Embed Size (px)
Citation preview

熊本大学学術リポジトリ
Kumamoto University Repository System
Title 肝内胆管癌と肝星細胞の相互作用に関する研究
Author(s) 岡部, 弘尚
Citation
Issue date 2010-03-25
Type Thesis or Dissertation
URL http://hdl.handle.net/2298/21744
Right

1
学位論文
Doctoral Thesis
肝内胆管癌と肝星細胞の相互作用に関する研究
(Role of hepatic stellate cells in intrahepatic cholangiocarcinoma)
岡部 弘尚
Hirohisa Okabe
熊本大学大学院医学教育部博士課程臨床医科学専攻消化器外科学
指導教員
馬場 秀夫 教授
熊本大学大学院医学教育部博士課程医学専攻消化器外科学
安東 由喜雄 教授
熊本大学大学院医学教育部博士課程医学専攻病態情報解析学
2010 年 3 月

2
目次
要旨 ・・・・・・・・・・・・・・・ 6
発表論文リスト ・・・・・・・・ 9
謝辞 ・・・・・・・・・・・・・・・・ 11
略語一覧 ・・・・・・・・・・・・ 12
第一章 研究の背景と目的 ・・・・・・・・・・・・ 13
1- 1. 肝内胆管癌の疫学と予後
1-1-1. 疫学とハイリスク因子
1-1-2. 治療成績
1- 2. 肝内胆管癌の腫瘍間質における特性
1- 3. 肝星細胞とは
1-3-1. 解剖・生理学的特徴
1-3-2. 肝の線維化における役割
1-3-3. 肝の線維化における myofibroblast の由来
1- 4. 癌関連線維芽細胞(Cancer-associated fibroblasts; CAFs)
1-4-1. CAF とは
1-4-2. 固形癌における CAF の臨床病理学的意義
1-4-3. 癌細胞と肝星細胞の相互作用に関する報告
1- 5. 今研究の目的

3
第二章 患者対象、実験材料と方法 ・・・・・・・・・・・・ 21
2- 1. 患者対象
2- 2. 免疫組織学的解析
2-2-1. 免疫染色手法 (パラフィン包埋組織)
2-2-2. 癌間質、慢性肝炎における myofibroblast の定量
2-2-3. 二重免疫染色 (凍結保存組織)
2- 3. 細胞培養
2- 4. 共培養モデル
2-4-1. 細胞浸潤アッセイ
2-4-2. 細胞増殖アッセイ
2-4-3. 細胞遊走アッセイ
2-4-3. サイトカインアレイ
2- 5. 培養上清を用いた実験
2-5-1. 培養上清の準備
2-5-2. 培養細胞の免疫染色
2-5-3. ウェスタン・ブロット法
2-5-4. ヒト臍帯静脈血管内皮細胞(HUVEC)チューブ形成アッセイ
2-5-5. アポトーシスアッセイ
2- 6. In vivo における腫瘍形成能の比較と組織学的評価
2- 7. 質量分析を用いた蛋白同定
2-7-1. SELDI-TOF MS 解析

4
2-7-2. Q-TOF/MS/MS 解析
2-7-3. 蛋白ビーズを用いた免疫沈降
2- 7. 統計学的解析方法
第三章 実験結果・・・・・・・・・・・・ 29
3- 1. 肝内胆管癌の免疫組織学的検討
3-1-1. 肝内胆管癌におけるαSMA、CD34 の発現
3-1-2. αSMA を用いた myofibroblast の定量化は有効である
3-1-3. 腫瘍間質の Myofibroblast とコラーゲン量は相関しない
3-1-4. 肝内胆管癌において、myofibroblast は生存予後に関与する
3-1-5. 肝内胆管癌の腫瘍間質に肝星細胞が存在する
3- 2. 培養上清を用いた胆管癌細胞と肝星細胞の相互作用の確認
3-2-1. 胆管癌細胞は肝星細胞を活性化する
3-2-2. 肝星細胞は胆管癌細胞の細胞増殖・浸潤能を促進する
3-2-3. 肝星細胞は胆管癌細胞の MAPK, Akt 経路を活性化する
3-2-4. 胆管癌と肝星細胞の相互作用によりチューブ形成は促進する
3-2-5. 肝星細胞は胆管癌細胞の抗癌剤感受性を抑制する
3-3. In vivo における肝星細胞と胆管癌細胞の相互作用
3-3-1. 肝星細胞は胆管癌細胞の腫瘍形成を促進する
3-3-2. 肝星細胞は胆管癌細胞の腫瘍形成において、腫瘍間質形成や血管新生
に関与しない

5
3- 4. 共培養モデルを用いた胆管癌と肝星細胞の相互作用
3-4-1. 共培養により胆管癌細胞の浸潤能は促進する
3-4-2. 共培養により胆管癌細胞の増殖能は促進する
3-4-3. 肝星細胞と胆管癌の相互作用に炎症性サイトカイン IL-1α ,IL-1β,C5a,G-CSF が関与する
3-5. 質量分析を用いた、肝星細胞と胆管癌の相互作用に関与する因子 CXCL5 の
同定
3-5-1. SELDI-TOF MS解析で得られた、胆管癌細胞と肝星細胞の相互作用に関
わる蛋白の質量は 8360Da である
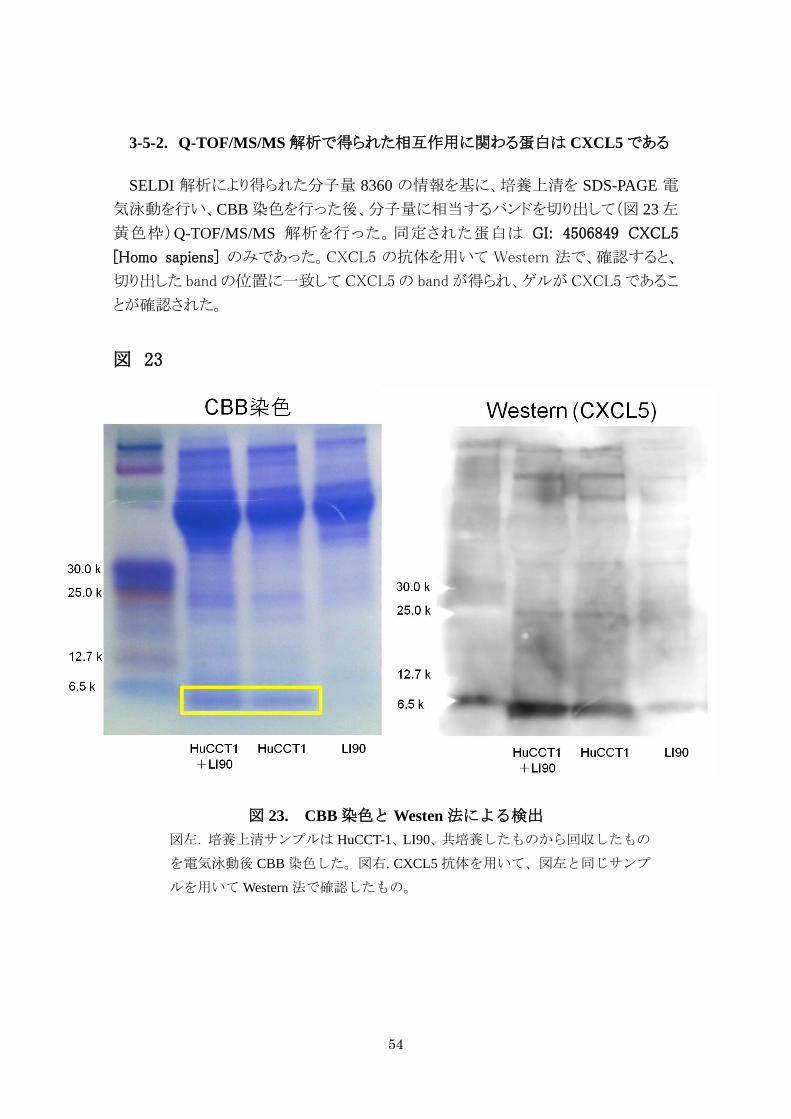
3-5-2. Q-TOF/MS/MS 解析で得られた相互作用に関わる蛋白は CXCL5 である
3-5-3. SELDI-TOF MS で得られた 8360 Da のピークは CXCL5 である
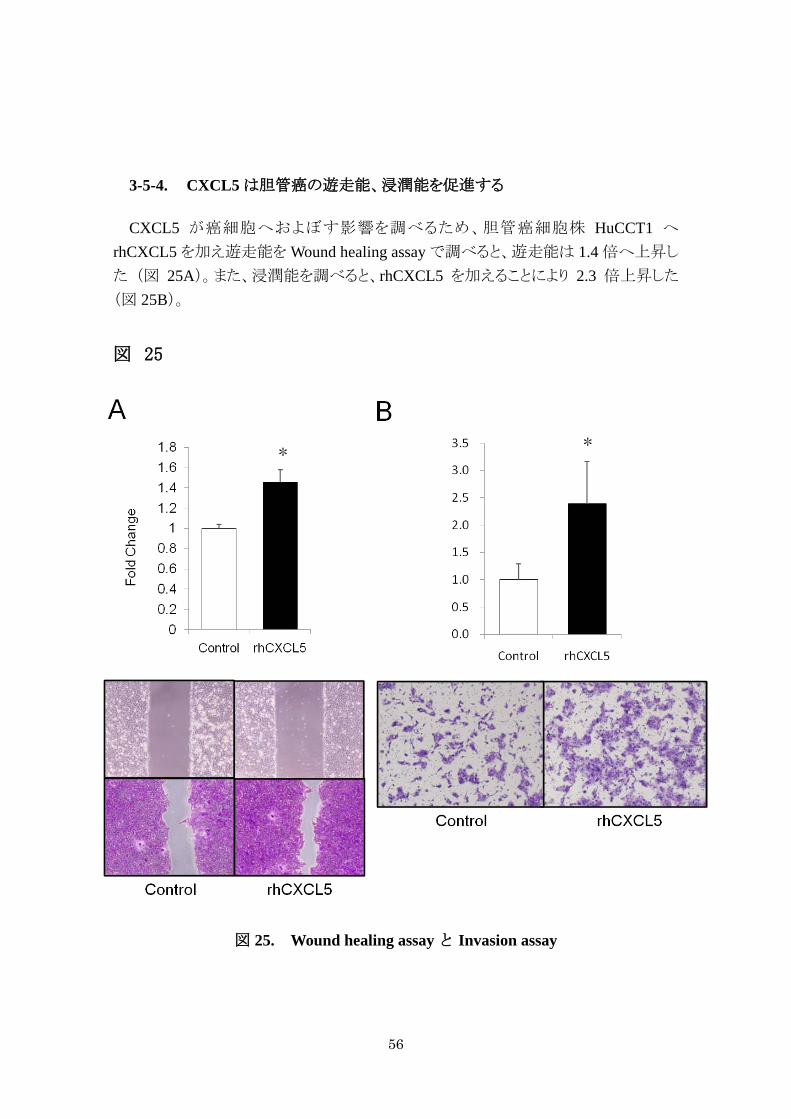
3-5-4. CXCL5 は胆管癌の遊走能、浸潤能を促進する
3-5-5. CXCL5 の発現は myofibroblast の存在と相関し、CXCL5 髙発現群は生
存予後不良である
第四章 考察 ・・・・・・・・・・・・ 54
第五章 結語 ・・・・・・・・・・・・ 57
第六章 参考文献 ・・・・・・・・・・・・ 58

6
要旨
【目的】 腫瘍間質の myofibroblast または Cancer associated fibroblasts (CAF) は、
腫瘍の増殖・浸潤・転移・血管新生を促進させ、癌に治療標的としての研究が
進められている。膵癌では、膵星細胞は癌間質において活性化し、myofibroblastへと分化し、癌の進行に関与することが知られているが、肝星細胞の肝癌、特
に肝内胆管癌(ICC)における存在やその意義は未だ明らかでない。ICC におけ
る myofibroblast の関連性、肝星細胞の存在を調べ、in vitro, in vivo における胆管
癌と肝星細胞の関連性とその相互作用に関わる因子を検索した。
【方法】1.肝切除を施行した ICC 46 例のサンプルを用いた。Myofibroblast の評
価として α-smooth muscle actin(α-SMA)による免疫染色を行い、臨床病理学的
因子や予後との関連を調べた。2.星細胞のマーカーとして知られる α-SMA、
grial fibrillary acidic protein (GFAP)、desmin の発現を、ICC の凍結組織で確認した。
3.胆管癌細胞株(HuCCT-1,RBE)と肝星細胞株(LI90)を用い、それぞれの
培養上清 (Conditioning medium: CM) を準備し、in vitro, in vivo での両者の癌の
進展における相互作用を調べた。4.相互作用に関与する因子について共培養
モデルと質量分析(DELDI-TOF MS, Q-TOF/MS/MS)を用いた手法で同定作業を
行った。5.同定された因子 CXCL5 について、細胞株における機能解析と肝内
胆管癌症例における発現意義の検証を行った。
【結果】 1. ICC における α-SMA の発現率は 0.69-10.6%(中央値 5.19%)であ
った。Α-SMA 高発現群は低発現群に比べて、女性・組織学的脈管浸潤を有意に
多く認め、α-SMA 高発現群は累積生存率、累積無再発生存率ともに低発現群よ
り有意に不良であった。多変量解析において、α-SMA 高発現(p=0.0045)とリ
ンパ節転移陽性(p=0.0127)は独立予後した不良因子であった。2.ICC 間質に
αSMA と GFAP または Desmin の共発現細胞を認め、これらは肝星細胞由来の
myofibroblast と考えられた。3.胆管癌細胞株の CM によって LI90 は活性化さ
れ、その増殖能は CM 濃度依存性に増加した。3. 逆に星細胞の CM によって、
胆管癌細胞株 2 種で、細胞増殖・浸潤能は CM 濃度依存性に増加し、細胞内シ
グナルの MAPK、Akt 経路の活性化がみられた。4.LI90 の CM によって、抗
癌剤に対するアポトーシスの抑制効果が得られた。5.癌細胞と星細胞の相互
作用により、HUVEC(ヒト臍帯静脈内皮細胞)のチューブ形成が促進された。
6.In vivo において、星細胞は胆管癌細胞の腫瘍形成を促進させた。7.
SELDI-TOF MS、Q-TOF/MS/MS を用いて、共培養モデルから得た培養上清の解
析により、胆管癌細胞と肝星細胞の相互作用に関わる因子として CXCL5 が同定
された。8.CXCL5 は胆管癌細胞の遊走能、浸潤能を促進した。ICC において、

7
癌部 CXCL5 の発現と myofibroblast の存在は相関し、CXCL5 髙発現群は予後不
良であった。
【考察】 ICC において myofibroblat はその予後に関与し、由来細胞として肝星細
胞が考えられた。しかし、ヒトにおいて肝星細胞を特定する優れたマーカーは
乏しく、その検出には今後のさらなる研究が必要である。共培養モデルと質量
分析を用いて同定された胆管癌細胞と肝星細胞の相互作用に関わる因子 CXCL5は肝内胆管癌症例において癌細胞が分泌し、CAF の存在と胆管癌の生存不良に
関与し、腫瘍の進展における癌間質相互作用に関わる重要な因子であると考え
られた。
【結論】 ICCの進展において、肝星細胞との相互作用は重要であり、相互作用に関わ
る因子として CXCL5 が同定された。

8
発表論文リスト
関連論文
1. Okabe H, Beppu T, Hayashi H, Ishiko T, Masuda T, Otao R, Horlad H, Shinriki S, Ueda M, Jono H, Ando Y, and Baba H. Hepatic stellate cells accerelate the malignant behavior of cholangiocarcinoma cells. Ann Surg Oncol 2010 (in press)
2. Okabe H, Beppu T, Hayashi H, Horino K, Masuda T, Komori H, Ishikawa S, Watanabe M, Takamori H, Iyama K, and Baba H. Hepatic stellate cell may promote progression of intrahepatic cholangiocarcinoma. Ann Surg Oncol 2009;16:2555-64
その他の論文 1. Hasita H, Komohara Y, Okabe H, Masuda T, Ohnishi K, Lei XF, Beppu T,
Baba H, Takeya M. The significance of alternatively activated macrophages in patients with intrahepatic cholangiocarcinoma. Cancer Sci 2010 (in press)
2. Beppu T, Hayashi N, Masuda T, Komori H, Horino K, Hayashi H, Okabe H, Baba Y, Kinoshita K, Akira C, Watanabe M, Takamori H, Baba H. FOLFOX enables high respectability and excellent prognosis for initially unresectable colorectal liver metastasis. Anticancer Res 2010;30:1015-20.
3. Komori H, Beppu T, Baba Y, Horino K, Imsung C, Masuda T, Hayashi H, Okabe H, Otao R, Watanabe M, Takamori H, Iyama K, Baba H. Histological liver injury and surgical outcome after FOLFOX followed by a hepatectomy for colorectal liver metastases in Japanese patients. Int J Clin Oncol 2010;15:263-70.
4. Hayashi H, Beppu T, Sugita H, Masuda T, Okabe H, Takamori H, Baba H. Serum HGF and TGF-beta1 levels after right portal vein embolization. Hepatol Res 2010;45:554-9.
5. Hayashi H, Beppu T, Okabe K, Masuda T, Okabe H, Ishiko T, Baba H. Therapeutic factors considered according to the preoperative splenic volume for a prolonged increase in platelet count after partial splenic embolization for liver cirrhosis. J Gastroenterol. 2010;45:554-9.
6. Masuda T, Beppu T, Horino K, Komori H, Hayashi H, Okabe H, Otao R, Horlad H, Ishiko T, Takamori H, Kikuchi K, Baba H. Preoperative tumor

9
marker doubling time is a useful predictor of recurrence and prognosis after hepatic resection of hepatocellular carcinoma. J Surg Oncol 2009 (in press)
7. Beppu T, Iwatsuki M, Okabe H, Okabe K, Masuda T, Hayashi H, Sugiyama S, Horino K, Komori H, Hayashi N, Takamori H, Ishiko T, Baba H. A new approach to percutaneous transhepatic portal embolization using ethanolamine oleate iopamidol. J Gastroenterol. 2010;45:211-7.
8. Hayashi H, Beppu T, Sugita H, Horino K, Komori H, Masuda T, Okabe H, Takamori H, Baba H. Increase in the serum bile acid level predicts the effective hypertrophy of the nonembolized hepatic lobe after right portal vein embolization. World J Surg. 33: 1933-40, 2009.
9. Imai K, Beppu T, Nakayama Y, Ishiko T, Horino K, Komori H, Masuda T, Hayashi H, Okabe H
Preoperative prediction of poorly differentiated components in small-sized hepatocellular carcinoma for safe local ablation therapy.
, Baba Y, Watanabe M, Takamori H, Awai K, Yamashita Y, Baba H.
J Surg Oncol. 100:121-6, 2009.
10. Masuda T, Beppu T, Horino K, Komori H, Hayashi H, Okabe HOccurrence of
hepatocellular carcinoma after hepatoblastoma resection in an adult with hepatitis C virus.
, Ootao R, Horlad H, Baba Y, Miyase S, Takamori H, Baba H.
Hepatol Res. 39:525-30, 2009.
11. Masuda T, Beppu T, Ishiko T, Horino K, Baba Y, Mizumoto T, Hayashi H, Okabe H
Intrahepatic dissemination of hepatocellular carcinoma after local ablation therapy.
, Horlad H, Doi K, Okabe K, Takamori H, Hirota M, Iyama K, Baba H.
J Hepatobiliary Pancreat Surg. 15:589-95, 2008.

10
謝辞
熊本大学大学院生命科学研究部消化器外科学 馬場秀夫 教授、熊本大学大学
院生命科学研究部病態情報解析学 安東由喜雄 教授のご指導の下、本研究を行い
ました。多くのご指導を頂き、深く感謝いたします。
熊本地域医療センター外科 石川晋之先生には研究全般における技術的なご指導
を頂きました。
熊本大学大学大学院生命科学研究部分子病理学 久保多津子実験補助員、消化
器外科学 谷口裕子実験補助員には病理組織学的検査の技術的なご指導を頂きま
した。
熊本大学大学医学部附属病院病理部 猪山賢一 先生には病理組織化学におけ
る多くのご指導を頂きました。
小沢医院 辻龍也 先生には臨床病理学的な詳細な情報収集のご協力を頂きまし
た。
また、病態情報解析学教室、消化器外科学教室の皆様には有形無形の多くのご協
力とご指導を頂きました。
最後に、熊本大学大学院生命科学研究部消化器外科学 別府透准教授には論文
の読み方、研究に対する姿勢など、直接的なご指導・ご鞭撻を頂きました。
心から感謝いたします。

11
略語一覧 ICC, intrahepatic cholangiocarcinoma HS cell, hepatic stellate cell α-SMA, α-smooth muscle actin GFAP, glial fibrillary acidic protein DAPI, 4′,6-diamidino-2-phenylindole OD, optical density CC, cholangiocarcinoma CM, conditioned media BMDCs, bone marrow derived-cells CAFs, cancer-associated fibroblasts TGF-β, transforming growth factor-β EGF, epidermal growth factor PDGF, platelet-derived growth factor bFGF, basic fibroblast growth factor VEGF, vascular endothelial growth factor HUVECs, human umbilical vein endothelial cells MAPK, mitogen-activated protein kinase HRP, horse radish peroxidase PFA, parafolm aldehyde CDDP, cisdiamine dichloroplatinum 5-FU, 5-Fluorouracil TUNEL, terminal deoxyribonucleotidyl transferase-mediated dUTP nick end labeling; MDV, mean densitometric value PVDF, polyvinylidene fluoride CXCL5, C-X-C ligand 5 CXCR2, C-X-C receptor 2 SELDI-TOF MS, Surface - Enhanced Laser Desorption and Ionization Time of Flight - Mass Spectrometry

12
第一章 研究の背景と目的
1- 1. 肝内胆管癌の疫学と予後
1-1-1. 疫学とハイリスク因子
肝内胆管癌 (ICC) の発生頻度は肝原発悪性腫瘍の中で肝細胞癌 (HCC) に次
いで 2 番目に高く、約 3~5 %と報告されている(2005 年 原発性肝癌調査報告)。ICCのハイリスク因子とされる疾病として、肝内結石症、先天性胆道拡張症、原発性硬化
性胆管炎、ウイルス性肝炎、肝吸虫がある 1-3。
1-1-2. 治療成績
ICC は HCC と異なり高率な肝転移や他臓器転移再発ため、治療成績は不良であり、
近年の根治的切除成績を見ても、5 年生存率は 21-48%と報告されている (Table 1)4-11。さらに、現在切除不能な ICC に対する標準的な化学療法は確立されておらず、
肝外胆管癌や胆嚢癌などの胆道癌に準じて化学療法が行われているのが現状である。
これまで ICC の多数例による prospective な検討は存在せず、本邦における後ろ向き
研究では、2000 年 4 月から 2003 年 3 月までの 3 年間に非切除治療を実施された
ICC54 例の 1 年生存率は 34.3%であった 12。
表 1. 肝内胆管癌の切除成績 著者 論文 年 症例数 5 年生存率
Weimann et al. Br J Surg 2000 95 21
Inoue et al. Surgery 2000 52 36
Nakagawa et al. W J Surg 2005 53 26.3
Tan et al. Ann Surg Oncol 2008 91 41
Konstadoulakis et al. Surgery 2008 54 25
Jonas et al. Ann Surg 2009 195 30.4
Choi et al. Ann Surg Oncol 2009 64 39.5
Cho et al. Ann Surg Oncol 2010 63 31.8
Ercolani et al. Ann Surg 2010 72 48

13
1- 2. 肝内胆管癌の腫瘍間質における特性
ICC は cholangiocyte を、HCC は hepatocyte をその起源とし、ICC は他の上皮癌同
様豊富な腫瘍間質を有し、HCC が被膜を有し、膨張性発育をその特徴とするのに対
して、浸潤性に発育し、被膜を有さない。 ICC、大腸癌肝転移、HCC 組織を
Malory-Azan 染色を用いて fibrotic stroma として腫瘍間質を比較すると、ICC と
大腸癌肝転移はほぼ等しく、HCC より著明に多いことが報告されている 13。
図 1
図1.肝内胆管癌と肝細胞癌の比較
上部写真は肝内胆管癌と肝細胞癌の肉眼
写真と病理組織写真。右図は肝内胆管癌
(CCC)と肝細胞癌(HCC)と大腸癌肝転移
(MLC)の間質量を比較したもの。
14
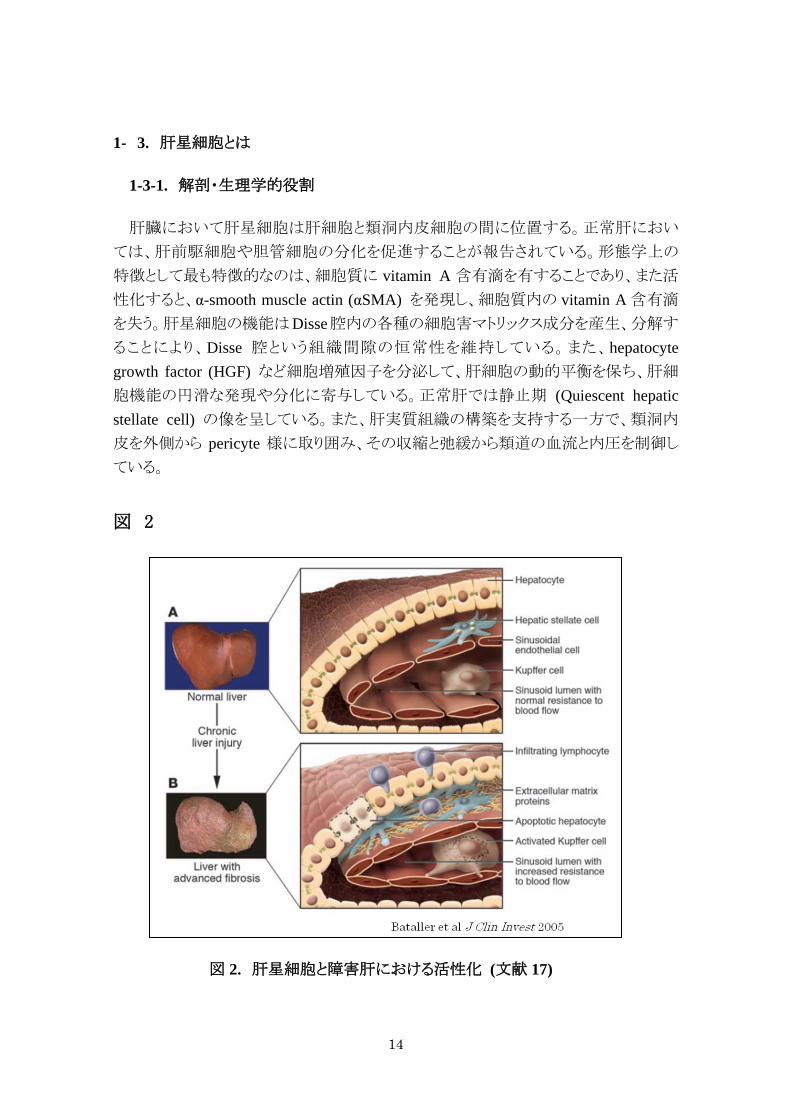
1- 3. 肝星細胞とは
1-3-1. 解剖・生理学的役割
肝臓において肝星細胞は肝細胞と類洞内皮細胞の間に位置する。正常肝におい
ては、肝前駆細胞や胆管細胞の分化を促進することが報告されている。形態学上の
特徴として最も特徴的なのは、細胞質に vitamin A 含有滴を有することであり、また活
性化すると、α-smooth muscle actin (αSMA) を発現し、細胞質内の vitamin A 含有滴
を失う。肝星細胞の機能はDisse腔内の各種の細胞害マトリックス成分を産生、分解す
ることにより、Disse 腔という組織間隙の恒常性を維持している。また、hepatocyte growth factor (HGF) など細胞増殖因子を分泌して、肝細胞の動的平衡を保ち、肝細
胞機能の円滑な発現や分化に寄与している。正常肝では静止期 (Quiescent hepatic stellate cell) の像を呈している。また、肝実質組織の構築を支持する一方で、類洞内
皮を外側から pericyte 様に取り囲み、その収縮と弛緩から類道の血流と内圧を制御し
ている。
図 2
図 2. 肝星細胞と障害肝における活性化 (文献 17)

15
1-3-2. 肝の線維化における役割
各種肝障害に伴う肝星細胞周囲の細胞外マトリックスの性状や立体構造の変化、障
害局所に産生された増殖因子、サイトカイン、ケモカインが肝星細胞のレセプターと結
合すると、肝星細胞は活性化あるいは分化して、形質が転換し、筋線維芽細胞化する。
筋線維芽細胞化した肝星細胞 (activated hepatic stellate cell)は特徴的な脂肪滴を失
い、α平滑筋アクチン (αSMA) の発現増強が生じる。また、以下の多彩な機能が亢進
する。①細胞外マトリックス成分の分解能の亢進、②細胞分裂・増殖能の亢進、③障
害局所への遊走能の亢進、④コラーゲン線維を主とした細胞外マトリックス成分の産
生能の増強、⑤白血球を障害局所に集めるケモカインの分泌、⑥収縮能の亢進、⑦ ①~⑥の作用を惹起するサイトカインやそれぞれのレセプターおよび ECM 成分、
ECM 成分の分解酵素やその阻害因子、線溶系酵素などの遺伝子の発現、増強。ま
た、肝星細胞の活性化に関係する代表的なサイトカインは肝星細胞の増殖や遊走に
関与する血小板由来増殖因子 platelet-derived growth factor (PDGF) 、増殖に関与
する transforming growth factor-β1 (TGF-β1) 、肝星細胞の収縮に関与するendothelin、ケモカインなどに分けられる 14-16。
図 3
図 3. 肝星細胞と障害肝における活性化 (文献 17)
16
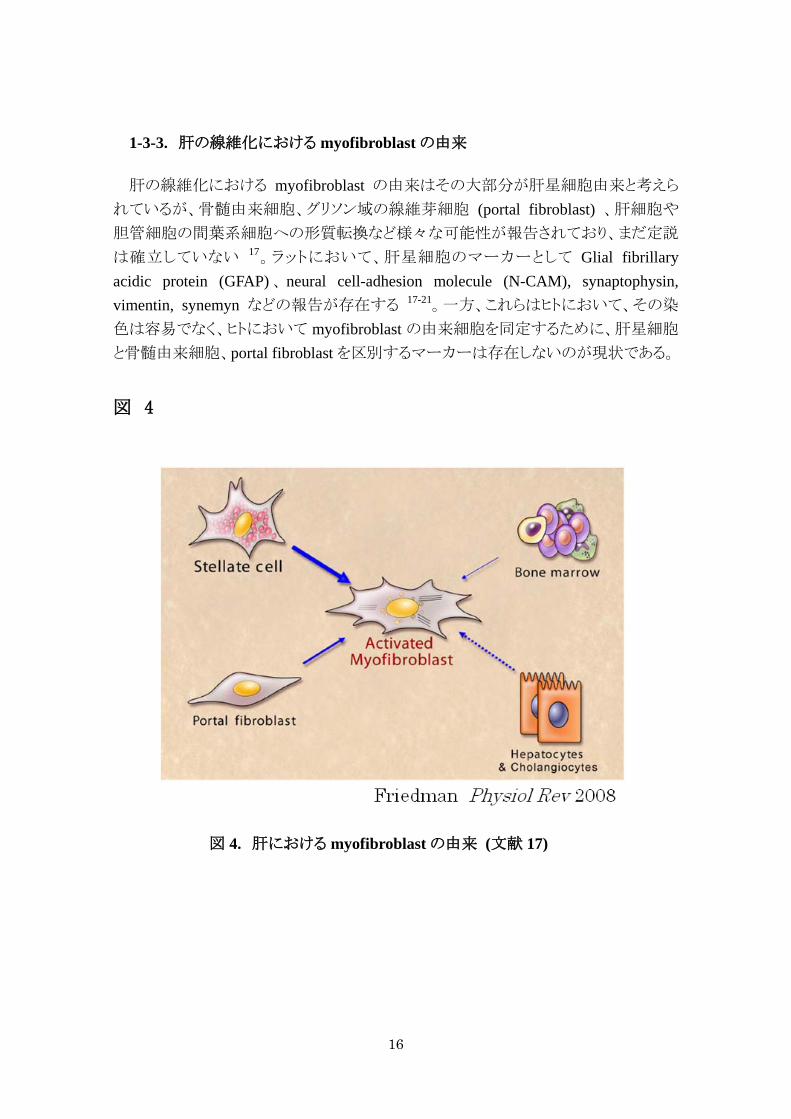
1-3-3. 肝の線維化における myofibroblast の由来
肝の線維化における myofibroblast の由来はその大部分が肝星細胞由来と考えら
れているが、骨髄由来細胞、グリソン域の線維芽細胞 (portal fibroblast) 、肝細胞や
胆管細胞の間葉系細胞への形質転換など様々な可能性が報告されており、まだ定説
は確立していない 17。ラットにおいて、肝星細胞のマーカーとして Glial fibrillary acidic protein (GFAP) 、 neural cell-adhesion molecule (N-CAM), synaptophysin, vimentin, synemyn などの報告が存在する 17-21。一方、これらはヒトにおいて、その染
色は容易でなく、ヒトにおいて myofibroblast の由来細胞を同定するために、肝星細胞
と骨髄由来細胞、portal fibroblast を区別するマーカーは存在しないのが現状である。
図 4
図 4. 肝における myofibroblast の由来 (文献 17)
17
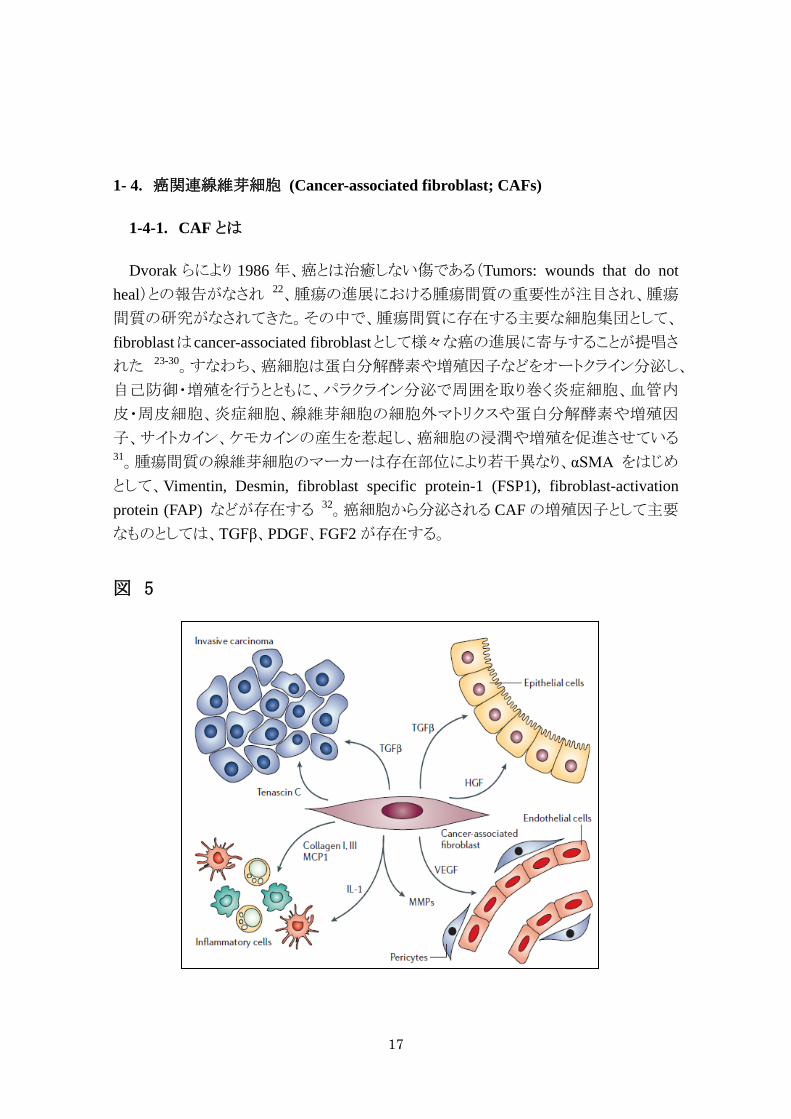
1- 4. 癌関連線維芽細胞 (Cancer-associated fibroblast; CAFs)
1-4-1. CAF とは
Dvorak らにより 1986 年、癌とは治癒しない傷である(Tumors: wounds that do not heal)との報告がなされ 22、腫瘍の進展における腫瘍間質の重要性が注目され、腫瘍
間質の研究がなされてきた。その中で、腫瘍間質に存在する主要な細胞集団として、
fibroblastはcancer-associated fibroblastとして様々な癌の進展に寄与することが提唱さ
れた 23-30。すなわち、癌細胞は蛋白分解酵素や増殖因子などをオートクライン分泌し、
自己防御・増殖を行うとともに、パラクライン分泌で周囲を取り巻く炎症細胞、血管内
皮・周皮細胞、炎症細胞、線維芽細胞の細胞外マトリクスや蛋白分解酵素や増殖因
子、サイトカイン、ケモカインの産生を惹起し、癌細胞の浸潤や増殖を促進させている 31。腫瘍間質の線維芽細胞のマーカーは存在部位により若干異なり、αSMA をはじめ
として、Vimentin, Desmin, fibroblast specific protein-1 (FSP1), fibroblast-activation protein (FAP) などが存在する 32。癌細胞から分泌される CAF の増殖因子として主要
なものとしては、TGFβ、PDGF、FGF2 が存在する。
図 5

18
図 5. 癌の進展における癌関連線維芽細胞の役割 (文献 31)
1-4-2. 固形癌における CAF の臨床的意義
乳癌 :乳癌においては線維芽細胞の80 %はαSMAを発現し、活性化されている 33-35。
45 名の乳癌患者において、免疫組織学的に腫瘍間質に myofibroblast を多く含む群
は癌細胞における Ki67, VEGF, bFGF の発現が高値で、生存率・無再発生存率が不
良であった 36。
結腸直腸癌 :Stage2、3 の 192 例の結腸直腸癌症例を用いて、免疫組織学的に
myofibroblast を定量化すると、αSMA 高発現群は低発現群に比べ無再発生存率が
不良であり、再発に寄与する因子の多変量解析で αSMA 高発現が独立因子となった。
また、原発巣と肝転移巣において αSMA の発現率は症例毎に非常に近似しており、
αSMA の発現率(間質の CAF)は腫瘍の保持する特性の一つである可能性が示唆さ
れた 37。
膵癌 :膵癌における αSMA 発現細胞は GFAP や desmin を発現し、膵星細胞由来で
あることが報告されており 38、CAF として fibronectin や collagen を産生し、癌の進展を
促進させている 39。近年、膵星細胞の CAF としての役割が注目され、ヒト組織より分
離した膵星細胞と膵癌細胞株を用いて、in vitro、in vivo で癌細胞の増殖・浸潤・転移
における細胞間の相互作用のメカニズムが少しずつ明らかにされてきている 40-48。膵
臓の慢性炎症において重要な PDGF や TGFβ などが相互作用に関与する重要な因
子として示されている。
肝細胞癌 :Schmitt らは腫瘍間質に存在する細胞における retinolbinding protein-1 (CRBP-1) の発現が低発現の群がより再発をきたし、活性化された肝星細胞が癌の進
展に関与する可能性を報告した 49。また、腫瘍間質の collagen-1 と MMP2 の mRNAの発現高値が早期再発と関係することや、腫瘍間質で collagen を発現する細胞の大
部分が αSMA を発現し、CD34 を発現しないことが報告されている 50。
1-4-3. 癌細胞と肝星細胞の相互作用に関する報告
肝星細胞の細胞株で樹立されているものは未だ 3 株しかなく、癌細胞との相互作用
を調べた報告はほとんど見られない。ラットを用いたmelanoma肝転移モデルにおいて、
GFAP、desmin を用いて、肝転移腫瘍内に肝星細胞が存在すること、また肝星細胞が
VEGF を産生し、血管新生を促すことが報告された 51-53。また、ヒト肝細胞癌細胞株と

19
ヒト組織より分離した肝星細胞を用いて、腫瘍形成や癌細胞の細胞内シグナルが肝星
細胞によって促進されることが示された 54。ヒト大腸癌細胞株と肝星細胞株 LI90 を用
いた in vivo における腫瘍形成においては、肝星細胞の分泌する SDF-1 が大腸癌の
chemokine receptor4 (CXCR4) に作用し、大腸癌細胞の増殖・浸潤能に関与するとの
報告もみられる 55。これらの報告において、肝星細胞は myofibroblast すなわち CAFとして癌の進展に加担している。
1- 5. 今研究の目的
これまで肝内胆管癌の進展において肝星細胞の関与を調べた報告はなく、また肝
星細胞と胆管癌細胞を用いた in vitro、in vivo における相互作用に関する報告もない。
我々は肝星細胞が肝の線維化において最も重要な細胞であるばかりでなく、胆管癌
の腫瘍間質の構成細胞であり、癌の進展を促進するとの仮説を立てた。本研究ではヒ
ト肝内胆管癌に肝星細胞が存在し癌の進展に関与するかについて調べ、また細胞を
用いた両者の相互作用とそれを制御する因子を解明することを目的とする。

20
第二章 患者対象、実験材料と方法
本実験を行うにあたり、さまざまな分子生物学的手法を用いたため、その方法を以
下に示す。臨床検体の取り扱いに関しては、倫理委員会に従って患者の同意のもと、
その保存・研究を行った。動物実験を行うにあたり、熊本大学 animal care guideline の
記載事項を順守した。
2- 1. 患者対象
免疫組織学的検討を行った ICC の患者対象は、1993 年~2008 年までに当院にて
肝切除を施行した 55 例のうち、肉眼的癌遺残を認める R2 症例を除いた 46 例を対象
とした。TMN 分類で、StageⅠが 22 例、Ⅱが 1 例、ⅢA が 5 例、ⅢC が 9 例であった。
年齢の中央値は 66 歳 (範囲:26 - 81 歳)、男性が 28 名、術後の平均観察期間は
40.5±34.1 ヶ月 (範囲:3 - 126.3 ヶ月)であった。
また、Myofibroblastが存在する陽性コントロールとして肝硬変の線維化の程度をあ
らわす Stage 分類(新犬山分類)を元に同時期に肝切除を施行した肝細胞癌 62症例の非癌部を評価・抽出した。新犬山分類では F0:線維化無し、F1:線維性
門脈域の拡大、F2:線維性架橋の形成、F3:小葉構造の破壊を伴う線維性架橋
の形成、F4:肝硬変と定義した。
2- 2. 免疫組織学的解析
2-2-1. 免疫染色手法 (パラフィン包埋組織)
肝内胆管癌組織は肝切除後 10 % ホルムアルデヒドにて 1 週間固定し、パラフィン
包埋を行った。3μm の厚さで薄切を行い、組織切片を作成した。Hematoxylin-eosin (HE) 染色と以下の免疫染色を行った。脱パラフィンを行い、120℃ 15 分 オートクレ
ーブで抗原の賦活化を行った。3%過酸化水素水にて内因性ペルオキシダーゼの除
去を 5 分間行った後、1 次抗体 (Mouse-monoclonalαSMA antibody (×100, DAKO), Mouse-monoclonal CD34 antibody (×50, DAKO), Goat-polyclonal CXCL5 antibody (×50, R&D) と 4℃ overnight で抗原抗体反応を行った。洗浄の後、2 次抗体

21
(mouse は Envision Plus (DAKO)、Goat は Anti-IgG HRP conjugated (×100, SantaCruz) ) と 1 時間反応させ、DAB にて発色を行い、ヘマトキシリン染色後封入を
行った。陰性コントロールに一次抗体の代わりに、マウス IgG を用いた。また、コラーゲ
ンの染色を行うため、同対象の症例組織切片を用いて、脱パラフィン後 Serius Red 染
色(Sigma, Japan)も施行した。
2-2-2. 癌間質、慢性肝炎における myofibroblast の定量
Myofibroblastの定量を行うために、αSMAの発現の定量を行った。ICCの腫瘍内か
ら 10 視野 (×200)をランダムに選択し評価を行った。解析ソフト VH Analyzer(KEYENCE Co., Japan)を用いて、αSMA 染色陽性領域を単一カラーに置換し、全視
野に占めるその面積比を算出した。
さらに、αSMA が Myofibroblast を検出し、これを定量化していることを確認す
るために、星細胞が myofibroblast として存在する慢性肝炎、肝硬変において検
証を行った。肝細胞癌切除症例のサンプルを、αSMA による免疫染色を行い、
背景肝における発現の定量化を行った。ICC における myofibroblast の定量化と
同様の方法で、全視野に占める myofibroblast の面積比を算出した。肝の線維化
の分類には新犬山分類を用いた 56。
表2. 新犬山分類 (慢性肝炎の肝組織診断基準 F0 線維化無し F1 門脈域の線維性拡大 F2 線維性の架橋形成 F3 小葉の歪みを伴う繊維性架橋形成 F4 肝硬変
2-2-3. 二重免疫染色 (凍結保存組織)
肝内胆管癌組織を肝切除後クリオモルドに包埋し、-80℃保存したものを使用
した。6μm の厚さで薄切を行い、4℃アセトン固定し風乾した。また、PBS にて
洗浄後、10 % PFA+1 % CaCl2 にて 2 分固定した。3 % BSA-TBS にて 15 分間ブ
ロッキング後、1 次抗体(Rabbit anti-GFAP antibody and mouse anti-αSMA antibody, rabbit anti-αSMA and Mouse anti-desmin antibody)を 1 時間反応させ、洗浄後暗室
にて 2 次抗体(Goat- anti mouse IgG (Hilyte fluor 555, 488), Goat-anti rabbit IgG (Hylite flour 555, 488))を 1 時間反応させた。洗浄後 DAPI にて核染色を行い、

22
封入を行った。
2- 3. 細胞培養
胆管癌細胞株 HuCCT-1, MEC は東北加齢研究所より、RBE は理研バイオリソ
ースセンターより入手した。肝星細胞株 LI90 は Japanese Collection of Research Bioresources (JCRB)より入手し 57、LX-2 は Scott L. Friedman (Division of Liver Disease, Department of Medicine, Mount Sinai School of Medicine, NewYork, NY)より分譲して頂いた 58。いずれも培養液は DMEM もしくは RPMI に 10%FBS を添
加し、5% CO2 下に 37 度で培養を行った。
2- 4. 共培養モデル
2-4-1. 細胞浸潤アッセイ
Matriegel invasion chamber (pore size 8μm、BD Biosciences, Japan)を用いて、
24well plate にて行った。まず肝星細胞株 LI90 または LX-2 細胞株を 2×104 個/wellで播いて、10%FBS を含む medium 下に overnight で培養し、750μl の同培養液に
交換した。そこに chamberをのせて、5×104個/wellのHuCCT1細胞株(または 1×105
個/well)を FBS free の medium 500μl で加えた。 コントロールは下層の 24well plate に細胞をいれず、10%FBS 入り medium のみとした。37 度で 22 時間培養し、
綿棒にて chamber の上面を拭き上げ細胞を除去した後、100%メタノール固定を
行い、1%トルイジンブルーにて染色を行い、chamber の下面へ浸潤している細
胞数を顕微鏡でカウントした。5 視野 (×200)でカウントした細胞数の平均個数
を比較した。同実験を 3 回行い、結果を確認した。
2-4-2. 細胞増殖アッセイ
6 well plate に HuCCT-1 を 1×105 個/well または MEC を 1×105 個/well 播いて、
Cell culture incer t (pore size1μm、BD Biosciences, Japan)を装着し、その中に LI90を 1×104 個播いて 10%FBS を含まない medium 下に 37℃で培養した。コントロ
ールは、Cell culture incert を用いない 6well plate とし、同数の癌細胞を準備した。
48 時間、72 時間、96 時間、120 時間に WST-8 (DOJIN Laboratories, Japan)を加え、
プレートリーダーにて OD を測定した。同実験を 3 回行い、結果を確認した。

23
2-4-3. 細胞遊走アッセイ
12 well plateにHuCCT-1を1×105個/well播いて、24時間後に scratchした。Scratchと同時に無血清培養液に Human recombinant CXCL5 (Goat, R&D)を 30 ng/ml 加え、
コントロールは Goat 血清を同濃度加え 12 時間後に判定した。判定時に 1%トル
イジンブルー染色を行い、遊走した細胞の面積を ImageJ (NIH)で定量化し、
Control との比を算出した。同実験を 3 回行い、結果を確認した。
2-4-4. サイトカインアレイ
HuCCT1 と LI90 を用いて、細胞増殖アッセイと同じ条件下に共培養を行い、
36 時間の時点で培養上清を回収し、HuCCT1、LI90、両者の共培養から抽出した
Conditioning medium を遠心濃縮キット(Millipore: 3 kDa 用)にて同量に濃縮
し、使用するまで-80 度保存した。36 種類の炎症に関連するサイトカイン・ケモ
カイン(下記)を検出するキット(R&D)を用いて、同量の Conditioning medium 3 種類をそれぞれニトロセルロースメンブレンに Overnight で反応させた。洗浄
の後、Streptavidin-HRP と 30 分間反応させ、発色は ECL Plus (GE Healthcare)で行
った。画像取り込みは Multigague software (FUJIFILM) にて行い、Spot の強度を
定量数値化し比較を行った。
表 3.サイトカイン・ケモカイン(36 種類)
RANTES (CCL5) CD40 Ligand (CD154) IL-2 I-309 (CCL1) IL-32α TNF-α GM-CSF MIP-1β (CCL4) IL10 (CXCL10) I-TAC (CXCL11) MIF IL-23 IL-1α IL-13 IL-17 IL-6 C5a IL-1ra IL-4 sICAM-1 SDF-1 (CXCL12) G-CSF CXCL1 SerpinE1 IP10 (CXCL10) sTREM-1 CCL3 IL-27 IL-12 p70 MCP-1 (CCL2) IL-17E IL-8 (CXCL8) IL-1β IL-16 (LCF) IL-5 INF-γ

24

25
2- 5. 培養上清を用いた実験
2-5-1. 培養上清の準備
LI90 細胞が 80%程度に増殖した後、培養液を無血清培養液に交換し、48 時間
後に回収した。1500rpm で 5 分間遠心して細胞を除去した後、Amicon Ultra filters (Millipore:3kDa 用)を用いて遠心(4000g、60 分間)を行った。培養上清の濃度
は Bradford assay にて確認し、使用するまで-80 度保存した。2 種類の胆管癌細胞
からの培養上清も同様に作成保存した。
2-5-2. 培養細胞の免疫染色
LI90 細胞をカバーガラス上に播き、8 時間後に培養上清を交換した。癌細胞
由来の Conditioning medium (CC-CM、0.25 μg/μl)と、Control として無血清培養液
に交換し 12 時間後に 4%PFA にて 15 分間固定した。3%BSA-TBS にて 15 分間の
ブロッキングを行い、αSMA(mouse-monoclonal、1:100)にて 1 時間室温にて反
応後、暗室で 2 次抗体(Hylite Fluor 555)と反応させスライドガラスに封入した。
Negative control として 1 次抗体の代わりにマウス IgG を用いて同様に染色を行
った。観察・撮影は FV300 蛍光顕微鏡 (Olympus) を用いた。
2-5-3. ウエスタン・ブロット法
胆管癌細胞の CM により星細胞が活性化するかを確認するために、6-well plate に星細胞を播いて 8 時間後に、培養液を無血清のものと CC-CM (0.5 μg/μg)に交換した。12 時間の incubation の後、細胞から蛋白を抽出した。 また、星細胞の CM を加えることにより胆管癌細胞の細胞内シグナルの変化
を確認するため、RBE 細胞株を用いて、6well plate に細胞を播き、80 % confluent になった時点で、培養液を無血清液に交換した。6 時間後、培養液を
肝星細胞培養上清 HS-CM (0.5μg/μg) または Control として無血清培養液に交
換し、5 分、15 分、30 分、60 分、120 分の時点でそれぞれ蛋白を回収した。 細胞からの蛋白抽出には lysis buffer [25 mM Tris (pH7.4), 100 mM NaCl,
1% Tween 20] を用いた。10 %ゲルで SDS-PAGE を行い、PVDF メンブレンへ
転写した。5% スキムミルク-TBSTで 1時間ブロッキングを行い、1次抗体と 4℃で 1 晩反応させた。洗浄の後、2 次抗体と 1 時間室温で反応させ、化学発色基質
(ECL;GE Healthcare)で発色した。
26
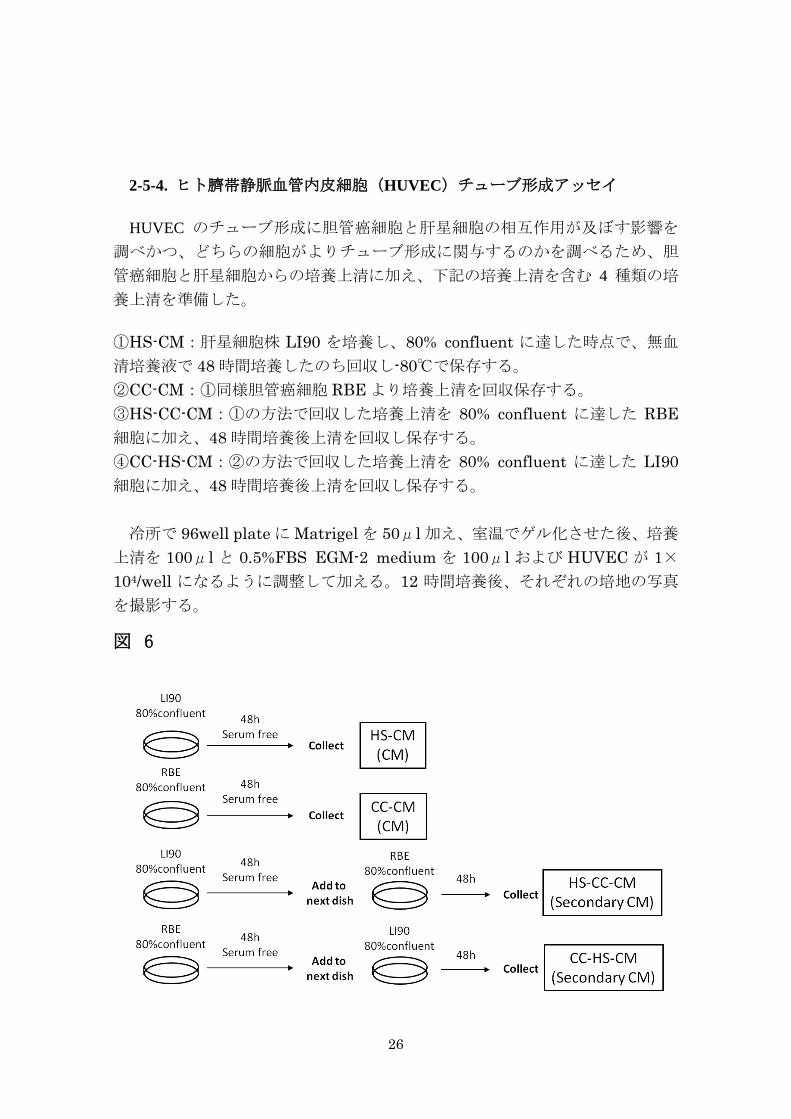
2-5-4. ヒト臍帯静脈血管内皮細胞(HUVEC)チューブ形成アッセイ
HUVEC のチューブ形成に胆管癌細胞と肝星細胞の相互作用が及ぼす影響を
調べかつ、どちらの細胞がよりチューブ形成に関与するのかを調べるため、胆
管癌細胞と肝星細胞からの培養上清に加え、下記の培養上清を含む 4 種類の培
養上清を準備した。
①HS-CM:肝星細胞株 LI90 を培養し、80% confluent に達した時点で、無血
清培養液で 48 時間培養したのち回収し-80℃で保存する。 ②CC-CM:①同様胆管癌細胞 RBE より培養上清を回収保存する。 ③HS-CC-CM:①の方法で回収した培養上清を 80% confluent に達した RBE細胞に加え、48 時間培養後上清を回収し保存する。 ④CC-HS-CM:②の方法で回収した培養上清を 80% confluent に達した LI90細胞に加え、48 時間培養後上清を回収し保存する。 冷所で 96well plate に Matrigel を 50μl 加え、室温でゲル化させた後、培養
上清を 100μl と 0.5%FBS EGM-2 medium を 100μl および HUVEC が 1×104/well になるように調整して加える。12 時間培養後、それぞれの培地の写真
を撮影する。
図 6

27
図 6. 胆管癌細胞株、肝星細胞株、および相互作用を行った培養上清の準備
2-5-5. アポトーシスアッセイ (TUNEL 法)
HuCCT-1 細胞を 1×105 個/well 8-well chamber に播き、1 晩 incubation した後、
培養液を肝星細胞培養上清 HS-CM (0.5μg/μg) または Control として無血清培養
液に交換し、それぞれ CDDP (50 μg/mL)、5FU (500 μg/mL)、Gemcitabine (500 μg/mL)を添加した。24 時間後、TUNEL 法 (Promega) にて染色を行い、アポト
ーシス細胞の数を数えその割合を算出した。Hematoxylin 染色で核染色を行った。
同実験は 3 回行い、カウントは無作為に 10 視野を数えた。
2- 6. In vivo における腫瘍形成能の比較と組織学的評価
6-8 週齢のヌードマウス (BALB/c nu/nu) の皮下に下記 3 群の細胞を注入し、
腫瘍形成能を比較した。各群 5 匹ずつのマウスを使用した。
1)胆管癌細胞 HuCCT1:3×106個/100 μL 2)肝星細胞 LI90:1.5×106個/100 μL 3)胆管癌細胞 HuCCT1:(3×106個+肝星細胞 LI90:1.5×106個) /100 μL
細胞を注入してから 7 週間後にマウスを sacrifice し、腫瘍を摘出して組織学的検
討を行うため凍結保存とパラフィン固定した。腫瘍体積は 1/2 × 長径 (mm) ×短径 (mm) × 厚さ (mm) で算出した。
パラフィン固定した組織は方法 2-2-1 と同様の方法で免疫染色を行い、1 次抗
体として Rat anti-mouse CD31 antibody (Biolegend) を用いた。間質の評価に Sirius Red も用いた。
2- 7. 質量分析を用いた蛋白同定
2-7-1. SELDI-TOF MS 解析
共培養モデルを用いて以下の 3 種類の CM を回収し濃縮したサンプルを
Surface - Enhanced Laser Desorption and Ionization Time of Flight - Mass Spectrometry (SELDI-TOF MS) (CIPHERGEN)を用いて解析した。CM10pH 7 と

28
Q10pH 7 の 2 種類のプロテインチップを使用し、4 通りの組み合わせの共培養か
ら得られた計 12 種類の CM を方法 2-5-1 の通り濃縮後測定に用いた。プロテイ
ンチップを 50 mM リン酸バッファーで 27℃ 5 分 2 回洗浄した後、CM 100μlをチップへ 90 分間反応させた。洗浄を 3 回繰り返した後、シナピン酸を反応さ
せ、SELDI で測定した。
2-7-2. Q-TOF/MS/MS 解析
HuCCT1、LI90、それらを共培養した CM をそれぞれ低分子量用グラディエン
トゲル (BIO-RAD)を用いて電気泳動を行い、CBB 染色を 90 分間行い、分子量
8360 と思われる band を切り出し、受託解析で Q-TOF/MS/MS 解析(北海道シス
テムサイエンス)を行った。NanoLC-ESI-還元アルキル化後にトリプシンを用い
たゲル内消化を行い、ペプチドを抽出し、濃縮後 Q-TOF/MS/MS で測定した。得
られたペプチド情報を Mascot でヒトデータベース検索を行った。
2-7-3. 蛋白ビーズを用いた免疫沈降
HuCCT1 と LI90 を共培養した CM を濃縮後解析に使用した。Dynabeads Protein A(Invitrogen)を 50 mM リン酸バッファーで 3 回洗浄した後、control IgG (Goat IgG, R&D)と Goat anti CXCL5 antibody (R&D)をビーズと 10 分間反応させた。50 mM リン酸バッファーで 4 回洗浄した後、サンプルとビーズをそれぞれ室温で 1時間 反応させた。ビーズを除去した上清を SELDI で測定した。
2- 8. 統計学的解析
統計学的解析には Stat View J-5.0 を用いた。群間比較には Student’s t 検定また
はχ2 検定を用い、p < 0.05 を有意な差とした。2 変量の相関検定には Spearmanの順位相関係数検定を用いた。生存率の算出には Kaplan-Meier 法を用い、単変
量解析には log-rank 法を用いた。Cox 比例ハザードモデルを用いて、生存に関す
る多変量解析を行った。多群の比較には ANOVA (Turkey-Kramer post hoc test) を用いた。
29
第三章 実験結果
3- 1. 肝内胆管癌の免疫組織学的検討
3-1-1. 肝内胆管癌におけるαSMA、CD34 の発現
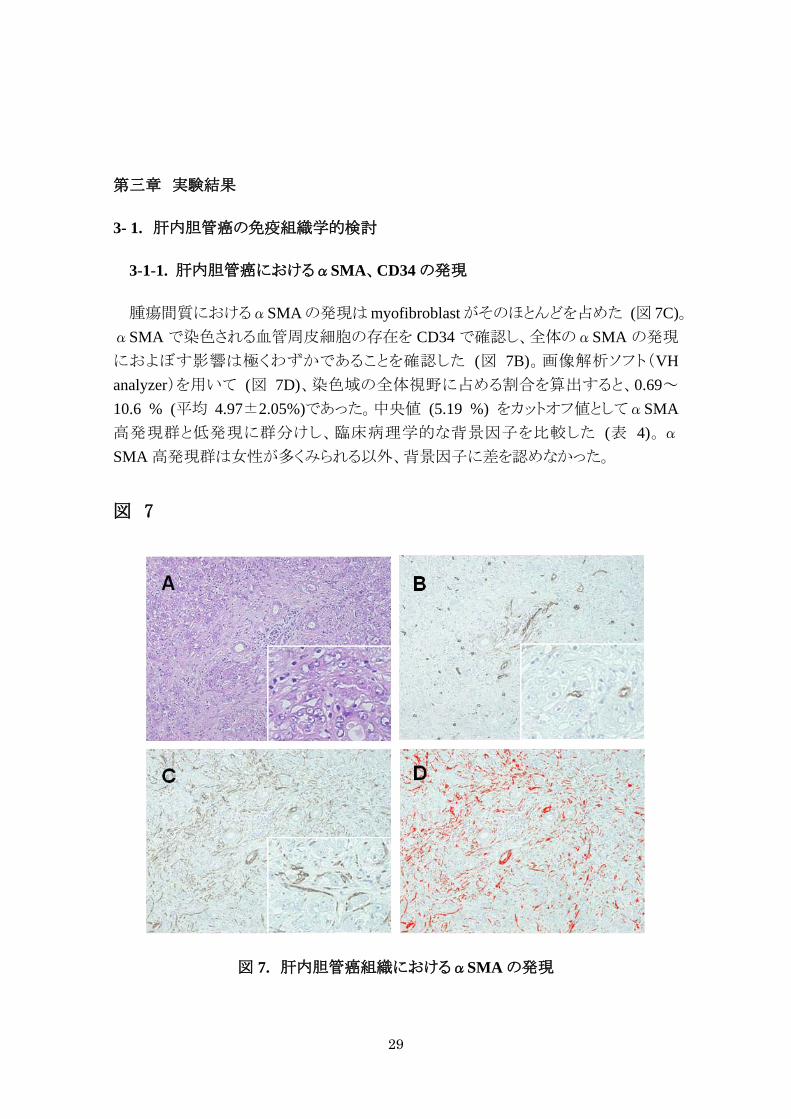
腫瘍間質におけるαSMA の発現は myofibroblast がそのほとんどを占めた (図 7C)。αSMA で染色される血管周皮細胞の存在を CD34 で確認し、全体のαSMA の発現
におよぼす影響は極くわずかであることを確認した (図 7B)。画像解析ソフト(VH analyzer)を用いて (図 7D)、染色域の全体視野に占める割合を算出すると、0.69~10.6 % (平均 4.97±2.05%)であった。中央値 (5.19 %) をカットオフ値としてαSMA高発現群と低発現に群分けし、臨床病理学的な背景因子を比較した (表 4)。αSMA 高発現群は女性が多くみられる以外、背景因子に差を認めなかった。
図 7
図 7. 肝内胆管癌組織におけるαSMA の発現
30
図 5. 上部写真は肝内胆管癌組織の免疫組織学的撮影。(A) HE、(B) CD34、(C) α
SMA、(D) αSMA の染色域を、ソフトを用いて単一色(赤)へ置換したものである。
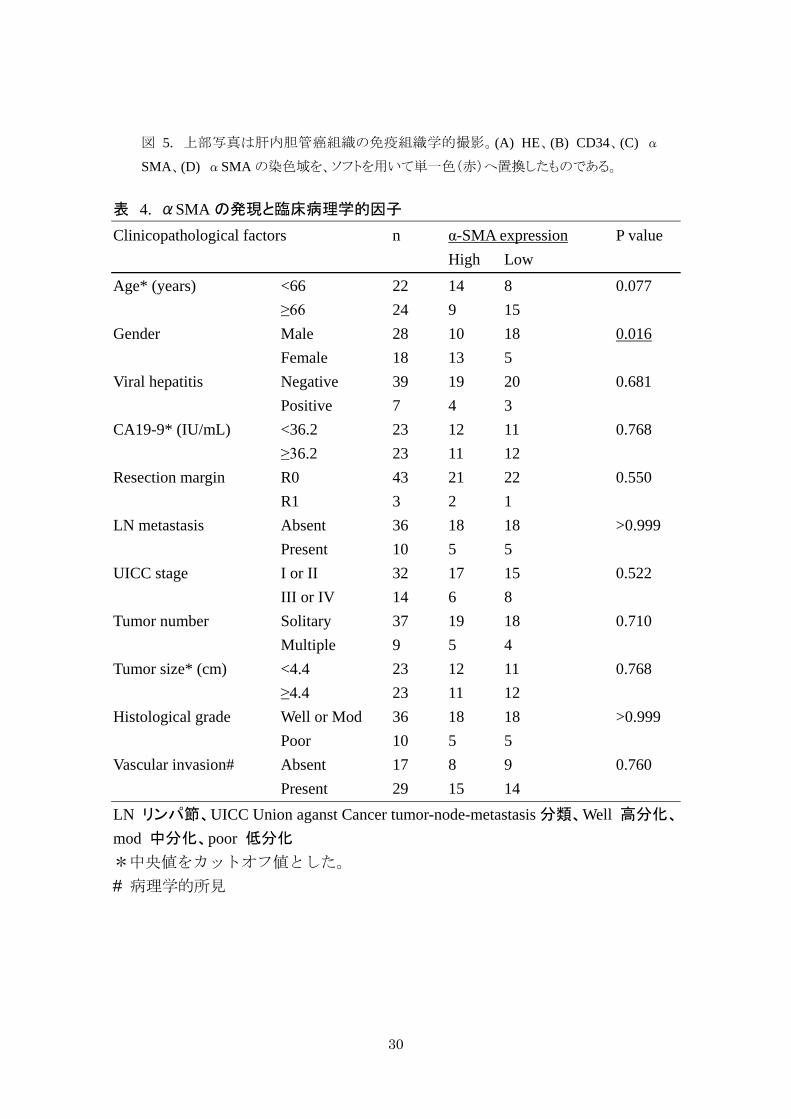
表 4. αSMA の発現と臨床病理学的因子
Clinicopathological factors n α-SMA expression P value High Low
Age* (years) <66 22 14 8 0.077 ≥66 24 9 15 Gender Male 28 10 18 0.016 Female 18 13 5 Viral hepatitis Negative 39 19 20 0.681 Positive 7 4 3 CA19-9* (IU/mL) <36.2 23 12 11 0.768 ≥36.2 23 11 12 Resection margin R0 43 21 22 0.550 R1 3 2 1 LN metastasis Absent 36 18 18 >0.999 Present 10 5 5 UICC stage I or II 32 17 15 0.522 III or IV 14 6 8 Tumor number Solitary 37 19 18 0.710 Multiple 9 5 4 Tumor size* (cm) <4.4 23 12 11 0.768 ≥4.4 23 11 12 Histological grade Well or Mod 36 18 18 >0.999 Poor 10 5 5
Vascular invasion# Absent 17 8 9 0.760 Present 29 15 14 LN リンパ節、UICC Union aganst Cancer tumor-node-metastasis 分類、Well 高分化、
mod 中分化、poor 低分化 *中央値をカットオフ値とした。 # 病理学的所見

31
3-1-2. αSMA を用いた miofibroblast の定量化は有効である
αSMA の染色域を定量化し、障害肝において線維化の程度との相関を評価した。
αSMA scoreはF0-1, F2, F3, F4症例においてそれぞれ0.29, 0.56, 2.33, 4.1 %であり、
線維化の程度に 応じて上昇した (図8)。
図 8 A
B
図 8. 肝の線維化とαSMA の発現強度の関係
32
A. 写真は肝硬変症例をαSMA で免疫染色したものである。B. αSMA の
発現強度と肝の線維化の程度の関係を示したもの。* は統計学的に有意差
があることを示す (* p < 0.05)。
3-1-3. 腫瘍間質の Myofibroblast とコラーゲン量は相関しない
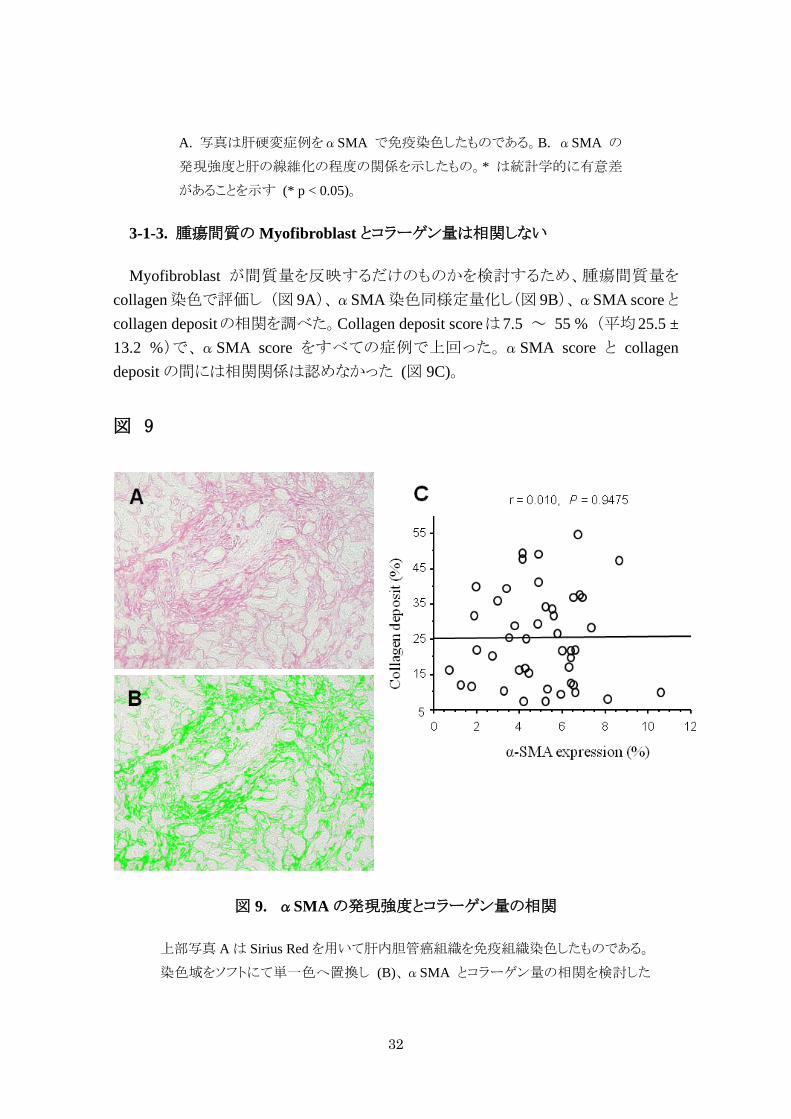
Myofibroblast が間質量を反映するだけのものかを検討するため、腫瘍間質量を
collagen 染色で評価し (図 9A)、αSMA 染色同様定量化し(図 9B)、αSMA score とcollagen depositの相関を調べた。Collagen deposit scoreは7.5 ~ 55 % (平均25.5 ± 13.2 %)で、αSMA score をすべての症例で上回った。αSMA score と collagen deposit の間には相関関係は認めなかった (図 9C)。
図 9
図 9. αSMA の発現強度とコラーゲン量の相関
上部写真 A は Sirius Red を用いて肝内胆管癌組織を免疫組織染色したものである。
染色域をソフトにて単一色へ置換し (B)、αSMA とコラーゲン量の相関を検討した

33
(C)。r は相関係数。
34
3-1-4. 肝内胆管癌において myofibroblast は生存予後に関与する
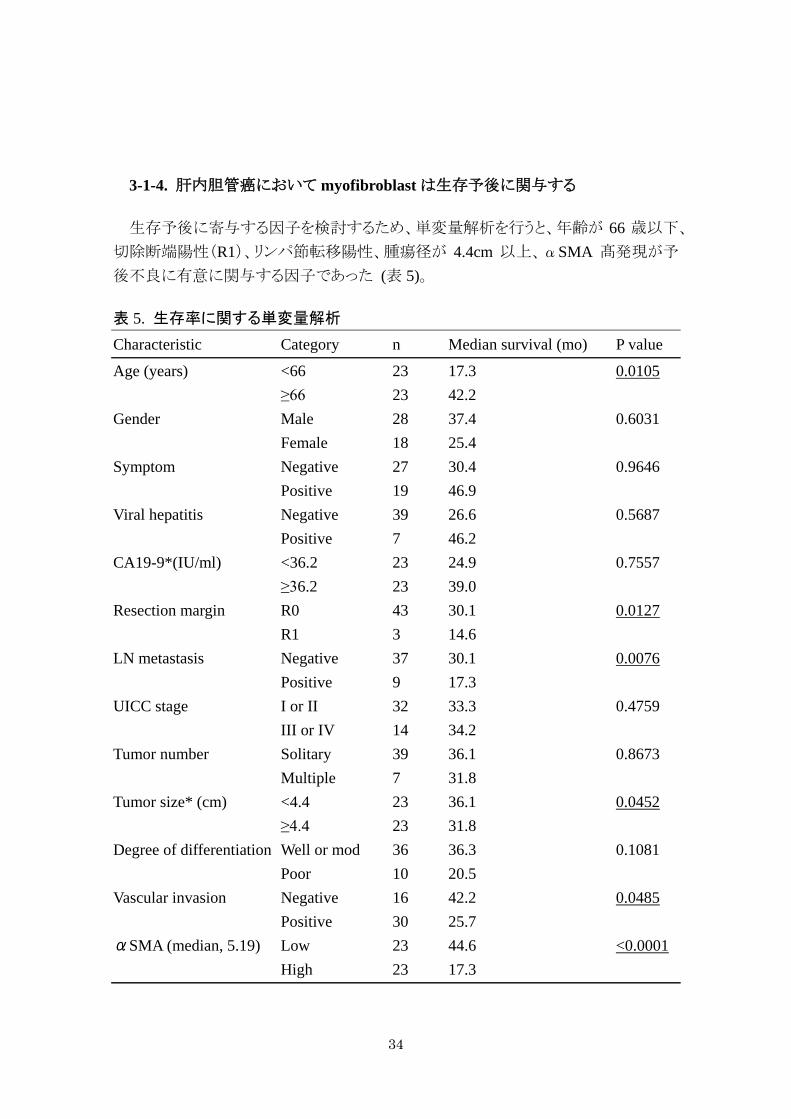
生存予後に寄与する因子を検討するため、単変量解析を行うと、年齢が 66 歳以下、
切除断端陽性(R1)、リンパ節転移陽性、腫瘍径が 4.4cm 以上、αSMA 髙発現が予
後不良に有意に関与する因子であった (表 5)。
表 5. 生存率に関する単変量解析
Characteristic Category n Median survival (mo) P value
Age (years) <66 23 17.3 0.0105 ≥66 23 42.2 Gender Male 28 37.4 0.6031 Female 18 25.4 Symptom Negative 27 30.4 0.9646
Positive 19 46.9 Viral hepatitis Negative 39 26.6 0.5687 Positive 7 46.2 CA19-9*(IU/ml) <36.2 23 24.9 0.7557 ≥36.2 23 39.0 Resection margin R0 43 30.1 0.0127 R1 3 14.6 LN metastasis Negative 37 30.1 0.0076 Positive 9 17.3 UICC stage I or II 32 33.3 0.4759
III or IV 14 34.2 Tumor number Solitary 39 36.1 0.8673 Multiple 7 31.8 Tumor size* (cm) <4.4 23 36.1 0.0452 ≥4.4 23 31.8 Degree of differentiation Well or mod 36 36.3 0.1081 Poor 10 20.5 Vascular invasion Negative 16 42.2 0.0485
Positive 30 25.7 αSMA (median, 5.19) Low 23 44.6 <0.0001 High 23 17.3

35
生存率に関する単変量解析にて有意差を認めた因子を用いて、多変量解析を行うと、
αSMA 髙発現とリンパ節転移陽性の 2 因子が生存予後不良に関与する独立因子で
あった (表 6)。αSMA 髙発現群は低発現群と比較して、無再発生存率も不良であっ
た (図 10)。
表 6. 生存率に関する多変量解析
Clinicopathological factors P Risk ratio (95% confidence interval)
High α-SMA expression 0.0015 9.48 (2.36–38.11) Positivity for LN metastasis 0.0109 5.49 (1.48–20.39) Age ≥66 0.3293 1.98 (0.50–7.79) Tumor size ≥ 4.4 0.7209 1.28 (0.33–4.88) R1 0.8575 1.16 (0.23–5.73)
図 10
図 10. 術後生存率と無再発生存率
A は術後生存率、B は術後無再発生存率を示したものである。
36
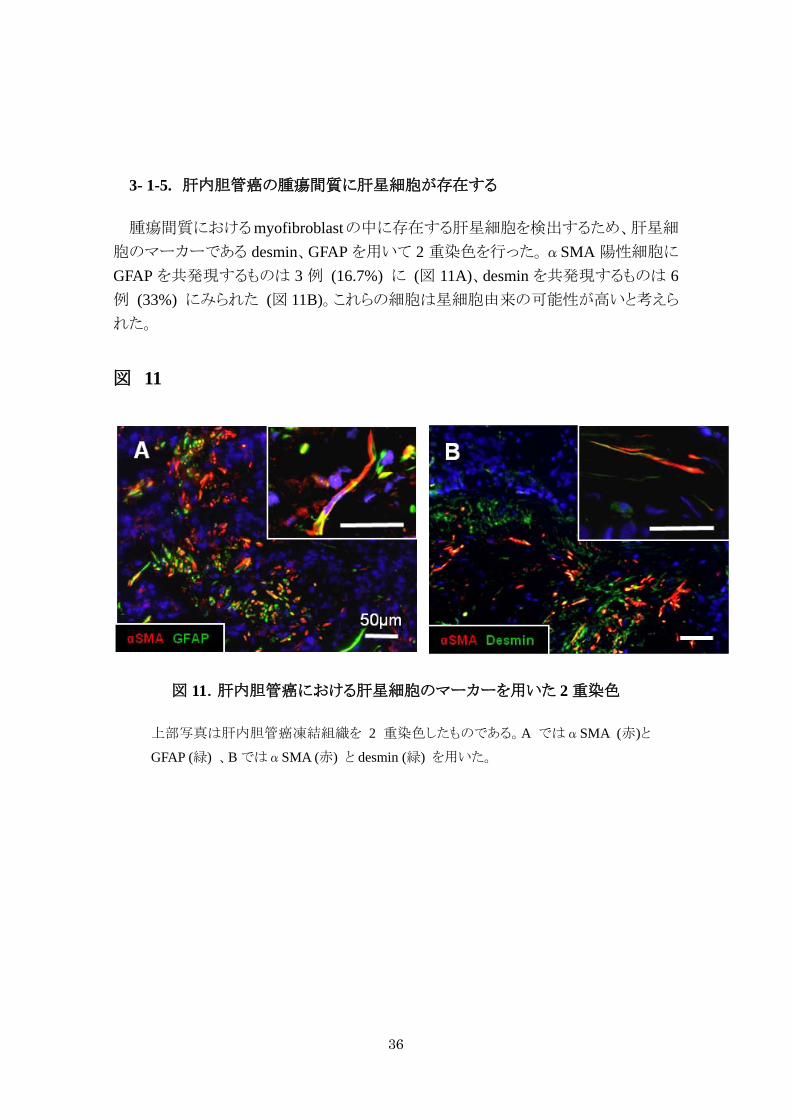
3- 1-5. 肝内胆管癌の腫瘍間質に肝星細胞が存在する
腫瘍間質におけるmyofibroblastの中に存在する肝星細胞を検出するため、肝星細
胞のマーカーである desmin、GFAP を用いて 2 重染色を行った。αSMA 陽性細胞に
GFAP を共発現するものは 3 例 (16.7%) に (図 11A)、desmin を共発現するものは 6例 (33%) にみられた (図 11B)。これらの細胞は星細胞由来の可能性が高いと考えら
れた。
図 11
図 11.肝内胆管癌における肝星細胞のマーカーを用いた 2 重染色
上部写真は肝内胆管癌凍結組織を 2 重染色したものである。A ではαSMA (赤)と
GFAP (緑) 、B ではαSMA (赤) と desmin (緑) を用いた。
37
3- 2. 培養上清を用いた胆管癌細胞と肝星細胞の相互作用の確認
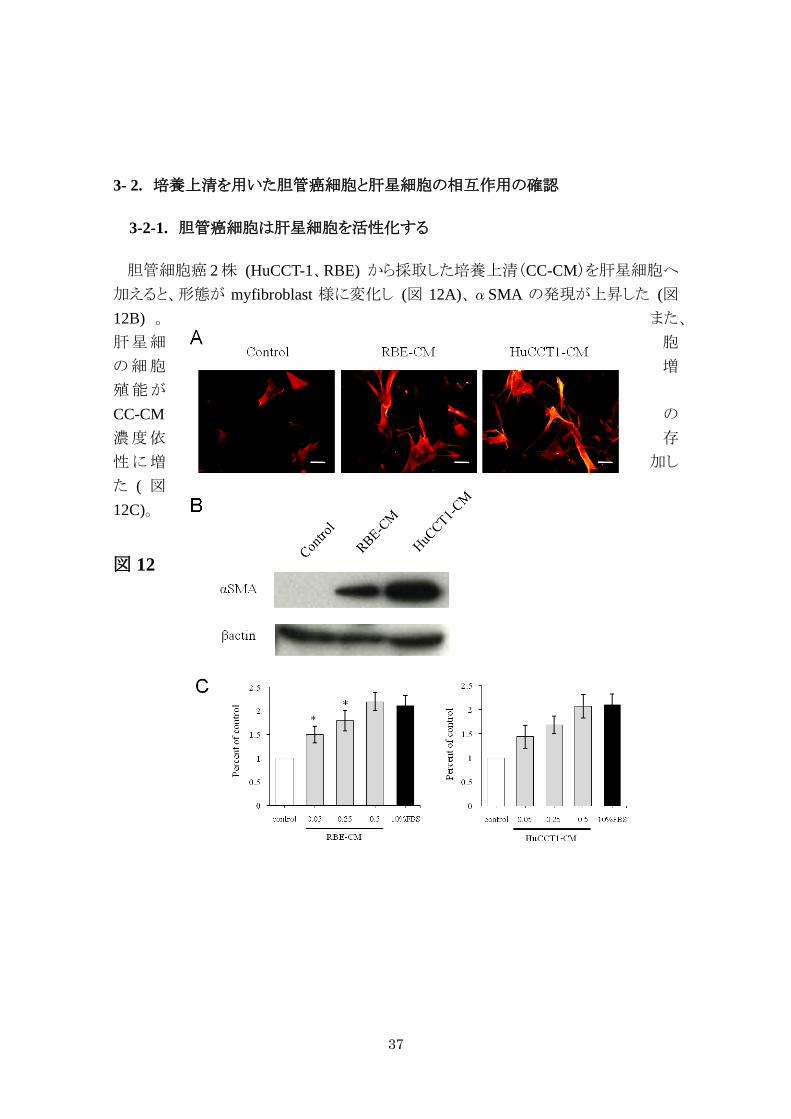
3-2-1. 胆管癌細胞は肝星細胞を活性化する
胆管細胞癌 2 株 (HuCCT-1、RBE) から採取した培養上清(CC-CM)を肝星細胞へ
加えると、形態が myfibroblast 様に変化し (図 12A)、αSMA の発現が上昇した (図12B) 。 また、
肝星細 胞
の細胞 増
殖能が
のCC-CM濃度依 存
性に増 加し
た ( 図
12C)。
図 12

38
図 12. 胆管癌細胞による肝星細胞の形質変化
(A) LI90 に胆管癌細胞株由来の培養上清を加えαSMA で免疫染色、(B) Western
でαSMAの発現を比較、(C) MTT assay、培養上清の濃度を 0.05、0.25、0.5 mg/ml
に調整し比較した。* は統計学的に有意差があることを示す (* p < 0.05)。
3-2-2. 肝星細胞は胆管癌細胞の細胞増殖・浸潤能を促進する
肝星細胞株 (LI90) から採取した培養上清(HS-CM)を胆管癌細胞株 2 種
(HuCCT-1、RBE) へ加えると、無血清培養液を control とした際、胆管癌細胞株の細
胞増殖能 (図 13A) ・浸潤能 (図 13B) が CC-CM の濃度依存性に増加した。
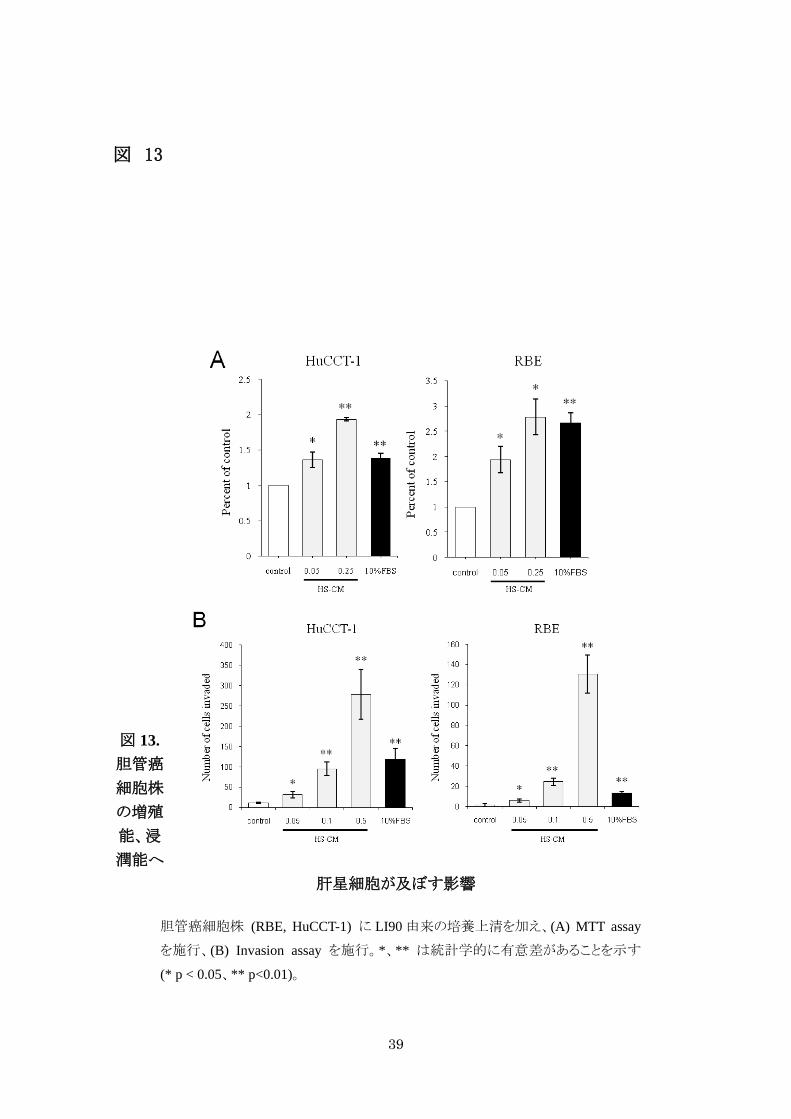
39
図 13
図 13. 胆管癌
細胞株
の増殖
能、浸
潤能へ
肝星細胞が及ぼす影響
胆管癌細胞株 (RBE, HuCCT-1) に LI90 由来の培養上清を加え、(A) MTT assay
を施行、(B) Invasion assay を施行。*、** は統計学的に有意差があることを示す
(* p < 0.05、** p<0.01)。
40
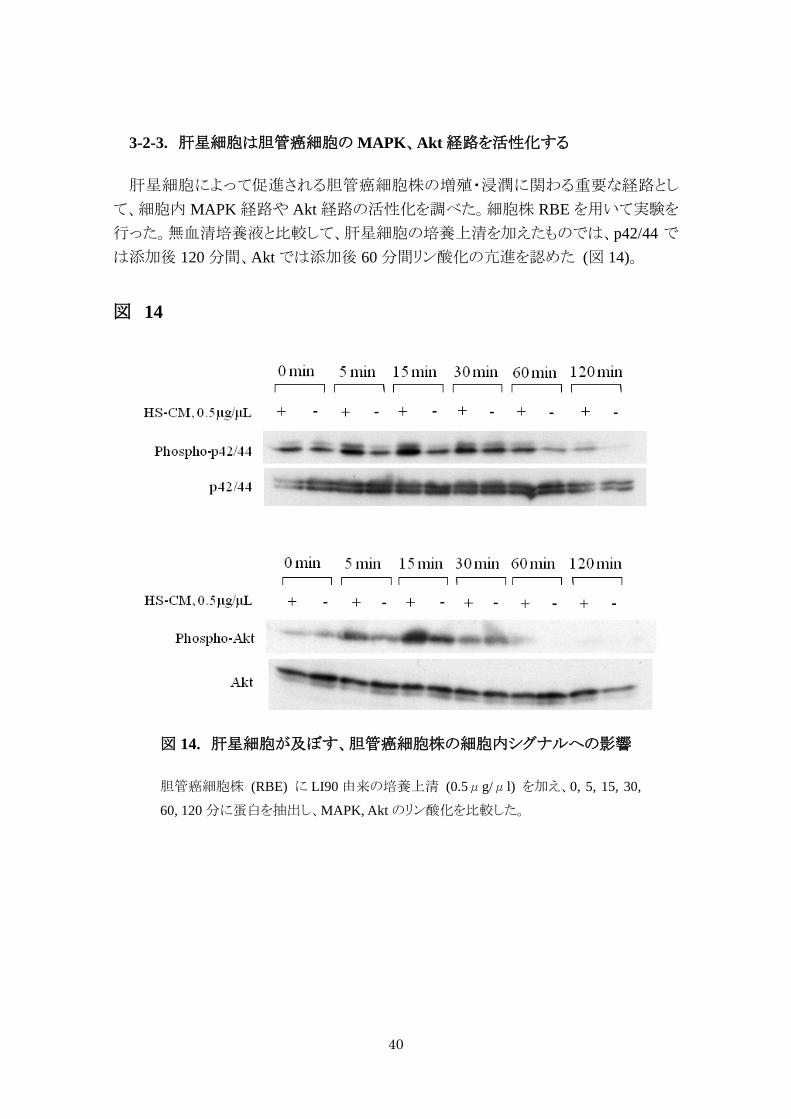
3-2-3. 肝星細胞は胆管癌細胞の MAPK、Akt 経路を活性化する
肝星細胞によって促進される胆管癌細胞株の増殖・浸潤に関わる重要な経路とし
て、細胞内 MAPK 経路や Akt 経路の活性化を調べた。細胞株 RBE を用いて実験を
行った。無血清培養液と比較して、肝星細胞の培養上清を加えたものでは、p42/44 で
は添加後 120 分間、Akt では添加後 60 分間リン酸化の亢進を認めた (図 14)。
図 14
図 14. 肝星細胞が及ぼす、胆管癌細胞株の細胞内シグナルへの影響
胆管癌細胞株 (RBE) に LI90 由来の培養上清 (0.5μg/μl) を加え、0, 5, 15, 30,
60, 120 分に蛋白を抽出し、MAPK, Akt のリン酸化を比較した。
41
3-2-4. 胆管癌と肝星細胞の相互作用によりチューブ形成は促進する
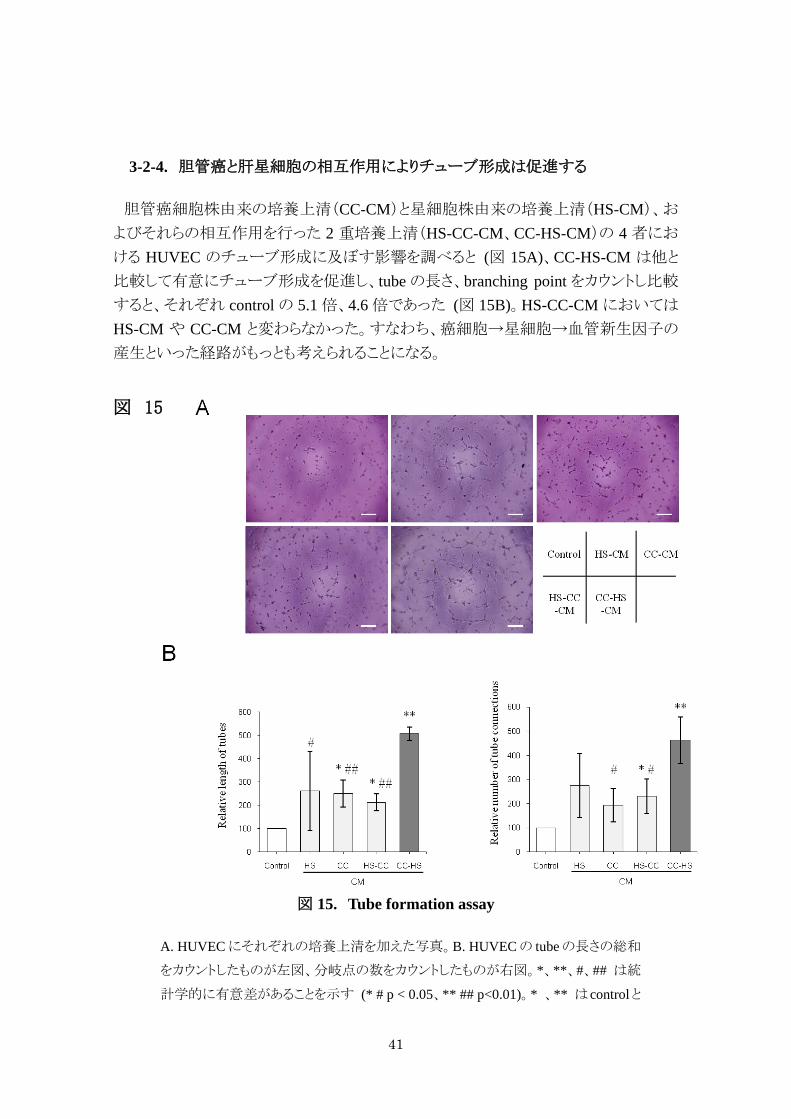
胆管癌細胞株由来の培養上清(CC-CM)と星細胞株由来の培養上清(HS-CM)、お
よびそれらの相互作用を行った 2 重培養上清(HS-CC-CM、CC-HS-CM)の 4 者にお
ける HUVEC のチューブ形成に及ぼす影響を調べると (図 15A)、CC-HS-CM は他と
比較して有意にチューブ形成を促進し、tube の長さ、branching point をカウントし比較
すると、それぞれ control の 5.1 倍、4.6 倍であった (図 15B)。HS-CC-CM においては
HS-CM や CC-CM と変わらなかった。すなわち、癌細胞→星細胞→血管新生因子の
産生といった経路がもっとも考えられることになる。
図 15
図 15. Tube formation assay
A. HUVEC にそれぞれの培養上清を加えた写真。B. HUVEC の tube の長さの総和
をカウントしたものが左図、分岐点の数をカウントしたものが右図。*、**、#、## は統
計学的に有意差があることを示す (* # p < 0.05、** ## p<0.01)。* 、** はcontrolと
42
比較。#、## は CC-HS-CM と比較。
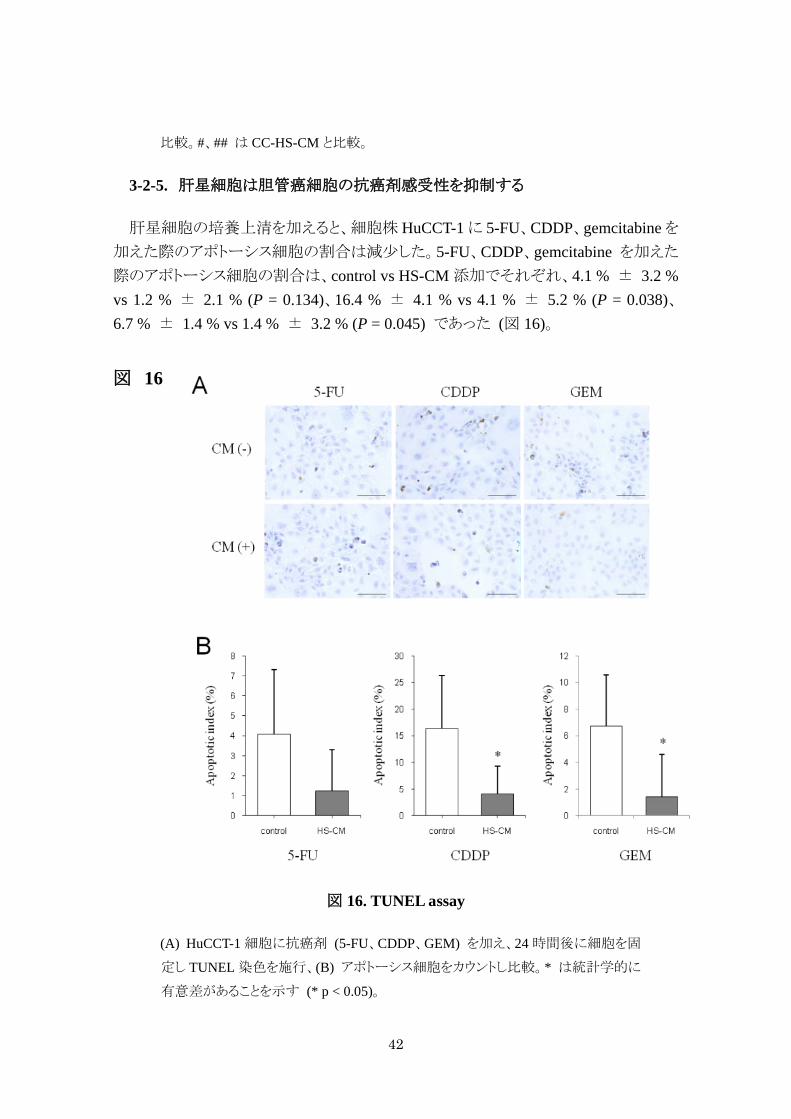
3-2-5. 肝星細胞は胆管癌細胞の抗癌剤感受性を抑制する
肝星細胞の培養上清を加えると、細胞株 HuCCT-1 に 5-FU、CDDP、gemcitabine を加えた際のアポトーシス細胞の割合は減少した。5-FU、CDDP、gemcitabine を加えた
際のアポトーシス細胞の割合は、control vs HS-CM 添加でそれぞれ、4.1 % ± 3.2 % vs 1.2 % ± 2.1 % (P = 0.134)、16.4 % ± 4.1 % vs 4.1 % ± 5.2 % (P = 0.038)、6.7 % ± 1.4 % vs 1.4 % ± 3.2 % (P = 0.045) であった (図 16)。
図 16
図 16. TUNEL assay
(A) HuCCT-1 細胞に抗癌剤 (5-FU、CDDP、GEM) を加え、24 時間後に細胞を固
定し TUNEL 染色を施行、(B) アポトーシス細胞をカウントし比較。* は統計学的に
有意差があることを示す (* p < 0.05)。

43
44
3-3. In vivo における肝星細胞と胆管癌細胞の相互作用
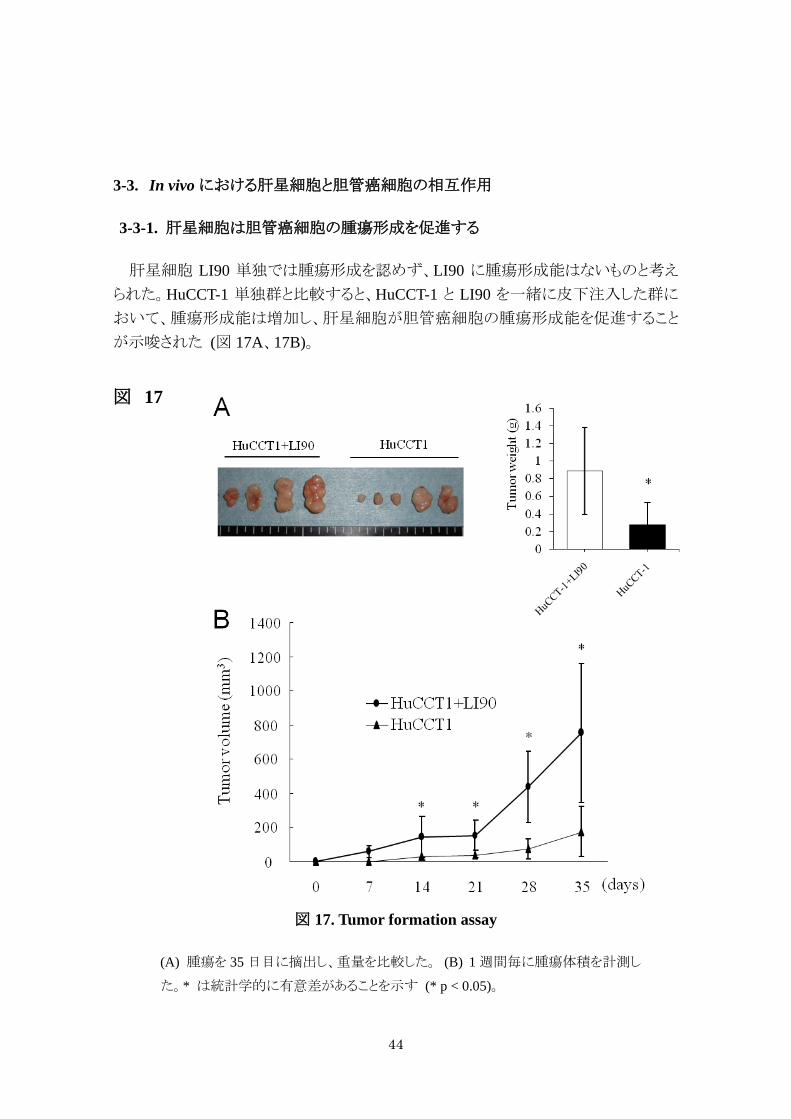
3-3-1. 肝星細胞は胆管癌細胞の腫瘍形成を促進する
肝星細胞 LI90 単独では腫瘍形成を認めず、LI90 に腫瘍形成能はないものと考え
られた。HuCCT-1 単独群と比較すると、HuCCT-1 と LI90 を一緒に皮下注入した群に
おいて、腫瘍形成能は増加し、肝星細胞が胆管癌細胞の腫瘍形成能を促進すること
が示唆された (図 17A、17B)。
図 17
図 17. Tumor formation assay
(A) 腫瘍を 35 日目に摘出し、重量を比較した。 (B) 1 週間毎に腫瘍体積を計測し
た。* は統計学的に有意差があることを示す (* p < 0.05)。

45
46
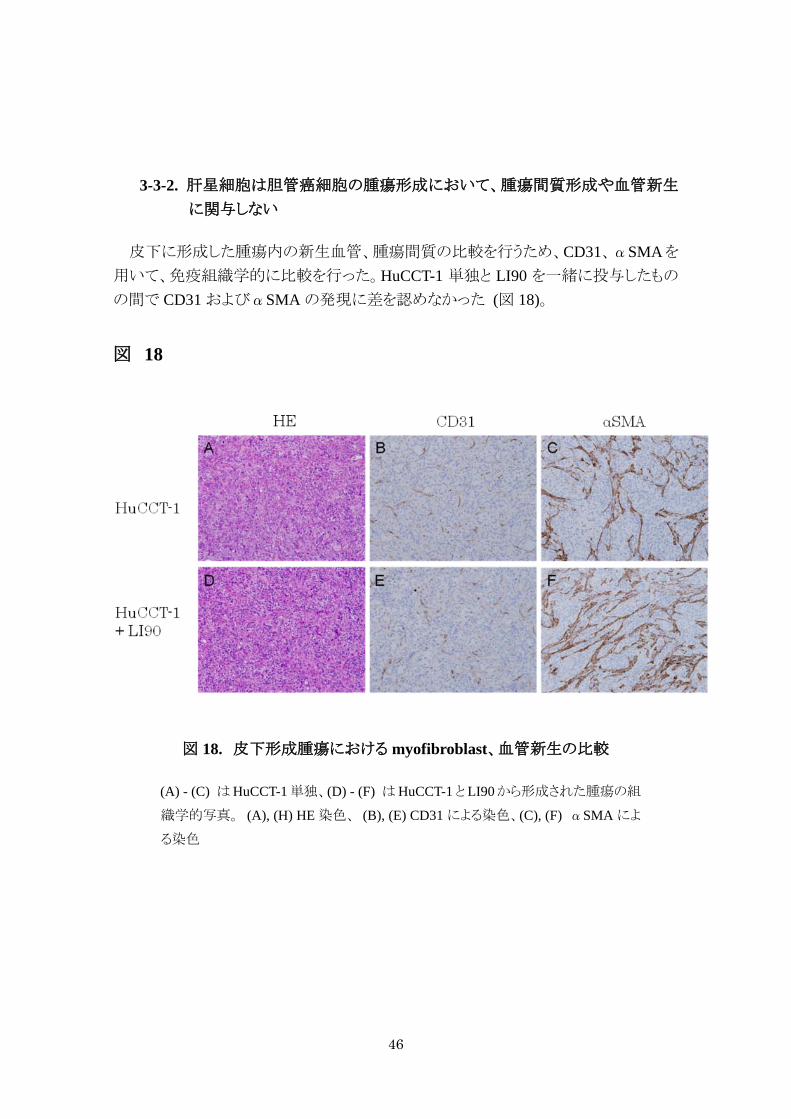
3-3-2. 肝星細胞は胆管癌細胞の腫瘍形成において、腫瘍間質形成や血管新生
に関与しない
皮下に形成した腫瘍内の新生血管、腫瘍間質の比較を行うため、CD31、αSMAを
用いて、免疫組織学的に比較を行った。HuCCT-1 単独と LI90 を一緒に投与したもの
の間で CD31 およびαSMA の発現に差を認めなかった (図 18)。
図 18
図 18. 皮下形成腫瘍における myofibroblast、血管新生の比較
(A) - (C) はHuCCT-1単独、(D) - (F) はHuCCT-1とLI90から形成された腫瘍の組
織学的写真。 (A), (H) HE 染色、 (B), (E) CD31 による染色、(C), (F) αSMA によ
る染色
47
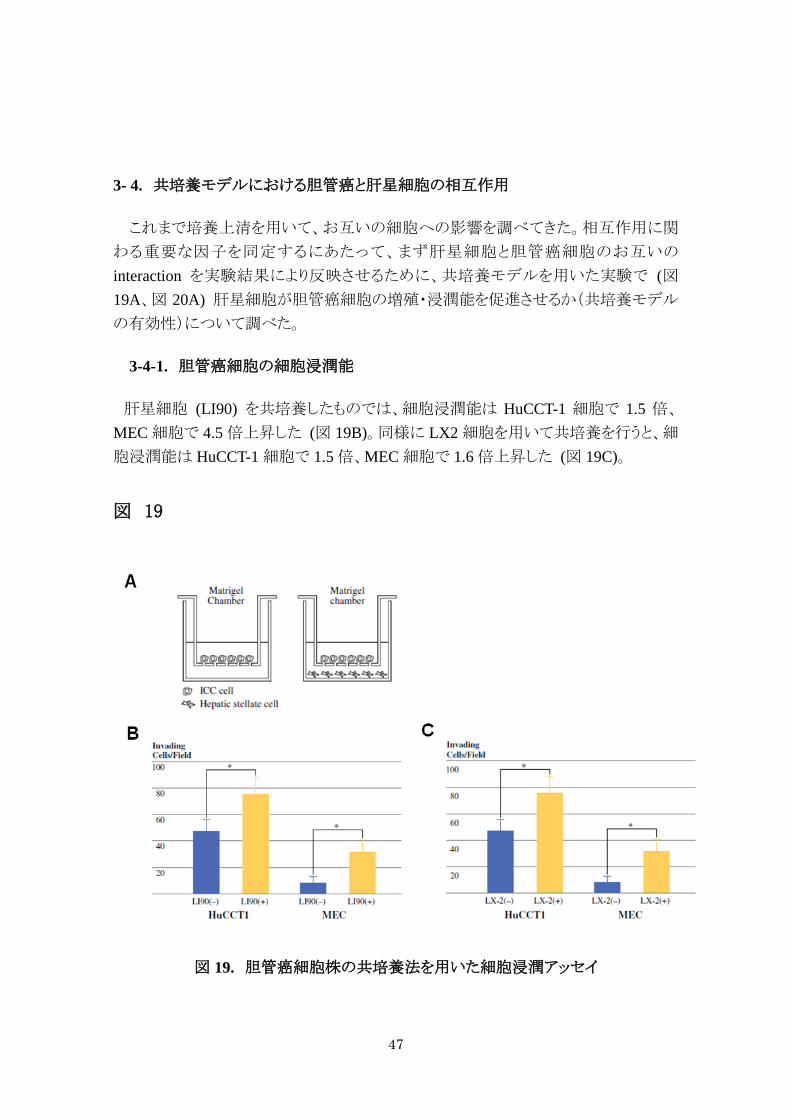
3- 4. 共培養モデルにおける胆管癌と肝星細胞の相互作用
これまで培養上清を用いて、お互いの細胞への影響を調べてきた。相互作用に関
わる重要な因子を同定するにあたって、まず肝星細胞と胆管癌細胞のお互いの
interaction を実験結果により反映させるために、共培養モデルを用いた実験で (図19A、図 20A) 肝星細胞が胆管癌細胞の増殖・浸潤能を促進させるか(共培養モデル
の有効性)について調べた。
3-4-1. 胆管癌細胞の細胞浸潤能
肝星細胞 (LI90) を共培養したものでは、細胞浸潤能は HuCCT-1 細胞で 1.5 倍、
MEC 細胞で 4.5 倍上昇した (図 19B)。同様に LX2 細胞を用いて共培養を行うと、細
胞浸潤能は HuCCT-1 細胞で 1.5 倍、MEC 細胞で 1.6 倍上昇した (図 19C)。
図 19
図 19. 胆管癌細胞株の共培養法を用いた細胞浸潤アッセイ
48
A. 共培養細胞浸潤アッセイ。B. 肝星細胞に LI90 を用いて、C では LX2 を
用いている。* は統計学的に有意差があることを示す (* p < 0.05)。
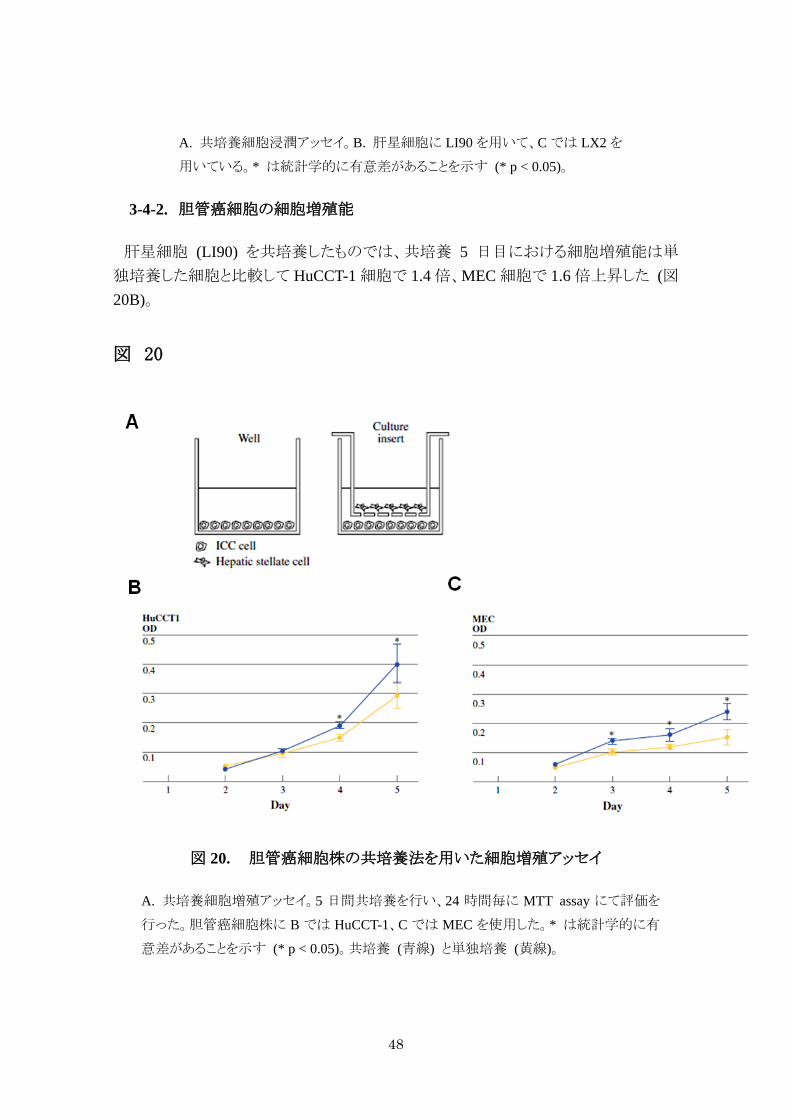
3-4-2. 胆管癌細胞の細胞増殖能
肝星細胞 (LI90) を共培養したものでは、共培養 5 日目における細胞増殖能は単
独培養した細胞と比較して HuCCT-1 細胞で 1.4 倍、MEC 細胞で 1.6 倍上昇した (図20B)。
図 20
図 20. 胆管癌細胞株の共培養法を用いた細胞増殖アッセイ
A. 共培養細胞増殖アッセイ。5 日間共培養を行い、24 時間毎に MTT assay にて評価を
行った。胆管癌細胞株に B では HuCCT-1、C では MEC を使用した。* は統計学的に有
意差があることを示す (* p < 0.05)。共培養 (青線) と単独培養 (黄線)。

49
50
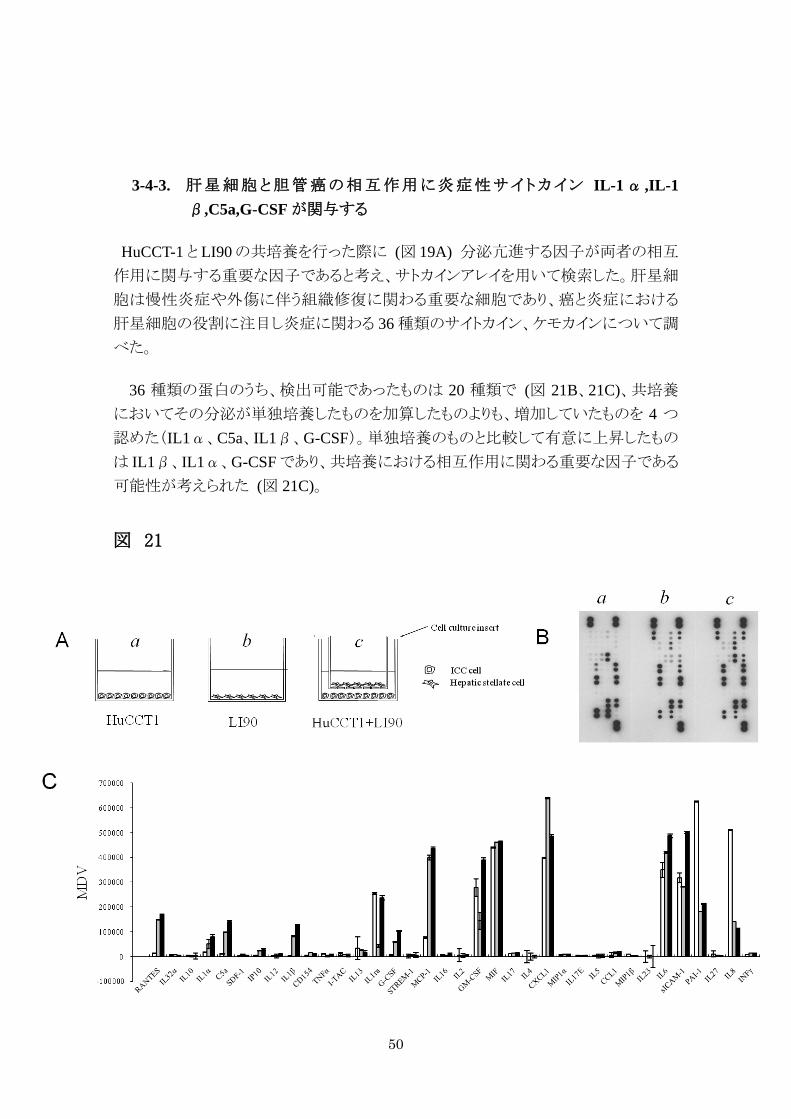
3-4-3. 肝星細胞と胆管癌の相互作用に炎症性サイトカイン IL-1α ,IL-1β,C5a,G-CSF が関与する
HuCCT-1 と LI90 の共培養を行った際に (図 19A) 分泌亢進する因子が両者の相互
作用に関与する重要な因子であると考え、サトカインアレイを用いて検索した。肝星細
胞は慢性炎症や外傷に伴う組織修復に関わる重要な細胞であり、癌と炎症における
肝星細胞の役割に注目し炎症に関わる 36 種類のサイトカイン、ケモカインについて調
べた。
36 種類の蛋白のうち、検出可能であったものは 20 種類で (図 21B、21C)、共培養
においてその分泌が単独培養したものを加算したものよりも、増加していたものを 4 つ
認めた(IL1α、C5a、IL1β、G-CSF)。単独培養のものと比較して有意に上昇したもの
は IL1β、IL1α、G-CSF であり、共培養における相互作用に関わる重要な因子である
可能性が考えられた (図 21C)。
図 21

51
図 21. サイトカインアレイ
A. 培養上清サンプルは HuCCT-1 (a)、LI90 (b)、共培養したもの (c) から回収
した。B. 3 つの培養上清を反応させたアレイチップの写真。C. 36 種類の分子
の集積値をソフトで定量化し示した。D. 集積値 (MDV)が c > ( a + b )を満たす
ものは 4 種類存在し、IL1α、C5a、IL1β、G-CSF であった。

52
3-5. 質量分析を用いた、肝星細胞と胆管癌の相互作用に関与する因子 CXCL5 の
同定
胆管癌細胞と肝星細胞の間接的な相互作用に関与する因子を網羅的に解析する
ために、質量分析を用いて検索を行った。結果 3-3-3 と同様に共培養によって著明に
分泌が上昇する因子が相互作用に関わる重要な因子であると考え、これを定量的に
まず SELDI-TOF MS で検索した。さらに、共培養の組み合わせを変えることにより抽
出された候補の絞り込みを行い、絞り込まれた分子量の蛋白の同定を行った。同定さ
れた CXCL5 の機能解析を細胞、ヒトサンプルを用いて検証した。
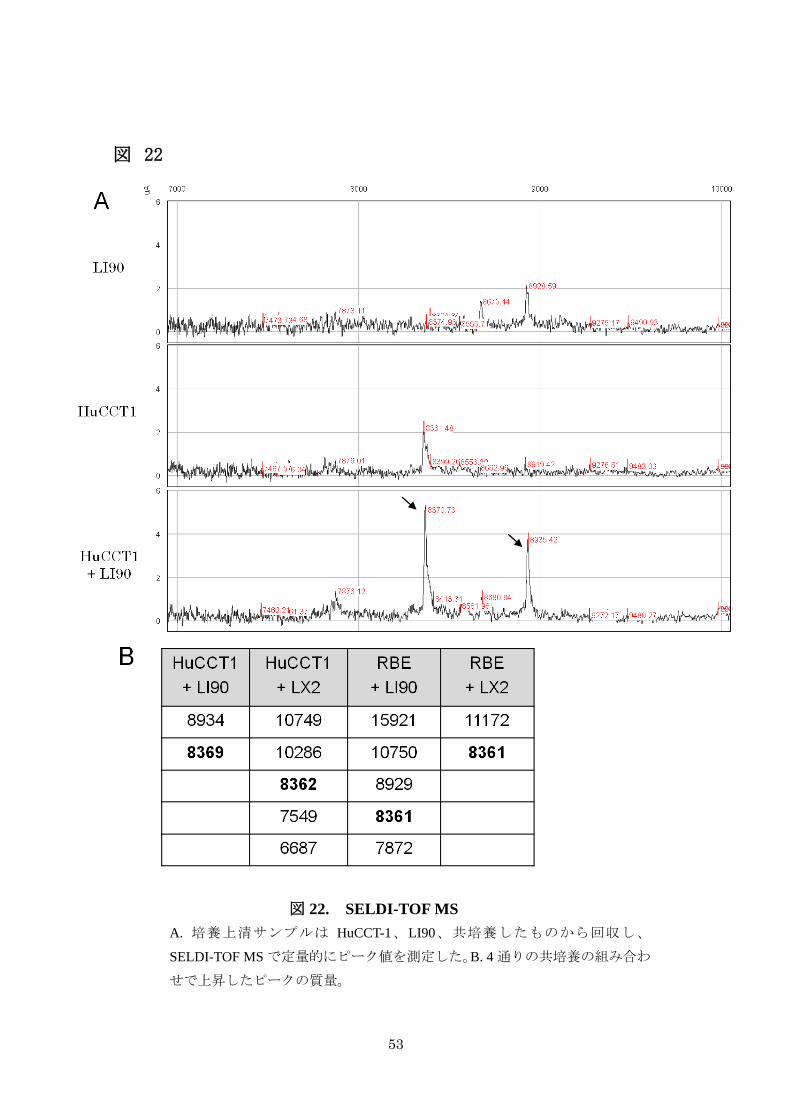
3-5-1. SELDI-TOF MS 解析で得られた、胆管癌細胞と肝星細胞の相互作用に
関わる蛋白の質量は 8360Da である
胆管癌細胞 2 株 (HuCCT1, RBE)と肝星細胞株 2 株 (LI90, LX2)を用いて、計 4通りの組み合わせで共培養を行い (HuCCT1×LI90、HuCCT1×LX2、RBE×LI90、RBE×LX2)、それぞれにつき、単独培養からの培養上清と共培養からの培養上清を
回収し、濃縮保存した。サンプル数は 3(単独培養の胆管癌細胞、肝星細胞と共培
養)×4(組み合わせの数)=12 個で、これを用いて SELDI で定量的にピークを比較し
た(図 22A)。4 通りの共培養の組み合わせで
共培養した培養上清のピーク値 > 2×( 胆管癌細胞を単独培養した培養上清
のピーク値+肝星細胞を単独培養した培養上清のピーク値 )
を満たすピークを検索すると CM10pH7 のチップが最も検出される蛋白が多く、計 14個のピークが検出された(図 22B)。いずれの共培養においても上昇する共通したピー
クを検索すると 8360Da の質量が得られた。
53
図 22
図 22. SELDI-TOF MS
A. 培養上清サンプルは HuCCT-1、LI90、共培養したものから回収し、
SELDI-TOF MS で定量的にピーク値を測定した。B. 4 通りの共培養の組み合わ
せで上昇したピークの質量。
54
3-5-2. Q-TOF/MS/MS 解析で得られた相互作用に関わる蛋白は CXCL5 である
SELDI 解析により得られた分子量 8360 の情報を基に、培養上清を SDS-PAGE 電
気泳動を行い、CBB 染色を行った後、分子量に相当するバンドを切り出して(図 23 左
黄色枠)Q-TOF/MS/MS 解析を行った。同定された蛋白は GI: 4506849 CXCL5
[Homo sapiens] のみであった。CXCL5 の抗体を用いて Western 法で、確認すると、
切り出した band の位置に一致して CXCL5 の band が得られ、ゲルが CXCL5 であるこ
とが確認された。
図 23
図 23. CBB 染色と Westen 法による検出
図左. 培養上清サンプルは HuCCT-1、LI90、共培養したものから回収したもの
を電気泳動後 CBB 染色した。図右. CXCL5 抗体を用いて、図左と同じサンプ
ルを用いて Western 法で確認したもの。

55
3-5-3. SELDI-TOF MS で得られた 8360 Da のピークは CXCL5 である
SELDI で得られたピークが CXCL5 であることを確認するために、Conntrol IgG と
CXCL5抗体を用いて、免疫沈降を行い、沈降後のサンプルを再度SELDIで検出した。
CXCL5 抗体で免疫沈降したサンプルは 8360 のピークの消失を認め、質量 8360 Daのピークが CXCL5 であることが確認された(図 24)。
図 24
図 24. 免疫沈降後のサンプルを SELDI-TOF MS で検出 Control IgG と CXCL5 抗体を用いて、蛋白ビーズを用いて培養上清と免疫沈降
を行った。培養上清に用いたサンプルは HuCCT1 と LI90 を共培養したもの。
56
3-5-4. CXCL5 は胆管癌の遊走能、浸潤能を促進する
CXCL5 が癌細胞へおよぼす影響を調べるため、胆管癌細胞株 HuCCT1 へ
rhCXCL5 を加え遊走能を Wound healing assay で調べると、遊走能は 1.4 倍へ上昇し
た (図 25A)。また、浸潤能を調べると、rhCXCL5 を加えることにより 2.3 倍上昇した
(図 25B)。
図 25
図 25. Wound healing assay と Invasion assay
* *
57
Control IgG と CXCL5 抗体を用いて、蛋白ビーズを用いて培養上清と免疫沈降
を行った。培養上清に用いたサンプルは HuCCT1 と LI90 を共培養したもの。
* は統計学的に有意差があることを示す (* p < 0.05)。
3-5-5. CXCL5 の発現は myofibroblast の存在と相関し、CXCL5 髙発現群は生
存予後不良である
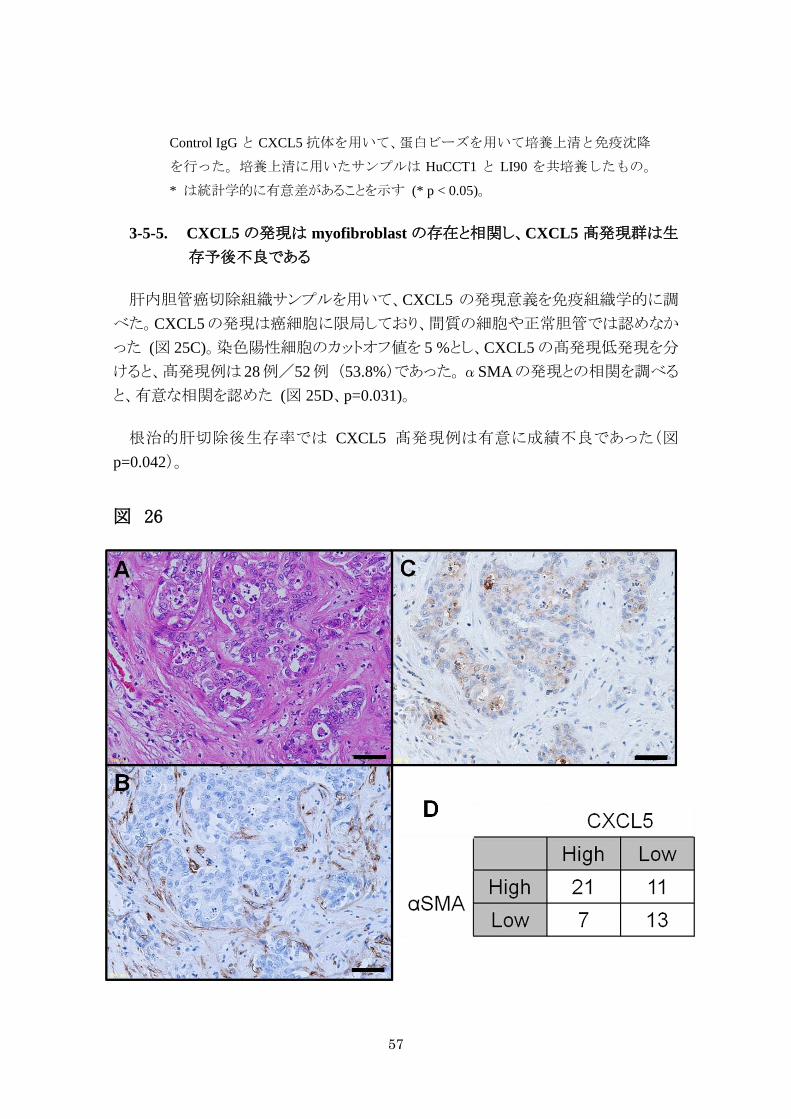
肝内胆管癌切除組織サンプルを用いて、CXCL5 の発現意義を免疫組織学的に調
べた。CXCL5の発現は癌細胞に限局しており、間質の細胞や正常胆管では認めなか
った (図 25C)。染色陽性細胞のカットオフ値を 5 %とし、CXCL5 の髙発現低発現を分
けると、髙発現例は 28例/52例 (53.8%)であった。αSMAの発現との相関を調べる
と、有意な相関を認めた (図 25D、p=0.031)。
根治的肝切除後生存率では CXCL5 髙発現例は有意に成績不良であった(図
p=0.042)。
図 26
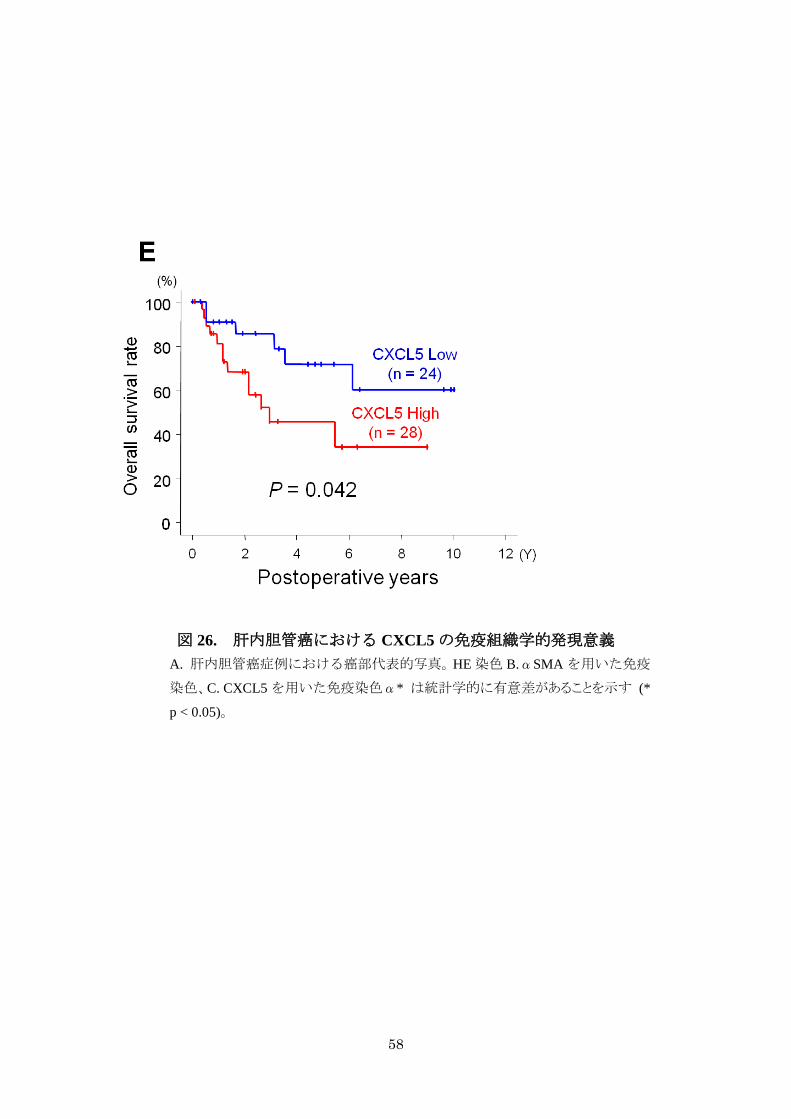
58
図 26. 肝内胆管癌における CXCL5 の免疫組織学的発現意義
A. 肝内胆管癌症例における癌部代表的写真。HE 染色 B.αSMA を用いた免疫
染色、C. CXCL5 を用いた免疫染色α* は統計学的に有意差があることを示す (*
p < 0.05)。

59
第四章 考察
これまで肝癌における肝星細胞に関する研究は極めて少なく、本研究の目的は肝
内胆管癌における肝星細胞の存在と、癌の進展における役割および肝星細胞と胆管
癌細胞の相互作用に関与する因子を明らかにすることにあった。
まずヒト切除サンプルを用いた免疫組織学的手法による myofibroblast の定量化によ
り、αSMA の髙発現群は切除後無再発・生存率が不良で、多変量解析では生存予
後に寄与する独立因子であり、癌の進展に myofibroblast または CAF が関与している
可能性が示唆された。Myofibroblast の定量化の有効性については、myofibroblast が関与することが知られる慢性肝炎・肝硬変のサンプルを用いて、検証を行い有効性が
確認された。αSMA の髙発現群と低発現群の間には臨床病理学的な差を認めず、
癌の悪性度に myofibroblast がどのように関与しているのかについては臨床的側面か
らは推測困難であった。発癌モデルマウスを用いた実験において、CAF は発癌の初
期より様々なサイトカインを通して癌の進展に関与し、血管新生やマクロファージの遊
走に関与していることが報告されている 59。また、大腸癌においてαSMAの発現は癌
の予後予測因子であるだけでなく、原発巣と転移巣において症例毎にその発現の程
度は非常に近似しており、癌の特性の一面を反映したものであることが推測される 37。
肝内胆管癌の腫瘍間質に存在する星細胞を検出するため、肝星細胞のマーカーで
ある GFAP、desmin を用いて、αSMA との共発現細胞を検索すると、少数ながら星細
胞と思われる細胞を認めた。ヒトにおいて肝星細胞を鋭敏に検出するマーカーは現在
コンセンサスが得られておらず、困難な理由として、門脈域の portal fibroblast 60 や骨
髄由来細胞 61 などの細胞集団を区別することが困難なことにある。ラットにおいて肝
星細胞を検出するのに有効な GFAP はヒトにおいて検出困難であることが知られてお
り、さらにGFAPの特性として、肝星細胞の活性化に伴いその発現は減弱することが報
告されており 62、癌間質における肝星細胞の検出率の低さはそういった影響が予想さ
れる。
これまで細胞株を用いた肝星細胞と胆管癌細胞の相互作用を検証した報告はな
く、我々は初めてそれらの in vitro、in vivo における相互作用を確認した。癌もしくは肝
星細胞の培養上清を用いた実験または共培養モデルを用いた実験では、肝星細胞
が胆管癌細胞の増殖・浸潤・抗癌剤に対するアポトーシスに関与する事が確認された。
これらの現象に関与する細胞内シグナルとしては MAPK や Akt の関与が示された。こ
れらのモデルはすべて分泌された因子の間接的な作用を確認したに過ぎず、直接接

60
触した状況下での癌細胞の phenotype の変化を調べることが今後必要であると考えら
れる。
膵癌において、中和抗体を用いた実験で膵癌細胞株と膵星細胞の相互作用に関
わる重要な因子としてPDGFが報告されており 39, 47、これは肝臓の線維化において肝
星細胞を活性化させる因子として最も重要な因子として知られることは興味深い。
CAF の腫瘍の血管新生に及ぼす影響に関して、胃癌または食道癌の細胞株を用い
た実験の報告がみられ、いずれの報告においても癌細胞からの分泌因子を受けて、
CAF から VEGF 等の血管新生因子が分泌されることの重要性が示されている 63, 64。
我々の in vitro における tube formation の実験でも、癌細胞からの培養上清を肝星細
胞に加え、それを回収した培養上清 (CC-HS-CM) が最も tube formation を促進させ
る結果であった。逆に肝星細胞からの培養上清を癌細胞に加え、これを回収したもの (HS-CC-CM) では同様の結果は得られず、これまでの報告を支持する結果である。
In vitro における腫瘍-間質細胞の相互作用による tube formation の促進効果につい
て、in vivo においても胆管癌細胞と肝星細胞を一緒に注入することで血管新生促進
作用が見られると期待したが、皮下腫瘍の免疫組織学的検討では胆管癌細胞単独と
の差はみられなかった。その理由として、1つは手技的な問題が考えられた。今回用
いた LI90 は JCRB より購入した細胞株であるが、細胞培養を開始してからその viableな状況は約 2 ヶ月程度しか維持できず、老化してしまう。凍結保存すると増殖活性が
極端に落ちてしまうため、実験の機会は非常に限られている。また、細胞が非常に大
きいため、必要な細胞を培養するための面積は膨大で、注入するための回収作業に
かなりの時間を要し、さらに皮下に注入する volume が大きくならざるを得ず、形成され
る腫瘍の形態にばらつきが生じやすい。つまり、迅速な均一な手技で実験を完遂する
には熟練を要する。肝星細胞として世界的なコンセンサスが得られ入手できる細胞株
は現在 3 株程度しかなく、今回分譲して頂いた LX2 も実験に用いているが、不死化さ
れていないのは LI90 のみであり、不死化されていない細胞株を用いて in vivo におけ
る腫瘍形成の促進効果を確認することを目的として LI90 を用いた。LX2 であれば、細
胞は小さく不死化されており、増殖速度も早いため、実験の手技的な安定性は得られ
やすいと考えられ、組織学的な比較検討も期待される。また、もう一つは皮下という異
種移植モデルであることも影響しているかもしれない。同所移植モデルが望ましい。
相互作用の実験において最も重要な関心事項は相互作用を左右する因子であり、
我々はサイトカインアレイを用いて、相互作用に関わる分泌因子の検索作業を行った。
その結果絞り込まれたのは 4 因子 (IL1α、C5a、IL-1β、G-CSF) であった。IL-1βに
関しては他の癌腫において腫瘍-間質相互作用に関する報告もこれまでみられ 41, 59,
65、線維化・炎症に関わる重要な因子である。

61
我々は相互作用を制御する分泌蛋白について網羅的に調べるために、
SELDI-TOF MS を用いた培養上清の解析を行った。共培養は間接的な相互作用で
あるため、分泌される蛋白で共培養によって上昇するものが重要な因子であると考え、
検索を行った。同定されたCXCL5については上皮細胞から分泌される好中球遊走因
子であることが知られており、肺癌、胃癌、食道癌、前立腺癌において癌の進展に寄
与するとの報告が見られる 66-72。肝内胆管癌組織では myofibroblast の存在と癌部
CXCL5 の発現は相関し、CXCL5 髙発現群は生存予後不良であることからも、CXCL5が CAF と癌細胞との相互作用に関与し、肝内胆管癌の進展に寄与する事が考えられ
る。CXCL5 の receptor は CXCR2 であり 73、CXCL8 の receptor としても知られる。上皮
細胞や血管内皮細胞、好中球における発現が報告されているものの、fibroblast や
myofibroblast における発現の報告は見られず 74、CAF が癌細胞の CXCL5 の分泌を
促進させ、本研究結果から肝内胆管癌の遊走や浸潤能を促進することからも、癌細胞
の進展を促進していることが考えられる。肺癌においては、IL1β刺激を行うと、癌細
胞の CXCL5 や IL8 の分泌が促進することが報告されており 71、今研究においても IL1βは共培養によって分泌が促進され、かつ肝星細胞が主要な産生細胞であり、肝星
細胞が分泌する IL1βにより胆管癌細胞の CXCL5 の産生が促進されることが予想さ
れる。
CAF あるいは myofibroblast は癌の進展において非常に重要な細胞集団であること
が報告され、治療標的としての研究が進められている。これらの多くの研究は、癌と炎
症の関連に注目した研究であり、これらの研究をいかに予防・診断・治療へと還元する
かが最も重要な点である 30, 75, 76。肝の線維化においては肝星細胞を治療標的とした
治療法も期待され 77、肝内胆管癌においても肝星細胞または肝星細胞と胆管癌細胞
の相互作用を治療標的とした治療戦略を構築していきたい。

62
第五章 結語
本研究において以下のことを明らかにした。
① 肝内胆管癌において肝星細胞由来の myofibroblast (CAF) は癌の発育進展に関
与する。
② 肝星細胞は胆管癌細胞によって活性化される。また一方、肝星細胞細胞によって、
胆管癌細胞の増殖能、浸潤能は促進され、抗癌剤に対するアポトーシスは抑制され
た。
③ 胆管癌細胞株の in vivo における腫瘍形成能は肝星細胞によって促進された。
④ 胆管癌細胞株と肝星細胞株の相互作用に関わるサイトカインとして IL1α、C5a、
IL-1β、G-CSF が抽出された。
⑤ 網羅的解析から、胆管癌細胞株と肝星細胞株の相互作用に関与する因子として
CXCL5 を同定した。
⑥ CXCL5 は胆管癌細胞の遊走・浸潤能を促進し、myoofibroblast との相互作用によ
り癌の進展に関与する可能性がある。
今後肝内胆管癌における CXCL5 の詳細な機能解析を行ってゆく。

63
第六章 参考文献
1. Ben-Menachem T. Risk factors for cholangiocarcinoma. Eur J Gastroenterol Hepatol 2007;19: 615-7. 2. Khan SA, Thomas HC, Davidson BR, Taylor-Robinson SD. Cholangiocarcinoma. Lancet 2005;366: 1303-14. 3. Shaib YH, El-Serag HB, Davila JA, Morgan R, McGlynn KA. Risk factors of intrahepatic cholangiocarcinoma in the United States: a case-control study. Gastroenterology 2005;128: 620-6. 4. Cho SY, Park SJ, Kim SH, et al. Survival analysis of intrahepatic cholangiocarcinoma after resection. Ann Surg Oncol;17: 1823-30. 5. Choi SB, Kim KS, Choi JY, et al. The prognosis and survival outcome of intrahepatic cholangiocarcinoma following surgical resection: association of lymph node metastasis and lymph node dissection with survival. Ann Surg Oncol 2009;16: 3048-56. 6. Inoue K, Makuuchi M, Takayama T, et al. Long-term survival and prognostic factors in the surgical treatment of mass-forming type cholangiocarcinoma. Surgery 2000;127: 498-505. 7. Jonas S, Thelen A, Benckert C, et al. Extended liver resection for intrahepatic cholangiocarcinoma: A comparison of the prognostic accuracy of the fifth and sixth editions of the TNM classification. Ann Surg 2009;249: 303-9. 8. Konstadoulakis MM, Roayaie S, Gomatos IP, et al. Fifteen-year, single-center experience with the surgical management of intrahepatic cholangiocarcinoma: operative results and long-term outcome. Surgery 2008;143: 366-74. 9. Nakagawa T, Kamiyama T, Kurauchi N, et al. Number of lymph node metastases is a significant prognostic factor in intrahepatic cholangiocarcinoma. World J Surg 2005;29: 728-33. 10. Tan JC, Coburn NG, Baxter NN, Kiss A, Law CH. Surgical management of intrahepatic cholangiocarcinoma--a population-based study. Ann Surg Oncol 2008;15: 600-8. 11. Weimann A, Varnholt H, Schlitt HJ, et al. Retrospective analysis of prognostic factors after liver resection and transplantation for cholangiocellular carcinoma. Br J Surg 2000;87: 1182-7. 12. Yonemoto N, Furuse J, Okusaka T, et al. A multi-center retrospective analysis of survival benefits of chemotherapy for unresectable biliary tract cancer.

64
Jpn J Clin Oncol 2007;37: 843-51. 13. Terada T, Makimoto K, Terayama N, Suzuki Y, Nakanuma Y. Alpha-smooth muscle actin-positive stromal cells in cholangiocarcinomas, hepatocellular carcinomas and metastatic liver carcinomas. J Hepatol 1996;24: 706-12. 14. Bataller R, Brenner DA. Liver fibrosis. J Clin Invest 2005;115: 209-18. 15. Mikula M, Proell V, Fischer AN, Mikulits W. Activated hepatic stellate cells induce tumor progression of neoplastic hepatocytes in a TGF-beta dependent fashion. J Cell Physiol 2006;209: 560-7. 16. Henderson NC, Iredale JP. Liver fibrosis: cellular mechanisms of progression and resolution. Clin Sci (Lond) 2007;112: 265-80. 17. Friedman SL. Hepatic stellate cells: protean, multifunctional, and enigmatic cells of the liver. Physiol Rev 2008;88: 125-72. 18. Cassiman D, van Pelt J, De Vos R, et al. Synaptophysin: A novel marker for human and rat hepatic stellate cells. Am J Pathol 1999;155: 1831-9. 19. Neubauer K, Knittel T, Aurisch S, Fellmer P, Ramadori G. Glial fibrillary acidic protein--a cell type specific marker for Ito cells in vivo and in vitro. J Hepatol 1996;24: 719-30. 20. Niki T, De Bleser PJ, Xu G, Van Den Berg K, Wisse E, Geerts A. Comparison of glial fibrillary acidic protein and desmin staining in normal and CCl4-induced fibrotic rat livers. Hepatology 1996;23: 1538-45. 21. Uyama N, Zhao L, Van Rossen E, et al. Hepatic stellate cells express synemin, a protein bridging intermediate filaments to focal adhesions. Gut 2006;55: 1276-89. 22. Dvorak HF. Tumors: wounds that do not heal. Similarities between tumor stroma generation and wound healing. N Engl J Med 1986;315: 1650-9. 23. Bhowmick NA, Neilson EG, Moses HL. Stromal fibroblasts in cancer initiation and progression. Nature 2004;432: 332-7. 24. Gregoire M, Lieubeau B. The role of fibroblasts in tumor behavior. Cancer Metastasis Rev 1995;14: 339-50. 25. Hanahan D, Weinberg RA. The hallmarks of cancer. Cell 2000;100: 57-70. 26. Hinz B, Phan SH, Thannickal VJ, Galli A, Bochaton-Piallat ML, Gabbiani G. The myofibroblast: one function, multiple origins. Am J Pathol 2007;170: 1807-16. 27. Joyce JA, Pollard JW. Microenvironmental regulation of metastasis. Nat Rev Cancer 2009;9: 239-52. 28. Liotta LA, Kohn EC. The microenvironment of the tumour-host interface.

65
Nature 2001;411: 375-9. 29. Littlepage LE, Egeblad M, Werb Z. Coevolution of cancer and stromal cellular responses. Cancer Cell 2005;7: 499-500. 30. Mantovani A, Allavena P, Sica A, Balkwill F. Cancer-related inflammation. Nature 2008;454: 436-44. 31. Mueller MM, Fusenig NE. Friends or foes - bipolar effects of the tumour stroma in cancer. Nat Rev Cancer 2004;4: 839-49. 32. Kalluri R, Zeisberg M. Fibroblasts in cancer. Nat Rev Cancer 2006;6: 392-401. 33. Barsky SH, Green WR, Grotendorst GR, Liotta LA. Desmoplastic breast carcinoma as a source of human myofibroblasts. Am J Pathol 1984;115: 329-33. 34. Durning P, Schor SL, Sellwood RA. Fibroblasts from patients with breast cancer show abnormal migratory behaviour in vitro. Lancet 1984;2: 890-2. 35. Sappino AP, Skalli O, Jackson B, Schurch W, Gabbiani G. Smooth-muscle differentiation in stromal cells of malignant and non-malignant breast tissues. Int J Cancer 1988;41: 707-12. 36. Surowiak P, Murawa D, Materna V, et al. Occurence of stromal myofibroblasts in the invasive ductal breast cancer tissue is an unfavourable prognostic factor. Anticancer Res 2007;27: 2917-24. 37. Tsujino T, Seshimo I, Yamamoto H, et al. Stromal myofibroblasts predict disease recurrence for colorectal cancer. Clin Cancer Res 2007;13: 2082-90. 38. Apte MV, Park S, Phillips PA, et al. Desmoplastic reaction in pancreatic cancer: role of pancreatic stellate cells. Pancreas 2004;29: 179-87. 39. Bachem MG, Schunemann M, Ramadani M, et al. Pancreatic carcinoma cells induce fibrosis by stimulating proliferation and matrix synthesis of stellate cells. Gastroenterology 2005;128: 907-21. 40. Algul H, Treiber M, Lesina M, Schmid RM. Mechanisms of disease: chronic inflammation and cancer in the pancreas--a potential role for pancreatic stellate cells? Nat Clin Pract Gastroenterol Hepatol 2007;4: 454-62. 41. Aoki H, Ohnishi H, Hama K, et al. Autocrine loop between TGF-beta1 and IL-1beta through Smad3- and ERK-dependent pathways in rat pancreatic stellate cells. Am J Physiol Cell Physiol 2006;290: C1100-8. 42. Bachem MG, Zhou S, Buck K, Schneiderhan W, Siech M. Pancreatic stellate cells--role in pancreas cancer. Langenbecks Arch Surg 2008;393: 891-900. 43. Hwang RF, Moore T, Arumugam T, et al. Cancer-associated stromal fibroblasts promote pancreatic tumor progression. Cancer Res 2008;68: 918-26.

66
44. Miyamoto H, Murakami T, Tsuchida K, Sugino H, Miyake H, Tashiro S. Tumor-stroma interaction of human pancreatic cancer: acquired resistance to anticancer drugs and proliferation regulation is dependent on extracellular matrix proteins. Pancreas 2004;28: 38-44. 45. Omary MB, Lugea A, Lowe AW, Pandol SJ. The pancreatic stellate cell: a star on the rise in pancreatic diseases. J Clin Invest 2007;117: 50-9. 46. Sato N, Maehara N, Goggins M. Gene expression profiling of tumor-stromal interactions between pancreatic cancer cells and stromal fibroblasts. Cancer Res 2004;64: 6950-6. 47. Vonlaufen A, Joshi S, Qu C, et al. Pancreatic stellate cells: partners in crime with pancreatic cancer cells. Cancer Res 2008;68: 2085-93. 48. Vonlaufen A, Phillips PA, Xu Z, et al. Pancreatic stellate cells and pancreatic cancer cells: an unholy alliance. Cancer Res 2008;68: 7707-10. 49. Schmitt-Graff A, Ertelt V, Allgaier HP, et al. Cellular retinol-binding protein-1 in hepatocellular carcinoma correlates with beta-catenin, Ki-67 index, and patient survival. Hepatology 2003;38: 470-80. 50. Faouzi S, Le Bail B, Neaud V, et al. Myofibroblasts are responsible for collagen synthesis in the stroma of human hepatocellular carcinoma: an in vivo and in vitro study. J Hepatol 1999;30: 275-84. 51. Olaso E, Salado C, Egilegor E, et al. Proangiogenic role of tumor-activated hepatic stellate cells in experimental melanoma metastasis. Hepatology 2003;37: 674-85. 52. Olaso E, Santisteban A, Bidaurrazaga J, Gressner AM, Rosenbaum J, Vidal-Vanaclocha F. Tumor-dependent activation of rodent hepatic stellate cells during experimental melanoma metastasis. Hepatology 1997;26: 634-42. 53. Olaso E, Vidal-Vanaclocha F. Use of tumor-activated hepatic stellate cell as a target for the preclinical testing of anti-angiogenic drugs against hepatic tumor development. Methods Mol Med 2003;85: 79-86. 54. Amann T, Bataille F, Spruss T, et al. Activated hepatic stellate cells promote tumorigenicity of hepatocellular carcinoma. Cancer Sci 2009;100: 646-53. 55. Matsusue R, Kubo H, Hisamori S, et al. Hepatic stellate cells promote liver metastasis of colon cancer cells by the action of SDF-1/CXCR4 axis. Ann Surg Oncol 2009;16: 2645-53. 56. Ichida F TT, Omata M. New Inuyama Classification; new criteria for histological assessment of chronic hepatitis. International Hepatology Communication 1996;6: 112-19.

67
57. Murakami K, Abe T, Miyazawa M, et al. Establishment of a new human cell line, LI90, exhibiting characteristics of hepatic Ito (fat-storing) cells. Lab Invest 1995;72: 731-9. 58. Xu L, Hui AY, Albanis E, et al. Human hepatic stellate cell lines, LX-1 and LX-2: new tools for analysis of hepatic fibrosis. Gut 2005;54: 142-51. 59. Erez N, Truitt M, Olson P, Hanahan D. Cancer-Associated Fibroblasts Are Activated in Incipient Neoplasia to Orchestrate Tumor-Promoting Inflammation in an NF-kappaB-Dependent Manner. Cancer Cell;17: 135-47. 60. Dranoff JA, Wells RG. Portal fibroblasts: Underappreciated mediators of biliary fibrosis. Hepatology;51: 1438-44. 61. Russo FP, Alison MR, Bigger BW, et al. The bone marrow functionally contributes to liver fibrosis. Gastroenterology 2006;130: 1807-21. 62. Carotti S, Morini S, Corradini SG, et al. Glial fibrillary acidic protein as an early marker of hepatic stellate cell activation in chronic and posttransplant recurrent hepatitis C. Liver Transpl 2008;14: 806-14. 63. Guo X, Oshima H, Kitmura T, Taketo MM, Oshima M. Stromal fibroblasts activated by tumor cells promote angiogenesis in mouse gastric cancer. J Biol Chem 2008;283: 19864-71. 64. Noma K, Smalley KS, Lioni M, et al. The essential role of fibroblasts in esophageal squamous cell carcinoma-induced angiogenesis. Gastroenterology 2008;134: 1981-93. 65. Cheng CY, Hsieh HL, Sun CC, Lin CC, Luo SF, Yang CM. IL-1 beta induces urokinase-plasminogen activator expression and cell migration through PKC alpha, JNK1/2, and NF-kappaB in A549 cells. J Cell Physiol 2009;219: 183-93. 66. Kuo PL, Chen YH, Chen TC, Shen KH, Hsu YL. CXCL5/ENA78 increased cell migration and epithelial-to-mesenchymal transition of hormone-independent prostate cancer by early growth response-1 /Snail signaling pathway. J Cell Physiol 2010. 67. Yanagawa J, Walser TC, Zhu LX, et al. Snail promotes CXCR2 ligand-dependent tumor progression in non-small cell lung carcinoma. Clin Cancer Res 2009;15: 6820-9. 68. Begley LA, Kasina S, Mehra R, et al. CXCL5 promotes prostate cancer progression. Neoplasia 2008;10: 244-54. 69. Park JY, Park KH, Bang S, et al. CXCL5 overexpression is associated with late stage gastric cancer. J Cancer Res Clin Oncol 2007;133: 835-40. 70. Miyazaki H, Patel V, Wang H, Edmunds RK, Gutkind JS, Yeudall WA.

68
Down-regulation of CXCL5 inhibits squamous carcinogenesis. Cancer Res 2006;66: 4279-84. 71. Pold M, Zhu LX, Sharma S, et al. Cyclooxygenase-2-dependent expression of angiogenic CXC chemokines ENA-78/CXC Ligand (CXCL) 5 and interleukin-8/CXCL8 in human non-small cell lung cancer. Cancer Res 2004;64: 1853-60. 72. Arenberg DA, Keane MP, DiGiovine B, et al. Epithelial-neutrophil activating peptide (ENA-78) is an important angiogenic factor in non-small cell lung cancer. J Clin Invest 1998;102: 465-72. 73. Lazennec G, Richmond A. Chemokines and chemokine receptors: new insights into cancer-related inflammation. Trends Mol Med 2010;16: 133-44. 74. Nirodi CS, Devalaraja R, Nanney LB, et al. Chemokine and chemokine receptor expression in keloid and normal fibroblasts. Wound Repair Regen 2000;8: 371-82. 75. Mantovani A. Cancer: inflammation by remote control. Nature 2005;435: 752-3. 76. Pietras K, Ostman A. Hallmarks of cancer: interactions with the tumor stroma. Exp Cell Res;316: 1324-31. 77. Sato Y, Murase K, Kato J, et al. Resolution of liver cirrhosis using vitamin A-coupled liposomes to deliver siRNA against a collagen-specific chaperone. Nat Biotechnol 2008;26: 431-42.





























