Embed Size (px)
Citation preview

Congreso del Gincuentenario
I Relate Oficial
Fisiologia y patologia respiratoriadel feto y recien nacido
DRA. ELIANA CERUTI *,
En este relate oficial, dado lo exiguo del tiem-po, solo me referire a algunos conceptos mas im-portantes de fisiologia respiratoria del feto y reciennacido incluyendo entre ellos la adaptacion circu-latoria.
En los ultimos 15 anos se ha despertado uninterns creciente en el estudio del recien nacido;se han ideado nuevas te"cnicas, entre ellas micro-tecnicas, y se han descartado antiguos prejuiciosque han hecho al feto parcialmente accesible alestudio fisiologico.
El pulmon fetal debe alcanzar un determina-do desarrollo para que la funcion que va a per-mitir la vida extrauterina pueda realizarse en for-ma satisfactory a, y como veremos mas adelante, elsurfactante pulmonar es el que facilita una funcionabsolutamente indispensable para la sobrevida ex-trauterina, cual es la estabilidad alveolar.
Circulation materna y fetal al final de la gestation
Aun cuando en el hombre la circulacion pla-centaria no ha sido tan intensamente estudiadacomo en algunos animales, especialmente monos,es muy probable que las diferencias no scan sig-nificativas. En esas especies la sangre materna lle-ga a los espacios intervellosos a traves de las ar-terias espirales, diseminandose en el centro de loscotiledones. El intercambio gaseoso se realiza enlos multiples capilares fetales expuestos a la san-gre materna. En este periodo de la gestacion secalcula que el espacio intervelloso contiene apro-ximadamente 250 ml. de sangre materna. La granfraccion de sangre que ha participado en el inter-cambio gaseoso, mas una pequena qua ha irrigado
* Profesor Asociado de Pediatria. Departamento de PediatriaArea Norte. Jefe Laboratorio Cardiorrespiratorio. HospitalRoberto del Rfo.
el endometrio (shunt materno), se juntan nueva-mente en las venas uterinas. Observaciones simi-lares se han efectuado en el hombre por medio deestudios cineangiograficos. Tambien las arteriasumbilicales presentan anastomosis en la superficiede la placenta fetal. Las arterias se dividen y en-tran en cada cotiled6n donde a su vez se subdividenen vasos mas pequenos y se distribuyen como ca-pilares vellosos. Todas las venas provenientes delos cotiledones se juntan en una sola, la vena um-bilical. No existen valvulas que aumenten la xesis-tencia vascular ni en los cotiledones ni en la venaumbilical. Si bien es cierto que la placenta humanaalcanza su maximo crecimiento a las 36 semanasde gestacion, las terminaciones de los capilares co-rionicos siguen creciendo hasta el termino del em-barazo.
Aherne y Dunnill estudiaron y dividieron elvolumen de la placenta en dos componentes mor-fologicos basicos; el parenquima, formado por lasvellosidades y los espacios intervellosos y la frac-cion no parenquimatosa formada por las capasdeciduales y corionica. Esta ultima fraccion corres-pondc a un 21% del volumen total (volumen pro-medio 488/ml., rango 391-723) y no participa enel intercambio gaseoso; sin embargo es importanteconsiderar que su metabolismo debe ser mante-nido por la circulacion materna o fetal. El paren-quima se divide en tres fracciones: el espacio in-tervelloso, cuyo volumen promedio es de 144 ml.es uno de los compatimientos mas susceptibles alos cambios que se producen por la perdida desangre durante el parto. Los capilares fetales no ex-ceden de 1 mm. de diametro en el septum interco-tiledonico ocupando solo un 12% del parenquima,lo que equivale a un volumen total de 45 ml. To-dos los estudios efecruados demuestran que en unmomento determinado existe mas sangre en el areamaterna que en la fetal.
Rev. Chilena Pediatria, Vol. 44, N1? 6, 1973. 503

El area total dc superficie de las vellosidadcsen la placenta normal se relaciona linealmentecon el volumen ptacentario; al termino de la gcs-tacion varia alredcdor de los 11 m2.
Respiration fetal
Las presiones parciales de los gases mas quesus concentraciones son fundamentales cuando scconsidera el intercambio gaseoso a traves de lasmembranas, incluyendo la placenta (fig. 1). Tan-to el oxigeno como el anhidrido carbonico pasana traves de la placenta por simple difusion. Se hademostrado que la presion parcial de oxigenoarterial (P*O2) de la sangre materna fluctua alrc-dedor de los 95 mmHg y la presion parcial de an-hidrido carbonico arterial (PaCO2) alrcdedor de
los 33 mmHg. La menor P.CO2 de la mujer ern-barazada, coinparada con la de un adulto normal,se explicaria por la presencia de una alcalosis res-piratoria asociada a hiperventilacion; se piensa queesta ultima estaria condicionada por la accion dela prostegerona sobre el centre respiratorio.
El flujo sangumeo de la arteria uterina se di-vide en dos fracciones; la mayor que entra a loscspacios intervellosos y la menor que irriga el mus-culo utcrino. Esta ultima fraction representa unshunt de tal manera que las tensiones de los gasesen la vena uterina, PO^: 33 mmHg y PCOs: 46mmHg, no representan realmente las tensiones delos gases de las venas placentarias maternas. Sinembargo, aun considerando la existencia de esteshunt, los valores de POz tan descendidos y dePCOi tan elevados presentes en la sangre venosa
DIAGRAMA DE LA CIRCULACION PLACENTARIA
ARTERIA
UTERINA
95PO.
PCC) : 332
SHUNT MATERNO
musculo uterino
SHUNT FETAL
VENA
UTERINA 2
ESPACIQ
INTERVELLQSQ
VEVA
UMBILICAL
P02 '-28
PCO :402
ARTERIA
UMBILICAL
Presion parcial ; mm Hg.
504

uterina, demuestran la presencia de un importanteintercambio gaseoso placentario.
A traves de punciones abdominales se ha po-dido conocer aproximadamente los valores pro-mcdio de los gases arteriales en el cspacio inter-velloso: la P8O2 fluctua alrededor de los 40-48mmHg y la PL,CO2 alrededor de los 38 mmHg. Lasangre de la vena umbilical tiene una PaO2 pro-medio de 28 mmHg y una P.CO2 promedio de40 mmHg; en cambio, la PBO2 de la arteria umbi-lical es de solo 15 mmHg, indicando que la san-gre que llega por la aorta descendente a los dife-rentes tejidos fetales es mas hipoxemica que lasangre de la vena umbilical.
Finalmente, es interesante consignar que Boeha demostrado la presencia de un shunt arterio-venoso fetal que puede constituir un factor quecontribuye a la ineficiencia del intercambio pla-centario.
La transferencia de oxigeno a traves de lamembrana placentaria es mas dificil de medir quela difusion de oxigeno pulmonar. Cada moleculade oxigeno desprendida de la hemoglobina debcatravesar el liquido intracelular y la membrana deleritrocito materno; posteriormente el plasma, elsincicio trofoblasto, la membrana basal, el endote-lio del capilar fetal, otra capa de plasma y final-mente la membrana y el liquido intracelular deleritrocito fetal, antes de entrar en contacto con lahemoglobina fetal. El intercambio gaseoso descritoocurre en csta forma solo en las areas placentariascon morfologia mas favorable. En muchas dreas,la difusion esta dificultada porque los capilares fe-tales estan separados del espacio intervelloso ma-terno por una masa variable de tejido que incluychasta celulas de Langhans. Ademas, la morfologiavascular placentaria cambia a mcdida quc la ges-tacion avanza: las vellosidades terminales se achi-
Curva de disociacion de la ox/hemoglobina fetal y maternapH : 7.40
temperaturo.-37 °C
100 r-
% Saturation02
— _ fetalmaterna
W 20 30 40 50 60 70 50 90 100 110 120
Fig. N? 2. — Curva de disociaci6n de la oxihcmoglobina fetal y mafcrna a un pH 7,40 y a una temperature dc 37?C. Notcsedesplazamicnfo dc la eurva fetal hacia hi ixquierda de la materna.
505
2.— Pediatria.

can, los capilares entre cada vellosidad aumentanen numero y se acercan aun mas a la superficiede la vellosidad. Finalmente el sincicio trofoblastoque tapiza los capilares llega a adelgazarse enextreme.
La difusion de oxigeno de la niadre al fetodebe estudiarse considerando la curva de disocia-cion de la oxihemoglobina (fig. 2). En los nivelesen los cuales existe una P0O2 elevada, la curva escasi plana, de manera que ocurren cambios mini-mos de la saturacion, aun cuando haya grandescambios de la P^Oa. Tncluso, cuando existe hipo-xemia matema, por hipoventilacion o por altera-ciones de la relation ventilacion/perfusion, la sa-turaci6n arterial a nivel de los espacios intervello-sos permanece en limites muy cercanos al normal.Cuando la PBO2 es inferior a los 40 mrnHg (partemedia o inferior de la curva), una pequena caidade la PaO2 facilita la liberation de una gran can-tidad de oxigeno desde la sangre hacia los tejidos;esto ultimo es precisamente lo que ocurre en loscapilares fetales.
Entre los factores que favorecen la difusiondel oxigeno desde la madre al feto deben mencio-narse:
a) Concentracion total de hemoglobina: se ha de-rnostrado que tanto la concentracion de ]a he-moglobina como su afuiidad por el oxigeno esmayor en el recien nacido que en el adulto.Posteriormente en el nino, tanto la concentra-cion como la afinidad por el oxigeno descien-den. En el feto la capacidad transportadora deoxigeno de la hemoglobina aumenta rapida-meente durante la gestacion, aunque mas len-tamente al termino de ella; de todas maneras,al termino del embarazo es mayor que Ja deladulto. La PaO2 fetal relativamente baja se pre-sume que actua como estimulo para la herna-topoyesis.
b) Tamano de los globulos rojos: otro factor im-portante en la respiration fetal es el tamanode los globulos rojos. El volumen corpuscularmedio es mayor durante el primer tercio dela gestacion. Por otro lado, el numero de ce-lulas rojas por unidad de volumen de sangrees menor durante esa misma etapa.
c) Diferencias entre las curvas de disociacion dela oxihemoglobina fetal y materna (Fig. 2).Se ha comprobado que a un mismo pH, la cur-va de disociacion de la hemoglobina fetal estadesplazado hacia la izquierda con respecto ala materna. Parte de esta diferencia reside enel contenido electrolitico, en el contenido debase y en la actividad de la anhidrasa carbo-nica del medio celular, mas bien que en lasdiferentes propiedades de la molecula de he-
moglobina fetal o materna. La diferencia enla position de ambas curvas condiciona que acualquiera PaO2 la sangre fetal tenga una ma-yor afinidad por el oxigeno que la sangre ma-terna.
d) Efecto de Bohr positivo: cuando en la sangrematerna aumenta la PaCO2 y la cantidad deacidos fijos, aumenta tambien la concentra-cion de iones (H+); esta acidification traeconsigo una disminucion de la afinidad de lahemoglobina por cl oxigeno. En estas condi-ciones se favorece la Iiberaci6n de oxigeno des-de la sangre materna a la fetal.
e) Efecto de Bohr negativo: cuando se realiza elintercarnbio gaseoso a nivel de los espaciosintervellosos disminuye la acidez de la sangrefetal, aumcntando la afinidad de esta por eloxigeno. En estas circunstancias se produceuna mayor difusion de oxigeno desde la san-gre materna.
Entre los factores que se oponen a la difusi6nde oxigeno desde la madre al feto deben mencio-narse la disminucion de la capacidad transporta-dora de oxigeno de la hemoglobina fetal y la ma-yor acidez de la sangre fetal que resulta en unadisminucion de la afinidad de esta por el oxigeno.La hemoglobina materna tiene una maxima capa-cidad transportadora de 1,34 mol. de oxigeno porgramo; en cambio, la hemoglobina fetal puedetransportar solo 1,26 mol. de oxigeno por gramo.En general, esta propiedad no es importante enlos rangos de saturacion comprometidos en la di-fuci6n placentaria.
Consumo de oxigeno
El intercarnbio de oxigeno a traves de la pla-centa esta mediado por todos los factores ante-riormcnte cxpucstos, pero realmente se relacionamaximamente con el consumo de oxigeno fetal.La P*O2 fetal relativamente baja favorece la crea-tion de una gradiente a traves de la membranaplacentaria, habiendose comprobado una gradien-te de difusion de 8 mmHg, que aproximadamentetransfiere 0,4 a 0,6 ml. de oxigeno por minuto ypor mmHg. de diferencia.
Aun cuando es evidente que la PO* del fetonormal in utero es bastante menor que la obser-vada en el recien nacido y en el adulto, se puedeconsiderar que la oxigenacion del feto es normal.Esto es posible gracias a la presencia de un debitocardiaco elevado que asegura un flujo sanguineoaumentado tanto a la placenta como a los tejidosfetales. Se presume ademas, que los tejidos feta-les tienen una red capilar mas rica y que la difu-
506

sion de oxigeno es mas rapida o bien que su uti-lization es mayor.
Difusion del anhldrico carhonico desde el fcto ala madre
La sangre fetal tiene menor afinidad por elCOs que la sangre materna, favoreciendose, dccste modo, la liberation de CO* desde el feto a lamadre. Durante el intercambio placentario, la san-gre fetal se- enriquece con O^ y tiende a liberarCOe; por el contrario, la sangre materna al perderO2 tienc mayor afinidad por el CO? (efecto Hal-dane).
Regulation de los gases arteriales y del pH jetales
Hasta el momento es imposible precisar todoslos mecanismos comprometidos en la regulationde los gases arteriales y del pH fetales. Existen evi-dencias de que el flujo sanguineo placentario tan-to materno como fetal y el area de contacto en-tre las dos circulaciones interfieren en el intercam-bio de O2 y COa umbilical. La reduccion de laconcentracion de hemoglobina fetal tendria unefecto similar; ambos reducen la cantidad total deOs transportado desde la placenta hacia los tejidosfetales.
La ventilation materna influye en la regula-cion de los gases arteriales del fcto. Si la madrehiperventila, la PaO2 del feto se eleva, disminuyen-do la PaCOi. La hipoventilacion materna actua ensentido opuesto. La hiperventilacion fisiologica dela madre durante el embarazo produce en ella unaalcalosis respiratoria que tiende a mantener el pHy la PaCO2 sanguineos y celulares del feto a nive-les cercanos a los observados en el adulto.
Respiration intrauterina
Aim cuando se han comprobado movimientosrespiratorios ritrnicos en el feto, no se ha demostra-do que el establecimiento de la respiration des-pues del nacimiento sea la continuation de unafuncion iniciada en la vida fetal. En apariencia es-tos movimientos muy escasos al termino del em-barazo, no lograrian producir un adecuado inter-cambio gaseosa pulmonar.
Regulacion termica del feto
Durante la vida intrauterina el intercambio decalor a traves de la placenta permite mantener latemperatura corporal del feto. En condiciones es-tables, la temperatura fetal es aproximadamente0,5?C. mayor que la de la madre. Experimentosen ovejas han demostrado que la alteration de laperfusion placentaria manifestada por la disminu-
cion de la P.O. y el aumento del pH del feto, esacompanada de un aumento de la gradiente detemperatura entre el feto y su madre. Cuando elfeto muerc su temperatura se iguala a la de lamadre.
DesarroUo del drbol vascular pulmonar
Durante el desarrollo fetal se observan cambioshistologicos progresivos del arbol vascular puhno-nar que se acentuan mas aun con el establecimien-to de la respiracion.
Proximo al cuarto mes de gestation las arte-rias y artcriolas pulmonares presentan un lumenmuy estrecho en relacion con el grosor de su pa-red. Aun cuando en cl ultimo periodo de la vidaintrauterina y durante los primeros dias de vidael lumen se agranda en forma progresiva, siem-pre es estrecho en relacion al diametro total delos vasos pulrnonares. A partir del decimo o duo-decimo dia de vida, la relacion lumen/diametrototal es muy semejante a la del adulto.
El arbol vascular fetal tiene caracteristicas es-peciales; alta resistencia y gran labilidad. Estascualidades dcpenden de las condiciones histoio-gicas ya expuestas, como son el desarrollo de unapared muscular extremadamente gruesa.
La expansion gaseosa pulmonar produce vaso-dilatacion independientemente de la composiciondel gas inhalado. Igualmcnte, los cambios en lacomposicion de los gases, ya sea de la sangre queirriga los pulmones fetales no ventilados o bien dela mezcla gaseosa que se usa para ventilar lospulmones, produce alteraciones en la resistenciavascular pulmonar. El aumento de la POs y lareduccion de la PCOsde la sangre perfundida o dela mezcla gaseosa usada para ventilar los pulmo-nes produce vasodilatacion pulmonar importante.
La resistencia vascular pulmonar disminuyecon rapidez una vez iniciada la expansion pulmo-nar, pero la presion de la arteria pulmonar per-manece elevada por mas tiempo, alcanzando va-lores aproximados a los del adulto solo a fines dela segunda semana de vida. Despues de una rapidacaida initial, la resistencia vascular pulmonar si-gue disminuyendo paulatinamente hasta el cuartou octavo ano de vida.
El flujo sanguineo pulmonar del feto es muybajo, 10 a 20% del debito derecho, y no es cons-tante; es influenciado por la administration dehistamina, acetilcolina y el grado de oxigenacionfetal. Cuando la saturation de Oa arterial del fetose eleva, se observa un significative aumento delflujo pulmonar. Si el pulmon materno es ventiladocon mezclas con baja concentracion de Oa, el flujosanguineo pulmonar disminuye en un 25%. Laasfixia fetal condiciona una intensa vasoconstric-cion del arbol vascular fetal.
507
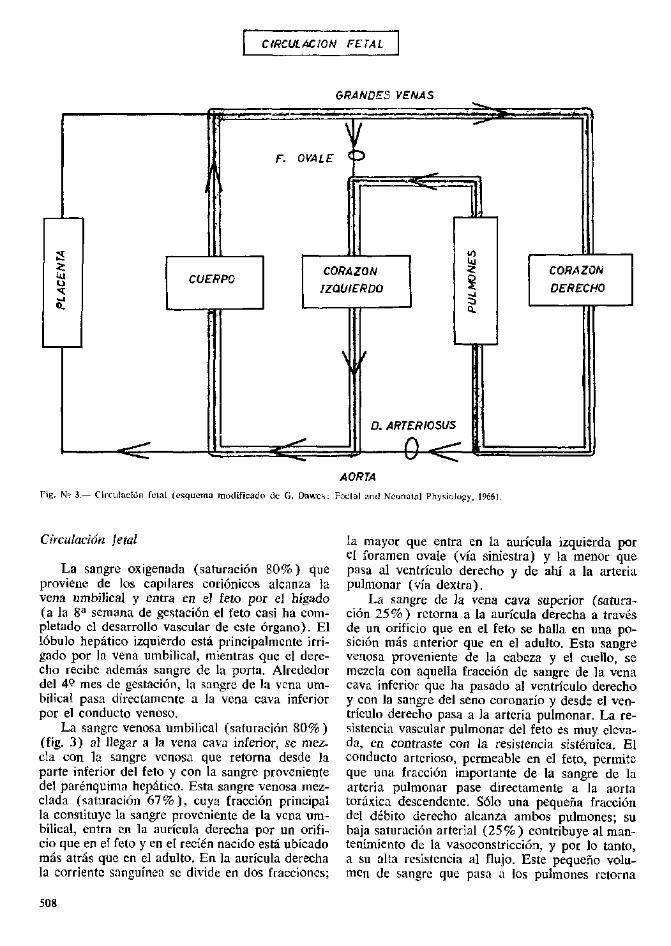
CtRCULACIQN FETAL
GRANDES VENAS
5
Ujo
MF. OVALE
CUERPOCORAZON
IZQUIERDO
D. ARTERIOSUS
•Q-
CORAZON
DERECHO
AORTA
Fig. N? 3.— Circulacion felal (esqucma modificado de G. Dawcs: Facial and Neonatal Physiology, 1966).
Circulocidn fetal
La sangre oxigenada (saturacion 80%) queproviene de los capilares corionicos alcanza lavena umbilical y entra en el feto por el higado(a la 8a semana de gestacion el feto casi ha com-pletado cl desarrollo vascular de este organo). Ellobulo hepatico izquierdo esta principalmcnte irri-gado por la vena umbilical, mientras que el dere-cho recibc ademas sangre de la porta. Alrededordel 41? mes de gestacion, la sangre de la vena um-bilical pasa directamcnte a la vena cava inferiorpor el conducto venoso.
La sangre venosa umbilical (saturacion 80%)(fig. 3) al llegar a la vena cava inferior, se mez-cla con la sangre venosa que retorna desde laparte inferior del feto y con la sangre provenientedel parenquima hepatico. Esta sangre venosa mez-clada (saturacion 67%), cuya fracci6n principalla constituye la sangre proveniente de la vena um-bilical, entra en la auricula derecha por un orifi-cio que en el feto y en el recien nacido esta ubicadomas atras que en el adulto. En la auricula derechala corrientc sangumea sc divide en dos fraccioncs;
la mayor que entra en la auricula izquierda porel foramen ovale (via siniestra) y la menor quepasa al ventriculo derecho y de ahi a la arteriapulmonar (via dextra).
La sangre de la vena cava superior (satura-cion 25%) retorna a la auricula derecha a travesde un orificio que en el feto se halla en una po-sicion mas anterior que en el adulto. Esta sangrevenosa proveniente de la cabeza y el cuello, semezcla con aquella fraccion de sangre de la venacava inferior que ha pasado al ventriculo derechoy con la sangre del seno coronario y desde el ven-triculo derecho pasa a la arteria pulmonar. La re-sistencia vascular pulmonar del feto es muy elcva-da, en contraste con la resistencia sistemica. Elconducto arterioso, permeable en el feto, permiteque una fraccion importante de la sangre de laarteria pulmonar pase directamente a la aortatoraxica descendente. Solo una pequena fracciondel debito derecho alcanza ambos pulmones; subaja saturacion arterial (25%) contribuye al man-tenimiento de la vasoconstriccion, y por lo tanto,a su alta resistencia al flujo. Este pequeno volu-mcn de sangre que pasa a los pulmones retorna
508

a la auricula izquierda por las venas pulmonares;aqui se mezcla con la sangre mejor oxigenada queha pasado desde la auricula derecha por el fora-men ovale. Del ventriculo izquierdo pasa a laaorta, a los vasos coronarios, y se dirige a la partesuperior del feto (cabeza y cuello). En la aortatoraxica descendente vuelve a mezclarse con lasangre probremente oxigenada que ha pasado porel conducto arterioso desde la arteria pulmonar.La sangre de la aorta descendente, con menorsaturation (58%) que la de la aorta ascendente,se distribuye en las visceras abdominales y en lasextremidades inferiores del feto, retornando a laplacenta por las arterias umbilicales.
Surfactante pulmonar
El surfactante pulmonar facilita una funcionabsolutamente indispensable para la sobrcvida ex-trauterina, cual es la estabilidad alveolar; los pul-mones pueden ser expandidos y las fuerzas quese oponen a esta expansion pueden ser sobrepasa-das. El surfactante pulmonar se desarrolla y ma-dura durante la vida fetal y cuando su sintesis sealtera, el recien nacido presenta el sindrome de di-ficultad respiratoria idiopatico.
En los primeros meses de la gestacion, los al-veolos son solidos y los pulmones no pueden serexpandidos; en cambio, en el ultimo periodo delembarazo los pulmones se expanden aun cuandolas presiones iniciales requeridas para distenderlosson mayores que las que se necesitan posteriormen-te. Aun mas, en este ultimo periodo, los pulmonespermanecen expandidos despues de la primcra in-flation. Este fenomeno solo puede ser explicadoconsiderando los factores que determinan la ten-sion superficial alveolar.
La tension superficial es un fenomeno que ocu-rre en la mterfase aire-liquido del alveolo pulmo-nar y es la resultante de las diferencias en la atrac-cion de las moleculas de las superficies liquiday gaseosa que tapizan el alveolo, Es decir, la ma-yor fuerza de retraction encontrada en el pulmoncon aire, se debe a la existencia dentro del alveolode esta interfase aire-liquido que significa la crea-tion de una energia de superficie. Los compo-nentes de la interfase definen la unidad de energia,tension superficial, y su magnitud se expresa endinas/cm. Ya en 1954 Radford habia calculadoque las fuerzas de tension superficial existentesen los pulmones eran suficientes para producirsu colapso total. Posteriormente Mead demostrola existencia en el pulmon de un determinado tipode substancias con notables propiedades tensoac-tivas y Clements propuso la existencia dentro delalveolo de un material que lo tapizaba y que de-terminaba que la tension superficial se alterabacon los cambios del area de superficie. Inmedia-tamente despues se publico la conclusion de que
en el pulmon del adulto y del recien nacido normalexiste una substancia con propiedades tensoactivasy que esta esta ausente en los pulmones de los re-cien nacidos con sindrome de membrana hialina.
El surfactante pulmonar o substancia tenso-activa es un complejo que tapiza el alveolo. Estacompuesto de una variedad de substancias, inclu-yendo hidratos de carbono y proteinas, pero prin-cipalmente lipidos. Entre estos ultimos el mas abun-dante (50-70%) es la lecitina que contiene acidosgrasos saturados tanto en el carbono a como enel [3. En el pulmon normal la fraction lipidica estapresente en una concentration de alrededor de 5mg/grn; aproximadamente 1/8 de la concentra-tion total de lipidos pulmonares.
El surfactante pulmonar tiene la propiedad dedisminuir la tension superficial y estabilizar el al-veolo. El alveolo cambia su radio durante el ciclorespiratorio; durante la inspiration el radio au-menta, con la espiracion disminuye. Los efectosdel cambio del radio alveolar se expresan mejorde acuerdo a la ley de Laplace que establece ba-sicamente que la tension superficial es inversamen-te proporcional al radio. Por lo tanto, cuando elradio disminuye durante la espiracion, la tensionsuperficial aumenta en forma importante y si noexistiera alguna substancia estabilizante que dismi-nuyera la tension superficial, el alveolo se colap-saria durante la espiracion.
El desarrollo del pulmon fetal involucra unacantidad de procesos tales como el desarrollo ana-tomico de los bronquios y alveoles, la formation ycrecimiento de los vasos sanguineos, especialmen-te dentro de las areas alveolares, la especializa-cion de las celulas alveolares tipo II y los cam-bios del epitclio alveolar. Sin embargo, la funcionpulmonar depende fundamentalmente de las subs-tancias tensoactivas, mas propiamente de la dipal-mitol-lecitina.
En el feto humano se ha demostrado la exis-tencia de 2 vias de sintesis de la lecitina. La pri-mera y mas importante es la catalizada por la fos-focolintransferasa segun la siguiente reaction:
Difosfato de cistidincolina + D-a-,3digliceridos > lecitina
Esta via de sintesis madura aproximadamen-te a las 35 semanas de gestacion y sintetiza prin-cipalmente Dipalmitol-Lecitlna.
La segunda via, mucho menos importante, esla catalizada por la mctiltransferasa segun la si-guiente reaction:
Fosfotidiletanolamina + 3 CH^(metionina) > lecitina
A traves de esta rcaccion se sintetiza principal-mente Palmitol-M/r/s-^/ Lecitina. Esta via ya es
509

posible identificarla alrededor de las 22 a 24 se-manas de la gestacion; su maduracion aumeata alte"rmino del embarazo y durante los primeros diasde vida. Es importante consignar que esta via esla iinica que permite nacer prematuramente y quees facilmente inhibida por acidosis, hipotermia ehipoxia.
La via de sintesis inas importante y la querealmente marca la maduracion del pulmon es laprimera de ellas, es decir, la que sintctiza dipal-mitol-lecitina. Esta via esta presente en mamife-ros tales como conejos, lauchas, ovejas, cabras, etc.Sin embargo, la segunda via, metilacion, practica-mente no tiene actividad en la mayoria de esasespecies y solo se activa dias o semanas despuesdel parto. De esta manera, solo los seres humanosy no otros primates pueden nacer prematuramente.
En todas las especies nombradas la primera viaalcanza su maduracion fetal durante la vida in-trauterina. En el feto humano su maduracion al-canza un 90% alrededor de las 35 semanas degestacion.
Es importante considerar que una informationadecuada del surfactante pulmonar ha permitidoun mejor entendimiento del desarrollo fetal. Elconocimiento de la lecitina fetal, a traves de ladetermination de los fosfolipidos que salen desdeel alveolo hacia el liquido amniotico, ha resultadoser un procedimiento muy util para predecir elestado de maduracion del pulmon fetal. Se ha com-probado que la relaci6n lecitina/esfingormeliiia enel liquido amniotico es de dos o superior a dos alas 35 semanas de gestacion. Relaciones menoresde dos predicen el nacimiento de ninos con gra-dos variables de sindrome de membrana hialina.
Es importante enfatizar que los estudios bio-qufmicos del pulmon fetal han sido los x>rimerosy mas importantes parametros para reconocer lamaduraci6n fetal y actualmente se consideran in-dicadores de la evaluation fuacional del feto. Lamas importante determinaci6n de la capacidad deun feto para una adecuada sobrevida extrauterinaesta dada por la capacidad que tienen sus pulmo-nes para ventilarse. Por lo demds, la correlationentre estos parametros y otras funciones funda-mentales, especialmente la maduracion del higadoy el S. N. C., ha demostrado ser excelente. Se hacomprobado que los mismos factores intrauterinosque aceleran la maduraci6n bioquimica del surfac-tante pulmonar aceleran tambien la maduracionneurologica.
Estableclmiento de la respiration
Cuando un nino nace es removido desdc unambiente caliente y humedo a un ambiente frioen el cual su peso corporal ya no es mantenidopor ningun liquido aislantc. Tambien el recien na-
cido nace un poco asfixiado y justamente en esemomento se le liga el cord6n umbilical. Adema"sde todos los hechos anteriores en el momentodel parto mismo el nifio es apretado y estrujado.Considerando todos estos cambios relativamenteviolentos es importante decidir cuales son los es-timulos esenciales que permiten al recie"n nacidoiniciar una respiration espontanea y ritmica y noes sorprendente que estos mecanismos hayan sidodiscutidos durante anos. Mucho se ha especuladoen relation a los factores que contribuyen al es-tablecimiento de la primera respiration. Atribuireste fenomeno solo a los estimulos propioceptivosparece muy improbable ya que durante el naci-miento se producen cambios significativos de laspresiones parciales de los gases arteriales y delpH. Es conocido ademas, que las variaciones dela temperatura ambiental y la presencia de esti-mulos tactiles, visuales y auditivos tienen tambienimportancia en el establecimiento de la respira-ci6n.
Considerando que el feto no sufre de hipoxe-mia y que su equilibrio acido-basico es similar alde la niadre, es posible pensar que la estimulacionrefleja del SNC a traves de los quimioreceptoresaorticos y carotideos, como resultado de la asfixia,sea uno de los principales estimulos para el esta-blecimiento de la respiration. No hay dudas quela caida de la P,O2 y del pH y el ascenso de laPaCO2 inducen movimientos respiratorios no s61oen el recien nacido sino tambien en el feto. Estosmovimientos irregulares en un primer momento,rapidamente son reemplazados por una respiraci6nde tipo regular una vez que tiende a normalizarsela oxigenacion y el equilibrio acido-basico del re-cie"n nacido. La respuesta a la asfixia depende dela presencia de los quimioreceptores a6rticos ycarotideos que se sabe son funcionalmente activosen el periodo neonatal inmediato. En la fig. 4 se
Tig. N; 4 .— Rcsultados promcdio de un estudio de 35 reci^anacidos en que se relacionan los cambios de la venti]aci6n conla prcsion parcial de oxigeno en el aire inspirado (PaO2). Seobserva como a medida que disminuye ]a PaO2 los recien naci-dos rcspondcn con un aumento significative y paralelo de laveniilaci6n. (de Brady, J. y Ceruti, E. J. Physiol. (London) JS4:
631, 1966).
510

muestra un experimento en el cual comprobamoscon la Dra. Brady justamente el hecho que acabode enunciar. Se observa que cuando se le admi-nistra a un recien nacido de solo 20 minutos devida una baja concentration de oxigeno en el aireinspirado (12% de O*), el nino responde con Tinaumento de la ventilation y que este aumento dela ventilation es mayor cuanto menor es la con-centracion de oxigeno en el aire inspirado.
Es improbable que la asfixia sea el unico es-timulo que contribuya a iniciar la respiration. Laestimulacion de los receptores termicos, inmedia-tamente despues del nacimiento es intensa. Calcu-los basados en el grado de enfriamiento de la pielde los recien nacidos durante los primeros minu-tos de vida, ban demostrado que a una tempera-tura ambiente de 21 a 25*?C., el recien nacido pier-de calor rapidamente, aproximadamente 600 cal.por minuto (200 cal/kg/min.). Dewes ha demos-trado experimentalmente que el frio por si solopuede ser usado para estimular la respiration. Esteautor comprobo en ovejas recien nacidas que cuan-do la temperatura rectal cac 2 a 3°C empiezan losmovimientos respiratorios. En la fig. 5 se observanlos registros de presion intraesofagica dcterminadosen un feto maduro (oveja) con circulation umbi-lical intacta y con los pulmones repletos de liquido.Se observa que cada vez que el feto se enfria conun ventilador se inician movimientos respiratorios(registros de presion intraesofagica).
La estimulacion de los receptores tactiles essecundaria; solo induce movimientos respiratoriosaislados incapaces de establecer una respirationritmica.
La oclusion del cordon umbilical, ha demos-trado no tener importancia, ya que pueden ini-ciarse movimientos respiratorios aun en presenciade una circulation umbilical intacta.
En relation a los factores mecanicos que inter-vienen en la primera respiration es importante des-tacar que la fuerza requcrida para la primera ins-piraci6n tiene que sobrepasar la viscosidad del
PRESION ESOFAGICA (mm Hg.)
VENTIUDOR
t con ventiladorI sin ventilador
Fig. N^ 5.— Registro de prusidn inlraesofagica en im fclo (ove-ja) maduro con circulacion umbilical intacta y pulmones re-pletos de liquido. Notese que al enfriar la cara del feto conun ventilador se inician movimientos respiratorios. (dc G. Da-
\vcs. Foetal and Neonatal physiology, 1966).
liquido contenido en las vfas aereas, las fuerzasde tensi6n superficial de los alve"olos y la resis-tencia tisular ela"stica y no elastica. El que las pre-siones requeridas para variar el tamano del alveo-lo, una vez establecida la primera respiration, seanfrancamente menores, hace pensar que la resis-tencia no es un factor importante. Al comenzarla primera respiration, antes de la entrada deaire a los pulmones, se desarrolla una alta presionnegativa intratoraxica de 40 a 60 cm. de agua. Amedida que la presion negativa va aumentando,los alveoles se abren y permiten la entrada deaire, aproximadamente 50 ml. Cuando el reciennacido aspira, solamcnte parte de este aire sale,quedando en los alveolos aproximadamente un30 a 40% del aire que primitivamente entro (ca-pacidad funcional residual). De esta manera, lapresion necesaria para la segunda inspiracion esmenor y asi va sucediendo sucesivamente, de modoque rapidamente despues de las primeras respira-ciones, el recien nacido desarrolla curvas presion/volumen semejantes a las del adulto, requiriendopresiones negativas de no mas de 5 cm. de aguapara expandir el pulmon.
De lo expuesto anteriormente se deduce que lamaxima rcsistencia initial esta condicionada porla gran tension superficial existente en los alveolos.La importancia del material surfactante, ha sidoadmirablemente bien demostrado por Dawes yReynolds. Se ha demostrado en el feto humano, yaen la 23^ semana de gestation, la presencia desurfactante pulmonar cuya cantidad va aumentan-do a medida que avanza el embarazo. Es proba-ble que no se disponga de ella en una cantidadsuficiente hasta las 30 a 35 semanas de gestation.En la fig. 6 se muestran las curvas presion/volu-men que se obtienen inflando los pulmones deovejas recien nacidas maduras (140 dias de ges-tation), e inmaduras (120 dias de gestation),dentro del primer dia de vida. En las primeras, lalinea continua muestra que es necesario aplicaruna presion de 18 cm. de agua (presion initial)antes de que un volumen significative de aire entreen los pulmones. La distension completa de estosse alcanza con una presion de 40 cm. de agua.En la deflation, aproximadamente se retiene, co-mo volumen residual, un cuarto del volumen ma-ximamente alcanzado. Es decir, ya con la primeradistensi6n pulmonar queda un determinado volu-men residual. Se observa ademas, en linea corta-da, que para la segunda respiration no se necesitauna presion initial y que el pulm6n se distiendehasta su capacidad maxima con una presion desolo 15 a 18 cm. de agua. La misma figura mues-tra la curva presion/volumen de un feto inmadu-ro de 120 dias dc gestation. Se observa que laprimera respiracion es igual a la del recien nacidoa termino, pero que al final de la primera defla-tion el pulm6n se colapsa totalmente no quedan-
511

Curvas presion volumen (oveja}:
VOLUMENPULMQNAR
% VOLUMENPULMONAR
100r- 100
50
10 20
Presion (
Feto a term/no (140 dt'as gestacion)
JO 20
Presion
30 40
Feto inmaduro (120 dias gestacion)
Fig. N? 6.— Curvas de prcsi6n/volumen obtenidas inflando los pulmones de fetos (oveja) maduros e inmaduros. K6tcse quela segimda respiraci6n (linca cortada) en el feto maduro no requiere dc una prcsion inicial como la primera y que el pul-mon sc distiende maMmamcnte con srtlo una presi6n de 15 a 20 cm. de agua. En el feto inmaduro se demuestra que parainiciar la segunda respiraci6n se requiere de una prcsi6n tan elcvada como la necesaria para la primera respiraci6n y que atfinal dc ella el pulmdn se colapsa lotalmente. (dc G. Dawcs. Foetal and Neonatal Physiology, 1966).
do volumen residual, a diferencia de lo ocurridoen el feto maduro. Se demuestra ademas, conlinea cortada, que para la segunda respiracion, elpulmon neccsita ser distendido completamente yque la'presion inicial requerida es tan grande comoen la primera respiracion, con el logico aumentodel trabajo respiratorio. Aiin mas, al medir losestractos de surfactante de ambos pulmones, Dawesy Reynolds comprobaron que los estractos delpulmon del recien nacido maduro alcanza una mu-cho menor tension superficial, y que los pulmonesdc los recien nacidos inmaduros se colapsan total-mente durante la espiracion porque en ellos per-siste una elevada tension superficial.
En resumen, los pulmones necesitan desarro-llar los espacios aereos y el surfactante pulmonarantes de que puedan ser ventilados y ofrecer asiuna minima resistencia a las primeras inspiracio-nes.
En relacion al destino posterior del liquido con-tenido en el pulmon fetal, tres vias parecen proba-bles: 1) expulsion del liquido durante el periodoexpulsive del parto; se ha determinado que a tra-
ves de esta via se elimina un volumen de mas omenos 7 a 42 ml.; 2) pasaje del liquido a la cir-culacion a traves de los capilares pulmonares; esprobable que el bajo contenido proteico del liqui-do intrapulmonar, en comparacion con el de lasangre, favorece su rapida absorcion tan prontocomo el flujo pulmonar aumenta, y 3) penetraciona la circulacion a traves de los linfaticos.
Una vez iniciada la respiracion, la distribuciondel aire inspirado es tan importante como la can-tidad total de aire inspirado o espirado en cadaciclo respiratorio. En el recien nacido normal, ladistribucion de la ventilacion es uniforme y la ca-pacidad funcional residual practicamente no cam-bia durante los primeros dias de vida lo que im-plica, entre otros factores, la presencia de unabaja tension superficial.
La iniciacion de la funcion respiratoria no soloimplica el establecimiento de una ventilacion ade-cuada sino tambien de una perfusion pulmonarque corresponda a la ventilacion alveolar con elobjeto de mantener una buena relacion ventila-cion/perfusion. Tambien la distribucion del flujo
512

sanguineo es tan importante como la cantidad ab-soluta de sangre que llega al alveolo. Se ha demos-trado que en el recien nacido normal no existeuna gradiente arterio-alveolar de COa significativa,sin embargo, la gradiente alveolo-arterial de Oaes tres veces superior a la del adulto, Este au-mento de la gradiente de Oa no esta condicionadopor la presencia de una ventilacion irregular, yaque se ha demostrado que esta es uniforme desdeel nacimiento, sino por la presencia de un shuntde derecha a izquierda, que en el recien nacidonormal puede llegar hasta un 25% del gasto car-diaco.
En el recien nacido los vasos pulmonares tie-nen paredes musculares mas gruesas y lumenesmas pequenos comparados con los del adulto.Cualquiera sea la causa de esta diferencia histolo-gica, los vasos pulmonares del recien nacido reac-cionan en forma diferente mostrando un tono va-somotor extremadamente active; vasoconstriccioncon la adrenalina y extrema vasodilatacion con lahistamina y acetilcolina. El aumento de la PaO2, ladisminuci6n de la PaCO2 el ascenso del pH y lainiciacion de la respiracion ritmica contribuyen ala vasodilataci6n pulmonar y por lo tanto, a ladisminucion de la resistencia vascular pulmonar.Estos factores, junto con el gasto cardiaco, deter-minan el flujo sanguineo pulmonar del recien na-cido. ,.
Adaptation circulatoria del recien nacido
La transicion entre la circulacion fetal y laadulta no se produce en forma abrupta sino gra-dual; el foramen ovale y el conducto arterioso per-manecen permeables durante cierto tiempo des-pues del nacimiento. Cuando se ocluye el cordonumbilical, la presion de la auricula izquierda so-brepasa la presion de la auricula derecha, favore-ciendose asi el cierre del foramen ovale. Este cierrcno ocurre inmediatamente; en recien nacidos nor-males se ha observado la persistencia de un shuntde izquierda a derecha por el foramen ovale du-rante las primeras horas de vida.
El conducto arterioso se contrae despues delnacimiento, pero permanece permeable durante 24a 72 horas de vida. Durante las primeras 6 a 8horas se produce un flujo bidireccional a travesdel ductus, que se influencia por los cambios depresiones que ocurren durante la respiracion yel llanto. Posteriormente pasa a constituirse unshunt de izquierda a derecha que persiste durantelas siguientes 15 a 36 horas de vida. Cuando a losrecien nacidos se Ics administra una baja concen-tracion de O:> en el aire inspirado se produce unavasoconstriccion pulmonar, la presion de la arte-ria pulmonar se eleva sobre la presion sistemica,invirtiendose el flujo a traves del ductus e insta-landose un shunt de derecha a izquierda como el
que existe en el feto. Esta respuesta ha servidopara demostrar que el principal estimulo para laconstriccion del ductus es la presencia de una ele-vada P»O2. Aun mas, se ha observado constricciondel ductus en presencia de altas concentracionesde O2, incluso despues de su denervacion.
Tanto el shunt a nivel del foramen ovale co-mo a nivel del ductus tienen efectos adversos. Losflujos bidireccionales, si son de cierta magnitud,reducen la efectividad del gasto cardiaco. Un au-mento del shunt de derecha a izquierda, junto conuna difusion alterada y la presencia de atelecta-sias, condiciona deterioro progresivo de la oxigc-nacion.
El balance entre la resistencia vascular siste-mica y puhnonar y el grado de constriccion delductus determinan, en ultimo termino, la magnitudy la direccion del shunt cardiovascular.
En el 37% de los recien nacidos normales exis-te un soplo sistolico "in crescendo'1 durante lasprimeras horas de vida. Este soplo se observa conmayor frecuencia en recien nacidos asfixiados, enprematures, y en nifios mantenidos a temperaturasambientales inferiores a la neutral (23 a 25?C). Sise considera que la produccion de un soplo de-pende de cierta velocidad critica del flujo sangui-neo, signo clasico de la persistencia del ductus enninos mayores, se presenta con muy poca frecuen-cia en los primeros meses de la vida, porque encsa epoca hay escasa diferencia de presiones entrela circulacion pulmonar y sistemica. Cuando elductus es ampliamente permeable, el volumen san-guineo que pasa por el puede ser grande, pero lavelocidad es muy pequena para lograr la produc-cion de un soplo audible. A medida que el ductusse va contrayendo, la velocidad aumenta y apa-rece el soplo.
Reflejos cardiovasculares y resplratorios
Diferentes autores en diferentes animates bansugerido que la presencia dc los quimioreceptoresarteriales, aorticos y carotideos, es fundamentaly asegura la primera respiracion profunda reque-rida para distcnder los pulmones. Para demostraren el recien nacido humano lo que otros autoreshabian comprobado en animales, con la Dra. Bra-dy planeamos una serie de experiencias destinadasa estudiar la actividad de los quimioreceptoresdesde el momento del nacimiento. Al mismo tiem-po pudimos estudiar algunos de los mecanismosde control de la ventilacion y circulacion del re-cien nacido. Demostramos que tanto los quimio-receptores perifericos como centrales son activosen el recien nacido. Los primeros entran en funcio-namiento aun cuando los centrales esten deprimi-dos por anoxia, hipercapnia, traumatismo o anes-tesia. Los quimioreceptores perifericos serian esen-ciales para la iniciacion de la primera respiracion,
513

It
Fig. N'.1 7.— Efecto de la adrninistraci6n de una clcvada con-centracion de oxigeno en cl aire inspirado (100%) en un recie'nnacido a t^rmino. Notese la calda brusca del volumen corrienteen reposo mienlras el nino rcspira 1DO?6 O2. (de Brady, J. y
cols. J. Physio! (London) 172: 332, 1964)).
£-:^*;£C^".V^^i^M^mFig. N? 8.— Porcentaje de cambio de la ventilacion cuando re-cicn nacidos respiran 21 y 12% de O2, tanto a lemperatura neu-tral como en un ambiente frio (28?C). Resultados promedio de10 nin.os. Notesc la abolicion de la hiperventilacion transitoriaque sc observa con la hipoxia en ambiente neutral, en los re-citn nacidos mantenidos a 28?C. (de Ceruti, E. Pediatrics 37:
556, 1966).
particularmente cuando los receptores centrales es-tan deprimidos.
Los quimioreceptores ya estan presentes enun embrion de 36 mm. y son tonicamente activosen el momento de nacer.
En la figura 7 se demuestra, en un recien na-cido de 1/2 hora, que al respirar una alta concen-tration de O:> (100%) inmediatamente disminuyela ventilacion.
Pudimos demostrar ademas, fig. 8, que duran-te la primera semana de vida los recien nacidosque se mantienen a temperatura neutral, es decir,a aquella temperatura ambiental en la cual el con-sumo de O2 es minimOj cuando respiran una bajaconcentration de O^ en el aire inspirado (12%),responden con una hiperventilacion transitoria se-guida de hipoventilacion. Observamos tambien queesta hiperventilaci6n transitoria ni siquiera se pre-senta cuando los recien nacidos son mantenidosa una temperatura ambiental mas fria, aunquede solo 28?C. Es decir, que la respuesta normalque observanios en el recien nacido con la hipoxiaes abolida por el Mo, comprobandose en este casosolo una depresion de la ventilacion. Quiero in-sistir, que en esta expcriencia, cuando hablo defrio me estoy refiriendo a una temperatura am-biental de 28*?C, temperatura en todo caso supe-rior a la que existe en todas muestras maternida-des. En esta misma experiencia demostramos queesta respuesta es peculiar al recien nacido menorde una semana, ya que la depresion de la venti-lacion con la hipoxia que se observa a 289C., no
Fig. Nv 9.— Efccto dc la hipoxia en un recien nacido durante el]er. y 7 dia de vida. N6tesc que despues de la primera semanade vida, la hipcrvcnlilacidn desencadenada por la hipoxia semantiene tanto a temperatura neutal como en un ambiente
friu (28?C). (de Ccruti, E. Pediatrics 37: 556, 1966).
se demuestra despues de la primera semana de vi-da cualquiera sea la temperatura ambiental (Fig.y).
De Jos estudios recientemente expuestos se de-duce que la efectividad de los reflejos se reduce
514
notablemente cuando disminuye la temperaturaambiental, por lo tanto, es muy importante recal-car que el recien nacido no debe ser expuesto atemperaturas ambientales inferiores a la neutral.
Como en el adulto, el centre respiratorio delrecien nacido es muy sensible al aumento de CO*y es estimulado tanto directa como indirectamentepor el aumento de la P^CO?. La PaCO2 elevadaque presentan los recien nacidos al nacer jugariaun rol muy importante en la primera respiracion.En la fig. 10 se observa que los recien nacidosresponden a la hipoxia con un aumento de la ven-tilation y que si a la hipoxia se agrega un 3% deCC>2 la hiperventilacion es aun mayor.
Finalmente observamos que en el recien naci-do la respuesta circulatoria a la hipoxia es dife-rente que en el adulto. En la fig. 11 se observaque la hipoxia moderada produce taquicardia; ade-mas disminucion del flujo sanguineo periferico,pero tambien moderada hipotension sistemica loque no se observa en el adulto. Por otro lado, laasfixia aguda produce vasoconstricci6n perifericae hipertension.
Equilibria dcido-bdsico del recien nacido y sitrelation con el ambiente termico
En el momento del nacimiento, la saturationde oxigeno arterial en la sangre umbilical varfa cn-
Fie. N? 10.— Efccto dc la adminis(raci6n de anhidrido carb6nicoc nipoxia, en 8 reci6n nacidos menores de 3 dias y 2 mayoresde una semana dc edad. N6tcse que el aumento de la venti-Iaci6n obscrvado al agregar 3% de CO2 al _airc inspirado (219&de O2) es similar en ambos grupos de ninos, en contrastc alpcqueiio aumento scguido de. dt-presi6n de la ventilacion quese demuestra en los menort's de 7 dias cuando respiran 12%de 02 y 3% de CO2, En estas uUimas condicioncs (hipoxia chipercapnia) los mayores de 13 dias responden con un aumen-to marcadi'simo de la ventilacion. (de Brady, J. y Ceruli, E. J.
Physiol. (London) 184: 631, 1966).
Fig. Nv 11.— C^mbios en la frecuencia cardfaca en 35 rceicn na-cidos (menurcs de 6 dias) al administrar difcrcntes concentra-ciones de O2 en el aire inspirado. N6tese el aumento signifi-cative dc la frecuencia cardiaca a medida que disminuye laPaO2. (de Brady, J. y Ceruti, E. J. Physiol. (London) 184: 631,
1966).
tre 0 y 70%, siendo el promedio de un 22%. Enun 25% de los recien nacidos normales la satu-racion arterial es menor de un 10%. En la fig. 12servan las variaciones de los gases arteriales y delequilibrio acido-b^sico que estudiamos en un grupode recien nacidos y prematures normales. Se de-muestra que la hipoxemia initial se acompana degrades variables de hipercapnia (PaCOa initial pro-medio: 58 mmHg), y de atidosis, (pH promedio:7,20). Estos hallazgos sugieren que en las etapasfinales del trabajo de parto y durante el parto mis-mo se produce una reduccion del intercambio deOa a traves de la placenta que lleva a grades va-riables de asfixia.
Inmediatamente despues del nacimiento se pro-duce una acentuacion de la acidosis que persistedurante varies minutos e incluso horas, a pesar dela mantencion de una temperatura ambiental ade-cuada, de la buena expansion pulmonar y de unarapida oxigenacion. La alteration de los gases ar-teriales y del pH en la sangre del cordon en elmomento del nacimiento son el resultado de unaalteration del intercambio placentario entre la ma-dre y el feto y no representan la adaptation delfeto a un periodo de hipoxia cronica. Mientrasmas tardio es el establecimiento de la respiracion,mayor es la disminucion de la P*O2 y mayor es laelevation dc la P,O2 y mayor es la elevacion de la
En experiencias rcalizadas con el Dr. Jorge To-rres en nuestro Departamento pudirnos comprobarque los recien nacidos mantenidos a temperaturasambientales inferiores a la neutral, es decir a lastemperaturas promedio de nuestras maternidades,rccuperan su balance acido-basico mucho mas tar-diamente que los ninos mantenidos a temperatura
515

740 .
xa 730
720
60
800
UJ
fM
OX
30
20
-14
-12
-8
0
95
85
75
65
55
T-—r i I i i i i i i t , • i i i T io o
J—I I i '• i i I • • • j i i i i i 1 i i i iT—i i I i i i i i i i i i i i
J 1 L
I I I I I I I I I I ) I I I I I 1 I I I
i I I I I I I I I 1 I 1 ' I I 1I I J I 1 I T I I I T i I I I I I T T
ic N° 12— Valores promedio de pH, prcsi6n parcial de CO2 arterial (PaCO2), presi6n parcial de oxigeno arterial (PaO2) yIxceso' de Base (EB) en un grupo dc 30 recten nacidos a t^r- mino y 30 prematures sanos (PTAEG) durante las pnmeras 72
Fig.
horas de vida.
516
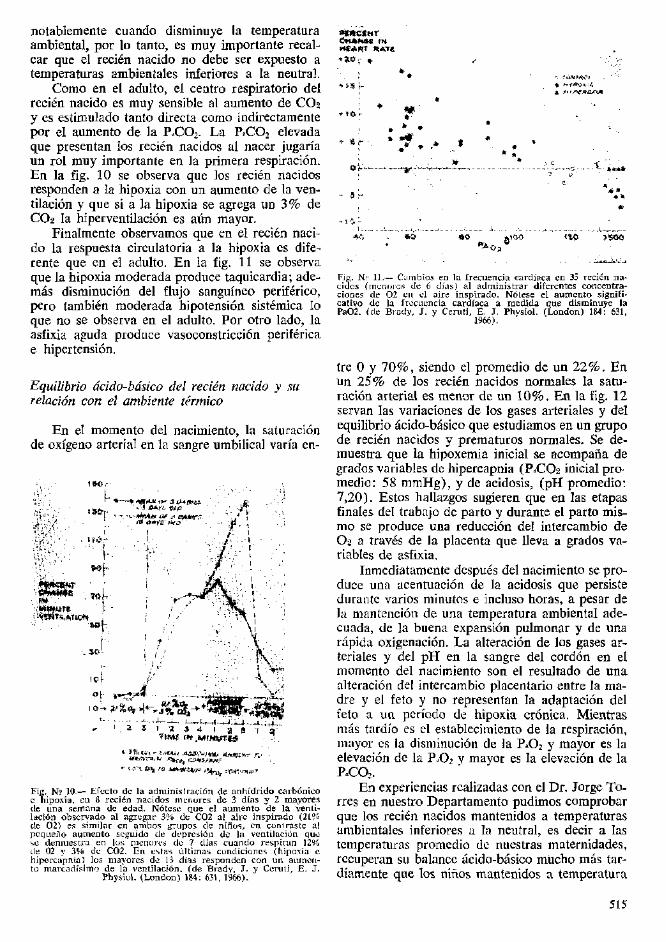
neutral (32 a 34°C). Hemos comprobado que elestablecimiento de condiciones acido-basicos nor-males en un lapso minimo de tiempo, depende deuna serie de factores, siendo los mas importantesla prematuridad, la medication materna y en granmedida la mantencion de los recien nacidos a unatemperatura ambiental optima, es decir, neutral,definiendo como temperatura ambiental neutralaquella en la cual el consumo de oxigeno es mi-nimo, vale decir, cuando el nino esta en condicio-nes metabolicas basales.
En la fig. 13 demostramos los valores prome-dio de pH, P.CO2 y Exceso dc Base (EB) en re-
cien nacidos a termino y prematures sanos com-parados con un grupo similar pero que se presen-taron asfixia al nacer o sindrome de dificultad res-piratoria. Ambos grupos de ninos eran mantenidosa la temperatura ambiental habitual de la mater-nidad (23 a 25<?C). En todos los grupos se ob-serva inicialmente un descenso del pH parcialmen-te compensado por hiperventilacion. Como en es-tos ninos se promediaron los valores encontrado?dentro de las primeras 6 horas de vida, no se al-canza a detectar la acidosis respiratoria inicial,ya que esta desaparece a la 2^ o 3a hora de vidapara ser reemplazada por una alcalosis respira-
Valores de pH,PacQ2. y Exceso de Base(promedlot error StarBandpromedio) en 39 recieh nacidos durante las primeras 72horas de vida -
• R N termino sanos• RN ttfrnmno I. R o---o Pr 1R
xa.
73
Horas de vida
74
73
72
71
45
40
35
30
25
20
-12
-10
-8
-6
-4
-2-0
12
Horas vida
Fig. N? 13.— Valores promedio de pH, PaCO2, y Exceso de base en 29 recie"n nacidos y prematures sanos y con insuficienciarespiratoria, durante las primeras 72 horas de vida, mantenidos en un arnbiente lermico inferior ai neutral (temperaturas am-bientales habitiiaJes de la Matcrnidad del Hospital J. J. A.). N6tese la scvera y mas prolongada acidosis metab61ica queprcsentan los prematures sanos en relaci6n a los rccicn nacidos a termino sanos y la aiin mas marcada y no compensada aci-dosis metabohca que dcmucstran ambos grupos con insuficiencia rcspiraroria. (de Ccruf i . E. y cols. Bol, M^d. Hosp. Mexico29: 757, 1972).
517

toria relativa (P*CO2 bajo). Como sc puede ob-servar, la alcalosis respiratoria sc combina conuna acidosis de tipo metabolico (deficit de basesignificative en todos los grupos). Sc demuestraademas, quo los prematures sanos prcsentan unaacidosis metabolica mas scvera y prolongada quelos recien nacidos a terrnino sanos y que en am-bos grupos la compensacion respiratoria ya escompleta a las 72 horas de vida. En la figura esposible observar ademas que en ambos grupos coninsuficiencia respiratoria, la acidosis metabolica esmas sevcra y prolongada quc en los ninos sanos yque no alcanza a ser compensada por hi respira-tion ni siquiera en cl 3er. dia de vida.
En la fig. 14 se demuestran los valores de lac-tacto, piruvato y exceso de lactato en los 4 gruposde ninos estudiados. Se destaca que los valoresiniciales de lactato y exceso de lactato son signifi-cativamente superiores en los prcmaturos sanosque en los recien nacidos a terrnino sanos pero, quede todas maneras, estan francamente mas elcvadosen ambos grupos de ninos con insuficiencia res-piratoria.
Con este estudio creemos poder demostrar ob-jetivamente que si los recien nacidos, particular-raente los ninos deprimidos al nacer o con sindro-me de dificultad respiratoria, se mantienen encondiciones termicas ambientales inferiores a laneutral, pierden calor rapidamente. Es decir, quctienen una muy limitada capacidad para mantenersu temperatura corporal, evidenciandose que enesta etapa de la vida, la production de calor esinsuficiente para equilibrar la perdida que se pro-duce como consecuencia de la alta conductanciatermica del recien nacido. La rapidez con que des-ciende la temperatura corporal, el nivel de equi-librio final y la distribution termica en los diferen-tes organos y tejidos dependen no solo de la tem-peratura y humedad ambientales sino de las gra-dientes de temperatura existentes entrc la pie! yel ambiente que rodea al recien nacido.
El recien nacido se comporta como un verda-dero homeotermo, cs decir, es capaz de aumentarel consume de O^ durantc el enfriamiento. Mien-tras mas pequeno es el recien nacido mayor esel consume de Oz a temperatura neutral y la tem-peratura critica tambien es mas alta y por supues-to, mayor sera el aumento de su metabolismocuando se enfrie. El recien nacido dispone de difc-rentes mecanismos para aumentar el consumo deO* y por lo tanto, la production de calor cuandocsta expuesto al frio: 1) prcsentan calofrios, quehan sido demostrados por algunos autores; otrosban determinado directamente un aumento del to-ne muscular y 2) presentan termogenesis quimica,no ligada a contracciones musculares, sino a unaumento de la produccion de calor en algunos te-jidos determinados, especialmente en la grasa par-da (± 15 grs.) la mayor parte de la cual esta si-
tuada en el cueilo, region interescapular, retroes-ternal y alrededor de los rinones. Se ha observadoque este tejido es influentiado por el frio y estu-dios post-mortem han demostradouna deplecion deli'pidos en la grasa parda de los recien nacidos ex-puesto al frio. La grasa parda cs ricamente vas-cularizada y tanto durante la expositi6n al friocomo despues de la infusion de catecolaminas (enel recien nacido se ha demostrado un aumento delas catecolaminas circulantes) el flujo sanguineoa traves de este tejido aumenta considerablemen-te. Asf, cl calor liberado per la exposition al friocs conducido por via sanguinea a proteger centresnerviosos vitales y tambien hacia la superficie cor-pora].
En nuestro estudio tambien comprobamos unaumento del consumo de oxigeno en todos losrecien nacidos estudiados, aun cuando e"ste es in-suficiente para mantener la temperatura corporalpor lo menos durante las primeras horas de vida.Esta incapacidad del recien nacido para equilibrarla perdida de calor estaria condicionada tanto porla cvaporacion de agua desde la superficie corpo-ral y los pulmones, como por la alta conductanciatermica del recien nacido que favorece la perdidade calor especialmente por irradiacion. Debe men-cionarse que en este estudio aun cuando los pre-matures se colocaron desde el nacimiento en in-cubadoras a temperaturas ambientales considera-das adecuadas. no lograron mantenerse a tempe-ratura neutral. Este hecho se podria explicar porlas perdidas permanentes de calor desde las pare-des de la pieza especialmente hacia las ventanas.
Los recien nacidos con asfixia o dificultad res-piratoria presentan una alteration de la termoge-nesis. Es sabido que la hipoxia e hipercapnia re-ducen la respuesta metabolica al frio y que la es-timulacion de los quimioreceptores perifericos quese produce justamente durante la hipoxia y la hi-percapnia puede llegar a abolir los calofrios, unade las respuestas a la exposicion al frio.
En estos ninos con insuficiencia respiratoriaapreciamos ademas una acidosis metabolica muchomas severa y prolongada que la presentada por losrecien nacidos normales. Tambien en todos ellosse demuestra una reduction dc la PaCO2 probable-mente como mecanismo compensatorio. Estos re-sultados sugieren que en los recien nacidos con in-suficiencia respiratoria se produciria una respuestarespiratoria y circulatoria inadecuada frente alstress que les rcprescnta el descenso de la tempera-tura ambiental, ya que en todos ellos la compen-sacion respiratoria fue incompleta todavia a las 72horas de vida. Es de suponer que en un reciennacido con alteraciones de la ventilation o de larelacion ventilaci<5n/perfusi6n, sea muy poco pro-bable alcanzar una adecuada compensacion respi-ratoria, Por otro lado, aun cuando la elimination
518
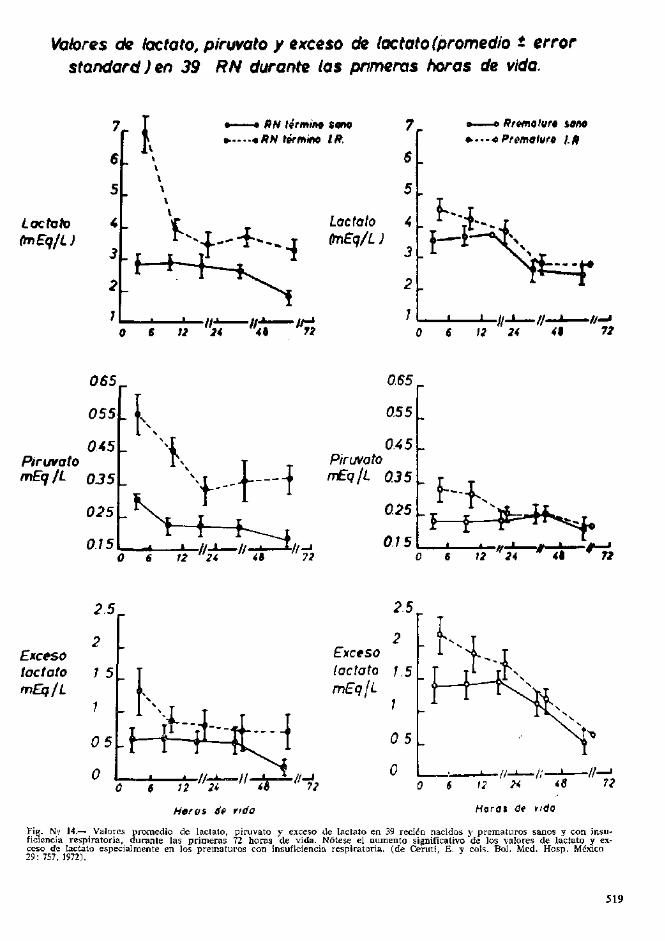
Vaiores cfe tactato, piruvato y exceso cte lactato (promedio t errorstandard)en 39 RN durante las pnmeras horas de vida.
Loctato
ffW flrmmo« fl W fermtfto
Lactato
> flremo/urd s0/>0'Premature I ft
065
055
045PiruvatomEq/L 035
025
015
r-
.1
- \-iS |
J
f II f
.1 1 //-I /
Piruvato
0.65
0.55
045
035
025
075<a 72 ;; 2*
Excesotoe to tomEq/L
25
0
nrfo
Fig. N? 14.— _ Valorcs proinedio dc lactato, piruvato y exceso de lactato en 39 reciiin nacidos y prematures sanos y con insu-ficiencia respiratoria, durante las primeras 72 haras de vida. N6tese el aiunento significativo de los valores de lactato y ex-ceso de lactato especialmente en IDS prematures con insuficiencia respiratoria. (de Certxti, E. y cols. Bol. Mcd. Hosp. Mexico29:757,1972).
519

de CO* por cl pulm6n sea adecuada, es dudosoque el debito cardiaco pueda aumentar lo sufi-ciente como para satisfacer la gran demanda poroxigeno que sc produce en condiciones de bajastemperaturas ambientales.
Conocidos los riesgos que implica la acidosis,parece necesario salvaguardar al recien nacido dedescensos excesivos del pH. En el recien nacidocon insuficiencia respiratoria, las necesidades me-tabolicas exceden con raucho la oferta de Os a lostejidos y al no existir una buena compensacionrespiratoria, la hipoxia y por consiguiente la aci-dosis se agravan aun mas.
Finalmente, creo del mdximo interns conside-rar en nuestro medio la incorporation a los cui-dados habituates del recien nacido, de la actionprotectora de temperaturas ambientales mas ele-vadas que las corrienteniente en uso, especialmen-te en los recien nacidos y prematures con asfixianeonatal o con sindrome de dificultad respiratoria.
REFERENCIAS
— Adams, F. H., Fugiwara, T., and Rowshan, G. Na-ture and origen of fluid in fetal lamb lung. J. Pediat.63; 881, 1963.
— Adamsons, K., Gandy, G. M., and James. L. S. In-fluence of thermal factors upon oxigen consumptionof the newborn infant, J. Pediat. 66: 495, 1965.
— Adamsons K. The role of thermal factors in fetaland neonatal life. Pediat. Clin. N. Amer. 13: 599,1966.
— Avcry, N. E., and Mead, J. Surface properties in re-lation fo atelectasia and hyaline membrane disease.Am. J, Dis. Child. 97: 517, 1969.
— Avery, M. E., Chernick, V., Dutton, R. E., and Per-mutt, S. Ventilatory response to inspired carbon dio-xide in infants and adults. J. Appl. Physiol. 18: 895,1963.
— Avery, M. E. The lung and its disorders in the new-born infant. VV. B. Saunders Company. Philadelphiaand London, 1964.
— Aherne W, and Dunnill M. S. Morphometry of thehuman placenta. Brit. M. Bull 22: 5, 1966.
— Aherne W. and Dunnill M. S. Cuantitative aspectsof placental structure. J. Path and Bact. 91: 123, 1966.
— Bartcls H., Moll W. and Metcaffe J. Phisiology of gasexchange in the humana placenta. Amer. J. Obstet.Gynec. 84: 1714, 1962.
-— Barcroft, Sir Joseph. Researches on Pre-natal Life,Vol. I. Spring field, III, Charles C. Thomas Cp.,1947.
Bertafanfy, F. D. and Leblond, D. P. Structure of res-piratory tissue. Lancet l .L: 1365, 1955.
Born, C. V. R., Dawes, G. S. MottJ, C., and Rcnnick,B, R. The constriction of tin ductus arteriosus causedby oxigen and by asphyxia in newborn lambs. J.Physiol. 132: 304, 1956.
Brady, J. P., and Toolcy, W. H. Cardiovascular andrespiratory reflexes in the newborn. Pediat. Clin. N.Amer. 13: 801, 1966.
Brack, K. Temperature regulation in the newborninfant. Biol. Nconat. 3: 65, 1961.
Burnard, E. D, A. murmur from the ductus in thenewborn baby. Brit. Med. J. 1: 806, 1958.
Burnard, E. D. The cardiac murmur in relation tosymptons in the newborn. Brit. Med. J. 1: 134,1959.
Campiche, iVl.t Cautler, A., Herndndez, Reymond, A.Electron microscope study of fetal development ofhuman lung. Pediatrics 32: 976, 1963.
Ceruti, E. Chemo receptor reflexes in the newborninfant. Effect of cooling in the response to hypoxia.Pediatrics 37: 556, 1966.
- Ceruti E., Torres T., Diaz A.t Auli G., Bailey P. yNunez N.— Efecto de la temperatura ambiental enla regulation del equilibrio acito base de R. N. determino y premature en los primeros dias de vida.Bol. Hosp. Inf. Mex. XXIX: 757, 1972.
- Clements /., Brown E., and Johnson R. Pulmonarysurface tension and the mucus lining of the lung. J.Appl. Physiol. 12, 262, 1958.
- Clements, J. A. Surface tension of lung extracts. Proc.Soc. Exptl. Biol. Med. 95: 170, 1957.
- Cotnro, A. H, Physiology of Respiration. ChicagoYear Book medical Publishers, Inc. 1965.
- Cook, C. D,, Cherry, R. B., O'Brien, D., Karlberg,P., and Smith, C. A. Studies of respiratory physiolo-gy in the newborn infant. I. Observations on normalpremature and full-term infants. J. Clin. Invest. 34:975, 1955.
- Cross, K, W,, and Warner, P. The effect of inhalationof high and low oxigen concentrations on the res-piration of the newborn infant J. Physiol. 114: 285,1951.
- Cross, K. W,, and Makom J. L. Evidence of carotidbody and sinus activity in newborn and foetal animals.J. Physiol. 118, 102, 1952.
520

Cross, K. W., and Oppe, T. E. The effect of inhala-tion of high and low concentrations of oxigen in therespiration of premature infants. J. Physiol. 117; 38,1952.
Dawcs, C. S., and Not!, L C. The vascular tone offoetal lung. J. Physiol. 164: 465, 1962.
Dawes, G. S., Mott, J. C,, Shelley, H, ]., and Stafford,A. Prolongation of survial time in asphyxiated inma-tufoetal lamba, J. Physiol. 168, 43, 1963.
Eldridge, P. L,, ffultgren, H. N., and Wigmore, M.E. Physiologic closure of ductus arteriosus in new-born infants. J. Clin. Invest.; 34: 987, 1955.
Fawcltt, }., Lind, J., and Wegelius, C. The first breath.Acta Paediat. (Suppl. 123) 49: 5, 1960.
Gandy, G. M.y Adamsons, K., Cunningham, N., Sil-vermann, W. A, and James, L. S. Thermal environmentand acid base homeostasis in human infants during thefirst few hours of life. J. Clin. Invest. 43: 751, 1964.
Gluck L., Motoyama E., Smits and KuJcovich M.The biochemical development of surface activity inmammalian lung I. The surface active phosplolipids,the separation and distribution of surface active leci-thin in the lung of developing rabbitt fetus. Pediat.Res. 1: 237, 1967.
• Gluck L., Sribney M. and Kulovich M. The bioche-mical development of surface activity in mammalianlung II. The biosynthesis of phospholipids in the lungof the developing rabbitt fetus and newborn Pediat.Res. 1: 247, 1967.
Gluck L, Landowne R. and Kulovich M. The bioche-mical develepment of surface activity in mammalianlung. III. Structural changes in lung lecithin duringdevelopment of the rabbitt fetus and newborn. Pediat.Res. 4: 312, 1970.
Gluck L., Kulovich M., Borer R., Brenner P., Ander-son G., and Spellacey W, Diagnosis of the respiratorydistress syndrome by amniocentesis. Amer. J. Obstetr.Gynec. 109, 440, 1971.
Gluck L., Kulovich M., Eidelman A., Cordero L.and Khazin A. The biochemical development of sur-face activity in mammalian lung. IV. Pulmonary le-cithin synthesis in the human fetus and newborn andthe etiology of the respiratory distress syndrome.Ped. Res. 6: 00, 1972.
Gould J., Gluck L., Kulovich M. The aceleration ofneurological maturation in high stress pregnancy andits relation to fetal lung maturity. Pediat. Res. 6:335, 1972.
— Cluck L. Surfactant 1972. Ped. Clin. N. A. 19:325, 1972.
Heehaber L, Loescher, H. H., and Westphal, F. Ei-ne Virkung des Progesterons and die Atmung. Arch.ges. Physiol. 250: 42, 1948.
James, L. S. Acidosis of newborn and its relation tobirth asphyxia. Acta Paediat. 49 (Suppl. 122): 17,1960.
James, L. S., and Rowe, R. D. The pattern of respon-se of pulmonary and systemic arterial pressures innewborn and older infants to short periods of hy-poxia. 1. Pediat. 51: 5, 1957.
James, L. S., and Adamsons, K. Respiratory physio-logy of the fetus and newborn infant. The N. Engl.I. Med. 271: 1352, 1964.
James, L. S. Onset of breathing and resuscitation.Pediat. Clin. N. Amer. 13: 621, 1966.
Karlberg, P. Physiology of prematurity. Trans. 2ndMacy Conference Lanman, I. T., ed.. The losiahMacy Ir. Foundation. New York, 1957.
Karlberg, P. Adaptive changes in immediate postnatalperiod, with particular reference to respiration. I.Pediat. 56: 585, 1960.
Karlberg, P., Charry, R. B., Escardo, F. E., and Koch,G. Respiratory studies in newborn infants, II. Pulmo-nary ventilation and mechanics of breathing in thefirst minutes of life, including onset of respiration.Acta. Paediat. 51: 121, 1962.
Klaus, M., Reiss, O. K., Tooloy, W. H., Weaver, K.H., and Clements. J. ^.Lung volumes in the newborninfant. Pediatrics 30: 111, 1962.
Lind, J., and Wegelius, C. Human fetal circulation:changes in the cardiovascular system at birth anddisturbances in the post-natal closure of the fora-men ovale and ductus arteriosus. Cold. Spr. Harb.Symp. Quant. Bid. 19: 309, 1954.
Loosli, C. G., and Potter, E. L. Pre-and post-nataldevelopment of the respiratory portion of the humanlung. Am. Rev. Resp. Dis. 80: 5, 1959.
Low, F. N. Pulmonary alveolar epithalium of Labo-ratory mammals and man. Anat. Rec. 117: 261, 1953.
Low, F. N., and Sampaiox, M. The pulmonary alveo-lars epithelium as an entodermal derivative. Anat.Rec. 127: 51, 1957.
Mitchell, S. Ductus artcriorus in the neonatal period.J. Pediat. 51: 12, 1957.
521
3.— Pediatria.

Mos, A. J. Emmanouilides, G., and Duffie, E. R.Closure of the ductus arteriosus in the newborn in-fant. Pediatrics 52: 25, 1963.
Mead J. Mechanical properties of lungs. Physiol. Rev.41: 281, 1961.
Mead J. Mechanical properties of lungs. Physiol. Rev.41: 281, 1961.
Nelson, N. M., Prod'hom, L. S., Cherry, R. B.r Lipsitz,P. J., and Smith, C. A. Pulmonary function in thenewborn infant II. Perfusion-Estimation by analysisof arterial-alveolar carbon dioxide difference. Pe-diatrics 30: 975, 1962.
Nelson, N. M., Prod'hom, L. S., Cherry, R. B., Lipsitz,P. J., and Smith, C. A. Pulmonary function in thenewborn infant.; Alveolar-arterial oxigen gradient. J.Appl. Physiol. 18: 534, 1963.
Netchman C. and Huisman T. Comparative studies ofoxygen equilibrie of human adult and blood red cellshemolyzates and suspensions. Clin. Chim Acta 10:165, 1964.
Nelson, N. M. Neonatal pulmonary function. Pediat.Clin. N. Amcr. 15: 769, 1966.
• Oliver, T. K., Davis, J. A., and Bates, C. D. Serialblood gas tensions and acid-base balance during thefirst hour of life in human infants. Acta. PaediatScand. 50: 346, 1961.
• Oliver, T. K., and Karlberg, P. Gaseous metabolismin newlyborn human infants. Amer. J. Dis. Child.105: 427, 1963.
- PaUle, R. E. Properties, function, and origin of alveo-lar lining layer. Nature (London) 175: 1125, 1955.
- Prod'hom, L. S., Levison, H., Cherry, R. B., Dror-baugh, J. E., Hubbell, J, P., and Smith, C. A. Ad-
justment of ventilation, intrupulmonary gas exchan-ge and acid-base balance during the first day of life.Pediatrics 33: 682, 1964.
Radford E. Method for estimating respiratory sur-face area of mammalian lungs from their physicalcharacteristics. Proc. Soc. Exper. Bio. S. Med. 87:58, 1954.
Ramsey E. Circulation in the intervillous space ofthe primate placenta. Amer. J. Obstet Gynec. 84:1649, 1962.
Ross, B. B. Comparison of boetal pulmonary fluidwith foetal plasma and ammiotic fluid. Nature (Lon-don) 199: 1100, 1963.
Rowe, R. D., and James, L. S. The normal pulmonaryarterial pressure during the first year of life. J. Pe-diat. 51: 1, 1957.
Setnikar, L, Agostoni, E., and Taglietti, A. The fe-tal lung, source of amniotic fluid. Proc. Soc. Exper.Biol. Med. 101: 842, 1959.
Silverman, W. A. Diagnosis and treatment: Use andmisuse of temperature and humidity in care of thenewborn infant. Pediatrics 34: 276, 1964.
• Strang, L. B. Alveolar and anatomical dead-spacein normal newborn infants studied by air washing oxi-gen breathing. Clin. Sci. 23: 129, 1962.
• Tooley, W. H,, Klaus, M., Weaver, K. H., and Cle-ments, J. A. The distribution of ventilation in normalnewborn infants. Am. J. Dis Child. 100: 731, 1960(Abstract).
• Weisbrot, I. M., James, L. S., Prince. C. E., Hola-day, D. A., and Apgar, V. Acid-base homeostasis ofthe newborn infant during the first twenty-four hoursof life. J. Pediat. 52: 395, 1958.
522