Embed Size (px)
Citation preview

1
CIIDIR IPN UNIDAD DURANGO
INFORME FINAL
ANALISIS DE SIMILITUD DE LAS ESPECIES DE CHAPULÍN EN PASTIZALES DE
DURANGO UTILIZANDO ANÁLISIS MULTIVARIADO
CLAVE SIP 20070837
ISAIAS CHAIREZ HERNÁNDEZ

2
INTRODUCCIÓN
Para conocer los habitats del chapulín es necesarion conocer su topografía,
vegetación, clima y suelo en relación con la distribución de los chapulines. En el aspecto
de la topogrfía, Samietz et al. (2005) realizaron un estudio con la especie Melanoplus
sanguinipes en un gradiente de altitud en laboratorio en California y encontraron que la
movilidad y reposo están determinado por la altitud de origen de los padres.
De acuerdo a Rodell (1977), la temperatura y la precipitación son consideradas los
componentes más importantes que modifican la dinámica poblacional de los chapulines, la
precipitación esta relacionada con la humedad del suelo y con el desarrollo del embrión y
tiene efecto sobre la cantidad y calidad de las plantas que consumen las ninfas y los adultos.
También la precipitación y la humedad tienen influencia en la mortalidad de los chapulines.
La temperatura determina las tazas de desarrollo, oviposición y eclosión, en particular, esta
última está en función de la acumulación de unidades calor.
Se ha reportado que la textura del suelo tiene relación con la oviposición de las
hembras de chapulín. Edwards (1965) considero tres diferentes tipos de suelo; arena, limo
y arcilla y tres niveles de humedad; saturada, intermedia y seca para determinar la
preferencia de la hembra de Melanoplus sanguinipes (Fabricius) para la oviposición,
cuando se le da a escoger libremente. La hembra prefirió el suelo arenoso y húmedo a todos
los otros sitios y evitó los suelos completamente secos. Cuando se le ofrecían los suelos sin
humedad, la hembra prefirió los suelos arenosos a finos.

3
Sin embargo, la vegetación es uno de los factores que más influencia tienen sobre
los chapulines. Capinera y Sechrist (1982) llevaron a cabo muestreos de composición
botánica y del complejo chapulín, en las planicies centrales de los Estados Unidos en el
Noroeste de Colorado y encontraron que el número total de chapulines estaba
significativamente relacionado con pastos de alto contenido de biomasa (sin pastoreo o con
bajo pastoreo), mientras el número de chapulines de la subfamilia Oedipodinae era
significativamente alto en pastizales de baja biomasa (Pastoreo alto o moderado).
Entre las herramientas estadísticas la regresión lineal por pasos se ha utilizado para
relacionar los factores ecológicos con la población de chapulín. Skinner y Child (2000)
utilizaron la regresión múltiple por pasos para determinar cuales variables de clima y suelo
explicaban mejor la población de chapulín. Capinera y Horton (1989) realizaron un estudio
en Montana, Wyoming, Colorado y Nuevo México utilizando análisis de correlación y
regresión. Los resultados mostraron que los niveles de infestación estaban relacionados con
precipitación, temperatura y textura del suelo. Sin embargo, el uso del análisis de similitud
y de componentes principales ha sido poco usado para asociar les chapulines con la
veegetación
El propósito de este estudio es conocer La asociación de las especies más
abundantes de chapulín en los pastizales de Durango con las especies vegetales con el
análisis de similitud los componentes principales

4
ANTECEDENTES
Ciclo biológico del chapulín
El chapulín es un Acrididae que se desarrolla principalmente en pastizales y cuya
distribución va desde las planicies de Estados Unidos hasta Guatemala (Kevan, 1977). Los
huevecillos de chapulín son depositados por la hembra a principios de Octubre y finales de
Noviembre eclosionan a finales de mayo y principios de junio y su máxima taza de eclosión
es a mediados de junio. Existen de 5 a 6 estados ninfales y el tiempo de huevo a adulto es
de 70 a 90 días (Glogoza y Weiss, 1997). La adulta y quinta etapa ninfal son las más
voraces y es cuando afecta significativamente los cultivos y pastizales. Se sabe que
condiciones específicas de suelo y clima y su relación con la biota algunas veces generan
condiciones ideales para el crecimiento explosivo de las poblaciones de chapulines, donde
más de dos chapulines por ft2, ocasionan daños de importancia económica en los cultivos y
pastizales.
Hábitos alimenticios
Los hábitos alimenticios del chapulín son diversos pero algo predecibles. Las
especies polífagas como los melanoplus, tiene un número muy extenso de plantas
hospederas, y las especies olifagas como Hypochlora alba se alimentan de pocas especies
de plantas en este caso Artemisia spp. Algunas especies comen estrictamente gramíneas,
arbustos o algunas otras comen una mezcla de ambos. También se alimentan de
desperdicios de plantas e insectos, el canibalismo es común. La mayoría de
Gomphorcerinae se alimenta de pasto, y muchos Oedopodinae. Catantopinae,
especialmente Melanoplus spp., se alimentan de una mezcla de pasto y arbustos pero

5
prefieren los arbustos. Las preferencias en el alimento están correlacionadas con las
estructuras de la boca. Los dientes incisivos de las mandíbulas son bruscos en especies que
prefieren gramíneas y puntiagudos en las especies que prefieren los arbustos; estados
intermedios son comunes (Isely, 1944). Existen otros factores que determinan las
preferencias en el alimento como son la dureza de la hoja, los vellos de la hoja y el
contenido de agua; también se cree que las diferencias en las preferencias alimenticias son
por las diferencias químicas tales como: Azucares, nitrógeno orgánico, taninos y otros. Aun
cuando existen algunas excepciones los chapulines prefieren plantas que son mas
adecuadas para su crecimiento, desarrollo y sobre vivencia. Finalmente los chapulines se
alimentan cuando la temperatura esta entre 15o y 30oC y emplean aproximadamente el 15%
de tiempo para alimentarse (Uvarov, 1977).
Distribución
Los mapas de distribución indican que la mayoría de las especies se restringen
áreas ecológicas especificas que están delimitadas por barreras específicas. Estas barrearas
incluyen cadenas montañosas, áreas de pastizales, planicies y desiertos. Especies benéficas
o dañinas son encontradas solamente en algunas de estas regiones, y esta información
puede ser utilizada como guía en muestreos. Estudios zoogeográficos son de vital
importancia para identificar las fronteras geográficas de las distribución de los chapulines.
En algunos casos las fronteras no están muy definidas, sin embargo, hay muchas especies
que tienen patrones de distribución que no pueden ser explicados solamente por
característica topográfica. El conocimiento de los patrones de elevación, vegetación y clima
relacionado con el complejo chapulín es un tema de mucho interés en ecología (Lockwood,

6
et al, 1988-92).
Daños a cultivos y pastizales por chapulín
Se ha considerado que de las plagas de insectos la de mayor pérdida económica en
muchas partes del mundo es el chapulín. En los Estados Unidos existen cerca de 400
especies de chapulín en los estados del oeste y se calcula que existe pérdida de forraje
estimadas en 393 millones de dólares anuales (Hewitt y Osanger, 1983), además se estima
perdida de pasto en Arizona debido al complejo chapulín de 600 libras en peso seco por
acre (Lockwood, A. et al 1988-92 “Boopedon nubilum Fact Sheet”) y se ha reportado que
una población densa de Melanoplus differentialis puede destruir un cultivo de plantas
jovenes de maíz en 3 o 4 días (Lockwood, A. et al 1988-92 “Melanoplus differentialis
Fact Sheet”). Por otro lado, McLeod (1994) establece que para tierras de cultivo los
umbrales económicos son de 8 o más adultos o 30 a 40 ninfas por yd2 y sugiere que los
umbrales para bordes de tierras de cultivo tales como lados de carreteras y agostaderos
adyacentes a tierras de cultivo son de 20 a 40 adultos o 50 a 75 ninfas por yd2 , mientras
que,(Belovsky, 2000), establece que densidades bajas de chapulines 6/yd2 (8/m2) pueden
causar daños considerables al forraje base (pérdidas arriba del 70%). Altos niveles de daño
ocurren si el forraje base tiene bajo potencial de abundancia (baja biomasa) y/o tiene baja
productividad (bajo recrecimiento). En el ámbito nacional los daños causados por chapulín
se estiman entre el 20 y 30 % de la producción cuando no se realizan acciones de control
(Cuellar, 1988 citado por García et al, 2000). Los estados más afectados son:
Aguascalientes, Chihuahua, Durango, Hidalgo, San Luis Potosí, Michoacán y Zacatecas

7
(García et al, 2000). Los daños en México son aproximados por que no se tiene un registro
de plaga de chapulín y cuantificación de daños en cultivos y pastizales.
Vegetación
Hewitt (1980) encontró que el florecimiento de las plantas era un indicador
relacionado con la eclosión del chapulín y que la temporada ideal para el control del
chapulín estaba asociada con el florecimiento de las plantas. Nerney (1957) encontró que
los habitas preferidos de las especies de chapulín que causan daño estaban asociados con
tierras pobres de pastizales dominadas por un bajo crecimiento de maleza y las poblaciones
de chapulín se incrementaban en los años con precipitación normal o arriba de lo normal y
disminuía en temporada de sequía. Por otro lado Capinera y Sechrist (1982) realizaron un
estudio de la asociación de plantas y el complejo chapulín en un campo experimental en el
norte de Colorado, donde encontraron 25 especies de chapulines, y un análisis de
correlación mostró que la subfamilia Cantantopinae mostraba una correlación de 0.739 y
la subfamilia Gomphocerinae una correlación de 0.691 con los pastizales.
Métodos espaciales y predictivos de plaga de chapulín
A fin de crear nuevas alternativas de solución al problema de predicción y control
de plagas de chapulín, muchos estudios se han realizado, relacionando el medio ambiente
con las densidades poblacionales de chapulín y otros explicando el fenómeno en base a los
datos históricos, por ejemplo Kemp (1987), utilizó cadenas de Markov de 2 estados para
predecir la probabilidad de plaga de chapulín en 56 condados de Montana. También,
Zimmerman (1999) utilizó el ArcView y aplicó cadenas de Markov de dos estados de los

8
mapas de infestaciones de chapulín del estado de Wyoming de 1944 a 1996 para predecir
dinámica de población futura. El sistema produjo mapas que eran capaces de hacer
probabilidades de predicciones anuales, duración de la infestación y probabilidades de
infestaciones por muchos años, las predicciones fueron muy cercanas a los datos reales.
Por otro lado David et al. (1993) utilizaron tres modelos binomiales de las poblaciones de
chapulines de Wyoming y Nuevo México, los modelos fueron la función de densidad de la
probabilidad de Poisson y los otros fueron basados en los modelos Nachman y regresión
binomial Logit. . Los resultados indicaron que la etapa ninfal y el año muestreado no
influenciaron los parámetros del modelo pero el lugar si. Lo tres modelos fueron probados
para determinar el menor error para predecir densidades poblacionales, los resultados
mostraron que no había diferencias entre modelos pero que se seleccionaba el modelo de
Poisson por su simplicidad. Otros estudios relacionan la densidad de chapulín con otros
factores, por ejemplo, Scoot (1994) relacionó clases de frecuencia de plagas de chapulín
en un período de 31 años en Wyoming con lo más común (moda) de vegetación,
precipitación, altitud, evaporación, forma y tipo de suelo. También, obtuvo un modelo
lineal donde la variable dependiente es años de infestación y las variables independientes
son; pendiente, profundidad y erosión del suelo, con (P<.001, r2=0.885). En otra orden de
ideas, Skinner y Child (2000) realizaron regresión múltiple y utilizaron el análisis
discriminante por pasos como una técnica de selección para reducir el número de variables.
Esto fue para identificar las variables de suelo y clima que mejor caracterizaron las
densidades poblacionales de chapulín, en el estado de Colorado en el período de 1995 y
1996. Se encontró en el análisis discriminante que la densidad de chapulín en 1995 fue la
más importante seguida por la precipitación de abril de 1993 y la temperatura media de

9
octubre del mismo año. Además, se encontró que las características del suelo no son
determinantes en los cambios de densidad de chapulín, lo anterior indica que existe
causalidad entre las variables climatológicas y la densidad del chapulín. En otros estudios,
(Anderson, 1973) relaciona la vegetación de sitios asociados con chapulín, Begon (1983)
relaciona la población de chapulín con el clima y Hilbert (1983) estudia la migración del
chapulín. También se han realizado estudios para obtener mapas de distribución con el uso
del programa GIS como, Bryceson (1989) en Australia uso datos multiespectrales de
Landsat para localizar camas de huevecillos de Langosta Australiana (Chortiocetes
terminifera Walker) y en Africa Tappan et al. (1991) utilizaron un proceso similar para
obtener mapas donde es más posible que chapulines y langostas se apareen, eclosionen y se
desarrollen. Con respecto al análisis de similitud se han hecho estudios en otras áreas sin
embargo en chapulines se han hecho pocos intentos.
Análisis de similitud
Para realizar un análisis de similitud primero se define la Unidad Taxonómica
Operacional (UTO) que pueden ser sitios, especies de chapulines o de plantas y luego las
características de las UTO. Los datos de las características se estandarizaran, restándole a
cada característica la media y dividiendo por la desviación estándar para evitar
ponderaciones. (Rohlf,1988).
Una vez estandarizados los datos se obtendrán las distancias entre UTO’s usando la
distancia taxonómica (Rohlf,1988).
σXxz i −=

10
y se construye la matriz de similitud con las distancias⎥⎥⎥
⎦
⎤
⎢⎢⎢
⎣
⎡
nnn
n
dd
dd
........
...
1
111
,
Se utilizara el método UPGA (Sneath, 1973) para obtener la similitud entre grupos
de UTO.
Componentes principales
Una vez que se ha encontrado grupos diferentes con la técnica de agrupamiento, se
utilizara la técnica estadística para verificar los resultados del análisis de similitud y el
desarrollo estadístico es el siguiente;
Sea X un vector aleatorio de n componentes donde n es el número de variables que
afectan la población de chapulín y β un vector de n componentes tal que β´β=1. La
varianza de β´X es
E(β´X)2 = E(β´XX´β´) =β´Σβ (1)
∑=
−=n
Kjkikij zz
nd
1)(1

11
Para determinar la combinación lineal β´X con máxima varianza, debemos
encontrar un vector β que satisfaga β´β = 1 y que maximicé (1).
Sea = β´∑β - λ(β´β-1) (2)
Donde λ es un multiplicador de Lagrange. El vector de derivadas parciales es
∂φ/∂β = 2Σβ - 2λβ (3)
Ya que β´Σβ y β´β tienen derivadas en toda la región que contiene β´β=1
(Anderson, 1985) un vector β que maximicé β´Σβ debe satisfacer que la expresión (3) sea
igual a cero; estos es
(Σ - λI)β = 0. (4)
Para obtener la solución de (4) con β´β=1 se debe tener Σ - λI singular en otra
palabras λ debe satisfacer |Σ - λI| = 0, esta función es un polinomio en λ de grado n. Si se
multiplica (4) en la izquierda por β¨ se obtiene β´Σβ = λβ´β = λ. Esto muestre que la
varianza es λ. Entonces para la máxima varianza se usa la raíz mas grande de λ en (4),
(Anderson, 1985). Luego se grafica el primer contra el segundo componente para verificar
si realmente existen estos grupos.

12
DEFINICION DEL PROBLEMA
México tiene aproximadamente 113.7 millones de has. bajo condiciones de clima
árido y semiárido, lo cual equivale a 58% de su superficie. La mayor parte de esta
superficie se encuentra utilizada como agostadero. Los pastizales predominantes son
perennes como el zacate Bouteloua gracilis, Bouteloua cultipendula y cola de zorra
(Pamanes, 1997). En México de las plagas de insectos la de mayor pérdida económica en
cultivos y pastizales es la del complejo chapulín. Los Estados que registran las mayores
pérdidas son: Tlaxcala, Durango, Chihuahua, San Luís Potosí, Puebla, Michoacán,
Zacatecas, Aguascalientes, Hidalgo y Veracruz.
Del territorio del estado de Durango (12,320,000 ha.), el 54%, posee un uso de suelo
de agostadero es decir 6,656,089 de has.. En su mayor parte, la historia del
aprovechamiento de estas tierras se resume en el completo agotamiento de recursos por la
falta de atención, sobrepastoreo, incendios, sequías y por el ataque de insectos entre los que
destaca para la entidad la plaga del chapulín. Para el semidesierto y los llanos de Durango
se tienen registradas 48 especies de chapulín donde destacan: Brachystola magna,
Boopedon nubilum, Taenipoda eques, y el complejo melanoplus que esta compuesto por al
menos 8 especies entre las que destacan M. differentialis, M. femurrubrum y M. thomasi
M. lakinus, P. nebrascensis (Rivera, 2000). En 1995 y 1996 el chapulín causó pérdidas
económicas en cultivos de fríjol de temporal del estado de Durango en los municipios de
Canatlán, San Juan del Río, Guadalupe Victoria, Villa Unión, Vicente Guerrero y Durango
(García, 1988), y se han reportado daños del 12% (36,000ha.) de la superficie total (Varela,

13
2000). De acuerdo con la información proporcionada por el departamento de sanidad
vegetal de SAGARPA y los Distritos de Desarrollo rural de Durango y la información
vertida en el simposio “Control de Plaga de Chapulín en Durango”, se estima que de no
tomar una acción oportuna y adecuada contra el complejo chapulín existirán pérdidas del
20 al 30 % y aun mayores en los cultivos de maíz, fríjol, avena y principalmente en
pastizales

14
JUSTIFICACION
En Durango el ciclo agrícola 1999 la superficie sembrada fue de 694,184 has.,
donde 321,352, 204,236 y 72,835 has. fueron de fríjol, maíz y avena respectivamente,
además se dedicaron 5,656,089 has. de pastos y praderas para la ganadería.. La región de
los llanos que es una región de pastizales, y cultivos, comprende los municipios de
Durango, Canatlán, P. Coronado, Gpe. Victoria, Nombre de Dios, Vicente Guerrero,
Poanas. En el ciclo agrícola 1999 la superficie sembrada de cultivos cíclicos fue de 262,858
has., donde 173,623 has. fueron fríjol y 48,398 has. de maíz y se dedicaron 802,924 has. de
pastos y praderas a la ganadería (INEGI, 2000), considerando que la producción promedio
de fríjol y de maíz de temporal por ha. es de 300 y 800 Kg. respectivamente. Entonces la
producción de fríjol y maíz en los municipios mencionados es de 52,086 y 38,718
toneladas. Entonces una pérdida de 20 al 30% de la producción por concepto de plaga de
chapulín, significa una perdida 10,572 a 15,625 toneladas de fríjol y de 7,743 a 11,615
toneladas de maíz. Considerando que la pérdida de pastos debido al complejo de chapulín
se estima en 600 libras en peso seco por acre, en Arizona (Loockwood, A. et al 1988-92
“Boopedon nubilum fact sheet”), una pérdida similar equivaldría a 600 kg. Por ha.,
entonces se tendrá una pérdida de 481,754 toneladas de pasto ante una plaga de chapulín en
estos municipios. El estado de Durango no cuenta con estudios ni antecedentes confiables
sobre la dinámica poblacional del chapulín razón por la cual no se tiene parámetros
establecido para tomar medidas de control de las plagas de chapulín. El presente estudio
pretende conocer por medio de muestreos y del análisis de similitud las especies de plantas
que se asocian con las especies mas abundantes de chapulín en los pastizales de Durango.

15
HIPOTESIS
La asociación de las especies mas abundantes de chapulín en los pastizales de
Durango con las especies vegetales se puede realizar con el análisis de similitud

16
OBJETIVO
Determinar la asociación de las especies mas abundantes de chapulín en los
pastizales de Durango con las especies vegetales con el análisis de similitud
OBJETIVOS PARTICULARES
Crear base de datos del complejo chapulines y tipos de vegetación del área de
estudio. Realizar análisis de similitud del complejo chapulín y la vegetación.

17
DELIMITACION TIEMPO Y ESPACIO
El estudio se va a realizar en 35 sitios de muestreo comprendidos entre la latitud 23
- 25 y longitud 104 – 105, se realizaran 4 muestreos que se llevaran a cabo de Agosto a
noviembre del 2007.

18
METODOLOGÍA
El estudio se realizó en la zona de pastizales del estado de Durango que comprende
los municipios de Durango, Canatlán, Panuco de Coronado, Gpe. Victoria, Nombre de
Dios, Vicente Guerrero, Poanas, San Juan del Rió, Rodeo, San Pedro del Gallo, San Luís
del Cordero, Nazas y parte de Peñón Blanco, estos municipios están localizados en la
latitud (23.916°, 25.983°) y longitud W (104.997°, 104.010°) aproximadamente. Se
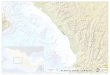
establecieron en los pastizales del área de estudio 35 sitios de muestreo (Figura 1. y Tabla
1.) es decir un sitio cada 8 Km. y utilizando el criterio del hábitat, el cual consiste según
Berry et al., (2000), en que cuando los hábitats o poblaciones son homogéneas durante
grandes distancias, la distancia entre puntos de muestreo puede ser mayor a ocho km. sin
detrimento de la calidad del análisis.
Figura 1. Distribución geográfica de los sitios de muestreo en los pastizales del estado de Durango.

19
Tabla 1. Coordenadas geográficas de los sitios de muestreo en los pastizales del estado de Durango.
Localidad Municipio Altitudmsnm
Lat. N Grados
Min. Seg. Long. W Grados
Min. Seg.
27 de Nov. Durango 1869 24 12 54.3 104 29 38.3 Carlos Real Durango 1871 24 16 17.2 104 26 16.9
Castillo Nájera Canatlán 1953 24 20 31.4 104 29 41.6Las Huertas P. de Coronado 1970 24 16 7.07 104 17 22.9* Madero P. de Coronado 1937 24 26 16.2 104 17 27.9 L Rivera Gpe. Victoria 1985 24 27 7 104 11 4
Rodríguez I Gpe. Victoria 1994 24 18 31 104 4 50 Zarco I Gpe. Victoria 2138 24 13 29.5 104 3 57.7
Zarco- Cieneguilla Poanas 2111 24 05 45.2 104 2 11 Berros - Saltito Poanas 1835 23 56 39.9 104 17 4.7 Berros - Tuitán Poanas 1854 23 58 1.9 104 16 22.1 18 de Agosto Poanas 2093 23 57 47.8 104 9 45.8
Zaragoza Poanas 1873 23 51 54.4 104 9 21 El Venado Nombre Dios 1743 23 51 56.3 104 16 51 La Ermita Nombre Dios 1930 23 53 14 104 19 56.2
Entrada Guadíana Durango 1809 23 59 46 104 22 33.4 Morcillo Durango 2001 24 6 43 104 42 17.2 Km 36 Durango 1923 24 15 57.6 104 41 57.2
Nicolás Bravo Canatlán 1920 24 23 4.6 104 41 44.9Km 86 Canatlán 1950 24 42 07.0 104 38 28.5Km 130 Canatlán 1747 24 59 36.8 104 30 35.3Km 153 Rodeo 1341 25 06 59.5 104 31 51.6Km 214 Rodeo 1562 25 30 51.6 104 44 30.7Km 188 Rodeo 1525 25 22 51.8 104 36 43.7Km 237 Hidalgo 1895 25 45 47.1 104 44 55.8Km 260 Hidalgo 1941 25 57 22.3 104 53 29.2Km 104 Hidalgo 1986 25 47 27.8 104 36 25.9Km 84 SPG 2011 25 49 02.3 104 25 11.5Km 76 SPG 1788 25 40 21.8 104 16 35.6Km 61 SPG 1643 25 32 42.2 104 17 32.6Km 29 SLC 1369 25 21 22.3 104 09 38.6Km 51 Nazas 1264 25 12 26.2 104 09 19.6Km 73 Nazas 1301 25 08 0 104 19 21.9Km 23 P. de Coronado 2128 24 30 52.8 104 27 22.3 Km 43 P. de Coronado 1926 24 39 14 104 28 13.1

20
Chapulín
Se realizarán 4 muestreos a partir de Agosto hasta Noviembre del 2007 35 sitios,
Tabla 1. Figura 1. Para la captura del complejo chapulín, se realizarán ochenta barridos de
red entomológica de un diámetro de 40 cm., por fecha de muestreo y por sitio, los
chapulines capturados se depositarán en frascos de plástico de 250 ml., con un contenido de
70 % de alcohol y 30 % de agua. Los especimenes se clasificacaranpor especie con la
ayuda de la clave del Departamento de Entomología de la Universidad Estatal de Nuevo
México (Richman, et al 1993), la clave del Departamento de Entomología de la
universidad Estatal de Colorado (Capinera y Sechrit,1982) y el sitio en Internet del
Departamento de Entomología de la Universidad Estatal de Wyoming
(http://www.sdvc.uwyo.edu/grasshopper/oeen.htm), y del Departamento de Entomología de
la Universidad Estatal del Norte de Dakota
(http://www.ndsu.nodak.edu/entomology/hopper/orthoptera_index.htm),
Se determinara la cantidad de chapulines por especie en el cuarto y quinto estado ninfal y
la etapa adulta.
Topografía
En cada sitio de muestreo se determinó la altitud, longitud y latitud por medio de un
geoposicionador satelital (GARMIN, eTrex personal navigator, 2000). Clima, suelo,
vegetación y altitud Se obtuvieron del sitio de Internet de CONABIO
http://conabioweb.conabio.gob.mx/ los mapas vectoriales de la República Mexicana de tipo
de suelo (SEMARNAP, Subsecretaria de Recursos Naturales, 1998), tipo de vegetación
(Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO), 1999),

21
isotermas (García y CONABIO, 1998) y precipitación total anual (García y CONABIO,
1998), la escala del mapa de tipo de suelo fué de 1:4 000 000 y el resto de 1:1 000 000.
Se obtuvieron del sitio de Internet de la Universidad de Michigan
http://glcf.umiacs.umd.edu/ los mapas de índice de vegetación, porcentaje herbáceas y
suelo desnudo; estos dos últimos mapas se generaron usando imágenes MODIS, con el
algoritmo de campos de vegetación continua, con el propósito de eliminar las nubes y las
sombras de las nubes, de cada mes con 500 metros de resolución, (Hansen et al., 2002,
2003).
El mapa de índice de vegetación de 250 metros de resolución se formó al tomar la
diferencia de la banda cerca de infrarrojo (0.7 a 1.1 mm., detecta la forma de la planta) y del
rojo (de 0.4 a 0.7 mm., lo absorbe la clorofila) entre la suma de ellos por 200 más 50 y se
usaron 16 días continuos con el fin de eliminar las nubes.
Finalmente del sitio de la Admisnistración Nacional de la Atmosfera y el Océano de
EU http://www.ngdc.noaa.gov/ se obtuvo el mapa de elevación del área de estudio con
proyección UTM y elipsoide WGS-84 se transformó a la proyección Cónica conforme de
Lamber y utilizando los comandos EXPAND y CONTRACT se logró que todos los mapas
tuvieran una resolución de 150x300 píxeles y el área de estudio quedo en la latitud
(24.0478947, 26.0035961) y longitud W (104.9476515, 103.997292) cada píxel en los
renglones es de 0.006519 grados de latitud y en las columnas de 0,006335 grados de
longitud .
De los 35 sitios de muestreo se obtuvieron los datos de longitud (grados); latitud
(grados); altitud (msnm); pendiente (porcentaje); temperatura (°C); precipitación (mm.);

22
tipo de vegetación; donde la escala se consideró tomando en cuenta el área de la cobertura
aérea de la vegetación (Bosque=6, Agricultura=5, Pastizal mediano abierto=4, Pastizal
amacollado y halófilo=3, Arbusto medíano=2, Arbusto bajo=1 y Ciudades y cuerpos de
agua=0); tipo de suelo; donde la escala se considero tomando en cuenta la cantidad de
materia orgánica del tipo de suelo (Ciudades y cuerpos de agua=0, Rendzina=1, Feozem=2,
Litosol=3, Fluvisol=4, Castañozem=5, Regosol=6, Vertisol=7, PLanosol=8, Xerosol=9,
Solonchartico=10, Yermosol=11, Chernozem=12, Cambisol=13 y Luvisol=14); índice de
vegetación ; porcentaje herbáceas y porcentaje de suelo desnudo para cada uno de los sitios,
la clasificación numérica en tipo de vegetación y suelo fué con el propósito de realizar el
análisis de similitud.
Vegetación
En cada sitio de muestreo se registraran parámetros como cobertura basal de las
gramíneas, la frecuencia, la distribución y variación de las especies utilizando la Línea
Canfield y la clasificación de vegetación de (Krebs, 1989 y COTECOCA, 1979) para la
determinación del tipo de vegetación se pedirá del apoyo del Herbario del CIIDIR
Durango.
Los datos colectados en cada sitio se copiarán en una ficha especial que contenga
los tipos de vegetación y se capturarán en una hoja de cálculo. Una vez que se tenga la
información de un año se realizarán los siguientes análisis estadísticos

23
Análisis de similitud
Primero se realizara un analisis de similtud en donde las UTO son los sitios de
muestreo y las características son los valores de los factores ecológicos, la presencia de las
especies de chapulín y de la especies de plantas. Segundo, se relizara un análisis en donde
las UTO serán las especies de chapulín y las caracteristicas serán los sitios donde estan
presentes. Finalmente se realizara un análisis donde las UTO serán las especies vegetales y
las características serán los sitios y la presencia de especies de chapulín Sneath, (1973).
Posterioremente se realizara el análisis de los componentes principales para contrastar con
el análisis de similitud.

24
RESULTADOS
Datos de campo
De los datos de los Sistemas de Información Geográfica se obtuvieron los datos
ecológicos de clima, suelo, vegetación y topogrfía (Tabla 2). De los muestreos de el
complejo chapulin se obtuvieron 45 especies (Tabla 3) de las lineas Canfield se obtuvieron
85 especies vegetales (Tabla 4).
Tabla 2. Localidad de los sitios de muestreo y los valores de las variables ambientales. Sitio Longitud Latitud Alt Pen Tem Pp Veg Ivn Herb Sd S Nicolás Bravo -104.6958 24.3846 1936 0 15 550 4 204 76 14 7 L Rivera -104.2615 24.4043 2019 2 17 450 4 201 61 34 3 La Ermita -104.3322 23.8872 1919 2 17 550 4 201 61 34 6 Madero -104.291 24.2711 1987 1 17 450 4 198 68 28 3 Castillo Nájera -104.4948 24.342 1940 2 17 450 4 197 71 17 7 Km 188 -104.6121 25.381 1525 2 17 450 2 194 80 18 9 Km 130 -104.5098 24.9935 1747 7 17 450 2 185 73 25 1 Las Huertas -104.2896 24.2686 1956 2 17 450 4 182 73 22 9 18 de Agosto -104.1513 23.95 1866 0 17 450 3 181 79 33 3 El Venado -104.2808 23.8656 1756 2 17 450 4 181 79 33 9 Km 23 -104.4561 24.5146 2128 2 15 550 4 171 59 28 2 Km 73 -104.3227 25.1333 1300 13 17 250 4 166 72 21 3 Rodríguez I -104.0916 24.31664 2060 1 17 450 4 165 69 32 1 Berros - Tuitán -104.2728 23.9671 1873 0 17 450 4 165 66 36 3 27 de Nov. -104.4939 24.215 1871 0 17 450 3 164 73 37 3 Km 86 -104.6412 24.7019 1950 2 17 450 4 163 70 29 3 Morcillo -104.7047 24.1119 2001 1 17 550 4 162 74 30 9 Km 43 -104.4703 24.6538 1926 2 15 550 4 162 64 34 3 Zarco- Cieneguilla -104.0363 24.0958 2132 6 17 450 4 157 62 29 1 Berros - Saltito -104.2846 23.9444 1810 1 17 450 4 154 66 37 9 Km 36 -104.6992 24.266 1911 0 15 550 4 152 72 23 2 Zaragoza -104.1558 23.8651 1866 1 17 450 3 152 72 23 3 Entrada Guadíana -104.3396 23.9522 1910 0 17 450 4 152 67 38 8 Carlos Real -104.438 24.2714 1872 1 17 450 3 151 63 34 7 Km 153 -104.531 25.1165 1341 2 19 350 1 146 59 42 1 Km 51 -104.1554 25.2072 1264 6 21 250 2 135 49 51 3 Km 29 -104.1607 25.3561 1369 0 21 250 1 121 70 28 9 Km 237 -104.7488 25.763 1895 1 17 350 2 114 68 31 1 Km 260 -104.8914 25.9561 1941 1 17 350 2 110 65 34 3 Km 84 -104.4198 25.8173 2011 2 19 350 1 98 45 55 9 Zarco I -104.066 24.2248 2139 1 17 450 2 74 66 33 1 Km 61 -104.2923 25.545 1643 1 21 250 1 72 57 42 6 Km 104 -104.581 25.791 1986 2 17 350 1 70 67 32 9 Km 76 -104.2765 25.6727 1788 2 19 250 1 59 44 56 9
Alt=Altitud, Pen=Pendiente, Tem=Temperatura, Pp=Precipitación, Veg=Tipo de vegetación, Ivn=Índice de vegetación normal, Herb=Porcentage de herbáceas, Sd=Porcentage de suelo desnudo y S=Tipo de suelo.

25
Los datos se capturaron en Excel y se pasaron al paquete estadístico STATISTICA
7 donde los datos de los renglones eran las características y las columnas las UTO. Los
datos se estandarizaron, restándole a cada característica la media y dividiendo por la
desviación estándar para evitar ponderaciones. Para obtener la matriz de distancia se utiliza
la distancia euclidiana (Sneath y Sokal,, 1973) y con esto se obtuvo una matriz diagonal de
distancias con las diagonales en cero debido a que es la distancia que hay de un sitio en
relación con el mismo. El siguiente paso fue obtener un dendrograma de sitios utilizando
el método UPGA (Sneath y Sokal,, 1973 ). Posteriormente se realizo un análisis de
componentes principales considerando los componentes que aportaran la mayor variancia el
primeroy el segundo.
Análisis de similitud y componente principales
Considerando los sitios como UTO se obtuvo el siguiente dendrograma

26
Figura 2. Dendrograma de sitios de muestreo.

27
Figura 3.- Primer contra segundo componente principal por sitios.
En el dendrograma por sitios Figura 2., se encontró que se divide en dos ramas
básicas, la rama de arriba se divide en tres subramas y encontramos en la primera a una
distancia de 1.7, los sitios de 18 de Agosto, Carlos Real, Entrada Guadiana y Librado
Rivera, Los cuales se encuentran en el área sur de estudio se encuentran entre una latitud de
1809 y 2093 m, y se encuentran entre la latitud 23° 57’ 8.2’’ y 24° 24’ 15.6’’ y la longitud
104° 3’ 15.5’’ y 104° 26’ 16.9’’, la vegetación que encontramos aquí es del tipo de
pastizales y tenemos las especies de el Melanoplus lakinus, Melanoplus sanguinippes, que
pertenecen a la subfamilia Catantopinae y al genero Melanoplus , los cuales causan mayor
daño a los cultivos y pastizales, además, encontramos algunas otras especies como
Psoloessa texana, Sphargemon humile de la subfamilia Oedipodinae y en menor
proporción de la subfamilia Gomphocerinae encontramos al Paropomala wyomigensis y

28
Opeia obscura. En esta subrama encontramos principalmente de la subfamilia
Catantopinae que se encuentra asociada básicamente a la vegetación de pastizales halófilo
y pastizal mediano arbosufrutescente en donde se encontraron las siguientes especies
Quelite (Heliantus palmeris), Romerillo (Bides leucantha), zacate mota (Chloris virgata),
Zacate navajita (Boutelova gracilis) y las mas predominantes son las dos ultimas.
En la siguiente subrama de la rama superior del dendrograma Figura 2., a una
distancia de 1.2, se encuentran una sub-subrama con los sitios Berros–Saltito, Berros–
Tuitan, Rodríguez, Madero km 43 y Zaragoza los cuales se encuentran entre una altitud de
1835 y 1994 m, latitud 23° 51’ 54.4’’ y 24° 39’ 14’’ y la longitud 104° 7’ 5.6’’ y 104° 28’
13.11’’, encontramos principalmente la especie del Trimerotropis Pallidipennis y
Sphargemon humile que pertenecen a la sub familia Oedipodinae y esto se debe
principalmente a la presencia de sitios de Matorrales. S encontraron escasamente algunas
otras especies como Melanoplus lakinus, Phoetaliotes nebrascencis y Melanoplus
sanguinipes de la subfamilia Catantopinae esto debido a la presencia de algunos sitios de
pastizales. En la primera rama, segunda subrama esta se encontró una sub-subrama a una
distancia de 1.6, donde están los sitios de Km 36, Km 86 y Morcillo los cuales se
encuentran entre una altitud de 1923 y 2001 m, latitud 24° 6’ 43’’ y 24° 42’ 7’’ y longitud
104° 38’ 28.5’’ y 104° 42’ 17.2’’, donde se encuentra las especies, Trimerotropis
Pallidipennis Melanoplus lakinus, Phoetaliotes nebrasencis que pertenecen y a la sub
familia Oedipodinae y y el genero melanoplus respectivamente, en estos sitios
encontramos la vegetación del tipo de pastizales mediano arbosufrutescente y abierto en
donde se encontraron las siguientes especies que son las mas predominantes zacate cola de

29
zorra (Enneapagon desvauxii), zacate flechita (Stipia neesiana), zacate mota (Chloris
virgata), zacate tres puntas (Aristidas adsencionis).
En la ultima subrama de la primear rama a una distancia de 1.5, encontramos los
sitios de Bermejillo km 84, Bermejillo km 104, km 237, km 214, km 260, Nazas km 51,
Nazas km 72 y SLC km 29 los cuales se encuentran entre una altitud de 1264 y 2001 m,
latitud 25° 9’ 37.8’’ y 25° 57’ 22.3’’ y longitud 104° 9’ 19.6’’ y 104° 53’ 29.2’’,
encontramos principalmente la especie del Trimerotropis Pallidipennis, Hippopedon
capito,Aulocara ellioti, que pertenecen principalmente a la subfamilia Oedipodinae esto es
debido a que tipo de vegetación es de matorrales. También se encontró de en cantidades
menores Melanoplus lakinus, Melanoplus sanguinipes que pertenecen a la subfamilia
Catantopinae y al genero melanoplus. La abundancia del los Oedipodinae en los matorrales
es debido a la facilidad de volar de estas especies pueden ir de un matorral a otro. La
vegetación asociada con estos sitios fue de Matorral Bajo subes pinoso y mediano espinoso,
en donde se encontraron las siguientes especies que son las mas predominantes Zárate mota
(Cloros virgaza), Zárate navajita (Boutelova gracilis),Gobernadora (Larrea tridentata),
zacate pajon (Sporobolus airoides).
.
En la rama inferior encontramos a una distancia de 3.0 los sitios de 27 de
Noviembre, Alamillo km 188, km 130, Nicolas Bravo, Zarco – Cieneguilla, El Venado, La
Ermita, Madero, SPG km 61, SPG km 76 y km 153 los cuales se encuentran entre una
altitud de 1643 y 2001 m, latitud 23° 51’ 14 ’’ y 25° 40’ 21.8’’ y longitud 104° 16’ 35.66’’
y 104° 41’ 44.9’’. Se encontraron principalmente la especie Melanoplus lakinus,

30
Phoetaliotes nebrascencis, Boopedon nubilum, , Trimerotropis Pallidipennis, Brachystola
magna, Tetigonido spp y Schystocercas spp, Estos pertenecen a la subfamilia Catantopinae,
Gomphocerinae, Oedipodinae, Familia Romaleida, Tetigonidae y Cirtacantacridinae
respectivamente, estos sitios están asociados con el tipo de vegetación de Matorrales
mediano espinoso y bajo subespinoso y también encontramos Pastizal mediano
arbosufrutescente en donde se encontró que las siguientes especies que son las mas
predominantes zacate tres puntas (Aristidas adsencionis) zacate navajita (Boutelova
gracilis), manto (Ipomea purpurea), huizache (Acacia tortusoso), gatuño (Mimosa
biuncifera).por lo que se dan las condiciones para esta variedad de especies. Y además son
de las que causan mayor daño a los cultivos.

31
Figura 4. Dendrograma de especies de chapulín muestreadas.

32
Figura 5.- Primer contra segundo componente principal por especies de chapulín.

33
En el dendrograma por especies Figura 4. encontramos dos ramas principales en la
primera rama podemos observar a una distancia de 4.0, especies como Melanoplus
lakinus, Boopedon nubilum, , Phoetaliotes nebrasencis,Trimerotropis Pallidipennis y
Tetigonido, estas especies son las que representan mayor densidad poblacional y están
asociadas a los sitios de muestreo donde la vegetación predominante son : zacate mota
(Chloris virgata), Zacate navajita (Boutelova gracilis), zacate cola de zorra (Enneapagon
desvauxii), zacate flechita (Stipia neesiana), zacate tres puntas (Aristidas adsencionis),
Huizache (Acacea tortuosa).
En la siguiente rama se observa que a su vez se divide en otras dos subramas la
primera a una distancia de 2.5 agrupa a las especies que se encuentran en menores
cantidades, aquí tenemos a las especies Taenipoda eques, Brachystola magna, Leprus
Wheeleri, , Aeloplides tumbulli, Opeia obscura, Mermiria bivittata, Mermiria picta,
Psoloessa texana, Bootettix argentatus, Melanoplus bivitattus, Dactylatum bicolor,
Trimerotropis incospicua, Trimerotropis pristinria, Schystocerca alutacea, Metator,
Melanoplus alas largas, Acrhlopitus, Syrbula admirabilis, Trimerotropis tolteca,
Chortophaga viridifasiciata, Arphia conspersa, Melanoplus occidentales, Arphia
Pseudonietana, Trimerotropis magnifica, Cordillacris, Ageotettix deorum, Trachyrhachys
aspera, Aeloplides tenuipenis, Trimeritropis fraterna, Tropidolophus formosus,
Paropomala wyomigensis, Hipoppedon capito ,Encoptholophus sordidus,Trimerotropis
latifaciata. Estas especies están asociadas con una mezcla entre pastizales y matorrales
donde las especies más importantes son: Árnica (Machaenthera pinnatifida), Chicoria
(Cichorium intybus), Gatuño (Mimosa biuncifera), Mal de ojo (Cymbopogon nardos),

34
Manto (Ipomea purpurea), Mezquite (Prosopis juliflora), Ocotillo(Fouquieria splendeis),
Sangre de grado(Jatropha dioica), Tasajillo(Opuntia leptocaulis), Zacate cola de zorra
(Enneapagon desvauxii), Zacate lobero (Lycurus phleoides), Zacate navajita(Boutelova
gracilis)Zacate popoton(Andropogon barbinodis), Zacate tres puntas(Aristidas
adsencionis).
.
En la segunda subrama de la segunda rama a una distancia de 3,0 encontramos las
especies Melanoplus sanguinipes, Aulocara ellioti, Conozoa texana, Sphargemon humile,
aquí se observa tenemos todo lo contrario a la primera subrama de la segunda rama rama,
aquí hay mas densidad pero principalmente de la subfamilia Oedipodinae y en menor
grado de la subfamilia Cantantopinae. Estas especies están asociadas principalmente a la
vegetación de tipo matorrales y escasamente pastizales donde las especies predominantes
son: Garambullo (Myrtillocactus geometrizans), Mancamula (Solanum rostratum),
Mezquitillo (Acacia constricta), Zacate mota (Chloris virgata), Zacate navajita (Boutelova
gracilis), Zacate pajon (Sporobolus tiroides).
Del análisis de componentes principales, Figura 3. se observa que los sitios Nazas
Km. 51, Nazas Km. 72, SLC Km. 29, Carr- Parral- Km. 153, Km. 214, se agrupan
alrededor de 0 en el primer componente y entre -600 y -200 en el segundo componente,
estos sitios coinciden a excepción de Km. 260, Km. 237, Bermejillo Km. 84, Bermejillo
Km. 104,con la subrama inferior de la rama superior del dendrograma por sitios Figura 2.

35
También se observa un grupo que se encuentra alrededor de -50 a 100 en el primer
componente y entre -200 hasta 400 en el segundo componente que es lo que corresponde a
los sitios de Nicolás Bravo, Madero, La Ermita, Castillo Nájera, Carlos Real, 27 de
Noviembre, Rodríguez, Librado Rivera, Madero Km. 23, Zarco, Zarco – Cieneguilla, Km.
130, SPG Km. 61, SPG Km. 76, coinciden con los sitios de la segunda rama del
dendrograma por sitios Figura 2, con excepción de Km. 188, El venado, Km. 153.
Se observa que un grupo se encuentra de 0 a -100 en el primer componente y de -
100 hasta 400 en el segundo componente que es lo correspondiente a los sitios de Entrada
Guadiana, Berros – Saltito, Berros – Tuitán, Km. 237, Las Huertas, Morcillo, 18 de Agosto,
Madero Km. 43, Bermejillo Km. 104, Bermejillo Km. 84, Km. 260, Km. 36, coinciden con
los sitios de la subrama superior de la primera rama del dendrograma de sitios Figura 2.
con excepción de Carlos Real, Librado Rivera, Zaragoza, Castillo Nájera, Zarco.
Se nota que el agrupamiento utilizando el análisis de grupo y el análisis de
componentes Principales coinciden a excepción de algunos sitios esto se debe a las
condiciones similares de topografía entre ellos.
De el análisis de componentes principales por especies Figura 6. se nota una nube
de puntos que se encuentra de -10 a 5 en el primer componente y de -10 hasta 0 en el
segundo componente que son lo correspondiente a la subrama superior de la segunda rama
del dendrograma por especies Figura 5. encontramos especies Taenipoda eques,

36
Brachystola magna, Leprus Wheeleri, , Aeloplides tumbulli, Opeia obscura, Mermiria
bivittata, Mermiria picta, Psoloessa texana, Bootettix argentatus, Melanoplus bivitattus,
Dactylatum bicolor, Trimerotropis incospicua, Trimerotropis pristinaria, Schystocerca
alutacea, Metator, Melanoplus alas largas, Acrohlopitus, Syrbula admirabilis,
Trimerotropis tolteca, Chortophaga viridifasiciata, Arphia conspersa, Melanoplus
occidentales, Arphia Pseudonietana, Trimerotropis magnifica, Cordillacris, Ageotettix
deorum, Trachyrhachys aspera, Aeloplides tenuipenis, Trimeritropis fraterna,
Tropidolophus formosus, Paropomala wyomigensis, Hipoppedon capito ,Encoptholophus
sordidus,Trimerotropis latifaciata.
También se observa que se agrupan las especies de Melanoplus lakinus, Boopedon
nubilum, Trimerotropis pallidipennis, Phoetaliotes nebrascencis, Tetigonido, estos los
encontramos desde 55 hasta -25 en el primer componente y de -15 a 140 en el segundo
componente y totalmente separadas del grupo anterior estas se encuentran relacionadas con
la rama superior del dendrograma de especies Figura 5.
Finalmente encontramos las especies Melanoplus Sanguinipes, Aulocara ellioti,
Conozoa texana, Sphargemon humile, estos los encontramos entre 10 y 25 en el primer
componente y de 0 a 15 en el segundo componente esto se encuentra relacionado con la
subrama inferior de la segunda rama del dendrograma de especies Figura 5. por lo que se
puede observar que se encuentra solo un poco mas retirada de la nube de puntos que
encontramos en el primer análisis.

37
Figura 6. Dendrograma de especies vegetales muestreadas.
De las ramas que presentas mas homogenidad es la inferior donde se agrupan las
especies; Cempoalillo, B. leucantha, C. virgata, A. adscensionis, A. tortuosa, P. juliflora,
A. barbonodis, R. roseum, H. palmeris, O. leocotricha, B. gracilis. Las cuales estan
asociadas con las especies mas abundantes de este estudio. Esto se refleja en el análisis de
principales componentes en donde las especies mencionadas se agrupan en el rango de -1.0
a -0.6 en el primer componente y en el rango de 0 a 0.4 en el segundo componente.

38
Figura 7.- Primer contra segundo componente principal por especies vegetales.
CONCLUSIONES
De el análisis de similitud y componentes principales se concluye que las especies
Melanoplus lakinus, Boopedon nubilum, Trimerotropis pallidipennis, Phoetaliotes
nebrascencis, conviven en los mismos sitios y están asociadas con vegetación del tipo
Pastizal mediano abierto y mediano arbosufrutescente y se encuentran principalmente en
los municipios de Poanas, Panuco de Coronado, Canatlán, Durango y Guadalupe Victoria y
Nombre de Dios. También se concluye que la especie Trimerotropis pallidipennis se

39
encuentra distribuida en ambos tipos de vegetación, Pastizal y Matorral. Las especies
Taenipoda eques, Brachystola magna, Leprus Wheeleri, , Aeloplides tumbulli, Opeia
obscura, Mermiria bivittata, Mermiria picta, Psoloessa texana, Bootettix argentatus,
Melanoplus bivitattus, Dactylatum bicolor, Trimerotropis incospicua, Trimerotropis
pristinaria, Schystocerca alutacea, Metator, Melanoplus alas largas, Acrohlopitus, Syrbula
admirabilis, Trimerotropis tolteca, Chortophaga viridifasiciata, Arphia conspersa,
Melanoplus occidentales, Arphia Pseudonietana, Trimerotropis magnifica, Cordillacris,
Ageotettix deorum, Trachyrhachys aspera, Aeloplides tenuipenis, Trimeritropis fraterna,
Tropidolophus formosus, Paropomala wyomigensis, Hipoppedon capito ,Encoptholophus
sordidus,Trimerotropis latifaciata que pertenecen en su mayoría al la subfamila
Oedipodinae y Gomphocerinae estan asociadas con la vegetación Matorral bajo
subespinoso, mediano subespinoso mediano arbosufrutescente.

40
BIBLIOGRAFÍA
Anderson T. W. (1985), An introduction to Multivariate Statistical Analysis. New
York:JOHN WILEY& SONS.
Anderson, N. L. 1973. The vegetation of rangeland sites associated with some grasshopper
studies in Montana. Bull. 668. Bozeman, MT: Montana State University and Montana
Agricultural Experiment Stattion.
Begon, M. 1983. Grasshopper populations and weather; the effects of insolation on
Chorthippus brunneus. Ecological Entomology 8: 361-370.
Belovsky, G.E., J.A. Lockwood, and K. Winks. 2000. In, Cunningham G.L. and M. W.
Sampson. Ed. 1996-2000. Grasshopper Integrated Pest Managment User Handbook.
USDA.
Bryceson, K.P. 1989. The use of Landsat MSS data to determine the distribution of
locust eggbeds in the Riverina region of the New South Wales, Australia. International
Journal Remote Sensing, 10(11): 1749-1762.

41
Capinera, J. L. y Schrit, T. S. 1982. GRasshoppers (Acrididade) of Colorado, Identification,
Biology and mangement. Colorado State University. Experiment Station Fort Collins.
Bulletin No. 584S.
Capinera, J. L., and D. R. Horton. 1989. Geographic variation in effects of weather on grasshopper infestation. Entomological Society of America 18(1): 8-14.
CoTeCoCA. 1979. SARH Durango.
David, E. L., Lockwood, J. A., Kemp, W. P. and Nolan, M. A. 1993. Estimating Densities
of Grasshopper (Orthoptera Acrididae) Assemblages Using Binomial Sampling.
Entomological Society of America, 22(4), 733-742.
Edwards, R. L. 1965. The Influence of Soil Moisture and Soil Type on the Oviposition Behavior of the Migratory Grasshopper, Melanoplus sanguinipes (Fabricius). The Canadían Entomology 97: 401-409.
García-Gutiérrez, C., S. Álvarez Amador, E. Rivera-García, H. Medrano-Roldán, N. Aviña-
Martínez, R. González-Betancourt y R. Álvarez-Zagoya. 2000. Control Biológico de
chapulín: Situación actual y posibilidades de uso de Bauveria bassiana. Simposio Control
de plaga de chapulín en Durango (Bioecología y Métodos para su control). Durango,
Durango, México

42
García-Gutiérrez, C., E. Rivera-García, S. Álvarez Amador e H. Medrano-Roldán. 1988.
Detección y combate químico-biológico de plaga de chapulín en Durango. UBAMARI.
Revista Hispanoamericana de Ciencia y Tecnología. Pág. 73-81.
Glogoza, P.A. y Weiss M.J. 1997. Grasshopper biology and management. NDSU Extension
Service E-272.
Hewitt, G. B. 1980. Plant Phenology as a Guide in Timing Grasshopper Control Efforts on
Montana Rangeland. Journal of Range Management, 33(4), 297-299).
Hewitt, G. B.y Onsager, J. A. 1983. Control of grasshopper and rangeland in the United
State a perspective. Journal of Range Management 36: 202-207
Hilbert, D. W y.Logan J. A. 1983. A population system model of the migratory
grasshopper(Melanoplus sanguinipes ). In: Lauenroth, W. K.; Skogerbee, G. V.; Flug, M.,
eds. Analyses of ecologicalsystems: state of the art in ecological modeling. Amsterdam:
Elseiver Scientific Publishing: 323-334
INEGI, 2000. Anuario Estadístico Durango Edición 2000.
Isely, F. B. 1944. Correlation between mandibular morphology and food specificity in
grasshoppers. Ann. Entomol. Soc. Amer. 37:47-67.

43
Kemp, W. P. 1987. Probability of Outbreak for Rangeland Grasshoppers(Orthoptera:
Acrididae) in Montana: Application of Markovian Principles. Journal of Economic
Entomology. 80(6): 1100-1105.
Kevan, 1977. Rev. Soc. Ent. Argentina 36:3-28
Krebs, C. J. 1989. Ecological Methodology, New York: Harper Collins Publisher.
Lockwood, Jeffrey A. J. McNairy Timothy, C. Larsen, John, and Zimmerman, Kiana.
1988-1992. Distribution Atlas for Grasshopper and the Mormon Cricket in Wyoming,
University of Wyoming in Cooperation with USDA, APHIS, PPQ and the Cooperative
Agricultural Pest Survey Program.
McLeod, M.J. 1994. Grasshopper management for South Dakota winter wheat pr
Poducers. ExEx 8106 August 1994 Plant Science. SDSU Plant Science.
Nerney ,N. J. 1957. Grasshopper infestions in relaction to range condition. Abstract of
paper presented at the meeting of the Arizona section, America society of range
management. Show low, Arizona, July 30-31. 1957.
Onsager, J, A. 1977. Comparison of Five Métodos for Estimang Density of Rangeland
Grasshopper. Jornal of Economic Entomology. 70(2): 187-190.

44
Pámanes - Garcia S.D. 1997. Iniciación, desarrollo y finalización en la producción de carne
de bovino. FMVZ-UJED. 1-4.
Richman, D. B. Sutherland, C. A. y Ferguson, D. J. 1993. A Manual of the grasshoppers of
New Mexico; Orhtoptera:Acrididae and Romaleidae. Cooperative Exrension Service. New
Mexico State University. Handbook No. 7
Rodell, C. F. 1977. A Grasshopper model for grassland ecosystem. Ecology 58: 227-245.
Rohlf, F. J. 1988. Numerical Taxonomic and Multivariate Analysis System. New York:
Exeter Publishing, LTD.
Scott, P. S. 1994. Spatial Analysis of Ecological Factors Related to Grasshopper
(Orhtoptera: Acrididae) Population Dynamic in Wyoming. Master Degree thesis in
Entomology. University of Wayoming.
Skinner, K.M. y R.D. Child. 2000. Multivariate Analysis of the Factors influencing changes
in Colorado grasshopper abundance. Journal of Othoptera Research. 9:103-110.
Sneath, P. H. A. and Sokal, R. R. 1973. Numerical Taxonomy. London: Freeman.

45
Tappan, G.G., D.G. Moore, y W.I. Knausenberg. 1991. Monitoring Grasshopper and
Locust habitats in Sahelian Africa using GIS and Remote Sensing Technology.
International Journal of Geographical Information System, 5(1):123-135
Samietz, J., M. A. Salser, and H. Dingle. 2005. Altitudinal variation in behavioral
thermoregulation: local adaptation vs. plasticity in California. Journal of Evolutionary
Biology 18(4): 1087 p.
Uvarov, B Watts. P. 1977. Grasshoppers and Locusts. A handbook of general Acridology.
Vol. 2. Centre for Overseas Pest Research, London. 613 pp.
Zimmerman, K.M. 1999. A Spatial Model for Markov Chain Analysis of Grasshopper
Population Dynamics in Wyoming. Master Degree in Computer Science. University of
Wyoming.

46

47
GLOSARIO
Biota: Conjunto de seres vivos de una región determinada.
Eclosión: Acción de abrirse un huevo.
Gramíneas: Plantas monocotildeas con flores.
Hospederas: Plantas que son el habitad del chapulin.
Ninfales: Estados intermedios que se presentan en los insectos holometabolos entre las
fases de larva e imagio.
Olífagas: tienen un limite variado de comida de ciertas especies y plantas.
Polífagas: Dicese de los organismos que se nutren de alimentos diversos.
Zoogeográficos: con características geográficas similares.

48
Tabla 3. Especies de chapulín encontradas en los sitios de muestreo Km
260 Km 84
Km 104
Km 237
Km 76
Km 61
Km 188
Km 29
Km 51
Km 73
Km 153
Km 130
B. nubilum 0 0 0 1 0 0 0 0 0 0 1 1B. magna 0 0 0 0 0 0 1 0 0 1 0 0H. trifasciatus 0 0 0 0 0 1 1 0 0 0 1 0M. lakinus 0 0 1 0 1 1 1 0 1 1 1 1M. occidentalis 0 0 0 0 0 0 0 0 0 0 0 0O. obscura 0 0 0 0 0 0 0 0 0 0 1 1O. pelidna 0 0 0 0 0 0 0 0 0 0 1 0P. wyomigensis 1 0 0 0 0 0 0 0 0 0 1 0P. virgata 0 0 0 0 0 0 0 0 0 0 0 0Var10 0 0 0 0 0 0 0 0 0 0 0 0P. nebrascensis 0 0 0 1 0 0 0 0 0 1 0 0P. texana 0 0 0 0 0 0 1 0 0 0 0 0Pygmy 0 0 0 0 1 0 1 0 0 1 1 0T. fontana 0 0 0 0 0 0 0 0 0 0 0 0T. modesta 0 0 0 0 0 0 0 0 1 0 1 0T. maritima 0 0 0 1 0 0 1 1 1 0 0 1T. pallidipennis 0 0 0 0 0 1 0 1 1 0 0 0T. californica 1 0 1 0 1 1 1 1 0 0 1 0T. kiowa 0 0 0 0 0 0 0 1 1 0 1 1T. aspera 0 0 0 0 0 0 0 0 0 0 0 0C. conspersa 0 0 0 0 0 0 1 0 0 0 0 0O. enigma 0 0 0 0 0 0 0 0 0 0 0 0X. corallipes 0 0 0 0 0 0 0 0 0 0 0 1M. bivittatus 0 0 0 0 0 0 0 0 0 0 0 0P. delicatula 0 0 0 0 0 0 0 0 0 0 0 0T. equs 0 0 0 0 0 0 0 0 0 1 0 1M. angustipennis 1 0 0 0 0 0 0 0 0 0 0 0
D. bycolor 0 0 0 0 0 0 0 0 0 0 0 0A. deorum 0 0 0 0 0 0 0 0 0 0 0 0C. neglectus 0 0 0 0 0 0 0 0 0 0 0 0M. picta 0 0 0 0 0 0 0 0 0 0 0 0S. occidentalis 0 0 0 0 0 0 0 0 0 0 0 0M. bivittata 0 0 0 0 0 0 0 0 0 0 1 0T. formosus 0 0 0 0 0 0 0 0 0 1 1 1S. alutacea 0 0 0 0 0 0 0 0 0 0 1 0M. kinecotti 0 0 0 0 0 0 0 0 0 0 0 0M. sanguinipes 0 0 0 0 0 0 0 0 0 0 0 0M. plattei 0 0 0 0 0 0 0 0 0 0 0 0M. pakardii 0 0 0 0 0 0 0 0 0 0 0 0M. devastator 0 0 0 0 1 0 0 0 0 0 0 0M. dawsoni 0 0 0 0 0 0 0 0 0 0 0 0S. arborea 0 0 0 0 0 0 0 0 0 1 0 0C. viridifasciata 0 0 0 0 0 0 0 0 0 0 0 0E. sordidus 0 0 0 0 0 0 0 0 0 0 0 0A. turnbulli 0 0 0 0 0 0 1 0 0 0 1 1

49
Continuación Tabla 3. Km
130 Km 86
Km 43
Km 23
L Rivera
N. Bravo
C. Najera Rodr C.
Real Madero
B. nubilum 1 1 1 1 1 1 1 0 1 1B. magna 0 0 0 1 1 1 1 0 0 1H. trifasciatus 0 0 0 1 0 0 0 0 0 1M. lakinus 1 1 1 1 1 1 1 1 1 1M. occidentalis 0 0 0 0 0 0 1 0 0 1O. obscura 1 0 0 0 0 0 1 1 1 1O. pelidna 0 0 0 0 0 0 0 0 0 0P. wyomigensis 0 0 0 0 1 0 1 0 1 1P. virgata 0 0 0 0 0 0 0 0 0 0Var10 0 0 0 0 0 1 0 0 0 0P. nebrascensis 0 1 1 1 1 1 1 0 1 1P. texana 0 1 0 0 0 1 0 1 1 1Pygmy 0 1 1 1 1 0 0 1 0 1T. fontana 0 0 0 0 0 1 0 0 0 0T. modesta 0 0 0 0 0 0 0 0 0 0T. maritima 1 0 0 1 0 0 0 0 0 0T. pallidipennis 0 0 0 0 0 0 0 0 0 0T. californica 0 1 0 0 0 0 0 1 1 0T. kiowa 1 1 0 1 0 0 1 1 1 0T. aspera 0 0 0 0 0 0 0 0 0 0C. conspersa 0 0 0 0 0 0 0 0 0 0O. enigma 0 0 0 0 0 0 0 0 0 0X. corallipes 1 0 0 0 0 0 0 0 0 0M. bivittatus 0 1 0 0 1 0 0 0 1 0P. delicatula 0 0 0 0 0 0 0 0 1 0T. equs 1 1 0 0 0 0 0 0 0 0M. angustipennis 0 0 0 0 0 0 0 0 0 0
D. bycolor 0 1 0 0 0 1 1 0 0 1A. deorum 0 0 0 0 0 0 0 0 0 0C. neglectus 0 0 0 0 0 0 0 0 0 0M. picta 0 0 0 0 0 0 0 0 0 0S. occidentalis 0 0 0 0 0 0 0 0 0 0M. bivittata 0 0 0 0 0 0 0 0 0 0T. formosus 1 1 0 0 0 0 0 0 0 0S. alutacea 0 0 0 0 0 0 0 0 0 0M. kinecotti 0 0 0 0 0 0 0 0 0 0M. sanguinipes 0 0 0 0 0 0 0 0 0 0M. plattei 0 0 0 0 0 0 0 0 0 0M. pakardii 0 0 0 0 0 0 0 0 0 0M. devastator 0 0 0 0 0 0 0 0 0 0M. dawsoni 0 0 0 0 0 0 0 0 0 0S. arborea 0 0 0 0 0 0 0 0 0 0C. viridifasciata 0 0 0 0 0 1 0 0 0 0E. sordidus 0 0 0 0 0 1 0 0 0 0A. turnbulli 1 0 0 0 0 0 0 0 0 0

50
Continuación Tabla 3. Las
Huer Km 36
Zarco
27-Nov
Morcillo
Zar-Cien
Berr-Tuit
E Guad
18-Ago
Berr-Salt
La Erm
El Ven
Zar
B. nubilum 0 0 1 1 1 1 0 1 1 1 1 1 1B. magna 0 0 0 1 1 1 0 1 1 0 1 1 0H. trifasciatus 0 0 0 0 0 0 0 0 1 0 0 0 0M. lakinus 0 1 1 1 1 1 0 1 1 1 1 1 1M. occidentalis 0 0 0 0 0 0 0 0 1 0 0 0 0O. obscura 0 1 1 0 0 0 1 1 1 1 1 1 1O. pelidna 0 0 0 0 0 1 0 1 1 0 0 1 0P. wyomigensis 0 1 1 1 0 0 1 1 1 0 0 0 1P. virgata 0 0 0 0 0 0 0 1 0 0 1 0 1Var10 0 0 0 0 0 0 0 0 0 0 0 0 0P. nebrascensis 0 1 0 1 1 1 0 1 1 1 1 1 1P. texana 1 0 1 0 1 1 1 1 1 1 1 1 1Pygmy 0 0 1 1 1 0 1 0 1 1 1 1 0T. fontana 1 0 0 0 0 1 1 1 1 0 1 1 0T. modesta 0 0 1 0 1 0 1 0 1 1 1 1 1T. maritima 1 0 0 0 0 1 0 1 1 1 0 0 0T. pallidipennis 1 0 0 0 0 0 0 0 0 0 0 0 0T. californica 0 1 0 0 0 0 1 1 0 0 1 1 1T. kiowa 1 0 1 0 0 1 1 1 0 1 1 1 1T. aspera 1 0 1 1 0 0 0 0 0 1 0 0 0C. conspersa 0 0 0 0 0 0 0 0 0 0 1 0 0O. enigma 0 0 0 0 0 0 0 0 0 1 0 0 0X. corallipes 0 0 0 0 0 0 0 1 0 1 1 1 0M. bivittatus 0 0 0 0 0 0 0 0 0 0 0 0 0P. delicatula 0 0 0 0 0 0 0 0 0 0 0 0 0T. equs 0 0 0 0 0 0 0 0 0 0 1 0 0M. angustipennis 0 0 0 0 0 0 0 0 0 0 0 0 0
D. bycolor 0 0 0 0 1 1 0 0 0 0 0 0 0A. deorum 0 0 0 0 0 0 0 0 0 0 0 1 0C. neglectus 0 0 0 0 0 0 0 0 0 0 1 1 0M. picta 0 0 0 0 0 0 0 0 0 0 0 1 0S. occidentalis 0 0 0 0 0 0 0 0 0 0 0 1 0M. bivittata 0 0 0 0 0 0 0 0 0 0 0 0 0T. formosus 0 0 0 0 0 0 0 0 0 0 0 0 0S. alutacea 0 0 0 0 0 0 0 0 0 0 0 0 0M. kinecotti 0 0 0 0 0 0 0 0 0 0 1 0 0M. sanguinipes 0 0 0 0 0 0 0 0 0 0 1 0 0M. plattei 0 0 0 0 0 0 0 0 0 0 1 0 0M. pakardii 0 0 0 0 1 0 0 0 0 0 0 0 0M. devastator 0 0 0 0 0 0 0 0 0 0 0 0 0M. dawsoni 0 0 0 0 0 0 0 0 0 0 0 0 1S. arborea 0 0 0 0 0 0 0 0 0 0 0 0 0C. viridifasciata 0 0 0 0 0 0 0 0 0 0 0 0 0E. sordidus 1 0 0 0 0 0 0 0 0 0 1 0 0A. turnbulli 0 0 0 0 0 0 0 0 0 0 1 0 0

51
Tabla 4. especies vegetales encontradas en los sitios de muestreo.
B.
adorata Agrito
nc. Artemisia
spp. Amole
nc Arbusto
desc M.
pinnatifida Bisnaga
nc Cabezona
nc Km 260 0 1 0 0 1 0 0 0Km 84 0 0 0 0 0 0 0 0Km 104 0 0 0 0 0 0 0 0Km 237 0 1 0 0 0 0 0 0Km 76 0 0 0 0 0 1 0 0Km 61 0 0 0 0 0 0 0 0Km 188 0 0 0 0 0 0 1 0Km 29 0 0 0 0 0 1 0 0Km 51 0 0 0 0 0 0 0 0Km 73 0 0 0 0 0 1 0 0Km 153 0 0 0 1 0 0 0 0Km 130 1 0 0 0 0 0 0 1Km 130 0 0 0 0 0 0 0 0Km 86 0 0 0 0 0 0 0 0 Km 43 0 0 0 0 0 0 0 0 Km 23 0 0 0 0 0 0 0 0 L Rivera 0 0 0 0 0 1 0 1 N. Bravo 0 0 0 0 0 0 0 0 C. Najera 0 0 0 0 0 0 0 0Rodr 0 0 0 0 0 0 0 0 C. Real 0 0 0 0 0 0 0 0 Madero 1 0 0 0 0 0 0 0Las Huer 0 0 0 0 0 0 0 0 Km 36 0 0 0 0 0 0 0 0 Zarco 0 0 0 0 0 0 0 027 No 0 0 0 0 0 0 0 0 Morcillo 0 0 0 0 0 0 0 0Zar-Cien 0 0 0 0 0 0 0 0Berr-Tuit 0 0 0 0 0 1 0 0 E Guad 0 0 0 0 0 0 0 018 Ag 0 0 0 0 0 0 0 1 Berr-Salt 0 0 0 0 0 0 0 0 La Erm 0 0 0 0 0 0 0 0 El Ven 1 0 0 1 0 3 0 3 Zar 0 0 0 0 0 2 0 2

52
Continuación Tabla 4.
O.
imbricata Cardo
1 Cardo
2 Cavalillo
nc Cebollino
nc Cempoalillo
nc Chayotillo
nc Chia nc
Chicoria nc
Km 260 0 0 0 0 0 0 0 0 0Km 84 0 0 0 0 0 0 0 0 0Km 104 0 0 0 0 0 0 0 1 0Km 237 0 0 0 0 0 0 0 0 0Km 76 1 0 0 0 0 0 0 0 0Km 61 1 0 0 0 0 0 0 0 0Km 188 1 0 0 0 0 0 0 0 0Km 29 1 0 0 0 0 0 0 0 0Km 51 1 0 0 0 0 0 0 0 1Km 73 0 0 0 0 0 0 0 0 1Km 153 0 0 0 0 0 1 0 0 0Km 130 1 0 0 0 0 0 0 0 0Km 130 0 1 0 1 0 1 0 0 0Km 86 0 1 0 1 0 1 1 0 0 Km 43 1 0 0 0 0 1 0 0 0 Km 23 0 0 0 0 0 1 0 0 0 L Rivera 0 0 0 0 0 1 0 1 0 N. Bravo 0 0 0 0 0 0 0 0 0 C. Najera 0 0 0 0 0 0 0 0 0Rodr 0 0 0 0 0 0 0 0 0 C. Real 0 0 0 0 0 0 0 0 0 Madero 0 0 0 0 0 0 0 0 0Las Huer 0 0 0 0 0 0 0 0 0 Km 36 0 0 0 0 0 0 0 0 0 Zarco 0 0 0 0 0 1 0 0 027 No 0 0 0 0 0 0 0 0 0 Morcillo 0 0 0 0 0 0 0 0 0Zar-Cien 0 0 0 0 0 0 0 0 0Berr-Tuit 0 0 0 0 1 1 0 0 0 E Guad 0 0 0 0 0 0 0 0 018 Ag 0 0 0 0 0 1 0 0 0 Berr-Salt 0 0 0 0 0 0 0 0 0 La Erm 0 0 0 0 0 0 0 0 0 El Ven 2 2 0 2 1 9 1 2 1 Zar 3 1 0 1 1 7 1 1 1

53
Continuación Tabla 4.
Encinilla
nc Escoba china
nc Garambullo
nc M.
biuncifera L.
tridentata H.
palmeris C.
pallida Guaco
nc Km 260 0 0 0 0 0 0 0 0Km 84 0 0 0 0 0 0 0 0Km 104 0 0 1 1 0 1 0 0Km 237 0 0 0 1 1 0 0 1Km 76 0 0 0 1 1 0 0 0Km 61 0 0 1 0 0 0 1 0Km 188 0 0 0 0 1 0 0 0Km 29 0 0 0 0 1 0 0 0Km 51 1 0 0 0 1 0 0 0Km 73 0 0 1 1 0 0 0 0Km 153 0 0 0 0 0 0 0 0Km 130 0 1 0 1 0 1 0 0Km 130 1 0 0 0 0 1 0 0Km 86 0 0 0 0 0 1 0 0 Km 43 0 0 0 0 0 1 0 0 Km 23 0 0 0 0 0 1 0 0 L Rivera 0 0 0 0 0 1 0 0 N. Bravo 0 0 0 0 0 0 0 0 C. Najera 0 0 0 0 0 0 0 0Rodr 0 0 0 0 0 0 0 0 C. Real 0 0 0 0 0 0 0 0 Madero 0 0 0 0 0 1 0 0Las Huer 0 0 0 0 0 0 0 0 Km 36 0 0 0 0 0 0 0 0 Zarco 0 0 0 0 0 0 0 027 No 0 0 0 0 0 0 0 0 Morcillo 0 0 0 0 0 0 0 0Zar-Cien 0 0 0 0 0 0 0 0Berr-Tuit 0 0 0 0 0 0 0 0 E Guad 0 0 0 0 0 0 0 018 Ag 0 0 0 1 0 0 0 0 Berr-Salt 0 0 0 0 0 0 0 0 La Erm 0 0 0 0 0 0 0 0 El Ven 1 1 2 4 0 7 0 0 Zar 1 0 1 1 1 4 1 0

54
Continuación Tabla 4.
V.
enceloides Hepazote de zorrillo
nc Hierba arbustiva
nc Hierba ceniza
nc Hierba de la golondrina
nc Hierbaniz
Km 260 0 0 1 0 0 0Km 84 0 0 0 0 0 0Km 104 0 0 0 0 0 0Km 237 0 0 0 0 0 0Km 76 0 0 0 0 0 0Km 61 0 0 0 1 0 0Km 188 1 0 0 0 0 0Km 29 0 0 0 0 0 0Km 51 0 0 0 0 0 0Km 73 0 0 0 0 0 0Km 153 0 0 0 0 0 0Km 130 1 0 0 0 0 0Km 130 0 0 0 0 0 0Km 86 0 0 0 0 0 0 Km 43 0 0 0 0 1 0 Km 23 0 0 0 0 0 0 L Rivera 0 0 0 0 1 0 N. Bravo 0 0 0 0 0 0 C. Najera 0 0 0 0 0 0Rodr 0 0 0 0 0 0 C. Real 0 0 0 0 0 0 Madero 0 0 0 0 0 1Las Huer 0 0 0 0 0 0 Km 36 0 0 0 0 0 0 Zarco 0 0 0 0 0 027 No 0 0 0 0 0 0 Morcillo 0 0 0 0 0 0Zar-Cien 0 0 0 0 0 0Berr-Tuit 0 0 0 0 0 0 E Guad 0 0 0 0 0 018 Ag 0 1 0 0 0 0 Berr-Salt 0 0 0 0 0 0 La Erm 0 0 0 0 0 0 El Ven 1 1 0 0 2 0 Zar 0 1 0 1 2 0

55
Continuación Tabla 4.
A.
tortuosa Huizachillo
nc S.salisifolia Lampotillo nc
C. nardus
S. rostratum
I. purpurea
P. incanum
P. juliflora
Km 260 0 0 0 0 0 1 0 0 0Km 84 0 0 0 0 0 0 0 0 0Km 104 1 0 0 0 0 1 1 0 1Km 237 0 0 0 0 0 0 0 0 0Km 76 1 0 0 0 1 1 0 0 1Km 61 0 0 0 0 0 0 1 1 1Km 188 0 0 0 0 0 0 1 0 0Km 29 0 0 0 0 0 0 0 1 0Km 51 1 1 0 0 0 0 0 0 1Km 73 0 0 0 0 1 0 1 0 1Km 153 0 0 0 1 0 0 0 0 0Km 130 1 0 0 1 0 0 0 0 1Km 130 1 0 0 0 0 0 0 0 1Km 86 1 0 0 1 0 0 0 0 0 Km 43 0 0 0 0 1 0 0 0 0 Km 23 1 0 1 0 1 0 1 0 0 L Rivera 1 0 0 0 1 1 1 0 1 N. Bravo 0 0 0 0 0 0 0 0 0 C. Najera 0 0 0 0 0 0 0 0 0Rodr 0 0 0 0 0 0 0 0 0 C. Real 0 0 0 0 0 0 0 0 0 Madero 1 0 1 0 0 0 0 0 0Las Huer 0 0 0 0 0 0 0 0 0 Km 36 0 0 0 0 0 0 0 0 0 Zarco 1 0 0 0 0 0 0 0 127 No 0 0 0 0 0 0 0 0 0 Morcillo 0 0 0 0 0 0 0 0 0Zar-Cien 0 0 0 0 0 0 0 0 0Berr-Tuit 1 0 0 0 0 1 1 0 0 E Guad 0 0 0 0 0 0 0 0 018 Ag 1 0 0 0 1 1 0 0 1 Berr-Salt 0 0 0 0 0 0 0 0 0 La Erm 0 0 0 0 0 0 0 0 0 El Ven 9 0 1 3 5 4 5 0 7 Zar 7 1 1 1 4 3 4 1 5

56
Continuación Tabla 4.
A.
constricta G.
mendocina O.migrodasys O. leucotricha
O. rastrera
F. splendes
Ojo de chanate nc
L. graveolens
Km 260 1 0 0 0 0 0 0 0Km 84 0 0 0 0 0 0 0 0Km 104 1 0 0 0 0 0 0 0Km 237 1 0 0 0 0 1 0 0Km 76 1 1 0 0 0 0 0 0Km 61 0 0 0 1 0 1 0 1Km 188 0 0 0 0 1 0 0 0Km 29 1 0 0 0 0 0 0 0Km 51 1 0 1 0 0 1 0 0Km 73 0 0 0 0 0 1 0 0Km 153 1 0 0 0 0 0 0 0Km 130 1 1 0 1 0 0 1 0Km 130 0 0 0 0 0 0 0 0Km 86 0 0 0 0 0 0 0 0 Km 43 0 0 0 1 0 0 0 0 Km 23 1 0 0 1 0 0 0 0 L Rivera 0 0 0 0 0 0 0 0 N. Bravo 0 0 0 0 0 0 0 0 C. Najera 0 0 0 0 0 0 0 0Rodr 0 0 0 0 0 0 0 0 C. Real 0 0 0 0 0 0 0 0 Madero 0 0 0 0 0 0 0 0Las Huer 0 0 0 0 0 0 0 0 Km 36 0 0 0 0 0 0 0 0 Zarco 0 0 0 1 0 0 0 027 No 0 0 0 0 0 0 0 0 Morcillo 0 0 0 0 0 0 0 0Zar-Cien 0 0 0 0 0 0 0 0Berr-Tuit 0 0 0 0 0 0 0 0 E Guad 0 0 0 1 0 0 0 018 Ag 0 0 0 0 0 0 0 0 Berr-Salt 0 0 0 0 0 0 0 0 La Erm 0 0 0 0 0 0 0 0 El Ven 4 1 0 5 0 1 1 0 Zar 2 0 1 4 0 2 0 1

57
Continuación Tabla 4.
C. glomeratum Pollo nc A. palmeris S. kali B. leucantha Salvilla nc J. dioica Siempre viva nc Tabacon ncKm 260 0 0 0 0 0 0 0 0 0Km 84 0 0 0 0 0 0 0 0 0Km 104 0 0 0 0 0 0 0 0 0Km 237 0 0 0 0 0 0 0 0 0Km 76 0 0 0 0 0 1 0 0 0Km 61 0 0 0 0 0 0 1 0 0Km 188 0 0 1 0 0 0 1 1 0Km 29 0 0 0 0 0 0 1 0 0Km 51 0 0 0 0 0 0 1 0 0Km 73 0 0 0 0 0 0 1 0 0Km 153 0 0 1 1 1 0 0 0 0Km 130 1 0 0 0 1 1 0 0 0Km 130 1 0 0 0 1 0 0 0 1Km 86 0 0 1 0 0 0 0 0 0 Km 43 0 0 1 0 1 1 0 0 0 Km 23 0 1 0 0 0 0 0 0 0 L Rivera 0 0 1 0 1 1 0 0 0 N. Bravo 0 0 0 0 0 0 0 0 0 C. Najera 0 0 0 0 0 0 0 0 0Rodr 0 0 0 0 0 0 0 0 0 C. Real 0 0 0 0 0 0 0 0 0 Madero 0 0 0 0 1 0 0 0 0Las Huer 0 0 0 0 0 0 0 0 0 Km 36 0 0 0 0 0 0 0 0 0 Zarco 0 0 0 0 1 0 0 0 027 No 0 0 0 0 0 0 0 0 0 Morcillo 0 0 0 0 0 0 0 0 0Zar-Cien 0 0 0 0 0 0 0 0 0Berr-Tuit 0 0 1 0 1 0 0 0 0 E Guad 0 0 0 0 0 0 0 0 018 Ag 0 0 0 0 1 0 0 0 0 Berr-Salt 0 0 0 0 0 0 0 0 0 La Erm 0 0 0 0 0 0 0 0 0 El Ven 2 1 5 1 8 3 1 0 1 Zar 0 1 4 0 5 2 2 0 0

58
Continuación Tabla 4. O. leptocaulis Tienguis nc Tomate de perro nc Torito nc S. eleagnifolum Tronadora nc Vara dulce nc
Km 260 0 0 0 0 0 0 0Km 84 0 0 0 0 0 0 0Km 104 0 1 0 0 0 0 0Km 237 0 0 0 0 0 0 1Km 76 1 0 0 0 1 0 0Km 61 1 0 0 0 0 0 0Km 188 1 0 0 0 0 0 0Km 29 1 0 0 0 1 0 0Km 51 1 0 0 0 0 0 0Km 73 1 0 0 0 0 0 0Km 153 0 0 0 0 1 0 0Km 130 0 0 1 0 0 0 0Km 130 0 0 0 0 0 0 0Km 86 0 0 0 0 0 0 0 Km 43 0 0 0 0 1 0 0 Km 23 0 0 0 0 0 1 0 L Rivera 0 0 1 1 1 0 0 N. Bravo 0 0 0 0 0 0 0 C. Najera 0 0 0 0 0 0 0Rodr 0 0 0 0 0 0 0 C. Real 0 0 0 0 0 0 0 Madero 0 0 0 0 0 0 0Las Huer 0 0 0 0 0 0 0 Km 36 0 0 0 0 0 0 0 Zarco 0 0 0 0 0 0 027 No 0 0 0 0 0 0 0 Morcillo 0 0 0 0 0 0 0Zar-Cien 0 0 0 0 0 0 0Berr-Tuit 0 0 1 0 1 0 0 E Guad 0 0 0 0 0 0 018 Ag 0 0 0 0 0 0 0 Berr-Salt 0 0 0 0 0 0 0 La Erm 0 0 0 0 0 0 0 El Ven 1 1 3 1 4 1 0 Zar 2 0 2 1 3 1 0

59
Continuación Tabla 4. P. oleracea Yucca spp. C. dactylon P. notatum B. curtipendula E. pulchellum E. desavauxi P. lividum
Km 260 0 0 0 0 0 0 0 0Km 84 0 0 0 0 0 0 0 0Km 104 0 0 1 0 1 0 0 0Km 237 0 0 0 0 0 0 0 0Km 76 0 0 0 0 0 0 1 0Km 61 0 0 0 0 1 0 1 0Km 188 0 0 0 0 0 0 0 0Km 29 0 0 0 0 0 0 0 0Km 51 0 1 0 0 1 1 0 0Km 73 0 0 0 0 0 0 1 0Km 153 0 0 0 0 1 0 0 0Km 130 0 0 0 0 0 0 0 1Km 130 0 0 0 0 1 0 1 0Km 86 0 0 0 0 0 0 1 0 Km 43 0 0 0 0 0 0 0 0 Km 23 0 0 0 1 0 0 0 0 L Rivera 0 0 0 0 0 0 0 0 N. Bravo 0 0 0 0 0 0 0 0 C. Najera 0 0 0 0 0 0 0 0Rodr 0 0 0 0 0 0 0 0 C. Real 0 0 0 0 0 0 0 0 Madero 0 0 0 0 0 0 0 0Las Huer 0 0 0 0 0 0 0 0 Km 36 0 0 0 0 0 0 0 0 Zarco 0 0 0 0 1 0 0 027 No 0 0 0 0 0 0 0 0 Morcillo 0 0 0 0 0 0 0 0Zar-Cien 0 0 0 0 0 0 0 0Berr-Tuit 1 0 0 0 0 0 0 0 E Guad 0 0 0 0 0 0 0 018 Ag 0 0 1 0 0 0 0 0 Berr-Salt 0 0 0 0 0 0 0 0 La Erm 0 0 0 0 0 0 0 0 El Ven 1 0 2 1 4 0 3 1 Zar 1 1 1 1 3 1 2 0

60
Continuación Tabla 4. S. neesiana A. tenuis L. phleoides Ch. virgata B. gracilis S. airoides S. leucopila A. barbinodis R roseum
Km 260 0 0 0 1 1 1 0 0 0Km 84 0 0 0 0 0 0 0 0 0Km 104 0 0 0 1 1 1 0 1 0Km 237 0 0 0 0 1 0 0 0 0Km 76 0 0 0 0 0 0 0 0 0Km 61 0 0 0 0 1 0 0 0 0Km 188 0 1 0 1 1 0 1 0 0Km 29 0 0 0 1 1 0 0 0 0Km 51 0 0 0 0 0 0 0 0 0Km 73 0 0 1 0 1 0 0 1 0Km 153 1 0 0 0 0 0 1 1 1Km 130 0 0 0 1 1 0 0 1 1Km 130 1 0 1 1 1 0 0 1 0Km 86 0 0 0 1 1 0 0 0 1 Km 43 0 0 0 1 1 1 0 1 1 Km 23 0 0 0 1 1 0 0 0 0 L Rivera 0 0 0 1 1 0 0 0 0 N. Bravo 0 0 0 0 0 0 0 0 0 C. Najera 0 0 0 0 0 0 0 0 0Rodr 0 0 0 0 0 0 0 0 0 C. Real 0 0 0 0 0 0 0 0 0 Madero 0 1 1 0 1 0 0 1 1Las Huer 0 0 0 0 0 0 0 0 0 Km 36 0 0 0 0 0 0 0 0 0 Zarco 0 0 1 0 1 0 0 1 127 No 0 0 0 0 0 0 0 0 0 Morcillo 0 0 0 0 0 0 0 0 0Zar-Cien 0 0 0 0 0 0 0 0 0Berr-Tuit 0 1 0 1 1 1 0 0 0 E Guad 0 0 0 0 0 0 0 0 018 Ag 0 1 0 1 1 0 0 1 1 Berr-Salt 0 0 0 0 0 0 0 0 0 La Erm 0 0 0 0 0 0 0 0 0 El Ven 2 2 3 9 11 3 1 8 6 Zar 0 2 1 6 8 2 0 3 4

61
Continuación Tabla 4. D. spicata B. laciniatus C. pauciflorus A. adscensionis
Km 260 0 0 0 0 Km 84 0 0 0 0 Km 104 0 0 0 1 Km 237 0 0 0 0 Km 76 0 0 0 0 Km 61 0 0 0 0 Km 188 0 0 0 0 Km 29 1 0 0 1 Km 51 1 0 0 1 Km 73 0 0 0 1 Km 153 0 1 1 1 Km 130 0 0 0 0 Km 130 0 0 0 1 Km 86 0 0 0 1 Km 43 0 0 0 1 Km 23 0 0 0 1 L Rivera 0 0 0 0 N. Bravo 0 0 0 0 C. Najera 0 0 0 0 Rodr 0 0 0 0 C. Real 0 0 0 0 Madero 0 1 0 0 Las Huer 0 0 0 0 Km 36 0 0 0 0 Zarco 0 0 0 1 27 No 0 0 0 0 Morcillo 0 0 0 0 Zar-Cien 0 0 0 0 Berr-Tuit 0 0 0 0 E Guad 0 0 0 0 18 Ag 1 0 1 1 Berr-Salt 0 0 0 0 La Erm 0 0 0 0 El Ven 1 1 2 9 Zar 2 0 1 6

62
TABLA DE CONTENIDO
INTRODUCCIÓN.............................................................................................................. 2 ANTECEDENTES ............................................................................................................. 4
Ciclo biológico del chapulín ......................................................................................... 4 Hábitos alimenticios ..................................................................................................... 4 Distribución................................................................................................................... 5 Daños a cultivos y pastizales por chapulín ................................................................. 6 Vegetación ..................................................................................................................... 7 Métodos espaciales y predictivos de plaga de chapulín ............................................ 7 Análisis de similitud ..................................................................................................... 9 Componentes principales ........................................................................................... 10
DEFINICION DEL PROBLEMA.................................................................................... 12 JUSTIFICACION............................................................................................................. 14 HIPOTESIS ...................................................................................................................... 15 OBJETIVO ....................................................................................................................... 16 OBJETIVOS PARTICULARES ...................................................................................... 16 DELIMITACION TIEMPO Y ESPACIO........................................................................ 17 METODOLOGÍA............................................................................................................. 18
Chapulín ...................................................................................................................... 20 Topografía ................................................................................................................... 20 Vegetación ................................................................................................................... 22 Análisis de similitud ................................................................................................... 23
RESULTADOS ................................................................................................................ 24 Datos de campo ........................................................................................................... 24 Análisis de similitud y componente principales ....................................................... 25
CONCLUSIONES............................................................................................................ 38 BIBLIOGRAFÍA.............................................................................................................. 40