Embed Size (px)
Citation preview

CAPITULO 4
Fenomenos de Transporte: Membranas
En este capıtulo presentaremos la difusion ionica a traves de membranas,la cual es fundamental para regular numerosos procesos en los seres vivos.
Como sabemos, las membranas son sistemas basicos para el funcionamentode las celulas vivas y de los organismos. Antes de entrar en los detalles delos procesos de transporte a traves de membranas, vamos a recordar algunosaspectos de la estructura y el funcionamiento de estos sistemas.
Cada celula esta rodeada por una membrana que controla su contenidoquımico y sirve para aislar la celula del ambiente, para transportar sustanciasy para transmitir mensajes.
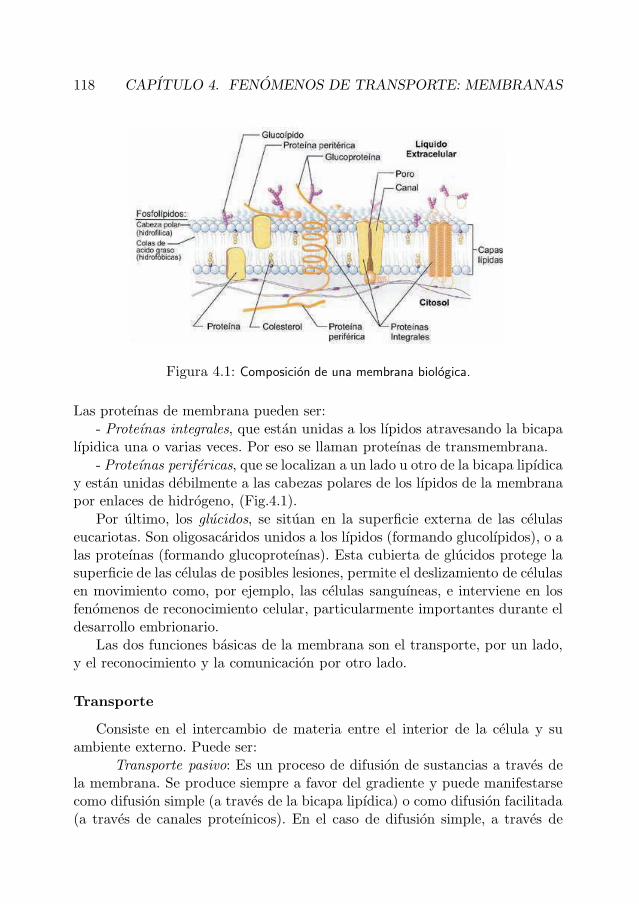
La membrana celular esta constituida por lıpidos, proteınas y glucidos enproporciones aproximadas de 40 %, 50 % y 10 %, respectivamente.
En la membrana de la celula eucariota existen tres tipos de lıpidos: fos-folıpidos, glucolıpidos y colesterol. Una parte de estas moleculas lipıdicas eshidrofila y la otra parte es hidrofoba por lo que cuando se encuentran en unmedio acuoso se orientan formando una bicapa lipıdica (Fig.4.1). La mem-brana tiene un cierto nivel de fluidez que depende de la temperatura y de sucomposicion. La presencia de lıpidos insaturados y de cadena corta favorecenel aumento de la fluidez, mientras que la presencia de colesterol endurece lasmembranas, reduciendo su fluidez y permeabilidad.
Las proteınas son los componentes de la membrana que desempenan lasfunciones especıficas de transporte y de comunicacion. Pueden girar alrededorde su eje y muchas de ellas pueden desplazarse lateralmente por la membrana.
118 CAPITULO 4. FENOMENOS DE TRANSPORTE: MEMBRANAS
Figura 4.1: Composicion de una membrana biologica.
Las proteınas de membrana pueden ser:- Proteınas integrales, que estan unidas a los lıpidos atravesando la bicapa
lıpidica una o varias veces. Por eso se llaman proteınas de transmembrana.
- Proteınas perifericas, que se localizan a un lado u otro de la bicapa lipıdicay estan unidas debilmente a las cabezas polares de los lıpidos de la membranapor enlaces de hidrogeno, (Fig.4.1).
Por ultimo, los glucidos, se situan en la superficie externa de las celulaseucariotas. Son oligosacaridos unidos a los lıpidos (formando glucolıpidos), o alas proteınas (formando glucoproteınas). Esta cubierta de glucidos protege lasuperficie de las celulas de posibles lesiones, permite el deslizamiento de celulasen movimiento como, por ejemplo, las celulas sanguıneas, e interviene en losfenomenos de reconocimiento celular, particularmente importantes durante eldesarrollo embrionario.
Las dos funciones basicas de la membrana son el transporte, por un lado,y el reconocimiento y la comunicacion por otro lado.
Transporte
Consiste en el intercambio de materia entre el interior de la celula y suambiente externo. Puede ser:
Transporte pasivo: Es un proceso de difusion de sustancias a traves dela membrana. Se produce siempre a favor del gradiente y puede manifestarsecomo difusion simple (a traves de la bicapa lipıdica) o como difusion facilitada(a traves de canales proteınicos). En el caso de difusion simple, a traves de

4.1. DIFUSION PURA A TRAVES DE MEMBRANAS 119
la bicapa lipıdica se realiza el transporte pasivo de moleculas lipıdicas, comolas hormonas esteroideas, una gran parte de anestesicos y sustancias apolarescomo el oxıgeno y el nitrogeno atmosferico. El transporte pasivo a traves decanales proteınicos da paso a los iones como el Na+, K+, Cl−, etc.
La difusion facilitada, por su parte, permite el transporte de pequenasmoleculas polares, como los aminoacidos que al no poder atravesar la bicapalipıdica, requieren que las proteınas transmembranosas faciliten su paso.
Transporte activo: En este proceso tambien actuan proteınas de mem-brana pero ahora, a diferencia de la difusion facilitada, estas requieren energıa,en forma de ATP, para transportar las moleculas al otro lado de la membra-na. Este tipo de transporte se utiliza cuando se realiza en contra del gradienteelectroquımico. Como ejemplos de transporte activo se pueden citar la bombade Na/K, y la bomba de Ca.
En este capıtulo vamos a estudiar con detalle los procesos de difusion simplea traves de la membrana, discutiendo tambien algunos aspectos de la difusionfacilitada.
Reconocimiento y comunicacion
Estos procesos se basan en la actuacion de las moleculas situadas en laparte externa de la membrana como receptoras de sustancias especıficas eimplican la presencia de unos receptores especıficos en la membrana celularcapaces de ser activados por estımulos que vengan del medio extracelular.Los receptores de membrana, ya estimulados, cambian su comportamiento ytransforman el estımulo extracelular en intracelular. Este actua sobre enzimaso factores intracelulares provocando una respuesta celular como, por ejemplo,la contraccion muscular, la secrecion glandular, la division celular, etc.
4.1. Difusion Pura a Traves de Membranas
En este capıtulo vamos a introducir los conceptos basicos para comprenderlos procesos fısicos que se realizan a traves de las membranas. Como punto departida, vamos a utilizar los fundamentos termodinamicos, introducidos en loscapıtulos anteriores.
4.1.1. Potencial del Equilibrio de Nernst
Tal y como hemos visto en el segundo capıtulo sobre Termodinamica, si enun sistema existe variacion de masa, entonces la variacion de la energıa libre

120 CAPITULO 4. FENOMENOS DE TRANSPORTE: MEMBRANAS
de Gibbs (2.8) viene dada por:
dG = −SdT + V dP +r∑
i=1
μidni, (4.1)
donde μi =(
∂G∂ni
)T,P,nj �=i
es el potencial quımico de un tipo de partıculas i,
que a continuacion vamos a denominar especie o componente i. Recordamosque este potencial mide la variacion de la energıa libre respecto al numero departıculas presentes.
En los ejemplos que vamos a estudiar, normalmente vamos a considerar eltransporte de cargas. Si una cantidad determinada de cargas (dq) se transportacontra el potencial electrico ψ, el trabajo realizado sera:
δWq = ψdq.
Teniendo en cuenta que existen otros tipos de trabajo como el trabajo δWP
realizado cuando un gas se comprime aplicando una presion P , que conlleva ala disminucion de su volumen dV
δWP = −PdV
y el trabajo que se ejerce transportando materia dn contra el gradiente deconcentracion
δWn = μdn,
el trabajo realizado por el sistema en el caso de r especies transportadas seexpresa finalmente con la siguiente ecuacion:
δW = −PdV +
r∑i=1
μidni + ψdq. (4.2)
Usando el Primer Principio de la Termodinamica que establece que la energıainterna de un sistema aumenta en dU si se suministra a este cierta cantidadde calor δQ = TdS y si se realiza sobre este cierta cantidad de trabajo δW ,
dU = δQ + δW,
obtenemos la ecuacion fundamental de Gibbs:
dU = TdS − PdV +
r∑i=1
μidni + ψdq. (4.3)

4.1. DIFUSION PURA A TRAVES DE MEMBRANAS 121
Finalmente, usando la relacion entre la energıa interna U y la energıa libre deGibbs:
G = U + PV − TS,
la ecuacion (4.3) en el caso de presencia de un potencial electrico ψ es:
dG = −SdT + V dP +r∑
i=1
μidni + ψdq.
Una ampliacion de esta ecuacion esta relacionada con el termino ψdq. La cargaelectrica por unidad de mol y de carga viene dada por la constante de FaradayF = NAq = 9,65 ·104C/mol, donde q es la magnitud de la carga de un electron(aproximadamente 1,602 · 1019 Coulombs) y NA es el numero de Avogadro.Ası, si un determinado ion posee una carga z, la carga total de n moles delmismo sera q = nzF . A partir de esta relacion, los dos ultimos terminos dela ecuacion (4.3), en el caso de que haya mas de un ion en la disolucion, sepueden expresar como
r∑i=1
μidni + ψ
r∑i=1
ziFdni =
r∑i=1
(μi + ziFψ)dni = μidni.
La cantidadμi = μi + ziFψ (4.4)
es el potencial electroquımico, que aparece en la mayorıa de los calculos elec-troquımicos.
Teniendo en cuenta la dependencia del potencial quımico μ de la concentra-cion a traves de la ecuacion (2.9), para el potencial electroquımico μ finalmenteobtenemos la siguiente expresion:
μ = μ0 + RT ln c + zFψ, (4.5)
siendo c la concentracion de la especie considerada.Vamos a considerar ahora un sistema a presion y temperatura constantes
que esta separado por una membrana semipermeable y que consiste en doscompartimentos int (interior) y ext (exterior). Vamos a considerar que los po-tenciales electricos en los dos compartimentos son ψint y ψext, respectivamente(Fig.4.2).
De acuerdo con la condicion del equilibrio, la energıa libre del sistema Gno experimenta cambios:
(dG)T,P = (dGint)T,P + (dGext)T,P = 0. (4.6)

122 CAPITULO 4. FENOMENOS DE TRANSPORTE: MEMBRANAS
Figura 4.2: Representacion de un sistema compuesto por dos fases y dividido por una
membrana semipermeable.
Supongamos que en los dos compartimentos hay distintos componentes i =1, 2, ..., como por ejemplo en el caso de una disolucion de una sal AB, formadapor dos componentes, con concentraciones distintas en las dos fases int y ext.Si dni es el numero de moles de la componente i, intercambiada desde elcompartimento ext hacia el compartimento int, entonces de la ecuacion (4.6),obtenemos la condicion de igualdad de los potenciales electroquımicos en losdos compartimentos, (4.4):
μ0i + RT ln cint
i + ziFψint = μ0i + RT ln cext
i + ziFψext,
donde las variables cinti , cext
i son las concentraciones del componente i en los doscompartimentos (int y ext), respectivamente, y μ0
i son los potenciales quımicosen ausencia de potencial electrico.
De aquı podemos obtener la ecuacion de Nernst para la diferencia delpotencial Δψ a traves de la membrana en funcion de la actividad quımicade la componente permeable en ambos compartimientos y en condiciones deequilibrio termodinamico, que se cumple para cada especie i:
ziF (ψint − ψext) = RT (ln cexti − ln cint
i ).
La ecuacion de Nernst entonces tiene la siguiente forma, valida para cada i:
Δψ ≡ (ψint − ψext) =RT
ziFln
cexti
cinti
. (4.7)
La ecuacion (4.7) puede ser transformada para calcular la diferencia de lasconcentraciones del ion permeable i, que se crea entre las dos fases, dada ladiferencia de potencial Δψ entre ambas:
cinti = cext
i e−ziFΔψ
RT .

4.1. DIFUSION PURA A TRAVES DE MEMBRANAS 123
La ecuacion de Nernst permite por una parte el calculo de la distribucion delos iones en funcion del potencial electrico y por otra parte, el calculo delpotencial electrico que se produce como consecuencia de la distribucion nouniforme de los iones.
Las ecuaciones deducidas en esta seccion se aplican solo en caso de equi-librio termodinamico. Esto significa que la ecuacion de Nernst no se puedeutilizar para calcular el potencial de una celula viva. En realidad, el potencialde membrana de una celula viva puede ser debido al potencial de difusiono generado por bombas de iones. Por otro lado, en general es bastante fre-cuente que algunos tipos de iones se distribuyen pasıvamente, y por lo tantoestan realmente en equilibrio. Esto ultimo permite el estudio basandose en unasuposicion bastante sencilla, como la ecuacion de Nernst.
Como ejemplo de aplicacion de esta teorıa se puede citar la concentracionde cloruro en la mayorıa de las celulas vivas. Debido a que la permeablidad demembrana es bastante alta y a que no existen bombas de cloruro, su distri-bucion es practicamente pasiva y determinada por la existencia del potencialde membrana. Por lo tanto, la ecuacion de Nernst permite el calculo de laconcentracion interna de los iones de cloruro si se conoce la concentracion decloruro fuera de la membrana y el potencial de membrana. Esta ecuacion tam-bien permite el calculo del potencial de la membrana de la celula, si se conocela concentracion de cloruro dentro y fuera de la membrana.
4.1.2. Ecuacion de Nernst-Planck
De los fundamentos de la Termodinamica, sabemos que el flujo de materiade la especie i a traves de una membrana expresa el numero de moles ni queatraviesan la unidad de area A de la membrana en unidad de tiempo y se definecon la expresion Ji = 1
Adnidt . Multiplicando el numerador y el denominador por
la velocidad vi de las partıculas y teniendo en cuenta que Avidt es el volumende las partıculas en el tiempo dt, se obtiene la siguiente expresion para el flujode la especie i a traves de la membrana:
Ji = civi,
denominando ci la concentracion de la especie i.Por otra parte, la velocidad que alcanzan las partıculas es proporcional a
la fuerza fi que actua sobre ellas vi = uifi, siendo ui la movilidad. Entonces,el flujo a traves de la membrana es:
Ji = ciuifi.

124 CAPITULO 4. FENOMENOS DE TRANSPORTE: MEMBRANAS
En el caso que consideramos, la fuerza depende del gradiente del potencialelectroquımico fi = −dμi
dx , de donde obtenemos:
Ji = −uicidμi
dx. (4.8)
Por otro lado, el potencial electroquımico de la especie i a temperatura ypresion constantes, se expresa mediante la ecuacion (4.5):
μi = μ0 + ziFψ + RT ln ci. (4.9)
Sustituyendo esta ultima relacion en la expresion del flujo, ec.(4.8), obtenemosla siguente ecuacion:
Ji = −uiciziFdψ
dx− uiRT
dci
dx, (4.10)
que se denomina ecuacion de Nernst-Planck. Esta ecuacion describe como elflujo de una especie ionica a traves de una membrana depende del gradientedel potencial y del gradiende de la concentracion. Es una ecuacion basica parala descripcion de los procesos de difusion pura a traves de membranas.
En el caso de que en el sistema existan flujos de varias especies distintas,como primera aproximacion, despreciando los flujos acoplados, el flujo resul-tante viene dado por la suma de cada especie por separado, lo que constituyeel principio de independencia.
4.1.3. Teorıa de Campo Constante
El modelo de campo constante fue propuesto originalmente por Goldman(1943) y mas tarde desarollado por Hodgkin y Katz (1949). Es el modelo massencillo para la integracion y el estudio posterior de la ecuacion de Nernst-Planck.
Supongamos un sistema constituido por una membrana de espesor Δx quesepara los compartimientos int (interior) y ext (exterior), presentados en lafigura 4.3.
Fijamos el orıgen x = 0 en la interfase entre la membrana y el comparti-miento ext. Segun la ecuacion de Nernst-Planck, el flujo total del componentei, Ji(x), a traves de la membrana en el punto x en su interior, se da mediantela siguiente expresion:
Ji(x) = −uici(x)ziFdψ(x)
dx− uiRT
dci(x)
dx. (4.11)

4.1. DIFUSION PURA A TRAVES DE MEMBRANAS 125
Figura 4.3: Representacion de una membrana de espesor Δx que separa dos comparti-
mentos.
Las variables (ci ≡ cinti , cext
i ), (ψ ≡ ψint
, ψext
) son las concentraciones y lospotenciales electrostaticos en el interior de la membrana, en las interfases conlos compartimentos int y ext respectivamente, (Fig.4.3).
Para obtener la expresion del flujo en funcion de las concentraciones en loscompartimentos que separa la membrana, o la diferencia del potencial a travesde la membrana, hay que hacer varias suposiciones:
1) El sistema esta en estado estacionario y el flujo Ji es constante paratodo componente i.
2) La movilidad es constante en el interior de la membrana.
3) El campo electrico a traves de la membrana es constante. Esto significaque la derivada del potencial se puede expresar como la variacion del potencial
en un intervalo Δx: dψdx = Δψ
Δx con Δψ = ψint − ψ
ext.
Integrando la ecuacion (4.11) entre los lımites x = 0 y x = Δx, con la
suposicion anterior de que ψ(x) es una funcion lineal de x (dψdx = const.),
obtenemos:
− 1
uiRT
∫ Δx
0dx =
∫ cinti
cexti
dci
Ji + uiziFΔψΔx ci
.

126 CAPITULO 4. FENOMENOS DE TRANSPORTE: MEMBRANAS
De aquı,
Δx
uiRT=
1
uiziFΔψΔx
lnJi + uiziF
ΔψΔx cext
i
Ji + uiziFΔψΔx cint
i
.
Despejando Ji, finalmente tenemos:
Ji = −uiziFΔψ
Δx
(cexti − cint
i eziFΔψ/RT
1 − eziFΔψ/RT
). (4.12)
Esta ecuacion da la forma explıcita del flujo de la especie i a traves de lamembrana en funcion de las concentraciones y los potenciales electrostaticosen el interior de la membrana en la interfase con el exterior.
Vamos ahora a relacionar las concentraciones y los potenciales en el interiorde la membrana con las mismas cantidades en el exterior. Podemos considerarque la interfase entre la membrana y el medio exterior es tal que cualquierespecie disuelta en uno de los medios, se disuelve en la fase contigua. Enequilibrio, la relacion entre las concentraciones de la especie i en las dos fasesviene dada por el coeficiente de reparto βi, definido a continuacion.
Si ademas suponemos que:1) El intercambio de iones entre la membrana y el exterior es mucho mas
rapido que el proceso de difusion a traves de la membrana, esto permitira tra-bajar en condiciones de equilibrio en las fases int y ext en todo instante yrelacionar las concentraciones en el interior y en el exterior de la membrana atraves del coeficiente de reparto.
2) El coeficiente de reparto βi de la componente i es constante e identicoen las interfases con el interior y el exterior. Entonces:
cexti
cexti
=cinti
cinti
= βi. (4.13)
Usando la condicion de equilibrio en cada una de las interfases, podemos apli-car la ecuacion de Nernst (4.7) para cada interfase, con lo que se obtiene elsiguiente sistema de ecuaciones:
ψext − ψext = −RT
ziFln
cexti
cexti
,
ψint − ψint = −RT
ziFln
cinti
cinti
. (4.14)
En consecuencia, tenemos:
Δψ ≡ ψint − ψ
ext= ψint − ψext ≡ Δψ. (4.15)

4.1. DIFUSION PURA A TRAVES DE MEMBRANAS 127
Ahora, introduciendo el parametro pi = uiRTΔx βi, que tiene el sentido de per-
meabilidad de la membrana a la especie i, y usando las ecuaciones (4.13-4.15),la ecuacion (4.12) se transforma en:
Ji = −piziFΔψ
RT
(cexti − cint
i expziFΔψ/RT
1 − expziFΔψ/RT
). (4.16)
Esta ecuacion de campo constante se denomina tambien ecuacion de flujo deGoldman-Hodgkin-Katz . A partir de ella se puede estudiar la dependencia delflujo de una componente a traves de la membrana, de las concentraciones encada uno de los compartimentos y de la diferencia de potencial existente entreellos, cuando el sistema esta en un estado estacionario.
En el caso en el que las concentraciones en el interior y en el exterior dela membrana sean iguales, sustituyendo cint
i = cexti = ci en la ecuacion (4.16),
obtenemos la siguiente expresion para los flujos:
Ji = −piciziF
RTΔψ.
Esta forma del flujo se puede interpretar, segun la ley de Ohm, como unacorriente que atraviesa un conductor electrico.
En el caso de ausencia de corriente de la componente i, Ji = 0, de laecuacion (4.16), obtenemos la ecuacion de Nernst:
cexti = cint
i eziFΔψ/RT .
Finalmente, cuando las dos concentraciones cexti , cint
i son distintas, cexti = cint
i ,la relacion entre el potencial y el flujo no es una recta, sino una curva conmayor pendiente cuando el flujo va desde el compartimento externo al com-partimento interno que en el caso contrario. Por consiguiente, la resistencia dela membrana al flujo del interior es menor en el caso en el que hay intercambiode materia ext → int que en el que hay intercambio de materia int → ext.

128 CAPITULO 4. FENOMENOS DE TRANSPORTE: MEMBRANAS
Problema.A partir de la ecuacion de Goldman-Hodgkin-Katz, (4.16), sabiendo el valordel flujo de la especie, expresar la concentracion de la especie en el compar-timento int en el exterior de la membrana en funcion de las demas variablesen el caso de valor pequeno de la diferencia del potencial.
Solucion.De la ecuacion (4.16), cuando Δψ � 1, desarrollando la exponencial hastael primer orden en Δψ, es decir, ex � 1 + x, obtenemos
JiRT (−ziFΔψ/RT ) = −piziFΔψ(cexti − cint
i − cinti ziFΔψ/RT ),
de donde
cinti =
picexti − Ji
pi(1 + ziFΔψRT )
.
4.2. Difusion Ionica a Traves de la Membrana
En esta seccion vamos a estudiar los fenomenos de difusion ionica. Es-tos procesos son fundamentales para explicar las diferencias de potencial demembrana que aparecen en las celulas. El objetivo sera presentar las distin-tas distribuciones ionicas que contribuiran a la generacion de potenciales demembrana. Esto se aplicara a lo largo del capıtulo al estudio de modelos degeneracion del potencial de membranas en los sistemas biofısicos.
Como punto de partida vamos a introducir el principio de electroneutrali-dad. Este principio establece que la suma de las cargas positivas es igual a lasuma de las cargas negativas en cualquiera de los compartimentos separadospor una membrana. Es un principio fundamental y se establece a nivel ma-croscopico. Sin embargo, las membranas biologicas son capaces de almacenarcargas en las superficies de contacto con algunos medios, actuando como con-densadores. No obstante, la magnitud de la carga es despreciable y el principiode la electroneutralidad es exacto en el lımite termodinamico.
4.2.1. Potenciales de Gibbs-Donnan
Consideramos el sistema de la figura 4.4, donde una membrana separados compartimentos en los cuales se introducen sales monovalentes A+X− yA+B− de la misma concentracion. Supongamos que los iones A+ y B− pueden

4.2. DIFUSION IONICA A TRAVES DE LA MEMBRANA 129
Figura 4.4: Esquema del sistema compuesto por las dos fases.
difundir a traves de la membrana, mientras que los iones X− se quedan confi-nados en el compartimento ext. Supongamos tambien que no existe potencialelectrico externo aplicado sobre el sistema.
En el caso de la figura 4.4, el anion B− se dirige hacia la parte exterior dela membrana (parte ext) arrastrando al anion A+ para satisfacer el principiode electroneutralidad. Esto conlleva un aumento de la concentracion de A+ enla parte ext y una disminucion en la parte int. Como consecuencia se generauna gradiente de concentraciones de iones A+, que se opone al flujo del mismoanion. En el equilibrio se alcanza una distribucion asimetrica de los dos ionesA+ y B− en ambos lados de la membrana. Esta distribucion asimetrica generauna diferencia de potencial electrostatico entre las dos partes, que se denominapotencial de Gibbs-Donnan.
En la parte exterior de la membrana la concentracion de A+ es mayor queen la parte interior y se compensa con el potencial electrostatico. Lo mismoocurre con el anion B−, pero de manera opuesta, debido a que su carga esnegativa.
Aplicando la condicion del equilibrio de Nernst, para la variable Δψ =ψint − ψext tenemos:
Δψ = −RT
Fln
cintA
cextA
y
Δψ =RT
Fln
cintB
cextB
.
Estas dos expresiones se cumplen si las respectivas razones entre las concen-traciones en el interior y en el exterior son iguales, lo que introduce la razon(o

130 CAPITULO 4. FENOMENOS DE TRANSPORTE: MEMBRANAS
tasa) de Donnan r:cintA
cextA
=cextB
cintB
= r.
En terminos de este parametro, el potencial de Gibbs-Donnan tiene la siguienteforma:
Δψ = −RT
Fln r. (4.17)
Si el ion indifundible X tiene su propia carga zX y su concentracion es cX , sepuede demostrar, que la razon de Donnan correspondiente es:
r = −R +√
R2 + 1, (4.18)
donde R = |zX |cX
2ceqy ceq es la concentracion de sal en equilibrio en el compar-
timento opuesto al compartimento donde se encuentra el ion X.Como se observa, la razon de Donnan es una funcion no lineal del parame-
tro R, aumentando con la concentracion de los iones X positivos y disminu-yendo con la misma para iones negativos.
El potencial de Gibbs-Donnan esta determinando basicamente por la canti-dad de componentes cargados de la celula no intercambiables, que provocan ladistribucion asimetrica de los demas iones. Si el sistema esta compuesto por io-nes X debilmente difundibles, entonces aparecera un flujo lento que tendera aigualar las concentraciones de estos iones en los dos lados de la membrana.El tiempo caracterıstico de este proceso es mucho mas grande que el tiempocaracterıstico de difusion de los demas iones.
Resumiendo, los potenciales de Gibbs-Donnan (4.17) son potenciales deequilibrio y aparecen cuando la presencia de un ion indifundible provoca ladistribucion asimetrica de los demas iones. Estos potenciales son una apro-ximacion muy buena para el estudio del potencial de membrana en sistemasbiologicos, aunque presentan una simplificacion por no tener en cuenta loscambios en el volumen celular, lo que conllevarıa alteraciones de todas lasconcentraciones intracelulares.
4.2.2. Creacion de Potenciales de Difusion
Consideramos el siguiente sistema, presentado en la figura 4.5. Supongamosque no existe potencial electrico externo aplicado sobre el sistema y que se haintroducido una sal en el compartimento izquierdo siendo la movilidad del ca-tion A+ menor que la del anion B−. Esto conlleva que haya una difusion masrapida del ion con mayor movilidad hacia el otro compartimento. Por otro la-do, los dos iones difunden con la misma velocidad para no violar el principio de

4.2. DIFUSION IONICA A TRAVES DE LA MEMBRANA 131
Figura 4.5: Representacion esquematica para la obtencion de los potenciales de difusion.
electroneutralidad. Esto significa que el ion mas rapido difunde mas despacio,frenado por el mas lento. En el mismo tiempo, el ion mas lento esta aceleradopor su ion opuesto. Como resultado, la difusion se realiza mediante una ve-locidad intermedia a la de los dos iones. Esta difusion crea una diferencia depotencial, que lleva el nombre de potencial de difusion.
Usando la ecuacion de Nernst-Planck (4.10) para los flujos de los cationes(+) y los aniones (−), podemos obtener la expresion del potencial de difusion:
J+ = −u+RTdc+
dx− u+z+c+F
dψ
dx, (4.19)
y
J− = −u−RTdc−dx
− u−z−c−Fdψ
dx. (4.20)
Para simplificar el analisis, vamos a considerar el caso de iones monovalentes,z+ = −z− = 1, c+ = c− = c en el exterior de la membrana y c+ = c− = c ensu interior.
Segun la condicion de electroneutralidad, tenemos:
J+ = J−.
De aquı, igualando la parte derecha de las ecuaciones (4.19,4.20) para los flujosde ambos iones, tenemos:
u+RTdc
dx+ u+cF
dψ
dx= u−RT
dc
dx− u−cF
dψ
dx.

132 CAPITULO 4. FENOMENOS DE TRANSPORTE: MEMBRANAS
Ahora, reorganizando e integrando entre los dos extremos de la membrana,tenemos:
(u+ + u−)F
∫ ψint
ψext
dψ = (u− − u+)RT
∫ cint
cext
dc
c,
lo que nos permite obtener la expresion final para la variacion del potencial:
Δψ = −u+ − u−
u+ + u−
RT
Fln
cint
cext . (4.21)
Suponiendo ahora que existe un equilibrio en las interfases y que la membranaes simetrica obtenemos, segun hemos visto en la seccion anterior, la ecuacionfinal para el potencial de difusion de una sal formada por iones monovalentes:
Δψ = −u+ − u−
u+ + u−
RT
Fln
cint
cext. (4.22)
Para iones de la misma movilidad, u+ = u−, Δψ = 0 y el potencial de difusiones cero. Esto significa que para tener un potencial de difusion es necesario queuno de los iones tenga una movilidad distinta que el otro.
Problema.Comparar el potencial de difusion antes y despues de aumentar la concen-tracion en el compartimento interior 2 veces.
Solucion.Segun la ecuacion (4.22), antes de variar los parametros, tenemos:
Δψantes =u+ − u−
u+ + u−
RT
Fln
cint
cext
Despues de variar la la concentracion tenemos:
Δψdespues =u+ − u−
u+ + u−
RT
Fln
2cint
cext,
de donde:Δψdespues
Δψantes= 1 +
ln 2
ln cint/cext.

4.2. DIFUSION IONICA A TRAVES DE LA MEMBRANA 133
4.2.3. Ecuacion de Goldman-Hodgkin-Katz
En una distribucion ionica que existe alrededor de la membrana de unaxon, hay que superar las aproximaciones triviales. Una forma de hacer estoes aplicar la teorıa del campo constante, (4.16). La aplicacion de esta teorıa auna distribucion de varios aniones y cationes monovalentes, da las siguientesecuaciones para los flujos:
J+ = −p+FΔψ
RT
[cext+ − cint
+ eFΔψ/RT
1 − eFΔψ/RT
]y
J− =p−FΔψ
RT
[cext− − cint
− e−FΔψ/RT
1 − e−FΔψ/RT
],
donde de nuevo los subındices + y − se refieren a los cationes y a los anionesrespectivamente y los parametros p± son sus permeabilidades.
De la condicion de electroneutralidad, sumando por separado sobre todoslos iones positivos y negativos:∑
+
J+ =∑−
J−, (4.23)
se obtiene: ∑+
(−p+)
[cext+ − cint
+ b
1 − b
]=∑−
(p−)
[cext− − cint
− /b
1 − 1/b
], (4.24)
donde el factor b = eFΔψ/RT .Por otro lado, despejando b, tenemos:
b =
∑+ p+cext
+ +∑
− p−cint−∑
+ p+cint+ +
∑− p−cext
−
. (4.25)
Comparando las dos expresiones para el factor b, obtenemos finalmente laexpresion del potencial de la membrana:
Δψ = −RT
Fln
∑+ p+cint
+ +∑
− p−cext−∑
+ p+cext+ +
∑− p−cint
−
. (4.26)
La ecuacion (4.26) es la famosa ecuacion de Goldman-Hodgkin-Katz (GHK)que expresa el potencial de la membrana en funcion de las concentraciones y

134 CAPITULO 4. FENOMENOS DE TRANSPORTE: MEMBRANAS
las permeabilidades de los distintos aniones y cationes monovalentes, que laatraviesan.
En un caso concreto de estudio, que es el caso del axon de una neurona,el potencial de membrana se debe principialmente a los iones de Na+,K+ yCl−. La ecuacion de GHK entonces se transforma en:
Δψ = −RT
Fln
pNacintNa + pKcint
K + pClcextCl
pNacextNa + pKcext
K + pClcintCl
. (4.27)
A partir de la ecuacion (4.27) se ve que si la permeabilidad de uno de los ioneses mucho mayor que la de los demas, los terminos correspondientes a los demasiones se pueden despreciar y el potencial determinado por la ecuacion de GHKse aproxima al potencial de Nernst del ion con la mayor permeabilidad.
La expresion (4.27) es aplicable a la descripcion del potencial generadopor la distribucion de iones de Na+,K+ y Cl− en membranas excitables enestado de reposo. En sistemas biologicos reales, la concentracion de estos ionesse mantiene constante gracias a la bomba Na+,K+-ATPasa y es interesantever como esta bomba de iones afecta al potencial de la membrana en mediosexcitables.
Se sabe que la bomba Na+,K+-ATPasa intercambia 3 iones de Na+ por2 iones de K+ y que el transporte debido al efecto de la bomba compensa ladifusion de los iones por el gradiente de concentracion.
Vamos a ver como se modifica la ecuacion de GHK en presencia de laNa+,K+-ATPasa. Sean JNa y JK los flujos de los iones de Na+ y K+ debidos
a la Na+,K+-ATPasa y sea d = − JNa
JK. En nuestro caso, la razon es d =
3/2. En el estado de reposo, la concentracion de Na+ en ambos lados de lamembrana es constante y por consiguinete JNa = −JNa.
Usando la condicion de la electroneutralidad (4.23),
JNa + JNa + JK + JK = JCl
e introduciendo la razon d, teniendo en cuenta que los dos primeros terminosse cancelan debido a la concentracion constante de Na+ en ambos lados dela membrana, obtenemos la siguiente relacion entre los flujos de los distintosiones:
1
dJNa + JK = JCl.
Como se observa, el efecto de la Na+,K+-ATPasa es la presencia del factor1d en comparacion con el caso de ausencia del flujo. Esto se puede interpretar

4.3. NOCIONES BASICAS DE LA ELECTROFISIOLOGIA 135
como una reduccion efectiva del flujo de los iones Na+. Si ahora sustitui-mos cada uno de los flujos por su expresion correspondiente de la ecuacionde campo constante, y despejando Δψ de la ecuacion resultante, obtenemosuna expresion analoga a (4.27), donde el termino relacionado con el ion Na+
esta modificado por el factor de la razon entre los flujos d:
Δψ = −RT
Fln
1dpNac
intNa + pKcint
K + pClcextCl
1dpNacext
Na + pKcextK + pClc
intCl
. (4.28)
La ecuacion (4.28) es la ecuacion GHK modificada por la presencia de lasbombas Na+,K+-ATPasa.
En el caso de d = 1, la ecuacion (4.28) se reduce a la ecuacion GHK. Enesta condicion se puede demostrar que la ATPasa es capaz de mantener lasconcentraciones de Na+ y de K+ y el potencial de la membrana estacionarios.De aquı se puede afirmar que el papel principal de la Na+,K+-ATPasa en elfuncionamento de la membrana esta basado en su capacidad de transportariones de Na+ y K+ gracias a la energıa metabolica.
4.3. Nociones Basicas de la Electrofisiologıa
En los capıtulos anteriores presentamos el estudio de los flujos ionicos y lageneracion del potencial de membrana desde el punto de vista termodinamico,describiendo el movimiento de los iones gracias al gradiente electroquımico.
En este capıtulo vamos a presentar los fenomenos desde el punto de vistade la Electrofisiologıa. Ahora los flujos ionicos son considerados como corrien-tes electricas que atraviesan la membrana y el potencial de la membrana seintroduce a partir de un circuito equivalente.
Esta teorıa clasica tiene su origen en los trabajos pioneros del estudio de laexcitabilidad del nervio y del musculo y tuvo su mayor exito con el modelo deHodgkin y Huxley del potencial de accion del axon gigante de calamar. Por susdescubrimientos importantısimos en el campo de la Neurofisiologıa, Hodgkiny Huxley fueron galadordonados en los anos 60 con el premio mas prestigiosoen el ambito de la investigacion, el premio Nobel.
4.3.1. Propiedades Electricas de las Membranas: Fuerza Elec-tromotriz
Consideramos el potencial de la membrana como una fuerza electromotrizque influye sobre el movimiento de los iones a traves de la membrana de formasimilar a como afecta a las cargas electricas a traves de un conductor.





























