Embed Size (px)
Citation preview
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
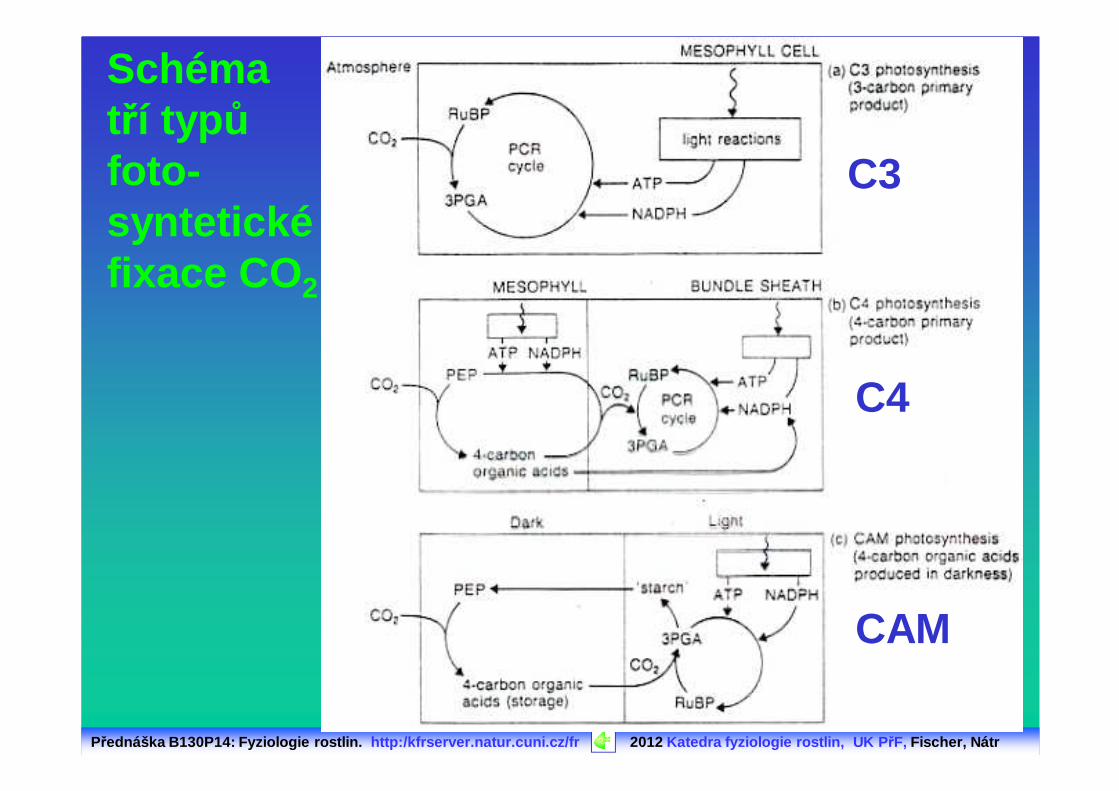
Schéma tří typůfoto -syntetickéfixace CO 2
C3
C4
CAM
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
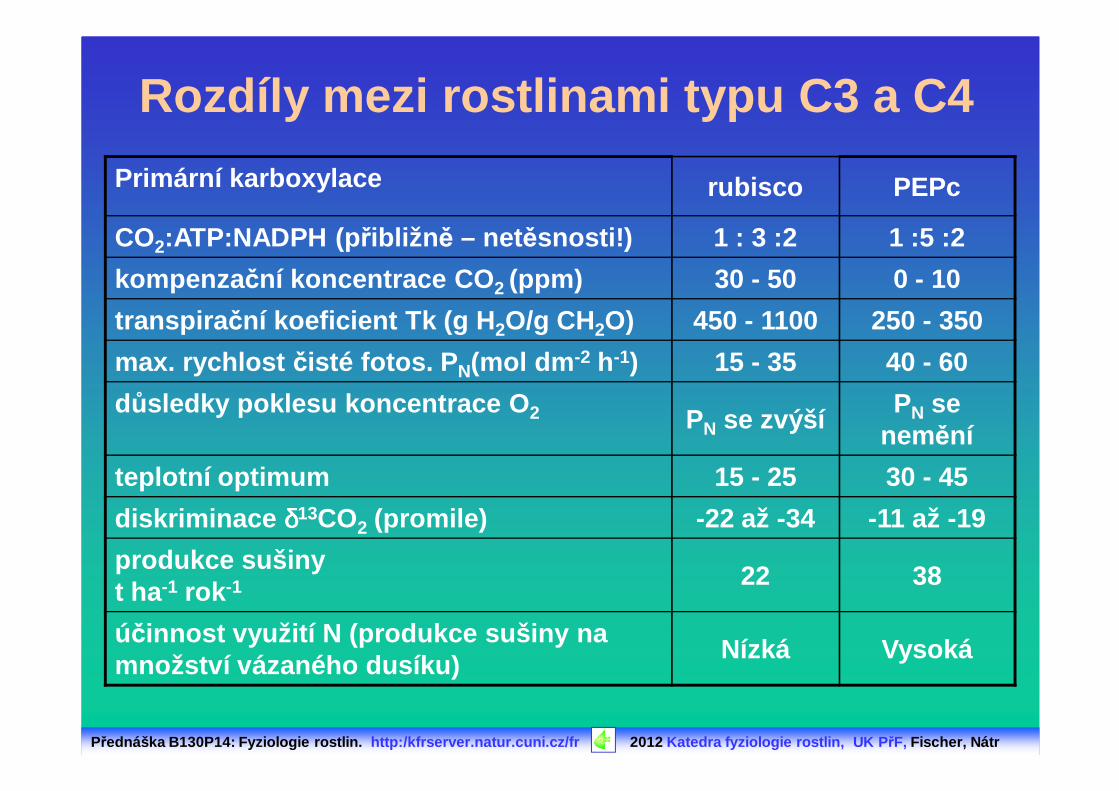
Rozdíly mezi rostlinami typu C3 a C4
Primární karboxylace rubisco PEPc
CO2:ATP:NADPH (p řibližn ě – netěsnosti!) 1 : 3 :2 1 :5 :2
kompenza ční koncentrace CO 2 (ppm) 30 - 50 0 - 10
transpira ční koeficient Tk (g H 2O/g CH2O) 450 - 1100 250 - 350
max. rychlost čisté fotos. P N(mol dm -2 h-1) 15 - 35 40 - 60
důsledky poklesu koncentrace O 2 PN se zvýšíPN se
nemění
teplotní optimum 15 - 25 30 - 45
diskriminace δδδδ13CO2 (promile) -22 až -34 -11 až -19
produkce sušinyt ha -1 rok -1 22 38
účinnost využití N (produkce sušiny na množství vázaného dusíku)
Nízká Vysoká
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
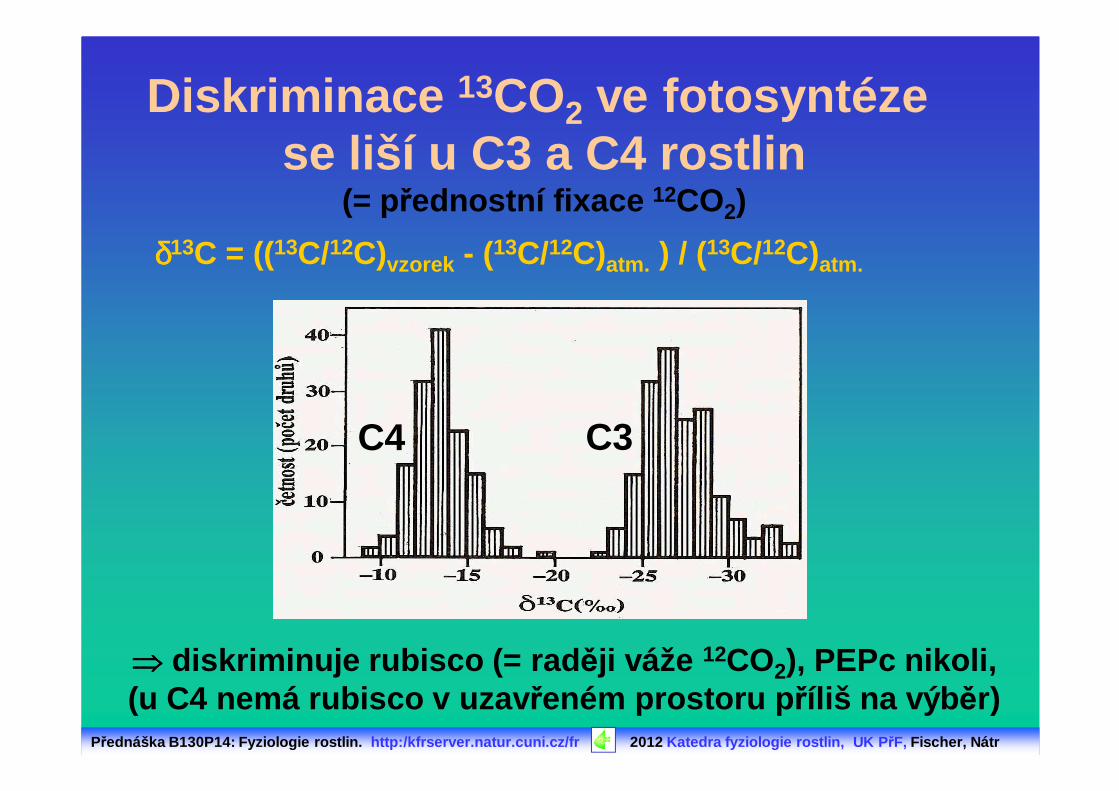
Diskriminace 13CO2 ve fotosyntéze se liší u C3 a C4 rostlin
(= přednostní fixace 12CO2)
δδδδ13C = ((13C/12C)vzorek - (13C/12C)atm. ) / (13C/12C)atm.
C3C4
⇒⇒⇒⇒ diskriminuje rubisco (= rad ěji váže 12CO2), PEPc nikoli,(u C4 nemá rubisco v uzav řeném prostoru p říliš na výb ěr)
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
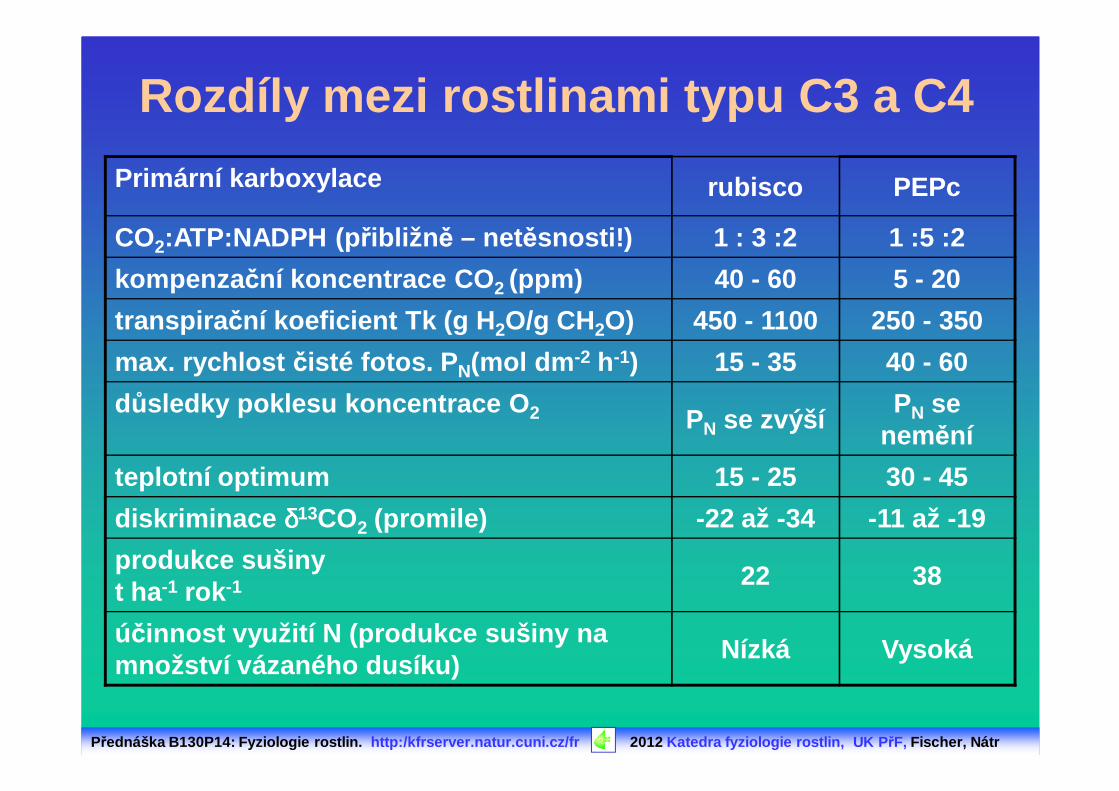
Rozdíly mezi rostlinami typu C3 a C4
Primární karboxylace rubisco PEPc
CO2:ATP:NADPH (p řibližn ě – netěsnosti!) 1 : 3 :2 1 :5 :2
kompenza ční koncentrace CO 2 (ppm) 40 - 60 5 - 20
transpira ční koeficient Tk (g H 2O/g CH2O) 450 - 1100 250 - 350
max. rychlost čisté fotos. P N(mol dm -2 h-1) 15 - 35 40 - 60
důsledky poklesu koncentrace O 2 PN se zvýšíPN se
nemění
teplotní optimum 15 - 25 30 - 45
diskriminace δδδδ13CO2 (promile) -22 až -34 -11 až -19
produkce sušinyt ha -1 rok -1 22 38
účinnost využití N (produkce sušiny na množství vázaného dusíku)
Nízká Vysoká

Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
Příklad rozdíl ů v účinnosti využítí vody, dusíku a zá ření C3 a C4
Rýže (C3) Kuku řice (C4)
Vody (g DM / kg vody) 76 144
Dusíku ( µµµµmol CO 2 s-1 / mmol N) 0,26 0,74
Záření (g DM / MJ pohlcené FAR) 2,2 3,3
Mitchell P. L., Sheehy J. E., New Phytol. 171:688, 2006
Účinnost využití dusíku = množství asimilovaného CO 2 na obsah dusíku v listu (v ětšina dusíku je vázaná v proteinech, u C3 rostlin j e až polovina protein ů tvo řena enzymem rubisco, u C4 mnohem mén ě, tedy dusík je efektivn ěji využit u C4)

Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
C3, C4, CAM
Fotoinhibice

Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
Fotoinhibicesoubor proces ů projevujících se snížením rychlosti fotosyntézy
při zvyšující se ozá řenosti
ROS:1O2
Fotopoškození (snížení po čtu f čních fotosystém ů)- fotosyntetických struktur (protein ů)
- UV světlo, oxidativní poškození (ROS, P680+, …)
- nastává i za nízké ozá řenosti, ale rychlá oprava – nenastává fotoinhibice
Fotoprotekce (snížení E p řenesené do RC … k fixaci CO 2)- předcházení fotopoškození (rychle vratné zm ěny)
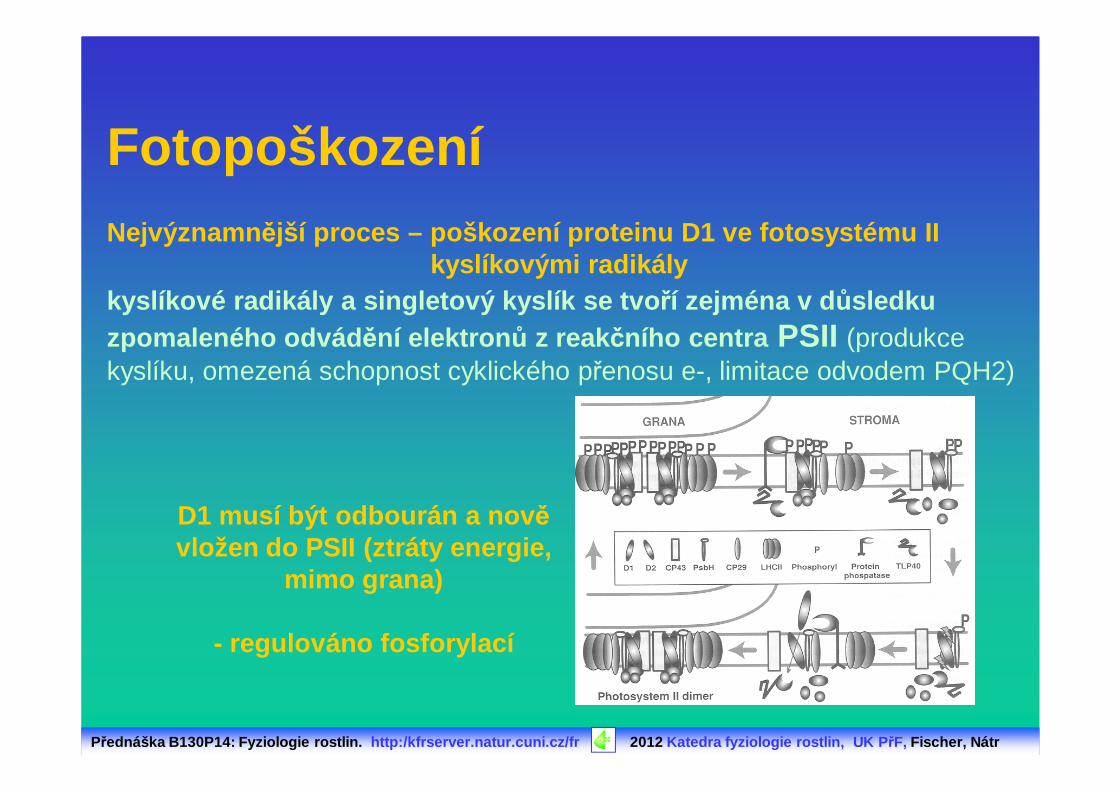
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
kyslíkové radikály a singletový kyslík se tvo ří zejména v d ůsledku zpomaleného odvád ění elektron ů z reakčního centra PSII (produkce kyslíku, omezená schopnost cyklického přenosu e-, limitace odvodem PQH2)
FotopoškozeníNejvýznamn ější proces – poškození proteinu D1 ve fotosystému II
kyslíkovými radikály
D1 musí být odbourán a nov ě vložen do PSII (ztráty energie,
mimo grana)
- regulováno fosforylací
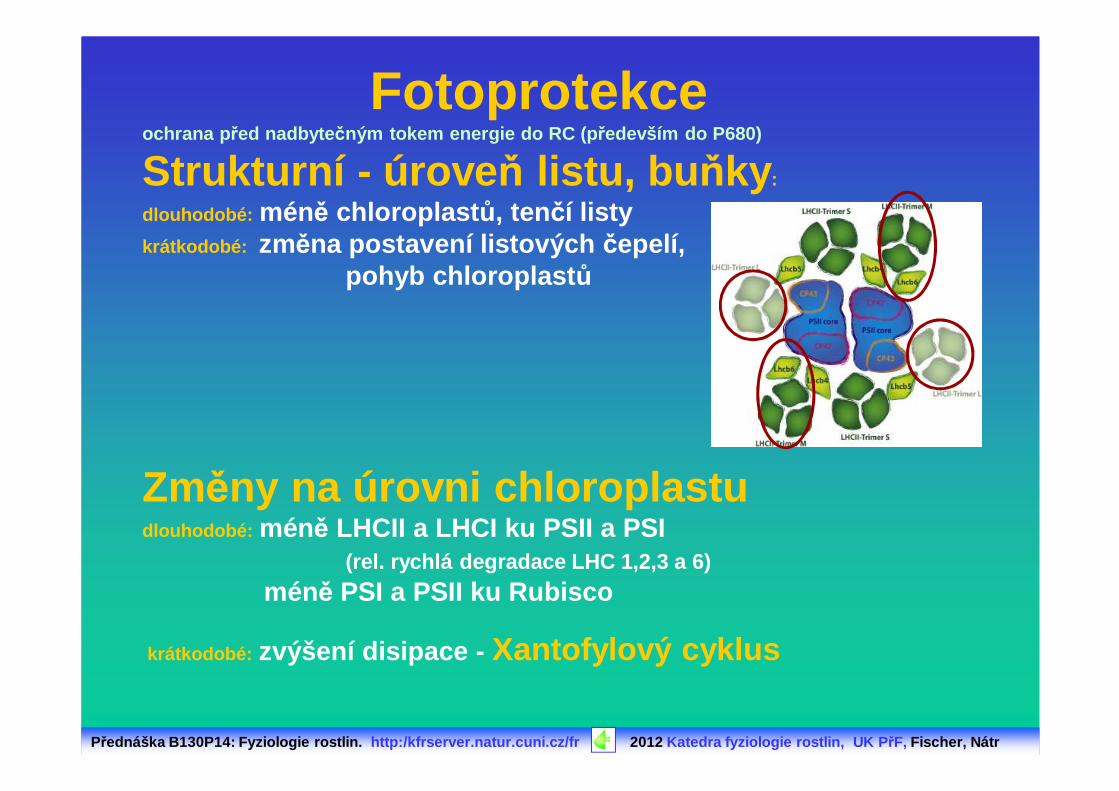
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
ochrana p řed nadbyte čným tokem energie do RC (p ředevším do P680)
Strukturní - úrove ň listu, bu ňky :
dlouhodobé: méně chloroplast ů, tenčí listykrátkodobé: změna postavení listových čepelí,
pohyb chloroplast ů
Změny na úrovni chloroplastudlouhodobé: méně LHCII a LHCI ku PSII a PSI
(rel. rychlá degradace LHC 1,2,3 a 6)
méně PSI a PSII ku Rubisco
krátkodobé: zvýšení disipace - Xantofylový cyklus
Fotoprotekce
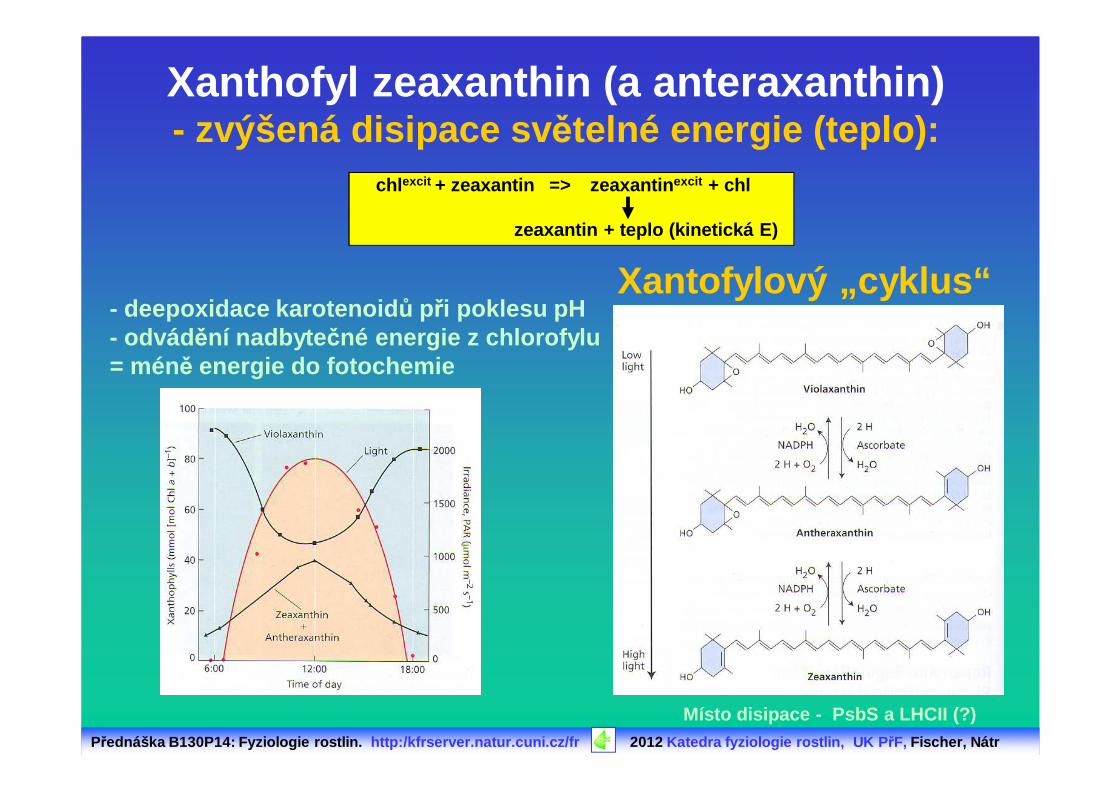
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
Xanthofyl zeaxanthin (a anteraxanthin)- zvýšená disipace sv ětelné energie (teplo):
chl excit + zeaxantin => zeaxantin excit + chl
zeaxantin + teplo (kinetická E)
- deepoxidace karotenoid ů při poklesu pH - odvád ění nadbyte čné energie z chlorofylu= méně energie do fotochemie
Xantofylový „cyklus“
Místo disipace - PsbS a LHCII (?)

Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
Fotoinhibice
Faktory ovliv ňující fotosyntézu

Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
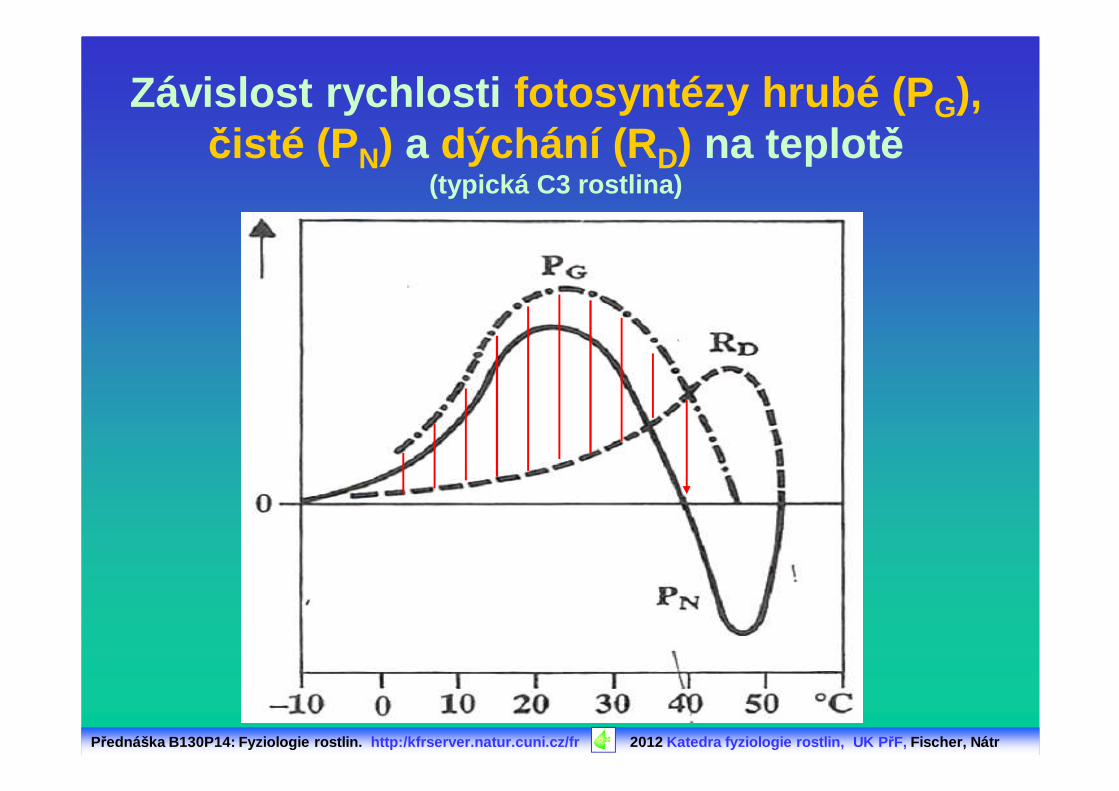
PG: celková (hrubá) fotosyntézaRL: fotorespiraceRD: dýchání (mitochondriální)
Rychlost fotosyntézy
- produkce O2- spotřeba CO2- produkce biomasy
PN: čistá (neto) fotosyntéza P N = PG - RD - RL
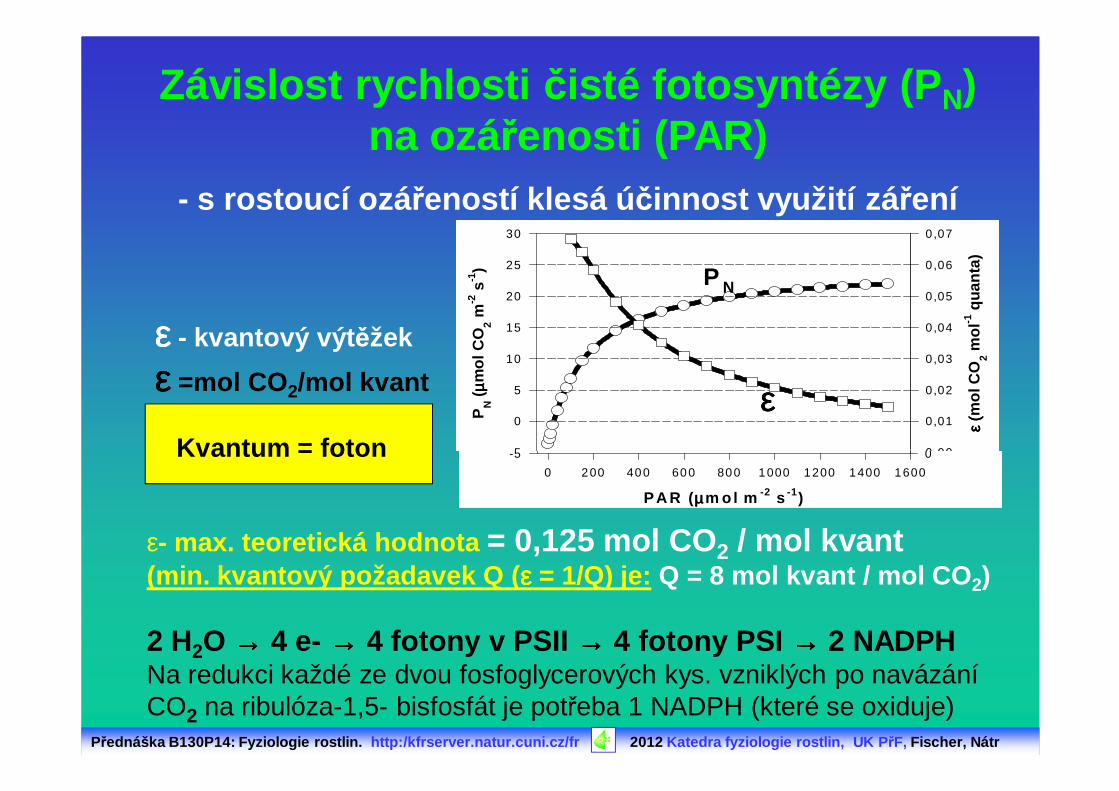
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
εεεε - kvantový výt ěžek
εεεε =mol CO 2/mol kvant
Kvantum = fotonP
N (
µµ µµmol
CO
2 m-2
s-1
)
-5
0
5
10
15
20
25
30
ε (
ε (
ε (
ε (m
ol C
O2 m
ol-1
qua
nta)
0,00
0,01
0,02
0,03
0,04
0,05
0,06
0,07
P N
εεεε
PA R ( µµµµm ol m -2 s -1)
0 200 400 600 800 1000 1200 1400 1600
-5 0
Závislost rychlosti čisté fotosyntézy (P N)na ozářenosti (PAR)
- s rostoucí ozá řeností klesá ú činnost využití zá ření
ε- max. teoretická hodnota = 0,125 mol CO 2 / mol kvant(min. kvantový požadavek Q ( εεεε = 1/Q) je: Q = 8 mol kvant / mol CO 2)
2 H2O →→→→ 4 e- →→→→ 4 fotony v PSII →→→→ 4 fotony PSI →→→→ 2 NADPHNa redukci každé ze dvou fosfoglycerových kys. vzniklých po navázání CO2 na ribulóza-1,5- bisfosfát je potřeba 1 NADPH (které se oxiduje)
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
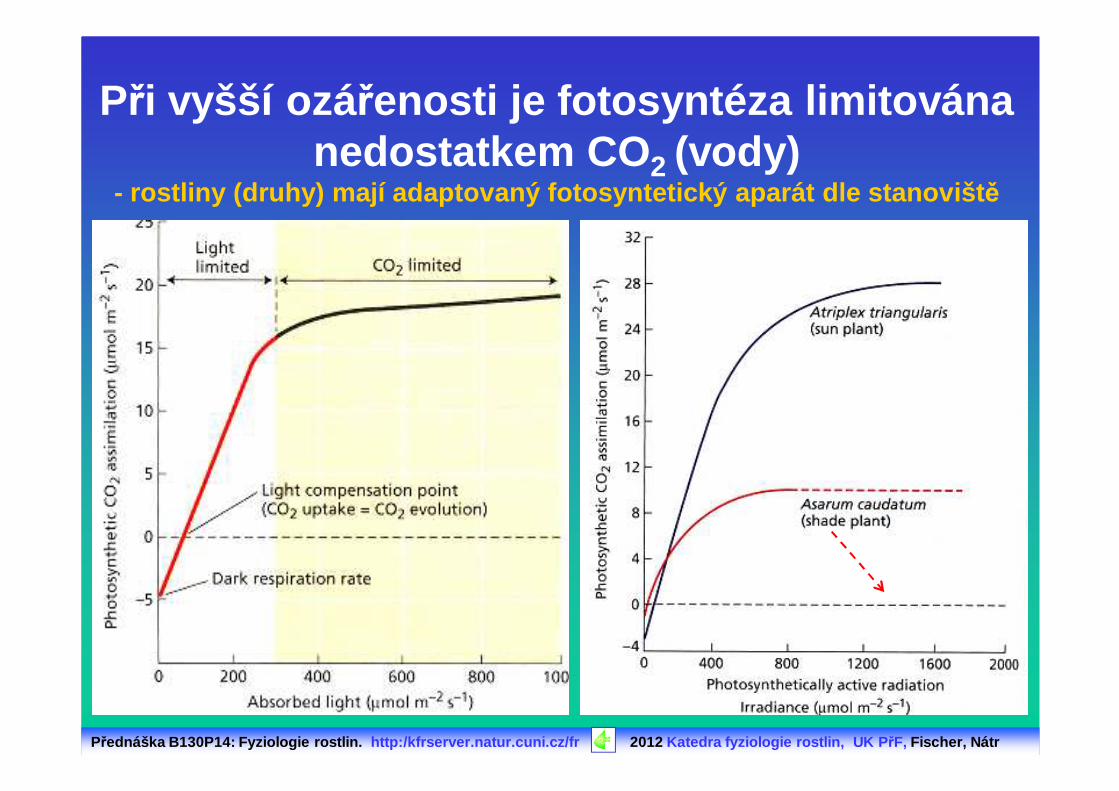
Při vyšší ozá řenosti je fotosyntéza limitovánanedostatkem CO 2 (vody)
- rostliny (druhy) mají adaptovaný fotosyntetický ap arát dle stanovišt ě

Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
500 W m-2 250 W m-2 125 W m-2
1 x 2 x 4 x 8 x větší plocha
Sluneční záření 1000 (W m-2)
1000 W m-2
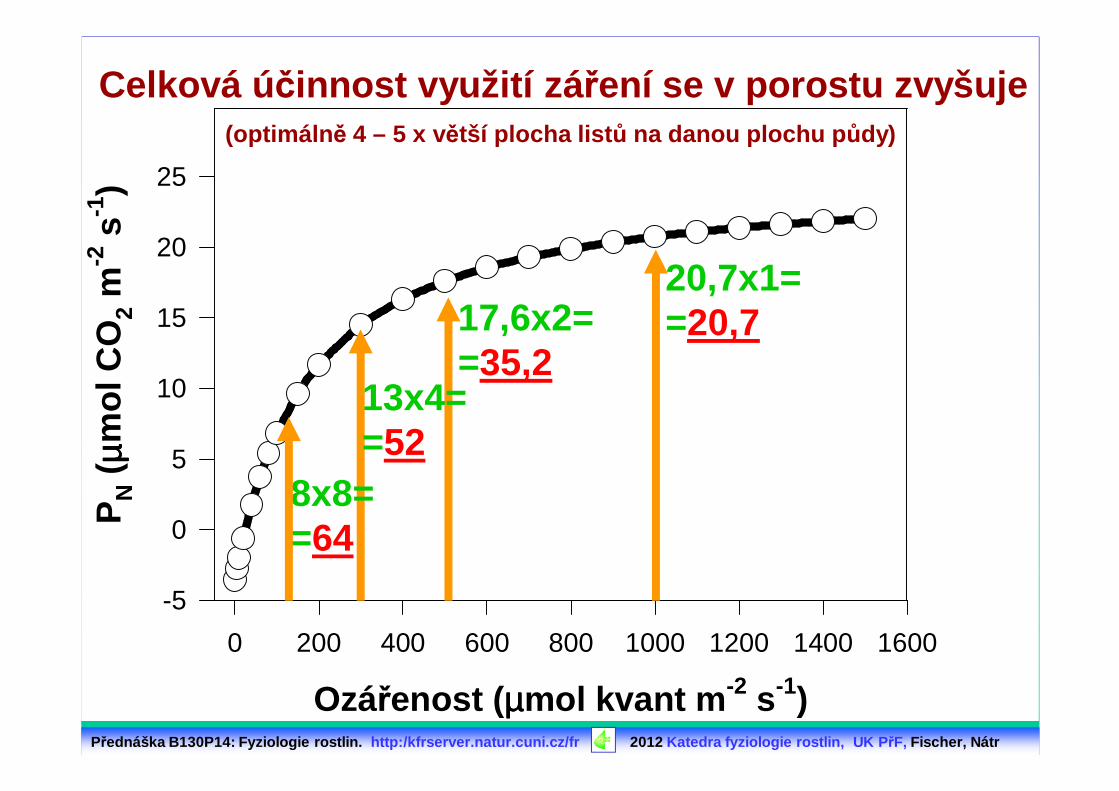
Rostliny p řirozen ě fotosyntetizují v porostu⇒⇒⇒⇒ výrazn ě vyšší ú činnost využití zá ření
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
Ozářenost ( µµµµmol kvant m -2 s-1)
0 200 400 600 800 1000 1200 1400 1600
PN (
µµ µµmol
CO
2 m
-2 s
-1)
-5
0
5
10
15
20
25
20,7x1==20,717,6x2=
=35,213x4==52
8x8==64
Celková ú činnost využití zá ření se v porostu zvyšuje(optimáln ě 4 – 5 x větší plocha list ů na danou plochu p ůdy)

Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
- rychlost fotosyntézy- rychlost fotorespirace- rychlost respirace- vodivost pr ůduch ů- vodní režim, ….
Vliv koncentrace CO 2
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
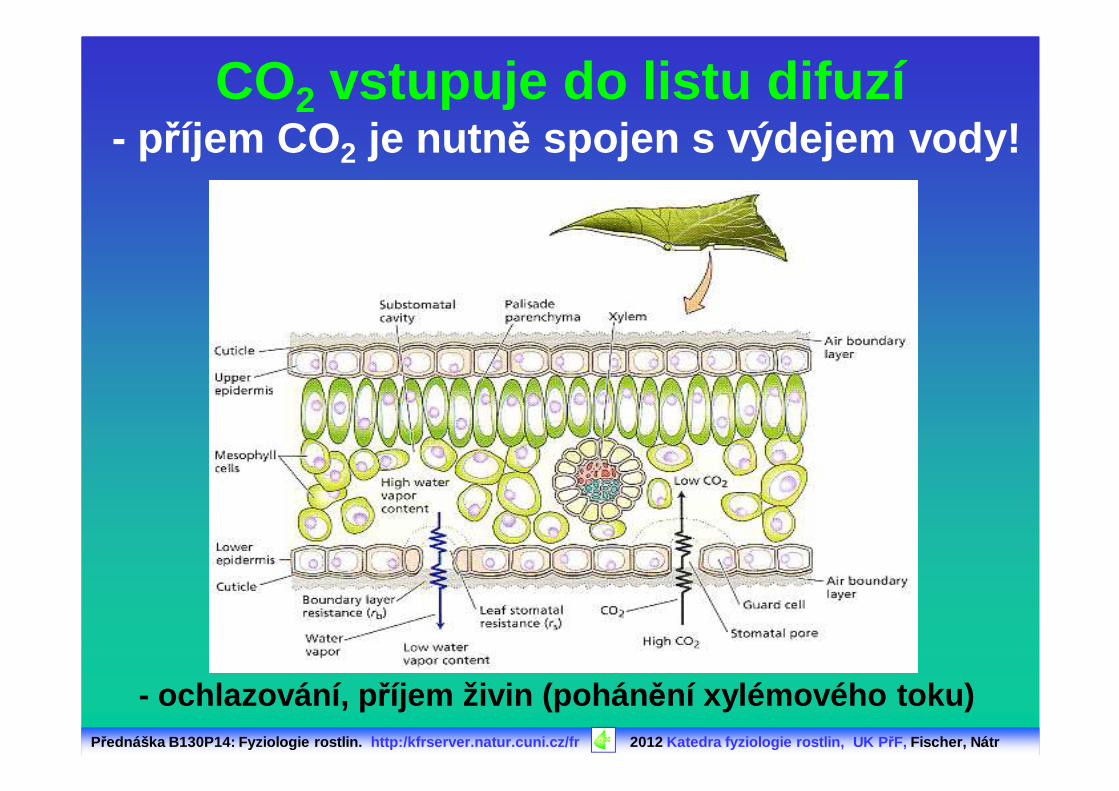
CO2 vstupuje do listu difuzí- příjem CO2 je nutn ě spojen s výdejem vody!
- ochlazování, p říjem živin (pohán ění xylémového toku)
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
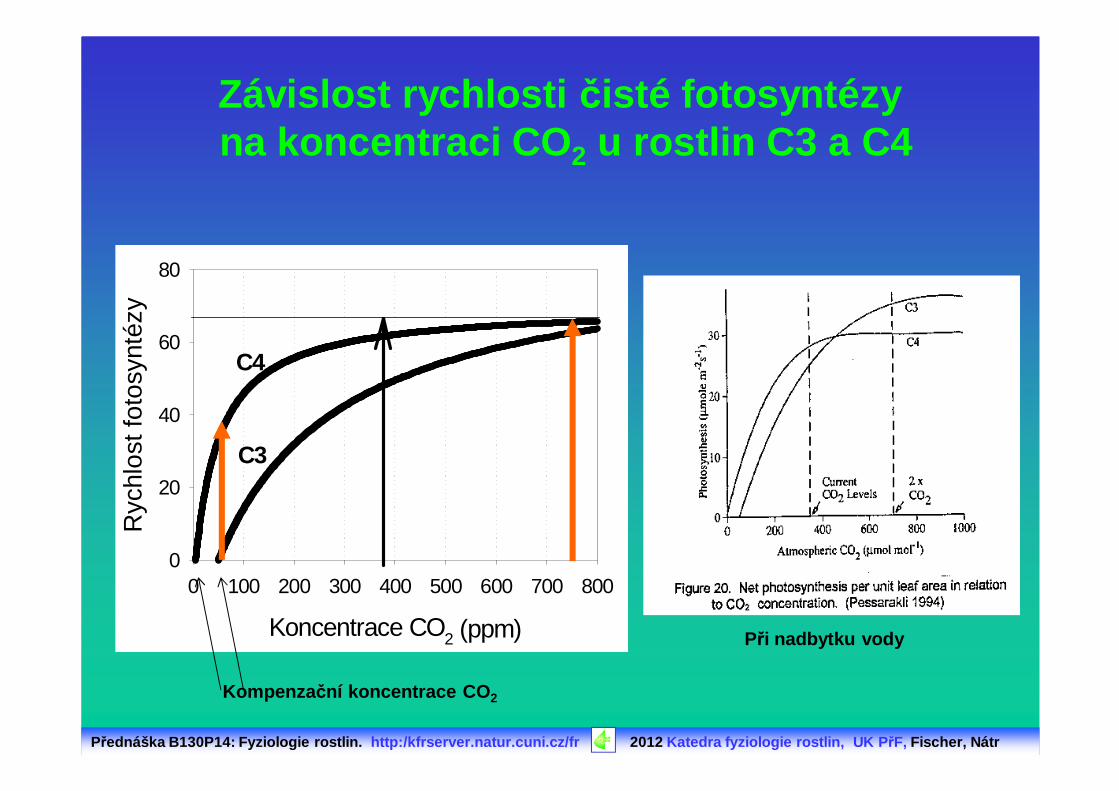
Závislost rychlosti čisté fotosyntézy na koncentraci CO 2 u rostlin C3 a C4
Koncentrace CO2 (ppm)
0 100 200 300 400 500 600 700 800
Ryc
hlos
t fot
osyn
tézy
0
20
40
60
80
C3
C4
Při nadbytku vody
Kompenza ční koncentrace CO 2

Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
Možné důsledky zvýšené koncentrace CO2
na porost s rostlinami C3 a C4
• změna konkuren čních vztah ů C3 x C4 ⇒⇒⇒⇒změna druhového složení rostlin
• změna „kvality“ biomasy (N/C aj. - C4 mén ě kvalitní)• změny spektra a množství šk ůdců a chorob,• změny spektra a množství býložravc ů (hmyzu),• změny spektra a množství predátor ů (ptáci aj.)
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
KultivarA
zuce
naIA
C 1
65IR
AT
104
ITA
186
Kin
. Pat
.
Sal
ump.
YR
L-39
grain373 grain664
Kultivar
Azu
cena
IAC
165
IRA
T 1
04
ITA
186
Kin
. Pat
.
Sal
ump.
YR
L-39
Kultivar - zm ěna pořadí
ITA
186
IR74
Sal
ump.
IR30
IR28
MG
L-2
IR46
Azu
cena
OS
4IR
AT
104
IR72
IR36
Kin
. Pat
.IR
64Y
RL-
39N
22IA
C 1
65
Rel
ativ
ní z
výše
ní (
664C
O2/
373C
O2,
%)
0
50
100
150
200
250
300
350
Kultivar - zm ěna pořadí
IR30
Sal
ump.
IR36
IR74
ITA
186
OS
4A
zuce
naIR
28K
in. P
at.
IR72
IRA
T 1
04Y
RL-
39M
GL-
2IR
46IR
64N
22IA
C 1
65
Rel
ativ
ní z
výše
ní (
664C
O2/
373C
O2,
%)
-25
0
25
50
75Výnos obilek na rostlinu Skliz ňový index
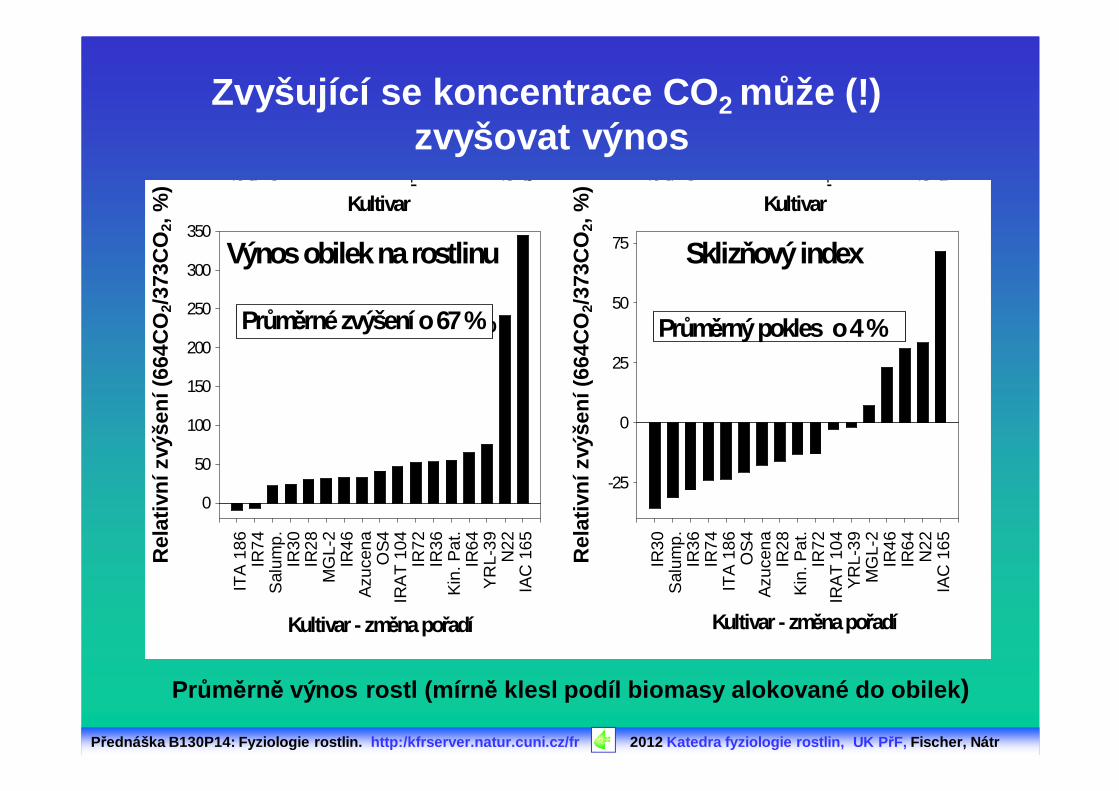
Průměrné zvýšení o 70 % Průměrné zvýšení o 23 %Průměrné zvýšení o 67 % Průměrný pokles o 4 %
Zvyšující se koncentrace CO 2 může (!) zvyšovat výnos
Průměrně výnos rostl (mírn ě klesl podíl biomasy alokované do obilek )
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
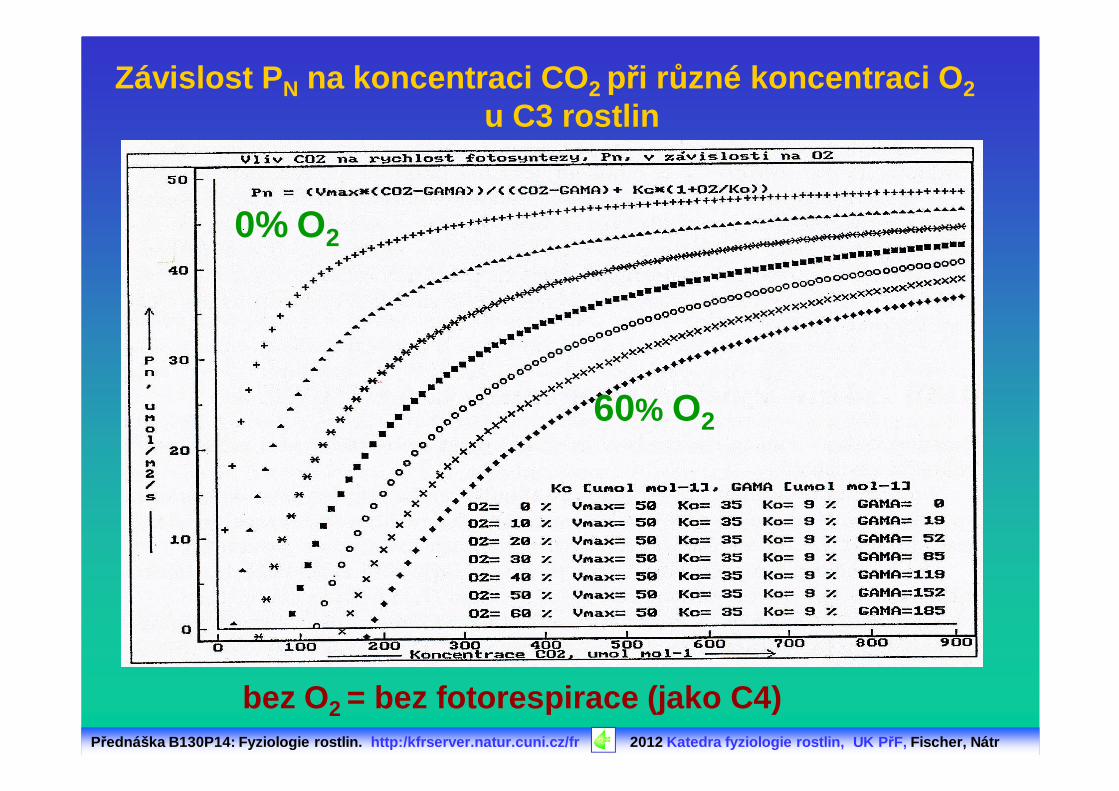
Závislost P N na koncentraci CO 2 při různé koncentraci O 2u C3 rostlin
bez O2 = bez fotorespirace (jako C4)
60% O2
0% O2
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
Závislost rychlosti fotosyntézy hrubé (P G), čisté (P N) a dýchání (R D) na teplot ě
(typická C3 rostlina)

Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
Faktory ovliv ňující fotosyntézu
Transport asimilát ů
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
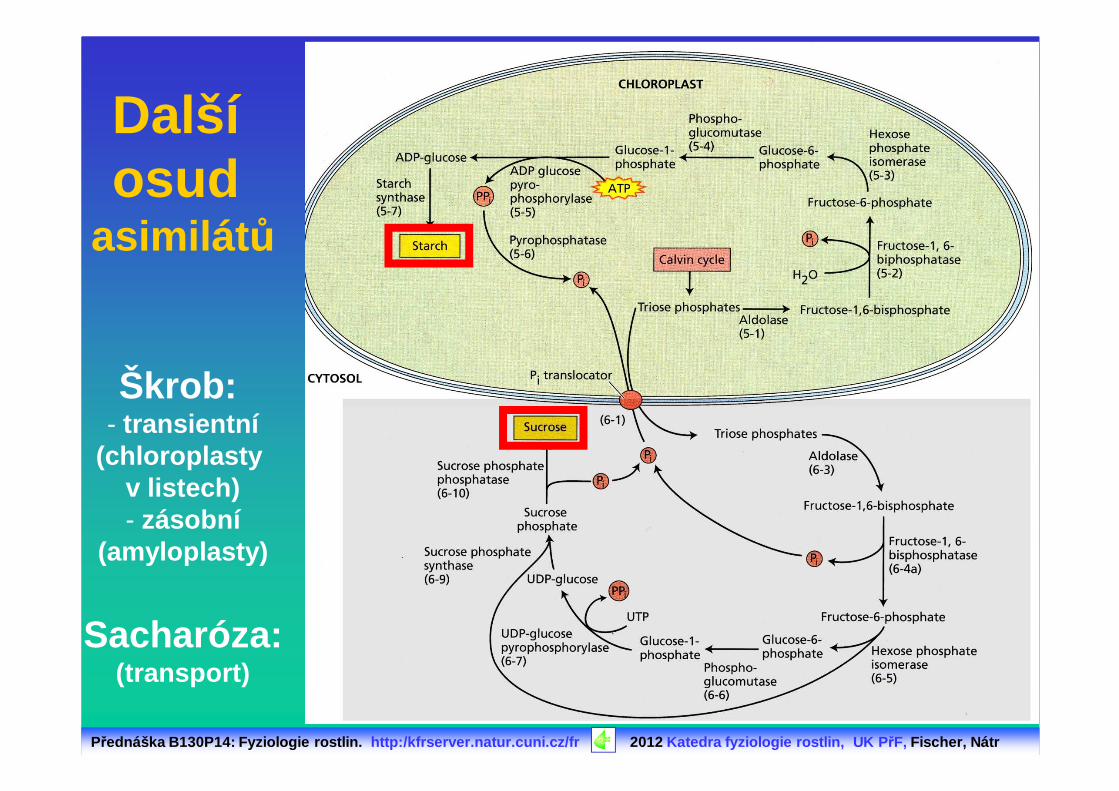
Další osud
asimilát ů
Škrob: - transientní
(chloroplasty v listech)- zásobní
(amyloplasty)
Sacharóza:(transport)
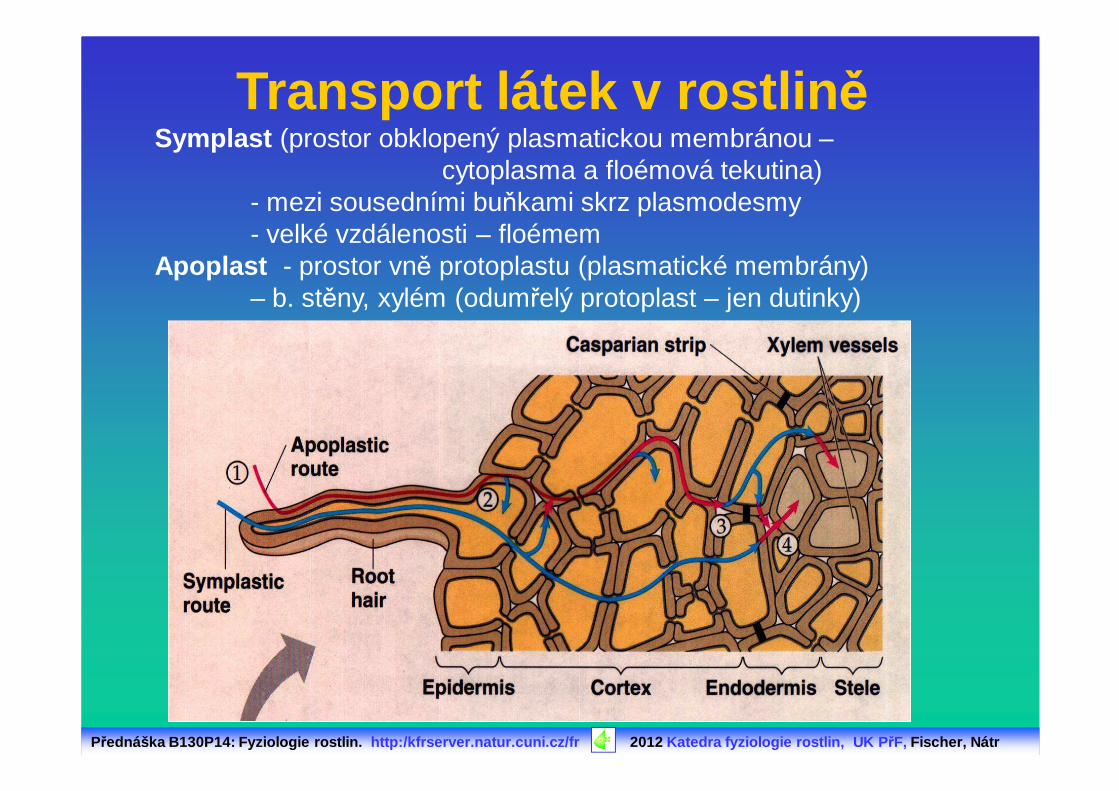
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
Transport látek v rostlin ěSymplast (prostor obklopený plasmatickou membránou –
cytoplasma a floémová tekutina)- mezi sousedními buňkami skrz plasmodesmy- velké vzdálenosti – floémem
Apoplast - prostor vně protoplastu (plasmatické membrány) – b. stěny, xylém (odumřelý protoplast – jen dutinky)
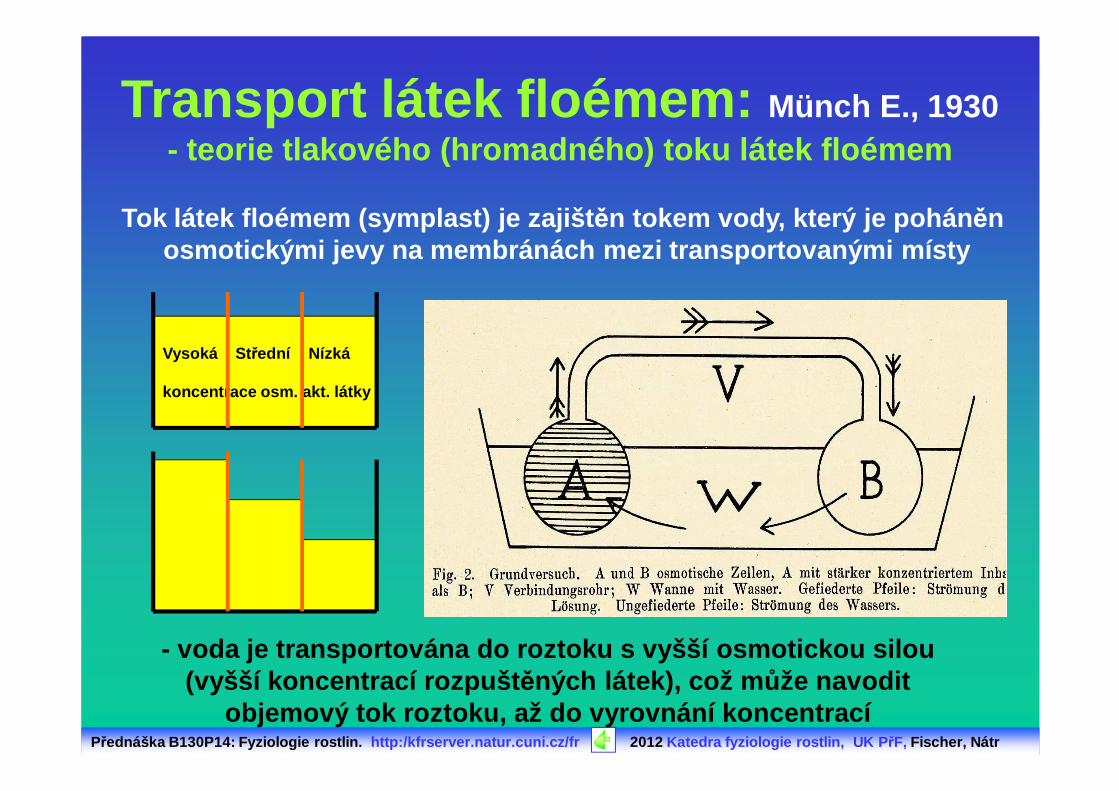
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
Transport látek floémem: Münch E., 1930- teorie tlakového (hromadného) toku látek floémem
- voda je transportována do roztoku s vyšší osmotick ou silou (vyšší koncentrací rozpušt ěných látek), což m ůže navodit
objemový tok roztoku, až do vyrovnání koncentrací
Vysoká St řední Nízká
koncentrace osm. akt. látky
Tok látek floémem (symplast) je zajišt ěn tokem vody, který je pohán ěn osmotickými jevy na membránách mezi transportovaným i místy
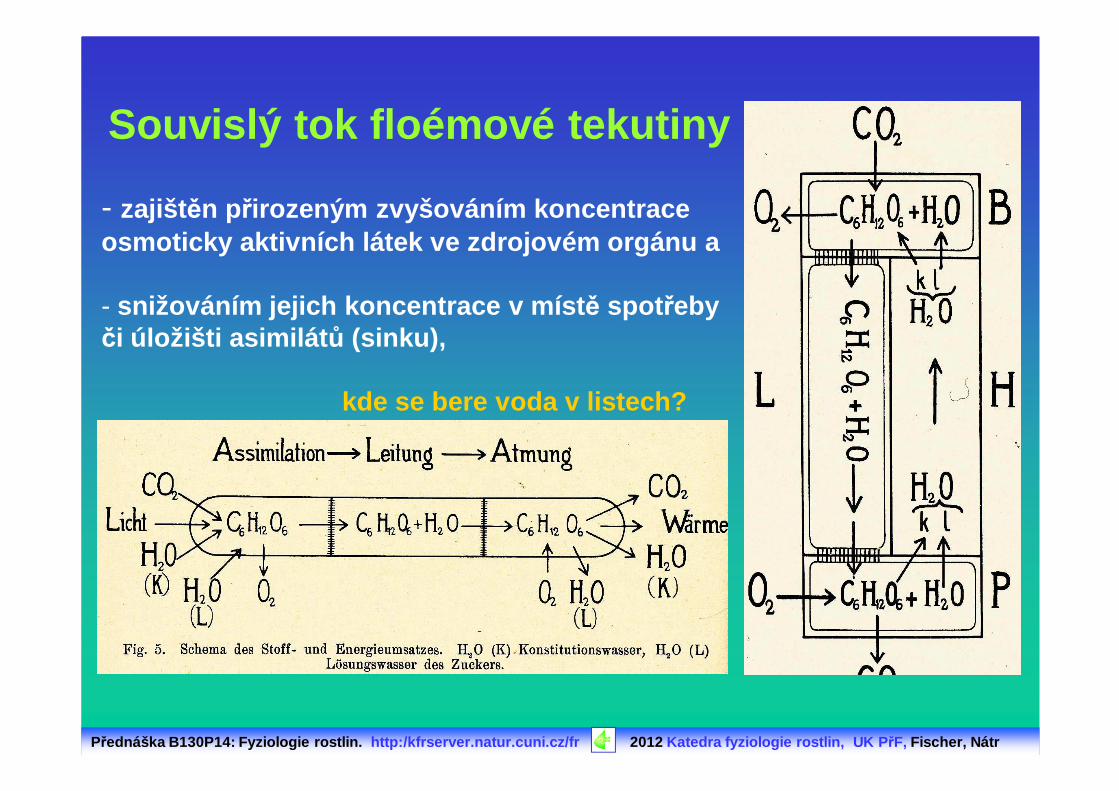
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
Souvislý tok floémové tekutiny
- zajišt ěn přirozeným zvyšováním koncentrace osmoticky aktivních látek ve zdrojovém orgánu a
- snižováním jejich koncentrace v míst ě spot řeby či úložišti asimilát ů (sinku),
kde se bere voda v listech?
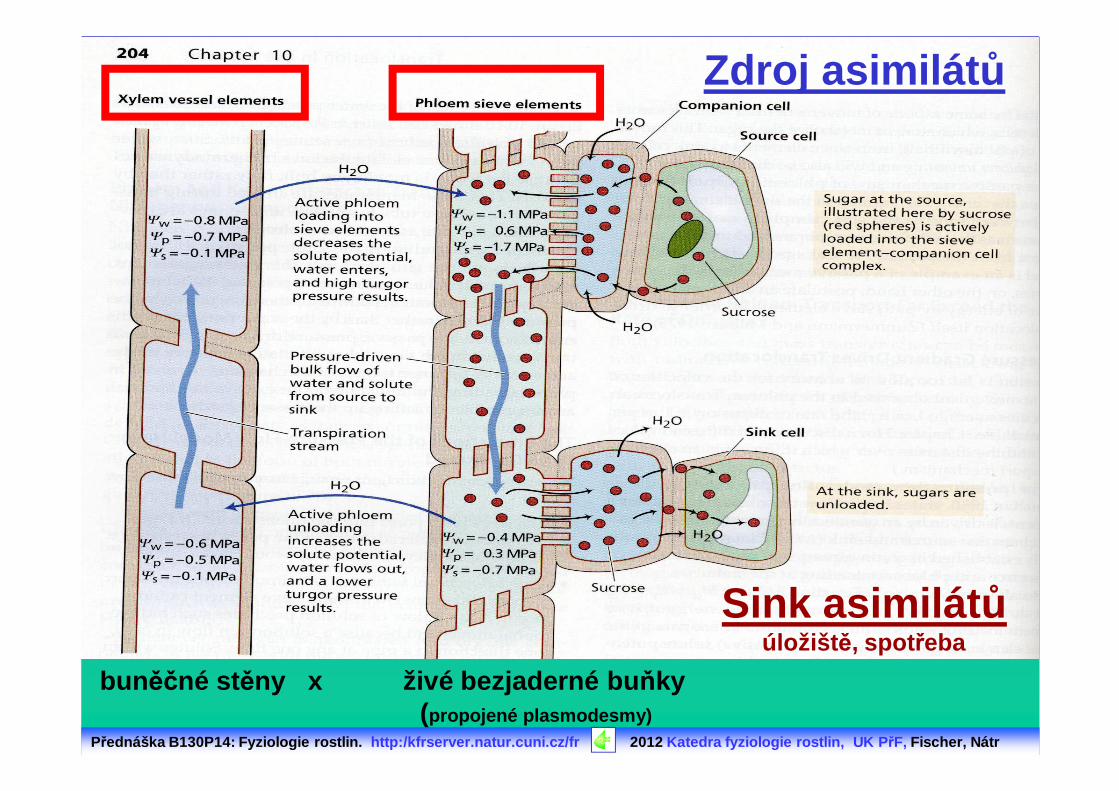
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
Zdroj asimilát ů
Sink asimilát ůúložišt ě, spot řeba
bun ěčné st ěny x živé bezjaderné bu ňky (propojené plasmodesmy)
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
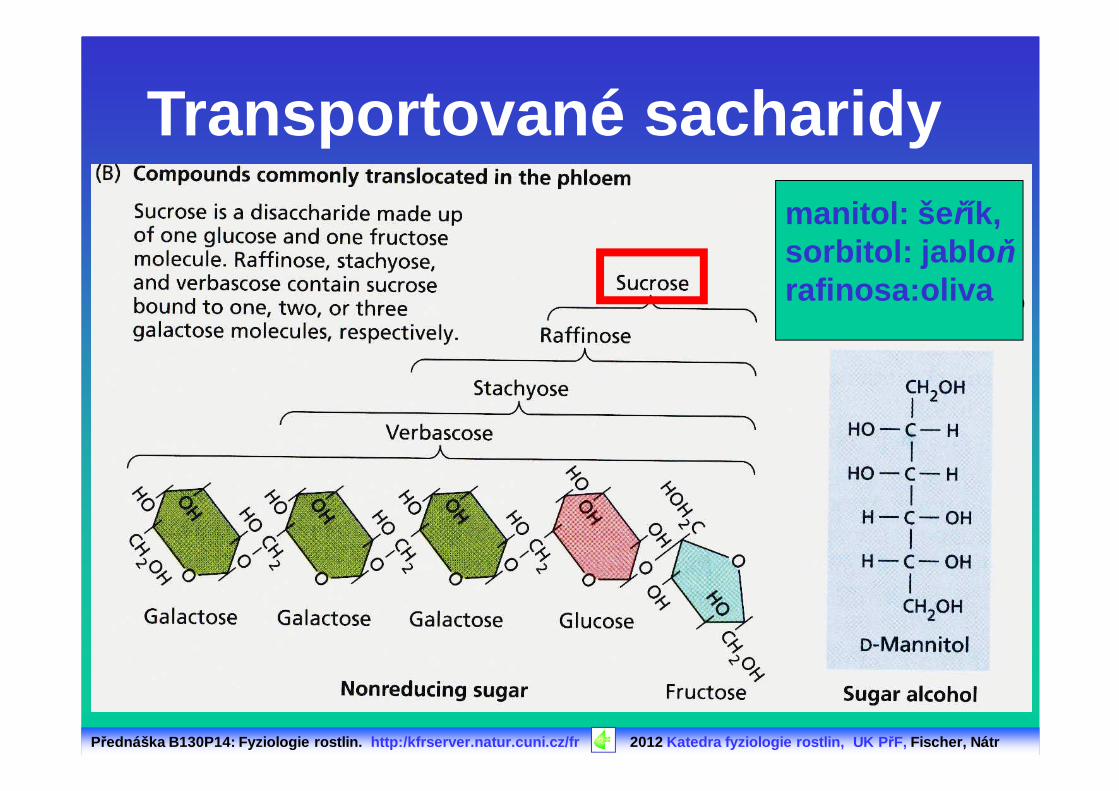
Transportované sacharidymanitol: šeřík,sorbitol: jablo ňrafinosa: oliva
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
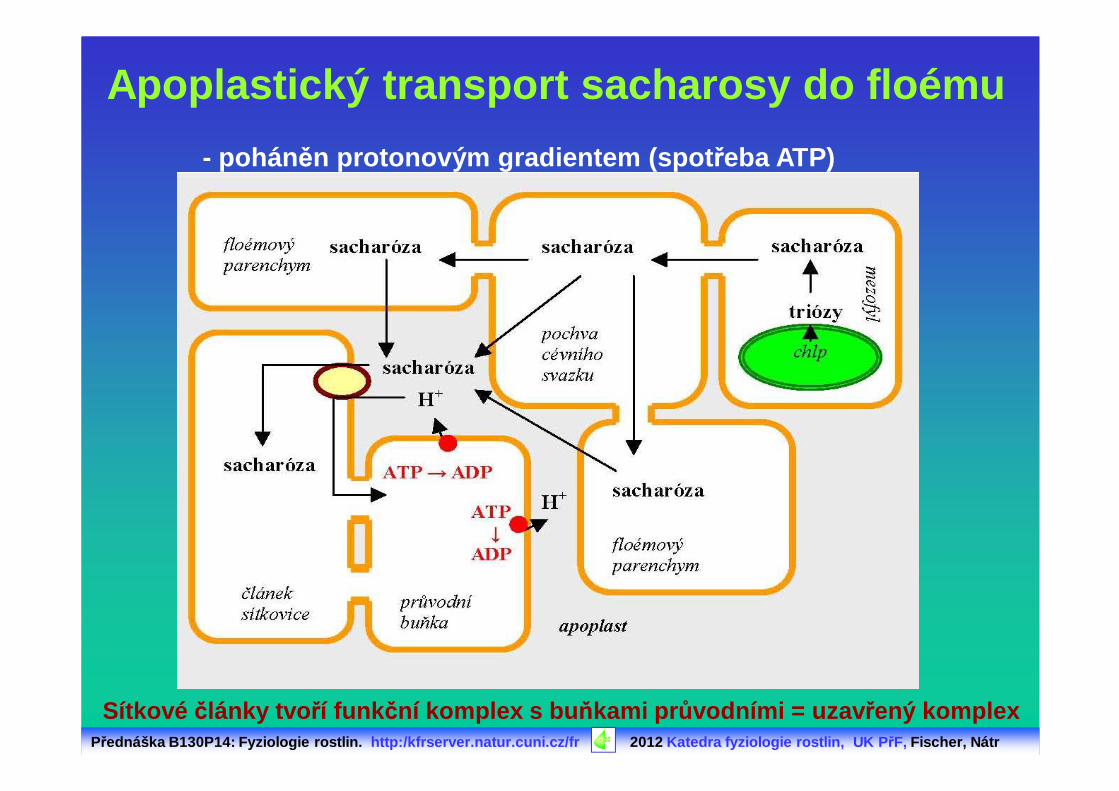
Sítkové články tvo ří funk ční komplex s bu ňkami pr ůvodními = uzav řený komplex
Apoplastický transport sacharosy do floému
- pohán ěn protonovým gradientem (spot řeba ATP)
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
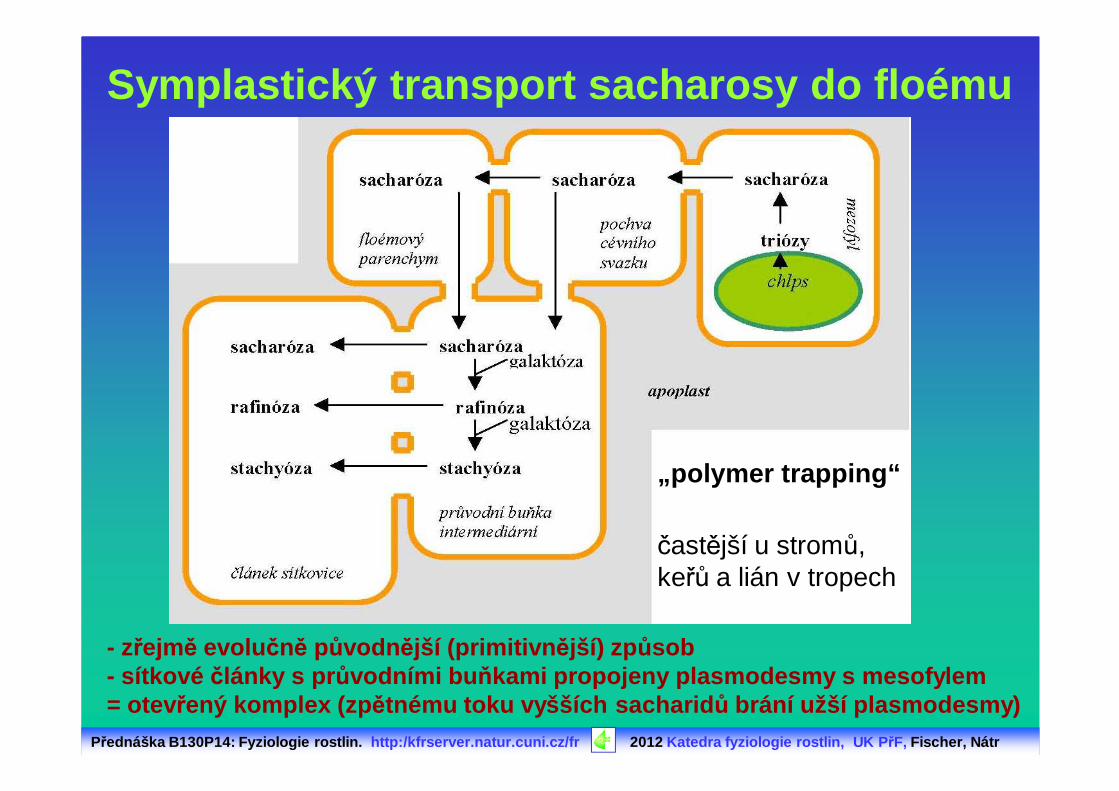
„polymer trapping“
častější u stromů, keřů a lián v tropech
- zřejmě evolu čně původn ější (primitivn ější) způsob- sítkové články s pr ůvodními bu ňkami propojeny plasmodesmy s mesofylem= otev řený komplex (zp ětnému toku vyšších sacharid ů brání užší plasmodesmy)
Symplastický transport sacharosy do floému
Přednáška B130P14: Fyziologie rostlin. http:/kfrserver.natur.cuni.cz/fr 2012 Katedra fyziologie rostlin, UK P řF, Fischer, Nátr
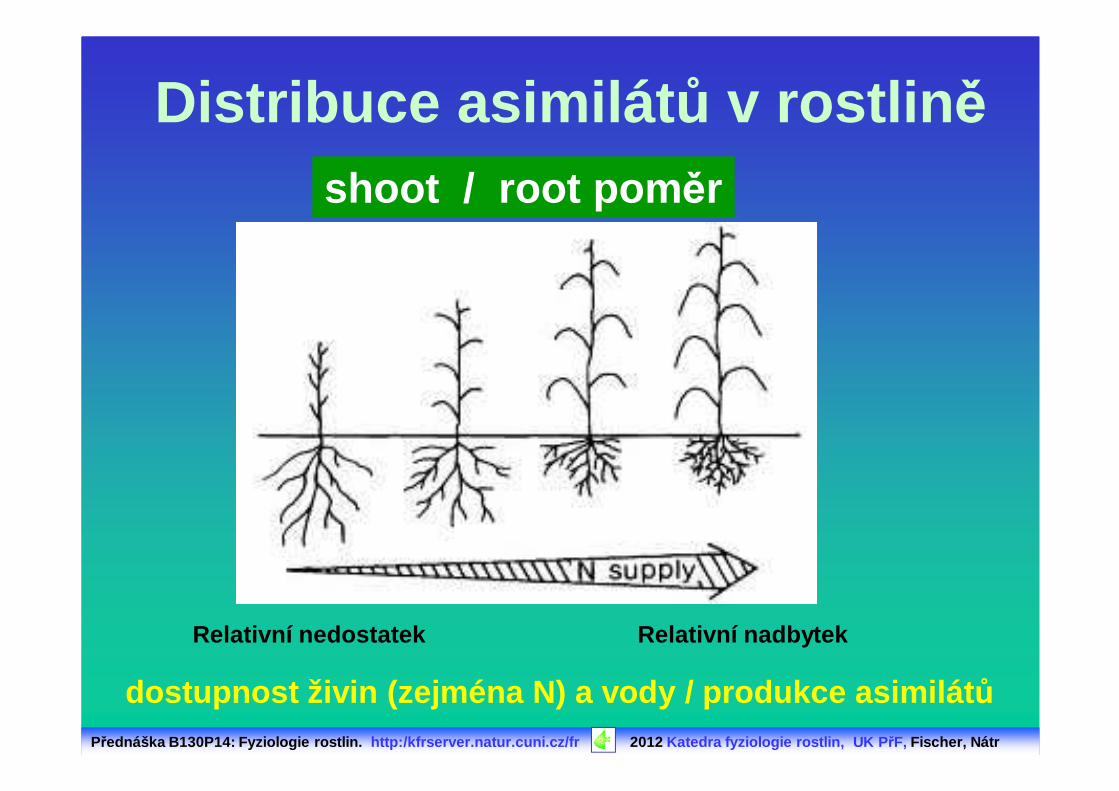
Distribuce asimilát ů v rostlin ěshoot / root pom ěr
Relativní nedostatek
dostupnost živin (zejména N) a vody / produkce asimil átů
Relativní nadbytek









![FIXACE BIOLOGICKÝCH VZORKŮ - CORE©m hodnocení jednotlivých fixátorů. 7)Výsledky vyhodnoťte a proveďte diskusi. DOPORUČENÁ LITERATURA: [1] BERNSTEIN, Matt A, Kevin Franklin](https://img.pdfslide.tips/doc/110x75/5cbfc02188c99337188bc1e8/fixace-biologickych-vzorku-core-m-hodnoceni-jednotlivych-fixatoru-7vysledky.jpg)



















