea sporilor din vase Roux cu ser fiziologic 0.8% astfel încât să avem în suspensie 108-1010
spori/ml păstrarea suspensiei la 0...4oCCultura de spori de mucegai concentrată poate fi livrata şi sub formă liofilizată .Culturile starter din
spori de mucegai se utilizează în industria cărnii şi laptelui.
3. CULTURI STARTER PENTRU BIOTEHNOLOGII FERMENTATIVE ALE PRODUSELOR DE ORIGINE VEGETALA
Ideea de a folosi ca maia(biomasă de celule de drojdie) o anumită specie de levuri datează de la sfârşitul secolului XIX şi apartine lui Christian Hansen. Ea are la bază teoriile lui Pasteur şi rezultatele cercetărilor proprii care dovedeau că există genuri de levuri utile şi altele dăunătoare proceselor fermentative.Numele lui se leagă de începuturile unei etape cu totul noi în biologia levurilor (levură de la lat.levere=a ridica; fr.levain=aluat dospit, care desemnează acele microorganisme care pot produce fermentaţia), precum şi o reforma fundamentală a tehnicii fermentative. Niciunul dintre predecesorii săi nu a lucrat cu culturi pure. El elaborează un procedeu pentru obţinerea culturilor, plecând de la o singură celulă de levură şi studiaza proprietăţile fiziologice ale tulpinilor pure obţinute în felul acesta.
Folosirea culturilor pure de drojdii şi a altor microorganisme (în special bacterii) a fost dezvoltată foarte rapid în special in industria berii, industria vinului, industria drojdiei de panificaţiei, practic în toate domeniile în care cercetătorii şi producătorii erau interesaţi de obţinerea unor fermentaţii controlate sau pentru obţinerea unui anumit produs principal de reacţie (fabricare oţetului).
Cele mai importante domenii în care folosirea culturilor pure este practic astăzi de neconceput sunt industria vinului şi berii.
3.1 UTILIZAREA CULTURILOR STARTER IN INDUSTRIA VINULUI
Fermentaţia spontană a mustului de struguri are loc sub influenţa drojdiilor care provin de pe struguri, deci nu are loc datorita unei singure specii, ci datorită unui ansamblu de specii a căror evoluţie este dependentă de concentraţia în alcool.
Fermentaţia este declanşată de drojdiile apiculate (în special Kloeckera apiculata)care produc ~4% volume alcool şi al căror număr reprezintă 70-80% din totalul drojdiilor aflate în mustul fermentat. După declanşarea fermentaţiei încep să predomine drojdiile eliptice (Saccaromyces ellipsoideus) ce pot produce 16% volume alcool şi care în plina fermentaţie alcoolică reprezintă 90% din totalul drojdiilor.La sfârşitul fermentatiei încep să acţioneze şi drojdiile Saccaromyces oviformis, care formează în mustul fermentat pină la 18-20% volume alcool.
Fermentaţia mustului de struguri poate fi însă controlată şi dirijată în două moduri: prin folosirea de culturi de producţie de drojdii indigene prin utilizarea de culturi starter de drojdii sub formă de culturi starter de producţie sau culturi
starter concentrate Avantajele folosirii celor două tipuri de culturi sunt următoarele :
40

- pornirea rapidă a fermentaţiei mustului chiar în condiţiile sulfitării cu doze mari de SO 2
impusă de starea recoltei - se modifică numeric şi calitativ microflora levurică din must, ceea ce face ca fermentarea
mustului să fie mai rapidă şi mai completă3.1.1 CULTURI DE PRODUCţIE DE DROJDII INDIGENE
In scopul obţinerii culturilor de producţie din drojdii indigene cu câteva zile înainte de începerarea campaniei de vinificare, se culege o cantitate de ~1500 kg struguri sănătoşi, copţi din care se obţine aproximativ 1200 l must.Circa 100 l se sulfitează la nivel de 5-10 g/hl SO2 şi se lasă în repaus 10-120 ore , într-un loc racoros pentru depunerea suspensiilor. Mustul limpede este transvazat într-un vas şi se depozitează la o temperatură care favorizează fermentaţia alcoolică(15-20 oC). Datorită procesului de selecţie exercitat de anhidrida sulfuroasă, procesul de fermentaţie este realizat de drojdiile eliptice. In momentul în care fermentaţia s-a declanşat, se adugă treptat restul de must (1100 l), care a fost sulfitat la obţinere cu 30-40 g/hl. Dupa 2-3 zile se obţine cultura primară(~ 12 hl), care este folosită pentru însămânţarea unei sarje mai mari de must sulfitat, din care după 2-3 zile de fermentaţie (cultură secundară) se folosesc cantităţi corespunzătoare pentru însămânţarea unei sarje mai mari de must(cultură de producţie).Această cultură de producţie este folosită pentru însămânţarea partidelor mari de must în vederea fermentaţiei.
3.1.2 CULTURI STARTER DE DROJDII PENTRU VINIFICATIECulturile starter se obţin prin selecţionarea celor mai valoroase drojdii din punct de vedere al
însuşirilor de fermentare.Obţinerea culturilor starter implică izolarea drojdiilor din habitatul lor natural şi multiplicarea pe
medii de cultură adecvate (must de struguri, must agarizat)Formele sub care se comercializează aceste culturi starter de drojdii sunt:
culturi starter în mediul lichid culturi starter pe mediul solid de gelatină sau geloză culturi starter liofilizate (drojdiile se liofilizează împreună cu mediul de protecţie )
Principalele culturi starter de drojdii sunt: - cultură starter de Saccaromyces ellipsoideus
- cultură starter de Saccaromyces oviformis - cultură starter de Saccaromyces pombe
Cultura starter de Saccaromyces ellipsoideus se utilizează pentru prelucrarea strugurilor avariaţi, a celor proveniţi din podgorii noi la care nu s-au identificat specii de drojdii valoroase, la obţinerea vinurilor spumante.Cultura starter de Saccaromyces oviformis este necesară pentru completarea fermentării.Cultura de producţie se adugă chiar la începutul fermentatiei, cu condiţia ca raportul Saccaromyces oviformis:Saccaromyces ellipsoideus să fie de 10:1.Cultura starter de Saccaromyces pombe realizează o dezacidifiere biologică , fără ca zahărul rezidual să fie fermentat.Această metodă se aplică pentru dezacidifierea musturilor care urmează să fie concentrate.
Pentru a fi utilizate în producţie culturile starter se pregătesc din inocul prin metoda pasajelor multiple. Se pot folosi şi culturi starter concentrate, care se conservă prin congelare sau prin uscare. Cultura starter concentrată conţine 3*1010 drojdii/g şi este păstrată sub atmosfera de CO2 sau N2. Inainte de folosire aceste concentrate se rehidratează în amestec apă/must sau soluţie 10% zahăr.Culturile concentrate se utilizează direct în producţie(fără pasaje).
4. ROLUL CULTURILOR STARTER ÎN ASIGURAREA CALITATII PRODUSELOR ALIMENTARE
4.1. Asigurarea inocuităţii produselor prin folosirea culturilor starterDintre toate avantajele folosirii culturilor starter, cel mai important este cel care asigură un anumit
grad de inocuitate produsului alimentar, datorită:— acizilor organici (şi în special acidului lactic) acumulaţi în mediu ;— substanţelor antibiotice şi bacteriocine ;
41

— competiţiei bacteriilor lactice cu microorganismele patogene şi cele de alterare în ceea ce priveşte consumul de substanţe nutritive ;— inhibării producţiei de amine biogene;— inhibării producţiei de nitrozamine.
Deosebit de importantă este asigurarea inocuităţii microbiologice a produselor alimentare la care s-au folosit culturi starter de bacterii lactice. Alături de materiile prime şi auxiliare de calitate ireproşabilă sau igiena producţiei şi personalului, prin folosirea de culturi starter de bacterii lactice se asigură o dominare numerică a acestora faţă de microflora naturală (spontană), inclusiv cea patogenă (salmonele, stafilococi, clostridii), care este împiedicată în dezvoltarea ei datorită unui sistem antagonic complex.
Sistemul antagonic include acizi organici (în special lactic şi acetic), antibiotice, peroxizi şi metaboliţi, nefiind exclusă nici competivitatea dintre bacteriile lactice pe de o parte şi cele de alterare şi patogene, pe de altă parte, pentru substanţele nutritive esenţiale din mediul în care se dezvoltă (produsul alimentar).
Acidificarea mediului este intensă şi rapidă mai ales în cazul în care substratul conţine un glucid uşor fermentescibil (lactoza—în cazul bacteriilor lactice tipice), datorită acumulării de acid lactic. In funcţie de tipul microflorei lactice, se pot acumula şi alţi acizi organici, în special acid acetic. Prin acu-mularea de acizi organici, pH-ul mediului (produsului alimentar) scade sub valoarea optimă de dezvoltare a bacteriilor de alterare şi a celor patogene.
2. Acţiunea faţă de stafilococi si salmoneleÎn preparatele de carne care au suferit un tratament termic necorespunzător (substerilizare), în
cîrnaţii şi salamurile crude semiuscate (aw > 0,92) sau în salamurile uscate (aw < 0,92), se poate dezvolta Staphylococcus aureus care elaborează enterotoxină, stabilă la căldură. In general, stafilococii proli-ferează şi produc enterotoxină în primele stadii ale fermentaţiei salamurilor şi cîrnaţilor cruzi, mai ales în stratul periferic.
Inhibarea dezvoltării stafilococilor este mărită dacă raportul bacterii lactice/stafilococi este mare şi dacă temperatura de fermentare-maturare este mai scăzută. S-a demonstrat experimental că, în prezenţă de cultură starter de bacterii lactice, numărul de stafilococi coagulazo-pozitivi din salamurile fermentate la 22 ... 24°C este mai redus iar producerea de enterotoxină este inhibată şi aceasta din cauză că, inocul de bacterii lactice (pediococi şi lactobacili) va controla fermentaţia, va accelera formarea de acid lactic şi astfel, indirect, va întîrzia dezvoltarea stafilococilor.
Prezenţa salmonelelor în cîrnaţii şi salamurile crude este mai rară. Bacteriile lactice componente ale culturilor starter au acţiune inhibitoare faţă de salmonele, efectul inhibitor fiind dependent de specie, suşă, raportul bacterii lactice/salmonele, temperatura de fermentare, intensitatea şi viteza acumulării acidului lactic.
Stafilococii şi salmonelele pot fi prezente şi în diverse produse lactate (produse lactate acide, smântână, brânză proaspătă de vaci, unt, brînzeturi fermentate cu pastă opărită sau neopărită, brânză telemea, îngheţată, lapte praf). La produsele lactate la care se folosesc culturi starter de bacterii lactice, s-a dovedit că există un antagonism evident între acestea, pe de o parte, şi stafilococi-salmonele, pe de altă parte In cazul culturilor starter de streptococi s-a dovedit că acţiunea de inhibare nu este direct corelată cu viteza de acumulare a acidului lactic .
In ceea ce priveşte formarea de toxine botulinice de către CI. botulinum tip A, B, E, F (toxine care sunt labile la căldură şi care se deosebesc între ele prin faptul că sunt neutralizate specific de către anticorpii omologi), aceasta este dependentă de condiţiile în care se dezvoltă microorganismul: potenţial redox, pH, activitatea apei; concentraţia NaCl şi azotit, temperatură. Experimental, s-a dovedit că prin folosirea culturilor starter de bacterii lactice, în prezenţa unui glucid fermentescibil adecvat (glucoza, zaharoza) se inhibă în mare măsură formarea de toxină botulinică. Dacă este prezent şi azotitul în concentraţie de 50—150 mg/kg, acţiunea de inhibare a formării toxinei este totală în cîrnaţii cruzi fermentaţi-maturaţi la 27°C.
In cazul produselor vegetale murate (fermentate lactic), incidenţa formării toxinei botulinice este extrem de rară, din cauza rapidei dezvoltări a bacteriilor producătoare de acid lactic, bacterii care au o bună toleranţă la concentraţii mai mari de NaCl (2—8%) şi acid lactic. Ca rezultat al respiraţiei ţesuturilor vegetale supuse murării şi a dezvoltării bacteriilor se creează un mediu anaerob şi prin urmare competiţia între bacteriile producătoare de toxine şi cele producătoare de acizi este cîştigata de acestea din urmă, care sunt tolerante la pH scăzut şi concentraţie mare de NaCl. In produsele vegetale murate,
42

dezvoltarea lui CI. botulinum şi producerea de toxină este inhibată la un pH < 4,6 şi la concentraţiile de NaCl existente în produs. La conservarea prin murare a produselor vegetale trebuie să se aibă însă în vedere că anumiţi compuşi pot acţiona favorabil sau defavorabil asupra bacteriilor lactice. De exemplu, în varză există substanţe care inhibă dezvoltarea bacteriilor gram-negative dăunătoare, dar nu şi pe cele lactice. Măslinele verzi tratate necorespunzător cu alcalii sau prin şoc termic, înainte de a fi saramurate, pot avea substanţe inhibitoare faţă de bacteriile lactice.
Avînd în vedere că produsele vegetale conţin azotaţi, aceştia sînt transformaţi în azotiţi, în timpul fermentaţiei lactice, care inhibă dezvoltarea microorganismelor formatoare de spori în produsele vegetale fermentate lactic. O contribuţie însemnată la capacitatea de conservare a produselor alimentare la care s-au folosit culturi starter o are şi peroxidul de hidrogen (H2O2) care este bacteriostatic şi bactericid, eficace în special faţă de microorganismele ce nu secretă catalază.
În acest context sunt mai sensibile la H2O2 bacteriile gram-negative (salmonele, bacteriile coliforme) în comparaţie cu cele gram-pozitive (stafilococi, clostridii). Peroxidul de hidrogen poate fi eficace prin el însăşi sau datorită formării unor compuşi inhibitori cu unele componente din produsul alimentar. De exemplu, în laptele crud, peroxidul de hidrogen reacţionează cu tiocianatul şi peroxidaza şi formează o serie de compuşi care au acţiune inhibitoare faţă de microorganismele de alterare
Având în vedere că anumite culturi starter pot forma H2O2 în condiţii de refrigerare fără a se dezvolta numeric şi fără a produce acid lactic, s-a ajuns la concluzia că aceste culturi starter pot fi utilizate pentru inhibarea microorganismelor psihrotrope în produsele alimentare depozitate în condiţii de refrigerare.Astfel, se cunoaşte că aproape 90% din bacteriile psihrotrope care alterează carnea sînt gram-negative, grupul cel mai important constituindu-l Pseudomonas— Achromobacter, atît pentru carnea proaspătă cât şi pentru cea refrigerată. S-a constatat că prin utilizarea unei culturi starter de Streptococus lactic şi Leuconostoc citrovorum în proporţie de 10% faţă de masa cărnii de vită tocată se împiedică dezvoltarea microorganismelor aerobe, fără a se ajunge la modificări de pH. De asemenea, prin folosirea de culturi starter de L. bulgaricus, L. lactis şi Pediococcus cerevisiae în carnea mărunţită la nivel de 5 • IO8 celule/g carne tocată şi păstrată în condiţii de refrigerare, s-a ajuns la o inhibare a bacteriilor psihrotrope, inhibare pusă pe seama H2 02 .
Cercetările au demonstrat că culturile starter lactice pot fi folosite şi ca microfloră de suplimentare în alimentaţia animalelor tinere (viţei, purcei) şi aceasta pe motivul că la viţei şi purcei lactobacilii şi streptococii joacă un rol important în digestia glucidelor, în special a lactozei, care este transformată în acid lactic ce joacă un dublu rol:— aciditatea lactică favorizează acţiunea pepsinei ce provoacă coagularea cazeinei laptelui, care în continuare este atacată de tripsină ce eliberează aminoacizii ;— aciditatea mediului intestinal blochează dezvoltarea microorganismelor cu putere patogenă, cum ar fi E. coli care se dezvoltă în mediu alcalin sau uşor acid.
Prin schimbarea habitatului şi datorită tratamentelor fizice, psihice, transportului sau înlocuirii laptelui matern cu lapte simulat, se modifică flora lactică la viţei şi purcei. Acest lucru se realizează şi prin tratament cu antibiotice. Datorită pierderii florei lactice, viţeii şi purceii pot căpăta diferite afecţiuni digestive care pot produce mortalitate de 3—10%. In condiţiile în care viţeii şi purceii nu sînt alăptaţi normal, alimentele de alăptare pot fi suplimentate cu culturi starter care trebuie să aibă o concentraţie mare de germeni viabili în condiţii normale de depozitare ale alimentelor de alăptare,. Se utilizează culturi starter de Str. lactis, Str. thermophilus, L. helveticus. L. acidophilus, îmbogăţite cu aminoacizi şi vitamine din grupul B. 3. Producerea de antibiotice şi bacteriocine
Anumiţi streptococi lactici şi lactobacili produc antibiotice în mediul în care se dezvoltă. De exemplu, Str. cremoris produce diplococcină strâns asociată de celulele respective, antibiotic care este inhibitor faţă de Staphylococcus aureus dar nu şi faţă de Eschcrichia coli.
Streptococcus lactis produce nizină, stabilă la căldură şi la pH acid, care are acţiune inhibitoare faţă de bacteriile gram-pozitive. Nizina nu stopează germinarea sporilor, dar inhibă dezvoltarea sporilor germinaţi. Lactobacillus plantarum produce lactolină, iar Lactobacillus brevis produce lactobrevină. Specii de Lactobacillus acidophilus produc lactocidină, acidofilină si acidolină. Acidofilina produsă în timpul dezvoltării lactobacililor în lapte este stabilă la căldură şi pH acid. Acidolină produsă în aceleaşi condiţii are acţiune faţă de bacteriile gram-pozitive şi negative.
43

Anumite specii de microorganisme produc bacteriocine care nu sînt clasificate ca antibiotice dar care au acţiune antibiotică. De exemplu, L. cremoris produce diacetil prin fermentarea citratului, care exercită acţiune inhibitoare faţă de bacteriile patogene şi cele de alterare. Având în vedere concentraţia relativ redusă a diacetilului în produsele alimentare, nu se poate vorbi de o acţiune inhibitoare a acestuia faţă de grupele de microorganisme menţionate. Efectul inhibitor al diacetilului este evident numai la o concentraţie a acestuia de 170 mg/kg, ceea ce ar însemna folosirea diacetilului ca aditiv de conservare, fapt ce nu este permis de legislaţia sanitară în vigoare, el fiind socotit un aditiv de aromatizare.
4. Producerea de amine biogeneDintre aminele biogene din produsele fermentate interesează în mod deosebit histamina şi
tiramina, care se formează în cantităţi mai mari datorită acţiunii histidin şi tirozin-decarboxilazelor produse de microorganisme (bacterii şi drojdii). S-a dovedit că în metabolismul normal al omului, tiramina este oxidată rapid de monoamina-acid-oxidaza (MAO), dar la persoanele tratate cu inhibitori MAO, tiramina se acumulează şi exercită un efect vasotonic puternic. Printr-un mecanism diferit şi histidina acţionează ca un vasopresor, în prezenţa inhibitorilor MAO.
In produsele de carne fermentate au fost găsite cantităţi relativ mari de tiramina şi histamina şi cantităţi mai mici de feniletilamină, triptamină, cadaverină, putresceină, în funcţie de durata fermentării. Produsele fermentate natural au un conţinut între 184 şi 534 mg tiramină/kg produs. Activitatea tirozin-decarboxilazică a microflorei naturale creşte paralel cu creşterea acidităţii produsului, fiind un mecanism de protecţie al bacteriilor faţă de acidificarea mediului. Microflora spontană are activitate decarboxilazică mai mare, fiind adaptată la temperaturi mai ridicate, la o activitate a apei aw mai redusă şi la o concentraţie de NaCl mai mare. Tirozindecarboxilaza din produsele de carne fermentate s-a dovedit a fi indusă în prezenţa tirozinei disponibile. Prin folosirea de culturi starter de Lactobacillus şi Pediococcus, se împiedică dezvoltarea microflorei cu activitate proteolitică (punerea în libertate de tirozină) şi decar-boxilazică şi deci se împiedică formarea unor cantităţi mari de tiramina. (Acelaşi lucru se poate spune şi despre acumularea celorlalte amine biogene.) Şi în cazul altor produse alimentare se înregistrează nivele mai scăzute de amine biogene dacă s-au folosit culturi starter.
În literatură sînt indicate următoarele nivele de tiramina: 0—2,17 g/g în brînza fermentată; 6—11 g/ml în bere; 0,2—12 g /ml în vin; 95—304 g/g în extract de drojdie; 0,9—13 mg/100 g în varză tocată şi murată.
5. Inhibarea producerii de nitrozamineÎn cazul cărnii, azotitul se adaugă pentru formarea culorii cărnii sărate destinată preparatelor din
carne, semiconservelor şi salamurilor crude şi pentru proprietăţile sale antimicrobiene. Azotitul rezidual poate însă reacţiona cu aminele secundare, conducînd la formarea de nitrozamine care au acţiune cancerigenă. Prin reducerea cantităţii de azotit rezidual se diminuează producţia de nitrozamine în produsele din carne dar, în acelaşi timp, se reduce şi capacitatea acestuia de a inhiba dezvoltarea lui CI. botulinum şi de formare a toxinei botulinice. Bacteriile lactice, avînd la dispoziţie un substrat fermentescibil, sunt agenţi eficace antibotulinici, deoarece produc acid lactic şi deci scad pH-ul sub valoarea optimă de dezvoltare a lui CI. botulinum, deci asigură stabilitatea produselor chiar în prezenţa unei cantităţi mai mici de azotit rezidual. Realizarea unui pH scăzut şi existenţa unui nivel de azotit rezidual de asemenea scăzut fac să scadă producţia de nitrozamine, mai ales la produsele care se supun frigerii şi coacerii (tabelul 40).
Conţinutul de nitrozamine în bacon după 25 zile de păstrare la 4°C
Specificaţie Azotit rezidual, mg/kg
NTG pH Conţinut de nitro-zamine, ppb
Fără adaos de cultură starter
Cu adaos de cultură starter
20-40
4-16
104 -105
106- 107
6,0-6,4
5,2-5,6
10-30
2- 9
Produsele vegetale conţin azotaţi şi azotiţi şi anumite bacterii lactice, incluzînd suşe de L.plantarum care pot reduce azotaţii la azotiţi. In această direcţie, s-a găsit că azotitul din varza murată
44

creşte de la un nivel iniţial foarte redus pînă la 108 mg/kg, după 5 zile de fermentare, după care se reduce aproape de nivelul iniţial. Din această cauză, în produsele vegetale fermentate lactic nu s-au pus în evidenţă nitrozamine, chiar dacă unul din partenerii de reacţie este prezent (aminele). La produsele vegetale fermentate, datorită pH-ului scăzut, azotitul rezidual este adus la nivele nepericuloase din punct de vedere al formării nitrozaminelor.
ALIMENTE FUNCŢIONALE
CAP I. IMBUNATATIREA VALORII NUTRITIVE SI IGIENICE A PRODUSELOR ALIMENTARE
Alimentaţia îşi pune amprenta asupra întregii existenţe umane, începând cu dezvoltarea embrionară, determinând starea de sănătate şi capacitatea de muncă. Ea constituie un factor cu acţiune permanentă care influenţează desfăşurarea proceselor metabolice, deoarece hrana reprezintă izvorul şi regulatorul proceselor de schimb. Dar între necesităţile organismului şi aportul alimentar nu există totdeauna un echilibru optim, acesta fiind perturbat de numeroşi factori sociali, economici, tehnologici, culturali, etc.
Dintre factorii care contribuie la un dezechilibru metabolic, în primul rand trebuie menţionat deficitul de substanţe nutritive, în special substanţe cu rol plastic, precum şi deficitul energetic , care determină o scădere a intensităţii proceselor metabolice , antrenează un deficit în alte substanţe care, în final, se reflectă asupra creşterii şi stării de sănătate a populaţiei.
Manifestările clinice ale deficitului de substanţe nutritive apar după un timp îndelungat de la folosirea unui regim alimentar necorespunzător.Acest lucru se explică prin faptul că deficitul de substanţe
45

din ratia alimentară, pe o perioadă determinată de timp, poate fi compensate pe baza proceselor biochimice de adaptare. In timp mai îndelungat, alimentaţia neechilibrată depăşeşte capacitatea de adaptare a organismului şi apar dereglări metabolice profunde care afectează sănătatea individului şi a unor colectivităţi.
Pentru ţările industrializate, creşterea calităţii şi cantităţii bunurilor alimentare puse la dispoziţie, creşterea nivelului de trai şi a facilităţilor de procurare, lipsa de cumpătare faţa de tentaţiile provate de însuşirile foarte atrăgătoare pe care le capată alimentele prin prelucrare, precum şi tendinţa de utilizare excesivă a produselor concentrate şi ultrarafinate, conduce , inevitabil, la un supraconsum. Toate acestea se asociază cu mutaţiile care s-au produs în viaţa sociala datorită civilizaţiei actuale, astfel: creşterea gradului de urbanizare, a condiţiilor de confort şi diminuarea efortului fizic. Ca urmare, pentru multe segmente de populaţie , asistăm la o supralimentaţie caracterizată prin aport mare energetic dar săracă în biocatalizatori.
Acest fenomen are loc pe fondul unei solicitări intelectuale mai intense, a creşterii stresului neuropsihic, a poluării mediului şi a alimentelor, procese care converg spre o mai mare solicitare de biocatalizatori: vitamine şi biolemente, care au rol de protecţie a organismului.
In aceste condiţii există orientarea de a echilibra şi mării valoarea nutritivă a dietei alimentare. Pe plan mondial se dezvoltă în acest moment proiecte biotehnologice de obţinere a unor alimente speciale FoSHU (Food for specified used), cunoscute şi sub denumirea de alimente funcţionale sau nutraceutice.
Impactul economico-social urmărit prin aceste proiect poate fi sintetizat în urmãtoareleefecte:
economice-selectarea ingredientelor după principiul celui mai avantajos raport dintre preţ/costuri/valoare nutritivă, în sensul minimizării preţului şi costurilor, maximizării valorii nutritive, pe baza unui model de calcul;-selectarea celor mai avantajoase ambalaje şi modalitãţi de ambalare din punct de vedere al stabilităţii la păstrare, al funcţionalităţii şi al igienei alimentului, corespunzătoare alimentelor funtionale realizate;-asigurarea celui mai convenabil echilibru al mesajului informaţional al etichetei privind menţiunile obligatorii;-motivarea opţiunii pentru alimentele de maximã siguranţă nutriţională, utilizând materii prime naturale, nepoluate sau cu substanţe antinutriţionale;-posibilitatea de a obţine cele mai potrivite produse alimentare/suplimente nutriţionale, la costuri şi preţuri mai scãzute faţã de cele practicate pe plan internaţional, în raport cu alocãrile financiare.
sociale-posibilitatea ca orice consumator să consume cu plăcere şi cu încredere alimentele funcţionale obţinute, în toate fazele activităţii sale ;-eliminarea suspiciunilor referitoare la prezenţa unor substanţe interzise în alimentele funcţionale;
Deşi vechimea alimentaţiei se confundã cu însăşi istoria umanităţii, o alimentaţie corectă, adecvată diferitelor condiţii de viaţă şi efort este o problemã de actualitate, care preocupă cercetătorii din multe domenii nutriţie, tehnologie alimentarã, igiena alimentaţiei, medicina muncii .
Această preocupare a contribuit la îmbogăţirea gamei produselor alimentare cu noi sortimente, suplimente alimentare, din dorinţa de a satisface cât mai complet necesităţile nutritive ale diferitelor grupe de consumatori, în funcţie de condiţiile fiziologice speciale, vârstã şi efort.
În acest sens, suplimentele alimentare semnifică produsele alimentare al căror scop este să suplimenteze dieta normală şi care reprezintă surse concentrate de nutrienţi sau alte substanţe cu efect nutriţional sau fiziologic, singure sau în combinaţii.
I.1 SUPLIMENTELE NUTRITIONALE
Concept şi importanţă in alimentaţie
Suplimentele alimentare pot fi definite drept acele produse alimentare care conţin un plus de nutrienţi cu o biodisponibilitate ridicată, destinate a acoperi necesităţile nutritive fiziologice specifice.
Potrivit Ordonanţei de urgenţã a Guvernului României nr. 97/21 iunie 2001 privind reglementarea producţiei, circulaţiei şi comercializãrii alimentelor, nutrienţii şi/sau suplimentele alimentare reprezintă substanţe nutritive, respective protide, lipide, glucide, vitamine, macro şi
46

microelemente minerale şi/sau preparate prelucrate sub formã de tablete, capsule, drajeuri, pulberi sau lichide care au în compoziţia lor macro şi micronutrienţi şi/sau alte substanţe comestibile, care sunt consumate în cantităţi definite, în mod suplimentar faţã de aportul alimentar obişnuit.
Este admis de către toţi nutriţioniştii cã funcţia alimentatiei, reprezentând schimbul permanent de informatie, energie şi substanţã, este, alături de respiraţie, cea mai strânsă legãturã a organismului uman cu mediul sãu de viaţã.
Analizând cele trei apecte ale acestui schimb se observă următoarele : schimbul de informaţie
se referã în special la structura alimentului, care trebuie sã fie cât mai apropiatã de posibilitãţile organismului de a-l prelucra digestiv şi metabolic, având o biodisponibilitate ridicatã;
schimbul de energie se referã, atât la conţinutul în nutrienţi bogaţi energetic, cât şi biodisponibili;
schimbul de substanţă se referã la conţinutul general de nutrienţi şi la echilibrul proporţional dintre aceştia.
Analizând în acest mod proprietãţile nutriţionale ale suplimentelor alimentare, se pot identifica şi substanţele şi produsele care nu pot fi utilizate, în aceste condiţii în alimentaţie.
Acestea sunt produsele greu de digerat şi/sau metabolizat, chiar dacã sunt bogate în nutrienţi. De asemenea, nu sunt indicate alimentele sau substanţele pentru prelucrarea cărora organismul uman consumă o cantitate crescută de enzime sau vitamine.
Un exemplu în acest sens este consumului de alcool etilic, care, deşi are un conţinut caloric foarte mare (care nu se conservã însã în organismul uman), este metabolizat cu un consum ridicat de vitamina B1, implicată în metabolismului glucidic, atât de necesar efortului neuromuscular.VitamiDe asemenea, o altă condiţie este adaptarea alimentaţiei la specificul activităţii depuse de organismul uman, individualizarea alimentaţiei, deoarece fiecare organism are o reactivitate biologicã proprie, ce se evidenþiazã mai ales în condiţii de efort sau boală.
I.2 CLASIFICAREA SUPLIMENTELOR NUTRITIONALE
În conformitate cu prevederile Ordinului MSF 282/2001 , suplimentele nutriţionale sunt reprezentate de :
vitamine şi elemente minerale în cantităţi care pe unitatea de consum (tabletã, capsulã, drajeu, linguriţã etc.) nu depãşesc de 3 ori dozele fiziologice zilnice recomandate pentru o persoanã în cazul vitaminelor hidrosolubile şi de 1,5 ori în cazul vitaminelor liposolubile şi al elementelor minerale;
proteine pure sau sub formã de concentrate proteice, izolate proteice şi hidrolizate proteice, aminoacizi şi amestecuri ale acestora;
uleiuri alimentare considerate dietetice (de exemplu: uleiuri din germeni de seminţe de cereale, uleiuri din ficat de peşte) şi acizi graşi polinesaturaţi esenţiali; fosfolipide( lecitine, cefaline, serinfosfatide, inozitolfosfatide) până la cantitãţi ale conţinutului de fosfor de 1 g/zi;
fibre alimentare până la cantitatea de 24 g/zi; lapte pentru copii, indiferent de vârsta de administrare, dar fãrã indicaţii terapeutice; produse pentru copii: lapte cu cereale şi/sau fructe, produse tip “ baby-food îndulcitori sintetici alimentari; produse pentru slăbit şi pentru menţinerea greutăţii corporale, fără indicaţii terapeutice; produse parabiotice: probiotice, prebiotice, simbiotice - alimente funcţionale care
regleazã flora microbianã intestinalã; produse pentru sportivi care conţin vitamine, elemente minerale, proteine, hidrolizate proteice,
aminoacizi, glucide uşor asimilabile, acizi graşi esenţiali şi alţi nutrienţi cu rol în susţinerea efortului sportiv, în refacerea post-efort, în dezvoltarea şi menţinerea masei musculare.
I.3 MODELAREA VALORII NUTRITIVE A ALIMENTELOR CU UTILIZARI NUTRITIONALE SPECIALE
Consumul de suplimente nutriţionale genereazã o serie de beneficii:
47

a. Efecte positive asupra sãnãtãţii, performanţelor fizice sau stãrii psihice : îmbunãtãţirea stãrii de sãnãtate sau bunãstare generalã şi/sau reducerea riscului unor boli; mentinerea sãnãtãţii şi dezvoltarea optimã a organismului; reglarea unor procese din organism, controlul unor funcţii ale organismului, reducerea duratei
convalescenţei, reducerea efectelor îmbãtrânirii;b. Beneficiu fiziologic şi/sau asigurarea protecţiei împotriva unor boli cronice; prevenirea
deficitului de vitamine şi de substanţe minerale; sunt considerate ca alimente pentru utilizare nutriţionalã particularã; sunt considerate ca alimente funcţionale; evaluarea se face dupã un protocol similar medicamentelor; pe etichetã nu se admit revendicãri referitoare la diagnosticarea, atenuarea, tratarea,
prevenirea sau vindecarea unor boli.Dintre efectele negative ale consumului de suplimente nutriţionale, datorate în special
toxicitãţii alimentelor funcţionale şi a nutraceuticelor, amintim: supradozarea principiilor active; interacţiunea cu alte componente din produs; prezenţa unor substanţe utilizate la extracţia sau formularea produsului de bazã.
Căile prin care un component poate afecta organismul uman sunt numeroase: reacţii adverse sau interrelaţii cu alte componente, care pot determina, fie distrugerea, fie
reducerea disponibilităţii nutrienţilor; interferenţe cu utilizarea sau digestia nutrienţilor; interferenţe sau interacţiuni cu substanţe terapeutice farmacologice.În funcţie de valoarea lor nutritivã, din punct de vedere al nutrienţilor de bazã, se poate realiza
gruparea alimentelor cu utilizări nutriţionale generale în: produse cu preponderenţã protidicã, lipidicã sau glucidicã.
Importanţa cunoaşterii valorii nutritive a produselor alimentare este dictatã şi de segmentarea accentuatã a necesitãţilor de consum alimentar, fãcând posibilã realizarea de alimente cu utilizãri nutriţionale specifice, care sã corespundã nevoilor anumitor grupe de consumatori (produse hipoglucidice pentru persoanele diabetice, produse îmbogãţite nutriţional pentru copii, produse hipocalorice pentru anumite diete, produse energizante pentru sportivi ş.a.).
La nivel macroeconomic, este înţeleasã ca o componentã a cererii globale, iar situaţia alimentarã a unei populaţii este adesea abordatã numai la nivelul mediilor naţionale obţinute prin metoda bilanturilor alimentare sau a contabilităţii naţionale. Consumul alimentar pe locuitor este principalul indicator general disponibil pentru a urmãri evolutia securitãţii alimentare mondiale, mãsurat la scarã nationalã de disponibilităţile enegetice alimentare medii (DEA) exprimate în kilocalorii. Experienţa internaţionalã demonstreazã cã în ţãrile în care DEA se situeazã în jurul valorii de 2700 Kcal, proporţia persoanelor subalimentate este micã.
Diferenţa în consumul alimentar este marcatã prin inegalitatea observatã în capacitatea de consum. Aceastã putere de consum este caracterizatã esenţial prin capacităţile productive din economiile de subzistenţã şi prin puterea de cumpãrare din economiile de piaţã.
S-a constatat cã pe mãsurã ce creşte produsul intern brut, creşte şi totalul de proteine din regimul alimentar (aceastã relaţie este valabilã şi pentru creşterea gradului de urbanizare). Pentru niveluri scãzute ale venitului, aproximativ 75% din kilocalorii provin din poliglucide, constatându-se un consum ridicat de porumb, orez, grâu, cartofi. Pe mãsurã ce venitul creşte, dieta devine mai complexã: creşte cantitatea de grãsimi (în special de naturã animalã), iar ponderea energiei provenite din poliglucide se reduce la 30%. Proporţia energiei din zaharuri creşte, iar contribuţia proteinelor rãmâne relative constantã, înregistrându-se însã un salt calitativ, prin trecerea de la consumul de proteine vegetale la cel de proteine animale.
În înţelegerea inegalitãţilor din consumul alimentar trebuie plecat şi de la premisa cã alimentaţia este un act social complex fondat pe satisfacerea cererii biologice, dar şi pe plãcerea de gust şi consum de simbol, de semnificaţie socialã. Nevoile de consum alimentar sunt nevoi reale, exprimate. De multe ori însã, cumpãrãtorul de produse alimentare are nevoie şi de satisfacţie, atât în procesul de achiziţie, cât şi în cel de consum. Marketerii, sociologii apreciazã cã un potenţial cumpãrãtor nu specificã întotdeauna ceea ce doreşte. El are nevoi neexprimate, care trebuie identificate. În plus, interesul unui cumpãrãtor depinde
48

şi de veniturile alocate alimentaţiei, de accesul la sortimentul de alimente oferit, de disponibilitatea şi preţul produselor nealimentare şi al serviciilor.
În general, cererea de mărfuri, este puternic înrâuritã de factori numeroşi şi diferiţi: economici (investiţii, ofertã, venituri, preţuri); demografici (numãrul şi structura populaţiei pe vârste şi sexe, profesii, grad de culturã şi
instruire); sociologici (modã, mediul social, mobilitate socio-profesionalã); organizatorici(calitatea activitãţii comerciale, acţiuni promoţionale, etc.); sezonieri şi conjuncturali.
Între obiectiv şi subiectiv, comportamentul alimentar este rezultatul factorilor: de naturã exogenã constituiþi din alimente propriu-zise având valoare nutritivã şi implicând
tonusul emotiv naturã endogenã de ordin fiziologic (senzaţia de foame sau sete), sensorial (sensibilitatea tactilã,
cea termicã, gustul) şi psihologic (semnificaţia simbolicã a alimentelor, funcţia socialã a zilelor festive).
În tot acest complex, fenomene ca instinct, dorinţã, agreabil sau dezagreabil, obişnuinţe diferã de la un individ la altul şi de la o populaþie la alta.
Produsele alimentare sunt incluse în categoria bunurilor de uz curent şi se caracterizeazã, pe lângã epuizarea lor continuă în procesul de consum, prin existenţa unui anumit termen de valabilitate (sau termen limitã de consum), prin anumite restricţii privind condiţiile de pãstrare, conservare, întrebuinţare. Ele deţin un loc bine determinat în cadrul consumului individual şi o anumitã stabilitate în consumul final. Faţã de producţie, care are un caracter sezonier, consumul produselor agricole (folosite în mare parte ca materii prime pentru industria alimentarã şi doar parţial consumate în stare naturalã) este mult mai uniform, înregistrând doar unele oscilaţiii în perioada de constituire a ofertei.
Ca expresie a nevoilor de consum, cererea reprezintã un permanent stimulator pentru productie, pentru lărgirea gamei sortimentale şi îmbunãtãţirea modului de satisfacere a consumului. Producţia realizata în vederea satisfacerii nevoilor de consum se confruntã, în cadrul pietei, cu cerintele societăţii, sub forma ofertei de mãrfuri.Oferta constituie un factor economic hotărâtor, fiind implicată major în structurarea şi volumul cererii. Prin amplificarea formelor de satisfacere a consumului, oferta de mãrfuri este, de asemenea, generatoare de nevoi.
Se considerã cã una din cele mi importante modalitãţi de care dispune o firmã pentru a avea succes pe piaţã o constituie diferenţierea eficientã a ofertei (prin produs, servicii care însoţesc produsul, personal, imagine). Pentru produsele alimentare diferenţierea este relativ dificilã. Pentru cele prelucrate, proiectarea nutriţională s-ar putea constitui într-o modalitate de diferenţiere, cu condiţia declarãrii reale a potenţialului energetic şi biologic, a aditivilor utilizaţi şi a respectãrii invariabilelor valenţe igienice. Mesajul informaţional, cuprinzând acele elemente ce vizeazã nutriţia, poate fi realizat numai de cãtre firme care pot garanta valorile minime pentru datele comunicate prin e t i c h e t ã sau a m b a l a j . Eventuala neconcordanţã dintre valoarea nutritivă declaratã a produsului şi cea realã, identificabilã de cãtre organismele de protecţie a consumatorului, pune în cauzã onestitatea, probitatea şi deci, reputaţia firmei producãtoare.
Proiectarea nutriţionalã modernã a alimentelor trebuie sã ia în considerare toate cele patru laturi inseparabile ale valorii nutritive :
1) valoarea psihosenzorială,2) valoarea energetică,3) valoarea biologică,4) valoarea igienică
iar modelarea si echilibrarea trebuie sã le vizeze pe toate, întrucât între ele existã interdependenţã. Trebuie avut în vedere faptul cã existã riscul ca modificând în sens pozitiv una din laturi (de exemplu valoarea biologicã prin adãugarea de sãruri solubile de fier) se poate modifica în sens negativ o altã laturã (scade valoarea psihosenzorialã, produsele cãpãtând un gust metalic sau astringent sau o culoare dezagreabilã).
49

Un alt exemplu îl constituie micşorarea valorii igienice prin îmbunãtãţirea valorii psihosenzoriale, în cazul adãugãrii în exces a unor aditivi alimentari.
Alimentele cãrora li s-a modelat valoarea nutritivã trebuie sã fie omogene şi stabile , sã poatã fi prelucrate şi obtinute în condiţii controlate.
Prin control trebuie sã se asigure constanţa dozelor indicate şi omogenitatea produselor obţinute, caracterul asimilabil al substanţei nutritive introduse, nealterarea caracteristicilor produsului, evitarea măririi exagerate a costului.
Eşecul în realizarea unor produse echilibrate nutritiv şi corect corelate cu necesitãţile segmentelor de populatie cãrora li se adreseazã are efecte grave asupra sãnãtãţii consumatorilor, generând situaţii de dezechilibru nutriţional, cu implicaţii majore pe termen lung. Pentru evitarea unor asemenea situaţii se impune un proces corect de asigurare a calitãţii noilor produse alimentare, a cãror valoare nutritivã a fost modelatã şi echilibratã.
Folosind previziunea tehnologicã, firma dispune de un instrument adecvat în determinarea duratei şi a volumului eforturilor necesare înnoirii gamei sale de produse. Ea obţine totdeauna informaţii asupra modalitãţilor de realizare a unor nivele calitative ale produselor în raport cu costurile acestora, cu strategiile de preţuri, cu schimbãrile în programul de distribuţie, cu oscilaţiile competitivitãţii lor pe piaţã. Esenţial pentru obiectul previziunii tehnologice nu este produsul ca atare, ci funcţiile lui, nevoile pe care urmeazã sã le satisfacã. Este perimatã practica de a avea reţete pe baza oricãror altor considerente decât cele nutriţionale. Astfel, mãrfurile alimentare actuale se caracterizeazã printr-un grad înalt de prelucrare , încorporând un volum din ce în ce mai mare de progres tehnico-ştiinţific.
Biotehnologia alimentarã modernã se diferenţiazã net de cea clasicã care urmãrea prioritar menţinerea sau ameliorarea calitãţii resurselor agroalimentare. Plecând de la necesitãţile obiective şi subiective de consum alimentar, se selecteazã materiile prime convenţionale şi neconvenţionale ce urmeazã a fi încorporate, aditivii alimentari corespunzãtori, tipurile de transformãri tehnologice eficiente pentru a ajunge la un produs finit cu proprietãţi identice sau cât mai apropiate de cele ale produsului proiectat.Utilizarea modelării în obţinerea, fabricarea şi monitorizarea continuã a proprietãţilor produselor alimentare, atât a celor de uz general, cât şi a celor destinate unor segmente speciale de consumatori, reprezintã nu numai un mijloc de validare a performanþelor producãtorilor, ci şi o modalitate sigurã de a evita orice risc în nutriţie.
I.4 INOCUITATEA PRODUSELOR NUTRITIONALE
Alimentaţia este o componentã indisolubilã a existenţei umane, omul ca organism heterotrof fiind nevoit sã preia din mediu substanţele nutritive necesare pentru desfãşurarea normalã a proceselor metabolice.
Cele cinci grupe de substante nutritive - glucidele, lipidele, protidele, substantele minerale şi vitaminele - nu sunt primite ca atare, ci sub forma unui complex, alimentul, ceea ce determinã şi o relaţie psihoafectivã cu acesta, relaţie ce poate deveni chiar determinantã în alegerea unui aliment din cele disponibile la un moment dat.
Caracterul de ingerabil al oricãrui aliment, contactul intern care se stabileşte între acesta şi organismul uman şi, mai ales, prin aliment, între organism şi mediul înconjurãtor, impune respectarea de cãtre producãtorii şi comercianţii acestor mãrfuri a unor restricţii, care sã asigure inocuitatea produselor consumate. Caracteristic produselor alimentare este marea diversitate de factori care pot influenţa inocuitatea acestora.
Alimentul este legãtura biologicã fundamentalã a omului cu mediul înconjurãtor. Orice perturbaţie a mediului îşi gãseşte, în alimente, mijlocitorul ideal pentru a ajunge la om.
50

Pentru a menţine starea de sãnãtate a populaţiei, produsele alimentare trebuie sã corespundã, în primul rând, sub aspectul calitãţii igienice (inocuitãţii) , deoarece neîndeplinirea acestei condiţii poate duce la îmbolnãviri mai mult sau mai puţin grave.
Factorii naturali de mediu acţioneazã în mod inevitabil asupra mãrfurilor alimentare pe întreg circuitul materie primã - produs finit. În diferite momente, relaţia aliment-mediu determinã modificãri care influenţeazã compoziţia şi salubritatea produsului destinat consumului uman. Produsele au o capacitate receptivã datoritã cãreia tot ceea ce mediul produce şi descarcã poate fi regãsit în acestea. Prin urmare, cele câteva perturbaţii ale mediului pot gãsi în alimente mijlocul ideal pentru a ajunge la om. Produsele alimentare vor avea o încãrcãturã minimã de poluanţi şi contaminanţi în mãsura în care agenţii de agresiune sunt cunoscuţi şi controlaţi în obţinerea, prelucrarea şi distribuţia fizicã a bunurilor.
Numeroasele substanţe toxice din alimente, care pot reduce sau chiar anihila inocuitatea acestora, pot fi sintetizate în urmãtoarele grupe:
substanţe antinutriţionale; substanţe toxice; produşi toxici; micotoxine; pesticide; metale şi metaloizi toxici; aditivi alimentari; microorganisme patogene.
Nominalizarea si poziţionarea corectã a tuturor poluanţilor şi contaminanţilor, precum şi a implicaţiilor acestora asupra organismului uman, impune lãrgirea orizontului valorii igienice a suplimentelor nutriţionale dincolo de inocuitatea clasicã (absenţa germenilor patogeni, a toxinelor bacteriene şi fungice, a unor otrãvuri şi contaminanţi chimici) a acestora. În present, ea trebuie sã aibã un conţinut complex care sã vizeze: absenţa sau limitarea strictã a unor poluanţi chimici generaţi de cãtre progresul tehnico-ştiinţific (pesticide, aditivi, antibiotice), a unor substanţe antinutriţionale sau perturbatoare ale metabolismului uman preexistente în diferite materii prime agroalimentare clasice, reconsiderarea poziţiei unor trofine energetice în plan cantitativ şi corelativ din punct de vedere igienic.
Standardizarea de produs soluţioneazã numai parţial şi indirect problema inocuitãţii alimentelor prin: admiterea într-un numãr foarte redus a compuşilor nocivi şi a impuritãţilor minerale, interzicerea prezenţei microorganismelor patogene şi a toxinelor acestora, admiterea unor bacterii condiţionat patogene într-un numãr foarte redus, dar fãrã a lua în evidenţã toate formele de contaminanţi ce pot interveni în fluxul tehnologic şi logistic normal.
Având în vedere toate aceste aspecte, definirea conceptualã a unor noţiuni cum sunt : puritatea, securitatea, siguranţa şi protecţia consumatorilor de suplimente nutriţionale capãtã o importanţã deosebitã.
Puritatea suplimentelor nutriţionale reprezintã gradul în care acestea sunt conforme cu specificaţiile de produs din punct de vedere al naturii şi proporţiei tuturor componentelor conţinute şi cu standardele în vigoare privind încadrarea în nivelurile maxime admise de contaminare fizicã, chimicã, microbiologicã, toxicologicã, radiologicã.
Pentru aceasta este necesar ca pe tot fluxul tehnologic sã se asigure un control strict din punct de vedere al apariţiei unor riscuri, sã se înregistreze valoarea parametrilor de lucru şi sã se pãstreze documente de control pentru fiecare lot fabricat. De asemenea, se impune ca producţia sã se realizeze din materii prime de calitate, garantate de furnizorul acestora prin buletine de analizã specifice, iar loturile de produse finite sã fie analizate în laboratoare acreditate conform specificaţiilor de produs şi cerinţelor normelor privind contaminarea microbiologicã, chimicã şi, în caz de necesitate, radioactivã. Comercializarea suplimentelor nutriţionale, în vederea asigurãrii puritãţii lor, impune ca acestea sã fie ambalate şi etichetate conform legislaţiei în vigoare, adicã în ambalaje originale şi cu eticheta în limba românã.
Securitatea consumatorului de suplimente nutriţionale se traduce prin cantitatea de alimente necesare unui individ, exprimate în unitãţi fizice, convenţionale (kilocalorii) şi în trofine (substanţe
51

nutritive), pentru a-şi asigura echilibrul fiziologic şi a-şi acoperi cele 3 raţii de consum: raţie de întreţinere; raţie de creştere; raţie de activitate.
Deşi i se conferã o sferã de cuprindere foarte amplã, totuşi, securitatea consumatorului de suplimente nutriţionale este, în primul rând, o problemã de nivel individual, a cãrei rezolvare este condiţionatã de disponibilitatea produselor, de posibilitatea de acces la acestea (determinatã, în general, de puterea de cumpãrare) şi de dorinţa de a avea o alimentaţie sãnãtoasã.
În acest sens, devine necesarã intervenþia autoritãtilor guvernamentale care, printr-o serie de mãsuri de politicã alimentarã şi nutriţionalã, pot contribui decisiv la realizarea securitãţii consumatorilor de suplimente nutriţionale.
Siguranţa consumatorilor de suplimente nutriţionale se referã la ansamblul mãsurilor necesare pentru a se asigura protecţia, stabilitatea şi salubritatea acestor produse, pornindu-se de la cultivare sau creştere, producţie ori fabricaţie şi pânã la consumul final.
Aceasta înseamnã cã în compoziţia suplimentelor nutriţionale nu trebuie sã se regãseascã elemente de naturã a afecta inocuitatea acestora: microorganisme patogene, substanţe chimice toxice (de exemplu, pesticide, metale grele, contaminanţi radioactivi), impuritãţi minerale neadmise sau a cãror proporţie se situeazã peste limitele maxime admise în standarde.În acest sens, producãtorii de alimente, trebuie sã comercializeze numai suplimentele nutriţionale care prezintã siguranţã, sunt salubre şi apte pentru consum, ei fiind obligaţi sã respecte parametrii privind proprietãţile organoleptice, fizice, chimice, microbiologice şi toxicologice ale alimentelor conform reglementãrilor legale în vigoare.
Astfel, producãtorii trebuie sã organizeze şi sã aplice controlul calitãţii igienice a suplimentelor nutriţionale, fabricate sau manipulate, în laboratoare proprii sau acreditate.
Protecţia consumatorilor de suplimente nutriţionale reprezintã ansamblul de mãsuri de ordin legislativ, administrativ şi financiar prin care acestora li se oferã încrederea cã pot beneficia de produse sigure, sãnãtoase şi de bunã calitate şi cã sunt protejaţi împotriva practicilor frauduloase de producere şi/sau comercializare a unor bunuri care nu respectã prevederile legale referitoare la securitatea şi sãnãtatea lor.
Altfel spus, o bunã protecţie a consumatorilor de suplimente nutriţionale nu se poate realiza în afara reglementãrii stricte a condiţiilor privind autorizarea unitãţilor care produc, prelucreazã, depoziteazã, transportã şi comercializeazã suplimente nutriţionale, a condiţiilor privind avizarea acestor tipuri de produse în scopul declarãrii lor ca fiind apte pentru consumul uman, respectiv a condiţiilor privind controlul unitãţilor care produc, prelucreazã, depoziteazã, transportã şi comercializeazã suplimente nutriţionale.
Cap.II PRODUSE PARABIOTICE
II.1 INTRODUCERE
Substanţele active biologic, administrate pentru a îmbunatăţi funcţionarea intestinală, pentru a regla compoziţia microflorei ca un tratament profilactic şi curativ al diferitelor afecţiuni infecţioase, pot fi clasificate în urmatoarele categorii:
Prebiotice Probiotice Simbiotice
Activitatea substanţelor probiotice si prebiotice constă în creşterea numărului bacteriilor producătoare de acid lactic, care sunt foarte importante pentru funcţionarea tractului gastrointestinal.
Probioticele
52

Probioticele sunt substanţe sau produse alimentare care conţin microorganisme vii: bacterii producătoare de acid lactic, bifido- şi lactobacterii, uneori drojdii, care, deasemenea, există în microflora intestinală normală a unei persoane sănătoase. Probioticele, ca substanţe, sunt frecvent folosite ca aditivi alimentari, mai ales în iaurturi şi alte produse lactate. Microorganismele existente în probiotice nu sunt patogene sau toxice, sunt în cantităţi adecvate; trecerea lor prin tractul gastrointestinal nu le modifică viabilitatea. Probioticele nu sunt văzute ca medicamente, ci ca substanţe benefice sănătăţii. Probioticele pot fi adăugate în produsele alimentare, sub forma aditivilor alimentari sau pot fi sub formă de tablete, conţinând bifido - şi lactobacterii sau combinaţii ale acestora. Scopul lor este normalizarea florei intestinale şi profilaxia îmbolnăvirilor.
Prebioticele
Prebioticele sunt mai ales alimente, care nu pot fi complet digerate, asigurând buna funcţionare a tubului digestiv şi îmbunătăţind starea generală a organismului. Aceast lucru se obţine prin creşterea, la nivelul tractului gastrointestinal, a uneia sau a mai multor grupe de microorganisme. Pentru a putea fi clasificat ca prebiotic, un component alimentar trebuie să nu poată fi digerat de către enzimele digestive umane. Ele furnizează substanţe nutritive selectiv, doar pentru grupe specifice de bacterii benefice, care trăiesc în intestinul gros. Alimentele care întrunesc aceste condiţii sunt carbohidraţii cu moleculă mică, cum ar fi inulina, lactuloza şi lactitol. Prebioticele sunt întâlnite în fulgii de porumb, terci, cicoare, pâine, usturoi, fasole, mazăre, anghinare, banane şi multe alte produse. Studiile au arătat o influenţă stimulatoare a carbohidraţilor simplii, mai ales cei conţinând fructoză, asupra bifido şi lactobacteriilor din intestinul gros.
Inulina - un polizaharid care poate fi gasit în tulpina şi rădăcina de dalie, anghinare, etc. Hidroliza lui produce fructoză. Au fost demonstrate: stimularea bifido şi lactobacteriilor, îmbunătăţirea absorbţiei de calciu care reduce riscul de osteoporoza, reglarea metabolismului lipidelor, reducerea riscului de arteroscleroză la nivelul sistemului cardiovascular.
Monozaharidele - incluzând glucoza, zaharoza, fructoza sau alte glicoproteine, care sunt principalele componente ale laptelui uman, sunt stimulanţi ai creşterii bifidobacteriilor.
Lactuloza - un dizaharid sintetic, care nu există în natură, în structura căruia fiecare moleculă de galactoză este legată de o molecula de fructoză. Lactuloza este transportată până la nivelul intestinului gros fără a fi modificată (doar aproximativ 0,25-2,00% este absorbită în intestinul subţire) şi constituie hrana perfectă pentru acele bacterii de care depinde funcţionalitatea adecvată a tractului gastrointestinal. Lactuloza a fost utilizată de mai bine de 40 de ani în pediatrie, pentru stimularea creşterii lactobacteriilor la nou-nascuţi. Metabolismul lactulozei produce acizi graşi cu molecula mică (acid lactic, butiric şi alţii) care determină scaderea pH-ului la nivelul intestinului gros. Utilizarea lactulozei ca sursă de carbohidraţi şi energie are ca efect creşterea cantitativă, în tubul digestiv, a bacteriilor benefice organismului, ceea ce duce la creşterea peristaltismului şi la reducerea produselor toxice ale catabolismului proteic ( amoniacului si altor compuşi cu azot). Amestecul de prebiotice şi probiotice formează grupul simbioticelor, substanţe benefice sănătătii, care stimulează în special creşterea şi activitatea lactobacteriilor şi bifidobacteriilor.
II.2 SCURT ISTORIC
Originea probioticelor datează din 1903 când în studiile lui Metchinikoff se descriu efectele benefice ale folosirii lactobacililor din iaurt de către om. Termenul de „probiotic” a fost folosit pentru prima dată în anul 1965 de către Lilly şi Stillwell, apoi de Parker în anul 1974 pentru a defini: „organismele sau substanţele care contribuie la echilibrul microbian al intestinului”. (Vanbelle şi colab., 1990)
În prezent, majoritatea cercetătorilor acceptă că probioticele sunt concentrate viabile şi atent selecţionate de bacterii acido – lactice, compuse adesea din tipuri de Lactobacillus acidophilus şi Streptococcus faecium, ori tipuri de Bacillus. Acestea sunt folosite pe scară largă în hrană, cu scopul de a preveni tulburările digestive sau pentru creşterea performanţelor zootehnice.
53

În alte studii, probioticele sunt definite ca „bacterii intestinale naturale care, după administrarea orală a dozelor, sunt capabile să se stabilească, eventual să se colonizeze în tractul digestiv şi să păstreze sau să determine creşterea florei naturale a tractului digestiv pentru a preveni colonizarea organismelor patogene şi pentru asigurarea inocuităţii hranei.
În ţările aparţinând CEE, în grupul produselor probiotice sunt incluse bacterii, drojdii, enzime şi alte substanţe cu rol probiotic, având la bază microorganisme aparţinând genurilor: Streptococcus, Lactobacillus, Bacillus, Aspergillus, Saccharomyces, Enterococcus, Pediococcus, enzime (lactoperoxidaza, glucanaza) şi extracte de rumen.
II.3 MICROFLORA INTESTINALǍ
Pe plan mondial, datorită interesului suscitat de problematica alimentelor probiotice, cercetările privind microflora probiotică implicată în procesul de sanogeneză au fost reluate şi continuate în mod intensiv, cunoscând noi direcţii de dezvoltare. Câteva produse probiotice de uz veterinar comercializate pe piaţa mondială sunt prezentate în Tabelul 2. Ţările producătoare de microorganisme sau produse probiotice sunt listate în Tabelele 3 .
Sistemul ecologic al microflorei intestinale a fost ilustrat de către Gedek în anul 1987 arătând că se găsesc circa 400-500 de tulpini microbiene care diferă în funcţie de specie , dar în general această microfloră poate fi clasificată în trei grupe şi anume
a. Grupul dominant (>90%, strict anaerob) ce însoţeşte flora subdominantă şi flora reziduală este compus din Bifidus bacteria, Lactobacili (Gram +) şi Bacteroidea (Gram -);
b. Microflora însoţitoare (<1% facultativ anaerobă) cuprinde alături de Escherichia coli (Gram -) şi Enterococi (Gram +);
c. Restul microflorei (<0,001%) este în principal compusă din Clostridia, Staphylococcus, Pseudomonas, Proteus şi drojdii ale genului Candida.
Studii la microscopul electronic au arătat că aproximativ 20% din tipurile de bacterii sunt încă în faza
de a fi identificate, obţinerea culturilor pure fiind greu de realizat. Colonizarea intestinală începe cu Escherichia coli, urmată de Streptococi, iar după câteva zile de Bifidobacterium, apoi Bacteroidea şi, în final, Lactobacillus, dar există mari diferenţe între unele specii de animale şi de om.
Starea de echilibru între diferitele specii bacteriene variază între duoden, ileon, colon şi cecum. Această variabilitate depinde de o mulţime de factori diferiţi ca: secreţiile digestive, pH gastric, secreţiile sărurilor biliare (motilitatea), integritatea mucoasei intestinale.
Microbiota din tractul intestinal uman cuprinde de asemenea diferite specii de microorganisme care pot fi grupate astfel:
- microbiota majoritară formată din Bifidobacterium(Bifidobacterium longum, infantis, brevis, bifidum) şi Bacteroides(Bact. Fragilis, vulgatus, distasonis, ovatus), care reprezintă 99% din microbiota totală. Bifodobacteriile sunt prezente în tractul digestiv de la naştere pe tot parcursul vieţii(Tabel 1)
- microbiota subdominantă formată din Lactobacillus (Lb. acidophilus , Lb. salivarius , Lb. fermenti), Enterococcus (Ec. Faecalis, Ec. Faecium) şi Enterobacteriaceae(E. coli)
- microbiota contaminantă care de fapt tranzitează tractul digestiv formată din specii de Bacillus , adesea sub formă sporulată, Staphilococcos, enterobacterii aparţinând genurilor: Klebsiella,Citrobacter, Proteus.
54

Tabel 1 Distributia bifidobacteriilor la om în funţie de vârsta
Segmentul de populaţie Specii minore Specii majore Sugari hrăniţi la sân Bifidobacterium bifidum Bifidobacterium longum,
infantis, breveSugari hrăniţi cu biberonul Bifidobacterium adolescentisCopii şi adolescenţi Bifidobacterium infantis,
breve, bifidum, longumAdulţi Bifidobacterium bifidum Bifidobacterium
adolescentis,longumPersone în vârsta Bifidobacterium
adolescentis,longum
La păsări, unde tranzitul este foarte scurt (2 – 5 ore), ecologia microbiană este direcţionată către microflora din guşă.
Cele mai întâlnite bacterii din tractul digestiv al porcilor şi păsărilor sunt listate în Tabelele 4 şi 5. (Vanbelle şi colab., 1990)
Tabelul 2. Produse probiotice comercializate pe piaţa mondială.
Produsul Conţine Recomandările producătorului Specia/Categoria beneficiară
Efecte
All – Lac Lactobacili Tineret porcin Reduce mortalitatea, creşte sporul mediu zilnic (smz)
Lacto – Sacc
Lactobacili, Saccharomyces sp.
Purcei Creşte smz, reduce incidenţa diareei şi mortalitatea
Toyocerin Bacillus toyoi (spori) Porci, păsări Reprimă dezvoltarea germenilor patogeni, atenuează stresul
Fralac – Streptococcus faecium Toate speciile Profilactic pentru diaree, promotor de creştere
55

Lbc Cocbactin Lactobacillus
acidophilus Tineret taurin Creşte smz
Oralin Enterococcus faecium Taurine, porcine, păsări
Îmbunătăţeşte performanţele productive
Protexin Streptococcus faecium, Streptococcus thermophilus, Lactobacillus acidophilus, Lactobacillus plantarum, Lactobacillus bulgaricus, Lactobacillus casei
Cabaline, ovine, porcine
Îmbunătăţeşte starea de sănătate şi performanţele productive
Probios Lactobacillus acidophilus, Lactobacillus plantarum, Lactobacillus casei
Viţei, porcine Creşte apetitul, producţia de lapte, reduce mortalitatea
Acosil – F.3
Orz încolţit, bacterii lactice
Vaci lapte, ovine şi scroafe gestante
Tabelul 3. Produse probiotice comercializate în Franţa.
Produsul probiotic
Microorganismele componente Destinaţia
Paciflor Bacillus cereus CIP 5832 Toate animalele Adjulact standard
Enterococcus sp., Lactobacillus helveticus, Lactobacillus acidophilus, Lactococcus lactis
Viţei, purcei
Adjulact 1000 Lactobacillus helveticus, Enterococcus sp. Viţei, purcei Adjulact 2000 Lactobacillus plantarum, Enterococcus sp. Viţei, purcei Yea – sacc Saccharomyces cerevisiae Poligastrice Fermacton Lactobacillus sp., Pediococcus sp., Enterococcus
faecium SF 368 Toate animalele
Bio – plus – porc
Lactobacillus sp., Pediococcus sp., Enterococcus faecium SF 368
Porci
Lyobacter P1 Lactobacillus plantarum, Lactobacillus rhamnosus, Enterococcus faecium
Toate animalele
Lyobacter SFL Enterococcus faecium SFL Toate animalele Multigerm Lactobacillus plantarum, Lactobacillus acidophilus,
Enterococcus faecium Porci
Biosaf SC 47 Saccharomyces cerevisiae SC 47 Toate animalele şi mai ales poligastrice
Bioplus 2B Bacillus licheniformis, Bacillus subtilis Toate animalele SF 68 Enterococcus faecium SF 68 Toate animalele Enteroferm Trei suşe de Lactobacillus, Enterococcus sp.,
Saccharomyces sp. Toate animalele
Degefermenti Lactobacillus acidophilus, Lactococcus lactis Toate animalele, probiotic, medicament
Bacteriolact Lactobacillus casei, Streptomyces thermophilus Probiotic, medicament, viţei, porci
56

Tabelul 4. Cele mai întâlnite bacterii din tractul digestiv al porcilor.
Nr. crt. Specia de microorganisme Nr. crt. Specia de microorganisme
1 Bacteroides ruminicola 11 Lactobacillus salivarius 2 Bacteroides uniformis 12 Peptostreptococcus productus 3 Bacteroides succinogenes 13 Proteus sp. 4 Butyrivibrio fibrisolvens 14 Ruminococcus flavefaciens 5 Clostridium perfringens 15 Selenomonas ruminantium 6 Escherichia coli 16 Streptococcus bovis 7 Eubacterium aerofaciens 17 Streptococcus equinus 8 Lactobacillus acidophilus 18 Streptococcus salivarius 9 Lactobacillus brevis 19 Veillonellae sp. 10 Lactobacillus cellobiosus 20 Lactobacillus fermentum
Tabelul 5. Cele mai întâlnite bacterii din tractul digestiv al păsărilor.
Nr. crt. Specia de microorganisme Nr. crt. Specia de microorganisme
1 Bacteroide sp. 9 Gemmiger formicilius
2 Bacteroides fragilis 10 Lactobacillus acidophilus
3 Bifidobacterium bifidus 11 Lactobacillus fermentum
4 Clostridium perfringens 12 Lactobacillus salivarius
5 Clostridium beijerinckii 13 Micrococcus sp.
6 Clostridium sp. 14 Streptococcus faecium
7 Eubacterium sp. 15 Streptococcus faecalis
8 Fusobacterium sp. 16 Ruminococcus obeum
II.3. MODUL DE ACTIUNE AL PROBIOTICELOR
În anul 1989 Raibaud şi Raynaud, au încercat să facă un rezumat privind mecanismul de acţiune al probioticelor, să explice creşterea performanţelor zootehnice şi îmbunătăţirea sănătăţii la om şi animale. Au fost emise o mulţime de ipoteze, asupra modului de acţiune a probioticelor, precum şi mai multe speculaţii dar, în principal trebuie avute în vedere următoarele:
Probioticele inhibă proliferarea bacteriilor patogene prin producerea de acizi organici şi prin reducerea pH;
Produc H2O2 şi previn aderarea bacteriilor patogene la peretele intestinului; Produc metaboliţi capabili să neutralizeze toxinele bacteriene „in situ” ori să inhibe producţia lor.
Datorită atenuării catabolismului intradigestiv, apare o reorientare a microflorei intestinale cu scopul reducerii absorbţiei substanţelor toxice cum ar fi NH3, amine, indoli precum şi diminuarea biotransformărilor sărurilor biliare şi ale acizilor graşi, în produşi toxici;
Prin enzimele proprii probioticele cresc utilizarea digestivă a hranei precum şi procesele de detoxifiere. Ameliorarea digestibilităţii alimentelor ingerate se realizează prin:
hidroliza lactozei de către β - galactozidază; degradarea β - glucanilor de către anumite suşe probiotice glucanolitice; stimularea activităţii enzimatice a microorganismelor endogene care permite o mai bună
asimilare a alimentelor; stimularea unor enzime propii celulelor epiteliale ale tractului digestiv (lactază, invertază,
maltază) Probioticele stimulează producţia de vitamine aparţinând în general grupului B şi
determină creşterea activităţii lactazei, invertazei şi maltazei; Se înmulţesc în tubul digestiv şi distrug bacteriile patogene;
57

Reduc catabolismul microbilor, tinzând către un echilibru mult mai bun între lactobacili; Stimulează imunitatea gazdei prin acţiune asupra celulelor implicate în imunitatea naturală şi în
imunitatea specifică. Se cunoaşte că fagocitoza este realizată, în principal, de macrofagi şi aceasta se constituie ca un
mecanism principal de apărare nespecific al organismului la pătrunderea unei substanţe străine. Probioticele stimulează deci activitatea macrofagilor.
Probioticele pot influenţa imunitatea nespecifică. Aceasta cuprinde două sisteme: un sistem care acţionează prin anticorpii secretaţi de limfocitele B (imunitate umorală) şi un alt sistem care acţionează prin intermediul direct al limfocitelor T (imunitate cu mediere celulară). Cele două sisteme comunică între ele prin intermediul substanţelor chimice denumite interleukine.
Creşterea răspunsului imunitar specific se traduce printr-o activare a limfocitelor T şi B ceea ce provoacă o creştere a nivelului de interleukine şi anticorpi circulanţi (imunoglobulinele M şi imunoglobulinele G).
Probioticele au şi efect asupra producţiei de anticorpi (în principal imunoglobulinele A) în lumenul intestinal. În contact cu antigenii prezenţi în conţinutul digestiv, imunoglobulinele A sunt foarte importante în tractul digestiv, constituindu-se ca o primă cale de apărare contra infecţiei. Imunoglobulinele A pot inhiba adeziunea bacteriilor patogene la suprafaţa mucoasei tractului digestiv prin:
- aglutinarea bacteriilor; - fixarea pe proteinele adezive, prezente la suprafaţa bacteriilor; - interferarea în complexul substanţe adezive/receptori celulari. Este important de amintit faptul privitor la adeziunea microorganismelor la animalele terestre unde
microbiologii deosebesc adeziunea nespecifică de adeziunea specifică. Cuvântul „adeziune” defineşte suprafaţa unei macromolecule, cunoscută a fi responsabilă de aderenţa specifică. Specificitatea adeziunii celulelor animale apare ca rezultat al compoziţiei chimice a structurilor responsabile de activitatea adezivă. Adezivii bacterieni în majoritatea lor sunt de natură proteică, în special la bacteriile Gram negative şi Gram pozitive. Aceste proteine de aderenţă, care diferă semnificativ între ele prin greutatea moleculară, compoziţia în aminoacizi şi structura cuaternară, se pare că recunosc şi leagă receptorii carbohidranţi de suprafaţa glicocalixului.
O a doua activitate de adeziune este exercitată de acizii lipoteicoici ai Streptococcus pyrogenes care s-a presupus că se ataşează pe membranele celulei gazdă prin intermediul resturilor de acizi graşi din acizi lipoteicoici prin interacţiuni hidrofobe.
Al treilea mecanism de adeziune specifică rezultă din depozitarea matricelor de polizaharide insolubile de către bacteria Streptococcus mutans, pe suprafaţa dinţilor, urmată de o lipire a polizaharidelor extracelulare la suprafaţa componentelor bacteriene.
In concluzie, multe activităţi adezive nu sunt cunoscute încă. Numărul şi puterea acestor interacţiuni variază de la caz la caz ca şi nivelul aderării unui microb la o varietate de suprafeţe. Toate acestea nu implică specificitate.
Numeroase studii de microscopie electronică au demonstrat claritatea apariţiei aderării, colonizarii ori asocierii bacteriilor în diferite părţi ale intestinului la o varietate de animale vertebrate, cum ar fi: şobolani, şoareci, porci, păsări, rumegătoare şi oameni. (Chan şi colab., 1984)
Referitor la colonizarea microbiană, conceptul iniţial a fost că probioticele trebuie să colonizeze mucoasa şi să se multiplice “in situ” în scopul de a produce substanţe ori metaboliţi în beneficiu lor. Faptul că numărul microorganismelor probiotice viabile este constant în fazele respective nu este suficient pentru a încheia colonizarea. Într-adevăr înmulţirea bacteriilor probiotice în timpul tranzitului lor în tubul digestiv este câteodată foarte limitată şi nu suficientă. Mulţi autori pretind că bacteria acidului lactic folosită ca probiotic aderă la celulele marginale cu cili intestinali. Această proprietate ar trebui să permită microorganismelor să prolifereze “in situ”. Însă datele experimentale arată că nu este o condiţie „sine qua non”. (Lee, 1980)
La şobolani şi şoareci holoxenici, dominantul Lactobacillus murinus nu aderă la celulele stomacale. Câteva tipuri de lactobacili aderă bine la celulele stomacale ale şobolanilor şi şoarecilor holoxenici, dar nu aderă la pereţii celulari ai puilor.
Probioticele inhibă aderarea enterobacteriilor patogene (E. coli)care scad considerabil numărul celulelor ce conţin imunoglobina A.
58

Locurile de aderare pentru Lactobacili şi E. coli nu sunt aceleaşi iar mecanismul exact de acţiune se referă fie la controlul direct al microorganismelor, fie la acţiunea indirectă prin stimularea microflorei autohtone şi a metabolismului lor. Imediat după naştere se pot implementa tipuri aparţinând microflorei autohtone normale. Este interesant a relata cercetările franceze recente efectuate pe copii cu Bifidobacterium longum, o bacterie anaerobă (Gram pozitivă) şi care este un locuitor normal al intestinului copiilor, adulţilor şi animalelor. Laptele fermentat de Bifidobacterium longum a dat rezultate mult mai bune la copii împotriva diareii, consecutiv folosirii antibioticului eritromicină decât iaurtul; mai mult decât atât laptele fermentat este activ împotriva infecţiei cu Rotavirus. Cel mai relevant fapt al acestui experiment este în primul rând acela că are o activitate pozitivă în reducerea Escherichia coli şi Clostridium perfringens, prin metaboliţii (glicoproteine) formaţi în timpul fermentării laptelui cu Bifidobacterium longum.
Ramond (1989) a relatat că tipurile de Bifidobacterium aderă la mucoasă şi la receptorii de β - glucosamină la fel ca şi unele tipuri patogene de E. coli. Se pare că Bifidobacteriumlongum este dificil de implementat, dacă numărul de germeni autohtoni este de 1011/g.
Aceste experimente arată clar că importanţă au atât viabilitatea şi natura receptorilor, cât şi unii metaboliţi formaţi din fracţiunea de β - cazeină în timpul fermentării laptelui, care pot acţiona împotriva bacteriilor patogene şi a câtorva infecţii virale. Lactobacili viabili din iaurt au şase proprietăţi (Antoine, 1989):
- supravieţuiesc în timpul tranzitului digestiv; - dezvoltă activitatea antidiareică la copii şi adulţi; - digeră lactoza; - desfăşoară o activitate probiotică la animale; - sunt capabili de a detoxifia prin reducerea activităţii nitroreductazei glucoronidazei şi
azotoreductazei; - administrarea iaurtului şi lactobacililor creşte nivelul interferonului la oameni.
Câteva tipuri de Bacillus licheniformis produc un antibiotic la şoriceii monoxenici care inhibă puternic Clostridium perfringens.
Nu a fost dovedit „in situ”, dacă probioticele produc antitoxine. Există o creştere a utilizării digestive a hranei, care a fost verificată pe şoricei şi oameni în experimente cu iaurt şi Saccharomyces boulardi.
Aceasta înseamnă că celulele vii de microorganisme, în timpul tranzitului intestinal pot modifica producţia de enzime în celulele marginale cu cili. Probioticele stimulează sistemul imunitar nespecific al animalelor. Există câteva indicaţii clare că la şoricei holoxenici ingerarea iaurtului proaspăt creşte nivelul imunoglobinei 2A sau creşte activitatea fagocitară şi limfolitică. La oameni, iaurtul induce de asemenea o creştere uşoară de γ - interferoni.
Concluzii: Referitor la modul de acţiune a probioticelor, au fost emise o mulţime de ipoteze, şi mai multe
speculaţii dar, în principal trebuie avute în vedere următoarele:
- Probioticele inhibă proliferarea bacteriilor patogene prin producerea de acizi organici şi prin reducerea pH;
- Produc H2O2 şi previn aderarea bacteriilor patogene la peretele intestinului; - Prin enzimele proprii probioticele măresc utilizarea digestivă a hranei precum şi procesele de
detoxifiere; - Probioticele stimulează producţia sistemului imunologic ca barieră de protecţie a tubului digestiv, fiind
demonstrat că administrarea de preparate cu Lactobacillus poate stimula producţia de γ - globuline, γ - interferoni şi intensifică activitatea macrofagelor responsabile de îndepărtarea agenţilor patogeni din organism;
- Probioticele stimulează producţia de vitamine aparţinând în general grupului B şi determină creşterea activităţii lactazei, sucrazei şi maltazei;
- Proliferează în tractul digestiv fiind în competiţie cu bacteriile patogene;
Utilizarea probioticelor în hrana animalelor are efecte benefice în următoarele activităţi: - Tulburări ale microflorei indigene: - disbacterioze neinfecţioase după doze mari cu antibiotice;
59

- stress (schimbări în alimentaţie, transport); - Dezvoltarea deficientă a microflorei; - Promovarea creşterii; - conversie mai înaltă a hranei; - sinteza vitaminelor; - predigestia proteinelor.
În plus, se pare că probioticele ar avea oarecare potenţial în tratamentul şi prevenirea bolilor coronariene şi encefalopatiilor, fiind demonstrat că administrarea de probiotice poate reduce nivelul colesterolului plasmatic la pui. De asemenea s-a sugerat că probioticele pot fi eficiente în combaterea unor forme de cancer, atât pe calea acţiunii sistemului imunitar cât şi prin inhibarea unor enzime ca β - glucoronidaza, β - glucozidaza şi nitroreductaza, care sunt implicate în carcinogeneza intestinală.
CARACTERISTICILE MICROORGANISMELOR PROBIOTICE
Pentru a putea fi încadrate în categoria microorganismelor probiotice, acestea trebuie să îndeplinească următoarele condiţii: - să producă substanţe utile organismului; - să facă parte sau să influenţeze favorabil flora digestivă normală a organismului; - să se regenereze şi să se dezvolte într-un timp cât mai scurt; - să colonizeze un timp cât mai îndelungat tractul digestiv; - să aibă stabilitate ridicată şi să poată fi obţinut cu uşurinţă; - ar fi de dorit ca stabilitatea microorganismelor probiotice să fie permanenta adică să rămână la
concentraţia corespunzătoare după prima administrare; în practică dacă se întrerupe pentru o anumită perioadă de timp administrarea probioticului, efectul benefic dispare.
Cele mai utilizate microorganisme pentru producerea probioticelor sunt: • Lactobacillus: acidophilus, bulgaricus, casei, helveticus, lactis, salivarium, plantarum • Streptococcus: termophylus, faecium, fecalis Dintre toate speciile amintite doar Streptococus termophylus şi Lactobacillus bulgaricus nu fac parte dintre microorganismele tubului digestiv.
II.4. BIOMASA DIN DROJDII CU ROL PROBIOTIC
În prezent cea mai utilizată tulpină de drojdie sub formă de produs probiotic este Saccharomyces cerevisiae. Sursa de carbon utilizată în procesul biotehnologic de obţinere de biomasă viabilă este etanolul, deoarece este disoponibil ca materie primă pură, este acceptabil ca ingredient alimentar şi este complet solubil în apă. Fiind un proces aerob, obţinerea biomasei viabile din drojdii pe etanol se realizează în condiţii aseptice, la concentraţii limitative de substrat şi la o rată înaltă de creştere, care asigură un randament mare de bioconversie în biomasă.
Cercetările au arătat că ingestia de Saccaromyces cerevisiae are efect protector asupra florei microbiene intestinale, inhibând dezvoltarea bacteriilor patogene, stimulează imunitatea organismului şi echilibrează flora saprofită intestinală, după tratamentul cu antibiotice şi chimioterapice.
Episoadele uşoare sau severe de diaree constituie principalul efect al tratamentului cu antibiotice. Forma cea mai frecventă a deranjamentelor intestinale este colita pseudomembranoasă. Printre căile ecologice de control ale acestor diarei ce rezultă din dezechilibrarea microflorei intestinale se numără mai multe microorganisme nepatogene care pot fi folosite pentru restabilirea echilibrului. Dintre microorganismele existente, Saccharomyces boulardii a fost cea mai studiată.(Bergogne-Bérézin, 1995)
Pe lângă datele obţinute din cercetări „in vitro”, experienţele efectuate pe cobai (infectaţi cu Clostridium difficile) au confirmat potenţialul lui S. boulardii în prevenirea efectelor patogene ale toxinelor A şi B(secretate de Clostridium difficele) şi în scăderea numărului de colonii ale microorganismelor patogene. În mod similar în cazurile pacienţilor cu manifestări grave de diaree, au fost administrate doze de S. boulardii, care au prevenit diaree şi colita pseudomembranoasă, determinând decontaminarea selectivă a tractului digestiv. (Corbon şi colab., 1995)
Efectele nedorite ale antibioticelor sunt legate de toxicitatea directă a anumitor produse asupra unor organe (ex. nefrotoxicitate, neurotoxicitate, hepatotoxicitate), de efectele farmacologice şi
60

metabolice (interacţiunea metabolică a macrofagelor cu teofilina) şi de efectele imunologice (alergii la penicilină şi alte antibiotice).
Printre efectele secundare ale antibioterapiei cele mai frecvente sunt accidentele digestive (prin diverse tipuri de molecule) de tipul diareelor de intensitate şi gravitate variabilă, care cedează în urma tratamentului. Unele forme mult mai severe necesită spitalizare, cum ar fi colitele pseudomembranoase asociate cu Clostridium difficile.
Practic, toate familiile de antibiotice provoacă, diarei de intensităţi variabile, în funcţie de modul de acţiune şi de spectrul de influenţă. Datorită stresului, al antibioterapiei sau al tratamentelor imunosupresoare, la pacienţii cu disfuncţii grave se constată anomalii profunde ale microflorei digestive. În acest context, o patologie severă asociată cu ingestia unor substanţe, este colita pseudomembranoasă în timpul căreia multiplicarea bacteriei Clostridium difficile este însoţită de producerea simultană a două toxine – o enterotoxină (A) şi o citotoxină (B) responsabile de leziunile stomacale şi de simptome severe.
Tratamentele specifice pentru eradierea eventualilor agenţi patogeni şi măsurile indispensabile de corecţie a disfuncţiilor electrolitice s-au propus a se asocia cu aportul de microorganisme nepatogene pentru restabilirea – cel puţin parţială – a unui nou echilibru intestinal.
III ASPECTE ALE UTILIZĂRII PROBIOTICELOR LA FABRICAREA PRODUSELOR LACTATE ACIDE
III.1 INTRODUCERE Timp de secole s-a considerat că prin consumul de produse lactate acide se îmbunătăţeşte starea de
sănătate, ceea ce ar conduce la creşterea longevităţii umane. În secolul XX, Eli Metchnikoff, câştigătorul Premiului „Nobel”pentru biologie, a furnizat un suport ştiinţific pentru afirmaţia anterioară. Studiul său a sugerat că lactobacilii ar putea contracara efectele putrefactive ale metabolismului gastrointestinal. De atunci, mai mulţi cercetători au abordat studiul efectelor consumului de produse lactate acide, şi în special al metabolismului bacteriilor lactice. Termenul „probiotic” se referă la un grup de microorganisme cu acţiune benefică asupra sănătăţii umane. În S.U.A., bacteriile probiotice includ bacterii lactice din speciile Lactobacillus şi Bifidobacterium şi sunt definite ca „organisme vii care, consumate într-o anumită cantitate, determină beneficii asupra sănătăţii, alături de beneficii nutriţionale”.Tendinţa consumatorilor a migrat de la alimente lipsite de grăsimi, cu un conţinut scăzut de sare şi colesterol, spre alimente „funcţionale” fortifiante şi suplimentate cu anumite ingrediente. Deoarece mulţi producători au ezitat să fabrice alimente funcţionale datorită reglementărilor severe, cei mai mulţi consumatori au apelat la suplimente nutriţionale.
Traiectul gastrointestinal uman reprezintă un ecosistem complex, care conţine, în mod natural, circa 400 de specii de bacterii. Mecanismele care reglează numărul, tipul şi activitatea metabolică a acestor tipuri de bacterii sunt deosebit de complexe. Totuşi, unele bacterii (din genul Lactobacillus şi Bifidobacterium) au o acţiune de prevenire faţă de bacteriile patogene. Ambele specii de bacterii Lactobacillus şi Bifidobacterium se regăsesc în cadrul florei ce populează traiectul gastrointestinal (cele din genul Bifidobacterium predomină în timpul copilăriei), dar numărul acestora se diminuează cu vârsta, consumul de alcool, administrarea de antibiotice, stresul şi alţi factori.Pentrua sprijini activitatea bacteriilor din flora normală, consumatorii pot introduce în traiectul gastrointestinal (prin consum oral) probioticele, întărind lupta organismului asupra potenţialelor bacterii dăunătoare. Activitatea metabolică a probioticelor poate influenţa pozitiv asupra raportului bacterii nedăunătoare/bacterii dăunătoare. S-a constatat o multitudine de efecte benefice asupra sănătăţii umane în urma consumului de bacterii probiotice.
Îmbunătăţirea digestiei lactozei la persoanele cu deficient de lactază şi prevenirea unor tipuri de boală diareică sunt două dintre cele mai cunoscute efecte ale consumului de bacterii probiotice. Pentru alte beneficii ale ingestiei de probiotice se aşteaptă confirmarea ştiinţifică. Cercetătorii au arătat că efectele benefice asupra sănătăţii variază în funcţie de cantitatea de probiotice ingerată şi de durata de
61

administrare. Mecanismele prin care probioticele influenţează microbiota traiectului gastrointestinal sunt considerate încă insuficient elucidate, dar s-au observat cel puţin patru mecanisme de acţiune: 1. Agenţii antibacterieni, incluzând acizii organici (cum ar fi acidul lactic şi acidul acetic) şi proteinele (bacteriocine), sunt produşi de bacteriile probiotice şi pot avea efect inhibitor asupra microflorei patogene din traiectul gastro-intestinal; 2. Laptele uman, care conţine componente bioactivatoare stimulează creşterea bifidobacteriilor din traiectul gastrointestinal al copiilor, poate modifica antagonismul bacterian şi poate împiedica creşterea bacteriilor aerobe nedorite; 3. Modularea sistemului imunitar (anticorpi, celule T, interferon) poate suprima activitatea patogenilor sau carcinogenilor;4. Bacteriile probiotice stimulează producţia lactazei în intestinul subţire, de aceea digestia lactozei este îmbunătăţită şi se previne utilizarea lactozei în colon.
Totuşi, mecanismul de acţiune al probioticelor nu este pe deplin elucidat, in condiţiile in care o mică parte din cantitatea de probiotice ingerată rămâne vie. Pentru a rămâne în viaţă, probioticele trebuie să reziste acţiunii acidităţii din stomac, să treacă de intestinul subţire ca, mai apoi să ajungă în colon. Cercetările au arătat că bacteriile probiotice nu aderă la peretele intestinal, ci ele se dezvoltă pe parcursul traiectului gastrointestinal. De aceea, se consideră că un consum zilnic de bacterii probiotice este, probabil, cea mai indicată cale de a menţine activitatea benefică a acestora.
III.2. CERCETĂRI ACTUALE ASUPRA MODULUI DE ACŢIUNE A PROBIOTICELOR DIN PRODUSELE LACTATE
Cercetătorii şi-au concentrat eforturile pentru investigarea şi caracterizarea componentelor produse de către culturile probiotice şi modul în care condiţiile de parcurgere a traiectului gastrointestinal afectează activitatea şi dezvoltarea probioticelor. O serie de cercetări au vizat bacteriocinele, care sunt peptide cu masă moleculară mică produse de către unele bacterii şi care exercită un efect bactericid asupra altor specii de bacterii.
Unii cercetători au studiat efectul consumului de iaurt fabricat cu ajutorul culturilor probiotice asupra traiectului gastrointestinal. S-a urmărit să se demonstreze modul în care este crescută rezistenţa la cancer şi infecţii după consumul de iaurt cu specii de Bifidobacterium. Cercetările au urmărit două aspecte: izolarea limfocitelor de la cobaii hrăniţi cu iaurt, identificarea şi urmărirea abilităţii limfocitelor de a secreta hormoni care să producă anticorpi. Cel de al doilea aspect a vizat creşterea limfocitelor direct pe culturile starter de iaurt şi cuantificarea funcţiilor imune ale componentelor rezultate.
Se consideră că există un efect sinergetic între componentele produselor lactate şi culturile probiotice, iar produsele lactate reprezintă un suport ideal pentru introducerea probioticelor în sistemul gastrointestinal.
Cercetările au confirmat abilitatea lactoferinei de a îmbunătăţi procesul de creştere al bifidobacteriilor, în mod direct prin mărirea numărului de celule vii şi indirect, prin reducerea bacteriilor intestinale producătoare de toxine.
Un criteriu esenţial în alegerea probioticelor pentru obţinerea produselor alimentare este acela de abilitate a microorganismelor de a supravieţui în timpul depozitării, în aşa fel încât să se asigure că vor ajunge în intestin celule viabile. Cel mai utilizat produs lactat cu probiotice este iaurtul care are un pH relativ scăzut. Abilitatea de creştere şi supravieţuire a diferitelor specii de probiotice depinde în mare măsură de nivelul de aciditate din mediu. În mod curent, au fost selectate pentru utilizare în produsele lactate acele specii de probiotice care să poată supravieţui în condiţiile de fabricare, depozitare şi distribuţie. Unii cercetători au dezvoltat o tehnică de îmbunătăţire a bifidobacteriilor în produsele lactate. Tehnologia implică capturarea bifidobacteriilor în gel de alginat de calciu.
După stabilirea ştiinţifică a efectelor consumului de culturi probiotice, utilizând produsele lactate ca suport, este necesar să fie identificate care dintre beneficiile asupra sănătăţii are un impact mai mare asupra consumatorilor.
La nivel mondial s-a creat o bază de date în care sunt indicate performanţele şi funcţionalitatea culturilor de probiotice în produsele lactate. Aceasta urmăreşte să furnizeze producătorilor informaţii despre culturile probiotice, în vederea selecţionării corecte a speciei de culturi indicate pentru un anumit produs lactat. Bacteriile probiotice se pot utiliza pentru o largă varietate de produse lactate, cum ar fi laptele, laptele acidofil, iaurtul şi brânza „Cottage”.
62

Dintre atributele ce pot fi selectate în baza de date, se pot enumera: activitatea lactazică, rezistenţa la antibiotice abilitatea de a creşte în prezenţa fructo-oligozaharidelor rezistenţa la aciditate, producţia inhibitorilor microbieni
Este demn de remarcat faptul că orice probiotic selectat pentru efectele sale asupra sănătăţii poate să nu fie un bun candidat pentru producţia industrială. În general, producătorii adaugă cultura de probiotice după pasteurizare şi înainte de ambalare. Este necesar să fie asigurate condiţii de sterilitate şi prevenirea contaminării produsului. Timpul şi temperatura de depozitare pot afecta de asemenea viabilitatea celulelor probiotice.La crearea unor noi produse lactate probiotice trebuie să se ţină seama de consideraţiile anterioare. Culturile de probiotice nu trebuie selectate doar pentru activitatea lor probiotică, ci şi în funcţie de adaptabilitatea la condiţiile de fabricare şi distribuţie.
III.3 PRODUSE DE TIP PROBIOTIC
Probioticele pentru consum uman pot fi de mai multe tipuri şi o parte dintre ele sunt prezentate în continuare.
PRODUSE TIP BABY-FOOD
Având în vedere că bifidobacteriile sunt sunt pseudodominante în tractul intestinal al sugarilor, utilizarea lor în produse probiotice este justificată de aspectele benefice aduse de acestea :
- competiţia antagonică cu bacteriile patogene- inhibarea reducerii NaNO3 la NaNO2
- îmbunătăţirea retenţiei azotului şi a creşterii în greutate a sugarilor - protecţia faţă de infecţiile enterice şi faţă de efectele secundare ale antibioticelor - ajutor în terapia tulburărilor intestinale
Produsele baby-food care folosesc bifodobacterii sunt: LACTANA B (Germania) sub formă de pulbere , care conţine Bifidobacterium bifidum, celule
viabile şi lactuloză(substanţă prebiotică0 FEMILACT (Cehia) care se obţine prin fermentarea unei smântâni slabe(12% grăsime) cu o
cultura mixtă formată din : Bifidobacterium bifidum , Lactobacillus acidophilus , Pediococcus acidilacti, în raport 1: 0.1 : 1 , la temperatura de 30 o C , până la aciditatea dorită , urmată de racire. In continuare se adaugă ulei vegetal , lactoză, proteine din zer şi vitamine , apoi amestecul se omogenizează şi se usucă prin pulverizare. Produsul finit , după reconstituire, conţine 0.25 % acid lactic, 10 8 - 10 9 bacterii viabile/ml .Dupa o depozitare timp de doua luni , numărul bacteriilor viabile se reduce cu un factor de 10.
PRODUSE LACTATE FERMENTATE DE TIP PROBIOTIC PENTRU COPII SI ADULTI
LAPTELE ACIDOFIL PENTRU COPIIProdusele care conţin Lactobacillus acidophilus sunt importante pentru că microorganismul menţine
balanţa microbiotei intestinale, având capacitate de supravieţuire în tractul intestinal.Trebuie avut în vedere că Lactobacillus acidophilus scade numeric dacă pH-ul mediului este mai mic de 5.
Laptele acidofil destinat copiilor se prepară din lapte pasteurizat la 95 o C/10 min , răcit la 37 o şi menţinut la această temperatură timp de 3-4 ore pentru germinarea sporilor, după care urmează o nouă pasteurizare /răcire şi inocularea cu 2-5 % cultură de producţie probiotică. Produsul are pH= 5.5-6.0 şi conţine un număr de aproximativ 3*10 9 bacterii viabile/ml. Dapă fermentare produsul este răcit la 5-10 o
C şi este rapid distribuit pentru că la depozitare numărul de celule viabile de Lactobacillus acidophilus se reduce .
63

LAPTELE ACIDOFIL DULCE Se prepară din lapte pasteurizat şi răcit, la care se adaugă un concentrat de Lactobacillus acidophilus ,
astfel încât să se obţina o concentraţie de celule vii de 5-10 6/ml. Produsul se depozitează la 5 o C , la această temperatură neavond loc fermentarea deci produsul rămâne dulce.
LAPTELE BIFIDUS Laptele bifidus tip 1
Tehnologia de preparare include :- standardizarea laptelui la conţinutul de grăsime dorit- standardizarea la concentraţia proteică dorită- omogenizarea- tratamentul termic la 85-120 o C timp de 5-30 min- inocularea cu 10 % cultură starter probiotică - termostatarea la 37 o C pâna la coagulare- răcirea
Cultura starter este formată din Bifidobacterium bifidum. Produsul finit poate fi prezentat ca un coagul sau sub formă de produs agitat(omogenizat0, anond pH-ul=4.3-4.7 şi un număr de 10 8- 10 9/ml. Acest număr scade după o depozitare de 1-2 săptămâni.
Se poate obţine şi un lapte bifidus cu gust de iaurt , prin fermentarea zerului şi a laptelui degresat îmbogăţit cu proteine , cu o cultură de Bifidobacterium bifidum, Bifidobacterium longum şi Bifidobacterium infantis. Produsul se obţine dintr-un amestec rezultat la ultrafiltrarea zerului dulce (factor de concentrare =8) cu lapte degresat ultrafiltrat (factor de concentrare=20 , amestec care se pasteurizează la 80 o C/30 min, este racit la 37 o C, îmbogăţit cu treonină 0.15, inoculat cu 2% cultură starter probiotică , apoi este termostatat la 37 o C/ 24 ore şi răcit la 4 o C.
Produsul finit are 15% substantă uscată , incluzând 7.3 % proteină şi 1.3 % grăsime , un pH= 4.7 , un nivel de acetaldehidă de 29-39 ppm.Produsul conţine de asemenea un număr de 10 9 celule viabile /ml. Acest număr scade după o depozitare de 21 zile la 10 7 celule viabile /ml.
Bibilografie : 1. Banu, C.- Biotehnologii in industria alimentară, Editura Tehnică, Bucureşti,1987 2. Banu, C.- Biotehnologii in industria alimentară, Editura Tehnică, Bucureşti,2000
64





























