Embed Size (px)
Citation preview

I
Caracterização biofísica dos 5000 m crol
Dissertação apresentada à Faculdade de Desporto da Universidade do Porto, no
âmbito do 2º Ciclo de estudos conducente ao grau de Mestre em Treino de Alto
Rendimento Desportivo – Natação, de acordo com o Decreto-lei nº 74/2006 de
24 de março.
Orientador: Professor Doutor Ricardo Fernandes
Coorientador: Professor Doutor João Paulo Vilas-Boas
Vânia Soares Neves
Porto, setembro de 2017

II
Neves, V. (2017). Caracterização biofísica dos 5000 m crol. Dissertação de 2º
ciclo em Treino de Alto Rendimento Desportivo. Faculdade de Desporto da
Universidade do Porto.
Palavras-Chave: NATAÇÃO, BIOENERGÉTICA, BIOMECÂNICA, TÉCNICA DE
CROL.

III
Agradecimentos
A concretização deste trabalho constitui um marco muito importante na minha
vida, que só foi possível concretizar com a preciosa contribuição, confiança e
encorajamento de várias pessoas, às quais não posso deixar de expressar os
meus sinceros e profundos agradecimentos:
- Ao Professor Doutor Ricardo Fernandes por tudo que me ensinou ao longo dos
anos, pela orientação prestada, disponibilidade e por me proporcionar uma visão
diferente de tudo que envolve a natação.
- Ao Professor Doutor João Paulo Vilas-Boas por todo o apoio e por ter
acreditado neste trabalho desde o primeiro momento.
- À Professora Doutora Susana Soares por todas as conversas sobre leituras
perdidas ou apaixonantes, por todos os momentos de desabafo e pela ajuda
prestada nas recolhas deste trabalho.
- Ao Mestre e Doutorando Rodrigo Zacca por todos os conselhos dados, pela
incansável disponibilidade, pela amizade ao longo de todo este percurso e pela
ajuda nas recolhas, tratamento de dados e estruturação do trabalho. Obrigada
por acreditar em mim, mais que eu mesma… “O Rio é seu… só tem de
acreditar!”.
- Ao Professor Doutor Luís Rama pela disponibilidade e ajuda nas recolhas feitas
em Coimbra e por me possibilitar conversar e reviver momentos do nosso
passado.
- A todos os nadadores que se disponibilizaram a participar neste trabalho, pois
sem o seu empenho e entrega nada seria possível.
- Aos meus pais, pela educação que tive, pelo apoio e acompanhamento
incansáveis e por tudo o que fizeram por mim desde o meu primeiro dia de vida.
A eles agradeço toda a amizade e amor, todas as lições de vida e chamadas de

IV
atenção, todas as conversas e conselhos, todos os momentos que vivemos e
todos aqueles que ainda estão por vir.
- Ao Tiago por todo o apoio e incentivo que sempre me deu, essenciais para esta
motivação e por querer chegar sempre mais longe. Por todas horas de mimo e
conversa e por todas as outras que perdemos, pela paciência nos momentos
mais difíceis e por todo o amor e carinho, pois já Gandhi dizia: “Onde há amor,
há vida”.
- A toda a minha família, pelo apoio constante ao longo de todo meu percurso
académico pois sem este suporte tudo seria mais complicado.
- A todos os treinadores que comigo se cruzaram ao longo de toda a minha
carreira desportiva, responsáveis por fizer nascer e crescer este “bichinho” da
natação que há dentro de mim.
- À Drequinhas, à Inesinha, ao Mi, ao Sérgio, à Cameira e ao Benny, meus
amigos de sempre (e para sempre) por todas as palavras de ânimo e
compreensão, por estarem sempre presentes e por serem quem são.

V
Índice
Índice de figuras ............................................................................................... VII
Índice de tabelas ............................................................................................... IX
Índice de equações ........................................................................................... XI
Resumo ........................................................................................................... XIII
Abstract ........................................................................................................... XV
Abreviaturas .................................................................................................. XVII
1. Introdução ................................................................................................ 1
2. Material e métodos ................................................................................. 11
2.1 Caracterização da amostra ................................................................. 11
2.2 Procedimento experimental ................................................................. 12
2.3 Análise estatística ............................................................................... 17
3. Resultados.............................................................................................. 19
3.1 Estudo 1 .............................................................................................. 19
3.2 Estudo 2 .............................................................................................. 21
3.3 Estudo 3 .............................................................................................. 24
4. Discussão ............................................................................................... 29
5. Conclusão .............................................................................................. 41
6. Sugestões para estudos futuros ............................................................. 43
7. Bibliografia .............................................................................................. 45

VI

VII
Índice de figuras
Figura 1: Diagrama síntese dos fatores determinantes do rendimento desportivo
do nadador (Fernandes & Vilas-Boas, 2006). .................................................... 6
Figura 2: Analisadores portáteis de lactato (Lactate Pro, Arkay, Inc., Quioto,
Japão) e de glicemia (One Touch Select Plus) utilizados nas avaliações. ....... 13
Figura 3: Avaliação dos parâmetros ventilatórios com o analisador portátil de
gases (K4b2, Cosmed, Roma, Itália) ................................................................ 14
Figura 4: Filmagem do teste de 5 x 1000 m com a camara HDR-CX160E, Sony,
Quioto, Japão ................................................................................................... 16
Figura 5: Cronofrequencimetro Finis (Califórnia, Estados Unidos) e utilizado para
controlo da FG dos nadadores ......................................................................... 17
Figura 6: Contribuição energética no teste 5 x 1000 m crol. ............................ 27
Figura 7: Exemplo de uma curva típica da modelagem realizada para a cinética
do VO2 à cada 1000m. .................................................................................... 33

VIII

IX
Índice de tabelas
Tabela 1: Principais resultados internacionais dos nadadores portugueses de
águas abertas entre 2012 e 2016. ...................................................................... 4
Tabela 2: Caracterização das principais características físicas e do background
do treino dos nadadores participantes no estudo 1. ......................................... 11
Tabela 3: Caracterização das principais características físicas e do background
do treino dos nadadores participantes no estudo 2. ......................................... 12
Tabela 4: Valores médios ± DP das variáveis bioenergéticas do estudo 1 e
respetiva comparação das médias dos dois géneros. ...................................... 19
Tabela 5: Média ± DP das variáveis biomecânicas do estudo 1 (n=16). .......... 20
Tabela 6: Valores médios ± DP das variáveis biomecânicas do estudo 1 e
respetiva comparação das médias dos dois géneros. ...................................... 21
Tabela 7: Média ± DP das variáveis bioenergéticas e biomecânicas avaliadas no
estudo 2............................................................................................................ 22
Tabela 8: Valores médios ± DP das variáveis bioenergéticas e biomecânicas em
cada um dos géneros no estudo 2. .................................................................. 23
Tabela 9: Fatores bioenergéticos e biomecânicos dos 5 x 1000 m crol do estudo
3 (n=2). ............................................................................................................. 25
Tabela 10: Correlação das variáveis bioenergéticas e biomecânicas com a
performance no estudo 3 ................................................................................. 26
Tabela 11: Valores de média ± DP da cinética VO2 do teste de 5 x 1000 m crol.
......................................................................................................................... 27
Tabela 12: Definição dos domínios de intensidade e zonas bioenergéticas de
treino correspondentes (Fernandes, 2013). ..................................................... 31

X

XI
Índice de equações
Equação 1 v = E ∙ (ep ∙ D-1) ........................................................................ 6
Equação 2 VO2(t) = A0 + Ap ∙ (1-e(t-TDp) τp⁄ ) .............................................. 14
Equação 3 Anala = b ∙ ⌊La-1⌋
basal ∙ M ......................................................... 15
Equação 4 C = E ∙ v-1 ................................................................................ 15
Equação 5 FG = 3 ∙ t3-1
............................................................................... 16
Equação 6 DC = v ∙ FG-1
........................................................................... 16

XII

XIII
Resumo
O futuro da natação em águas abertas, entendido como a intenção de
desenvolver a capacidade de desempenho desses nadadores específicos,
depende do aumento do conhecimento científico sobre esse tema. O nosso
objetivo passa por identificar e caracterizar as determinantes biofísicas em
5000m crol. Trinta e quatro nadadores foram avaliados numa competição e 10
num protocolo de 5 x 1000 m para a avaliação da concentração sanguínea de
lactato ([La-]), glicemia e variáveis biomecânicas. Após isso, dois nadadores
realizaram 5 x 1000 m, onde se incluiu a análise das variáveis ventilatórias e
absorção de oxigénio (cinética VO2). A velocidade média (v) para a prova de
5000 m foi de 1.31 ± 0.08 m∙s-1, com 35.0 ± 3.2 ciclos∙min-1 para a frequência
gestual (FG), 2.49 ± 0.3 m∙ciclo-1 para distância por ciclo (DC), 3.26 ± 0.5 m2∙s-
1.ciclo-1 para o índice braçada (IB), 3.0 ± 1.6 mmol∙L-1 para [La-] e 115 ± 21 mg∙dL-
1 para glicemia. No teste de 5 x 1000 m, a média da v foi de 1.29 ± 0.01 m∙s-1,
FG foi de 33.0 ± 3.4 ciclos∙min-1, DC foi de 2.62 ± 0.3 m∙ciclo-1, IB foi de 3.39 ±
0.4 m2∙s-1∙ciclo-1, [La-] foi 2.3 ± 1.1 mmol∙L-1 e a glicemia foi de 96 ± 20 mg∙dL-1.
Durante o teste de 5 x 1000 m onde se utilizou o analisador de gás portátil, os
valores variaram entre 1.30 ± 0.03 m∙s-1 para v, 34.7 ± 1.1 ciclos∙min-1 para FG,
2.47 ± 0.04 m∙ciclo-1 para DC, 3.20 ± 0.08 m2∙s-1∙ciclo-1 para IB, 2.0 ± 0.3 mmol∙L-
1 para [La-] e 92 ± 9 mg∙dL-1 para a glicemia. VO2, ventilação pulmonar e
frequência cardíaca não mudaram em função do tempo. O gasto energético e o
custo energético variaram em torno de 576 ± 70 kJ e 0.6 ± 0.07 kJ∙m-1 com a
fonte aeróbia representando ~ 99,5%. Os resultados permitem concluir que, em
exercícios de longa distância com intensidade baixa a moderada, a contribuição
energética aeróbia desempenha um papel fundamental na demanda de energia
dos nadadores e que ocorre um estado de equilíbrio fisiológico associado a
ajustes biomecânicos.
Palavras-chave: NATAÇÃO, FISIOLOGIA, BIOMECÂNICA, TÉNCICA DE CROL

XIV

XV
Abstract
The future of open water swimming, understood as the intention to develop the
performance capacity of these specific swimmers, depends on the increase of
scientific knowledge on this topic. Our purpose was to identify and characterize
the biophysical determinants in 5000 m front crawl. Thirty-four swimmers were
evaluated in a formal competition and 10 in a 5 x 1000 m protocol for the
assessment of blood lactate concentrations ([La-]), blood glucose and
biomechanical variables. Afterwards, two swimmers performed 5 x 1000 m
including the analysis of ventilatory variables and oxygen uptake (VO2) kinetics.
Mean velocity (v) for the 5000 m event was 1.31 ± 0.08 m.s-1, with 35.0 ± 3.2
cycles.min-1 for stroke rate (SR), 2.49 ± 0.3 m.cycle-1 for stroke length (SI), 3.26
± 0.5 m2.s-1.cycle-1 for stroke index (SI), 3.0 ± 1.6 mmol.L-1 for [La-] and 115 ± 21
mg.dL-1 for blood glucose. In the 5 x 1000 m test the mean v was 1.29 ± 0.01 m.s-
1, SR was 33.0 ± 3.4 cycles.min-1, SL was 2.62 ±0.3 m.cycle-1, SI was 3.39 ± 0.4
m2.s-1.cycle-1, [La-] were 2.3 ± 1.1 mmol.L-1 and blood glucose were 96 ± 20
mg.dL-1. During the 5 x 1000 m test using the portable gas analyzer, values
ranged around 1.30 ± 0.03 m.s-1 for v, 34.7 ± 1.1 cycles.min-1 for SR, 2.47 ± 0.04
m.cycle-1 for SI, 3.20 ± 0.08 m2.s-1.cycle-1 for SI, 2.0 ± 0.3 mmol.L-1 for [La-] and
92 ± 9 mg.dL-1 for blood glucose. VO2, pulmonary ventilation and heart rate did
not change as function of time. The energy expenditure and energy cost ranged
around 576 ± 70 kJ and 0.6 ± 0.07 kJ.m-1 with the aerobic pathway accounting
for ~99.5%. Results allow us to conclude that in long distance exercises at low to
moderate intensity, the aerobic energy contribution plays a fundamental role in
the energy demand of the swimmers and that a physiological steady state occur
associated to biomechanical adjustments.
Key words: SWIMMING, PHYSIOLOGY, BIOMECHANICS, FRONT CRAWL

XVI

XVII
Abreviaturas
% FCmáx Percentagem da frequência cardíaca máxima
[La-] Concentração sanguínea de lactato
[La-]basal Concentração sanguínea de lactato em repouso
± Mais ou menos
♀ Género feminino
♂ Género masculino
A Amplitude do consumo de oxigénio
Aer Componente energética aeróbia
Anaala Componente energética anaeróbia alática
Anala Componente energética anaeróbia lática
b Equivalente energético
bpm Batimentos por minuto
C Custo energético
cm Centímetros
D Arrasto hidrodinâmico
DC Distância percorrida por ciclo
DP Desvio-padrão
Ė Dispêndio energético
ep Eficiência mecânica propulsiva
FC Frequência cardíaca
FG Frequência gestual
FINA Fédération Internationale de Natation
FPN Federação Portuguesa de Natação
IB Índice de braçada
kJ Kilojoule
km Quilómetros

XVIII
LAN Limiar Anaeróbio
LEN Ligue Européenne de Natation
M Massa corporal
min Minutos
ml Mililitros
MLSS Máxima fase estável de lactato
O2 Oxigénio
R Quociente respiratório
s Segundos
SPSS Statistical Package for the Social Sciences
t Tempo
τ Constante de tempo
TD Tempo de atraso
v Velocidade
VE Ventilação pulmonar
VO2 Consumo de oxigénio
VO2basal Consumo de oxigénio em repouso

1
1. Introdução
A natação é uma modalidade desportiva individual, contínua e cíclica. Como
disciplina desportiva é uma atividade multidisciplinar, podendo ser extremamente
rica ao nível da experimentação motora e variação de práticas desportivas
aquáticas (Canossa et al., 2007). É também uma modalidade mista, uma vez
que o rendimento desportivo do nadador está intimamente dependente de
fatores bioenergéticos e biomecânicos (Fernandes & Vilas-Boas, 2006).
Tradicionalmente são associadas a esta modalidade quatro disciplinas: natação
pura, polo aquático, natação sincronizada e saltos para a água, às quais
podemos acrescentar, na atualidade, a natação de águas abertas. Para além
destas disciplinas, podemos ainda relacionar a esta modalidade, a natação de
salvamento aquático e a natação como segmento do triatlo.
Embora as origens da natação de águas abertas remontem ao século XIX,
podendo mesmo ser confundidas com as origens da natação, é uma das mais
recentes disciplinas da Fédération Internationale de Natation (FINA). Aliás, este
órgão que dirige a natação a nível internacional reconheceu oficialmente a
natação de águas abertas como disciplina em 1986 (Kruger et al., 2011), tendo
o primeiro Campeonato do Mundo sido realizado em Perth, em 1991, com a
prova de 25 km, cidade que em 1988 acolheu também a primeira prova mundial
de 5 km. A prova de 10 km teve a sua primeira edição em Fukuoka, em 2001.
Esta disciplina acabou por conquistar definitivamente o seu lugar com a
integração da prova de 10 km nos Jogos Olímpicos de Pequim, em 2008.
Atualmente são disputados títulos mundiais e europeus nas provas individuais
de 5, 10 e 25 km e a prova coletiva de 4 x 1250 m.
Para além da organização de Jogos Olímpicos e de Campeonatos do Mundo, a
FINA define os regulamentos de todas as competições e promove anualmente a
10 km Marathon Swimming World Cup, constituída por várias etapas disputadas
em diversos países, e o Open Water Swimming Grand Prix, composta por
múltiplas etapas de distâncias que variam entre 15 e 88 km. Os Campeonatos
da Europa são organizados pela Ligue Européenne de Natation (LEN), que
supervisiona os desportos aquáticos na Europa e se encontra filiada à FINA. A

2
LEN também organiza uma competição de elite, a Len Cup, disputada em várias
etapas com distâncias que vão dos 5 aos 25 km.
A natação de águas abertas ocorre num ambiente distinto da natação pura, uma
vez que a dinâmica do meio que os nadadores podem encontrar é muito variada,
sendo isto, por si só, um elemento desafiador das capacidades de cada um
(Borges, 2015). A natação de águas abertas apresenta aspetos singulares que
a convertem numa disciplina com caráter próprio e com necessidades de
preparação diferentes (Swift, 2003). Existem competições disputadas em
diferentes espaços (rios, lagos, albufeiras, mar e pistas), com água salgada ou
doce, em águas paradas ou com correntes e com temperaturas que podem variar
entre os 16 e os 31ºC (Borges, 2015).
As provas de águas abertas podem apresentar diferentes tipos de partidas (com
ou sem plataforma e com ou sem ordem pré-estabelecida), diferentes
configurações e número variado de voltas e de boias, variando também as
condições de abastecimento, número de nadadores a participar, estratégias e
velocidades de nado. As estratégias pré-definidas podem fazer a diferença entre
uma prova bem conseguida ou não e, mesmo sendo discutidas e definidas com
os treinadores, as táticas bem-sucedidas requerem determinação e reajustes no
decorrer da prova.
Para além destas características, parece que outras a diferenciam da natação
pura: a distância percorrida, as referências visuais, o nado em grupo, o contacto
físico, o contorno de boias e a flutuabilidade. Devido a esta enorme variedade de
fatores, a natação de águas abertas representa a mais diversa e fisiologicamente
desafiadora disciplina da FINA (Shaw et al., 2014). Face a tudo isto, a FINA criou
um comité de natação de águas abertas, o Open Water Swimming Committee,
que administra a disciplina e colabora de perto com o comité médico para
garantir a segurança dos nadadores.
Podemos afirmar que, tendo em conta as particularidades da disciplina, os
nadadores de águas abertas parecem ter características muito específicas,
nomeadamente uma grande disponibilidade temporal e mental para o treino,
espirito de sacrifício, resiliência e uma personalidade forte, assim como ter

3
objetivos bem definidos a curto e a longo prazo, e ser inteligentes do ponto de
vista tático e estratégico, aspetos fundamentais quer em treino quer em
competição (Borges, 2015). Nadadores de águas abertas têm de ser capazes de
se adaptar e lidar com o inesperado (Munatones, 2008), e devem ter uma boa
capacidade aeróbia associada à capacidade de nadar a velocidades elevadas
durante longos períodos de tempo (VanHeest et al., 2004). Esses nadadores
parecem ser diferentes dos nadadores de natação pura em relação à
antropometria (sendo mais baixos e mais leves) e à fisiologia (VanHeest et al.,
2004; Zamparo et al., 2005).
Em Portugal, a disciplina tem vindo a desenvolver-se de forma consistente,
tentando acompanhar o que se passa a nível internacional com o aumento do
número de campeonatos organizados pela Federação Portuguesa de Natação
(FPN), bem como o número de participantes (Borges, 2015; FPN, 2017). A FPN,
em conjunto com as respetivas federações internacionais, organiza uma das
mais prestigiadas etapas da 10Km Swimming World Cup, em Setúbal, que teve
a sua primeira edição em 2007 e se tem realizado anualmente desde então, e
uma Len Cup, prova com 10 km, que teve em novembro de 2015 a sua estreia,
no Funchal. De referir que a prova de apuramento olímpico, a Marathon
Swimming Olympic Games Qualification Tournament, para os Jogos Olímpicos
Londres’ 2012 e Rio de Janeiro’ 2016, foram ambas realizadas em Portugal, mais
precisamente em Setúbal.
No que respeita às atividades das seleções nacionais, a irregularidade tem sido
uma constante, ficando muito aquém das necessidades inerentes ao alto
rendimento desportivo (Borges, 2015). Pese embora estas adversidades e a
tentativa de possibilitar maior e melhor contacto internacional a nadadores
portugueses por parte da FPN, as prestações internacionais dos nadadores
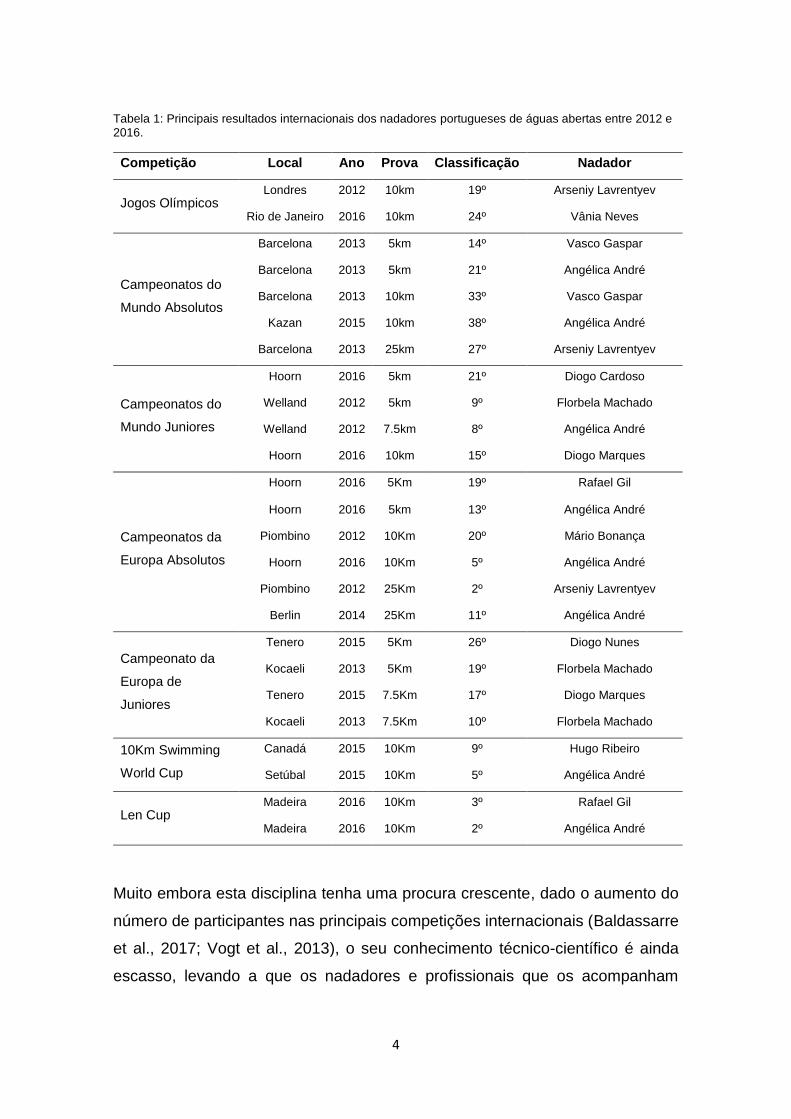
portugueses têm sido de boa qualidade, como podemos constatar na Tabela 1,
onde são apresentadas as melhores classificações obtidas nas principais
competições internacionais no decorrer do último ciclo olímpico (2012-2016),
destacando-se as duas participações olímpicas em Londres’ 2012 e Rio de
Janeiro’ 2016.
4
Tabela 1: Principais resultados internacionais dos nadadores portugueses de águas abertas entre 2012 e 2016.
Competição Local Ano Prova Classificação Nadador
Jogos Olímpicos Londres 2012 10km 19º Arseniy Lavrentyev
Rio de Janeiro 2016 10km 24º Vânia Neves
Campeonatos do
Mundo Absolutos
Barcelona 2013 5km 14º Vasco Gaspar
Barcelona 2013 5km 21º Angélica André
Barcelona 2013 10km 33º Vasco Gaspar
Kazan 2015 10km 38º Angélica André
Barcelona 2013 25km 27º Arseniy Lavrentyev
Campeonatos do
Mundo Juniores
Hoorn 2016 5km 21º Diogo Cardoso
Welland 2012 5km 9º Florbela Machado
Welland 2012 7.5km 8º Angélica André
Hoorn 2016 10km 15º Diogo Marques
Campeonatos da
Europa Absolutos
Hoorn 2016 5Km 19º Rafael Gil
Hoorn 2016 5km 13º Angélica André
Piombino 2012 10Km 20º Mário Bonança
Hoorn 2016 10Km 5º Angélica André
Piombino 2012 25Km 2º Arseniy Lavrentyev
Berlin 2014 25Km 11º Angélica André
Campeonato da
Europa de
Juniores
Tenero 2015 5Km 26º Diogo Nunes
Kocaeli 2013 5Km 19º Florbela Machado
Tenero 2015 7.5Km 17º Diogo Marques
Kocaeli 2013 7.5Km 10º Florbela Machado
10Km Swimming
World Cup
Canadá 2015 10Km 9º Hugo Ribeiro
Setúbal 2015 10Km 5º Angélica André
Len Cup Madeira 2016 10Km 3º Rafael Gil
Madeira 2016 10Km 2º Angélica André
Muito embora esta disciplina tenha uma procura crescente, dado o aumento do
número de participantes nas principais competições internacionais (Baldassarre
et al., 2017; Vogt et al., 2013), o seu conhecimento técnico-científico é ainda
escasso, levando a que os nadadores e profissionais que os acompanham

5
tenham necessidade de olhar para evidências cientificas de modalidades
semelhantes e procurar implementá-las ao ambiente competitivo das águas
abertas.
De facto, a elaboração de trabalhos científicos tem vindo a adquirir elevada
importância no que respeita à perceção do fenómeno desportivo, contribuindo
para a compreensão e otimização dos fatores que influenciam o rendimento
desportivo. A avaliação e controlo de treino tem sido apresentada como uma
tarefa primordial no processo de treino (Vilas-Boas, 1989), sendo definida como
um conjunto de tarefas que permitem a avaliação dos resultados e adequação
dos programas e exercícios de treino, bem como o desenvolvimento dos fatores
de rendimento e objetivação do processo de treino (Fernandes et al., 2009).
Na literatura foram apresentadas, ao longo dos anos, diferentes propostas de
modelos influenciadores do rendimento desportivo em natação pura. Fernandes
(1999) verificou que, embora não exista unanimidade científica relativamente à
classificação, parece existir uma tendência para uma estruturação complexa dos
fatores determinantes do rendimento, sugerindo um modelo que inter-relaciona
aspetos genéticos, biomecânicos, bioenergéticos, psicológicos e contextuais, e
que se constituem determinantes no rendimento desportivo do nadador (Figura
1). Complementarmente, é de sublinhar que os parâmetros determinantes da
performance estão interligados de forma diferente em provas de velocidade (50-
100m), meia-distância (200-400m), longa distância (800-1500m) (Aspenes &
Karlsen, 2012; di Prampero et al., 1974; Maglischo, 2003) e em águas abertas
(Aspenes & Karlsen, 2012).

6
Figura 1: Diagrama síntese dos fatores determinantes do rendimento desportivo do nadador (Fernandes &
Vilas-Boas, 2006).
Não obstante o rendimento desportivo ser um fenómeno multifatorial, assume-
se a possibilidade de o predizer a partir do perfil biofísico do desportista (Barbosa
et al., 2006). As determinantes biofísicas relacionadas à performance dos
nadadores são um dos pontos mais atrativos para a comunidade científica,
sendo consensual que uma abordagem biofísica é um elemento que enaltece a
performance e permite ao nadador atingir níveis mais elevados em competição
(Barbosa et al., 2010; Vilas-Boas, 2010). Entende-se por biofísica o domínio
cientifico onde se exercita o estudo dos fenómenos biológicos na perspetiva da
física (Vilas-Boas, 2000) e tem como objetivos primários o incremento da
economia e/ou eficiência do nadador (Barbosa et al., 2006). A performance de
um nadador pode ser expressa por:
Equação 1 v = E ∙ (ep ∙ D-1)
onde v é a velocidade de nado, Ė é o dispêndio energético, ep expressa a
eficiência mecânica propulsiva e D representa o arrasto hidrodinâmico oposto ao
deslocamento do nadador. Esta equação, tradicionalmente denominada por
“equação geral da performance do nadador”, foi proposta por di Prampero et al.
(1974) e Pendergast et al. (1977), evidenciando que o resultado final do nadador
é diretamente dependente da energia disponível total (Ė), da sua eficiência

7
técnica (ep ∙ D-1) e da análise hidrodinâmica a que se sujeita (técnica e
morfologicamente).
Vários investigadores têm avaliado o Ė dos nadadores através da quantificação
energética das fontes aeróbias e anaeróbias (Toussaint & Beek, 1992; Olbrecht,
2000; Zamparo et al., 2011), nomeadamente através da determinação do
consumo de oxigénio (VO2) como medida do metabolismo aeróbio e da
concentração sanguínea de lactato ([La-]) como indicador do metabolismo
anaeróbio lático (Fernandes, 2013). Em exercícios a intensidades moderadas, a
contribuição alática parece ser negligenciável (Vilas-Boas, 1996). A análise da
cinética do VO2 tem sido considerada uma ferramenta útil quer para o treino quer
para o diagnóstico em natação (Fernandes & Vilas-Boas, 2012; Reis et al.,
2012b), particularmente para melhorar a compreensão da tolerância ao
exercício, do efeito que parâmetros tradicionais de função fisiológica e dos
efeitos que indicadores de eficiência/economia de nado (Bentley et al., 2007) têm
na resposta do VO2 durante o nado (Reis et al., 2012a; Rodríguez et al., 2003;
Sousa et al., 2011).
Também as medições da [La-] se têm tornado uma pratica comum no diagnóstico
da performance e no controlo do treino, como já referido. Esta variável fornece
evidencias quanto ao stress fisiológico num determinado evento (Vescovi et al.,
2011) e é tido como indicador da ativação do uso de hidratos de carbono como
combustível do metabolismo aeróbio (Beneke et al., 2009). O musculo
esquelético também extrai glicose do sangue para manter a ação dos hidratos
de carbono como fonte energética durante o exercício (Sylow et al., 2016). A
intensidade e a duração do exercício são determinantes no consumo de glicose
pelo musculo esquelético (Sylow et al., 2016), logo o exercício constitui um forte
indutor de alterações glicémicas, quer devido ao aumento da captação periférica
de glicose (Hardin et al., 1995), quer devido às respostas hormonais (Robergs &
Roberts, 2000). Apesar disso a investigação cientifica que recai sobre a
concentração sanguínea de glicose (ou glicemia) em desportos competitivos é
ainda muito escassa.

8
O sucesso do nadador é também determinado pela sua capacidade de se
propulsionar enquanto reduz a resistência hidrodinâmica ao seu deslocamento
(Fernandes, 2013). Uma vez que a eficiência propulsiva e o arrasto
hidrodinâmico são de difícil e dispendiosa avaliação, a sua análise habitualmente
recai sobre os parâmetros biomecânicos gerais, frequência gestual (FG) e
distância percorrida por ciclo (DC). A relação entre a FG, a DC e a v, de acordo
com a distância percorrida, tem sido um dos maiores pontos de interesse na
investigação biomecânica (Barbosa et al., 2010; Figueiredo et al., 2013). Outro
indicador de grande utilidade para medir a habilidade técnica do nadador é o
índice de braçada (IB; Costill et al., 1985), assumindo-se que quanto mais
elevado o IB mais adequada mecanicamente será a técnica do nadador (Vilas-
Boas, 2013). Apesar de reconhecida a importância da avaliação destas variáveis
no desempenho desportivo do nadador, a sua investigação é praticamente
inexistente em águas abertas.
Como referido anteriormente, a investigação cientifica em águas abertas é
extremamente escassa, não existindo nenhum estudo que tenha feito uma
análise aprofundada em distâncias oficiais (5, 7.5 e 10 km), sendo o mais
aproximado a análise de intensidade de nado correspondente ao limiar
anaeróbio analisado através de algumas repetições de 30 min de nado contínuo
aquando da realização do teste da máxima fase estável de [La-] (Dekerle et al.,
2005; Pelarigo et al., 2016). Sendo crucial que os treinadores conheçam e
entendam as alterações que podem acontecer em exercícios prolongados, bem
como as consequências dessas alterações, na presente dissertação realizou-se
uma caracterização biofísica, i.e., uma caracterização combinada de fatores
bioenergéticos e biomecânicos, na distância de 5000 m livres, distância mais
curta em provas oficiais de águas abertas, habitualmente realizada na técnica de
crol.
Para o propósito acima descrito efetuamos três estudos: (i) avaliação em
situação competitiva numa prova de 5000 m livres, na fase de qualificação do
campeonato nacional de longa distância, onde foram analisados a [La-], a
glicemia, as variáveis biomecânicas FG, DC e IB, e a performance; (ii) avaliação
num protocolo experimental de 5x1000 m a v constante, da [La-], da glicemia e

9
da FG, DC e IB; (iii) análise da cinética do VO2, ventilação pulmonar (VE), cálculo
do Ė e do C (custo energético), quociente respiratório (R), das variáveis
biomecânicas, [La-] e glicemia em 5 x 1000 m.
Sabendo que em esforços contínuos de 30 min a uma velocidade constante o
VO2, a [La-], o Ė e o C usualmente apresentam valores estáveis durante todo o
esforço (Pelarigo et al., 2016), e que o equilíbrio ou diminuição da [La-] é
frequentemente observada em exercícios prolongados a intensidade constante
(Beneke, 2003; Beneke et al., 2011), colocou-se a hipótese que existe
estabilidade nas variáveis bioenergéticas num esforço de 5000 m crol. É também
esperado que, apesar dos fatores bioenergéticos apresentarem valores
constantes, o mesmo não se observe com os fatores biomecânicos, onde, devido
ao aparecimento da fadiga, ocorra um aumento da FG e uma diminuição da DC,
que consequentemente conduz a uma redução do IB, para que os nadadores
consigam manter uma velocidade constante. Assim, não descartamos a hipótese
de se poder observar, ao longo de um esforço prolongado superior a 1h de
duração, uma diminuição da v de nado (devido ao aparecimento de fadiga
muscular), associada a uma estabilização da FG e diminuição quer da DC quer
do IB.

10
11
2. Material e métodos
2.1 Caracterização da amostra
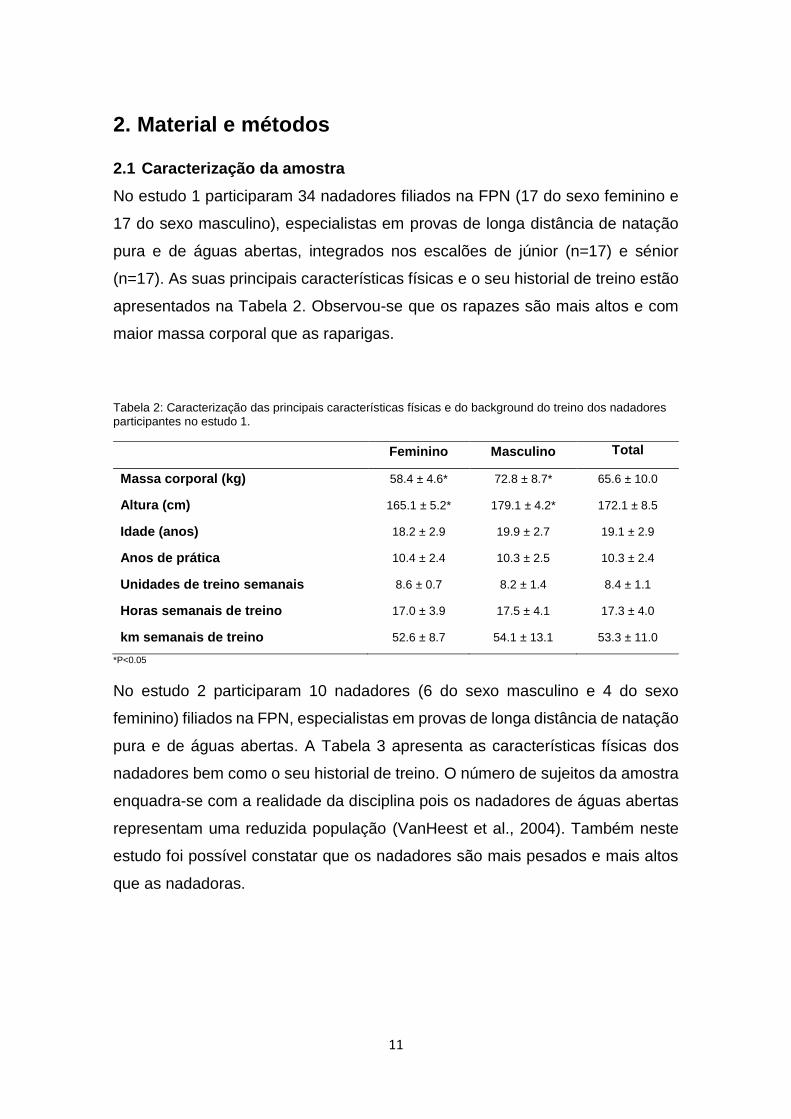
No estudo 1 participaram 34 nadadores filiados na FPN (17 do sexo feminino e
17 do sexo masculino), especialistas em provas de longa distância de natação
pura e de águas abertas, integrados nos escalões de júnior (n=17) e sénior
(n=17). As suas principais características físicas e o seu historial de treino estão
apresentados na Tabela 2. Observou-se que os rapazes são mais altos e com
maior massa corporal que as raparigas.
Tabela 2: Caracterização das principais características físicas e do background do treino dos nadadores participantes no estudo 1.
Feminino Masculino Total
Massa corporal (kg) 58.4 ± 4.6* 72.8 ± 8.7* 65.6 ± 10.0
Altura (cm) 165.1 ± 5.2* 179.1 ± 4.2* 172.1 ± 8.5
Idade (anos) 18.2 ± 2.9 19.9 ± 2.7 19.1 ± 2.9
Anos de prática 10.4 ± 2.4 10.3 ± 2.5 10.3 ± 2.4
Unidades de treino semanais 8.6 ± 0.7 8.2 ± 1.4 8.4 ± 1.1
Horas semanais de treino 17.0 ± 3.9 17.5 ± 4.1 17.3 ± 4.0
km semanais de treino 52.6 ± 8.7 54.1 ± 13.1 53.3 ± 11.0
*P<0.05
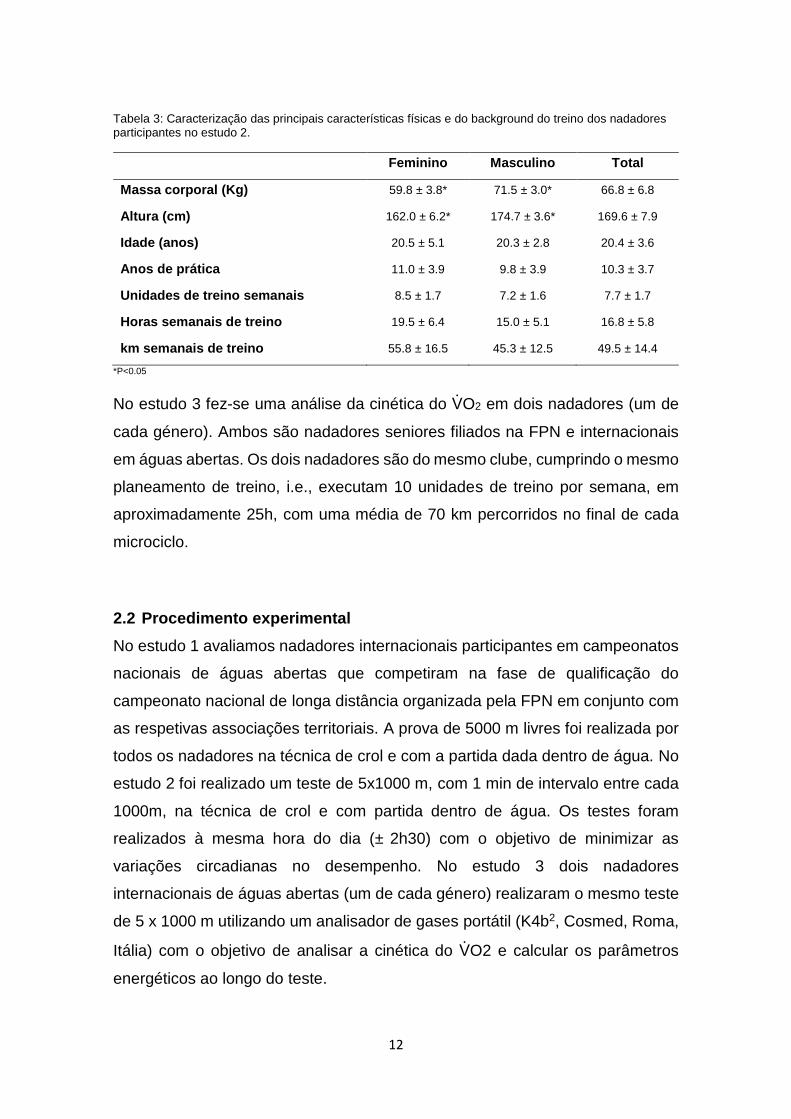
No estudo 2 participaram 10 nadadores (6 do sexo masculino e 4 do sexo
feminino) filiados na FPN, especialistas em provas de longa distância de natação
pura e de águas abertas. A Tabela 3 apresenta as características físicas dos
nadadores bem como o seu historial de treino. O número de sujeitos da amostra
enquadra-se com a realidade da disciplina pois os nadadores de águas abertas
representam uma reduzida população (VanHeest et al., 2004). Também neste
estudo foi possível constatar que os nadadores são mais pesados e mais altos
que as nadadoras.
12
Tabela 3: Caracterização das principais características físicas e do background do treino dos nadadores participantes no estudo 2.
Feminino Masculino Total
Massa corporal (Kg) 59.8 ± 3.8* 71.5 ± 3.0* 66.8 ± 6.8
Altura (cm) 162.0 ± 6.2* 174.7 ± 3.6* 169.6 ± 7.9
Idade (anos) 20.5 ± 5.1 20.3 ± 2.8 20.4 ± 3.6
Anos de prática 11.0 ± 3.9 9.8 ± 3.9 10.3 ± 3.7
Unidades de treino semanais 8.5 ± 1.7 7.2 ± 1.6 7.7 ± 1.7
Horas semanais de treino 19.5 ± 6.4 15.0 ± 5.1 16.8 ± 5.8
km semanais de treino 55.8 ± 16.5 45.3 ± 12.5 49.5 ± 14.4
*P<0.05
No estudo 3 fez-se uma análise da cinética do VO2 em dois nadadores (um de
cada género). Ambos são nadadores seniores filiados na FPN e internacionais
em águas abertas. Os dois nadadores são do mesmo clube, cumprindo o mesmo
planeamento de treino, i.e., executam 10 unidades de treino por semana, em
aproximadamente 25h, com uma média de 70 km percorridos no final de cada
microciclo.
2.2 Procedimento experimental
No estudo 1 avaliamos nadadores internacionais participantes em campeonatos
nacionais de águas abertas que competiram na fase de qualificação do
campeonato nacional de longa distância organizada pela FPN em conjunto com
as respetivas associações territoriais. A prova de 5000 m livres foi realizada por
todos os nadadores na técnica de crol e com a partida dada dentro de água. No
estudo 2 foi realizado um teste de 5x1000 m, com 1 min de intervalo entre cada
1000m, na técnica de crol e com partida dentro de água. Os testes foram
realizados à mesma hora do dia (± 2h30) com o objetivo de minimizar as
variações circadianas no desempenho. No estudo 3 dois nadadores
internacionais de águas abertas (um de cada género) realizaram o mesmo teste
de 5 x 1000 m utilizando um analisador de gases portátil (K4b2, Cosmed, Roma,
Itália) com o objetivo de analisar a cinética do VO2 e calcular os parâmetros
energéticos ao longo do teste.

13
Nos estudos 1 e 2 as variáveis bioenergéticas avaliadas foram a [La-] e a
glicemia. Já no estudo 3, para além dessas variáveis, avaliou-se a cinética do
VO2, a frequência cardíaca (FC), o Ė e o C. Para determinar a [La-] e a glicemia
foi retirada uma amostra de sangue capilar da ponta do dedo do nadador e
quantificada num analisador portátil de lactato (Lactate Pro, Arkay, Inc., Quioto,
Japão) e num analisador portátil de glicose sanguínea (One Touch Select Plus)
(Figura 2). No estudo 1 a recolha foi feita em repouso e nos 1º e 3º min após a
conclusão da prova para se obter o valor mais elevado das variáveis. Já nos
estudos 2 e 3 a recolha foi feita em repouso, durante os intervalos do teste de 5
x 1000 m e ao 3º e 5º min de recuperação. Não foram feitas quaisquer restrições
quanto ao pequeno almoço dos sujeitos ou suplementação de hidratos de
carbono durante o teste. A FC foi controlada continuamente, em intervalos de 5
s, durante todo o exercício, através de um cardiofrequencimentro (Polar NS400,
Polar Eletro Oy, Kempele, Finlândia), fornecendo um indicador mais exato do
stress a que o nadador está sujeito (Fernandes, 2013).
Figura 2: Analisadores portáteis de lactato (Lactate Pro, Arkay, Inc., Quioto, Japão) e de glicemia (One
Touch Select Plus) utilizados nas avaliações.
Os valores das trocas gasosas, particularmente do VO2, foram gravados
respiração-a-respiração através de um analisador de gás portátil (K4b2, Cosmed,
Roma, Itália), o qual se conectou ao nadador por um snorkel e uma válvula

14
respiratória de baixa resistência hidrodinâmica (Aquatrainer, Cosmed, Roma,
Itália, Figura 3, Ribeiro et al., 2016). Foram considerados apenas os valores de
VO2 compreendidos entre a média ± 4 DP, a cada 10 s e com três suavizações
(de Jesus et al., 2014). Para os parâmetros ventilatórios estudados, o valor de
cada patamar foi obtido pela média do último minuto do tempo de nado. A
cinética do VO2 foi modelada pela seguinte função exponencial:
Equação 2 VO2(t) = A0 + Ap ∙ (1 - e(t-TDp) τp⁄ )
onde VO2(t) representa o VO2 por unidade de tempo (ml·min-1), A0 é a amplitude
basal de VO2 (ml·min-1), Ap é a amplitude do VO2 (ml·min-1), TD é o tempo de
atraso do VO2 (s), e 𝜏 é a constante de tempo (s). Foi utilizada esta função uma
vez que a intensidades moderadas de exercício são descritas por modelações
monoexponenciais (Ozyener et al., 2001).
Figura 3: Avaliação dos parâmetros ventilatórios com o analisador portátil de gases (K4b2, Cosmed,
Roma, Itália)
O Ė é assumido como sendo o somatório das componentes energéticas aeróbia
(Aer) e anaeróbia lática (Anala). A contribuição Aer foi calculada através da
multiplicação do integral no tempo do VO2 de repouso (VO2basal) pela relação de

15
tempo nos intervalos de tempo apropriados (mLO2) e, em seguida, expressa em
kJ assumindo um equivalente de energia de 20.9 kJ·LO2-1 (Figueiredo et al.,
2011; Sousa et al., 2014; Zamparo et al., 2011). A contribuição Anala foi estimada
através da energia proveniente da produção de ácido lático (equação 3):
Equação 3 Anala = b ∙ [La-]basal ∙ M
onde [La-]basal é o lactato de repouso, b é o equivalente energético para a
acumulação de lactato no sangue capilar (2.7 ml O2 mM-1·kg-1) (di Prampero et
al., 1978; Thevelein et al., 1984) e M (kg) é a massa corporal do nadador. Neste
caso, a contribuição do sistema anaeróbio alático foi desconsiderada (Vilas-
Boas, 1996). As variáveis foram expressas em kJ assumindo um equivalente de
energia de 20.9 kJ·lO2-1 (Zamparo et al., 2011).
O C representa a energia despendida para se percorrer uma determinada
distância (di Prampero, 1986; Pendergast et al., 2003; Zamparo et al., 2001)
sendo considerado o declive da regressão linear obtida através da relação de Ė
com a v correspondente a cada repetição do teste (Fernandes et al., 2006).
Equação 4 C = E ∙ v-1
Nos três estudos, as variáveis biomecânicas sujeitas a avaliação foram a v, a
FG, a DC e o IB. A análise foi feita com recurso a imagens capturadas por duas
câmaras de vídeo (HDR-CX160E, Sony, Quioto, Japão, Figura 4) e a
cronómetros com cronofrequencímetros. As câmaras foram colocadas na lateral
da piscina, de modo a permitir a visualização do eixo longitudinal do nadador e
um mínimo de três ciclos de membros superiores (MS) em cada percurso de 50
m.

16
Figura 4: Filmagem do teste de 5 x 1000 m com a camara HDR-CX160E, Sony, Quioto, Japão
A FG foi calculada pelo quociente entre o número de ciclos realizados pelos
membros superiores numa determinada distância pelo tempo gasto no percurso
(Maglischo, 1999) sendo utilizada a seguinte equação:
Equação 5 FG = 3 ∙ t3-1
onde t3 é o tempo gasto para completar 3 ciclos de MS. Neste caso particular, a
FG foi determinada recorrendo-se a cronofrequencímetro de base 3 (3x100M
Stopwatch, Finis, California, Estados Unidos, Figura 5), que permite o seu calculo
automaticamente através da análise de três ciclos. A DC é considerada como
um dos fatores com maior preponderância para se alcançar o sucesso em
competição (Toussaint et al., 2006) e foi calculada através da razão entre v e FG
(Craig & Pendergast, 1979):
Equação 6 DC = v ∙ FG-1
O IB foi definido pelo produto da DC com a v (Costill et al., 1985).
Os tempos finais e parciais de cada nadador foram obtidos por cronometragem
manual (3x100M Stopwatch, Finis, Califórnia, Estados Unidos) e a v foi calcula

17
através do quociente entre a distância percorrida e o tempo gasto a percorrer
essa distância.
Figura 5: Cronofrequencimetro Finis (Califórnia, Estados Unidos) e utilizado para controlo da FG dos nadadores
2.3 Análise estatística
A análise estatística dos dados foi efetuada na versão 24.0 do programa
Statistical Package for the Social Sciences (SPSS) e na versão 7.0 do GraphPad
Prism. Na análise exploratória de dados foram calculados parâmetros de
tendência central (média) e dispersão (desvio padrão) para todas as variáveis.
Foram verificados os pressupostos de normalidade através do teste de Shapiro-
Wilk. Posteriormente realizou-se uma análise inferencial onde se recorreu ao
teste ANOVA para medidas repetidas, para comparar as variáveis nas repetições
de 1000 m (excluindo o efeito do género, utilizando este como covariável). O
teste Post Hoc de Bonferroni foi utilizado para identificar diferenças de pares
sempre que fosse obtido um valor de F significativo. Para a comparação entre
géneros recorremos ao Teste-T para medidas independentes. Coeficientes de
correlação parcial (0.40 pobre, 0.40 - 0.75 bom e 0.75 excelente; Fleiss, 1986)
foram utilizados para quantificar o grau de associação entre as variáveis
estudadas e a performance dos nadadores. Foi estabelecido um nível de

18
significância de α = 5% dado ser o valor mais convencionado para pesquisas na
área das ciências humanas. Utilizamos o procedimento estatístico bootstrapping
com 1000 amostras para calcular os parâmetros do modelo monoexponencial,
onde a média, o desvio padrão e o coeficiente de variação foram calculados para
estimar cada parâmetro.
19
3. Resultados
3.1 Estudo 1
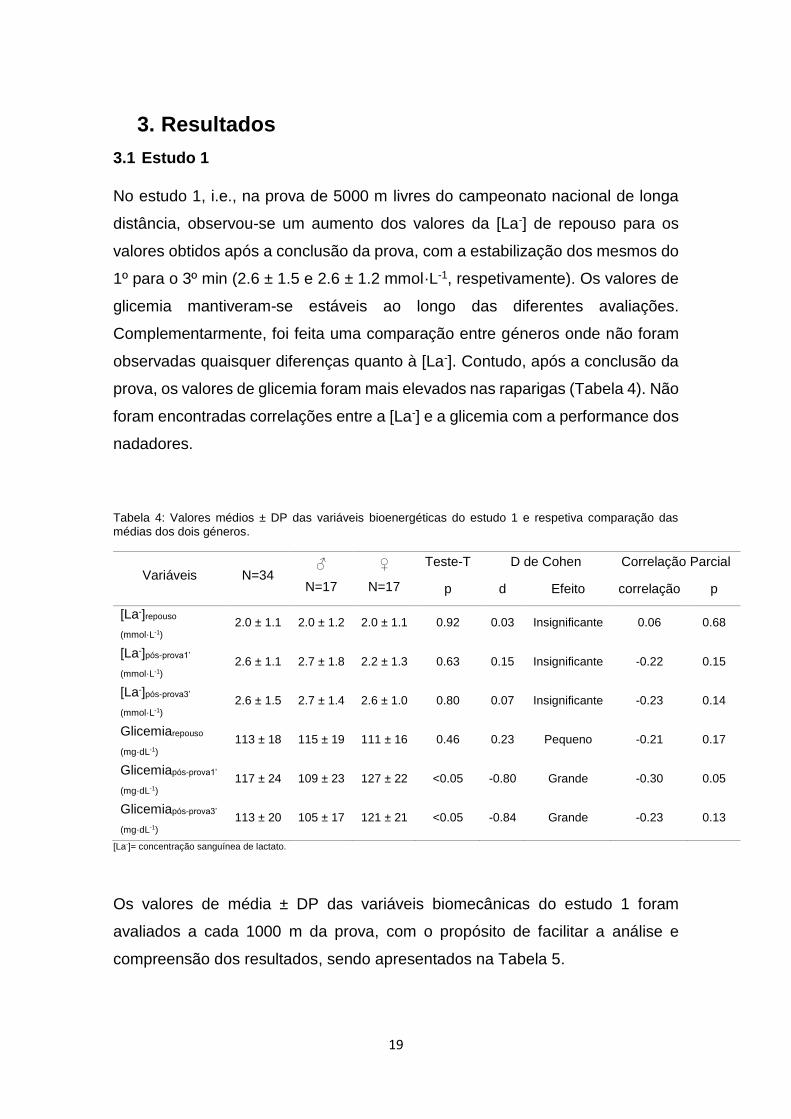
No estudo 1, i.e., na prova de 5000 m livres do campeonato nacional de longa
distância, observou-se um aumento dos valores da [La-] de repouso para os
valores obtidos após a conclusão da prova, com a estabilização dos mesmos do
1º para o 3º min (2.6 ± 1.5 e 2.6 ± 1.2 mmol·L-1, respetivamente). Os valores de
glicemia mantiveram-se estáveis ao longo das diferentes avaliações.
Complementarmente, foi feita uma comparação entre géneros onde não foram
observadas quaisquer diferenças quanto à [La-]. Contudo, após a conclusão da
prova, os valores de glicemia foram mais elevados nas raparigas (Tabela 4). Não
foram encontradas correlações entre a [La-] e a glicemia com a performance dos
nadadores.
Tabela 4: Valores médios ± DP das variáveis bioenergéticas do estudo 1 e respetiva comparação das médias dos dois géneros.
Variáveis N=34 ♂
N=17
♀
N=17
Teste-T D de Cohen Correlação Parcial
p d Efeito correlação p
[La-]repouso
(mmol·L-1)
2.0 ± 1.1 2.0 ± 1.2 2.0 ± 1.1 0.92 0.03 Insignificante 0.06 0.68
[La-]pós-prova1’
(mmol·L-1) 2.6 ± 1.1 2.7 ± 1.8 2.2 ± 1.3 0.63 0.15 Insignificante -0.22 0.15
[La-]pós-prova3’
(mmol·L-1) 2.6 ± 1.5 2.7 ± 1.4 2.6 ± 1.0 0.80 0.07 Insignificante -0.23 0.14
Glicemiarepouso
(mg·dL-1) 113 ± 18 115 ± 19 111 ± 16 0.46 0.23 Pequeno -0.21 0.17
Glicemiapós-prova1’
(mg·dL-1) 117 ± 24 109 ± 23 127 ± 22 <0.05 -0.80 Grande -0.30 0.05
Glicemiapós-prova3’
(mg·dL-1) 113 ± 20 105 ± 17 121 ± 21 <0.05 -0.84 Grande -0.23 0.13
[La-]= concentração sanguínea de lactato.
Os valores de média ± DP das variáveis biomecânicas do estudo 1 foram
avaliados a cada 1000 m da prova, com o propósito de facilitar a análise e
compreensão dos resultados, sendo apresentados na Tabela 5.
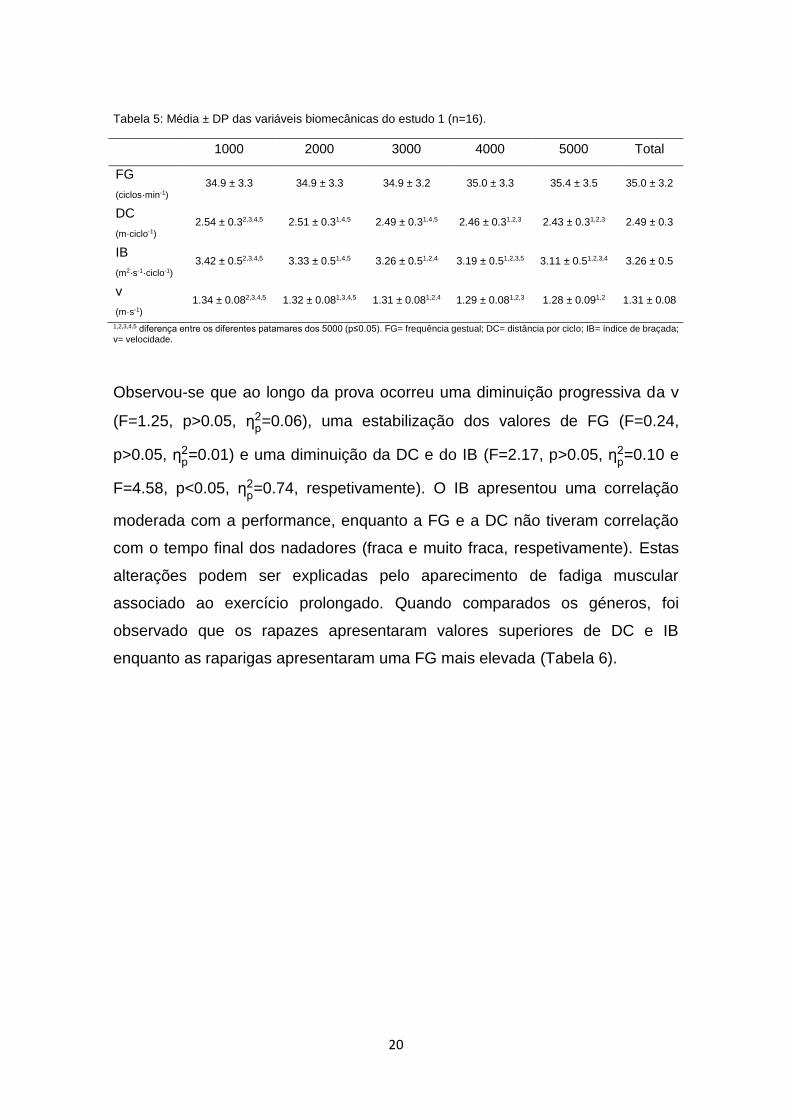
20
Tabela 5: Média ± DP das variáveis biomecânicas do estudo 1 (n=16).
1000 2000 3000 4000 5000 Total
FG
(ciclos·min-1)
34.9 ± 3.3 34.9 ± 3.3 34.9 ± 3.2 35.0 ± 3.3 35.4 ± 3.5 35.0 ± 3.2
DC
(m·ciclo-1)
2.54 ± 0.32,3,4,5 2.51 ± 0.31,4,5 2.49 ± 0.31,4,5 2.46 ± 0.31,2,3 2.43 ± 0.31,2,3 2.49 ± 0.3
IB
(m2·s-1·ciclo-1)
3.42 ± 0.52,3,4,5 3.33 ± 0.51,4,5 3.26 ± 0.51,2,4 3.19 ± 0.51,2,3,5 3.11 ± 0.51,2,3,4 3.26 ± 0.5
v
(m·s-1)
1.34 ± 0.082,3,4,5 1.32 ± 0.081,3,4,5 1.31 ± 0.081,2,4 1.29 ± 0.081,2,3 1.28 ± 0.091,2 1.31 ± 0.08
1,2,3,4,5 diferença entre os diferentes patamares dos 5000 (p≤0.05). FG= frequência gestual; DC= distância por ciclo; IB= índice de braçada; v= velocidade.
Observou-se que ao longo da prova ocorreu uma diminuição progressiva da v
(F=1.25, p>0.05, ƞp2=0.06), uma estabilização dos valores de FG (F=0.24,
p>0.05, ƞp2=0.01) e uma diminuição da DC e do IB (F=2.17, p>0.05, ƞ
p2=0.10 e
F=4.58, p<0.05, ƞp2=0.74, respetivamente). O IB apresentou uma correlação
moderada com a performance, enquanto a FG e a DC não tiveram correlação
com o tempo final dos nadadores (fraca e muito fraca, respetivamente). Estas
alterações podem ser explicadas pelo aparecimento de fadiga muscular
associado ao exercício prolongado. Quando comparados os géneros, foi
observado que os rapazes apresentaram valores superiores de DC e IB
enquanto as raparigas apresentaram uma FG mais elevada (Tabela 6).
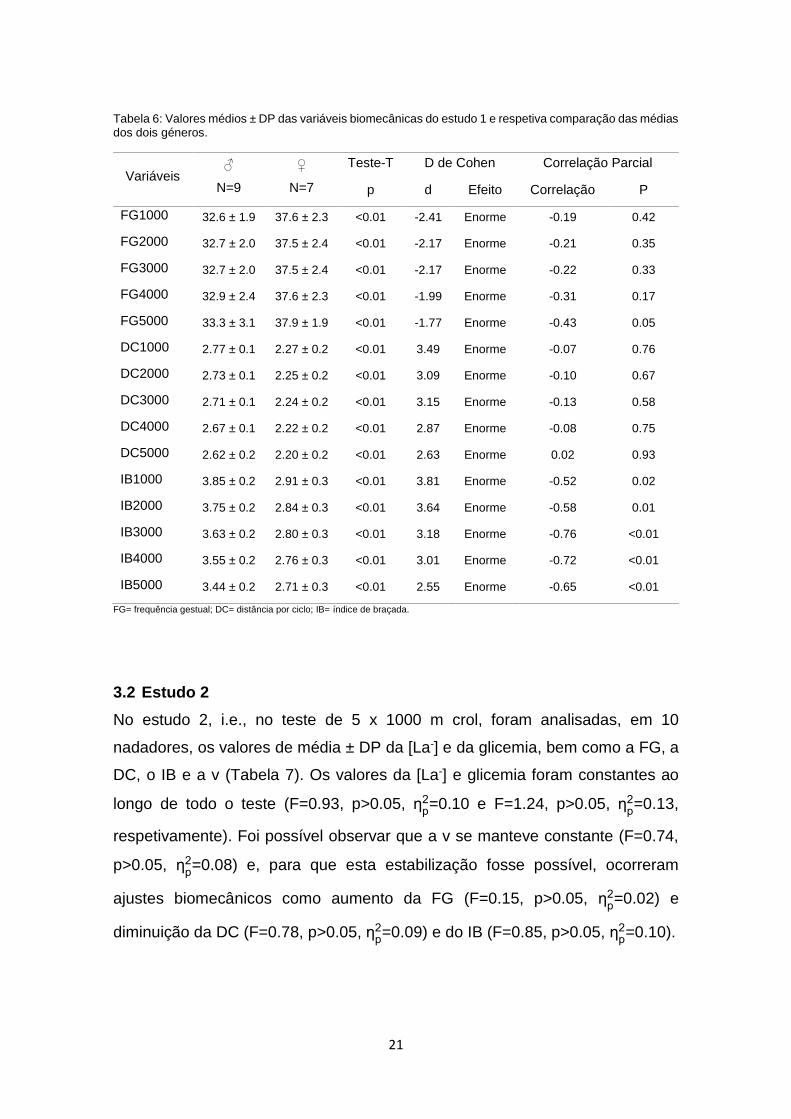
21
Tabela 6: Valores médios ± DP das variáveis biomecânicas do estudo 1 e respetiva comparação das médias dos dois géneros.
Variáveis ♂
N=9
♀
N=7
Teste-T D de Cohen Correlação Parcial
p d Efeito Correlação P
FG1000 32.6 ± 1.9 37.6 ± 2.3 <0.01 -2.41 Enorme -0.19 0.42
FG2000 32.7 ± 2.0 37.5 ± 2.4 <0.01 -2.17 Enorme -0.21 0.35
FG3000 32.7 ± 2.0 37.5 ± 2.4 <0.01 -2.17 Enorme -0.22 0.33
FG4000 32.9 ± 2.4 37.6 ± 2.3 <0.01 -1.99 Enorme -0.31 0.17
FG5000 33.3 ± 3.1 37.9 ± 1.9 <0.01 -1.77 Enorme -0.43 0.05
DC1000 2.77 ± 0.1 2.27 ± 0.2 <0.01 3.49 Enorme -0.07 0.76
DC2000 2.73 ± 0.1 2.25 ± 0.2 <0.01 3.09 Enorme -0.10 0.67
DC3000 2.71 ± 0.1 2.24 ± 0.2 <0.01 3.15 Enorme -0.13 0.58
DC4000 2.67 ± 0.1 2.22 ± 0.2 <0.01 2.87 Enorme -0.08 0.75
DC5000 2.62 ± 0.2 2.20 ± 0.2 <0.01 2.63 Enorme 0.02 0.93
IB1000 3.85 ± 0.2 2.91 ± 0.3 <0.01 3.81 Enorme -0.52 0.02
IB2000 3.75 ± 0.2 2.84 ± 0.3 <0.01 3.64 Enorme -0.58 0.01
IB3000 3.63 ± 0.2 2.80 ± 0.3 <0.01 3.18 Enorme -0.76 <0.01
IB4000 3.55 ± 0.2 2.76 ± 0.3 <0.01 3.01 Enorme -0.72 <0.01
IB5000 3.44 ± 0.2 2.71 ± 0.3 <0.01 2.55 Enorme -0.65 <0.01
FG= frequência gestual; DC= distância por ciclo; IB= índice de braçada.
3.2 Estudo 2
No estudo 2, i.e., no teste de 5 x 1000 m crol, foram analisadas, em 10
nadadores, os valores de média ± DP da [La-] e da glicemia, bem como a FG, a
DC, o IB e a v (Tabela 7). Os valores da [La-] e glicemia foram constantes ao
longo de todo o teste (F=0.93, p>0.05, ƞp2=0.10 e F=1.24, p>0.05, ƞ
p2=0.13,
respetivamente). Foi possível observar que a v se manteve constante (F=0.74,
p>0.05, ƞp2=0.08) e, para que esta estabilização fosse possível, ocorreram
ajustes biomecânicos como aumento da FG (F=0.15, p>0.05, ƞp2=0.02) e
diminuição da DC (F=0.78, p>0.05, ƞp2=0.09) e do IB (F=0.85, p>0.05, ƞ
p2=0.10).
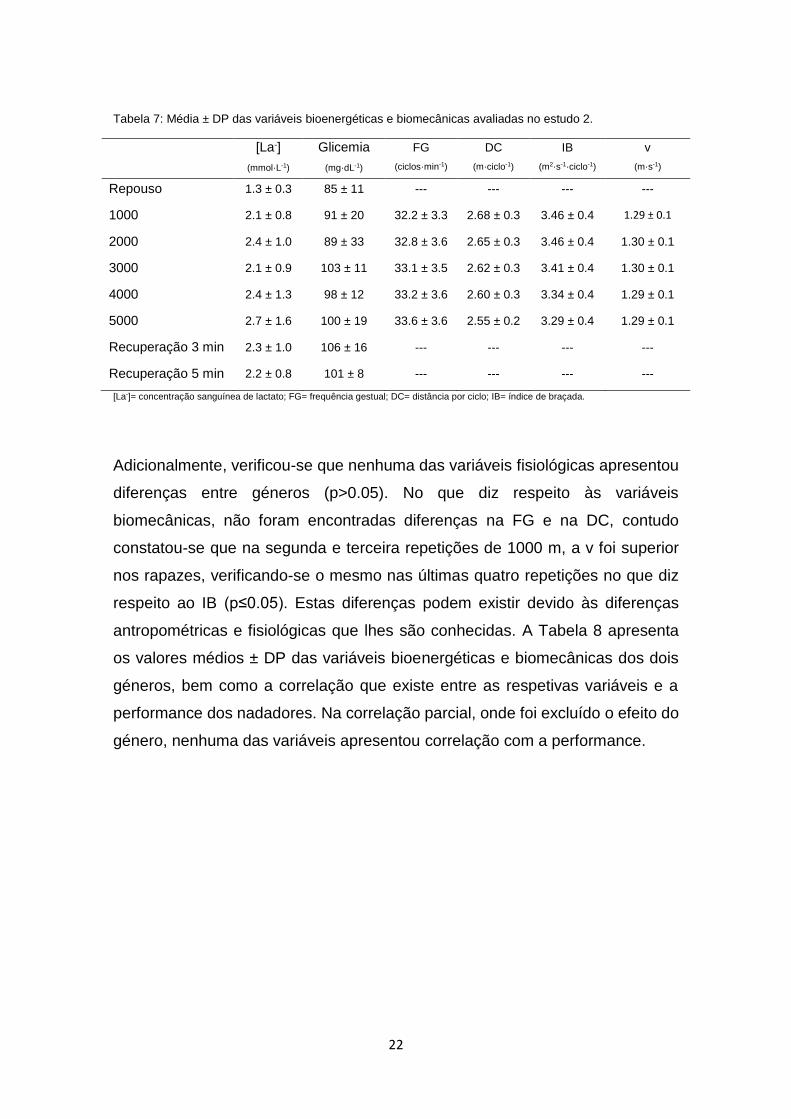
22
Tabela 7: Média ± DP das variáveis bioenergéticas e biomecânicas avaliadas no estudo 2.
[La-]
(mmol·L-1)
Glicemia
(mg·dL-1)
FG
(ciclos·min-1)
DC
(m·ciclo-1)
IB
(m2·s-1·ciclo-1)
v
(m·s-1)
Repouso 1.3 ± 0.3 85 ± 11 --- --- --- ---
1000 2.1 ± 0.8 91 ± 20 32.2 ± 3.3 2.68 ± 0.3 3.46 ± 0.4 1.29 ± 0.1
2000 2.4 ± 1.0 89 ± 33 32.8 ± 3.6 2.65 ± 0.3 3.46 ± 0.4 1.30 ± 0.1
3000 2.1 ± 0.9 103 ± 11 33.1 ± 3.5 2.62 ± 0.3 3.41 ± 0.4 1.30 ± 0.1
4000 2.4 ± 1.3 98 ± 12 33.2 ± 3.6 2.60 ± 0.3 3.34 ± 0.4 1.29 ± 0.1
5000 2.7 ± 1.6 100 ± 19 33.6 ± 3.6 2.55 ± 0.2 3.29 ± 0.4 1.29 ± 0.1
Recuperação 3 min 2.3 ± 1.0 106 ± 16 --- --- --- ---
Recuperação 5 min 2.2 ± 0.8 101 ± 8 --- --- --- ---
[La-]= concentração sanguínea de lactato; FG= frequência gestual; DC= distância por ciclo; IB= índice de braçada.
Adicionalmente, verificou-se que nenhuma das variáveis fisiológicas apresentou
diferenças entre géneros (p>0.05). No que diz respeito às variáveis
biomecânicas, não foram encontradas diferenças na FG e na DC, contudo
constatou-se que na segunda e terceira repetições de 1000 m, a v foi superior
nos rapazes, verificando-se o mesmo nas últimas quatro repetições no que diz
respeito ao IB (p≤0.05). Estas diferenças podem existir devido às diferenças
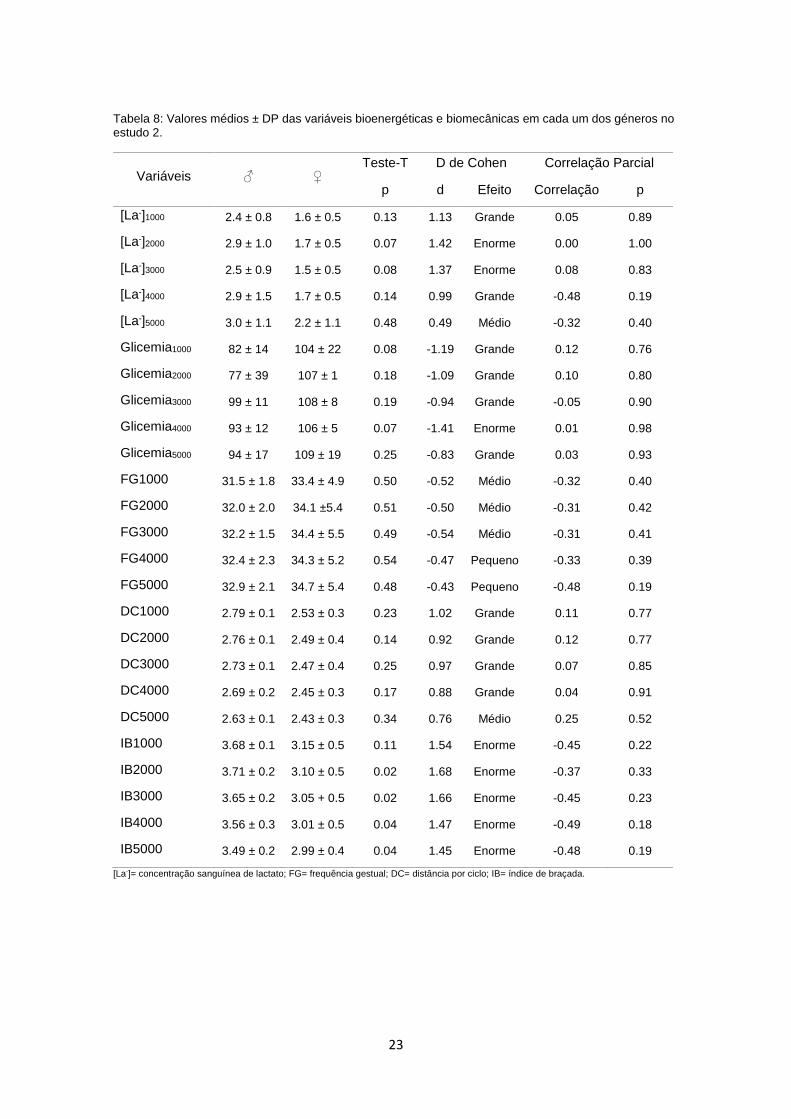
antropométricas e fisiológicas que lhes são conhecidas. A Tabela 8 apresenta
os valores médios ± DP das variáveis bioenergéticas e biomecânicas dos dois
géneros, bem como a correlação que existe entre as respetivas variáveis e a
performance dos nadadores. Na correlação parcial, onde foi excluído o efeito do
género, nenhuma das variáveis apresentou correlação com a performance.
23
Tabela 8: Valores médios ± DP das variáveis bioenergéticas e biomecânicas em cada um dos géneros no estudo 2.
Variáveis ♂ ♀ Teste-T D de Cohen Correlação Parcial
p d Efeito Correlação p
[La-]1000 2.4 ± 0.8 1.6 ± 0.5 0.13 1.13 Grande 0.05 0.89
[La-]2000 2.9 ± 1.0 1.7 ± 0.5 0.07 1.42 Enorme 0.00 1.00
[La-]3000 2.5 ± 0.9 1.5 ± 0.5 0.08 1.37 Enorme 0.08 0.83
[La-]4000 2.9 ± 1.5 1.7 ± 0.5 0.14 0.99 Grande -0.48 0.19
[La-]5000 3.0 ± 1.1 2.2 ± 1.1 0.48 0.49 Médio -0.32 0.40
Glicemia1000 82 ± 14 104 ± 22 0.08 -1.19 Grande 0.12 0.76
Glicemia2000 77 ± 39 107 ± 1 0.18 -1.09 Grande 0.10 0.80
Glicemia3000 99 ± 11 108 ± 8 0.19 -0.94 Grande -0.05 0.90
Glicemia4000 93 ± 12 106 ± 5 0.07 -1.41 Enorme 0.01 0.98
Glicemia5000 94 ± 17 109 ± 19 0.25 -0.83 Grande 0.03 0.93
FG1000 31.5 ± 1.8 33.4 ± 4.9 0.50 -0.52 Médio -0.32 0.40
FG2000 32.0 ± 2.0 34.1 ±5.4 0.51 -0.50 Médio -0.31 0.42
FG3000 32.2 ± 1.5 34.4 ± 5.5 0.49 -0.54 Médio -0.31 0.41
FG4000 32.4 ± 2.3 34.3 ± 5.2 0.54 -0.47 Pequeno -0.33 0.39
FG5000 32.9 ± 2.1 34.7 ± 5.4 0.48 -0.43 Pequeno -0.48 0.19
DC1000 2.79 ± 0.1 2.53 ± 0.3 0.23 1.02 Grande 0.11 0.77
DC2000 2.76 ± 0.1 2.49 ± 0.4 0.14 0.92 Grande 0.12 0.77
DC3000 2.73 ± 0.1 2.47 ± 0.4 0.25 0.97 Grande 0.07 0.85
DC4000 2.69 ± 0.2 2.45 ± 0.3 0.17 0.88 Grande 0.04 0.91
DC5000 2.63 ± 0.1 2.43 ± 0.3 0.34 0.76 Médio 0.25 0.52
IB1000 3.68 ± 0.1 3.15 ± 0.5 0.11 1.54 Enorme -0.45 0.22
IB2000 3.71 ± 0.2 3.10 ± 0.5 0.02 1.68 Enorme -0.37 0.33
IB3000 3.65 ± 0.2 3.05 + 0.5 0.02 1.66 Enorme -0.45 0.23
IB4000 3.56 ± 0.3 3.01 ± 0.5 0.04 1.47 Enorme -0.49 0.18
IB5000 3.49 ± 0.2 2.99 ± 0.4 0.04 1.45 Enorme -0.48 0.19
[La-]= concentração sanguínea de lactato; FG= frequência gestual; DC= distância por ciclo; IB= índice de braçada.

24
3.3 Estudo 3
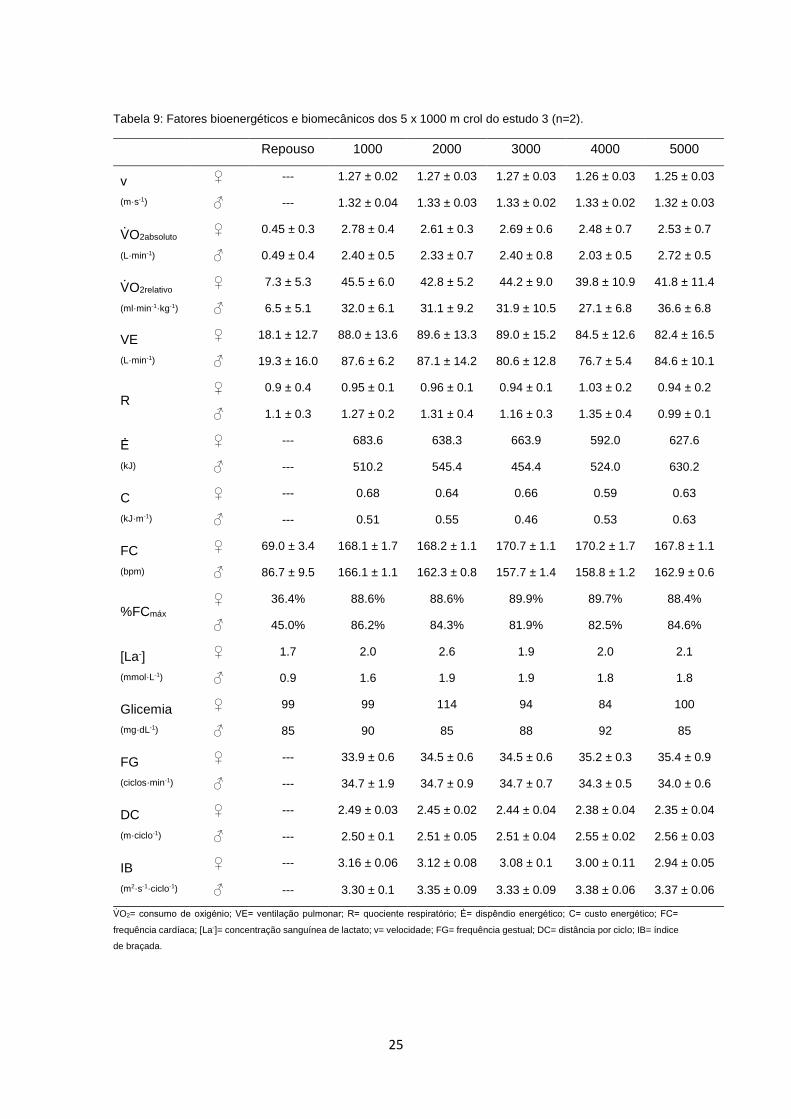
Como já referido anteriormente, neste estudo avaliou-se a cinética do VO2,
juntamente com as variáveis ventilatórias (VE e R), a [La-], a glicemia e as
variáveis biomecânicas (Tabela 9). Foram também calculados o Ė e o C, bem
como a contribuição energética em cada 1000 m do teste. Os valores basais das
variáveis bioenergéticas, i.e., os valores encontrados em repouso, foram
semelhantes entre os dois casos estudados.
As variáveis ventilatórias e bioenergéticas não sofreram alterações no decorrer
dos 5 x 1000 m crol. O VO2 (absoluto e relativo) não sofreu mudanças em função
do tempo (F=1.37, p>0.05, ƞp2=0.58) e o mesmo sucedeu com os valores de VE
(F=1.90, p>0.05, ƞp2=0.66), R (F=1.96, p>0.05, ƞ
p2=0.66) e FC (F=0.24, p>0.05,
ƞp2=0.19). Constatou-se que, tanto o C, como o Ė, apresentaram um
comportamento idêntico, i.e., mantiveram-se contantes ao longo de todo o teste
(F=0.52, p>0.05, ƞp2=0.34 e F=0.49, p>0.05, ƞ
p2=0.33, respetivamente). Quanto
às variáveis biomecânicas, o comportamento foi semelhante, com todas a
variáveis a apresentarem valores constantes ao longo das 5 x 1000 m (F=2.14,
p>0.05, ƞp2=0.68 para a v, F=0.15, p>0.05, ƞ
p2=0.13 para a FG, F=0.13, p>0.05,
ƞp2=0.12 para a DC e F=0.31, p>0.05, ƞ
p2=0.23 para o IB).
25
Tabela 9: Fatores bioenergéticos e biomecânicos dos 5 x 1000 m crol do estudo 3 (n=2).
Repouso 1000 2000 3000 4000 5000
v
(m·s-1)
♀ --- 1.27 ± 0.02 1.27 ± 0.03 1.27 ± 0.03 1.26 ± 0.03 1.25 ± 0.03
♂ --- 1.32 ± 0.04 1.33 ± 0.03 1.33 ± 0.02 1.33 ± 0.02 1.32 ± 0.03
VO2absoluto
(L·min-1)
♀ 0.45 ± 0.3 2.78 ± 0.4 2.61 ± 0.3 2.69 ± 0.6 2.48 ± 0.7 2.53 ± 0.7
♂ 0.49 ± 0.4 2.40 ± 0.5 2.33 ± 0.7 2.40 ± 0.8 2.03 ± 0.5 2.72 ± 0.5
VO2relativo
(ml·min-1·kg-1)
♀ 7.3 ± 5.3 45.5 ± 6.0 42.8 ± 5.2 44.2 ± 9.0 39.8 ± 10.9 41.8 ± 11.4
♂ 6.5 ± 5.1 32.0 ± 6.1 31.1 ± 9.2 31.9 ± 10.5 27.1 ± 6.8 36.6 ± 6.8
VE
(L·min-1)
♀ 18.1 ± 12.7 88.0 ± 13.6 89.6 ± 13.3 89.0 ± 15.2 84.5 ± 12.6 82.4 ± 16.5
♂ 19.3 ± 16.0 87.6 ± 6.2 87.1 ± 14.2 80.6 ± 12.8 76.7 ± 5.4 84.6 ± 10.1
R ♀ 0.9 ± 0.4 0.95 ± 0.1 0.96 ± 0.1 0.94 ± 0.1 1.03 ± 0.2 0.94 ± 0.2
♂ 1.1 ± 0.3 1.27 ± 0.2 1.31 ± 0.4 1.16 ± 0.3 1.35 ± 0.4 0.99 ± 0.1
Ė
(kJ)
♀ --- 683.6 638.3 663.9 592.0 627.6
♂ --- 510.2 545.4 454.4 524.0 630.2
C
(kJ·m-1)
♀ --- 0.68 0.64 0.66 0.59 0.63
♂ --- 0.51 0.55 0.46 0.53 0.63
FC
(bpm)
♀ 69.0 ± 3.4 168.1 ± 1.7 168.2 ± 1.1 170.7 ± 1.1 170.2 ± 1.7 167.8 ± 1.1
♂ 86.7 ± 9.5 166.1 ± 1.1 162.3 ± 0.8 157.7 ± 1.4 158.8 ± 1.2 162.9 ± 0.6
%FCmáx
♀ 36.4% 88.6% 88.6% 89.9% 89.7% 88.4%
♂ 45.0% 86.2% 84.3% 81.9% 82.5% 84.6%
[La-]
(mmol·L-1)
♀ 1.7 2.0 2.6 1.9 2.0 2.1
♂ 0.9 1.6 1.9 1.9 1.8 1.8
Glicemia
(mg·dL-1)
♀ 99 99 114 94 84 100
♂ 85 90 85 88 92 85
FG
(ciclos·min-1)
♀ --- 33.9 ± 0.6 34.5 ± 0.6 34.5 ± 0.6 35.2 ± 0.3 35.4 ± 0.9
♂ --- 34.7 ± 1.9 34.7 ± 0.9 34.7 ± 0.7 34.3 ± 0.5 34.0 ± 0.6
DC
(m·ciclo-1)
♀ --- 2.49 ± 0.03 2.45 ± 0.02 2.44 ± 0.04 2.38 ± 0.04 2.35 ± 0.04
♂ --- 2.50 ± 0.1 2.51 ± 0.05 2.51 ± 0.04 2.55 ± 0.02 2.56 ± 0.03
IB
(m2·s-1·ciclo-1)
♀ --- 3.16 ± 0.06 3.12 ± 0.08 3.08 ± 0.1 3.00 ± 0.11 2.94 ± 0.05
♂ --- 3.30 ± 0.1 3.35 ± 0.09 3.33 ± 0.09 3.38 ± 0.06 3.37 ± 0.06
VO2= consumo de oxigénio; VE= ventilação pulmonar; R= quociente respiratório; Ė= dispêndio energético; C= custo energético; FC=
frequência cardíaca; [La-]= concentração sanguínea de lactato; v= velocidade; FG= frequência gestual; DC= distância por ciclo; IB= índice
de braçada.
26
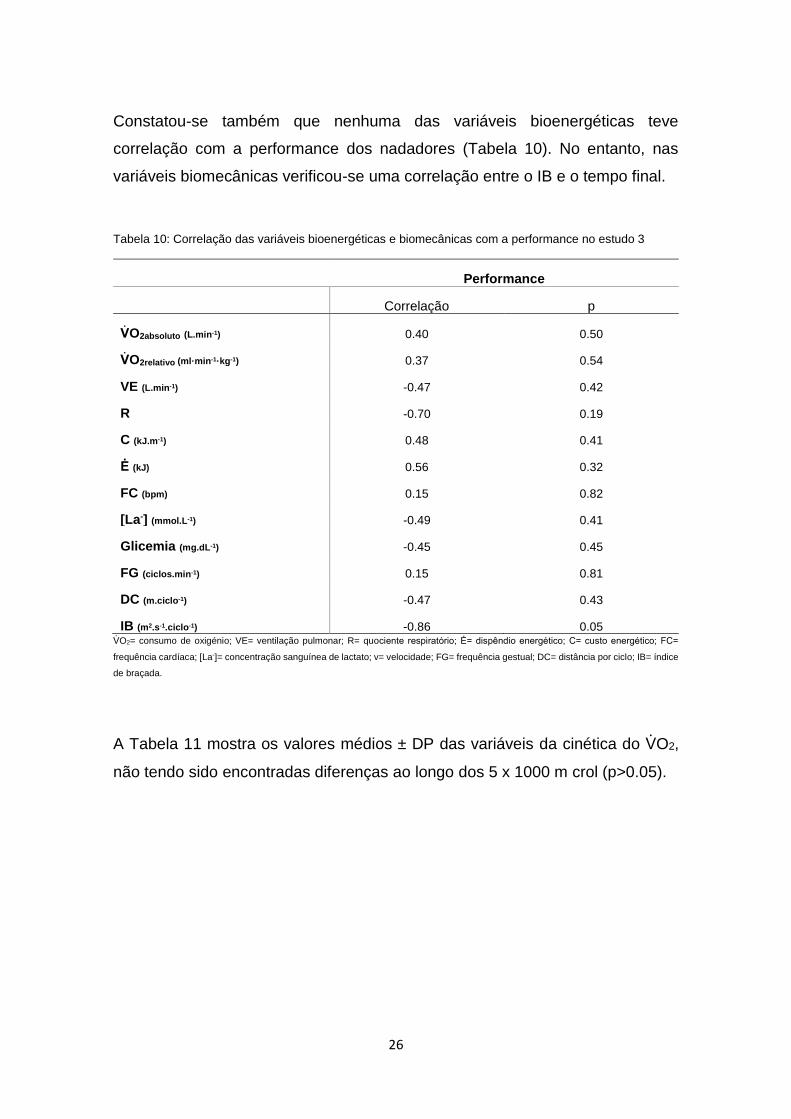
Constatou-se também que nenhuma das variáveis bioenergéticas teve
correlação com a performance dos nadadores (Tabela 10). No entanto, nas
variáveis biomecânicas verificou-se uma correlação entre o IB e o tempo final.
Tabela 10: Correlação das variáveis bioenergéticas e biomecânicas com a performance no estudo 3
Performance
Correlação p
VO2absoluto (L.min-1) 0.40 0.50
VO2relativo (ml·min-1·kg-1) 0.37 0.54
VE (L.min-1) -0.47 0.42
R -0.70 0.19
C (kJ.m-1) 0.48 0.41
Ė (kJ) 0.56 0.32
FC (bpm) 0.15 0.82
[La-] (mmol.L-1) -0.49 0.41
Glicemia (mg.dL-1) -0.45 0.45
FG (ciclos.min-1) 0.15 0.81
DC (m.ciclo-1) -0.47 0.43
IB (m2.s-1.ciclo-1) -0.86 0.05 VO2= consumo de oxigénio; VE= ventilação pulmonar; R= quociente respiratório; Ė= dispêndio energético; C= custo energético; FC=
frequência cardíaca; [La-]= concentração sanguínea de lactato; v= velocidade; FG= frequência gestual; DC= distância por ciclo; IB= índice
de braçada.
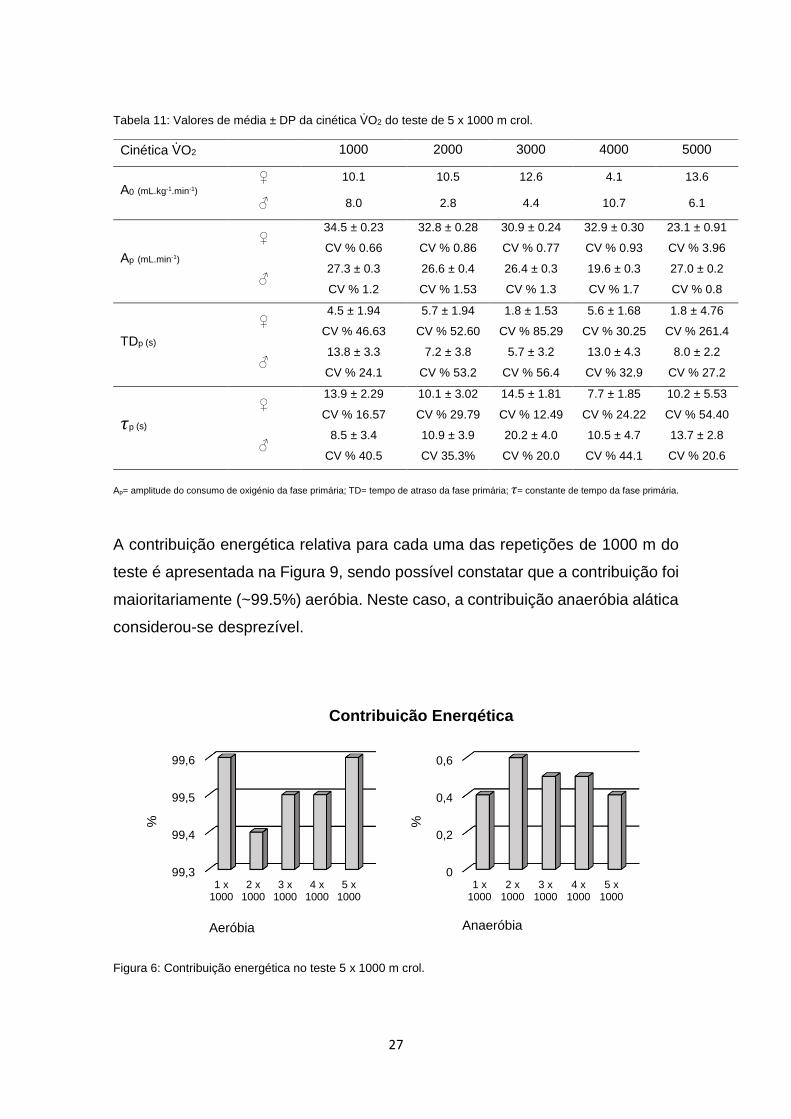
A Tabela 11 mostra os valores médios ± DP das variáveis da cinética do VO2,
não tendo sido encontradas diferenças ao longo dos 5 x 1000 m crol (p>0.05).
27
Tabela 11: Valores de média ± DP da cinética VO2 do teste de 5 x 1000 m crol.
Ap= amplitude do consumo de oxigénio da fase primária; TD= tempo de atraso da fase primária; 𝜏= constante de tempo da fase primária.
A contribuição energética relativa para cada uma das repetições de 1000 m do
teste é apresentada na Figura 9, sendo possível constatar que a contribuição foi
maioritariamente (~99.5%) aeróbia. Neste caso, a contribuição anaeróbia alática
considerou-se desprezível.
Figura 6: Contribuição energética no teste 5 x 1000 m crol.
99,3
99,4
99,5
99,6
1 x1000
2 x1000
3 x1000
4 x1000
5 x1000
%
Aeróbia
0
0,2
0,4
0,6
1 x1000
2 x1000
3 x1000
4 x1000
5 x1000
%
Anaeróbia
Cinética VO2 1000 2000 3000 4000 5000
A0 (mL.kg-1.min-1) ♀ 10.1 10.5 12.6 4.1 13.6
♂ 8.0 2.8 4.4 10.7 6.1
Ap (mL.min-1)
♀ 34.5 ± 0.23
CV % 0.66
32.8 ± 0.28
CV % 0.86
30.9 ± 0.24
CV % 0.77
32.9 ± 0.30
CV % 0.93
23.1 ± 0.91
CV % 3.96
♂ 27.3 ± 0.3
CV % 1.2
26.6 ± 0.4
CV % 1.53
26.4 ± 0.3
CV % 1.3
19.6 ± 0.3
CV % 1.7
27.0 ± 0.2
CV % 0.8
TDp (s)
♀ 4.5 ± 1.94
CV % 46.63
5.7 ± 1.94
CV % 52.60
1.8 ± 1.53
CV % 85.29
5.6 ± 1.68
CV % 30.25
1.8 ± 4.76
CV % 261.4
♂ 13.8 ± 3.3
CV % 24.1
7.2 ± 3.8
CV % 53.2
5.7 ± 3.2
CV % 56.4
13.0 ± 4.3
CV % 32.9
8.0 ± 2.2
CV % 27.2
𝜏p (s)
♀ 13.9 ± 2.29
CV % 16.57
10.1 ± 3.02
CV % 29.79
14.5 ± 1.81
CV % 12.49
7.7 ± 1.85
CV % 24.22
10.2 ± 5.53
CV % 54.40
♂ 8.5 ± 3.4
CV % 40.5
10.9 ± 3.9
CV 35.3%
20.2 ± 4.0
CV % 20.0
10.5 ± 4.7
CV % 44.1
13.7 ± 2.8
CV % 20.6
Contribuição Energética

28

29
4. Discussão
A rendimento desportivo na natação é influenciada por diversos fatores, tendo
os bioenergéticos e os biomecânicos elevada importância (Fernandes et al.,
2008). Assim, tratando-se de uma modalidade técnica, é prudente fazer uma
avaliação das características biomecânicas em paralelo com as respostas
fisiológicas (Pyne et al., 2000). O propósito desta dissertação foi apresentar uma
análise global das alterações fisiológicas e biomecânicas na prova de 5000 m e
num teste experimental de 5 x 1000 m, ambas na técnica de crol, a uma
intensidade correspondente à máxima fase estável de lactato (MLSS). O MLSS
é definido como a maior [La-] e maior carga de trabalho que pode ser mantida ao
longo do tempo sem acumulação continua de lactato, sendo considero o padrão
de ouro para prever o desempenho aeróbio e o limiar anaeróbio (Beneke &
Duvillard, 1996; Beneke, 2003; Faude et al., 2009; Pelarigo et al. 2011). É medida
em exercícios prolongados a intensidade constante e define uma intensidade de
exercício suportável pelo tempo acima referido (Baron et al., 2008), tal como
acontece nos estudos desta dissertação.
Ao analisarmos globalmente os valores por nós obtidos é possível constatar que
todas as variáveis bioenergéticas avaliadas nos três estudos apresentaram
valores estáveis durante as avaliações, algo que pode ser explicado pela
intensidade constante e moderada a que quer a prova de 5000 m quer o teste
de 5 x 1000 m foram realizados. Já os valores biomecânicos apresentaram um
comportamento diferente, observando-se no estudo 1 uma diminuição da v, DC
e IB e uma estabilização da FG ao longo dos 5000 m, enquanto que a v foi
constate e aconteceu um aumento da FG e uma diminuição da DC e do IB no
teste de 5 x 1000 m dos estudos 2 e 3. Sabemos que a intensidade e duração
do exercício provocam alterações quer fisiológicas quer biomecânicas nos
nadadores, levando a adaptações fisiológicas e reajustes biomecânicos devido
ao aparecimento de fadiga (Pelarigo et al., 2016).

30
Na presente dissertação, os valores de VO2 foram medidos diretamente
respiração-a-respiração em função do tempo para uma intensidade de nado
constante e moderada. O comportamento das trocas gasosas (especialmente o
VO2) e o estado de acidez do sangue (monitorizado através da medição da
[La-]), em exercícios de carga constante, são utilizados para definir quatro
domínios de intensidade: baixa-moderada, pesada, severa (Burnley & Jones,
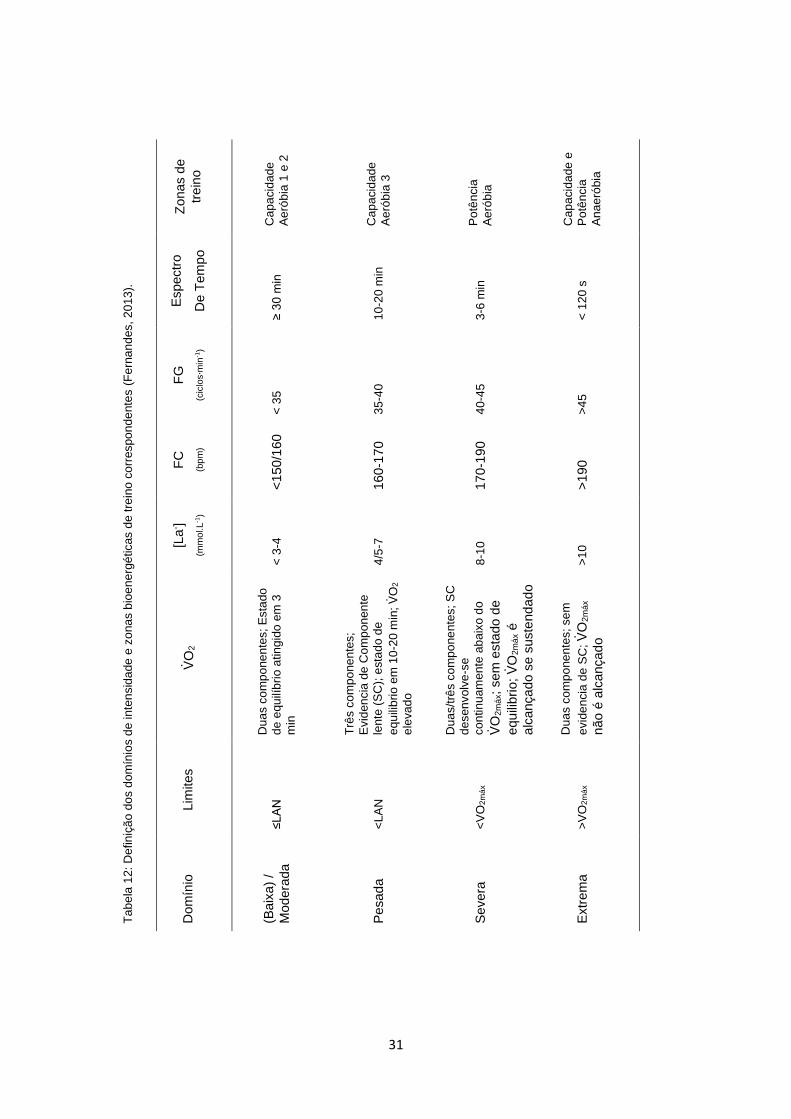
2007) e extrema (Hill et al., 2002). A Tabela 12 apresenta um resumo dos quatro
domínios de intensidade do exercício, os parâmetros que delimitam os seus
limites, as respostas fisiológicas e biomecânicas e respetivo espectro de tempo
e as zonas bioenergéticas de treino (Fernandes, 2013).
Nadar a uma intensidade continua e baixa a moderada depende da capacidade
de extrair, distribuir e utilizar O2, da capacidade em manter a exercício dentro do
limiar anaeróbio (LAN) e da capacidade em realizar o exercício eficientemente
com o menor C possível (Fernandes, 2013). O estudo da cinética do VO2 incide
nos mecanismos fisiológicos responsáveis pela resposta dinâmica do VO2 ao
exercício e subsequente recuperação (Sousa et al., 2014), sendo descrita como
um processo com três fases (Poole & Erickson, 2011; Whipp & Ward, 1990): (i)
fase I ou cardiodinâmica, está associada a um aumento abrupto do VO2 como
resultado do aumento do débito cardíaco e, consequentemente, do fluxo
sanguíneo pulmonar e tem uma duração ~15-20 s (Whipp, 1987); (ii) fase II ou
fase fundamental, primária ou componente rápida, reflete a cinética do VO2 do
músculo e tem uma constante de tempo de 30-45 s (Burnley & Jones, 2007); (iii)
fase III ou fase de estabilização, corresponde ao estado de equilíbrio do VO2 que
é atingido ao fim de 2-3 min (Xu & Rhodes, 1999). A analise da cinética do VO2
é uma ferramenta útil para o processo de treino e para a avaliação dos
nadadores (Fernandes & Vilas-Boas, 2012; Reis et al., 2012b) especialmente
para uma melhor compreensão da tolerância ao exercício e do efeito que
variáveis fisiológicas têm na resposta do VO2 ao exercício (Rodriguez et al., 2003;
Reis et al., 2012a; Sousa et al., 2011).
31
T
ab
ela
12:
Defin
içã
o d
os d
om
ínio
s d
e in
ten
sid
ade
e z
ona
s b
ioe
ne
rgé
ticas d
e tre
ino
co
rrespo
nd
ente
s (
Fe
rna
nd
es, 2
01
3).
Zonas d
e
trein
o
Ca
pa
cid
ade
Ae
rób
ia 1
e 2
Ca
pa
cid
ade
Ae
rób
ia 3
Po
tên
cia
Ae
rób
ia
Ca
pa
cid
ade
e
Po
tên
cia
An
ae
róbia
Espectr
o
De T
em
po
≥ 3
0 m
in
10
-20
min
3-6
min
< 1
20
s
FG
(cic
los
∙min
-1)
< 3
5
35
-40
40
-45
>4
5
FC
(bp
m)
<150/1
60
160-1
70
170-1
90
>190
[La
- ]
(mm
ol.L
-1)
< 3
-4
4/5
-7
8-1
0
>1
0
VO
2
Dua
s c
om
po
ne
nte
s;
Esta
do
de
equ
ilíb
rio a
tin
gid
o e
m 3
min
Trê
s c
om
po
ne
nte
s;
Evid
en
cia
de
Co
mp
on
en
te
len
te (
SC
); e
sta
do d
e
eq
uili
brio
em
10-2
0 m
in; V
O2
ele
va
do
Dua
s/t
rês c
om
po
ne
nte
s;
SC
de
sen
volv
e-s
e
co
ntin
uam
en
te a
ba
ixo
do
VO
2m
áx; sem
esta
do
de
equ
ilibri
o;
VO
2m
áx é
alc
ança
do s
e s
uste
nd
ad
o
Dua
s c
om
po
ne
nte
s; se
m
evid
en
cia
de
SC
; V
O2m
áx
não é
alc
ançad
o
Lim
ites
≤LA
N
<L
AN
<V
O2
má
x
>V
O2
má
x
Dom
ínio
(Baix
a)
/
Modera
da
Pesad
a
Severa
Extr
em
a

32
No estudo 3, os valores de VO2 foram constantes, tal como esperado, sendo
corroborados por estudos anteriores que referem que em intensidades baixas a
moderadas é esperado um estado de equilíbrio fisiológico do VO2 (Baron et al.,
2005; Pelarigo et al., 2016; Whipp & Ward, 1990). Os valores de VO2 por nós
encontrados são inferiores aos obtidos por Dekerle et al. (2005) e Pelarigo et al.
(2016), num protocolo de 30 min na técnica de crol a carga constante, o que
pode ser justificado por: (i) elevado treino aeróbio a que os nadadores da nossa
amostra são sujeitos no seu dia-a-dia e ao seu nível de especialização, uma vez
que se tratam de nadadores internacionais de águas abertas; (ii) a uma amostra
reduzida (n=2), a diferenças antropométricas entre os nadadores e à duração do
teste (≈ 1h10).
Verificou-se também que, para a intensidade a que o teste de 5x1000 m crol foi
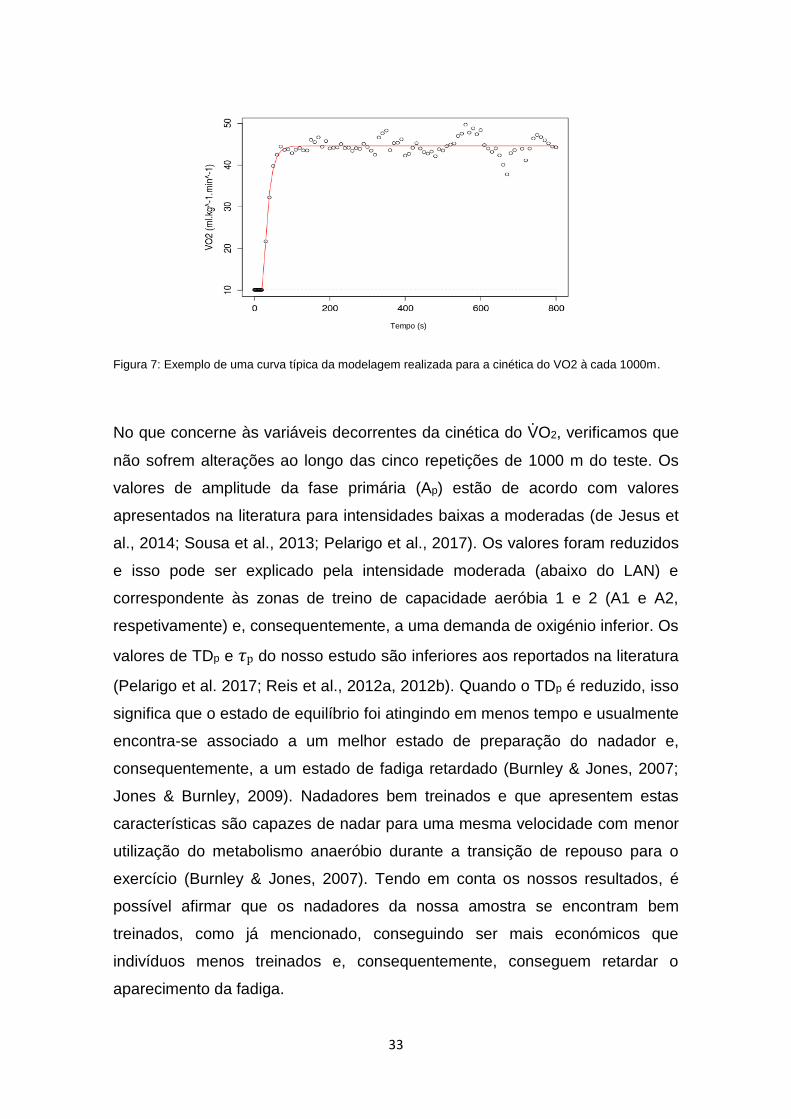
executado, a cinética do VO2 pôde ser descrita considerando-se a existência de
uma componente rápida, que reflete um processo de controlo fisiológico
subjacente até se atingir um estado de equilíbrio (Burnley & Jones, 2007),
levando-nos a utilizar um ajuste monoexponencial (Figura 7).
Consequentemente, a utilização de um ajuste biexponencial tornou-se
desnecessária, sendo usual a sua utilização em exercícios de intensidade
superior (pesada e severa) onde ocorre uma fase denominada de componente
lenta. Ao analisarmos estes resultados surgiu uma questão importante: será que
o facto deste teste ser intermitente diferencia os valores de VO2 e da sua cinética
de uma prova de 5000 m (nado continuo) dentro da mesma intensidade?
Contudo, ao fazermos uma revisão da literatura constatamos que estudos
anteriores encontraram valores similares de VO2 em exercícios intermitentes
(testes de limiar anaeróbio, limiar ventilatório e limiar de FC) e contínuos (testes
de MLSS) (Pelarigo et al., 2017), contudo é claramente necessária uma maior
investigação neste domínio.
33
Figura 7: Exemplo de uma curva típica da modelagem realizada para a cinética do VO2 à cada 1000m.
No que concerne às variáveis decorrentes da cinética do VO2, verificamos que
não sofrem alterações ao longo das cinco repetições de 1000 m do teste. Os
valores de amplitude da fase primária (Ap) estão de acordo com valores
apresentados na literatura para intensidades baixas a moderadas (de Jesus et
al., 2014; Sousa et al., 2013; Pelarigo et al., 2017). Os valores foram reduzidos
e isso pode ser explicado pela intensidade moderada (abaixo do LAN) e
correspondente às zonas de treino de capacidade aeróbia 1 e 2 (A1 e A2,
respetivamente) e, consequentemente, a uma demanda de oxigénio inferior. Os
valores de TDp e 𝜏p do nosso estudo são inferiores aos reportados na literatura
(Pelarigo et al. 2017; Reis et al., 2012a, 2012b). Quando o TDp é reduzido, isso
significa que o estado de equilíbrio foi atingindo em menos tempo e usualmente
encontra-se associado a um melhor estado de preparação do nadador e,
consequentemente, a um estado de fadiga retardado (Burnley & Jones, 2007;
Jones & Burnley, 2009). Nadadores bem treinados e que apresentem estas
características são capazes de nadar para uma mesma velocidade com menor
utilização do metabolismo anaeróbio durante a transição de repouso para o
exercício (Burnley & Jones, 2007). Tendo em conta os nossos resultados, é
possível afirmar que os nadadores da nossa amostra se encontram bem
treinados, como já mencionado, conseguindo ser mais económicos que
indivíduos menos treinados e, consequentemente, conseguem retardar o
aparecimento da fadiga.
Tempo (s)

34
A VE, sendo o volume de ar que entra e sai dos pulmões, mantive-se estável ao
longo dos 5 x 1000 m crol. Estes dados estão em concordância com estudos
anteriores (Baron et al., 2005; Pelarigo et al., 2016) que reportam valores
similares em ciclismo e natação (respetivamente). Esta estabilidade aconteceu,
pois a variação da VE ocorre em simultâneo com a variação do VO2 e este
manteve-se constante. Já o R, é o quociente entre o volume de CO2 expirado e
de O2 consumido, indicando o tipo de substrato que está a ser utilizado em
determinado esforço (Wilmore et al., 2008). De acordo com a literatura,
aproximadamente 75% do O2 consumido é convertido em CO2, portanto, em
repouso, o R varia entre os 0.78 e os 0.80 (Neto et al., 2001), valores ligeiramente
inferiores aos obtidos no nosso estudo. Essa diferença pode ser explicada pelo
facto de os nossos valores de repouso serem considerados os valores
imediatamente antes do inicio do teste (pré-exercício). Após o inicio do exercício
os valores de R aumentaram e estabilizaram próximos da unidade. Estes valores
sugerem uma maior utilização de CHO, facto diretamente associado a uma maior
solicitação de fibras musculares tipo II (Robergs & Roberts, 2000; Wilmore &
Costill, 2001), fibras altamente recrutadas em exercícios de endurance, como o
caso do nosso teste de 5 x 1000 m crol. As diferenças de valores que foram
observadas também podem ser justificada pela razão sinal-ruído, que foi
diminuindo ao longo do teste.
A [La-] manteve-se constante, tal como esperado em exercícios de intensidades
moderadas, onde ocorre um estado de equilíbrio fisiológico na [La-] (Baron et al.,
2005; Whipp & Ward, 1990), ou seja, o lactato é produzido e removido em
proporções iguais (Beneke et al., 2011; Billat et al., 2003). Os valores
encontrados (2.6 mmol·L-1 no estudo 1, 2.4 mmol·L-1 no estudo 2 e 2.0 mmol·L-
1 no estudo 3) são similares aos encontrados na literatura, que descrevem uma
variação de 1.9 a 3.3 mmol·L-1, em nado a intensidades moderadas (Baron et
al., 2005; Dekerle et al., 2005; Fernandes et al., 2011; Pelarigo et al., 2011). Já
Olbrecht (2000) considera que, dependendo do tipo de nadador, os valores
lácticos esperados para a capacidade aeróbia devem rondar 1-2.5 mmol·L-1.
Distâncias mais longas são caracterizadas por velocidades mais reduzidas e isso
pode explicar os valores mais baixos de [La-] encontrados. Isto pode acontecer

35
devido a uma maior percentagem de fibras musculares de contração lenta, que
aumentam a remoção de lactato sanguíneo após o exercício (Avlonitov, 1996).
Complementarmente, constatámos que as raparigas da nossa amostra
apresentaram (nos estudos 1 e 2) valores inferiores de [La-], quando comparadas
aos rapazes. Esta diferença corrobora estudos recentes (Crewther et al., 2006;
Greco et al., 2007) e pode ser explicada por uma menor massa muscular das
raparigas (Crewther et al., 2006) e uma maior concentração de testosterona nos
rapazes (di Prampero et al., 2011). Os valores mais elevados da rapariga no
estudo 3, podem ser explicados por uma maior intensidade na realização do
teste de 5 x 1000 m, corroborado pelo valores de %FCmáx.
Também os valores de glicemia foram estáveis e são corroborados pela literatura
onde se afirma que a manutenção da glicemia é importante para evitar a
glicogenólise e atenuar distúrbios homeostáticos, como a hipoglicemia. O
comportamento glicémico é, em grande parte, determinado pela intensidade do
exercício (Silva & Azevedo, 2010), i.e., a variação dos valores de glicemia é
influenciada pela intensidade do exercício, pelo nível físico dos sujeitos, pela
capilarização, concentração e dimensões das mitocôndrias (McArdle et al.,
2011). A ingestão de CHO durante exercício prolongado retarda a fadiga
(Murray, 2007); em exercícios prolongados o glicogénio muscular esgota-se e é
necessário a ingestão de CHO exógeno (Kumstat et al., 2016). O aumento da
glicemia indica melhor performance e nadadores com maior declínio têm,
usualmente, prestações inferiores (Hara & Muraora, 2015), tal como verificamos
com os nadadores que estudámos. A monitorização e compreensão do
comportamento glicémico pode ser uma ferramenta extremamente útil para os
treinadores, fornecendo informações sobre as respostas orgânicas dos seus
nadadores ao exercício. Apesar disso, é necessário ter em consideração que o
comportamento glicémico é muito individual (Hara & Muraora, 2015).
A FC foi controlada continuamente em intervalos de 5s e, após um aumento
substancial no início do exercício, manteve-se estável ao longo de todo o
protocolo. A FC pré-exercício eleva-se a níveis mais altos do que os de repouso
(70-80bpm), conhecido como a resposta antecipatória ao estímulo (McArdle et

36
al., 2011), que explica os dados obtidos no momento por nós designado de
repouso e que nada mais é do que um momento pré-exercício. Nos estudos
pioneiros que avaliaram a FC em natação, foi afirmado que é alcançado um
plateau de FC para nadadores bem treinados 90s após o inicio do exercício
(Treffene et al., 1980), tal como acontece no nosso estudo. Segundo Fernandes
(2013), os valores da FC em intensidades moderadas são, habitualmente,
inferiores a 160bpm, valor inferior à média do nosso estudo (165bpm). Contudo,
a literatura mostra-nos que, em exercícios prolongados, a FC pode aumentar,
atingindo valores de 150-178bpm (Dekerle et al., 2005; Fernandes et al., 2011;
Pelarigo et al., 2016).
A performance em natação é definida como emergindo da conversão do Ė em
trabalho propulsivo a uma determinada eficiência energética. O C é definido
como a energia despendida para percorrer uma determinada distância (Barbosa
et al., 2010; diPrampero et al., 1974, 2011; Pendergast et al., 2003; Zamparo et
al., 2011). O calculo de C permite determinar a participação energética aeróbia
e anaeróbia e avaliar quantitativamente o nível de adequação mecânica global
do gesto técnico (Silva et al., 2006). Assim sendo, tanto o Ė como o C diferem
com a intensidade do exercício e, consequentemente, com a sua duração
(diPrampero et al., 2011). O Ė, calculado utilizando o C, é muito importante uma
vez que a um C inferior os nadadores mais económicos conseguem nadar a uma
menor percentagem da sua potência aeróbia máxima e poupar substratos
energéticos (Zamparo et al., 2005). Para o calculo do Ė e, consequentemente,
do C, foram utilizados os valores de VO2 e [La-]. Os valores encontrados foram
constantes ao longo do teste de 5 x 1000 m, o que pode acontecer devido à
qualidade técnica dos nadadores da amostra, i.e., quanto mais proficientes
tecnicamente menor será o custo energético (Wakayoshi et al., 1993; Silva et al.,
2006), e à estabilidade de velocidade que se verificou no teste, quanto maior a
variação de velocidade, maior o Ė e o C (Barbosa et al., 2006; Toussaint &
Hollander, 1994).
Grande parte, se não mesmo quase a totalidade, da contribuição energética nos
5000 m estudados foi proveniente do metabolismo aeróbio, contribuindo com

37
mais de 99% do Ė total. Aliás, nadar a intensidades definidas dentro da
capacidade aeróbia 1 e 2, a contribuição energética parece depender quase
exclusivamente do sistema energético aeróbio, sendo a depleção de substrato,
alterações de hidratação ou problemas de termorregulação, a razão da
finalização do exercício em longas distâncias (Fernandes, 2013). Do nosso
conhecimento apenas Pelarigo et al. (2017) estudaram as contribuições
anaeróbia e aeróbia num exercício a intensidades próximas do MLSS, tendo
obtido resultados que vêm consolidar os nossos. Os restantes estudos
estudaram essa contribuição energética durante esforços máximos (Ogita et al.,
1996, 1999; Sousa et al., 2014). Estas evidências realçam que a contribuição
energética em intensidades na e próximas da MLSS são principalmente e quase
exclusivamente controladas pelo sistema oxidativo (Pelarigo et al., 2017).
A avaliação de fatores biomecânicos tem sido considerada bastante relevante
para o treino e para a descrição do desempenho (Figueiredo et al., 2011;
Keskinen & Komi, 1993; Termin & Pendergast, 2000), sendo a DC aceite como
um elemento fundamental para o sucesso do nadador (Costill et al., 1985;
Figueiredo et al., 2011) e o IB a expressão da capacidade do nadador se
deslocar a uma determinada v com o menor número possível de braçadas
(Costill et al., 1985). O comportamento das variáveis biomecânicas na prova de
5000 m no campeonato nacional de longa distância, onde a v, a DC e o IB
diminuíram ao longo da prova, enquanto a FG se manteve estável, está de
acordo com o que se encontra descrito na literatura (Alberty et al., 2009; Craig &
Pendergast, 1979; Craig et al., 1985). A diminuição de v, DC e IB pode estar
associada ao desenvolvimento da fadiga muscular (Chollet et al., 1997; Keskinen
& Komi, 1993; Pendergast et al., 1979), que conduz a uma diminuição do output
de energia mecânica e, consequentemente, leva a uma redução da v e da DC
(Toussaint et al., 2006), isto é, um nadador fatigado perde, gradualmente, a
capacidade de produzir força para combater a resistência oposta ao seu
deslocamento (Alberty et al., 2009). Durante uma prova, há uma diminuição da
v e da DC enquanto a FG se mantém estável, aumentando no final da prova
devido ao aparecimento de cansaço (Alberty et al., 2009; Baron et al., 2005;

38
Craig & Pendergast, 1979; Craig et al., 1985; Invernizzi et al., 2014), tal como
observamos no campeonato nacional de longa distância.
Os estudos 2 e 3 mostram que para haver uma manutenção da v nos 5 x 1000
m há um aumento da FG e uma diminuição da DC e do IB, resultados que são
justificados pelos trabalhos de nado continuo de 30 min (Alberty et al., 2009;
Dekerle et al., 2005; Pelarigo et al., 2016). Aliás, em águas abertas, as
estratégias utilizadas para se manter uma v constante passam por manter um
equilíbrio metabólico (Pelayo et al., 2007), o que se verificou, e já foi
anteriormente discutido, sendo necessárias adaptações biomecânicas uma vez
que exercícios de longa duração têm uma fadiga periférica associada
(Figueiredo et al., 2014). Os parâmetros FG e DC representam o estado técnico
de um nadador e sofrem alterações com o aparecimento de fadiga (Mujika et al.,
2000). A FG nos 5 x 1000 m aumentou progressivamente ao longo das
repetições de 1000 m ao mesmo tempo que a DC e o IB foram diminuindo
continuamente. Os nossos resultados nos estudos 2 e 3 estão em concordância
com dados anteriores, que descrevem uma diminuição da DC e do IB e um
aumento da FG com o passar do tempo, quando o exercício é realizado a uma v
constante (Dekerle et al., 2005; Figueiredo et al., 2014; Pelarigo et al., 2011).
Vários investigadores demonstraram que a intensidade de nado e a fadiga
podem interferir com os parâmetros da braçada adotados pelos nadadores, com
o passar do tempo (Dekerle et al., 2005; Oliveira et al., 2012a, 2012b; Pelarigo
et al., 2011). Maior FG esta associada a um maior C, independentemente da
massa corporal e da v (Barbosa et al., 2007). Estes dados vêm reforçar os
nossos resultados, uma vez que os nadadores da nossa amostra, para
conseguirem manter a v, apresentaram uma adaptação biomecânica que
aconteceu sobretudo devido ao aparecimento da fadiga devido ao exercício
prolongado.
Como curiosidade, a prova de 5000 m teve uma v média de 1.31 ± 0.08 m.s-1 e
o teste de 5 x 1000 m de 1.30 ± 0.07 m.s-1. Estes valores são ligeiramente
inferiores aqueles encontrados por Zingg et al., (2014) que ao analisarem provas
de 5 km verificaram uma v média de 1.47 ± 0.20 m.s-1. Em provas internacionais
de 10 km a v média apresentada foi 1.35 ± 0.09 m.s-1 para mulheres e 1.45 ± 0.10

39
m.s-1 para homens (Vogt et al., 2013). Esta diferença nos valores pode ser
explicada pela disparidade do nível competitivo dos nadadores em causa, mas
também pela desigualdade da importância das competições, uma vez que o
nosso estudo analisou nadadores portugueses na fase de qualificação do
campeonato nacional de longa distância, enquanto Zingg et al., (2014) e Vogt et
al., (2013) estudaram nadadores internacionais em provas da FINA World Cup.
Os fatores biomecânicos são muito influenciados pelo género (Barbosa et al.,
2006, 2008) onde as mulheres apresentam valores mais elevados de FG e os
homens de DC, IB e v, tal como aconteceu nos nossos resultados. Estas
diferenças entre géneros podem ser explicadas por diferenças antropométricas
e fisiológicas. O sexo masculino apresentar valores mais elevados de IB e DC,
indo de acordo com o que estudos anteriores revelam (Chollet et al., 2000; Greco
et al., 2007; Pelayo et al., 1996; Seifert et al., 2007). Isto pode ser devido a uma
maior capacidade dos rapazes em aplicar mais potência na braçada, em superar
um maior arrasto, causado por uma maior altura e maior área de secção
transversa (Toussaint et al., 2006) e pelas dimensões superiores dos MS
(Barbosa et al., 2008). Outras justificações podem ser a sua maior capacidade
aeróbia, maior massa muscular e maior concentração de androgénio (VanHeest
et al., 2004).

40

41
5. Conclusão
As conclusões que podemos retirar desta dissertação enaltecem a importância
do estudo de fatores biofísicos na avaliação da performance dos nadadores em
longa distância, visando a capacidade aeróbia do nadador. Assim, as conclusões
desta dissertação são:
i) Existe uma relação entre o perfil bioenergético e biomecânico dos nadadores;
ii) Um esforço de 5000 m na técnica de crol é realizado no domínio de
intensidade baixa-moderada e é caracterizado por uma cinética do VO2 com
três fases, existindo um equilíbrio metabólico, com o nadador rearranjando a
sua técnica (combinações entre FG e DC) para manter a sua homeostase e
ritmo de nado no final do esforço;
iii) Em exercícios prolongados há um aumento da FG e diminuição da DC e do
IB associados ao aparecimento da fadiga muscular, sendo necessário o treino
técnico para otimizar estes ajustes biomecânicos tornando o nadador mais
eficiente e, consequentemente, mais económico;
iv) Os nadadores de águas abertas realizam as suas provas em intensidades
próximas do LAN ou até mesmo ao LAN. Esta informação torna-se uma boa
ferramenta para a prescrição do treino destes nadadores, i.e., os treinadores
devem potencializar a base aeróbia dos nadadores sem descurar as restantes
zonas de treino;
v) A ingestão de CHO em exercícios prolongados, como é o caso das provas de
águas abertas, é fundamental para retardar o aparecimento de fadiga e
possibilitar uma melhor resposta às variações de intensidade que existem no
desenrolar de uma prova. Tendo em consideração que o comportamento
glicémico é muito individual cada nadador deverá pensar, estudar e idealizar
o seu abastecimento com base nas suas características específicas;

42

43
6. Sugestões para estudos futuros
Esta dissertação fornece aos treinadores de natação algumas evidencias iniciais
e objetivas para a compreensão do comportamento biomecânico e bioenergético
de um teste de 5 x 1000 m. Estudos com foco na disciplina de águas abertas são
muito escassos e, apesar do primeiro passo já ter sido dado neste trabalho com
a caracterização biofísica, continuam a existir ainda muitas falhas e lacunas,
sendo propósito do nosso grupo reduzi-las em estudos futuros, nomeadamente:
i) Realizando um teste de 5 x 1000 m em meio natural (nomeadamente
uma pista) para caraterização biofísica dos nadadores em situação
real de águas abertas;
ii) Replicar o teste anterior tendo como foco a prova de 10 km, ou seja,
fazendo um teste de 10 x 1000 m. Sendo esta a prova do calendário
olímpico, é a distância com mais enfase em competições nacionais e
internacionais;
iii) Estudo em situação de competição de águas abertas, com controlo da
velocidade e das estratégias de nado mais utilizadas. Em seguida,
reproduzir essas estratégias em protocolos experimentais, com
caracterização biofísica do esforço, com o objetivo de perceber qual a
mais económica dentro daquelas que são mais utilizadas pelos
principais nadadores nacionais e internacionais;
iv) Procurar perceber de que forma o fator psicológico tem influencia no
resultado final, estudando o seu peso no controlo emocional ao longo
da prova, nas tomadas de decisão e nas estratégias utilizadas.

44

45
7. Bibliografia
Alberty, M., Sidney, M., Pelayo, P., & Toussaint, H. M. (2009). Stroking
characteristics during time to exhaustion tests. Medicine and Science in
Sports and Exercise, 41(3), 637-644.
Aspenes, S. T., & Karlsen, T. (2012). Exercise-training intervention studies in
competitive swimming. International Journal of Sports Medicine, 42(6), 527-
543.
Avlonitou, E. (1996). Maximal lactate values following competitive performance
varying according to age, sex and swimming style. The Journal of Sports
Medicine and Physical Fitness, 36(1), 24-30.
Baldassarre, R., Bonifazi, M., Zamparo, P., & Piacentini, M. F. (2017).
Characteristics and Challenges of Open-Water Swimming Performance: A
Review. International Journal of Sports Physiology and Performance, 1-35.
Barbosa, T. M., Bragada, J. A., Reis, V. M., Marinho, D. A., Carvalho, C., & Silva,
A. J. (2010). Energetics and biomechanics as determining factors of swimming
performance: updating the state of the art. Journal of Science and Medicine in
Sport, 13(2), 262-269.
Barbosa, T. M., Fernandes, R. J., Keskinen, K. L., & Vilas-Boas, J. P. (2008). The
influence of stroke mechanics into energy cost of elite swimmers. European
Journal of Applied Physiology, 103(2), 139-149.
Barbosa, T. M., Garrido, M. F., & Bragada, J. (2007). Physiological adaptations
to head-out aquatic exercises with different levels of body immersion. The
Journal of Strength & Conditioning Research, 21(4), 1255-1259.
Barbosa, T., Keskinen, K., & Vilas-Boas, J. P. (2006). Factores biomecânicos e
bioenergéticos limitativos do rendimento em natação pura
desportiva. Motricidade, 2(4).

46
Baron, B., Dekerle, J., Depretz, S., Lefevre, T., & Pelayo, P. (2005). Self selected
speed and maximal lactate steady state speed in swimming. The Journal of
Sports Medicine and Physical Fitness, 45(1), 1.
Baron, B., Noakes, T. D., Dekerle, J., Moullan, F., Robin, S., Matran, R., & Pelayo,
P. (2008). Why does exercise terminate at the maximal lactate steady state
intensity?. British Journal of Sports Medicine, 42(10), 828-833.
Beneke, R. (2003). Methodological aspects of maximal lactate steady state—
implications for performance testing. European Journal of Applied
Physiology, 89(1), 95-99.
Beneke, R., & von Duvillard, S. P. (1996). Determination of maximal lactate
steady state response in selected sports events. Medicine and Science in
Sports and Exercise, 28(2), 241-246.
Beneke, R., Heck, H., Hebestreit, H., & Leithäuser, R. M. (2009). Predicting
maximal lactate steady state in children and adults. Pediatric Exercise
Science, 21(4), 493-505.
Beneke, R., Leithäuser, R. M., & Ochentel, O. (2011). Blood lactate diagnostics
in exercise testing and training. International Journal of Sports Physiology and
Performance, 6(1), 8-24.
Bentley, D. J., Newell, J., & Bishop, D. (2007). Incremental exercise test design
and analysis. International Journal of Sports Medicine, 37(7), 575-586.
Billat, V. L., Sirvent, P., Py, G., Koralsztein, J. P., & Mercier, J. (2003). The
concept of maximal lactate steady state. International Journal of Sports
Medicine, 33(6), 407-426.
Borges, J. M. (2015). Águas abertas, o outro lado da natação. Actas do 37º
Congresso Técnico Científico da APTN. Anadia.
Burnley, M., & Jones, A. M. (2007). Oxygen uptake kinetics as a determinant of
sports performance. European Journal of Sport Science, 7(2), 63-79.

47
Canossa, S., Fernandes, R. J., Carmo, C., Andrade, A., & Soares, S. M. (2007).
Ensino multidisciplinar em natação: reflexão metodológica e proposta de lista
de verificação. Motricidade, 3(4).
Chollet, D., Chalies, S., & Chatard, J. C. (2000). A new index of coordination for
the crawl: description and usefulness. International Journal of Sports
Medicine, 21(01), 54-59.
Chollet, D., Delaplace, C., Pelayo, P., Tourny, C., & Sidney, M. (1997). Stroking
characteristic variations in the 100-m freestyle for male swimmers of differing
skill. Perceptual and Motor Skills, 85(1), 167-177.
Costill, D. L., Bennett, A., Branam, G., & Eddy, D. (1973). Glucose ingestion at
rest and during prolonged exercise. European Journal of Applied
Physiology, 34(6), 764-769.
Costill, D. L., Kovaleski, J., Porter, D., Kirwan, J., Fielding, R., & King, D. (1985).
Energy expenditure during front crawl swimming: predicting success in middle-
distance events. International Journal of Sports Medicine, 6(05), 266-270.
Craig, A. B., & Pendergast, D. R. (1979). Relationships of stroke rate, distance
per stroke, and velocity in competitive swimming. Medicine and Science in
Sports and Exercise, 11(3), 278-283.
Craig, A. B., Skehan, P. L., Pawelczyk, J. A., & Boomer, W. L. (1985). Velocity,
stroke rate, and distance per stroke during elite swimming
competition. Medicine and Science in Sports and Exercise, 17(6), 625-634.
Crewther, B., Keogh, J., Cronin, J., & Cook, C. (2006). Possible stimuli for
strength and power adaptation. International Journal of Sports
Medicine, 36(3), 215-238.
De Jesus, K., Guidetti, L., Vilas-Boas, J. P., Baldari, C., & Fernandes, R. J.
(2014). Which are the best VO2 sampling intervals to characterize low to
severe swimming intensities?. International Journal of Sports
Medicine, 35(12), 1030-1036.

48
Dekerle, J., Pelayo, P., Clipet, B., Depretz, S., Lefevre, T., & Sidney, M. (2005).
Critical swimming speed does not represent the speed at maximal lactate
steady state. International Journal of Sports Medicine, 26(07), 524-530.
di Prampero, P. E. (1986). The energy cost of human locomotion on land and in
water. International Journal of Sports Medicine, 7(02), 55-72.
di Prampero, P. E., Meyer, M., Cerretelli, P., & Piiper, J. (1978). Energetics of
anaerobic glycolysis in dog gastrocnemius. European Journal of Applied
Physiology, 377(1), 1-8.
di Prampero, P. E., Pendergast, D. R., Wilson, D. W., & Rennie, D. W. (1974).
Energetics of swimming in man. European Journal of Applied
Physiology, 37(1), 1-5.
di Prampero, P. E., Pendergast, D., Zamparo, P. (2011). Swimming economy
(Energy Cost) and Efficiency. In L. Seifert, D., Chollet & I. Mujika (Eds.), World
Book of Swimming: From Science to Performance (297-312). New York: Nova.
Faude, O., Kindermann, W., & Meyer, T. (2009). Lactate threshold
concepts. International Journal of Sports Medicine, 39(6), 469-490.
Fernandes, R. (1999). Caracterização fisiológica, cineantropométrica e
psicológica do nadador pré-júnior. Porto: R. Fernandes. Tese de Mestrado
apresentada à Faculdade de Ciências do Desporto e Educação Física da
Universidade do Porto.
Fernandes, R. (2013). Biophysical characterization of low to extreme Swimming
intensities: seminar.
Fernandes, R. J., Billat, V. L., Cruz, A. C., & Colaço, P. J. (2006). Does net energy
cost of swimming affect time to exhaustion at the individual's maximal oxygen
consumption velocity?. The Journal of Sports Medicine and Physical
Fitness, 46(3), 373.
Fernandes, R. J., Keskinen, K. L., Colaço, P., Querido, A. J., Machado, L. J.,
Morais, P. A., & Boas, J. V. (2008). Time Limit at VO2max Velocity in Elite
Crawl Swimmers. International Journal of Sports Medicine, 29(02), 145-150.

49
Fernandes, R. J., Sousa, M., Machado, L., & Vilas-Boas, J. P. (2011). Step length
and individual anaerobic threshold assessment in swimming. International
Journal of Sports Medicine, 32(12), 940-946.
Fernandes, R., & Vilas-Boas, J. P. (2006). Tempo limite à intensidade mínima
correspondente ao consumo máximo de oxigénio: novos desenvolvimentos
num parâmetro de recente investigação em natação. Motricidade, 2(4).
Fernandes, R., & Vilas-Boas, J. P. (2012). Time to exhaustion at the VO2max
velocity in swimming: a review. Journal of Human Kinetics, 32, 121-134.
Fernandes, R., Oliveira, E., & Colaço, P. (2009). Bioenergetical assessment and
training control as useful tools to improve performance in cyclic sports. Journal
of Contemporary Athletics, 41(1), 51-72.
Figueiredo, P., Nazario, R., Sousa, M., Pelarigo, J. G., Vilas-Boas, J. P., &
Fernandes, R. (2014). Kinematical analysis along maximal lactate steady state
swimming intensity. Journal of Sports Science and Medicine, 13(3), 610.
Figueiredo, P., Pendergast, D. R., Vilas-Boas, J. P., & Fernandes, R. J. (2013).
Interplay of biomechanical, energetic, coordinative, and muscular factors in a
200 m front crawl swim. BioMed Research International, 2013.
Figueiredo, P., Zamparo, P., Sousa, A., Vilas-Boas, J. P., & Fernandes, R. J.
(2011). An energy balance of the 200 m front crawl race. European Journal of
Applied Physiology, 111(5), 767-777.
Fleiss, J. L. (1986). Reliability of measurement. The Design and Analysis of
Clinical Experiments, 1-32.
Greco, C. C., Pelarigo, J. G., Figueira, T. R., & Denadai, B. S. (2007). Effects of
gender on stroke rates, critical speed and velocity of a 30-min swim in young
swimmers. Journal of Sports Science and Medicine, 6(4), 441.
Hara, R., & Muraoka, I. (2015). Open water swimming performance. In Sports
Performance (pp. 313-322). Springer Japan.
Hardin, D. S., Azzarelli, B. I. A. G. I. O., Edwards, J. E. F. F., Wigglesworth, J. A.
N. E. T., Maianu, L. I. D. I. A., Brechtel, G. I. N. G. E. R., & Garvey, W. T.

50
(1995). Mechanisms of enhanced insulin sensitivity in endurance-trained
athletes: effects on blood flow and differential expression of GLUT 4 in skeletal
muscles. The Journal of Clinical Endocrinology and Metabolism, 80(8), 2437-
2446.
Hill, D. W., Poole, D. C., & Smith, J. C. (2002). The relationship between power
and the time to achieve. VO (2max). Medicine and Science in Sports and
Exercise, 34(4), 709-714.
Invernizzi, P. L., Limonta, E., Bosio, A., Scurati, R., Veicsteinas, A., & Esposito,
F. (2014). Effects of a 25-km trial on psychological, physiological and stroke
characteristics of short-and mid-distance swimmers. The Journal of Sports
Medicine and Physical Fitness, 54(1), 53-62.
Jones, A. M., & Burnley, M. (2009). Oxygen uptake kinetics: an underappreciated
determinant of exercise performance. International Journal of Sports
Physiology and Performance, 4(4), 524-532.
Keskinen, K. L., & Komi, P. V. (1993). Stroking characteristics of front crawl
swimming during exercise. Journal of Applied Biomechanics, 9(3), 219-226.
Kruger, M., Saayman, M., & Ellis, S. (2011). A motivation based typology of open-
water swimmers. South African Journal for Research in Sport, Physical
Education and Recreation, 33(2), 59-79.
Kumstát, M., Rybářová, S., Thomas, A., & Novotný, J. (2016). Case Study:
Competition Nutrition Intakes During the Open Water Swimming Grand Prix
Races in Elite Female Swimmer. International Journal of Sport Nutrition and
Exercise Metabolism, 26(4), 370-376.
Maglischo, E. W. (1999). Nadando ainda mais rápido: padrão de referência para
o nadador profissional.
Maglischo, E. W. (2003). Swimming fastest. Human Kinetics.
McArdle, W. D., Katch, F. I., & Katch, V. L. (2011). Fisiologia do Exercício –
Nutrição, Energia e Desempenho Humano. Brasil: Nova Guanabara.

51
Mujika, I, Goya, A., Padilla, S., Grijalba, A., Gorostiaga, E., & Ibanez, J. (2000).
Physiological responses to a 6-d taper in middle-distance runners: influence of
training intensity and volume. Medicine and Science in Sports and
Exercise, 32(2), 511-517.
Munatones, S. (2008). Seven essentials of open Water success. In Swimming
World Magazine online.
Murray, B. (2007). Hydration and physical performance. Journal of American
College of Nutrition, 26(sup5), 542s-548s.
Neto, T. L., Tebexreni, A. S., & Tambeiro, V. L. (2001). Aplicações práticas da
ergoespirometria no atleta. Revista da Sociedade de Cardiologia, 11(3-695),
705.
Ogita, F., Hara, M., & Tabata, I. (1996). Anaerobic capacity and maximal oxygen
uptake during arm stroke, leg kicking and whole body swimming. Acta
Physiological, 157(4), 435-441.
Ogita, F., Onodera, T., & Tabata, I. (1999). Effect of hand paddles on anaerobic
energy release during supramaximal swimming. Medicine and Science in
Sports and Exercise, 31(5), 729-735.
Olbrecht, J. (2000). The science of winning.
Oliveira, M. F. M., Caputo, F., Dekerle, J., Denadai, B. S., & Greco, C. C. (2012a).
Stroking parameters during continuous and intermittent exercise in regional-
level competitive swimmers. International Journal of Sports Medicine, 33(09),
696-701.
Oliveira, M. F., Caputo, F., Lucas, R. D., Denadai, B. S., & Greco, C. C. (2012b).
Physiological and stroke parameters to assess aerobic capacity in
swimming. International Journal of Sports Physiology and Performance, 7(3),
218-223.
Özyener, F., Rossiter, H. B., Ward, S. A., & Whipp, B. J. (2001). Influence of
exercise intensity on the on‐and off‐transient kinetics of pulmonary oxygen
uptake in humans. The Journal of Physiology, 533(3), 891-902.

52
Pelarigo, J. G., Denadai, B. S., & Greco, C. C. (2011). Stroke phases responses
around maximal lactate steady state in front crawl. Journal of Science and
Medicine in Sport, 14(2), 168-e1.
Pelarigo, J. G., Fernandes, R. J., Ribeiro, J., Denadai, B. S., Greco, C. C., &
Vilas-Boas, J. P. (2017). Comparison Of Different Methods For The Swimming
Aerobic Capacity Evaluation. The Journal of Strength & Conditioning
Research.
Pelarigo, J. G., Greco, C. C., Denadai, B. S., Fernandes, R. J., Vilas-Boas, J. P.,
& Pendergast, D. R. (2016). Do 5% changes around maximal lactate steady
state lead to swimming biophysical modifications?. Journal of Human
Movement Science, 49, 258-266.
Pelayo, P., Alberty, M., Sidney, M., Potdevin, F., & Dekerle, J. (2007). Aerobic
potential, stroke parameters, and coordination in swimming front-crawl
performance. International Journal of Sports Physiology and
Performance, 2(4), 347-359.
Pelayo, P., Mujika, I., Sidney, M., & Chatard, J. C. (1996). Blood lactate recovery
measurements, training, and performance during a 23-week period of
competitive swimming. European Journal of Applied Physiology and
Occupational Physiology, 74(1), 107-113.
Pendergast, D. R., Di Prampero, P. E., Craig, A. B., Wilson, D. R., & Rennie, D.
W. (1977). Quantitative analysis of the front crawl in men and
women. European Journal of Applied Physiology, 43(3), 475-479.
Pendergast, D., Cerretelli, P., & Rennie, D. W. (1979). Aerobic and glycolytic
metabolism in arm exercise. European Journal of Applied Physiology, 47(4),
754-760.
Pendergast, D., Zamparo, P., Di Prampero, P. E., Capelli, C., Cerretelli, P.,
Termin, A., & Mollendorf, J. (2003). Energy balance of human locomotion in
water. European Journal of Applied Physiology, 90(3-4), 377-386.

53
Plano de Alto Rendimento. Consultado a 21 Agosto 2017, disponível em
http://fpnatacao.pt/federacao/documentos/planos-de-alto-rendimento
Poole, D. C., & Erickson, H. H. (2011). Highly athletic terrestrial mammals: horses
and dogs. Comprehensive Physiology.
Pyne, D. B., Boston, T., Martin, D. T., & Logan, A. (2000). Evaluation of the
Lactate Pro blood lactate analyser. European Journal of Applied
Physiology, 82(1-2), 112-116.
Reis, J. F., Alves, F. B., Bruno, P. M., Vleck, V., & Millet, G. P. (2012b). Oxygen
uptake kinetics and middle distance swimming performance. Journal of
Science and Medicine in Sport, 15(1), 58-63.
Reis, J. F., Alves, F. B., Bruno, P. M., Vleck, V., & Millet, G. P. (2012a). Effects
of aerobic fitness on oxygen uptake kinetics in heavy intensity
swimming. European Journal of Applied Physiology, 112(5), 1689-1697.
Relatórios e Contas FPN. Consultado a 21 de Agosto de 2017, disponível em
http://fpnatacao.pt/federacao/documentos/relat%C3%B3rio-e-contas
Ribeiro, J., Figueiredo, P., Guidetti, L., Alves, F., Toussaint, H., Vilas-Boas, J. P.,
& Fernandes, R. J. (2016). AquaTrainer® snorkel does not increase
hydrodynamic drag but influences turning time. International Journal of Sports
Medicine, 37(04), 324-328.
Robergs, R. A., & Roberts, S. (2000). Fundamental principles of exercise
physiology: for fitness, performance, and health. McGraw-Hill College.
Rodriguez, F. A., Keskinen, K. L., Keskinen, O. P., & Malvela, M. (2003). Oxygen
uptake kinetics during free swimming: a pilot study. IX Biomechanics and
Medicine in Swimming, 379-384.
Seifert, L., Chollet, D., & Chatard, J. C. (2007). Kinematic changes during a 100-
m front crawl: effects of performance level and gender. Medicine and
Science in Sports and Exercise, 39(10), 1784-1793.

54
Shaw, G., Koivisto, A., Gerrard, D., & Burke, L. M. (2014). Nutrition
considerations for open-water swimming. International Journal of Sport
Nutrition and Exercise Metabolism, 24(4), 373-381.
Silva, A. J.; Reis, V. M.; Marinho, D.; Carneiro, A. L.; Novaes, J.; Aidar, F. J.
(2006). Economia de nado: factores determinantes e avaliação. Revista
Brasileira de Cineantropometria e Desempenho Humamo, 8(3), 93-99.
Silva, A. S., Azevedo, W. K. C. (2007). Comportamento Glicêmico em
Treinamento de Natação com carater Aeróbio e Anaeróbio. Journal of Physical
Education, 137 (s.n.): 26-32.
Sousa, A. C., Figueiredo, P., Oliveira, N. L., Oliveira, J., Silva, A. J., Keskinen, K.
L., & Fernandes, R. J. (2011). V˙ O2 Kinetics in 200-m Race-Pace Front Crawl
Swimming. International Journal of Sports Medicine, 32(10), 765-770.
Sousa, A. C., Vilas-Boas, J. P., & Fernandes, R. J. (2014). Kinetics and Metabolic
Contributions Whilst Swimming at 95, 100, and 105% of the Velocity
at. BioMed Research International, 2014.
Sousa, A., Figueiredo, P., Zamparo, P., Vilas-Boas, J. P., & Fernandes, R. J.
(2013). Anaerobic alactic energy assessment in middle distance
swimming. European Journal of Applied Physiology, 113(8), 2153-2158.
Swift, W. (2003). Australian Swimming Coaches and Teachers. In Gold Coast.
Queensland: Association Convention.
Sylow, L., Kleinert, M., Richter, E. A., & Jensen, T. E. (2016). Exercise-stimulated
glucose uptake — regulation and implications for glycaemic control.
Nature Reviews Endocrinology, 13, 133.
Termin, B., & Pendergast, D. R. (2000). Training using the stroke frequency-
velocity relationship to combine biomechanical and metabolic
paradigms. Journal of Swimming Research, 14.
Thevelein, X., Daly, D., & Persyn, U. (1984). Measurement of total energy use in
the evaluation of competitive swimmers. In: N. Bachl, L. Prakop, R. Suckert
(eds.). Current Topics in Sports Medicine, pp. 668-676.

55
Toussaint, H. M., & Beek, P. J. (1992). Biomechanics of competitive front crawl
swimming. Sports Medicine, 13(1), 8-24.
Toussaint, H. M., & Hollander, A. P. (1994). Energetics of competitive
swimming. Sports Medicine, 18(6), 384-405.
Toussaint, H. M., Carol, A., Kranenborg, H., & Truijens, M. J. (2006). Effect of
fatigue on stroking characteristics in an arms-only 100-m front-crawl
race. Medicine and Science in Sports and Exercise, 38(9), 1635-1642.
Treffene, R. J., Dickson, R., Craven, C., Osborne, C., Woodhead, K., & Hobbs,
K. (1980). Lactic acid accumulation during constant speed swimming at
controlled relative intensities. Journal of Sports Medicine and Physical
Fitness, 20(3), 244-254.
Vanheest, J. L., Mahoney, C. E., & Herr, L. (2004). Characteristics of elite open-
water swimmers. The Journal of Strength and Conditioning Research, 18(2),
302-305.
Vescovi, J. D., Rupf, R., Brown, T. D., & Marques, M. C. (2011). Physical
performance characteristics of high‐level female soccer players 12–21 years
of age. Scandinavian Journal of Medicine and Science in Sports, 21(5), 670-
678.
Vilas-Boas, J. P. (1989). Controlo do treino em Natação: considerações gerais,
rigor e operacionalidade dos métodos de avaliação. Comunicação
apresentada às Jornadas Técnicas Galaico-Durienses de Natação. Corunha,
Espanha.
Vilas-Boas, J. P. (1996). Speed fluctuations and energy cost of different
breaststroke techniques. Biomechanics and Medicine in Swimming VII, 7,
167-171.
Vilas-Boas, J. P. (2000). Aproximação biofísica ao desempenho e ao treino de
nadadores. Revista Paulista de Educação Física, 14(2), 107-117.

56
Vilas-Boas, J. P. (2010). The Leon Lewillie Memorial Lecture: Biomechanics and
Medicine in Swimming. Past, Present and Future. In PL Kjendlie, R. Stallman,
& J. Cabri (Eds.) Biomechanics and Medicine in Swimming XI, 12-19.
Vilas-Boas, J. P. (2013). O índice de braçada como indicador do nível de
adequação mecânica da técnica em natação. Kinesis, 6(1).
Vogt, P., Rüst, C. A., Rosemann, T., Lepers, R., & Knechtle, B. (2013). Analysis
of 10 km swimming performance of elite male and female open-water
swimmers. Springerplus, 2(1), 603.
Wakayoshi, K., Yoshida, T., Ikuta, Y., Mutoh, Y., & Miyashita, M. (1993).
Adaptations to six months of aerobic swim training. International Journal of
Sports Medicine, 14(07), 368-372.
Whipp, B. J. (1987). Dynamics of pulmonary gas exchange. Circulation, 76(6 Pt
2), VI18-28.
Whipp, B. J., & Ward, S. A. (1990). Physiological determinants of pulmonary gas
exchange kinetics during exercise. Medicine and Science in Sports and
Exercise, 22(1), 62-71.
Wilmore, J. H., Costill, D. L., & Kenney, W. L. (2008). Physiology of Sport and
Exercise (4th ed. Ed.). Champaign, IL: Human Kinetics.
Wilmore, J., & COSTILL, D. (2001). Physiology of sport. Kiev.: Olympiiskaya
literatura.
Xu, F., & Rhodes, E. C. (1999). Oxygen uptake kinetics during exercise. Sports
Medicine, 27, 313-327.
Zamparo, P., Bonifazi, M., Faina, M., Milan, A., Sardella, F., Schena, F., &
Capelli, C. (2005). Energy cost of swimming of elite long-distance
swimmers. European Journal of Applied Physiology, 94(5-6), 697-704.
Zamparo, P., Capelli, C., & Pendergast, D. (2011). Energetics of swimming: a
historical perspective. European Journal of Applied Physiology, 111(3), 367-
378.

57
Zamparo, P., Perini, R., Peano, C., & di Prampero, P. E. (2001). The self selected
speed of running in recreational long distance runners. International Journal of
Sports Medicine, 22(08), 598-604.
Zingg, M. A., Rüst, C. A., Rosemann, T., Lepers, R., & Knechtle, B. (2014).
Analysis of swimming performance in FINA World Cup long-distance open
water races. Extreme Physiology and Medicine, 3(1), 2.