Embed Size (px)
Citation preview

NeuroImage 59 (2012) 238–247
Contents lists available at ScienceDirect
NeuroImage
j ourna l homepage: www.e lsev ie r.com/ locate /yn img
Cortico-striatal disconnection within the cingulo-opercular network in schizophreniarevealed by intrinsic functional connectivity analysis: A resting fMRI study
Pei-Chi Tu a,b,c, Jen-Chuen Hsieh d,e, Cheng-Ta Li c,d, Ya-Mai Bai c,f, Tung-Ping Su b,c,f,⁎a Department of Medical Research and Education, Taipei Veterans General Hospital, Taipei 112, Taiwanb Institute of Neuroscience, School of Life Science, National Yang-Ming University, Taipei 112, Taiwanc Department of Psychiatry, Taipei Veterans General Hospital, Taipei 112, Taiwand Institute of Brain Science, National Yang-Ming University, Taipei, Taiwane Laboratory of Integrated Brain Research, Department of Medical Research and Education, Taipei Veterans General Hospital, Taiwanf Department of Psychiatry, Faculty of Medicine, National Yang-Ming University, Taipei, Taiwan
⁎ Corresponding author at: Office of the Vice SuperinteHospital, No. 201, Sec. 2, Shih-Pai Road, Taipei 112, Taiwa
E-mail address: [email protected] (T.-P. Su).
1053-8119/$ – see front matter © 2011 Elsevier Inc. Aldoi:10.1016/j.neuroimage.2011.07.086
a b s t r a c t
a r t i c l e i n f oArticle history:Received 20 April 2011Revised 22 June 2011Accepted 28 July 2011Available online 3 August 2011
Keywords:Cingulo-opercular networkSchizophreniaFunctional connectivityCortico-striatal disconnection
The cingulo-opercular network (CON) is a newly defined control network responsible for various cognitiveprocesses that have been consistently found to be impaired in schizophrenia. The aim of this study was to usefunctional connectivity magnetic resonance imaging (fcMRI) to test the hypothesis that schizophrenia isassociated with functional disconnection within the CON. Thirty subjects with schizophrenia and thirtyhealthy controls were enrolled in the study. Each subject received resting fMRI scanning, clinical evaluationsand cognitive examinations. The CON of each subject was derived by calculating the functional connectivitymap of a seed in the dorsal anterior cingulate (dACC). A between-group comparison was performed using arandom effect analysis. Further network analyses with multiple regions of interest (ROIs) were performed tocharacterize the pattern of functional disconnection within the entire CON. Using the dACC seed in healthycontrols, we derived the CON, which includes the following anatomical structures: the dACC; the bilateralanterior prefrontal, inferior parietal and anterior insular cortices; the putamen; the thalamus; and thecerebellum. Compared with healthy controls, schizophrenic patients showed significantly reduced functionalconnectivity in the bilateral putamens. Further network analysis demonstrated widespread cortico-striataldisconnection within the CON of schizophrenic patients. The disconnections correlated with negativesymptom severity. Behavioral regression revealed that cortico-striatal functional connectivity predicted 2-back working memory performance in healthy controls, but not in schizophrenic patients. Our findingssuggest that schizophrenia is associated with cortical–striatal disconnection within the CON. The resultprovides a network basis for the cortico-striatal disconnection hypothesis of schizophrenia.
ndent, Taipei Veterans Generaln. Fax: +886 2 28757555.
l rights reserved.
© 2011 Elsevier Inc. All rights reserved.
Introduction
Schizophrenia is a devastating psychiatric disorder accompanied byauditory hallucination, delusion and cognitive impairment. The under-lying etiology is largely unknown, but it is thought to involve someaberration of neural circuits during the neural developmental process(Andreasen et al., 1998; Pettersson-Yeo et al., 2011). Despite thesehypotheses, there has been no direct method to evaluate the functionalintegrity of these circuits in living human subjects until recently.Developments in resting-state functional connectivity MRI (fcMRI)(Biswal et al., 1995; Greicius et al., 2003) provide an excellent tool toevaluate the functional integrity of neural circuits and thus allowsthe testing of several disconnection hypotheses for schizophrenia(Pettersson-Yeo et al., 2011). Recent studies by means of the fcMRI
technique found that the brain is organized into numerous large-scalefunctional networks, and the regions within the same functionalnetwork show coherent, low-frequency (b0.1 Hz) fluctuations duringthe resting state, as measured by blood-oxygen-level-dependent(BOLD) imaging signals. Functional connectivity is defined by thetemporal correlation of these low-frequencyfluctuations betweenbrainregions, and the increase or decrease of functional connectivity isthought to play an important role in various neuropsychiatric disorders.Greicius et al. (2004) were the first to show that functional connectivitywithin the default mode network (DMN) is disrupted in patients withAlzheimer's disease (AD) when compared to healthy controls. Connec-tivity disruptions were subsequently reported for a number ofneuropsychiatric disorders including autism (Kennedy and Courchesne,2008), attention deficit hyperactivity disorder (Castellanos et al., 2008),depression (Greicius et al., 2007), and schizophrenia (Bluhmet al., 2007;Garrity et al., 2007;Whitfield-Gabrieli et al., 2009; Zhouet al., 2007) (seeGreicius, 2008 for a review). Two recent fcMRI studies from the samegroup (Seeley et al., 2009; Zhou et al., 2010) showed that distinct types

239P.-C. Tu et al. / NeuroImage 59 (2012) 238–247
of dementia, including Alzheimer's dementia (AD) and frontotemporaldementia, map to the degeneration of distinct networks defined byfcMRI. These studies provide evidence that different neuropsychiatricdisorders may be caused by specific patterns (hyper- or hypo-connectivity) in specific large-scale networks. If true, then understand-ing the nature of network dysfunction in specific illnesses will providean excellent endophenotype for further genetic imaging studies.
Among the various networks defined by fcMRI, the DMN ishypothesized to play an important role in the pathogenesis ofschizophrenic disorders. The DMN includes several regions in theposterior cingulate cortex (PCC) and the medial prefrontal cortex(mPFC)and bilateral inferior parietal regions that were commonlyfound to have decreased activities during various goal-directed tasks(Shulman et al., 1997). Raichle et al. (2001) demonstrated that thesebrain regions are in their baseline statewhen subjects restwith their eyesclosed. A regional decrease represented the presence of ongoingfunctionality in the resting state and that functionality was attenuatedonly when resources were temporarily reallocated during goal-directedbehaviors. Because schizophrenia is characterized by symptoms ofdelusion and hallucinations that are proposed to be associated withdisturbances in inner awareness, it was hypothesized that schizophreniamaybe causedby thedysfunctionof theDMN.However, previous studieson this topic produced inconsistent findings. Some studies (Bluhm et al.,2007; Camchong et al., 2011) found decreased functional connectivity,whereas others (Zhouet al., 2007; Liu et al., in press;Whitfield-Gabrieli etal., 2009) found either increased functional connectivitywithin the DMNor no difference (Repovs et al., 2011). The exact cause of theseinconsistencies remains unclear, but it has been observed that patientsin the early phase of the illness show increased functional connectivitywithin the DMN, whereas those with a longer-duration illness showeddecreased functional connectivity. This finding raised the possibility thatDMN connectivitymay depend on the stage of the illness andmay not bea core feature of schizophrenia. Compared with the DMN, functionalconnectivity in other important large-scale functional networks is rarelyexplored, and it is less clear whether other disruptions of functionalconnectivity exist.
Abnormal prefrontal cortical function during cognitive tasks is themost robust physiological feature of schizophrenia (Broomeet al., 2009;Glahn et al., 2005; Lewis and Anderson, 1995), providing hints that thenetwork signature of schizophrenia may lay in networks important forexecutive control. Several important large-scale networks involved inattention and executive control were identified following the concep-tual proof of the DMN and include the dorsal attention network (Fox etal., 2005), the cingulo-opercular network (CON) (Dosenbach et al.,2007; Seeley et al., 2007) and the fronto-parietal network (FPN)(Dosenbach et al., 2007; Seeley et al., 2007). Among these, the CON is aparticularly important candidate network for schizophrenia. The CONincludes the anatomical structures of the dACC, the anterior insula/operculum, the anterior prefrontal cortex, the inferior parietal cortex,the basal ganglion, the thalamus and the cerebellum and was firstidentified in a meta-analysis (Dosenbach et al., 2006) of fMRI studiesinvolving control-demanding tasks. Later, a functional connectivityanalysis (Dosenbach et al., 2007) confirmed this finding. The authors ofthis study proposed that the brain regions exhibiting control signalsacross many tasks are grouped into two distinct “fronto-parietal” and“cingulo-opercular” networks, the primary roles of the dual networkarchitecture of top-down control (Dosenbach et al., 2007, 2008). Theauthors also suggested that theCONoperates on a longer timescale thanthe FPN and provides “set initiation”, stable “set-maintenance” and“error monitoring” for the duration of task blocks.
Previous structural and functional imaging studies have providedpreliminary evidence that the CON may play an important role inschizophrenia. Two recent meta-analyses of VBM studies (Glahn et al.,2008; Fornito et al., 2009) consistently identified the insula and thedACC as the two hubs of the CON with the highest ranking associationswith the structural deficits of schizophrenia. A recent VBM study
(Palaniyappan et al., 2010) showed that structural deficits within theCON were also found to correlate with reality distortion in schizophre-nia. Furthermore, fMRI studies using control-demanding tasks such asworking memory (Glahn et al., 2005; Becerril et al., 2011) andantisaccade (Polli et al., 2005; Tu et al., 2006) tasks consistently foundblunted activation of the network. Finally, two recent functionalconnectivity analyses (Tu et al., 2010; White et al., 2010) during task-based fMRI studies identified aberrant functional connectivity of theCON in schizophrenia. Previously, we identified functional disconnec-tion with right dACC in schizophrenia during an antisaccade task (Tu etal., 2010). Thesefindings suggested that theCONplays an important rolein the pathogenesis of schizophrenia. One resting state fcMRI study(Repovs et al., 2011) investigated the CON in schizophrenic patient, butdid not find any significant abnormalities. However, this study onlycompared the averaged functional connectivity within the CON and itwas possible that some region specific disconnections were neglectedduring the averaging process. A detailed intrinsic functional connectiv-ity analysis of this network will be helpful to clarify the issue.
The aim of the present study was to examine the functionalconnectivity of CON in schizophrenia using resting-state fMRI. Basedon our previous functional connectivity studies using task-based fMRI,we hypothesized that schizophrenia would be associated with reducedfunctional connectivity within the CON. We used a seed in the dACCbased on the coordinates of a previous meta-analysis (Dosenbach et al.,2006) of fMRI studies to derive the CON in schizophrenic patients andhealthy controls and to perform a between-group comparison in abrain-wise manner. Next, we undertook a ROI-based approach usingmultiple hub regions within the network to further delineate thefunctional connectivity of the core regions of these networks. Finally, abehavioral regression of working memory performance was performedto further delineate the functional role of the CON in schizophrenia andhealthy.
Materials and method
Subjects
The schizophrenic sample group was composed of thirty out-patients and in-patients from the Taipei Veterans General Hospital inTaiwan (Table 1). Diagnoses were confirmed by structured clinicalinterviews for DSM-IV (First et al., 1997) and evaluated using the MiniInternational Neuropsychiatric Inventory Plus (M.I.N.I.) (Sheehan et al.,1998). Participants were screened to exclude those with substanceabuse or dependencewithin the preceding sixmonths, a history of headinjury resulting in a sustained loss of consciousness and/or cognitivesequelae, neurological illness, or any disorder affecting cerebralmetabolism. With the exception of one unmedicated patient, thesepatients had been using a variety of atypical antipsychotic medicationsprior to participation in the experiment, including paliperidone ER(N=14), aripiprazole (Abilify) (N=9), seroquel (N=2), haldol(N=2), clozapine (N=1) and lodopine (N=1). Thirty age-, gender-,and handedness-matched healthy controls were recruited by adver-tisement. These participants were screened by an experiencedpsychiatrist using MINI to exclude subjects with major psychiatricillnesses. Additionally, those candidateswith a family history of an axis Idisorder in first-degree relatives, including schizophrenia, majordepression or bipolar disorder, were excluded. The clinical status ofschizophrenic patients was characterized using the Positive andNegative Syndrome Scale (PANSS) (Kay et al., 1987), and the cognitiveperformance of each participant was evaluated by 1-back and 2-backworking memory tasks. The tasks were chosen because schizophreniapatients have been consistently shown to have deficits in workingmemory tasks, and the CON has been found to be consistently activatedby the task. All procedures were approved by the institutional reviewboard of the Taipei Veterans General Hospital, and all participants
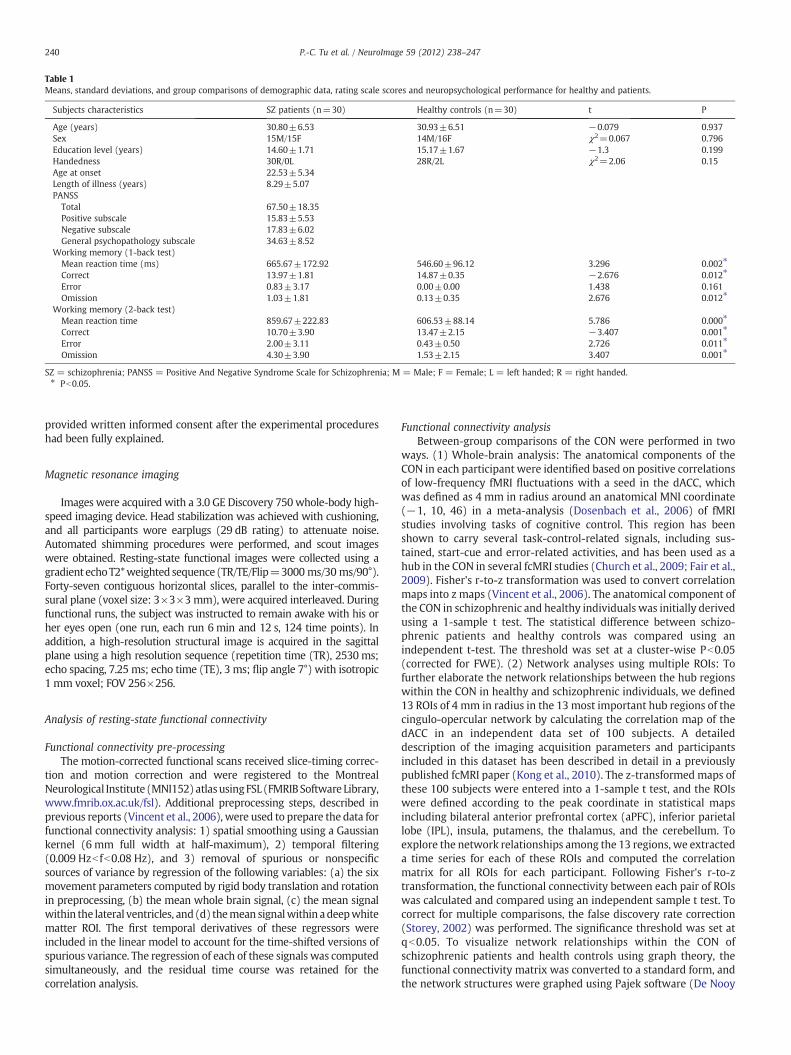
Table 1Means, standard deviations, and group comparisons of demographic data, rating scale scores and neuropsychological performance for healthy and patients.
Subjects characteristics SZ patients (n=30) Healthy controls (n=30) t P
Age (years) 30.80±6.53 30.93±6.51 −0.079 0.937Sex 15M/15F 14M/16F χ2=0.067 0.796Education level (years) 14.60±1.71 15.17±1.67 −1.3 0.199Handedness 30R/0L 28R/2L χ2=2.06 0.15Age at onset 22.53±5.34Length of illness (years) 8.29±5.07PANSS
Total 67.50±18.35Positive subscale 15.83±5.53Negative subscale 17.83±6.02General psychopathology subscale 34.63±8.52
Working memory (1-back test)Mean reaction time (ms) 665.67±172.92 546.60±96.12 3.296 0.002⁎
Correct 13.97±1.81 14.87±0.35 −2.676 0.012⁎
Error 0.83±3.17 0.00±0.00 1.438 0.161Omission 1.03±1.81 0.13±0.35 2.676 0.012⁎
Working memory (2-back test)Mean reaction time 859.67±222.83 606.53±88.14 5.786 0.000⁎
Correct 10.70±3.90 13.47±2.15 −3.407 0.001⁎
Error 2.00±3.11 0.43±0.50 2.726 0.011⁎
Omission 4.30±3.90 1.53±2.15 3.407 0.001⁎
SZ = schizophrenia; PANSS = Positive And Negative Syndrome Scale for Schizophrenia; M = Male; F = Female; L = left handed; R = right handed.⁎ Pb0.05.
240 P.-C. Tu et al. / NeuroImage 59 (2012) 238–247
provided written informed consent after the experimental procedureshad been fully explained.
Magnetic resonance imaging
Images were acquired with a 3.0 GE Discovery 750whole-body high-speed imaging device. Head stabilization was achieved with cushioning,and all participants wore earplugs (29 dB rating) to attenuate noise.Automated shimming procedures were performed, and scout imageswere obtained. Resting-state functional images were collected using agradient echoT2*weighted sequence (TR/TE/Flip=3000ms/30 ms/90°).Forty-seven contiguous horizontal slices, parallel to the inter-commis-sural plane (voxel size: 3×3×3mm), were acquired interleaved. Duringfunctional runs, the subject was instructed to remain awake with his orher eyes open (one run, each run 6 min and 12 s, 124 time points). Inaddition, a high-resolution structural image is acquired in the sagittalplane using a high resolution sequence (repetition time (TR), 2530 ms;echo spacing, 7.25 ms; echo time (TE), 3 ms; flip angle 7°) with isotropic1 mm voxel; FOV 256×256.
Analysis of resting-state functional connectivity
Functional connectivity pre-processingThe motion-corrected functional scans received slice-timing correc-
tion and motion correction and were registered to the MontrealNeurological Institute (MNI152) atlasusingFSL (FMRIBSoftware Library,www.fmrib.ox.ac.uk/fsl). Additional preprocessing steps, described inprevious reports (Vincent et al., 2006), were used to prepare the data forfunctional connectivity analysis: 1) spatial smoothing using a Gaussiankernel (6 mm full width at half-maximum), 2) temporal filtering(0.009 Hzb fb0.08 Hz), and 3) removal of spurious or nonspecificsources of variance by regression of the following variables: (a) the sixmovement parameters computed by rigid body translation and rotationin preprocessing, (b) the mean whole brain signal, (c) the mean signalwithin the lateral ventricles, and(d) themean signalwithin adeepwhitematter ROI. The first temporal derivatives of these regressors wereincluded in the linear model to account for the time-shifted versions ofspurious variance. The regression of each of these signals was computedsimultaneously, and the residual time course was retained for thecorrelation analysis.
Functional connectivity analysisBetween-group comparisons of the CON were performed in two
ways. (1) Whole-brain analysis: The anatomical components of theCON in each participant were identified based on positive correlationsof low-frequency fMRI fluctuations with a seed in the dACC, whichwas defined as 4 mm in radius around an anatomical MNI coordinate(−1, 10, 46) in a meta-analysis (Dosenbach et al., 2006) of fMRIstudies involving tasks of cognitive control. This region has beenshown to carry several task-control-related signals, including sus-tained, start-cue and error-related activities, and has been used as ahub in the CON in several fcMRI studies (Church et al., 2009; Fair et al.,2009). Fisher's r-to-z transformation was used to convert correlationmaps into z maps (Vincent et al., 2006). The anatomical component ofthe CON in schizophrenic and healthy individuals was initially derivedusing a 1-sample t test. The statistical difference between schizo-phrenic patients and healthy controls was compared using anindependent t-test. The threshold was set at a cluster-wise Pb0.05(corrected for FWE). (2) Network analyses using multiple ROIs: Tofurther elaborate the network relationships between the hub regionswithin the CON in healthy and schizophrenic individuals, we defined13 ROIs of 4 mm in radius in the 13 most important hub regions of thecingulo-opercular network by calculating the correlation map of thedACC in an independent data set of 100 subjects. A detaileddescription of the imaging acquisition parameters and participantsincluded in this dataset has been described in detail in a previouslypublished fcMRI paper (Kong et al., 2010). The z-transformed maps ofthese 100 subjects were entered into a 1-sample t test, and the ROIswere defined according to the peak coordinate in statistical mapsincluding bilateral anterior prefrontal cortex (aPFC), inferior parietallobe (IPL), insula, putamens, the thalamus, and the cerebellum. Toexplore the network relationships among the 13 regions, we extracteda time series for each of these ROIs and computed the correlationmatrix for all ROIs for each participant. Following Fisher's r-to-ztransformation, the functional connectivity between each pair of ROIswas calculated and compared using an independent sample t test. Tocorrect for multiple comparisons, the false discovery rate correction(Storey, 2002) was performed. The significance threshold was set atqb0.05. To visualize network relationships within the CON ofschizophrenic patients and health controls using graph theory, thefunctional connectivity matrix was converted to a standard form, andthe network structures were graphed using Pajek software (De Nooy

Fig. 1. The dACC seed used to derive the CON in this study.
241P.-C. Tu et al. / NeuroImage 59 (2012) 238–247
et al., 2005). The relationships between seeds were graphed using theKamada–Kawai graphing algorithm (Kamada and Kawai, 1989) basedon energy minimization of the networks.
Control analysisTo further confirm the specificity of our findings, we chose the
primary sensorimotor network for control analysis. In this analysis,the motor network was derived for each participant by calculating thefunctional connectivity map of a seed in the left primarymotor cortex,defined as 4 mm in radius according to the anatomical landmark inthe atlas, and between-group comparisons were performed byindependent sample t tests with the threshold set at a cluster-wisePb0.05 (corrected for FWE).
Regression analyses of functional connectivity on working memoryperformance and clinical variables
Behavioral correlation analyses were performed to understand thefunctional consequence of the functional connectivity differences wefound. The functional connectivity values were calculated for each pairof ROIs that showed a significant difference between groups and wereused to perform a regression analysis on the performance of 1-back and2-back tests as measured by the response latency and correction rate.The association between functional connectivity and cognitive perfor-mance, adjusting both variables for age, was investigated usingmultipleregression analyses. For these analyses, functional connectivity valueswere the dependent variables, and cognitive performance and agewerecovariates. An interaction term (performance by group)was included inthemodel to testwhether the slopeof the relationshipdifferedby group.If a significant correlation in the combined groupwas found, regressionswere computed for each group separately. The correlative relationshipwas considered to be significant at Pb0.05 (uncorrected for multiplecomparisons). One regression model was constructed per seed-pair percognitive variable in the regression analysis.
The correlation between functional connectivity and clinical variablesin schizophrenia was performed using the partial correlation methodwith age as a covariate of no interest. The correlative relationship wasconsidered to be significant at Pb0.05 (uncorrected for multiplecomparisons).
Results
Demographic, clinical and neuropsychological characteristics
There were no significant differences in age, gender or educationlevel between healthy controls and the patients with schizophrenia(Table 1). Compared to healthy controls, the schizophrenic patientshad significantly worse performances in 1-back and 2-back workingmemory tests as characterized by a longer response time and fewercorrect responses (Table 1).
Brain-wise analysis
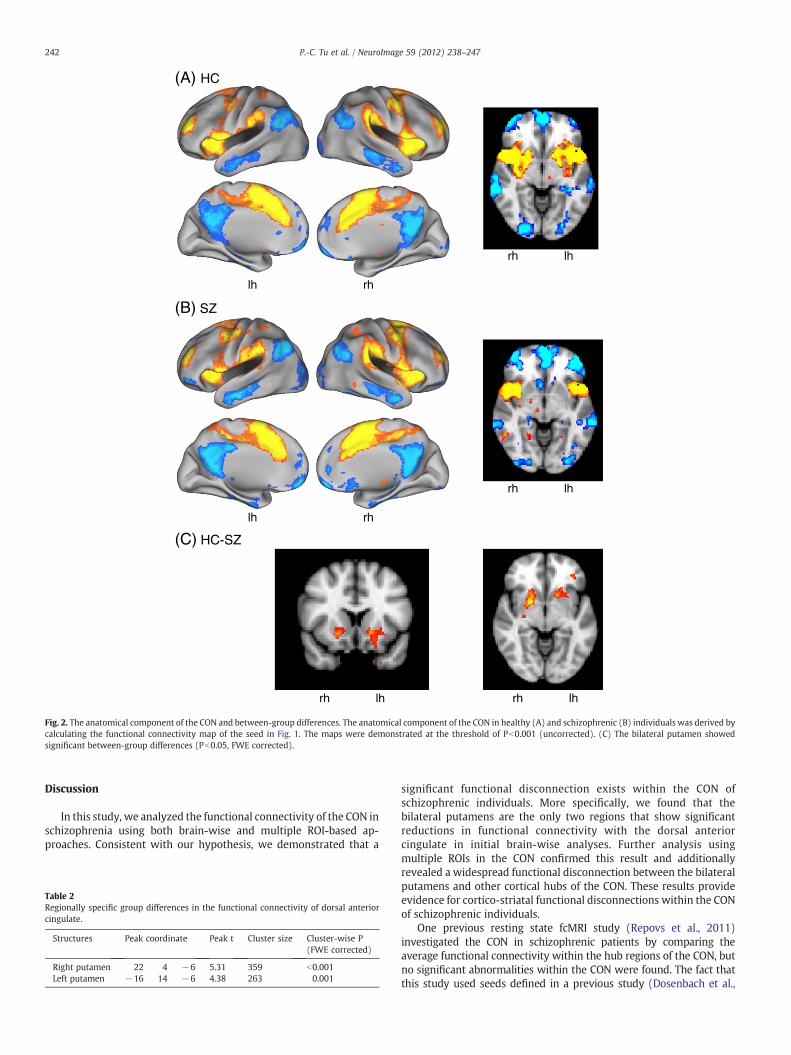
Using the dACC as the seed, we robustly derived the cingulo-opercular network in both healthy and schizophrenic individuals. Theanatomical structures were largely consistent with previous fMRI orfunctional connectivity studies and included the bilateral aPFC, dACC,anterior insula/operculum, IPL, rostral putamen, lateral thalamus andcerebellum (Fig. 1). Schizophrenic patients showed a similar patternof network distribution in the cortical areas. Between-group compar-isons revealed only two regions that survive the threshold of Pb0.05(FWE correction): the left and right putamen (Fig. 2, Table 2). Theyshowed significant lower functional connectivity in schizophrenicpatients. There was no region that showed increased functionalconnectivity to the dACC seed at the chosen threshold.
We also used a seed in the left motor cortex to perform a controlanalysis and did not find any region showing a significant differencebetween healthy and schizophrenic patients.
ROI-based analysis
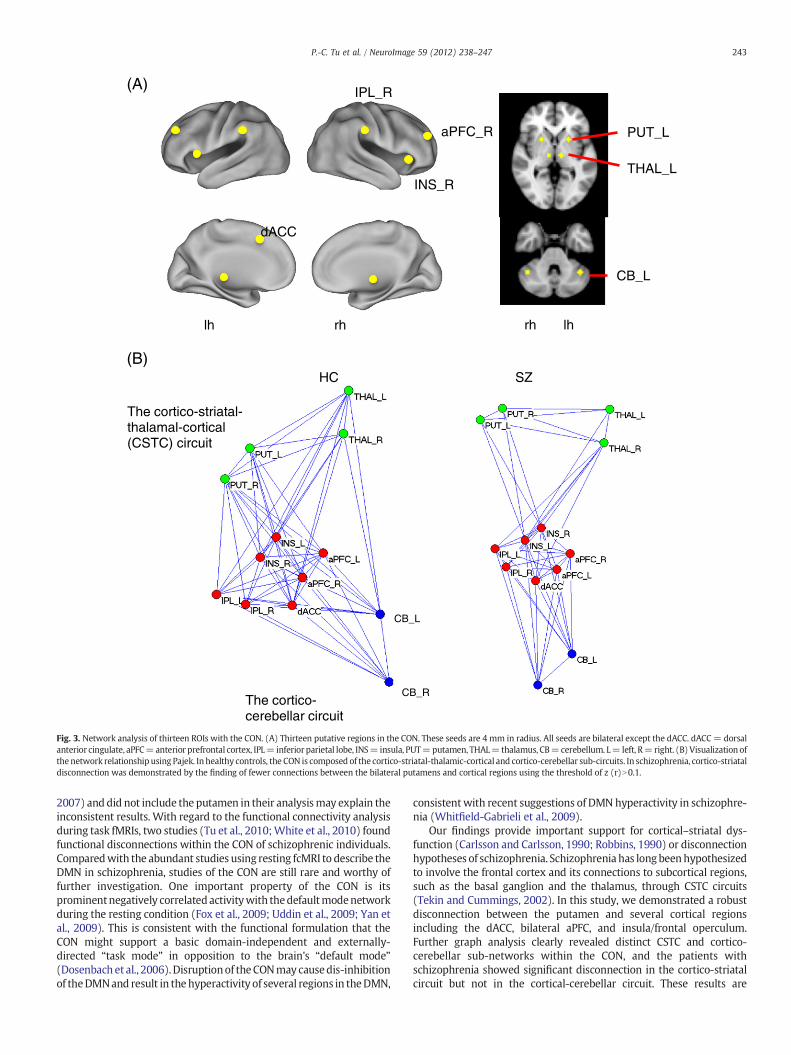
Thirteen ROIs, defined according to the functional connectivity mapof the dACC seed, were selected for analysis (Fig. 3), and the detailedMNI coordinates are described in Supplementary Table 1. Among thesefunctional connectivity values, 15 of the 16 pairs showing a significantbetween-group difference included the left or right putamen (Pb0.05,FDR corrected; Table 3). The ROIs showing decreased functionalconnectivity with the left putamen included the dACC, bilateral aPFC,bilateral insula, and left IPL. The ROIs showing significantly reducedfunctional connectivity with the right putamen included the dACC,bilateral aPFC, bilateral insula, bilateral IPL and left cerebellum(Table 4).The results revealed a pattern of prominent cortico-striatal disconnectwithin the CON of schizophrenic individuals.
To better visualize the network relationships of the CON in HC andSZ, we performed a graph analysis and showed that the CON wascomposed of a core cortical network interconnected with cortico-striatal-thalamic-cortical (CSTC) and cortico-cerebellar sub-circuits(Fig. 4). In the schizophrenic patients, prominent cortico-striataldisconnections were demonstrated by fewer cortico-striatal connec-tions within the CON using a significance threshold of z(r)N0.1. Thethresholds were set strictly for visualization purposes, and allfunctional connectivities showing significant statistical differencesrefer to the results of Tables 3 and 4.
Regression analysis of cortico-striatal connection on working memoryperformance and clinical variables
Regression analysis revealed that greater functional connectivityin two of the cortico-striatal circuits, the dACC/right putamen and theright IPL/right putamen, was predictive of more efficient workingmemory performance, as demonstrated by shorter 2-back latencies(Table 5 and Fig. 4). These relations differed significantly by group,reflecting the finding that controls showed strong and significantrelations (Pb0.05, uncorrected for multiple comparisons). However,the result did not survive the Bonferroni correction for multiplecomparisons and should be cautiously interpreted.
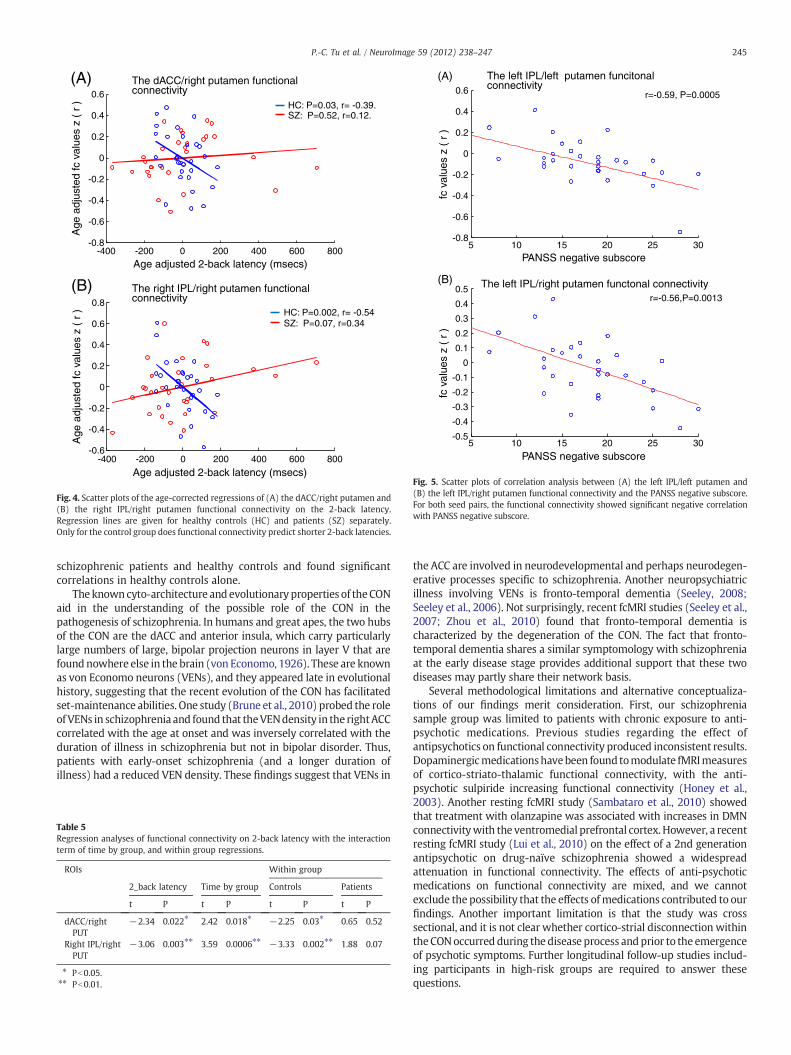
We also found a significant negative correlation between cortico-striatal connectivity and negative symptom severity. The lowerfunctional connectivity between two pairs of the ROIs: the leftputamen/left IPL (r=−0.59, P=0.0005) and the right putamen/leftIPL (r=−0.56, P=0.0013) predicted higher PANSS negative subscore(Fig. 5). The correlationwas still significant after correction for the effectof age using partial correlation approach, and correction for multiplecomparison by the Bonferroni method. There is no significantcorrelation between cortico-striatal connectivity and duration of illnessor other clinical variables.
(A) HC
(B) SZ
(C) HC-SZ
lh rh
rh lh
lh rh
rh lh
rh lh rh lh
Fig. 2. The anatomical component of the CON and between-group differences. The anatomical component of the CON in healthy (A) and schizophrenic (B) individuals was derived bycalculating the functional connectivity map of the seed in Fig. 1. The maps were demonstrated at the threshold of Pb0.001 (uncorrected). (C) The bilateral putamen showedsignificant between-group differences (Pb0.05, FWE corrected).
242 P.-C. Tu et al. / NeuroImage 59 (2012) 238–247
Discussion
In this study, we analyzed the functional connectivity of the CON inschizophrenia using both brain-wise and multiple ROI-based ap-proaches. Consistent with our hypothesis, we demonstrated that a
Table 2Regionally specific group differences in the functional connectivity of dorsal anteriorcingulate.
Structures Peak coordinate Peak t Cluster size Cluster-wise P(FWE corrected)
Right putamen 22 4 −6 5.31 359 b0.001Left putamen −16 14 −6 4.38 263 0.001
significant functional disconnection exists within the CON ofschizophrenic individuals. More specifically, we found that thebilateral putamens are the only two regions that show significantreductions in functional connectivity with the dorsal anteriorcingulate in initial brain-wise analyses. Further analysis usingmultiple ROIs in the CON confirmed this result and additionallyrevealed a widespread functional disconnection between the bilateralputamens and other cortical hubs of the CON. These results provideevidence for cortico-striatal functional disconnections within the CONof schizophrenic individuals.
One previous resting state fcMRI study (Repovs et al., 2011)investigated the CON in schizophrenic patients by comparing theaverage functional connectivity within the hub regions of the CON, butno significant abnormalities within the CON were found. The fact thatthis study used seeds defined in a previous study (Dosenbach et al.,
HC SZ
lh rh rh lh
(A)
(B)
aPFC_R
INS_R
IPL_R
PUT_L
THAL_L
CB_L
dACC
The cortico-striatal-thalamal-cortical (CSTC) circuit
The cortico-cerebellar circuit
CB_L
CB_R
Fig. 3. Network analysis of thirteen ROIs with the CON. (A) Thirteen putative regions in the CON. These seeds are 4 mm in radius. All seeds are bilateral except the dACC. dACC= dorsalanterior cingulate, aPFC=anterior prefrontal cortex, IPL= inferior parietal lobe, INS= insula, PUT=putamen, THAL=thalamus, CB=cerebellum. L= left, R=right. (B)Visualization ofthenetwork relationship using Pajek. Inhealthy controls, the CON is composed of the cortico-striatal-thalamic-cortical and cortico-cerebellar sub-circuits. In schizophrenia, cortico-striataldisconnection was demonstrated by the finding of fewer connections between the bilateral putamens and cortical regions using the threshold of z (r)N0.1.
243P.-C. Tu et al. / NeuroImage 59 (2012) 238–247
2007) and did not include the putamen in their analysismay explain theinconsistent results. With regard to the functional connectivity analysisduring task fMRIs, two studies (Tu et al., 2010;White et al., 2010) foundfunctional disconnections within the CON of schizophrenic individuals.Comparedwith the abundant studies using resting fcMRI to describe theDMN in schizophrenia, studies of the CON are still rare and worthy offurther investigation. One important property of the CON is itsprominentnegatively correlatedactivitywith thedefaultmodenetworkduring the resting condition (Fox et al., 2009; Uddin et al., 2009; Yan etal., 2009). This is consistent with the functional formulation that theCON might support a basic domain-independent and externally-directed “task mode” in opposition to the brain's “default mode”(Dosenbach et al., 2006).Disruptionof theCONmay causedis-inhibitionof theDMNand result in thehyperactivity of several regions in theDMN,
consistentwith recent suggestions of DMN hyperactivity in schizophre-nia (Whitfield-Gabrieli et al., 2009).
Our findings provide important support for cortical–striatal dys-function (Carlsson and Carlsson, 1990; Robbins, 1990) or disconnectionhypotheses of schizophrenia. Schizophrenia has longbeenhypothesizedto involve the frontal cortex and its connections to subcortical regions,such as the basal ganglion and the thalamus, through CSTC circuits(Tekin and Cummings, 2002). In this study, we demonstrated a robustdisconnection between the putamen and several cortical regionsincluding the dACC, bilateral aPFC, and insula/frontal operculum.Further graph analysis clearly revealed distinct CSTC and cortico-cerebellar sub-networks within the CON, and the patients withschizophrenia showed significant disconnection in the cortico-striatalcircuit but not in the cortical-cerebellar circuit. These results are
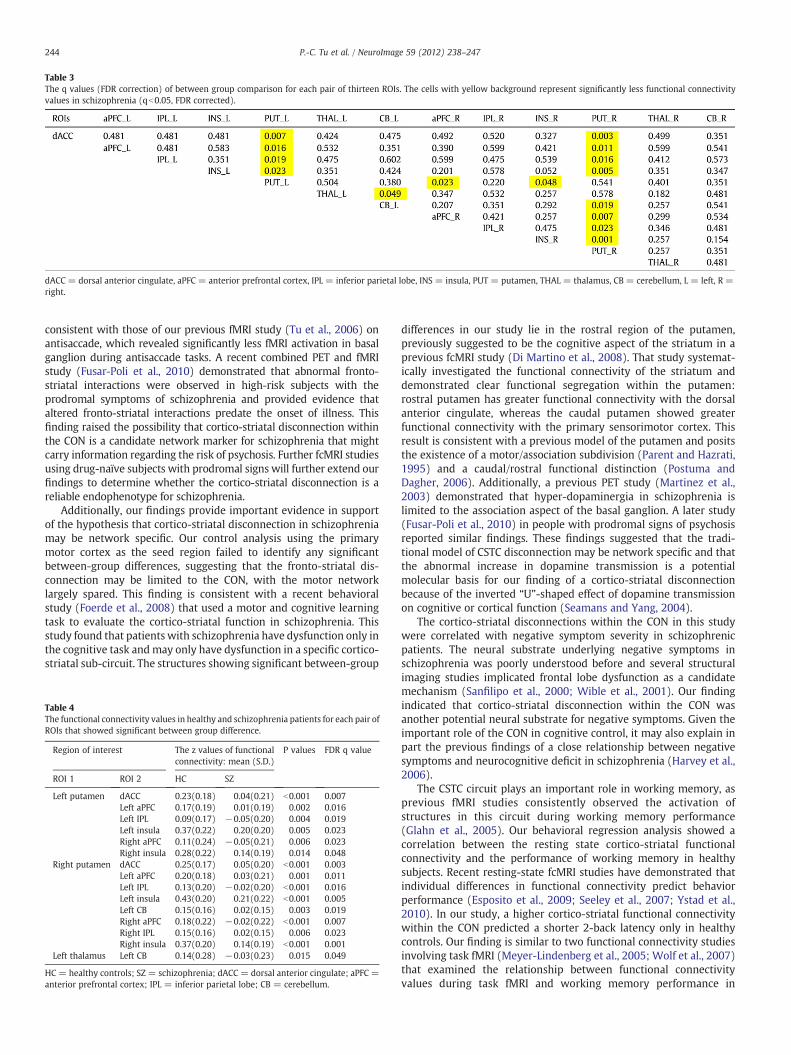
Table 3The q values (FDR correction) of between group comparison for each pair of thirteen ROIs. The cells with yellow background represent significantly less functional connectivityvalues in schizophrenia (qb0.05, FDR corrected).
dACC = dorsal anterior cingulate, aPFC = anterior prefrontal cortex, IPL = inferior parietal lobe, INS = insula, PUT = putamen, THAL = thalamus, CB = cerebellum, L = left, R =right.
244 P.-C. Tu et al. / NeuroImage 59 (2012) 238–247
consistent with those of our previous fMRI study (Tu et al., 2006) onantisaccade, which revealed significantly less fMRI activation in basalganglion during antisaccade tasks. A recent combined PET and fMRIstudy (Fusar-Poli et al., 2010) demonstrated that abnormal fronto-striatal interactions were observed in high-risk subjects with theprodromal symptoms of schizophrenia and provided evidence thataltered fronto-striatal interactions predate the onset of illness. Thisfinding raised the possibility that cortico-striatal disconnection withinthe CON is a candidate network marker for schizophrenia that mightcarry information regarding the risk of psychosis. Further fcMRI studiesusing drug-naïve subjects with prodromal signs will further extend ourfindings to determine whether the cortico-striatal disconnection is areliable endophenotype for schizophrenia.
Additionally, our findings provide important evidence in supportof the hypothesis that cortico-striatal disconnection in schizophreniamay be network specific. Our control analysis using the primarymotor cortex as the seed region failed to identify any significantbetween-group differences, suggesting that the fronto-striatal dis-connection may be limited to the CON, with the motor networklargely spared. This finding is consistent with a recent behavioralstudy (Foerde et al., 2008) that used a motor and cognitive learningtask to evaluate the cortico-striatal function in schizophrenia. Thisstudy found that patients with schizophrenia have dysfunction only inthe cognitive task andmay only have dysfunction in a specific cortico-striatal sub-circuit. The structures showing significant between-group
Table 4The functional connectivity values in healthy and schizophrenia patients for each pair ofROIs that showed significant between group difference.
Region of interest The z values of functionalconnectivity: mean (S.D.)
P values FDR q value
ROI 1 ROI 2 HC SZ
Left putamen dACC 0.23(0.18) 0.04(0.21) b0.001 0.007Left aPFC 0.17(0.19) 0.01(0.19) 0.002 0.016Left IPL 0.09(0.17) −0.05(0.20) 0.004 0.019Left insula 0.37(0.22) 0.20(0.20) 0.005 0.023Right aPFC 0.11(0.24) −0.05(0.21) 0.006 0.023Right insula 0.28(0.22) 0.14(0.19) 0.014 0.048
Right putamen dACC 0.25(0.17) 0.05(0.20) b0.001 0.003Left aPFC 0.20(0.18) 0.03(0.21) 0.001 0.011Left IPL 0.13(0.20) −0.02(0.20) b0.001 0.016Left insula 0.43(0.20) 0.21(0.22) b0.001 0.005Left CB 0.15(0.16) 0.02(0.15) 0.003 0.019Right aPFC 0.18(0.22) −0.02(0.22) b0.001 0.007Right IPL 0.15(0.16) 0.02(0.15) 0.006 0.023Right insula 0.37(0.20) 0.14(0.19) b0.001 0.001
Left thalamus Left CB 0.14(0.28) −0.03(0.23) 0.015 0.049
HC = healthy controls; SZ = schizophrenia; dACC = dorsal anterior cingulate; aPFC =anterior prefrontal cortex; IPL = inferior parietal lobe; CB = cerebellum.
differences in our study lie in the rostral region of the putamen,previously suggested to be the cognitive aspect of the striatum in aprevious fcMRI study (Di Martino et al., 2008). That study systemat-ically investigated the functional connectivity of the striatum anddemonstrated clear functional segregation within the putamen:rostral putamen has greater functional connectivity with the dorsalanterior cingulate, whereas the caudal putamen showed greaterfunctional connectivity with the primary sensorimotor cortex. Thisresult is consistent with a previous model of the putamen and positsthe existence of a motor/association subdivision (Parent and Hazrati,1995) and a caudal/rostral functional distinction (Postuma andDagher, 2006). Additionally, a previous PET study (Martinez et al.,2003) demonstrated that hyper-dopaminergia in schizophrenia islimited to the association aspect of the basal ganglion. A later study(Fusar-Poli et al., 2010) in people with prodromal signs of psychosisreported similar findings. These findings suggested that the tradi-tional model of CSTC disconnection may be network specific and thatthe abnormal increase in dopamine transmission is a potentialmolecular basis for our finding of a cortico-striatal disconnectionbecause of the inverted “U”-shaped effect of dopamine transmissionon cognitive or cortical function (Seamans and Yang, 2004).
The cortico-striatal disconnections within the CON in this studywere correlated with negative symptom severity in schizophrenicpatients. The neural substrate underlying negative symptoms inschizophrenia was poorly understood before and several structuralimaging studies implicated frontal lobe dysfunction as a candidatemechanism (Sanfilipo et al., 2000; Wible et al., 2001). Our findingindicated that cortico-striatal disconnection within the CON wasanother potential neural substrate for negative symptoms. Given theimportant role of the CON in cognitive control, it may also explain inpart the previous findings of a close relationship between negativesymptoms and neurocognitive deficit in schizophrenia (Harvey et al.,2006).
The CSTC circuit plays an important role in working memory, asprevious fMRI studies consistently observed the activation ofstructures in this circuit during working memory performance(Glahn et al., 2005). Our behavioral regression analysis showed acorrelation between the resting state cortico-striatal functionalconnectivity and the performance of working memory in healthysubjects. Recent resting-state fcMRI studies have demonstrated thatindividual differences in functional connectivity predict behaviorperformance (Esposito et al., 2009; Seeley et al., 2007; Ystad et al.,2010). In our study, a higher cortico-striatal functional connectivitywithin the CON predicted a shorter 2-back latency only in healthycontrols. Our finding is similar to two functional connectivity studiesinvolving task fMRI (Meyer-Lindenberg et al., 2005; Wolf et al., 2007)that examined the relationship between functional connectivityvalues during task fMRI and working memory performance in
r=-0.59, P=0.0005
r=-0.56,P=0.0013
fc v
alue
s z
( r
)fc
val
ues
z (
r )
PANSS negative subscore
PANSS negative subscore
The left IPL/left putamen funcitonal connectivity
The left IPL/right putamen functonal connectivity
(A)
(B)
0.6
0.4
0.2
0
-0.2
-0.4
-0.6
-0.8
0.5
0.4
0.3
0.2
0.1
0
-0.1
-0.2
-0.3
-0.5
-0.4
5 10 15 20 25 30
5 10 15 20 25 30
Fig. 5. Scatter plots of correlation analysis between (A) the left IPL/left putamen and(B) the left IPL/right putamen functional connectivity and the PANSS negative subscore.For both seed pairs, the functional connectivity showed significant negative correlationwith PANSS negative subscore.
-400 -200 0 200 400 600 800-0.6
-0.4
-0.2
0
0.2
0.4
0.6
0.8
-400 -200 0 200 400 600 800-0.8
-0.6
-0.4
-0.2
0
0.2
0.4
0.6HC: P=0.03, r= -0.39.SZ: P=0.52, r=0.12.
HC: P=0.002, r= -0.54SZ: P=0.07, r=0.34
(A)A
ge a
djus
ted
fc v
alue
s z
( r
)A
ge a
djus
ted
fc v
alue
s z
( r
)
Age adjusted 2-back latency (msecs)
Age adjusted 2-back latency (msecs)
The right IPL/right putamen functional connectivity
The dACC/right putamen functional connectivity
(B)
Fig. 4. Scatter plots of the age-corrected regressions of (A) the dACC/right putamen and(B) the right IPL/right putamen functional connectivity on the 2-back latency.Regression lines are given for healthy controls (HC) and patients (SZ) separately.Only for the control group does functional connectivity predict shorter 2-back latencies.
245P.-C. Tu et al. / NeuroImage 59 (2012) 238–247
schizophrenic patients and healthy controls and found significantcorrelations in healthy controls alone.
The knowncyto-architecture andevolutionaryproperties of theCONaid in the understanding of the possible role of the CON in thepathogenesis of schizophrenia. In humans and great apes, the two hubsof the CON are the dACC and anterior insula, which carry particularlylarge numbers of large, bipolar projection neurons in layer V that arefoundnowhere else in the brain (von Economo, 1926). These are knownas von Economo neurons (VENs), and they appeared late in evolutionalhistory, suggesting that the recent evolution of the CON has facilitatedset-maintenance abilities. One study (Brune et al., 2010) probed the roleof VENs in schizophrenia and found that theVENdensity in the rightACCcorrelated with the age at onset and was inversely correlated with theduration of illness in schizophrenia but not in bipolar disorder. Thus,patients with early-onset schizophrenia (and a longer duration ofillness) had a reduced VEN density. These findings suggest that VENs in
Table 5Regression analyses of functional connectivity on 2-back latency with the interactionterm of time by group, and within group regressions.
ROIs Within group
2_back latency Time by group Controls Patients
t P t P t P t P
dACC/rightPUT
−2.34 0.022⁎ 2.42 0.018⁎ −2.25 0.03⁎ 0.65 0.52
Right IPL/rightPUT
−3.06 0.003⁎⁎ 3.59 0.0006⁎⁎ −3.33 0.002⁎⁎ 1.88 0.07
⁎ Pb0.05.⁎⁎ Pb0.01.
the ACC are involved in neurodevelopmental and perhaps neurodegen-erative processes specific to schizophrenia. Another neuropsychiatricillness involving VENs is fronto-temporal dementia (Seeley, 2008;Seeley et al., 2006). Not surprisingly, recent fcMRI studies (Seeley et al.,2007; Zhou et al., 2010) found that fronto-temporal dementia ischaracterized by the degeneration of the CON. The fact that fronto-temporal dementia shares a similar symptomology with schizophreniaat the early disease stage provides additional support that these twodiseases may partly share their network basis.
Several methodological limitations and alternative conceptualiza-tions of our findings merit consideration. First, our schizophreniasample group was limited to patients with chronic exposure to anti-psychotic medications. Previous studies regarding the effect ofantipsychotics on functional connectivity produced inconsistent results.Dopaminergicmedicationshavebeen found tomodulate fMRImeasuresof cortico-striato-thalamic functional connectivity, with the anti-psychotic sulpiride increasing functional connectivity (Honey et al.,2003). Another resting fcMRI study (Sambataro et al., 2010) showedthat treatment with olanzapine was associated with increases in DMNconnectivitywith the ventromedial prefrontal cortex. However, a recentresting fcMRI study (Lui et al., 2010) on the effect of a 2nd generationantipsychotic on drug-naïve schizophrenia showed a widespreadattenuation in functional connectivity. The effects of anti-psychoticmedications on functional connectivity are mixed, and we cannotexclude the possibility that the effects ofmedications contributed to ourfindings. Another important limitation is that the study was crosssectional, and it is not clear whether cortico-strial disconnection withintheCONoccurred during the disease process andprior to the emergenceof psychotic symptoms. Further longitudinal follow-up studies includ-ing participants in high-risk groups are required to answer thesequestions.

246 P.-C. Tu et al. / NeuroImage 59 (2012) 238–247
Conclusions
In conclusion, our findings indicate that schizophrenia is associatedwith a functional disconnectionwithin the cortico-striatal circuits of theCON. The disconnections correlate with the severity of negativesymptoms. Further investigations of the CON in high-risk groups arerequired to establish the possible network signature of schizophrenia.
Supplementarymaterials related to this article can be found onlineat doi:10.1016/j.neuroimage.2011.07.086.
Acknowledgment
The study was supported by grants from Taipei Veterans GeneralHospital (V99C1-040) and National Science Council (NSC99-2628-B-010-021-MY2).
References
Andreasen, N.C., Paradiso, S., O'Leary, D.S., 1998. “Cognitive dysmetria” as an integrativetheory of schizophrenia: a dysfunction in cortical–subcortical–cerebellar circuitry?Schizophr. Bull. 24, 203–218.
Becerril, K.E., Repovs, G., Barch, D.M., 2011. Error processing network dynamics inschizophrenia. Neuroimage 54, 1495–1505.
Biswal, B., Yetkin, F.Z., Haughton, V.M., Hyde, J.S., 1995. Functional connectivity in themotor cortex of resting human brain using echo-planar MRI. Magn. Reson. Med. 34,537–541.
Bluhm, R.L., Miller, J., Lanius, R.A., Osuch, E.A., Boksman, K., Neufeld, R.W., Theberge, J.,Schaefer, B., Williamson, P., 2007. Spontaneous low-frequency fluctuations in theBOLD signal in schizophrenic patients: anomalies in the default network.Schizophr. Bull. 33, 1004–1012.
Broome, M.R., Matthiasson, P., Fusar-Poli, P., Woolley, J.B., Johns, L.C., Tabraham, P.,Bramon, E., Valmaggia, L., Williams, S.C., Brammer, M.J., Chitnis, X., McGuire, P.K.,2009. Neural correlates of executive function and working memory in the ‘at-riskmental state’. Br. J. Psychiatry 194, 25–33.
Brune, M., Schobel, A., Karau, R., Benali, A., Faustmann, P.M., Juckel, G., Petrasch-Parwez,E., 2010. Von Economo neuron density in the anterior cingulate cortex is reduced inearly onset schizophrenia. Acta Neuropathol. 119, 771–778.
Camchong, J., Macdonald 3rd, A.W., Bell, C., Mueller, B.A., Lim, K.O., 2011. Alteredfunctional and anatomical connectivity in schizophrenia. Schizophr. Bull. 37,640–650.
Carlsson, M., Carlsson, A., 1990. Schizophrenia: a subcortical neurotransmitterimbalance syndrome? Schizophr. Bull. 16, 425–432.
Castellanos, F.X., Margulies, D.S., Kelly, C., Uddin, L.Q., Ghaffari, M., Kirsch, A., Shaw, D.,Shehzad, Z., Di Martino, A., Biswal, B., Sonuga-Barke, E.J., Rotrosen, J., Adler, L.A.,Milham, M.P., 2008. Cingulate–precuneus interactions: a new locus of dysfunctionin adult attention-deficit/hyperactivity disorder. Biol. Psychiatry 63, 332–337.
Church, J.A., Fair, D.A., Dosenbach, N.U., Cohen, A.L., Miezin, F.M., Petersen, S.E.,Schlaggar, B.L., 2009. Control networks in paediatric Tourette syndrome showimmature and anomalous patterns of functional connectivity. Brain 132, 225–238.
De Nooy, W., Mrvar, A., Batageli, V., 2005. Exploratory Network Analysis with Pajek.Cambridge UP, New York.
DiMartino, A., Scheres, A., Margulies, D.S., Kelly, A.M., Uddin, L.Q., Shehzad, Z., Biswal, B.,Walters, J.R., Castellanos, F.X., Milham, M.P., 2008. Functional connectivity ofhuman striatum: a resting state FMRI study. Cereb. Cortex 18, 2735–2747.
Dosenbach, N.U., Visscher, K.M., Palmer, E.D., Miezin, F.M., Wenger, K.K., Kang, H.C.,Burgund, E.D., Grimes, A.L., Schlaggar, B.L., Petersen, S.E., 2006. A core system forthe implementation of task sets. Neuron 50, 799–812.
Dosenbach, N.U., Fair, D.A., Miezin, F.M., Cohen, A.L., Wenger, K.K., Dosenbach, R.A., Fox,M.D., Snyder, A.Z., Vincent, J.L., Raichle, M.E., Schlaggar, B.L., Petersen, S.E., 2007.Distinct brain networks for adaptive and stable task control in humans. Proc. Natl.Acad. Sci. U.S.A. 104, 11073–11078.
Dosenbach, N.U., Fair, D.A., Cohen, A.L., Schlaggar, B.L., Petersen, S.E., 2008. A dual-networks architecture of top-down control. Trends Cogn. Sci. 12, 99–105.
Esposito, F., Aragri, A., Latorre, V., Popolizio, T., Scarabino, T., Cirillo, S., Marciano, E.,Tedeschi, G., Di Salle, F., 2009. Does the default-mode functional connectivity of thebrain correlate with working-memory performances? Arch. Ital. Biol. 147, 11–20.
Fair, D.A., Cohen, A.L., Power, J.D., Dosenbach, N.U., Church, J.A., Miezin, F.M., Schlaggar,B.L., Petersen, S.E., 2009. Functional brain networks develop from a “local todistributed” organization. PLoS Comput. Biol. 5, e1000381.
First, M., Spitzer, R., Gibbon, M., Williams, J., 1997. Structured Clinical Interview forDSM-IV Axis I Disorders, Research Version, Patient Edition with Psychotic Screen(SCID-I/P W/PSY SCREEN). Biometrics Research, New York State PsychiatricInstitute, New York.
Foerde, K., Poldrack, R.A., Khan, B.J., Sabb, F.W., Bookheimer, S.Y., Bilder, R.M., Guthrie,D., Granholm, E., Nuechterlein, K.H., Marder, S.R., Asarnow, R.F., 2008. Selectivecorticostriatal dysfunction in schizophrenia: examination of motor and cognitiveskill learning. Neuropsychology 22, 100–109.
Fornito, A., Yucel, M., Dean, B., Wood, S.J., Pantelis, C., 2009. Anatomical abnormalities ofthe anterior cingulate cortex in schizophrenia: bridging the gap betweenneuroimaging and neuropathology. Schizophr. Bull. 35, 973–993.
Fox, M.D., Snyder, A.Z., Vincent, J.L., Corbetta, M., Van Essen, D.C., Raichle, M.E., 2005.The human brain is intrinsically organized into dynamic, anticorrelated functionalnetworks. Proc. Natl. Acad. Sci. U.S.A. 102, 9673–9678.
Fox, M.D., Zhang, D., Snyder, A.Z., Raichle, M.E., 2009. The global signal and observedanticorrelated resting state brain networks. J. Neurophysiol. 101, 3270–3283.
Fusar-Poli, P., Howes, O.D., Allen, P., Broome, M., Valli, I., Asselin, M.C., Grasby, P.M.,McGuire, P.K., 2010. Abnormal frontostriatal interactions in people with prodromalsigns of psychosis: a multimodal imaging study. Arch. Gen. Psychiatry 67, 683–691.
Garrity, A.G., Pearlson, G.D.,McKiernan, K., Lloyd, D., Kiehl, K.A., Calhoun, V.D., 2007. Aberrant“defaultmode” functional connectivity in schizophrenia. Am. J. Psychiatry 164, 450–457.
Glahn, D.C., Ragland, J.D., Abramoff, A., Barrett, J., Laird, A.R., Bearden, C.E., Velligan, D.I.,2005. Beyond hypofrontality: a quantitative meta-analysis of functional neuroim-aging studies of working memory in schizophrenia. Hum. Brain Mapp. 25, 60–69.
Glahn, D.C., Laird, A.R., Ellison-Wright, I., Thelen, S.M., Robinson, J.L., Lancaster, J.L.,Bullmore, E., Fox, P.T., 2008. Meta-analysis of gray matter anomalies inschizophrenia: application of anatomic likelihood estimation and network analysis.Biol. Psychiatry 64, 774–781.
Greicius, M., 2008. Resting-state functional connectivity in neuropsychiatric disorders.Curr. Opin. Neurol. 21, 424–430.
Greicius, M.D., Krasnow, B., Reiss, A.L., Menon, V., 2003. Functional connectivity in theresting brain: a network analysis of the default mode hypothesis. Proc. Natl. Acad.Sci. U.S.A. 100, 253–258.
Greicius, M.D., Srivastava, G., Reiss, A.L., Menon, V., 2004. Default-mode networkactivity distinguishes Alzheimer's disease from healthy aging: evidence fromfunctional MRI. Proc. Natl. Acad. Sci. U.S.A. 101, 4637–4642.
Greicius, M.D., Flores, B.H., Menon, V., Glover, G.H., Solvason, H.B., Kenna, H., Reiss, A.L.,Schatzberg, A.F., 2007. Resting-state functional connectivity in major depression:abnormally increased contributions from subgenual cingulate cortex and thalamus.Biol. Psychiatry 62, 429–437.
Harvey, P.D., Koren, D., Reichenberg, A., Bowie, C.R., 2006. Negative symptoms and cognitivedeficits: what is the nature of their relationship? Schizophr. Bull. 32, 250–258.
Honey, G.D., Suckling, J., Zelaya, F., Long, C., Routledge, C., Jackson, S., Ng, V., Fletcher,P.C., Williams, S.C., Brown, J., Bullmore, E.T., 2003. Dopaminergic drug effects onphysiological connectivity in a human cortico-striato-thalamic system. Brain126, 1767–1781.
Kamada, K., Kawai, S., 1989. An algorithm for drawing general undirected graphs. Inf.Process. Lett. 31, 7–15.
Kay, S.R., Fiszbein, A., Opler, L.A., 1987. The positive and negative syndrome scale(PANSS) for schizophrenia. Schizophr. Bull. 13, 261–276.
Kennedy, D.P., Courchesne, E., 2008. Functional abnormalities of the default networkduring self- and other-reflection in autism. Soc. Cogn. Affect. Neurosci. 3, 177–190.
Kong, J., Tu, P.C., Zyloney, C., Su, T.P., 2010. Intrinsic functional connectivity of theperiaqueductal gray, a resting fMRI study. Behav. Brain Res. 211, 215–219.
Lewis, D.A., Anderson, S.A., 1995. The functional architecture of the prefrontal cortexand schizophrenia. Psychol. Med. 25, 887–894.
Liu, H., Kaneko, Y., Ouyang, X., Li, L., Hao, Y., Chen, E.Y., Jiang, T., Zhou, Y., Liu, Z., in press.Schizophrenic patients and their unaffected siblings share increased resting-stateconnectivity in the task-negative network but not its anticorrelated task-positivenetwork. Schizophr. Bull.
Lui, S., Li, T., Deng, W., Jiang, L., Wu, Q., Tang, H., Yue, Q., Huang, X., Chan, R.C., Collier,D.A., Meda, S.A., Pearlson, G., Mechelli, A., Sweeney, J.A., Gong, Q., 2010. Short-termeffects of antipsychotic treatment on cerebral function in drug-naive first-episodeschizophrenia revealed by “resting state” functional magnetic resonance imaging.Arch. Gen. Psychiatry 67, 783–792.
Martinez, D., Slifstein, M., Broft, A., Mawlawi, O., Hwang, D.R., Huang, Y., Cooper, T.,Kegeles, L., Zarahn, E., Abi-Dargham, A., Haber, S.N., Laruelle, M., 2003. Imaginghuman mesolimbic dopamine transmission with positron emission tomography.Part II: amphetamine-induced dopamine release in the functional subdivisions ofthe striatum. J. Cereb. Blood Flow Metab. 23, 285–300.
Meyer-Lindenberg, A.S., Olsen, R.K., Kohn, P.D., Brown, T., Egan, M.F., Weinberger, D.R.,Berman, K.F., 2005. Regionally specific disturbance of dorsolateral prefrontal–hippocampal functional connectivity in schizophrenia. Arch. Gen. Psychiatry 62,379–386.
Palaniyappan, L., Mallikarjun, P., Joseph, V., White, T.P., Liddle, P.F., 2010. Realitydistortion is related to the structure of the salience network in schizophrenia.Psychol. Med. 1–8.
Parent, A., Hazrati, L.N., 1995. Functional anatomy of the basal ganglia. I. The cortico-basal ganglia-thalamo-cortical loop. Brain Res. Brain Res. Rev. 20, 91–127.
Pettersson-Yeo, W., Allen, P., Benetti, S., McGuire, P., Mechelli, A., 2011. Dysconnectivityin schizophrenia: where are we now? Neurosci. Biobehav. Rev. 35, 1110–1124.
Polli, F.E., Barton, J.J., Cain, M.S., Thakkar, K.N., Rauch, S.L., Manoach, D.S., 2005. Rostraland dorsal anterior cingulate cortex make dissociable contributions duringantisaccade error commission. Proc. Natl. Acad. Sci. U.S.A. 102, 15700–15705.
Postuma, R.B., Dagher, A., 2006. Basal ganglia functional connectivity based on a meta-analysis of 126 positron emission tomography and functional magnetic resonanceimaging publications. Cereb. Cortex 16, 1508–1521.
Raichle, M.E., MacLeod, A.M., Snyder, A.Z., Powers, W.J., Gusnard, D.A., Shulman, G.L.,2001. A default mode of brain function. Proc. Natl. Acad. Sci. U.S.A. 98, 676–682.
Repovs, G., Csernansky, J.G., Barch, D.M., 2011. Brain network connectivity inindividuals with schizophrenia and their siblings. Biol. Psychiatry 69, 967–973.
Robbins, T.W., 1990. The case of frontostriatal dysfunction in schizophrenia. Schizophr.Bull. 16, 391–402.
Sambataro, F., Blasi, G., Fazio, L., Caforio, G., Taurisano, P., Romano, R., Di Giorgio, A., Gelao,B., Lo Bianco, L., Papazacharias, A., Popolizio, T., Nardini, M., Bertolino, A., 2010.Treatment with olanzapine is associated with modulation of the default modenetwork in patients with schizophrenia. Neuropsychopharmacology 35, 904–912.

247P.-C. Tu et al. / NeuroImage 59 (2012) 238–247
Sanfilipo,M., Lafargue, T., Rusinek, H., Arena, L., Loneragan, C., Lautin, A., Feiner, D., Rotrosen,J., Wolkin, A., 2000. Volumetric measure of the frontal and temporal lobe regions inschizophrenia: relationship to negative symptoms. Arch. Gen. Psychiatry 57, 471–480.
Seamans, J.K., Yang, C.R., 2004. The principal features and mechanisms of dopaminemodulation in the prefrontal cortex. Prog. Neurobiol. 74, 1–58.
Seeley, W.W., 2008. Selective functional, regional, and neuronal vulnerability infrontotemporal dementia. Curr. Opin. Neurol. 21, 701–707.
Seeley, W.W., Carlin, D.A., Allman, J.M., Macedo, M.N., Bush, C., Miller, B.L., Dearmond, S.J.,2006. Early frontotemporal dementia targets neurons unique to apes and humans.Ann. Neurol. 60, 660–667.
Seeley, W.W., Menon, V., Schatzberg, A.F., Keller, J., Glover, G.H., Kenna, H., Reiss, A.L.,Greicius, M.D., 2007. Dissociable intrinsic connectivity networks for salienceprocessing and executive control. J. Neurosci. 27, 2349–2356.
Seeley, W.W., Crawford, R.K., Zhou, J., Miller, B.L., Greicius, M.D., 2009. Neurodegen-erative diseases target large-scale human brain networks. Neuron 62, 42–52.
Sheehan, D.V., Lecrubier, Y., Sheehan, K.H., Amorim, P., Janavs, J., Weiller, E., Hergueta, T.,Baker, R., Dunbar, G.C., 1998. The Mini-International Neuropsychiatric Interview(M.I.N.I.): the development and validation of a structured diagnostic psychiatricinterviewforDSM-IVand ICD-10. J. Clin.Psychiatry59 (Suppl. 20), 22–33(quiz34–57).
Shulman, G.L., Corbetta, M., Buckner, R.L., Raichle, M.E., Fiez, J.A., Miezin, F.M., Petersen,S.E., 1997. Top-down modulation of early sensory cortex. Cereb. Cortex 7, 193–206.
Storey, J.D., 2002. A direct approach to false discovery rates. J. R. Stat. Soc. Ser. B Stat.Methodol. 64, 479–498.
Tekin, S., Cummings, J.L., 2002. Frontal–subcortical neuronal circuits and clinicalneuropsychiatry: an update. J. Psychosom. Res. 53, 647–654.
Tu, P.C., Yang, T.H., Kuo, W.J., Hsieh, J.C., Su, T.P., 2006. Neural correlates of antisaccadedeficits in schizophrenia, an fMRI study. J. Psychiatr. Res. 40, 606–612.
Tu, P., Buckner, R.L., Zollei, L., Dyckman, K.A., Goff, D.C., Manoach, D.S., 2010. Reducedfunctional connectivity in a right-hemisphere network for volitional ocular motorcontrol in schizophrenia. Brain 133, 625–637.
Uddin, L.Q., Kelly, A.M., Biswal, B.B., Xavier Castellanos, F., Milham, M.P., 2009.Functional connectivity of default mode network components: correlation, antic-orrelation, and causality. Hum. Brain Mapp. 30, 625–637.
Vincent, J.L., Snyder, A.Z., Fox, M.D., Shannon, B.J., Andrews, J.R., Raichle, M.E., Buckner,R.L., 2006. Coherent spontaneous activity identifies a hippocampal–parietalmemory network. J. Neurophysiol. 96, 3517–3531.
von Economo, 1926. Eine neue art spezialzellen des lobus cinguli und lobus insulae. Z.Gesamte Neurol. Psychiatr. 100, 706–712.
White, T.P., Joseph, V., Francis, S.T., Liddle, P.F., 2010. Aberrant salience network(bilateral insula and anterior cingulate cortex) connectivity during informationprocessing in schizophrenia. Schizophr. Res. 123, 105–115.
Whitfield-Gabrieli, S., Thermenos, H.W., Milanovic, S., Tsuang, M.T., Faraone, S.V.,McCarley, R.W., Shenton, M.E., Green, A.I., Nieto-Castanon, A., LaViolette, P., Wojcik,J., Gabrieli, J.D., Seidman, L.J., 2009. Hyperactivity and hyperconnectivity of thedefault network in schizophrenia and in first-degree relatives of persons withschizophrenia. Proc. Natl. Acad. Sci. U.S.A. 106, 1279–1284.
Wible, C.G., Anderson, J., Shenton, M.E., Kricun, A., Hirayasu, Y., Tanaka, S., Levitt, J.J.,O'Donnell, B.F., Kikinis, R., Jolesz, F.A., McCarley, R.W., 2001. Prefrontal cortex,negative symptoms, and schizophrenia: an MRI study. Psychiatry Res. 108,65–78.
Wolf, D.H., Gur, R.C., Valdez, J.N., Loughead, J., Elliott, M.A., Gur, R.E., Ragland, J.D., 2007.Alterations of fronto-temporal connectivity during word encoding in schizophre-nia. Psychiatry Res. 154, 221–232.
Yan, H., Zuo, X.N., Wang, D., Wang, J., Zhu, C., Milham, M.P., Zhang, D., Zang, Y., 2009.Hemispheric asymmetry in cognitive division of anterior cingulate cortex: aresting-state functional connectivity study. Neuroimage 47, 1579–1589.
Ystad, M., Eichele, T., Lundervold, A.J., Lundervold, A., 2010. Subcortical functionalconnectivity and verbal episodic memory in healthy elderly—a resting state fMRIstudy. Neuroimage 52, 379–388.
Zhou, Y., Liang, M., Tian, L., Wang, K., Hao, Y., Liu, H., Liu, Z., Jiang, T., 2007. Functionaldisintegration in paranoid schizophrenia using resting-state fMRI. Schizophr. Res. 97,194–205.
Zhou, J., Greicius, M.D., Gennatas, E.D., Growdon, M.E., Jang, J.Y., Rabinovici, G.D., Kramer, J.H.,Weiner, M., Miller, B.L., Seeley, W.W., 2010. Divergent network connectivity changes inbehavioural variant frontotemporal dementia and Alzheimer's disease. Brain 133,1352–1367.





























