Embed Size (px)
Citation preview

CAPÍTULO 1.
LA ORGANIZACIÓN DEL CUERPO DE LAS PLANTAS VASCULARES
Las plantas con semillas (Espermatofitas) constituyen en la actualidad el grupo más numeroso e
importante del reino vegetal y sus integrantes forman, por ese motivo, la mayor parte de la
vegetación actual de la tierra. Por esta razón, y por el hecho de ser las plantas más especializadas
tanto en su organización externa (morfología) como interna (anatomía) comenzamos con ellas este
curso.
La enorme variabilidad que se observa en la organización de las Espermatofitas queda evidenciada
si comparamos por ejemplo, un roble con una lenteja de agua, una palmera con una gramínea, un
cactus con un girasol. Si bien la apariencia de estas plantas es muy diferente, la organización de sus
cuerpos sigue un plan o modelo que puede reconocerse en todos los ejemplos citados.
Lo característico en la organización de este modelo es la presencia de tres órganos básicos: tallo,
hojas y raíz. Tal división en órganos responde a un criterio de conveniencia ya que la planta
constituye de hecho una unidad. El límite entre tallo y hoja es particularmente dudoso y por esto
algunos autores prefieren reunirlos bajo un concepto más amplio: el de brote o vástago.
La raíz constituye la parte subterránea del eje de la planta, especializada en la absorción de
sustancias. Se presenta comúnmente como un sistema radical a consecuencia de la ramificación de
una raíz originariamente única que recibe el nombre de raíz principal, careciendo de hojas. Las
ramificaciones o raíces laterales tienen un origen profundo o interno (endógeno) desde la raíz
principal.
El brote es generalmente aéreo y también se ramifica dando ramas laterales que tienen un origen
superficial o externo (exógeno). Se forman a partir de yemas axilares, pequeños órganos cubiertos
por escamas que se localizan en el tallo, en el ángulo o axila determinado por la inserción de cada
hoja. Las ramas laterales pueden volver a ramificarse formando el todo un sistema caulinar.
En síntesis, podemos establecer que el modelo estructural del cuerpo vegetativo de las plantas con
semillas consta de una parte subterránea, el sistema radical, y partes aéreas (tallo y hojas) que en
conjunto constituyen el sistema caulinar. El cuello de la raíz es el límite entre ambos.
Cuando el cuerpo de un vegetal está diferenciado en raíz, tallo y hojas se lo llama cormo y
Cormofitas a las plantas que lo poseen. Son ejemplos todas las Plantas Vasculares (Traqueofitas)
que comprenden además de las Espermatofitas, las Pteridofitas (ej. helechos) que carecen de
semillas.

En contraposición a las Cormofitas están las Plantas Avasculares o Talofitas. Se entiende por talo a
un cuerpo vegetativo que no está dividido en los tres órganos fundamentales, raíz, tallo y hojas y por
lo tanto no responde al modelo de organización de las Cormofitas. Comprende las algas, los hongos,
los musgos, las hepáticas.
BIBLIOGRAFÍA Troll, W. 1959. Allgemeine Batanik. I-XVI, 1-927, fig. 1-699- Ferdinand Enke Verlag, Sttutgart Leyenda Modelo estructural de una planta vascular; co, cotiledón; h, hipocótilo; r, raíz principal; rl, raíz lateral; t, tallo; y, yemas axilares; yt, yema terminal. Modificado por Troll siguiendo a Sachs

CAPÍTULO 2
SEMILLAS Y PLÁNTULAS
Las Espermatofitas o plantas con semillas comprenden las Angiospermas y las Gimnospermas. Son
Angiospermas las plantas que poseen las semillas encerradas en un fruto. A ellas pertenecen las
Dicotiledóneas y las Monocotiledóneas. La diferencia entre ambas se establece fundamentalmente
debido a que, en las primeras, el embrión posee dos hojas embrionarias o cotiledones, mientras que
en la segunda, hay una sola. Ejemplos de Dicotiledóneas zapallo, papa, algarrobo, roble, quebracho;
Ejemplos de Monocotiledóneas, lirio, yuca, bananero, azucena, maíz, arroz, trigo, especies de
orquídeas, entre muchas otras).
Las Gimnospermas en cambio tienen las semillas desnudas, es decir no encerradas en un fruto; el
embrión posee de uno a varios cotiledones (Ej. Pino, ciprés, cedro, araucaria, ginkgo, cicas, entre
otras)
1. SEMILLAS
Es la estructura típica de diseminación de las Espermatofitas. Posee una cubierta de protección,
comúnmente dura y resistente, el tegumento seminal o episperma que encierra el embrión. El
embrión es una planta en miniatura en estado de vida latente o de letargo, la cual tiene
representados los tres órganos fundamentales de la planta adulta, esto es raíz, tallo y hojas, por la
radícula, el hipocótilo y el/ o los cotiledón/cotiledones respectivamente. El hipocótilo y la radícula
forman el eje radícula-hipocótilo, en uno de cuyos extremos se encuentra el ápice meristemático de
la raíz y en el otro el ápice meristemático del tallo. Dichos ápices están encargados del desarrollo del
sistema radical y del brote de la futura planta.
En la germinación, el embrión crece convirtiéndose en una plántula. Dado que en este tiempo no
puede aún realizar la fotosíntesis por no poseer clorofila, debe nutrirse de sustancias de reservas de
manera heterótrofa. Las reservas se almacenan en un tejido especial que es el endosperma,
ubicado entre el embrión y el episperma. Es el caso del ricino, por ejemplo.
En otras semillas el endosperma es consumido por el embrión durante el desarrollo de las semillas
por lo que las sustancias de reserva en la semilla madura se acumulan en el mismo embrión. Ej. La
semilla de poroto.
Se distinguen, varios tipos de semillas:

Semillas endospermadas: poseen endosperma, un tejido especial que almacena las
sustancias de reserva. (Endosperma: tejido exclusivo de Angiospermas, que se origina a
partir de la fertilización de una gameta masculina con la célula media, binucleada, del saco
embrionario; la dotación cromosómica de las células del endosperma es 3n).
Semillas exendospermadas: Las sustancias de reserva son almacenadas por el embrión. El
endosperma se consume durante el desarrollo de la semilla.
Semillas protálicas: Son las semillas de Gimnosperma. El tejido reservante es el protalo o
gametofito femenino (dotación cromosómica de sus células es: n)
Semillas perispermadas: el principal tejido de reserva es el tejido de la nucela del óvulo
(tejido de la planta madre, por lo que la dotación cromosómica 2n). Es un tejido materno que
persiste en la semilla, no es consumido por el desarrollo del endosperma, y acumula
reservas. En estas semillas generalmente también desarrolla el endosperma pero es
reducido en relación al perisperma.
Principales sustancias de reserva de las semillas
Hidratos de carbono: (almidón y hemicelulosas). El almidón se reserva en los plástidos. Las
hemicelulosas se acumulan en las paredes celulares
Proteínas: Se almacenan en los cuerpos proteicos
Lípidos: Se almacenan en los cuerpos lipídicos
Nutrientes minerales: se almacenan principalmente en los cristales globoides que son
inclusiones dentro de la matriz proteica de los cuerpos proteicos. Se trata de la sal del mio-
inositol-6-fosfato de K y Mg. Los principales minerales que se acumulan en la semilla son P,
K, Mg, y también Ca, Fe, Mn.
El Fe se acumula como fitoferritina; éta es una molécula que frecuentemente se encuentra
en los plástidos de los tejidos del embrión (también se la encuentra en los plástidos de las
hojas).
Ejemplos de semillas endospermadas:
Ricino o castor: La semilla deriva de un óvulo anátropo. Embrión central, recto, eje radícula-
hipocótilo corto. El ápice meristematico del brote no tiene primordios foliares desarrollados.
Dos cotiledones muy delgados, no reservantes, con un sistema procambial visible.

Episperma con una carúncula; se identifica la región del rafe. Endosperma abundante,
reservante principalmente de proteínas y lípidos.
Maíz: La semilla es la típica semilla de una Gramínea con formaciones específicas que
caracterizan a las semillas de esta familia. El episperma esta unido al pericarpo, por lo cual
se habla comúnmente de grano y no de semilla. La semilla deriva de un óvulo hemi-
anátropo. El embrión desplazado hacia un lado. El cotiledón es grande cubre lateralmente al
embrión y tiene forma de escudo, de ahí el nombre de escutelo con el que se lo conoce.
Esta ubicado entre el eje y el endosperma. En la gémula o plumula se diferencia el ápice
meristemático del brote cubierto por el coleoptilo (interpretado como un catafilo), y varios
primordios foliares. La radícula está cubierta por una estructura especial denominada
coleorriza.
Cebolla: La semilla deriva de un óvulo anátropo. Embrión central con un eje corto; un
cotiledón alargado, enrollado en su extremo con una vaina que envuelve el ápice
meristemático del brote. El ápice meristemático del brote no tiene primordios foliares
desarrollados.
Ejemplo de una semilla exendospermada:
Poroto: Deriva de un óvulo campilótropo. En el episperma se observan claramente la cicatriz
o hilo, el micrópilo, la lente. La lente es una formación característica de las semillas de
Leguminosas. Los tejidos que conforman el episperma tambien son caracteristicos de la
familia. No hay endosperma en la semilla madura. El embrión, durante el desarrollo,
consume el endosperma originado a partir de la doble fecundación. Las reservas se
acumulan en los tejidos embrionarios. El eje es alargado y en el se visualiza claramente el
nudo cotiledonar y el epicótilo. La gémula está contituida por el ápice meristemático del
brote y cubierta por primordios foliares y hojás muy jóvenes.
2. PLÁNTULAS
Se denomina plántula a la pequeña planta que se produce por el desarrollo del embrión durante la
germinación.
El primer órgano en desarrollarse es la radícula, de la cual deriva el sistema radical de la planta
adulta. Los cotiledones están al servicio de la nutrición de la plántula. En los primeros estados de
desarrollo, el embrión no puede alimentarse de manera autótrofa dado que no posee clorofila.

Depende por lo tanto de los nutrientes orgánicos almacenados en el tejido de reserva o en los
mismos cotiledones, siendo por ende su nutrición heterótrofa.
Algunos ejemplos:
Ricino: El hipocótilo se alarga elevando los cotiledones por encima del nivel de la tierra, por
lo que la germinación es epigea. Hasta que el endosperma se consume en su totalidad, los
cotiledones permanecen cubiertos por el episperma; en esta etapa, los cotiledones son
haustoriales. Cuando las reservas se agotan, los cotiledones emergen a la luz, y se vuelven
clorofílicos. La nutrición entonces continúa en forma heterótrofa, siendo los cotiledones los
primeros órganos fotosintetizantes.
Cebolla: La nutrición autótrofa y heterótrofa son simultáneas durante la germinación. El
único cotiledón crece alargándose durante la germinación. Su porción apical permanece
dentro del episperma y funciona como haustorio nutriéndose del endosperma hasta que este
se agota, mientras la parte basal progresivamente se expone a la luz y se vuelve clorofílica.
La germinación es epigea, aunque lo que se alarga más notablemente no es el hipocótilo
como en el ricino sino el cotiledón.
Maíz: el cotiledón o escutelo funciona totalmente como haustorio durante toda su vida, es
decir que está dedicado por completo a la nutrición heterótrofa, permaneciendo en el interior
de la semilla. El hipocótilo no crece por lo que el cotiledón permanece bajo tierra. La
germinación es, por lo tanto, hipogea. Se desarrolla la gémula, principalmente dando lugar al
alargamiento del coleoptilo y al despliegue de las hojas fotosintetizantes. Que emergen a
través del coleoptilo. A la raíz primaria derivada de la radícula del embrión, se agregan otras
raíces que se originan en otros lugares del eje, por lo que se trata de raíces adventicias.
Phaseolus multiflorus/ Phaseolus vulgaris: En estas dos especies de porotos, la germinación
es hipogea y epigea, respectivamente. En la primera de las especies, las hojitas de la
gémula emergen rápidamente a la luz, convirtiéndose en órganos clorofílicos. Los
cotiledones en cambio no emergen, siendo su función exclusivamente reservante; cuando
terminan de ceder sus reservas, mueren. Otros ejemplos de semillas exendospermadas con
germinación hipogea son las semillas de arveja, haba, taco de reina, castaño silvestres,
entre muchas otras.

En la segunda de las especies, la germinación es epigea, es decir, el hipocótilo se alarga y
los cotiledones, luego de ceder la mayor parte de las reservas, se vuelven clorofílicos. Otros
ejemplos de semillas exendospermadas con germinación epigea son: zapallo, girasol,
sauce, entre muchas otras.
En el cedro y en el pino, la germinación es epigea. El hipocótilo se alarga y eleva los
cotiledones sobre el nivel del suelo. Los cotiledones son varios y están dispuestos en un
verticilo, rodeando el ápice meristemático del brote. Los cotiledones permanecen dentro de
la semilla, y en la medida que avanza la germinación, las bases se exponen a la luz,
volviéndose verde progresivamente, en tanto las puntas quedan hasta el final, absorbiendo
nutrientes del protalo. Finalmente, los nutrientes se agotan y los cotiledones se despliegan.
BIBLIOGRAFÍA
Troll, W. 1959. Allgemeine Batanik. I-XVI, 1-927, fig. 1-699- Ferdinand Enke Verlag, Sttutgart
Leyenda
SEMILLAS Y PLÁNTULAS. Abreviaturas: ca, carúncula; cl, coleoptilo; co, cotiledones; cr, coleorriza;
e, endosperma; e, endosperma; ep, epicotilo; g, gémula; h, hipocótilo; ho, hojas; pi, pilorriza; pr,
protalo; ra, radícula; raíz


CAPÍTULO 3
EL CUERPO DE LA PLANTA: MORFOLOGÍA EXTERNA
I. El brote
a. La hoja
b. El tallo
II. II. La raíz
a. La hoja
El término hoja se aplica generalmente a las hojas normales o nomofilos encargados de la
fotosíntesis. Sin embargo, el concepto es mucho más amplio, comprendiendo todos los apéndices
laterales del eje que tienen un crecimiento apical definido.
Se presentan únicamente en las Cormofitas, variando su estructura, forma y función de acuerdo con
la posición que ocupan sobre el eje de la planta
Basándose en estas variaciones, se ha establecido una clasificación de los diferentes tipos de
órganos foliares
Organización de la hoja
Hay un plan básico de organización de la hoja que se reconoce en todos los tipos. Los nomofilos
sirven como modelo dado que en ellos normalmente están representadas todas las partes de una
hoja aunque con distinto grado de desarrollo según las plantas.
En un nomofilo se distinguen fundamentalmente dos partes: una distal y una proximal respecto al eje
La parte distal está constituida por: el limbo o lámina, verde, aplanada y con frecuencia muy delgada.
La lámina puede ser entera, indivisa o variablemente dentada o lobada, en las hojas simples. O bien
puede estar dividida en unidades denominadas folíolos que se insertan a lo largo de un eje o raquis
(el cual a veces es muy corto o queda reducido a un punto) en las hojas compuestas (Ej. rosa,
trébol). El pecíolo o pie une la lámina al tallo cuando la hoja es peciolada. Cuando el pecíolo falta, la
lámina se denomina sésil.
La parte proximal está representada por la base de la hoja que comúnmente es ensanchada y
aplanada respecto al pecíolo. En ciertas plantas da lugar a una vaina que envuelve el tallo: esta
puede ser cerrada, cuando los bordes laterales se sueldan, o abierta, si se tocan o superponen sin
soldarse. En otros casos la base foliar está representada por las estípulas que son dos pequeñas
formaciones foliosas dispuestas una a cada lado del pecíolo. En algunas plantas (Poligonáceas) las
estípulas se presentan soldadas constituyendo la ócrea, formación que rodea al tallo por encima de
la inserción de cada hoja.
Venación:
El limbo de las hojas se encuentra recorrido por haces (venas o nervios). Por lo común, las venas
más gruesas son más o menos sobresalientes por la cara inferior o envés de la hoja (cara abaxial)
mientras que aparecen como surcos por la cara superior o haz (cara adaxial).
Una hoja puede tener una, dos o más venas. Ejemplos de hojas con una sola vena, uninervadas
encontramos en las Coníferas (Gimnospermas). Hojas con muchas venas, plurinervadas, son
comunes en helechos y Angiospermas.
Los casos más frecuentes en Angiospermas corresponden a: (i) Venación reticulada, común entre
Dicotiledóneas (aunque también se presentan en algunos helechos y Monocotiledóneas): hay una
vena principal de la que parten venas laterales. Se anastomosan entre sí por medio de venas más
pequeñas que divergen entre ellas formando una red (de allí el nombre de reticulada). (2) Venación
estriada o paralela, característica en Monocotiledóneas (también se presenta en algunas
Dicotiledóneas). Los haces principales se disponen paralelos en el sentido longitudinal de la lámina.
Convergen entre sí sólo en el extremo apical o en ambos extremos del limbo foliar. Las venas
principales se hallan intercomunicadas lateralmente por otras pequeñas.
Tanto la venación reticulada como la paralela son de tipo cerrado porque las venas se anastomosan
entre sí. (Cabe aclarar que siempre la última ramificación siempre queda abierta) En contraposición
a ello encontramos el tipo abierto donde las venas terminan libremente sin anastomosarse; se
encuentran en helechos y Gimnospermas. Un caso particular de venación abierta es la dicotómica
(repetidamente bifurcada)) que observamos en Ginkgo (Gimnospermas) y en algunos helechos.
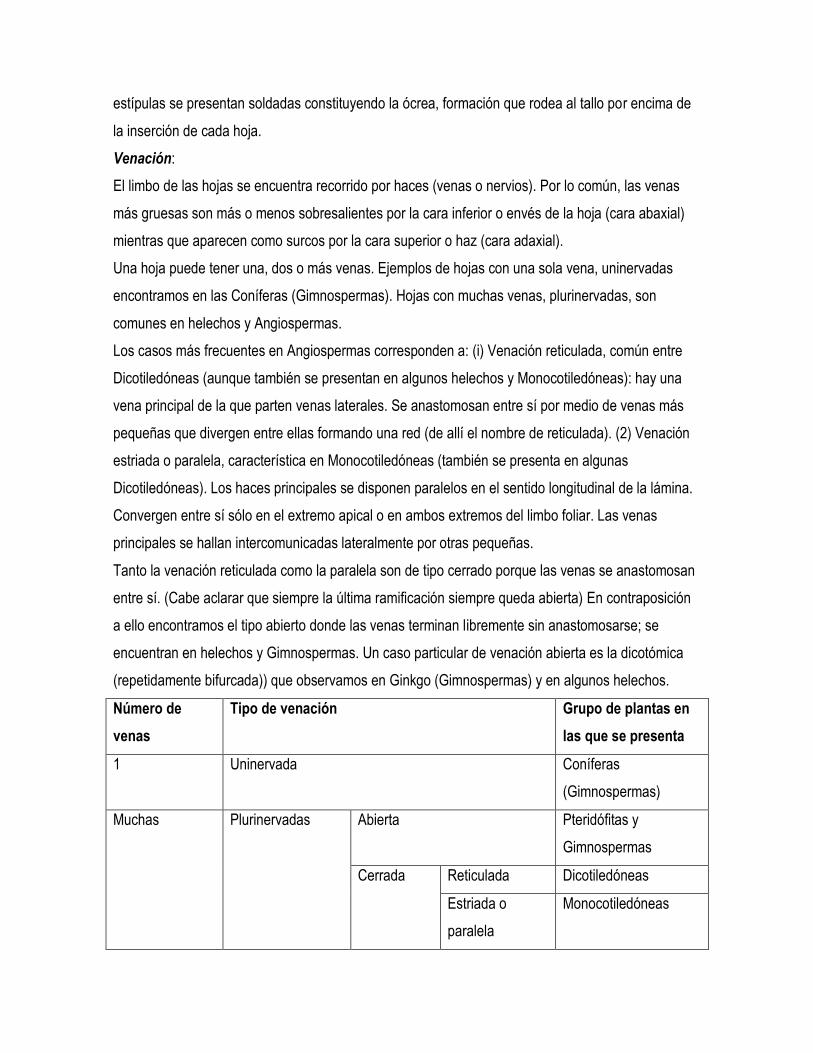
Número de
venas
Tipo de venación Grupo de plantas en
las que se presenta
1 Uninervada Coníferas
(Gimnospermas)
Muchas Plurinervadas Abierta Pteridófitas y
Gimnospermas
Cerrada Reticulada Dicotiledóneas
Estriada o
paralela
Monocotiledóneas

En el brote de las plantas vasculares se puede diferenciar una parte vegetativa y otra reproductiva,
ambas provistas de órganos foliares.
En la región vegetativa se distinguen los siguientes tipos de hojas:
Cotiledones: son las hojas del embrión que pueden o no persistir en la planta adulta, siendo
variable su número y función.
Catafilos: (del griego cata, abajo y filo, hoja). De estructura más sencilla que los nomofilos.
Son las hojas ubicadas por debajo de los nomofilos. En el eje principal se ubican por debajo
de los nomofilos y por encima de los cotiledones. En las ramas laterales, son las primeras
hojas de la rama. En los árboles, los catafilos son las escamas que cubren las yemas
invernantes.
Nomofilos: (del griego nomo, normal y filo, hoja) son las hojas normales cuya funciones
principales son la fotosíntesis.
En la región reproductiva se diferencian.
Hipsofilos (del griego hipso, cima y filo, hoja) son las hojas que se insertan en niveles altos
de la planta, por encima de los nomofilos. Se presentan como brácteas de las flores o
inflorescencias, con diferentes coloraciones y funciones (protección, atracción de insectos,
etc. )
Antofilos (del griego anto, flor y filo, hoja) son las hojas modificadas que componen la flor:
sépalos, pétalos, estambres y carpelos. Sépalos y pétalos son hojas estériles; estambres y
carpelos son hojas fértiles.
Secuenciación de hojas
Tres ejemplos:
I. Vicia faba L.”haba” (Dicotiledóneas)
II. Zea mays L. “maíz” (Monocotiledóneas)
III. Malus baccata (L.) Borkh (Dicotiledóneas leñosas)
Haba: Se observa una secuencia de hojas que comienza con los cotiledones (hipogeos),
siguen los catafilos y finalmente los nomofilos. Por encima hipsofilos y antofilos. Los catafilos
están formados fundamentalmente por la parte basal de la hoja, que en este grupo
taxonómico (Leguminosas) presenta estípulas. Forman una estructura reducida, la cual, en

su parte apical termina en tres dientes: los cuales se interpretan como las estípulas (los
laterales) y la parte distal reducida (el diente central). En los nomofilos la lámina es
compuesta: en los nomofilos inferiores. presentan dos folíolos, los ubicados más arriba en
cambio, 5-y más folíolos. Entre estos dos tipos extremos hay formas intermedias.
Maíz: Al cotiledón (hipogeo) le siguen los nomofilos. Entre ambos se observa el coleóptilo,
que algunos autores interpretan como la vaina del primer catafilo, el cual carece de lámina.
Los nomofilos están constituidos por una lámina sésil, es decir, no hay pecíolo. La parte
proximal de la hoja forma una vaina abierta. En la cara adaxial, en el límite entre lámina y
vaina se encuentra la lígula, que es un pequeño apéndice membranoso.
En otras monocotiledóneas puede haber uno o varios catafilos, en los nudos inferiores, a los
que siguen, en los nudos subsecuentes, los nomofilos. Estos catafilos están constituidos por
la vaina foliar; en estos catafilos la lámina está reducida o ausente y la proporción de la
lámina aumenta en los sucesivos catafilos, es decir, se presentan formas intermedias entre
los catafilos reducidos casi exclusivamente a la vaina foliar y los nomofilos con la lámina
completamente desarrollada.
Malus baccata. Esta planta es perenne. Después de cada período vegetativo forma yemas
invernantes, que entran en actividad recién el período siguiente. Tales yemas están
cubiertas por catafilos (en este caso, hojas inferiores de cada rama) que aquí cumplen la
función de proteger el meristema apical de la rama frente a las condiciones climáticas
adversas de invierno y del ataque de los insectos. En su estructura son similares a los
catafilos del haba, anchas escamitas con tres dientes en la parte superior, los dos laterales
representan las estípulas y el diente central representa la parte distal de la hoja. Cuando la
yema entra en actividad, se desarrollan hojas en una secuencia que va desde los catafilos a
los nomofilos, pasando por formas intermedias en las cuales hay un cierto desarrollo de la
lámina y de las estípulas. Los otros tipos de hojas, hipsofilos y antofilos desarrollan por
encima de los nomofilos.
En cuanto a los hipsofilos, llamados también brácteas, son las hojas en cuya axila nace una flor o
una inflorescencia. Se encuentran diferentes estructuras en diferentes plantas:
Formados por la base foliar, a veces reducida a apéndices escuamiformes (brácteas
escuamiformes, ej. frutilla) o bien muy desarrollada; un ejemplo de este último caso es la

hoja blanca que acompaña la inflorescencia de la cala, la cual está formada por la base
foliar, y la parte distal solo está representada por un apéndice filiforme. Tales brácteas muy
desarrolladas son comunes en Monocotiledóneas
Formados exclusivamente por la lámina, faltando la base foliar (brácteas laminares)
Similares a los nomofilos, de los cuales no se diferencian (brácteas foliosas)
Filotaxis
Se entiende por filotaxis al estudio de la ordenación o disposición de las hojas alrededor del eje,
incluyéndose todos los tipos de hojas: cotiledones, catafilos, hipsofilos y antofilos.
Fundamentalmente se puede diferenciar dos tipos de disposición: verticilada y alterna.
Disposición verticilada: está caracterizada porque en cada nudo se insertan dos o más
hojas. El conjunto de hojas constituye un verticilo. Por lo tanto se encuentran verticilos
dímeros, trímeros o polímeros. Las hojas verticiladas ocupan posiciones en el espacio que
cumplen las siguientes reglas: (i) regla de equidistancia: las hojas de un mismo verticilo
están separadas entre sí por distancias iguales; (ii) regla de alternancia: en verticilos
sucesivos, las hojas de cada nudo, se ubican exactamente en la mitad de la distancia que
separa entre sí las hojas del verticilo anterior. Se denominan ortósticos a las líneas rectas
imaginarias que pasan por los puntos de inserción de las hojas superpuestas. En la
disposición verticilada en general, el número de ortósticos es igual al doble del número de
hojas que nacen en cada nudo. En el caso particular de verticilos dímeros se habla de
ordenación opuesta, cruciforme o decusada. Aquí las dos hojas de un verticilo están
separadas entre sí por un ángulo de 180° y alternan con las del verticilo siguiente en un
ángulo de 90°. Existen por lo tanto, cuatro ortósticos. En la mayoría de las flores, los
antofilos tienen disposición verticilada que cumplen en general con las reglas de
equidistancia y alternancia. Los verticilos florales toman nombres particulares: cáliz,
formados por sépalos; corola, formada por pétalos; androceo, formados por estambres, y
gineceo, formados por carpelos.
Disposición alterna: se caracteriza por presentar una sola hoja en cada nudo. Se distinguen:
Disposición dística: cuando las hojas se ubican a lo largo del eje, según dos
ortósticos. El ángulo que separa a dos hojas sucesivas o ángulo de divergencia es
de 180°, es decir, ½ de la circunferencia (360°).

o Disposición dispersa o espiralada: cuando las hojas se disponen a lo largo del eje
siguiendo una espiral. La línea espiralada imaginaria que se inicia en la inserción de
una hoja y que une las bases de las hojas sucesivas, hasta alcanzar la base de una
hoja situada sobre el mismo ortóstico que la primera, se denomina espira generatriz.
Las hojas se ordenan a lo largo de la espira generatriz, con un ángulo de
divergencia característico. Este ángulo es menor de 180°, y el valor se expresa por
medio de fracciones de circunferencias que siguen la Serie de Fibonacci. En esta
serie, las fracciones siguen un orden en el cual, cada numerador y denominador es
igual a la suma de los dos valores que le preceden: ½; 1/3; 2/5; 3/8; 5/13; 8/21, etc.
Los ángulos respectivos son 120°; 144°; 135°; 138° 27”; 137° 8”., etc.
¿Cómo se obtiene la fracción? Supongamos que una planta tiene una filotaxis de
2/5. La forma práctica de obtener la fracción filotáxica que le es propia, es
seleccionar dos hojas que se encuentren en el mismo ortóstico; se cuenta el número
de vueltas al trazar la espira generatriz, para llegar de una a otra; el número de
vueltas es el numerador de la fracción y el número de hojas encontradas es el
denominado de la fracción.
La filotaxis de una determinada planta no es necesariamente constante y, a menudo, cambia. En
Monocotiledóneas es característica la disposición dística iniciándose con el cotiledón que se dispone
formando un ángulo de 180° con la primera hoja. Esta disposición se continúa con las hojas
subsiguientes. Esta disposición dística puede más arriba transformarse en disposición dispersa. Y al
llegar a la flor, la disposición de las hojas (antofilos) se vuelve verticilada. Hay otras variantes. En
Dicotiledóneas, los dos cotiledones se disponen en un verticilo, y a partir de allí la filotaxis puede
seguir verticilada, decusada, dística o dispersa, Y dentro de la dispersa, el ángulo de divergencia
puede cambiar.
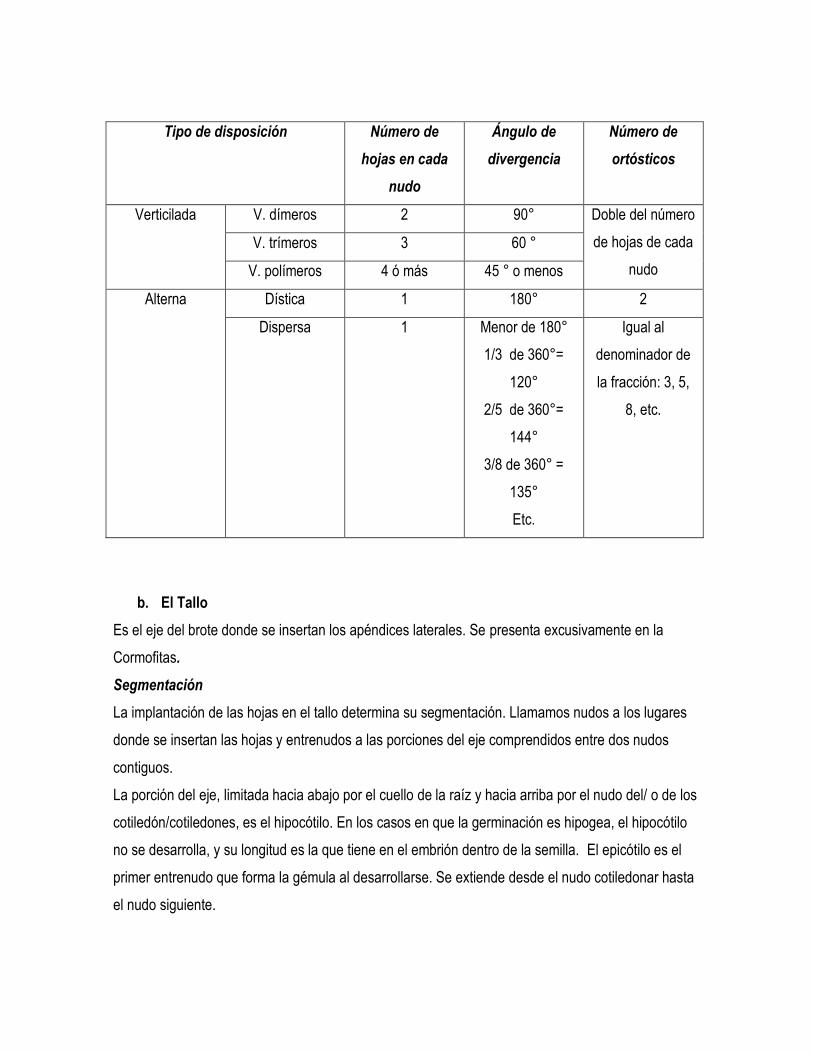
Tipo de disposición Número de
hojas en cada
nudo
Ángulo de
divergencia
Número de
ortósticos
Verticilada V. dímeros 2 90° Doble del número
de hojas de cada
nudo
V. trímeros 3 60 °
V. polímeros 4 ó más 45 ° o menos
Alterna Dística 1 180° 2
Dispersa 1 Menor de 180°
1/3 de 360°=
120°
2/5 de 360°=
144°
3/8 de 360° =
135°
Etc.
Igual al
denominador de
la fracción: 3, 5,
8, etc.
b. El Tallo
Es el eje del brote donde se insertan los apéndices laterales. Se presenta excusivamente en la
Cormofitas.
Segmentación
La implantación de las hojas en el tallo determina su segmentación. Llamamos nudos a los lugares
donde se insertan las hojas y entrenudos a las porciones del eje comprendidos entre dos nudos
contiguos.
La porción del eje, limitada hacia abajo por el cuello de la raíz y hacia arriba por el nudo del/ o de los
cotiledón/cotiledones, es el hipocótilo. En los casos en que la germinación es hipogea, el hipocótilo
no se desarrolla, y su longitud es la que tiene en el embrión dentro de la semilla. El epicótilo es el
primer entrenudo que forma la gémula al desarrollarse. Se extiende desde el nudo cotiledonar hasta
el nudo siguiente.

En las yemas se visualiza la segmentación del eje en nudos y entrenudos, pero estos últimos están
muy acortados. Las hojas aparecen por debajo del ápice propiamente dicho, como primordios
foliares, muy próximos entre sí. El desarrollo de las yemas es el desarrollo del tallo y de las hojas.
Crecimiento
En longitud: Los entrenudos se van alargando debido al crecimiento intercalar que
ocurre por encima de cada nudo, en la base de los entrenudos. La duración de este
crecimiento no es ilimitada, sino que, después de un alargamiento dado que varía
según la especie y las condiciones ambientales, el crecimiento cesa. Por otra parte,
el crecimiento intercalar no alcanza la misma proporción en todos los entrenudos, lo
cual determina que la longitud de los mismos varíe a lo largo del tallo. Esta variación
se manifiesta en un número grande de plantas y en ramas de plantas leñosas a
través de una secuencia tal que, en la base del eje los entrenudos son más cortos,
en la región intermedia cada vez más largos y finalmente en el ápice otra vez más
corto. En las plantas que tienen hojas en roseta, los entrenudos prácticamente no se
desarrollan por crecimiento intercalar y las hojas aparecen muy próximas sobre el
eje. Es importante destacar aquí que la mayoría de la flores se desarrollan como
brotes arrosetados con entrenudos extremadamente cortos. La flor es un brote que
luego de la formación de un determinado número de órganos foliares (sépalos,
pétalos, estambres y carpelos) termina su crecimiento.
En espesor: La yema terminal es la responsable del crecimiento en espesor del eje,
llamado engrosamiento primario o crecimiento de fortificación, que se produce por
debajo del ápice caulinar antes que comience el alargamiento de los entrenudos. La
yema en su totalidad toma, debido a ello, la forma de un cono, de allí la expresión
de cono vegetativo, siendo el ápice caulinar propiamente dicho la parte terminal de
ese cono. La magnitud del engrosamiento primario producido en la yemaestá
relacionado con el estado de desarrollo de la planta: en la juventud es escaso,
aumenta con la madurez hasta llegar a un máximo. Cuando la planta se acondiciona
para formar flores, otra vez disminuye. Esta característica determina la forma del
eje, más delgado en la base, engrosado en la región intermedia y en la parte
superior, nuevamente adelgazado.

Es interesante detacar que los órganos laterales (hojas y raíces adventicias) muestran un
aumento en tamaño que tienen relación directa con el aumento de espesor del eje. El crecimiento de
fortificación está muy extendido en Monocotiledóneas, aunque también en Dicotiledóneas herbáceas
y helechos. En Dicotiledóneas leñosas, el engrosamiento primario o crecimiento de fortificación
aparece enmascarado por un crecimiento secundario, el cual alcanza su máxima intensidad en la
base del eje y disminuye gradualmente hacia el ápice.
III. La raíz
La raíz es un órgano generalmente subterráneo, especializado en la fijación de la planta en el
sustrato y en la absorción de agua y sustancias nutricias.
Las Cormofitas por poseer raíces son rizófitas. Sólo las Psilotales carecen de ellas y si bien hay
otras Cormofitas que no presentan raíces se debe a que sufrieron la pérdida de este órgano durante
el desarrollo. Las Talofitas nunca forman raíces, es decir, son arrizofitas.
En las rizófitas, la raíz se origina durante la germinación a partir de la radícula del embrión, es la raíz
principal, primaria o embrionaria. Las que aparecen posteriormente en cualquier otro lugar de la
planta constituyen las raíces adventicias. Las ramificaciones de la raíz reciben el nombre de raíces
laterales. Según el desarrollo alcanzado por la raíz principal y las raíces adventicias, se pueden
diferenciar dos tipos de sistemas radicales:
Sistema alorrizo: está constituido exclusivamente por la raíz principal que crece mucho y sus
ramificaciones laterales, y forman un sistema radical primario. Este sistema se encuentra en
Dicotiledóneas y Gimnospermas. Estas plantas, sin embargo, bajo condiciones especiales pueden
originar raíces adventicias. Por ejemplo, cuando una rama o trozo de rama llega al suelo; constituye
un método de multiplicación vegetativa de gran aplicación en jardinería y fruticultura.
Sistema homorrizo está constituido principalmente por raíces adventicias que complementan o
reemplazan totalmente la función de absorción y sostén del sistema radical primario. La raíz primaria
puede no crecer o hacerlo en grado variable. Este tipo de sistema radical ocurre, como regla general
en Monocotiledóneas, en Pteridófitas y también en muchas Dicotiledóneas herbáceas. Se presenta
asimismo en algunas Dicotiledóneas leñosas.
BIBLIOGRAFÍA
Troll, W. 1959. Allgemeine Batanik. I-XVI, 1-927, fig. 1-699- Ferdinand Enke Verlag, Sttutgart
Leyenda
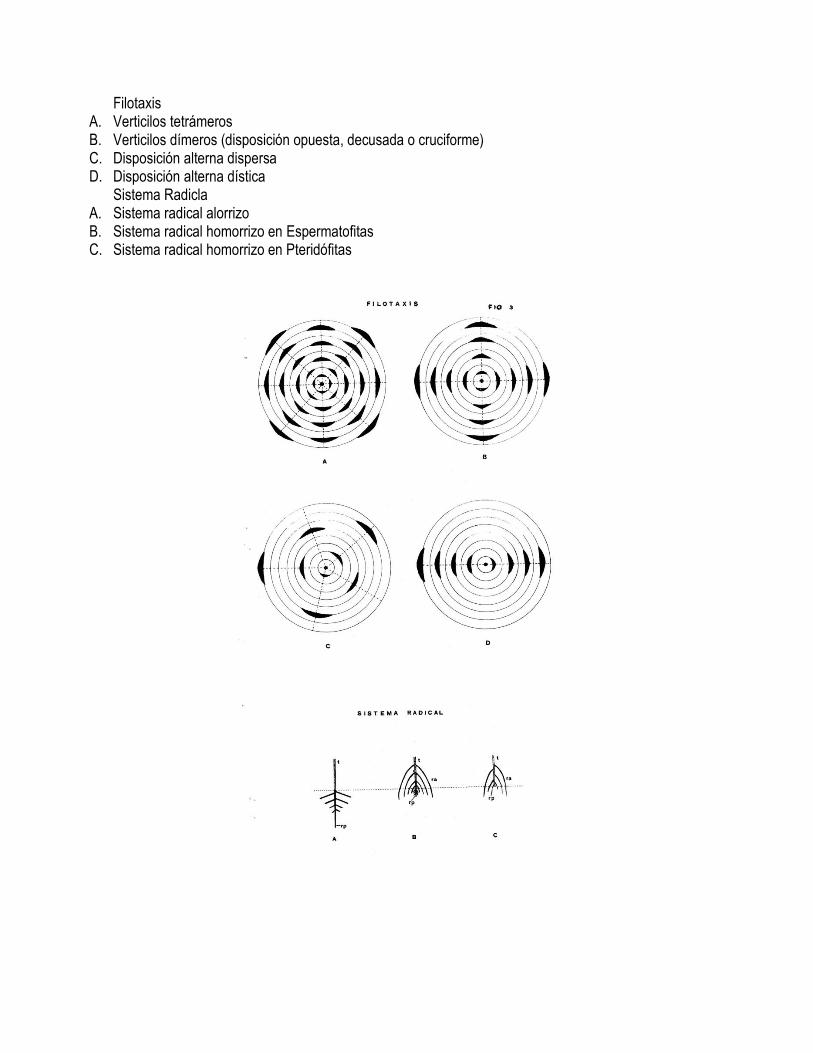
Filotaxis A. Verticilos tetrámeros B. Verticilos dímeros (disposición opuesta, decusada o cruciforme) C. Disposición alterna dispersa D. Disposición alterna dística
Sistema Radicla A. Sistema radical alorrizo B. Sistema radical homorrizo en Espermatofitas C. Sistema radical homorrizo en Pteridófitas

CAPÍTULO 4
RAMIFICACIÓN DEL EJE E INFLORESCENCIA
1. Ramificación del eje
Por ramificación se entiende la producción de ramas a partir de un eje originalmente único. Ramas
de segundo o tercer orden se forman a partir de las de orden inferior, formando el conjunto un
sistema de ramificación.
En Cormofitas, tanto el tallo como la raíz pueden ramificarse, originando un sistema de ramificación
caulinar o radical.
La ramificación del brote se produce de dos maneras
(i) Por ramificación de la yema terminal, es la ramificación dicotómica, que se encuentra en
algunas Pteridofitas (Psilotum, Lycopodium, Selaginella).La característica fundamental
consiste en que el punto vegetativo se divide en dos, originando dos ramas en el
extremo del eje. En el momento en que el ápice se dispone a dividirse, se inhibe su
potencialidad de crecimiento, la cual se transmite a dos puntos que quedan a ambos
lados del ápice. En consecuencia, en lugar del punto vegetativo inicial se han formado
dos nuevos, los que en un momento determinado entran en actividad y comienzan a
originar primordios foliares.
Hay dos casos de dicotomía. Puede ocurrir que los dos nuevos puntos de crecimiento
originen ramas de igual vigor, dando como resultado una ramificación isótoma o
isotomía, que se presenta por ejemplo en algunas especies de Lycopodium. Por el
contrario, las ramas hijas son de distinto vigor en la ramificación anisótoma o
anisotomía. En este caso, cuando las ramas débiles nacen a un lado y al otro,
aparentan tener una posición lateral. Ejemplo de este tipo de ramificación se encuentra
en las especies trepadoras de Lycopodium y Selaginella.
De lo dicho se pueden deducir las dos reglas básicas de la dicotomía: Las ramas se
originan en la yema terminal; y las ramas no están relacionadas con las hojas
ii. Por actividad de yemas axilares: ramificación lateral, que es el tipo de ramificación de
Espermatofitas. Entre las Pteridófitas también se presenta este tipo de ramificación en
Equisetum. La característica fundamental es que las ramas no se forman en la región
apical, sino por debajo de ésta, en regiones laterales del cuerpo de las plantas, hecho al

que debe su nombre. El resultado es un sistema con un eje único y ramas laterales que
son de menor desarrollo que el eje principal.
Las yemas que darán origen a ramas laterales se encuentran en las axilas de las hojas,
por lo que se habla también de una ramificación axilar. La hoja que en su axila lleva el
primordio de la rama recibe el nombre de hoja axilante u hoja tectriz. En general, la
yema axilar está situada en el plano medio o mediana de la hoja axilante.
Las primeras hojas de una rama lateral se denominan profilos. En las Dicotiledóneas se
encuentran generalmente en número de dos, a ambos lados de la mediana de la hoja
axilante. En Monocotiledóneas, como regla general, se encuentra un solo profilo en
posición lateral, o, lo que es más frecuente, en posición adosada. En este último caso
está ubicado entre la rama lateral a la que pertenece, y el eje madre, con el dorso
dirigido hacia este último.
De lo dicho se puede deducir las dos reglas básicas de la ramificación lateral: las ramas
se originan en yemas axilares y la ramificación está estrechamente relacionada con las
hojas.
Yemas accesorias y supernumerarias
En numerosas plantas, en lugar de haber una sola yema en la axila de la hoja, hay varias.
En este caso podemos distinguir una yema principal y otras accesorias o supernumerarias.
Su posición con respecto a la yema principal es variable. Si se encuentran a ambos lados de
la misma, sobre el plano transversal, se denominan colaterales, caso frecuente en
Monocotiledóneas. Cuando se encuentran sobre el plano medio, son yemas seriales,
disposición que es frecuente en Dicotiledóneas. Si las yemas accesorias están por encima
de la yema principal son seriales ascendentes, en caso contrario, seriales descendientes.
Por lo general, las yemas axilares no se desarrollan sino en aquellos casos en que se pierde
la yema principal. Por esta razón son consideradas reservorios de órganos.
Las yemas accesorias tienen gran significación en la región reproductiva de la planta, ya que
en este caso generalmente se desarrollan dando como resultado mayor número de flores y
por lo tanto mayor producción de frutos y semillas
Sistema de ramificación
Como arriba se menciona, la ramificación puede ser dicotómica o lateral. Basándose en esto
se establecen dos sistemas de ramificación:

Sistema dicotómico: aquel originado por ramificación dicotómica. En este sistema se puede
diferenciar dos tipos: isotómico y anisotómico.
Sistema monopodial: aquel que se origina por ramificación lateral. Dentro de éste podemos
distinguir el sistema monopódico y el simpódico. En el tipo monopódico, la yema terminal
tiene crecimiento ilimitado y produce numerosas hojas, en cuyas axilas se desarrollan ramas
de segundo, tercero, cuarto .(…) orden que a su vez pueden ramificarse nuevamente. En el
tipo simpódico, la yema terminal cesa su desarrollo ya sea porque el ápice vegetativo pierde
su funcionalidad o porque se transforma en eje de una inflorescencia. El crecimiento
continúa con el desarrollo de ramas laterales de órdenes sucesivos.
En el tipo simpódico se pueden distinguir monocasios, dicasios, pleocasios.
Dicasio: se obtiene cuando un eje con hojas decusadas (con verticilos dímeros)
finaliza su crecimiento, y el mismo se continúa por dos ramas que nacen en la axila
de las hojas del último nudo más próximo al ápice; esto se repite en las ramas de
órdenes sucesivos
Monocasio: cuando es una sola la rama que continúa el crecimiento. Cuando las
ramas de diferentes órdenes se originan siempre del mismo lado, se forma un
monocasio escorpioide, mientras que si las ramas nacen alternativamente de un
lado y del otro, el monocasio es helicoide.
Pleocasio: se origina cuando las ramas provienen de tres o más yemas laterales del
mismo nudo, en plantas con hojas en verticilos trímeros o polímeros.
Inflorescencia
El sistema de ramificación que se resuelve en flores se denomina inflorescencia. Se pueden
distinguir los tipos de ramificación monopódico y simpódico aunque en inflorescencia se los llama
racemosos y cimoso, respectivamente.
Clasificación: Se distinguen las inflorescencias simples y complejas. Las primeras, tienen flores
como ramas de primer orden, las segundas en cambio producen ramas de primer orden, que no son
flores, y que pueden volver a ramificarse nuevamente. Las ramas laterales constituyen las
inflorescencias parciales
Inflorescencias simples: pueden ser cerradas o abiertas, según el eje termine o no en flor.
Las formas mas importantes son:

Racimo: constituido por un eje principal y ramificaciones de primer orden
consistentes en flores pedunculadas.
Espiga: es semejante al racimo pero en ella las flores no tienen pedúnculo (son
flores sésiles) y salen directamente de la axila de las brácteas.
Amento: es un tipo especial de espiga en el que el eje principal no es rígido y por lo
tanto pende.
Espádice: es una espiga en la que el eje principal es grueso. Se asocia con la
presencia de una espata (hipsofilo o bráctea en la que la base foliar esta muy
desarrollada y la parte distal es reducida)+
Capítulo: el eje es grueso, forma un disco dilatado. En la base de la inflorescencia
hay un involucro, compuesto por numerosos hipsofilos. Una bráctea acompaña
también a cada flor.
Umbela: deriva de un racimo, pero aquí el eje tiene entrenudos extremadamente
cortos y las brácteas están arrosetadas. Las flores aparentan salir de un mismo
punto.
Corimbo: semejante a un racimo, con los pedúnculos florales de diferentes
longitudes. Los inferiores son más largos y se van acortando a medida que se
acercan al ápice. En consecuencia, todas las flores llegan a la misma altura.
Inflorescencias complejas: son aquellas que en lugar de flores laterales, presentan inflorescencias
parciales. Y estas pueden ser racemosas o cimosas. Tanto el eje principal como el eje de las
inflorescencias parciales racemosas, pueden ser abiertos o cerrados. Los ejes de las inflorescencias
parciales cimosas es siempre cerrado.
Inflorescencias complejas con inflorescencias parciales racemosas. Entre ellas tenemos:
Racimo doble: Las inflorescencias parciales son racimos simples abiertos o cerrados. Y se
pueden aquí agrupar todas las inflorescencias que resultan de reemplazar las flores por
inflorescencias simples racemosas
Panícula o panoja: las inflorescencias parciales son racimos (como los racimos dobles). La
diferencia consiste en que en la panícula, las inflorescencias parciales de la base tienen

mayor número de flores y ramificaciones de órdenes más elevados que se van reduciendo
hacia el ápice. En la panícula, tanto el eje principal como los laterales, son ejes cerrados.
Inflorescencias complejas con inflorescencias parciales cimosas. Entre ellas, debemos
considerar:
Tirso: equivale a un racimo doble, pero con inflorescencias parciales cimosas en lugar de
racemosas. Estas pueden ser monocasios, dicasios o pleocasios.
Cimoides: a los cimoides se puede llegar partiendo de tirsos cerrados, en los que solo se
desarrollan las inflorescencias parciales que están en el nudo inmediatamente por debajo de
la flor terminal. Si las inflorescencias parciales son dos y son dicasios, toda la inflorescencia
es un dicasio; si son dos monocasios, es un dicasio de monocasios.
Cuando falta una de las dos inflorescencias parciales, se llega a los monocasios helicoides o
escorpioides.
Si las inflorescencias parciales son más de dos, y están dispuestas en verticilos, se llega a
un pleocasio; en estos las inflorescencias parciales pueden ser pleocasios, dicasios o
monocasios.
Complejidad Inflorescencias parciales Tipos de inflorescencia
Inflorescencias simples Racimo
Espiga
Amento
Espádice
Capítulo
Umbela
Corimbo
Inflorescencias complejas Con inflorescencias parciales racemosas Racimo doble
Otras combinaciones posibles de
inflorescencias simples
Panícula
Con Inflorescencias parciales cimosas Tirsos
Cimoides
BIBLIOGRAFÍA
Troll, W. 1959. Allgemeine Batanik. I-XVI, 1-927, fig. 1-699- Ferdinand Enke Verlag, Sttutgart
Leyenda

Ramificación del eje e Inflorescencias. Abreviaturas: a, b, profilos; c, rama madre; d, rama lateral; e,
hoja tectriz o axilante; m, plano medio; t, plano transversal en Sicotiledóneas (1) y en
Monocotiledóneas (2) y (3) posición lateral y adosada, respectivamente.
INFLORESCENCIAS SIMPLES

INFLORESCENCIAS COMPLEJAS

CAPÍTULO 5
MODIFICACIONES DEL CUERPO DE LAS PLANTAS VASCULARES
1. Duración de la vida de las Plantas Vasculares
En las Espermatofitas la duración de la vida puede estar o no condicionada al momento en que
aparecen las flores. De acuerdo con esto, distinguimos:
Plantas que florecen y fructifican una sola vez y mueren después que las semillas maduraron.
Son las plantas anuales, bienales o plurienales
Plantas que florecen y fructifican más de una vez en la vida. Son las plantas perennes. Son las
plantas en que a la floración/fructificación no le sigue la muerte.
Las plantas anuales son aquellas que desarrollan durante un período que no pasa de un
año y luego que maduran sus frutos, muere. Se agotan después de producir semillas
donde acumulan las reservas. Ej. Girasol, maíz etc.
Las plantas bienales se caracterizan porque su desarrollo se completa en dos años. En
el primer año germinan y forman un cuerpo vegetativo, acumulando en el tallo, raíz u
hojas una cantidad importante de reservas y luego pasan un período de reposo invernal.
En la primavera del año siguiente se forma nuevas hojas y se forman las flores, y se
producen semillas. En este caso, las reservas acumuladas durante el primer año se
utilizan en el segundo año de vida de la planta. En el segundo año de vida, las reservas
se acumulan en las semillas (Ej. zanahoria, remolacha, cebolla).
Las plantas plurienales son plantas en las que el desarrollo del cuerpo vegetativo dura
más de dos años al cabo de los cuales recién florecen y mueren luego de fructificar. Ej.
Algunas palmeras, la pita (agave o henequén); en esta última especie la fase vegetativa
puede durar 10-100 años; cuando fructifican, todas las reservas acumuladas en los
brotes, se agotan, y sólo permanecen las contenidas en las semillas.
Las plantas anuales y bienales pueden, bajo condiciones climáticas determinadas,
extender su período vegetativo a más de un año, transformándose en plurienales.
Las plantas perennes se caracterizan porque la duración de la vida de la planta no está
condicionada a la floración. Viven tres o más años y, lo que es más importante, no agotan
sus reservas en la formación de las semillas sino que parte de ellas se conservan en el
mismo cuerpo vegetativo. De esta manera pueden repetirse varios ciclos de vida en los que
cada período vegetativo va seguido de producción de flores y semillas.

La duración de las plantas perennes varía según las especies. Son perennes las plantas leñosas
como los árboles y arbustos y también las plantas herbáceas con órganos subterráneos persistentes
como el lirio, la papa.
2. Modificaciones del cuerpo de las plantas vasculares
Las modificaciones del cuerpo de las plantas están en relación con el medio ambiente en el que se
desarrolla la planta, la acumulación de reservas, la existencia de un mecanismo de multiplicación
vegetativa.
Modificaciones independientes de la acumulación de reservas
Hoja
Espinas foliares: Se transforma toda la hoja, o una parte de ella. En Berberis, acacia
(Robinia pseudoacacia) y algarrobo, por ej, son las dos estípulas las que se transforman en
espinas.
Zarcillos foliares: son órganos filiformes simples o ramificados que tienen la propiedad de
responder al contacto con determinadas superficies, enrollándose; de este modo fijan el tallo
trepador o rastrero de la planta. Se originan por transformación de la lámina foliar, o de otra
parte de la hoja; en el zapallo, los zarcillos resultan de hojas reducidas a la nervadura
media; en la arveja, la hoja es compuesta, y son los folíolos apicales los que se transforman
en zarcillos; en la leguminosa Lathyrus aphaea, toda la hoja se transforma en zarcillo y la
fotosíntesis la realiza las estípulas que crecen notablemente.
Filodios: el pecíolo dilatado y laminar que reemplaza la lámina en sus funciones; se presenta
en especies australianas del género Acacia; en este caso, la lámina está muy reducida o
ausente.
Tallo
Espinas caulinares: En Crataegus, en Prunus spinosa. Se diferencian de las espinas foliares
porque se originan de una yema axilar; la foliación es reducida o ausente, pero en muchos
casos se identifican los nudos y entrenudos.
Zarcillos caulinares: son producidos por una yema axilar en el género Passiflora; en la vid,
cuya ramificación es simpódica, los zarcillos son producidos por transformación de la yema
terminal de cada rama.

Braquiblastos: Algunas plantas presentan macroblastos y braquiblastos (ramas con
entrenudos cortos). En Berberis vulgaris, los macroblastos no producen hojas normales sino
espinas, y en sus axilas nacen braquiblastos que llevan los nomofilos. En algunas especies
de pino, en los macroblastos se forman hojitas escamosas pardas, de cuyas axilas salen los
braquiplastos y éstos son los que llevan las hojas aciculares.
Cladodios y filóclados: Los cladodios son ramas comprimidas, a veces laminares, siempre
clorofílicas porque deben realizar la fotosíntesis, ya que las hojas son generalmente
rudimentarios y de corta vida. En los miembros de las Cactáceas, también hay cladodios en
el espárrago plumoso. Los filóclados son ramas en forma de hoja, con crecimiento limitado,
y sobre ellas se insertan las flores; el clásico ejemplo se encuentra en el género Ruscus (R.
hypoglossum L.).
Estolones: son ramas laterales de crecimiento horizontal, más o menos delgadas. Pueden
ser epigeos o hipogeos. De acuerdo a su sistema de ramificación pueden ser simpódicos o
monopódicos. En frutilla son monopódicos; en Ajuga reptans son simpódicos.
Raíz
Espinas radicales: En algunas palmeras (Acanthorrhiza sp.) las raíces adventicias se
transforman en espinas
Raíces contráctiles: son estrías que se forman en la superficie de tubérculos, bulbos,
rizomas, o raíces napiformes. La función es llevar la planta a posiciones mas profundas o
favorecer la dispersión. La zanahoria es un ejemplo de una raíz con contracciones.
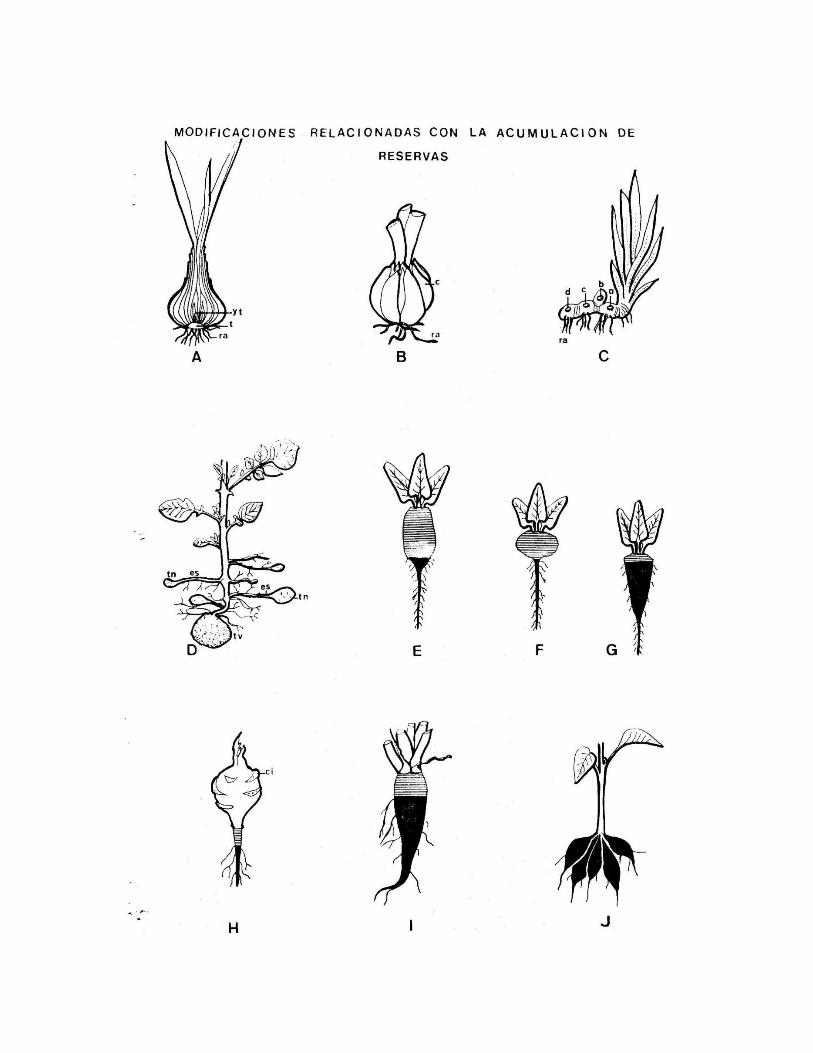
Modificaciones relacionadas con la acumulación de reservas
Hoja: La reserva se hace en las hojas o en determinadas partes de la hoja.
Bulbo tunicado: En la cebolla, la reserva se hace en la vaina de nomofilos. El tallo es corto,
y las vainas son cerradas. El bulbo se forma al final del primer año de vida de la planta.
Bulbo escamoso: la reserva se acumula en catafilos. Ej Lilium
Tallo
Rizoma: son tallos reservantes, subterráneos con crecimiento horizontal que carecen de
nomofilos, pero presentan catafilos. No son raíces porque presentan nudos y entrenudos.
De acuerdo al sistema de ramificacion se distinguen rizomas monopódicos y simpódicos.

Los monopódicos crecen con crecimiento horizontal ilimitado, son perennes, y nunca
emergen a la superficie; en cambio, cada año dan ramas que crecen verticalmente, asoman
a la superficie, dan nomofilos y florecen. Los brotes aéreos se comportan como las plantas
anuales, es decir mueren luego de producir semillas. Los rizomas de crecimiento simpódico
en cambio, dan ramas de crecimiento horizontal, que cada año emergen dando brotes
aéreos con hojas y flores. La parte aérea de la rama muere, luego de producir semillas, pero
queda la base horizontal con yemas y el crecimiento continúa el año siguiente. El rizoma
determina un monocoasio o un dicasio según produzca cada año una o dos ramas. Ej. Lirio
Tubérculo: Tienen crecimiento limitado, a diferencia del rizoma.
En la papa el tubérculo se forma en la porción terminal de estolones subterráneos,
comprendiendo varios entrenudos; las hojas están reducidas a escamas que se
caen muy pronto y dejan la cicatriz; en su axila desarrollan las yemas (los “ojos” de
la papa). En el tubérculo de papa se distinguen dos polos: uno corresponde a la
cicatriz que marca la unión al resto del estolón; el otro polo corresponde a la yema
terminal del estolón. El tubérculo permanece bajo tierra en la estación desfavorable,
con sus yemas latentes, las cuales brotan en la primavera dando lugar a un brote
aéreo que produce flores.
En el colinabo (Brassica oleraceae var. gongyloides), no se desarrollan estolones, y
los tubérculos se forman en segmentos elevados del tallo con nomofilos.
En el rabanito se diferencian dos variedades; en una la reserva se acumula en el
hipocótilo y en menor medida en la raíz; en la otra sólo en el hipocótilo.
En la remolacha existen diferencias entre la forrajera, la roja y la azucarera, en la
forrajera, en el hipocótilo las reservas se acumulan en el hipocótilo y en la raíz, en la
roja, en el hipocótilo, en la azucarera, en la raíz. El rabanito (planta anual) y la
remolacha (planta bienal) utilizan las reservas acumuladas el mismo año o el año
siguiente respectivamente.
Raíz
Tubérculos radicales: Se originan por transformación de raíces adventicias. No tienen hojas
y por ello no tienen yemas axilares.
Raíz napiforme: Se originan por transformación de la raíz principal o primaria


















![Cuerpo Real, Cuerpo Imaginario [Sami-Ali]](https://img.pdfslide.tips/doc/110x75/563dba74550346aa9aa5c9ce/cuerpo-real-cuerpo-imaginario-sami-ali.jpg)










