Embed Size (px)
Citation preview
S U N A R N O
FISIOLOGI hewan

FISIOLOGI HEWAN


FISIOLOGI HEWAN
SUNARNO
MADINA2016

Hak Cipta 2016, pada penulis
Dilarang mengutip sebagian atau seluruh isi bukuini dengan cara apapun,Termasuk dengan cara penggunaan mesinfotokopi, tanpa izin sah dari penerbit
Cetakan 2016-1
2016.01 MDMDr. Sunarno, S.Si., M.SiFISIOLOGI HEWAN
Hak Penerbit pada CV Madina, Semarang
Desain cover dan layout oleh Saifur Rijal
Dicetak di Pinter Berkah Berjaya
CV MADINAKantor Pusat� � : Jl. Bulusan XI/5� � � Perum Korpri Tembalang SemarangTel/Fax� � : (024) 76482660Email� � � : [email protected]

Segala puji bagi Allah Tuhan Yang Maha Kuasa yang telah
memberikan bimbingan dan hidayah sehingga Buku Fisiologi dapat
diselesaikan. Buku ini memuat materi tentang metabolisme
karbohidrat, metabolisme lemak, metabolisme protein, dan siklus
Kreb dengan pembahasan yang komprehenship. Semoga buku ini
dapat digunakan untuk melengkapi referensi yang sudah ada dan
sebagai acuan bagi mahasiswa yang ingin mengembangkan keilmuan
bidang Fisiologi.
Kritik dan saran sangat penulis harapkan untuk perbaikan buku
ini. Akhirnya penulis berharap semoga buku ini dapat bermanfaat
sebesar-besarnya bagi pembaca sekalian.
Tahun 2016
Penulis
KATA PENGANTAR


Halaman JudulKata PengantarDaftar Isi
BAB I. Metabolisme Karbohidrat 1.1 Sistem Energi 1.2 Penggunaan dan Jalur Energi Pakan 1.3 Karbohidrat 1.4 Fungsi Karbohidrat 1.5 Karbohidrat Sebagai Sumber Energi Paling Penting Bagi Tubuh 1.6 Klasifikasi dan Sumber Karbohidrat 1.7 Jalur Pemrosesan Karbohidrat Di Dalam Tubuh 1.8 Pencernaan Karbohidrat 1.9 Pencernaan dan Metabolisme Karbohidrat 1.10 Pencernaan dan Metabolisme Karbohidrat pada Ruminansia 1.11 Kontrol Metabolisme Karbohidrat 1.12 Defisiensi Karbohidrat 1.13 Hubungan Konsumsi Karbohidrat dengan Karier Dentis
DAFTAR ISI
vvi
1245
68
222424
315356
58

1.14 Soal – Soal Latihan
BAB II. Metabolisme Lemak 2.1 Pengertian Lemak 2.2 Klasifikasi dan Nomenklatur Asam Lemak 2.3 Asam Lemak Esensial 2.4 Lemak Netral 2.5 Sifat Fisik Lemak dan Reaksi Trigliserida 2.6 Fungsi Lipid 2.7 Lemak Majemuk dan Lipid Turunan 2.8 Aktivitas Vitamin D 2.9 Pencernaan, Absorbsi, Transportasi, dan Metabolisme Lemak 2.10 Jalur Pengangkutan Lemak Dalam Darah 2.11 Oksidasi Asam Lemak 2.12 Transport Asam Lemak Ke Dalam Matriks Mitokondria 2.13 Oksidasi Asam Lemak Jenuh: Asetil CoA, NADH, FADH Terbentuk Pada Setiap Kali2
Oksidasi 2.14 Oksidasi Sempurna Asam Palmitat 2.15 Oksidasi Asam Lemak Tak Jenuh 2.16 Oksidasi Lemak Dengan Nomor Atom Karbon Ganjil 2.17 Proses Ketogenesis 2.18 Asetoasetat adalah Merupakan Salah Satu Bahan Bakar Yang Utama Dalam Jaringan 2.19 Biosintesis Asam Lemak 2.20 Metabolisme Lemak Pada Hewan Ruminansia 2.21 Faktor – Faktor Yang Mempengaruhi Produksi Volatile Fat Acid Dalam Rumen 2.22 Kelainan Lemak dan Diagnosis Kelainan Lemak 2.23 Klasifikasi Klinis Hiperlipidemia 2.24 Gejala Hiperlipidemia 2.25 Patofisiologi Penyakit Jantung Koroner 2.26 Soal – Soal Latihan
viiidaftar isi
58
6167697071727276
767982
83
88899091
9294
95103107
108109110111113

BAB III. Metabolisme Protein 3.1 Pengertian Protein 3.2 Pencernaan Protein 3.3 Absorbsi dan Transportasi Asam-Asam Amino 3.4 Metabolisme Protein dan Asam-Asam Amino 3.5 Katabolisme Asam Amino yang Mengandung Nitrogen 3.6 Metabolisme Protein pada Hewan Poligastrik (Ruminansia) 3.7 Jalur Metabolisme Amonia 3.8 Soal-Soal Latihan
BAB IV. Siklus Kreb 4.1 Pengertian Siklus Kreb 4.2 Reaksi Siklus Kreb 4.3 Pembentukan dan Oksidasi pada Isositrat 4.4 Pembentukan dan Penguraian α-Ketoglutarat menjadi 4.5 Pembentukan GTP 4.6 Oksidasi Suksinat menjadi Oksaloasetat 4.7 Koenzim pada Siklus Kreb 4.8 Regulasi pada Siklus Kreb 4.9 Prekursor pada Asetil KoA 4.10 Senyawa Intermidiet pada Siklus Kreb dan Reaksi Anaplerotik 4.11 Soal-Soal Latihan
Daftar PustakaGlosarium
115121124125130
136
144145
147152153
154154155156161164168
171
173179
fisiologi hewanix


1.1 Sistem Energi
Setiap gerak molekul selalu membutuhkan energi. Energi
diperoleh melalui makanan. Energi diperlukan secara berkelanjutan
untuk mendukung proses-proses yang terjadi di dalam tubuh supaya
makhluk hidup tetap mampu bertahan hidup. Energi dikaitkan dengan
kemampuan makhluk hidup untuk melakukan kerja. Energi tidak
dapat diciptakan atau dimusnahkan, namun energi dapat berubah
bentuk. Beberapa bentuk energi, antara lain energi kimia, mekanik,
cahaya dan lain-lain. Energi yang dihasilkan dari proses pemecahan
makanan digunakan untuk menghasilkan senyawa kimia yang
dikenal dengan nama adenosin triphosfat (ATP) melalui proses
respirasi seluler, kebalikan dari proses fotosintesis. Energi yang
dihasilkan dapat disimpan sebagai cadangan sumber energi di dalam
sel-sel otot. Energi yang dihasilkan saat hidrolisis ATP merupakan
sumber energi yang dapat digunakan oleh s el-sel otot untuk
melakukan kerja.
METABOLISME KARBOHIDRAT
1

1.2 Penggunaan dan Jalur Energi Pakan
Energi tersimpan di dalam karbohidrat, lemak, dan protein dari
bahan makanan. Semua bahan tersebut mengandung karbon (C) dan
hidrogen (H) dalam bentuk yang dapat dioksidasi menjadi
karbondioksida (CO ) dan molekul air (H O) yang menunjukan 2 2
energi potensial untuk hewan. Jumlah energi panas yang diproduksi
ketika pakan dioksidasi secara sempurna oleh oksigen dapat diukur
dengan alat ”bomb calorimeter” dan disebut Energi Bruto (EB) dari
pakan. Prinsip dari pengukuran EB pakan adalah konversi energi
dalam pakan (karbohidrat, lemak, protein) menjadi energi panas
dengan cara oksidasi zat makanan tersebut melalui pembakaran.
Bomb calorimeter dapat digunakan untuk mengukur energi bruto dari
pakan secara utuh (whole food) atau dari bagian-bagian pakan
(misalnya glukosa, pati, selulosa), jaringan hewan dan ekskreta
(feses, urin). Nilai energi bruto dari suatu bahan pakan tergantung dari
proporsi karbohidrat, lemak, dan protein yang dikandung bahan
pakan tersebut. Air dan mineral tidak menyumbang energi pakan
tersebut. Nilai energi bruto tidak menunjukkan apakah energi tersebut
tersedia untuk hewan atau tidak tersedia, tergantung dari kecernaan
bahan pakan tersebut.
Persentase EB yang dapat dimanfaatkan oleh tubuh hewan dan
digunakan untuk mendukung proses metabolisme tergantung pada
Gambar 1. Perubahan adenosin triphosfat (ATP) menjadi adenosin diphosfat (ADP) dan fosfat bukan organik (Pi) dan energi
fisiologi hewan2

kemampuan hewan untuk mencerna bahan pakan. Pencernaan
mencerminkan proses fisika dan kimia yang terjadi dalam saluran
pencernaan dan menyebabkan terjadinya degradasi senyawa kimia
kompleks pakan menjadi molekul kecil yang diabsorbsi dan
digunakan oleh sel-sel tubuh hewan.
Energi yang dapat diabsorbsi disebut Energi Dapat Dicerna
(EDD). Pada hewan non ruminansia, kehilangan energi dapat terjadi
melalui urin, yaitu limbah yang mengandung nitrogen dan senyawa
lain yang tidak dioksidasi oleh tubuh hewan. Untuk hewan
ruminansia selain melalui urin, kehilangan energi terjadi melalui
pembentukan gas methan. EDD dikurangi energi yang hilang melalui
urin (non-ruminansia) atau urin dan methan (ruminansia) disebut
Energi Metabolis (EM) pakan. Selama proses metabolisme zat
makanan, terjadi kehilangan energi yang disebut Heat Increament.
Sisa energi dari pakan yang tersedia bagi hewan digunakan untuk
keperluan hidup pokok (maintenance) dan produksi dan disebut
sebagai energi netto (EN).
Gambar 2. Jalur energi pakan dalam tubuh hewan
3metabolisme karbohidrat

Energi pakan yang dikonsumsi hewan dapat digunakan dalam 3
cara: (1) energi yang tersedia digunakan untuk aktivitas; (2) energi
dikonversi menjadi panas; dan (3) energi disimpan dalam jaringan
tubuh. Kelebihan energi pakan yang dikonsumsi setelah kebutuhan
pertumbuhan normal dan metabolisme tubuh terpenuhi biasanya akan
disimpan sebagai lemak. Kelebihan energi tersebut tidak dapat
dibuang oleh tubuh hewan.
1.3 Karbohidrat
Karbohidrat adalah senyawa organik yang mengandung atom
karbon (C), hidrogen (H), oksigen (O), dan pada umumnya unsur
hidrogen akan berikatan dengan oksigen untuk menghasilkan H O. 2
Secara kimia karbohidrat adalah turunan dari aldehida atau keton dari -polihidrida alkohol (mempunyai gugus hidroksil atau OH lebih dari
satu). Di dalam tubuh karbohidrat dapat dibentuk dari beberapa
asam amino dan sebagian dari gliserol lemak melalui jalur
glukoneogenesis. Akan tetapi sebagian besar karbohidrat diperoleh
dari bahan makanan yang dikonsumsi sehari-hari, terutama sumber
bahan makanan yang berasal dari tumbuh-tumbuhan.
Sumber karbohidrat dalam bentuk glikogen, hanya dijumpai
pada otot dan hepar, sedangkan karbohidrat dalam bentuk laktosa
dijumpai di dalam susu. Pada tumbuh-tumbuhan, karbohidrat di
bentuk dari hasil reaksi CO dan H O melalui proses fotosintesis di 2 2
dalam sel-sel tumbuhan yang mengandung hijau daun (klorofil)
dengan bantuan cahaya matahari. Matahari merupakan sumber dari
seluruh kehidupan, tanpa matahari tanda-tanda dari kehidupan tidak
akan dijumpai. Reaksi fotosintesis yang menghasilkan karbohidrat
4 fisiologi hewan

adalah sebagai berikut:
Proses fotosintesis, klorofil pada tumbuh-tumbuhan akan
menyerap dan menggunakan energi cahaya matahari untuk
membentuk karbohidrat dengan bahan utama CO dari udara dan 2
molekul air (H O) yang berasal dari tanah. Energi kimia yang 2
terbentuk akan disimpan di dalam daun, batang, umbi, buah dan biji-
bijian.
1.4 Fungsi Karbohidrat
Karbohidrat mempunyai peranan penting dalam menentukan
karakteristik bahan makanan, seperti rasa, warna dan tekstur.
Fungsi karbohidrat di dalam tubuh adalah:
a. Fungsi utamanya sebagai sumber energi (1 gram karbohidrat
menghasilkan 4 kalori) bagi kebutuhan sel-sel jaringan tubuh.
Sebagian dari karbohidrat diubah langsung menjadi energi
untuk aktifitas tubuh, dan sebagian lagi disimpan dalam bentuk
glikogen di hepar dan di otot. Ada beberapa jaringan tubuh
seperti sistem saraf dan eritrosit, hanya dapat menggunakan
energi yang berasal dari karbohidrat saja.
b. Melindungi protein agar tidak teroksidasi.
c. Kebutuhan tubuh akan energi merupakan prioritas pertama;
bila karbohidrat yang di konsumsi tidak mencukupi untuk
kebutuhan energi tubuh dan jika tidak cukup terdapat lemak di
6 CO + 6 H O C H O + 6O2 2 6 6 212Cahaya Matahari
Chlorophil
Gambar 3. Proses fotosintesis pada tumbuhan
5metabolisme karbohidrat

dalam makanan atau cadangan lemak yang disimpan di dalam
tubuh, maka protein akan menggantikan fungsi karbohidrat
sebagai penghasil energi. Dengan demikian protein akan
meninggalkan fungsi utamanya sebagai zat pembangun.
Apabila keadaan ini berlangsung terus menerus, maka keadaan
kekurangan energi dan protein tidak dapat dihindari lagi.
d. Membantu metabolisme lemak dan protein dengan demikian
dapat mencegah terjadinya ketosis dan pemecahan protein yang
berlebihan.
e. Di dalam hepar berfungsi untuk detoksifikasi zat-zat toksik
tertentu.
f. Beberapa jenis karbohidrat mempunyai fungsi khusus di
dalam tubuh. Laktosa misalnya berfungsi membantu
penyerapan kalsium, ribosa merupakan komponen penting
penyusun asam nukleat.
g. Selain itu beberapa golongan karbohidrat yang tidak dapat
dicerna, mengandung serat (dietary fiber) berguna untuk
pencernaan dan memperlancar defekasi.
1.5 Karbohidrat Sebagai Sumber Energi Paling Penting Bagi
Tubuh
Karbohidrat atau hidrat arang adalah suatu zat gizi yang
mempunyai fungsi utama sebagi sumber energi. Karbohidrat untuk
setiap gramnya menghasilkan 4 kalori. Walaupun lemak
menghasilkan energi lebih besar, namun karbohidrat lebih banyak
dikonsumsi sehari-hari sebagai bahan makanan pokok. Alasan
fisiologis mengapa karbohidrat begitu penting untuk dikonsumsi
6 fisiologi hewan

adalah bahwa karbohidrat merupakan penyedia sumber energi
pertama kali bagi tubuh melalui pembentukan oksaloasetat sehingga
proses oksidasi-fosforilasi yang melibatkan transport elektron
dengan pembentukan senyawa intermidiet dapat terjadi secara
bertahap dan silih berganti.
Di negara sedang berkembang karbohidrat dikonsumsi sekitar
70-80% dari total kalori, bahkan pada daerah-daerah miskin bisa
mencapai 90%. Sedangkan pada negara maju karbohidrat dikonsumsi
hanya sekitar 40-60%. Hal ini disebabkan sumber bahan makanan
yang mengandung karbohidrat lebih murah harganya dibandingkan
sumber bahan makanan seperti lemak maupun protein. Sumber
karbohidrat banyak ditemukan pada serealia (beras, gandum, jagung,
kentang dan sebagainya), serta pada biji-bijian yang tersebar luas di
alam. Contoh bahan makanan sumber karbohidrat (kandungan
karbohidrat per 100 gram bahan) dapat dilihat pada tabel berikut ini:
Tabel 1. Sumber karbohidrat hewani dan nabati
7metabolisme karbohidrat

1.6 Klasifikasi dan Sumber Karbohidrat
Karbohidrat merupakan sumber energi bagi sebagian besar
makhluk hidup, terutama hewan tingkat tinggi. Karbohidrat berasal
dari tumbuh-tumbuhan, merupakan hasil fotosintesis yang
melibatkan klorofil, cahaya matahari, karbondioksida dan molekul
air. Hasil dari proses fotosintesis menghasilkan karbohidrat atau gula
sederhana (C H O ), yaitu glukosa.6 12 6
Secara umum, karbohidrat dikelompokkan menjadi,
monosakarida (gula sederhana), disakarida (dua molekul
monosakarida), oligosakarida (tiga sampai 6 molekul monosakarida,
dan polisakarida (lebih dari 6 molekul monosakarida) dan mixed
polisakarida. Monosakarida, disakarida dan oligosakarida biasanya
disebut kelompok sugar, sedangkan polisakarida yang terdiri dari
homoglukan (arabinan, xilan, glukan, fruktan, galaktan, mannan dan
glukosamin) dan heteroglukan (pektin, hemiselulosa, gum, musilago
asam, asam hialuronik dan kondroitin) disebut kelompok 'non sugar'.
Dalam ilmu Gizi, penggolongan karbohidrat yang paling
sering dipakai adalah berdasarkan jumlah molekulnya. Karbohidrat
yang paling sederhana adalah monosakarida yang memiliki 5-6
karbon, terdiri dari heksosa (mengandung 6 buah karbon): glukosa,
fruktosa, galaktosa dan pentosa (mengandung 5 buah karbon) :
ribosa, arabinosa, xilosa. Karbohidrat disakarida terdiri atas sukrosa,
maltosa dan laktosa. Karbohidrat yang lebih komplek adalah
polisakarida, yang amilum, dekstrin, glikogen dan selulosa.
Kelompok karbohidrat; monosakarida (gula sederhana),
disakarida (dua molekul monosakarida), oligosakarida (tiga sampai 6
molekul monosakarida, dan polisakarida (lebih dari 6 molekul
8 fisiologi hewan

monosakarida) dan mixed polisakarida, dijelaskan sebagai berikut:
a. Monosakarida
Karbohidrat yang paling sederhana (simple sugar) merupakan
bahan baku metabolisme yang siap digunakan sebagai sumber energi
bagi tubuh. Monosakarida larut di dalam air dan rasanya manis,
sehingga secara umum disebut juga gula. Penamaan kimianya
selalu berakhiran osa. Dalam Ilmu Gizi hanya ada tiga jenis
monosakarida yang penting yaitu, glukosa, fruktosa dan
galaktosa. Gula aldosa dan ketosa mempunyai rumus molekul yang
sama tetapi berbeda struktur molekulnya. Salah satu gula sederhana
yang siap digunakan oleh sel-sel tubuh adalah glukosa dalam bentuk
D-glukosa. Glukosa merupakan hasil akhir dari pencernaan pati,
sukrosa, maltosa atau laktosa. Tingkat kemanisan glukosa separuh
dari sukrosa.
Gambar 4. Jenis monosakarida
Gambar 5. Struktur molekul L-glukosa dan D-glukosa
O
C
C
C
C
C
CH
H
H
OH
H
H
OH
HO
H
HO
HO
1
2
3
4
5
6
2
O
C
C
C
C
C
CH
H
OH
H
OH
OH
OH
H
HO
H
H
2
L-Glucose D-Glucose
9metabolisme karbohidrat

Glukosa dan fruktosa merupakan karbohidrat jenis
monosakarida. Dua jenis karbohidrat ini terdapat melimpah sebagai
monosakarida bebas dan berperan sebagai sumber energi penting
bagi tubuh. Glukosa adalah molekul sederhana yang bersifat mudah
larut dalam air dan mudah diserap oleh sel-sel epitel usus dan
ditarnsport secara langsung ke seluruh sel-sel tubuh sebagai sumber
energi. Glukosa dapat mengalami polimerisasi membentuk
polisakarida. Glukosa banyak dijumpai di alam, terutama pada buah-
buahan, sayur-sayuran, madu, sirup jagung dan tetes tebu. Di dalam
tubuh glukosa berasal dari hasil akhir pencemaan amilum, sukrosa,
maltosa dan laktosa. Glukosa terdapat di dalam aliran darah
(disebut kadar gula darah) dan berfungsi sebagai sumber energi bagi
seluruh sel-sel dan jaringan tubuh. Terdapat kekhususan pada
sistem saraf pusat, yaitu sel-sel saraf hanya dapat menggunakan
glukosa sebagai sumber energi dan tidak mampu melakukan
penyimpanan. Kadar gula darah dalam tubuh sekitar 80-120 mg%.
Kadar gula darah dapat meningkat melebihi normal dan kondisi ini
disebut hiperglikemia dan banyak dijumpai pada penderita diabetes
mellitus (DM).
Tabel 2. Derajat rasa manis karbohidrat monosakarida dan disakarida
10 fisiologi hewan

Gula alkohol terdapat di alam dan dapat dibuat secara sintetis
dengan menggunakan sarbitol, manitol, dulkito atau inositol. Sarbitol
terdapat dalam beberapa jenis buah dan dapat dibuat dari glukosa
dengan enzim aldosa reduktase dengan mengubah gugus aldehida
dari glukosa menjadi alkohol (CH OH). Sarbitol banyak digunakan 2
dalam minuman dan kue, khusus untuk penderita diabetes melitus
(DM). Tingkat kemanisannya hanya 60% dibanding sukrosa,
diabsorpsi lebih lambat dan di sel-sel hepar diubah menjadi glukosa.
Sorbitol tidak mudah dimetabolisme oleh bakteri, tidak menimbulkan
karies gigi, dan dapat digunakan sebagai bahan untuk membuat
permen karet. Manitol dan dulkitol adalah gula alkohol yang dibuat
dari monoskarida manosa dan galaktosa. Manitol terdapat dalam
nenas, asparagus, ubi jalar, wortel. Manitol juga dapat diperoleh dari
ekstrak rumput laut. Inositol adalah alkohol siklik yang menyerupai
glukosa, terutama bentuk esternya. Inositol dengan asam pitat
menghambat absorbsi kalsium (Ca) dan zat besi (Fe).
Fruktosa disebut juga sebagai gula buah atau levulosa
mempunyai rumus bangun sama dengan glukosa tetapi memiliki
struktur molekul yang berbeda. Karbohidrat ini merupakan gula
sederhana jenis monosakarida yang paling manis, banyak dijumpai
pada mahkota bunga, madu dan hasil hidrolisa dari gula tebu. Di
dalam tubuh fruktosa didapat dari hasil pemecahan sukrosa. Gula
sederhana yang lain adalah galaktosa, manosa dan pentosa, ribosa
atau dioksiribosa. Galaktosa tidak terdapat di alam bebas dan hanya
terdapat dalam tubuh sebagai hasil pencernaan laktosa.
11metabolisme karbohidrat

b. Disakarida
Disakarida dibentuk dari dua unit monosakarida yang saling
berikatan satu sama lain dengan ikatan glikosida (glycocide). Ikatan
terjadi antara atom carbon nomor 1 dengan atom C nomor 4 dengan melepas H O. Disakarida dengan unit monosakarida yang saling 2
berikatan tersebut dapat dipecah kembali menjadi 2 monosakarida
melalui proses hidrolisis. Disakarida dibagi menjadi sukrosa
(sakarosa), maltosa, laktosa, dan trehalosa. Sumber disakarida, antara
lain tebu (100% mengandung sukrosa), bit, gula, nira (50%
mengandung sukrosa) dan lain-lain.
Sukrosa adalah gula yang sering digunakan untuk kebutuhan
sehari-hari, sehingga lebih sering disebut gula meja (table sugar) atau
gula pasir dan disebut juga invert sugar. Sukrosa atau sakarosa
merupakan gula bit atau gula tebu yang banyak terdapat dalam buah,
sayur dan madu. Apabila gula ini dicerna atau dihidrolisis akan
menghasilkan satu unit glukosa dan satu unit fruktosa.
Maltosa mempunyai 2 (dua) molekul monosakarida yang
terdiri dari dua molekul glukosa. Maltosa atau gula malt tidak
terdapat di alam bebas dan merupakan hasil pemecahan pati.
Pencernaan atau hidrolisis dari gula maltosa juga akan menghasilkan
2 unit glukosa. Di dalam tubuh maltosa didapat dari hasil pemecahan
amilum dan lebih mudah dicema, rasanya lebih enak dan nikmat.
Dengan iodium amilum akan berubah menjadi warna biru.
Laktosa atau gula susu merupakan disakarida yang terdapat
dalam susu yang mempunyai 2 (dua) molekul monosakarida yang
terdiri dari satu molekul glukosa dan satu molekul galaktosa. Laktosa
kurang larut di dalam air. Laktosa hanya terdapat pada susu sehingga
12 fisiologi hewan

disebut juga gula susu, seperti susu sapi (4-5%) dan air susu ibu (4-
7%). Laktosa yang mengalami proses fermentasi oleh sejumlah
mikroorganisme termasuk Streptococcus lactis akan dikonversi
menjadi asam laktat, misalnya dalam memproduksi yakult melalui
peran Laktobacillus casei sirota atau memproduksi yogurt melalui
peran Steptococcus thermophilus dan Lactobactillus bulgaricus.
Laktosa dapat menimbulkan intolerance (laktosa intolerance)
disebabkan kekurangan enzim laktase sehingga kemampuan untuk
mencerna laktosa berkurang. Kelainan ini dapat dijumpai pada bayi,
anak dan orang dewasa, baik untuk sementara (temporer) maupun
secara tetap (permanen). Gejala yang sering dijumpai adalah
diare, kembung, flatus dan kejang perut. Defisiensi laktase pada
bayi dapat menyebabkan gangguan pertumbuhan, karena bayi sering
diare. Terapi diet dapat dilakukan dengan pemberian formula rendah
laktosa seperti Almiron, Isomil, Prosobee dan Nutramigen, dan AI
110 bebas laktosa. Formula rendah laktosa tidak boleh diberikan
terlalu lama (maksimum tiga bulan), karena laktosa diperlukan untuk
pertumbuhan sel-sel otak. Setelah tiga bulan, laktosa diberikan
secara bertahap sesuai dengan pertumbuhan anak.
Kadar laktosa dalam susu sapi adalah 6,8 g per 100 ml, dalam
ASI 4,8 g. Orang kulit berwarna tidak tahan terhadap susu sapi karena
sifat negatif dari enzim laktase sehingga menimbulkan kondisi
”lactosa-intolerance”. Oleh karena itu laktosa tidak bisa dicerna dan
tetap tinggal dalam saluran pencernaan dan mempengaruhi
mikroorganisme yang tumbuh, sehingga mengakibatkan munculnya
mual, kembung, kejang perut dan diare. Trihalosa merupakan
disakarida yang terdiri dari 2 unit molekul glukosa seperti halnya
13metabolisme karbohidrat

maltosa.
c. Oligosakarida
Oligosakarida terdiri atas polimer 2-10 monosakarida yang
terdiri unit-unit glukosa dan fruktosa yang dihubungi dengan ikatan
glikosida (α, β). Contoh gula oligosakarida adalah rafinosa,
stakinosa, fruktan dan verbaskosa. Oligosakarida banyak terdapat
dalam biji tumbuh-tumbuhan dan kacang-kacangan dan tidak dapat
dipecah oleh enzim-enzim pencernaan melainkan dengan proses
fermentasi. Fruktan sebagai oligosakarida banyak terdapat dalam
tanaman serealia, bawang merah, bawang putih dan asparagus.
Fruktan terdiri atas beberapa unit fruktosa yang terikat dengan 1
molekul glukosa. Karbohidrat jenis ini tidak dapat dicerna dan
difermentasi.
d. Polisakarida Pati
Polisakarida merupakan senyawa karbohidrat kompleks,
dapat mengandung lebih dari 60.000 molekul monosakarida yang
tersusun membentuk rantai lurus ataupun bercabang. Polisakarida
rasanya tawar (tidak manis), tidak seperti monosakarida dan
disakarida. Contoh polisakarida adalah; amilum (pati) dekstrin dan
glikogen. Polisakarida, diklasifikasikan sebagai: heteropolisakarida
dan homopolisakarida. Terdapat sebagai struktur dasar dari sel,
hampir diseluruh jaringan, di mukus, beberapa hormon, enzim-
enz im, bahan-bahan darah , dan za t -za t kekeba lan .
Homopolisakarida: glikogen, tersedia di jaringan hewan yang
menyerupai pati tanaman dan merupakan simpanan energi jangka
pendek. Hanya hepar dan ginjal yang dapat melepaskan glukosa
14 fisiologi hewan

untuk masuk ke darah. Glikogen hepar adalah glukosa terpenting.
Proses pembentukan glikogen disebut glikogenesis (glycogen
synthetase). Proses pemecahan glikogen disebut glikogenolisis
(glycogen phosphorylase).
Pati merupakan simpanan karbohidrat yang terdapat pada
tumbuh-tumbuhan dan merupakan sumber karbohidrat utama bagi
manusia. Jumlah unit glukosa dan susunannya dalam satu jenis pati
berbeda satu sama lainnya bergantung jenis tanaman asalnya. Bentuk
butiran pati mempunyai karateristik tersendiri dalam hal daya larut,
daya mengentalkan dan rasa. Ada 2 jenis pati, yaitu amilosa dan
amilopektin. Amilosa mempunyai rantai panjang unit glukosa yang
tidak bercabang; sedangkan amilopektin adalah polimer yang
susunannya bercabang-cabang dengan 15-30 unit glukosa. Pada
setiap cabang dan rantai glukosa terikat melalui ikatan tertentu, yang
kemudian dapat dipecah dalam proses pencernaan. Beras banyak
mengandung amilosa, sedangkan beras ketan mengandung banyak
amilopektin. Pencernaan pati secara umum akan menghasilkan
dekstrin, maltosa dan glukosa sebagai produk akhir.
Pati (amilum) merupakan sumber energi utama bagi orang
dewasa di seluruh penduduk dunia, terutama di negara sedang
berkembang oleh karena di konsumsi sebagai bahan makanan
pokok. Disamping itu bahan pangan kaya akan amilum juga
mengandung protein, vitamin, serat dan beberapa zat gizi penting
lainnya. Amilum merupakan karbohidrat dalam bentuk cadangan
bagi tumbuh-tumbuhan dalam bentuk granul yang dijumpai pada
umbi dan akarnya. Umbi-umbian, serealia dan biji-bijian merupakan
sumber amilum yang berlimpah ruah oleh karena mudah didapat
15metabolisme karbohidrat

untuk di konsumsi. Jagung, beras dan gandum kandungan amilumnya
lebih dari 70%, sedangkan pada kacang-kacangan sekitar 40%.
Amilum tidak larut di dalam air dingin, tetapi larut di dalam air
panas membentuk cairan yang sangat pekat seperti pasta; fenomena
ini disebut "gelatinisasi".
Pati (amilum) dapat dipisahkan dengan air panas menjadi 2
fraksi, yaitu amilosa yang mempunyai sifat larut dengan air panas
serta mempunyai struktur rantai yang lurus dan amilopektin yang
mempunyai sifat tidak larut dengan air panas serta mempunyai
sruktur rantai yang bercabang. Peranan perbandingan amilosa
dan amilopektin terlihat pada serelia; contohnya beras. Semakin
kecil kandungan amilosa atau semakin tinggi kandungan
amilopektinnya, semakin lekat nasi tersebut. Pulut sedikit sekali
amilosanya (1-2%), beras mengandung amilosa >2%. Berdasarkan
kandungan amilosanya, beras (nasi) dapat dibagi menjadi 4 golongan:
1. amilosa tinggi 25-33%
2. amilosa menengah 20-25%
3. amilosa rendah 09-20%
4. amilosa sangat rendah < 9%
Secara umum penduduk di negara-negara Asean, khususnya
Flipina, Malaysia, Thailand dan Indonesia menyenangi nasi dengan
kandungan amilosa medium, sedangkan Jepang dan Korea
menyenangi nasi dengan amilosa rendah.
Dekstrin merupakan produk antara dari proses pencernaan pati
atau dihasilkan melalui hidrolisis pati secara parsial. Molekulnya
lebih sederhana, lebih mudah larut di dalam air, dengan yodium akan
berubah menjadi wama merah. Dekstrin biasanya digunakan untuk
16 fisiologi hewan

'tube feeding'. Cairan glukosa untuk infus merupakan campuran
dekstrin, maltosa, glukosa dan air. Campuran ini molaritasnya lebih
besar dari sukrosa dan glukosa, pengaruh osmolarnya lebih rendah
sehingga tidak mudah menimbulkan diare. Dekstrin maltosa
merupakan produk hidrolisis pati secara parsial yang memiliki sifat
tidak mudah terfermentasi namun mudah dicerna dan biasanya
digunakan untuk makanan bayi.
Glikogen merupakan polisakarida yang disimpan dalam tubuh,
terutama pada sel-sel hepar dan otot. Karbohidrat jenis ini terdiri dari
unit-unit glukosa dalam bentuk rantai lebih bercabang daripada
amilopektin dan lebih mudah dipecah. Tubuh manusia hanya
menyimpan 350 g glikogen; 2/3 di otot dan 1/3 di hepar. Glikogen
merupakan "pati hewani", terbentuk dari ikatan 1000 molekul, larut di
dalam air (pati nabati tidak larut dalam air) dan bila bereaksi dengan
yodium akan menghasilkan warna merah. Glikogen terdapat pada
otot hewan, manusia dan ikan. Glikogen di otot hanya digunakan
sebagai sumber energi otot tersebut, sedangkan glikogen yang di
hepar digunakan untuk keperluan semua bagian tubuh yang
membutuhkan. Kelebihan glukosa yang melebihi batas kemampuan
simpan (glikogen) akan diubah menjadi lemak dan disimpan dalam
jaringan lemak. Pada waktu hewan disembelih, terjadi kekejangan
(rigor mortis) dan kemudian glikogen dipecah menjadi asam laktat
selama post mortum. Glikogen disimpan di dalam hepar dan otot
sebagai cadangan energi, yang sewaktu-waktu dapat diubah
kembali menjadi glukosa bila dibutuhkan. Sumber glikogen terdapat
pada kecambah, serealia, susu dan sirup jagung (26%).
17metabolisme karbohidrat

e. Polisakarida Non-Pati (Mixed Polysakaride)
Polisakarida non-pati (serat) terdiri atas 2 kelompok, yaitu serat
yang larut dalam air dan tidak laut dalam air. Serat yang larut dalam
air, contohnya: pektin, gum, mukilase, glukan dan alga; sedangkan
serat yang tidak larut dalam air adalah: selulosa, hemiselulosa, lignin,
selobiosa.
Selulosa hampir 50% berasal dari tumbuh-tumbuhan sebagai
bagian yang terpenting dari dinding sel tumbuh-tumbuhan. Selulosa
berikatan erat secara fisik dan kimia dengan hemiselulosa dan lignin.
Selulosa tidak dapat dicerna oleh tubuh manusia, oleh karena
tidak ada enzim untuk memecah selulosa. Meskipun tidak dapat
dicerna, selulosa berfungsi sebagai sumber serat yang dapat
memperbesar volume dari feses, sehingga akan memperlancar
defekasi. Dahulu serat digunakan sebagai indeks dalam menilai
kualitas makanan, semakin tinggi kandungan serat dalam
makanan maka nilai gizi makanan tersebut semakin buruk. Akan
tetapi pada dasawarsa terakhir ini, para ahli sepakat bahwa serat
(dietary fiber) merupakan komponen penyusun diet manusia yang
sangat penting. Tanpa adanya serat, mengakibatkan terjadinya
konstipasi (susah buang air besar), haemorhoid (ambeyen),
divertikulosis, kanker pada usus besar, appendisitis, diabetes
penyakit jantung koroner dan obesitas. Fungsi serat adalah:
1. Mencegah penyakit jantung koroner
Kolesterol telah lama diduga sebagai penyebab
terjadinya aterosklerosis yang akhirnya berakibat timbulnya
penyakit jantung koroner. Produk akhir metabolisme kolesterol
adalah asam empedu. Serat yang berasal dari makanan
18 fisiologi hewan

sesampainya di saluran pencernaan akan mengikat asam
empedu. Dalam keadaan terikat, asam empedu bersama-sama
serat dikeluarkan dalam bentuk feses. Dengan demikian
semakin banyak serat dimakan, maka semakin banyak lemak
dan kolesterol yang dikeluarkan.
Masyarakat yang mengkonsumsi makanan tinggi
serat (terutama serat dari seralia dan kacang-kacangan),
cenderung memiliki kadar kolesterol dalam darah yang
rendah serta angka kematian akibat penyakit jantung
koroner lebih rendah dibandingkan dengan orang Eropa yang
konsumsi serat relatif lebih sedikit. Penelitian lain
menunjukkan bahwa suku pedalaman Masai di Afrika yang
hidupnya berburu dan suku Venda di Afrika Selatan yang
hidupnya bercocok tanam, tidak ditemukan adanya penyakit
jantung koroner, walaupun mereka mengkonsurnsi lemak
hewan dalam jumlah yang tinggi, yaitu masing-masing 300
gram dan 126 gram per hari. Hal tersebut disebabkan mereka
makan jagung yang tidak digiling rata-rata 494 gram per hari,
yang kadar seratnya diperkirakan 5,7 gram. Dibandingkan
dengan orang Inggris yang rata-rata hanya mengkonsumsi
serat sebanyak 0,5 gram per hari.
2. Mencegah kanker pada usus besar
Kanker pada usus besar (kolon) diakibatkan masuknya benda-
benda asing ke dalam usus besar, benda-benda asing tersebut
akan diubah sifatnya menjadi karsinogenik. Adanya serat
kasar yang melalui kolon, mengakibatkan lingkungan mikroba
terganggu sehingga aktifitas mikroba tersebut berkurang.
19metabolisme karbohidrat

3. Mencegah penyakit diabetes
Pemyataan ini didukung oleh suatu penelitian yang
dilakukan di Capetown, yang menunjukkan bahwa pada
penduduk yang mengkonsumsi serat rata-rata 6,5 gram per
hari ditemukan banyak kasus diabetes sebanyak 3,6 %, lebih
besar dibandingkan dengan penduduk yang makan serat rata-
rata 24,8 gram per hari dengan persentase kasus hanya 0,05%.
4. Mencegah penyakit divertikular
Konsumsi serat yang cukup akan menghasilkan feses yang
lembut sehingga dengan konstraksi otot yang rendah (<10
mmHg) feses dapat dikeluarkan dengan lancar. Apabila
konsumsi serat berkurang, maka volume kotoran menjadi kecil-
kecil dan keras (seperti feses kambing), sehingga untuk
membuangnya membutuhkan kontraksi otot yang lebih besar
(tekanan bisa mencapai >90 mm Hg). Apabila tekanan kuat
tersebut berlangsung berulang ulang setiap hari dalam jangka
waktu yang lama, maka otot- otot kolon menjadi lelah dan
lemah. Keadaan ini menyebabkan penyakit "divertikular",
yaitu penonjolan bagian luar usus berbentuk bisul yang
kadang- kadang disertai peradangan yang dapat
menimbulkan infeksi.
5. Mencegah kegemukan
Dengan adanya serat, maka penyerapan karbohidrat, lemak
dan protein menjadi berkurang. Jika hal ini dilakukan secara
teratur dan berkesinambungan, maka kegemukan dapat
dihindari. Serat mampu memberikan perasaan kenyang dalam
waktu yang cukup lama. Sumber serat yang baik adalah
20 fisiologi hewan

sayuran, buah-buahan, serealia dan kacang-kacangan.
Memakan sayuran dan buah-buahan dalam jumlah yang
banyak, mempunyai fungsi ganda, yaitu disamping sebagai
sumber serat juga merupakan sumber vitamin dan mineral,
yang semua itu sangat dibutuhkan untuk memelihara
kesehatan tubuh manusia. Produk makanan hewani, seperti
daging, ikan susu dan telur serta bahan-bahan olahannya,
umumnya mengandung serat dalam jumlah yang sedikit.
Sayuran yang banyak mengandung serat adalah, bayam,
kangkung, buncis, daun beluntas, daun singkong, kacang
panjang, daun katuk, daun kelor, sawi, kecipir, kol dan lain-lain.
Buah-buahan yang banyak mengandung tinggi serat adalah,
alpukat, belimbing, srikaya, cempedak, nangka, durian, jeruk,
kedondong, kemang, mangga, nenas dan sebagainya. Seralia
yang kaya serat adalah beras, jagung, jali dan jewawut. Beras
giling mernpunyai kadar serat dan vitamin (khususnya vitamin
B) lebih rendah dari beras tumbuk, karena itu memilih beras
sebaiknya jangan yang terlalu bersih (putih). Kacang-
kacangan yang banyak mengandung serat adalah kacang bogar,
kacang merah, kacang ijo, kedele, serta kacang-kacangan
lainnya.
Hewan ruminansia selulosa dicerna dalam saluran pencernaan
oleh enzim selulase menghasilkan selobiosa, lalu dihidrolisis menjadi
glukosa oleh selobiase. Enzim selulase dihasilkan oleh mikroba
rumen dan retikulum ruminansia. Hasil akhir dari pencernaan
selulosa adalah asam-asam lemak bebas (VFA) yang terdiri dari
asetat, propionat dan butirat, dengan hasil sampingan antara lain
21metabolisme karbohidrat

berupa gas metan, dan CO yang akan digunakan dalam metabolisme 2
energi pada hewan ruminansia.
Lignin merupakan polimer yang mengandung protein sulit
dicerna. Lignin sangat tahan terhadap degradasi kimia dan enzimatik.
Lignin sering digunakan sebagai indikator di dalam eksperimen studi
kecernaan pada hewan ruminansia karena sifatnya yang tidak larut
tersebut. Lignin bukan karbohidrat, tetapi sangat berhubungan erat
dengan senyawa- senyawa kabohidrat. Kulit kayu, biji, bagian
serabut kasar, batang dan daun mengandung lignin yang berupa
substansi kompleks oleh adanya lignin dan polisakarida yang lain.
Kadar lignin akan bertambah dengan bertambahnya umur tanaman.
Adapun, selobiosa tidak terdapat bebas di alam, mempunyai ikatan β-
(1,4). Ikatan tersebut tidak dapat dipecah oleh enzim yang dihasilkan
oleh mamalia kecuali oleh enzim yang disintesis oleh
mikroorganisme retikulorumen.
1.7 Jalur Pemrosesaan Karbohidrat Di Dalam Tubuh
Dari sekian banyak jenis karbohidrat, polisakarida dan
disakarida merupakan jenis karbohidrat utama yang terdapat dalam
diet makanan. Polisakarida seperti kanji dan glikogen terdiri dari unit-
unit glukosa yang terikat dengan ikatan glikosida. Disakarida terdiri
dari sukrosa dan Iaktosa. Pencernaan karbohidrat bermula di dalam
mulut. Enzim ptialin atau amilase liur akan menghidrolisis kanji atau
glikogen menjadi dekstrin dan maltosa. Di dalam perut, amilase liur
menjadi tidak aktif karena pengaruh HCl yang disekresikan oleh sel-
sel lambung. Dekstrin dan maltosa selanjutnya akan masuk ke
duodenum usus halus, merangsang pankreas untuk memproduksi dan
22 fisiologi hewan

mensekresikan enzim yang bersifat alkalis, yaitu amilase.
Amilase pankreas ini lebih aktif daripada amilase liur. Oleh
peran amilase dekstrin dan maltosa akan dihidrolisis menghasilkan
disakarida dan sejumlah kecil monosakarida. Selanjutnya enzim-
enzim yang dihasilkan oleh sel-sel pada duodenum atau bagian
permukaan usus halus, seperti maltase, sukrase, laktase dan
isomaltase (disakaridase) akan menghidrolisis unit-unit disakarida
menjadi monosakarida. Monosakarida ini selanjutnya akan diserap
dengan sistem transport aktif melalui sel-sel epitel yang terdapat pada
bagian permukaan usus halus, terutama di bagian yeyunum dan
akhirnya akan masuk ke dalam aliran darah. Jika daya absorbsi sel-sel
epitel pada bagian yeyenum sudah mencapai ambang batas kapasitas,
maka absorbsi akan berlanjut pada bagian ileum. Monosakarida yang
telah terabsorbsi selanjutnya diangkut menuju ke hepar melalui
pembuluh portal. Sedangkan disakarida yang terabsorbsi akan di
bawa dari pembuluh portal menuju ke hepar dan masuk ke dalam
sirkulasi darah untuk diedarkan ke sel-sel di seluruh tubuh.
Di dalam hepar, monosakarida dapat digunakan untuk
menghasilkan energi, diubah menjadi glikogen untuk cadangan
energi, diubah menjadi karbohidrat bentuk lain atau gula-gula
pentosa. Selain itu dalam kondisi tertentu, monosakarida dapat
diubah menjadi lemak atau asam-asam amino melalui jalur
transaminasi di dalam siklus Kreb. Glukosa yang dimobilisasi dari
dalam sel-sel hepar masuk ke dalam sistem sirkulasi menuju sel-sel di
jaringan perifer untuk digunakan sebagai sumber energi.
Glukosa merupakan monosakarida tunggal yang digunakan
oleh tubuh untuk menghasilkan energi. Hepar mempunyai enzim
23metabolisme karbohidrat

yang dapat mengubah monosakarida (galaktosa dan fruktosa)
menjadi glukosa. Glukosa dapat disimpan dalam hepar dan otot
dalam bentuk glikogen. Glikogen hepar dapat digunakan sewaktu-
waktu apabila kadar glukosa darah menurun. Berbeda dengan
glikogen hepar, glikogen otot tidak dapat digunakan sewaktu-waktu
karena glikogen otot tidak mudah masuk ke dalam darah, namun
demikian glikogen otot dapat digunakan mencukupi kebutuhan
energi di otot. Enzim hepar yang mampu mengubah monosakarida
adalah glukosa-6-phospatase yang dapat mengubah glikogen
menjadi glukosa, tetapi otot tidak mempunyai enzim ini. Oleh karena
itu, hepar memainkan peranan penting dalam metabolisme
karbohidrat dan sangat berperan dalam meregulasi kadar glukosa
dalam plasma darah.
1.8 Pencernaan Karbohidrat
Karbohidrat yang terdapat pada makanan dapat
dikelompokkan:
1. Karbohidrat yang tersedia (available carbohydrate), yaitu
karbohidrat yang dapat dicerna, diserap serta dimetabolisme
sebagai karbohidrat.
2. Karbohidrat yang tidak tersedia (unvailable carbohydrate),
yaitu karbohidrat yang tidak dapat dihidrolisa oleh enzim –
enzim pencernaan manusia, sehingga tidak dapat diabsorpsi.
1.9 Pencernaan dan Metabolisme Karbohidrat
a. Pencernaan di Mulut
Pencernaan karbohidrat sudah dimulai sejak makanan masuk
24 fisiologi hewan
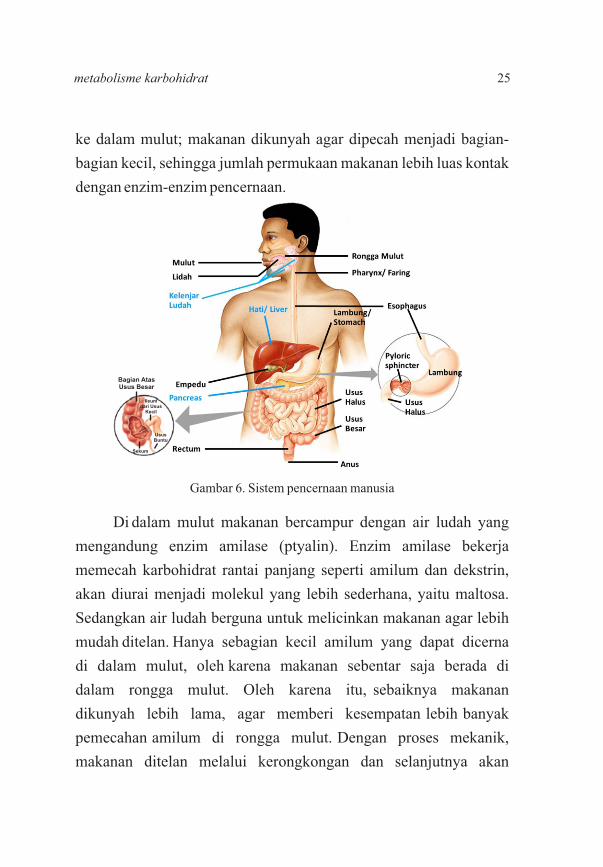
ke dalam mulut; makanan dikunyah agar dipecah menjadi bagian-
bagian kecil, sehingga jumlah permukaan makanan lebih luas kontak
dengan enzim-enzim pencernaan.
Di dalam mulut makanan bercampur dengan air ludah yang
mengandung enzim amilase (ptyalin). Enzim amilase bekerja
memecah karbohidrat rantai panjang seperti amilum dan dekstrin,
akan diurai menjadi molekul yang lebih sederhana, yaitu maltosa.
Sedangkan air ludah berguna untuk melicinkan makanan agar lebih
mudah ditelan. Hanya sebagian kecil amilum yang dapat dicerna
di dalam mulut, oleh karena makanan sebentar saja berada di
dalam rongga mulut. Oleh karena itu, sebaiknya makanan
dikunyah lebih lama, agar memberi kesempatan lebih banyak
pemecahan amilum di rongga mulut. Dengan proses mekanik,
makanan ditelan melalui kerongkongan dan selanjutnya akan
Gambar 6. Sistem pencernaan manusia
25metabolisme karbohidrat
Bagian AtasUsus Besar
Ileumdari Usus
Kecil
Usus Buntu
Sekum
memasuki lambung.
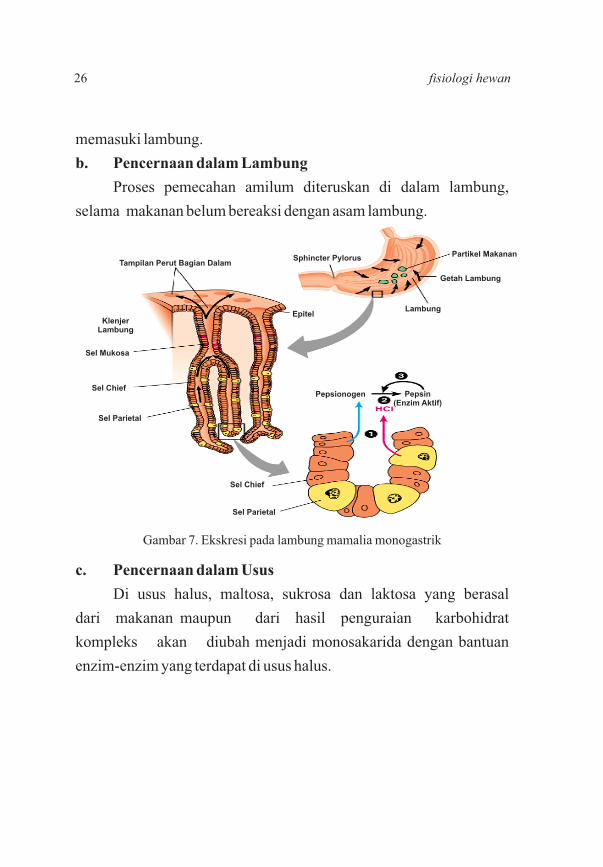
b. Pencernaan dalam Lambung
Proses pemecahan amilum diteruskan di dalam lambung,
selama makanan belum bereaksi dengan asam lambung.
c. Pencernaan dalam Usus
Di usus halus, maltosa, sukrosa dan laktosa yang berasal
dari makanan maupun dari hasil penguraian karbohidrat
kompleks akan diubah menjadi monosakarida dengan bantuan
enzim-enzim yang terdapat di usus halus.
Gambar 7. Ekskresi pada lambung mamalia monogastrik
26 fisiologi hewan
Tampilan Perut Bagian Dalam
KlenjerLambung
Sel Mukosa
Sel Chief
Sel Parietal
Sel Chief
Sel Parietal
Pepsin(Enzim Aktif)
Pepsionogen
EpitelLambung
Getah Lambung
Partikel MakananSphincter Pylorus
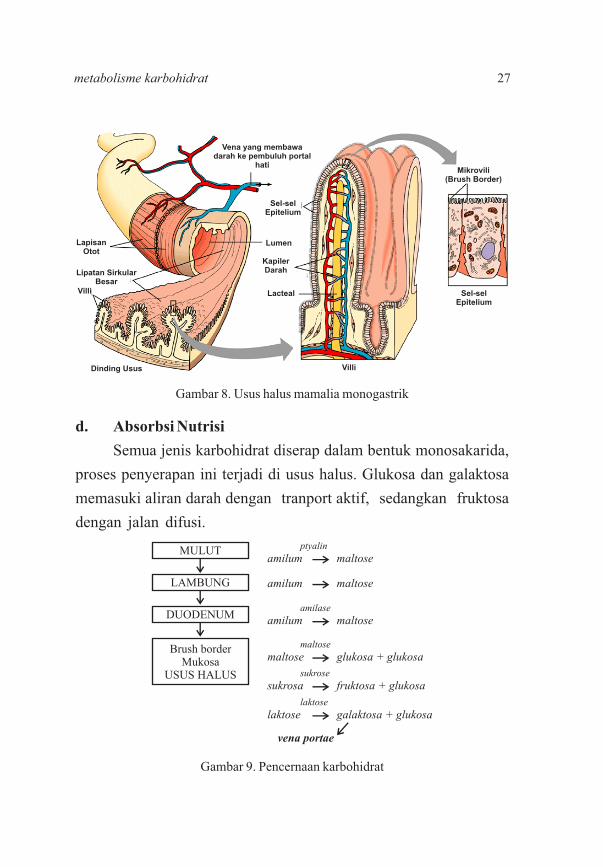
d. Absorbsi Nutrisi
Semua jenis karbohidrat diserap dalam bentuk monosakarida,
proses penyerapan ini terjadi di usus halus. Glukosa dan galaktosa
memasuki aliran darah dengan tranport aktif, sedangkan fruktosa
dengan jalan difusi.
Gambar 8. Usus halus mamalia monogastrik
Gambar 9. Pencernaan karbohidrat
27metabolisme karbohidrat
Dinding Usus
Villi
Lipatan SirkularBesar
LapisanOtot
Vena yang membawadarah ke pembuluh portal
hati
Lumen
Sel-selEpitelium
KapilerDarah
Lacteal
Villi
Sel-selEpitelium
Mikrovili(Brush Border)

Karbohidrat diserap dalam bentuk disakarida. Hal ini
dibuktikan dengan dijumpainya maltosa, sukrosa dan laktosa
dalam urin apabila mengkonsumsi gula dalam jumlah banyak.
Akhirnya berbagai jenis karbohidrat diubah menjadi glukosa
sebelum diikutsertakan dalam proses metabolisme. Berdasarkan
urutan, yang paling cepat di absorpsi adalah galaktosa, glukosa
dan terakhir fruktosa.
e. Metabolisme Nutrisi
Setelah melalui dinding usus halus, glukosa akan menuju
ke hepar melalui vena portae. Sebagian karbohidrat ini diikat di
dalam hepar dan disimpan sebagai glikogen, sehingga kadar gula
darah dapat dipertahankan dalam batas-batas normal (80-120 mg%).
Karbohidrat yang terdapat dalam darah, praktis dalam bentuk
glukosa, oleh karena fruktosa dan galaktosa akan diubah terlebih
dahulu sebelum memasuki pembuluh darah. Apabila jumlah
karbohidrat yang dimakan melebihi kebutuhan tubuh, sebagian
besar (2/3) akan disimpan di dalam otot dan selebihnya di dalam
hepar sebagai glikogen. Kapasitas pembentukan glikogen ini sangat
terbatas (maksimum 350 gram), dan jika penimbunan dalam
bentuk glikogen ini telah mencapai batasnya, kelebihan
karbohidrat akan diubah menjadi lemak dan disimpan di jaringan
lemak. Bila tubuh memerlukan kembali energi tersebut, simpanan
glikogen akan digunakan terlebih dahulu, disusul mobilisasi
lemak. Jika dihitung dalam jumlah kalori, simpanan energi
bentuk lemak jauh melebihi jumlah simpanan dalam bentuk
glikogen.
28 fisiologi hewan

Sel-sel tubuh yang sangat aktif dan memerlukan banyak energi,
mendapatkan energi dari hasil pembakaran glukosa yang diambil
dari aliran darah. Kadar gula darah akan diisi kembali dari
cadangan glikogen yang ada di dalam hepar. Kalau energi yang
diperlukan lebih banyak lagi, timbunan lemak dari jaringan lemak
akan digunakan. Dalam jaringan lemak diubah ke dalam zat antara
yang dialirkan ke hepar. Disini zat antara itu diubah menjadi
glikogen, mengisi kembali cadangan glikogen yang telah
dipergunakan untuk meningkatkan kadar gula darah. Peristiwa
oksidasi glukosa di dalam jaringan-jaringan terjadi secara bertahap
dan pada tahap-tahap itulah energi dilepaskan sedikit demi sedikit,
untuk dapat digunakan selanjutnya.
Melalui suatu deretan proses-proses kimiawi, glukosa dan
glikogen diubah menjadi asam piruvat. Asam piruvat ini merupakan
zat antara yang sangat penting dalam metabolisme karbohidrat.
Asam piruvat dapat segera diolah lebih lanjut dalam suatu proses
pada "siklus Krebs". Dalam proses siklis ini dihasilkan CO dan H O 2 2
dan terlepas energi dalam bentuk persenyawaan yang mengandung
tenaga kimia yang besar yaitu ATP (Adenosin trifosfat). ATP ini
mudah sekali melepaskan energinya sambil berubah menjadi ADP
(Adenosindifosfat). Sebagian dari asam piruvat dapat diubah
menjadi "asam laktat". Asam laktat ini dapat keluar dari sel-sel
jaringan dan memasuki aliran darah menuju ke hepar.
29metabolisme karbohidrat
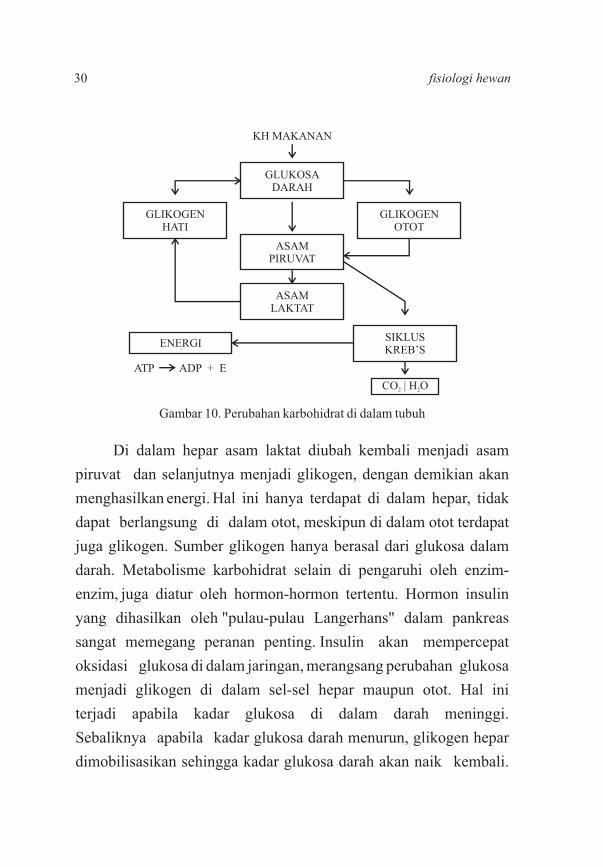
Di dalam hepar asam laktat diubah kembali menjadi asam
piruvat dan selanjutnya menjadi glikogen, dengan demikian akan
menghasilkan energi. Hal ini hanya terdapat di dalam hepar, tidak
dapat berlangsung di dalam otot, meskipun di dalam otot terdapat
juga glikogen. Sumber glikogen hanya berasal dari glukosa dalam
darah. Metabolisme karbohidrat selain di pengaruhi oleh enzim-
enzim, juga diatur oleh hormon-hormon tertentu. Hormon insulin
yang dihasilkan oleh "pulau-pulau Langerhans" dalam pankreas
sangat memegang peranan penting. Insulin akan mempercepat
oksidasi glukosa di dalam jaringan, merangsang perubahan glukosa
menjadi glikogen di dalam sel-sel hepar maupun otot. Hal ini
terjadi apabila kadar glukosa di dalam darah meninggi.
Sebaliknya apabila kadar glukosa darah menurun, glikogen hepar
dimobilisasikan sehingga kadar glukosa darah akan naik kembali.
Gambar 10. Perubahan karbohidrat di dalam tubuh
30 fisiologi hewan

Insulin juga merangsang glukoneogenesis, yaitu mengubah lemak
atau protein menjadi glukosa.
Beberapa horrnon yang dihasilkan oleh hipofisis dan
kelenjar suprarenal merupakan pengatur-pengatur penting dari
metabolisme karbohidrat. Enzim sangat diperlukan pada proses-
proses kimiawi metabolisme zat-zat makanan. Vitamin-vitamin
secara tidak langsung berpengaruh pada metabolisme karbohidrat
ini. Tiamin (vitamin B1) diperlukan dalam proses dekarboksilase
karbohidrat. Kekurangan vitamin B1 akan menyebabkan
terhambatnya enzim-enzim dekarboksilase, sehingga asam piruvat
dan asam laktat tertimbun di dalam tubuh. Penyakit yang ditimbulkan
akibat defisiensi vitamin B1 itu dikenal sebagai penyakit beri-beri.
1.10 Pencernaan dan Metabolisme Karbohidrat pada
Ruminansia
a. Pencernaan Karbohidrat dalam Rumen
Terdapat perbedaan mendasar antara ruminansia dan
monogastrik dalam metabolisme karbohidrat, yaitu: jalur
metabolisme dan produk akhir yang dihasilkan. Tanaman makanan
hewan mengandung: 20-30% BK selulosa, 14-20% BK hemiselulosa,
dan kurang dari 10% BK pektin dimana 2-12% BK adalah lignin.
Gambar 11. Kegiatan ruminansi pada hewan ruminansia
31metabolisme karbohidrat

Ruminansia mempunyai mikroorganisme di dalam
retikulorumen yang mensekresikan enzim-enzim sehingga dapat
mencerna makanan yang masuk. Bagian terbesar karbohidrat terdiri
dari: yang mudah larut (gula dan pati) dan yang sukar larut (selulosa
dan hemiselulosa, misal hijauan dan limbah serat). Keduanya ini
difermentasikan oleh mikroba rumen membentuk asam lemak bebas
(volatil fat acid) atau VFA di dalam rumen dan retikulum. Pemecahan
karbohidrat menjadi VFA terjadi di rumen terdiri dari 2 tahap: 1).
hidrolisis ekstraseluler dari karbohidrat kompleks (selulosa,
hemiselulosa, pektin) menjadi oligosakarida rantai pendek terutama
disakarida (selobiosa, maltosa, pentosa) dan gula-gula sederhana.
Pemecahan oligosakarida dan gula-gula sederhana menjadi
VFA oleh aktivitas enzim intraseluler. Komposisi VFA terbanyak di
dalam cairan rumen adalah: asam asetat, propionat dan butirat,
sedangkan yang dalam jumlah kecil: asam format, isobutirat, valerat,
isovalerat dan kaproat. Pemecahan protein oleh bakteri juga
menghasilkan asam lemak berantai cabang yang terdapat dalam
jumlah kecil tersebut. Dalam pencernaan ini dihasilkan pula produk
ikutan berupa beberapa gas: metan (CH ), CO2 dan H2; yang 4
d i k e l u a r k a n d a r i t u b u h m e l a l u i p r o s e s e r u k t a s i
(belching/bersendawa). Sejumlah kecil karbohidrat yang dicerna dan
sebagian dari polimer karbohidrat yang lolos dari fermentasi mikroba
di perut depan akan masuk ke usus halus, dicerna dan diserap. Asam
lemak bebas (VFA) yang dominan (asam asetat, propionat, dan
butirat) akan diserap melalui dinding rumen, masuk ke dalam
sirkulasi darah dan di transportasikan ke jaringan tubuh hewan.
32 fisiologi hewan
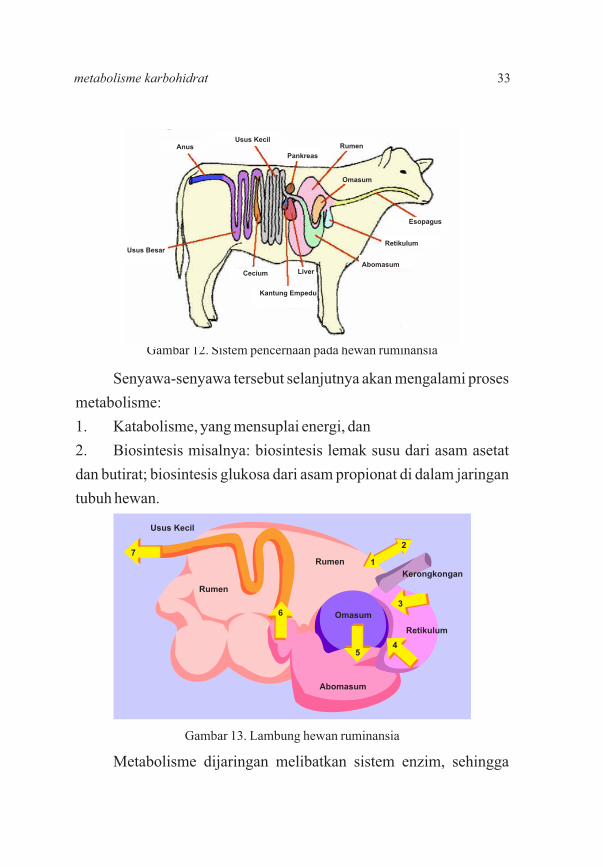
Senyawa-senyawa tersebut selanjutnya akan mengalami proses
metabolisme:
1. Katabolisme, yang mensuplai energi, dan
2. Biosintesis misalnya: biosintesis lemak susu dari asam asetat
dan butirat; biosintesis glukosa dari asam propionat di dalam jaringan
tubuh hewan.
Metabolisme dijaringan melibatkan sistem enzim, sehingga
Gambar 12. Sistem pencernaan pada hewan ruminansia
Gambar 13. Lambung hewan ruminansia
33metabolisme karbohidrat

produk akhir metabolisme dapat dimanfaatkan. Ruminansia dapat
mensintesis glukosa dari asam propionat di dalam rumen, dan
fungsinya sebagai energi (monogastrik: glukosa adalah sumber
energi utama) maka glukosa di jaringan menjadi terbatas (di dalam
darah: 40-70 mg%, sedang monogastrik 100 mg%). namun pada
hewan baru lahir (pre-ruminan) sama dengan monogastrik, glukosa
dalam darah: 100-120 mg%.
b. Pencernaan Karbohidrat di dalam Usus Ruminansia
Karbohidrat tercerna (pati, selulosa dan hemi selulosa) dan
polisakarida seluler dari mikroba yang lolos dari fermentasi rumen,
akan masuk ke dalam usus sebagai digesta, jumlahnya 10-20% dari
karbohidrat yang dicerna. Jumlah selulosa atau pati yang tahan dari
degradasi rumen dipengaruhi oleh pakan itu sendiri atau prosesing.
Misalnya pati dari jagung giling dapat dicerna ± 20% nya di usus
halus oleh enzim yang sama dengan monogastrik. Pencernaan pati di
usus halus menghasilkan energi yang dapat digunakan oleh induk
lebih efisien daripada didegradasi oleh mikroba rumen, dimana akan
hilang sebagai CH atau panas. Selulosa, hemiselulosa dan pati yang 4
lolos dari usus halus difermentasi di dalam sekum menjadi VFA, CO 2
dan CH dengan jalur yang sama dengan di dalam rumen. VFA yang 4
terbentuk di sekum ini di serap masuk ke dalam sirkulasi dan
digunakan di jaringan, hal yang sama terjadi di dalam rumen.
c. Energi yang Dihasilkan dari Pencernaan Karbohidrat
(Glukosa)
Dari dua tahap proses pencernaan karbohidrat didalam rumen,
34 fisiologi hewan

dihasilkan sumber energi berupa ATP seperti berikut :
Tahap1:
- Heksosa (senyawa-senyawa yang mempunyai atom karbon 6
buah) menghasilkan Piruvat + 4 (H) + 2 ATP
- Pentosa (senyawa-senyawa yang mempunyai atom karbon 5
buah) menghasilkan 1.67 Piruvat + 1.67 (H) + 1.67 ATP
Tahap 2:
- 2 Piruvat + 2H O ® 2 Asam Asetat + 2 CO2 + 2 H2 + 2 ATP2
- 2 Piruvat + 8 (H) ® 2 Asam Propionat + 2 H2O + 2 ATP
- 2 Piruvat + 4 (H) ® Asam butirat + 2 H2 + 2 CO2 + 2 ATP
Energi yang dihasilkan tersebut akan digunakan untuk hidup
pokok dan sintesis protein mikroba. Dengan cara demikian, mikroba
akan memperbanyak diri, sehingga pada gilirannya mikroba –
mikroba tersebut dapat dimanfaatkan sebagai sumber protein yang
bernilai hayati tinggi.
Gambar 14. Proses digesti pada hewan ruminansia
35metabolisme karbohidrat
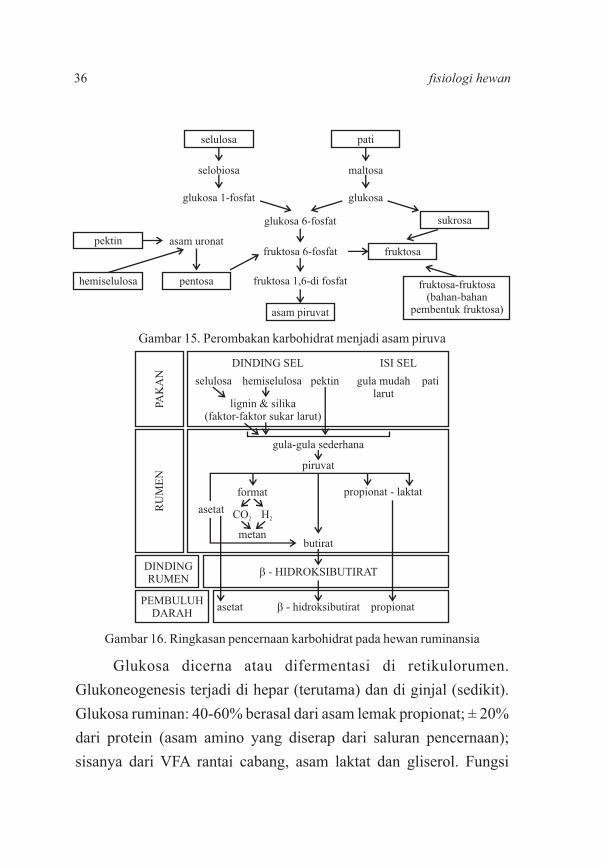
Glukosa dicerna atau difermentasi di retikulorumen.
Glukoneogenesis terjadi di hepar (terutama) dan di ginjal (sedikit).
Glukosa ruminan: 40-60% berasal dari asam lemak propionat; ± 20%
dari protein (asam amino yang diserap dari saluran pencernaan);
sisanya dari VFA rantai cabang, asam laktat dan gliserol. Fungsi
Gambar 15. Perombakan karbohidrat menjadi asam piruva
Gambar 16. Ringkasan pencernaan karbohidrat pada hewan ruminansia
36 fisiologi hewan

metabolit glukosa pada ruminansia, meliputi:
a. Sumber utama energi di jaringan saraf terutama di otak dan sel-
sel darah merah.
b. Untuk metabolisme otot dan produksi glikogen (persendian
energi di otot, hepar).
c. Prekursor utama dari pembentukan laktosa dan gliserol
(komponen lemak susu) dan suplai nutrisi untuk janin pada
hewan yang sedang laktasi. Kebutuhan glukosa ini meningkat
pada akhir kebuntingan.
Untuk pembentukan NADPH yang diperlukan untuk sintesis
asam lemak rantai panjang. NADPH berasal dari oksidasi glukosa
melalui jalur pentosa-fosfat
d. Pembentukan Adenosin Triphosfat (ATP)
Adenosin triphos fat (ATP) dihasilkan melalui beberapa jalur
reaksi, yaitu:
(a) Sistem anaerobik alaktat (jalur glikolisis)
(b) Sistem anaerobik laktat (glikolisis anaerobik/pembentukan
asam laktat)
(c) Sistem aerobik (reaksi oksidasi yang memerlukan oksigen)
e. Jalur Reaksi Anaerobik Alaktat (Jalur Glikolisis Alaktat)
Sistem anaerobik alaktat dikenal dengan sistem fosfagen.
Sistem ini mampu menghasilkan energi dengan intensitas yang
tinggi. Melalui reaksi ini fosfat terurai dari fosfokreatin disertai
dengan lepasnya kreatin. Fosfokreatin disimpan dalam sel-sel otot.
Fosfat yang dihasilkan digunakan untuk membentuk molekul ATP.
37metabolisme karbohidrat

f. Jalur Reaksi Anaerobik Laktat (Jalur Glikolisis Laktat)
Jalur reaksi ini menghasilkan energi dari proses katabolisme
molekul glukosa yang diperoleh dari karbohidrat tercerna atau
glikogen yang disimpan dalam sel-sel otot atau hepar.
Dalam reaksi ini tidak memerlukan oksigen untuk memecah
glukosa. Reaksi ini menghasilkan asam laktat sebagai akibat
rendahnya ketersediaan ATP dalam sel dan menghasilkan kondisi
kelelahan (fatigue), apabila kandungan asam laktat dalam darah yang
semakin meningkat.
g. Jalur Reaksi Siklus Kreb (Oksidasi-Fosforilasi)
Reaksi siklus Kreb diawali dengan terjadinya dekarboksilasi
piruvat menjadi asetil KoA dengan katalisasi enzim piruvat
dekarboksilase. Asetil KoA merupakan senyawa kunci yang
memegang peranan penting bagi siklus Kreb. Banyak enzim dengan
berbagai macam kofaktor yang terlibat dalam proses oksidasi-
fosforilasi dengan melibatkan rantai transport elektron. Hasil akhir
dari siklus Kreb adalah energi (FADH NADH dan GTP), CO dan 2, 2
H O.2
asam piruvat ketersediaan oksigen
oksigen tak adaatau tak cukup
oksigen adaataucukup
respirasianaerobik
respirasiaerobik
Gambar 17. Lintasan respirasi aerobik - anaerobik
38 fisiologi hewan
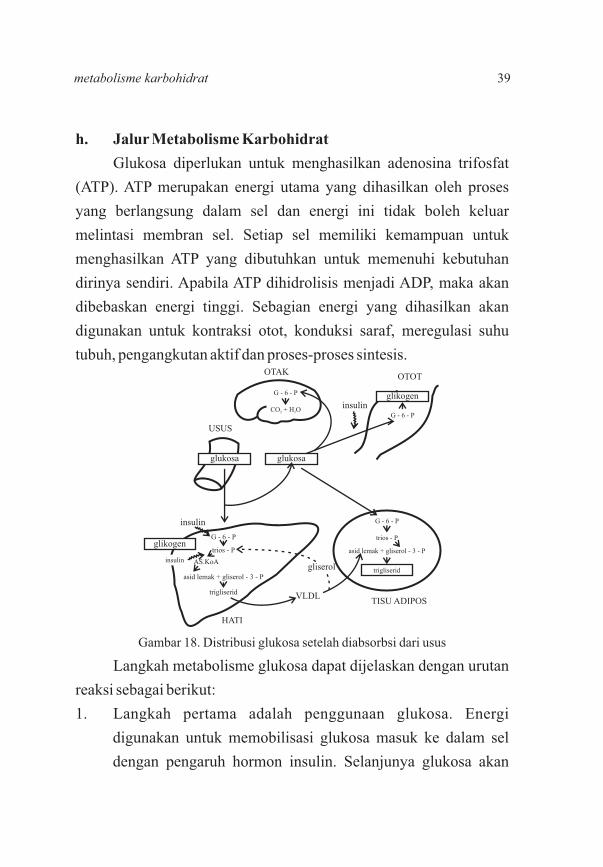
h. Jalur Metabolisme Karbohidrat
Glukosa diperlukan untuk menghasilkan adenosina trifosfat
(ATP). ATP merupakan energi utama yang dihasilkan oleh proses
yang berlangsung dalam sel dan energi ini tidak boleh keluar
melintasi membran sel. Setiap sel memiliki kemampuan untuk
menghasilkan ATP yang dibutuhkan untuk memenuhi kebutuhan
dirinya sendiri. Apabila ATP dihidrolisis menjadi ADP, maka akan
dibebaskan energi tinggi. Sebagian energi yang dihasilkan akan
digunakan untuk kontraksi otot, konduksi saraf, meregulasi suhu
tubuh, pengangkutan aktif dan proses-proses sintesis.
Langkah metabolisme glukosa dapat dijelaskan dengan urutan
reaksi sebagai berikut:
1. Langkah pertama adalah penggunaan glukosa. Energi
digunakan untuk memobilisasi glukosa masuk ke dalam sel
dengan pengaruh hormon insulin. Selanjunya glukosa akan
Gambar 18. Distribusi glukosa setelah diabsorbsi dari usus
39metabolisme karbohidrat

dikatabolisme oleh enzim heksokinase menjadi glukosa 6-
fosfat dengan melibatkan hidrolisis ATP menjadi ADP dan Pi.
Heksokinase
Glukosa + ATP Glukosa 6-Phospat +
ADP + Pi
Glukosa 6-fosfat merupakan senyawa perantara untuk berbagai
jalur metabolisme glukosa. Ada dua jalur utama proses
pemecahan glukosa-6-fosfat, yaitu: jalur glikolisis atau jalur
Embden-Meyerhof dan jalur pentosa fosfat atau jalur Shunt.
Glukosa-6-fosfat dapat diubah menjadi glikogen, proses ini
memerlukan sistem enzim yang rumit.
2. Langkah kedua adalah jalur glikolisis yang mengubah satu
molekul glukosa (6C) menjadi 2 molekul asam piruvat (3C).
Proses ini bersifat anaerob, yaitu tidak memerlukan oksigen
dan terjadi di dalam sitoplasma sel. Koenzim nikotinamida
adenina dinukleotida (NAD) merupakan molekul pembawa
yang menerima hidrogen yang dibebaskan selama glikolisis.
Jalur glikolisis menghasilkan dua molekul ATP untuk setiap
molekul glukosa. Proses glikolisis dapat ditulis sebagai berikut:
C H 0 + 2 NAD + 2ADP + Pi � � � � � � � � �2C H 0 + 6 12 6 3 4 3
2NADH + 2ATP2
Nasib asam piruvat tergantung pada ketersediaan oksigen dan
ATP. Jika ketersediaan oksigen dan ATP tidak memenuhi syarat
40 fisiologi hewan

maka piruvat akan diubah menjadi asam laktat. Dengan
ketersediaan oksigen yang optimal, piruvat akan mengalami
proses oksidasi-karboksilasi menghasilkan asetil (2C). Asetil
selanjutnya akan bereaksi dengan koenzim A dengan bantuan
ATP dan vitamin B membentuk asetil Coenzim A. Selanjutnya
asetil CoA akan masuk ke dalam siklus Kreb. Jalur siklus Kreb
terdiri dari proses-proses kimia yang terlibat dalam proses
oksidasi secara lengkap asetil CoA. Proses ini memerlukan
oksigen (aerob) dan berlangsung dalam organel mitokondria
sel. NAD bertindak sebagai penerima atom hidrogen
membentuk NADH . Dalam jalur ini elektron diangkut dalam 2
bentuk hidrogen (elektron + proton) dan bukan sebagai elektron
bebas.
3. Jalur terakhir dari proses oksidasi-fosforilasi yang melibatkan
rantai transport elektron akan menghasilkan energi (FADH , 2
NADH, GTP), karbondioksida (CO ) dan molekul air (H O)2 2
Gambar 19. Jalur metabolisme glukosa
41metabolisme karbohidrat

Respirasi seluler adalah proses yang melibatkan sistem rantai
transport elektron dan proses oksidasi-fosforilasi. Hidrogen yang
berasal dari NADH akan memasuki sistem transport elektron, dengan 2
melibatkan enzim sitokrom oksidase (enzim yang mengandungi
besi). Rantai transport elektron terjadi dengan melibatkan proses
42
Gambar 20. Proses glikolisis
fisiologi hewan

reduksi dan oksidasi secara bertahap hingga akhirnya dihasilkan
molekul oksigen. Selama proses rantai transport elektron, energi
bebas elektron digunakan untuk mengubah ADP menjadi ATP
(oksidasi-fosforilasi). Sebanyak 36 ATP dihasilkan dari rantai
transport elektron. Satu molekul glukosa yang dioksidasi secara
lengkap menghasilkan total 38 ATP (2 ATP dihasilkan dalam jalur
glikolisis dan 36 ATP dalam proses rantai transport elektron dan
proses oksidasi-fosforilasi (C H O + 6O > 6CO + 6H 0 + Energi: 38 6 12 6 2 2 2
ATP). Kurang lebih 40% energi yang dihasilkan melalui proses
glikolisis dan proses rantai transport elektron dan proses oksidasi-
fosforilasi disimpan dalam bentuk ATP dan sisanya hilang selama
proses katabolisme berlangsung.
Jalur pentosa fosfat atau heksosa monofosfat Shunt adalah jalur
sekunder yang berlangsung juga pada semua jaringan tubuh, namun
43metabolisme karbohidrat
Gambar 21. Sistem transport elektron dalam siklus asam sitrat (siklus kreb)

tidak menghasilkan energi dalam bentuk ATP. Namun demikian, jalur
metabolisme ini merupakan jalur bagi mekanisme sintesis gula
ribosa, yaitu karbohidrat yang merupakan komponen dalam
nukleotida (DNA, RNA). Selain itu jalur Shunt ini juga menghasilkan
NADPH yang digunakan pembentukan lipid dan membantu menjaga
keutuhan (integrasi) membran sel darah merah.
i. Siklus Kreb
Siklus Kreb merupakan akhir dari jalur katabolisme glukosa,
lemak dan asam-asam amino. Glukosa, lemak dan asam-asam amino
di dalam siklus Kreb akan dioksidasi menghasilkan CO , H O dan 2 2
menghasilkan sejumlah besar energi dalam bentuk ATP. Beberapa
senyawa intermidiet dalam siklus Kreb dapat dikonversi menjadi
glukosa melalui proses yang disebut glukoneogenesis dan beberapa
dapat dikonversi menjadi asam-asam amino tertentu melalui proses
transaminasi. Energi yang dilepaskan dari makanan kurang lebih
90%, melalui jalur siklus Kreb.
Reaksi siklus Kreb terjadi di dalam mitokondria dengan
melibatkan proses oksidasi fosforilasi dan transport elektron yang
menghasilkan sejumlah energi. Reaksi oksidasi dalam siklus Kreb
sesungguhnya merupakan reaksi dehidrogenasi, dimana enzim akan
mengkatalisis proses pelepasan 2 atom hidrogen untuk kosubstrat
penerimanya, yaitu Nikotinamid Adenin Denukleotida (NAD) atau
Flavin Adenin Denukleotida (FAD). NADH dan FADH2 dalam
bentuk reduksi selanjutnya akan dioksidasi dengan oksigen melalui
rantai transport elektron. Selain itu siklus Kreb juga menghasilkan
banyak karbondioksida melalui reaksi dekarboksilasi.
44 fisiologi hewan

Siklus Kreb dimulai dengan kondensasi asetil KoA dengan
oksaloasetat untuk membentuk sitrat. Asetil KoA dihasilkan dari
berbagai macam sumber, seperti asam-asam lemak, glukosa dan
asam-asam amino tertentu. Asam piruvat dihasilkan dari glukosa
melalui proses glikolisis dan selanjutnya dikonversi menjadi asetil
KoA. Asam piruvat merupakan penghubung proses glikolisis yang
terjadi dalam sitoplasma dengan siklus Kreb yang terjadi dalam
mitokondria.
Piruvat dehidrogenase adalah enzim komplek yang terdapat
dalam organel mitokondria dan merupakan salah satu sistem
multienzim dengan berbagai macam kofaktor yang berperan dalam
konversi asam piruvat menjadi asetil KoA. Enzim-enzim yang lain
adalah piruvat dekarboksilase, dihidrolipoil dehidrogenase dan
dihidrolipoil transasetilse. Adapun kofaktor-kofaktornya, antara lain
Koenzim A, TPP, Mg2+, NAD, FAD dan asam lipoid. Berikut reaksi
konversi asam piruvat menjadi asetil KoA yang dikatalisis oleh enzim
piruvat dehidrogenase:
45metabolisme karbohidrat
Gambar 22. Proses dekarboksilasi piruvat
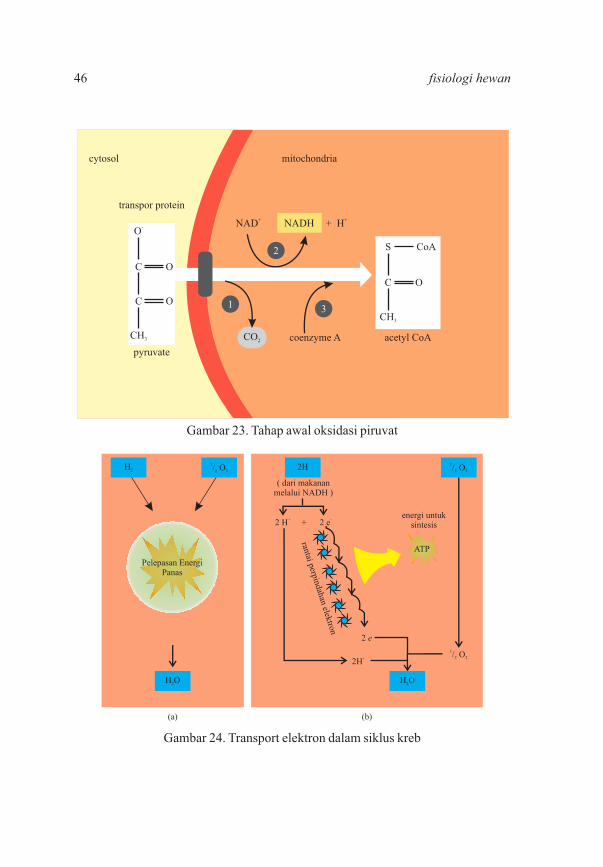
Gambar 24. Transport elektron dalam siklus kreb
Gambar 23. Tahap awal oksidasi piruvat
46 fisiologi hewan

Produk bersih dekarboksilasi dan dehidrogenasi adalah
konversi piruvat menjadi asetil CoA, dimana terjadi konversi NAD + +menjadi NADH dan H . Konversi piruvat menjadi asetil KoA
merupakan reaksi yang bersifat irreversibel yang menghasilkan
energi karena terjadi reoksidasi NADH oleh transport elektron yang
menghasilkan 3 molekul ATP. Dalam siklus terdapat beberapa enzim
yang terlibat dalam siklus tersebut, antara lain:
1. Enzim sitrat sintetase: enzim yang mengkatalisis pembentukan
sitrat dari asetil CoA dan oksaloasetat. Enzim ini diregulasi
secara negatif oleh ATP.
2. Sitrat kemudian mengalami isomerisasi menjadi isositrat oleh
enzim akonitase melalui penambahan gugus OH.
3. Isositrat dehidrogenase: enzim yang mengkonversi perubahan
isositrat menjadi α ketoglutarat melalui proses dekarboksilasi
dan dehidrogenasi. Molekul CO dilepaskan dan NAD 2
direduksi menjadi NADH. Reaksi ini dimodulasi secara positif
oleh ADP dan diregulasi secara negatif oleh NADH (NADH
yang diproduksi pertama kali dalam siklus Kreb).
4. Alfa ketoglutarat dehidrogenase: enzim yang mengkonversi
perubahan α ketoglutarat menjadi suksinil CoA melalui proses
karboksilasi dan dehidrogenasi dengan melibatkan sistem
multienzim dengan berbagai macam kofaktor.
Gambar 25. Proses oksidasi alfa ketoglutara
47metabolisme karbohidrat
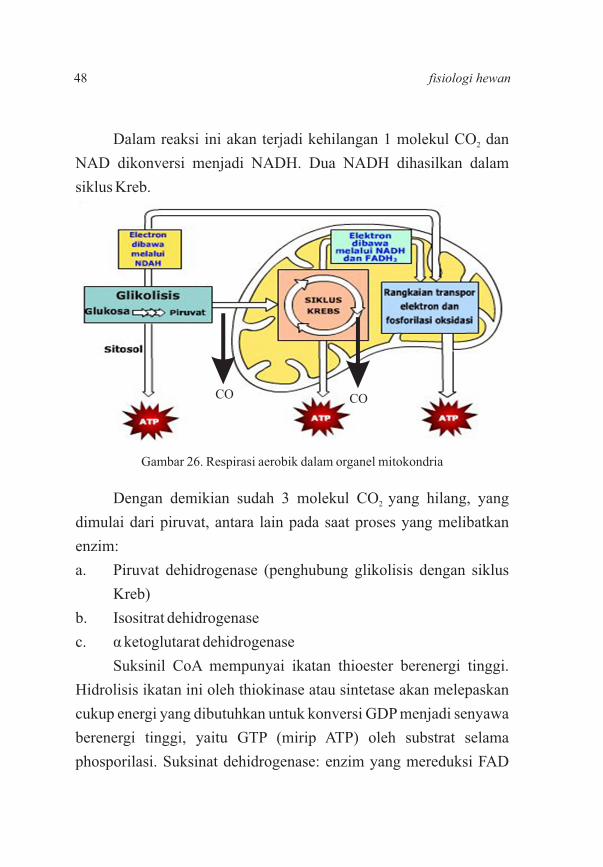
Dalam reaksi ini akan terjadi kehilangan 1 molekul CO dan 2
NAD dikonversi menjadi NADH. Dua NADH dihasilkan dalam
siklus Kreb.
Dengan demikian sudah 3 molekul CO yang hilang, yang 2
dimulai dari piruvat, antara lain pada saat proses yang melibatkan
enzim:
a. Piruvat dehidrogenase (penghubung glikolisis dengan siklus
Kreb)
b. Isositrat dehidrogenase
c. α ketoglutarat dehidrogenase
Suksinil CoA mempunyai ikatan thioester berenergi tinggi.
Hidrolisis ikatan ini oleh thiokinase atau sintetase akan melepaskan
cukup energi yang dibutuhkan untuk konversi GDP menjadi senyawa
berenergi tinggi, yaitu GTP (mirip ATP) oleh substrat selama
phosporilasi. Suksinat dehidrogenase: enzim yang mereduksi FAD
Gambar 26. Respirasi aerobik dalam organel mitokondria
48 fisiologi hewan
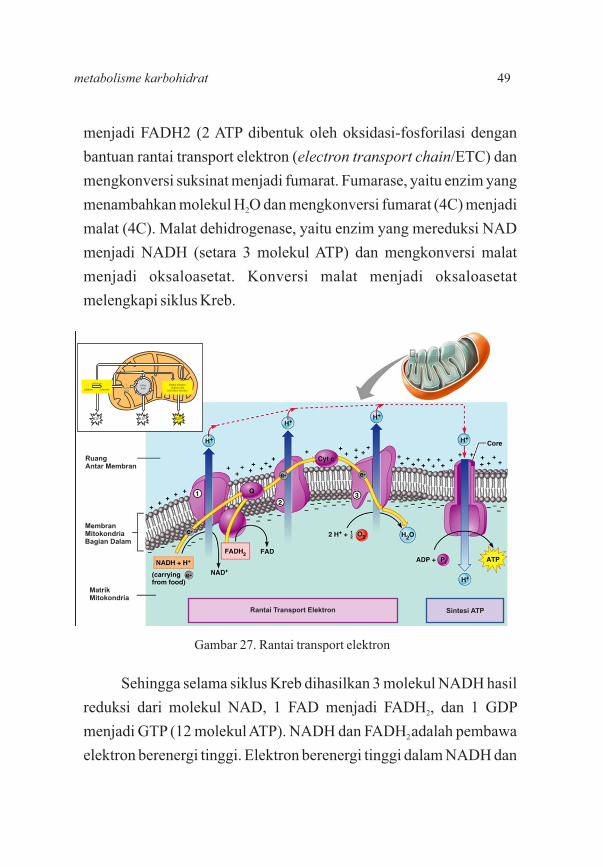
menjadi FADH2 (2 ATP dibentuk oleh oksidasi-fosforilasi dengan
bantuan rantai transport elektron (electron transport chain/ETC) dan
mengkonversi suksinat menjadi fumarat. Fumarase, yaitu enzim yang
menambahkan molekul H O dan mengkonversi fumarat (4C) menjadi 2
malat (4C). Malat dehidrogenase, yaitu enzim yang mereduksi NAD
menjadi NADH (setara 3 molekul ATP) dan mengkonversi malat
menjadi oksaloasetat. Konversi malat menjadi oksaloasetat
melengkapi siklus Kreb.
Sehingga selama siklus Kreb dihasilkan 3 molekul NADH hasil
reduksi dari molekul NAD, 1 FAD menjadi FADH , dan 1 GDP 2
menjadi GTP (12 molekul ATP). NADH dan FADH adalah pembawa 2
elektron berenergi tinggi. Elektron berenergi tinggi dalam NADH dan
Gambar 27. Rantai transport elektron
49metabolisme karbohidrat
RuangAntar Membran
MembranMitokondriaBagian Dalam
MatrikMitokondria
Rantai Transport Elektron Sintesi ATP

FADH dilewatkan setahap demi setahap ketingkat energi yang lebih 2
rendah dan akhirnya diterima oksigen (O ). Oksidasi 1 molekul asetil 2
CoA dalam siklus Kreb akan menghasilkan total energi 12 molekul
ATP, dengan rincian sebagai berikut:
a. 3 molekul ATP dalam reaksi yang melibatkan isositrat
dehidrogenase
b. 3 molekul ATP dalam reaksi yang melibatkan α ketoglutarat
dehidrogense
c. 1 molekul GTP (ATP) dalam reaksi yang melibatkan suksinil
CoA sintetase
d. 2 molekul ATP dalam reaksi yang melibatkan suksinat
dehidrogenase
e. 3 molekul ATP dalam reaksi yang melibatkan malat
dehidrogenase
Beradasarkan perhitungan katabolisme 1 molekul glukosa akan
dihasilkan 1 molekul asetil CoA. Karena setiap asetil CoA
menghasilkan 12 molekul ATP, sehingga total ATP selama siklus
Kreb adalah 24 molekul ATP.�� Oksidasi lengkap glukosa akan menghasilkan karbondioksida
(CO ) dan molekul air (H O). Proses ini terjadi melalui kombinasi 2 2
reaksi berurutan jalur glikolisis dan siklus Kreb.
C H O + O � 6CO + 6 H O + Energi6 12 6 2 2 2
Rincian jumlah ATP yang dihasilkan pada jalur glikolisis dan siklus
Kreb:
50 fisiologi hewan
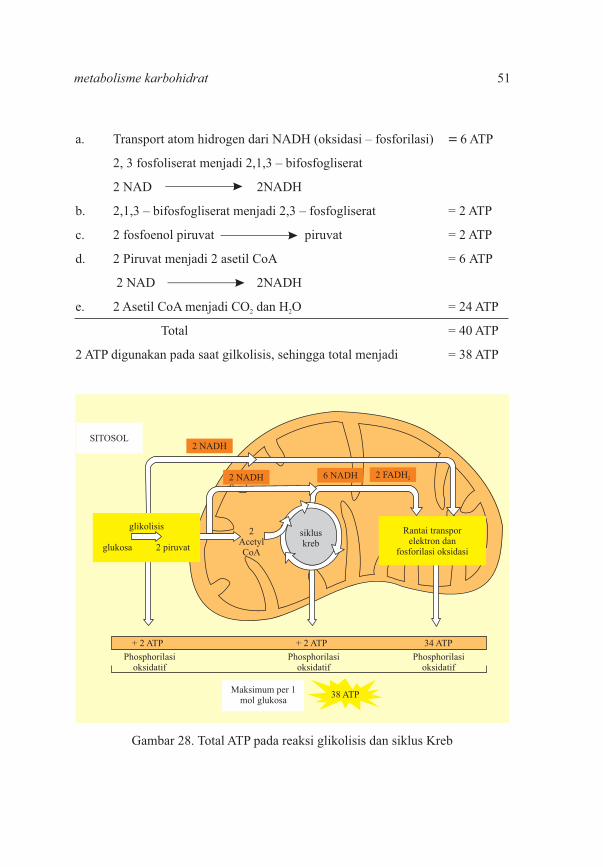
a. Transport atom hidrogen dari NADH (oksidasi – fosforilasi)� =�6 ATP
2, 3 fosfoliserat menjadi 2,1,3 – bifosfogliserat
2 NAD� � � 2NADH
b. 2,1,3 – bifosfogliserat menjadi 2,3 – fosfogliserat = 2 ATP
c. 2 fosfoenol piruvat� � piruvat = 2 ATP
d. 2 Piruvat menjadi 2 asetil CoA� = 6 ATP
2 NAD 2NADH
e. 2 Asetil CoA menjadi CO dan H O�� � � = 24 ATP2 2
Total� � � � � � = 40 ATP
2 ATP digunakan pada saat gilkolisis, sehingga total menjadi = 38 ATP
Gambar 28. Total ATP pada reaksi glikolisis dan siklus Kreb
51metabolisme karbohidrat

j. Reaksi Anaplerotik
- Asetil Coa + Oksaloasetat� � � Sitrat
(2C) (4C) (6C)
- Asetik CoA dihasilkan melalui oksidasi asam lemak dan
katabolisme asam amino yang merupakan turunan piruvat
hasil proses glikolisis.
- Jumlah asetil CoA dan oksaloasetat selalu mengarah pada
proses keseimbangan dengan membentuk sitrat.
- Keseimbangan tersebut penting agar pembentukan senyawa
intermidiet dapat berlanjut secara bertahap dalam siklus Kreb
hingga akhirnya terbentuk kembali oksaloasetat (replenish
oxaloacetat).
- Oksaloasetat, fumarat, suksinil CoA dan α ketoglutarat dapat
dibentuk dari asam-asam amino tertentu melalui jalur
transaminasi
Asam-asam amino (kerangka karbon 4)
Oksaloasetat atau senyawa intermidier lain dalam siklus Kreb
k. Reaksi Bersambung Anaplerotik
Mekanisme yang paling penting untuk menjamin suplai
oksaloasetat dalam siklus Kreb adalah karboksilasi piruvat oleh
enzim piruvat karboksilase melalui pembentukan asetil CoA. Satu
molekul ATP digunakan dalam reaksi ini. Enzim piruvat karboksilase
meregulasi secara positif perubahan piruvat menjadi asetil CoA.
52 fisiologi hewan
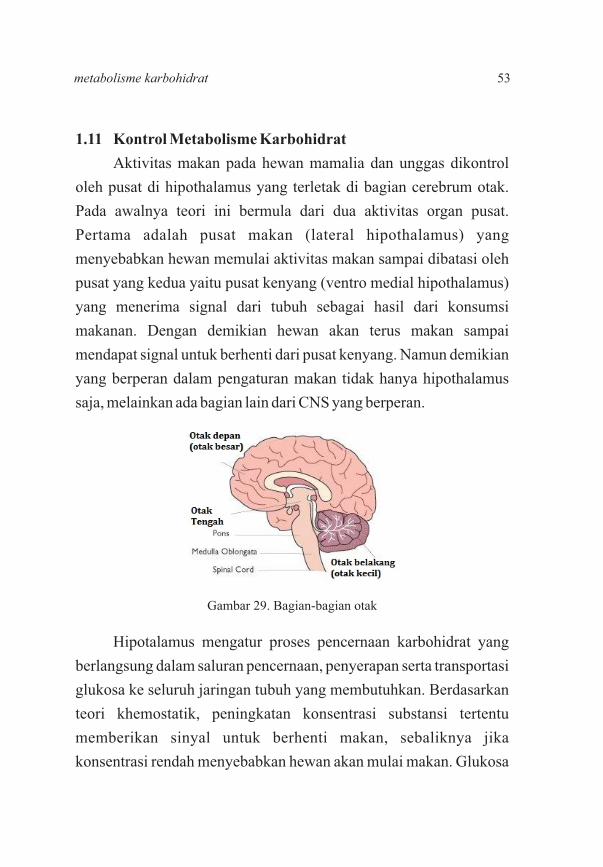
1.11 Kontrol Metabolisme Karbohidrat
Aktivitas makan pada hewan mamalia dan unggas dikontrol
oleh pusat di hipothalamus yang terletak di bagian cerebrum otak.
Pada awalnya teori ini bermula dari dua aktivitas organ pusat.
Pertama adalah pusat makan (lateral hipothalamus) yang
menyebabkan hewan memulai aktivitas makan sampai dibatasi oleh
pusat yang kedua yaitu pusat kenyang (ventro medial hipothalamus)
yang menerima signal dari tubuh sebagai hasil dari konsumsi
makanan. Dengan demikian hewan akan terus makan sampai
mendapat signal untuk berhenti dari pusat kenyang. Namun demikian
yang berperan dalam pengaturan makan tidak hanya hipothalamus
saja, melainkan ada bagian lain dari CNS yang berperan.
Hipotalamus mengatur proses pencernaan karbohidrat yang
berlangsung dalam saluran pencernaan, penyerapan serta transportasi
glukosa ke seluruh jaringan tubuh yang membutuhkan. Berdasarkan
teori khemostatik, peningkatan konsentrasi substansi tertentu
memberikan sinyal untuk berhenti makan, sebaliknya jika
konsentrasi rendah menyebabkan hewan akan mulai makan. Glukosa
Gambar 29. Bagian-bagian otak
53metabolisme karbohidrat

merupakan indikator yang menentukan kenyang atau lapar bagi
hewan. Jika konsentrasi glukosa darah rendah dan disuntik dengan
insulin maka hewan akan merasa lapar. Sebaliknya setelah makan
konsentrasi glukosa akan meningkat dan hewan akan berhenti makan.
Mekanisme pengaturan gula darah diilustrasikan pada Gambar 30.
Insulin dan glukagon meregulasi produksi glukosa ruminansia
dan mengontrol homeostasis kadar glukosa dalam darah. Insulin
diproduksi oleh sel-sel β pulau Langerhans pankreas yang berfungsi
menurunkan produksi glukosa dari propionat dan prekursor glukosa
lainnya. Walaupun mempunyai peran yang berbeda, glukagon dan
insulin secara sinergi mendukung proses glukogenesis dari prekursor
glukosa dan meregulasi pelepasan glukosa hasil metabolisme
glikogen di sel hepar ke sirkulasi darah.
Reseptor glukosa diduga terletak di hipothalamus.
Hipotalamus dapat memonitor kadar glukosa baik di pembuluh vena
maupun arteri. Penelitian yang lain menunjukan bahwa reseptor
tersebut terdapat pada saluran pencernaan dan hepar. Sebagai bukti
bahwa jika glukosa disuntikan di usus atau di sistem portal hepatik
menyebabkan menurunnya intake pakan yang lebih besar
dibandingkan jika disuntikan di sirkulasi periperal.
Dugaan lain yang mengatur komunikasi saluran pencernaan
dan otak adalah hormon peptida cholecystokinin. Hormon ini
dikeluarkan jika asam amino dan asam-asam lemak mencapai
duodenum, dan ini merupakan kerja hipothalamus.
Tanpa bantuan hormon, kadar gula darah akan mengalami
fluktuasi yang besar. Kadar gula darah akan segera meningkat
sesudah makan, dan sebaliknya bila tidak ada asupan makanan
54 fisiologi hewan

pada periode tertentu, kadar gula darah akan turun sangat rendah.
Untuk mencegah terjadinya fluktuasi yang membahayakan ini, tubuh
akan meregulasi glukosa darah dengan menggunakan hormon insulin
dan glukagon. Hormon insulin disekresikan oleh sel-sel β pankreas
apabila kadar gula darah meninggi (hiperglikemia), yang biasanya
terjadi sesudah rnakan, seperti nasi, roti, gula, dan lain sebagainya.
Peninggian kadar gula darah ini, akan merangsang sekresi insulin
dari sel-sel β di pulau Langerhans pankreas. Sekresi Insulin ini
berlangsung dalam dua fase, pada fase pertama kadar insulin
melonjak tinggi seketika. Hal ini terjadi 10 menit sesudah kenaikan
kadar gula darah, dan dimungkinkan karena ada simpanan insulin
dalam granula. Kemudian terjadi fase ke dua yang bersifat lambat,
berlangsung selama lebih dari 10 menit sampai 2 jam. Dalam jam
pertama sesudah makan, gula darah meningkat sampai 160,11
mg%, dan kemudian menurun lagi berkat pengaruh insulin, sehingga
2 jam sesudah makan kadar gula darah normal kembali, yakni 120
mg%. Insulin akan merangsang pengambilan glukosa oleh jaringan
dan kemudian memecahnya menjadi energi, menyimpannya dalam
bentuk glikogen dan mengubahnya menjadi lemak. Dengan proses
tersebut diatas, kadar gula darah akan menurun dan kembali
normal 2 sampai 2 ½ jam sesudah makan.
Sebaliknya bila kadar gula darah rendah, hormon glukagon
yang dihasilkan sel-sel α pulau Langerhans pankreas akan
menstimulasi sintesa glukosa dari asam amino, menyebabkan
terlepasnya glikogen dari hepar, yang akan rneninggikan kadar
gula darah. Jadi, aktifitas hormon insulin dan glukagon berlawanan
satu sama lain. Ada juga hormon lain yang dapat membantu
55metabolisme karbohidrat

meninggikan kadar gula darah, salah satu yang paling penting
adalah epinefrin (adrenalin) yang merangsang pembebasan
glukosa dari glikogen. Hormon epinefrin ini akan disekresikan
pada situasi dimana tubuh dalam keadaan stress ataupun dalarn
keadaan bahaya. Peningkatannya akan menaikkan kadar gula
darah, yang akan membantu tubuh untuk berkelahi atau berlari
mengambil langkah seribu.
Kasus diabetes melitus dengan defesiensi insulin, glukosa tidak
dapat masuk ke dalam sel-sel, sehingga kadar gula darah meninggi,
namun timbunan glukosa tersebut tidak dapat dimanfaatkan
untuk menghasilkan energi untuk keperluan sel-sel yang
membutuhkannya. Glukosa yang tertumpuk itu dibuang melalui
ginjal ke dalam urine, sehingga terjadi glukosuria. Karena glukosa
tidak dapat dipergunakan sebagai penghasil energi, maka lemak dan
protein lebih banyak dipecah untuk menghasilkan energi yang
dibutuhkan, sehingga terjadi peningkatan glukoneogenesis.
Peningkatan pemecahan asam lemak akan menghasilkan benda-
benda keton (keton bodies), sehingga bila keton bodies ini meninggi
dalam darah (ketosis) akan mengakibatkan penurunan pH darah,
sehingga terjadi asidosis.
1.12 Defisiensi Karbohidrat
Manusia membutuhkan karbohidrat dalam jumlah tertentu
setiap harinya. Walaupun tubuh tidak membutuhkan dalam jumlah
yang khusus, kekurangan karbohidrat yang sangat parah akan
menimbulkan masalah. Diperlukan sekitar 2 gram karbohidrat per
kg berat badan sehari untuk mencegah terjadinya ketosis.
56 fisiologi hewan

Secara keseluruhan tubuh harus mempertahankan
keseimbangan tertentu dalam utilisasi karbohidrat, lemak dan
protein sebagai sumber energi. Jika asupan karbohidrat ditiadakan,
maka cadangan lemak dalam jaringan adiposa akan dimobilisasi
sedemikian cepatnya, sehingga tubuh tidak dapat mengoksidasi
karbohidrat seluruhnya menjadi CO dan H O. Sebagian dari hasil 2 2
pemecahan lemak itu akan diubah menjadi substansi yang disebut
dengan keton bodies. Walaupun tubuh dapat menggunakan keton
bodies ini sebagai penghasil energi dan dieksresikan melalui urine,
produksi dalam jumlah besar akan terjadi penumpukan keton
bodies di dalam darah dan mengakibatkan terjadinya ketosis. Hal ini
sangat berbahaya dan dapat terjadi pada penderita diabetes
mellitus yang tidak terkontrol. Jumlah asupan karbohidrat juga
mempengaruhi penggunaan protein sebagai penghasil energi. Jika
asupan karbohidrat rendah, tubuh akan memecah asam amino
untuk menghasilkan energi dan mensintesa glukosa tubuh,
sehingga jaringan yang membutuhkan gula ini akan mampu
menjalankan fungsinya. Oleh karena sebagian protein tubuh
digunakan untuk tujuan ini, maka sedikit karbohidrat dapat
menyebabkan pemecahan dari jaringan otot untuk menghasilkan
energi.
Gejala yang timbul akibat asupan karbohidrat yang rendah
adalah fatigue, dehidrasi, mual, nafsu makan berkurang, dan tekanan
darah kadang-kadang turun dengan mendadak sewaktu bangkit dari
posisi berbaring (hipotensi ortostatik). Asupan karbohidrat yang
adekuat, penting untuk mempertahankan cadangan glikogen yang
dibutuhkan pada aktifitas fisik jangka panjang. Peningkatan glikogen
57metabolisme karbohidrat

otot dengan adanya proses penumpukan karbohidrat akan menambah
stamina 30-60 menit lebih lama.
1.13 Hubungan Konsumsi Karbohidrat dengan Karies Dentis
Hubungan antara konsumsi karbohidrat dengan terjadinya
karies dentis ada kaitannya dengan pembentukan plak pada
permukaan gigi. Plak terbentuk dari sisa-sisa makanan yang melekat
di sela-sela gigi. Plak ini akhirnya akan ditumbuhi bakteri yang dapat
mengubah pH rongga mulut menurun sampai dengan pH 4,5. Pada
keadaan demikian maka struktur email gigi akan terlarut (email
tidak larut pada pH 5,41). Konsumsi gula murni (permen, coklat,
karamel) sering, akan menyebabkan keasaman rongga mulut menjadi
permanen, sehingga semakin banyak email yang terlarut. Kerusakan
email yang parah, disebut dengan karies dentis. Dari berbagai
penelelitian sukrosa (gula bit dan gula tebu) mempunyai efek
kariogenik lebih tinggi dibandingkan dengan fruktosa, glukosa dan
maltosa. Sedangkan karbohidrat kompleks seperti amilum dan
dekstrin, efek kariogeniknya tidak ada sama sekali.
1.14 SOAL -SOAL LATIHAN
1. Jelaskan mengapa karbohidrat merupakan sumber energi
paling penting bagi tubuh, lebih penting dibanding lemak dan
protein ?
2. Melalui jalur anaerobik glukosa dapat dikonversi menjadi
gliserol yang merupakan penyusun komponen lemak, mengapa
demikian ?
3. Sebutkan beberapa enzim yang terlibat dalam proses glikolisis
58 fisiologi hewan

didalam sitosol, jelaskan fungsinya masing – masing !
4. Asam piruvat merupakan produk akhir proses glikolisis.
Sebutkan perbedaan kondisi fisiologis yang menyebabkan
piruvat mengalami konversi menjadi asam laktat dan
mengalami dekarboksilasi menjadi asetil CoA dengan bantuan
kofaktor enzim A ?
5. Sebutkan total ATP yang dihasilkan dari proses glikolisis dan
siklus Kreb ! Berikan penjelasan terhadap jawaban yang anda
berikan !
6. Jelaskan perbedaan kondisi fisiologis yang ditimbulkan
sebagai akibat asupan karbohidrat yang rendah dan asupan
kerbohidrat adekuat !
7. Perubahan asam piruvat menjadi asetil CoA menghasilkan
NADH yang setara dengan 3 molekul ATP, namun demikian
NADH belum dapat digunakan sebagai sumber energi,
mengapa demikian ?
8. Jelaskan kontrol kadar gula darah dalam kondisi normal
(homeostasis) dan kondisi patologis !
9. Jelaskan mekanisme insulin dalam menurunkan kadar gula
darah ?
10. Terakumulasinya glukosa dalam darah akan menyebabkan
suatu kondisi patologis yang disebut diabetes melitus.
Berkaitan dengan kondisi patologis ini terdapat 2 tipe diabetes,
yaitu diabetes melitus tergantung insulin (insulin dependent
diabetes melitus) dan diabetes melitus tidak tergantung insulin
(non – insulin dependent diabetes melitus). Berikan penjelasan
terhadap 2 kondisi tersebut !
59metabolisme karbohidrat


2.1 Pengertian Lemak
Lemak disebut juga lipid, dua istilah yang berbeda namun pada
dasarnya memiliki esensi yang sama. Lipid bersifat lebih umum,
sedangkan lemak mempunyai arti yang lebih khusus. Lipid adalah
senyawa heterogen yang terdiri atas lemak dan minyak yang terdapat
pada makanan/tubuh manusia. Lipid mempunyai sifat tidak larut
dalam pelarut non-polar, seperti etanol, eter, kloroform, dan benzena.
Adapun lemak (triasil gliserol) merupakan senyawa biologis yang
terdiri atas tiga asam lemak utama yang masing-masing berikatan
dengan satu molekul gliserol. Lipid dan lemak dalam proses
metabolisme merupakan senyawa penting sebagai sumber energi
selain karbohidrat.
Selain sebagai sumber energi, lemak atau lipid juga berfungsi
untuk melindungi organ tubuh, berperan dalam pembentukan sel,
sebagai sumber asam lemak esensial, senyawa pengikat dan
pengangkut vitamin yang larut dalam lemak. Senyawa ini juga
METABOLISME LIPID
2

berfungsi sebagi prekursor hormon dan berperan dalam pensinyalan
sel, penghematan protein, memberi rasa kenyang, sebagai pelumas,
dan memelihara suhu tubuh. Lemak atau lipid merupakan komponen
utama membran sel dalam jaringan tubuh. Senyawa ini banyak
ditemukan dalam bentuk berasosiasi dengan air. Sel-sel hati memiliki
cadangan lipid yang berfungsi sebagai sumber energi dengan
kandungan energi dua kali lipat lebih besar dibanding glikogen dan
protein. Selama mengalami proses fisiologis, lipid akan mengalami
reduksi secara total untuk menghasilkan energi maksimal yang
digunakan untuk proses oksidasi. Lipid juga berperan penting dalam
menjamin ketersedian air selama kondisi hidrasi.
Lemak yang terdapat di dalam tubuh berasal dari makanan dan
hasil biosintesis oleh sel-sel hati. Berdasarkan sumbernya, lemak
dikelompokkan menjadi 3 macam:
a. Lemak sederhana
Lemak sederhana juga disebut lemak netral, terdiri dari
monogliserida, digliserida, dan trigliserida, ester sterol, ester
non-sterol, ester vitamin A dan D.
b. Lemak majemuk (compound lipids)
Jenis lemak yang termasuk dalam kelompok lemak majemuk
adalah fosfolipid dan lipoprotein
c. Lemak turunan (derived lipids)
- asam lemak (fatty acids)
- sterol (kolesterol, ergosterol, dan sebagainya)
- hormon steroid, vitamin D, garam empedu
- karotenoid dan vitamin. A, E, K
62 fisiologi hewan

Berdasarkan fungsi biologis, lemak dapat diklasifikasikan
sebagai berikut:
1. Lemak simpanan (trigliserida) dalam jaringan, merupakan
simpanan energi utama dalam tubuh dan sebagai sumber zat
gizi esensial
2. Lemak struktural (fosfolipid dan kolesterol) dalam jaringan
lunak dan otak, terdapat dengan konsentrasi yang tinggi
Secara klinis, dikenal berbagai macam lemak tubuh, seperti
kolesterol, trigliserida (lemak netral), lipoprotein, dan asam lemak.
a. Kolesterol
Kolesterol adalah jenis lemak yang paling banyak ditemukan.
Kolesterol merupakan komponen utama pada struktur membran sel
tubuh dan sel saraf. Kolesterol merupakan bahan perantara untuk
pembentukan sejumlah komponen penting, seperti vitamin D (untuk
membentuk dan mempertahankan tulang), hormon seks (estrogen,
testosteron, dan progesteron) dan asam empedu (untuk fungsi
pencernaan ). Kolesterol tubuh berasal dari hasil pembentukan di
dalam tubuh (sekitar 500 mg/hari) dan dari makanan yang dimakan.
Pembentukan kolesterol di dalam tubuh, terutama terjadi di hati (50%
total sintesis) dan sisanya di usus, kulit, dan semua jaringan yang
mempunyai sel-sel berinti. Berbagai macam makanan yang banyak
mengandung kolesterol, meliputi daging (sapi, unggas), ikan, dan
produk susu. Makanan yang berasal dari daging hewan biasanya
banyak mengandung kolesterol, namun makanan yang berasal dari
tumbuh-tumbuhan sedikit/tidak mengandung kolesterol.
63metabolisme lipid

b. Trigliserida
Trigliserida atau triasilgliserol merupakan cadangan energi
yang sangat besar, baik dalam bentuk tereduksi atau anhidrat.
Oksidasi sempurna trigliserida menghasilkan energi 9 kkal/gram,
lebih banyak dibanding energi yang dihasilkan karbohidrat dan
protein yang rata-rata hanya 4 kkal/gram. Trigliserida banyak
ditemukan dalam kondisi tereduksi dengan tingkat reduksi yang
tinggi. Selain itu, trigliserida bersifat non-polar, sehingga dominan
terdapat dalam bentuk anhidrat, sedangkan protein dan karbohidrat
bersifat lebih polar, sehingga mudah mengalami hidratasi. Satu gram
karbohidrat (glikogen) dapat mengikat kurang lebih 2 gram molekul
air, sedangkan trigliserida anhidrat mampu mengikat dan menyimpan
energi, enam kali lebih banyak dibanding glikogen yang terhidratasi.
Hal ini merupakan bukti bahwa trigliserida merupakan bentuk
simpanan energi yang lebih utama dibanding glikogen. Dalam tubuh,
sel adiposa merupakan sel yang memiliki fungsi khusus, yaitu sebagai
tempay untuk sintesis dan penyimpanan trigliserida serta
memobilisasi trigliserida menjadi molekul sumber energi yang akan
dipindahkan ke jaringan lain oleh darah.
Trigliserida dalam bentuk ester gliserol banyak terdapat di alam
(98-99%) , baik sebagai lemak atau minyak. Trigliserida disintesis
dari tiga asam lemak dan gliserol. Jika terdapat satu asam lemak yang
berikatan dengan gliserol, maka akan terbentuk monogliserida.
Trigliserida dalam tubuh mempunyai peran penting sebagai sumber
energi. Di dalam tubuh trigliserida disimpan dalam bentuk lemak.
Apabila sel membutuhkan sumber energi, enzim lipase dalam sel
adiposa akan memecah trigliserida menjadi gliserol dan asam lemak,
64 fisiologi hewan

kemudian kedua senyawa ini masuk ke dalam pembuluh darah.
Setelah sampai pada sel target di dalam tubuh yang membutuhkan,
kedua senyawa tersebut digunakan untuk menghasilkan energi,
karbondioksida (CO ), dan air (H O). 2 2
c. Lipoprotein
Secara umum lemak tidak larut dalam air, sehingga secara
otomatis juga tidak larut dalam plasma darah. Agar lemak dapat
diangkut ke dalam peredaran darah, maka lemak harus dibuat larut,
yaitu dengan cara mengikatkannya pada protein yang larut dalam air.
Ikatan antara lemak (kolesterol, trigliserida, dan fosfolipid) dengan
protein membentuk lipoprotein (dari kata lipo = lemak dan protein).
Dengan demikian, lipoprotein adalah molekul komplek yang
merupakan ikatan antara lipid dan protein yang berfungsi dalam
proses transpor dari jaringan ke jaringan yang bersifat spesifik target
atau spesifik protein reseptor sel. Selain itu, lipoprotein berfungsi
mengangkut lemak dari tempat pembentukannya menuju tempat
penggunaannya. Ada beberapa jenis lipoprotein, antara lain:
kilomikron, VLDL (very low density lypoprotein), IDL (intermediate
density lypoprotein), LDL (low density lipoprotein) dan HDL (high
density lipoprotein)
d. Asam Lemak
Asam lemak merupakan senyawa hidrokarbon yang memiliki
rantai panjang dengan gugus karboksilat pada ujungnya. Asam lemak
memiliki empat peran utama. Pertama, asam lemak merupakan unit
penyusun fosfolipid dan glikolipid. Molekul-molekul ampifatik ini
65metabolisme lipid

merupakan komponen penting bagi membran biologi, baik membran
sel atau membran organel. Kedua, banyak protein dimodifikasi oleh
ikatan kovalen asam lemak, yang menempatkan protein-protein
tersebut ke lokasi-lokasinya pada membran. Ketiga, asam lemak
merupakan molekul sumber energi. Asam lemak disimpan dalam
bentuk triasilgliserol, yang merupakan ester gliserol yang tidak
bermuatan. Triasilgliserol disebut juga lemak netral atau trigliserida.
Keempat, derivat asam lemak berperan sebagai hormon dan
messenger intraseluler.
Asam lemak jarang terdapat bebas dan biasanya banyak
teradapat dalam bentuk ikatan ester dan merupakan asam organik.
Karakteristik asam lemak adalah memiliki rantai hidrokarbon lurus,
disatu ujung mempunyai gugus karboksil (COOH) dan diujung lain
mempunyai gugus metil (CH ) serta mempunyai rantai dengan atom 3
karbon (C) berjumlah genap, antara 4-22 atom C. Berdasarkan jumlah
atom karbon, asam lemak dibedakan menjadi 4 macam:
1. Asam lemak rantai pendek (short chain fatty acids): atom
karbon kurang dari 6
2. Asam lemak rantai sedang (medium chain fatty acids): atom
karbon antara 8-12
3. Asam lemak rantai panjang (long chain fatty acids): atom
karbon antara 14-18
4. Asam lemak rantai sangat panjang (very long chain fatty acids):
atom karbon lebih dari 20
Lemak hewani dan sebagian minyak nabati mengandung asam
lemak rantai panjang. Ikan banyak mengandung asam lemak rantai
sangat panjang Titik cair asam lemak meningkat dengan bertambah
66 fisiologi hewan

panjangnya rantai karbon (C). Lemak memiliki beberapa ciri spesifik
sesuai dengan struktur molekul yang dimiliki. Rantai asam lemak
alifatik adalah hidrofobik. Lemak secara umum akan membentuk
agregat untuk meminimalisasi daerah pada gugus hidrofobik
terekspose oleh pelarut. Unit paling sederhana dari molekul lemak
dalam air adalah misel; yaitu gliserol yang bersifat polar (gugus
kepala yang terdapat pada bagian permukaan) dan gugus hidrofobik
yang terdapat pada bagian dalam. Dalam molekul air, lemak
mempunyai struktur yang multilapis (multilayer)
2.2 Klasifikasi dan Nomenklatur Asam Lemak
Nama asam lemak secara sistematis berasal dari nama
hidrokarbon induknya dengan mensubstitusikan oat untuk akhiran a
terakhir. Misalnya, asam lemak jenuh C disebut asam oktadekanoat 18
sebab hidrokarbon induknya adalah oktadekana. Suatu asam lemak
C dengan satu ikatan rangkap disebut asam okta desinoat, dengan 18
dua ikatan rangkap disebut okta dienoat, dengan tiga ikatan rangkap,
okta trinoat. Simbol 18:0 menyatakan suatu asam lemak C tanpa 18
ikatan rangkap, sedangkan 18:2 menandakan adanya dua ikatan
rangkap. Nomor atom karbon pada asam lemak dimulai dari ujung
karboksil.
H C – (CH ) – CH – CH – C – OH3 2 n 2 2
Atom karbon kedua dan ketiga sering disebut sebagai α dan β.
Gugus metil pada ujung distal rantai disebut karbon ω. Posisi ikatan
rangkap diperlihatkan oleh simbol Δ diikuti oleh nomor superskrip.
67metabolisme lipid

Misalnya sis-Δ 9 berarti terdapat ikatan rangkap sis antara atom
karbon 9 dan 10; trans-Δ ² berarti terdapat ikatan rangkap trans antara
atom karbon 2 dan 3. Sebaliknya posisi ikatan rangkap dapat
dinyatakan dengan cara menghitung dari ujung distal, dengan atom
karbon ω (karbon metil) sebagai atom karbon nomor 1. struktur asam
lemak ω-3 misalnya, diperlihatkan di sebelah kiri. Asam lemak
terionisasi pada pH fisiologis, jadi lebih tepat bila asam lemak disebut
menurut bentuk karboksilatnya : misalnya palmitat atau
heksadekanoat.
Asam lemak jenuh (saturated fatty acid) memiliki rantai
karbon (C) mengikat semua atom hidrogen (H). Asam lemak tidak
jenuh (unsaturated fatty acids) memiliki satu atau lebih atom C yang
mempunyai ikatan rangkap yang dapat diikat oleh tambahan atom H.
Asam lemak tidak jenuh terdiri atas:
1. Asam lemak tidak jenuh tunggal (mono unsaturated fatty
acids): yaitu mempunyai satu ikatan rangkap
2. Asam lemak tidak jenuh ganda (poly unsaturated fatty acids):
mempunyai dua atau lebih ikatan rangkap
Lipid hewan merupakan asam lemak jenuh, sebagai contoh
asam palmitat yang memiliki karbon 16 (C16) dan asam stearat yang
memiliki karbon 18 (C18). Komposisi susu, 10% adalah asam lemak
rantai pendek, misalnya asam butirat yang memiliki aton karbon 4
(C4). Titik cair asam lemak tidak jenuh lebih rendah daripada asam
lemak jenuh. Asam lemak tidak jenuh mencair pada suhu ruang.
Minyak nabati terdiri dari asam palmitat, stearat, oleat, linoleat,
kecuali minyak kelapa dan minyak kelapa sawit yang banyak terdiri
dari asam lemak rantai sedang yang memiliki atom karbon 8-14 (C8-
68 fisiologi hewan

C14). Asam lemak dapat dinyatakan dengan nomenklatur umum,
istilah kimia atau nomenklatur pendek, yaitu dengan cara jumlah
atom C diikuti oleh titik dua (:) dan jumlah ikatan rangkap, kemudian
diantara 2 kurung ditulis posisi ikatan rangkap I terhadap ujung metil
(CH ). Asam linoleat mempuny 18 atom C, mempunyai 2 ikatan 3
rangkap dan posisi ikatan rangkap di atom C ke 6 dari ujung metil
(CH ) ditulis 18:2 (n-6), sekarang sering ditulis 18:2(ω-6).3
2.3 Asam Lemak Esensial
Burr dan Burr, (1929) menjelaskan tentang keberadaan asam
lemak esensial yang tidak dapat disintesis oleh tubuh, dan asam lemak
ini harus didatangkan dari luar tubuh melalui intake bahan makanan.
Asam lemak esensial mempunyai peranan vital dalam mendukung
proses metabolisme untuk keberlangsungan makhluk hidup. Contoh
asam lemak esensial adalah asam. linoleat (18:2 ω-6) dan asam
linolenat (18:3 ω-3). Kedua asam lemak ini dibutuhkan untuk
Gambar 30. Contoh nomenklatur asam lemak
69metabolisme lipid

pertumbuhan dan fungsi normal semua jaringan. Turunan asam
lemak dari kedua asam lemak esensial tersebut, yaitu asam
arachidonat (20: 4 ω-6) dari asam linoleat dan eikosapentaenoat
(20:5 ω-3) dan dokosaheksaenoat atau DHA (22:6 ω-3) berasal dari
asam linolenat. Asam lemak omega-3 mempunyai peranan penting
dalam ilmu gizi, seperti alpha-asam linolenat (C 18:3 ω-3) serta
turunannya, yaitu asam eikosapentaenoat (C 20:5 ω-3)/EPA dan asam
dokosaheksaenoat (C 22:6 ω-3)/DHA. Plankton laut banyak
mengandung asam lemak ω-3. Ikan dapat merubah asam linolenat
menjadi EPA dan DHA; ω-3 dpt mengimbangi kerja asam
arachidonat yang dapat menyebabkan trombosis dan artritis bila
produksi metabolitnya bertumpuk; membersihkan plasma dari
lipoprotein khilomikron dan very low density lipoprotein (VLDL)
menurunkan produksi trigliserida dan apolipoprotein β di dalam hati
yang merupakan bagian utama lipid dan protein dalam VLDL;
dihubungkan dengan pencegahan penyakit jantung koroner dan
arthritis.
2.4 Lemak Netral
Lemak netral adalah lemak yang padat pada suhu kamar dan
berubah menjadi minyak pada saat mencair. Sebagian besar lemak di
alam dalam bentuk trigliserida. Trigliserida adalah ester gliserol,
yaitu alkohol trihidrat dan asam lemak dan lebih tepat disebut
'triasilgliserol'; bila ke tiga asam didalam triglisrida adalah asam
lemak yang sama maka disebut trigliserida sederhana.
70 fisiologi hewan

2.5 Sifat Fisik Lemak dan Reaksi Trigliserida
Trigliserida mempunyai berat jenis lebih rendah daripada air.
Sifat trigliserida ditentukan oleh proporsi dan struktur kimia asam
lemak yang membentuknya. Titik cair dan tingkat kepadatannya
tergantung pada panjang rantai dan tingkat kejenuhannya. Semakin
banyak rantai pendek dan ikatan tidak jenuh semakin lunak lemak
tersebut. Sebaliknya, semakin banyak asam lemak jenuh rantai
panjang, seperti asam palmitat (C16:0) dan asam stearat (C18:0)
maka lemak hewan akan semakin padat. Safonifikasi adalah proses
hidrolisis dengan menggunakan bahan alkali, seperti sabun (foam),
dapat mengganggu jantung. Hidrogenasi adalah proses hidrolisis
lemak tak jenuh menjadi asam lemak jenuh dan proses ini biasanya
digunakan dalam kegiatan komersial, misalnya perubahan minyak
menjadi margarin dan shortening (padat).
Gambar 31. Struktur molekul trigliserida (ester gliserol)
Gambar 32. Proses hidrogenasi asam lemak tidak jenuh
71metabolisme lipid

Proses ketengikan terjadi apabila molekul oksigen terikat pada
ikatan rangkap, sehingga membentuk peroksida aktif, seperti
butylated hydroxy anisole (BHA) dan butylated hydroxy toluene
(BHT) memperpanjang masa simpan lemak.
2.6 Fungsi Lipid
Setiap satu gram lemak menghasilkan energi 9 kilokalori,
dalam bentuk cadangan energi 50% di subkutan; 45% sekeliling
rongga perut, dan 5% intramuskuler. Sumber asam lemak esensial,
seperti asam lemak linoleat dan linolenat Alat angkut vitamin yang
larut dalam lemak: vitamin A, D, E, K. Lipid sebagai cadangan energi
yang digunakan setelah karbohidrat dan sebelum protein Lipid
memiliki fungsi:
1. Memberi rasa kenyang dan kelezatan, dengan memperlambat
sekresi asam lambung dan memperlambat pengosongan
lambung
2. Sebagai pelumas: pelumas dan membantu pengeluaran sisa
pencernaan
3. Memelihara suhu tubuh: mengisolasi tubuh dan mencegah
kehilangan panas
4. Pelindung organ: menyelubungi organ-organ tubuh, seperti
jantung, hati dan ginjal
2.7 Lemak Majemuk dan Lipid Turunan
Fosfolipid terdapat di setiap sel hidup dan fosfolipid dibentuk
dalam sel-sel hati. Fosfolipid merupakan trigliserida dengan posisi
karbon ketiga ditempati oleh gugus fosfat dan gugus basa yang
72 fisiologi hewan

mengandung nitrogen. Gugus basa menentukan nama fosfolipid,
contoh fosfatidilkolin (lesitin); fosfatidilserin mempunyai gugus
serin sebagai gugus basa, fosfatidil etinolamin, fosfatidil inositol dan
lain-lain.
Fungsi fospolid adalah membentuk membran sel dan sebagai
alat angkut protein dalam darah. Fosfolipid mempunyai sifat khusus,
yaitu polar, nonpolar dan amfilitik. Gugus fosfat bermuatan negatif (-
), basa bermuatan (+) dan polar bersifat menarik air (hidrofilik);
gugus asam bersifat tidak bermuatan, nonpolar tidak dapat menarik
air (hidropobik), tetapi menarik lemak. Di membran sel, fosfolipid
membentuk dua lapisan, yaitu rantai asam lemak menghadap ke
dalam sel dan gugus polar pada permukaan luar. Sifat amfilitik punya
peran biologis dalam membran sel, sehingga sel mempunyai daya
tarik yang sama terhadap zat yang larut dalam air dan zat yang larut
dalam lemak.
Nomenklatur Umum Istilah Khusus N.Pendek Sumber
Asam lemak jenuh rantai pendek o Butirat Butanoat 4:0 Mentega
o Kaproat Heksanoat 6:0 Mentega
Asam lemak jenuh rantai sedang
o Kaprilat Oktanoat 8:0 Minyak kelapa
o Kaprat Dekanoat 10:0 Kelapa sawit Asam lemak jenuh rantai panjang
o Laurat Dodekanoat 12:0 Minyak kelapa
o Miristat Tetradekanoat 14:0 Mentega, M. kelapa
o Palmitat Heksadekanoat 16:0 Minyak kelapa, Minyak tumbuhan
o Stearat Oktadekanoat 18:0 Lemak hewan, minyak tumbuhan
Asam lemak tidak jenuh tunggal o Oleat Asam 9 -
oktadekanoat 18:1 (n-9)
Lemak, minyak zaitun
Asam lemak tidak jenuh ganda
o Linoleat Asam 9,12 - oktadekanoat
18:2 (ώ-6)
Minyak jagung, kapas
o Arachidonat Asam 5,8,11,14 - eikodekanoat
20:4 (ώ-6)
Minyak kacang tanah
o Linolenat Asam 9,12,15 - oktadekanoat
18:3 (ώ-3)
Minyak kacangkedelai, kecambah
o Eikosapentaenoat Asam 5,8,11,14,17 - eikodekanoat
20:3 (ώ-3)
Minyak ikan
o Dokosaheksanoat (DHA)
Asam 4,7,10,13,16, 19 - heksadekanoat
22:6 (ώ-3)
Air Susu Ibu, minyak ikan
Tabel 3. Klasifikasi asam lemak
73metabolisme lipid

Fosfolipid berperan sebagai sabun yang akan mampu
membentuk emulsi untuk memecah lemak menjadi tetesan halus
yang mengambang dalam air dan akan membentuk lemak lain dalam
keadaan mengambang dalam darah dan cairan tubuh lain. Lesitin
(fosfatidilkolin) adalah merupakan ikatan antara asam fosfat dan basa
yang mengandung N-kolin; berfungsi memudahkan pengangkutan
dan penggunaan asam lemak dengan menggunakan enzim lesitin-
kolesterol asiltransferase sebagai katalisator. Lesitin banyak
digunakan di industri pangan untuk pembuatan margarin, keju dan
permen. Fosfolipid lain adalah sefalin (struktur sama dengan lesitin)
yang dapat membentuk tromboplastin; lipontol (mengandung
senyawa inositol yang mempunyai fungsi mirip vitamin);
spingomielin (amino alkohol komplek) yang menyusun mielin di
jaringan syaraf otak. Glikolipid adalah merupakan trigliserida dengan
posisi atom karbon ketiga diisi oleh gugus CHO dan gugus basa,
sebagai contoh serebrosid dan angliosid. Sterol memiliki struktur
Gambar 33. Struktur molekul phospatidilkolin
74 fisiologi hewan

cincin yang komplek, yaitu steroid dan terdapat di makanan.
Kolesterol banyak dikenal oleh masyarakat, terutama yang
berkaitan dengan penyebab penyakit jantung. Untuk tubuh yang
normal kebutuhan tubuh terhadap kolesterol adalah konstan, apabila
berlebih maka akan menimbulkan dampak yang membahayakan
terhadap fungsi jaringan atau organ tubuh. Di dalam tubuh kolesterol
mempunyai fungsi ganda, yaitu: kolesterol merupakan komponen
utama membran struktural sel dan saraf. Hati sebagai kelenjar
terbesar di dalam tubuh berfungsi dalam meregulasi kadar kolesterol
dalam tubuh dengan melakukan sintesis dan menyimpan kolesterol
sebagai cadangan energi. Selain fungsi pertama, kolesterol
merupakan bahan untuk pembentukan steroid yang dalam proses
pembentukan asam empedu, asam folat, hormon-hormon adrenal
korteks, estrogen, androgen dan progesteron. Dalam jumlah yang
berlebihan, kolesterol dalam tubuh akan berbahaya, terutama kalau
berada dalam plasma darah, karena dapat menimbulkan penyempitan
pembuluh (aterosklerosis) sehingga menimbulkan penyakit jantung
(Penyakit Jantung Koroner) dan rusaknya pembuluh darah otak
serebrovaskular. Kolesterol dalam tubuh merupakan hasil sintesis di
hati dengan menggunakan bahan baku, berupa CHO, N dan lemak.
Gambar 34. Struktur molekul sterol
75metabolisme lipid

2.8 Aktivitas Vitamin D
Kolesterol dan ergosterol (kolesterol tanaman) merupakan
prekursor vitamin D. Di mukosa usus halus kolesterol akan diubah
menjadi 7-dehidrokolesterol, provitamin kolekalsiferol (vitamin D-
3) dan disimpan dilapisan lemak bawah kulit (sub kutan). Lipoprotein
merupakan gabungan molekul lipid dan protein pengikat lipid
tersebut disintesa di hati. Penyusun lipoprotein adalah protein (25-
33.3%); protein pengikat lipid ini berfungsi mengangkut lipid dalam
plasma menuju ke jaringan-jaringan di seluruh tubuh yang
membutuhkan sebagai sumber energi, sebagai komponen sel atau
sebagai prekursor metabolit aktif. Dikenal ada 4 jenis lipoprotein:
kilomikron, very low density lipoprotein (VLDL), low density
lipoprotein (LDL), dan high density lipoprotein (HDL)
2.9 Pencernaan, Absorpsi, Transportasi, dan Metabolisme
Lemak
Trigliserida merupakan sebagian besar lemak makanan dan
sebagian besar akan terabsorbsi (95%) oleh sel-sel epitel usus masuk
ke dalam sistem sirkulasi darah. Sebelum dicerna, lemak atau
trigliserida akan mengalami emulsifikasi. Emulsifikasi terjadi di usus
halus dibantu oleh garam empedu. Hormon kolesistokinin
memberikan sinyal atau isyarat kepada kantung empedu untuk
mensekresikan asam dan garam empedu. Asam dan garam empedu
berfungsi untuk menarik molekul dalam proses emulsifikasi. Lemak
yang sudah dipecah akan menjadi bagian-bagian kecil didalam cairan
tubuh yang disebut dengan kilomikron. Enzim lipase yang
dikeluarkan oleh sel-sel permukaan usus halus yang menghadap ke
76 fisiologi hewan

lumen dan pankreas akan mencerna lemak yang sudah diemulsifikasi.
Tahap awal penggunaan lemak sebagai sumber energi adalah
hidrolisis triasilgliserol oleh lipase yang akan menghasilkan gliserol
dan asam lemak. Aktivitas lipase sel adiposa diatur oleh beberapa
hormon. Epinefrin, norepinefrin, glukagon dan hormon
adrenokortikotropik mengaktifkan adenilat siklase di dalam sel
adiposa dengan cara memicu reseptor-reseptor. Peningkatan kadar
AMP siklik merangsang protein kinase A, yang akan mengaktifkan
lipase dengan cara fosforilasi. Jadi epinefrin, norepinefrin, glukagon
dan hormon adrenokortikotropik bersifat menginduksi lipolisis. AMP
siklik adalah messenger pada pengaktifan lipolisis di jaringan adiposa
seperti juga pada pengaktifan pemecahan glikogen. Insulin
menghambat proses lipolisis.
Gliserol yang terbentuk pada lipolisis mengalami fosforilasi
dan dioksidasi menjadi dihidroksiaseton fosfat, yang selanjutnya
mengalami isomerisasi menjadi gliseraldehida 3-fosfat. Zat antara ini
terdapat baik pada jalur glikolisis dan glukoneogenesis. Dengan
demikian, gliserol dapat diubah menjadi piruvat atau glukosa di hati,
tempat enzim-enzim yang diperlukan. Proses kebalikannya dapat
terjadi melalui reduksi dihidroksiasetonfosfat menjadi gliserol 3-
fosfat. Hidrolisis oleh fosfatase akan menghasilkan gliserol. Jadi,
gliserol dan zat-zat antara glikolisis dapat saling mudah mengalami
interkonversi.
77metabolisme lipid

Absorpsi lipid berlangsung di yeyenum usus halus bagian
membran mukosa dengan cara difusi pasif. Perbedaan konsentrasi
terjadi dengan cara, protein pengikat asam lemak mengikat asam
lemak yang masuk sel dan esterifikasi kembali asam lemak menjadi
monogliserida; produk utama yang melintasi membran mukosa usus
halus. Sebelum diabsorpsi, kolesterol akan mengalami esterifikasi
kembali yang dikatalis oleh asetil- Coenzim A dan kolesterol
asetiltransferase.
Kilomikron adalah lipoprotein yang paling besar, mengangkut
lipid dari saluran cerna ke seluruh tubuh. Kilomikron mengemulsi
lemak sebelum masuk ke dalam aliran darah. Fungsi kilomikron ini
hampir sama dengan kegiatan lesitin dan asam lemak dalam usus
halus untuk mengemulsi lemak makanan selama pencernaan. Dalam
proses emulsifikasi yang mengelilingi tetesan lemak adalah air.
Adapun kilomikron yang mengalami emulsi adalah kilomikron
lemak yang dikelilingi oleh protein, kolesterol dan fosfolipid. Very
Low Density Lipoprotein (VLDL) dibentuk di dalam sel-sel hati,
mempunyai densitas yang sangat rendah, dan umumnya merupakan
trigliserida. Ketika VLDL meninggalkan hati, lipoprotein-lipase
Hasil Pencernaan Lipid Absorbsi
Gliserol
Diserap langsung ke dalam darah Asam lemak rantai pendek
Asam lemak rantai menengah
Asam lemak rantai panjang Diubah menjadi trigliserida di dalam sel –
sel usus halus Monotrigliserida
Trigliserida Membentuk kilomikron, masuk ke dalam
pembuluh limfe, kemudian ke dalam aliran
darah
Kolesterol
Phospolipid
Tabel 4. Absorbsi lemak ke dalam aliran darah
78 fisiologi hewan

memecah trigliserida yang ada di VLDL, mengikat kolesterol yang
ada di lipoprotein lain di aliran darah, dengan cara sisi/muatan negatif
nya (-) berikatan dengan trigliserida, sisi/muatan positifnya berikatan
dengan VLDL. Low Density Liporpotein (LDL) adalah lipoprotein
berdensitas rendah. LDL terdiri dari kolesterol yang bersirkulasi
dalam tubuh dan dibawa ke sel-sel otot, lemak, dan sel-sel lain.
Kolesterol yang banyak terdapat dalam bentuk LDL. LDL dalam
jumlah yang berlebihan akan menumpuk dan menimbulkan plak
(plague), bercampur dengan protein dan ditutupi sel-sel otot dan
kalsium sehingga menimbulkan aterosklerosis. HDL di produksi oleh
sel-sel hati dan usus halus, dan masuk ke aliran darah. HDL
mengambil kolesterol dan fosfolipid yang terdapat dalam darah
kemudian menyerahkannya ke lipoprotein lain untuk dibawa ke hati
untuk diedarkan kembali atau dikeluarkan dari tubuh.
Tubuh mengatur kadar lipoprotein melalui beberapa cara:
o Mengurangi pembentukan lipoprotein dan mengurangi jumlah
lipoprotein yang masuk ke dalam darah
o Meningkatkan/menurunkan kecepatan pembuangan
lipoprotein dari dalam darah
2.10 Jalur Pengangkutan Lemak dalam Darah
Lemak dalam darah diangkut dengan dua cara, yaitu melalui
jalur eksogen dan jalur endogen.
Jalur eksogen
Trigliserida dan kolesterol yang berasal dari makanan dalam
usus dikemas dalam bentuk partikel besar lipoprotein, yang disebut
kilomikron. Kilomikron ini akan membawanya ke dalam aliran
79metabolisme lipid
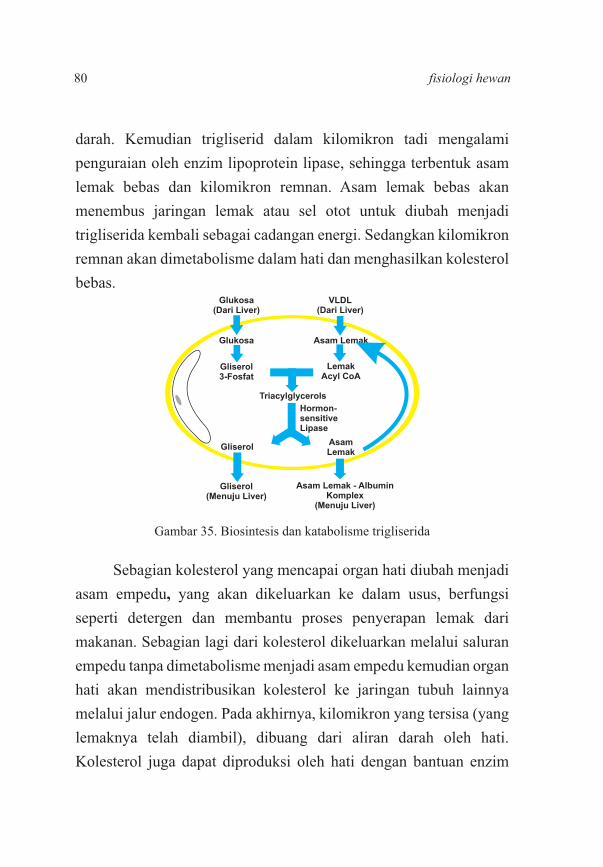
darah. Kemudian trigliserid dalam kilomikron tadi mengalami
penguraian oleh enzim lipoprotein lipase, sehingga terbentuk asam
lemak bebas dan kilomikron remnan. Asam lemak bebas akan
menembus jaringan lemak atau sel otot untuk diubah menjadi
trigliserida kembali sebagai cadangan energi. Sedangkan kilomikron
remnan akan dimetabolisme dalam hati dan menghasilkan kolesterol
bebas.
Sebagian kolesterol yang mencapai organ hati diubah menjadi
asam empedu, yang akan dikeluarkan ke dalam usus, berfungsi
seperti detergen dan membantu proses penyerapan lemak dari
makanan. Sebagian lagi dari kolesterol dikeluarkan melalui saluran
empedu tanpa dimetabolisme menjadi asam empedu kemudian organ
hati akan mendistribusikan kolesterol ke jaringan tubuh lainnya
melalui jalur endogen. Pada akhirnya, kilomikron yang tersisa (yang
lemaknya telah diambil), dibuang dari aliran darah oleh hati.
Kolesterol juga dapat diproduksi oleh hati dengan bantuan enzim
Gambar 35. Biosintesis dan katabolisme trigliserida
80 fisiologi hewan

yang disebut HMG Koenzim-A reduktase, kemudian dikirimkan ke
dalam aliran darah.
Jalur endogen
Pembentukan trigliserida dalam hati akan meningkat apabila
makanan sehari-hari mengandung karbohidrat yang berlebihan. Hati
mengubah karbohidrat menjadi asam lemak, kemudian membentuk
trigliserida, trigliserida ini dibawa melalui aliran darah dalam bentuk
Very Low Density Lipoprotein (VLDL). VLDL kemudian akan
dimetabolisme oleh enzim lipoprotein lipase menjadi IDL
(Intermediate Density Lipoprotein). Kemudian IDL melalui
serangkaian proses akan berubah menjadi LDL (Low Density
Lipoprotein) yang kaya akan kolesterol. Kira-kira ¾ dari kolesterol
total dalam plasma normal manusia mengandung partikel LDL.
Gambar 36. Metabolisme lipid
81metabolisme lipid
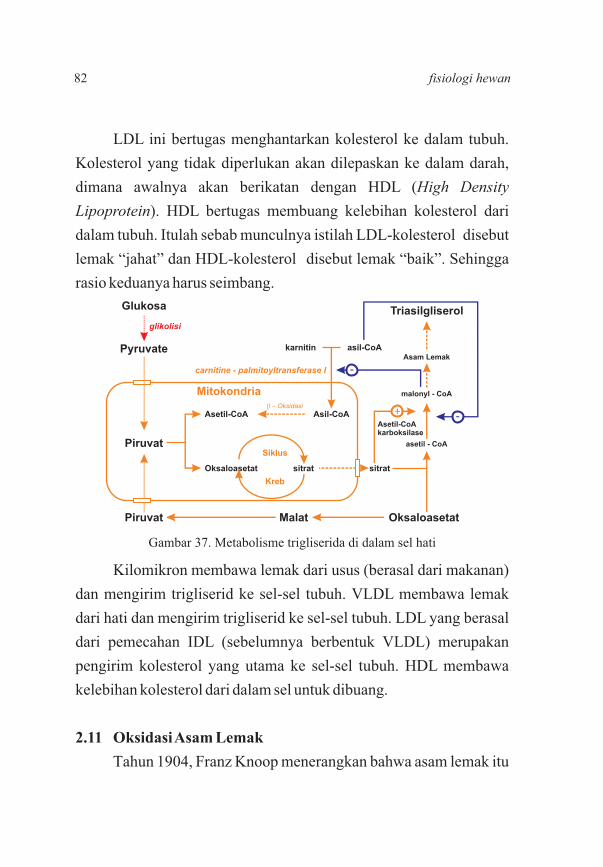
LDL ini bertugas menghantarkan kolesterol ke dalam tubuh.
Kolesterol yang tidak diperlukan akan dilepaskan ke dalam darah,
dimana awalnya akan berikatan dengan HDL (High Density
Lipoprotein). HDL bertugas membuang kelebihan kolesterol dari
dalam tubuh. Itulah sebab munculnya istilah LDL-kolesterol disebut
lemak “jahat” dan HDL-kolesterol disebut lemak “baik”. Sehingga
rasio keduanya harus seimbang.
Kilomikron membawa lemak dari usus (berasal dari makanan)
dan mengirim trigliserid ke sel-sel tubuh. VLDL membawa lemak
dari hati dan mengirim trigliserid ke sel-sel tubuh. LDL yang berasal
dari pemecahan IDL (sebelumnya berbentuk VLDL) merupakan
pengirim kolesterol yang utama ke sel-sel tubuh. HDL membawa
kelebihan kolesterol dari dalam sel untuk dibuang.
2.11 Oksidasi Asam Lemak
Tahun 1904, Franz Knoop menerangkan bahwa asam lemak itu
Gambar 37. Metabolisme trigliserida di dalam sel hati
82 fisiologi hewan

dipecah melalui oksidasi pada karbon-β. Kemudian pada tahun 1949
Eugene Kennedy dan Lehninger menerangkan bahwa terjadinya
oksidasi asam lemak di mitokondria. Di mana asam lemak sebelum
memasuki mitokondria mengalami aktivasi. Adenosin trifosfat (ATP)
memacu pembentukan ikatan tioester antara gugus karboksil asam
lemak dengan gugus sulfhidril pada CoA. Reaksi pengaktifan ini
berlangsung di luar mitokondria dan dikatalisis oleh enzim asil CoA
sintetase (tiokinase asam lemak)
Paul Berg membuktikan bahwa aktivasi asam lemak terjadi
dalam dua tahap. Pertama, asam lemak bereaksi dengan ATP
membentuk asil adenilat. Dalam bentuk anhidra campuran ini, gugus
karboksilat asam lemak diikatkan dengan gugus fosforil AMP. Dua
gugus fosforil lainnya dari ATP dibebaskan sebagai pirofosfat. Gugus
sulfhidril dari CoA kemudian bereaksi dengan asila adenilat yang
berikatan kuat dengan enzim membentuk asil CoA dan AMP.
R – C + ATP R – C – AMP + Ppi
Asam lemak Asil adenilat
R – C – AMP + H- S – CoA R – C – S – CoA + AMP Asil CoA
2.12 Transport Asam Lemak ke Dalam Matriks Mitokondria
Asam lemak diaktifkan di luar membran mitokondria, proses
oksidasi terjadi di dalam matriks mitokondria. Molekul asil CoA
rantai panjang tidak dapat melintasi membran mitokondria, sehingga
diperlukan suatu mekanisme transport khusus. Asam lemak rantai
panjang aktif melintasi membran dalam mitokondria dengan cara
mengkonjugasinya dengan karnitin, suatu senyawa yang terbentuk
dari lisin. Gugus asil dipindahkan dari atom sulfur pada CoA ke gugus
83metabolisme lipid

hidroksil pada karnitin dan membentuk asil karnitin. Reaksi ini
dikatalisis oleh karnitin transferase I, yang terikat pada membran di
luar mitokondria.
R – C – S – CoA + H C – N – CH – C – CH – C HS – CoA + H C – N – CH – C – CH – C3 2 2 3 2 2
Asil CoA Karnitin Asil Karnitin
Selanjutnya, asil karnitin melintasi membran dalam
mitokondria oleh suatu translokase. Gugus asil dipindahkan lagi ke
CoA pada sisi matriks dari membran yang dikatalisis oleh karnitin asil
transferase II. Akhirnya karnitin dikembalikan ke sisi sitosol oleh
translokase menggantikan masuknya asil karnitin yang masuk.
Molekul asil CoA dari sedang dan rantai pendek dapat menembus
mitokondria tanpa adanya karnitin. Kelainan pada transferase atau
translokase atau defisiensi karnitin dapat menyebabkan gangguan
oksidasi asam lemak rantai panjang, Kelainan tersebut diatas
ditemukan pada kembar identik yang menderita kejang otot disertai
rasa nyeri yang dialami sejak masa kanak-kanak. Rasa nyeri
diperberat oleh puasa, latihan fisik, atau diet tinggi lemak; oksidasi
asam lemak adalah proses penghasil energi utama pada ketiga
keadaan tersebut. Enzim glikolisis dan glikogenolisis dalam keadaan
normal.
84 fisiologi hewan
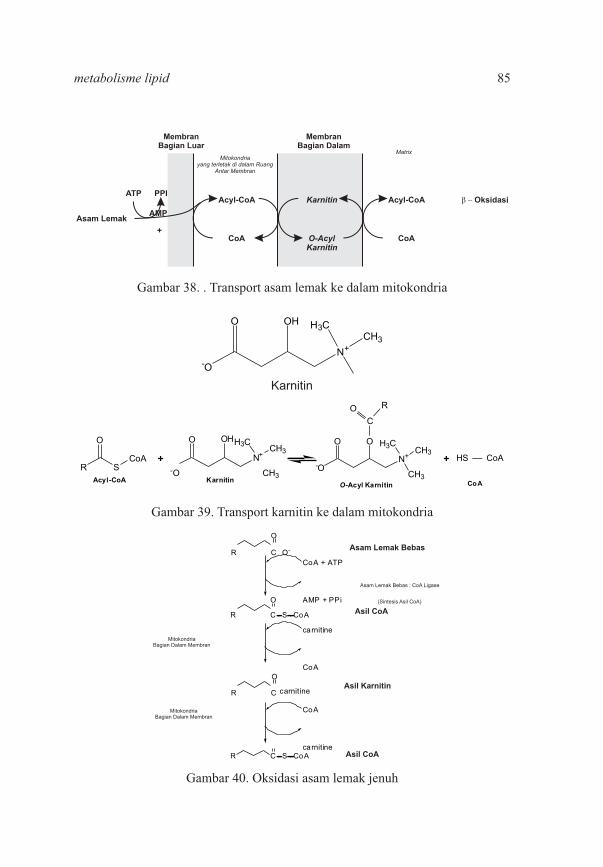
Gambar 38. . Transport asam lemak ke dalam mitokondria
Gambar 39. Transport karnitin ke dalam mitokondria
Gambar 40. Oksidasi asam lemak jenuh
85metabolisme lipid
MembranBagian Luar
MembranBagian Dalam
Asam Lemak Bebas
Asam Lemak Bebas : CoA Ligase
(Sintesis Asil CoA)
Asil CoA
Asil Karnitin
Asil CoA
MitokondriaBagian Dalam Membran
MitokondriaBagian Dalam Membran
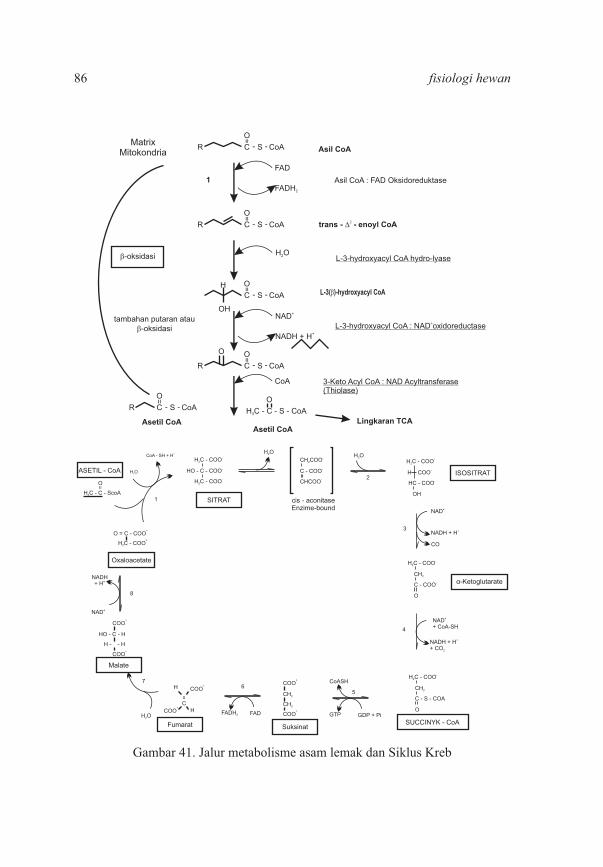
Gambar 41. Jalur metabolisme asam lemak dan Siklus Kreb
86 fisiologi hewan
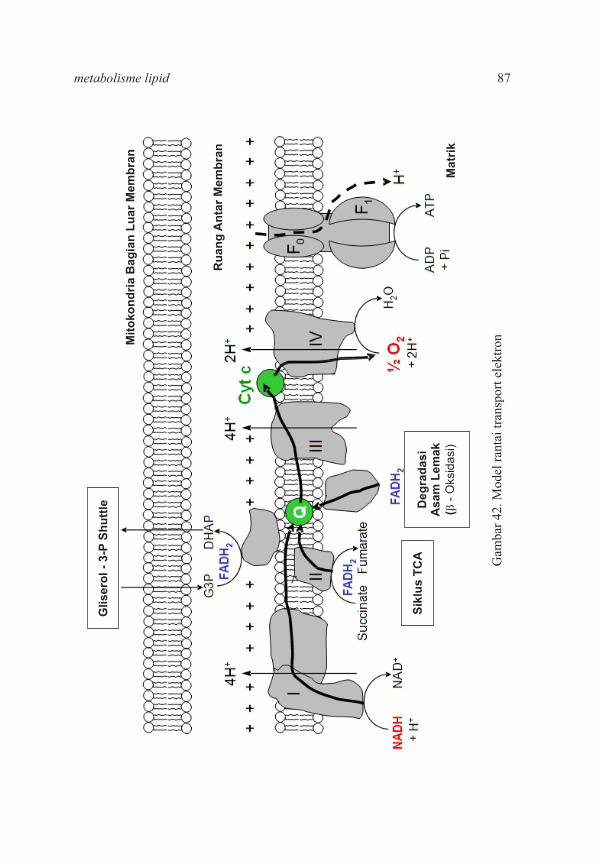
Gam
bar
42. M
odel
ran
tai
tran
spor
t el
ektr
on
87metabolisme lipid

2.13 Oksidasi Asam Lemak Jenuh: Asetil Coa, NADH dan
FADH2
Asil CoA jenuh dipecah melalui urutan empat reaksi yang
berulang, yaitu oksidasi oleh flavin adenin dinukleotida (FAD),
hidrasi oleh NAD dan tiolisis oleh CoA. Rantai asil diperpendek
dengan dua atom karbon sebagai hasil dari keempat reaksi tadi dan
terjadi pembentukan FADH , NADH dan asetil CoA. Reaksi pertama 2
pada tiap daur pemecahan adalah oksidasi asil CoA oleh asil CoA
dehidrogenase yang menghasilkan satu enoil CoA dengan ikatan
rangkap trans antara C-2 dan C-3.
Asil CoA + E – FAD → trans - Δ² - Enoil CoA + E – FADH2
Langkah selanjutnya adalah hidrasi ikatan ganda antara C-2 dan
C-3 oleh enoil CoA hidratase.
Trans - Δ² - Enoil CoA + H O ↔ L-3-hydroksiasil CoA. 2
Hidrasi enoil CoA membuka jalan bagi reaksi oksidasi kedua,
yang mengubah gugus hidroksil pada C-3 menjadi gugus keto dan
menghasilkan NADH. Oksidasi ini dikatalisis oleh L-3-hidroksiasil
CoA dehidrogenase.
L-3-hidroksiasil CoA + NAD ↔ 3-ketoasil CoA + NADH + H+
Langkah akhir adalah pemecahan 3-ketoasil CoA oleh gugus
tiol dari molekul CoA lain, yang akan menghasilkan asetil CoA dan
suatu asil CoA rantai karbonnya dua atom karbon lebih pendek.
88 fisiologi hewan

Reaksi ini dikatalisis oleh β-ketotiolase.
3- ketoasil CoA + HS-CoA ↔ asetil CoA + asil CoA.
(karbon-karbon n)�� (karbon-karbon n-2)
Asil CoA yang memendek selanjutnya mengalami daur
oksidasi berikutnya, yang diawali dengan reaksi yang dikatalisis oleh
asil CoA dehidrogenase. Rantai asam lemak yang mengandung 12
sampai 18 karbon dioksidasi oleh asil CoA dehidrogenase rantai
panjang. Asil CoA dehidrogenase untuk rantai sedang mengoksidasi
ranta asam lemak yang memiliki 14 sampai 4 karbon, sedangkan asil
CoA dehidrogenase untuk rantai pendek hanya bekerja pada rantai 4
dan 6 karbon. Sebaliknya, β-ketotiolase, hidroksiasil dehidrogenase,
dan enoil CoA hidratase memiliki spesifitas yang luas berkenaan
dengan panjangnya gugus asil.
2.14 Oksidasi Sempurna Asam Palmitat
Kita dapat menghitung energi yang dihasilkan dari oksidasi
suatu asam lemak. Pada tiap daur reaksi, asil CoA diperpendek dua
karbon dan satu FADH NADH dan asetil CoA terbentuk. 2,
C – asil CoA + FAD + NAD + H O + CoA → C – asil CoA + FADH + n 2 n-2 2
NADH + asetil CoA + H
Pemecahan palmitoil CoA (C -asil CoA ) memerlukan tujuh daur 16
reaksi. Pada daur ketujuh, C ketoasil CoA mengalami tiolisis menjadi 4
dua molekul asetil CoA. Dengan demikian stoikiometri oksidasi
89

palmitoil CoA menjadi.
Palmitoil CoA + 7 FAD + 7 NAD +7 CoA + & H O→ 8 asetil CoA + 7 2
FADH + 7 NADH + 7 H 2
Dua setengah ATP akan terbentuk per NADH yang dioksidasi
pada rantai pernafasan, sedangkan 1,5 ATP akan terbentuk untuk tiap
FADH . Jika diingat bahwa oksidasi asetil CoA oleh daur asam sitrat 2
menghasilkan 10 ATP, maka jumlah ATP yang terbentuk pad oksidasi
palmitoil CoA adalah 10,5 dari 7 FADH , 17,5 dari 7 NADH dan 80 2
dari 8 molekul asetil CoA, sehingga jumlah keseluruhannya adalah
108. Dua ikatan fosfat energi tinggi dipakai untuk mengaktifkan
palmitat, saat ATP terpecah menjadi AMP dan 2 P . Jadi oksidasi i
sempurna satu molekul asam palmitat menghasilkan 106 ATP.
2.15 Oksidasi Asam Lemak Tak Jenuh
Oksidasi asam lemak tak jenuh reaksinya sama seperti reaksi
oksidasi asam lemak jenuh. Hanya diperlukan tambahan dua enzim
lagi yaitu isomerase dan reduktase untuk memecah asam-asam lemak
tak jenuh. Oksidasi asam palmitoleat atau asam lemak C yang 16
memiliki ikatan rangkap antara C-9 dan C-10 ini diaktifkan dan
diangkut melintasi membran dalam mitokondria dengan cara yang
sama dengan asam lemak jenuh. Selanjutnya palmitoleil CoA
mengalami tiga kali pemecahan dengan enzim-enzim yang sama
seperti oksidasi asam lemak jenuh. Enoil CoA-sis-Δ³ yang terbentuk
pada ketiga kali jalur oksidasi bukanlah substrat bagi asil CoA
dehidrogenase. Adanya ikatan rangkap antara C-3 dan C-4
90 fisiologi hewan

menghalangi pembentukan ikatan rangkap lainnya antar C-2 dan C-3.
Kendala ini dapat diatasi oleh suatu reaksi yang mengubah posisi dan
konfigurasi dari ikatan rangkap sis-Δ³. Suatu isomerase mengubah
ikatan rangkap ini menjadi ikatan rangkap trans-Δ². Reaksi- reaksi
berikutnya mengikuti reaksi oksidasi asam lemak jenuh saat enoil
CoA-trans-Δ² merupakan substrat yang reguler.
Satu enzim tambahan lagi diperlukan untuk oksidasi asam
lemak tak jenuh jamak. Misalnya asam lemak tak jenuh jamak C18
yaitu linoleat, dengan ikatan rangkap sis-Δ9 dan sis Δ12. Ikatan
rangkap sis-Δ³ yang terbentuk setelah tiga daur oksidasi-β, diubah
menjadi ikatan rangkap trans-Δ² oleh isomerase tersebut di atas,
seperti pada oksidasi palmitoleat. Ikatan rangkap sis-Δ¹²-linoleat
menghadapi masalah baru. Asil CoA yang dihasilkan oleh empat daur
oksidasi β mengandung ikatan rangkap rangkap sis-Δ4. Enzim
dehidrogenase pada spesies ini oleh asil CoA dehidrogenase
menghasilkan zat antara 2,4-dienoil yang bukan substrat bagi enzim
berikutnya pada jalur oksidasi β. Kendala ini dapat diatasi oleh 2,4-
dienoil-CoA reduktase, suatu enzim yang menggunakan NADH
untuk mereduksi zat antara 2,4-dienoil menjadi enoil CoA-sis-Δ³.
Isomerase tersebut di atas kemudian mengubah enoil CoA-sis-Δ³
menjadi bentuk trans, suatu zat antara yang lazim pada oksidasi-
β.Jadi ikatan rangkap yang letaknya pada atom C nomor ganjil
ditangani oleh isomerase dan ikatan rangkap yang terletak pada atom
C nomor genap ditangani oleh reduktase dan isomerase.
2.16 Oksidasi Asam Lemak dengan Nomor Atom Karbon Ganjil
Asam lemak yang memiliki jumlah karbon ganjil merupakan
91metabolisme lipid

spesies jarang. Asam lemak ini dioksidasi dengan cara yang sama
seperti oksidasi asam lemak dengan jumlah atom karbon genap,
kecuali pada daur akhir degradasi akan terbentuk propionil CoA dan
asetil CoA, bukan dua molekul asetil CoA. Unit tiga karbon aktif
pada propionil CoA memasuki daur asam sitrat setelah diubah
menjadi suksinil CoA.
2.17 Proses Ketogenesis
Asetil CoA yang terbentuk pada oksidasi asam lemak akan
memasuki daur asam sitrat hanya jika pemecahan lemak dan
karbohidrat terjadi secara berimbang. Karena masuknya asetil CoA
ke dalam daur asam sitrat tergantung pada tersedianya oksaloasetat
untuk pembentukan sitrat. Tetapi konsentrasi oksaloasetat akan
menurun jika karbohidrat tidak tersedia atau penggunaannya tidak
sebagaimana mestinya. Oksaloasetat dalam keadaan normal dibentuk
dari piruvat. Pada puasa atau diabetes, oksaloasetat dipakai untuk
membentuk glukosa pada jalur glukoneogenesis dan demikian tidak
tersedia untuk kondensasi dengan asetil CoA. Pada keadaan ini asetil
CoA dialihkan ke pembentukan asetoasetat dan D-3-hidroksibutirat.
Asetoasetat, D-3-hidroksibutirat dan aseton disebut dengan zat keton.
Asetoasetat dibentuk dari asetil CoA dalam tiga tahap. Dua molekul
asetil CoA berkondensasi membentuk asetoasetil CoA. Reaksi yang
dikatalisis oleh tiolase ini merupakan kebalikan dari tahap tiolisis
pada oksidasi asam lemak. Selanjutnya astoasetil CoA bereaksi
dengan asetil CoA dan air untuk menghasilkan 3-hidroksi-3-
metilglutaril CoA (HMG-CoA) dan CoA. Kondensasi ini mirip
dengan kondensasi yang dikatalisis oleh sitrat sintase. Keseimbangan
92 fisiologi hewan

yang tidak menguntungkan bagi pembentukan asetoasetil CoA
diimbangi oleh reaksi ini, yang keseimbangannya menguntungkan
karena hidrolisis ikatan tioester, 3-Hidroksi-3-metilglutaril CoA
kemudian terpecah menjadi asetil CoA dan asetoasetat.
Hasil dari keseluruhan reaksi adalah: 2 Asetil CoA + H O 2
+Asetoasetat +2 CoA H , 3-hidroksibutirat terbentuk melalui reduksi
asetoasetat di matriks mitokondria. Rasio hidroksibutirat terhadap +
asetoasetat tergantung pada rasio NADH/NAD di dalam
mitokondria. Karena merupakan asam keto-β, asetoasetat secara
lambat mengalami dekarboksilasi spontan menjadi aseton. Bau
aseton dapat dideteksi dalam udara pernafasan seseorang yang kadar
asetoasetat dalam darahnya tinggi.
93metabolisme lipid
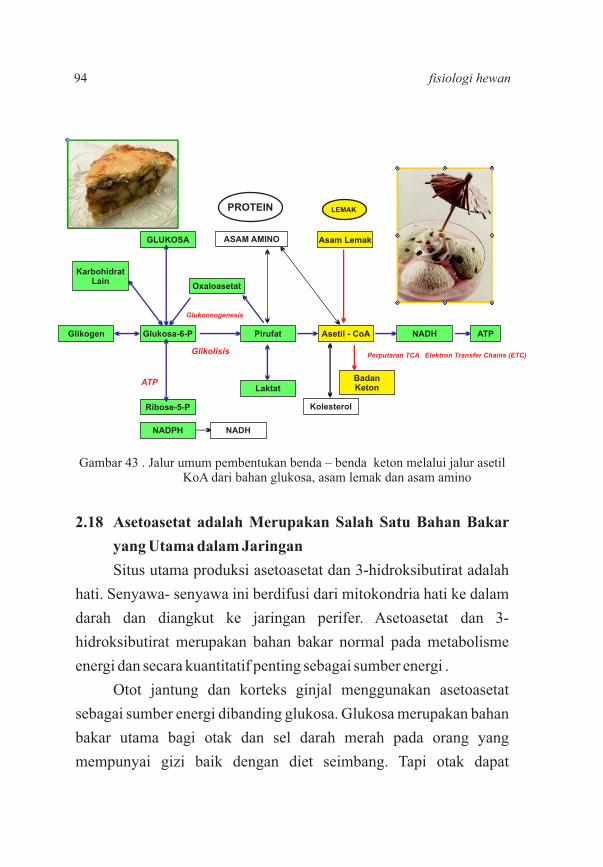
2.18 Asetoasetat adalah Merupakan Salah Satu Bahan Bakar
yang Utama dalam Jaringan
Situs utama produksi asetoasetat dan 3-hidroksibutirat adalah
hati. Senyawa- senyawa ini berdifusi dari mitokondria hati ke dalam
darah dan diangkut ke jaringan perifer. Asetoasetat dan 3-
hidroksibutirat merupakan bahan bakar normal pada metabolisme
energi dan secara kuantitatif penting sebagai sumber energi .
Otot jantung dan korteks ginjal menggunakan asetoasetat
sebagai sumber energi dibanding glukosa. Glukosa merupakan bahan
bakar utama bagi otak dan sel darah merah pada orang yang
mempunyai gizi baik dengan diet seimbang. Tapi otak dapat
Gambar 43 . Jalur umum pembentukan benda – benda keton melalui jalur asetil KoA dari bahan glukosa, asam lemak dan asam amino
94 fisiologi hewan

beradaptasi dan menggunakan asetoasetat dalam keadaan kelaparan
dan diabetes. Pada kelaparan berkepanjangan, 75% bahan bakar yang
diperlukan oleh otak didapat dari asetoasetat. Asetoasetat dapat
diaktifkan melalui pemindahan CoA dari suksinil CoA dalam suatu
reaksi yang dikatalisis oleh suatu CoA transferase spesifik.
Kemudian, asetoasetil CoA dipecah oleh tiolase menjadi dua molekul
asetil CoA, yang selanjutnya memasuki daur asam sitrat. Hati dapat
membekali organ-organ lain dengan asetoasetat karena hati tidak
memiliki CoA transferase spesifik ini.
Asam lemak dilepaskan oleh jaringan adiposa dan diubah
menjadi unit-unit asetil oleh hati, yang kemudian mengeluarkannya
sebagai asetoasetat. Kadar asetoasetat yang tinggi dalam darah
menandakan berlimpahnya unit asetil yang menyebabkan
berkurangnya laju lipolisis di jaringan adiposa.
2.19 Biosintesis Asam Lemak
Biosintesis asam lemak terjadi di beberapa jaringan dalam
tubuh, terutama di jaringan yang mempunyai kapasitas menyimpan
cadangan energi dalam jumlah besar, seperti di hati dan otot. Selain
sebagai cadangan energi, biosintesis asam lemak penting dalam
rangka untuk menyusun elemen structural membrane sel,
pembentukan hormone steroid dan sebagai molekul-molekul yang
mampu memberi sinyal (signaling molecules). Contoh biosintesis
asam lemak adalah pembentukan asam palmitat dari asetil CoA:
8 asetil CoA → Asam palmitat
95metabolisme lipid

Sintesis asam lemak bukan merupakan kebalikan dari jalur
pemecahannya. Sintesis asam lemak lebih merupakan seperangkat
reaksi, yang menunjukkan prinsip bahwa jalur sintesis dan jalur
pemecahan dalam system biologis biasanya berbeda. Beberapa ciri
penting jalur biosintesis asam lemak adalah:
1. Sintesis berlangsung di luar mitokondria, oksidasi terjadi di
dalam matriks mitokondria.
2. Zat antara pada sintesis asam lemak berikatan kovalen dengan
gugus sulfhidril pada protein-pembawa asil (ACP), sedangkan
zat antara pada pemecahan asam lemak berikatan dengan
koenzim A.
3. Enzim-enzim yang terlibat dalam proses sintesis asam lemak
pada organisme yang lebih tinggi tergabung dalam suatu rantai
polipeptida tunggal, yang disebut sintase asam lemak.
Sebaliknya, enzim-enzim pemecahan tampaknya tidak saling
berikatan.
4. Rantai asam lemak yang sedang tumbuh, diperpanjang dengan
cara penambahan berturut-turut unit dua karbon yang berasal
dari asetil CoA. Donor aktif unit dua karbon pada tahap
perpanjangan adalah malonil-ACP. Reaksi perpanjangan
dipacu oleh pelepasan Co . 2
5. Reduktor pada sintesis asam lemak adalah NADPH, sedangkan
oksidator pada pemecahan asam lemak adalah NAD dan FAD.
6. Perpanjangan rantai oleh kompleks sontase asam lemak
terhenti setelah terbentuknya palmitat (C16). Perpanjangan
rantai lebih lanjut dan penyisipan ikatan rangkap oleh sistem
enzim yang lain.
96 fisiologi hewan
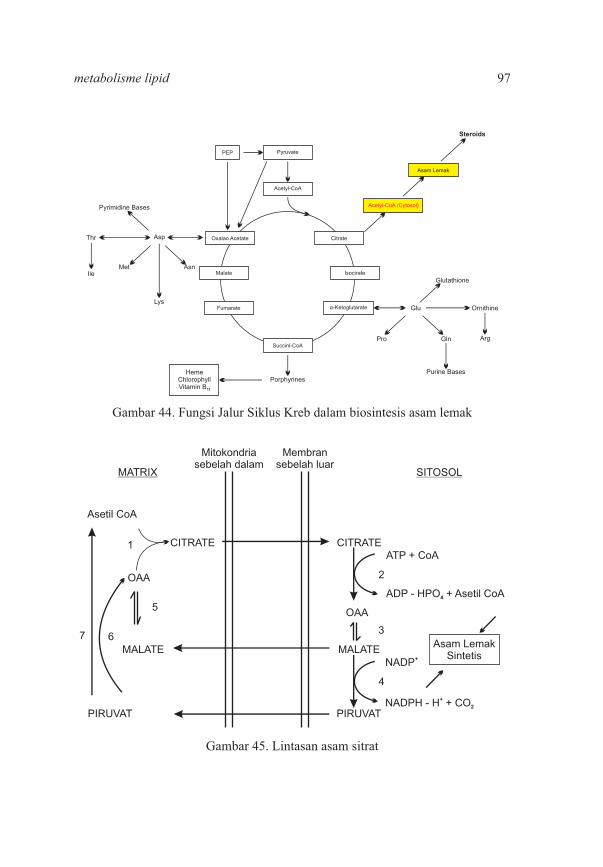
Gambar 44. Fungsi Jalur Siklus Kreb dalam biosintesis asam lemak
Gambar 45. Lintasan asam sitrat
97metabolisme lipid
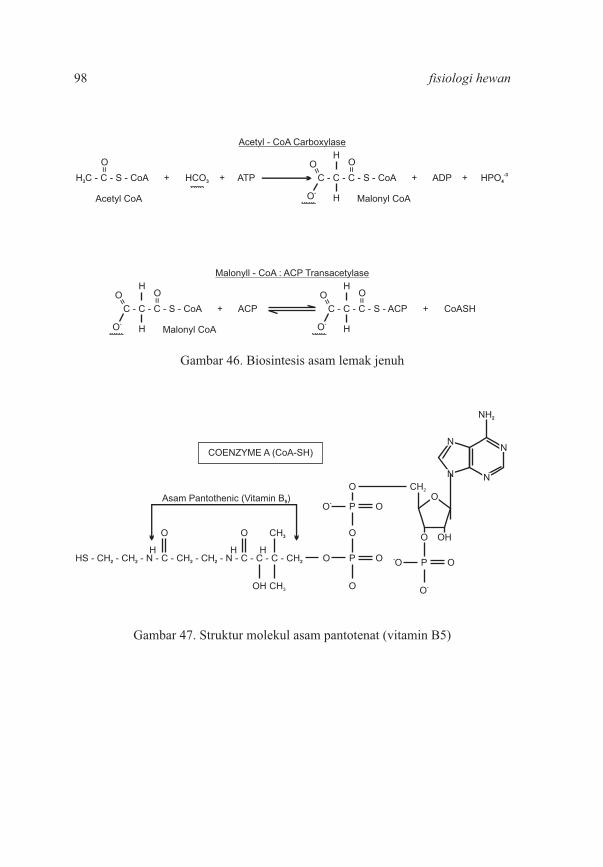
Gambar 46. Biosintesis asam lemak jenuh
Gambar 47. Struktur molekul asam pantotenat (vitamin B5)
98 fisiologi hewan
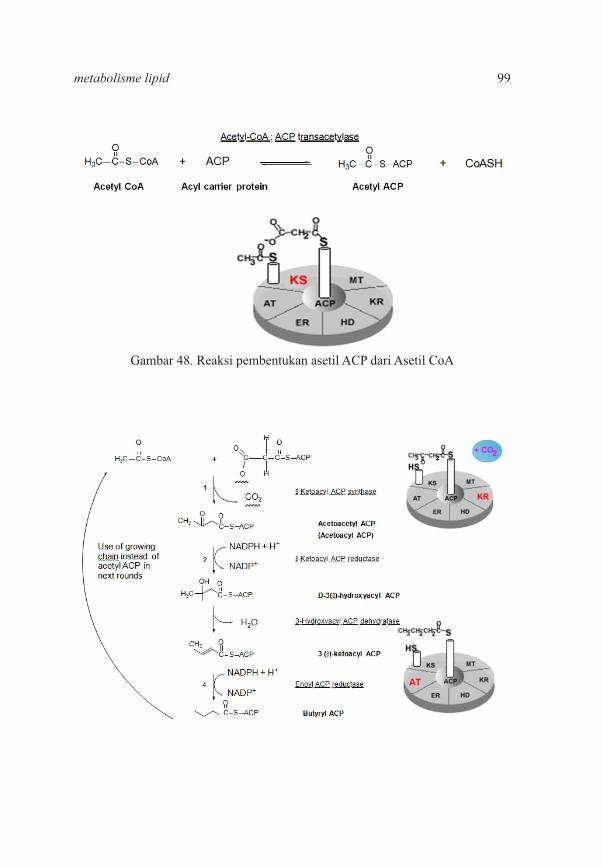
Gambar 48. Reaksi pembentukan asetil ACP dari Asetil CoA
99metabolisme lipid
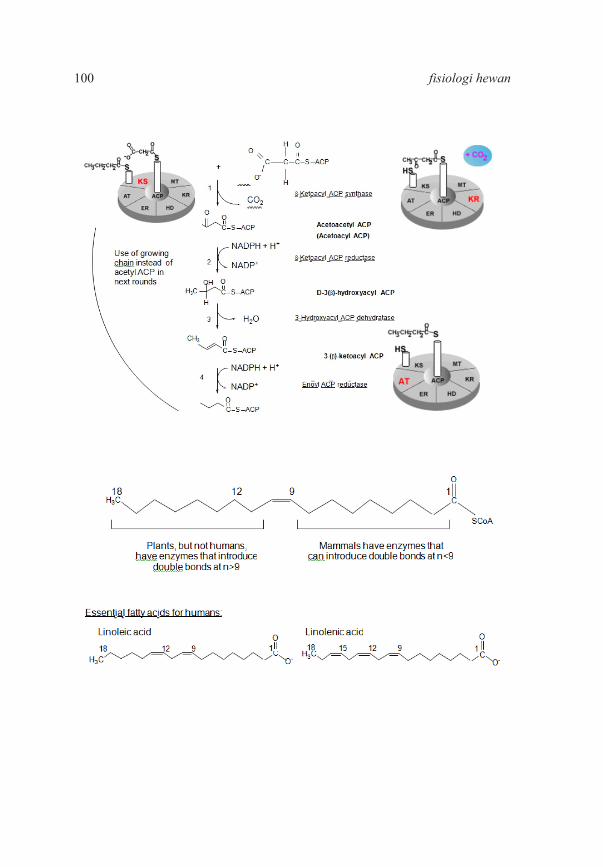
100 fisiologi hewan
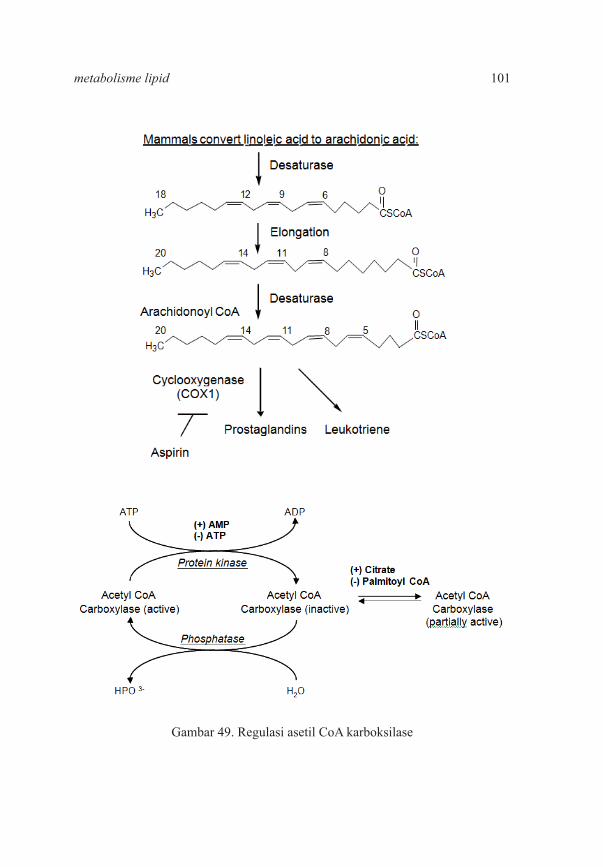
Gambar 49. Regulasi asetil CoA karboksilase
101metabolisme lipid

Gambar 50. Grafik hubungan antara asetil CoA karboksilase dengan kadar asam
Gambar 51 . Sintesis phospolipid dan trigliserida
102 fisiologi hewan

2.20 Metabolisme Lemak pada Hewan Ruminansia
Pada monogastrik, trigliserida dikonversi menjadi
monogliserida lalu menjadi asam lemak bebas dan gliserol,
membentuk misel dan masuk ke pembuluh darah, menjadi
kilomikron (trigliserida) lalu ke limpa, atau menjadi asam lemak
rantai pendek atau menengah langsung ke portal darah. Pada
ruminansia, lesitin dikonversi menjadi lisolesitin, bercampur dengan
partikel digesti dan garam-garam empedu membentuk misel lalu
masuk ke pembuluh darah, membentuk kilomikron (trigliserida,
lesitin dan lipoprotein) selanjutnya masuk ke limpa. Tidak ada asam
lemak rantai pendek atau menengah yang langsung ke portal darah.
Pakan hijauan dan biji-bijian umumnya berbentuk lemak tidak
jenuh. Hidrolisis lipid yang teresterifikasi oleh lipase asal mikroba
akan membebaskan asam-asam lemak bebas, sehingga galaktosa dan
103metabolisme lipid

gliserol akan difermentasi menjadi VFA. Hasil samping dari
pembentukan VFA akan menghasilkan gas methane. Metan
merupakan produk sampingan dalam proses fermentasi
karbohidrat/gula secara anaerob. Metan merupakan energi yang
terbuang. Bakteri metanogen akan menggunakan H yang terbentuk 2
dari konversi asam piruvat menjadi asam asetat untuk membentuk
metan dan juga dari dekomposisi format, atau metanol. Dalam
pembentukan metan oleh mikroorganisme, terlibat pula peran asam
folat dan vitamin B12. Pembentukan metan dapat dikurangi dengan
cara:
§ Menambahkan asam lemak tidak jenuh ke dalam ransum.
§ Menggunakan feed additive seperti choloform, chloral hidrat
dan garam tembaga.
Secara umum mekanisme pembentukan VFA (asam asetat,
propionat dan butirat) melalui asam piruvat terjadi melalui beberapa
cara, yaitu: (1) Oksidasi asam piruvat menjadi asam asetat melalui
Asetil CoA atau asetil fosfat (sebagai tahap antara); (2) fiksasi CO 2
dari fosfoenol piruvat membentuk oksaloasetat dan propionat serta
reduksi akrilat menjadi asam propionat, terutama terjadi pada hewan
yang mengkonsumsi ransum tinggi biji-bijian (laktat tinggi); (3)
kebalikan dari β-oksidasi dengan melibatkan asam asetat untuk
membentuk asam butirat dan melibatkan malonil-CoA seperti pada
sintesis asam lemak rantai panjang.
Volatile Fat Acid yang diabsorbsi dari retikulorumen akan
mengalami perombakan dan oksidasi menghasilkan energi yang
dapat digunakan untuk biosintesis lemak atau glukosa. Setiap jenis
Volatile Fat Acid mempunyai kemampuan menghasilkan energi yang
104 fisiologi hewan

berbeda-beda. Proporsi Volatile Fat Acid yang mengalami oksidasi di
dalam jaringan tubuh hewan ruminansia masing – masing adalah,
asam asetat 50%, asam butirat 67% dan asam propionat 25%.
Metabolisme asam propionat dan butirat terjadi di hati. Kurang lebih
60% asetat dimetabolisme di jaringan perifer (otot dan adiposa) dan
hanya 20% di metabolisasi di hati. Pada hewan yang sedang laktasi,
asam asetat digunakan untuk sintesis lemak air susu diambing.
Penyerapan VFA tergantung pada perbedaan antara
konsentrasinya di dalam cairan rumen dan di dalam sel-sel epitel atau
darah. Proporsi penyerapan VFA adalah 75% diserap langsung dari
retikulum masuk ke dalam darah, 20% diserap dari abomasum-
omasum dan 5% lolos masuk ke usus halus untuk diserap masuk ke
darah. Laju penyerapan VFA dari rumen meningkat sejalan dengan
penurunan pH cairan rumen. Asam butirat dari rumen akan melalui
dinding rumen untuk masuk ke dalam darah untuk dikonversi
menjadi β-hidroksibutirat, sedangkan asam propionat akan
dikonversi menjadi asam laktat. Hal ini terjadi karena peran enzim-
enzim tertentu yang ada di dalam sel-sel epitel. β-hidroksibutirat
dapat digunakan sebagai sumber energi bagi sejumlah jaringan,
misal: otot kerangka atau hati.
Untuk menghasilkan energi dalam bentuk ATP, volatile fat
acids selanjutnya akan mengalami proses oksidasi melalui beberapa
macam reaksi, antara lain: oksidasi asam propionat setelah konversi
menjadi glukosa melalui jalur glukonegenesis yang menghasilkan 17
mol ATP/mol asam propionat, Oksidasi langsung asam propionat
menghasilkan 18 mol ATP/mol asam propionat, konversi asam butirat
menjadi β- hidroksibutirat yang menghasilkan 2 mol ATP serta
105metabolisme lipid

konversi asam asetat menajdi asetil CoA yang menghasilkan 10 mol
ATP/mol asetat.
Sebagian besar asam lemak esensial akan rusak oleh karena
proses biohidrogenasi, namun hewan tidak mengalami defisiensi.
Sebagian kecil asam lemak esensial yang lolos dari proses di dalam
rumen tersebut, sudah dapat memenuhi kebutuhan hewan. Mikroba
rumen juga mampu mensintesis beberapa asam lemak rantai panjang
dari propionat dan asam lemak rantai cabang dari kerangka karbon
asam-asam amino valin, leusin dan isoleusin. Asam-asam lemak
tersebut akan bergabung dengan lemak susu dan lemak tubuh
ruminansia.
Kebanyakan lipid pada hewan ruminansia masuk ke duodenum
sebagai asam lemak bebas dengan kandungan asam lemak jenuh yang
tinggi. Monogliserida yang dominan pada monogastrik, pada
ruminansia akan mengalami hidrolisis di dalam rumen, sehingga
sangat sedikit terdapat pada rumen. Hewan ruminansia yang masih
muda mempunyai kemampuan untuk mengkonversi glukosa menjadi
asam – asam lemak, namun ketika rumen berfungsi, kemampuan itu
hilang dan asetat menjadi sumber karbon utama yang digunakan
untuk mensintesis asam-asam lemak. Asetat akan didifusi masuk ke
dalam darah dari rumen dan dikonversi di jaringan menjadi asetil –
CoA, dengan energi berasal dari hidrolisis ATP menjadi AMP. Jalur
ini terjadi di tempat penyimpanan lemak tubuh yaitu jaringan adiposa
(di bawah kulit, jantung dan ginjal). Konversi asetil – CoA menjadi
asam-asam lemak rantai panjang sama terjadinya antara ruminan dan
monogastrik.
106 fisiologi hewan
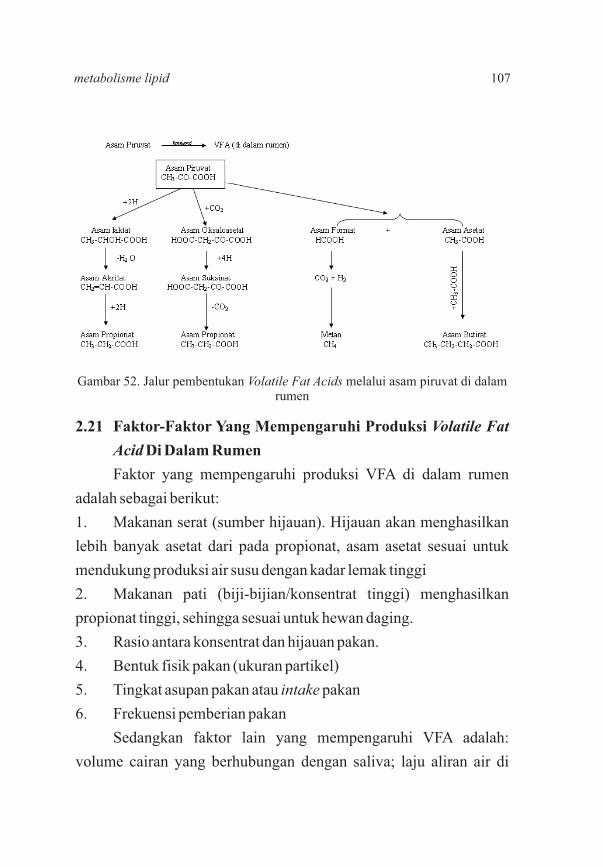
2.21 Faktor-Faktor Yang Mempengaruhi Produksi Volatile Fat
Acid Di Dalam Rumen
Faktor yang mempengaruhi produksi VFA di dalam rumen
adalah sebagai berikut:
1. Makanan serat (sumber hijauan). Hijauan akan menghasilkan
lebih banyak asetat dari pada propionat, asam asetat sesuai untuk
mendukung produksi air susu dengan kadar lemak tinggi
2. Makanan pati (biji-bijian/konsentrat tinggi) menghasilkan
propionat tinggi, sehingga sesuai untuk hewan daging.
3. Rasio antara konsentrat dan hijauan pakan.
4. Bentuk fisik pakan (ukuran partikel)
5. Tingkat asupan pakan atau intake pakan
6. Frekuensi pemberian pakan
Sedangkan faktor lain yang mempengaruhi VFA adalah:
volume cairan yang berhubungan dengan saliva; laju aliran air di
Gambar 52. Jalur pembentukan Volatile Fat Acids melalui asam piruvat di dalam rumen
107metabolisme lipid

dalam darah. Konsentrasi VFA rumen diatur oleh keseimbangan
antara produksi dan penyerapan. Konsentrasi meningkat setelah
makan, sehingga akibatnya pH menurun. Jika glikogen meningkat,
asam propionat, asam butirat atau asam format meningkat, pH akan
menurun 4.5-5 dan pH rendah akan menghambat pertumbuhan
bakteri selulolitik, sehingga akan menghambat pencernaan hijauan.
Derajad keasaman rumen normal yang sesuai untuk pertumbuhan
mikroba adalah 6-7. Derajad keasaman ini ditentukan oleh kapasitas
produksi saliva dan penyerapan VFA.
2.22 Kelainan Lemak Dan Diagnosis Kelainan Lemak
Dilakukan pemeriksaan darah untuk mengukur kadar
kolesterol total. Untuk mengukur kadar kolesterol LDL, HDL dan
trigliserida, sebaiknya penderita berpuasa dulu minimal selama 12
jam.
Hiperlipidemia
Yang dimakud dengan hiperlipidemia adalah suatu keadaan yang
ditandai oleh peningkatan kadar lipid/lemak darah. Berdasarkan
jenisnya, hiperlipidemia dibagi menjadi 2, yaitu:
Hiperlipidemia primer
Banyak disebabkan oleh karena kelainan genetik. Biasanya kelainan
ini ditemukan pada waktu pemeriksaan laboratorium secara
kebetulan. Pada umumnya tidak ada keluhan, kecuali pada keadaan
yang agak berat tampak adanya xantoma (penumpukan lemak di
bawah jaringan kulit).
108 fisiologi hewan

Hiperlipidemia sekunder
Pada jenis ini, peningkatan kadar lipid darah disebabkan oleh
suatu penyakit tertentu, misalnya: diabetes melitus, gangguan tiroid,
penyakit hepar dan penyakit ginjal. Hiperlipidemia sekunder bersifat
reversibel (berulang). Ada juga obat –obatan yang menyebabkan
gangguan metabolisme lemak, seperti: Beta – blocker, diuretik,
kontrasepsi oral (estrogen, gestagen).
2.23 Klasifikasi Klinis Hiperlipidemia
Klasifikasi klinis hiperlipidemia dalam hubungannya dengan
penyakit jantung koroner:
· Hiperkolesterolemia yaitu kadar kolesterol meningkat dalam
darah .
· Hipertrigliseridemia yaitu kadar trigliserida meningkat dalam
darah.
· Hiperlipidemia campuran yaitu: kadar kolesterol dan
trigliserida meningkat dalam darah.
Penyebab hiperlipidemia, meliputi penyebab primer, yaitu
Tabel 5. Kadar lemak darah normal
109metabolisme lipid

faktor keturunan (genetik) dan penyebab sekunder, seperti usia, jenis
kelamin, riwayat keluarga dengan hiperlipidemia, obesitas atau
kegemukan. Menu makanan yang mengandung asam lemak jenuh
seperti mentega, margarin, whole milk, es krim, keju, daging
berlemak. Penyebab lainnya adalah kurang melakukan olah raga,
penggunaan alcohol, merokok, diabetes yang tidak terkontrol dengan
baik, gagal ginjal, dan kelenjar tiroid yang kurang aktif. Obat-obatan
tertentu dapat mengganggu metabolisme lemak seperti estrogen,
pilKB, kortikosteroid, diuretik tiazid (pada keadaan tertentu)
Sebagian besar kasus peningkatan kadar trigliserida dan
kolesterol total bersifat sementara dan tidak berat, dan terutama
merupakan akibat dari makan lemak. Pembuangan lemak dari darah
pada setiap orang memiliki kecepatan yang berbeda. Seseorang bisa
makan sejumlah besar lemak hewani dan tidak pernah memiliki kadar
kolesterol total lebih dari 200 mg/dl, sedangkan yang lainnya
menjalani diet rendah lemak yang ketat dan tidak pernah memiliki
kadar kolesterol total dibawah 260 mg/dl. Perbedaan ini tampaknya
bersifat genetik dan secara luas berhubungan dengan perbedaan
kecepatan masuk dan keluarnya lipoprotein dari aliran darah.
2.24 Gejala Hiperlipidemia
Biasanya kadar lemak yang tinggi tidak menimbulkan gejala.
Kadang-kadang, jika kadarnya sangat tinggi, endapan lemak akan
membentuk suatu penumpukan lemak yang disebut xantoma di dalam
tendo (urat daging) dan di dalam kulit. Kadar trigliserida yang sangat
tinggi (sampai 800 mg/dl atau lebih) bisa menyebabkan pembesaran
hati dan limpa dan gejala-gejala dari pankreatitis (misalnya nyeri
110 fisiologi hewan

perut yang hebat).
Hiperlipidemia dapat meningkatkan resiko terkena
aterosklerosis, penyakit jantung koroner, pankreatitis (peradangan
pada organ pankreas), diabetes melitus, gangguan tiroid, penyakit
hepar dan penyakit ginjal. Yang paling sering adalah resiko terkena
penyakit jantung. Tidak semua kolesterol meningkatkan resiko
terjadinya penyakit jantung. Kolesterol yang dibawa oleh LDL
(disebut juga kolesterol jahat) menyebabkan meningkatnya resiko;
kolesterol yang dibawa oleh HDL (disebut juga kolesterol baik)
menyebabkan menurunnya resiko dan menguntungkan. Lalu, apakah
kadar trigliserida yang tinggi meningkatkan resiko terjadinya
penyakit jantung atau stroke, masih belum jelas. Kadar trigliserida
darah diatas 250 mg/dl dianggap abnormal, tetapi kadar yang tinggi
ini tidak selalu meningkatkan resiko terjadinya aterosklerosis
maupun penyakit jantung koroner. Kadar trigliserid yang sangat
tinggi (sampai lebih dari 800 mg/dl) bisa menyebabkan pankreatitis
(gangguan pada organ pankreas).
2.25 Patofisiologi Penyakit Jantung Koroner
Tubuh sendiri memproduksi kolesterol sesuai kebutuhan
melalui hati. Bila terlalu banyak mengkonsumsi makanan yang
mengandung kolesterol, maka kadar kolesterol dalam darah bisa
berlebih (disebut hiperkolesterolemia). Kelebihan kadar kolesterol
dalam darah akan disimpan di dalam lapisan dinding pembuluh darah
arteri, yang disebut sebagai plak atau ateroma (sumber utama plak
berasal dari LDL-kolesterol. Sedangkan HDL membawa kembali
kelebihan kolesterol ke dalam hati, sehingga mengurangi
111metabolisme lipid

penumpukan kolesterol di dalam dinding pembuluh darah). Ateroma
berisi bahan lembut seperti keju, mengandung sejumlah bahan lemak,
terutama kolesterol, sel-sel otot polos dan sel-sel jaringan ikat.
Apabila makin lama plak yang terbentuk makin banyak, akan
terjadi suatu penebalan pada dinding pembuluh darah arteri, sehingga
terjadi penyempitan pembuluh darah arteri. Kejadian ini disebut
sebagai aterosklerosis (terdapatnya aterom pada dinding arteri, berisi
kolesterol dan zat lemak lainnya). Hal ini menyebabkan terjadinya
arteriosklerosis (penebalan pada dinding arteri dan hilangnya
kelenturan dinding arteri). Bila ateroma yang terbentuk semakin
tebal, dapat merobek lapisan dinding arteri dan darah akan menjadi
membeku (trombus) yang dapat menyumbat aliran darah dalam arteri
tersebut.
Tersumbatnya aliran darah di arteri dapat menyebabkan
berkurangnya aliran darah serta suplai zat – zat penting seperti
oksigen ke daerah atau organ tertentu seperti jantung. Bila kondisi ini
terjadi pada arteri koronaria yang berfungsi mensuplai darah ke otot
jantung (istilah medisnya miokardium), maka suplai darah jadi
berkurang dan menyebabkan kematian di daerah tersebut (disebut
sebagai infark miokardia). Konsekuensinya adalah terjadinya
serangan jantung dan menyebabkan timbulnya gejala berupa nyeri
dada yang hebat (dikenal sebagai angina pectoris). Keadaan ini yang
disebut sebagai Penyakit Jantung Koroner (PJK).
RANGKUMAN
Asam lemak merupakan sekelompok senyawa hidrokarbon
yang berantai panjang debgan gugus karboksil pada ujungnya.
112 fisiologi hewan

1. Proses oksidasi asam lemak merupakan proses yang terjadi di
mitokondria di mana terjadi pada atom karbon beta sehingga
disebut beta oksidasi. Setiap kali oksidasi menghasilkan asetil
CoA, NADH dan FADH . 2
2. Proses ketogenesis merupakan proses pembentukan badan-
badan keton. Pada manusia proses ini terjadi akibat katabolisme
lemak dan karbohidrat yang tidak seimbang. Proses
ketogenesis sering terjadi pada keadaan kelaparan dan Diabetes
Melitus yang tak terkontrol.
3. Asetoasetat merupakan salah satu bahan bakar dalam jaringan.
4. Proses sintesis asam lemak terjadi di luar mitokondria, dimana
proses ini terjadi memerlukan suatu NADPH sebagai reduktor
2.26 SOAL SOAL LATIHAN
1. Dengan melihat jumlah atom karbon, asam lemak
menghasilkan energi (ATP) dengan jumlah yang lebih besar
dibandingkan karbohidrat. Mengapa demiki- an ?
2. Sebutkan fungsi asam lemak bagi tubuh !
3. Jelaskan mekanisme transport asam lemak ke dalam matrik
mitokondria !
4. Jelaskan mengapa lipolisis dapat menyebabkan kondisi
ketoasidosis darah yang mengarah pada kondisi hipoksia !
5. Terangkan proses sintesis kolesterol di dalam sel hati dengan
katalisator enzim HMG CoA – reduktase !
6. Terakumulasinya kolesterol dalam darah akan menyebabkan
kondisi yang disebut aterosklerosis. Dampak apa yang
kemungkinan dapat terjadi akibat kondisi tersebut ?
113metabolisme lipid

7. Kecerdasan sel sangat dibutuhkan untuk mendukung
kemantapan fungsi sistem tubuh dan kondisi homeostasis.
Senyawa yang mempunyai peran penting dalam mendukung
kecerdasan sel adalah lemak atau lipid. Berikan penjelasan
saudara berkaitan dengan pernyataan tersebut !
8. Jelaskan mekanisme proses katabolisme dan anabolisme lemak
di dalam sel tubuh !
9. Sebutkan macam–macam penyebab terjadinya hiperlipidemia !
10. Jelaskan proses pembentukan lemak pada air susu hewan yang
sedang laktasi !
114 fisiologi hewan

3.1 Pengertian Protein
Protein berasal dari bahasa Yunani, “protos” that mean: the
prime inpoertance artinya terutama atau penting. G Mulder
menemukan bahwa senyawa ini ditemukan pada semua organisme
hidup baik pada tumbuhan maupun hewan. Protein merupakan
komponen terbesar pada jaringan tubuh hewan dan tumbuhan.
Protein merupakan senyawa polipeptida, dimana setiap peptide
tersusun atas asam amino sebagai komponen terkecil. U n s u r
penyusun protein adalah atom C (karbon), H (hydrogen), O (oksigen)
dan N (nitrogen). Dengan kata lain protein adalah senyawa organik
yang merupakan polimer asam-asam amino. Asam amino adalah
senyawa yang tersusun atas gugus karboksil (COOH) dan gugus amin
(NH ). Struktur asam amino sebagai berikut :2
METABOLISME PROTEIN
3

Berdasarkan jenisnya, asam amino terdiri dari asam amino
esensial, non esensial dan semiesensial.
1. Asam amino esensial (indispensable amino acid), yaitu asam
amino yang tidak dapat disintesis oleh tubuh, harus diperoleh
dari luar (makanan).
2. Asam amino non esensial (dispensable amino acid), yaitu asam
amino yang dapat disintesis di dalam tubuh, dari suplai
nitrogen.
3. Asam amino semi esensial (conditionally essencial), yaitu
asam amino yang pada kondisi tertentu tidak dapat dibentuk
oleh tubuh.
Protein memiliki fungsi yang sangat vital dalam proses fisiologis di
dalam tubuh hewan. Fungsi protein, antara lain:
1. Sebagai sumber energy dan membentuk struktur sel
2. Membentuk sistem pertahanan tubuh atau sebagai antibodi
3. Mempertahankan keseimbangan cairan dalam tubuh
Gambar 53. Struktur asam amino
116 fisiologi hewan

4. Mempertahankan keseimbangan asam basa di dalam tubuh
5. Membentuk senyawa tubuh yang esensial, seperti hormon,
enzim dan hemoglobin
Protein Sebagai Sumber Energi
Protein digunakan sebagai sumber energi bila suplai
karbohidrat dan lemak berkurang. Karbohidrat dan lemak adalah
”protein-sparing”, artinya they spare amino acids from being burned
for energy and allow them to be used for protein syntesis.
Protein membentuk struktur sel
Sel tersusun atas protein dimana protein berfungsi untuk
menunjang proses pertumbuhan dan pemeliharaan sel. Selain itu
protein mempunyai fungsi kolagen, yaitu suatu protein fibrosa di
dalam tubuh yang merupakan komponen jaringan ikat (connective
tissue). Protein yang membentuk struktur sel adalah protein yang
Gambar 53. Struktur asam amino
117metabolisme protein

berfungsi struktural, yaitu memberi bentuk spesifik sel serta berperan
dalam proses transport bahan-bahan baku metabolisme, mineral dan
vitamin melintasi membran sel serta penerimaan signal dari ligan
ekstraseluler. Selain itu protein struktural juga berperan dalam proses
pergerakan sel, pembelahan, transport produk sintesis dari organel-
organel yang terdapat dalam sel, memperkuat permukaan sel,
menjaga supaya sel bersatu satu sama lain serta berperan dalam
komunikasi antar sel.
Protein struktural selain terdapat pada membran sel, juga
terdapat pada cairan intraseluler. Protein yang terdapat pada membran
sel dapat berupa saluran (channel), transporter (carrier), reseptor,
pemandu atau tempat bertaut unsur struktural sitoskelet glikokalik.
Berdasarkan integrasi pada membran sel, protein struktural membran
dibedakan menjadi protein integran (integral protein), protein
jangkar (anchor protein) dan protein yang terdapat pada bagian tepi
membran sel (periferal protein). Adapun protein yang terdapat pada
cairan intraseluler adalah sitoskelet. Sitoskelet adalah jaringan kerja
tiga dimensi yang tersusun atas protein filamen dan tubul. Sitoskelet
terdiri atas jaringan mikrotubul, mikrofilamen dan filamen antara.
Sitoskelet berperan dalam pergerakan kromosom selama pembelahan
sel, lokomosi sel, pengaturan posisi dan pergerakan organel dalam
sel, kontraksi otot, denyut silia, pada sel-sel oviduk dan sel-sel trakea,
denyut flagella, seperti pada sel spermatozoa, transport
neurotransmitter pada sel syaraf, mempertahankan konstruksi sel dan
mempertahankan posisi nukleus dalam sel.
Protein yang membentuk struktur semua sel meliputi; rambut,
kuku, kulit dan tulang. Adapun protein motorik berperan
118 fisiologi hewan

membalikkan energi dalam kerja mekanik, mengaktifkan proses
seperti pembelahan sel, kontraksi otot, dan pergerakan sperma.
Protein sebagai sistem pertahanan tubuh
Protein membentuk antibodi yang bertanggung jawab untuk
melawan infeksi dan benda asing yang masuk dalam tubuh. Antibodi
adalah protein yang dihasilkan oleh sel B limfosit sebagai respon
adanya antigen. Antibodi imunoglobulin (Ig A, Ig M dan Ig E) adalah
protein. Protein yang berperan dalam mekanisme pertahanan tubuh
non spesifik, seperti enzim lisozim yang dihasilkan oleh kelenjar air
mata atau sel-sel epitel berlapis banyak semu bersilia pada batang
tenggorokan (trakea). Enzim ini berperan dalam mencerna dinding
sel bakteri, sehingga membunuh banyak bakteri yang memasuki
saluran respirasi bagian atas dan pembukaan di sekitar mata. Adapun
protein yang berperan dalam mekanisme pertahanan tubuh secara
spesifik, adalah antibodi yang dihasilkan oleh sel-sel limfosit T
sitotoksik maupun sel-sel plasma limfosit B.
Protein mengatur keseimbangan cairan tubuh
Protein darah dapat menghasilkan tekanan onkotik yaitu
tekanan untuk mempertahankan agar cairan tubuh tetap berada pada
kompartemennya. Cairan tubuh terdiri dari cairan intraseluler (di
dalam sel) dan cairan ekstraseluler (interseluler dan intravaskuler).
Defisiensi protein di dalam darah akan menyebabkan odema, karena
tekanan onkotik yang dihasilkan oleh protein tidak mampu
mempertahankan cairan untuk tetap berada dalam kompartemen,
sehingga cairan akan keluar dan menyebabkan odema.
119metabolisme protein

Protein mengatur keseimbangan asam basa
� Protein darah dapat berperan sebagai buffer, karena pH darah
ditentukan oleh konsentrasi ion hidrogen. Semakin tinggi konsentrasi
ion hidrogen semakin rendah pH (semakin asam = pH lebih rendah
dari 7).
Protein membentuk senyawa tubuh esensial
Protein yang berfungsi membentuk senyawa tubuh yang
esensial adalah protein fungsional. Protein fungsional: protein yang
berfungsi dalam proses regulasi, katalisasi, seperti pertumbuhan,
perkembangan maupun proses-proses metabolisme yang terjadi
dalam tubuh serta mekanisme sistem pertahanan tubuh terhadap
substansi asing dari luar tubuh yang membahayakan, baik mekanisme
pertahanan tubuh yang non spesifik maupun spesifik. Contoh protein
fungsional adalah hormon, enzim dan hemoglobin. Hormon adalah
suatu senyawa yang dihasilkan oleh tubuh yang tersusun atas protein
yang mempunyai fungsi sebagai regulator proses metabolisme,
pertumbuhan atau perkembangan. Hormon diproduksi oleh sel-sel di
seluruh kelenjar endokrin maupun eksokrin di seluruh tubuh, baik
melibat aksis hipotalamus-hipofisis organ target ataupun tidak.
Contoh kelenjar endokrin yang berada dalam tubuh, seperti
hipothalamus, hipofisis, kelenjar tiroid dan paratiroid, adrenalin,
gonad (ovarium dan testis), kelenjar asinus pada pulau-pulau
Langerhans pankreas dan lain-lain. Contoh kelenjar eksokrin adalah
kelenjar asinus pankreas serta kelenjar lambung. Sedangkan enzim
merupakan protein yang mengkatalisasi reaksi kimia di dalam tubuh
agar dapat menghasilkan produk metabolit yang optimal untuk
120 fisiologi hewan

kebutuhan hidup pokok, maintenance atau homeostasis. Enzim
terdapat dalam cairan intraseluler, baik yang terdapat dalam organel-
organel bermembran, seperti lisosom, peroksisom, mitokondria
maupun terdapat bebas dalam sitosol. Selain itu enzim juga terdapat
dalam nukleoplasma nukleus sel. Hemoglobin adalah komponen
darah yang tersusun atas protein dan zat besi yang berperan dalam
pengikatan oksigen pada saat inhalasi dan transportasi menuju ke sel
seluruh tubuh.
Selain hormon, enzim dan hemoglobin juga terdapat protein
fungsional lain yang berperan sebagai alat transport zat gizi dalam
darah, seperti protein transferin yang berperan dalam transport zat
besi, protein pengikat asam folat (folate binding proteine), retinol
binding protein, lipoprotein dan lain-lain
3.2 Pencernaan Protein
Tidak seperti karbohidrat, protein tidak mengalami pencernaan
di mulut. Pencernaan protein dimulai di lambung dengan melibatkan
enzim pepsin. Enzim pepsin berfungsi mengkatalisis perombakan
protein menjadi komponen-komponen penyusunnya yang lebih kecil,
yaitu proteosa, pepton, polipeptida-polipeptida, dipeptida dan asam
amino. Di dalam lambung, pencernaam protein melibatkan HCl yang
dihasilkan oleh sel-sel epitel lambung.
Dalam lambung HCl berfungsi dalam mengaktifkan proenzim
(pepsinogen) yang dihasilkan oleh sel-sel pada bagian permukaan
lambung yang menghadap ke lumen menjadi enzim pepsin,
mendenaturasi protein sehingga protein mudah dipecah oleh enzim,
mengoptimumkan pH yang cocok untuk aktivitas pepsin dalam
121metabolisme protein
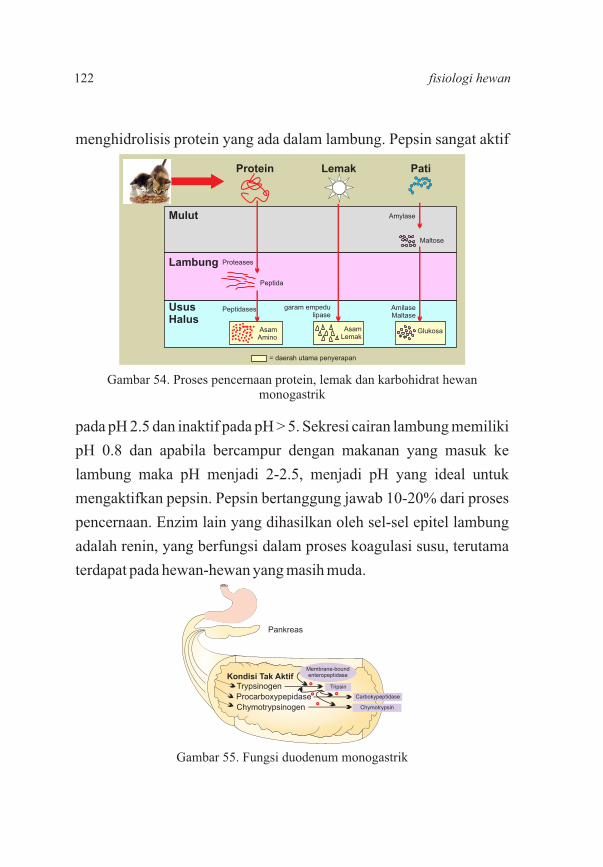
menghidrolisis protein yang ada dalam lambung. Pepsin sangat aktif
pada pH 2.5 dan inaktif pada pH > 5. Sekresi cairan lambung memiliki
pH 0.8 dan apabila bercampur dengan makanan yang masuk ke
lambung maka pH menjadi 2-2.5, menjadi pH yang ideal untuk
mengaktifkan pepsin. Pepsin bertanggung jawab 10-20% dari proses
pencernaan. Enzim lain yang dihasilkan oleh sel-sel epitel lambung
adalah renin, yang berfungsi dalam proses koagulasi susu, terutama
terdapat pada hewan-hewan yang masih muda.
Gambar 54. Proses pencernaan protein, lemak dan karbohidrat hewan monogastrik
Gambar 55. Fungsi duodenum monogastrik
122 fisiologi hewan
Pankreas
Kondisi Tak AktifTrypsinogen
Procarboxypepidase
Chymotrypsinogen
Membrane-boundenteropeptidase
Tripsin
Carbokypeptidase
Chymotrypsin
+
+ +
+

Pencernaan selanjutnya terjadi di usus halus, tepatnya pada
bagian duodenum dengan melibatkan enzim-enzim endopeptidase
dan eksopeptidase. Pada bagian ini pemecahan protein berlangsung
lebih efektif. Berbeda dengan pencernaan di lambung yang
melibatkan HCl sehingga bersifat asam, pencernaan di duodenum
bersifat alkali. Sel-sel asinus pankreas menghasilkan proenzim
tripsinogen dan kemotripsinogen sebagai respons masuknya protein
di dalam usus yang kemudian proenzim tersebut berubah menjadi
enzim tripsin dan kemotripsin karena suasana yang alkali. Dalam
duodenum polipeptida-polipeptida kecil diubah menjadi komponen
asam-asam aminonya atau menjadi peptida kecil (fragmen yang
panjangnya hanya dua atu tiga asam amino) dengan katalisasi enzim
tripsin atau kemotripsin. Tripsin dan kemotripsin bersifat spesifik
untuk ikatan peptida yang berdekatan dengan asam amino tertentu.
Dengan demikian pepsin memutuskan polipeptida tertentu menjadi
rantai-rantai yang lebih pendek. Selain enzim tripsin atau
kemotripsin, terdapat enzim-enzim lain yang bekerja mengkatalisasi
polipeptida atau peptida menjadi komponen penyusunnya, seperti
karboksipept idase , aminopept idase dan d ipept idase .
Karboksipeptidase dihasilkan oleh sel-sel asinus pankreas dalam
bentuk inaktif, seperti halnya tripsin dan kemotripsin. Enzim-enzim
tripsin, kemotripsin dan karboksipeptidase adalah pankreatik
protease. Di dalam lumen usus halus enzim ini bersama-sama dengan
tripsin dan kemotripsin diaktifkan oleh enteropeptidase, baik secara
langsung atau tidak langsung. Enzim enteropeptidase dihasilkan oleh
sel sel epitel usus halus, seperti halnya aminopeptidase.
Selanjutnya karboksipeptidase berfungsi mengkatalisis
123metabolisme protein

pemecahan polipeptida menjadi komponen penyusunnya, yaitu
asam-asam amino yang dimulai pada bagian ujung polipeptida yang
memiliki gugus karboksil yang bebas, sedangkan aminopeptiadase
bekerja dengan arah yang sebaliknya, yaitu pemecahan polipeptida
menjadi asam-asam amino dimulai pada bagian ujung yang memiliki
gugus amina. Melalui proses yang sinergi antara enzim tripsin,
kemotripsin, karboksipeptidase dan aminopeptidase pencernaan atau
hidrolisis protein dapat berjalan secara sempurna. Enzim lainnya
yang terlibat dalam pencernaan protein adalah dipeptidase. Enzim ini
dihasilkan oleh sel-sel epitel yang melekat pada bagian glikokaliks
permukaan luar sel usus yang berfungsi dalam mempercepat
pencernaan dengan cara memecah peptida-peptida kecil menjadi
asam-asam amino yang siap untuk diabsorbsi oleh sel-sel epitel usus
dan masuk ke dalam sistem peredaran darah.
3.3 Absorbsi dan Transportasi Asam-Asam Amino
Asam-asam amino hasil perombakan protein terdiri atas L-
asam amino (asam-asam amino Lefo) dan D-asam amino (asam-asam
amino dektro). Asam-asam amino tersebut akan diabsorbsi oleh sel-
sel epitel usus halus dan selanjutnya masuk ke dalam sistem
peredaran darah. Absorbsi asam-asam amino lefo terjadi lebih cepat
dibandingkan dengan asam-asam amino dektro dan terjadi secara
selektif. Absorbsi asam-asam amino oleh sel- sel epitel usus halus
terjadi melalui transport aktif dengan melibatkan protein transport
membran (permease/karier permease/transporter) yang dikopel
dengan sumber energi ATP.
Asam-asam amino yang terabsorbsi dan masuk ke sistem
124 fisiologi hewan
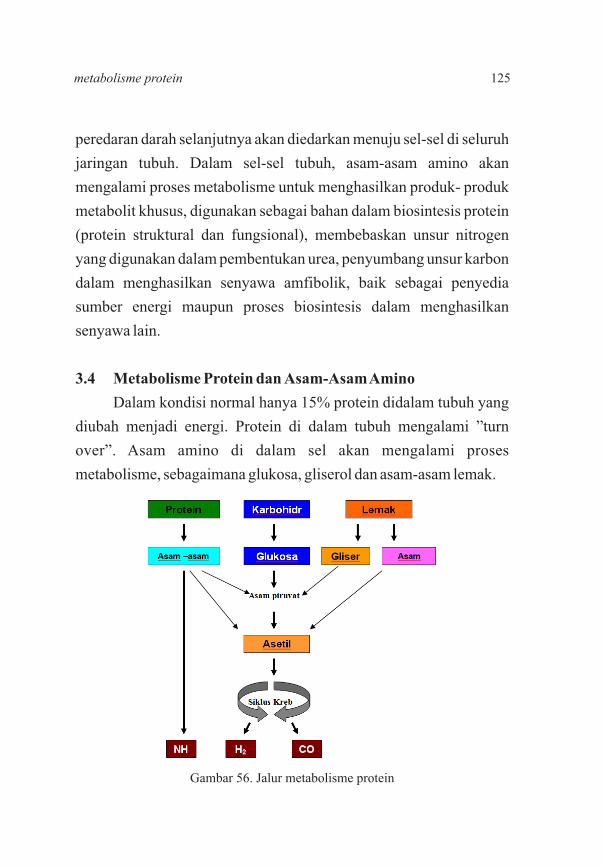
peredaran darah selanjutnya akan diedarkan menuju sel-sel di seluruh
jaringan tubuh. Dalam sel-sel tubuh, asam-asam amino akan
mengalami proses metabolisme untuk menghasilkan produk- produk
metabolit khusus, digunakan sebagai bahan dalam biosintesis protein
(protein struktural dan fungsional), membebaskan unsur nitrogen
yang digunakan dalam pembentukan urea, penyumbang unsur karbon
dalam menghasilkan senyawa amfibolik, baik sebagai penyedia
sumber energi maupun proses biosintesis dalam menghasilkan
senyawa lain.
3.4 Metabolisme Protein dan Asam-Asam Amino
Dalam kondisi normal hanya 15% protein didalam tubuh yang
diubah menjadi energi. Protein di dalam tubuh mengalami ”turn
over”. Asam amino di dalam sel akan mengalami proses
metabolisme, sebagaimana glukosa, gliserol dan asam-asam lemak.
Gambar 56. Jalur metabolisme protein
125metabolisme protein

Metabolisme asam-asam amino di dalam sel meliputi proses
katabolisme dan anabolisme. Melalui proses katabolisme di dalam
sel, unsur nitrogen yang terdapat dalam asam-asam amino akan
dilepaskan untuk membentuk urea (CO(NH ) ). Selain itu dari proses 2 2
katabolisme asam-asam amino juga dilepaskan unsur karbon yang
digunakan untuk pembentukan senyawa amfibolik.
Proses anabolisme asam-asam amino akan menghasilkan
protein dengan melibatkan 20 macam asam-asam amino dasar, yaitu
asam-asam amino esensial dan non esensial. Asam amino esensial
meliputi; arginin histidin, ilein, leusin, listin, metionin, phenilalanin,
thirosin, triptopan, maupun valin. Asam-asam amino ini tidak dapat
disintesis oleh sel-sel tubuh dan harus didatangkan dari luar melalui
makanan. Dari 10 macam asam-asam amino tersebut, asam amino
histidin bersifat semi esensial, dimana asam-asam amino ini dapat
disintesis oleh sel-sel tubuh tetapi tidak mencukupi untuk mendukung
proses metabolisme. Adapun asam-asam amino yang bersifat non
essensial adalah; alanin, asparagin, sistein, glutamat, glutamin,
prolin, serin, tirosin, hidroksiprolin dan hidroksilisin.
Gambar 57. Bagan katabolisme dan anabolisme asam amino
fisiologi126

Protein dalam tubuh bersifat dinamis, selalu ada proses
biosintesis dan degradasi. Dengan mengukur jumlah nitrogen yang
masuk dan keluar, kondisi metabolisme protein di dalam tubuh dapat
diperkirakan, oleh karena nitrogen dalam tubuh terdapat pada protein.
Nitrogen masuk ke dalam tubuh lewat makanan dan keluar dari dalam
tubuh dalam bentuk urin, keringat dan feses. Apabila asam-asam
amino terdapat dalam jumlah yang berlebih di dalam tubuh, maka
asam-asam amino akan digunakan sebagai bahan untuk sintesis
protein dan sintesis produk khusus, seperti serotonin. Asam-asam
amino yang tersisa akan dikatabolisme menghasilkan senyawa urea
melalui penyumbangan unsur nitrogen dan menghasilkan senyawa
amfibolik melalui pelepasan atom karbon. Senyawa amfibolik ini
merupakan senyawa yang terlibat dalam siklus asam sitrat (siklus
Kreb) dan selanjutnya digunakan dalam proses biosintesis lemak
maupun sintesis glikogen. Beberapa senyawa amfibolik, antara lain
oksaloasetat, fumarat, suksinil Koa dan α-ketoglutarat. Senyawa
amfibolik ini bersifat glukogenik karena dapat digunakan untuk
sintesis glukosa.
Hasil katabolisme kerangka karbon asam amino dapat
menghasilkan asam piruvat yang selanjutnya akan diubah menjadi
asam oksaloasetat yang bersifat glukogenik. Selain asam piruvat,
hasil katabolisme asam amino juga digunakan untuk membentuk
asetil Koa tanpa melalui piruvat dan bersifat ketogenik karena dapat
membentuk benda-benda keton (senyawa keton). Berdasarkan
katabolisme kerangka karbon asam amino, terdapat beberapa sifat
asam amino, antara lain:
1. Asam amino yang murni bersifat ketogenik, seperti leusin dan
127metabolisme protein

lisin
2. Asam amino yang membentuk fragmen ketogenik dan fragmen
glukogenik (bersifat campuran), seperti ilein, phenilalanin dan
triptopan
3. Asam amino yang murni bersifat glukogenik, seperti arginin,
histidin, metionin, treonin dan valin.
Keseimbangan nitogen di dalam tubuh dikatakan positif
apabila nitrogen yang masuk ke dalam tubuh lebih besar dibandingan
nitrogen yang keluar. Dengan demikian produk dari proses
katabolisme protein dapat digunakan secara optimal untuk
mendukung proses sintesis protein. Hal ini biasanya terjadi pada saat
proses penyembuhan luka atau infeksi penyakit, pada masa
pertumbuhan atau kehamilan. Keseimbangan nitrogen dikatakan
negatif, apabila produk yang dihasilkan dari proses katabolisme
sedikit yang digunakan untuk proses sintesis dan sebagian besar
digunakan untuk pemeliharaan atau adaptasi. Sebagai contoh pada
saat tubuh mengalami kelaparan atau sakit. Dalam kondisi normal
atau sehat keseimbangan nitrogen berada pada kondisi yang
homeostasis.
Hewan tingkat tinggi dan manusia tidak dapat mensintesis
protein dari asam amino esensial dan asam amino tersebut harus di
datangkan dari luar tubuh, karena sel tidak memiliki kemampuan
untuk melakukan sintesis asam amino tersebut. Berbeda dengan asam
amino non esesial, sel mempunyai kemampuan untuk mensintesis
protein dari bahan asam amino tersebut. Protein yang terdapat pada
sel hewan memiliki nilai biologis lebih tinggi dibanding protein
nabati, oleh karena komposisi asam aminonya lebih lengkap dan
fisiologi128

kadarnya lebih tinggi, terutama asam amino esensialnya.
Senyawa amfibolik dapat disintesis menjadi senyawa lain
melalui proses anabolisme dan dapat dikatabolisme atau dioksidasi.
Sebagai contoh senyawa intermedier dalam siklus Kreb, yaitu α-
ketoglutarat dapat digunakan untuk sintesis glutamat. Demikian pula
asam keto dan gugus amino melalui proses oksidasi dapat
menghasilkan asam amino tertentu. Berikut contoh biosintesis asam
amino non esensial dengan sesama asam amino esensial, dengan
senyawa amfibolik, dan asam amino essensial:
a. Biosintesis asam amino non esensial dari asam amino non
esensial lain
Glutamat prolin hidroksiprolin
Serin glisin
b. Biosintesis asam amino esensial dari senyawa amfibolik
Alfa ketoglutarat glutamat glutamin
Piruvat alanin
Oksaloasetat aspartat
D3 Fosfogliserat serin
Kholin/glioksilat glisin
c. Biosintesis asam amino esensial dari asam amino esensial
Phenilalanin tirosin
Lisin hidroksilisin
Senyawa α ketoglutarat yang merupakan anggota siklus asam
sitrat dapat diubah menjadi asam amino melalui reaksi transaminasi
(transfer gugus amino) yang dikatalis oleh enzim tranaminase (amino
transferase). Alfa ketoglutarat yang merupakan asam keto yang
bereaksi dengan gugus amino (amino alanin) menghasilkan asam
129metabolisme protein

amino glutamat. Alanin kehilangan gugus aminonya dan berubah
menjadi piruvat (asam keto). Kalau alfa ketoglutarat bereaksi dengan
aspartat, maka akan menghasilkan oksaloasetat.
Contoh biosintesis asam amino non esensial dari senyawa
amfibolik melalui proses transaminasi adalah, biosintesis asam
amino aspartat dari oksaloasetat, alanin dari piruvat serta glutamin
dari glutamat. Biosintesis glutamin dari glutamat dapat dilihat pada
reaksi berikut:
3.5 Katabolisme Asam Amino yang Mengandung Nitrogen
Katabolisme asam amino (asam amino nitrogen) dibagi
menjadi beberapa tahap, yaitu: transaminasi, deaminasi oksidatif,
transport amonia dan siklus urea.
Tahap Transaminasi
Transaminasi adalah proses pemindahan gugus amin dari satu
asam amino untuk membentuk asam amino baru. Proses transaminasi
melibatkan enzim transaminase atau amino-transferase dengan
kofaktor enzim piridoksal fosfat. Contoh proses transaminasi adalah
pembentukan sistin dari metionin. Transaminase yang di dapatkan
pada seluruh jaringan pada hewan mamalia adalah alanin
Gambar 57. Bagan katabolisme dan anabolisme asam amino
fisiologi hewan130
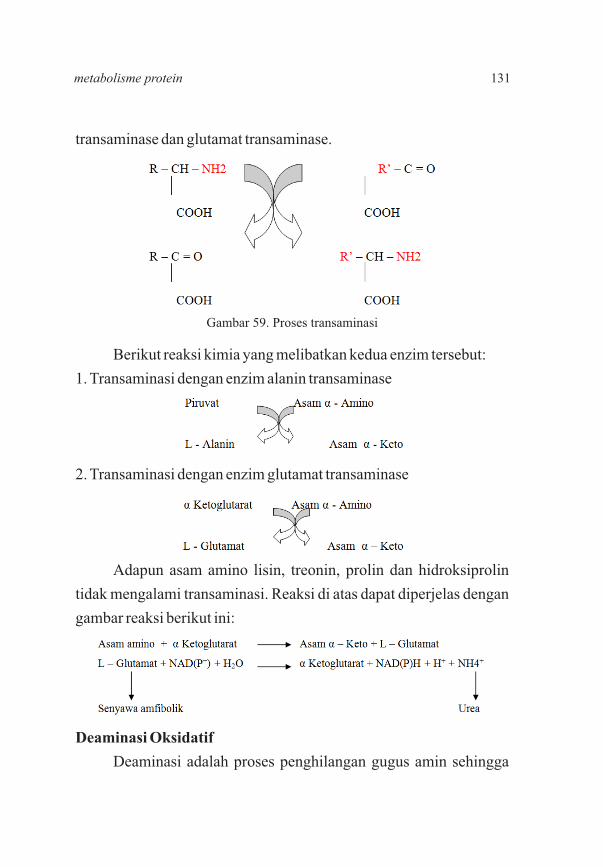
transaminase dan glutamat transaminase.
Berikut reaksi kimia yang melibatkan kedua enzim tersebut:
1. Transaminasi dengan enzim alanin transaminase
2. Transaminasi dengan enzim glutamat transaminase
Adapun asam amino lisin, treonin, prolin dan hidroksiprolin
tidak mengalami transaminasi. Reaksi di atas dapat diperjelas dengan
gambar reaksi berikut ini:
Deaminasi Oksidatif
Deaminasi adalah proses penghilangan gugus amin sehingga
Gambar 59. Proses transaminasi
131metabolisme protein

akan membentuk senyawa baru. Proses deaminasi membutuhkan
oksigen yang diperlukan untuk oksidasi dengan melibatkan enzim
dehidrogenase. Contoh proses deaminasi adalah pembentukan niasin
dari triptopan. Proses ini membebaskan atom hidrogen dan amonia
(NH ). Berikut contoh reaksi deaminasi oksidatif yang melibatkan 3
enzim L-glutamat dehidrogenase:
Proses deaminasi oksidatif yang melibatkan enzim L-glutamat
dehidrogenase bersifat reversibel. Enzim L-glutamat dehidrogenase
bersifat sebagai enzim pengendali, yaitu sebagai inhibitor allosterik
yang berkaitan dengan pembentukan ATP, GTP dan NADH dan pada
sisi lain bersifat aktivator allosterik yang berkaitan dengan
pembentukan ADP dan GDP. Enzim ini dijumpai dalam sitoplasma
dan mitokondria pada sel yang terdapat di seluruh jaringan tubuh.
Transport Amonia
Amonia bersifat toksik, sehingga tidak diangkut dalam bentuk
bebas di luar jaringan hati (hepar). Transport amonia terjadi pada
sebagian besar jaringan tubuh, dengan mekanisme sebagai berikut:
- Glutamin sintetase akan mengkatalisis perubahan amonia
menjadi glutamin yang bersifat nontoksik
Glutamat + NH + + ATP Glutamin + H O + ADP + Pi4 2
Glutamin sintetase
- Glutamat berasal dari α ketoglutarat yang merupakan senyawa
fisiologi hewan132

intermedier dalam siklus Kreb melalui reaksi transaminasi
dengan asam-asam amino lain.
- Glutamin diangkut oleh darah dalam sistem pembuluh menuju
ke hati, ginjal dan usus.
- Dalam hati glutamin dihidrolisis untuk melepas ammonia yang
akan masuk siklus urea+
Glutamin + H O Glutamat + Nh42
Glutaminase
- Dalam ginjal glutaminase membebaskan amonia untuk
diekskresikan dengan kelebihan asam dari darah dalam rangka
untuk mencapai kesetimbangan asam basa.
Mekanisme transport amonia juga terjadi pada otot dengan
mekanisme yang spesifik. Dalam otot, 30% dari asam amino nitrogen
yang dibentuk dari proses katabolisme protein dikirim ke hati melalui
pembuluh darah dalam bentuk alanin dengan suatu siklus yang
disebut dengan ”glukosa-alanin”. Siklus ini diawali dari proses
glikolisis pada otot yang menghasilkan asam piruvat. Piruvat
selanjutnya mengalami proses transaminasi menghasilkan alanin
dengan proses sebagai berikut:
Asam piruvat + glutamat alanin + α ketoglutarat
Glutamat yang digunakan pada reaksi transaminasi tersebut dibentuk
dari α ketoglutarat dan NH4+ berasal dari katabolisme asam-asam
amino dengan dikatalisasi enzim glutamat dehidrogenase.
Alanin hasil transaminasi piruvat dalam otot kemudian
diangkut menuju ke hati dan kemudian terjadi reaksi
sebaliknya, yaitu:
α ketoglutarat + alanin glutamat + piruvat
133metabolisme protein

Glutamat yang terbentuk dalam sel hati kemudian mengalami
deaminasi oksidatif membentuk amonia dan α ketoglutarat. Amonia
selanjutnya akan diubah menjadi urea melalui siklus urea. Piruvat
yang terbentuk merupakan metabolit yang dapat digunakan untuk
sintesis glukosa melalui proses glukoneogenesis di hati, dan
selanjutnya glukosa akan dimobilisasi ke dalam pembuluh darah dan
kembali lagi ke otot. Pada kasus kelaparan sumber utama
glukoneogenesis di hati adalah asam-asam amino hasil pemecahan
protein otot, karena otot merupakan jaringan tubuh yang kaya akan
protein. Banyak protein dalam otot yang mengalami proses
katabolisme menghasilkan asam piruvat, dimana asam piruvat ini
kemudian diangkut ke hati sebagai alanin.
Sintesis urea (CO(NH ) ) berasal dari ammonia, 2 2
karbondioksida (CO ) dan asam amino asparagin. Melalui reaksi 2
Gambar 60. Siklus glukosa-alanin
fisiologi hewan134

endorgenik, setiap sintesis satu molekul urea memerlukan 3 molekul
ATP dengan bantuan enzim glutamat dehidrogenase dan karbamoil
fosfat sintetase melalui reaksi yang terkenal dengan siklus urea. Urea
yang dihasilkan akan masuk ke dalam sirkulasi darah dan
diekskresikan keluar tubuh dalam bentuk urin melalui ginjal.
Beberapa enzim yang berperan dalam siklus urea antara lain:
karbamoil fosfat sintetase, ornitin transkarbamoilase, asam
argininosuksinat sintetase, arginino suksinase dan arginase. Reaksi
yang melibatkan enzim karbamoil fosfat sintetase dan ornitin
transkarbamoilase terjadi di dalam matriks mitokondria di sel hati,
sedangkan reaksi yang melibatkan enzim asam argininosuksinat
sintetase, arginino suksinase dan arginase terjadi di dalam sitosol sel
hati.
Amonia yang merupakan bahan dasar sintesis urea berasal dari
hasil katabolisme asam amino nitrogen di jaringan tubuh. Selain itu
amonia juga dihasilkan dari hasil kerja bakteri usus terhadap sisa-sisa
protein atau asam amino dalam makanan dan urea hasil sekresi sel-sel
usus. Pada kasus sirosis hati, diketahui bahwa kadar amonia darah
meningkat, lebih-lebih apabila disertai perdarahan gastrointestinal.
Ammonia dalam jumlah yang melampaui ambang batas bersifat
toksik terhadap sistem saraf dan dapat menyebabkan timbulnya
beberapa gangguan, seperti gangguan penglihatan, gangguan bicara,
flapping tremor, koma sampai kematian. Pada kasus kerusakan ginjal
berat biasanya ditandai dengan terjadinya uremia, yaitu
meningkatnya kadar ureum.
135metabolisme protein

3.6 Metabolisme Protein pada Hewan Poligastrik
(Ruminansia)
a. Karakteristik Pakan yang Menentukan Intake
Ruminansia dapat mencerna pakan kasar, karena memiliki
kemampuan dalam menfermentasi serat. Proses fermentasi adalah
proses yang lambat, serat pakan diproses dalam waktu yang lama di
dalam saluran pencernaan untuk memperoleh zat-zat yang dapat
dicerna. Jika terlalu banyak bahan yang tidak dapat dicerna maka
intake akan menurun. Intake dipengaruhi oleh kapasitas rumen,
reseptor dinding rumen menyampaikan sinyal dari isi rumen ke otak.
Tapi kapasitas maksimum dan pakan yang mengisi rumen tidak dapat
diketahui secara pasti.
Pakan yang voluminous (bulky) seperti hay (silase), akan
mengisi rumen dengan jumlah lebih banyak dari pada konsentrat jika
rumput tersebut dipotong-potong. Berdasarkan kapasitas rumennya,
hewan ruminasia makan dalam jumlah yang konstan. Hal ini
dibuktikan dengan beberapa eksperimen. Pada pakan dengan
kandungan air yang berbeda-beda mempengaruhi kapasitas rumen
serta intake pakan. Pemberian pakan dengan kandungan air yang
tinggi dapat menurunkan intake bahan kering (BK) bila dibandingkan
dengan pakan kandungan air rendah. Intake makanan pada
ruminansia berbeda dengan intake pada monogastrik. Hal tersebut
dapat dilihat dari metabolisme glukosa. Pada ruminansia glukosa
yang diserap dalam saluran pencernaan relatif sedikit dan level
glukosa dalam darah pun rendah. Hal ini ada keterkaitannya dengan
kebiasaan makan ruminansia. Mekanisme intake pada ruminansia
berhubungan dengan proses penyerapan volatile fat acid (VFA)
fisiologi hewan136

dalam rumen. Penyerapan asetat dan propionat oleh dinding rumen
dapat menurunkan intake konsentrat oleh ruminansia. Hal ini
menunjukan bahwa terdapat reseptor-reseptor dalam lumen/dinding
retikulo-rumen. Proses penyerapan VFA ke dalam hepatit vena portal
juga menurunkan intake. Hal ini dilakukan dengan cara pengiriman
sinyal dari hati ke hipotalamus. Butirat mempengaruhi intake dalam
level lebih rendah bila dibandingkan dengan asetat dan propionat,
karena butirat dimetabolisme menjadi aceta.
Pakan dengan kandungan BK tinggi berpengaruh terhadap
intake. Pada ruminansia intake dipengaruhi oleh tingkat penyerapan
dan bentuk pakan. Persentase daya cerna dan tingkat konsumsi tidak
hanya dipengaruhi oleh proporsi dinding sel pakan tetapi bentuk fisik
dari dinding sel tersebut. Hijauan yang digiling, struktur dinding
selnya rusak sehingga proses pencernaannya lebih cepat dan tingkat
konsumsi meningkat. Partikel hijauan yang digiling tersebut berjalan
dengan cepat meninggalkan rumen, sehingga rumen cepat kosong.
Hal ini yang menyebabkan terjadi peningkatan konsumsi. Bagian
daun dapat dicerna dan dikonsumsi lebih tinggi dibandingkan
batang, karena dinding sel pada daun lebih mudah dihancurkan dari
pada batang. Hewan yang diberi daun dapat mengkonsumsi lebih dari
40% BK per hari bila dibandingkan dengan pemberian batang.
Kekurangan zat makanan tertentu pada ransum dapat
menurunkan aktivitas mikroba rumen sehingga tingkat konsumsi
menurun. Beberapa nutrien yang berpengaruh terhadap intake adalah
protein, sulfur, phospor, sodium dan kobal. Silase yang mengandung
produk-produk fermentasi yang tinggi. Namun pada silase dengan
kandungan amonia yang tinggi, karena prosesnya yang kurang baik,
137metabolisme protein

akan menurunkan tingkat konsumsi, walaupun silase tersebut
memiliki dinding sel yang mudah dicerna. Disamping itu bentuk fisik
silase juga mempengaruhi tingkat konsumsinya, silase dari hijauan
yang digiling terlebih dahulu memiliki tingkat konsumsi yang lebih
tinggi.
b. Pengaruh Hewan terhadap Tingkat Konsumsi
Kapasitas rumen merupkan faktor yang menentukan tingkat
konsumsi hewan ruminansia. Kapasitas rumen berbagai hewan
ruminansia berbeda sehingga konsumsi hewan ruminansia berbeda-
beda. Konsumsi hewan ruminansia ditentukan oleh bobot badan 0.75
metabolik (BB ). Jumlah konsumsi pada sapi lebih besar dari pada
domba per unit bobot metabolik. Contoh, sapi denga berat 300 kg,
yang diberi pakan pakan mengandung 11 MJ ME/Kg BK akan 0.75
mengkonsumsi sekitar 90 g BK per Kg BB per hari (6.3
kg/ekor/hari). Domba (40 Kg) akan mengkonsumsi 60 g BK per kg 0.75
BB per hari (0.96 g per ekor). Hewan gemuk memiliki konsumsi
yang seimbang, dengan kata lain tidak bertambah sesuai dengan
pertambahan BB. Hal ini dikarenakan lemak abdomen yang dideposit
menurunkan volume rumen, bisa juga efek metabolisme. Hewan
dengan kandungan daging yang tinggi memiliki jumlah konsumsi per
BB metabolik yang tinggi. Hal ini dapat ditunjukan oleh hewan yang
mengalami pertumbuhan konpensasi karena pemberian makan yang
dibatasi dan kandungan zat makanan dalam ransum yang rendah.
Pada hewan yang bunting, ada dua hal yang berlawanan
mempengaruhi konsumsi. Peningkatan kebutuhan nutrisi fetus
meningkatkan konsumsi ransum. Sedangkan, akibat lain dari
fisiologi hewan138

kebuntingan adalah menurunkan kapasitas rumen karena
pertumbuhan foetus yang semakin besar. Hal ini menyebabkan
terjadinya penurunan konsumsi terutama jika pakan terdiri dari
hijauan saja.
Konsumsi pada ruminansia berhubungan dengan fase laktasi.
Awal laktasi, sapi perah kehilangan bobot badannya. Hal ini terganti
pada fase akhir laktasi, dimana produksi susu mulai turun dan
konsumsi bahan kering meningkat. Konsumsi energi bruto pada sapi
laktasi 50% lebih tinggi dari pada sapi yang tidak laktasi.
c. Faktor Lingkungan yang Mempengaruhi Konsumsi
Tingkat konsumsi hewan ruminansia yang digembalakan di
pastura atau padang penggembalaan dipengaruhi oleh komposisi
kimia dan daya cerna hijauan serta struktur dan distribusi hijauan di
padang penggembalaan tersebut. Konsumsi hewan di padang
penggembalaan tergantung dari :
1. Ukuran renggutan (kuantitas/jumlah bahan kering yang dapat
diperoleh dalam satu gigitan)
2. Kecepatan renggutan (jumlah gigitan dalam satu menit)
3. Waktu yang dibutuhkan untu merumput
Sebagai contoh, sapi (600 kg) memiliki ukuran renggutan 0.6 g
BK, dengan kecepatan gigi 60 kali per menit dan mendapatkan
hijauan 36 g BK per menit atau 2.16 kg BK perjam. Untuk
mendapatkan konsumsi 16 kg BK perhari maka sapi harus merumput
selama 16/2.16 = 7.4 jam per hari. Sapi perah biasanya merumput
selama 8 jam perhari. Sapi dapat mengkonsumsi dalam jumlah
banyak bila ukuran gigitan dan kecepatan gigitan tinggi yang dapat
139metabolisme protein

terjadi jika distribusi hijauan merata. Hijauan diusahakan pendek (12
x 15 cm) dan tebal agar ukuran gigitan maksimum. Hewan lebih
menyukai bagian daun dari pada batang karena batang lebih sulit
dicerna. Hewan juga lebih menyukai hijauan berwarna hijau (masih
segar) dari pada hijauan yang layu. Tidak semua hijauan di makan
hewan karena hijauan yang berduri dan terkontaminasi feses tidak
disukai hewan.
Pada kondisi pastura yang baik hewan dapat mengkonsumsi
hijauan sebanyak mungkin. Tetapi pada kondisi pastura yang buruk
hewan hanya akan mengkonsumsi hijauan yang mudah dicerna dan
dapat dimetabolis. Suhu lingkungan sangat berpengaruh terhadap
tingkat konsumsi. Pada temperatur di bawah temperatur netral hewan
akan meningkatkan konsumsi dan pada suhu di atas suhu netral
hewan akan menurunkan konsumsi. Sapi Bos Taurus akan o
menurunkan 2% konsumsinya setiap kenaikan suhu 1 C di atas suhu o
rata-rata 25 C. Panjang hari juga mempengaruhi tingkat konsumsi.
Semakin pendek hari maka tingkat konsumsi pada domba semakin
menurun. Panjang hari tidak terlalu berpengaruh pada tingkat
konsumsi sapi. Kondisi kesehatan hewan berpengaruh terhadap
tingkat konsumsi. Hewan yang sakit cenderung menurunkan tingkat
konsumsinya. Hal tersebut dikarenakan daya serap saluran
pencernaan terhadap zat makanan menurun, serta akibat sistem
kekebalan tubuh hewan menurun karena adanya parasit yang masuk.
Agar dapat menghasilkan performan produksi yang tertinggi,
hewan memerlukan nutrien. Nutrien ini dibutuhkan untuk hidup
pokok (maintenance) dan berbagai produksi. Faktor yang harus
diperhatikan adalah jumlah makanan yang diberikan, semakin
fisiologi hewan140

banyak jumlah makanan yang dikonsumsi setiap hari, akan semakin
memberikan kesempatan untuk menghasilkan produksi tinggi.
Peningkatan produksi yang diperoleh dari konsumsi makanan yang
lebih tinggi biasanya berkaitan dengan peningkatan efesiensi proses-
proses produksi, sehingga proporsi kebutuhan pokok menurun
sedangkan produksi meningkat.
Proses makan (feeding) adalah aktivitas yang komplek, yang
meliputi mencari makanan, mengamati, pergerakan, aktifitas
sensorik, memakan dan mencerna. Dalam saluran pencernaan
makanan dan zat-zat makanan diserap dan dimetabolismekan. Semua
proses ini dapat mempengaruhi konsumsi makanan dalam jangka
pendek (short term basis). Namun demikian perlu diperhatikan
bahwa, pada hewan dewasa kebutuhan pokoknya (berat tubuhnya)
relatif konstan, walaupun makanan tersedia ad libitum. Dengan
demikian konsep jangka pendek-jangka panjang dalam mengontrol
konsumsi harus diperhatikan. Walaupun sistem kontrol ini sama pada
setiap jenis hewan, namun ada perbedaan antar spesies yang
tergantung pada pada struktur dan fungsi saluran pencernaannya.
Mekanisme kontrol konsumsi makanan dilakukan sebagai berikut:
1. Level metabolik: konsentrasi zat-zat makanan, metabolit atau
hormon dapat menstimulir sistem syaraf pusat (CNS=Central
Nervous System) yang menyebabkan hewan mulai atau
berhenti makan.
2. Level sistem pencernaan: jumlah digesta dapat ditentukan yang
dapat dicerna oleh hewan.
3. Pengaruh external: misalnya iklim
141metabolisme protein

d. Penggunaan protein atau asam amino pada hewan
Ruminansia
Keberadaan mikroba di dalam rumen, mengakibatkan
metabolisme protein pada ruminansia berbeda dengan monogastrik.
Mikroba mempunyai kemampuan mensintesis semua asam amino
termasuk asam-asam amino yang dibutuhkan oleh induk semang. Hal
ini menunjukkan bahwa kualitas protein tidak menjadi unsur mutlak
dalam ransum ruminansia, sehingga pemberian garam ammonium
atau urea sudah mencukupi kebutuhan hewan ruminansia akan
protein. Penggunaan protein pakan yang dicerna oleh ruminansia:
- Protein pakan didegradasi menjadi peptida oleh protease di
dalam rumen. Peptida dikatabolisasi menjadi asam amino
bebas lalu menjadi amonia, asam lemak dan Co . 2
- Produk degradasi yang terbentuk dalam rumen, terutama
amonia, digunakan oleh mikroba bersama-sumber energy
untuk mensintesis protein dan bahan-bahan sel mikroba seperti
bahan sel yang mengandung N dan asam nukleat.
- Bagian amonia bebas akan diserap masuk ke pembuluh darah
hewan dan ditransformasikan menjadi urea di dalam liver.
Sebagian besarnya tidak dapat digunakan oleh hewan dan
diekresikan ke dalam urin.
Sel-sel mikroba (bakteri dan protozoa) mengandung protein
sebagai komponen utama, bersama protein pakan melalui omasum
dan abomasum dan usus halus. Sel-sel pakan yang dicerna
mengandung protein 70-80%, 30-40% adalah protein kurang larut.
Protein hijauan dicerna dalam rumen sebesar 30-80%. Jumlah ini
tergantung kepada waktu tinggal di dalam rumen dan tingkat
fisiologi hewan142

pemberian makan.
Pencernaan dan penyerapan mikroba dan protein pakan terjadi
di usus halus hewan (ruminan dan monogastrik) oleh protease. Asam
amino esensial bagi semua jenis hewan. Komposisi asam-asam
amino yang mencapai usus akan sangat tergantung kepada jenis
protein, kuantitas dan kualitas sumber protein pensuplai. Hewan
ruminan tergantung pada protein mikroba dan protein pakan yang
lolos dari pencernaan dalam rumen untuk mensuplai asam amino
esensial.
e. Degradasi Protein di Dalam Rumen
Degradasi protein di dalam rumen dilakukan oleh mikroba
(bakteri, protozoa, fungi). Sekitar 40% bakteri rumen memilik
aktivitas proteolitik. Bakteri ini memiliki emzim prptease yang terikat
pada permukaan sel dan siap kontak dengan subtrat/pakan. Selain
bakteri, protozoa juga memiliki kemampuan sebagai protease
intraseluler, sehingga protozoa juga berperan dalam degradasi protein
di dalam rumen. Efektivitas aktivitas mikroba dalam rumen
dibutuhkan kondisi yang optimal, misalnya pH 5-6 dengan o
temperatur sekitar 39 C.
Berbeda halnya dengan perombakan karbohidrat oleh mikroba
yang lebih banyak menghasilkan VFA rantai pendek, proses
deaminasi asam amino membentuk rantai cabang VFA seperti asam-
asam isobutirat, isovalerat, isoleusin, 2-metilbutirat. Rantai-rantai
cabang tersebut adalah faktor pertumbuhan untuk mikroba dan
merupakan materi awal untuk membentuk cabang VFA rantai
panjang yang merupakan karakteristik mikroba. Sebagian kecil VFA
143metabolisme protein

berasal dari degradasi asam amino. Mikroba mendegradasi asam
amino seperti sistein, asam glutamat dan serin menjadi asam piruvat
akan ditransformasi lanjut menjadi asetat, propionat dan butirat.
Amonia hasil perombakan asam amino adalah sumber nutrien
bagi bakteri. Bakteri ini akan menggunakan amonia bersama dengan
karbohidrat mudah larut untuk membentuk asam amino yang
dibutuhkan dalam rangka memenuhi kebutuhan proteinnya sendiri.
Sebanyak 50-80% N mikroba berasal dari amonia rumen, sedangkan
30% protein berasal dari sumber selain amonia seperti peptida dan
asam-asam amino.
Protozoa tidak dapat menggunakan amonia. Nitrogen diperoleh
dari bakteri yang dicerna melalui peran enzim protease intraseluler.
Sumber fosfat dan sulfur perlu disuplai dalam ransum untuk sintesis
protein. Fosfat untuk sintesis asam nukleat, sedang sulfur untuk
sintesis metionin dan sistein pada protein mikroba.
3.7 Jalur Metabolisme Amonia
Protein pakan akan masuk ke dalam aliran darah sebagai asam
amino bebas. Jadi asam-asam amino tersebut akan mengikuti 3 jalur
metabolisme yaitu: 1) mereka akan digunakan untuk sintesis protein
mikroba rumen; 2) mereka akan menjadi prekursor bagi
Tabel 7. Spesifikasi zat penghasil energi bagi asam amino
fisiologi hewan144

pembentukan senyawa-senyawa yang mengandung nitrogen seperti
asam-asam nukleat, keratin, kholin, dan tiroksin; atau 3) mereka akan
didegradasi, nitrogen akan diekskresikan sebagai urea dan kerangka
karbon masuk ke metabolisme energi. Beberapa metoda untuk
mengurangi efek fermentasi bagi protein pakan. Pemanasan akan
mengurangi kelarutan protein, sehingga akan menurunkan efek
fermentasi dalam rumen. Formaldehid juga akan mengurangi
kelarutan protein, melalui pembentukan kompleks antara aldehida
dan grup amino bebas dari protein seperti yang terjadi pada reaksi
Maillard. Formaldehida berlebihan akan menghambat aktifitas
selulolitik dari mikroba rumen.
Enkapsulasi protein dengan bahan yang tidak larut dalam
rumen namun larut dalam abomasum/usus halus. Contohnya:
enkapsulasi metionin dengan lemak terhidrogenasi, yang akan pecah
dengan adanya lipase dan empedu di duodenum. Untuk menghasilkan
energi yang berasal dari protein pada ruminansia, beberapa pintu
masuk yang dapat dilalui oleh protein adalah: piruvat, asetil CoA, -
ketoglutarat dan suksinil CoA.
3.8 SOAL SOAL LATIHAN
1. Sebutkan macam-macam protein yang terdapat di dalam sel
tubuh !
2. Protein tersusun atas asam-asam amino tertentu. Berdasarkan
sumber dan fungsinya di dalam tubuh, asam amino dibagi
menjadi asam amino esensial dan asam amino non- esensial.
Sebutkan macam-macam asam amino yang termasuk ke dalam
2 kelompok tersebut !
145metabolisme protein

3. Sebutkan enzim-enzim yang terlibat proses katabolisme dan
anabolisme protein !
4. Dalam kondisi tubuh mengalami defisiensi ATP, protein
mempunyai fungsi sebagai sumber energi. Jelaskan proses
dalam tubuh berkaitan dengan fungsi protein tersebut !
5. Jelaskan tahapan proses dalam siklus glukosa-alanin !
6. Jelaskan perbedaan proses deaminasi dan transaminasi, berikan
contoh !
7. Sebutkan enzim-enzim yang terlibat dalam siklus urea, baik
yang bekerja pada sitosol maupun mitokondria pada sel hati!
8. Sebutkan beberapa sifat asam amino berdasarkan katabolisme
kerangka karbon yang dimiliki !
9. Dengan mengukur jumlah nitrogen yang masuk dan keluar,
kondisi metabolisme protein di dalam tubuh dapat
diperkirakan. Mengapa demikian ?
10. Apa yang terjadi dengan protein pada sel-sel otot (otot rangka)
apabila tubuh dalam kondisi kelaparan ?
fisiologi hewan146

4.1 Pengertian Siklus Kreb
Siklus Kreb atau asam trikarboksilat bertanggung jawab
terhadap 32 ATP yang dibentuk dari oksidasi bahan baku
metabolisme. Jalur untuk oksidasi asam-asam lemak, glukosa, asam-
asam amino, asetat dan badan-badan keton semuanya menghasilkan
asetil KoA, yang merupakan substrat untuk siklus asam
trikarboksilat. Ketika gugus asetil berkarbon dua diaktifkan melalui
proses oksidasi menjadi 2 molekul CO , energi disimpan sebagai 2
NADH, FAD(2H) dan GTP. NADH dan FAD(2H) secara bergantian
mendonasikan elektron-elektron ke O melalui rantai transport 2
elektron dengan menghasilkan ATP dari oksidasi phosporilasi.
Dengan demikian siklus asam trikarboksilat merupakan pusat
pembentukan energi dari respirasi selular.
Dalam siklus trikarboksilat, oksidasi dekarboksilasi α-
ketoglutarat dikatalisis oleh banyak subunit komplek α-ketoglutarat
dehidrogenase yang mengandung koenzim thiamin -pirofosfat, lipoat
SIKLUS KREB
4

dan FAD. Selain itu juga terdapat enzim komplek lain yaitu komplek
piruvat dehidrogenase (PDC) yang mengkatalisis oksidasi piruvat
menjadi asetil KoA, yang merupakan penghubung antara jalur
glikolisis dengan siklus asam trikarboksilat/tricarboxilicacid (TCA).
Gugus asetil berkarbon dua adalah sumber terakhir elektron +
yang dipindahkan ke NAD dan FAD, selain itu juga sebagai sumber
karbon yang berasal dari molekul CO yang dihasilkan. Oksaloasetat 2
digunakan dan dibentuk dalam setiap putaran siklus trikarboksilat.
Bagaimanapun, ketika sel menggunakan senyawa-senyawa
intermidiet pada siklus asam trikarboksilat untuk reaksi biosintesis,
karbon-karbon pada oksaloasetat harus diganti oleh reaksi
anaplerotik (pengisian kembali), seperti reaksi piruvat karboksilase.
Siklus asam trikarboksilat terjadi dalam mitokondria, dimana tahapan
proses yang terjadi dikoordinasi secara ketat oleh laju rantai transport
elektron dan oksidasi phosporilasi melalui regulasi umpan balik yang
merefleksikan tingkat kebutuhan terhadap ATP. Laju siklus asam
trikarboksilat ditingkatkan ketika ketersediaan ATP di dalam sel
meningkat melalui respon beberapa enzim untuk tingkat ADP, rasio +
NADH/NAD , laju oksidasi FAD(2H) atau konsentrasi ion kalsium 2+
(Ca ). Sebagai contoh, isositrat dehidrogenase adalah enzim
allosterik yang diaktifkan oleh ADP.
Ada dua konsekuensi umum terjadinya gangguan fungsional
pada siklus asam trikarboksilat yaitu: ketidakmampuan untuk
membentuk ATP dari oksidasi bahan baku metabolisme dan
terjadinya akumulasi prekursor siklus asam trikarboksilat (TCA).
Sebagai contoh penghambatan oksidasi piruvat dalam TCA akan
menyebabkan terjadinya proses reduksi piruvat menjadi laktat yang
fisiologi hewan148

dapat menyebabkan asidosis laktat (lactic acidosis). Kondisi yang
paling umum mengarah pada terjadinya gangguan fungsi siklus TCA
adalah berkurangnya oksigen secara relatif untuk menerima elektron
dalam rantai trasport elektron.
Resistensi Glukosa
Resistensi glukosa pada penderita diabetes diawali oleh suatu
kondisi kadar gula dalam darah yang tinggi, produksi insulin yang
tinggi dan produksi ATP yang tinggi. Selanjutnya secara perlahan
produksi ATP yang berlebih akan menyebabkan pemblokiran saluran
ion kalsium membran sel yang berakibat pada konsentrasi kadar
kalsium dalam cairan sel yang tinggi. Konsentrasi kalsium yang
tinggi di dalam sel akan menyebabkan terakumulasinya kalsium
sehingga dapat mengganggu metabolisme sel melalui inaktivasi
enzim melalui jalur sistem cascade karena kalsium mempunyai peran
sebagai second messenger. Sehingga substrat metabolik di dalam sel
akan semakin tinggi dan akhirnya akan mengganggu metabolisme.
Apabila jumlah ATP yang dihasilkan dalam siklus Kreb berlebih akan
menyebabkan piruvat karboksilase terblokir sehingga jalur
metabolisme glukosa, lemak dan protein akan terhenti. Terhentinya
jalur metabolisme tersebut akan menyebabkan glukosa dalam cairan
sel jenuh dan menyebabkan terhentinya transport glukosa dan
muncullah kondisi terjadinya resistensi glukosa. Selain itu
kedudukan reseptor dalam membran sel akan beralih orientasi dengan
mengikat komponen lain selain insulin karena komponen tersebut
mempunyai struktur yang mirip dengan insulin, seperti reseptor
membentuk ikatan dengan lipoprotein maupun glikoprotein. Dua
117siklus kreb

kondisi tersebut pada akhirnya menyebabkan terjadinya resistensi sel
terhadap insulin.
Pada kasus diabetes melitus tipe dua, resistensi insulin akan
menyebabkan pemenuhan energi beralih orientasi ke lemak. Lemak
akan mengalami metabolisasi menghasilkan asam-asam lemak dan
gliserol dan selanjutnya menjadi asetil KoA. Metabolisme lemak
pada penderita diabetes akan menyebabkan terbentuknya banyak
badan-badan keton hasil perombakan asetil KoA menjadi asetoasetat
(badan-badan keton) sehingga menyebabkan kondisi ketoasidosis
yang menyebabkan inaktivasi enzim-enzim terutama piruvat
karboksilase, sehingga piruvat tidak dapat masuk ke dalam siklus
Kreb. Kondisi ini menyebabkan pembentukan ATP menjadi
terganggu.
Pemberian protein pada penderita diabetes melitus juga tidak
akan menyelesaikan masalah. Protein dalam jaringan otot akan
dimetabolisasi dan melalui proses glukoneogenesis akan diubah
menjadi asam-asam amino, mengalami transaminasi dan deaminasi
menjadi bentuk senyawa-senyawa intermidiet di dalam sel. Namun
demikian terakumulasinya banyak senyawa intermidiet dalam siklus
Kreb hasil deaminasi ini akan menyebabkan kondisi pemblokiran
piruvat karboksilase yang menyebabkan kegagalam perubahan
piruvat menjadi asetil KoA. Rendahnya asetil KoA pada akhirnya
akan menyebabkan produksi ATP terganggu. Selain itu
terakumulasinya asam amino dalam sel dalam jumlah yang berlebih
akan menyebabkan enzim-enzim yang berperan dalam siklus urea
mengalami inaktivasi yang menyebabkan keberadaan amonia
semakin tinggi. Kondisi lain juga akan muncul yaitu terbentuknya
fisiologi hewan150

badan-badan keton. Kondisi pada akhirnya akan mempercepat
terjadinya kasus diabetes/privalensi terkena diabetes yang tinggi pada
pasien. Perubahan derajad keasaman yang disebabkan oleh asidosis,
alkalosis, terakumulasi prekursor metabolit dan lain-lain akan
menyebabkan inaktivasi enzim atau berubahnya orientasi enzim
terhadap substrat.
Reaksi enzim : E + S ES E + produk
Pemberian obat pada penderita diabetes harus diamati secara
terus menerus persatuan waktu untuk melihat trend kenaikan dan
penurunan kembali kadar glukosa darah. Orang diabetes mempunyai
tanda-tanda cepat haus, frekuensi buang urin tinggi, mempunyai
nafsu makan yang tinggi, cepat mengalami kelelahan, kekuatan otot
menjadi lemah, mudah mengantuk setelah makan, anggota tubuh
tremor dan cepat pingsan. Tanda yang lain adalah terbentuknya
badan-badan keton dalam darah, kadar gula darah tinggi dan
terdapatnya glukosa dalam urin. Kadar elektrolit, BUN dan kreatinin
yang abnormal juga merupakan tanda-tanda diabetes. Orang yang
mempunyai kakek nenek terkenan diabetes, saudara perempuan
diabetes, cenderung mempunyai privalensi yang tinggi terkena
diabetes. Demikian juga orang yang obesitas juga mempunyai
privalensi yang sama. Orang gemuk tidak efisien dalam mengambil
oksigen dan cenderung terengah-engah. Orang gemuk butuh 9 kali
dalam pengambilan oksigen untuk kepentingan metabolisme lemak
dalam rangka mencukupi kebutuhan ATP, sedangkan pada orang
normal kebutuhan ATP lebih banyak dicukupi dari glukosa dengan
151siklus kreb

membutuhkan pengambilan oksigen lebih efisien yaitu 3 kali untuk
metabolisme glukosa.
4.2 Reaksi Siklus Kreb
Dalam siklus TCA, gugus asetil berkarbon-2 (asetoasetat) pada
asetil KoA dioksidasi menjadi 2 molekul CO . Fungsi siklus TCA 2
adalah untuk menyediakan energi dari oksidasi ini, yang secara
prinsip menyesuaikan dengan adanya proses rantai transport elektron +
dari senyawa intermidiet dalam siklus TCA menjadi NAD dan FAD.
Delapan elektron akan didonasikan oleh gugus asetil dan akhirnya
akan terbentuk tiga molekul NADH dan satu molekul FAD. Secara
bertahap ATP akan dibentuk dari oksidasi phosporilasi ketika NADH
dan FAD(2H) mendonasikan elektron-elektron ini ke oksigen melalui
rantai transport elektron. Awalnya gugus asetil akan digabungkan ke
dalam sitrat yang merupakan senyawa intermidiet pada TCA. Sitrat
secara progresif melalui siklus TCA akan diubah menjadi
oksaloasetat. Sitrat melalui reaksi secara bertahap akan dioksidasi
oleh 4 enzim dehidrogenase, yaitu isositrat dehidrogenase, α-
ketoglutarat dehidrogenase, suksinat dehidrogense, dan malat +
dehidrogenase yang memindahkan elektron ke NAD atau FAD.
Isomerase akonitase menyusun kembali elektron-elektron dalam
isositrat, kadang-kadang membentuk isositrat untuk memfasilitasi +
transfer elektron ke NAD . Meskipun tidak ada oksigen yang masuk
ke siklus TCA, dua molekul CO yang dihasilkan mempunyai lebih 2
banyak atom oksigen daripada gugus asetil. Atom oksigen ini pada
akhirnya diturunkan oleh gugus karbonil pada asetil KoA, diturunkan
oleh dua molekul air yang ditambah oleh fumarase/sitrat sintase serta
fisiologi hewan152

2-PO yang ditambah GDP.4
Keseluruhan energi yang dihasilkan dari siklus TCA adalah 3
NADH, 1 FAD(2H) dan 1 GTP. Ikatan phospat energi tinggi pada
GTP merupakan hasil phosporilasi substrat yang dikatalisis oleh
enzim suksinat tiokinase (suksinil KoA sintetase). Ketika NADH dan
FAD(2H) melakukan reoksidasi dalam rantai transport elektron,
kurang lebih 2,5 ATP dibentuk untuk setiap NADH dan 1,5 ATP untuk
FAD(2H). Konsekuensinya, seluruh energi bersih dari siklus TCA
dan oksidasi phosporilasi adalah sekitar 10 ikatan phospat energi
tinggi untuk setiap gugus asetil yang dioksidasi.
4.3 Pembentukan dan Oksidasi pada Isositrat
Siklus Kreb dimulai dengan terjadinya kondensasi pada gugus
asetil dan oksaloasetat. Gugus asetil dan okaloasetat yang telah
diaktifkan akan membentuk senyawa intermidiet sitrat berkarbon-6,
dengan reaksi yang dikatalisis oleh enzim sitrat sintase. Karena
oksaloasetat dibentuk sekali untuk setiap putaran siklus TCA,
oksaloasetat tidak dipersiapkan sebagai substrat dalam siklus TCA
atau sebagai sumber elektron atau karbon.
Tahap selanjutnya dalam siklus TCA adalah gugus hidroksil
(alkohol) pada sitrat akan dipindahkan ke karbon yang berdekatan
dan selanjutnya akan dioksidasi untuk membentuk gugus keto.
Isomerisasi sitrat akan membentuk isositrat yang dikatalisis oleh
enzim akonitase. Enzim isositrat dehidrogenase mengkatalisis
oksidasi gugus alkohol dan secara bertahap memutus gugus karboksil
untuk melepaskan CO (oksidasi dekarboksilasi).2
153siklus kreb

4.4 Pembentukan dan Penguraian α-Ketoglutarat menjadi
Suksinil KoA
Tahapan selanjutya dalam siklus TCA adalah oksidasi
dekarboksilasi α-ketoglutarat menjadi suksinil KoA yang dikatalisis
oleh komplek α-ketoglutarat dehidrogenase. Komplek α-ketoglutarat
dehidrogenase mengandung koenzim thiamin pirofosfat, asam lipoik
dan FAD. Dalam reaksi ini, satu gugus karboksil pada α-ketoglutarat
dilepaskan sebagai CO , dan gugus keto yang berdekatan dioksidasi 2
menjadi asam, dan berkombinasi dengan KoASH untuk membentuk
suksinil KoA. Energi dari reaksi ini disimpan dan secara prinsip
mempunyai peran dalam reduksi pada NADH, dengan sumber energi
yang berasal dari ikatan thioester energi tinggi pada suksinil KoA.
4.5 Pembentukan GTP
Energi dari ikatan thioester suksinil KoA digunakan untuk
membentuk GTP dari GDP dan Pi dalam reaksi yang dikatalisis oleh
suksinat thiokinase. Suksinat thiokinase juga dikenal dengan sebutan
suksinil KoA sintetase. Dua nama ini mempunyai arti enzim yang
berperan dalam reaksi umpan balik secara langsung, yaitu konversi
suksinat menjadi thioester suksinil KoA, yang menyediakan energi
dari GTP. Enzim sintase, misalnya sitrat sintase berbeda dari sintetase
yang memutus ikatan phospat energi tinggi pada ATP, UTP, CTP atau
GTP, sedangkan sintase tidak. Reaksi ini adalah contoh phosporilasi
tingkat substrat. Phosporilasi tingkat substrat merupakan
pembentukan ikatan fosfat berenergi tinggi dimana awalnya tidak
membutuhkan kehadiran oksigen (dengan kata lain bukan oksidasi
phosporilasi). Ikatan fosfat energi tinggi pada GTP mempunyai
fisiologi hewan154

energi yang ekuivalen dengan ATP dan dapat digunakan secara
langsung untuk reaksi yang membutuhkan energi, seperti síntesis
protein.
4.6 Oksidasi Suksinat menjadi Oksaloasetat
Tahap selanjutnya dalam siklus TCA, dua karbon akan
menyediakan elektron dan dilepaskan sebagai CO . Dua pasang 2
+elektron ini akan dipindahkan menuju ke 2NAD , dan akan dihasilkan
satu GTP. Tambahan dua pasang elektron berasal dari asetil KoA yang
tersisa dalam siklus TCA sebagai bagian suksinat dan selanjutnya +
akan memindahkan dua pasang elektron ke FAD dan NAD dan
menambahkan H O, karena itu oksaloasetat terbentuk kembali.2
Urutan reaksi perubahan suksinat menjadi oksaloasetat dimulai
dengan oksidasi suksinat menjadi fumarat. Elektron tunggal
ditransfer dari dua gugus metilen (-CH -) yang berdekatan pada 2
suksinat ke FAD yang berikatan dengan suksinat dehidrogenase, dan
membentuk ikatan ganda pada fumarat. Dari reduksi FAD yang
berikatan dengan enzim (enzym bond FAD), elektron – elektron akan -
diloloskan ke dalam rantai transport elektron. Gugus OH dan proton
dari H O ditambahkan pada ikatan ganda pada fumarat, sehingga 2
merubah fumarat menjadi malat. Dalam reaksi akhir TCA, gugus
alkohol pada malat dioksidasi menjadi gugus keto melalui donasi +
elektron – elektron ke NAD . Dengan adanya pembentukan kembali
oksaloasetat, siklus TCA menjadi lengkap atau sempurna; dimana
ikatan kimia berenergi, karbon dan elektron – elektron didonasikan
oleh gugus asetil untuk dikonversi menjadi CO , NADH, FAD(2H), 2
GTP dan panas.
155siklus kreb

4.7 Koenzim pada Siklus Kreb
Enzim-enzim yang bekerja pada siklus TCA bergantung pada
peran penting koenzim untuk dapat melakukan fungsi katalitiknya.
Isositrat dehidrogenase dan malat dehidrogenase menggunakan +
NAD sebagai koenzim dan suksinat dehidrogenase menggunakan
FAD. Sitrat sintase mengkatalisis reaksi yang menggunakan substrat
turunan KoA, misalnya asetil KoA. Komplek α-ketoglutarat
dehidrogenase menggunakan thiamin pirophospat, lipoat dan FAD +
sebagai koenzim dan NAD serta KoASH sebagai substrat. Setiap
koenzim-koenzim ini mempunyai bentuk dan struktur yang unik dan
masing-masing mempunyai peran penting dalam siklus TCA.
+FAD dan NAD +
FAD dan NAD adalah dua koenzim yang berperan sebagai
penerima atau akseptor elektron. Muncul pertanyaan, mengapa FAD +
digunakan dalam beberapa reaksi dan NAD dalam reaksi yang lain?
Berdasarkan bentuk dan struktur uniknya kemampuan FAD dan +
NAD mempunyai aksi sebagai akseptor elektron namun bekerja
pada berbagai tipe reaksi yang berbeda serta mempunyai perbedaan
peran fisiologis yang dimainkan dalam sel. FAD mampu menerima .
elektron tunggal (H ) dan membentuk senyawa intermidiet
berelektron tunggal setengah reduksi. Dengan demikian FAD
berpartisipasi dalam reaksi yang melibatkan transfer elektron tunggal
secara bebas dari dua atom yang berbeda, seperti yang terjadi dalam
pembentukan ikatan ganda (misalnya: suksinat menjadi fumarat) dan
pembentukan ikatan disulfida (misalnya lipoat menjadi lipoat
disulfida dalam reaksi α-ketoglutarat dehidrogenase. Sebaliknya,
fisiologi hewan156

+ -NAD menerima sepasang elektron dalam bentuk ion hidrida (H ),
yang menarik karbon pada sisi yang berlawanan yang bermuatan
positif pada cincin piridin. Contoh reaksi ini adalah oksidasi alkohol
menjadi keton oleh malat dehidrogenase dan isositrat dehidrogenase.
Cincin nikotinamid menerima ion hidrida dari ikatan C-H, dimana
hidrogen alkoholik dilepaskan ke dalam medium dalam bentuk +
proton yang bermuatan positif (H ).
Radikal bebas yang berupa bentuk elektron tunggal pada FAD
sangat reaktif dan FADH dapat kehilangan elektronnya melalui
pendedahan dengan air atau inisiasi pada reaksi berantai. Sebagai
konsekuensinya FAD harus melakukan kerja secara ketat dengan
melakukan ikatan kovalen dengan enzim, sementara pada sisi lain
FAD menerima dan menstransfer elektron-elektron menuju gugus
lain yang terikat pada enzim. Karena FAD berinteraksi dengan
beberapa gugus fungsional pada rantai samping asam amino dalam 0
sisi aktifnya, E untuk FAD yang berikatan dengan enzim mempunyai +
variasi lebih besar dan dapat lebih besar atau lebih rendah dari NAD . +
Sebaliknya NAD dan NADH lebih berperan sebagai substrat dan
produk daripada sebagai koenzim.
NADH mempunyai peranan sebagai regulator dalam
menyeimbangkan energi metabolisme dan kemampuan ini tidak
dimiliki oleh FAD(2H) karena FAD(2H) masih menyisakan +
perlekatan pada enzimnya. NAD bebas berikatan dengan
dehidrogenase dan direduksi menjadi NADH. NADH dapat
berikatan dan menghambat dehidrogenase yang berbeda.
Konsekuensinya, enzim-enzim oksidatif dikendalikan oleh rasio +
NADH/NAD dan tidak membentuk NADH yang lebih cepat dari
157siklus kreb

NADH hasil reoksidasi dalam rantai transport elektron. Regulasi
pada siklus TCA dan jalur-jalur lain pada oksidasi bahan baku +
metabolisme oleh rasio NADH/NAD adalah bagian mekanisme
untuk mengkoordinasi laju oksidasi bahan baku metabolisme untuk
mengatur laju ketersediaan ATP.
Peran KoA dalam Siklus TCA
KoASH adalah koenzim asilasi yang berperan dalam reaksi
melalui pembentukan ikatan thioester antara sulfur (S) pada KoASH
dan gugus asil (seperti asetil KoA, suksinil KoA). Perbedaan ikatan
thioester dari tipe ikatan ester oksigen adalah adanya atom S, bukan
O, karena ikatan ini tidak membagi elektron-elektronnya dan ikatan
ini berperan dalam pembentukan resonansi. Konsekuensinya bentuk
ikatan thioester dengan struktur kimiawi yang mengandung sulfur
adalah karbon karbonil, α-karbon dan β-karbon pada gugus asil dalam
ikatan thioester KoA. Karbon karbonil, α-karbon dan β-karbon pada
gugus asil dalam ikatan thioester KoA dapat diaktifkan agar masing-
masing dapat berperan dalam berbagai tipe-tipe reaksi yang berbeda
(reaksi sitrat sintase, gugus metil α-karbon diaktifkan untuk
kondensasi dengan oksaloasetat). Bentuk kondensasi lain yang
terjadi pada ikatan thioester yaitu ikatan berenergi tinggi yang
mempunyai sejumlah besar reaksi negatif pada saat reaksi hidrolisis
(kurang lebih -13 kkal/mol).
Energi hasil pemutusan ikatan thioester berenergi tinggi pada
suksinil KoA dan asetil KoA digunakan dalam lintasan yang berbeda
pada siklus TCA. Ketika ikatan suksinil KoA diputus oleh suksinat
thiokinase, energi digunakan secara langsung untuk mengaktifkan
fisiologi hewan158

enzim yang mengikat phospat yang dipindahkan ke GDP. Sebaliknya,
ketika ikatan thioester pada asetil KoA diputus dalam reaksi yang
melibatkan sitrat sintase, energi yang dilepaskan memberikan
sejumlah besar reaksi negatif -7,7 kkal/mol. Sejumlah besar reaksi
negatif untuk pembentukan sitrat membantu berjalannya reaksi-
reaksi yang terjadi dalam siklus TCA.
Komplek α-Asamketo Dehidrogenase
Komplek α-ketoglutarat dehidrogenase merupakan salah satu
dari tiga anggota famili komplek α-asamketo dehidrogenase. Enzim
lain yang merupakan anggota dari famili ini adalah komplek piruvat
dehidrogenase, dan komplek rantai bercabang asam amino α-
asamketo dehidrogenase. Setiap komplek enzim-enzim ini adalah
spesifik untuk struktur α-asamketo yang berbeda. Rangkaian reaksi
dalam siklus TCA yang dikatalisis oleh komplek-komplek enzim,
seperti α-asamketo adalah dekarboksilasi (yaitu melepaskan gugus
karboksil dalam bentuk CO ). Gugus keto dioksidasi menjadi asam 2
karboksilik dan kemudian dikombinasikan dengan KoASH untuk
membentuk thioester asil KoA (suksinil KoA).
Komplek α-asamketo dehidrogenase adalah komplek enzim
besar yang disusun oleh banyak subunit-subunit (multipel subunits).
Enzim-enzim yang merupakan komplek α-asamketo dehidrogenase
terdiri enzim E1, E2 dan E3. E1 adalah α-asamketo dekarboksilase
yang terdiri dari thiamin pirophospat (TPP), enzim ini memutuskan
gugus karboksil pada α-asamketo. E2 adalah transasilase yang terdiri
lipoat. Enzim ini menstransfer bagian asil pada α-asamketo dari
thiamin menjadi KoASH. E3 adalah dehidrolipoil dehidrogenase
159siklus kreb

yang terdiri FAD, enzim ini menstranfer elektron-elektron dari lipoat +
tereduksi ke NAD .
Thiamin Pirophospat dalam Komplek α-Ketoglutarat
Dehidrogenase
Thiamin pirophospat disintesis dari thiamin oleh penambahan
pirophospat. Gugus pirophospat berikatan dengan magnesium yang
berikatan dengan rantai samping asam amino pada enzim. Fungsi
umum thiamin pirophospat adalah memutuskan ikatan karbon-
karbon yang selanjutnya menjadi gugus keto. Dalam α-ketoglutarat,
piruvat, rantai bercabang komplek α-asam keto dehidrogenase,
karbon fungsional pada cincin thiazol akan membentuk ikatan
kovalen dengan karbon α-keto dan gugus asam karboksilik yang
berdekatan. Thiamin pirophospat juga merupakan koenzim untuk
transketolase dalam jalur pentosa phospat, dimana thiamin
pirophospat akan memutuskan ikatan karbon-karbon selanjutnya
menjadi gugus keto.
Lipoat
Lipoat adalah koenzim yang ditemukan hanya pada komplek-
komplek α-asam keto dehidrogenase. Lipoat disintesis dalam tubuh
manusia dari karbohidrat dan asamasam amino dan tidak
memerlukan prekursor vitamin. Lipoat dilekatkan pada enzim
transasilase melalui gugus karboksilnya, yang berikatan secara
kovalen dengan bagian terminal-NH pada lisin dalam protein. Lipoat 2
terdiri gugus disulfida yang menerima elektron-elektron ketika
lipoat berikatan dengan fragmen asil pada α-ketoglutarat. Lipoat
fisiologi hewan160

dapat bertindak seperti lengan panjang-CH yang fleksibel yang 2
mampu bereaksi dengan enzim dekarboksilase untuk mengambil
fragmen asil dari thiamin dan memindahkan thiamin kepadanya
untuk mengaktifkan tempat yang mengandung ikatan KoASH.
Komplek ini kemudian melanjutkan reaksi dengan dehidrolipoil
dehidrogenase untuk memindahkan elektron-elektron dari gugus
sulfidril lipoil ke FAD.
FAD dan Dehidrolipoil Dehidrogenase
FAD pada dehidrolipoil dehidrogenase menerima elektron-
elektron dari gugus sulfidril lipoil dan menstransfernya untuk +
berikatan dengan NAD . Dengan demikian FAD menerima dan
memindahkan elektron-elektron tanpa meninggalkan tempat
ikatannya pada enzim. Secara langsung reaksi ini merupakan hasil
interaksi antara FAD dengan gugus tertentu pada enzim, yang
mengubah potensial reduksi dan secara keseluruhan melepaskan
energi dari pemutusan dan oksidasi pada α-ketoglutarat.
4.8 Regulasi Pada Siklus Kreb
Oksidasi asetil KoA pada siklus TCA dan penyimpanan
cadangan energi dalam bentuk NADH dan FAD(2H) adalah sangat
penting untuk pembentukan ATP dalam banyak jaringan di dalam
tubuh. Perubahan dalam ketersediaan bahan baku metabolisme, jenis-
jenis bahan baku metabolisme dalam darah atau laju ketersediaan
ATP diregulasi oleh sel-sel dengan memelihara homeostasis ATP
(konsentrasi ATP selalu konstan). Laju pada siklus TCA, untuk semua
jalur oksidasi bahan baku metabolisme, secara prinsip diregulasi dan
161siklus kreb

selalu berkaitan dengan laju rantai transport elektron, yang diregulasi
oleh rasio ATP/ADP dan laju ketersediaan ATP.
Dua messenger utama yang memberi informasi terhadap laju
ketersediaan ATP pada siklus TCA adalah : (a) kedudukan
phosporilasi ATP, yang direfleksikan dalam bentuk ATP atau ADP dan +
(b) kedudukan reduksi pada NAD , yang direfleksikan dalam rasio +
NADH/NAD . Dalam sel terutama mitokondria adalah tempat
keberadaan adenin nukleotida (AMP, ADP dan ATP) dan merupakan +
tempat keberadaan NAD (NAD dan NADH) yang secara relatif
bersifat konstan. Dengan demikian, apabila laju ketersediaan ATP
ditingkatkan akan menghasilkan penurunan sejumlah kecil
konsentrasi ATP dan meningkatnya ADP. Demikian halnya, +
peningkatan oksidasi NAD oleh rantai transport elektron akan
meningkatkan jalur yang menghasilkan NADH. Dalam kondisi
fisiologis dibawah normal, siklus TCA dan jalur oksidatif lain
merespon secara cepat terjadinya peningkatan kebutuhan ATP namun
konsentrasi ATP tidak berubah secara signifikan.
Regulasi Sitrat Sintase
Dalam jalur regulasi umpan balik, tahap pertama pada jalur ini
adalah meregulasi aliran prekursor kedalam jalur alternatif jika
produk tidak dibutuhkan. Sitrat sintase adalah enzim pertama pada
siklus TCA yang merupakan enzim sederhana yang tidak mempunyai
regulator allosterik. Laju sitrat sintase secara prinsip dikendalikan
oleh konsentrasi oksaloasetat, substrat sitrat sintase, dan konsentrasi
sitrat, yang merupakan inhibitor produk yang berkompetisi dengan
oksaloasetat. Keseimbangan oksaloasetat-malat menguntungkan
fisiologi hewan162

malat, sehingga konsentrasi oksaloasetat sangat rendah di dalam +
mitokondria dan dibawah sitrat sintase. Ketika rasio NADH/NAD
menurun, rasio oksaloasetat terhadap malat meningkat. Ketika
isositrat dehidrogenase diaktifkan, konsentrasi sitrat menurun,
dengan demikian mengurangi penghambatan produk pada sintrat
sintase. Peningkatan konsentrasi oksaloasetat dan penurunan sitrat
meregulasi kemampuan merespon sitrat sintase untuk kondisi yang
dimapankan oleh rantai transport elektron dan oksidasi phosporilasi. +
Dalam hati, rasio NADH/NAD membantu menentukan apakah asetil
KoA masuk ke dalam siklus TCA atau pergi masuk ke jalur alternatif
untuk sintesis badan-badan keton.
Regulasi Allosterik pada Isositrat Dehidrogense
Isositrat dehidrogenase merupakan salah satu enzim yang
dipersiapkan untuk pembatasan laju tahapan pada siklus TCA dan
secara allosterik diaktifkan oleh ADP dan dihambat oleh NADH.
Dalam kondisi tanpa kehadiran ADP, enzim secara positif
memperlihatkan kooperatif dengan isositrat yang berikatan pada
salah satu subunitnya, sedangkan subunit yang lain dikonversi
menjadi konformasi yang aktif. Dalam kondisi kehadiran ADP,
semua subunit enzim akan menjadi bentuk konformasi yang aktif dan
enzim isositrat sintase akan melakukan ikatan terhadap substrat lebih
cepat lagi. Konsentrasi isositrat ditemukan dalam matriks
mitokondria, sejumlah kecil perubahan ADP dalam matriks
mitokondria dapat menghasilkan sejumlah besar laju reaksi isositrat
dehidrogenase. Berubahnya sebagian kecil konsentrasi produk, +
NADH, kosubstrat dan NAD juga berpengaruh terhadap laju enzim
163siklus kreb

isositrat dehidrogenase yang lebih besar daripada enzim
nonallosterik.
Regulasi α-Ketoglutarat Dehidrogenase
Komplek α-ketoglutarat dehidrogenase meskipun bukan enzim
allosterik adalah enzim yang dihambat oleh NADH dan suksinil KoA
dan mungkin juga dihambat oleh GTP. Dua enzim α-ketoglutarat
dehidrogenase dan isositrat dehidrogenase merespon secara langsung
untuk merubah konsentrasi relatif ADP dan meningkatkan laju
NADH yang dioksidasi oleh transport elektron. Dua enzim ini juga 2+
diaktifkan oleh ion kalsium (Ca ). Dalam kontraksi otot jantung, dan
mungkin juga jaringan otot lainnya, pelepasan kalsium dari retikulum
sarkoplasmik selama kontraksi otot menyediakan penambahan
aktivasi pada enzim-enzim ini ketika ATP secara cepat dihidrolisasi.
Regulasi Siklus Senyawa Intermidiet TCA
Regulasi pada siklus TCA menyediakan dua fungsi: siklus TCA
menjamin bahwa NADH adalah dibentuk secara cepat dalam jumlah
yang cukup untuk memelihara homeostasis dan NADH meregulasi
konsentrasi senyawa-senyawa intermidiet pada siklus TCA. Sebagai
contoh, dalam hati, menurunnya laju isositrat dehidrogenase akan
meningkatkan konsentrasi sitrat yang menstimulasi pengeluaran
sitrat menuju sitosol.
4.9 Prekursor pada Asetil KoA
Senyawa-senyawa yang masuk ke dalam siklus TCA adalah
senyawa intermidiet yang dapat diubah ke dalam malat atau
fisiologi hewan164

oksaloasetat. Senyawa-senyawa yang masuk seperti asetil KoA
adalah dioksidasi menjadi CO . Senyawa-senyawa yang masuk 2
sebagai senyawa intermidiet dalam siklus TCA mengisi senyawa-
senyawa intermidiet yang digunakan dalam jalur biosintesis, seperti
glukoneogenesis atau sintesis heme, tetapi tidak dapat secara penuh
dioksidasi menjadi Co .2
Sumber-Sumber Asetil KoA
Asetil KoA adalah senyawa utama yang menyediakan
kovergensi untuk jalur utama oksidasi bahan baku metabolisme.
Asetil KoA dibentuk secara langsung dari oksidasi β asam-asam
lemak dan degradasi badan-badan keton β-hidroksibutirat dan
asetoasetat. Asetil KoA juga dibentuk dari asetat, yang berasal dari
diet makanan atau dari oksidasi etanol. Glukosa dan karbohidrat-
karbohidrat lain akan masuk ke jalur glikolisis, yaitu jalur umum
untuk semua sel, dan kemudian akan dioksidasi menjadi asam
piruvat. Piruvat selanjutnya dioksidasi menjadi asetil KoA oleh
komplek piruvat dehidrogenase. Sejumlah asam-asam amino, seperti
leusin dan isoleusin adalah juga dioksidasi menjadi asetil KoA.
Dengan demikian, oksidasi akhir pada asetil KoA menjadi CO dalam 2
siklus TCA adalah tahap terakhir dalam semua jalur utama pada
oksidasi bahan baku metabolisme.
Komplek Piruvat Dehidrogenase
Komplek piruvat dehidrogenase/piruvat dehidrogenase
complex (PDC) akan mengoksidasi piruvat menjadi asetil KoA, yang
merupakan penghubung antara glikolisis dengan siklus TCA. Dalam
165siklus kreb

otak, ketergantungan terhadap oksidasi glukosa menjadi CO untuk 2
mencukupi kebutuhan ATP sangat penting. Regulasi PDC adalah
kehidupan dan PDC adalah enzim yang bertanggung jawab terhadap
soal kematian.
Struktur Komplek Piruvat Dehidrogenase
PDC tergolong ke dalam famili komplek α-asamketo
dehidrogenase, dimana bentuk struktural dan katalitiknya terbagi ke
dalam komplek α–ketoglutarat dehidrogenase dan dicabangkan
dengan rantai komplek α–asam keto dehidrogenase. PDC
mempunyai 3 subunit katalitik yang sama: (1) subunit-subunit piruvat
karboksilase yang berikatan dengan thiamin pirophospat (E1); (2)
subunit-subunit transasetilase yang berikatan dengan lipoat (E2); (3)
subunit-subunit dehidrolipoil dehidrogenase yang berikatan dengan
FAD (E3). Meskipun enzim E1 dan E2 dalam PDC secara relatif
spesifik untuk piruvat, namun kedua enzim tersebut juga seperti
dehidrolipoil dehidrogenase yang juga berperan seperti halnya semua
komplek α-asam dehidrogenase. Hal yang perlu ditambahkan adalah
bahwa selain tiga jenis subunit ini, komplek PDC masih mempunyai
satu tambahan subunit yang lain, yaitu subunit katalitik protein X
yang disebut transasetilase. Setiap komponen fungsional pada
komplek PDC menghadirkan sekian banyak bentuk kopiannya
(seperti komplek PDC bovine jantung mempunyai 30 subunit E1, 60
subunit E2 dan 6 subunit E3 dan X). Enzim E1 adalah tetramer dengan
dua jenis subunit yang berbeda, yaitu α dan β.
fisiologi hewan166

Regulasi Komplek Piruvat Dehidrogenase
Aktivitas komplek piruvat dehidrogenase secara prinsip
dikontrol oleh piruvat dehidrogenase kinase dan reaksi
dephosporilasi yang dikatalisis oleh piruvat dehidrogenase
phospatase yang mengaktifkannya. Kedua enzim tersebut meregulasi
subunit-subunit dalam komplek PDC dan bertindak hanya terhadap
komplek PDC. PDC kinase menstransfer phospat dari ATP ke gugus
serin hidroksil spesifik (ser-OH) pada piruvat dekarboksilase (E1).
PDC phospatase memindahkan gugus-gugus phospat ini melalui
proses hidrolisis. Phosporilasi yang baru saja terjadi pada serin pada
subunit α PDC E1 dapat menurunkan aktivitasnya lebih dari 99%.
PDC kinase hadir dalam bentuk komplek isozim yang spesifik
jaringan yang mempunyai banyak variasi sifat regulasi.
PDC kinase dapat dihambat oleh ADP dan piruvat. Dengan
demikian, ketika ATP secara cepat tersedia akan menghasilkan
peningkatan ADP, atau ketika aktivasi glikolisis terjadi maka akan
segera meningkatkan konsentrasi piruvat. PDC kinase dihambat dan
PDC yang tersisa yang masih aktif (bentuk yang tidak mengalami 2+
phosporilasi). PDC phospatase memerlukan Ca selama kontraksi
cepat yang mengaktifkan phospatase, karena itu meningkatkan
sejumlah aktivitas yang tidak diphosporilasi oleh PDC.
PDC juga diregulasi melalui penghambatan oleh produk PDC
sendiri, seperti asetil KoA dan NADH. Penghambatan ini bersifat
kuat yang meregulasi penghambatan produk karena ikatan mereka
dengan PDC menstimulasi phosporilasi menjadi bentuk yang tidak +
aktif. Substrat-substrat enzim, KoASH dan NAD antagonis dengan
penghambatan produk ini. Dengan demikian ketika semakin besar
167siklus kreb

suplai pada asetil KoA untuk siklus TCA disiapkan untuk menjamin
berjalannya oksidasi asam lemak, asetil KoA dan NADH meningkat
dan menurun secara dramatis ketika terjadi sintesis selanjutnya oleh
PDC. PDC dapat juga secara cepat diaktifkan melalui mekanisme
yang melibatkan insulin yang memainkan peran penting dalam sel-sel
adiposit. Dalam banyak jaringan, insulin mungkin secara perlahan-
lahan dalam waktu yang lama akan meningkatkan sejumlah komplek
piruvat dehidrogenase. Laju pada jalur oksidasi bahan baku
metabolisme yang memberi suplai substart ke dalam siklus TCA
adalah juga ditingkatkan ketika ketersediaan ATP menjadi
meningkat. Insulin, hormon-hormon lain dan kontrol diet mempunyai
kemampuan dalam menyediakan bahan baku metabolisme untuk
jalur oksidasi ini.
4.10 Senyawa Intermidiet pada Siklus Kreb dan Reaksi
Anaplerotik
Senyawa Intermidiet adalah Prekursor untuk Jalur Biosintesis
Senyawa-senyawa intermidiet pada siklus TCA menyediakan
prekursor-prekursor untuk bermacam-macam jalur yang berbeda dan
terjadi dalam beberapa jenis sel yang berbeda. Ini penting terutama
peran pusat metabolik pada hati. Siklus TCA dalam hati sering
disebut ”open cycle” karena adanya pengeluaran yang tinggi
senyawa-senyawa intermidiet. Selama puasa, prekursor-prekursor
glukoneogenik akan diubah menjadi malat yang meninggalkan
mitokondria untuk glukoneogenesis di sitosol. Hati juga
menggunakan senyawa-senyawa intermidiet pada siklus TCA untuk
sintesis kerangka karbon pada asam-asam amino. Suksinil KoA akan
fisiologi hewan168

dipindahkan dari siklus TCA untuk membentuk heme dalam sel-sel
hati dan sumsum tulang. Dalam otak, α-ketoglutarat akan diubah
menjadi glutamat dan kemudian menjadi γ-asam aminobutirat
(GABA) yang merupakan neurotransmiter. Dalam otot rangka, α-
ketoglutarat diubah menjadi glutamin yang kemudian ditransportasi
melalui darah menuju jaringan lainnya.
Reaksi Anaplerotik
Pemindahan banyak senyawa-senyawa intermidiet dari siklus
TCA memindahkan 4 karbon yang digunakan untuk membentuk
oksaloasetat selama setiap putaran dalam siklus TCA. Dengan
kekosongan aksaloasetat, oksaloasetat tidak mungkin secara terus
menerus disediakan dari oksidasi asetil KoA. Agar siklus TCA tetap
dapat berjalan, sel mempunyai kemampuan dalam mensuplai
senyawa-senyawa intermidiet dengan karbon empat dalam jumlah
yang cukup dari degradasi karbohidrat atau asam-asam amino
tertentu untuk mengkompensasi untuk laju pemindahan oksaloasetat.
Jalur atau reaksi yang mengosongkan senyawa-senyawa intermidiet
pada siklus TCA diartikan sebagai anaplerotik dan kemudian
melakukan pengisian kembali (”filling up).
Piruvat Karboksilase adalah Enzim Anaplerotik Utama
Piruvat karboksilase adalah salah satu enzim anaplerotik utama
dalam sel. Enzim ini mengkatalisis penambahan CO pada piruvat 2
untuk membentuk oksaloasetat. Seperti banyak enzim karboksilase,
piruvat karboksilase mengandung biotin yang membentuk ikatan
kovalen intermidiet dengan CO dalam reaksi yang memerlukan ATP 2
169siklus kreb

2+dan Mg . CO2 yang diaktifkan kemudian ditransfer menuju piruvat
untuk membentuk gugus karboksil pada oksaloasetat. Piruvat
karboksilase banyak ditemukan pada banyak jaringan, seperti hati,
otak, adiposit, fibroblast dimana fungsinya adalah anaplerotik. Enzim
ini konsentrasinya tinggi di dalam sel hati dan kortek ginjal, dimana
keberadaannya secara kontinue memindahkan oksaloasetat dan malat
dari siklus TCA untuk masuk ke jalur glukoneogenik.
Piruvat karboksilase diaktifkan oleh asetil KoA dan dihambat
oleh turunan-turunan asetil KoA dalam konsentrasi yang tinggi.
Konsentrasi oksaloasetat akan dihabiskan melalui pengeluaran
senyawa-senyawa intermidiet dalam siklus TCA, laju reaksi sitrat
sintase yang menurun dan kenaikan konsentrasi asetil KoA. Asetil
KoA kemudian mengaktifkan piruvat karboksilase untuk sintesis
oksaloasetat dalam jumlah yang banyak.
Degradasi Asam Amino Membentuk Senyawa Intermidiet dalam
Siklus Kreb
Jalur untuk oksidasi beberapa asam-asam amino mengubah
kerangka karbonnya ke dalam senyawa intermidiet karbon-4 dan
karbon-5 pada siklus TCA yang dapat membentuk kembali
oksaloasetat. Karbon-karbon alanin dan serin dapat masuk melalui
piruvat karboksilase. Dalam semua mitokondria jaringan (kecuali
pada hati), oksidasi pada dua rantai asam amino bercabang, seperti
isoleusin dan valin menjadi suksinil KoA membentuk rute
anaplerotik utama. Dalam hati senyawa-senyawa lain membentuk
propionil KoA (misalnya methionin, timin dan rantai panjang yang
ganjil atau rantai asam-asam lemak bercabang) juga masuk dalam
fisiologi hewan170

siklus TCA sebagai suksinil KoA. Dalam banyak jaringan, glutamin
diambil dari darah, diubah menjadi glutamat dan kemudian
dioksidasi menjadi α-ketoglutarat membentuk rute anaplerotik utama
lainnya. Tetapi siklus TCA tidak dapat melakukan suplai kembali
dengan senyawa-senyawa intermidiet oleh oksidai asam-asam lemak
rantai panjang yang genap atau oksidasi badan-badan keton yang
hanya membentuk asetil KoA. Dalam siklus TCA, dua karbon adalah
hilang dari sitrat sebelum suksinil KoA dibentuk dan selanjutnya
tidak ada konversi bersih karbon asetil menjadi oksaloasetat.
4.11 SOAL – SOAL LATIHAN
1. Jelaskan jumlah ATP bersih yang dihasilkan pada siklus Kreb !
2. Jelaskan enzim-enzim yang terlibat selama reaksi Siklus Kreb
berlangsung !
3. Jelaskan peran enzim piruvat dehidrogenase !
4. Jelaskan aksi enzim transasetilase dan phosphatase !+
5. Rasio NADH/NAD dalam hati merupakan indikator utama
efektivitas proses siklus Kreb, jelaskan !
171siklus kreb


Almatsjer., S. 2002. Prinsip Dasar Ilmu Gizi. PT. Gramedia Pustaka
Utama, Jakarta.
Anonim. 2005. Pectin. (24 September http://www.wikipedia.com
2007)
Arjmandi, B. H., Craigh, J., Nathani, S., Reeves, R. D. 1992. Soluble
Dietary Fiber and Cholesterol Influence In Vivo Hepatic and
Intestinal Cholesterol-Biosyntehesis in Rats. J Nutr., 122:1559
– 1565.
Bagian Gizi R.S Dr. Cipto Mangunkusomo dan Persatuan Ahli Gizi
Indonesia. 1996. Penuntun Diet. Ed. Kedua Penerbit PT
Gramedia Pustaka Utama, Jakarta.
Champe, P. C and R. A. Harvey. 1994. Lippincott's Illustrated ndReviews: Biochemistry . Ed. 2 ., page: 171 – 186.
Chaplin, M. 2006. Pectin. [Online]. www.waterandscience.com
Donald, S. and McLaren. 1981. Nutrition and its Disorders. Third
Edition Churchill Livingstone Edinburgh, London, Melbourne
DAFTAR PUSTAKA

and New York.
Eleanor, R and Williams. 1984. Nutrition, Principles, Issues, and
Applications. McGraw-Hill Book., Company, New York
copyright Fergus M.Clydesdale: Food Nutrition and Health.
1995. The AVI Publishing Company Inc. WeStport,
Connecticut.
Fahmy, S. 2007. Prostate cancer cells killed by pectin in Fruits and
V e g e t a b l e s .
http://www.medicalnewstoday.com/articles/80268.php (23
Agustus 2007)
Fernandez, M. L., Wilson, T. A., Conde, K., Vergara – Jimenez and M.
Nicolosi R. J. 1994. Citrus Pectin and Cholesterol Interact to
Regulate Hepatic Cholesterol Homeostasis and Lipoprotein
Metabolism - A Dose – Response Study in Guinea – Pigs. Am. J.
Clin Nutr., 59:869-878.
Fernandez, M. L., Wilson, T. A., Conde, K., Vergara – Jimenez and M.
Nicolosi R. J. 1999. Hamster and Guinea Pigs Differ in Their
Plasma Lipoprotein Cholesterol Distribution when Fed Diets
Varying in Animal Protein, Soluble Fiber, or Cholesterol
Content. J. Nutr., 129 : 1323-1332.th
Fruton, J. S., and S. Simmonds. 1963. Biochemstry. Ed. 5 . John
Wiley and Sons, Inc., NewYork. pp. 619-633.
Hexeberg, S., Hexeberg, E., Willumsen, N and R. K. Berge. 1994. A
Study On Lipid-Metabolism In Heart And Liver Of Cholesterol
- Fed and Pectin – Fed Rats. Br. J. Nutr., 71:181-192.
Ismadi, M., dan S. D. Ismadi. 1993. Biokimia Satu Pendekatan
Berorientasi Kasus. Jilid 2, Edisi keempat. Gajahmada
fisiologi hewan174

University Press. th
Lesson, S and J. D. Summers. 2001. Nutrition of the Chicken. 4 Ed.
University Books. Guelph, Ontario, Canada .
Judd, P. A and A. S. Truswell. 1985. The Hypocholesteroleamic
Effects Of Pectins In Rats. Br. J. Nutr., 53: 409-425.
Lehninger, A., Nelson, D and M. M. Cox. 1993. Principles of nd
Biochemistry . 2 Ed.
McDonald, P., Edwards, R. A., Greenhalgh, J. F. D and C. A. Morgan. th
2002. Animal Nutrition. 6 Ed. Prentice Hall. London . th
Murray, R. K. Harper's Biochemistry. 2000. 25 Ed. Appleton and th
Lange. America: Stryer, L .1995. Biochemistry, 4 Ed. page 603
– 623.Moerdowo, R.M. 1989 . Spektrum Diabetes Mellitus.
Penerbit Djambatan, Jakarta.
Marks, D. B., Maraks, A. D and C. M. Smith. 1996. Basic Medical
Biochemistry. William and Walkins A Waverly Company,
Baltimore.
Marounek, M., Synytsya, A., Capikova, J and K. Sirotek. 2005. Assay
Of Availability Of Amidated Pectins For Colonic
Microorganisms (In Czech). Chem, Listy., 99:591-593.
Mayes, P. A. 1995. Biokimia Harper's. Edisi 22. Hartono A,
penerjemah. Jakarta : Penerbit buku kedokteran EGC.
Terjemahan dari : Harper's Biochemistry.
Montgomery, R., Dryer, R. L., Conway, T and W. Spector, A. A. 1983.
Biokimia suatu Pendekatan Berorientasi Kasus. Jilid 2, Edisi 4.
Gajah Mada University.
Muchtadi, D., Sri Palupi, N dan M, Astawan. 1993. Metabolisme Zat
Gizi, Sumber, Fungsi dan Kebutuhan Bagi Manusia. Jilid 2.
175siklus kreb

Pustaka Sinara Harapan. Jakarta:
Nangia – Makker, P., Hogan, V., Honjo, Y., Baccarini, S., Tait, L.,
Bresalier, R and A. Raz. 2002. Inhibiton Of Human Cancer Cell
Growth And Metastasis In Nude Mice By Oral Intake Of
Modifies Citrus Pectin. Journal of the National Cancer Inst.,
94/24:1854 – 1862.
Pastuszewska, B., Kowalczyk, J and A. Ochtabinska. 2000. Dietary
Carbohydrates Affect Caecal Fermentation and Modify
Nitrogen Excretion Pattern In Rats I. Studies With Protein –
Free Diets. Arch. Anim. Nutr., 53: 207-225.
Piliang, W. G., Djojosoebagio, S dan A. L. Haj. 2006. Fisiologi
Nutrisi. Vol. 1, Ed. Rev.. IPB Press, Bogor.
Ramunson, M. 1993. Nutrition Communication In The Post –
Epidemiological Transition: Lessons Leorned from the US
Experience. Widyakarya Nasional Pangan dan Gizi. LIPI.
R a v n s k o v U . 2 0 0 3 . T h e C h o l e s s t e r o l M y t h s .
http://www.ravnskov.nu/cholesterol.htm. (21 Desember
2005).
Sediaoetama, A. D. 1989. Ilmu Gizi. Penerbit Dian Rakyat, Jakarta .
Stryer, L. 2000. Biokimia. Sadikin, M. penerjemah; Zahir S. S,
Setiadi E. Edisi 4, volume 2. Penerbit Buku Kedokteran EGC,
Jakarta: Terjemahan dari: Biochemistry.
Terpstra, A. H. M., Lapre, J. A., De Vries, H. T and A. C. Beynen.
1998. Dietary Pectin With High Viscosity Lowers Plasma And
Liver Cholesterol Concentration And Plasma Cholesteryl Ester
Transfer Protein Activity In Hamsters. J. Nutr., 128: 1944 –
1949.
fisiologi hewan176

Terpstra, A. H. M., Lapre, J. A, De Vries, H. T and A. C. Beynen. 2002.
The Hypocholesterolemic Effect Of Lemon Peels, Lemon
Pectin, And The Waste Stream Material Of Lemon Peels In
Hybrid F1B Hamsters. Eur. J. Nutr., 41: 19-26.nd
Williams, S. R. 1973. Nutrion and Diet Therapy. 2 Ed. The CV.
Mosby Company. Saint Louis.
Winarno, F. G. 1984. Kimia Pangan dan Gizi. Penerbit PT. Gramedia.
Wirahadikusumah, M. 1985. Biokimia Metabolisme Energi,
Karbohidrat dan Lipid. Bandung. Penerbit ITB.
177siklus kreb


Adenilat siklase : enzim yang mengubah ATP menjadi AMP
siklik sebagai respons terhadap suatu sinyal
kimiawi
Aldehida : molekul organic dengan gugus karbonil yang
terletak pada ujung kerangka karbon
AMP siklik : adenosine monofosfat siklik, molekul
(cAMP, cyclic berbentuk cincin yang dibuat dari ATP yang
AMP) merupakan molekul penyisalan intraseluler
umum (second messenger) pada sel
eukariotik
Androgen : hormone steroid utama pada seks jantan,
misalnya testosterone, yang menstimulasi
perkembangan dan pemeliharaan system
reproduktif jantan dan sifat – sifat seks
sekunder
Antibodi : immunoglobulin pengikat antigen yang
GLOSARIUM

dihasilkan oleh sel limfosit B, berfungsi
sebagai efektor dalam suatu respons imun
Antigen : makromolekul asing yang bukan merupakan
bagian dari organisme inang dan yang
memicu munculnya respons imun
Arteriosklerosis : penyakit kardiovaskuler yang disebabkan
oleh pembentukan kerak keras di dalam arteri
Asam amino : molekul organic yang memiliki gugus
karboksil maupun gugus amino. Asam amino
berfungsi sebagai monomer protein
Asam amino : asam amino yang tidak dapat disintesis
sendiri oleh sel – sel tubuh hewan atau
manusia sehingga harus tersedia dalam
makanan. Ada 9 asam amino esensial yang
dibutuhkan oleh tubuh, yaitu histidin, lisin,
leusin, isoleusin, metionin, valin, treonin,
venilalanin, triptofan
Asam amino non : asam amino yang dapat disintesis oleh sel –
esensial sel tubuh hewan atau manusia dari suplai
nitrogen. Ada 11 asam amino esensial yang
dibutuhkan oleh tubuh, yaitu alanin, arginin,
asparagin, asam aspartat, asam glutamat,
glutamin, glisin, serin, prolin, sistein, tirosin
Asam amino semi : asam amino yang pada kondisi tertentu tidak
esensial dapat disintesis oleh sel – sel tubuh. Ada 4
asam amino semi esensial, yaitu argin,
glutamine, sistein, tirosin
fisiologi hewan180

Asam lemak : asam karboksilik dengan rantai karbon
(fatty acid) panjang. Asam lemak bervariasi panjang,
jumlah dan lokasi ikatan gandanya; tiga asam
lemak berikatan dengan satu molekul gliserol
akan membentuk lemak
Asam lemak jenuh (saturated fatty acid) asam lemak dimana
semua karbon dalam ekor hidrokarbon
dihubungkan oleh ikatan tunggal, sehingga
memaksimumkan jumlah atom hydrogen
yang dapat berikatan dengan kerangka
karbon
Asam lemak tak jenuh (unsaturated fatty acid) asam lemak yang
memiliki satu atau lebih ikatan ganda antara
karbon – karbon dalam ekor hidrokarbon.
Ikatan tersebut mengurangi jumlah atom
hydrogen yang terikat ke kerangka karbon
Asetil CoA (asetyl CoA) senyawa awal atau pembuka untuk siklus
Krebs dalam respirasi seluler; terbentuk dari
suatu fragmen asam piruvat yang terikat pada
suatu koenzim
Asetilkolin salah satu neurotransmitter yang paling umum; berfungsi
dengan cara mengikatkan diri pada reseptor
dan mengubah permeabilitas membran
pascasinaptik terhadap ion spesifik, yang
selanjutnya akan membuat membran
mengalami depolarisasi atau hiperpolarisasi
ATP (adenosine triphosphate = adenosin trifosfat) nukleosida
181siklus kreb

tr ifosfat mengandung adenin yang
mengeluarkan energi bebas ketika ikatan
fosfatnya dihidrolisis. Energi ini digunakan
untuk menggerakkan reaksi endergonik
(yang memerlukan energi) dalam sel
ATP sintase (ATP synthase) kumpulan beberapa protein membran
yang ditemukan di dalam krista mitokondria
yang berfungsi dalam kemiosmosis dengan
rantai t ransport elektron terdekat ,
menggunakan energi dar i gradien
konsentrasi ion hidrogen untuk membuat
ATP. ATP sintase menyediakan suatu tempat
persinggahan yang dilalui ion hidrogen yang
akan berdifusi ke dalam matriks mitokondria
Deaminasi adalah proses penghilangan gugus amin sehingga akan
membentuk senyawa baru. Proses deaminasi
membutuhkan oksigen yang diperlukan
untuk oksidasi dengan melibatkan enzim
dehidrogenase.
Difusi kecenderungan spontan suatu bahan untuk berpindah
menuruni gradien konsentrasinya dari daerah
dengan konsentrasi yang lebih tinggi ke
konsentrasi yang lebih rendah
Difusi yang dipermudah (facilitated diffusion) aliran molekul atau
ion secara spontan, yang terikat ke suatu
protein pembawa spesifik, melewati suatu
membran biologis menuruni gradien
fisiologi hewan182

konsentrasinya
Disakarida : gula yang terdiri atas dua monomer
monosakarida yang dihubungkan melalui
sintesis dehidrasi (pembentukan air)
Energi : kapas i t as a tau kemampuan un tuk
melaksanakan pekerjaan dengan cara
menggerakkan materi melawan gaya yang
berlawanan
Enzim : suatu kelas protein yang berfungsi sebagai
katalis, agen kimiawi yang mengubah laju
suatu reaksi tanpa dikonsumsi oleh reaksi itu
Epinefrin : hormon yang dihasilkan sebagai respons
terhadap stress, juga disebut adrenalin
Eritrosit : sel darah merah, mengandung hemoglobin,
yang berfungsi mengangkut oksigen di
dalam sistem sirkulasi atau sistem peredaran
Estrogen : hormon seks steroid betina yang utama, yang
dihasilkan di dalam ovarium oleh folikel
yang sedang berkembang selama paruh
pertama siklus estrus dan dalam jumlah yang
lebih kecil oleh korpus luteum selama paruh
kedua siklus estrus. Estrogen merangsang
perkembangan dan pemeliharaan sistem
reproduksi betina dan ciri – ciri seks sekunder
Fermentasi : proses katabolik yang membuat sejumlah
tertentu ATP dari glukosa tanpa rantai
transport elektron dan yang menghasilkan
183siklus kreb

produk akhir yang khas, seperti etil alkohol
atau asam laktat
Filamen : komponen sitoskeleton atau kerangka sel
intermediet yang meliputi semua filamen yang
ukurannya berada diantara ukuran
mikrotubula dan mikrofilamen
Fosfolipid : molekul yang menyusun bilayer bagian
(phospholipid) dalam membran biologis, yang memiliki
kepala hidrofilik dan polar, dan ekor
hidrofobik yang non – polar. Fosfolipid
merupakan trigliserida dengan posisi karbon
ketiga ditempati oleh gugus fosfat dan gugus
basa yang mengandung nitrogen
Fosforilasi : produksi ATP dengan menggunakan energi
oksidatif yang diperoleh dari reaksi redoks pada suatu
rantai transport elektron
Fotosintesis : pengubahan energi cahaya menjadi energi
kimiawi yang disimpan dalam bentuk
glukosa atau senyawa organik lainnya,
terjadi pada tumbuhan, algae, dan prokariot
tertentu
Glikogen : polisakarida cadangan yang terdiri atas
monomer glukosa yang bercabang banyak
dan yang ditemukan dalam sel hati dan otot;
pada manusia atau hewan
Glikokalik : manter longgar pada bagian permukaan luar
(glycocalyx) sel hewan yang terbuat dari oligosakarida
fisiologi hewan184

yang lengket
Glikolisis : pemecahan glukosa menjadi piruvat.
Glikolisis adalah suatu jalur metabolisme
yang terjadi pada semua sel hidup berfungsi
sebagai titik permulaan untuk fermentasi
atau respirasi aerobik
Glukagon : hormon peptida yang disekresikan oleh sel –
(glucagon) sel endokrin pankreas yang berfungsi untuk
menaikkan kadar gula darah, yang
merupakan hormon yang berlawanan dengan
fungsi insulin
Gugus amino : gugus fungsional yang terdiri dari satu atom
(amino group) nitrogen yang berikatan dengan dua atom
hidrogen; dapat bertindak sebagai basa
dalam suatu larutan, yang menerima ion
hidrogen dan mendapatkan suatu muatan
positif
Gugus hidroksil : gugus fungsional yang terdiri atas suatu atom
(hydroxyl group) hidrogen yang digabungkan dengan atom
oksigen oleh ikatan kovalen polar. Molekul
yang memiliki gugus fungsional ini menjadi
larut dalam air dan disebut alkohol
Gugus karboksil : gugus fungsional yang ditemukan pada asam
(carboxyl group) organik, terdiri atas suatu atom karbon
tunggal yang berikatan dengan suatu atom
oksigen dan juga berikatan dengan sebuah
gugus hidroksil
185siklus kreb

Hemoglobin : protein mengandung besi dalam sel darah
merah yang berikatan secara reversibel
dengan oksigen
Hidrofilik : mempunyai afinitas terhadap air
Hidrolisis : proses kimiawi yang melisiskan atau
memutuskan molekul dengan cara
penambahan air; suatu proses yang sangat
esensial dalam pencernaan
Hiperlipidemia : adalah suatu keadaan yang ditandai oleh
peningkatan kadar lipid/lemak darah
Homeostasis : kondisi fisiologis yang mantap dalam tubuh
Hormon : salah satu diantara banyak jenis sinyal
kimiawi yang beredar pada semua organisme
multiseluler yang dibentuk dalam sel – sel
terspesialisasi, yang beredar dalam cairan
tubuh dan mengkoordinasikan berbagai
bagian organisme dengan cara berinteraksi
dengan sel – sel target
Hidrofobik : yang mempunyai ketidaksukaan atau
menolak air; cenderung bersatu atau
membentuk butiran di dalam air
Inositol : (inositol trifosfat atau IP3) messenger kedua
yang berfungsi sebagai intermidiet diantara
hormon non steroid tertentu dan messenger
ketiga, suatu peningkatan dalam konsentrasi 2+
Ca sitoplasmik
Karbohidrat : gula (monosakarida) atau salah satu dari
fisiologi hewan186

dimernya (disakarida) atau polimernya
(polisakarida)
Karotenoid : pigmen aksesoris berwarna kuning dan
jingga di dalam kloroplast tumbuhan; dengan
cara menyerap panjang gelombang cahaya
yang tidak dapat diserap oleh klorofil,
pigmen ini akan memperluas spektrum
warna sehingga dapat menggerakkan
fotosintesis
Karsinogenik : agen kimia yang bersifat menyebabkan
kanker
Kelenjar endokrin : kelenjar tak bersaluran yang mensekresikan
(endocrine gland) hormon secara langsung ke dalam aliran
darah
Klorofil : pigmen hijau yang terdapat dalam kloroplast
tumbuhan; klorofil a dapat berpartisipasi
langsung dalam reaksi terang, yang
mengubah energi cahaya matahari menjadi
energi kimiawi
Koenzim : molekul organik yang berfungsi sebagai
kofaktor. Sebagian besar vitamin berfungsi
sebagai koenzim di dalam reaksi metabolik
yang penting
Kofaktor : setiap molekul atau ion non protein yang
diperlukan agar suatu enzim bisa berfungsi
dengan baik. Kofaktor dapat berikatan secara
permanen dengan tempat aktif enzim atau
187siklus kreb

bisa berikatan secara longgar dengan substrat
selama katalisis
Kolesterol : steroid yang menyusun komponen esensial
membran sel hewan dan bertindak sebagai
molekul prekursor untuk sintesis steroid lain
yang penting secara biologis
Lemak (triasil : senyawa biologis yang terdiri atas tiga asam
gliserol) lemak utama yang diikatkan dengan satu
molekul gliserol
Lemak netral : adalah lemak yang padat pada suhu kamar
dan berubah menjadi minyak pada saat
mencair
Lesitin : adalah merupakan ikatan antara asam fosfat
(fosfatidilkolin) dan basa yang mengandung N – kolin;
berfungsi memudahkan pengangkutan dan
p e n g g u n a a n a s a m l e m a k d e n g a n
menggunakan enzim lesitin – kolesterol
asiltransferase sebagai katalisator
Lignin : bahan keras yang terkubur dalam matriks
selulosa dinding sel tumbuhan berpembuluh
yang berfungsi sebagai adaptasi yang penting
untuk mendukung spesies terestrial
Lipid : salah satu keluarga senywa, yang meliputi
lemak, fosfolipid, dan steroid yang tidak larut
dalam air
Lipoprotein : protein yang terikat dengan lipid; meliputi
lipoprotein berdensitas rendah (LDL) dan
fisiologi hewan188

lipoprotein berdensitas tinggi (HDL) yang
mengangkut lemak dan kolesterol dalam
darah
Matriks : substansi dimana sel – sel jaringan hewan
ekstraseluler berada; terdiri atas protein dan polisakarida
(ECM, extrasellular
matriks)
Matriks : komponen non hidup pada jaringan ikat,
yang terdiri atas suatu anyaman serat yang
terkubur dalam bahan dasar homogen yang
bisa berbentuk cairan, seperti jeli atau
padatan
Matriks : ruangan mitokondria yang terbungkus oleh
mitokondria membran dalam dan mengandung enzim dan
substrat untuk siklus Krebs
Metabolisme : totalitas proses kimiawi suatu organisme,
yang terdiri atas jalur – jalur katabolik dan
anabolik
Mikrofilamen : protein aktin berbentuk batang padat dalam
sitoplasma hampir semua sel eukariotik,
yang membentuk sebagian sitoskelet dan
yang bekerja secara sendirian atau bersama –
sama dengan miosin untuk menyebabkan
kontraksi sel
Mikrotubul : protein tubulin berbentuk batang berlubang
dalam sitoplasma hampir semua sel
eukariotik dan pada silia, flagela dan
189siklus kreb

sitoskelet
Mitokondria : organel pada sel eukariotik yang berfungsi
sebagai tempat respirasi sellular
Monosakarida : karbohidrat yang paling sederhana, yang
secara sendirian tetap aktif atau yang
berfungsi sebagai monomer untuk disakarida
dan polisakarida. Juga dikenal sebagai gula
sederhana, rumus molekul monosakarida
umumnya adalah kelipatan CH O2
+NAD : koenzim yang ditemukan dalam semua sel
(nicotinamide yang membantu enzim memindahkan
adenine elektron selama reaksi redoks metabolisme
dinucleotide)
Oksidasi : hilangnya elektron dari suatu substansi yang
terlibat dalam reaksi redoks
Ovarium : struktur yang menghasilkan gamet betina dan
hormon reproduktif
Pati (starch, : polisakarida cadangan pada tumbuhan yang
polisakarida) keseluruhannya terdiri atas glukosa
Polipeptida : polimer atau rantai yang terdiri dari banyak
asam amino yang diikatkan bersama oleh
ikatan peptida
Polisakarida : polimer yang terdiri dari sampai lebih dari
sejuta monosakarida, terbentuk melalui
sintesis kondensasi
Protein kinase : suatu enzim yang memindahkan gugus fosfat
dari ATP ke suatu protein
fisiologi hewan190

Protein : polimer biologis tiga domensi yang dibangun
dari kumpulan 20 jenis monomer yang
berbeda yang disebut asam amino
Reduksi : perolehan elektron oleh suatu substansi yang
terlibat dalam reaksi redoks
Respirasi sellular : jalur katabolik yang paling umum dan paling
efisien untuk menghasilkan ATP, dimana
oksigen dikonsumsi sebagai reaktan bersama
– sama dengan bahan bakar organik.
Respirasi seluler merupakan proses yang
melibatkan sistem rantai transport elektron
dan proses oksidasi – fosforilasi.
Ribosa : komponen gula pada RNA
Ruminansia : hewan seperti sapi atau domba dengan
lambung komplek yang dibagi dalam
beberapa ruangan (poligastrik) yang
dikhususkan untuk pola makan herbivora
Second messenger : molekul atau ion non protein kecil yang larut
(messenger kedua) dalam air, misalnya ion kalsium atau AMP
siklik, yang merelai sinyal ke bagian dalam
sel sebagai respons terhadap sinyal yang
diterima oleh protein reseptor sinyal
Safonifikasi : a d a l a h p r o s e s h i d r o l i s i s d e n g a n
menggunakan bahan alkali
Selulosa : polisakarida struktural dinding sel yang
terdiri atas monomer glukosa yang diikat
dengan ikatan β – 1, 4 – glikosidik
191siklus kreb

Serat (fiber) : suatu jenis sel berlignin yang memperkuat
xilem angiospermae dan berfungsi untuk
memberikan dukungan mekanis; sel
slerenkima pipih dan runcing yang
umumditemukan dalam bentuk berkas
Siklus Krebs : siklus kimiawi yang melibatkan delapan
tahapan yang menyempurnakan perombakan
metabolik molekul glukosa menjadi
karbondioksida; terjadi di dalam mitokondria
Siklus aerobik : proses metabolisme yang terjadi secara
berulang dengan membutuhkan oksigen,
mengacu pada proses seluler yang
memerlukan oksige
Siklus anaerobik : proses metabolisme yang terjadi secara
berulang tanpa memerlukan oksigen
mengacu pada proses seluler tanpa oksigen
dan mungkin akan teracuni oleh kehadiran
oksigen
Sistem limbik : sekelompok nukleus (kumpulan badan sel
(limbic system) saraf) pada bagian bawah otak depan
mamalia yang berinteraksi dengan kortek
serebri dalam penentuan emosi; meliputi
hipokampus dan amigdala
Sistem saraf : suatu subdivisi dari sistem saraf motoris
otonom vertebrata yang mengatur lingkungan
internal; terdiri atas divisi simpatik dan
parasimpatik
fisiologi hewan192

Sistem saraf pusat : pada hewan vertebrat otak dan sumsum
(CNS, central tulang belakang (medula spinalis)
nervous system)
Sistem saraf : cabang divisi motoris sistem saraf tepi
somatis (somatic vertebrata yang terdiri atas neuron motoris
nervous system) yang membawa sinyal ke otot rangka sebagai
respons terhadap rangsangan eksternal
Sistem saraf tepi : neurons sensoris dan motoris yang
(PNS, peripheral berhubungan dengan sistem saraf pusat
nervous system)
Sitokrom : protein yang mengandung besi, suatu
(sitokrom P – 45 konponen rantai transport elektron dalam
oksidase) mitokondria
Sitoplasma : keseluruhan isi sel selain nukleus, dan
dibungkus oleh membran plasma
Sitoskeleton : jaringan kerja yang terdiri mikrotubula,
mikrofilamen, dan filamen intermidiet yang
bercabang di seluruh sitoplasma dan
melakukan berbagai fungsi mekanis dan
transport
Sitosol : bagian semicair sitoplasma
Testis : organ reproduksi jantan atau gonad, dimana
sperma dan hormon reproduksi dihasilkan
Tirosin kinase : enzim yang mengkatalisis transfer gugus
fosfat dari ATP ke asam amino tirosin dalam
suatu protein substrat
Transport aktif : pergerakan suatu substansi melewati suatu
193siklus kreb

membran biologis melawan gradien
konsentrasi atau elektrokimiawi dengan
bantuan input energi dan protein transport
spesifik
Transport pasif : difusi suatu bahan melewati membran
biologis
Transaminasi : adalah proses pemindahan gugus amin dari
satu asam amino untuk membentuk asam
amino baru. Proses transaminasi melibatkan
enzim transaminase atau amino-transferase
dengan kofaktor enzim piridoksal fosfat
fisiologi hewan194

FISIOLOGI hewan Buku ini membahas tentang konsep fisiologi yang dapat digunakan sebagai referensi dalam proses perkuliahan dan sebagai acuan bagi mahasiswa dalam mengembangkan keilmuan di bidang Fisiologi. Pembahasan isi buku ini dimulai dengan metabolism karbohidrat yang berisi penjelasan tentang energi, penggunaan dan jalur energi, jenis, fungsi, klasifikasi, dan sumber karbohidrat, jalur pemrosesan, pencernaan, dan metabolism karbohidrat di dalam tubuh. Metabolisme lemak berisi penjelasan tentang jenis lemak, klasifikasi, nomenklatur lemak, sifat, dan fungsi lemak, pencernaan, absorbs, transportasi, dan metabolism lemak. Selanjutnya, dibahas pula tentang metabolisme protein yang berisi penjelasan tentang pengertian protein, pencernaan protein, absorbs dan transportasi asam-asam amino, metabolisme protein dan asam-asam amino. Dalam upaya melengkapi pembahasan tentang metabolism karbohidrat, lemak, dan protein, buku ini berisi penjelasan pula tentang siklus Krebs sebagai proses lanjutan dari ketiga metabolism tersebut.
Dr. Sunarno, S.Si, M.Si dilahirkan di Klaten (Jawa Tengah), 22 September 1973. Ia menyelesaikan pendidikan Sarjana di Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Diponegoro, Semarang (1997), Magister Bidang Fisiologi dan Biologi Perkembangan di Pasca Sarjana Institut Teknologi Bandung (2003), dan Doktor dalam Bidang Ilmu – Ilmu Faal dan Khasiat Obat di Sekolah Pasca Sarjana Institut Pertanian Bogor (2012). Ia sekarang.
Dosen di Bidang Fisiologi pada Fakultas Sains dan Matematika, Universitas Diponegoro.Selain aktif terlibat dalam pengembangan dan pemahaman sains, penelitian dan pengabdian masyarakat, validasi riset khusus vector dan reservoir penyakit, ia juga telah menerbitkan beberapa artikel di jurnal nasional terakreditasi maupun jurnal internasional.
MadinaJl. Bulusan XI/5 Semarang
(024) 76482660





























