Embed Size (px)
Citation preview

CARACTERIZAÇÃO DAS BOMBAS DE PRÓTONS VACUOLARES
E SEU PAPEL NAS RESPOSTAS ADAPTATIVAS DE PLANTAS
AO ESTRESSE HÍDRICO.
MICHELLE GUEDES CATUNDA
UNIVERSIDADE ESTADUAL DO NORTE FLUMINESE
DARCY RIBEIRO
CAMPOS DOS GOYTACAZES
JANEIRO DE 2008

CARACTERIZAÇÃO DAS BOMBAS DE PRÓTONS VACUOLARES
E SEU PAPEL NAS RESPOSTAS ADAPTATIVAS DE PLANTAS
AO ESTRESSE HÍDRICO.
MICHELLE GUEDES CATUNDA
“Tese apresentada ao Centro de
Biociências e Biotecnologia da Universidade
Estadual do Norte Fluminense Darcy Ribeiro,
como parte das exigências para obtenção do
título de Doutor em Biociências”.
Orientador: Arnoldo Rocha Façanha
CAMPOS DOS GOYTACAZES – RJ
JANEIRO DE 2008

ii
Dedico esta tese aos meus pais, meu irmão, meu esposo,
mas principalmente ao meu filho Daniel a quem ensino e
com quem aprendo a viver e a ser feliz a cada dia que
amanhece.

iii
AGRADECIMENTOS
Costumo dizer que minha vida é preenchida por seis grandes amores, aos quais
gostaria muito de agradecer...
• Deus, que me deu a vida e essa oportunidade maravilhosa de ser feliz;
• Meu filho, Daniel, que desde que chegou a esse mundão me ensinou o
real significado de duas nobres palavras: vida e felicidade!
• Meu esposo, grande amor e companheiro de todos os momentos;
• Minha mãe, exemplo de mulher, meu pai, o melhor pai do mundo, e
meu irmão, sempre o meu caçulinha.
Aos meus amores, o meu muito obrigada por cada minuto de amor,
atenção, dedicação e confiança... Sem vocês por perto, nada disso estaria
sendo concretizado agora!
Agradeço também ao meu orientador, Arnoldo Rocha Façanha pela
imensa paciência e por cada discurso apaixonado pela ciência... Prometo
que vou levar um pouquinho dessa paixão para sempre comigo!
À professora Anna Okorocova, ao professor Lev Okorocov e ao
professor Fábio Olivares pelos ensinamentos oportunos.
Ao pessoal do LQFPP, LMGV e LFBM, que sempre estiveram de
portas abertas para a utilização dos equipamentos necessários para o
desenvolvimento desse trabalho.
À UENF e à FAPERJ pelo suporte financeiro.
Aos meus amigos, que riram de mim e que riram comigo... Que graça
tem a vida sem amigos???
Inga, Rosivany e Rosane... Para sempre no meu coração!!
Ana Paula (cunhadona!!), Tati Felice, Nathália, Fabiana, Alena, Juares,
Daniel, Jorge André, Geórgia, Josimara, Flávia Emenegilda, ... Obrigada!!!
Ao pessoal do campo que me ajudou a cultivar e a colher tiririca, por
mais esquisito que isso lhes parecesse!!

iv
Ao pessoal do LBCT, Giovana, Bia e Márcia Adriana (ex-LBCT) pelo
carinho e pela força no preparo das amostras para microscopia!!
Ao amigo Noil pelo suporte no microscópio... Jamais teria alinhado
aquele “trem” sem a sua ajuda!!!
À Dona Maria pelo bolo de banana espetacular quando eu estava
grávida do Daniel!!!!!
Os meus alunos do CEDERJ por terem me permitido vivenciar a
experiência didática, e ao pessoal do CEDERJ, pólo Macaé, Aninha,
Giselli, Ada, Claudinha, Sílvio, Maurício, ..., obrigada pela força sempre!!
Enfim... A todos os que estiveram por perto, me acompanhando,
torcendo por mim ou mesmo rindo comigo, o meu super-hiper-mega muito
obrigada!!! Vocês fizeram dessa fase da minha vida uma passagem muito
mais agradável!

v
SUMÁRIO página
RESUMO........................................................................ vii
ABSTRACT.................................................................... viii
1. INTRODUÇÃO........................................................... 01
2. REVISÃO DE LITERATURA...................................... 03
2.1. H+-ATPase vacuolar........................................... 03
2.2. H+-PPase vacuolar............................................. 05
2.3. Estresse nas plantas.......................................... 09
2.3.1. O estresse hídrico...................................... 11
2.4. Tiririca (Cyperus rotundus)................................ 12
2.5. Cana-de-açúcar (Híbrido interespecífico de
Saccharum L.)................................................................
16
2.6. Milho (Zea mays).............................................. 17
3. OBJETIVO GERAL..................................................... 19
3.1. OBJETIVOS ESPECÍFICOS.............................. 19
4. MATERIAL E MÉTODOS........................................... 20
4.1 Plantas................................................................. 20
4.2 Preparação da fração microssomal..................... 21
4.3 Purificação das vesículas de tonoplasto e
membrana plasmática....................................................
22
4.4 Determinação das atividades PPásicas e
ATPásicas......................................................................
23
4.5 Monitoramento do gradiente de prótons ............. 23
4.6 Determinação da condutância estomática e da
transpiração....................................................................
23
4.7 Determinação da fluorescência da clorofila a...... 24
4.8 Determinação do teor de clorofila........................ 24
4.9 Microscopia eletrônica de transmissão e
imunocitoquímica............................................................
24
4.10 Microscopia eletrônica de varredura................. 25
5. RESULTADOS E DISCUSSÃO.................................. 26
5.1 Atividade hidrolítica e de transporte de H+ da P- 26

vi
ATPase, da V-ATPase e da PPase em plantas de C.
rotundus..........................................................................
5.2 Condutância estomática e da transpiração......... 41
5.3 Fluorescência da clorofila a................................ 42
5.4 Teores de clorofila............................................... 45
5.5 Microscopia eletrônica de varredura................... 46
5.6 Microscopia eletrônica de transmissão............... 48
5.7 Atividade hidrolítica e de transporte de H+ da P-
ATPase, da V-ATPase e da PPase em colmos de
Saccharum spp...............................................................
50
6. CONCLUSÕES.......................................................... 53
7. BIBLIOGRAFIA........................................................... 55

vii
RESUMO A Cyperus rotundus é uma espécie que possui alto vigor reprodutivo e
rusticidade, que expressam sua alta capacidade de sobreviver a
situações de estresse abiótico. O turgor celular é parcialmente
controlado pelas bombas de prótons vacuolares, que geram gradiente
de H+ e facilitam o transporte secundário de íons e água para dentro do
vacúolo. A análise das bombas de prótons de C. rotundus retiradas de
seu ambiente natural exibiram uma hidrólise de PPi de cerca de 30%
em relação a hidrólise do ATP, enquanto que o gradiente de prótons
gerado por ambas as bombas foram muito similares. Esses dados
sugerem que, em condição natural de campo, as bombas de prótons
vacuolares funcionam como em outras espécies vegetais e que a V-
ATPase é a principal bomba. Analizando o desenvolvimento dessa
planta daninha sob condição de estresse hídrico, observou-se que as
atividadedes hidrolíticas de ambas as bombas foram progressivamente
inibida com o estresse hídrico. Após o restabelecimento da irrigação, a
atividade tanto de hidrólise, quanto de transporte de H+ pela PPase é
estimulada, estímulo esse que diminui para níveis próximos ao controle
48h após o restabelecimento da irrigação. A atividade da V-ATPase foi
pouco alterada, sugerindo que a capacidade de manter o turgor da
célula vem do trabalho em conjunto das duas bombas vacuolares. A
PPase funciona como um sistema back-up para a ATPase, suprindo o
meio citossólico de Pi para síntese de ATP pela ATPase. Em cana-de-
açúcar, análise de data mining no banco de dados SUCEST sugere que
os genes que codificam para as bombas de prótons vacuolares são
expressos em altos níveis em tecido de casca e os dados apresentados
no presente trabalho confirmam a atividade alta das bombas de prótons
nesse tecido. Esses dados são discutidos em relação ao possível papel
dessas bombas no metabolismo da cana-de-açúcar.

viii
ABSTRACT Cyperus rotundus is a weed that has high reproductive vigor and
rusticity that express high capacity to cope with abiotic stress. The cell
turgor is partially controlled by vacuolar H+-pumps, which generate a H+-
gradient that energize the uptake of ions and water into the vacuole.
The analysis of the vacuolar H+-pumps of plants of Cyperus rotundus
harvested from their natural environment exhibited a PPi hydrolysis
about 30% of their ATP hydrolysis, while the H+-gradients coupled to
either PPi and ATP hydrolytic activities were quite similar. These data
suggest that, on field conditions, the vacuolar proton pumps function just
like other plants and the V-ATPase is the main proton pump. Analyzing
this weed development under drought stress the activity of both
tonoplast H+-pumps were progressively inhibited. After rewatering, the
H+-PPase activity exhibited a striking stimulation, but this activation
declines to the level of control plants 48h after rehydration. In contrast,
the V-ATPase activity was only barely changed, suggesting that the
vacuolar turgor capacity of C. rotundus could be regulated by both
proton pumps working together during drought stress. It is likely that H+-
PPase function as backup system on ATP synthesis. On sugar cane
study, analyses of data mining in SUCEST bank (sugarcane EST
Project) suggest that the genes encoding this vacuolar H+-pumps are
expressed in higher levels in stem bark and our results confirm and
extend the data mining analysis of SUCEST bank suggesting that the
proton pumps are induced in the active form in stem bark of sugarcane.
These data are discussed in relation to a possible role of these pumps
in the yet underestimated metabolism of the stem bark of sugarcane.

1
1 - INTRODUÇÃO
A população mundial está aumentando em uma taxa alarmante e
espera-se que alcance aproximadamente seis bilhões de pessoas até o
final do ano de 2050. Por outro lado, a produção de alimentos está
diminuindo devido ao efeito de vários estresses bióticos e abióticos.
Assim, minimizar estas perdas é uma grande área de interesse para
todas as nações que precisam lidar com as exigências crescentes do
alimento. A herbivoria, a competição por plantas invasoras, o frio, a
salinidade e a seca são, entre os estresses principais, aqueles que
prejudicam o crescimento e a produtividade de plantas (Mahajan e
Tuteja, 2005).
Dados divulgados pela Empresa Brasileira de Pesquisa
Agropecuária (EMBRAPA) relatam que dos 3,3 bilhões de hectares
considerados aptos para a agricultura no mundo, cerca de 28% estão
sujeitos a estresse hídrico por déficit de água, 22% a estresses
minerais e 12% a estresse por excesso de água. Com isso, pode-se
dizer que o desenvolvimento de uma agricultura sustentável em áreas
tropicais deve estar apoiado em estratégias de desenvolvimento de
plantas mais adaptadas às condições de estresse ambiental,
associadas a práticas de manejo de água e nutrientes que conduzam a
ganhos de eficiência e desempenho das culturas.
O estresse hídrico é, sem dúvida, o maior limitador da
produtividade de plantas cultivadas em todo o mundo (Bohnert et al.,
1995). A identificação e compreensão dos mecanismos de tolerância ao
estresse hídrico são de grande importância para o desenvolvimento de
novas cultivares (van Rensburg, 1994; Ingram e Bartels, 1996).
Vários estudos têm sido feitos com o objetivo de identificar
padrões fisiológicos que possam ser usados como critérios para a
seleção de resistência ao estresse hídrico (Blum et al., 1996; Dure et
al., 1989).
Diferente do que ocorre nos laboratórios, as plantas no campo
estão sujeitas a incidência de vários estresses simultâneos. De fato,

2
dados recentes de varredura de expressão gênica mostram que os
mecanismos de tolerância a esses estresses apresentam vários
componentes que se sobrepõem (Bohnert et al.., 2001). Nesse projeto
será estudada a ação da ATPase e da H+-PPase vacuolar nos
mecanismos de adaptação ao estresse hídrico. Embora cada enzima
seja específica na utilização de seus respectivos substratos (Rea, et al.,
1992), ambas catalisam a translocação eletrogênica de prótons do
citossol para o lúmem do vacúolo para gerar um gradiente
eletroquímico de prótons. Esta força próton motriz promove a energia
para o transporte secundário de vários íons e metabólitos através da
membrana (Rea et al., 1992; Sze et al., 1992). No tonoplasto, a V-
ATPase é a mais abundante bomba de prótons, sendo
extraordinariamente importante para manter a homeostase iônica e o
metabolismo celular (Lüttge e Ratajczak, 1997). A H+-PPase vacuolar é
uma bomba transportadora de prótons que têm suas atividade e/ou
expressão modulados durante os mais diversos estresses ambientais.
Rea e Poole (1993) citados por Maeshima (2000) indicaram a
importância desta enzima em células vegetais sob condições de
estresse energético, com isso, exerce função importante nas
estratégias de sobrevivência de plantas expostas a condições limitantes
de suprimento de ATP.
Plantas resistentes ao estresse hídrico são capazes de manter
seu turgor mesmo com potencial hídrico baixo, aumentando o número
de moléculas de solutos dentro da célula (Bray et al., 2000).
Para que as plantas sejam capazes de sobreviver as constantes
alterações do ambiente e ao mesmo tempo manter suas condições
metabólicas em um nível ótimo, a expressão e a atividade das bombas
de próntos deve ser muito bem reguladas (Gaxiola et al., 2007).
A planta daninha tiririca (Cyperus rotundus L.) e a cultivada
cana-de-açúcar (Saccharum sp) foram selecionadas para objeto de
estudo devido sua rusticidade e alta capacidade de proliferação.

3
2 - REVISÃO DE LITERATURA
Bombas de prótons são proteínas integrais da membrana que
funcionam a partir da hidrólise de moléculas de ATP, no caso das
ATPases, e PPi, no caso das PPases. Efetuam o transporte ativo dos
prótons através da membrana da célula e possuem como objetivos a
manutenção da composição iônica intracelular, a importação de solutos
contra o gradiente de concentração, o balanço da pressão osmótica dos
dois lados da membrana celular e também, a manutenção do potencial
da membrana celular.
As bombas podem funcionar contra o gradiente de concentração,
bombeando os prótons e, neste caso, consumindo energia. No entanto,
as ATPases podem também funcionar de forma reversa, sintetizando
ATP a partir de ADP e Pi, quando permitem a passagem dos prótons a
favor do gradiente de concentração.
Pedersen e Carafoli (1987) baseados em considerações de
estrutura e mecanismo de ação, agruparam as ATPases em 3 classes
principais: P, V e F. Essas ATPases estão distribuídas nas diversas
membranas celulares e sua função básica é a hidrólise ou síntese de
ATP e o transporte de íons. Além desses sistemas existe um quarto tipo
de bomba de prótons que foi posteriormente identificado, as H+-
pirofostatases (PPase).
Células vegetais são as únicas que contém vacúolos, que
ocupam grande parte de seu volume celular, e as principais enzimas,
presentes no tonoplasto, responsáveis pela acidificação do vacúolo
central são a V-ATPase e a PPase (Sze 1985, Taiz, 1992; Rea e Poole,
1993,). Essas 2 bombas de prótons fornecem o gradiente eletroquímico
de H+ que energiza o transporte de outros metabólitos (Rea e Sanders,
1987).
2.1 – H+-ATPase Vacuolar
A H+-ATPase vacuolar (V-ATPase) é a enzima responsável pela
acidificação do vacúolo central. Também está presente no Golgi e em
vários outros sistemas de endomembranas das células vegetais

4
(Ratajczak et al., 1999). A maior parte dos processos de transporte
secundário são impulsionados por um gradiente eletroquímico de
prótons gerado pela V-ATPase, incluindo reações de osmoregulação,
homeostase, armazenamento, defesa vegetal e muitas outras funções.
Ela exerce um importante papel no controle do pH extra e intra cellular
nas células eucarióticas, que é essencial para o bom funcionamento
dos processos celulares (Yao et al., 2007).
A V-ATPase de vegetais é estruturalmente muito semelhante a
de outros seres eucariotos e suas subunidades são codificadas por
uma família de múltiplos genes, que desempenha um importante papel
na regulação da expressão gênica e na separação das isoformas das
V-ATPase para as diferentes organelas (Taiz, 1992). É uma enzima
multimérica que exibe uma estrutura molecular complexa. Mais de 10
diferentes subunidades da V-ATPase já foram descritas para algumas
plantas (DuPont e Morrissey, 1992; Ward e Sze, 1992). Três dessas
subunidades, que foram encontradas em todas as V-ATPases
estudadas até o momento parecem ser mais importantes para o seu
funcionamento, são elas a subunidade calalítica A (~ 70kDa) e a
subunidade regulatória B (~ 60kDa), ambas localizadas na periferia da
membrana no domínio V1 e o canal de prótons hidrofóbico, formado
pela subunidade C (~ 16kDa) que formam o domínio V0, integral da
membrana (Rockel et al., 1998). No tonoplasto, a V-ATPase é a mais
abundante, atingindo 6,5 – 35% da proteína total do vacúolo de
diferentes espécies (Xiao et al., 2007). Sua estrutura pode ser
visualizada na figura 1.

5
Figura 1 - Modelo esquemático de uma V-ATPase de planta (Drory e
Nelson, 2006; Gaxiola et al., 2007)
Devido a sua função como uma bomba de H+ no tonoplasto, a V-
ATPase está envolvida no transporte de metabólitos para dentro do
vacúolo. Por outro lado, em condições de estresse ambiental, a V-
ATPase funciona como uma enzima de resposta a estresses sofrendo
modificações moderadas na expressão de suas subunidades e
modulação de estrutura (Dietz et al., 2001). Visto que está envolvida em
adaptações ecofisiológicas em níveis moleculares, a V-ATPase foi,
também denominada como uma “eco-enzima” (Lüttge et al., 1995, ).
Pouca atenção tem sido dada para a função da V-ATPase em
situação de estresse hídrico. Dietz et al.(2001) alertavam para a
necessidade de estudos da atividade e expressão da V-ATPase em
plantas submetidas ao déficit hídrico.
2.2 - H+-PPase Vacuolar
Uma característica própria de plantas é a existência de vias
metabólicas paralelas que usam tanto nucleotídeos quanto pirofosfato
como fontes alternativas de energia. Uma delas é a H+-ATPase, a qual
está presente em todos os eucarióticos e a outra é a Pirofosfatase
(PPase) que tem sido encontrada em todas as plantas e em algumas
bactérias fototróficas (Rea e Poole, 1993) e protozoários. Já está bem

6
estabelecido que a membrana vacuolar (tonoplasto) de células vegetais
possui 2 bombas de prótons, a H+-ATPase e a PPase. São sugeridas
duas hipóteses para a existência de duas bombas numa mesma
membrana: que a PPase agiria como um sistema back-up para a H+-
ATPase nas condições em que o fornecimento de ATP fosse limitado
ou que a PPase é reversível e pode usar o gradiente de H+ do trans-
tonoplasto para sintetizar PPi (Leigh et al., 1992).
A localização da PPase é ainda controvertida. Quando ela foi
primeiramente isolada em tonoplasto, acreditava-se que ela fosse uma
enzima marcadora de vacúolo (Maeshima, 1990), porém estudos
recentes têm mostrado que a PPase é encontrada em membrana
plasmática (Long et al. 1995; Robinson et al.. 1996), complexo de Golgi
e retículo endoplasmático.
Por algum tempo já se suspeitava da existência de PPase
associada à membrana. Foi em meados dos anos 70, que Karlsson
(1975) demonstrou uma atividade PPase estimulada por K+ e Walker e
Leigh (1981) caracterizaram uma PPase dependente de Mg2+. Somente
em 1985 é que finalmente identificaram que o transporte de H+
dependente de PPi era mediado por uma PPase e não uma V-ATPase
capaz de usar PPi em algumas circunstâncias. Seguidas
caracterizações mostraram que essa PPase diferia da V-ATPase em
vários aspectos: A PPase era estimulada pelo K+ enquanto que a V-
ATPase era seletivamente ativada por halógenos; nem a hidrolise nem
o transporte de prótons pela PPase são inibidos por nitrato; finalmente
foi mostrado que a PPase é específica para MgPPi (ou Mg2PPi) como
substrato enquanto que a V-ATPase não mostra nenhuma atividade
em relação a esse composto.
As atividades tanto de transporte de prótons dependente de PPi
quanto a de hidrólise estimulada por K+ e /ou insensível a molibidato
tem sido demonstrada em membranas vacuolares da maioria dos
principais tipos de plantas vasculares (plantas monocotiledôneas,
dicotiledôneas, C3, C4 e CAM) como também em seus prováveis
ancestrais, a algas clarófitas. Mais ainda, a PPase é o componente
principal das membranas vacuolares e é capaz de gerar um gradiente

7
de prótons de magnitude similar ou superior do que a V-ATPase
(Maeshima e Yoshida, 1989). A estimativa de abundância sugere que
essas enzimas constituem de 1% (em Beta vulgaris) e 5-10% (em Mung
bean) da proteína total da membrana vacuolar (Maeshima e Yoshida,
1989).
A membrana vacuolar preparada a partir de várias espécies de
plantas, incluindo musgos, samambáias, e algas, apresenta atividades
PPásicas em adição a V-ATPase (Nakanishi e Maeshima, 1998).
Notáveis exemplos de PPases transportadoras ou conservadoras de
energia são as PPases reversíveis transportadoras de prótons
encontradas em cromatóforos de bactérias não sulfurosas,
Rhodospirillum rubrum, as PPases transportadoras de Prótons
vacuolares de células vegetais e PPase associada a membrana de
animal e mitocondria de levedura.
Essa última, porém, apresenta padrões de sensibilidade a
inibidores bem diferentes das duas primeiras (Baykov et al., 1993).
Baseado nestes resultados considera-se que as PPases associadas a
mitocôndria pertencem a um grupo distinto de PPases (Baykov et al.,
1993).
A PPase está amplamente distribuída entre as plantas
superiores, algas e bactérias fotossintéticas, Rhodospirillum rubrum.
Embora Lichko e Okorokov (1984) tenham demonstrado uma atividade
de transporte de prótons dependente de PPi em frações enriquecidas
de tonoplasto de Saccharomyces carlsbergenesis, a H+-PPase é ainda
considerada restrita a plantas (Rea et al., 1992) e certos procariotos.
A PPase consiste de um único peptídeo com uma massa
molecular de aproximadamente 73KDa (Nakanishi e Maeshima, 1998),
embora tenham sido encontrados valores de pesos moleculares
diferentes em várias espécies, 67,000 (Beta vulgaris) e 73,000 (Vigna
radiata) para a mesma subunidade. Essa subunidade mostra-se não
somente necessária, mas também suficiente para o transporte de H+
dependente de PPi. A subunidade que se liga ao substrato funciona
como um monômero durante a hidrólise de PPi (Rea e Poole, 1993),
embora tenha sido proposto uma estrutura dimérica para a PPase

8
funcional nativa (Sato et al., 1991). A estrutura primária da PPase foi
deduzida a partir do cDNA de Arabdopsis thaliana, cevada, beterraba,
tabaco e arroz, no entanto, poucos resíduos característicos foram
determinados (Takasu et al.., 1997). A estrutura da PPase pode ser
visualizada na figura 2.
A abundância e a ubiqüidade da PPase em plantas necessita um
fornecimento regular de PPi citossólico e uma taxa de ação de massa
PPi:Pi em favor da translocação de prótons. O nível de PPi em tecidos
vegetais varia na faixa de 5-39 nmol/g de peso fresco (Smyth et al..,
1984) e esse PPi parece está quase que limitado ao citossol. Já foi
calculado em folhas de espinafre e protoplasto de mesófilo de trigo o
PPi citossólico de 200 a 300 µM (Weiner et al. , 1987), enquanto que
em outras organelas como vacúolo e cloroplasto de alga, Chara, foi
encontra <1 e 2-3µM respectivamente, contra 193µM para o citossol
(Takeshige e Takawa, 1989). Considerando uma estequiometria de H+:
PPi igual a 1, considera-se que a energia livre liberada pela hidrólise do
PPi citossólico exceda o mínimo teórico requerido para energização
vacuolar.
Figura 2 – Modelo de H+-PPase de Streptomyces coelicolor (Gaxiola et al.., 2007)

9
Gaxiola (2001) mostrou que a PPase é codificada por um único
gene e, provavelmente por esse motivo, é mais fácil de ser sintetizada
que a V-ATPase em condições de estresse metabólico.
2.3 - Estresse nas Plantas
O termo estresse é definido como qualquer fator físicoquímico ou
ambiental capaz de produzir uma tensão prejudicial ao organismo vivo
(Levitt, 1980) e a definição mais aceita para estresse ambiental é, uma
força adversa ou uma condição que inibe o funcionamento normal de
um sistema biológico como as plantas (Jones e Jones, 1989). Na
natureza o estresse, normalmente, não acontece isoladamente, mas
sim em conjunto com outros, assim, existem diversas vias para que as
plantas sejam capazes de sobreviver a uma situação de estresse
(Mahajan e Narendra, 2005).
As plantas são constantemente expostas a uma série de fatores
de estresses como, baixas temperaturas, salinidade, seca, alagamento,
calor, estresses oxidativos e toxidade por metais pesados, sendo que
os estresses abióticos são a principal causa da queda na produtividade
mundial, causando perdas de centenas de milhões de dólares por ano
(Bray et al.., 2000).
As plantas respondem ao estresse ambiental de maneira
complexa e dinâmica. Um número crescente de evidências mostra que
as membranas possuem um papel central na percepção do estresse,
respondendo ao estímulo ambiental (e hormonal) através da alteração
do bombeamento de íons e da conformação das proteínas ligadas a
membrana e suas atividades. As membranas participam diretamente ou
indiretamente, determinando a compartimentalização de precursores,
intermediários, produtos, e efetores de sistemas enzimáticos não
ligados à membrana, influenciando o pH. Em resposta a situações de
estresses, vários genes são regulados de modo a minimizar os efeitos
do estresse em questão (Shinozaki et al., 2003).
Amzallag e Lerner (1994) dividem a resposta ao estresse em 2
grupos principais: os de resposta rápida, que está relacionada a um
programa de defesa preexistente na planta e ou outro denominado

10
adaptação, onde o tempo influencia de maneira crucial na elaboração
dessa resposta. A adaptação é uma resposta prolongada durante a
qual as plantas ajustam sua fisiologia as condições ambientais numa
maneira orientada pelo meio.
Quando uma planta responde ao estresse através de sua
resistência (resposta pré-adaptativa) ela expressa um programa
preexistente que lhe permite sobreviver ao estresse mantendo (mais ou
menos) seu programa original de desenvolvimento, onde o estresse é
considerado como o gatilho da expressão genética.
A resistência reflete a capacidade da planta de expressar, sob
estresse, seu programa de desenvolvimento original. Em geral, a
expressão deste programa preexistente ocorre relativamente rápido, em
48h de exposição ao estresse. A reação não é específica ao estresse
particular e rapidamente estabiliza ao nível atual. A diminuição do
crescimento é proporcional a intensidade do estresse e inversamente
proporcional a tolerância (capacidade pré-adaptativa) da planta.
Durante a adaptação a planta estabelece um novo programa de
desenvolvimento como função das condições precisas de estresse. No
início do processo de adaptação existe uma considerável diminuição no
crescimento, que é comumente mais importante do que a
preadaptação. Uma vez a planta esteja adaptada, a velocidade de
crescimento aumenta, e pode atingir níveis similares a média de
velocidade de crescimento relativo (RGR – relative growth rate)
comparada as plantas controle de mesma idade. Um aumento da RGR
no final do processo de adaptação indica que o fim do estresse à
planta. A resposta adaptativa inclui modifições do balanço hormonal,
processos metabólicos e expressão de genoma. Em contraste a
resposta pré-adaptativa não é completamente pré-programada no
genoma.
Enquanto a resposta adaptativa não é programada, a
capacidade de responder ao estresse por adaptação é geneticamente
controlada. Nem toda espécie possui essa capacidade de adaptação.
Dentro de uma espécie capaz de se adaptar, nem todo o cultivar é

11
capaz . E mais ainda, dentro de uma espécie, nem todo o cultivar que
se adapta o faz com a mesma extensão (Amzallag e Lerner, 1994).
Quando ocorrem severas mudanças nas condições ambientais,
a célula pode responder de uma maneira específica e rápida
selecionando o aumento ou diminuição da expressão de genes
específicos. Os genes cuja expressão é aumentada durante o tempo de
estresse, são, presumivelmente críticos para a adaptação do organismo
à condição adversa; um exame dos genes ativados em resposta ao
estresse podem mostrar-se útil no entendimento da resposta biológica
de plantas em condições de estresse. Os sistemas genéticos que
respondem ao estresse são de interesse não somente devido ao seu
papel de apoiar a planta sob estresse, mas também por sua utilidade no
estudo dos eventos moleculares que controlam o nível quantitativo da
expressão gênica (Matters e Scandallos, 1986).
Nem toda resposta metabólica é considerada deletéria ou
prejudicial, e o maior desafio para os bioquímicos é distinguir as
respostas que representam um sintoma prejudicial e aquelas que são
verdadeiramente adaptativas, ou seja, que favorecem o crescimento
continuado durante a recuperação ao estresse. Na verdade para
qualquer variável no metabolismo deve existir uma faixa ótima, onde
abaixo dela a planta pode sofrer privação, e exceder essa faixa pode
levar a toxicidade (DeRocher e Bohnert, 1993).
2.3.1 - O estresse hídrico
O estresse hídrico pode ser causado por excesso ou por falta de
água. O estresse hídrico mais comum é o causado por falta de
disponibilidade de água para as plantas. A remoção de água das
membranas rompe a estrutura normal da bicamada lipídica e a
membrana plasmática fica extremamente porosa, podendo levar a um
desarranjo das proteínas presentes na membrana, o que contribui para
a perda de sua integridade e seletividade e também, na perda da
capacidade enzimática (Crowe et al., 1992; Hoekstra et al.., 2001;
Mahajan e Narendra, 2005). Além dos danos à membrana, o estresse
hídrico pode causar redução na atividade de proteínas citossólicas ou

12
de organelas. A alta concentração de eletrólitos durante a desidratação
pode causar rompimento no metabolismo celular (Liu e Zhu, 1998;
Yanqiong et al.., 2007).
Um outro efeito fisiológico causado pelo estresse hídrico é a
redução do crescimento vegetativo das plantas. A parte aérea é,
normalmente, mais sensível que as raízes e a redução da expansão da
parte aérea em situação de estresse hídrico é benéfica para a planta,
tendo em vista a redução na transpiração. Já o crescimento radicular
recebe um estímulo de crescimento, com o objetivo de explorar novas
áreas do solo a procura de água (Liu e Zhu, 1998).
O aumento da temperatura ou a rápida queda na umidade
resultam em uma condição de déficit hídrico para as plantas. A primeira
resposta de todas as plantas ao déficit hídrico é, em geral, o
fechamento dos estômatos, para evitar a perda de água por
transpiração (Mansfeld e Atkinson, 1990). O fechamento estomático
pode ser resultado da evaporação direta da água das células guarda
sem envolvimento metabólico, sendo conhecido como fechamento
hidropassivo, ou pode ser dependente do metabolismo, envolvendo
fluxo de íons que causam o fechamento estomático.
2.4 - Tiririca (Cyperus rotundus L.)
As plantas daninhas verdadeiras são, segundo Fisher (1973), as
que apresentam rusticidade e grande vigor vegetativo e reprodutivo,
tendo capacidade de sobreviver, crescer e reproduzir em condições
extremas de ambiente, como seca, encharcamento, altas e baixas
temperaturas, solos com problemas de salinidade, alcalinidade e
acidez. Além disso, são resistentes a pragas e doenças. De acordo com
Cardenas et al.. (1972), as principais características das plantas
daninhas são: ciclo de vida semelhante ao da cultura, plasticidade
populacional, germinação desuniforme (mecanismo de sobrevivência
em função da dormência), produção de inibidores (efeitos alelopáticos
ou teletóxidos) e produção de grande número de sementes (reprodução
sexuada) e/ou estruturas reprodutivas (reprodução assexuada).

13
Os danos causados pelas invasoras tendem a ser medido,
através do número de culturas que afetam, pela dificuldade de controle,
e pelo número de países em que ocorrem. A Cyperus rotundus, que
pode ser visualizada na figura 3, é considerada uma das espécie mais
nociva entre as plantas daninhas (Holm et al., 1977), sendo inclusive
citada no Guinnes Book (1994), como infestante de 52 importantes
culturas em 92 países. É uma planta altamente competitiva e de difícil
controle químico e/ou mecânico, ocorrendo com freqüência nas regiões
compreendidas entre as latitudes 30º Norte a 35º Sul (Junqueira Neto
et al.., 1982). No Brasil a Cyperus rotundus ocorre em toda extensão
territorial. É a invasora mais conhecida no país, possui ampla
distribuição, rápida disseminação, crescimento e desenvolvimento, alta
capacidade de competição e rusticidade. Pode ser ainda hospedeira
alternativa para fungos como e Fusarium spp, insetos, ácaros e para
diversas espécies de nematóides, além de causar efeitos alelopáticos,
produzindo substâncias que afetam a germinação, a brotação e o
desenvolvimento de outras espécies (Kissmann, 1991; Lorenzi, 1994).
A Cyperus rotundus é altamente beneficiada pelos cultivos
intensivos do solo, o que favorece a sua multiplicação e dispersão e,
além disso, é resistente a grande maioria dos herbicidas registrados
(Forster e Cerdeira, 1993; Sharma e Gupta, 2007). Em conseqüência
destas características e de sua rápida propagação vegetativa, esta
ciperácea passa a dominar áreas agrícolas com quase total
exclusividade.
A Cyperus rotundus é uma planta de metabolismo C4 (Mercado,
1979), e por este motivo é beneficiada por alta intensidade luminosa e
temperatura elevada (Holm et al., 1977) mantendo uma alta atividade
fotossintética nestas condições, com ausência aparente de
fotorespiração (Taiz e Zeiger, 1995; Guimarães, 1993). Essas
características fazem com que essa planta daninha se torne muito mais
competitiva nas regiões tropicais. Muitas culturas de importância
econômica são afetadas pela Cyperus rotundus, como a cana-de-
açúcar, o milho, o feijão, o arroz, a soja e hortaliças.

14
Na cana-de-açúcar, cerca de 1 milhão de hectares plantados
com esta cultura apresentam infestação de Cyperus rotundus, no Brasil
(Kissmann, 1991). Os prejuízos decorrem de competição, durante todo
o ciclo, mas especialmente é na fase inicial da cultura e nas reformas
que a invasora ocasiona maiores problemas. Pela exudação de
substâncias químicas de efeito alelopático a Cyperus rotundus inibe a
brotação de gemas e o perfilhamento da cana, o que resulta em
estandes muito baixos (Lorenzi, 1983; Durigan, 1991). Na Colômbia
Cruz et al. (1971), demonstraram que uma competição inicial por 10
dias reduziu em 10% a produtividade da cana-de-açúcar, enquanto a
competição por 30 dias reduziu em 30%. Experimentos conduzidos na
Argentina, em 1965, citados por Kissmann (1991), mostraram que em
casos extremos de infestação há uma queda de até 75% na colheita de
cana e uma redução de 65% na produção de açúcar.

15
Figura 3 – A Cyperus rotundus L.
Rede de tubérculos

16
2.5 - Cana-de-açúcar (híbrido interespecífico de Saccharum L.)
Originária do sudeste da Ásia, onde é cultivada desde épocas
remotas, a exploração canavieira assentou-se, no início, sobre a
espécie S. officinarum. O surgimento de várias doenças e de uma
tecnologia mais avançada exigiram a criação de novas variedades, as
quais foram obtidas pelo cruzamento da S. officinarum com as outras
quatro espécies do gênero Saccharum e, posteriormente, através de
recruzamentos com as ascendentes (Doorembos e Kassam, 1979)).
Os trabalhos de melhoramento persistem até os dias atuais e
conferem a todas as variedades em cultivo uma mistura das cinco
espécies originais e a existência de cultivares ou variedades híbridas.
A importância da cana-de-açúcar pode ser atribuída à sua
múltipla utilização, podendo ser empregada in natura, sob a forma de
forragem, para alimentação animal, ou como matéria prima para a
fabricação de rapadura, melado, aguardente, açúcar e álcool (Orlando
Filho, 1983) .
O Brasil encontra-se em primeiro lugar no ranking mundial da
produção de cana-de-açúcar (Saccharum spp.), atualmente com uma
área total cultuvada de aproximadamente 5.400.000 há e uma
produção anual da ordem de 400.000.000 de toneladas (cana colhida).
O estado de Alagoas com aproximadamente 7,7% da área colhida do
Brasil (Agrianual, 2007). Com essa produção a cultura da cana-de-
açúcar demonstra ser de grande importância para a economia do
Brasil.
As características dos cultivares influenciam a eficiência
fotossintética da cana, além das variações climáticas que prevalecem
durante o desenvolvimento da cultura. A fotossíntese é correlacionada
negativamente com a largura das folhas e positivamente com a sua
espessura. A temperatura e a disponibilidade de água no solo, dos
fatores climáticos, são os mais importantes para a produção de cana-
de-açúcar (Vitória Filho e Christoffoleti, 2004). Sendo a cana-de-açúcar
uma planta de metabolismo fotossintético C4, é considerada altamente
eficiente na conversão de energia radiante em energia química.

17
Para o aprimoramento do cultivo da cana-de-açúcar, existe a
necessidade de melhor entendimento sobre o metabolismo do processo
fotossintético. É necessário conhecer-se mais e melhor o sistema de
transporte e de acúmulo dos metabólicos, principalmente sacarose, o
mais valioso produto da cana-de-açúcar .
2.6 – Milho (Zea mays L.)
O milho é uma das mais importantes plantas comerciais com
origem nas Américas. Há indicações de que sua origem tenha sido no
México, América Central ou Sudoeste dos Estados Unidos. É uma das
culturas mais antigas do mundo, havendo provas, através de
escavações arqueológicas e geológicas, e através de medições por
desintegração radioativa, de que é cultivado há pelo menos 5.000 anos.
Logo depois do descobrimento da América, foi levado para a Europa,
onde era cultivado em jardins, até que seu valor alimentício tornou-se
conhecido.
A importância econômica do milho é caracterizada pelas
diversas formas de sua utilização, que vai desde a alimentação animal
até a indústria de alta tecnologia. Na realidade, o uso do milho em grão
como alimentação animal representa a maior parte do consumo desse
cereal, isto é, cerca de 70% no mundo. Nos Estados Unidos, cerca de
50% é destinado a esse fim, enquanto que no Brasil varia de 60 a 80%,
dependendo da fonte da estimativa e de ano para ano (Fanceli e
Dourado Neto, 2000)
O milho é uma planta característica de clima tropical, ou seja,
exige calor e umidade para produzir satisfatoriamente e proporcionar
rendimentos compensadores (Resende et al., 2000).
Em plantas de milho, o déficit hídrico afeta praticamente todos os
aspectos relacionados ao desenvolvimento das plantas, reduzindo a
área foliar, diminuindo a fotossíntese e afetando vários outros
processos, além de alterar o ambiente físico das culturas, por modificar
o balanço de energia do sistema (Bergamaschi, 1992).

18
As restrições causadas pela baixa disponibilidade de água do
solo ou pela alta demanda evaporativa ativam certos mecanismos
fisiológicos que permitem aos vegetais escapar ou tolerar essas
limitações climáticas, modificando seu crescimento e desenvolvimento,
e até mesmo atenuando as reduções na produção final.
O milho é cultivado em regiões cuja precipitação varia de 300 a
5.000 mm anuais, sendo que a quantidade de água consumida por uma
lavoura de milho durante o seu ciclo está em torno de 600 mm. Dois
dias de estresse hídrico no florescimento diminuem o rendimento em
mais de 20%, quatro a oito dias diminuem em mais de 50% (Magalhães
e Durães, 2006).
Segundo Kramer (1969), os efeitos causados pelo déficit hídrico
são devidos às modificações na anatomia, morfologia, fisiologia e
bioquímica das plantas.

19
3 - OBJETIVO GERAL
Caracterizar a nível celular, bioquímico e molecular as enzimas
transdutoras de energia em espécies vegetais que expressem
diferentes suscetibilidades ao estresse hídrico. A abordagem visa
atender a grande demanda de conhecimento básico da área e
desenvolver ferramentas de suporte a programas de geração de
variedades mais adaptadas e tolerantes a condições de estresses
abióticos (extremos de seca).
3.1 - OBJETIVOS ESPECÍFICOS
1. Gerar conhecimento científico básico sobre o funcionamento e
distribuição dos sistemas de transdução de energia da planta
daninha Cyperus rotundus, e da planta cultivada Saccharum spp.,
as quais apresentam impressionante tolerância a estresses
ambientais e das quais pouco se conhece sobre sua energética
celular, além de estabelecer comparação entre a planta daninha e a
cultivada Zea mays ;
2. Identificar padrões de distribuição, abundância e atividade das
bombas de prótons V-ATPase e PPase que possam ser usados
como indicadores para fenotipagem de plantas tolerantes ao
estresse hídrico.
3. Elucidar mecanismos envolvidos na ativação de vias alternativas do
metabolismo energético que utilizam pirofosfato ao invés de ATP,
durante a adaptação ao estresse hídrico em plantas visando o futuro
desenvolvimento de variedades adaptadas de culturas importantes.

20
4 - MATERIAL E MÉTODOS
O presente trabalho foi realizado em duas fases, a primeira com plantas
retiradas de uma área experimental do campo e a segunda com plantas
cultivadas em casa de vegetação e submetidas ao estresse hídrico.
As médias dos dados climatológicos obtidos pelo Setor de Irrigação e
Agrometeorologia da Universidade Estadual do Norte Fluminense na
estação evapotranspirométrica em Campos dos Goytacazes, RJ, entre
março de 2002 e dezembro de 2003 e março de 2004 e dezembro de
2005 , foram: precipitação, 620 mm; umidade relativa, 79,8 %;
temperatura máxima, 28,3ºC; temperatura mínima, 18,9 ºC e
temperatura média, 24,5 ºC.
A segunda fase foi realizada em casa de vegetação, também na
Unidade de Apoio à Pesquisa, no campus da Universidade Estadual do
Norte Fluminense Darcy Ribeiro – UENF, no município de Campos dos
Goytacazes, RJ.
4.1 – Plantas
Na primeira fase foram utilizadas plantas adultas de Cyperus
rotundus coletadas em campo aberto, na Unidade de Apoio à Pesquisa,
no campus da Universidade Estadual do Norte Fluminense Darcy
Ribeiro – UENF, no município de Campos dos Goytacazes, RJ, região,
segundo Andreazzi (1997), localizada ao Norte do Estado do Rio de
Janeiro a uma altitude de 13 m do nível médio do mar, tendo como
coordenadas geográficas 21º 45´15” de latitude Sul e 41º 19´28” de
longitude Oeste. O clima da região, conforme W. Koppen é classificado
como Awi, com temperatura média em torno de 24 ºC e precipitação
pluvial de 932 mm por ano.
Na segunda fase foram utilizados tubérculos de C. rotundus,
colmos de cana-de-açúcar e sementes de milho, que foram plantados
em vasos com capacidade para 8 litros, contendo uma mistura de
substrato e areia na proporção de 2 :1.
As características químicas da mistura de areia e substrato
utilizada são apresentadas no Quadro 1.

21
Quadro 1 - – Resultado da análise química do solo utilizado
Características Químicas
pH Ca2+ Mg2+ Al3+ Na+ K P
H2O cmolc.dm3 mg.dm3
7,2 2,2 2,3 0,0 0,26 1,518 18,70
Análise realizada no Laboratório de Solos da UFRRJ em
Campos dos Goytacazes, RJ.
As plantas cultivadas foram irrigadas diariamente e após 15 a 20
dias da germinação, foram selecionadas para os procedimentos
experimentais.
O monitorando do estresse hídrico foi feito por quantidade (%) de
água no solo através do peso fresco comparado ao peso seco de três
amostras de 25 cm3 de solo, e após o estresse atingir o nível de 70% de
ressecamento do solo, a irrigação foi re-estabelecida e mantida
constante por 5 dias (120h).
A parte vegetal utilizada das plantas de C. rotundus e de milho foi
o pseudocaule (como indicado na figura 3). Também foram utilizados
colmos de cana-de-açúcar (Saccharum spp.), de onde foram separadas
casca (parte mais externa) e miolo para fracionamento celular.
No local do experimento, as temperaturas máxima, mínima e
média do ar e a umidade relativa (UR) foram monitoradas, entre as 8h e
às 17h, por meio de um termohigrômetro modelo 450, Spectrum
Technologies. A umidade do solo foi monitorada por um sensor Spectrum
Technologies.
4.2 - Preparacão da fração microssomal
A metodologia aplicada permite que de uma só vez, partindo-se
de uma mesma amostra, as frações de vesículas de membrana
plasmática e tonoplasto sejam isoladas. A fração microssomal contendo
vesículas de membrana plasmática e de tonoplasto foram isoladas de
pseudocaule das plantas de C. rotundus e do colmo de Saccharum spp,
através de centrifugação diferencial, essencialmente como descrito por
Giannini e Briskin (1987), com algumas modificações. Foram cortadas e
pesadas cerca de 15 a 20g de tecido e estes foram então

22
homogeneizadas em meio tamponado, usando grau e pistilo. O tampão
de extração é composto de sacarose 250 mM, glicerol a 10 %, 5 mM
DTT, 5 mM EDTA, 0,4 % de PVP-40, 100 mM KCl, 0,3 % BSA, PMSF 1
mM, Benzamidina 1mM, Tris-HCl (pH 8,0) 100 mM, na relação peso de
tecido/volume de tampão de 1:2. As soluções usadas na preparação
foram mantidas no gelo e toda a manipulação foi realizada a 4°C. O
homogenato resultante foi filtrado através de quatro camadas de gaze e
submetido a centrifugação a 3.000 x g durante dez minutos para a
remoção de células não rompidas, e núcleos. O sobrenadante foi
submetido a nova centrifugação a 100.000 x g por 30 minutos. O
precipitado dessa centrifugação é solubilizado em solução tampão
contendo: glicerol a 10 %, DTT 5 mM, PMSF 1 mM, Benzamidina 1mM,
Tris-HCl 100 mM pH 7,6 e EDTA 5 mM.
4.3 - Purificação das vesículas de tonoplasto e membrana
plasmática
A purificação das vesículas de membrana plasmática, foi
realizada essencialmente como descrito por Serrano (1990). 1,5 mL da
fração microssomal foi aplicada sobre um gradiente descontínuo
bifásico de sacarose nas concentrações de 25/46 % p/p, contendo
ainda: Tris-HCl 100 mM pH 7,6, EDTA 5 mM, DTT 5mM, PMSF 1mM e
Benzamidina 1mM. O gradiente com a fração microssomal foi
submetido a uma centrifugação de 100.000 x g em um rotor SW40
(Beckman), durante 2 horas. Após a centrifugação a banda contendo as
vesículas de membrana plasmática purificadas localizava-se na
interface entre 30 e 46 %, enquanto as vesículas de tonoplasto situava-
se na interface de 10 e 25 % ou 10 e 30 %. As bandas foram coletadas
e congeladas em nitrogênio líquido e armazenadas a –70ºC até
utilização. A proteína total contida na preparação foi dosada pelo
método clássico descrito por Lowry et al.. (1951) e Bradford (1976).

23
4.4 - Determinação das atividades PPásicas e ATPásicas
As atividades PPásicas e ATPásica foram determinadas
colorimetricamente, segundo o método clássico descrito por Fiske e
Subbarrow (1925). A reação foi iniciada com a adição da proteína e
parada através da adição de ácido tricloroacético em baixas
temperaturas, para uma concentração final de 10 % (v/v). A
composição do meio de reação foi de Tris , 50 mM em pH 6,5 para
frações de membrana plasmática e 7,0 para frações de tonoplasto;
MgSO4, 3 mM, KCl 100 mM, ATP 1 mM para as ATPases e PPi 1 mM
para as PPases e 0,03mg.ml-1 de proteína. Como inibidores da
atividade ATPásica foram utilizados 0,2 mM de vanadato no caso de
vesículas de membrana plasmática (Bowman et. al., 1988) A hidrólise
de PPi foi aferida através de sua dependência por K+.
4.5 - Monitoramento do gradiente de prótons
O gradiente de próton foi monitorado pelo decréscimo da
fluorescência da sonda fluorescente metacromática, 9-amino-6-cloro-2-
metoxiacridina (ACMA), excitada com um feixe de λ 415 nm e a
emissão captada a 485 nm. O meio de reação era composto de 250mM
sacarose, 10 mM de Tris pH 6.5 ou 7,0, KCl 100 mM, ACMA 1,3 µM,
MgSO4 1 ou 3 mM e ATP 3 mM ou PPi 0,4mM e 50 µg de proteína. O
gradiente foi dissipado com NH4Cl 1,5 mM.
4.6 - Determinação da condutância estomática e da
transpiração
A taxa fotossintética líquida (A, µmol m-2 s-1), a condutância
estomática (gs, mol m-2 s-1) e a transpiração foram determinadas nas
plantas de C. rotundus mantidas sob tratamento, por meio do sistema
portátil de medição das trocas gasosas, modelo LI-6200 (LI-COR). Para
tanto, foi utilizada uma câmara de 0,25L com área de medição de
aproximadamente 7,5 cm2. Todas as medidas foram efetuadas entre 7h
e 14h com intervalos de 1h nos dias estabelecidos de estresse por
déficit hídrico, utilizando sempre a luz ambiente.

24
4.7 - Determinação da fluorescência da clorofila a
A variável da fluorescência emitida pela clorofila a foi
determinada, nas mesmas folhas que as trocas gasosas, por meio de
um fluorímetro de luz modulada modelo Mini-PAM (WALZ).
As folhas das plantas foram adaptadas ao escuro, com o uso de
pinças, por 30 minutos, para que os centros de reação, do PSII,
adquiram a condição de “abertos” (todos os aceitadores primários
oxidados) e a perda de calor seja mínima. A fluorescência inicial (F0) foi
obtida com luz modulada de baixa intensidade (< 0,1 µmol m-2 s-1) para
não induzir efeito na fluorescência variável. A fluorescência máxima
(Fm) foi determinada por um pulso de luz saturante de 0,3 s de duração,
com freqüência de 20000 Hz. A fluorescência variável (FV) foi
determinada pela diferença entre F0 e Fm. Esse pulso permite o
fechamento dos centros de reação do PSII. Com os valores de FV e Fm
foi obtida a relação FV/Fm.
4.8 - Determinação do teores de clorofila
Os teores de clorofila foram determinados a partir do início do
tratamento pelo estresse por déficit hídrico, por meio do medidor portátil
de clorofila modelo SPAD-502, Minolta, Japão.
4.9 - Microscopia Eletrônica de Transmissão e
Imunocitoquímica
As amostras de plantas submetidas ao estresse hídrico e após
restabelecimento da irrigação foram fixadas em solução de
glutaraldeído 2,5 % em tampão fosfato por 24 horas a temperatura
ambiente. Após a fixação primária, as amostras foram lavadas por três
vezes em tampão fosfato para remoção do glutaraldeído residual e
desidratadas em série etanólica crescente. Amostras destinadas a
microscopia eletrônica de transmissão (MET) foram pós-fixadas por 1
hora em solução de tetróxido de ósmio 1% em água, lavadas por três
vezes em tampão fosfato e só então sofreram desidratação em séria
etanólica crescente. Após a desidratação as amostras foram embebidas

25
em resina UNICRIL durante um período de 72h, sendo mantidas em
estufa para polimerização da resina. Para visualização do material
antes de ser levado ao MET foram obtidas seções semi-finas (0,7 - 1
µm) utilizando um ultramicrótomo Reichert-Jung Ultracut S (Leica) e
facas de vidro preparadas em um Knife maker II Reichert-Junga partir
dos blocos obtidos. Seções semi-finas coradas com azul de toluidina
solução 0,1% foram examinadas em um microscópio ótico Axioplan
(Zeiss). Seções ultra-finas para observação da imunocitoquímica ao
MET foram recolhidas em grades de níquel e incubadas com anticorpo
anti-PPase, subsequentemente contrastadas em acetato de uranila
solução 5% em água por 20 minutos e citrato de chumbo por 5 a 7
minutos e então examinados em um microscópio eletrônico de
transmissão Zeiss modelo EM 900 (Carls Zeiss).
4.10 - Microscopia Eletrônica de Varredura
Amostras de raízes e mesocótilos fixadas em glutaraldeído e
tetróxido de ósmio da mesma forma como descrito para amostras
preparadas para MET, foram lavadas por três vezes em tampão fosfato
e desidratadas em série etanólica crescente. Em seguida, amostras
selecionadas foram transferidas para o equipamento Critical Point
Drying Apparatus (Mod CPD 030, Bal-tec), e após secagem, foram
metalizadas utilizando o Automatic Sputter Coater SCD 050, Bal-tec.
Finalizado este processo, as amostras estavam prontas para
observação no microscópio eletrônico de varredura DSEM 962 (Zeiss).

26
5 - RESULTADOS E DISCUSSÃO
5.1 - Atividade hidrolítica e de transporte de H+ da P-ATPase,
da V-ATPase e da H+-PPase em plantas de C. rotundus
O trabalho teve início com a caracterização das atividades das
bombas de prótons de membrana plasmática e de vacúolo da planta
daninha C. rotundus, crescidas em condições de campo.
A Figura 4 mostra os resultados de atividade hidrolítica e a
Figura 5 de transporte de H+ pelas bombas P-ATPase, V-ATPase e
PPase isoladas de plantas crescidas no campo, ou seja, os resultados
encontrados relatam a atividade dessas bombas em condições de
campo para tal espécie vegetal.
Figura 4 - Atividade hidrolítica das bombas de prótons em vesículas
enriquecidas de membrana plasmática e de vacúolo de
plantas de C. rotundus crescidas em condições de campo
(média representativa de cinco experimentos).
P-ATPase V-ATPase V-PPase0.0
0.5
1.0
1.5
2.0
2.5
3.0
Ati
vid
ade
µµ µµm
ol.m
g-1
.min
-1

27
Figura 5 – Transporte de prótons pelas bombas ATP/PPi dependente
de vesículas de membrana plasmática e tonoplasto de
plantas em condições de campo. P-ATPase (a), V-ATPase
(b) e PPase (c). Fluorescência do ACMA na presença de 50
µg de proteína de membrana.
Os experimentos mostraram que a atividade da V-ATPase de
membranas enriquecidas de tonoplasto na C. rotundus, foi, em média
de 1,6 µmol.mg ptn-1.min-1 e a atividade da PPase, de 0,5 µmol.mg ptn-
1.min-1 (Figura 4), apresentando, portanto valores superiores aos
encontrados na literatura para Vigna unguiculata (Otoch et al., 2000) e
para Suaeda salsa (Han et al., 2005), porém, o balanço na atividade
entre as duas bombas de prótons vacuolares está coerente com o
padrão estabelecido para a maioria das preparações de tecido vegetal,
em que a V-ATPase, que é a principal bomba vacuolar, podendo gerar
um gradiente de H+ através da membrana vacuolar de magnitude
parecida ou maior que PPase (Perotti et al.,1994; Giannini e Briskin,
1987; Rea e Sanders, 1987), como é possível observar na figura 5.
Em condições ótimas para a planta, a atividade da V-ATPase
supera a da PPase (Nakanishi e Maeshima, 1998). Segundo Maeshima
(2000), uma maior quantidade da enzima PPase está presente em
0 200 400 600 8000
20
40
60
80
100
120
a
bc
Tempo (segundos)
Flu
ore
scên
cia
(%)

28
tecidos jovens, ou em condições de estresse energético. Esta fase do
experimento foi realizada com plantas adultas não estressadas. Tendo
o conhecimento de que a C. rotundus possui característica de
tolerância a condições adversas do ambiente, esperava-se encontrar
um padrão diferenciado do descrito para outras plantas mais sensíveis
a alterações ambientais, com uma atividade hidrolítica e de transporte
pela PPase mais alto que a da V-ATPase, mostrando um padrão
diferenciado da C. rotundus, porém, os valores encontrados para as
atividades das bombas de prótons PPase e V-ATPase da C. rotundus
seguem o padrão descrito na literatura para outras plantas.
Quando o assunto é estresse abiótico, muitos grupos, com
muitos resultados contraditórios, têm estudado os efeitos do estresse
salino nas bombas de prótons vacuolares (V-ATPase e PPase). Otoch
et al. (2001) mostraram que a atividade de transporte dependente de
PPi sofre uma redução na germinação de sementes de Vigna
unguiculata crescidas sob condição de estresse salino. Qiu et al. (2007)
mostraram que em vesículas enriquecidas de tonoplasto de S. salsa a
atividade hidrolítica e de transporte de prótons da V-ATPase
aumentaram quando as plantas foram submetidas ao tratamento com
estresse salino em comparação com plantas controle. Russak e Klobus
(2007) mostraram que plantas submetidas ao estresse salino
apresentaram aumento na atividade da V-ATPase. Porém, estudos de
energética associados ao estresse hídrico estão limitados a poucos
grupos.
Com o objetivo de buscar características diferenciadas na
bioenergética da C. rotundus quando submetidas à situação de
estresse por déficit de água, realizamos experimentos em casa de
vegetação.
Os resultados apresentados a seguir mostram o comportamento
das bombas de prótons de membrana plasmática e vacuolares da C.
rotundus sob condição de déficit hídrico.
A figura 6 mostra a atividade de hidrólise da bomba de prótons
de membrana plasmática P-ATPase. Uma importante característica da

29
P-ATPase é que ela funciona como um indicador do funcionamento
celular e sua atividade é afetada por diversos fatores ambientais
(Serrano, 1990; Palmgren, 1998). Sendo assim, é possível detectar a
presença de estresse em nível celular através do funcionamento da P-
ATPase.
Observou-se que durante o estresse hídrico a atividade
hidrolítica dessa bomba cai, chegando a aproximadamente 60% de
inibição em relação ao controle, indicado no eixo 0 (zero) da figura 7, e
após 24h do restabelecimento da irrigação a atividade retorna a níveis
próximos ao do controle, indicando o final do período de estresse.
controle dh.30% dh.50% dh.70% ri.10min ri.24h ri.48h ri.120h0
1
2
3
hidr
ólis
e A
TP
µµ µµm
ol.m
g-1
.min
-1
Figura 6 – Atividade hidrolítica da bomba de próton P-ATPase em
vesículas enriquecidas de membrana plasmática de plantas
de C. rotundus submetidas a déficit de água (dh) e após o
restabelecimento da irrigação (ri) (média representativa de
três experimentos).

30
dh.30% dh.50% dh.70% ri.10min ri.24h ri.48h ri.120h
-60
-40
-20
0
20
40
60
inib
ição
/est
ímu
lo (
%)
Figura 7 - Porcentagem de inibição ou estímulo da atividade hidrolítica
(µmol.mg-1.min-1) da bomba de prótons P-ATPase dos
tratamentos em relação ao controle.
Os resultados que seguem (figuras 8, 9, 10, 11 e 12) mostram a
atividade hidrolítica das bombas V-ATPase e PPase, principal foco do
trabalho apresentado. Observou-se que ambas bombas sofreram
inibição da atividade de hidrólise conforme o aumento do estresse
hídrico, indicado pelo aumento na porcentagem de ressecamento do
solo.
O resultado apresentado na figura 12 indica claramente que a V-
ATPase é a bomba de prótons mais inibida quando comparada a
PPase. Esse resultado sugere um déficit no suprimento de ATP,
levando a uma redução na quantidade de substrato para ser hidrolisado
pela V-ATPase. Corbineau et al. (2004), mostraram que o estresse
hídrico leva a um déficit energético avaliado pelos baixos níveis de
ATP, ADP e NTP, nucleotídeos responsáveis pelo armazenamento de
energia em suas ligações químicas para utilização nos diversos
processos celulares.
O suprimento de PPi, que é subproduto de muitos processos
metabólicos, parece ter sido menos afetado, já que a atividade da
PPase sofreu uma inibição cerca de 40% menor que a V-ATPase
quando comparadas ao controle (figura 12).

31
controle dh.30% dh.50% dh.70% ri.10min ri.24h ri.48h ri.120h0
1
2
hidr
ólis
e A
TP
µµ µµm
ol.m
g-1
.min
-1
Figura 8 – Atividade hidrolítica da bomba de próton V-ATPase em
vesículas enriquecidas de tonoplasto de plantas de C.
rotundus submetidas a déficit de água (dh) e após o
restabelecimento da irrigação (ri) (média representativa de
três experimentos).
dh.30% dh.50% dh.70% ri.10min ri.24h ri.48h ri.120h
-100
-60
-20
20
60
100
inib
ição
/est
imu
laçã
o (
%)
Figura 9 - Porcentagem de inibição ou estímulo da atividade
hidrolítica (µmol.mg-1.min-1) da bomba de prótons V-
ATPase dos tratamentos em relação ao controle.

32
controle dh.30% dh.50% dh.70% ri.10min ri.24h ri.48h ri.120h0
1
2
hid
rólis
e P
Pi
mm
ol.m
g-1
.min
-1
Figura 10 – Atividade hidrolítica das bomba de próton PPase em
vesículas enriquecidas de tonoplasto de plantas de C.
rotundus submetidas a déficit de água (dh) e após o
restabelecimento da irrigação (ri) (média representativa de
três experimentos).
dh.30% dh.50% dh.70% ri.10min ri.24h ri.48h ri.120h
-120
-80
-40
0
40
80
120
inib
ição
/est
imu
laçã
o(%
)
Figura 11 - Porcentagem de inibição ou estímulo da atividade hidrolítica
(µmol.mg-1.min-1) da bomba de prótons PPase dos
tratamentos em relação ao controle.

33
dh. 30% dh. 70% ri. 24h ri. 120h
-100
-50
0
50
100V-ATPasePPase
inib
ição
/est
imu
laçã
o (
%)
Figura 12 – Comparação entre a porcentagem de inibição ou estímulo
da atividade hidrolítica (µmol.mg-1.min-1) das bomba de
prótons V-ATPase e PPase dos tratamentos em relação ao
controle.
24h após o restabelecimento da irrigação, é possível observar
um incremento de aproximadamente 20% da atividade da V-ATPase e
de 100% da PPase, quando comparadas ao controle (figura 12), o que
sugere que a capacidade de recuperação da PPase é muito superior
que a da V-ATPase.
A explicação para os resultados apresentados pode ter como
base o fato da enzima PPase ser um polipeptídeo único, codificada por
um único gene (Sarafian et al.., 1992), sendo assim, sua resposta de
expressão após o término do estresse com a finalidade de restabelecer
o metabolismo celular, torna-se mais rápida. Além disso, também é
possível discutir a partir do fato de que após o término do estresse
hídrico, que provocou um estresse energético com conseqüente
depleção nos níveis de ATP, o substrato mais abundante presente no
citossol é o PPi, então, 24h após o restabelecimento da irrigação, a
atividade PPásica superou a ATPásica.
A maior atividade PPásica acarreta na hidrólise do pirofosfato
(PPi) e aumenta a produção de fosfato inorgânico, que, juntamente com
o ADP, é substrato para formação de ATP. Na figura 12 é possível

34
observar que aos 5 dias após o restabelecimento da irrigação (120h) a
atividade da V-ATPase encontra-se em níveis semelhantes ao controle
e a da PPase é reduzida a um nível pouco maior que o controle. Nesta
situação, a planta, provavelmente, está retomando sua atividade
energética normal, onde o suprimento de ATP aos poucos está sendo
normalizado. É possível que após mais alguns dias a atividade da V-
ATPase seja recuperada e volte a ser prioritária em relação a PPase.
Na figura 13, é possível observar o comportamento das plantas
de C. rotundus no período do estresse hídrico e após o
restabelecimento da irrigação. É possível observar que mesmo após
cinco dias sem irrigação, as folhas da planta se mantêm túrgidas e os
sintomas de déficit hídrico aparecem apenas aos 11 dias sem irrigação,
onde é possível observar sintomas de clorose e murcha. Após o
restabelecimento da irrigação, a recuperação da planta é bastante
rápida, onde após 24h do restabelecimento da irrigação as folhas
retomam seu aspecto de turgescência e os sintomas da desidratação
sofrida podem ser observados apenas em algumas folhas.
Em 1971 e 1977, Gaff descreveu um grupo de plantas que
chamou de ressurection plants, que são um grupo de plantas que
possuem a habilidade de tolerar longos períodos de dessecação e
“ressuscitar” após o restabelecimento da irrigação. Plantas com tais
características podem utilizar diferentes mecanismos fisiológicos e
bioquímicos de adaptação à dessecação e de retorno ao padrão
metabólico normal após a re-hidratação (Bewley e Krochko, 1982;
Navari-Izzo e Rascio, 1999).
No decorrer do experimento foi possível observar que cerca de 3
dias após o restabelecimento da irrigação, novas brotações foram
observadas, provavelmente como uma estratégia da espécie em se
reproduzir enquanto as condições do ambiente permitem (dados não
apresentados).
A C. rotundus não foi descrita como uma ressurection plant,
porém, suas características de recuperação de seu estado metabólico

35
após a re-hidratação, como pode ser visualizado na figura 13, sugerem
características próximas a essas plantas.
Figura 13 – Plantas de C. rotundus. Controle (A), 5º dia de estresse
hídrico (B), 11º dia de estresse hídrico (C) e 24h após o
restabelecimento da irrigação (D).

36
Para facilitar a comparação com uma planta cultivada de
metabolismo C4, experimentos com milho (Zea mays) foram conduzidos
nas mesmas condições do experimento com a planta daninha C.
rotundus. Os dados comparativos para a atividade hidrolítica das
bombas de prótons vacuolares (V-ATPase e PPase) podem ser
observados na tabela 1.
Tabela 1 – Comparação entre a atividade hidrolítica das bombas de
prótons vacuolares de C. rotundus e Z. mays em situação de déficit hídrico (dh) e após o restabelecimento da irrigação (re-irri). Valores representativos (± SD) de três experimentos.
controle dh 30% dh 70% re-irri 24h
V-ATPase Hidrólise ATP (µµµµmol.mg-1.min-1)
C. rotundus 1,77 ±0,15 1,16 ±0,18 0,54 ±0,03 1,93 ±0,08
Zea mays 0,14 ±0,04 0,09 ±0,04 0,05 ±0,03 0,13 ±0,05
V-PPase Hidrólise PPi (µµµµmol.mg-1.min-1)
C. rotundus 0,92 ±0,07 0,93 ±0,07 0,63 ±0,03 1,95 ±0,05
Zea mays 0,1 ±,050 0,05 ±0,02 0,03 ±0,02 0,09 ±0,03
Os dados apresentados na tabela 1 mostram a diferença
metabólica existente entre a planta daninha e a cultivada, onde os
valores de atividade hidrolítica obtidos em preparações de plantas
controle, plantas sob déficit hídrico e plantas re-irrigadas de C. rotundus
superam os de Z. mays, sugerindo uma energética diferenciada, o que
sustentaria a condição de rusticidade da C. rotundus.
Nas figuras 14 e 15 é possível observar a atividade de transporte
de prótons pelas bombas vacuolares.
Normalmente o funcionamento de uma bomba de prótons é
avaliado por sua capacidade de hidrolisar substrato acoplado a
capacidade de transportar prótons e com isso favorecer o transporte
secundário.

37
Figura 14 – Bomba de próton ATP dependente de vesículas de
tonoplasto de plantas irrigadas diariamente (a), 30% dh (b),
70% dh (c) e 24h após restabelecimento da irrigação (d).
Fluorescência do ACMA na presença de vesículas de
tonoplasto (50 µg de proteína de membrana).
Figura 15 – Bomba de próton PPi dependente e permeabilidade da
membrana de vesículas de tonoplasto de plantas irrigadas
diariamente (a), 30% dh (b), 70% dh (c) e após
restabelecimento da irrigação (d). Fluorescência do ACMA
na presença de vesículas de tonoplasto (50 µg de proteína
de membrana).
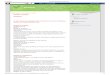
38
Figura 16 – Porcentagem de inibição ou estímulo da velocidade inicial
do transporte de prótons pelas bombas de próton V-ATPase
e PPase da membrana de vesículas de tonoplasto dos
tratamentos em relação ao controle.
Com os resultados apresentados nas figuras 14, 15 e 16, é
possível observar que a atividade de transporte de prótons da V-
ATPase parece estar em uma situação desacoplada de sua atividade
hidrolítica, pois sua atividade de transporte, mesmo com plantas sob
déficit hídrico de 70% apresentou-se maior que a da PPase, enquanto
que sua atividade hidrolítica foi 40% menor que a da PPase.
Matsuura-Endo et al. (1992) apresentaram dados em que o
tratamento com estresse por baixas temperaturas afetou a atividade
hidrolítica e de transporte da V-ATPase; Darley et al. (1995) mostraram
que sob condição de estresse por baixas temperaturas a V-ATPase
não foi afetada em plântulas de Mung bean e que no caso apresentado
a principal bomba de prótons foi a PPase; Colombo e Cerana (1993)
mostraram um incremento na atividade PPásica em suspensão de
células de cenoura sob tratamento com NaCl e Rea e Poole (1993)
mostraram a importância dessa bomba em células de plantas sob
condição de anoxia e estresse por baixas temperaturas. As diferenças
encontradas nos resultados dos trabalhos supracitados são atribuídas
5º dia 11º dia re-irrigada
-100
-80
-60
-40
-20
0
20
40
60
80
100V-ATPasePPase
% e
stím
ulo
/in
ibiç
ãoV
0

39
ao fato de que as condições de crescimento para as diversas plantas
submetidas aos estresses variados são diferentes, favorecendo, a
variabilidade dos fenômenos celulares.
Os resultados seguintes mostram que a bomba de prótons V-
ATPase apresentou-se pouco menos inibida, em relação a PPase
(figuras 17 e 18) quando comparada aos resultados citados
anteriomente, o que pode ser atribuído ao fato dos experimentos terem
sido conduzidos em diferentes épocas do ano, porém, este fato não
comprometeu as conclusões do trabalho.
Como a H+-PPase vacuolar é a principal enzima capaz de
hidrolisar PPi presente nas membranas do vacúolo, o estímulo
observado deve estar relacionado com a ativação desta enzima
induzida pelo re-estabelecimento da irrigação, sustentando a hipótese
de que existe um possível acoplamento entre as duas bombas de
prótons vacuolares, onde o gradiente eletroquímico gerado pela H+-
PPase energizaria a reversão do ciclo catalítico da H+-ATPase,
favorecendo a síntese de ATP (Façanha & de Meis, 1998). Assim, a
indução deste sistema regenerador de ATP durante o re-
estabelecimento da irrigação pode ser parte da resposta adaptativa da
planta ao estresse hídrico a que foi submetida.
Importante ressaltar que os dados apresentados mostram que as
duas bombas de prótons vacuolares trabalham em conjunto para
manter os processos fisiológicos da planta e superar o estresse a que
foi submetida.

40
Figura 17 - Atividade hidrolítica das bombas de prótons em vesículas
enriquecidas de membrana plasmática e de vacúolo de
plantas de C. rotundus submetidas a estresse hídrico (média
representativa de três experimentos).
5º dia 11º dia re-irrigada
-100
-80
-60
-40
-20
0
20
40
60
80
100V-ATPasePPase
inib
ição
/est
imu
laçã
o (
%)
Figura 18 - Porcentagem de inibição ou estímulo da atividade hidrolítica
(µmol.mg-1.min-1) das bombas de prótons V-ATPase e PPase
dos tratamentos em relação ao controle.
P-ATPase V-ATPase PPase0.0
0.5
1.0
1.5
2.0
2.5controle5º dia11º diare-irrigada
Ati
vid
ade
µµ µµm
ol.m
g-1
.min
-1

41
No presente trabalho, os resultados encontrados sugerem que
as plantas de C. rotundus possuem um metabolismo adaptado para
resistência ao estresse por déficit de água, pois mesmo após 11 dias
sem irrigação, e apesar das plantas apresentarem sintomas de clorose
severa e murcha, suas bombas de prótons vacuolares ainda possuíam
a capacidade de hidrolisar os substratos ATP e PPi, possivelmente com
a finalidade de manter as atividades celulares das plantas. Porém,
pode-se observar que houve um decréscimo na atividade de ambas as
bombas com o passar dos dias, sendo que a PPase foi a enzima que
sofreu maior inibição, quando comparada a V-ATPase. Esses
resultados corroboram com dados de literatura que apresentam a V-
ATPase como uma enzima extraordinariamente importante para manter
a homeostase iônica e o metabolismo celular. Lüttge et al. (1995)
demonstraram que em condições de estresse ambiental a V-ATPase
funciona como uma enzima de resposta de estresse, sofrendo
pequenas modificações na expressão de subunidades. Observamos
também que após o restabelecimento da irrigação a porcentagem de
estímulo em relação ao controle da atividade hidrolítica da PPase
superou em aproximadamente 80% a da V-ATPase, o que nos sugere a
rápida recuperação dessa enzima objetivando a recuperação do
metabolismo da planta.
5.2 - Determinação da condutância estomática e da
transpiração
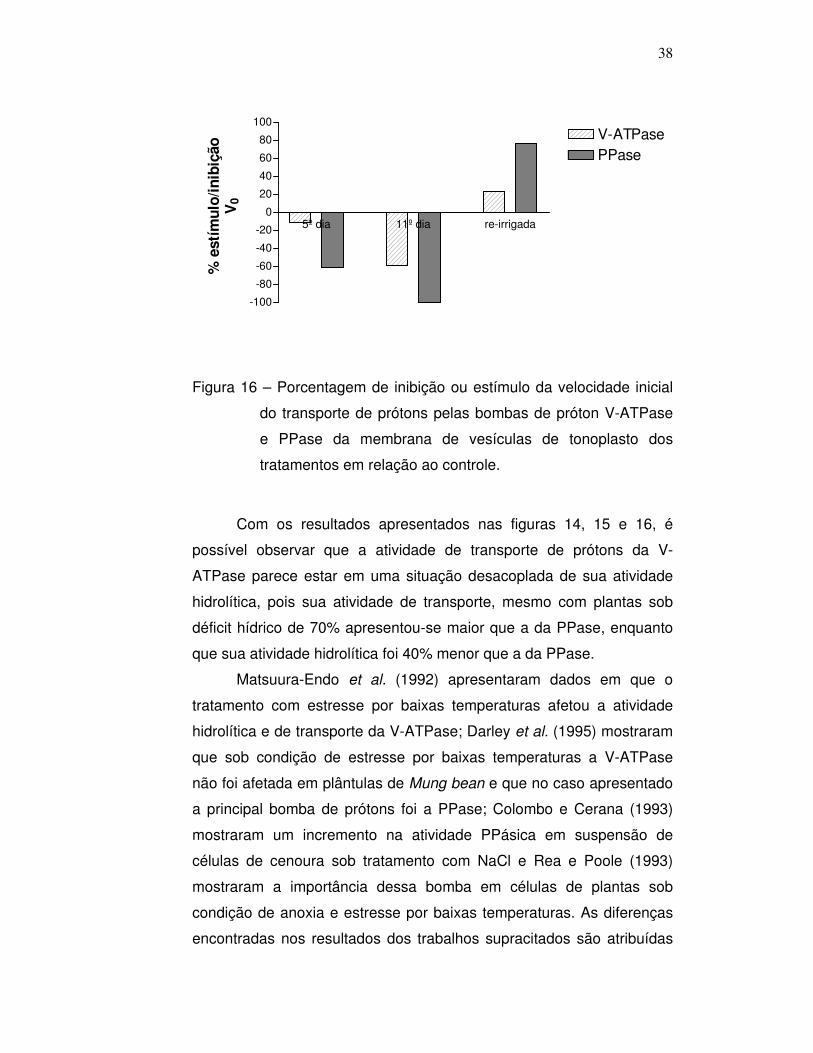
Nas figuras 19, 20 e 21 é possível observar os resultados da
taxa fotossintética líquida, transpiração e condutância estomática das
plantas submetidas ao estresse hídrico quando comparado às plantas
controle. É possível observar que as plantas sob estresse hídrico
possuem taxa de transpiração e condutância estomática mais baixas
que as plantas controle. Esse resultado vai de encontro aos dados
encontrados na literatura, onde em situação de déficit hídrico as plantas
fecham os estômatos. Esse fechamento estomático evita a transpiração
excessiva, prevenindo a perda de água pela planta, porém,
compromete a fotossíntese, uma vez que com os estômatos fechados,

42
as plantas não absorvem o CO2 da atmosfera, substrato para
ocorrência da fotossíntese. Aparentemente, a C. rotundus segue o
mesmo padrão das mais diversas espécies vegetais citadas na
literatura, onde sob déficit hídrico a condutância estomática decresce,
assim como a assimilação de carbono e conseqüentemente, a
fotossíntese. A visualização desse fechamento estomático foi possível
utilizando a técnica de microscopia de varredura, que será apresentada
no seguimento do trabalho.
5.3 - Determinação da fluorescência da clorofila a
A fluorescência da clorofila a indica a quantidade de energia que
não é aproveitada pela planta para a realização dos processos
fotoquímicos. Nos dados apresentados na figura 22, pode-se observar
que as diferenças entre as plantas controle e as submetidas ao
estresse somente apareceram no 11º dia, e os valores apresentados
são pouco menores que a faixa considerada normal para plantas não
submetidas ao estresse. Segundo Jakl e Bolhàr-Nordenkampf (1991),
a eficiência quântica (Fv/Fm) pode variar em uma faixa de 0,75 a 0,85
em plantas não submetidas a estresses. A diminuição da relação Fv/Fm
pode ser usada para estudar o efeito de estresse causado pela
aplicação de herbicidas (Eullaffroy, 2003).
Esses resultados sugerem a resistência fisiológica dessa planta
daninha ao estresse ambiental a que foi submetida.
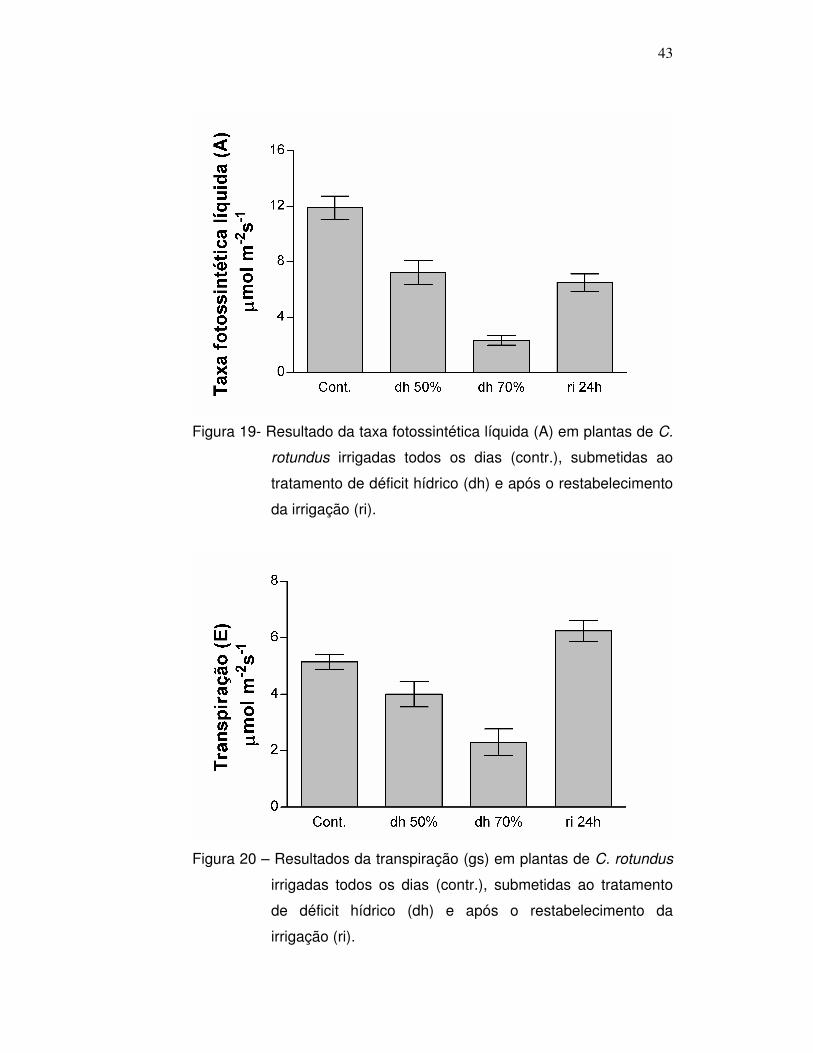
43
Figura 19- Resultado da taxa fotossintética líquida (A) em plantas de C.
rotundus irrigadas todos os dias (contr.), submetidas ao
tratamento de déficit hídrico (dh) e após o restabelecimento
da irrigação (ri).
Figura 20 – Resultados da transpiração (gs) em plantas de C. rotundus
irrigadas todos os dias (contr.), submetidas ao tratamento
de déficit hídrico (dh) e após o restabelecimento da
irrigação (ri).
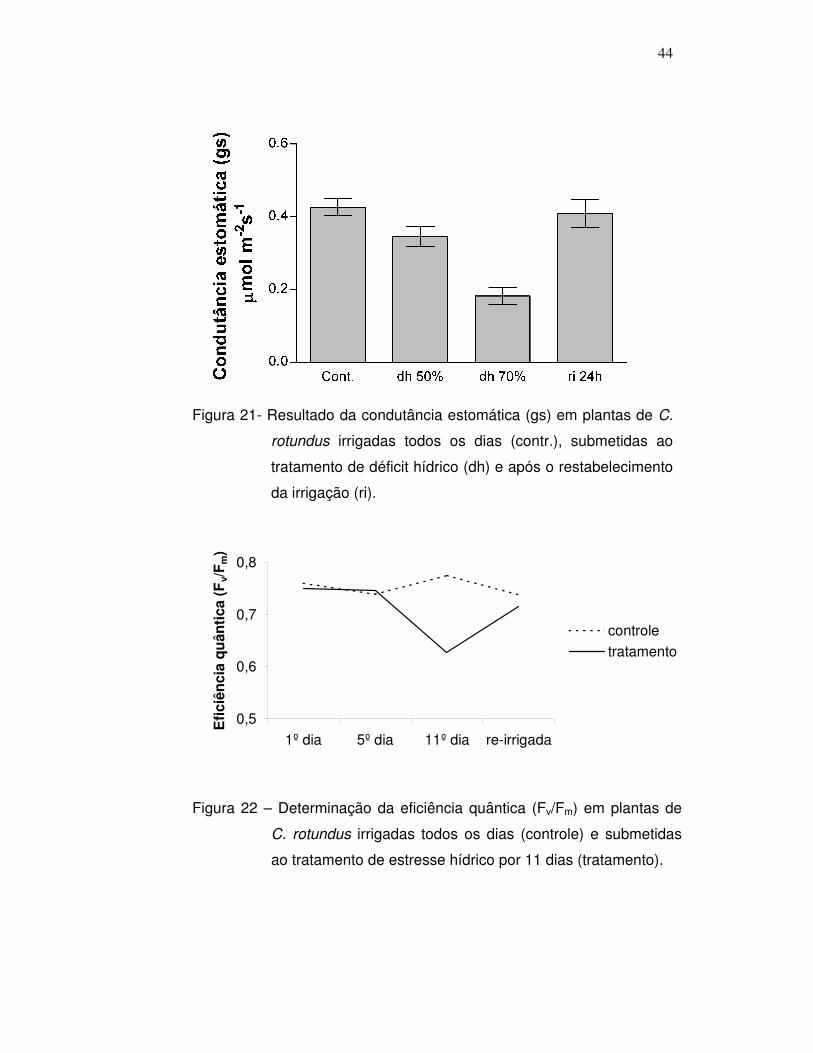
44
Figura 21- Resultado da condutância estomática (gs) em plantas de C.
rotundus irrigadas todos os dias (contr.), submetidas ao
tratamento de déficit hídrico (dh) e após o restabelecimento
da irrigação (ri).
0,5
0,6
0,7
0,8
1º dia 5º dia 11º dia re-irrigada
Efi
ciên
cia
qu
ânti
ca (
Fv/
Fm
)
controle
tratamento
Figura 22 – Determinação da eficiência quântica (Fv/Fm) em plantas de
C. rotundus irrigadas todos os dias (controle) e submetidas
ao tratamento de estresse hídrico por 11 dias (tratamento).

45
5.4 - Determinação do teor de clorofila
Os dados apresentados para teor de clorofila na figura 23,
mostram a redução nas plantas estressadas em relação às plantas
controle ao longo dos dias de estresse. Quando comparados aos dados
de eficiência quântica apresentados na figura 22 esse decréscimo no
teor de pigmentos pode sugerir uma capacidade de adaptação da C.
rotundus às condições adversas de modo que a quantidade de fótons
captada pelas clorofilas do complexo antena que se mantiveram
inalteradas é o suficiente para manter os valores de fluorescência da
clorofila a nas plantas estressadas em relação ao controle até o 7º dia
após a suspensão da irrigação. No 11º dia, o estresse hídrico severo
contribuiu para redução do teor de pigmentos, o que também afetou o
rendimento quântico, porém, após o re-estabelecimento da irrigação,
com o ajuste do metabolismo da planta, o teor de pigmentos voltou a
aumentar, acompanhado do rendimento quântico da fotossíntese.
15
20
25
30
35
40
1º dia 5º dia 11º dia re-irrigada
Teo
r d
e p
igm
ento
s
controle
tratamento
Figura 23 – Determinação do teor de pigmentos fotossintéticos em
plantas de C. otundus irrigadas todos os dias (controle) e
submetidas ao tratamento de estresse hídrico por 11 dias
(tratamento).

46
5.5 – Microscopia Eletrônica de Varredura
O déficit hídrico tem efeito em diversos processos fisiológicos
das plantas, visto que o estresse geralmente aumenta a resistência
difusiva ao vapor de água, mediante fechamento dos estômatos,
reduzindo a transpiração e, conseqüentemente, o suprimento de CO2
para a fotossíntese. Muitos desses efeitos refletem mecanismos de
adaptação das plantas ao ambiente (Nogueira, 1997).
Mendonça (2000) estudando os estômatos de plantas daninhas
monocotiledôneas observou em Cyperus rotundus L. a presença de
apenas uma fileira de estômatos nos bordos foliares da superfície
adaxial, e densidade estomática de 135 estômatos/mm2 na superfície
abaxial.
Procurou-se observar o comportamento estomático das plantas
de C. rotundus conforme os tratamentos de estresse hídrico a que
estavam sendo submetidas.
Observou-se após o preparo das amostras que nas plantas
controle, que recebiam irrigação diária, os estômatos apresentavam-se
abertos (figura 24 A e B), facilitando as trocas hídricas e gasosas,
porém, nas plantas submetidas ao estresse hídrico, os estômatos
estavam fechados (figura 24 C e D), o que faz com que a planta não
realize as trocas gasosas necessárias para o processo fotossintético,
com conseqüente perda de biomassa no decorrer do tempo. Pudemos
observar também que 24h após o restabelecimento da irrigação (figura
24 E e F), os estômatos ainda não estavam abertos como no controle,
porém, espera-se que após o ajustamento da planta a nova situação
(final do estresse hídrico), os estômatos voltem a abrir normalmente.
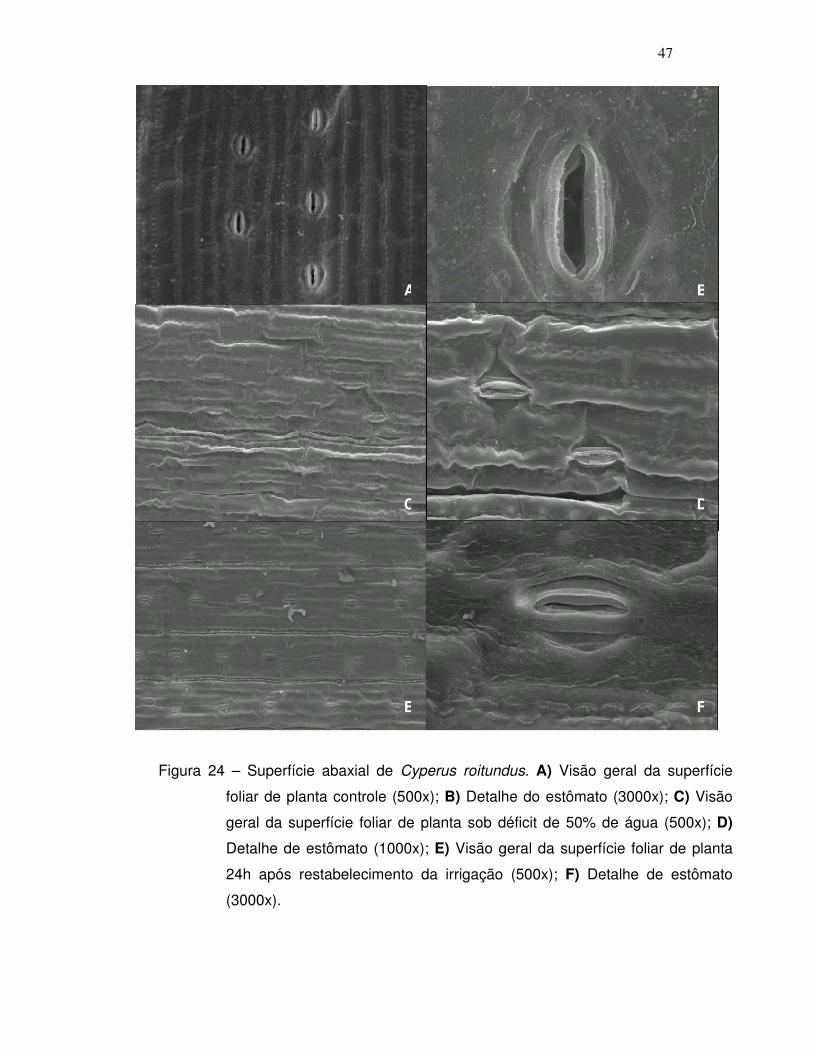
47
Figura 24 – Superfície abaxial de Cyperus roitundus. A) Visão geral da superfície
foliar de planta controle (500x); B) Detalhe do estômato (3000x); C) Visão
geral da superfície foliar de planta sob déficit de 50% de água (500x); D)
Detalhe de estômato (1000x); E) Visão geral da superfície foliar de planta
24h após restabelecimento da irrigação (500x); F) Detalhe de estômato
(3000x).
A B
C D
E F

48
5.6 – Microscopia Eletrônica de Transmissão
Plantas tolerantes ao estresse hídrico são capazes de manter
sua pressão de turgor em situação de baixo potencial hídrico
aumentando o número de moléculas de soluto dentro das células
(Radin, 1983; Bray et al..,2000). Esse transporte de solutos acontece
por intermédio das bombas de prótons. Uma vez que a avaliação visual
do experimento mostrou que os sintomas do estresse, como a murcha
e a clorose das folhas, apareceram somente após um período de 11
dias sem irrigação e que após o restabelecimento da irrigação, a
recuperação da planta foi bastante rápida (figura 13), sugerindo uma
maior ativação das bombas vacuolares, ou mesmo um incremento na
expressão das mesmas, como mostrado por Gaxiola et al.. (2001).
Esperava-se encontrar a marcação ao redor da membrana
vacuolar das células das plantas, uma vez que a bomba de prótons
PPase encontra-se distibuída por tal membrana. Esperava-se, também,
encontrar uma maior marcação nas amostras coletadas de plantas re-
irrigadas, um vez que os dados bioquímicos apresentados apontavam
essa etapa como a de maior atividade da bomba PPase. No entanto, as
preparações obtidas não permitiram observar com clareza as estruturas
celulares. A imunomarcação para PPase permitiu observar que a
presença da bomba de prótons PPase é maior nas plantas controle
(figura 25), sofrendo uma redução durante o estresse (figura 26) e
apresentando uma quantidade ainda menor que o controle nas plantas
re-irrigadas (figura 27), sugerindo que a regulação dessas bombas de
prótons acontecem de maneira pós transcripcional.

49
Figura 25 – Micrografia eletrônica de transmissão com imunomarcação anti PPase em plantas controle (20000x).
Figura 26 – Micrografia eletrônica de transmissão com imunomarcação
anti PPase em plantas sob estresse hídrico (20000x).

50
Figura 27 – Micrografia eletrônica de transmissão com imunomarcação
anti PPase em plantas re-irrigadas (20000x).
5.7 - Atividade hidrolítica e de transporte de H+ da P-ATPase,
da V-ATPase e da H+-PPase em colmos de Saccharum spp.
Os resultados apresentados a seguir mostram uma
caracterização inicial e preliminar das atividades das bombas de
prótons de membrana plasmática e de vacúolo em colmos da planta
Saccharum spp., cultivadas em condições de campo. Tal espécie foi
escolhida para o experimento por ser uma das plantas mais importantes
economicamente para o Brasil.
A figura 28 mostra os resultados de atividade hidrolítica e a
figura 29 de transporte de H+ pelas bombas P-ATPase, V-ATPase e
PPase isoladas de casca de colmo plantas crescidas em condições de
campo, ou seja, os resultados encontrados relatam a atividade dessas
bombas em tal tecido.

51
Figura 28 - Atividade hidrolítica das bombas de prótons em vesículas
enriquecidas de membrana plasmática e de vacúolo de
casca de colmo de plantas cultivadas em condições de
campo (média representativa de três experimentos).
Figura 29 – Bomba de próton ATP/PPi dependente de vesículas de
membrana plasmática e tonoplasto de casca de colmo de
plantas cultivadas em condições de campo. P-ATPase (a),
V-ATPase (b) e PPase (c). Fluorescência do ACMA na
presença de 50 µg de proteína de membrana.
P-ATPase V-ATPase PPase0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Ati
vid
ade
µµ µµm
ol.m
g-1
.ptn
-1

52
Os resultados apresentados na figura 28 sugerem o padrão de
atividade hidrolítica para casca de colmo de cana-de-açúcar, onde a
bomba de prótons PPase apresentou atividade mais alta que as
demais, resultado esse que se confirmou no transporte de prótons
(figura 29).
É possível que esse resultado esteja relacionado com o
transporte de solutos, pois esta espécie vegetal é conhecida por sua
alta atividade de transporte de solutos e síntese de açúcares. O tecido
externo do colmo (que denominamos de casca), possui atividade
fotossintética e, possivelmente alta atividade de biossíntese de
açúcares, o que justificaria a alta atividade da PPase, que estaria
funcionando utilizando-se do PPi liberado pelas reações de biossíntese.

53
6 – CONCLUSÕES
• Em condições de campo a planta daninha Cyperus rotundus
apresentou um padrão de atividade hidrolítica e de transporte
de prótons semelhante ao apresentado na literatura para as
demais plantas, onde a atividade hidrolítica é maior para a V-
ATPase e o gradiente de H+ apresenta magnitude
semelhante ao da PPase;
• Durante o estresse hídrico, observou-se que as plantas de C.
rotundus apresentaram sintomas de déficit hídrico somente
aos 11 dias sem irrigação e que após 24h do seu
restabelecimento, as folhas retomam seu aspecto de
turgescência, semelhante ao que acontece com as
ressurection plants;
• Após restabelecimento da irrigação a bomba de prótons
PPase mostrou-se superior a V-ATPase tanto em hidrólise de
substrato quanto em transporte de prótons, sugerindo que o
gradiente de prótons gerado através da membrana vacuolar
pela hidrólise do PPi poderia estar sendo usado para síntese
de ATP, para que os processos celulares da planta fossem
mantidos;
• Os parâmetros fisiológicos avaliados demonstram que a
planta daninha C. rotundus segue os mesmos padrões das
mais diversas espécies vegetais de metabolismo C4, onde
sob déficit hídrico, a condutância estomática, a eficiência
quântica e o teor de pigmentos decrescem;
• As avaliações morfológicas utilizando a técnica de
microscopia de varredura comprovam o fechamento
estomático durante o estresse hídrico;
• As avaliações da quantificação e localização da bomba de
prótons PPase, utilizando a técnica de imunomarcação e
visualização em microscópio eletrônico de transmissão
mostraram que a presença das bombas de prótons PPase é

54
maior nas plantas controle, sofrendo redução durante o
estresse e não superando a quantidade encontrada no
controle mesmo após o restabelecimento da irrigação,
sugerindo que a regulação seja pós transcripcional;
• As avaliações efetuadas em tecido de cana-de-açúcar
mostraram ser maior a atividade de hidrólise e transporte de
prótons pelas bombas vacuolares em casca e que, neste
caso, a PPase foi a que apresentou maior atividade,
sugerindo que seu funcionamento pode estar relacionado
com a síntese de ATP, utilizando o gradiente gerado pela
hidrólise do PPi, para ser utilizado durante os processos de
biossíntese de açúcares

55
7 - BIBLIOGRAFIA
Amzallag, G.N., Lerner, H.R. (1994) Adaptation versus pre-existing
resistance: an intergenotype analysis of the response of Sorghum
bicolor to salinity. Journal of Plant Science. 42,125-141
Baykov, A. A., Dubnova, E.B., Bakuleva, N.P., Eytushenko, O.A., Zhen,
R., Rea, P. A. (1993) Differential sensitivity of membrane-
associated pyrophosphatases to inhibition by diphosphonates and
fluoride delineates two classes of enzyme. FEBS Letters, 327, 199-
202.
Bergamaschi, H. (1992) Desenvolvimento de déficit hídrico em culturas.
In: Bergamaschi, H. (Coord.). Agrometeorologia Aplicada à
Irrigação. Porto Alegre. Editora da Universidade/UFRGS. p. 25-32.
Bewley, J.D., Krochko, J.E. (1982). Dessication tolerance. In: Lange,
O.L., Nobel, P.S., Osmond, C.B., Ziegler, H. (Eds.), Encyclopedia
of Plant Physiology. New series, vol. 12B. Springer, Berlin, pp.
325–378.
Boddey, R. (1993) Green energy from sugar-cane. Chem Ind-London
(10): 355-358.
Bohnert, H.J., Nelson, D.E., Jensen, R.G. (1995) Adaptations to
environmental stress, Plant Cell 7, 1099 - 1111.
Bohnert, H.J., Ayoubi, P., Borchert, C., Bressan, R. A., Burnap, R.L.,
Cushman, J.C., Cushman, M. A., Deyholos, M., Galbraith,
R.F.D.W. (2001) A genomics approach towards salt stress
tolerance. Plant Physiology and Biochemistry 39, 295-311.
Bowman EJ, Siebers A, Altendorf K (1988) Bafilomicins: A class of
inhibitors of membrane ATPases from microorganisms, animal
cells, and plant cells. Proc Natl Acad Sci (USA) 85: 7972
Bray, E.A.; Bailey-Serres, J.; Weretilnyk, E. (2000) Responses to abiotic
stresses, in: W. Gruissem, B. Buchannan, R. Jones (Eds.),
Biochemistry and Molecular Biology of Plants, American Society of
Plant Biologists, Rockville, MD, pp. 158–1249.

56
Bradford, M.M. (1976) A rapid and sensive method for the quantitation
of microgram quantities of protein utilizing the principle of the
protein-dye binding. Anal Biochem 72, 248-254.
Cardenas, J., Barea, J.L., Rivas, J., Morena, C.G. (1972) Purification
and properties of nitrite reductase from spinach leaves. FEBS
Letters, 23, 131-135.
Colombo, R., Cerana, R. (1993) Enhanced activity of tonoplast
pyrophosphatase in NaCl-grown cells of Dacus carota, J. Plant
Physiol. 142, 226-229
Crowe JH, Hoekstra FA, Crowe LM (1992) Anhydrobiosis. Annu Rev
Physiol 54: 579–599.
Cruz, R., Cardenas, J., Lagos, E., Franco, H., Carmona, C., Vargas, C.,
Riveros, G. (1971) El coquito y su control. Instituto Colombiano
Agropecuario, N42.
Darley, C.P.; Davies, J.M.; Sanders, D. (1995) Chill-lnduced changes in
the activity and abundance of the vacuolar proton pumping
pyrophosphatase from Mung Bean hypocotyls. Plant Physiology,
V. 109: 659-665.
Dietz, K.J., Tavakoli, N., Kluge, C., Mimura, T., Sharma, S.S., Harris,
G.C., Chardonnens, A.N., Golldack, D. (2001) Significance of the
V-type ATPase for the adaptation to stressful growth conditions
and its regulation on the molecular and biochemical level. Journal
of Experimental Botany 52, 1969-1980.
DeRocher, E..J. and Bohnert, H.J. (1993). Development and
environmental stress employ different mechanisms in the
expression of a plant gene family. Plant Cell 5, 1611-1625.
Doorembos, J.; Kassam, A.H. (1979) Efectos del agua sobre el
rendimiento de los cultivos. Roma: FAO, p. 212 (FAO. Riego y
Drenaje, 33).
Drory, O. and Nelson, N. (2006) The emerging structure of vacuolar
ATPases. Physiology (Bethesda) 21, 317–325.

57
DuPont, F.M., Morrissey, P.J. (1992) Subunit composition and Ca2+-
ATPase activity of the vacuolar ATPase from barley roots. Archives
of Biochemistry and Biophysics, v.294, p.341-346.
Durigan, J.C. (1991); Manejo da tiririca (Cyperus rotundus L.) antes e
durante a implantação da cultura da cana-de-açúcar (Saccharum
spp.), Universidade Estadual Paulista, Faculdade de Ciências
Agrárias e Veterinárias. Tese de Livre Docência, Jaboticabal, SP,
336p.
Dure, L. I. 1993. Structural motifs in Lea proteins. p. 91-103. In T. J.
Close, and E. A. Bray (eds.) Plant Responses to Cellular
Dehydration during Environmental Stress. Amer. Soc. Plant
Physiol., Rockville, MD.
EMBRAPA. (1981) Fruticultura de Clima Tropical; programas nacionais
de pesquisa em fruticultura de clima tropical. Brasília,
Departamento de Informação e Documentação, 198p.
Eullaffroy e Vernet, (2003) The F684/F735 chlorophyll fluorescence
ratio: a potencial tool for rapid detection and determination of
herbicide phytotoxicity in algae. Water Research, 37, 1983-1990.
Façanha AR, de Meis L (1998) Reversibility of H+-ATPase and H+-
Pyrophosphatase in tonoplast vesicles from maize coleoptiles and
seeds. Plant Physiol 116: 1487
Fancelli, A. L. e Dourado Neto, D. (2000) Produção de milho. Guaíba:
Agropecuária, 360p.
Fisher, H.H. (1973) Conceito de erva daninha. In: Warren, G.F., Willian,
R.D., Fisher, H.H., Sacco, J. da C., Lamar, R.V., Alberto, C.A.;
Curso Intensivo de Controle de Ervas Daninhas, Universidade
Federal de Viçosa, 11-15.
Fiske C.F., Subbarow Y. (1925) The colorometric determination of
phosphorus. J Biol Chem 66: 375

58
Forster, R., Cerdeira, A.L. (1993); Multiplicação Vegetativa da Tiririca
(Cyperus rotundus L.). EMBRAPA – CNPDA, Jaguariúna, SP.
Gaff, D.F. (1971). Desiccation-tolerant flowering plants in Southern
Africa. Science 174, 1033–1034.
Gaff, D.F. (1977). Desiccation tolerant vascular plants of Southern
Africa. Oecologia 31, 93–109.
Gaxiola, R.A., Li, J., Undurraga, S., Dang, L.M., Allen, G.J., Alper, S.L.
(2001) Drought and salt tolerant plants result from overexpression
of the AVP1 H+-pump. PNAS, 98, 11444-11449.
Gaxiola, R. A.; Palmgrenb, M. G.; Schumacher, K. (2007) Plant proton
pumps. FEBS Letters 581 (2007) 2204–2214.
Giannini, J.L., Briskin, D.P. (1987) Proton transport in plasma
membrane and tonoplast vesicles from red beet (Beta vulgaris L.)
storage tissue. Plant Physiol 84: 613.
Guimarães, S.C. (1993); Influencia da idade de corte e de aditivos na
ação do Glyphosate sobre a tiririca (Cyperus rotundus L.), In:
Congresso Brasileiro de Herbicidas e Plantas Daninhas, 19.
Resumos..., Londrina, PR.
GUINNES BOOK (1994); O Grande Livro dos Recordes.
Han, N.; Shao, Q.; Lu, C.M.; Wang, B.S. (2005) The leaf tonoplast V-
H+ATPase activity of a C3 halophyte Suaeda Salsa is enhanced by
salt stress in a Ca-dependent mode. Journal of Plant Physiology, v.
162, p. 267-274.
Hoekstra FA, Golovina EA, Buitink J (2001) Mechanisms of plant
desiccation tolerance. Trends Plant Sci 6: 431–438.
Holm, L.G.; Plucknett, D.L.; Pancho, J.V. and Herberger, J.P. (1977)
Cyperus rotundus L. In: The world´s worst weeds: Distribution and
biology. University Press of Hawaii, Honolulu, p. 8-24.
Ingram, J., Bartels, D. (1996) The molecular basis of dehydration
tolerance in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol.
47:377-403.

59
Jakl, T.; Bolhar-Nordenkampf, H. R (1991) Energy conversion
efficiency and energy partitioning of white lupins (Lupinus albus L.)
Bioresource Technology, Volume 36, Issue 2, Pages 193-197.
Janicka-Russak, M. and Klobus, G. (2007) Modification of plasma
membrane and vacuolar H+-ATPases in response to NaCl and
ABA. J. Plant Physiol. 164, 295–302.
Jones, H.G.; Jones, M.B. (1989) Introduction: some terminology and
common mechanisms, in: H.G. Jones, T.J. Flowers, M.B. Jones
(Eds.), Plants Under Stress, Cambridge university Press,
Cambridge, pp. 1–10.
Junqueira Neto, A., Resende, P.M. de, Alcântara, E.N. (1982);
Herbicidas na cultura do feijão (Phaseolus vulgaris L.) e sua
influência na produção de grãos e no controle de plantas
daninhas. In: Reunião Nacional de Pesquisa do Feijão – I, Anais,
Goiânia, EMBRAPA – CNPAF, p. 223-224.
Karlsson, J. (1975) Membrane-bound potassium and magnesium ion-
stimulated inorganic pyrophosphatase from roots and cotyledons of
sugar beet (Beta Vulgaris L.). Biochimica et Biophysica Acta (BBA)
- General Subjects, 399, 356-363.
Kramer, P. J. (1969) Plant and soil water relationships: a modern
synthesis. New York, McGraw Hill. 482 p.
Kissmann, K.G. (1991); Plantas infestantes e nocivas. São Paulo:
BASF BRASILEIRA S.A., tomo 1: p. 167-173.
Leigh, R.A., Pope, A.J., Jennings, I.R., Sanders, D. (1992) Kinetics of
the vacuolar H+-pyrophosphatase. The roles of magnesium,
pyrophosphate, and their complexes as substrates, activators, and
inhibitors. Plant Physiol 100: 1698-1705.
Levitt, J. (1980) Responses of plants to environmental stress, 2nd ed,
Academic Press, New York, 365-434.
Lichko, L.P., Okorokov, L. A. (1984) Some properties of membrane-
bound, solubilized and reconstituted into liposomes H+-ATPase of
vacuoles of Saccharomyces carlsbergensis . FEBS Letters, 174,
233-237.

60
Liu, J.; Zhu, J.K. (1998) A calcium sensor homolog required for plant
salt tolerance, Science 280 1943–1945.
Long, A.R., Williams, L.E., Nelson, S.J. and Hall, J.L. (1995)
Localization of membrane pyrophosphatase activity in Ricinus
communis seedlings. J. Plant Physiol 146, 629–638.
Lorenzi, H. (1983); Plantas daninhas e seu controle na cultura da cana-
de-açúcar. In: Reunião Técnica Agronômica. Anais...:
COOPERSUCAR, Piracicaba, p. 59-73.
Lorenzi, H. (1994) Manual de identificação e controle de plantas
daninhas: plantio direto e convencional. 4º edição – Nova Odessa,
SP.
Löwry, R., Rockel, B., Kirsch, M., Ratajczak, R., Hörtensteiner, S.,
Martinoia, E., Lüttge, U., Lowry, O.H., Rosebrough, N.J., Farr,
A.L., Randall, R.J. (1951) Protein measurement with the Folin
phenol reagent. J Biol Chem 193: 265.
Lüttge, U., Ratajczak, R., Rausch, T., Rockel, B. (1995) Stress
responses of tonoplast proteins: an example for molecular
ecophysiology and the search for eco-enzymes. Acta Bot Neerl 44:
343-362.
Lüttge, U., Ratajczak, R. (1997) The physiology, biochemistry and
molecular biology of the plant vacuolar ATPase. In RA Leigh, D
Sanders, eds, The Plant Vacuole: Advances in Botanical
Research, Vol. 25. Academic Press, Oxford, pp 253-296
Maeshima, M., Yoshida, S. (1989) Purification and properties of the V-
PPase from mung bean. J. Biol. Chem 264, 20068-20073
Maeshima M (1990) Development of vacuolar membranes during
elongation of cells in mung bean hypocotyls. Plant Cell Physiol 31:
311-317
Maeshima, M. (2000) Vacuolar H+-pyrophosphatase. Biochim Biophys
Acta, 1465, 37-51.
Magalhães, P.C. e Durães, F.O.M. (2006) Fisiologia da produção de
milho. Circular Técnica 76, EMBRAPA - Sete Lagoas, MG.

61
Mahajan, S.; Narendra, T. (2005) Cold, salinity and drought stresses: An
overview Archives of Biochemistry and Biophysics 444, p. 139–
158.
Mahajan, S., Tuteja, N. (2005) Cold, salinity and drought stresses: An
overview. Archives of Biochemistry and Biophysics, Volume 444,
p. 139-158.
Matsuura-Endo, C.; Maeshima, M.; Yoshida, S. (1992) Mechanism for
the decline of V-ATPase activity in mung bean hypocotyls during
chilling. Plant Physiol 100: 718-722.
Matters, G.L., Scandalios, J.G. (1986) Effect of the free radical-
generating herbicide paraquat on the expression of the superoxide
dismutase (Sod) genes in maize. Biochimica et Biophysica Acta
(BBA) - General Subjects, 882, 29-38.
Mendonça, C.G. (2000) Algumas características da superfície foliar de
diversas plantas daninhas monocotiledôneas. Botucatu, 89p.
Dissertação (M.S.)
Mercado, B.L. (1979); A monograph on Cyperus rotundus L. Biotrop
(Bulletin 15), 63p.
Nakanishi, Y.,Maeshima, M. (1998) Molecular Cloning of Vacuolar H+-
Pyrophosphatase and Its Developmental Expression in Growing
Hypocotyl of Mung Bean. Plant Physiol. 116: 589-597.
Navari-Izzo, F., Rascio, N. (1999). Plant responses to water deficit
conditions. In: Pessarakli, M. (Ed.), Handbook of Plant and Crop
Stress. Marcel Dekker Inc., New York, pp. 231–270.
Orlando Filho, J. (1983) Nutrição e adubação de cana-de-açúcar no
Brasil. Piracicaba: Instituto do Açúcar e do álcool, p. 369.
Otoch, M.L.O.; Sobreira, A.C.M.; Aragão, M.E.F.; Lima, M.G.S.; Melo,
D.F. (2001) Salt modulation of vacuolar H+-ATPase and H+-
Pyrophosphatase activities in Vigna unguiculata. J. Plant Physiol.
158. 545–551
Palmgren, M.G. (1991) Regulation of plant plasma membrane H+-
ATPase activity, Physiol. Plant 83, p.314–323.

62
Pedersen, P.L., Carafoli, E. (1987) Ion motive ATPases. I. Ubiquity,
properties, and significance to cell function. Trends in Biochemical
Sciences, 12,146-150.
Perotti, E.; Gavin, O.; Widmer, F.; Chanson, A. (1994) Purification and
functional reconstitution of the tonoplast pyrophosphate-dependent
proton pump from Rubus hispidus cell cultures. Plant Science, v.
103, p. 25-31.
Qiu, N.; Chen, M.; Guo, J.; Bao, H.; Ma, X.; Wang, B. (2007) Coordinate
up-regulation of V-H+-ATPase and vacuolar Na+/H+ antiporter as a
response to NaCl treatment in a C3 halophyte Suaeda salsa. Plant
Science, Volume 172, Issue 6, Pages 1218-1225
Radin, W.J. (1983) in Limitations to Efficient Water Use in Crop
Production, eds Taylor, H.M., Jordan, W.R. e Sinclair, T.R. (ASA-
CSSA-SSSA, Madison, WI), 267-676
Ratajczak, R., Hinz, G. and Robinson, D.G. (1999) Localization of
pyrophosphatase in membranes of cauliflower inflorescence cells.
Planta 208, 205–211.
Ratajczak, R. (2000) Structure, function and regulation of the plant
vacuolar H+.-translocating ATPase. Biochimica et Biophysica Acta,
v.1465 p. 17-36.
Rea, P.A., Sanders, D. (1987) Tonoplast energization: two H+ pumps,
one membrane. Physiol Plant 71: 131-141.
Rea, P.A., Kim, Y., Sarafian, V., Poole, R.J., Davies, J.M., Sanders, D.
(1992) Vacuolar H+-translocating pyrophosphatases: a new
category of ion translocase. Trends Biochem. Sci. 17, 348-353.
Rea, P.A., Poole, R. (1993) Vacuolar H+-translocating pyrophosphatase.
Annu Rev Plant Physiol Plant Mol Biol 44 – 157-180.
Resende, M., França, G.E. e Couto, L. (2000) Cultivo do milho irrigado.
Sete Lagoas: Embrapa Milho e Sorgo, 39p. (Embrapa Milho e
Sorgo.Circular Técnica, 6).

63
Robinson, D.G., Haschke, H.P., Hinz, G., Hoh, B., Maeshima, M., Marty,
F. (1996) Immunological detection of tonoplast polypeptides in the
plasma membrane of pea cotyledons. Planta 198: 95-103.
Rockel, B. Lüttge, U., Ratajczak, R. (1998) Changes in message
amount of V-ATPase subunits during salt-stress induced C3-CAM
transition in Mesembryanthemum crystallinum. Plant Physiology
and Biochemistry, v. 36, p. 567-573.
Russak, M.J.; Klobus, G. (2007) Modification of plasma membrane and
vacuolar H+-ATPases in response to NaCL and ABA. Journal of
Plant Physiology, v. 164, p. 295-302.
Sarafian, V., Ki.m, Y., Poole, R.J., Rea, P.A. (1992) Molecular cloning
and sequence of cDNA encoding the pyrophosphate-energized
vacuolar membrane proton pump of Arabidopsis thaliana. Proc
Natl Acad Sci USA 89: 1775-1779.
Sato, M.H., Maeshima, M., Ohsumi, Y., Yoshida, M. (1991) Dimeric
structure of H+- translocating pyrophosphatase from pumpkin
vacuolar membranes. FEBS Letters, Volume 290, Issues 1-2, 23
September 1991, Pages 177-180.
Serrano, R. (1990) Plasma membrane ATPase, in: C. Larsson, I.M.
Moller (Eds.), The Plant Plasma Membrane, Structure, Function
and Molecular Biology, Springer, Berlin, pp. 127–153.
Sharma, R.; Gupta, R. (2007) Cyperus rotundus extract inhibits
acetylcholinesterase activity from animal and plants as well as
inhibits germination and seedling growth in wheat and tomato. Life
Sciences, Volume 80, Issues 24-25, 30, Pages 2389-2392.
Shinozaki, K.; Yamaguchi-Shinozaki, K.; Seki, M. (2003) Regulatory
network of gene expression in the drought and cold stress
responses. Current Opinion in Plant Biology, 6:410–417.
Smyth, D.A., Wu, M.X., Black, C.C. (1984) Phosphofructokinase and
fructose 2,6-bisphosphatase activities in developing corn seedlings
(Zea mays L.) Plant Science Letters, Volume 33, Issue 1, January
1984, Pages 61-70.

64
Sze, H. (1985) H+-Translocating ATPases: advances using membrane
vesicles. Annual Review of Plant Physiology. V. 36, P. 175-208.
Sze, H., Ward, M.W., Lai, S., Perera, I. (1992) Vacuolar type H+-
translocating ATPases in plant endomembranes: subunit
organization and multigene families. J. Exp. Biol 172, 123-135
Taiz, L., Zeiger, E. (1995) Plant Physiology.-. Benjamin/Cummings
Publishing Co., Inc. Redwood City. pp 120
Taiz, L. (1992) The Plant Vacuole. J. exp. Biol. 172, 113-122.
Takasu, A., Nakanishi, Y., Yamauchi, T. and Maeshima, M. (1997)
Analysis of the substrate binding site and carboxyl terminal region
of vacuolar H+-pyrophosphatase of mung bean with peptide
antibodies. J. Biochem. 122, 883–889.
Takeshige, K., Tazawa, M. (1989) Determination of the inorganic
pyrophosphate leve1 and its subcellular localization in Chara
corallina. J Biol Chem 264 3262-3266.
van Rensburg, L. (1994) Adaptive significance of photosynthetic and
metabolic regulation in Nicotiana tabacum L. plants during drought
stress. Ph.D. Diss., Potchefstroom University for Christian Higher
Education, Potchefstroom, South Africa.
Victoria Filho, R.; Christoffoleti, P.J. (2004) Manejo de plantas daninhas
e produtividade da cana-de-açúcar. Visão Agrícola, Piracicaba, p.
32-37.
Walker, R.R., Leigh, R.A. (1981) Mg"-dependent, cation-stimulated
inorganic pyrophosphatase associated with vacuoles isolated from
storage roots of red beet (Beta vulgaris). Planta 153: 150-155.
Ward, J.M., Sze, H. (1992) Proton transport activity of the purified
vacuolar H+-ATPase from oats. Plant Physiol 99: 925-931.
Weiner, H.; Stitt, M.; Heldt, H.W. (1987) Subcellular compartmentation
of pyrophosphate and alkaline phosphatase in leaves. Biochim
Biophys Acta 893: 13.
Yanqiong, L.; Xingliang, L.; Shaowei, Z.; Hong, C.; Yongjie, Y.;
Changlong, M.; Jun, L. (2007) Drought-resistant physiological

65
characteristics of four shrub species in arid valley of Minjiang
River, China. Acta Ecologica Sinica, Volume 27, Issue 3, Pages
870-877
Yao, G.F.; Feng, H.T.; Cai, Y.L.; Qi, W.L. and Kong, K.M. (2007)
Characterization of vacuolar-ATPase and selective inhibition of
vacuolar-H(+)-ATPase in osteoclasts. Biochemical and Biophysical
Research Communications, Volume 357, Issue 4, 15, Pages 821-
827.
Yoshida, S., Matsuura-Endo, C., Etani, S. (1989) Impairment of the
tonoplast H+-ATPase as an initial physiological response of the
cells to chilling in mung bean. Plant Physiol 89, 634-642.
Xiao, Y.T.; Xiang, L.X.; Shao, J.Z. (2007) Vacuolar H+-ATPase. The
International Journal of Biochemistry & Cell Biology, In Press,
Corrected Proof, Available online 22 August 2007.