Embed Size (px)
Citation preview

Original Article
Canine mesenchymal stem cells show antioxidantproperties against thioacetamide-induced liver injuryin vitro and in vivo
Luiz Fernando Quintanilha,1,4 Taro Takami,1 Yoshikazu Hirose,1 Koichi Fujisawa,2
Yasuhiko Murata,1 Naoki Yamamoto,1,3 Regina Coeli dos Santos Goldenberg,4 Shuji Terai1
and Isao Sakaida1,2
1Department of Gastroenterology and Hepatology, 2Center for Reparative Medicine, Yamaguchi UniversityGraduate School of Medicine, 3Yamaguchi University Health Administration Center, Yamaguchi University, Ube,Japan; and 4Carlos Chagas Filho Biophysics Institute, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil
Aim: To overcome current limitations of therapy for liverdiseases, cell-based therapies using mesenchymal stemcells (MSC) have been attempted through basic and clinicalapproaches. Oxidative stress is a crucial factor in hepatology,and reactive oxygen species (ROS) are well-established mol-ecules responsible for its deleterious effects. The antioxidantproperties of MSC were recently demonstrated, and thereforewe examined the antioxidant activity of canine MSC (cMSC),their effects on isolated hepatocytes in vitro and their cura-tive potential against thioacetamide (TAA)-induced liver injuryin vivo.
Methods: To evaluate the ability of cMSC to challenge oxi-dative stress, cell viability, cytotoxicity and ROS were mea-sured in cultured cMSC treated with TAA. Also, cMSC wereco-cultured with hepatocytes in the same injury condition,and the ROS level was measured exclusively in hepatocytes.Finally, to verify the curative potential of cMSC, 2.0 × 106 cellsor phosphate-buffered saline were injected systemically innon-obese diabetic/severe combined immunodeficiency mice
that received TAA injections twice a week for 13 weeks. Wethen evaluated histological parameters, serum injury markersand redox homeostasis.
Results: cMSC overcame TAA-induced oxidative stress invitro, as shown by increased viability and lower cytotoxicityand ROS levels. Moreover, hepatocytes co-cultured withcMSC also showed decreased cellular ROS. The in vivo studyshowed that mice treated with cMSC presented with anameliorated histological pattern, suppressed fibrosis, lowerserum injury marker levels and better oxidative parameters.
Conclusion: We concluded that cMSC injection reduceTAA-induced liver injury through antioxidant activities andhepatoprotective effects, showing a curative potential in liverdiseases.
Key words: liver, mesenchymal stem cells, NF-E2-relatedfactor 2, oxidative stress, reactive oxygen species
INTRODUCTION
LIVER DISEASES ARE highly prevalent in the popula-tion worldwide. Currently, despite different alterna-
tives that have been tested, the standard treatment forend-stage chronic liver disease that is available and
effective is whole liver transplantation. However, livertransplantation has serious limitations such as donorscarcity, immunological incompatibilities, high cost,and significant morbidity and mortality associated withthe procedure.1–3 Additionally, considerable long-termside-effects have been reported.4–7 Given the inherentlimitations of this treatment, alternative therapies areurgently needed.
In recent years, cell-based therapy, especially therapyusing bone marrow cells (BMC), has emerged as analternative to improve damaged liver function. Anincreasing number of studies have been publishedshowing evidence of therapeutic effects of BMC in liverdiseases,8–15 including clinical trials worldwide.16–20
Correspondence: Dr Taro Takami, Department of Gastroenterologyand Hepatology, Yamaguchi University Graduate School of Medicine,1-1-1 Minami-Kogushi, Ube, Yamaguchi 755-8505, Japan. Email:[email protected] 24 February 2013; revision 19 June 2013; accepted 8 July2013.
bs_bs_banner
Hepatology Research 2014; 44: E206–E217 doi: 10.1111/hepr.12204
© 2013 The Japan Society of HepatologyE206

The interest in this particular cell niche comes fromprevious reports showing the presence of donor-derivedcells in the liver of bone marrow transplant recipi-ents.21,22 This observation, which has been proven inanimal models,23,24 showed potential cross-talk betweenBMC and the liver under certain conditions. Among thedifferent cell types found in bone marrow, mesenchy-mal stem cells (MSC) have shown promising results intissue regeneration.8,12,14,25 These cells can be easily iso-lated from the patient, cultured, expanded and used asan autologous cell-based therapy.
Although promising results have been shown, impor-tant questions remain. For example, no consensus existsabout the mechanisms of liver repair by BMC infusion.This topic constitutes one of the most debated issues inregenerative medicine.
Recently, oxidative stress has been shown to be animportant factor in liver diseases such as liver fibrosis,cirrhosis, viral hepatitis, hepatocellular carcinoma andothers.26–30 Oxidative stress is partly generated by reac-tive oxygen species (ROS), which are produced by dif-ferent pathways such as NAD(P)H oxidases, xenobioticmetabolism, mitochondrial leakage and cytochromeP450 activity, which lead to hepatocyte damagethrough lipid peroxidation and alkylation of proteins,nucleic acids and lipids.31–33 Although the liver itselfhas an efficient antioxidant defense system, sometimesthis system is not sufficient to repair the damage and/oran imbalance exists between oxidative stress elimina-tion and production. MSC were recently reported tohave an antioxidant ability that may contribute to oxi-dative stress resolution.34 Importantly, NF-E2-relatedfactor 2 (Nrf2) has emerged as a crucial transcriptionfactor that is capable of inducing a large array ofenzymes involved in oxidative stress resolution.35,36
Maintenance of the cellular redox balance by Nrf2 hasmultiple activation pathways and has been shownto be essential in combating many inflammatorydiseases.37–43 Some molecules such as all-trans retinoicacid (ATRA) and tert-butylhydroquinone (t-BHQ) haveshown the ability to significantly reduce (ATRA) orinduce (t-BHQ) Nrf2 functions, which modify theexpression of antioxidant response element (ARE)-driven genes.44,45
Thioacetamide (TAA) is one the most popular chemi-cal toxins used worldwide to generate experimental liverinjury.46,47 Its toxicity results from its biotransformationby a mixed-function oxidase system (e.g. cytochromeP450 enzymes and FAD monooxygenases), which leadsto the formation of reactive metabolites includingROS.48–53 ROS production resulting from TAA adminis-
tration is related to the consequences of oxidativedamage including lipid peroxidation.54,55
Given the above concerns and the recent evidence forthe effectiveness of cell-based therapy in liver diseasesinvolving oxidative stress, we hypothesized that MSCcould ameliorate the deleterious effects of TAA-inducedoxidative stress injury in liver. In this study, we testedthe ability of canine MSC (cMSC) to overcome TAA-induced oxidative stress in vitro and verified whetherthese cells could protect against oxidative stress damagein isolated hepatocytes. In addition, we evaluatedwhether cMSC could reduce the effects of TAA-inducedchronic injury in vivo. An important note is that fewstudies have used cells derived from medium-sizedanimals. Results from such studies will be important forsupporting new clinical trials.
METHODS
Animals and ethics
ALL ANIMALS USED in this study were maintainedand used in accordance with the Animal Care
Guidelines of Yamaguchi University. Non-obesediabetic/severe combined immunodeficiency (NOD/SCID) mice were purchased from Kyudo (Saga, Japan).
Cells and culture conditionsCanine bone marrow-derived MSC (Cyagen, Sunnyvale,CA, USA) were seeded onto 10-cm dishes (Iwaki, Tokyo,Japan) and cultured in OriCell Mesenchymal Stem CellGrowth Medium (Cyagen) supplemented with 10%fetal bovine serum (FBS; Life Technologies, GrandIsland, NY, USA), penicillin (100 U/mL; Life Technolo-gies) and streptomycin (100 μg/mL; Life Technologies)in a 5% CO2 incubator at 37°C. After seeding, non-adherent cells were removed when the medium wasreplaced. The culture medium was changed every 2 days.Cells from the fourth to sixth passages were used in thisstudy.
Cellular characterizationAdherent cells were dissociated with 0.05% trypsin-ethylenediaminetetraacetic acid (Life Technologies) andresuspended in Dulbecco’s modified Eagle’s medium(DMEM; Life Technologies) containing 10% FBS. Then,they were washed once with phosphate-buffered saline(PBS; Life Technologies) and incubated in PBS contain-ing 2% canine serum (AbD Serotec, Oxford, UK) for20 min on ice. After incubation, cells were incubatedfor 20 min on ice with monoclonal antibodies against
Hepatology Research 2014; 44: E206–E217 Antioxidant activity of mesenchymal cells in liver E207
© 2013 The Japan Society of Hepatology

CD11b (AbD Serotec), CD14-PE-Cy7 (BD Biosciences,San Jose, CA, USA), CD29-PE (Abcam, Cambridge, UK),CD34-PE (Abcam), CD44-PE-Cy7 (Biolegend, SanDiego, CA, USA), CD45-e-fluor (ebioscience, San Diego,CA, USA), CD90-APC (ebioscience) or CD133-PercP-eFluor 710 (ebioscience). Secondary detection of theCD11b antibody was performed using goat polyclonalsecondary antibody to mouse IgG–H&L (DyLight 488;Abcam). Isotype-identical antibodies were used as con-trols. Flow cytometry analyses were performed utilizingGallios equipment (Beckman Coulter, Danvers, MA,USA). Propidium iodide (PI; Sigma-Aldrich, St Louis,MO, USA) was used to exclude dead cells from analyses.Assessment of each sample was performed at least intriplicate. Data were analyzed using Kaluza software(Beckman Coulter). To confirm the differentiationpotential, the cells were grown in osteogenic andadipogenic canine differentiation media (Cell Applica-tions, San Diego, CA, USA) in accordance with themanufacturer’s instructions. After 2 weeks, lipid drop-lets were observed following oil red O staining (Sigma-Aldrich), and deposition of bone mineral was observedfollowing alizarin red staining (AppliChem, Darmstadt,Germany).
CM-DiI labelingCultured cMSC were CM-DiI stained following themanufacturer’s instructions. In brief, cMSC were sus-pended in 2 μM CM-DiI (Molecular Probes, Eugene,OR, USA) and incubated for 5 min at 37°C followed byan additional incubation at 4°C for 15 min. Then,labeled cells were washed three times and resuspendedin PBS. Cell labeling was confirmed with fluorescencemicroscopic observation.
Monoculture systemTo evaluate the ability of cMSC to challenge TAA-induced oxidative stress, cMSC were seeded onto 96-or six-well plates (Corning, NY, USA) at a density of1.0 × 105 cells/cm2 in DMEM supplemented with 10%FBS, penicillin (100 U/mL) and streptomycin (100 μg/mL) in a 5% CO2 incubator at 37°C. Non-adherentcells were removed by washing with PBS. The mediumwas replaced with non-supplemented DMEM withor without 50 mM TAA with additional overnightincubation.
Cells in 96-well plates were washed three times, andthe viability was measured indirectly using CellTiter 96AQueous One Solution (Promega, Madison, WI, USA).Cytotoxicity was measured with lactate dehydrogenase(LDH) quantification using the Cytotox 96 Non-
Radioactive Cytotoxicity Assay (Promega) according tothe manufacturer’s instructions. Values were normalizedusing media exposed to the same culture conditionswithout cells. Finally, to quantify cellular ROS, cells insix-well plates were stained with the CellRox Reagent(Molecular Probes) following the manufacturer’sinstructions. Stained live cells were quantified usingGallios equipment. Additionally, to reduce or increasethe antioxidant response by Nrf2, cMSC were pretreatedfor 1 h with non-supplemented DMEM containing10 μM ATRA (Sigma-Aldrich) or 10 μM t-BHQ (Sigma-Aldrich), respectively. ATRA is an inhibitor, and t-BHQis an inducer of ARE-driven gene induction that is medi-ated by Nrf2. After incubation, the same volume ofnon-supplemented DMEM was added (final concentra-tion, 50 mM TAA). After overnight incubation, we per-formed a viability test, LDH quantification and ROSmeasurement as described above.
Co-culture systemFor co-culture, primary green fluorescent protein (GFP)positive murine hepatocytes were isolated fromC57BL/6 Tg14 (act-EGFP) OsbY01 mice as previouslydescribed56 (with modifications). In brief, livers wereperfused via the portal vein with pre-warmed liver per-fusion medium (Life Technologies) at a flow rate of6 mL/min for 5 min with additional perfusion with0.05% type IV collagenase (Sigma-Aldrich) containing20 U/mL deoxyribonuclease I (Sigma-Aldrich) at 37°C.Whole livers were carefully harvested, the gall bladderwas removed and further mechanical digestion was per-formed in a glass dish. The released cells were filteredthrough 100-μm nylon mesh (BD Falcon, San Jose, CA,USA) and washed twice with centrifugation at 50 g at4°C for 1 min. Finally, the pellet was resuspended inPBS and diluted in stock isotonic Percoll solution (GELifesciences, Uppsala, Sweden) with further centrifuga-tion at 60 g for 10 min. The pellet was washed twicewith PBS, and the viability and cell number were deter-mined with a Trypan blue exclusion test (MolecularProbes). Then, approximately 3–5 × 105 isolated hepa-tocytes were seeded onto collagen I-coated six-wellplates (Life Technologies) using rodent hepatocyteplating medium (Zenbio, Durham, NC, USA) in a 5%CO2 incubator at 37°C. After 4 h, the same numberof CM-DiI-labeled cMSC was seeded in experimentalplates (monoculture was used as control). After over-night incubation, non-adherent cells were removed bywashing with PBS, and the medium was replaced withrodent hepatocyte maintenance medium (Zenbio) withor without 50 mM TAA.
E208 L. F. Quintanilha et al. Hepatology Research 2014; 44: E206–E217
© 2013 The Japan Society of Hepatology

After overnight incubation, hepatocytes in monocul-ture or direct co-culture were analyzed. For ROS quan-tification, six-well plates were stained with the CellRoxDeep Red Reagent for 30 min. After staining, cells wereharvested, centrifuged for 1 min at 50 g, and resus-pended in PBS containing 1% FBS and 2 μg/mL PIfor flow cytometry analysis. The cellular ROS levelsof PI negative live GFP positive cells were analyzed.Figure 3(a–c) illustrates our experimental strategy.
Experimental model of liver injuryFor liver injury induction, 6-week-old female NOD/SCID mice (n = 16) were given TAA (Sigma-Aldrich)injection (250 mg/kg i.p.) twice a week for 13 weeks.Beginning on the 10th week, 2.0 × 106 cMSC diluted in200 μL PBS (cell-treated group; n = 8) or the samevolume of PBS only (non-treated group; n = 8) wereslowly injected weekly via the tail vein using a metal hubneedle (31/2″/2) and a 250-μL syringe (Hamilton,Reno, NV, USA). Three days after the last cMSC/PBSinjections, mice were killed.
Biochemical analysesSerum alanine aminotransferase (ALT), aspartate ami-notransferase (AST) and LDH were measured in dupli-cate using an automated analyzer for clinical chemistry(SPOTCHEM EZ SP-4430; Arkray, Kyoto, Japan).
Histological stainingParaffin-embedded liver samples were sectioned (5 μm)and stained with hematoxylin–eosin and Sirius redaccording to standard protocols.
Fibrosis levelHistomorphometry was performed using an imagingsystem coupled to a fluorescence microscope (BiorevoBZ9000; Keyence, Osaka, Japan). The fibrosis index wasestimated by the percent of the area that was stainedwith Sirius red of the total area of the histological fields,which were examined with a BZ Analyzer II (Keyence).
Hyaluronic acid (HA), procollagen N-terminalpeptide (PIIIP) and AST/ALT ratio are valuable markersfor fibrosis measurement.57 Here, serum HA and PIIIPlevels were measured by using an enzyme-linkedimmunosorbent assay (ELISA; Mybiosource.com, SanDiego, CA, USA) in accordance with the manufacturer’sinstructions.
Antioxidant effects of cMSCTotal antioxidant activity was measured in serumsamples using an antioxidant assay kit (Cayman Chemi-
cals, Ann Arbor, MI, USA), which is based on theability of all antioxidant components in the sampleto inhibit the oxidation of ABTS (2,2′-Azino-di-[3-ethylbenzthiazoline sulphonate]) to ABTS+ bymetmyoglobin. The amount of ABTS+ produced wasmonitored by reading the absorbance at 750 nm usingInfinite M200 (Tecan, Männedorf, Switzerland). Thecapacity of antioxidants in the sample to prevent ABTSoxidation was compared with that of TROLOX, a water-soluble tocopherol analog. The result is presented as thepercent of the maximum antioxidant activity.
Lipid peroxidation was assessed in liver tissues using amalondialdehyde (MDA) assay kit (Abcam), which isbased on colorimetric quantification of MDA, a naturalproduct of lipid peroxidation in the samples.
Statistical analysisData were analyzed using Student’s paired t-test or one-way ANOVA as appropriate. Values of P < 0.05 were con-sidered statistically significant. Data are presented as themean 1 standard deviation.
RESULTS
Cellular characterization
THE CELLS USED in this study adhered to plasticand showed homogeneous distribution with a
fibroblastoid shape (Fig. 1a). Flow cytometry analysesshowed that cultured cMSC were positive for CD29,CD44 and CD90. On the other hand, these cellswere negative for the pan-leukocyte marker CD45 andthe monocyte/macrophage marker CD11b (Fig. 1b).These cells also had the potential to differentiate intoadipogenic and osteogenic lineages (Fig. 1c,d), indicat-ing a typical MSC phenotype.58
cMSC have a high capacity to overcomeTAA-induced oxidative stress in vitroSurprisingly, when cultured in medium containing TAA,cMSC exhibited an elevated ability to resist this condi-tion by showing lower levels of LDH release (cytotoxic-ity) into the culture medium (P < 0.001), consistentwith better viability (MTS assay, P < 0.001) and, also,they showed a lower cellular ROS level (P < 0.001).Additionally, because the Nrf2 pathway is considered tobe an important factor in oxidative stress protection andresolution,44 we used ATRA to abrogate and t-BHQ toinduce the antioxidant effects mediated by Nrf2. Inthe presence of ATRA, pre-treated cMSC showed higherLDH release (P < 0.05), lower cell viability (P < 0.001)
Hepatology Research 2014; 44: E206–E217 Antioxidant activity of mesenchymal cells in liver E209
© 2013 The Japan Society of Hepatology
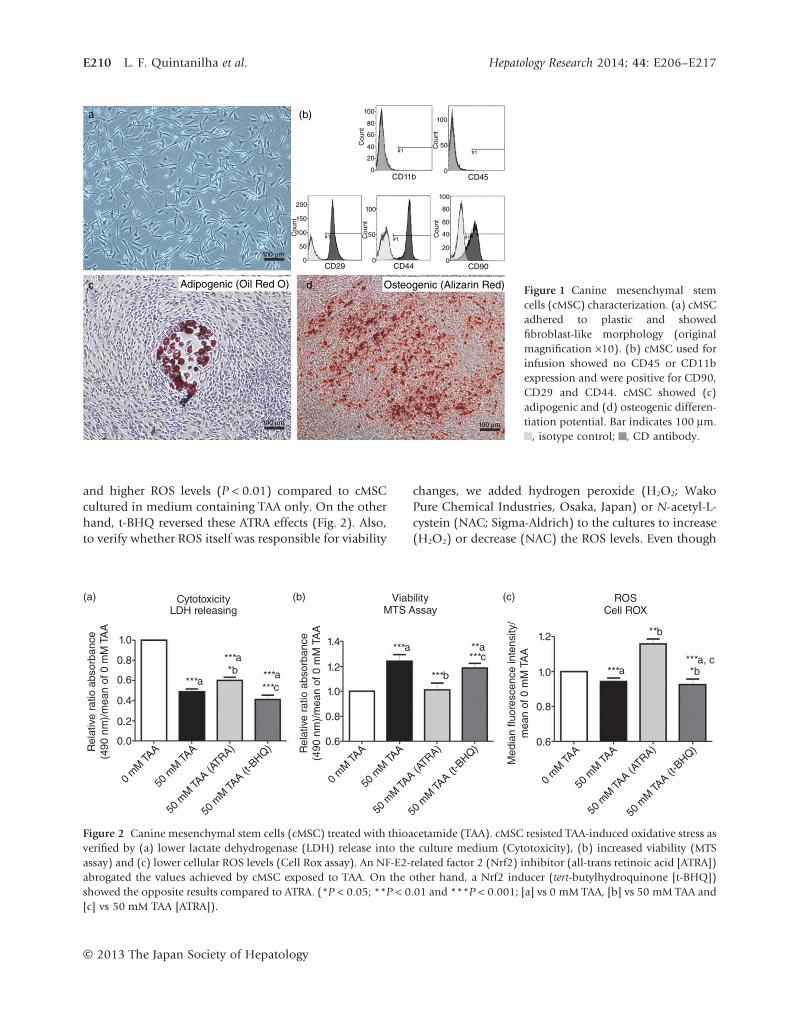
and higher ROS levels (P < 0.01) compared to cMSCcultured in medium containing TAA only. On the otherhand, t-BHQ reversed these ATRA effects (Fig. 2). Also,to verify whether ROS itself was responsible for viability
changes, we added hydrogen peroxide (H2O2; WakoPure Chemical Industries, Osaka, Japan) or N-acetyl-L-cystein (NAC; Sigma-Aldrich) to the cultures to increase(H2O2) or decrease (NAC) the ROS levels. Even though
a
100 µm
100 µm 100 µm
c dAdipogenic (Oil Red O) Osteogenic (Alizarin Red)
(b) 100100
50
0
80
60
#1
#1 #1 #1
#140C
ount
Cou
nt
Cou
nt
Cou
ntC
ount
20isotype control
CD antibody
200
150
100
100
100
80
60
40
20
0
50
0
50
0
0CD11b
CD29 CD44 CD90
CD45
Figure 1 Canine mesenchymal stemcells (cMSC) characterization. (a) cMSCadhered to plastic and showedfibroblast-like morphology (originalmagnification ×10). (b) cMSC used forinfusion showed no CD45 or CD11bexpression and were positive for CD90,CD29 and CD44. cMSC showed (c)adipogenic and (d) osteogenic differen-tiation potential. Bar indicates 100 μm.
, isotype control; , CD antibody.
(a) CytotoxicityLDH releasing
Viability
MTS AssayROS
Cell ROX
1.0 1.4
1.2
1.2
1.0
0.8
0.6
1.0
0.8
0.6
***a
***a***a
***a
**b
*ba, c***
***b
***c**a
***a
***c
*b0.8
0.6
0.4
0.2
0.0
0 m
M TAA
50 m
M TAA
50 m
M TAA (A
TRA)
50 m
M TAA (t
-BHQ)
0 m
M TAA
50 m
M TAA
50 m
M TAA (A
TRA)
50 m
M TAA (t
-BHQ)
0 m
M TAA
50 m
M TAA
50 m
M TAA (A
TRA)
50 m
M TAA (t
-BHQ)R
ela
tive r
atio a
bsorb
ance
(490 n
m)/
mea
n o
f 0 m
M T
AA
Rela
tive r
atio a
bsorb
ance
(490 n
m)/
mea
n o
f 0 m
M T
AA
Media
n fl
uore
scence inte
nsity/
mean o
f 0
mM
TA
A
(b) (c)
Figure 2 Canine mesenchymal stem cells (cMSC) treated with thioacetamide (TAA). cMSC resisted TAA-induced oxidative stress asverified by (a) lower lactate dehydrogenase (LDH) release into the culture medium (Cytotoxicity), (b) increased viability (MTSassay) and (c) lower cellular ROS levels (Cell Rox assay). An NF-E2-related factor 2 (Nrf2) inhibitor (all-trans retinoic acid [ATRA])abrogated the values achieved by cMSC exposed to TAA. On the other hand, a Nrf2 inducer (tert-butylhydroquinone [t-BHQ])showed the opposite results compared to ATRA. (*P < 0.05; **P < 0.01 and ***P < 0.001; [a] vs 0 mM TAA, [b] vs 50 mM TAA and[c] vs 50 mM TAA [ATRA]).
E210 L. F. Quintanilha et al. Hepatology Research 2014; 44: E206–E217
© 2013 The Japan Society of Hepatology

ROS levels changed as expected, no direct associationwas observed with viability at the time points tested(data not shown).
cMSC protect hepatocytes against oxidativestress in vitroTo evaluate whether cMSC could exert antioxidanteffects on hepatocytes, we co-cultured both types ofcells (CM-DiI-labeled cMSC and GFP positive murinehepatocytes; Fig. 3a,b). Cellular ROS in GFP positivehepatocytes was evaluated with flow cytometric analysis(Fig. 3c). Interestingly, hepatocyte monoculture in TAAshowed lower levels of cellular ROS. The co-culturesystem induced further reduction in the cellular ROSlevel in hepatocytes treated with 50 mM TAA (Fig. 3d).
Cell-therapy ameliorates TAA-inducedliver injuryBiochemical analyses were performed to verify theextent of liver injury in TAA-treated NOD/SCID mice bymeasuring serum ALT, AST and LDH levels. The cell-treated group showed reduced levels of serum ALT(non-treated group vs cell-treated group, 356.1 1 48.1 vs
286.4 1 69.3 U/L; P < 0.05) and AST (non-treated groupvs cell-treated group, 553.0 1 174.9 vs 372.8 1 71.1 U/L;P < 0.05). LDH also tended to be lower in the cell-treated group, but the difference was not statisticallysignificant (non-treated group vs cell-treated group,880.0 1 164.9 vs 695.6 1 305.8 U/L; P = 0.06) (Fig. 4).
In accordance, tissues harvested from non-treatedmice showed more necrotic areas and increased inflam-matory infiltration compared to the cell-treated group(Fig. 4d,e).
Fibrosis quantificationMorphometric analysis showed a decrease in the livercollagen content in the cell-treated group (5.1 1 2.9%)compared to the non-treated group (8.3 1 1.7%)(Fig. 5a,b). Moreover, the cell-treated group exhibitedbetter indirect fibrosis parameters such as HA (non-treated vs cell-treated group, 40.0 1 7.8 vs 29.7 111.6 ng/mL, P < 0.05; Fig. 5c), PIIIP (non-treated vscell-treated group, 71.7 1 3.6 vs 56.9 1 14.0 pg/mL,P = 0.06; Fig. 5d) and AST/ALT ratio (non-treated vscell-treated group, 1.5 1 0.3 vs 1.3 1 0.03, P < 0.05;Fig. 5e).
a
(c) (d)
1.0
***a
***a,b
0.8
0.6
0.4
0.2
0.0
0 mM TAA 50 mM TAAM
onoc
ultur
e
Mon
ocult
ure
Co-cu
lture
Cell ROX
Med
ian
fluor
esce
nce
inte
nsity
/m
ean
of M
onoc
ultu
re 0
mM
TA
A
[A] GFP
100 µm 100 µm
GFP
[B] Cell Rox [B] Cell Rox
40
D B
C C
30
10
5
0100 101
Cell Rox
All 1,405 80.5 100.00 1.49C 101 0.58 7.19 4.89
102 103
100 101 102 103
100 101
Cell Rox102 103
20
Cou
nt
Cou
nt
10
5
0
Cou
nt
10
0
Strategy:
GFP+ROS
Monoculture Co-culture
b
Gate Number %Total %Gated X-MedAll 1,834 22.98 100.00 0.38C 3 0.04 0.16 7.05
Gate Number %Total %Gated X-Med
Figure 3 Co-culture with canine mes-enchymal stem cells (cMSC) protectedhepatocytes from oxidative stress.Murine green fluorescent protein (GFP)positive hepatocytes (green) were cul-tured alone or in co-culture withCM-DiI-labeled cMSC (red). (a) Brightfield and (b) fluorescence microscopyof the co-culture system are shown. (c)Strategy to analyze GFP positive cells.(d) Cell ROX analysis of GFP positivecells revealed that hepatocytes havereduced reactive oxygen species (ROS)levels when cultured in the presenceof thioacetamide (TAA). When co-cultured with cMSC, they showed afurther decrease in intracellular ROS(***P < 0.001; [a] vs monoculturewithout TAA and [b] vs monoculturewith TAA). Bar indicates 100 μm.
Hepatology Research 2014; 44: E206–E217 Antioxidant activity of mesenchymal cells in liver E211
© 2013 The Japan Society of Hepatology

Cell therapy improves redox homeostasisBased on our in vitro results, we investigated whethercMSC transplantation could ameliorate oxidative stressin animals with chronic TAA infusions. High total anti-oxidant activity was sustained in sera collected fromthe cell-treated group, whereas the non-treated groupshowed a clear decrease in this ability (non-treatedgroup vs cell-treated group, 26.9 1 19.2% vs 61.4 113.0% of maximum values; P < 0.001, Fig. 6a). In addi-tion, lipid peroxidation was lower in liver tissuesharvested from cell-treated mice, indicating lower sus-ceptibility to oxidation in tissue (non-treated group vscell-treated group, 23.8 1 4.2 vs 19.6 1 3.0 nmol/mg;P < 0.05, Fig. 6b).
DISCUSSION
MURINE EXPERIMENTAL MODELS are commonlyused to test new therapies for hepatic diseases,59
including cell-based therapy using bone marrow-derived cells, which have shown promising results.8–15
Among the different cell populations found in bonemarrow, MSC have shown beneficial effects against liverdisease.8,12,14,25,60 Furthermore, MSC have advantagessuch as multiple tissue sources, fast proliferation, pos-sible use in autologous transplantation and in vitromanipulation. Also, MSC were recently shown topromote an antioxidant response in injured liver.34
Despite good results in basic studies and clinicaltrials,61–63 the mechanism of action of these cells is stillbeing discussed. Recently, many studies have linked oxi-
dative stress and development of liver diseases such asviral hepatitis, cirrhosis, hepatocellular carcinoma andothers.26–29 Here, we examined whether the antioxidantpotential demonstrated by MSC has effects in reducingTAA-induced liver injury.
Thioacetamide is a drug that is widely used in animalmodels.59 Because biotransformation of TAA producesoxidative damage associated with liver injury and thisdrug is usually used for systemic infusion, we examinedif MSC, which are also usually injected systemically,could provide resistance to the toxic effects produced byTAA. Surprisingly, rather than resistance alone, cMSCshowed a high level of tolerance to TAA (Fig. 2). Addi-tionally, when cMSC were pretreated with ATRA ort-BHQ, they showed opposite responses regarding cyto-toxicity, viability and ROS accumulation (Fig. 2). Con-sidering that ATRA inhibits and t-BHQ induces Nrf2effects both in vitro and in vivo,44,45 these results indicatethat cMSC have high antioxidant activity in vitroand suggest that the Nrf2 pathway may be involvedin this process. Consistent with this hypothesis,Mohammadzadeh et al. recently showed that inducedoverexpression of Nrf2 by MSC was able to promotereduction of cell death in hypoxia, serum deprivationand oxidative stress conditions. In this study, MSCwith transient overexpression of Nrf2 presented bettercell viability and reduced apoptosis levels.64 Moreover,Gorbunov et al. showed that MSC treated with lipopoly-saccharide, which induces inflammatory responsesincluding release of ROS, induce a number ofadaptive responses including induction and nuclear
(a)
d e
ALT
* *
AST LDH
p=0.06
Non-treated Cell-treated Non-treated Cell-treated Non-treated Cell-treated
400
400
300
200
500
600 1000
900
800
700
600
350
300U/L
U/L
U/L
250
200
(b) (c)Figure 4 Canine mesenchymal stemcells (cMSC) transplantation protectsagainst thioacetamide (TAA)-inducedliver injury. The injury markers (a)alanine aminotransferase (ALT), (b)aspartate aminotransferase (AST) and(c) lower lactate dehydrogenase (LDH)were measured to evaluate the extentof liver injury. The cell-treated groupshowed lower levels in all measure-ments. (d–e) Representative figuresshowing higher levels of parenchymaldisarrangement, diffuse cellular fataccumulation and necrosis in non-treated samples (d) compared withsamples from the cell-treated group(e) (original magnification ×20;*P < 0.05).
E212 L. F. Quintanilha et al. Hepatology Research 2014; 44: E206–E217
© 2013 The Japan Society of Hepatology

a
Non-treated
(b) (c)
(d) (e)
2.0*
1.5
1.0
0.5
0.0
Fibrosis Index
PIIIPELISA
AST/ALT ratio
*
*
Non-treated Cell-treated
p=0.0680
60
40
20
0
Non-treated
Non-treated
Cell-treated
Cell-treatedNon-treated Cell-treated
10 50
40
30
ng/m
lR
atio
pg/m
l
20
% s
tain
ed a
rea 8
6
4
2
0
Srtum Hyaluronic AcidELISA
Cell-treated
Figure 5 Fibrosis quantification. (a–b)Morphometry analysis of Sirius red-stained liver samples showed that thecell-treated group had lower levelsof fibrosis (original magnification ×4).Complementarily, serum collectedfrom cell-treated mice presented lowermean values for (c) hyaluronic acid(HA), (d) procollagen N-terminalpeptide (PIIIP) and (e) aspartate ami-notransferase (AST)/alanine amino-transferase (ALT) ratio (*P < 0.05).
(a)
****
80
60
30
20
10
0
40
20
0
Non-treated Cell-treated Non-treated Cell-treated
Rela
tive a
ntioxid
ant activity
MD
A e
quiv
ale
nt (n
mol/m
g)
(b)Total antioxidant activity
ELISA
Lipid peroxidation
MDA assay
Figure 6 Antioxidant activity in cell-treated mice. (a) Total antioxidantactivity was higher in the cell-treatedgroup compared to the non-treatedgroup (***P < 0.001). Additionally,(b) lipid peroxidation in liver tissuewas lower in the cell-treated groupcompared to the non-treated group(*P < 0.05).
Hepatology Research 2014; 44: E206–E217 Antioxidant activity of mesenchymal cells in liver E213
© 2013 The Japan Society of Hepatology

translocation of redox response elements such asnuclear factor-κB and Nrf2. They suggested that theprosurvival pathways that are activated in MSC in vitrocould be a part of an adaptive response employed bystromal cells under injury conditions.65
A direct and specific effect of ROS in viability wasruled out using H2O2 and NAC in cultures. As expected,these molecules increased (H2O2) and decreased (NAC)intracellular ROS, but no direct relationship betweenviability and ROS levels was seen at the time pointstested (data not shown). Additionally, to assess whethercMSC could potentially prevent oxidative stress in livercells, we utilized a co-culture model with murine hepa-tocytes and cMSC. In this experiment, we found a lowerROS level in co-cultured murine hepatocytes treatedwith TAA (Fig. 3), suggesting a hepatoprotective effect ofcMSC via antioxidant activity. Using a mouse primer forNrf2 with no cross-reactivity against canine samples insilico, we verified the higher amount of mRNA inco-cultured hepatocytes (Fig. S1). However, unexpect-edly, monocultured hepatocytes showed higher ROSlevels when TAA was absent from the culture medium,suggesting that hepatocytes have a mechanism similarto cMSC in the presence of TAA. The underlying mecha-nisms are now under investigation.
Our above in vitro results motivated us to test celltherapy using cMSC in TAA-induced liver injury inNOD/SCID mice. In chronic TAA-induced injury, theanimals that received cMSC infusions by tail veinshowed better results for the biochemical parameters.The serum injury markers (ALT, AST and LDH) werereduced with successive cell infusions, suggesting pro-tection of hepatocytes from necrosis and apoptosis(Fig. 4). Because ALT and AST are enzymes that revealhepatocyte damage, these results strongly support our invitro findings showing that cMSC have hepatoprotectiveeffects against TAA-induced injury. We cannot rule outthe possibility that infused cMSC may act systemically toaid the liver in its recovery. Consistent with our resultsand considering the possibility that Nrf2 may beinvolved in this process, Xu et al.66 demonstrated adelayed ALT decrease in sera from Nrf2-knockout miceafter treatment with hepatotoxin. Because Nrf2 is crucialfor induction of expression of a wide range of antioxi-dant genes, antioxidant activity may be essential forpromoting liver regeneration.
As already discussed, oxidative stress plays an impor-tant role in liver injury, and some authors have recentlydemonstrated that cell-based therapy can be an effectivetreatment. Recently, Cho et al. have shown that MSChave an antioxidant potential to ameliorate acute liver
injury induced by carbon tetrachloride.34 In a murinemodel of carbon tetrachloride-induced acute liverinjury, they found increased Nrf2 activity and lowerROS, ALT and AST levels in animals treated with synge-neic MSC.
Okuyama et al. reported that transgenic mice withhigh expression of thioredoxin, a small redox-activeprotein with antioxidant effects, showed not only ame-liorated liver injury but also decreased liver fibrosis.67,68
Consistent with this result, we showed that the possibleantioxidant activity of cMSC reduced necrotic andinflammatory areas (Fig. 4d,e) and fibrosis levels bymeasuring of different parameters (Fig. 5). We alsofound higher concentration of matrix metalloproteinase9 in liver tissues harvested from cell-treated group whatcan in part explain the results found in fibrosis analyses(Fig. S2).
In this present study, we confirmed that animals inthe cell-treated group had better redox homeostasis byshowing higher total serum antioxidant activity andlower lipid peroxidation in liver tissues (Fig. 6). ThecMSC infusions seemed to sustain normal overall totalantioxidant activity in these animals, which may explainthe decreased lipid peroxidation (Fig. 6b), serum injurymarkers (Fig. 4a–c) and histological findings in vivo(Figs 4,5). At this juncture, we can clearly see that cMSCcan act efficiently in combating oxidative stress in liver.
As far as we know, this study is the first to use acomplete approach (in vitro + in vivo) to evaluate therole of antioxidant activity in ameliorating liver injuryusing cells from a medium-sized animal. These resultsreveal potent antioxidant activity and hepatoprotectiveeffects of cMSC in vitro and in vivo and support morestudies examining the antioxidant activity of stem cellsto combat liver diseases.
In conclusion, we showed that cMSC can protecthepatocytes by reducing ROS damage induced by TAAboth in vivo and in vitro. These results suggest a potentialfor MSC treatment in several hepatic diseases.
ACKNOWLEDGMENTS
WE THANK MS Mariko Yamada, Ms IhokoFujimoto, Ms Yoko Fukusumi, Ms Hiromi Kurose
and Ms Kumie Ohta for technical experimental supportand Mr Bruno Diaz Paredes for graphic works. Thisstudy was supported by Grants-in-Aid for scientificresearch from the Japan Society for the Promotion ofScience (JSPS) (nos. 22390150 to S. T. and 24790703 toT. T.); Ministry of Health, Labor and Welfare, health andlabor sciences research grants; and the Japan Science and
E214 L. F. Quintanilha et al. Hepatology Research 2014; 44: E206–E217
© 2013 The Japan Society of Hepatology

Technology Agency (JST), the project of realization ofregenerative medicine (Highway program).
REFERENCES
1 Nussler A, Konig S, Ott M et al. Present status and perspec-tives of cell-based therapies for liver diseases. J Hepatol2006; 45: 144–59.
2 Stravitz RT. Critical management decisions in patients withacute liver failure. Chest 2008; 134: 1092–102.
3 Lee SW, Wang X, Chowdhury NR, Roy-Chowdhury J.Hepatocyte transplantation: state of the art and strategiesfor overcoming existing hurdles. Ann Hepatol 2004; 3:48–53.
4 Chung H, Kim KH, Kim JG, Lee SY, Yoon YH. Retinalcomplications in patients with solid organ or bone marrowtransplantation. Transplantation 2007; 83: 694–9.
5 Francoz C, Belghiti J, Durand J. Indications of liver trans-plantation in patients with complications of cirrhosis. BestPract Res Clin Gastroenterol 2007; 21: 175–90.
6 Patel H, Vogl DT, Aqui N et al. Posttransplantlymphoproliferative disorder in adult liver transplantrecipients: a report of seventeen cases. Leuk Lymphoma2007; 48: 885–91.
7 Tamsel S, Demirpolat G, Killi R et al. Vascular complica-tions after liver transplantation: evaluation with DopplerUS. Abdom Imaging 2007; 32: 339–47.
8 Cho KA, Ju SY, Cho SJ et al. Mesenchymal stem cellsshowed the highest potential for the regeneration ofinjured liver tissue compared with other subpopulations ofthe bone marrow. Cell Biol Int 2009; 33: 772–7.
9 Ishikawa T, Terai S, Urata Y et al. Administration offibroblast growth factor 2 in combination with bonemarrow transplantation synergistically improves carbon-tetrachloride-induced liver fibrosis in mice. Cell Tissue Res2007; 327: 463–70.
10 Higashiyama R, Inagaki Y, Hong YY et al. Bone marrow–derived cells express matrix metallopeptidases and con-tribute to regression of liver fibrosis in mice. Hepatology2007; 45: 213–22.
11 Sakaida I, Terai S, Yamamoto N et al. Transplantation ofbone marrow cells reduces CCl4-induced liver fibrosis inmice. Hepatology 2004; 40: 1304–11.
12 Yan Y, Xu W, Qian H et al. Mesenchymal stem cells fromhuman umbilical cords ameliorate mouse hepatic injury invivo. Liver Int 2009; 29: 356–65.
13 Ali G, Sadia M, Mohsin K et al. Nitric oxide augmentsmesenchymal stem cell ability to repair liver fibrosis.J Transl Med 2012; 10: 75.
14 Hardjo M, Miyazaki M, Sakaguchi M et al. Suppression ofcarbon tetrachloride-induced liver fibrosis by transplanta-tion of a clonal mesenchymal stem cell line derived fromrat bone marrow. Cell Transplant 2009; 18: 89–99.
15 Maeda M, Takami T, Terai S, Sakaida I. Autologous bonemarrow cell infusion suppress tumor initiation in
hepatocarcinogenic mice with liver cirrhosis. J GastroenterolHepatol 2012; 27: 104–11.
16 Chernykh ER, Starostina NM, Paltsev AI et al. Autologousbone marrow cells in treatment of cirrhosis of the liver. BullExp Biol Med 2007; 144: 640–5.
17 Couto BG, Goldenberg RC, da Fonseca LM et al. Bonemarrow mononuclear cell therapy for patients with cirrho-sis: a Phase 1 study. Liver Int 2011; 31: 391–400.
18 Lyra AC, Soares MB, da Silva LF et al. Infusion of autolo-gous bone marrow mononuclear cells through hepaticartery results in a short-term improvement of liver functionin patients with chronic liver disease: a pilot randomizedcontrolled study. Eur J Gastroenterol Hepatol 2010; 22:33–42.
19 Saito T, Okumoto K, Haga H et al. Stem cells and develop-ment potential therapeutic application of intravenousautologous bone marrow infusion in patients with alco-holic liver cirrhosis. Stem Cells Dev 2011; 20: 1503–10.
20 Terai S, Ishikawa T, Omori K et al. Improved liver func-tion in patients with liver cirrhosis after autologousbone marrow cell infusion therapy. Stem Cells 2006; 24:2292–8.
21 Theise ND, Nimmakayalu M, Gardner R et al. Liver frombone marrow in humans. Hepatology 2000; 32: 11–6.
22 Alison MR, Poulsom R, Jeffery R et al. Hepatocytes fromnon-hepatic adult stem cells. Nature 2000; 406 (6793):257.
23 Petersen BE, Bowen WC, Patrene KD et al. Bone marrow asa potential source of hepatic oval cells. Science 1999; 284(5417): 1168–70.
24 Theise ND, Badve S, Saxena R et al. Derivation of hepato-cytes from bone marrow cells in mice after radiation-induced myeloablation. Hepatology 2000; 31: 235–40.
25 Salem HK, Thiemermann C. Mesenchymal stromal cells:current understanding and clinical status. Stem Cells 2010;28: 585–96.
26 Clément S, Pascarella S, Negro F. Hepatitis C virus infec-tion: molecular pathways to steatosis, insulin resistanceand oxidative stress. Viruses 2009; 1: 126–43.
27 Tanikawa K, Torimura T. Studies on oxidative stress in liverdiseases: important future trends in liver research. Med MolMorphol 2006; 39: 22–7.
28 Ivanov AV, Smirnova OA, Ivanova ON, Masalova OV,Kochetkov SN, Isaguliants MG. Hepatitis C virus proteinsactivate NRF2/ARE pathway by distinct ROS-dependentand independent mechanisms in HUH7 cells. PLoS ONE2011; 6: e24957.
29 Zhu R, Wang Y, Zhang L, Guo Q. Oxidative stress and liverdisease. Hepatol Res 2012; 42: 741–9.
30 Cash WJ, McCance DR, Young IS et al. Primary biliary cir-rhosis is associated with oxidative stress and endothelialdysfunction but not increased cardiovascular risk. HepatolRes 2010; 40: 1098–106.
31 Parola M, Robino G. Oxidative stress-related moleculesand liver fibrosis. J Hepatol 2001; 35: 297–306.
Hepatology Research 2014; 44: E206–E217 Antioxidant activity of mesenchymal cells in liver E215
© 2013 The Japan Society of Hepatology

32 De Minicis S, Brenner DA. NOX in liver fibrosis. ArchBiochem Biophys 2007; 462: 266–72.
33 Weber LW, Boll M, Stampfl A. Hepatotoxicity and mecha-nism of action of haloalkanes: carbon tetrachloride as atoxicological model. Crit Rev Toxicol 2003; 33: 105–36.
34 Cho KA, Woo SY, Seoh JY, Han HS, Ryu KH. Mesenchymalstem cells restore CCl4-induced liver injury by anantioxidative process. Cell Biol Int 2012; 36: 1267–74.
35 Jaiswal AK. Nrf2 signaling in coordinated activation ofantioxidant gene expression. Free Radic Biol Med 2004; 36:1199–207.
36 Klaassen CD, Reisman SA. Nrf2 the rescue: effects of theantioxidative/electrophilic response on the liver. ToxicolAppl Pharmacol 2010; 244: 57–65.
37 Hochmuth CE, Biteau B, Bohmann D, Jasper H. Redoxregulation by Keap1 and Nrf2 controls intestinal stem cellproliferation in Drosophila. Cell Stem Cell 2011; 8: 188–99.
38 Surh YJ, Kundu JK, Na HK. Nrf2 as a master redox switch inturning on the cellular signaling involved in the inductionof cytoprotective genes by some chemopreventivephytochemicals. Planta Med 2008; 74: 1526–39.
39 Brigelius-Flohe R, Flohe L. Basic principles and emergingconcepts in the redox control of transcription factors.Antioxid Redox Signal 2011; 15: 2335–81.
40 Hayes JD, McMahon M. NRF2 and KEAP1 mutations: per-manent activation of an adaptive response in cancer.Trends Biochem Sci 2009; 34: 176–88.
41 Kensler TW, Wakabayashi N, Biswal S. Cell survivalresponses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu Rev Pharmacol Toxicol 2007; 47:89–116.
42 Lee JM, Johnson JA. An important role of Nrf2-AREpathway in the cellular defense mechanism. J Biochem MolBiol 2004; 37: 139–43.
43 Taguchi K, Motohashi H, Yamamoto M. Molecular mecha-nisms of the Keap1-Nrf2 pathway in stress response andcancer evolution. Genes Cells 2011; 16: 123–40.
44 Magesh S, Chen Y, Hu L. Small molecule modulatorsof Keap1-Nrf2-ARE pathway as potential preventive andtherapeutic agents. Med Res Rev 2012; 32: 687–726.
45 Wang XJ, Hayes JD, Henderson CJ, Wolf CR. Identificationof retinoic acid as an inhibitor of transcription factor Nrf2through activation of retinoic acid receptor alpha. Proc NatlAcad Sci U S A 2007; 104: 19589–94.
46 Li X, Benjamin IS, Alexander B. Reproducible productionof thioacetamide-induced macronodular cirrhosis in therat with no mortality. J Hepatol 2002; 36: 488–93.
47 Ledda-Collumbano GM, Coni P, Curto M et al. Inductionof two different modes of cell death, apoptosis and necro-sis, in rat liver after single dose of thioacetamide. Am JPathol 1991; 139: 1099–109.
48 Zaragoza A, Andres D, Sarrion D, Cascales M. Potentiationof thioacetamide hepatotoxicity by phenobarbital pretreat-ment in rats. Inducibility of FAD monooxygenase systemand age effect. Chem Biol Interact 2000; 124: 87–101.
49 Sanz N, Diez-Fernandez C, Andres D, Cascales M. Hepato-toxicity and aging: endogenous antioxidant systems inhepatocytes from 2-, 6-, 12-, 18- and 30-month-old ratsfollowing a necrogenic dose of thioacetamide. BiochimBiophys Acta 2002; 1587: 12–20.
50 Chilakapati J, Korrapati MC, Hill RA, Warbritton A,Latendresse JR, Mehendale HM. Toxicokinetics and toxicityof thioacetamide sulfoxide: a metabolite of thioacetamide.Toxicology 2007; 230: 105–16.
51 Chilakapati J, Shankar K, Korrapati MC, Hill RA,Mehendale HM. Saturation toxicokinetics of thioace-tamide: role in initiation of liver injury. Drug Metab Dispos2005; 33: 1877–85.
52 Andres D, Sanchez-Reus I, Bautista M, Cascales M. Deple-tion of Kupffer cell function by gadolinium chlorideattenuates thioacetamide-induced hepatotoxicity. Expres-sion of metallothionein and HSP70. Biochem Pharmacol2003; 66: 917–26.
53 Hajovsky H, Hu G, Koen Y et al. Metabolism and toxicity ofthioacetamide and thioacetamide S-oxide in rat hepato-cytes. Chem Res Toxicol 2012; 25: 1955–63.
54 Lotkova H, Cervinkova Z, Kucera O, Rousar T, Krivakova P.S-Adenosylmethionine exerts a protective effect againstthioacetamide-induced injury in primary cultures of rathepatocytes. Altern Lab Anim 2007; 35: 363–71.
55 Stanková P, Kucera O, Lotková H, Roušar T, Endlicher R,Cervinková Z. The toxic effect of thioacetamide on rat liverin vitro. Toxicol In Vitro 2010; 24: 2097–103.
56 Seglen PO. Preparation of isolated rat liver cells. MethodsCell Biol 1976; 13: 29–83.
57 Kawada N. Evolution of hepatic fibrosis research. HepatolRes 2011; 41: 199–208.
58 Dominici M, Le Blanc K, Mueller I et al. Minimal criteriafor defining multipotent mesenchymal stromal cells. TheInternational Society for Cellular Therapy position state-ment. Cytotherapy 2006; 8: 315–7.
59 Tuñón MJ, Alvarez M, Culebras JM, González-Gallego J. Anoverview of animal models for investigating the pathogen-esis and therapeutic strategies in acute hepatic failure.World J Gastroenterol 2009; 15: 3086–98.
60 Iwamoto T, Terai S, Hisanaga T et al. Bone-marrow-derivedcells cultured in serum-free medium reduce liver fibrosisand improve liver function in carbon-tetrachloride-treatedcirrhotic mice. Cell Tissue Res 2013; 351: 487–95.
61 Takami T, Terai S, Sakaida I. Novel findings for thedevelopment of drug therapy for various liver diseases:current state and future prospects for our liver regenerationtherapy using autologous bone marrow cells for decom-pensated liver cirrhosis patients. J Pharmacol Sci 2011; 115:274–8.
62 Takami T, Terai S, Sakaida I. Stem cell therapy in chronicliver disease. Curr Opin Gastroenterol 2012; 28: 203–8.
63 Terai S, Sakaida I. Autologous bone marrow cell infusiontherapy for liver cirrhosis patients. J Hepatobiliary PancreatSci 2011; 18: 23–5.
E216 L. F. Quintanilha et al. Hepatology Research 2014; 44: E206–E217
© 2013 The Japan Society of Hepatology

64 Mohammadzadeh M, Halabian R, Gharehbaghian A et al.Nrf-2 overexpression in mesenchymal stem cells reducesoxidative stress-induced apoptosis and cytotoxicity. CellStress Chaperones 2012; 17: 553–65.
65 Gorbunov NV, Garrison BR, McDaniel DP et al. Adaptiveredox response of mesenchymal stromal cells to stimula-tion with lipopolysaccharide inflammagen: mechanismsof remodeling of tissue barriers in sepsis. Oxid Med CellLongev 2013; 2013: 186795.
66 Xu W, Hellerbrand C, Köhler UA et al. The Nrf2 transcrip-tion factor protects from toxin-induced liver injury andfibrosis. Lab Invest 2008; 88: 1068–78.
67 Okuyama H, Nakamura H, Shimahara Y et al. Over-expression of thioredoxin prevents acute hepatitis causedby thioacetamide or lipopolysaccharide in mice. Hepatology2003; 37: 1015–25.
68 Okuyama H, Nakamura H, Shimahara Y et al. Over-expression of thioredoxin prevents thioacetamide-inducedhepatic fibrosis in mice. J Hepatol 2005; 42: 117–23.
SUPPORTING INFORMATION
ADDITIONAL SUPPORTING INFORMATION maybe found in the online version of this article at the
publisher’s website:
Figure S1 Relative quantification of NF-E2-relatedfactor 2 (Nrf2) mRNA in hepatocytes in co-cultureshowed higher values when compared to samples frommonoculture under thioacetamide (TAA) condition(*P < 0.05).Figure S2 Enzyme-linked immunoassay revealed thatliver tissues harvested from cell-treated group presentedhigher concentration of matrix metalloproteinase 9(*P < 0.05).
Hepatology Research 2014; 44: E206–E217 Antioxidant activity of mesenchymal cells in liver E217
© 2013 The Japan Society of Hepatology





























