-
8/9/2019 Captulo II - estructura de las membranas biolgicas
1/6
Federico Rivadeneira
1
C APTULOII
Estructura de las membranas biolgicas
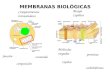
L A BICAPA LIPDICA: ESTRUCTURA Y COMPOSICIN Como sabemos existe
una gran diversidad de fosfolpidos pero una caracterstica que los
rene es su
naturaleza anfiptica. O sea, poseen una cabeza polar constituida
por un glicerol y un fosfato que puede
formar enlaces de hidrgeno con el agua y una cola hidrocarbonada
hidrofbica de 12 a 24 carbonos unidos
a un glicerol 3P. Como sabemos, estas cadenas pueden ser
saturadas o insaturadas, provocando la presencia
de dobles enlaces la disminucin del PF e incrementando el mismo
mediante el largo de la cadena;esto es
debido a la mayor cantidad de interacciones de Van der Waals que
se producen.
Respecto a la composicin relativa de cadenas saturadas e
insaturadas podemos decir que la
cantidad de cadenas insaturadas en las membranas bacterianas es
inversamente proporcional a la
temperatura, o sea, al aumentar la temperatura disminuye la
cantidad de cidos grasos insaturados
produciendo este efecto un aumento del PF. Esto no ocurre en
clulas animales que poseen un ambiente
ms regulado por lo que la composicin vara poco con la
temperatura externa.
Los glicerofosfolpidos son los componentes
mayoritarios de las membranas celulares y estn esterificados
en los C1 y C2 del glicerol-3-fosfato. Las combinaciones de
acilos
unidos al C1 y C3 incrementa la variedad de los mismos y
adems aumenta esta diversidad al unirse un OH de otro
compuesto al grupo fosfato en el C3 (serina, etanolamina,
colina, etc.)
Un segundo grupo lo constituyen los esfingolpidos
formados a partir de un ncleo de esfingosina conformado por
un alcohol de cadena larga que contiene un grupo amino donde
se puede unir un cido graso y un hidroxilo en C1 donde se
puede unir una azcar para formar la cabeza
polar. Un cido graso (imagen) en el grupo
amino del C2 conforma un esfingolpido
llamado ceramida; precursor de la mayora de
los esfingolpidos que los podemos clasificar en esfingomielinas,
cerebrsidos y ganglisidos.
Otra molcula es el colesterol que ubica su funcin hidroxilo a la
cara acuosa de lamembrana bajando la interaccin entre las cadenas
hidrocarbonadas con su parte no polar y
disminuyendo por esto el PF.
La estabilidad de la bicapa lipdica es en gran parte debido a
fuerzas de Van der Waals
entre las cadenas hidrocarbonadas y por la misma razn de que
esta es la mayor barrera
energtica, es difcil que un extremo polar pase hacia el otro
lado de la membrana. Por esto,
la bicapa posee una fluidez restringida y una composicin
diferente tanto del lado externo como del interno.
-
8/9/2019 Captulo II - estructura de las membranas biolgicas
2/6
Federico Rivadeneira
2
Fluidez de la bicapa lipdicaLa fluidez depende de la composicin
fosfolipdica de la membrana y su PF oscila entre 30 y 40 C.
PROTENAS DE MEMBRANA: DISTINTOS TIPOS Y FUNCIN Como sabemos las
protenas de membrana las podemos clasificar como protenas
integrales de
membrana y protenas perifricas. Estas ltimas se caracterizan por
poseer dos propiedades; la primera es
que se pueden disociar en un medio suave como mediante un cambio
en la fuerza inica del medio o
mediante la extraccin de medios quelantes; la segunda es que se
separan sin estar asociadas a lpidos y que
se puede disolver en pH neutro. Esto indica que se mantienen
unidas mediante uniones dbiles y que no se
encuentran asociadas fuertemente a lpidos (Citocromo C y
espectrina son ejemplos).
En cambio las protenas integrales de membrana no se disocian tan
fcilmente por lo que se
mantienen unidas a lpidos generalmente y por lo mismo, son
insolubles en agua.
Mo del o de m os aic o fluid o
Este modelo implica el movimiento lateral de las protenas tanto
perifricas como integrales en la
bicapa. Este movimiento en algunos casos se ve limitado por la
asociacin de las mismas con el
citoesqueleto.
Tran s p o rte a trav s de membrana s
Debido a la naturaleza hidrofbica de la membrana y a su estrecha
separacin las molculas polares
o muy grandes no pueden atravesar la membrana por simple difusin
por lo que las protenas cumplen esta
funcin. Estas pueden ser protenas transportadoras o carriers que
se unen al soluto a transportar y
mediante un cambio conformacional lo transportan. Otro tipo son
las protenas de canal que no se unen al
soluto y generan un canal hidroflica permitiendo el paso de
solutos de cierto tamao y carga.
El transporte, a su vez, puede ser pasivo (sin aporte endgeno de
energa) o activo (con un aporte
endgeno de energa), debido a que las molculas difunden
pasivamente mediante gradientes de
concentracin, elctricos o ambos a la vez.
Tran s p o rte de m o lcula s pequea s El transporte de estas
molculas puede ser pasivo (difusin simple y difusin facilitada) o
activo.
1. Difusin simple: Este tipo de difusin ocurre mediante un
gradiente de concentracin y el mismopuede ser cuantificado
mediante:
Siendo D el coeficiente de difusin y L la longitud de la
membrana a atravesar por el soluto. El
cambio de energa libre depende directamente del gradiente de
concentracin y en caso de no ser unelectrolito queda determinado
por:
Otra magnitud de importancia es el coeficiente de permeabilidad
de la membrana que es el cociente
entre el coeficiente de difusin D y L.
-
8/9/2019 Captulo II - estructura de las membranas biolgicas
3/6
Federico Rivadeneira
3
2. Difusin facilitada: La difusin facilitada tambin depende de
los gradientes de concentracinpero la diferencia principal es que
est mediada por protenas integrales de membrana. Este tipo
de transporte puede ser a la vez antiporte o simporte. En el
caso del transporte pasivo, la
difusin facilitada posee un lmite superior de velocidad
determinado por la cantidad de
protenas presentes. Por lo que la velocidad puede ser descripta
por:
Tra s po rte activ o El transporte activo, al actuar contra un
gradiente necesita energa y protenas de membrana. El
mismo puede ser Primario, Secundario o de Translocacin de
grupo.
1. T ransporte activo primario : Requiere un aporte primaria de
energa como la hidrlisis del ATP,transporte de electrones o
fotones, ejemplos son la Na +-K+ ATPasa, la Ca ++ ATPasa o el
transporte
de H + por la bacteriorrodopsina.1.1 Actividad Na +-K+ ATPasa:
La concentracin de K + intracelular es mayor que la extracelular y
la
de Na + extracelular es mayor que la intracelular; esto indica
que existe un gradiente de
concentracin de K + hacia el LEC y viceversa. Por esto mismo la
concentracin es mantenida
por la enzima que por cada molcula de ATP hidrolizada extrae 3
Na + por cada 2 K + que
entran a la clula.
Es una protena integral compuesta por dos subunidades de
estequiometria a 2b2. La
subunidad mayor a posee los sitios de unin para los iones e
hidroliza el ATP mientras que la
b ms pequea es una glicoprotena de funcin desconocida.
2. Transporte activo secundario: En este tipo de transporte la
fuente de energa son gradientesinicos y de potencial a travs de
membrana. O sea, son sistemas de transporte que se hayan
acoplados a otros transportes como en el caso del transporte de
glucosa al epitelio intestinal. En
este caso la Na +-K+ ATPasa acta haciendo salir Na + al LEC y
luego, debido al gradiente de
concentracin establecido la glucosa utiliza la energa liberada
para entrar al epitelio desde el
lumen.
3. Translocacin de grupo: Este tipo involucra una reaccin
enzimtica donde una azcar esfosforilado al ser transportado
mediante un PEP donor de P - (fosfotransferasa dependiente de
PEP), este proceso transporta en E . coli alrededor de diez
tipos de azcares.
P o tencial de membrana
El potencial de membrana, de acuerdo al tipo celular vara de
-20mV a -200mV. En este caso la
bomba de sodio y potasio juega un papel importante pero tambin
existen los canales de fuga de K + que
actan a favor del gradiente electroqumico. Esta no es la nica
funcin de la bomba, pues tambin tomaparte en el transporte activo
de azcares y aminocidos y en el control del volumen celular. Por
esto mismo,
la actividad de la bomba que necesita de ATP consume
aproximadamente un tercio de la energa disponible.
Siendo posible que se inhiba la misma mediante la ouabana, hecho
que hace reventar a las clulas.
-
8/9/2019 Captulo II - estructura de las membranas biolgicas
4/6
Federico Rivadeneira
4
Tran s p o rte de i o ne s a trav s de la s membrana s bi o
lgica s
Muchos de los sistemas de transporte activo son acoplados ya
sean unidireccionales o de
intercambio mediante el aprovechamiento de la energa liberada
mediante el transporte a favor de ungradiente electroqumico.
La mayora de estos transportes involucran el cotransporte con Na
+ y su gradiente del LEC hacia el
citoplasma que es mantenido por la bomba sodio potasio que
extrae el sodio hacia afuera de la clula.Transporte que es
realizado mediante la unin del Na + y el soluto a diferentes
posiciones de la protena.
Regulacin del pH intracelular:El mantenimiento del pH es vital;
las enzimas lisosmicas actan a pH 5 y las citoslicas a un pH
7,2. Este equilibrio est tambin mediado por transporte acoplado
al gradiente de Na +. Estos son:
1. Intercambiador de Na +-H+ que extrae protones al ingresar Na
+.2. Intercambiador de Cl - - HCO3- impulsado por Na + que acopla
la entrada de Na + con HCO 3-.
Estos se ven afectados por el pH citoslico. Tambin existe un
transportador impulsado por Na +
unidireccional que acopla la entrada de uno o varios HCO 3-,
proceso que se activa cuando disminuye
el potencial de membrana.
C anale s inic os
Los canales selectivos forman conductos de comunicacin
hidroflicos directamente entre el
citoplasma y el LEC, por lo mismo deben ser altamente
selectivos. Este es el caso de las porinas que son
protenas formadoras de canales en membranas de bacterias,
mitocondrias y cloroplastos, los cuales sonrelativamente
grandes.
En el caso de los canales de iones inorgnicos (Na +, K+, Ca++ y
Cl-) simplemente los llamamos canales
inicos y son transportadores pasivos que cumplen su funcin a
favor de un gradiente electroqumico a una
tasa mayor que cualquier otro transportador. Estos canales no
estn permanentemente abiertos sino quepueden ser regulados ya sea,
mediante un ligando, mediante el potencial de membrana,
mecnicamente o
por la fosforilacin y defosforilacin de sus protenas.
TRANSPORTE DE MACROMOLCULAS Y PARTCULAS: ENDOCITOSIS Y F
AGOCITOSIS Las sustancias de alto peso molecular no pueden entrar
libremente a la clula por lo que utilizan un
sistema de endomembranas que nombramos generalmente como
endocitosis. El mismo, podemos dividirlo
en dos tipos morfolgica y funcionalmente diferentes: pinocitosis
y fagocitosis. Tambin existe un
mecanismo de transporte transcelular que implica a las vesculas
que no se fusionan con lisosomas sino que
lo hacen con la membrana liberando las sustancias que contienen
en el LEC.
1. Fagocitosis: La fagocitosis es un proceso activo que implica
la invaginacin de la membrana entorno a la sustancia a ingerir y la
posterior estrangulacin de la misma formando el fagosoma (de
50 a 400nm) que por transporte microtubular se dirige a la regin
del Golgi fusionndose con los
lisosomas primarios para formar los lisosomas secundarios.
2. Pinocitosis: En este caso la clula invagina porciones de la
membrana para incorporar LEC consolutos, proceso acompaado por un
gran movimiento citoplasmtico en su alrededor que
-
8/9/2019 Captulo II - estructura de las membranas biolgicas
5/6
Federico Rivadeneira
5
forma las vacuolas o macropinosomas que al ingresar en hilera al
citoplasma van disminuyendo
su tamao para luego fusionarse y ser finalmente digeridos como
en la fagocitosis.
Existe otra clase de vesculas (50-250nm) que quiz formen parte
del transporte intracelular
llamadas vesculas revestidas. Sin embargo otras se generan
continuamente en ciertas regiones llamadas
depresiones revestidas que constituyen el 2% de la superficie de
la misma. Estas vesculas revestidas pierden
tempranamente el revestimiento constituido por una unidad bsica,
el complejo llamado trisquelin,
formado por tres polipptidos de clatrina con tres polipptidos
menores formando un poliedro pentagonal.
E nd o cit os i s mediada p o r recept o r:
La endocitosis mediada por receptor es un proceso especfico
donde la macromolcula se une a
receptores en el lado extracelular de la membrana y en una regin
de depresin revestida produciendo un
transporte de mayor velocidad relativa respecto a la
pinocitosis.
Este es el proceso por el cual se incorpora colesterol y Fe2+ al
interior celular mediante las LDL y la
transferrina respectivamente. La LDL se une a receptores
expresados en la membrana en funcin del
metabolismo de colesterol entonces la membrana se invagina y
estrangula formando el endosoma que siguela misma va fagoctica.
L A MEMBRANA DEL ERITROCITO La electroforesis con gel de
poliacrilamida y SDS revela la presencia de 15
bandas proteicas de las cuales tres, la espectrina, glicoforina
y la banda 3
constituyen ms del 60% del peso de las protenas de esta
membrana.
La espectrina es una protena perifrica citoplasmtica que
constituye el
25% de la masa total de protenas total, esta es la encargada de
mantener la
estructura de la clula y la forma bicncava. La misma es un
heterodmero formado
por dos grandes subunidades similares. Estos heterodmeros
consisten en dos
cadenas polipeptdicas antiparalelas y flexibles y . Esta protena
se enlaza a
filamentos cortos de actina y la banda 4.1 formando un complejo
de unin.
Esta red de espectrina es unida mediante una protena de unin
a
membrana llamada banda 3, mientras que la 4.1 se une a la
espectrina, a la actina y al dominio citoplasmtico de la
banda 3 y la glicoforina.
Esta ltima es una posee unos 16 oligosacridos
orientados hacia la zona externa, cuya funcin no se conoce.
La banda 3 es una protena transmembrana de paso mltiple con 14
pasos, que transporta el CO 2
mediante un intercambiador de CO 3H- por Cl -.
-
8/9/2019 Captulo II - estructura de las membranas biolgicas
6/6
Federico Rivadeneira
6