Embed Size (px)
Citation preview

UE5 Ronéo n°5 - Cours n° 6 Page 1 sur 18
UE5 IMMUNOLOGIE
Pr Carcelain
Le 27 octobre de 10h30 à 12h30
Ronéotypeur: Margaux Dumas
Roneolecteur: Alexandra Hemet
COURS 6
Origine et différenciation des
lymphocytes T, TCR et tolérance
centrale
La prof met à disposition son mail pour d’éventuelles questions:
[email protected] Le cours n’a pas du tout changé par rapport à celui de
l’année dernière.
La prof n’a pas relu la ronéo, cependant elle nous informe que l’ensemble des cours
d’immunologie peut être trouvé sur le site de l’assim sous forme de livre numérique.
Livre Immunologie Fondamentale et Immunopathologie, L2/L3 médecine, Elsevier
Masson.

UE5 Ronéo n°5 - Cours n° 6 Page 2 sur 18
Sommaire
I- Les lymphocytes T dans le système immunitaire
II- Le récepteur à l’antigène des lymphocytes T = TCR
III- Les organes lymphoïdes primaires
IV- La maturation centrale des lymphocytes T
1. Le réarrangement des gènes du TCR 2. La sélection positive 3. La sélection négative
V- Sortie du thymus des lymphocytes T et circulation
périphérique VI- La numération lymphocytaire
VII- Les lymphocytes T non classiques

UE5 Ronéo n°5 - Cours n° 6 Page 3 sur 18
Introduction (Partie dite à l’oral par la prof)
Ce cours est axé sur les lymphocytes T, leur origine et leurs différenciations.
Il y a 2 types de différenciations :
Différenciation centrale : Dans la moelle : la cellule T n’est pas une cellule finie, elle est
produite en pré-lymphocytes T dans la moelle, pareil pour le B
Différenciation périphérique : Dans la moelle osseuse pour le lymphocyte B et dans le
thymus pour le lymphocyte T. Avant cette différenciation, les lymphocytes ne sont pas
prêt à l’action (Par exemple on sait que les LB ne sont pas capables de produire des
anticorps, il faut d’abord qu’ils deviennent plasmocytes avant de pouvoir les produire). Il
faut une maturation une fois que l’antigène est rencontré
Il y a donc deux différenciations : une pour avoir une cellule compétente, et l’autre pour avoir
une cellule effectrice
Ce qui caractérise un lymphocyte T comme un lymphocyte B c’est un récepteur spécifique à
l’antigène : T Cell Receptor (TCR) pour les LT et BCR pour les LB. Chaque cellule B et chaque
cellule T va avoir la particularité d’avoir un récepteur à l’antigène différent de sa voisine. Pour
réussir à avoir des cellules qui ont des récepteurs différents, le hasard va intervenir, mais cela va
avoir pour conséquence de créer des cellules qui ne vont pas être utilisables car elles ne vont pas
reconnaitre les molécules qui vont présenter l’antigène et car certaines cellules créées vont être
contre soi puisque c’est du hasard -> Processus aléatoire. Il va donc falloir faire des sélections
de cellules et avoir une tolérance pour ne pas réagir contre le soi en particulier
I – Les lymphocytes T dans le système immunitaire
Rôle du système immunitaire
L’intérêt principal de ce système immunitaire est de distinguer le soi du non soi, dans
l’idée de répondre contre le non soi mais de ne surtout pas répondre contre le soi. Le
système immunitaire reconnaît un contexte de danger.
On distingue donc :
o Le non soi avec danger : Cela va engendrer la production de cytokines dû à des
parasites, micro organismes et virus, mais également des greffes (appelé l’autre). Si la
compatibilité n’est pas complète (au niveau des molécules du CMH qu’on hérite
chacun du père et de la mère) alors on risque une réaction contre le greffon.
o Soi modifié et danger : cellules tumorales, la tumeur est du soi au départ.
o Non soi sans danger : le fœtus par exemple, sans inflammation ou rien d’autre autour.
On a besoin d’une situation de danger pour faire réagir le système immunitaire. Nous
sommes ici dans une situation de tolérance.
o Soi sans danger : Tolérance également.
Il faut donc des cellules spécifiques, avec des récepteurs à l’antigène spécifique, qui composent
le système immunitaire et qui peuvent reconnaître uniquement ce qu’il faut.
Les réponses immunes sont coordonnées et coopérantes.
(La prof a fait un rappel sur les cellules de l’immunité)
Les cellules immunes sont divisées entre l’immunité innée et l’immunité spécifique. Ces
deux systèmes vont coopérer : l’immunité spécifique se met en marche si la réponse de
l’immunité innée n’est pas suffisante.

UE5 Ronéo n°5 - Cours n° 6 Page 4 sur 18
Immunité Innée : les cellules qui la composent sont « prêtes » elles peuvent agir
immédiatement.
On y retrouve :
Les phagocytes : monocytes/macrophages/Polynucléaires Neutrophiles qui sont les
circulants.
Les Lymphocytes NK : des cellules cytotoxiques qui vont tuer des cellules
infectées ou transformées.
Cellules dendritiques : aussi des phagocytes, mais également présentatrices d’Ag
aux cellules de l’immunité spécifique. Elles digèrent en petits morceaux l’Ag
(comme les autres cellules présentatrices d’Ag) et le présentent sur les molécules
du CMH.
Immunité spécifique :
Deux types de lymphocytes T (LT) : les CD4+ et les CD8+. Ils vont réguler les
réponses immunes. Ils possèdent un récepteur spécifique à l’Ag (qui reconnaît aussi la
molécule du CMH). Le CD4 a pour rôle spécifique la régulation des réponses immunes
en libérant des cytokines pour transmettre des signaux aux autres cellules : CD4 est le
« chef d’orchestre » de la réponse immunitaire. Tandis que CD8 a pour rôle d’appliquer
sa cytotoxicité sur les cellules infectées ou anormales.
Les Lymphocytes B (LB) : E n d e v e n a n t p l a s m o c y t e s , i l s sécrètent des
anticorps.
Les lymphocytes B concernent donc des pathogènes qui sont extracellulaires alors que les
cellules cytotoxiques comme les LT concernent les pathogènes en intracellulaire.
Ce groupe de cellules de l’immunité spécifique est caractérisé par la présence d’un récepteur à
l’Ag : chaque LB et LT a un récepteur unique pour avoir le plus possible de reconnaissance.
Seules quelques cellules sont dirigées contre le même Antigène. L’expansion clonale est la
prolifération des cellules afin d’en avoir plus d’une qui soit spécifique contre un AG donné
pour pouvoir y répondre. Elles doivent ensuite donner lieu aux cellules mémoires, qui vont
permettre de réagir beaucoup plus vite au cours d’une seconde attaque.
Il y a des récepteurs pour reconnaître spécifiquement des épitopes, Ces récepteurs sont crées par
recombinaison, il y a une diversité importante.
Le récepteur des LT (TCR) reconnaît des petits fragments de protéines qui ont été
produits à l’intérieur des cellules qui sont présentées sur les molécules du CMH
(Complexe Majeur d’Histocompatibilité).
Le récepteur des LB reconnaît l’Ag directement (soit soluble soit sur une cellule).
Pour avoir des Anticorps spécifiques, il y a besoin d’une maturation de 21 jours des LB et de 7
jours pour les lymphocytes T.
Il n’y a pas beaucoup de lymphocytes B et de lymphocytes T spécifique d’un antigène, c’est pour
cela que des qu’il y a une cellule spécifique on va devoir la multiplier pour en avoir plusieurs :
c’est l’expansion clonale.
Notion de clone : 1 antigène ↔ 1 lymphocyte

UE5 Ronéo n°5 - Cours n° 6 Page 5 sur 18
Les Lymphocytes T représentent 75 à 85% des lymphocytes sanguins.
Comme vu précédemment les
lymphocytes sont caractérisés par leur
TCR mais également par l’expression
d’une molécule associée : la molécule
CD3 (qui caractérise un LT).
Un récepteur à l’Ag des cellules T est
composé de deux chaînes. Ces chaînes
sont des glycoprotéines codées par des
gènes en situation germinale.
-Le TCR αβ est majoritaire (>90%)
-Le TCR γδ est minoritaire (<10%)
Les cellules auxquelles on va s’intéresser et qui sont majoritaires sont les LT
classiques, conventionnels, présents dans le sang en majorité. Ils expriment un récepteur avec les
chaînes α et β, soit CD4+ soit CD8+.
CD4 : Fonction auxiliaire avec sécrétion de cytokines pour faire maturer les autres
cellules du système immunitaire.
CD8 : cytotoxiques.
De façon minoritaire il existe également des lymphocytes dits non classiques sur
lesquels on reviendra.
II- Le recepteur T pour l’antigene (TCR)
La structure du complexe CD3-TCR : il n’y a jamais de TCR sans CD3 dans les
situations normales.
Le TCR : Les deux chaînes (reliées par
un pont disulfure) sont l’assemblage de
segments différents, dont une partie
constante au niveau C-ter et une partie
variable au niveau de l’extrémité N-ter.
Tout au bout des chaînes (N-ter), on
trouve la région dite hypervariable.
Aucun LT n’a de régions hypervariables
identiques, et c’est à cet endroit qu’aura
lieu la reconnaissance.
En intra-cytoplasmique il n’y a rien
d e spécifique ce qui montre que cette
molécule est entièrement dédiée vers
l’extérieur et donc vers la
reconnaissance.

UE5 Ronéo n°5 - Cours n° 6 Page 6 sur 18
Le but de la reconnaissance de la cellule est d’activer la cellule afin d’activer la
prolifération, la sécrétion de cytokines etc…
L’association avec la molécule CD3 (composée de l’association de 5 chaînes différentes
possibles) permet une présence également en intracellulaire, puisque cette dernière est beaucoup
plus dirigée vers l’intracellulaire. Elle ne va pas reconnaître l’Ag mais va permettre la
transduction du signal. Lorsque le TCR a reconnu l’Ag, il va interagir en transmembranaire
au niveau des résidus phosphorylés, et activer des motifs d’activation (ITAM) de la molécule
CD3. Une fois qu’il y a cette activation, des kinases vont être recrutées qui vont phosphoryler
ITAM puis entrainer une cascade de réactions à l’intérieur de la cellule.
On a donc bien besoin d’un TCR et d’une molécule CD3.
Les TCR ont une structure du type « immunoglobulin like »
Cette structure a été initialement décrite en observant la structure d’une immunoglobuline. Il
s’agit en fait d’un domaine conservé de 110 acides aminés, chacun reliés par un pont
disulfure avec une structure interne de feuillets β. On parle de structure en tonneau.
Beaucoup de molécules ont ce type de structure en immunologie comme CD3, CD4, CD8…
Les molécules CD4 et CD8:
Associées au TCR et à CD3, il y a beaucoup d’autres molécules qui vont servir à
l’activation périphérique.
Quand le LT reconnaît son Ag présenté par le CMH, les deux cellules doivent restées collées
l’une à l’autre pendant un moment de façon stable, notamment pour avoir le temps de s’activer.
Pour stabiliser cette reconnaissance on a plein de molécules qui vont agir autour, et c’est ce
phénomène qu’on appelle la synapse immunologique. Dans cette synapse sont présentes des
molécules qui permettent l’adhésion dont le CD3, le TCR et le CD4 ou le CD8. Ces
molécules CD4 et CD8 servent à deux choses : à stabiliser l’interaction car elles interagissent
avec les molécules constantes du CMH et à participer à la signalisation intracellulaire en
recrutant des kinases.
Le complexe majeur d’histocompatibilité (CMH)
Il existe sur certaines cellules humaines des molécules qui sont différentes d’un sujet à l’autre.
Pour les molécules du CMH : on en reçoit la moitié du père et l’autre de la mère, et de par les
combinaisons au niveau des frères et sœurs on aura donc une chance sur quatre d’avoir les
mêmes.
C’est un complexe puisque qu’il y a plus de 200 gènes codants pour des produits très divers.
On dit qu’il est majeur car les produits sont à l’origine de différences allogéniques importantes
entre individus de la même espèce.
D’autre part, on parle d’histocompatibilité puisqu’elles sont à l’origine de rejet de greffe entre
sujets incompatibles par exemple.
Fonctions principales : Présenter des peptides du soi ou du non soi. Une molécule HLA
(CMH) n’est jamais vide, pour qu’elle soit stable elle doit toujours avoir quelque chose à
présenter : c’est un système qui permet de montrer à l’extérieur ce qu’il se passe à l’intérieur
de la cellule (peptides qui proviennent soit de la cellule, soit de ce qu’elle a ingéré).
Intéressant également pour la sélection thymique (quel TCR on va garder ou non)
Elles sont importantes en périphérie pour l’action du système immunitaire et elles vont
aussi modifier l’activité cytotoxique des cellules NK. Au sein du CMH on retrouve les
molécules de la classe I et celles de la classe II.

UE5 Ronéo n°5 - Cours n° 6 Page 7 sur 18
Elles ne vont pas présenter aux mêmes cellules, et n’ont pas exactement la même
structure. On a un sillon de présentation dans lequel va se nicher le peptide. Ces
molécules sont codées par le chromosome 6 et elles sont nombreuses.
Il existe plusieurs molécules de classe II : P, R Q et plusieurs molécules de classe I : A, B et C.
Selon la molécule, on va présenter une séquence du peptide différente en fonction de la
composition chimique en acides aminés de la « poche » de la molécule HLA (elle même
dépendante de sa séquence).
D’où la nécessité d’avoir beaucoup de molécules du HLA pour présenter beaucoup de
choses, et on ne va pas tous présenter la même chose en fonction de nos séquences.
On reçoit un chromosome du père et un chromosome de la mère, il s’agit d’une
transmission en haplotypes (un du père et un de la mère), ce qui explique les 4 situations
différentes pour les enfants. Les recombinaisons sont rares.
À la surface des cellules, on co-exprime, l’ensemble des molécules : classe I et
classe II. Cela va dépendre des cellules, mais les expressions ne s’excluent pas les unes des
autres.
Les cellules n’expriment pas toutes leur classe II mais elles expriment toutes leur classe I.
À l’intérieur du sillon, on a un
peptide de petite taille qui
rentre dans la poche pour la
classe I (présence de ponts
salins qui vont limiter la taille
du peptide).
Pour la classe II, on a un plus
long peptide, qui déborde de la
poche, >13aa (mais les
chiffres peuvent varier).
CMH et HLA sont des
termes différents mais
qualifiant les mêmes
molécules, elle varie donc
entre les deux.
Le TCR avec son extrémité hypervariable à l’extrémité, reconnaît le peptide présent dans la
poche du CMH, ainsi le TCR ne reconnaît pas que le peptide mais également le CMH. Il
reconnaît le peptide dans le contexte du CMH, plus précisément de notre CMH. C’est ce qu’on
appelle la reconnaissance restreinte par le CMH.
Le TCR du LT mature
o Les TCR des LT CD4 reconnaissent des peptides de 12 à 15 acides aminés (long) présentés
par le CMH de classe II présent sur les CPA, provenant de la dégradation intracellulaire
(par voie endosomale) de protéines extracellulaires. La cellule dendritique, le LB et le
macrophage sont les cellules présentatrices d’Ag (CPA).
o Les TCR des LT CD8 reconnaissent des peptides de 9 acides aminés présentés par le
CMH de classe I sur toutes les cellules nucléées (d’où la nécessité que toutes les cellules
expriment les molécules de classe I), provenant de la synthèse de protéines intracellulaires de
pathogènes (cellule anormale).

UE5 Ronéo n°5 - Cours n° 6 Page 8 sur 18
III- Les organes lymphoïdes primaires : Moelle osseuse et Thymus
Organes et tissus lymphoïdes
o Organe souche : la moelle osseuse, qui va donner l’ensemble des précurseurs.
o Lymphoïdes primaires ou centraux : la moelle osseuse et le thymus, qui vont
permettre la maturation et la différenciation des précurseurs (LB dans la moelle, LT dans le
thymus).
o Organes et tissus lymphoïdes secondaires ou périphériques : les cellules naïves vont
circuler vers ces organes et tissus. Ils sont nombreux (ganglions lymphatiques, rate, tissu
lymphatique associé aux bronches, amygdales, ganglions mésentériques, plaques de
Peyer, tissu lymphoïde urogénital…) qui sont le siège des réactions immunitaires.
La moelle osseuse (MO)
Elle est présente dans les os longs et le squelette axial. Elle est très irriguée, les précurseurs
vont partir de la moelle pour aller vers le thymus.
Toutes les cellules sanguines sont dérivées des cellules souches de la moelle, seules capables
d’auto-renouvellement ou auto-régénération.
Elle assure 3 fonctions :
o Maintien d’un contingent de cellules souches et production de cellules hématopoïétiques
o Maturation et différenciation des pro-lymphocytes B en LB matures aptes à coloniser les
organes lymphoïdes secondaires
o Hébergement des LB activés par l’Ag en provenance des organes secondaires et qui se
transforment en plasmocytes sécréteurs d’Ac (anticorps) à longue durée de vie.
Le thymus
C’est un organe lymphoïde primaire qui sera colonisé par les progéniteurs T.
Il est bilobé, thoracique en position médiastinale (soit médian) et derrière le sternum.
Il est composé de lobes et chaque lobe est
divisé en lobules. Ces lobules sont recouverts
d’une capsule avec des invaginations de la
capsule formant des septums. On retrouve le
cortex en périphérie (avec thymocytes
matures) et la medulla au centre (avec les
cellules matures)
Composition cellulaire du Thymus :
On observe des thymocytes qui sont les précurseurs de LT, et les plus représentés. Il va
falloir sélectionner les bons LT, ainsi l’une des caractéristique du thymus est qu’à l’intérieur de
celui-ci il va y avoir beaucoup de pertes, et donc beaucoup de cellules macrophagiques qui vont
éliminer les cellules mortes.
On retrouve également des cellules épithéliales corticales, des cellules dendritiques et des
cellules épithéliales médullaires. Ces trois types de cellules vont participer à l’interaction avec
le récepteur à l’Ag, elles sont donc importantes et sont toutes les trois des cellules stromales.
UE5 Ronéo n°5 - Cours n° 6 Page 9 sur 18
Fonctions :
Maturation des progéniteurs T pour produire des LT CD4+ ET CD8+ naïfs qui vont
migrer par la suite en périphérie
S’il n’y a pas de thymus il n’y a pas de LT naïfs (syndrome de Di Georges)
Production journalière moyenne environ de 108
de LT par jour, mais ces chiffres dépendent
de l’âge.
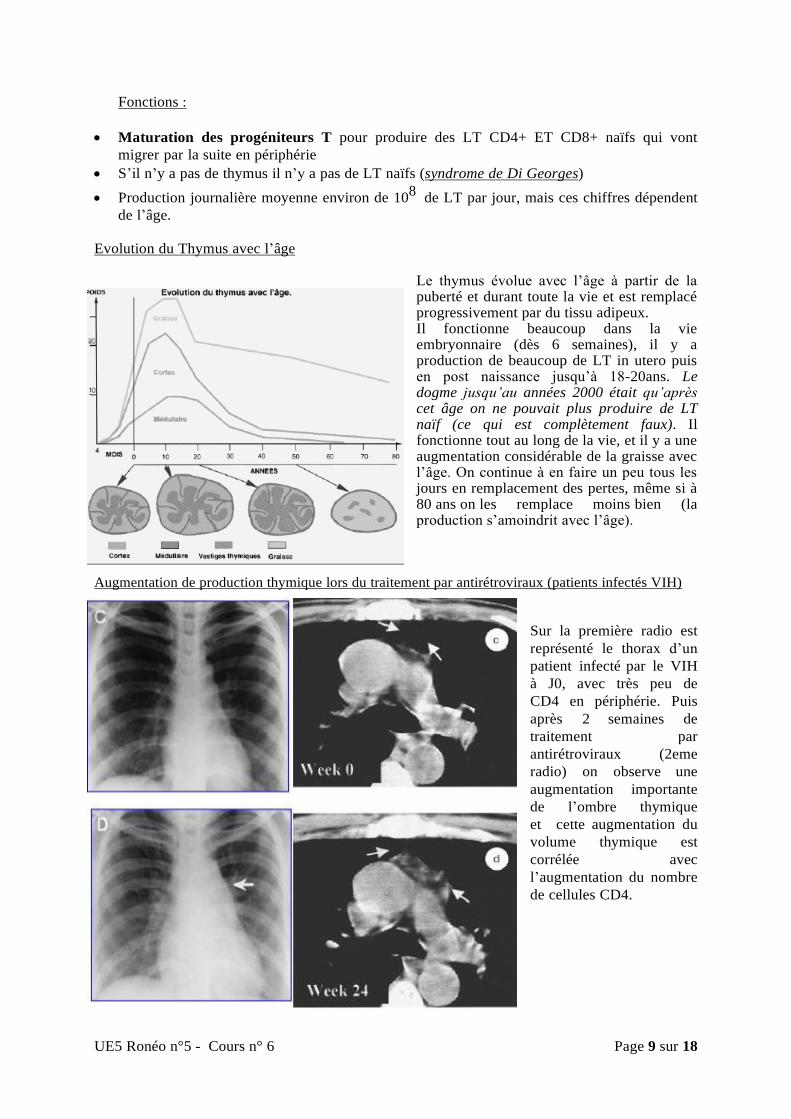
Evolution du Thymus avec l’âge
Le thymus évolue avec l’âge à partir de la puberté et durant toute la vie et est remplacé progressivement par du tissu adipeux. Il fonctionne beaucoup dans la vie embryonnaire (dès 6 semaines), il y a production de beaucoup de LT in utero puis en post naissance jusqu’à 18-20ans. Le dogme jusqu’au années 2000 était qu’après cet âge on ne pouvait plus produire de LT naïf (ce qui est complètement faux). Il fonctionne tout au long de la vie, et il y a une augmentation considérable de la graisse avec l’âge. On continue à en faire un peu tous les jours en remplacement des pertes, même si à 80 ans on les remplace moins bien (la production s’amoindrit avec l’âge).
Augmentation de production thymique lors du traitement par antirétroviraux (patients infectés VIH)
Sur la première radio est
représenté le thorax d’un
patient infecté par le VIH
à J0, avec très peu de
CD4 en périphérie. Puis
après 2 semaines de
traitement par
antirétroviraux (2eme
radio) on observe une
augmentation importante
de l’ombre thymique
et cette augmentation du
volume thymique est
corrélée avec
l’augmentation du nombre
de cellules CD4.

UE5 Ronéo n°5 - Cours n° 6 Page 10 sur 18
IV- Maturation centrale des Lymphocytes T
Étapes importantes de la maturation centrale des lymphocytes T o Multiplication intense des thymocytes, qui précède une élimination très importante au cours
des sélections
o Acquisition des marqueurs de surface spécifiques de lignée : exemple CD3 spécifique des LT
o Acquisition des récepteurs spécifiques à l’Ag par recombinaison somatique au hasard : TCR
o Sélections (positive et négative) des LT compétents et ne reconnaissant pas le soi
o Sortie des organes lymphoïdes de LT naïfs
Facteurs nécessaires à la maturation centrale des cellules lymphoïdes
Ce sont des facteurs de survie importants. L’environnement du thymus doit les produire, il y
en a beaucoup, la prof ne les cite pas tous, mais affirme que les principaux et ceux qui sont à
retenir sont les IL7 et IL2. Ces deux cytokines (=interleukines) sont utilisées en clinique.
Chez des patients cancéreux on a utilisé l’IL2 pour faire proliférer les lymphocytes et pour le
VIH on a également utilisé IL2 et IL7 afin de faire remonter les LTCD4.
Pour la maturation des cellules ayant lieu dans le thymus, les cellules reçoivent les signaux de
survie (pour qu’elles ne meurent pas en route) via l’interaction du TCR et par ces facteurs
qui sont synthétisés par le thymus.
On a également besoin du réseau des cellules épithéliales qui permet la maturation des LT et qui sont:
o Les cellules épithéliales nourricières (cortex externe) qui sécrètent beaucoup d’IL7,
facteur permettant la prolifération et la survie
(la prof insiste encore une fois sur le fait que la prolifération est importante puisque par la suite
nous allons perdre beaucoup de LT)
o Les cellules épithéliales du cortex thymique (cTEC) pour la sélection positive
o Les cellules épithéliales médullaires (mTEc) pour la sélection négative (enlever les mauvais
TCR qui reconnaissent le soi)
Au début de la maturation, les cellules sont doublement négatives, elles n’expriment ni le CD4 ni le CD8 (avec DN=double négatif sur le schéma ci-contre) ce sont des thymocytes qui peuvent encore devenir des LT des LB ou des cellules dendritiques. Ce qui signe l’entrée dans la lignée T c’est
l’acquisition, en même temps que le TCR, du
CD3.
Au stade DN2, elles peuvent encore se
différencier en lymphocytes B ou T
(théoriquement), puis elles deviennent double
positives.
Ensuite a lieu la sélection positive
(représentée dans le deuxième cadre du
schéma en partant du haut) : on séléctionne le TCR qui reconnaît soit la classe I, soit la classe II
des molécules HLA.
Une cellule qui reconnaît la classe II devient CD4, une autre qui reconnaît la classe I devient
CD8.
Au cours de cette sélection positive on a reconnaissance des HLA, et on finit avec
CD4 simple positive ou CD8 simple positive. La sélection négative arrive en dernier avec la reconnaissance du soi, si la cellule le reconnaît elle meurt, sinon elle finit sa maturation et part en
périphérie. Il y a présence de macrophages qui vont phagocyter tout ce qu’on a éliminé.

UE5 Ronéo n°5 - Cours n° 6 Page 11 sur 18
1. Le réarrangement des gènes du TCR
Le but est de constituer ces chaines
uniques, avec leur partie extrêmement
variable en N-ter. On veut une séquence
unique afin qu’un TCR n’ait pas la même
séquence que son « voisin ».
Au départ on a une association de gènes
variables : de diversité D de jonction J et de
constante C. Pour chaque chaîne, si on
prend β comme exemple, on va
sélectionner dans une batterie de gènes en
position germinale différents gènes :
On a par exemple 100 V différents, et
même chose pour les D. Chaque LT va
prendre un D un J et un C (constant)
uniques parmi tous ce qu’on lui propose.
On réarrange la chaine β et ensuite α. Particularité : Lorsqu’on réarrange α, la partie de chaine δ est à l’intérieur et donc il faut exciser tout le locus delta.
Réarrangement des chaînes alpha et beta :
Les gènes sont en situation germinale (initiale,
originelle, sans qu’on ait encore rien touché) dans
le noyau du thymocyte.
Dans la diapo on a l’exemple des chaines alpha et
beta, la prof a préféré nous expliquer le
réarrangement du beta. Parmi tous les V on va
donc en choisir un au hasard via une enzyme.
Ensuite cette enzyme va choisir un D, couper ce
qu’il y a entre les deux et les coller. Elle associe
donc le V au D, puis par le même mécanisme
elle va associer ce VD à un J unique. On se
retrouve donc avec un gène réarrangé particulier,
un gène recombiné.
Se produit ensuite une subtilité supplémentaire :
Il y a deux systèmes d’enzymes qui interviennent, les plus connus sont les RAG
(recombinase activating gene) qui coupent et recollent. Intervention également d’endonucléases
et de TdT (terminal desoxynucleotidyl transferase), qui, lorsque l’on va couper, vont enlever
quelques acides aminés et en rajouter quelques uns (qui sont non codés par le génome) de
façon complètement aléatoire ce qui réduit d’autant plus les possibilités de se retrouver avec les
mêmes séquences,
À l’extrémité du V on est dans une région hypervariable, le J et le D sont aussi variables et le
C est constant. Entre l’extrémité du V et celle du C c’est hypervariable, et c’est le domaine qui
va interagir avec le complexe CMH/peptide.

UE5 Ronéo n°5 - Cours n° 6 Page 12 sur 18
Tout ce qui est enlevé lors des réarrangements est des segments d’ADN inutiles.
Ils vont se circulariser dans la cellule (plus précisément dans le noyau de la cellule thymique),
ils forment ainsi ce qu’on appelle des cercles d’excision (en particulier quand on excise le
locus delta). Dans toutes les cellules on excise le même locus delta. Ces cercles restent en
position épisomales dans le noyau. Lorsque la cellule se réplique, ils ne se répliquent pas.
Quand elle se divise, c’est uniquement la première cellule naïve qui conserve le cercle
d’excision, les autres cellules mémoires qui resteront ne l’auront pas.
Ces cercles sont donc des bons marqueurs de la production thymique. En effet
pour évaluer la production de LT naïfs on peut quantifier ces cercles par PCR quantitative.
Cette nouvelle production thymique est importante puisqu’elle permet d’augmenter la
spécificité et donc d’élargir le « répertoire » des LT.
Par exemple pour un malade du VIH, la prolifération des LT qui lui restent est un bon début
mais ne permet pas d’augmenter sa spécificité qui est très faible. La production de nouveaux
LT naïfs par le thymus lui permet d’élargir le répertoire des LT.
Ces marqueurs ont permis de réfuter le dogme qui persistait jusqu’aux années 2000, en
prouvant que le thymus était capable de produire des LT naïfs tout au long de la vie.
Dans ce tableau les chiffres ne sont pas à retenir
« ils changent tout le temps ». On peut juste voir
les différents éléments, soit les segments V de
variabilité, les segments D uniquement pour beta,
les nucléotides enlevés et ajoutés, les segments de
jonction J etc.
Au total on combine une diversité combinatoire (en associant une chaîne alpha à une chaîne beta)
et une diversité fonctionnelle, qui donne une diversité totale de 1018
, en pratique c’est pas vraiment le bon chiffre, on trouve toujours plus qu’en réalité, il faut donc retenir que c’est
environ 1013
à 1015
TCR différents qui sont produits.
L’extrémité terminale du TCR a une structure tridimensionnelle.
Au niveau de cette région hypervariable sont présentes les molécules CDR
(complémenter determining regions), au nombre de 3 (CD1, CD2, et CD3). Ces régions
interagissent de plus avec la poche à peptide et le peptide lui-même. Entre les CDR (replis) il
y a les FR (framework region) qui présentent peu de variabilité. Les interactions ont lieu entre
les acides aminés des CDR (paratope) et l’Ag (épitope)
Le site de fixation est donc défini par des séquences hypervariables qui sont les CDR.
Au total, les LT naïfs ont chacun un TCR unique, créés au hasard par recombinaison de gène
ce qui permet la reconnaissance d’un grand nombre de peptides différents. Mais :
reconnaissant les molécules du CMH de l’hôte ou pas (sélection positive)
reconnaissant du non soi ou du soi (sélection négative)
Il faut donc éliminer les thymocytes ayant un TCR ne reconnaissant pas les molécules du
CMH de l’hôte et/ou reconnaissant le soi, ce qui va induire des pertes très importantes.

UE5 Ronéo n°5 - Cours n° 6 Page 13 sur 18
2. La sélection positive
Le TCR est fait, avec chaine α et β (si la cellule n’y arrive pas alors elle meurt).
Dans le thymus se produit la sélection positive, où l’on a besoin des cellules épithéliales
corticales. Ces cellules sont des mauvaises présentatrices d’Ag, elles présentent les Ag du soi
mais ce qui nous intéresse c’est qu’elles expriment très bien les molécules du CMH.
À ce stade, si on n’a pas de reconnaissance du CMH/peptide on meurt, il n’y aura pas de signal
de survie donné. En revanche si la cellule reconnaît, la reconnaissance sera faible mais un signal
de survie sera donné. Si on reconnaît le CMH de classe I on survit et on diminue l’expression
du CD4 pour avoir une CD8 simple positive, et inversement pour CD4.
Les CMH de classe I et II permettent ainsi la sélection positive des cellules CD8 et
CD4 respectivement, et on a toujours la notion de restriction par le CMH.
3. La sélection négative
À cette étape on va enlever les LT qui réagissent contre les Ag du soi dans le thymus.
On est donc dans une situation particulière puisqu’on est dans un tissu particulier et qu’il faut
faire réagir les LT contre l’ensemble des Ag du soi de l’organisme. Cela est possible grâce
aux cellules épithéliales médullaires, et plus précisément grâce à un système génétique appelé
Aire (auto-immune regulatory gene) qui va faire ré-exprimer la majorité des gènes du soi,
par exemple du cordon ombilical, des poumons, du foie, du pancréas, des glandes salivaires
etc… globalement plus de 80% (imparfait).
Bien évidemment certaines cellules vont y échapper, mais la tolérance centrale (le fait
d’éviter de répondre contre le soi) est majeure à hauteur de 80% mais n’est pas complète. Par
la suite, en périphérie, des cellules régulatrices très importantes vont contrôler les LT auto-
réactifs.
La cellule dendritique peut également avoir ce rôle après avoir phagocyté les cellules
épithéliales. Tout cela aboutit à la présentation sur classe I et classe II de protéines du soi de tissus
même éloignés du thymus. Il y a un engagement fort du récepteur, et pour le TCR qui reconnaît le
soi, il y a un signal hyper fort, voire trop fort, qui entraîne la mort de la cellule. Avec la sélection
négative, il y a donc perte des cellules auto-réactives.

UE5 Ronéo n°5 - Cours n° 6 Page 14 sur 18
CD =Cluster de différenciation, qui permet
de caractériser les différents stades de
différenciation.
On retrouve au départ une cellule souche,
puis un précurseur thymocyte qui devient
DN1 et 2 qui sont multipotentes (peuvent
encore devenir LB, LT, NK…) encore, puis
les pro-T (donnent essentiellement T, ou NK
possible), et enfin la DP (double positif)
avec la molécule CD3 réelle, on est
définitivement engagé dans la lignée T.
Il y a encore une toute petite subtilité
concernant le réarrangement du TCR, il se
fait en deux phases : D’abord on réarrange
la chaine beta, puis on l’associe avec une
pré-chaine alpha, qui est une pseudo chaine alpha ayant une très faible variabilité. Cette pseudo
chaine sert juste à constituer un TCR pour qu’il y ait des signaux de survie afin qu’il puisse rester
et proliférer encore un peu, et ensuite on va faire la vraie chaîne alpha.
On peut utiliser toutes ces molécules CD pour les caractériser mais on y reviendra à la fin.
V- Sortie du thymus des lymphocytes T et circulation périphérique
Circulation des lymphocytes empruntant les vaisseaux sanguins et lymphatiques
Les organes lymphoïdes primaires (moelle osseuse et thymus), permettent la production
des cellules naïves. Celles-ci vont sortir et rejoindre la circulation à la recherche de leur Ag. (Un
bébé à la naissance a essentiellement des cellules naïves, car elles n’ont pas encore rencontré
d’Ag). Parallèlement, la cellule dendritique capte l’Ag et l’apporte au ganglion (gg) proximal,
c’est-à-dire le plus proche. Les ganglions sont présents dans tout le corps, il y en a énormément.
La cellule dendritique le met donc dans le ganglion, et les cellules naïves vont pouvoir le
rencontrer en entrant dans le ganglion (puis le ganglion gonfle).
La cellule naïve peut également rentrer au niveau des MALT à travers les veinules à
endothélium élevé. Elle peut aussi rentrer dans la rate à travers la zone marginale.

UE5 Ronéo n°5 - Cours n° 6 Page 15 sur 18
Ensuite on a les organes lymphoïdes secondaires, qui permettent la différenciation et
l’activation, puis les cellules effectrices repartent en périphérie par le canal thoracique,
dans le sang. Elles reviennent ainsi à l’endroit où était la cellule dendritique du départ. Ainsi
les cellules doivent circuler dans le sang et passer en permanence dans les organes lymphoïdes.
Pour cela, la cellule va se retrouver sur les cellules endothéliales qui sont hautes, larges et
assez épaisses. Elle va rouler sur ces cellules hautes, s’accrocher doucement et s’il ne se
passe rien, la cellule continue son chemin en s’accrochant de façon réversible et pas très forte.
En revanche, s’il y a une inflammation,
un deuxième type de molécule d’adhésion
(pas forcément à apprendre) est augmenté,
donc la cellule va s’arrêter, il y a fixation
du LT naïf, puis activation dans le
ganglion. Ce ganglion produit des
chimiokines (sous la forme d’un gradient),
qui sont des protéines solubles, servant de
messagers, et qui jouent un rôle de
chimio attracteur. En effet, elles vont
interagir avec les récepteurs aux
chimiokines, attraper les cellules
associées, et les entrainer dans le sens du
gradient, en passant par diapédèse à
travers les cellules endothéliales.
Il existe un autre type de molécule soluble intervenant dans le système immunitaire,
qui sont les cytokines (interleukines), ayant un rôle de messagers. (« active toi » « prolifère »
etc…)
VI- Numération Lymphocytaire
CD28/B7 et CD40L/CD40 sont
nécessaires à la co activation
puisque le premier signal par CD3
et CD4 ou CD8 n’est pas suffisant,
il en faut un deuxième.
Le récepteur aux cytokines est
important pour la prolifération. Les
molécules d’adhésion sont
nécessaires à la diapédèse ainsi que
pour la synapse au début.
Ces molécules sont toutes en
surface, elles peuvent être des
marqueurs d’activation, de
cytotoxicité.
d’activation, de cytotoxicité.

UE5 Ronéo n°5 - Cours n° 6 Page 16 sur 18
On utilise la cytométrie de flux pour les étudier:
Les cellules passent une à une, dans des gaines de liquide, afin de les numérer et de déterminer
leurs marqueurs. Elles passent devant un laser, celui-ci excite la cellule et grâce à cela on
récupère deux paramètres : la taille et la structure de la cellule (granulosité). À cela on peut
ajouter la présence d’un ou de plusieurs Ac monoclonaux fluorescents, de fluorescence
différente (pour détecter la présence d’une molécule particulière). La cellule va ré-émettre en
fonction de sa granulosité. On a un signal proportionnel à sa granulosité, un signal émis « en
face » qui sera proportionnel à sa taille, et enfin un troisième signal qui va correspondre au
fluorochrome, et qui traduit la présence d’un marqueur ( par exemple CD3 en rouge, CD4 en
vert etc…). Ces signaux sont récupérés par différents photomultiplicateurs. Tous ces
fluorochromes correspondent à des molécules de surface, ainsi on peut déterminer ce qu’une
cellule possède à sa surface en fonction de ce qu’elle ré-émet.
Cette technique peut être réalisée pour beaucoup de cellules, on l’utilise typiquement pour
les LT CD4 et les CD8 (on utilise CD45 qui marque tous les leucocytes). On peut aller jusqu’à
15 marqueurs différents. L’analyse se fait sur sang total ce qui signifie que c’est rapide.
Puis on analyse les résultats sur le cytomètre, on regarde ainsi sur un graphique la
répartition des cellules étudiées en fonction de la taille (petites, grosses) et de leur granulosité
(importante ou non).
Sur le graphique on distingue les
lymphocytes qui sont peu granuleux,
les monocytes granuleux et les
PNn (polynucléaires neutrophiles)
hyper granuleux. Etant donné qu’on
s’intéresse aux LT, on va
sélectionner le groupe de
lymphocytes afin de les étudier
plus spécifiquement. On va donc
analyser les différents marqueurs
en fonction des couleurs, et cette
analyse sera faite deux par deux.
À partir de ces résultats il est
donc possible de compter les
cellules et de fournir une réponse au
patient concernant la proportionnalité
de ces cellules (en 20-30 minutes).
La prof met en diapo un tableau
de proportions de répartition des
cellules qu’elle ne demande pas d’apprendre.
Au niveau des proportions on voit
donc qu’on a environ 1/3 de CD8
et 2/3 de CD4.

UE5 Ronéo n°5 - Cours n° 6 Page 17 sur 18
On peut analyser dans le sang,
dans la moelle osseuse (où il y aura
beaucoup plus de LB), dans le
thymus (majoritairement des LT) dans
les ganglions et dans la rate il y a un
équilibre un peu particulier mais
avec plus de LB.
Chez l’enfant de moins d’un an, on a de façon caricaturale une hyperlymphocytose dès la
naissance. Et à l’opposé, chez le sujet âgé, se forme une pseudo lymphopénie qui n’est
cependant pas systématique et qui peut se voir quantitativement.
VII- Lymphocytes T non conventionnels
Les lymphocytes T avec un récepteur à l’Ag de type γẟ
Les lymphocytes T γẟ découverts il y a longtemps, ont un récepteur à l’antigène
composé de l’association d’une chaine γ et d’une chaine ẟ. Ils sont soit doubles négatives
CD 4-CD 8-, soit simples positives CD 8+ CD 4-.
Ce sont des cellules qui sont le plus souvent présentes dans les tissus et les muqueuses où
elles ont une action particulière, essentiellement anti pathogène. Elles ont une diversité
beaucoup plus restreinte (pas autant de V, de D de J) et elles sont restreintes par le CMH
non classique CD1. Elles reconnaissent les épitopes conservés au sein des pathogènes et ont
un mode d’action rapide, proche de l’immunité innée, elles ne nécessitent pas de
maturation longue, elles agissent tout de suite.
Elles sont capables de cytotoxicité et de sécrétion de cytokines, capables de réguler les
réponses immunes.
Les lymphocytes NKT
Les lymphocytes NKT expriment les marqueurs T CD3 et TCR ainsi que quelques
marqueurs des cellules NK comme CD56 et CD16. Ils ont un TCR quasi invariant avec
une chaine α V14-J18 et une diversité β mais qui est restreinte.
Les lymphocytes NKT reconnaissent des lipides présentés par la molécule de CMH non
classique CD1d. Ils ont une fréquence sanguine variable et faible et une distribution
tissulaire hétérogène. Ce sont des cellules effectrices cytotoxiques qui ont un rôle dans
la régulation des réponses immunitaires notamment dans des anomalies associées à
des pathologies auto-immunes, inflammatoires ou encore tumorales.

UE5 Ronéo n°5 - Cours n° 6 Page 18 sur 18
Conclusion : Les points clefs
o La majorité des lymphocytes T circulants ont un récepteur à l’antigène composé
des chaines α et β .
o Le TCR est au sein d’un complexe : TCR/CD3/CD4 ou CD8.
o Le TCR reconnaît le complexe CMH/peptide, notion de restriction par CMH.
o Le TCR des LT CD4 reconnaît des peptides de 12-25 acides aminés, présentés
par le CMH II des cellules présentatrices provenant de la dégradation
intracellulaire de protéines extracellulaires.
o Les TCR des LT CD8 reconnaît des peptides de 9 acides aminés, présentés par le
CMH I des cellules nucléées, qui proviennent de la synthèse des protéines
intracellulaires de pathogènes.
o Le thymus est l’organe de différenciation centrale des lymphocytes T :
multiplication, formation TCR, sélection TCR adéquats.
o Les TCR sont formés par la recombinaison au hasard de segments de gènes.
o Il y a une sélection positive des thymocytes présentant un TCR reconnaissant le
CMH et une sélection négative des thymocytes présentant un TCR reconnaissant le
soi.
o Il existe des populations de lymphocytes T non classiques, minoritaires, ayant
une fonction se rapprochant de celle des cellules innées.





















![3[1]. tcr-y_bcr](https://img.pdfslide.tips/doc/110x75/556e58bfd8b42a2c658b5305/31-tcr-ybcr.jpg)







