Embed Size (px)
Citation preview

Instituto de Pesquisas Jardim Botânico do Rio de Janeiro
Escola Nacional de Botânica Tropical Programa de Pós-graduação em Botânica
Crescimento clonal em Neoregelia cruenta na Restinga de
Jurubatiba: estrutura populacional, plasticidade morfo-anatômica e
integração fisiológica.
DULCE GILSON MANTUANO
Rio de Janeiro – 2008

Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical Programa de Pós-graduação em Botânica
Crescimento clonal em Neoregelia cruenta (R. Graham) L.B. Smith
(Bromeliaceae) na Restinga de Jurubatiba: estrutura populacional,
plasticidade morfo-anatômica e integração fisiológica.
DULCE GILSON MANTUANO
Tese apresentada ao Programa de Pós-
graduação em Botânica da Escola Nacional de
Botânica Tropical do Instituto de Pesquisas
Jardim Botânico do Rio de Janeiro, como parte
dos requisitos para a obtenção do título de
Doutora em Botânica.
Orientadora.: Dra. Cecília Golçaves Costa
Co-orientador.: Dr. Gustavo Martinelli
Rio de Janeiro – RJ
2008

ii
Crescimento clonal em Neoregelia cruenta (R. Graham) L.B. Smith
(Bromeliaceae) na Restinga de Jurubatiba: estrutura populacional,
plasticidade morfo-anatômica e integração fisiológica.
Dulce Gilson Mantuano
Tese apresentada ao Programa de Pós-Graduação em Botânica da Escola
Nacional de Botânica Tropical do Instituto de Pesquisas Jardim Botânico do Rio
de Janeiro, como parte dos requisitos necessários à obtenção do grau de Doutora
em Botânica.
______________________________________ Dra. Cecília Golçaves Costa _______________________________________ Dra. Cláudia Franca Barros _______________________________________ Dr.Fábio Rubio Scarano
_______________________________________ Dr. Flavio Nunes Ramos
_______________________________________ Dra. Maria Regina Torres Boeger
Rio de Janeiro, RJ – Brasil Fevereiro de 2008

iii
FICHA CATALOGRÁFICA
Mantuano, Dulce Gilson.
M291 Crescimento clonal em Neoregelia cruenta na Restinga de Jurubatiba : estrutura populacional, plasticidade morfo-anatômica e integração fisiológica. – Rio de Janeiro, 2008.
vii, 104 f. : il. Tese (doutorado) – Instituto de Pesquisas Jardim Botânico do Rio
de Janeiro/Escola Nacional de Botânica Tropical, 2008. Orientadora: Dra. Cecília Gonçalves Costa. Bibliografia. 1. Bromeliaceae. 2. Neoregelia cruenta. 3. Clonalidade. 4.
Restinga de Jurubatiba. I. Título. II. Escola Nacional de Botânica Tropical.
CDD 584.22

iv
Agradecimentos
Misteriosas são as razões que nos levam ao caminho árduo e tortuoso da busca do
conhecimento. Mais insondáveis ainda são as razões que levam pessoas ao nosso redor a
nos apoiarem e estimularem nesta escolha. Gostaria de agradecer a todos que facilitaram e
conspiraram para que essa tese fosse realizada e que neste intento tornaram este caminho
mais prazeroso.
De início gostaria de agradecer a minha orientadora, a pesquisadora Cecília
Gonçalves Costa, por manter em si sempre aceso o desejo da descoberta e ser capaz de
aceitar desafios. Esteve à todo momento aberta a minha opinião e desta forma me manteve
criativa e estimulada durante todo este estudo.
Agradeço ao meu co-orientador Gustavo Martinelli por me fazer estudar bromélias
e ter o prazer de descobrir o quão fascinantes elas são.
Os meus alunos Rodrigo Gadelha e Natalia Lorenzo que me mostraram que ensinar
e aprender são verbos que têm origem comum. Dividir com eles o prazer da conquista foi
inestimável.
Os meus amigos e colegas de Jardim Botânico, alunos da ENBT, em especial nas
pessoas de Marcelo da Costa Souza e Adriana Lobão, que por serem tão incríveis e
interessantes me fazem sentir que escolher ser botânico é poder participar de um mundo de
pessoas fantásticas.
À minha amiga espongóloga, Suzi Menezes Ribeiro, por compartilhar as
experiências da vida acadêmica e ter a melhor risada do mundo.
Agradeço a CAPES pela bolsa de estudos que permitiu a realização desta Tese, e
aos outros apoios financeiros recebidos durante a minha formação. Aos funcionários que
me receberam na secretaria da ENBT sempre com muito carinho e atenção: Márcia
Aparecida Betim Demby, Abílio Cardoso Lopes Filho, Januzia Fernandes Ferreira e Nilson
Guedes de Freitas.
Sou parte das minhas irmãs Ana e Clara e dos meus pais Heloisa e Carlos, portanto
este trabalho também é deles.
Por fim, sou grata ao David Angst por ter a coragem de vir me encontrar. O seu
companheirismo foi importante nos trabalhos de campo e na tranqüilidade e incentivo que
me trouxe a todo o momento. O seu amor foi fundamental por colocar felicidade em minha
vida e sonhos em meus pensamentos.

v
Resumo
Neoregelia cruenta é uma bromélia-tanque com metabolismo CAM e é amplamente
distribuída nas vegetações arbustivas abertas das planícies arenosas costeiras no sudeste do
Brasil. Nestas localidades atua como espécie-focal aumentando o conteúdo nutricional e a
retenção hídrica do solo, agindo como local de germinação para outras espécies. Apesar da
heterogeneidade ambiental das restingas, N. cruenta ocorre em abundância em ambos
micro-hábitats: exposto a irradiância total, ocorrendo fora das ilhas de vegetação;
parcialmente exposto a radiância total, ocorrendo na borda das ilhas de vegetação;
sombreado, ocorrendo dentro da ilha de vegetação. Os objetivos deste estudo foram (i)
examinar como os micro-hábitats afetam o crescimento e estrutura populacional de N.
cruenta, (ii) se a plasticidade fenotípica em parâmetros morfo-anatômicos podem ajudar a
explicar a alta abundância nos três micro-hábitats estudados e, (iii) buscar indícios da
integração fisiológica intraclonal. N. cruenta produziu novos rametes essencialmente
através da reprodução clonal. Rametes conectados crescendo sombreados apresentaram as
maiores taxas de crescimento, porém isto não refletiu nos maiores valores de recrutamento
entre os micro-hábitats, porque o tamanho dos rametes também foram os maiores. Os
rametes expostos foram fortemente afetados pelas condições ambientais e tiveram o
crescimento populacional negativo. Rametes conectados expostos em parte a irradiância
total foram capazes de sustentar o maior número de rametes por agregação e, apresentaram
a maior taxa de crescimento populacional, sugerindo que ambientes heterogêneos são
favoráveis ao crescimento clonal de N. cruenta. A disponibilidade hídrica foi o fator
preponderante na indução de variação nas características anatômicas estudadas, tais como,
alta densidade de tricomas peltados na face adaxial e aumento da espessura do parênquima
aqüífero concomitantemente ao estresse hídrico. Caracteres anatômicos não variaram em
resposta a condições de luminosidade, provavelmente porque rametes expostos e
parcialmente expostos estiveram fotoinibidos, como sugerido pelo decréscimo no conteúdo
de clorofila. Suculência foliar foi similar entre rametes parentais e filiais no micro-hábitats
heterogêneo, apesar da condutância estomática ser estatisticamente diferente. Este
resultado e o maior número de rametes sustentados por agregado sugere que a integração
intraclonal permite o movimento acropétalo do fluxo do xilema e resulta num aumento do
desempenho do agregado de rosetas clonais de N. cruenta.

vi
Abstract
Neoregelia cruenta is a tank bromeliad which presents CAM metabolism and is
widespread in the open scrub vegetation on Southeast Brazilian coastal sandy plains. In
this area it acts as nurse-plant increasing soil nutricional status and water retention and
acting as germination place to other species. Althought restingas presents high
environmental heterogeneity, N. cruenta has high abundances irrespective to micro-habitat:
exposed to full irradiance, outside vegetation islands; partially exposed to full irradiance, at
the border of vegetation islands; shaded, inside vegetation islands. This study purposes
were (i) to test if popultational growth and structure is affected by micro-habitats; (ii) to
test if phenotypic plasticity in morphological and anatomical parameters might help to
explain the high abundance in all three studied sites; (iii) to colect evidences of intraclonal
physiological integration. New ramets were stablished only through clonal reproduction.
Shaded conected ramets presented highest values of growth. However, it did not reflect in
the highest values of recruiting among micro-habitats because ramets size were also the
highest. Exposed ramets were strongly affected and had negative populational growth.
Conected ramets partially exposed to full irradiance were able to sustain larger number of
ramets per agregation, and presented the highest populational growth rate, suggesting that
heterogeneous habitat is favourable to N. cruenta clonal growth. Water availability was the
main factor to induce variation in the studied anatomical parameters such as high density
of peltate trichomes in adaxial surface and increasing in thickness of water parenchyma in
regard to irradiance exposure. Anatomical traits did not response to light condition,
probably because partially esposed and exposed ramets were photoinhibited, as shown in
decrease of chlorophyll content. Suculence were similar among mother and young ramets
in heterogenous micro-habitat, although stomatal condutance were statisticaly different.
This results and the higher number of ramets sustained per agregation suggest that
intraclonal integration allow xylem sap to flow acropetaly and leads to an increase of the
agregational fitness of conected ramets of N. cruenta..

vii
Sumário Agradecimentos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . iv Resumo. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .v Abstract. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . vi Capítulo 1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 01 1. Introdução. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 02
1.1. Especialização intraclonal em plantas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .07
1.2. Modelo do mecanismo fisiológico envolvido na especialização morfo-funcional
intraclonal. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 09
1.3. Modelo da especialização intraclonal em um ambiente de restinga. . . . . . . . . . . .10
1.4. Área de estudo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .13
1.5. Caracterização ambiental dos fragmentos clonais sob moitas de vegetação,
parcialmente sombreados e em área desnuda. . . . . . . . . . . . . . . . . . . . . . . . . . . . .15
1.6. Espécie de estudo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
Capítulo 2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24 2. Estrutura e Crescimento Populacional da bromélia clonal Neoregelia cruenta na
Restinga de Jurubatiba
2.1 Introdução . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .26
2.2 Material e Métodos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
2.2.1 Área de estudo. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .27
2.2.2 Amostragem da população. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
2.2.3 Delineamento esperimental e análise estatística. . . . . . . . . . . . . . . . . . . . . .30
2.3 Resultados. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
2.4 Discussão . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
Capítulo 3 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .40 3. Plasticidade intraclonal na anatomia foliar de Neoregelia cruenta em três micro-habitats
da Restinga Aberta de Clúsia
3.1 Introdução . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .42

viii
3.2 Material e Métodos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .43
3.3 Resultados . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .46
3.4 Discussão . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 54
Capítulo 4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .56 4. Variação intraclonal nos atributos morfológicos foliares, conteúdo de pigmentos e
condutância estomática de Neoregelia cruenta crescendo em três micro-habitats da
Restinga Aberta de Clúsia Aberta de Clúsia.
4.1 Introdução . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .58
4.2 Material e Métodos
4.2.1. Atributos morfológicos: Área Foliar, Espessura, Suculência e Massa
Específica Foliar . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .64
4.2.2. Pigmentos Fotossintéticos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .65
4.2.3. Condutância Estomática (Gs). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 67
4.2.4. Análise dos dados . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .68
4.3 Resultados
4.3.1. Atributos morfológicos: Área Foliar, Espessura, Suculência e Massa
Específica Foliar . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
4.3.2. Pigmentos Fotossintéticos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74
4.3.3. Condutância Estomática (Gs). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .77
4.4 Discussão
4.4.1. Atributos morfológicos: Área Foliar, Espessura, Suculência e Massa
Específica Foliar . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 80
4.4.2. Pigmentos Fotossintéticos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83
4.4.3. Condutância Estomática (Gs). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .85
Capítulo 5 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88 5. Conclusões gerais . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 89
Referências Bibliográficas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .93

1
Capítulo 1
Introdução geral

2
1. Introdução
A maioria dos ambientes naturais não é homogênea, mas sim um mosaico formado
com a combinação de manchas de recurso. Água, nutrientes e luz, assim como outros
fatores bióticos e abióticos que influenciam a distribuição e crescimento das plantas, não
são distribuídos uniformemente no espaço (Levin 1992). Diferenças significativas na
disponibilidade dos recursos podem ocorrer mesmo em pequena escala e, manchas
favoráveis e não-favoráveis de recursos podem ocorrer com diferentes tamanhos e
formatos.
A heterogeneidade ambiental pode, ainda, ser alterada por fatores bióticos e, assim
como o ambiente físico, determinar a composição e a ocorrência da vegetação.
Freqüentemente em ambientes estressantes, a amenização das condições abióticas
provocada por determinada(s) espécie(s) é um fator de ação fundamental na estruturação
da comunidade e na freqüência de ocorrência de outras espécies (Yarranton & Morrison
1974). A ação destas, também chamadas espécies-focais ou espécies-chave, pode ser
responsável até mesmo pela manutenção de um valor relativamente superior de diversidade
na vegetação, que na sua ausência seria fortemente afetado (Scarano 2002). Podem
aumentar a taxa de germinação, o crescimento, a sobrevivência e a distribuição geográfica
de plantas, que a elas se associam pela amenização de uma condição ambiental limitante
(p. ex. temperatura superficial do solo, Franco & Nobel 1989) ou ainda pelo saldo positivo
do conjunto de diversas alterações ambientais proporcionadas pela espécie-focal (para
revisão veja Callaway 1995). Desta maneira, nestas interações criam-se agregados
vegetacionais que ampliam a heterogeneidade ambiental.
A vegetação de restingas cresce sobre as planícies arenosas costeiras e, embora
apresentem uma taxa de endemismo baixa quando comparada a outras vegetações
tropicais, têm grande abundância e diversidade de espécies, com estruturas oligárquicas,

3
isto é, com eqüitabilidade baixa (Araujo 2000, Assunção & Nascimento 2000, Cordeiro,
2005, Pimentel 2002, Pereira, Cordeiro & Araujo 2004, Mantuano, 2003, Ormond 1960,
Zaluar 2002, Scarano, 2002). Existem algumas hipóteses para explicar a razão desta
fisionomia predominar nas restingas: (1) grande parte da flora advém de outras
comunidades igualmente diversas, a saber, Mata Atlântica e Cerrado, ou com grande
número de endemismos, Caatinga (Rizzini 1979, Araujo 2000). A similaridade florística,
ou de pares filogenéticos, existente entre as espécies da restinga e a mata atlântica,
certamente se relaciona com a proximidade entre os dois ecossistemas. Embora as
condições abióticas sejam muito distintas entre estas vegetações, espécies que previamente
apresentavam adaptações ao estresse hídrico e nutricional, tais como epífitas na mata
atlântica (p.ex.: espécies das famílias Bromeliaceae e Clusiaceae), puderam contribuir com
a composição florística das restingas. Já os outros dois ecossistemas comentados
anteriormente, partilham com as restingas o estresse hídrico como fator preponderante, isto
é, as espécies vegetais presentes nestes locais teriam adaptações prévias a resistirem e
colonizarem o ambiente recente das áreas costeiras (Braz 2002, Arruda 2003, Ferri 1955).
Na formação geológica das planícies de restinga contribuíram eventos de transgressão e
regressão marinhas, que vinham acompanhados de alterações climáticas (Suguio & Tessler
1984). Durante estes períodos, barreiras geográficas que isolam os três ecossistemas foram
anuladas ou reduzidas, e assim a introdução de espécies pôde ocorrer. Portanto, a grande
diversidade da vegetação costeira atual seria resultado da sua origem em múltiplos
ecossistemas.
(2) Em um ambiente em formação como as restingas, as espécies dos ecossistemas
originais que possuíssem fenótipo plástico poderiam contornar as diferenças ambientais, e
colonizar a costa (Rôças, Barros & Scarano 1997, Rôças, Scarano e Barros 2001,
Mantuano, Barros & Scarano 2006). Contudo, o tempo geológico recente da formação das

4
restingas (Holoceno; Martin et al 1993, Suguio & Tessler 1984) não teria permitido grande
número de especiações locais, refletidos no baixo endemismo.
(3) Modelos sucessionais para algumas comunidades da vegetação de restinga
relatam que algumas espécies-chave ou focais estão aptas a colonizarem a areia desnuda e
modificarem as condições ambientais, proporcionando sítio de germinação e
estabelecimento para outras espécies (Lacerda & Hay 1977, 1978, Hay & Lacerda 1980,
1984, Mantovani 2002, Andrade 1992, Zaluar 1997, 2002, Zaluar & Scarano, 2000). Este
mecanismo diminuiria a pressão seletiva sobre a entrada de espécies, uma vez que estas
podem desfrutar das condições ambientais amenizadas espacialmente próximas às
espécies-focais (Archer et al 1988, Archer 1989, Yarranton & Morrison 1974).
Vale ressaltar que os três mecanismos não são excludentes e podem estar correndo
concomitantemente.
Neoregelia cruenta (R. Graham) L.B. Smith é uma bromélia terrestre que pode ter
papel facilitador na comunidade da Restinga Aberta de Clúsia, isto é, pode amenizar o
ambiente (Hay & Lacerda 1980, Hay, Lacerda & Tan 1981, Lacerda & Hay 1982)
proporcionando local para germinação de outras plantas (Fialho & Furtado 1993, Zaluar
com. pess.). Tal como outras espécies da família, N. cruenta é capaz de se reproduzir
através do crescimento vegetativo (Benzing 2000). Assim, como espécie iniciadora de
ilhas de vegetação, N. cruenta não precisaria passar pelos processos de germinação e
estabelecimento na areia nua. O recrutamento está, majoritariamente, associado à
reprodução assexuada. De fato, Reinert (1995), Mantovani (2002) e Sampaio (2004)
relatam não terem observado, ou observado em rara freqüência plântulas de bromélias no
solo das restingas. Além das observações feitas neste estudo, que corroboram esta
afirmação. Todos os estudos tiveram duração de quatro anos e, portanto,
consecutivamente, permitem concluir que mesmo anos favoráveis em séries temporais

5
longas não são capazes de recrutar novos indivíduos de bromélias da restinga através da
reprodução sexuada, embora a formação de fruto e sementes ocorra anualmente.
Na comunidade Arbustiva Aberta de Clúsia ocorre outra importante espécie-
facilitadora, a Clusia hilariana. De porte arbustivo-arbóreo (média de altura para o
PARNA Jurubatiba entre 4-5 metros; Mantuano 2003), possui metabolismo CAM (Franco
et al. 1996, 1999). Estas duas características permitem que esta espécie se desenvolva bem
sob a restrição hídrica do solo arenoso das planícies costeiras, e que alcance porte,
avantajado se comparado com a maioria das outras espécies simpátricas. Assim, o espaço
formado abaixo da sua copa permite o desenvolvimento de outras espécies (Zaluar 2002,
Zaluar & Scarano 2000). Inclusive é possível encontrar uma relação direta e positiva entre
o tamanho da moita e a diversidade encontrada nas comunidades (Andrade 1992 para
Maricá; Zaluar 2002 para Macaé).
A heterogeneidade ambiental espacial nas restingas tem várias fontes de origem, e
já foram estudadas por vários autores. A proximidade com a costa forma um gradiente de
influência salina e deposição de salsugem no solo, que se ameniza para o interior do
continente (Pimentel 2002, Cordeiro 2005, Pereira, Cordeiro & Araujo 2004, e Reinert
1995 para Neoregelia cruenta). A formação geológica marcou a topografia em cordões
arenosos sucessivos e paralelos à linha do mar. Existe assim uma outra fonte de variação
ambiental entre os cordões (Pimentel 2002, Sampaio 2004, e Reinert 1995 para Neoregelia
cruenta). A proximidade com as lagoas costeiras também forma um gradiente ambiental de
disponibilidade hídrica (Pimentel 2002), bem como os regimes de alagamento dos “braços”
de lagoa (Kurtz, dados não publicados). Vale lembrar que outra forma de variação
ambiental ocorre em escala temporal, e também afeta processos na restinga (Mattos et al.
2004).
As influências de fatores bióticos são ainda uma outra fonte de variação ambiental.

6
Um exemplo marcante são as alterações impostas pela ocorrência de moitas de vegetação,
nas quais C. hilariana pode estar presente ou não. Sob a copa desta espécie é possível
encontrar redução da incidência luminosa, aumento do material particulado no solo (que
por sua vez aumenta a retenção hídrica e melhora as condições nutricionais), diminuição da
velocidade dos ventos (que diminui o déficit hídrico por impedirem a remoção da camada
limítrofe) e cobertura de folhedo sobre o solo (que diminui as temperaturas superficiais e
reduz a evaporação).
A heterogeneidade ambiental conceitua uma distribuição não-uniforme dos recursos
no espaço, e pode ocorrer tanto em variáveis acima do nível do solo, como abaixo dele. A
influência da distribuição dos recursos no tempo e no espaço é crítica para organismos
sésseis como os vegetais, e tem função fundamental em processos e fenômenos ecológicos
importantes (Levin 1992). Para plantas com crescimento clonal, a distribuição dos recursos
no espaço é ainda mais fundamental uma vez que estas apresentam estruturas de
forrageamento horizontal e, não somente vertical como a grande parte das plantas não-
clonais (Stuefer 1997, Hutchings & Wijesinghe 1997, Lovett Doust & Lovett Doust 1982).
A relação entre o tamanho das manchas de recursos e o tamanho que o conjunto de clones
conectados alcança, infere sobre como estes últimos “percebem” o ambiente, isto é, se
heterogêneo ou não. Em outras palavras, a heterogeneidade ambiental está condicionada ao
espaço ocupado e/ou utilizado pelo indivíduo.
Na vegetação Arbustiva Aberta de Clúsia, os conjuntos de rosetas conectadas
(estolão funcional) de Neoregelia cruenta podem ocorrer inteiramente no interior das
moitas de vegetação e inteiramente sobre a areia sem vegetação. Mas mais que isso, o
tamanho das rosetas e conexões da bromélia N. cruenta permitem que um único conjunto
conectado de rosetas possa ocupar a situação de borda da moita de vegetação, local em que
estão sujeitas à complementaridade das condições ambientais do interior e exterior das

7
moitas de vegetação.
O objetivo deste estudo foi analisar a variação intraclonal de características
morfológicas, anatômicas e fisiológicas das folhas de Neoregelia cruenta e o crescimento
clonal desta espécie-focal frente à variação ambiental gerada pelos agregados
vegetacionais dominados por C. hilariana; o que nos permitiu entender (1) como esta afeta
a sua dinâmica populacional, (2) sobre a potencialidade plástica desta espécie em se
aclimatar anatomicamente à heterogeneidade ambiental e (3) como o ambiente afeta a
estrutura e funcionamento das rosetas. Este tese propõe um modelo ecofisiológico pelo
qual os rametes clonais poderiam forragear ambientes heterogêneos e homogêneos, e
discute a confrontação do modelo com os paradigmas ecológicos e evolutivos conhecidos
para a população de N. cruenta na vegetação da Comunidade Arbustiva Aberta de Clúsia.
1.1. Especialização intraclonal em plantas
Plantas podem ajustar a aquisição de recursos do ambiente, modificando a biomassa
investida em cada órgão (Larcher 2000). Desta maneira mantém um equilíbrio funcional
entre as estruturas subterrâneas de aquisição de água e nutrientes e as partes aéreas
responsáveis pela fixação de carbono e exportação de fotossintatos. Este equilíbrio
funcional é estudado como a razão entre o peso das raízes e o peso das estruturas acima do
solo, tais como folhas e ramos (functional equilibrium Brouwer 1963, 1983).
Reconhecidamente, plantas crescendo em condições ideais tendem a apresentar valores
desta razão próximos a um (Taiz & Zeiger 1998, Larcher 2000, Nobel 1980). Órgãos de
reserva e alocação em reprodução podem afetar este balanço, contudo são, em geral,
excluídos da análise quando o objetivo é pesquisar o estado morfofuncional da aquisição
de recurso pelo indivíduo.
Quando os recursos são escassos no local, ou existe algum tipo de restrição

8
temporal, a biomassa investida é maior, ou seja, um indivíduo crescendo em localidades
áridas possui valores da razão parte subterrânea / parte aérea acima de um e, em situações
com sombra extrema, apresenta valores decimais. É importante perceber que na variação
ambiental existe um certo grau de acoplamento entre: +luz-água e –luz+água. Desta forma,
somente sendo um arranjo não-fatorial, a razão pode ser descritiva do funcionamento
vegetal. Em verdade, este acoplamento não é obrigatório e, sem dúvida existem locais
aonde esta premissa não é verdadeira. Para tanto, este fator estimador tem as suas
limitações práticas.
O investimento de biomassa direcionado ao aumento da eficiência de aquisição dos
órgãos relacionados ao recurso limitante é uma assertiva apenas válida para plantas não-
clonais, ou plantas clonais sujeitas à ambiente homogêneo. Intrigantemente, plantas clonais
em cujos clones conectados estão sujeitas a condições heterogêneas investem biomassa nos
órgãos localizados nos recursos não-limitados. Assim, se especializam em obter o recurso
abundante. Novamente, a noção de complementaridade é fundamental porque ao final o
indivíduo clonal (composto de dois ou mais clones conectados) precisa manter um balanço
funcional. Em outras palavras, a aquisição dos recursos pelo somatório das biomassas dos
clones investidas em parte subterrânea precisa equivaler à capacidade de metabolização do
somatório das biomassas investidas nas partes aéreas. Este fenômeno de inversão da
especialização dentro de conjuntos de clones é conhecido por Division of Labour (Stuefer,
During & de Kroon 1994, Stuefer, de Kroon & During 1996, Stuefer 1996, Alpert &
Stuefer 1997, Hutchings & Wijesingh 1997).
Clones (ou rametes) crescendo conectados em ambientes heterogêneos podem
apresentar fitness (expresso em taxa de crescimento ou fixação de carbono) maior que
clones de igual tamanho crescendo em ambientes homogêneos. Os órgãos quando
especializados na função de adquirir o recurso local acabam por criar um excedente de

9
produção, tal qual modelos econômicos de produção prevêem, por isso a analogia com
Division of Labour (Josef 1997). Este excedente de produção prevê que ambientes
heterogêneos, portanto, são favoráveis ao crescimento de plantas clonais capazes de
translocar recursos (Hutchings & Wijesinghe 1997).
1.2. Modelo do mecanismo fisiológico envolvido na especialização morfofuncional
intraclonal
O ajuste do balanço funcional do investimento em biomassa em cada órgão é
direcionado e estimulado pelas condições ambientais em que o vegetal cresce. Os ramos
dentro de um mesmo indivíduo em uma planta não-clonal ajustam as taxas de crescimento
e relação disponibilidade de luz, permitindo o melhor forrageamento do ambiente luminoso
através do crescimento diferencial. Ao receber mais radiação os ramos têm um aumento da
taxa transpiracional foliar e recebem ao final maior aporte de xilema. Dois mecanismos,
então, passam a estimular e subsidiar a aceleração do crescimento: (1) o maior aporte de
água e nutrientes e (2) as regiões da fronde com altas taxas de transpiração recebem maior
volume de hormônio de crescimento da parte aérea (Citocininas) que é produzido nas
raízes e carreado pelo xilema. Recentemente, alguns estudos mostraram que as taxas de
assimilação fotossintética são afetadas pela ação deste hormônio (Pons Jordi & Kuiper
2001).
A produção dos fotossintatos é distribuída no vegetal através do floema e segue
algumas premissas para as prioridades de alocação. Tipicamente o fluxo segue uma via
entre produção e consumo (Davies 1987, Taiz & Zeiger 1998) e, tecidos em
desenvolvimento são beneficiados. Órgãos reprodutores e de reserva também apresentam
fluxo direcionado, provavelmente induzido por hormônios. O investimento em crescimento
das partes subterrâneas pode estar envolvido com uma sinalização associada ao teor de

10
nitrogênio absorvido pelo solo, que é redirecionado para um sinal hormonal (em grande
parte associado à ação das auxinas) e induz o fluxo do transporte ativo do floema. Embora,
o mecanismo completo de direcionamento do fluxo do floema não seja completamente
conhecido na literatura, o envolvimento do nitrogênio como um sinalizador do ambiente é
aceito amplamente (Lamber et al. 1998, Taiz & Zeiger 1998).
Assim, para o mecanismo de especialização intraclonal é necessário haver
ambientes que promovam taxas transpiracionais e taxas de absorção de nitrogênio
diferenciadas entre clones. O mecanismo que segue abaixo é um modelo comparativo da
fisiologia de plantas não-clonais crescendo sob condições ambientais específicas e, é
proposto um modelo de funcionamento da especialização intraclonal. As premissas desta
proposição levam em consideração que há a translocação de recursos nos feixes vasculares
entre os clones e, que alterações de ajuste nas características morfofuncionais
(aclimatação) podem alterar o modelo (Figura 1.1).
1.3. Modelo da especialização intraclonal em um ambiente de restinga
As bromélias crescendo na condição de borda das moitas de vegetação foram
relatadas como tendo um possível papel na expansão da área de cobertura das moitas na
comunidade Arbustiva Aberta de Clúsia (Mantovani 2002). Nesta situação o conjunto de
rosetas está em parte sujeito à alta irradiância e altas taxas de evaporação e, em parte
sujeita a menores irradiâncias luminosas, temperaturas e demandas evaporativas. Rosetas
que estejam expostas podem estar sendo subsidiadas pela aquisição de água de rosetas em
situações protegidas. O transporte de água, caso exista a translocação através do estolão de
conexão, pode estar sendo direcionado por diferenças nas taxas de transpiração.
Nas comunidades abertas das planícies costeiras do sudeste, das quatro bromélias
mais freqüentes nas restingas Neoregelia cruenta, Aechmea nudicaulis, Tillandsia stricta e

11

12
Baixa irradiância luminosa Baixa demanda evaporativa do ar
Alta irradiância luminosa Remoção da camada limítrofe
Longo período de retenção de água no tanque
Curto tempo de retenção de água no tanque
Vriesea neoglutinosa (Cogliatti-Carvalho et al. 2001), as três primeiras apresentam
metabolismo CAM (Martin 1994, Medina et al. 1977, Griffiths 1998, Reinert 1995).
Portanto, maximizam a abertura estomática durante o período noturno, período em que as
diferenças na temperatura foliar são menores. Contudo, as taxas de transpiração não
necessariamente são similares entre o interior e o exterior das moitas de vegetação, haja
vista que a proteção do dossel diminui a ação de ventos e a remoção da camada limítrofe
de circulação de ar na superfície foliar. Assim, mesmo tendo metabolismo CAM, é
possível que rametes crescendo na condição borda-exposto sejam sustentados pela
aquisição de água dos rametes na condição borda-protegido. O tempo de retenção da água
no copo e área de contato entre a superfície foliar são maiores também nos rametes nas
situações interior e borda-protegido. O modelo que segue abaixo é um mecanismo
hipotético do funcionamento de conjuntos de rosetas de bromélias CAM clonais
conectadas forrageando as condições ambientais criadas pelo sombreamento por moitas de
vegetação no solo arenoso de restinga. As premissas desta proposição levam em
Figura 1.2. Modelo de translocação de água e nutrientes através do xilema de feixes
vasculares do estolão entre rosetas de bromélias clonais submetidas à condições ambientais
heterogêneas (movimento acropétalo).

13
1.4. Área de Estudo
Os ecossistemas de restingas são formados por um mosaico de vegetações que
crescem sobre a planície arenosa e por lagoas costeiras. O substrato das planícies costeiras
é oriundo da deposição de sedimentos marinhos e fluviais, que ocorreram intensamente
durante os eventos de regressão e transgressão marinhas no período do Quaternário, indo
até o Holoceno (Martin et al. 1993). A deposição destes sedimentos em faixas de areia
alongadas delimitou as lagoas costeiras e gerou no perfil topográfico uma série de cordões
arenosos seguidos de depressões.
O estudo foi desenvolvido no Parque Nacional da Restinga de Jurubatiba (Figura
1.3) que está situado no litoral norte do estado do Rio de Janeiro, entre as coordenadas
22º00’-22º23’S; 41º15’-41º45’O. O macroclima da região é classificado como Aw –
Tropical Chuvoso segundo a escala de Köppen (1948). A temperatura média anual é de
25,6ºC, a temperatura máxima (29,7ºC) ocorre em janeiro e a mínima (20,0ºC) no mês de
julho (Henriques et al. 1986). A pluviosidade é fortemente sazonal, marcando o verão
chuvoso e o inverso seco. A pluviosidade média anual está entre as isoietas de 1100 e
1300mm, sendo a mínima mensal de 41mm no inverno e a máxima de 189mm no verão
(Araujo et al. 1998). A passagem de frentes frias pela região durante os meses de inverno
pode amenizar temporariamente o período de seca (Dau 1960). A disponibilidade hídrica
para a vegetação, entretanto, pode ser afetada pelas características de retenção do solo e
pela acessibilidade das espécies ao lençol freático. Os solos arenosos, como os das
planícies costeiras, apresentam tipicamente baixa capacidade de retenção de água. Assim,
nos períodos após as chuvas, quando o solo está saturado, as plantas podem ter acesso a
uma grande quantidade de água, contudo, nas horas seguintes ocorre um rápido decaimento
no teor hídrico do solo. O déficit hídrico no solo é acentuado durante os meses de julho e
agosto (Henriques et al. 1986). O lençol freático é superficial e aflora durante o período da

14
estação chuvosa nas depressões topográficas da planície.
O efeito da sazonalidade também pode ser observado na incidência de ventos. O
vento de maior freqüência para a região é o alísio de nordeste (ENE) que incide durante
todo o ano, mas é predominante em janeiro. O segundo vento de maior freqüência e
intensidade é o sudoeste (SSO). Este vento, decorrente da chegada de frentes frias, é
responsável pelas tempestades e ocorrência das mais baixas temperaturas na região. Suas
maiores intensidades são observadas nos meses de julho e agosto, quando alcança
4,3m/seg. Os outros ventos incidentes na região possuem uma freqüência bastante menor,
sendo uma ordem de grandeza abaixo destes (Dau 1960, Martin et al. 1993).
A composição florística das restingas está intimamente associada à ação dos fatores
ambientais, que apresentam uma ampla escala de variação temporal e espacial, chegando
muitas vezes a extremos limitantes à ocorrência das espécies. A combinação destes fatores,
que podem ainda variar em freqüência e em intensidade, determina a comunidade vegetal
predominante, formando um mosaico de ambientes.
A vegetação da região compreende pelo menos doze comunidades vegetais
distintas, ocorrendo de comunidades aquáticas nas lagoas até matas de restinga (Araujo &
Henriques 1984, Araujo et al. 1998). Os estudos de dinâmica populacional foram
desenvolvidos na comunidade Arbustiva Aberta de Clúsia (sensu Araujo 1998) que ocorre
adjacente à Lagoa Comprida (Figura 1.3). O material foliar coletado para mensuração de
parâmetros morfológicos, quantificação de pigmentos e para estudos anatômicos teve
origem mais ampla e foi aleatoriamente amostrado nas comunidades de Clúsia distribuídas
em toda a extensão do Parque Nacional da Restinga de Jurubatiba. Esta formação ocorre
sobre os cordões arenosos internos e é constituída de moitas densas de tamanhos variados,
intercalados por solo desnudo, onde a cobertura vegetal é esparsa (Araujo & Henriques
1984, Araujo 1998). A ocorrência de folhedo está restrita as manchas de vegetação. As

15
moitas têm formato aproximadamente hemisférico e variam de tamanho de <1m2 até
>100m2 (Andrade 1992, Zaluar 1997). As moitas menores são as de maior freqüência
(<5m2), contudo, somente a partir de 5m2 o número de espécies aumenta conforme o
volume (Andrade 1992). Na maior parte das moitas, o dossel é dominado apenas pela copa
de Clusia hilariana, que está posicionada no centro e atinge a altura média de 4 metros.
Entretanto, algumas moitas não apresentam a ocorrência de C. hilariana. Neste caso, o
dossel misto é dominado por Protium icicariba, Myrsine parvifolia, Eugenia umbeliflora,
Myrcia lundiana e Erythroxylum sp., que também são espécies freqüentes nas moitas
juntamente com C. hilariana (Pimentel 2002). Nas moitas de pequeno porte as espécies
mais freqüentes são Allagoptera arenaria, Pilosocereus arrabidae, Aechmea nudicaulis e
Neoregelia cruenta. Estas espécies também ocorrem com alta freqüência nas moitas
dominadas por C. hilariana.
1.5. Caracterização ambiental dos fragmentos clonais sob moitas de vegetação,
parcialmente sombreados e em área desnuda.
Para este estudo foram estabelecidas três categorias de micro-hábitats na
comunidade Arbustiva Aberta de Clúsia, de acordo com o grau de sombreamento e
associação com a espécie-focal C. hilariana. Os rametes conectados de Neoregelia cruenta
estudados ocorreram em três micro-hábitats classificados como (1) exterior – rosetas
crescendo ao sol pleno sobre a areia desnuda, (2) borda – rosetas crescendo parcialmente
sombreadas por estarem sob a copa de C. hilariana ou sombreadas por um período do ciclo
diário, (3) interior – rosetas crescendo dentro de moitas de vegetação completamente
sombreadas por dossel dominado por C. hilariana (Figuras 1.4a-c).
O fator ambiental que tem a maior amplitude de variação entre os três micro-
hábitats é, sem dúvida, a condição luminosa. A situação interior apresenta máximo de

16
radiação incidente de aproximadamente 200µmol.m-2.s-1, com alguma variação entre as
moitas de cerca de 50µmol.m-2.s-1. Indivíduos crescendo parcialmente sombreados estão
sujeitos a um ambiente fortemente heterogêneo quanto ao fornecimento de luz. O sentido
de orientação em relação ao centro da moita e a posição em relação à linha de borda da
vegetação determina ao final a quantidade total de energia luminosa recebida. Os máximos
variaram entre 400-800µmol.m-2.s-1. A radiação que atinge os indivíduos completamente
expostos é expressiva com máximos entre 1750-2000µmol.m-2.s-1 em dias sem
nebulosidade (Figura 1.5a). A temperatura do solo e, conseqüentemente, a velocidade de
evaporação da umidade do solo acompanham a quantidade de radiação recebida e,
portanto, no microclima do interior das moitas as temperaturas da superfície do solo são
menores do que as da borda, e estas por sua vez, menores que o exterior (Figura 1.5b). O
teor de água encontrado no solo é maior no interior das moitas de C. hilariana devido a
maior proporção de matéria particulada além da matriz arenosa. A capacidade de campo no
solo (retenção hídrica máxima após a saturação do solo) sob C. hilariana adulta é de 15%
(g H20 retida em 100g de solo). Enquanto o valor de capacidade de campo encontrado para
solo sem cobertura é de apenas 1%. Apesar de a amplitude de temperatura variar
significativamente entre o microclimas, é provável que este fato tenha pouca atuação sobre
Neoregelia cruenta. Embora seja uma espécie herbácea e cresça próximo ao solo, é capaz
de formar tanque de reserva de água com a base das folhas da roseta. As temperaturas
medidas na água do tanque das rosetas mostram que mesmo estando sujeitas ao
aquecimento a sua amplitude é muito menor (máximo de temperatura do solo exterior =
50ºC, máximo de temperatura encontrado na água do tanque de rosetas crescendo no
exterior = 32ºC; Figura 1.5c). É provável que esta seja uma proteção significativa para o
meristema central, e que situações de seca e excesso de radiação, em anos atípicos, seja um
perigo potencial para a sobrevivência dos indivíduos, como documentado por Reinert

17
(1995, pág.72).
A temperatura ambiente no microclima sob as moitas com Clusia hilariana e C.
fluminensis estudadas para a restinga de Cabo Frio é menor do que no espaço desnudo
(Dau 1960). Embora a temperatura ambiente não tenha sido medida para a vegetação
arbustiva aberta de Clúsia do Parque Nacional da Restinga de Jurubatiba é bastante
provável que o comportamento seja similar. A umidade relativa do ar varia com as
características atmosféricas, mas nas restingas as maiores flutuações são oriundas da
variação diária de temperatura (Dau, 1960), à exceção de Cabo Frio que possui incidência
constante de massas de ar seco associadas ao padrão climático local (Barbière, 1984). Em
Jurubatiba na estação seca as amplitudes diárias podem ser de até 70% (Figura 1.5d). A
diferença encontrada na umidade relativa do ar no interior e exterior das moitas de
vegetação dependerá, principalmente, da movimentação atmosférica. As médias tendem a
ser maiores no interior dos agregados vegetacionais que têm efeito de tamponamento na
variação diária de temperatura e em mudança das características do ar. Dau (1960)
observou médias mais altas em moitas dominadas por espécies de Clusia, seguidas de
moitas com o dossel misto e por fim, o menor valor é relativo à umidade no espaço
desnudo. Estes padrões para temperatura, umidade relativa do ar e evaporação foram
observados durante o dia, e tendem a se inverter durante a noite, pela proteção que a
cobertura vegetal confere à incidência de ventos (Dau 1960).
Os micro-hábitats interior e exterior foram categorizados como homogêneos porque
os fragmentos clonais (conjunto de rosetas conectadas) crescem sob a mesma condição
ambiental, enquanto o micro-habitat borda foi considerado heterogêneo porque os rametes
clonais conectados crescem sujeitos a condições distintas de luz, temperatura, umidade
relativa, disponibilidade de nutrientes no copo e no solo, e demanda evaporativa do ar (veja
Figura 2.1).
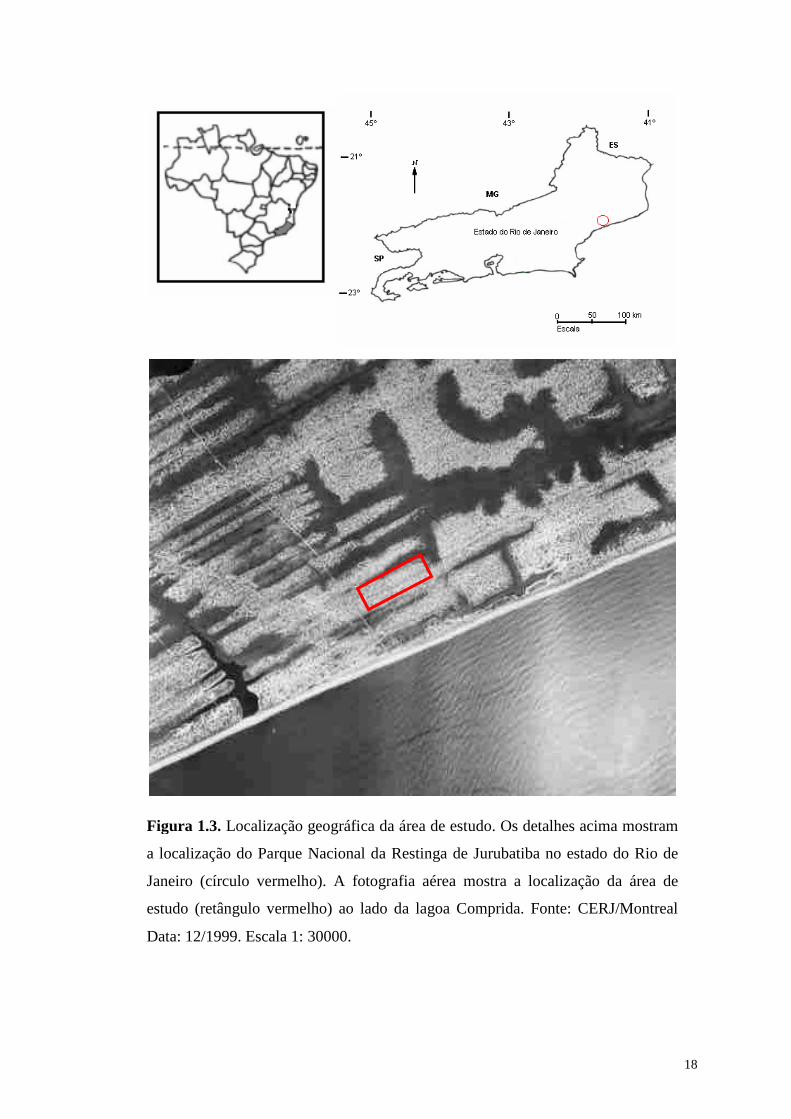
18
Figura 1.3. Localização geográfica da área de estudo. Os detalhes acima mostram
a localização do Parque Nacional da Restinga de Jurubatiba no estado do Rio de
Janeiro (círculo vermelho). A fotografia aérea mostra a localização da área de
estudo (retângulo vermelho) ao lado da lagoa Comprida. Fonte: CERJ/Montreal
Data: 12/1999. Escala 1: 30000.

19
Fig
ura
1.4.
Neo
reg
elia
cru
en
ta (R
. G
raha
m)
L.B
.Sm
ith (
Bro
mel
iace
ae)
cres
cend
o em
trê
s di
stin
tos
mic
ro-h
ábita
ts c
riado
s pe
lo
posi
cion
amen
to d
os in
diví
duos
em
rel
ação
as
moi
tas
de v
eget
ação
na
Com
unid
ade
Arb
ustiv
a A
bert
a de
Clú
sia
do
Par
que
Nac
iona
l da
Res
tinga
de
Juru
batib
a.
a. E
xpos
to a
sol
ple
no (
Ext
erio
r); b. P
arci
alm
ente
som
brea
do e
pa
rcia
lme
nte
expo
sto
(Bor
da
); c
. Som
brea
do
(Int
erio
r).
a
b
c

20
Figura 1.5. Variáveis climáticas estimadas para caracterizar os micro-hábitats da Restinga
Aberta de Clúsia, no Parque Nacional da Restinga de Jurubatiba, RJ. a. Radiação
fotossinteticamente ativa (µmol.m-2.s-1); b. Temperatura à superfície do solo (ºC); c.
Temperatura medida na água contida no tanque das rosetas de Neoregelia cruenta; d.
Umidade relativa do ar (%). As mensurações foram feitas em setembro/2007.
a b
c d

21
1.6. Espécie de estudo
Nas restingas fluminenses ocorrem cerca de 55 espécies da família Bromeliaceae,
entretanto apenas 9 têm ampla distribuição no Estado (isto é, ocorrem em pelo menos 8 das
10 regiões fitogeográficas definidas por Araujo, 2000), dentre elas encontramos Neoregelia
cruenta. Esta bromélia ocorre nos ecossistemas de Restingas da Costa Atlântica Sudeste
(Araujo, 2000), entre os estados do Espírito Santo e São Paulo. No Estado do Rio de
Janeiro ocorre em São João da Barra, Jurubatiba (Macaé/Carapebus/Quissamã), Barra de
São João, Cabo Frio/Arraial do Cabo, Marica, Jacarepaguá (fitoregião que abrangia as
Restingas da cidade do Rio de Janeiro), Grumari e Marambaia (Araujo, 2000). Só não
ocorre na Reserva Biológica da Praia do Sul, município de Angra dos Reis que
compreende ambiente insular (Nunes-Freitas et al., 2006). Embora com distribuição
restrita ao sudeste brasileiro (Figura 1.6), N. cruenta pode ser encontrada em abundância
localmente, principalmente crescendo terrestre nas comunidades arbustivas abertas (Araujo
& Henriques, 1984).
N. cruenta é uma bromélia-tanque clonal com crescimento simpodial definido, de
metabolismo CAM, pertencente à subfamília Bromelioideae. A reprodução assexuada
produz rosetas perenes monocárpicas e o período de floração ocorre no início da estação
chuvosa (Outubro – Dezembro) e a frutificação no final da mesma estação (Janeiro –
Março). Pode crescer ocupando os hábitos epifítico e terrestre, nestes casos com
crescimento ortotrópico e plagiotrópico, respectivamente. O hábito preferencial de N.
cruenta é terrestre, sendo epífita eventual, em especial nas matas de Restinga. Na
localidade de Maricá grande parte das rosetas de N. cruenta é terrestre (82-92%), com
pequena porção epífita (8-18%; Lacerda & Hay, 1982). O mesmo ocorre para a Restinga
Aberta de Clúsia no Parque Nacional da Restinga de Jurubatiba.

22
N. cruenta possui raízes fixadoras e, provavelmente, parcialmente funcionais; o
conjunto das bainhas foliares forma tanque bem desenvolvido e apresenta tricomas
peltados de absorção, principalmente na base dos limbos foliares (classe tipo III; Benzing,
2000; Pittendrigh, 1948). A arquitetura desta espécie motivou estudos sobre o possível
aprisionamento de nutrientes oriundos da salsugem que seriam incorporados através da
água do tanque e conduzidos ao solo com a senescência foliar (Lacerda & Hay, 1977;
1978; Hay & Lacerda, 1980). Contudo, a melhora no teor nutricional do solo, expresso no
teor dos nutrientes em si, ou na Capacidade de Troca Catiônica, relacionou-se
positivamente com o teor de matéria orgânica (Hay & Lacerda, 1980; Mantovani, 2002). A
salsugem contribui para a entrada de nutrientes nas restingas (Hay & Lacerda, 1984),
porém o aprisionamento deste, expresso pelo teor do cátion Na+, é independente da
espécie, sendo inclusive similar entre três diferentes espécies de bromélias (Mantovani,
2002). Entretanto, N. cruenta parece contribuir significativamente para o conteúdo
nutricional do solo, em especial os macronutrientes K e P, uma vez que altera do teor de
matéria orgânica, e favorece a retenção da água que lava o salsugem aprisionado na
vegetação.
As alterações das condições edáficas e microclimáticas proporcionadas por N.
cruenta foram usadas como argumento do papel facilitador que esta espécie tem em
ecossistemas de Restingas (Lacerda & Hay, 1977; 1978; Hay & Lacerda, 1980; Mantovani,
2002). Outro papel ecológico focal relatado para N. cruenta é como sítio de germinação de
sementes para espécies formadoras de moitas como Erythroxylum ovalifolium Pey. em
Maricá (Andrade, 1992; Fialho & Furtado, 1993), e Clusia hilariana Schl. em Macaé
(Zaluar & Scarano, 2000; Zaluar, 2002). Motivados pela abundância de N. cruenta nas
vegetações de Restinga do sudeste, ou pelo seu possível papel facilitador, esta espécie
recebeu considerável atenção na literatura (Lacerda & Hay, 1977; 1978; 1982; Hay &

23
Lacerda, 1980; Hay, Lacerda & Tan, 1981; Mantovani & Iglesias, 2001; Mantovani, 2002;
Reinert, 1995; Fernandes, Chaloub & Reinert, 2002, Fialho & Furtado,1993; Zaluar &
Scarano, 2000). Além dos trabalhos que tratam de fauna associada ao tanque (Oliveira,
Rocha & Bagnall, 1994; Oliveira & Rocha, 1997).
Figura 1.6. Distribuição geográfica da bromélia-tanque Neoregelia cruenta (R. Graham)
L.B.Smith, evidenciando o padrão “Costa Atlântica Sudeste/Sul” sensu Araujo (2000).
60 55 50 45 40 35 30 25 20 65 70 75 80 85 90 95 100 105
0 5 10
15 20
25 30
5 10 15
20 25
30 35

24
Capítulo 2
Variação espacial na reprodução
clonal de Neoregelia cruenta

25
2. Estrutura e crescimento populacional da bromélia clonal Neoregelia cruenta na
Restinga de Jurubatiba
Mantuano, D.G.; Costa, C.G. & Martinelli, G.
Abstract
Clonal reproduction is a common feature in herbaceous plants and has been shown to be important for recruitment and population maintenance. Although numerous phenological responses have been shown to be associated with seasonality and local climate factors in the tropics, little is known about how it affects levels of clonal reproduction. I examined the temporal and spatial variation in clonal reproduction for a endemic nurse-plant, Neoregelia cruenta (R. Graham) L.B. Smith (Bromeliaceae). I adressed the following questions (1) How do vegetative growth and clonal reproduction vary among seasons?; (2) Does clonal reproduction differs between homogenous microhabitats (fully exposed to sun light and fully shaded) and hetegeneous habitats (partially exposed)?; and if so (3) Has heterogeneous michohabitat higher population growht rates?. Rosettes (or ramets) demography, phenological status and size classes were followed in 30 ramets clumps for 18 months. Ramets vegetative growth rates and mortality were found to be highly influenced by seasonality. During the transition from wet to dry season there was a significant decrease in ramets growth rate. Highest number of conected ramets (4,3 per clump) and frequency of ramets in population (72,3%) were found in partially exposed microhabitat. Although fully exposed ramets produced the highest number of new ramets, high mortality imposed a negative ramets populational growth to this microhabitat (-0,38 ramet per clump.year-1). Shaded and partially exposed microhabitat presented similar rates (0,80 and 0,90 ramets per clump.year-1, respectively). Results from this study suggest that vegetative growth and clonal reproduction of N. cruenta responds negatively to light levels increase. In shaded conditions, carbon investments shift from clonal reproduction to vegetative growth.
Resumo
Reprodução clonal é uma característica encontrada comumente em plantas herbáceas e tam sido referida como importante para o recrutamento e manutenção populacional. Embora numerosas respostas fenológicas tenham sido associadas a sazonalidade e a fatores climáticos locais nos trópicos, pouco se sabe sobre como estes fatores afetam a intensidade da reprodução clonal. Neste estudo se examinou a variação temporal e espacial na reprodução clonal de uma planta-focal endêmica, Neoregelia cruenta (R. Graham) L.B. Smith (Bromeliaceae). Foram estudadas as seguintes questões: (1) Como o crescimento vegetativo e a reprodução clonal varia entre estações?; (2) A reprodução clonal difere entre micro-hábitats homogêneos (exposto ao sol pleno e sombreado) e micro-hábitat heterogêneo (parcialmente exposto)?; e em caso afirmativo, se (3) O micro-hábitat heterogêneo apresenta a maior taxa de crescimento populacional?. A demografia dos rametes, o status fenológico e as classes de tamanho foram acompanhadas em 30 agregados de rametes conectados por 18 meses. As taxas de crescimento vegetativo dos rametes e a mortalidade foram fortemente influenciadas pela sazonalidade. Durante a transição da estação chuvosa para a estação seca ocorreu significativo decréscimo das taxas de crescimento de rametes. O maior número de rametes conectados (4,3 por agregado) e a maior freqüência de rametes na população (72,3%) foram encontrados no micro-hábitat parcialmente exposto. Apesar dos rametes expostos produzirem o maior número de novos rametes, a alta mortalidade impôs uma taxa de crescimento populacional negativa para este micro-hábitat (-0,38 rametes por agregado.ano-1). Micro-hábitats sombreado e parcialmente exposto apresentaram taxas similares (0,80 e 0,90 rametes por agregado.ano-1, respectivamente). Os resultados destes estudo sugerem que o crescimento vegetativo e a reprodução clonal de N. cruenta responde negativamente ao aumento dos níveis de luminosidade. Na condição sombreada, o investimento de carbono é transferido da reprodução clonal para o crescimento vegetativo.
Palavras-chaves: Neoregelia cruenta, reprodução clonal, luz, variação espacial, Restinga.
Key words: Neoregelia cruenta, clonal reproduction, light, spatial variation, Restinga.

26
2.1. Introdução
O crescimento clonal em plantas resulta no recrutamento de novos indivíduos,
geneticamente idênticos, que tem a sua fase inicial de desenvolvimento subsidiada pelo
indivíduo parental. Neoregelia cruenta é uma bromélia clonal que ocorre em abundância
nas restingas do Rio de Janeiro (Araujo, 2000). Nestas localidades pode ter papel focal
para a vegetação, tendo sido documentada como um possível sítio de germinação para
outras espécies lenhosas simpátricas (Fialho & Furtado 1993), espécie iniciadora de
sucessão (Andrade, 1992) e, capaz de enriquecer o teor nutricional do solo (Lacerda &
Hay, 1977; 1978; Hay & Lacerda, 1980).
As restingas são um ambiente constituído de um mosaico de tipos de vegetações
definidos principalmente pelo arranjo dos fatores ambientais estressantes que ocorrem nas
localidades, tais como salinidade, alagamento e seca (Scarano, 2002). Em contribuição à
heterogeneidade ambiental, a comunidade Arbustiva Aberta de Clúsia é formada por
moitas de vegetação separadas por areia nua. Populações de N. cruenta são abundantes no
interior e borda das moitas de clúsia. Apesar de menos freqüente, também é possível
observar conjuntos de rosetas conectadas (doravante denominados segmentos clonais)
ocorrendo sobre a areia sem cobertura. Uma importante questão relacionada a ambientes
com recursos distribuídos em manchas ou com flutuações sazonais é como estes fatores
afetam a estrutura e dinâmica da população. Os estudos que descrevem este fenômeno para
espécies de reprodução clonal ainda são escassos, especialmente para bromélias nos
trópicos (Garcia-Franco & Rico-Gray 1995, Sampaio, 2004; Souza, 2004, Villegas 2001).
A integração fisiológica e especialização intraclonal entre indivíduos conectados
são fenômenos que permitem a colonização de ambientes limitantes e heterogêneos (Alpert
& Mooney, 1986; Stuefer, During & de Kroon, 1994; Hutchings & Wijesinghe, 1997). De
fato, plantas clonais são capazes de produzirem mais biomassa e exibir maior crescimento

27
em localidades que indivíduos clonais conectados estejam submetidos a condições
ambientais distintas (Hutchings & Wijesinghe, 1997). A escala da variação ambiental e o
tamanho dos segmentos clonais, e o comprimento do estolão (espaçador) são determinantes
na percepção da heterogeneidade ambiental pelos indivíduos conectados.
Os objetivos deste estudo são: (1) descrever a estrutura e a dinâmica da população
de N. cruenta, (2) quantificar os efeitos da variação temporal e espacial do ambiente nas
taxa de crescimento vegetativo e da reprodução clonal desta espécie numa restinga do
Norte Fluminense e, (3) verificar se o ambiente heterogêneo produz um crescimento
favorável de modo a subsidiar a hipótese de que ocorre integração fisiológica entre as
rosetas.
2.2. Material e Métodos
2.2.1 Área de estudo
O estudo foi desenvolvido na comunidade classificada como Arbustiva Aberta de
Clúsia da vegetação de Restinga do segundo cordão arenoso próximo à Lagoa Comprida
no Parque Nacional de Jurubatiba - RJ (22º00’-22º23’ S; 41º15’-41º45’ O). O clima da
região é classificado como Aw – Tropical Chuvoso (Köppen, 1948). A precipitação anual é
de 1164mm, com duas estações distintas, o verão quente e úmido (out-abr) e o inverno
seco (mai-set). A temperatura média anual é 25,6ºC, com média mensal máxima de 30,8ºC
e mínima de 20,2ºC (para mais detalhes veja tópico 1.4).
2.2.2. Amostragem da população
Um segmento clonal foi considerado como um conjunto de rosetas (ou rametes) que
permanecem conectados por um estolão funcional. Esta categoria foi admitida como a
unidade amostral mais amplamente utilizada na dinâmica populacional de estudos de

28
espécies que apresentam reprodução vegetativa. Os segmentos clonais que distavam menos
de 50cm de outro foram considerados indivíduos genéticos idênticos. Cada roseta (ou
ramete) conectado, que possui potencial para existência individual após a fragmentação,
foi considerado um indivíduo fisiológico. O tamanho mínimo de um segmento clonal foi
estabelecido como sendo de dois rametes vivos e o número máximo não foi limitado. As
rosetas parentais e filiais foram determinadas a partir da direção do crescimento de novas
rosetas no segmento. As conexões entre os rametes são aparentes e, em geral são
encontradas ao nível do solo. Foram consideradas na amostragem as rosetas parentais
mortas, indicadas pelos vestígios de rametes em conexões que persistiram.
A quantificação do crescimento populacional foi realizada em dez moitas de
vegetação selecionadas por apresentaram simultaneamente segmentos clonais de
Neoregelia cruenta nos três micro-hábitats: interior, borda e exterior (para descrição
abiótica destes, veja tópico 1.5). Os micro-hábitats interior e exterior foram categorizados
como ambientes homogêneos, i.e. onde todas as rosetas do segmento clonal estavam
expostas à mesma condição ambiental, respectivamente, sombra e maior disponibilidade de
água e nutrientes, e alta exposição luminosa e restrição de água e nutrientes. O micro-
hábitat borda foi caracterizado como ambiente heterogêneo, tendo sido selecionados
segmentos cujo crescimento das rosetas jovens tinha orientação radial em relação ao centro
da moita de vegetação e, portanto, estavam expostos a condições de maior luminosidade do
que os rametes parentais (Figura 2.1). As diferenças microclimáticas foram delineadas
experimentalmente para manter discrepâncias de exposição luminosa entre rametes
parentais e filiais na situação de borda, muito embora, esta categoria tenha apresentado
uma variação ampla de regimes de radiação, dependendo, principalmente, da direção
cardeal do fragmento em relação ao centro da moita, espécie lenhosa do dossel sobre o
segmento e altura da moita (que influenciou o tempo de duração da sombra). A taxa de

29
crescimento populacional foi estimada pela média do número de novos rametes produzidos
por segmentos anualmente diminuídos do número de rametes que senesceram.
Os segmentos foram mensurados e acompanhados por dezoito meses, com
medições em intervalos de três meses. Foi estimada a proporção de rametes crescendo em
cada habitat, e dentro do fator micro-hábitats foram comparados: o comprimento do
estolão, o comprimento da maior folha, a área da roseta, o número de rosetas conectadas
por segmento, e o número de rosetas produzidas por número de rametes vivos no
segmento.
A proporção de segmentos clonais crescendo entre os três micro-hábitats na
população de N. cruenta de Jurubatiba foi obtida através de quatro transectos de 50 metros,
aonde foram registrados todos os segmentos clonais em 5 metros de distância para cada
lado. O estudo foi conduzido nos anos de 2006-2007.
Figura 2.1. Esquema de moitas de vegetação selecionadas para o acompanhamento do
crescimento e dinâmica populacional das rosetas da bromélia clonal Neoregelia cruenta no
Parque Nacional da Restinga de Jurubatiba. Micro-hábitats homogêneos: (1) Interior e (3)
Exposto; micro-habitat heterogêneo: (2) Borda – ramete parental sombreado e ramete filial
exposto.
(1) (2) (3)
Cobertura da moita
(1) (2) (3)
Cobertura da moita

30
2.2.3. Delineamento experimental e análise estatística
A amostragem de transectos seguiu a orientação paralela à linha da costa no
segundo cordão arenoso, para se evitar a influência mais significativa da salsugem e as
variações do perfil topográfico marcada pelas regressões e transgressões marinhas na
formação das planícies de restinga. Para compreender a influência destes fatores na
ecofisiologia de N. cruenta consulte Reinert (1995). Agregados de vegetação (moitas) que
tiveram simultaneamente segmentos clonais crescendo nas três condições de micro-habitat
foram selecionados como a unidade amostral, desta forma é possível incluir na variação
dos dados proporções eqüitativas e por tanto comparáveis, de diferenças na história de
formação das moitas (Zaluar, 2002; Andrade 1992) e diferenças ambientais entre
localidades (Pimentel, 2002).
A normalidade dos dados foi testada usando Kolmogorov-Smirnov (adaptado de
Zar, 1996). O comprimento do estolão entre os micro-hábitats foi comparado usando
curvas de freqüência. As comparações entre os parâmetros de tamanhos de rametes,
número de rametes por segmento, e a proporção do número de rametes produzidos por
rametes vivos foram realizadas utilizando ANOVA “one-way”, seguido pelo teste de
Tukey para evidenciar as diferenças de médias, quando os dados foram paramétricos. O
teste de Kruskal-Wallis, seguido pelo teste de Dunn, foi usado quando os dados foram não-
paramétricos, em especial para os dados de incremento nos parâmetros de tamanho
(comprimento da maior folha e área da roseta). Não foram utilizadas transformações para a
normalização dos dados, porque os dados de crescimento apresentaram comportamento
tipicamente não-paramétrico. As análises foram feitas com o auxílio do software Statistica
6.0, e os gráficos com o software Origin 4.1.

31
2.3. Resultados
Espécies vegetais com reprodução clonal muitas vezes apresentam forrageamento
horizontal do espaço, subsidiado pela produção de novos meristemas. Desta forma, o
aumento do crescimento vegetativo em manchas favoráveis de recurso e, inversamente o
decréscimo em manchas desfavoráveis, pode estar representado na distribuição
populacional de clones no espaço. A população de N. cruenta apresentou o maior número
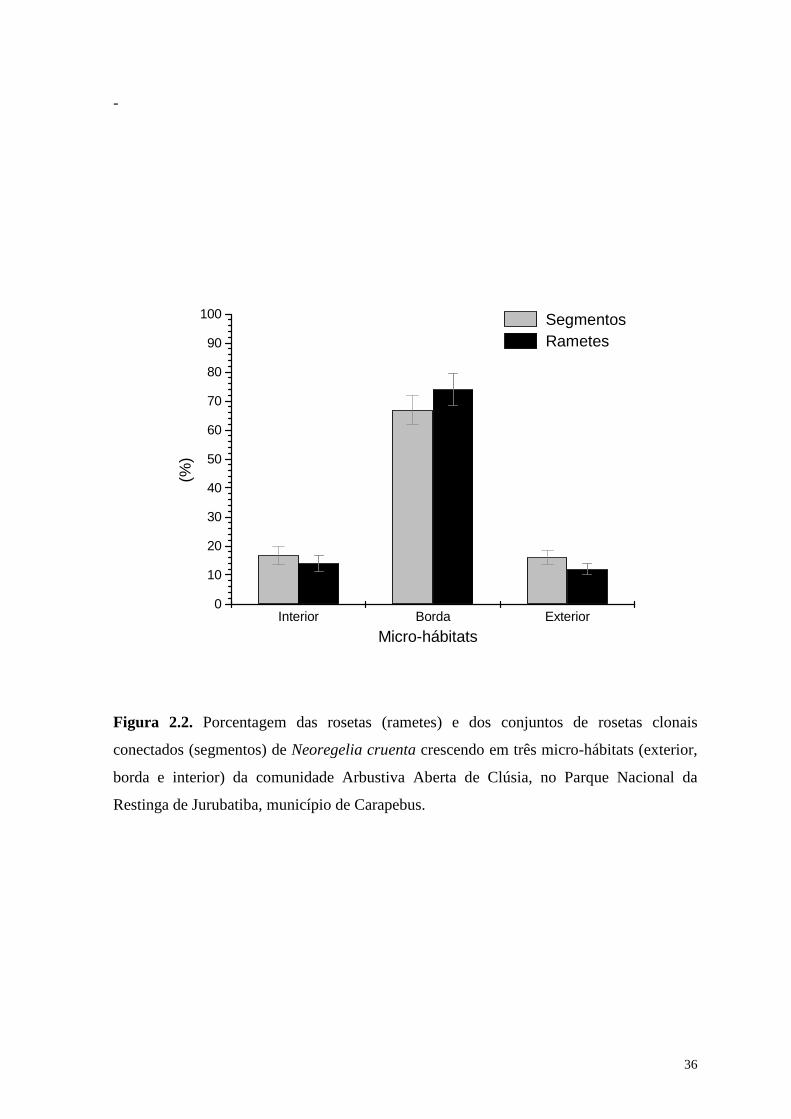
de segmentos clonais (67,7%) no ambiente heterogêneo da borda das moitas de vegetação.
O mesmo padrão foi encontrado para o número de rametes (72,3%; Figura 2.2). Foram
amostrados 161 segmentos, que congregaram 412 rametes. Os segmentos e rametes que
crescem nas condições de interior de moita de vegetação e na areia nua (exterior) tiveram
contribuições entre 15 e 18% do total de amostras.
O recrutamento de novos indivíduos em populações de bromélias crescendo nos
ambientes marginais da Mata Atlântica é, principalmente, oriundo do crescimento clonal
(Scarano, 2000). Esta propriedade populacional, nas espécies de reprodução vegetativa,
não se distingue da taxa de crescimento do indivíduo, embora o direcionamento do
investimento nas diferentes partes da arquitetura da planta possa variar. O tamanho das
rosetas de N. cruenta na localidade de estudo é afetado pelos micro-hábitats, atingindo as
maiores classes, tanto no comprimento da maior folha quanto na área de roseta, no interior
da moita (Figura 2.3A e 2.3B).
No crescimento clonal, o subsídio dado pelo indivíduo parental durante a fase de
estabelecimento diminui significativamente a mortalidade nas fases juvenis. Entretanto,
limita a ocorrência deste ao raio do comprimento do estolão. Esta característica adquire,
portanto, relevância na dinâmica da população, tendo sido inclusive identificados dois
extremos em estratégias de crescimento: falange e guerrilha, categorizados de acordo com
a distância do conector (Lovett Doust, 1981; Lovett Doust & Lovett Doust, 1982; Sampaio

32
et al., 2002). A distância e direcionamento dos novos estolões (ou conectores) influenciam
o comportamento de forrageamento do ambiente pela espécie clonal e podem ser uma
característica plástica, ou sem variação, quando determinada geneticamente ou canalizada
(de Kroons & Hutchings, 1995). O estolão de N. cruenta não variou com o ambiente. Duas
classes modais puderam ser igualmente identificadas na distribuição de freqüência nos três
micro-hábitats (8 e 14cm). O somatório da ocorrência das duas modas foi sempre maior
que 80% do total da distribuição.
A heterogeneidade ambiental pode favorecer o desempenho de segmentos clonais,
resultando em uma maior taxa de incorporação de biomassa, uma vez que os indivíduos
podem translocar fotossintatos e água e nutrientes. O número de rosetas que permanecem
conectadas nos segmentos de N. cruenta nas bordas das moitas de vegetação, onde os
indivíduos parentais e jovens estão submetidos à microclimas distintos, foi
significativamente maior do que no interior e exterior (Figura 2.3C). Apesar deste
resultado, o número de brotos produzidos na borda foi menor, sugerindo que os indivíduos
permanecem por mais tempo conectados (Figura 2.3D).
Os segmentos do exterior apresentaram maior média de produção de novos
indivíduos, o que em face ao menor número de rosetas por segmento, indica uma baixa
taxa de sobrevivência dos jovens ou o encurtamento do ciclo de vida das rosetas nesta
situação. As taxas de crescimento populacional foram negativas no exterior, enquanto
apresentaram valores positivos nas situações borda e interior (Figura 2.4).
Os resultados mostram que o microclima afeta a estrutura e a dinâmica das
subpopulações de N. cruenta e, além disso, que a marcante sazonalidade na pluviosidade
do clima da região pode ser significativa para a taxa de crescimento desta espécie. Os
menores valores de incremento no tamanho da roseta foram observados na estação seca (2o
trimestre; Tabela 2.1), independente do micro-habitat.

33
2.4. Discussão
Plantas que colonizam ambos ambientes sombreados, e expostos à forte radiação,
apresentam um conjunto de aclimatações, tradicionalmente relatadas como morfótipos
heliófitos e esciófitos (“sun” e “shade plants’, Bradshaw 1965, Boardman, 1977). Os
valores de comprimento foliar e área da roseta encontrados para os clones de N. cruenta
corroboram a capacidade de responder plasticamente na morfologia em relação à
incidência de radiação luminosa. De fato, este é um fator ambiental freqüentemente
relatado como estressante no ambiente de restinga, especialmente quando associado à
baixa retenção hídrica do solo (Reinert 1995; Mattos 1998). A necessidade de aclimatação
de N. cruenta ao ambiente luminoso, junto com as baixas taxas de crescimento,
encontrados neste estudo para o micro-habitat exposto, sugere que este fator seja
preponderante na freqüência de ocorrência dos indivíduos. Sampaio, Picó & Scarano
(2005), estudando Aechmea nudicaulis encontraram influência de variações nas
características do ambiente sobre a taxa de crescimento da população. Quando comparadas
as espécies, nos micro-habitats exposto e interior, N. cruenta apresenta taxa de crescimento
positiva no interior da moita e negativa na areia desnuda (Figura 2.4). Já A. nudicaulis teve
tendência à taxas de crescimento negativo em ambas as situações, embora com grande
variação (Sampaio 2004).
O comprimento do estolão, ao contrário das características da parte aérea das
rosetas, não sofreu alterações entre os microambientes estudados. Duas possíveis causas
podem ser responsáveis por este resultado: que esta seja uma característica espécie
específica, ou que haja uma canalização dos dados. O surgimento de novos rametes por
crescimento vegetativo, da mesma forma que o recrutamento sexuado, pode depender de
anos favoráveis. No recrutamento via sementes estes anos influenciam fortemente os
períodos de germinação e estabelecimento. Na reprodução vegetativa o vigor dos

34
indivíduos parentais determina se haverá o investimento na produção de novos brotos e em
que quantidade. Por fim, em N. cruenta o comprimento do estolão reflete a idade da planta,
uma vez que é formado conforme o crescimento do vegetal, tendo inclusive as marcas das
inserções das folhas que foram emitidas. As duas classes de comprimento de estolão
encontradas podem refletir anos favoráveis, aonde o excedente de carbono foi investido em
novos indivíduos produzidos por brotamento.
O tempo de vida da roseta e o número de brotos formados anualmente mantém os
segmentos clonais de N. cruenta em agregados de, em média, quatro indivíduos (mín.1 e
máx.9). O maior número de rosetas por segmento encontrado na situação de borda sugere
que as rosetas estejam integradas fisiológicamente melhorando o desempenho do segmento
como um todo e aumentando o número de clones que podem ser sustentandos, isto é, que
os rametes sejam capazes de se especializar no microclima local e translocar os recursos,
gerando uma melhoria no balanço fisiológico. Curiosamente, a maior taxa de crescimento
ocorreu no interior, mas a necessidade de fixação de mais carbono para atingir a classe de
tamanho adulta, mostra que apesar de mésico, para o crescimento da população de uma
espécie clonal, o ambiente pode não ser o mais favorável. As maiores taxas de incremento
na localidade de menor luminosidade sugerem que esta espécie pode passar por períodos
de fotoinibição, corroborados pela clorose comumente encontrada nas folhas (mensurações
do teor de clorofila são apresentados no tópico 4.3.2 e 4.4.2) e pelos resultados encontrados
por Reinert (1995).
Apesar de outros estudos com bromeliáceas relatarem que nesta família é comum a
reprodução vegetativa ser estimulada após o florescimento da roseta (Negrelle & Muraro,
2006; Sampaio et al., 2002; Sampaio, Picó & Scarano, 2005; Siqueira Filho & Machado,
2001), e mesmo que a reprodução assexuada seja estimulada em plantas clonais justamente
em anos desfavoráveis como estratégia à manutenção do recrutamento; a reprodução

35
assexuada em N. cruenta faz parte do crescimento vegetativo do indivíduo e, portanto, está
ligado ao balanço de carbono do ramete parental, pelo menos até a formação de raízes e a
abertura do copo. Os resultados deste estudo sugerem a possibilidade de um trade-off entre
o crescimento vegetativo da roseta e a reprodução clonal, isto é, localidades mais
estresssantes (exterior) estimularam o investimento em crescimento clonal, enquanto em
localidade mais protegida e favorável (interior) o crescimento e manutenção da roseta foi
maximizado. Isto é, a reprodução clonal em N. cruenta não parece ser uma simples
extensão do crescimento vegetativo, mas sim uma das alterações na arquitetura da roseta,
juntamente com outras respostas de aclimatação (abertura do copo e comprimento das
folhas) à variação ambiental.
A sub-população de N. cruenta, uma espécie-focal da vegetação, no micro-habitat
exposto, não apresentou crescimento populacional positivo (Figura 2.4). Uma vez que a
germinação e estabelecimento de plântulas desta espécie é improvável na areia nua
(Mantovani, 2002), os segmentos clonais que são hoje encontradas nesta situação se
originaram de indivíduos recrutados no interior das moitas de vegetação, ou são
remanescentes de moitas de vegetação dominadas por Clúsia que senesceram e regrediram
(Mantuano, 2003; Zaluar, 2002).
36
-
Figura 2.2. Porcentagem das rosetas (rametes) e dos conjuntos de rosetas clonais
conectados (segmentos) de Neoregelia cruenta crescendo em três micro-hábitats (exterior,
borda e interior) da comunidade Arbustiva Aberta de Clúsia, no Parque Nacional da
Restinga de Jurubatiba, município de Carapebus.
Interior Borda Exterior0
10
20
30
40
50
60
70
80
90
100
Micro-hábitats
(%)
Segmentos Rametes

37
Figura 2.3. Média (± DP) do (A) comprimento da maior folha, da (B) área de abertura da
roseta, do (C) número de rosetas vivas conectadas por segmento, e do (D) número brotos
produzidos por roseta adulta de Neoregelia cruenta, em três micro-hábitats (exterior, borda
e interior) da comunidade Arbustiva Aberta de Clúsia, no Parque Nacional da Restinga de
Jurubatiba, município de Carapebus. Letras mostram diferenças estatísticas (ANOVA one-
way; p<0,05). Dados coletados em Abril e Julho de 2006.

38
Figura 2.4. Média (± DP) da taxa de crescimento avaliada pelo número de novos rametes
decrescido do número de rametes que senesceram em segmentos clonais de Neoregelia
cruenta, em três micro-hábitats (exterior, borda e interior) da comunidade Arbustiva
Aberta de Clúsia, no Parque Nacional da Restinga de Jurubatiba, município de Carapebus.
Letras mostram diferenças estatísticas (Kruskal-wallis; p<0,05). Dados coletados em Abril
e Julho de 2006.
Exterior
Borda
Interior
-0,6 -0,4 -0,2 0,0 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8
Mic
ro-h
ábita
t
Taxa de crescimento (ramete/segmento.ano-1)
a
b
b

39
Tabela 2.1. Incremento em comprimento da maior folha e área de roseta nos primeiro e
segundo trimestres (intervalos de 89 e 86 dias, respectivamente). Os dados foram obtidos
no final da estação chuvosa (dez/2005-mar/2006) e início da estação seca (abr/2006 –
jul/2006), no Parque Nacional da Restinga de Jurubatiba, município de Carapebus.
Incremento em comprimento da roseta (cm.dia-1)
Incremento em área de roseta (cm2.dia-1)
Estação chuvosa Estação Seca Estação chuvosa Estação Seca
Exterior 0,27 (±0,08) 0,19 (±0,02) 3,24 (±0,23) 1,98 (±0,09)
Borda 0,40 (±0,09) 0,13 (±0,04) 2,26 (±0,18) 0,86 (±0,12)
Interior 0,27 (±0,04) 0,08 (±0,02) 2,44 (±0,17) 1,83 (±0,09)

40
Capítulo 3
Plasticidade intraclonal e variação interclonal
na anatomia foliar de Neoregelia cruenta

41
3. Plasticidade intraclonal e variação interclonal na anatomia foliar de Neoregelia cruenta (R. Graham) L.B. Smith (Bromeliaceae) em três micro-hábitats da Restinga Aberta de Clúsia
Mantuano, D.G.; Martinelli, G. & Costa, C.G.
Abstract
Neoregelia cruenta (R. Graham) L.B. Smith (Bromeliaceae) is a CAM tank bromeliad, endemic of southeast “restinga”, i.e, the open scrub vegetation of Brazilian coastal plains. Clonal reproduction is a common feature in herbaceous plants and has been shown to be important for recruitment and population maintenance in this species. N. cruenta population is subject to different levels of light exposure depending on the indivual position related to vegetation clumps. Conected ramets are subjected to three microhabitats: (i) exposed to full irradiance, outside vegetation islands; (ii) partially exposed to full irradiance, at the border of vegetation islands; (iii) shaded, inside vegetation islands. We examined the enviromental effect on leaf anatomy of N. cruenta variation both within intraclonal and interclonal. Leaves of two conected ramets of five replicas in each microhabitat were sampled. Anatomical parameters were thickness of leaf blade, thickness of chlorophyllian parenchyma, thickness and number of layer of water-storage parenchyma, thickness of adaxial and abaxial epidermis, density and position of trichomes, and stomatal density. All leaves of N. cruenta shared same anatomical features irrespective of microhabitat. Thicker water-storage parenchyma and leaf blade were found on both conected ramets in exposed microhabitat and the exposed ramet of partially exposed microhabitat, related to the shorter time of tank water storage. Intraclonal variation was statistically significative only at partially exposed microhabitat. Both phenotypic plasticity within conected ramets and genetic variation among conected ramets migh help to explain the high abundance of N. cruenta along heterogenous environment of “restinga” vegetation.
Resumo
Neoregelia cruenta (R. Graham) L.B. Smith (Bromeliaceae) é uma bromélia tanque de metabolismo CAM endêmica das restingas do sudeste, i.e, nas vegetações arbustivas abertas das planícies costeiras do Brasil. Reprodução clonal é uma característica comum em plantas herbáceas e tem sido relatada como importante para o recrutamento e manutenção populacional desta espécie. As populações de N. cruenta estão sujeitas a distintos níveis de exposição à luz, dependendo da posição do indivíduo em relação a moita de vegetação. Rametes conectados estão sujeitos a três micro-hábitats: (i) exposto a irradiância total, ocorrendo fora da ilha de vegetação; (ii) parcialmente exposto a irradiância total, ocorrendo na borda das ilhas de vegetação; (iii) sombreado, ocorrendo dentro das ilhas de vegetação. Este estudo examinou o efeito do ambiente sobre a variação intra e interclonal na anatomia foliar de N. cruenta. Folhas de dois rametes conectados em cinco réplicas em cada micro-hábitat foram amostradas. Os parâmetros anatômicos utilizados foram espessura da lâmina foliar, espessura do parênquima clorofiliano, espessura e número de camadas do parênquima aqüífero, espessuras das epidermes abaxial e adaxial, densidade e posição dos tricomas, densidade de estômatos. Todas as folhas de N. cruenta mostraram estruturas anatômicas similares, independentes do micro-hábitat. Parênquima aqüífero e lâmina foliar mais espessos foram encontrados em ambos rametes do micro-hábitat exposto e no ramete exposto do micro-hábitat parcialmente exposto, relacionando-se como o encurtamento do tempo de estoque da água no tanque. A variação intraclonal foi estatisticamente significativa apenas para o micro-hábitat parcialmente exposto. Ambos plasticidade fenotípica dentro dos rametes conectados e variação genética entre os clones podem explicar a alta abundância de N. cruenta através do ambiente heterogêneo da vegetação de restinga.
Palavras-chaves: Neoregelia cruenta, plasticidade, reprodução clonal, anatomia foliar, Restinga. Key words: Neoregelia cruenta, plasticity, clonal reproduction, foliar anatomy, Restinga.

42
3.1. Introdução
A diversidade dos ambientes que cercam o complexo Mata Atlântica, que é seguido
de grande diversidade nas comunidades vegetais, estimulou vários estudos acerca da
variação morfofuncional de espécies que são distribuídas amplamente entre os vários
hábitats (Scarano 2002). Estes estudos mostram que as variações em características
estruturais e funcionais entre populações podem subsidiar a ocorrência e a abundância
destas espécies (Rôças 1997, 2001, Scarano et al 2001, 2002, 2005a, 2005b, Vieira 1995).
O ambiente de restinga ocupa as planícies arenosas costeiras contíguas à costa
atlântica e apresentam uma forte heterogeneidade ambiental. Alagamento, salinidade
regime de ventos e topografia são alguns dos fatores abióticos que imprimem variação
espacial a esse ambiente. As comunidades arbustivas abertas recobrem grande parte deste
ambiente (Araujo & Henriques 1984, Henriques Araujo & Hay 1986) e, por apresentarem
áreas cobertas por vegetação e áreas desnudas, apresentam micro-hábitats distintos quanto
à incidência de radiação luminosa. Mantuano et al. (2006) acreditam que espécies de
ampla distribuição e abundantes deverão ser capazes de lidar com os micro-hábitats dentro
das populações em comunidades abertas de restinga.
Neoregelia cruenta é uma espécie-focal na vegetação Arbustiva Aberta de
Clusia, na qual aumenta o teor de nutrientes, a capacidade de troca catiônica e o conteúdo
de matéria orgânica do solo (Lacerda & Hay 1977, 1978, Hay & Lacerda 1980, Mantovani
2002) e, é sítio de germinação de outras espécies (Fialho & Furtado 1993, Zaluar com.
pess.). Esta bromélia foi relatada como uma possível iniciadora de moitas de vegetação
(Zaluar & Scarano 2000) e age, possivelmente, expandindo a área de ocupação das moitas
(Mantovani 2002).
Assim como outras espécies do gênero, N. cruenta é capaz de se reproduzir
assexuadamente através do crescimento vegetativo do estolão (Benzing 2000). Esta

43
característica permite recrutar novos indivíduos sobre a areia nua. Local em que a
deficiência hídrica severa e altas temperaturas são limitantes a germinação de sementes.
Embora, N. cruenta seja freqüente em áreas expostas é também encontrada em abundância
sombreada por moitas de vegetação.
Plasticidade fenotípica define a capacidade de expressão de fenótipos distintos
induzidos pelo ambiente a partir de indivíduos geneticamente idênticos (Bradshaw 1965,
Schlichting 1986). O crescimento clonal de N. cruenta na comunidade aberta das restingas
sujeita conjuntos de rosetas clonais à ambientes diferenciados em incidência de radiação
luminosa e disponibilidade hídrica. O objetivo deste estudo foi entender se a alta
freqüência de ocorrência de N. cruenta nos micro-hábitats da Comunidade Arbustiva
Aberta de Clúsia é subsidiada por variação na anatomia foliar entre indivíduos crescendo
expostos ao sol pleno e protegidos sobre o dossel das moitas de vegetação e/ou pela a
plasticidade fenotípica intraclonal.
3.2. Material e métodos
A variação anatômica nas folhas de Neoregelia cruenta foi estimada com amostras
de cinco segmentos clonais coletados em cada um dos três micro-hábitats (descritos na
seção 1.5). Nestes segmentos foram amostrados dois indivíduos adultos com conexão
aparente e funcional categorizados como ramete parental e ramete filial. A escolha dos
rametes foi direcionada para evidenciar a discrepância nas condições de luminosidade no
micro-hábitat borda, e para a similaridade entre as rosetas crescendo nos micro-hábitats
exposto e interior, respectivamente, alta e baixa irradiância. Discos foliares com 1cm de
diâmetro foram retirados da porção central do limbo e imediatamente fixados em solução
de Glutaraldeído 2,5% em tampão fosfato. No laboratório as amostras foram lavadas em
água destilada abundantemente e em seguida emblocados em Agarose 6% (Ruzin, 1999).
Neste procedimento não houve desidratação do material, e em consonância durante o

44
emblocamento não houve a necessidade de infiltrar o material. A Agarose não teve a
função de homogeneização da densidade como nos blocos feitos a partir de parafina ou
historesina. Neste procedimento o emblocamento tem apenas a função de sustentar o
material para o corte. Secções de 5-7µm do limbo foram obtidas com Vibrátomo de
deslize. Neste procedimento as secções são obtidas com a utilização de lâminas de barbear
que deslizam sobre o material com o auxílio de micro vibração criada pelo aparelho. Os
cortes se separam do invólucro de sustentação ao serem cortados e imergirem numa cuba
com água destilada. Em seguida as secções foram lavadas em Hipoclorito de Sódio 5%, e
em água Acética, e por fim coradas com Safrablau. Lâminas semipermanentes foram
montadas com gelatina glicerinada e Fenol (Sass, 1940).
O material coletado para a descrição do estolão de conexão entre as rosetas de N.
cruenta foi coletado em fixador Glutaraldeído 2,5% em tampão fosfato. No laboratório o
material foi desidratado em série alcoólica, com intervalos de 3 horas entre cada solução, e
ao final imerso em álcool 95% por 24 horas. A infiltração em historesina (Leica) foi feita
com solução de infiltração e álcool 95% em proporção de 1:1 por 24 horas resfriado em
aproximadamente 15ºC. O material foi então lavado em historesina pura por duas vezes,
em 48 horas, sempre resfriado. Em seguida foi emblocado com endurecedor e
polimerizado em dessecador com sílica gel (o procedimento foi adaptado para materiais
volumosos conforme manual do fabricante Leica Historesin Embedding Kit 7022). O bloco
foi seccionado em micrótomo de deslize, com espessuras entre 8-10µm. As lâminas foram
coradas com azul de Toluidina O 0.1% em placa de aquecimento e em seguidas lavadas em
abundância com água destilada para retirar o excesso de corante (Ruzin, 1999). As lâminas
foram secas e montadas em Permount.
A descrição e mensuração dos mesofilos foi feita utilizando microscópio ótico
(Olympus) acoplado a câmera digitalizadora e software de processamento de imagens

45
Image Pro-plus.
Os parâmetros selecionados foram: espessura foliar total, espessura do parênquima
clorofiliano, espessura do parênquima aqüífero, número de camadas do parênquima
aqüífero e comprimento da primeira camada de células do aqüífero adjacente ao
parênquima clorofiliano.
A observação da epiderme foi feita em dissociações com a solução de Jefrey (1:1
Ácido Crômico 10% e Ácido Nítrico 10%) por 4-6 horas em estufa a 55-60ºC. As
epidermes foram coradas com safranina e montadas em água glicerinada 50%. As
lamínulas foram lutadas com esmalte incolor. Neste procedimento os tricomas peltados
característicos na subfamília não puderam ser observados. Outras técnicas de dissociação
também se mostraram ineficientes na preservação dos tricomas. Apesar disso, a
mensuração do número de tricomas por área de epiderme, bem como a relação do número
de tricomas por estômato, puderam ser feitas com a contagem das cicatrizes das inserções
da base do tricoma no tecido.
A disposição dos tricomas peltados na folha e o seu posicionamento em relação aos
estômatos foram observados através de observações em Microscópio Eletrônico de
Varredura. Para isso, o material coletado com fixador glutaraldeído em tampão fosfato. No
laboratório as amostras foram lavadas em água destilada para a retirada do fixador, em
seguida desidratadas em séria acetólica e, por fim infiltradas com infiltrados em Tetróxido
de Ósmio. O material foi seco com Ponto Crítico de dióxido de carbono, montado em
suportes (Stubs) com fita de carbono e cola de prata. Ao final o material foi recoberto com
fina camada de ouro com auxílio de metalizador, e observado num microscópio eletrônico
de varredura (Evo40, JEOL). O material processado neste caso foi apenas de amostras da
situação borda, aonde a folha do ramete parental foi categorizado como amostra de sombra
baixa irradiância, aprox. 200µmol PPFD) e a folha do ramete filial foi categorizada como

46
amostra de sol (alta irradiância, aprox. 1700 µmol PPFD).
A variação anatômica intraclonal foi estimada pelas diferenças numéricas nos pares
de rametes amostrados entre os três micro-hábitats. Detalhadamente, uma amostra
composta de cinco medições foi feita para cada folha amostrada de um par de rametes
conectados num segmento clonal, isto é, foram feitas uma amostra da folha do ramete
parental e uma do filial. A diferença entre a média calculada destas medidas representou a
variação intraclonal.
A comparação estatística entre os micro-hábitats foi feita utilizando ANOVA “one-
way”, seguida do Teste de Tukey para evidenciar a diferenças entre as médias quando elas
existiram (Zar, 1996). As diferenças nas contagens dos estômatos e tricomas entre a
situação exposto e sombreado na situação Borda, assim como a relação entre o número de
tricomas por estômato, foi testada usando o Teste “T” Student (Zar, 1996).
3.3. Resultados
A anatomia foliar de Neoregelia cruenta apresentou estrutura comum a todas as
amostras estudadas: folhas hipostomáticas, epiderme uniestratificada com tricomas
peltados, presença de cristais estegmatas na cutícula da parede periclinal externa da
epiderme adaxial, presença de parênquima aqüífero com células em paliçada apresentando
espessamento helicoidal nas paredes, parênquima clorofiliano com células tendendo a
isodiamétricas e presença de diafragma com parênquima baciforme preenchendo canais
aeríferos formados pela junção das cavidades subestomáticas (Figura 3.1a-f, 3.3a-d).
Apesar de não terem sido encontradas diferenças qualitativas, foram observadas,
entretanto, alterações quantitativas. A espessura total do mesofilo e a espessura total do
parênquima aqüífero mostraram uma tendência ao espessamento acompanhando o aumento
do grau de exposição luminosa quando analisados entre micro-hábitat e entre rametes
(Figura 3.1a-f), embora não tenha sido encontrada diferença estatística significativa.

47
(Tabela 3.1). O espessamento total do mesofilo deve-se a contribuição do parênquima
aqüífero, uma vez que o parênquima clorofiliano não variou com o grau de exposição
luminosa (Tabela 3.1). O comprimento da primeira célula do parênquima apresentou
valores diferentes estatisticamente entre os rametes parentais e filiais nas situações borda e
exterior (Tabela 3.1). Assim, a variação individual, isto é, entre os conjuntos de rametes
conectados de N. cruenta, foi maior que a variação entre os micro-hábitats, não podendo
prever um ajuste anatômico que subsidie a ocorrência nos três micro-hábitats.
Entretanto, quando os rametes parentais e filiais foram analisados em pares,
Neoregelia cruenta apresentou plasticidade fenotípica intraclonal significativa no ambiente
heterogêneo (borda das moitas de vegetação), e valores de variação menores nos ambientes
homogêneos (interior e exterior de moitas de vegetação) (Tabela 3.2.). A espessura foliar
total e a espessura do parênquima aqüífero foram maiores no ramete filial (borda-exposto)
do que no ramete parental (borda-protegido). Já a altura do primeiro estrato do parênquima
aqüífero próxima ao clorofiliano teve variação menor que os outros dois micro-hábitats
(Tabela 3.2). O espessamento do parênquima aqüífero pode ser explicado pelo aumento do
número de estratos do tecido (Figura 3.2). Novamente o parênquima clorofiliano não
apresentou alterações entre rametes parentais e filiais nas três situações de exposição
luminosa.
Os estômatos e os tricomas ocorrem próximos na epiderme abaxial e ocorreram de
1-2 tricomas por estômatos (Figura 3.3b e Tabela 3.3), nos quais houve sobreposição
parcial dos escudos. Na epiderme adaxial os tricomas foram dispostos uniformemente e
recobriram área maior, com pouco ou nenhuma sobreposição dos escudos (Figura 3.3a). As
folhas de rosetas crescendo no interior apresentaram menor densidade de tricomas na face
adaxial que nas rosetas do exterior (Tabela 3.3).

48
Figura 3.1. Anatomia foliar de Neoregelia cruenta em três diferentes micro-hábitats. a.
Mesofilo do ramete parental crescendo sobre a areia nua; b. Mesofilo do ramete filial
crescendo sobre a areia nua; c. Mesofilo do ramete parental crescendo protegido na borda
da moita de vegetação; d. Mesofilo do ramete filial crescendo exposto na borda da moita
de vegetação; e. Mesofilo do ramete parental crescendo sombreado no interior da moita de
vegetação; f. Mesofilo do ramete filial crescendo sombreado no interior da moita de
vegetação. Os pares de ramete parental e filial amostrados possuem estolão funcional de
conexão. Barra = 100µm. Material coletado na Restinga Aberta de Clúsia do Parque
Nacional da Restinga de Jurubatiba, município de Carapebus.
c d
e f
a b

49
Tabela 3.1. Média da espessura foliar total, espessura do parênquima clorofiliano,
espessura do parênquima aqüífero, e comprimento da primeira célula do parênquima
aqüífero adjacente ao parênquima clorofiliano em Neoregelia cruenta em situações de
interior de moita de vegetação, borda e exterior na Restinga aberta de Clúsia no Parque
Nacional da Restinga de Jurubatiba, Carapebus/RJ. Valores medidos em µm. Letras
mostram diferenças estatísticas usando ANOVA “one way” seguida do Teste de Tukey.
Espessura foliar total
Espessura do Parênquima Clorofiliano
Espessura do Parênquima
Aqüífero
Altura do primeiro estrato do
Parênquima Aqüífero
Ramete parental 1541,71 670,24 621,88 218,27
Ramete filial 1436,75 649,02 575,93 218,62
Ramete parental 1309,07 598,89 536,20 164,44
Ramete filial 1503,50 620,26 695,32 252,33
Ramete parental 1292,24 672,94 403,09 183,82
Ramete filial 1467,33 692,87 500,75 248,79
Exterior
Borda
Interior

50
Tabela 3.2. Diferença entre as mensurações foliares realizadas em duas rosetas
conectadas em cada micro-habitat. Valores medidos em µm. Letras mostram diferenças
estatísticas usando ANOVA “one way” seguida do Teste de Tukey.
Espessura Foliar Total
Espessura do
Parênquima Clorofiliano
Espessura do
Parênquima Aqüífero
Altura primeiro estrato do
Parênquima Aqüífero
Exterior 118,2a 61,2a 150,0a 115,6a
Borda 234,7b 44,4a 183,0b 92,7b
Interior 176,5c 52,8a 166,5a 104,1a
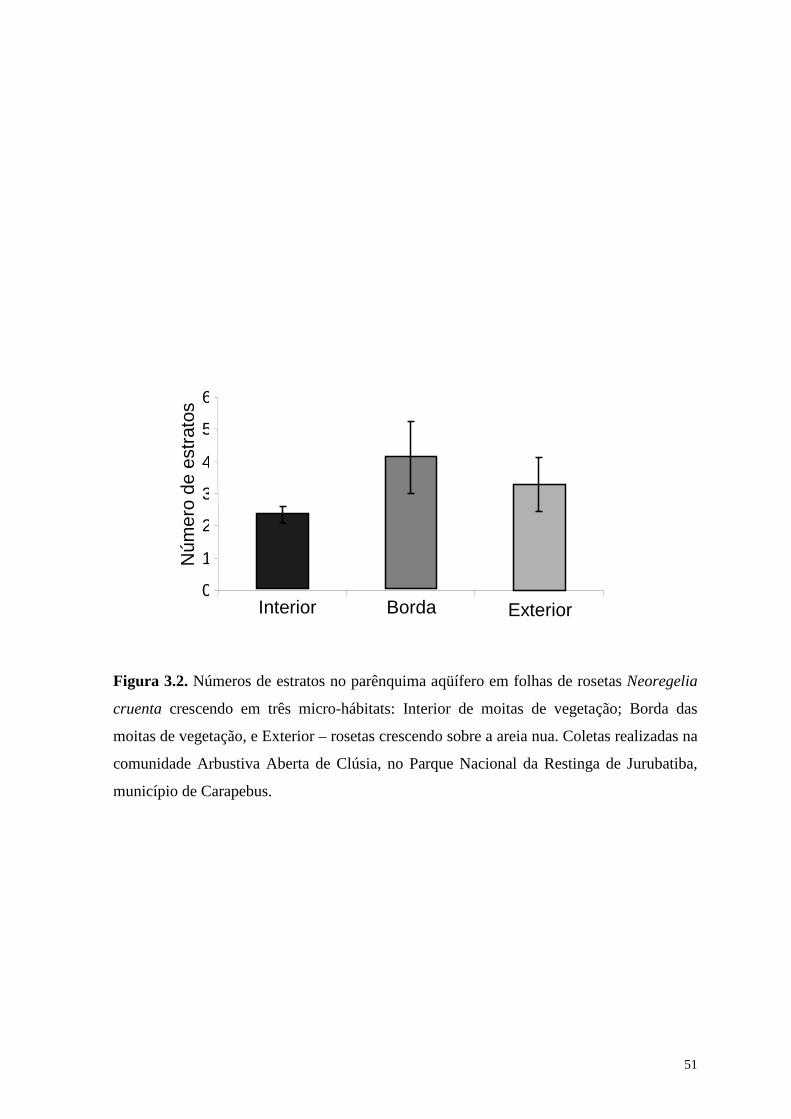
51
Figura 3.2. Números de estratos no parênquima aqüífero em folhas de rosetas Neoregelia
cruenta crescendo em três micro-hábitats: Interior de moitas de vegetação; Borda das
moitas de vegetação, e Exterior – rosetas crescendo sobre a areia nua. Coletas realizadas na
comunidade Arbustiva Aberta de Clúsia, no Parque Nacional da Restinga de Jurubatiba,
município de Carapebus.
Núm
ero
de e
stra
tos
0
1
2
3
4
5
6
Interior Borda Exterior
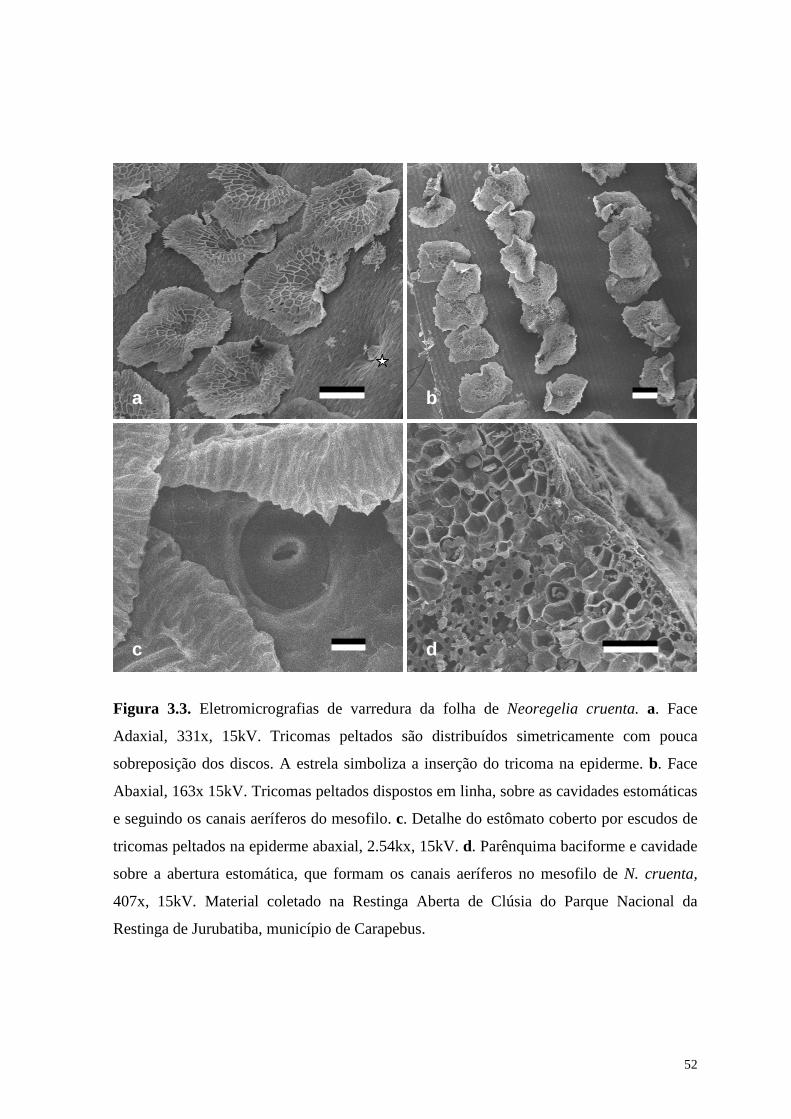
52
Figura 3.3. Eletromicrografias de varredura da folha de Neoregelia cruenta. a. Face
Adaxial, 331x, 15kV. Tricomas peltados são distribuídos simetricamente com pouca
sobreposição dos discos. A estrela simboliza a inserção do tricoma na epiderme. b. Face
Abaxial, 163x 15kV. Tricomas peltados dispostos em linha, sobre as cavidades estomáticas
e seguindo os canais aeríferos do mesofilo. c. Detalhe do estômato coberto por escudos de
tricomas peltados na epiderme abaxial, 2.54kx, 15kV. d. Parênquima baciforme e cavidade
sobre a abertura estomática, que formam os canais aeríferos no mesofilo de N. cruenta,
407x, 15kV. Material coletado na Restinga Aberta de Clúsia do Parque Nacional da
Restinga de Jurubatiba, município de Carapebus.
a b
c d

53
Tabela 3.3. Médias do número de tricomas, número de estômatos e da razão entre tricomas
e estômatos por mm2 de folha de Neoregelia cruenta em dois micro-hábitats: interior de
moitas de vegetação (baixa irradiância) e exposto crescendo sob areia nua (alta
irradiância). Os valores denotam diferença entre mensurações em duas rosetas conectadas
(ramete parental e ramete filial) em cada micro-habitat. Letras mostram diferenças
estatísticas usando Teste “t” Student (n=8). Material coletado na Restinga Aberta de Clúsia
no Parque Nacional da Restinga de Jurubatiba, município de Carapebus.
Densidade de tricomas
(#/mm2)
Densidade estomática
(#/mm2)
Razão estômatos/tricomas
Abaxial Adaxial Abaxial Abaxial
Interior (baixa irradiância)
26,38 ±(4,13)a 8,31 ±(7,74)a 30,44 ±(5,54)a 1,16 ±(0,18)a
Exposto (alta irradiância) 34,38 ±(4,26)a 25,56 ±(4,55)b 43,94±(8,59)a 1,28 ±(0,22)a

54
3.4. Discussão
Alterações na anatomia foliar ligadas à quantidade de energia luminosa recebida
são amplamente conhecidas na literatura como síndrome de folhas de sol e sombra (sun e
shade leaves Boardman 1977, Wylie 1951, Jackson 1967). Na situação sombreada os
tecidos fotossintéticos do mesofilo tendem à captar o máximo de energia luminosa e em
locais expostos ao sol pleno, tendem ao desenvolvimento de estratégias que minimizem a
perda d’água. As alterações encontradas na anatomia foliar de Neoregelia cruenta sugerem
que a disponibilidade hídrica, fator ligado à estratificação e alongamento celular do
parênquima aqüífero concomitantemente ao aumento do grau de exposição, é
preponderante, enquanto a forte amplitude de radiação luminosa não provocou alterações.
O ambiente luminoso nas restingas tende a um excesso de luz para fotossíntese
(Mattos et al. 2004). Valores perto de 2000µmol PPFD podem ser medidos em dias de
verão. O baixo teor de clorofila encontradas nas rosetas crescendo expostas (dados
apresentados no item quatro desta tese) e os valores de Fv/Fm medidos por Reinert (1995)
para N. cruenta em Maricá indicam que o excesso de luz pode provocar fotoinibição
crônica nesta espécie. Neste caso, a rosetas crescendo sombreadas no interior das moitas
apresentariam a assimilação fotossintética ideal, enquanto as rosetas crescendo na borda e
na situação exposta experimentam excesso de luz.
Contribuem para a não variação dos tecidos fotossintéticos do mesofilo, os ajustes
observados na abertura do copo. Havendo a modificação do ângulo de inserção das folhas
o total de energia que incide é menor (Nobel 1991), diminuindo a necessidade de ajustes
morfofuncionais.
Ajustes na estrutura da epiderme e do parênquima paliçádico podem alterar a
interação óptica da luz com a superfície da folha e as camadas adjacentes (Vogelmann
1993, Vogelmann & Martin 1993). A disposição do parênquima aqüífero na face adaxial

55
pode aumentar a difusão dos raios luminosos que chegam ao parênquima clorofiliano.
Assim, o espessamento do parênquima aqüífero pode contribuir para ambas as situações
ambientais: deficiência hídrica e excesso luminoso.
As diferenças na densidade e disposição dos tricomas peltados entre a epiderme
adaxial e abaxial sugerem que os tricomas desempenham funções distintas no balanço
hídrico (Benzing & Burt 1970, Benzing Seemann & Renfrow 1978, Pierce et al. 2001). A
proximidade com os estômatos na face abaxial e a sobreposição dos escudos aumenta a
resistência da camada limítrofe da superfície foliar. Por outro lado, a distribuição uniforme
na face adaxial aumenta a área de retenção de filme de água sobre a superfície foliar
auxiliando a sua absorção. De fato, folhas crescendo no interior apresentaram menos
tricomas na face adaxial do que as folhas de rosetas crescendo no exterior. O aumento da
temperatura no interior das rosetas expostas (dados apresentados na figura 1.4) certamente
aumenta a taxa de evaporação da água retida no tanque, em bromélias expostas ao sol
pleno. O maior número de tricomas na face foliar voltada para o copo sugere que haja
aceleração da taxa de absorção compensando o encurtamento do tempo de retenção de
água.
A população de Neoregelia cruenta estudada apresentou variação quantitativa em
alguns doa tecidos foliares (especialmente nos envolvidos no balanço hídrico) entre os
conjuntos de rosetas clonais nos distintos micro-hábitats. A amplitude de variação entre os
clones foi maior e abrangeu as alterações intraclonais provocadas por diferenças micro-
climáticas, quando rosetas parentais e filiais foram analisadas separadamente. A
plasticidade fenotípica intraclonal, medida pela diferença encontrada entre os pares de
rosetas com a mesma carga genética, foi significativa. Tanto a variação genética entre
clones, quanto a capacidade de expressão plástica no fenótipo podem estar subsidiando a
alta freqüência de ocorrência de N. cruenta, espécie-focal na vegetação.

56
Capítulo 4
Desacoplamento da condutância estomática
de rametes conectados em micro-hábitat heterogêneo

57
4. Variação intra e interclonal nos atributos morfológicos foliares, conteúdo de
pigmentos e condutância estomática de Neoregelia cruenta (R. Graham) L.B. Smith
(Bromeliaceae) crescendo em três micro-hábitats da Restinga Aberta de Clúsia.
Mantuano, D.G.; Martinelli, G. & Costa, C.G.
Abstract
Clonal plant species can be considered as populations of interconnected ramets which are basically identical in form and function, and potentially independent from each other. Experimental studies suggest that an intra-clonal specialization of ramets with different roles can increase the performance of clonal systems under hetrogeneous condition. This paper explores structural and functional variation of rosettes of Neoregelia cruenta (R. Graham) L.B. Smith (Bromeliaceae)¸ a CAM tank bromeliad, under three environmental conditions in a tropical vegetation. The main question asked in this study is if heterogenous habitats are able to induce diferent transpiration rates on connected ramets. We assumed it is a potential mechanism to induce intraclonal specialization. N. cruenta plays an important ecological role, increasing soil nutrient content and initiating vegetation islands, at sandy coastal plains in Southeast Brazil. In field conditions clonal clumps can be found shaded or fully exposed to sun light (homeogeneous microhabitats), and partially exposed to full sun light (heterogeneous microhabitat). We compared morphological and physiological parameters among intraclonal and interclonal rosettes in all microhabitats. Leaf area and lenght of leaf blade vary strongly, decreasing towards to light exposition. Leaf suculence and specific leaf weight were similar between connected ramets but lower on exposed microhabitat. Chlorophyll and carotenoids contents decrease as light gradient enhance. This results suggest that, although acclimation features could be observed, exposed ramets (entire clonal clump of exposed microhabitat and half clump of partially exposed microhabitat) are under light stress conditions. Stomatal condutance were different betweem rosettes of heterogeneous habitat, contributing to confirm that directional xylem flow between ramets is likely to occurs. Resumo
Espécies clonais podem ser consideradas como populações de rametes interconectados que são essencialmente idênticos em forma e função, e potencialmente independentes um do outro. Estudos experimentais sugerem que uma especialização intraclonal com distintas funções pode aumentar a performance do sistema clonal sobre condições heterogêneas. Este estudo explora a variação estrutural e funcional de rosetas de Neoregelia cruenta (R. Graham) L.B. Smith (Bromeliaceae)¸ uma bromélia-tanque CAM, sobre três condições ambientais em uma vegetação tropical. A questão central deste estudo foi se hábitats heterogêneos são capazes de induzir diferentes taxas de transpiração em rametes conectados. Assumiu-se que este é um mecanismo em potencial para a indução da especialização intraclonal. N. cruenta tem um importante papel ecológico, aumentando o conteúdo nutricional do solo e iniciando moitas de vegetação, nas restingas do sudeste do Brasil. Em condições de campo agregados clonais podem ser encontrados sombreados ou completamente expostos ao sol (micro-hábitats homogêneos), e parcialmente expostos ao sol pleno (micro-hábitat heterogêneo). Comparou-se parâmetros morfológicos e fisiológicos entre rosetas intraclonais e interclonais em todos os micro-hábitats. Área foliar e comprimento do limbo foliar variaram fortemente, decrescendo de acordo com a exposição à luz. Suculência foliar e peso específico foliar foram similares entre rametes conectados, mas menores no micro-hábitat exposto. Conteúdo de clorofila e carotenóides diminuiu enquanto o gradiente luminoso aumentou. Este resultado sugere que, embora caracteres de aclimatação tenham sido observados, rametes expostos (o agregado clonal inteiro no micro-hábitat exposto e a metade do agregado no micro-hábitat parcialmente exposto) estão sobre estresse luminoso. A condutância estomática foi distinta entre rosetas do hábitat heterogêneo, corroborando a idéia de um fluxo direcional do xilema entre rametes.
Palavras-chaves: Neoregelia cruenta, especialização intraclonal, morfologia foliar, condutância estomática, reprodução clonal. Key words: Neoregelia cruenta, intraclonal specialization, leaf morphology, stomatal condutance, clonal reproduction.

58
4.1. Introdução
As restingas são ambientes costeiros onde espécies lenhosas de porte
arbustivo/arbóreo crescem sobre planícies arenosas oriundas do depósito marinho (Araújo
& Henriques 1984). As plantas que ocorrem neste ecossistema estão submetidas a diversos
fatores de estresse. O substrato tipicamente arenoso possui baixa retenção hídrica e pode
conferir pouca disponibilidade de água às plantas. Além disto, as espécies vegetais da
restinga sofrem influência do estresse salino devido a sua proximidade com o mar. A
incidência de salinidade pode ainda ter como efeito indireto o agravamento da baixa
captação de água pelas raízes. Altas temperaturas na superfície do solo desnudo, grandes
amplitudes térmicas diárias e alta incidência de radiação luminosa são fatores estressantes
relatados para ambientes costeiros (Crawford 1989) e também são observados na restinga
(Franco et al 1984, Lacerda et al. 1993).
Os fatores de estresse podem limitar a ocorrência das espécies, principalmente em
ambientes adversos. A associação com espécies focais é um fenômeno que permite às
espécies ocorrerem em locais onde os fatores ambientais, de outra forma, as restringiriam
(Bertness & Callaway 1994, Callaway 1992, 1995) Neste tipo de interação, a ocorrência da
espécie-focal altera as condições ambientais restritivas, subsidiando a germinação e o
estabelecimento das espécies beneficiárias. Este mecanismo é particularmente comum em
ambientes áridos, locais aonde a embebição das sementes é fortemente limitada
(Munzbergova & Ward, 2002; Jordan & Nobel, 1979; Franco & Nobel, 1988, 1989; Gass
& Barnes, 1998; Yeaton, 1978; Valient-Banuet et al.,1991; Valiente-Banuet, Vite &
Zavala-Hurtado, 1991; Valiente-Banuet & Ezcurra, 1991; Fulbright, Kuti & Tipton,1995;
Barnes & Archer, 1996).
A restinga é um ecossistema que apresenta intensa adversidade ambiental, contudo
esta combinação de fatores limitantes às plantas não impediu a fixação de uma grande

59
riqueza de espécies (Araujo & Henriques 1984, Pereira et al. 2001). Zaluar & Scarano
(2000) propõe que a germinação e o estabelecimento de diversas espécies lenhosas nas
restingas são facilitados pela ocorrência de espécies-focais na vegetação (com destaque
para C. hilariana) que aumentam a disponibilidade de umidade e nutrientes, e diminui a
temperatura superficial do solo. Desta forma, a estruturação da comunidade depende de
espécies-chaves (Scarano, 2002). Contudo, além do seu papel ecológico, é preciso entender
a biologia das espécies focais porque, em última análise, serão os fatores que influenciam a
estrutura populacional destas espécies que influenciará a dinâmica da comunidade.
Habitualmente, as relações ecológicas das plantas focais, e como estas interações
são reguladas por alterações microclimáticas, são o foco de estudos na área. (Franco &
Nobel, 1989, 1990; Franco-Pizaña et al., 1996; Fulbright, Kuti & Tipton, 1995; Valiente-
Banuet & Ezcurra, 1991; Vetaas, 1992; Forseth, Wait & Casper, 2001; Flores-Martinez,
Ezcurra & Sánchez-Colón, 1998). Apesar das espécies-focais terem sido documentadas
como de ampla capacidade de aclimatação, tolerantes ao estresse e com estruturas
xeromórficas, os processos fisiológicos que subsidiam sua colonização nestes ambientes
adversos têm sido pouco estudados (veja Berg et al., 2004). Esta tese propõe que o
crescimento vegetativo, através da reprodução clonal, pode ser importante para o
estabelecimento de novos indivíduos de Neoregelia cruenta; e admite-se que esta tem
papel facilitador na comunidade, subsidiando a germinação (Fialho & Furtado, 1993) e
melhorando as condições edáficas (Hay & Lacerda, 1980; Hay, Lacerda & Tan, 1981,
Lacerda & Hay, 1982). Contudo, apesar de o recrutamento em espécies focais ser
importante, o estabelecimento estará condicionado à capacidade de tolerância ao estresse e
aclimatação.
O estresse é responsável por causar e modelar respostas morfológicas, anatômicas e
fisiológicas nas plantas, que visam diminuir o efeito da adversidade sobre o seu

60
desempenho fisiológico e reprodutivo (Larcher 2000). A alteração nos atributos foliares e
fisiologia das plantas que alguns fatores ambientais estressantes geram e os mecanismos
pelos quais atuam já são conhecidos e podem subsidiar as premissas deste estudo.
As plantas podem adotar várias características para lidar com o déficit hídrico, tais
como: modificações na estrutura anatômica da folha, regulação estomática, investimento
no sistema radicular, área foliar reduzida, aumento da suculência foliar, redução da massa
específica foliar, metabolismo CAM, entre outros (Larcher 2000). O estresse hídrico, em
geral, pode ser menos prejudicial aos indivíduos com área foliar reduzida ou induzir a
produção de novas folhas com área menor. Isto se deve, além da diminuição da superfície
de evapotranspiração, à menor diferença de umidade e temperatura entre a folha e o
ambiente. Folhas maiores podem apresentar a camada envolvente de ar mais espessa e que
se renova mais lentamente, assim à transferência de calor da folha para o ar fica
dificultada. Como resultado, as folhas concentram mais calor apresentando temperaturas
superiores às do ar e incrementando as taxas transpiracionais (Nobel 1991). Entretanto,
esta relação entre o estresse hídrico e a área foliar não é obrigatória, sendo fortemente
influenciada pela abertura estomática (Valladares e Pearcy 1997) e por características
morfológicas da folha, como por exemplo, a presença de cavidade estomática e pêlos
(Larcher 2000). As espécies que resistem à deficiência hídrica possuem mecanismos
capazes de manter um alto potencial hídrico ou mantêm uma grande quantidade de água
nas suas células para evitar a desidratação (Larcher 2000). A produção de tecidos e folhas
suculentas é uma estratégia que permite às plantas estocar o recurso limitante e resistir aos
períodos de estiagem.
A incidência de radiação luminosa é indispensável à fotossíntese e, portanto, à
ocorrência das plantas. A absorção e a utilização da radiação fotossinteticamente ativa
pode aumentar até alcançar um ótimo de eficiência pelo aparato fotossintético. Entretanto,

61
o excesso de radiação introduz uma quantidade de energia fotoquímica na folha maior que
a capacidade de utilização dessa energia na fotossíntese, resultando na fotoinibição
(Chazdon 1992). Uma irradiância extremamente forte é capaz de destruir os pigmentos
fotossintéticos e as estruturas dos tilacóides. Um dos primeiros locais a sofrer danos é o
centro de reação do fotossistema II. Quando isso acontece, o transporte de elétrons do
processo fotossintético é interrompido e a eficiência do fotossistema II (Fv/Fm) é
diminuída (Osmond 1994, Larcher 2000). O aporte excessivo de radiação é dissipado por
fluorescência e na forma de calor. Uma parte da dissipação da energia é feita pelos
pigmentos carotenóides que previnem o aparato fotossintético de danos fotooxidativos
(Demmig-Adams & Adams, 1996a). Assim, plantas submetidas ao excesso de radiação
tendem a apresentar uma maior quantidade de carotenóides (em especial de xantofilas) do
que plantas sombreadas (Demmig-Adams & Adams 1996a). Devido a esta proteção que os
carotenóides conferem ao aparato fotossintético sob estresse luminoso, a razão entre o
conteúdo de carotenóides e o total de clorofilas é maior em plantas que habitam ambientes
sujeitos a este estresse (Demmig-Adams & Adams 1996b). A clorofila, como não poderia
deixar de ser, é também um pigmento que apresenta variações no conteúdo de acordo com
a exposição luminosa. A clorofila do tipo a é mais abundante e é o único tipo capaz de
converter energia luminosa em química. A clorofila b é um pigmento acessório
responsável por captar energia luminosa em outra faixa dos comprimentos de onda e
transferi-los para a clorofila a (Taiz & Zeiger 1998). A razão entre as clorofilas a e b é
maior nas plantas expostas a forte radiação luminosa (Murchie & Horton 1997, Lacher
2000).
A maioria dos ambientes apresenta uma combinação de fatores ambientais
estressantes atuando juntos na determinação da distribuição geográfica das espécies bem
como na modelagem das características anatômicas e fisiológicas (Chapin 1991). O

62
excesso de radiação luminosa e altas temperaturas são fatores ambientais que muitas vezes
estão correlacionados. Uma conseqüência grave para as plantas expostas a altas
temperaturas é a desnaturação de proteínas e alterações nas biomembranas que podem
levar à morte celular. As membranas dos tilacóides são sensíveis ao calor e sendo afetadas
levam, conseqüentemente, a diminuição da fotossíntese. O calor afeta o fotossistema II e
pode ter efeito aditivo com o excesso de radiação, levando também a fotoinibição (Larcher
2000). Um mecanismo que ajuda a evitar a perda massiva de água é a transpiração noturna,
no mecanismo fotossintético conhecido como Metabolismo Ácido das Crassuláceas
(CAM). Neoregelia cruenta, assim como outros membros da família bromeliaceae,
apresenta este ciclo fotossintético. Nesta espécie, ele aparentemente é obrigatório (Reinert,
1995) e, portanto, mesmo em condições amenas os indivíduos não são capazes de realizar
o ciclo C3, muito embora, plantas CAM tenham todo o aparato bioquímico necessário para
isto, haja vista que é possível encontrar o ciclo C3-CAM, aonde as condições ambientais
podem determinar a forma de atuação dos estômatos e a atividade da PEP-carboxilase.
Embora, eficiente na restrição à perda d’água, o metabolismo CAM, pode ser
competitivamente desvantajoso em ambientes de baixa luminosidade em relação à plantas
C3, porque para a mesma fixação de carbono, plantas CAM necessitam de
aproximadamente 10 % mais energia luminosa (Winter & Smith, 1996).
Embora, folhas de mecanismo CAM abram os estômatos à noite também estão
sujeitas a transpirarem num fluxo de massa que é proporcional à diferença da concentração
de vapor d’água no interior da cavidade estomática e o ambiente e, mesmo em baixas
temperaturas e luminosidade pode sofrer restrição estomática. Isto pode ser visto em
Neoregelia cruenta em anos atípicos, de seca excessiva, quando a água estocada no tanque
fica reduzida (Reinert, 1995).
As plantas que estão sujeitas a ambientes com deficiência nutricional podem

63
apresentar um maior investimento de biomassa nas raízes e na manutenção de sua
longevidade para aumentar a capacidade de absorção de nutrientes do solo. O estresse
nutricional causa o desenvolvimento de plantas com características xerófitas porque
mantêm a relação entre a concentração de fotossintatos e nutrientes alta (Arens 1958).
Estes vegetais apresentam nanismo, folhas esclerófilas e longevas, baixas taxas
fotossintéticas e crescimento lento (Chapin 1991, Reich & Ellsworth 1992). Um outro
mecanismo que permite evitar o estresse nutricional é a retranslocação de nutrientes de
folhas senescentes para as mais jovens (Taiz & Zeiger, 1998). Os indivíduos desenvolvidos
sob a falta de nutrientes, principalmente de nitrogênio, possuem menor quantidade de
clorofila e massa específica foliar maior (Chapin 1980).
A característica apresentada pelas plantas em resposta ao estresse ambiental pode
ser a mesma para mais de um fator ambiental (Chapin 1991). A massa específica foliar
pode ser influenciada através de seus dois componentes: densidade e espessura, por fatores
ambientais distintos (Witkowski & Lamont 1991). Especificamente a massa específica
foliar é positivamente relacionada com a incidência de radiação luminosa e positivamente
relacionada com a disponibilidade de água; o aumento da massa específica foliar com a
irradiância é resultado de um aumento da espessura foliar causado pelo desenvolvimento
dos tecidos fotossintéticos do mesófilo, enquanto o aumento da massa específica foliar
com o déficit hídrico é resultado da produção de paredes celulares mais espessas e células
menores e mais compactas (Witkowski & Lamont 1991, Niinemets 2001).
O objetivo deste estudo foi identificar as diferenças existentes nos atributos
morfológicos foliares (massa específica foliar, suculência, e área foliar), no conteúdo de
pigmentos (clorofila total, razão clorofila a/b e a razão clorofila/carotenóides) e na
condutância estomática (Gs), causadas pelos microclimas à que estão sujeitas as
subpopulações de Neoregelia cruenta na Restinga Aberta de Clúsia, aonde atua como

64
espécie-focal iniciando a formação de moitas de pequeno porte.
4.2 Materiais e Métodos
4.2.1. Atributos morfológicos: Área Foliar, Suculência e Massa Específica Foliar.
A coleta das folhas para o estudo da Área Foliar (m2), Suculência Foliar (g/m2) e
Massa Específica Foliar (g/m2) foi realizada na estação seca em duas coletas consecutivas
nos anos de 2006 e 2007. Os parâmetros foliares estruturais foram medidos durante a
estação de menor pluviosidade porque as diferenças nas condições micro-climáticas ficam
mais pronunciadas neste período e, portanto, similaridades encontradas nesta estação se
confirmarão na época chuvosa. Parâmetros estruturais foliares, como os medidos aqui, não
variam durante a longevidade foliar e, portanto, apenas uma amostragem anual foi
realizada.
As coletas foram feitas segmentos de rosetas clonais de Neoregelia cruenta
crescendo em micro-hábitats classificados em três categorias: Interior (1) das moitas de
vegetação, com baixa irradiância luminosa; Borda (2) das moitas de vegetação, com
regime de irradiância luminosa variável; Exposto (3) ao sol pleno, com altas taxas de
irradiância luminosa (para melhor definição consulte também o tópico 1.5).
Foram amostrados aleatoriamente quinze segmentos clonais em cada micro-hábitat.
De cada segmento foram amostradas duas rosetas maduras, uma parental e uma filial.
Amostras foram retiradas invariavelmente a partir da quinta folha contadas do centro da
roseta para o exterior. Durante a coleta foram retiradas quatro sub-amostras foliares, com o
auxílio de um cortador circular de área 0,63cm2. As folhas e os discos foliares foram
etiquetados e levados para o laboratório. Medidas de comprimento e largura do limbo
foliar,.comprimento e largura da bainha foliar e a área foliar foram mensurados em 90

65
amostras.
Os discos foliares foram hidratados em uma bandeja cobertos por papel absorvente
embebido em água destilada por aproximadamente 48 horas. Após este período, os discos
foram pesados para a obtenção do peso saturado. Posteriormente os discos foram levados à
estufa a 60ºC e desidratados por 72 horas, e foram novamente pesados para a obtenção do
peso seco. Para as medidas de peso seco e saturados a mesma balança analítica foi
utilizada. Após esses procedimentos, os índices de Suculência e Massa Específica Foliar
puderam ser calculados pelas relações abaixo:
Suculência Foliar (g/m2) = Peso Fresco (g) – Peso Seco (g) Área Foliar (m2) Massa Específica Foliar (g/m2) = Peso Seco (g) Área Foliar (m2)
A relação matemática para o cálculo da suculência foi extraída de Mantovani
(1999) e a relação que estima a Massa Específica Foliar pode ser encontrada em vários
estudos (Witkowski & Lamont 1991, Reich et al. 1991, Reich et al. 1999, Niinemets 2001)
e é derivada do inverso da relação que estima Área Específica Foliar extraída de Larcher
(2000). Foram observadas as recomendações metodológicas feitas por Garnier et al.
(2001).
4.2.2. Pigmentos Fotossintéticos
Para o estudo dos pigmentos fotossintéticos presentes nas folhas de N. cruenta nos
três micro-hábitats foram feitas duas amostragens anuais no período da estação seca, citado
anteriormente. Embora, o valor do conteúdo de clorofila possa sofrer variações sazonais

66
(Soave & Silva 1993), Mantuano (2003) trabalhando com a mesma espécie em três
condições de luminosidade para o mesmo local, encontrou as maiores diferenças na
estação seca e, portanto, mesmo para conteúdo de pigmentos, caráter de variação em curto
espaço de tempo, a amostragem foi feita apenas no período de maior estresse ambiental.
As mensurações dos pigmentos fotossintéticos foram realizadas com dois discos
foliares (de 0,63cm2 de área cada) retirados das mesmas folhas utilizadas na amostragem
dos parâmetros morfológicos foliares. Os discos, imediatamente depois de coletados,
foram colocados em frascos livres de luz com volume conhecido de álcool absoluto. No
laboratório os discos foram fracionados em pequenos pedaços. Após o período de, em
média, vinte e quatro horas, as frações dos discos apresentaram perda total da coloração
verde e, portanto, a clorofila foi considerada extraída. A determinação do conteúdo de
pigmentos foi determinada por espectrofotometria (MacKinney 1941,Vernon 1960) com
sub-amostras de 1ml. As soluções com os extratos de pigmentos fotossintéticos em álcool
foram medidas em três diferentes absorbâncias (470, 648,6 e 664,2nm) em
espectrofotômetro (Beckman Du 520, Alemanha) e as concentrações de pigmentos por
volume de solução (µg/ml) foram calculadas pelas seguintes fórmulas (Lichtenthaler
1987):
Clorofila a= 13,36A664,2 – 5,19A648,6
Clorofila b= 24,43A648,6 – 8,12A664,2
Clorofila a+b= 5,24A664,2 + 22,24A648,6
C x+c= 1000A470 – 2,13Ca – 97,64Cb /209
Em seguida com os valores estimados para a concentração dos pigmentos foram

67
correlacionados com a área dos discos amostrados e os dados foram expressos em mg/cm2
por mililitro de extração.
4.2.3. Condutância Estomática (Gs)
As medidas de condutância estomática (fator estimador da quantidade de
transpiração realizada por área amostrada de folha) foram realizadas com um Porômetro
(AP4, DeltaT, UK) aparelho portátil que detecta a velocidade das variações na umidade
relativa do ar em uma câmara confinada e suprida com ar seco. A condutância estomática é
um parâmetro que informa sobre a quantidade de cavidades estomáticas abertas, e pode
indicar a taxa de transpiração das folhas quando comparadas na mesma temperatura e
umidade relativa do ar. Como N. cruenta apresenta disposição dos estômatos em linha
(veja capítulo 2), a câmara selecionada para medição foi a linear, disponível no
equipamento.
A amostragem foi feita em um ciclo diário, com medições em intervalos de, em
média, três horas, e duração de trinta minutos (6:30; 9:30; 12:30; 15:30; 18:30; 23:30). As
mensurações foram realizadas durante a estação seca em julho/2006.
A amostragem foi realizada em cinco pares de rosetas (roseta parental e roseta
filial), para cada situação de micro-habitat (descritos anteriormente) em cinco moitas de
vegetação na comunidade da Restinga Aberta de Clúsia, próximo à lagoa Comprida.
Neoregelia cruenta apresenta o mecanismo fotossintético do tipo CAM e, portanto, os
maiores valores de condutância serão referidos ao período noturno.
4.2.4. Análise dos dados

68
As análises descritivas dos parâmetros morfológicos foliares (área foliar, suculência
e massa específica foliar) e dos parâmetros que indicam a composição dos pigmentos na
folha (clorofila total, razão clorofila a/b e carotenóides totais) apresentaram tendência a
“anormalidade”. Embora alguns conjuntos de dados tenham se encaixado numa
distribuição normal (teste de Kolmogorov-Smirnov; adaptado de Zar 1996), a maioria não
teve este comportamento, tendo ainda o coeficiente de variação dos parâmetros estado
entre 15 e 30%. Assim, optou-se por tratar estes dados com estatística descritiva e não-
paramétrica.
A comparação dos atributos foliares entre as espécies, e ainda dos pigmentos
fotossintéticos entre as estações seca e chuvosa, foi feita utilizando o teste gráfico nocthed
box plot proposto por McGill, Tukey & Larsen (1978). Neste gráfico as barras inferiores e
superiores indicam os valores mínimos e máximos respectivamente; os limites inferiores e
superiores do box indicam os quartis de 25 e 75% respectivamente; e a constrição lateral
do box indica o intervalo de confiança da mediana, que por sua vez está posicionada no
estrangulamento desta constrição. Os símbolos asteriscos e círculos representam os valores
discrepantes que alcançam 1,5 e 3,0 vezes à distância entre os quartis de 25 e 75%,
respectivamente. As diferenças estatísticas indicadas por este teste ocorrem quando não há
sobreposição dos intervalos de confiança da mediana. Os gráficos foram desenhados com o
auxilio do software Systat 7.0.1.
A comparação condutância estomática (Gs) entre os rametes parentais e filiais nos
três tipos de condições ambientais foi feita com ANOVA com duas fontes de variação
(“two-way”). A diferenciação das médias após a ANOVA foi feita pelo teste de Tukey
destinado a delineamentos com números amostrais desiguais (Zar 1996). Os testes
estatísticos foram feitos com o auxílio do software Statistica 5.0.
A área foliar foi relacionada com as demais medidas tomadas para o tamanho das

69
folhas através da regressão linear.
4.3. Resultados
4.3.1. Atributos morfológicos: Área Foliar, Suculência e Massa Específica Foliar
A área foliar apresentou bastante variação entre os micro-hábitats estudados (200-
500.10-3m-2). Embora a maior parte da amplitude dos dados deva-se a diferenças entre os
micro-hábitats exterior e interior, dentro dos segmentos clonais a diferença entre o ramete
parental e filial foi maior no tratamento borda (Figura 4.1). O comprimento do limbo foliar
foi o melhor estimador da área foliar (R2=0,801, p<0,001). A diferença encontrada entre as
rosetas da situação borda, e os micro-hábitats exterior e interior, não somente se referiu à
área das folhas, mas também foi possível observar que o ângulo de inserção das folhas na
roseta diminuiu, isto é, a roseta teve a abertura do copo restringida, com o aumento da
luminosidade (Figura 2.2B).
A suculência foliar apresentou amplitude entre 0,12 e 0,23 g/cm2. Os indivíduos
crescendo no interior e na borda das moitas apresentaram valores similares entre as folhas
amostradas no ramete parental e filial, nos dois anos seguidos. Já os resultados obtidos para
os indivíduos crescendo no exterior foram estatisticamente diferentes entre as duas rosetas
conectadas no ano de 2007, mas não no ano anterior. Neste sentido, a roseta filial do
segmento clonal crescendo sob sol pleno em 2006 teve suculência similar à amplitude que
abrangeu todas as outras situações amostrais. Os segmentos clonais do ambiente exposto
apresentaram menor suculência que as outras situações amostrais (Figura 4.2)
A massa específica foliar, isto é, a quantidade de gramas contidas em uma unidade
de área, apresentou valores entre 0,022 e 0,041. Novamente o padrão encontrado para a
suculência foliar se repetiu e não houve diferenças significativas entre os rametes parentais

70
e filiais e entre os micro-hábitats nos tratamentos borda e interior, nos dois anos. Na
situação exterior os dois segmentos clonais amostrados em 2006 e o ramete filial
amostrado em 2007 apresentaram valores consideravelmente menores do que a amplitude
do restante dos dados. Apenas as folhas do ramete parental coletadas em 2007
apresentaram valores similares ao restante dos dados (Figura 4.3).

71
Figura 4.1. Representação em Nocthed box plot do parâmetro Comprimento do Limbo
Foliar (cm) de Neoregelia cruenta crescendo nas situações de Exterior (sol pleno), Borda
(regime semi-exposto, na margem das moitas de vegetação) e Interior (sombreadas no
interior das moitas de vegetação). A diferença estatística significativa é alcançada quando
não há sobreposição dos intervalos de confiança (α=95%, p<0,05). Os asteriscos são
pontos discrepantes que representam valores maiores que uma vez e meia a amplitude dos
dados e os círculos valores maiores que três vezes a amplitude. Dados representam
amostragens em Janeiro 2008, na Restinga Aberta de Clúsia do Parque Nacional da
Restinga de Jurubatiba, município de Carapebus.
Interior
Microhabitat
0
10
20
30
40
50
60
70
80
90
Cm
prim
ento
do
Lim
bo F
olia
r (c
m)
Borda Exterior
Ramete parental Ramete filial

72
Figura 4.2. Representação em Nocthed box plot do parâmetro Suculência Foliar (g/cm2)
de Neoregelia cruenta crescendo nas situações de Exterior (sol pleno), Borda (regime
semi-exposto, na margem das moitas de vegetação) e Interior (sombreadas no interior das
moitas de vegetação). A diferença estatística significativa é alcançada quando não há
sobreposição dos intervalos de confiança (α=95%, p<0,05). Os asteriscos são pontos
discrepantes que representam valores maiores que uma vez e meia a amplitude dos dados e
os círculos valores maiores que três vezes a amplitude. Dados representam amostragens em
Julho de 2006 e 2007, na Restinga Aberta de Clúsia do Parque Nacional da Restinga de
Jurubatiba, município de Carapebus.
Borda Exterior Interior
Microhabitat
0.10
0.15
0.20
0.25
Suc
ulên
cia
Fol
iar
(g/c
m2 )
Ramete parental Ramete fillial 2006
Interior
Microhabitat
Borda Exterior
Ramete parental Ramete filial 2007

73
Figura 4.3. Representação em Nocthed box plot do parâmetro Massa Específica Foliar
(g/cm2) de Neoregelia cruenta crescendo nas situações de Exterior (sol pleno), Borda
(regime semi-exposto, na margem das moitas de vegetação) e Interior (sombreadas no
interior das moitas de vegetação). A diferença estatística significativa é alcançada quando
não há sobreposição dos intervalos de confiança (α=95%, p<0,05). Os asteriscos são
pontos discrepantes que representam valores maiores que uma vez e meia a amplitude dos
dados. Dados representam amostragens em Julho de 2006 e 2007, na Restinga Aberta de
Clúsia do Parque Nacional da Restinga de Jurubatiba, município de Carapebus.
Interior
Microhabitat Borda Exterior
Ramete parental Ramete filial
Microhabitat
0.020
0.025
0.030
0.035
0.040
0.045
Pes
o E
spec
ífico
Fol
iar
(g/c
m2 )
Ramete parental
Ramete filial 2006
Borda Exterior Interior
2007

74
4.3.2. Pigmentos Fotossintéticos
O conteúdo total de clorofila das folhas variou entre os valores de 0,5 e 3,0 mg/cm2
(Figura 4.4a). Analisando as diferenças entre os tratamentos podemos observar que a maior
amplitude de diferença ocorre entre os micro-hábitats exterior, completamente exposto à
radiação luminosa e, interior, inteiramente protegido (para conferir a diferença na radiação
incidente consulte tópico 1.5). Também é marcante a diferença entre os rametes parental e
filial nas situações borda e exterior. Neoregelia cruenta apresentou, de maneira geral,
baixos valores de conteúdo de clorofila, quando comparados a outras espécies de restinga,
porém equiparáveis a Aechmea nudicaulis, também espécie da família Bromeliaceae
(Mantuano, 2003).
A amplitude da razão clorofila a/b ficou entre os valores de 1 e 6, com as medianas
ente os valores de 3-4 (Figura 4.4b). As situações exterior e borda tiveram razões similares
entre os micro-hábitats e entre os rametes conectados. A situação interior apresentou
valores similares entre os rametes, porém distinto dos outros dois micro-hábitats
Neoregelia cruenta apresenta pigmentos carotenóides que, em verdade, fazem parte
das características de adorno dos indivíduos e, provavelmente tem função de atração de
espécies polinizadoras ou dispersoras, que são próprias da espécie, tal como o ápice foliar
avermelhado. O conteúdo de carotenóides, portanto, não está restrito a funções
fotossintéticas. Contudo, as amostragens foram feitas na porção mediana do limbo foliar e
admite-se que nesta porção a presença de carotenóides esteja relacionada apenas com a
interação da folha com a luz. A amostragem excluiu os indivíduos de N. cruenta variedade
rubra presentes no local.
Para tanto, os valores do teor de carotenóides totais variou entre 0,1 e 0,6mg/cm2
(Figura 4.4c). Os maiores valores foram encontrados nas folhas crescendo no interior de
moitas de vegetação, e nestes caso não houve diferenças entre os rametes parentais e filiais.

75
Os valores da situação exposto foram menores, mas igualmente não apresentaram
diferenças dentro dos segmentos clonais. Na situação borda a concentração deste pigmento
foi distinta entre os rametes. A variação do teor de carotenóides esteve relacionada com a
luz interceptada pelas folhas mostrando relação desta característica com o aparato
fotoprotetor.

76
Figura 4.4. Teor de pigmentos encontrados em Neoregelia cruenta crescendo nas
situações de Exterior (sol pleno), Borda (regime semi-exposto, na margem das moitas de
vegetação) e Interior (sombreadas no interior das moitas de vegetação). a. Teor de clorofila
total em mg por ml de extração em álcool por área foliar; b. Razão clorofila a/b; c. Teor de
carotenóides totais em mg por ml de extração em álcool por área foliar. A diferença
estatística significativa é alcançada quando não há sobreposição dos intervalos de
confiança (α=95%, p<0,05). Os asteriscos são pontos discrepantes que representam valores
maiores que uma vez e meia a amplitude dos dados e os círculos valores maiores que três
vezes a amplitude. Dados representam amostragens em Janeiro 2008, estação chuvosa.
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Clo
rofil
a to
tal (
mg/
ml/c
m2 ) Ramete parental
Ramete filial
Car
oten
óide
s to
tais
(m
g/m
l/cm
2 )
Interior
Microhabitat
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Borda Exterior
1
2
3
4
5
6
7
8
9
Raz
ão C
loro
fila
a/b
a
b
c

77
4.3.3. Condutância Estomática (Gs)
A maior parte da umidade transpirada através da abertura estomática de N. cruenta
durante a amostragem ocorreu invariavelmente durante os períodos da noite, embora, tenha
ocorrido uma transpiração residual durante o início da manhã e o final da tarde. Este fato
contribui para a caracterização de N. cruenta como CAM obrigatória.
As transpirações estomáticas foram estatisticamente diferentes entre as três
situações microclimáticas estudadas (Figura 4.5). Os indivíduos crescendo protegidos no
interior da moita de vegetação apresentaram os menores valores de condutância
estomática. Os que foram amostrados crescendo na situação borda apresentaram valores
intermediários e, os crescendo no exterior apresentaram as maiores médias.
A regulação estomática em Neoregelia cruenta responde significativamente à
mudanças microclimáticas, como pode ser observado na diferença entre as três situações.
Este resultado indica que esta espécie pode restringir a perda d’água com eficiência em
pequenas mudanças de umidade relativa do ar. Embora, estes dados tenham sido
amostrados durante a estação seca, momento que há a maior amplitude entre os micro-
hábitats e, seja possível que o padrão não se mantenha durante a estação chuvosa, é
importante considerar que as diferenças irão ocorrer também nos momentos de mudança
das massas atmosféricas, isto é, a situação interior de moita tenderá a manter as
características das massas atmosféricas por um período mais longo, do que o exterior, uma
vez que o dossel da moita protege da incidência de ventos. Por tanto, as diferenças
encontradas neste estudo para a estação seca, devem ter correspondência em outros
períodos do ano.
A condutância estomática, que reflete a taxa de transpiração, apresentou diferenças
entre os rametes parental e filial apenas na situação borda. Os segmentos crescendo no
interior e exterior, não apresentaram diferenças intraclonais significativas (Figura 4.6).

78
Figura 4.5. Condutância estomática de Neoregelia cruenta crescendo nas situações de
Exterior (sol pleno), Borda (regime semi-exposto, na margem das moitas de vegetação) e
Interior (sombreadas no interior das moitas de vegetação). As medidas foram realizadas em
duas rosetas conectadas (ramete filial e parental). As comparações foram feitas com
ANOVA (“two-way”; p<0,05), asteriscos denotam diferenças significativas. Os dados
foram coletados em Julho de 2006, na Restinga Aberta de Clúsia do Parque Nacional da
Restinga de Jurubatiba, município de Carapebus.
Con
dutâ
ncia
Est
omát
ica
(mm
ol.m
-2.s
-1)
Tempo (horas)
0 200 400 600 800
1000 1200 1400 1600 Ramete filial Exterior
0 200 400 600 800
1000 1200 1400 1600
12:00 15:00 18:00 21:00 24:00 03:00 06:00 09:00 12:00 15:00 0 200 400 600 800
1000 1200 1400 1600
Borda
Interior
Ramete parental
Ramete filial Ramete parental
*
*

79
Figura 4.6. Diferença na condutância estomática de pares de rosetas conectadas de
Neoregelia cruenta (ramete filial e ramete parental) crescendo nas situações de Exterior
(sol pleno), Borda (regime semi-exposto, na margem das moitas de vegetação) e Interior
(sombreadas no interior das moitas de vegetação). Os dados foram coletados em Julho de
2006, na Restinga Aberta de Clúsia do Parque Nacional da Restinga de Jurubatiba,
município de Carapebus.
12:00 15:00 18:00 21:00 24:00 03:00 06:00 09:00 12:00 -200 -100
0 100 200 300 400 500 600
Exterior Borda Interior
Dife
renç
a G
s (f
ilial
-par
enta
l; m
mol
.m-2
.s-1
)
Tempo (horas)
Ramete parental
Ramete filial

80
4.4. Discussão
4.4.1. Atributos morfológicos: Área Foliar, Suculência e Massa Específica Foliar
A forte variação encontrada nos valores de área foliar parece estar relacionada
majoritariamente com a incidência de luz. Embora, a superfície foliar tenha relação direta
com a área de perda d’água na evapotranspiração e a velocidade de transpiração seja ainda
influenciada pela diferença de temperatura entre a folha e o ar (Larcher 2000); a eficiente
regulação estomática, bem como a presença de tricomas peltados associados ao poro
estomático, a presença de parênquima aqüífero, a capacidade de reservar água livre no
tanque e o metabolismo CAM, sugerem que em Neoregelia cruenta possa haver um
desacoplamento dos índices de área foliar e a disponibilidade de água. Mais além, é
possível que nas folhas desta espécie haja uma setorização das influências ambientais, ou
seja, que a parte da bainha das folhas que forma o tanque se desenvolva em área em locais
de estresse hídrico, contudo a parte do limbo tenho padrão invertido e sofra incremento em
área em locais com menos incidência luminosa, entretanto com menos restrição hídrica
haja vista a subseqüente menor transpiração. Por tanto, o aumento do comprimento foliar
como componente de maior contribuição para o aumento da área foliar do micro-hábitat
exterior em direção ao interior da moita de vegetação pode permitir ao vegetal ajustar o
fornecimento de luz sem, contudo, alterar o balanço hídrico.
Uma forma adicional de ajustar o fornecimento de radiação luminosa, sem alterar a
área foliar, é diminuindo o ângulo de inserção das folhas nos ramos (Larcher, 2000). As
rosetas de N. cruenta expostas à maior radiação tiveram a área de abertura da roseta
reduzida (veja resultados capítulo 2).
A suculência encontrada nas rosetas do interior das moitas são altas e não houve
diferença entre os jovens e adultos, porque o fornecimento de água nesta micro-localidade

81
não é restrito. Nas bordas das moitas de vegetação apesar das diferenças climáticas entre os
dois indivíduos conectados amostrados, os valores medidos são similares. Os atributos
morfológicos foliares avaliados neste estudo apresentaram padrões similares entre os
micro-hábitats interior e borda. Sugerindo que as rosetas que crescem expostas na borda
das moitas de vegetação não estão passando por restrições hídricas e por tanto, não
apresentaram valores similares aos das rosetas de segmentos clonais expostos. É possível
que a conexão vascular que há entre as rosetas possa subsidiar o ramete exposto com
fornecimento de água absorvida na localidade sombreada. É importante observar que
houve variação nos valores expressos entre as situações interior e exterior, permitindo
concluir que Neoregelia cruenta possui um potencial plástico para o ajuste nas
características morfológicas foliares em condições de estresse hídrico e excesso de
luminosidade, porém na situação borda ele não foi expresso.
A suculência reflete o status hídrico da folha durante o seu desenvolvimento
(Larcher 2000) e, nesta espécie é, em grande parte, a capacidade do parênquima aqüífero
em acumular água por unidade de área foliar. Este parâmetro reflete o balanço entre a taxa
de aquisição de água regulada pela disponibilidade hídrica do ambiente e a capacidade de
absorção da roseta e, a taxa de perda regulada pela transpiração estomática e cuticular
(Nobel 1980, Levitt 1989). Nas rosetas crescendo expostas, os segmentos clonais de 2006 e
o ramete parental de 2007 apresentaram os menores valores, refletindo que estão sujeitos
às maiores taxas de transpiração e períodos de restrição hídrica. O ramete filial de 2007
que apresenta a mesma carga genética que o ramete parental e ocorre em condições
ambientais similares, teve valores muito discrepantes de suculência, sendo mais próximos
aos encontrados na situação mésica do interior da moita de vegetação. Neoregelia cruenta
tem reprodução assexuada anual ou sub-anual. As folhas dos rametes jovens amostrados
em 2007 neste estudo têm, provavelmente, menos de um ano de emissão. É possível que os

82
valores encontrados reflitam características juvenis das folhas ou a não exposição aos
períodos de restrição hídrica.
A massa específica foliar no exterior sofreu uma diminuição significativa no ramete
filial crescendo no exterior em 2007 e nos dois segmentos clonais amostrados em 2006.
Enquanto os outros valores (borda e interior; ramete parental e filial) foram similares. O
padrão observado coincide com o encontrado para a suculência foliar, à exceção do ramete
parental do ano de 2007 que teve comportamento atípico. Novamente é possível que a
conexão funcional que une as duas rosetas esteja subsidiando o ramete filial com água e
nutrientes e o ramete parental com fotossintatos.
A massa específica foliar pode integrar dois mecanismos diferentes de
xeromorfismo foliar representados pela alteração dos componentes espessura e densidade
(Witkowski & Lamont 1991, Niinemets 1996, 2001). A espessura foliar é um componente
influenciado pela incidência de radiação solar e pode conferir alta capacidade de
assimilação por unidade de área (Chazdon & Kaufmann 1993, Niinemets 2001). Em geral,
folhas submetidas a forte irradiância apresentam folhas com reduzidas área foliar e
incremento na espessura, que é um reflexo do aumento de camadas e alongamento das
células do parênquima clorofiliano no mesofilo (Boardman 1977, Björkman 1981). A
densidade é influenciada pela disponibilidade hídrica a qual a planta tem acesso (Niinemets
2001). A escassez de água durante a divisão e expansão celular em vegetais gera estruturas
anatômicas com células de lúmen reduzido, compactadas e com paredes celulares espessas.
Esta conformação anatômica tem baixa taxa de assimilação por unidade de área por causa
do decréscimo de compostos assimilativos na folha (Niinemets 2001) e aumento da
resistência a difusão de gases (Reich et.al. 1999, Niinemets 2001). A relação entre a massa
específica foliar e a espessura medidas em uma folha pode apontar qual mecanismo está
atuando no xeromorfismo foliar. Contudo, é importante ressaltar que para espécies que

83
apresentem tecidos de armazenamento de água, a relação entre a espessura foliar total e a
massa específica foliar contém a espessura do parênquima aqüífero com fonte de variação
adicional. Mantuano (2003) estudando a mesma espécie concluiu que a espessura não tem
relação com a massa específica foliar. Mais além, resultados de estudos anatômicos,
apresentados no capítulo três desta tese, indicam que a espessura foliar está mais
fortemente relacionada com a estratificação do parênquima aqüífero do que o clorofiliano
e, sugerem que esta é influenciada pela disponibilidade hídrica e não pela disponibilidade
de radiação luminosa.
4.4.2. Pigmentos Fotossintéticos
O conteúdo e a composição dos pigmentos fotossintéticos nos vegetais são
características fortemente influenciadas pela quantidade e qualidade da radiação luminosa
que incide sobre as folhas. Adicionalmente, todas as adaptações em diferentes condições
luminosas também têm importância sobre os efeitos secundários da radiação, tais como,
altas temperaturas e estresse hídrico periódico (Larcher 2000). Muitos estudos têm tratado
das adaptações ao excesso e a falta de radiação luminosa (para revisão veja Boardman
1977, Björkman 1981). O teor de clorofila nas folhas é um dos atributos que tem intensa
relação com a incidência luminosa e a capacidade fotossintética apresentada pela planta,
muito embora a relação entre esses parâmetros apresente uma gama de respostas possíveis
à variação da radiação luminosa (Murchie & Horton 1997). Tipicamente, o conteúdo de
clorofila por área de folha tende a aumentar de acordo com a intensidade de luz, mostrando
o desenvolvimento do parênquima clorofiliano no mesofilo exposto a forte irradiância.
A incidência luminosa é alta nos ambientes de restingas e, muitas vezes causam
fotoinibição nas taxas fotossintéticas apresentadas pelos vegetais (Reinert 1995, Mantuano
2003). Um dos sintomas da síndrome de fotoinibição é a ocorrência de clorose, ou seja,

84
diminuição no conteúdo de clorofila que é desnaturada pelo excesso de energia não
dissipado (Long et al 1994). A forte variação no conteúdo de clorofila total no gradiente
entre o ambiente exposto e protegido mostra que este parâmetro é fortemente afetado pela
luz. Haja vista a intensidade luminosa a que estão sujeitas as rosetas de N. cruenta nas
restingas fluminenses, a redução da clorofila provavelmente não é uma característica
relacionada à pobreza nutricional, mas sim um sintoma da fotoinibição crônica. Por ser um
sintoma e não um caráter expresso pelo vegetal, a diferença encontrada na situação borda
apenas expressa o delineamento experimental, e não pode ser relacionada com a integração
intraclonal.
A razão entre a clorofila a e b nas espécies vegetais apresenta altos valores
associados a ambientes com forte incidência de radiação luminosa (Boardman 1977,
Demmig-Adams & Adams 1996b, Murchie & Horton 1997). A clorofila do tipo a é a única
envolvida na fotossíntese diretamente. Os pigmentos acessórios, como a clorofila do tipo b,
captam a energia luminosa e a transferem por ressonância para a clorofila a. Assim,
quando a planta está sombreada apresenta quantidades, relativamente, maiores de clorofila
b, como uma estratégia a otimização na interceptação e absorção de energia luminosa e, de
outra forma, apresenta valores relativos altos de clorofila a em ambientes iluminados para
manter altas capacidades fotossintéticas (Taiz & Zeiger 1998, Larcher 2000). As diferenças
na razão clorofila a/b podem se originar da proporção entre os números de complexos
antenas (com maior teor de clorofila b) e dos centros de reação do fotossistema II (com
maior teor de clorofila a) (Demmig-Adams & Adams 1996b). O mecanismo que determina
respostas na composição das clorofilas a e b é, então, uma menor ênfase na captação de luz
em proporção ao aparato fotossintético. Entretanto, este mecanismo não é capaz de
explicar a relação entre o ambiente e a composição dos pigmentos em todos os casos.
Provavelmente outros mecanismos que também influenciam a composição dos pigmentos

85
fotossintéticos devem atuar conjuntamente. A determinação da razão clorofila a/b expressa
pela condição ambiental de forte radiação luminosa pôde superar a predisposição genética
de 28 espécies britânicas (Rosevear et al. 2001), muito embora um estudo nos trópicos
tenha relatado que espécies de estágio sucessional secundário desenvolvidas sob altas
irradiâncias não tenham mostrado alteração neste atributo (Krause et al. 2001). A razão
clorofila a/b não sofreu alteração em N. cruenta neste estudo. Este resultado pode estar
ligado ao regime de incidência luminosa das restingas, no qual a situação interior de moitas
de vegetação pode não estar representando um ambiente sombreado verdadeiramente. Isto
é, a incidência luminosa que incide sobre rosetas crescendo nesta situação já é suficiente
para manter altas taxas fotossintéticas (veja resultados do crescimento apresentados no
capítulo dois desta tese) e a diferença de radiação para os micro-hábitats borda e exterior
representam apenas excedente.
O teor de carotenóides nas plantas tende a ser maior quanto mais intensa é a
exposição a radiação luminosa. A presença de carotenóides, principalmente de xantofilas,
nas folhas confere ao aparato fotossintético proteção ao estresse luminoso através da
dissipação da energia de excitação (Demmig-Adams & Adams 1996a e b). Os resultados
encontrados neste estudo corroboram a afirmativa de excesso de luminosidade na condição
exterior, uma vez que esta condição apresentou os maiores valores de concentração de
carotenóides.
4.4.3. Condutância Estomática
Espécies que apresentam o ciclo fotossintético CAM apresentam a abertura
estomática preferencialmente à noite, período em que a temperatura das folhas diminui por
causa da extinção da incidência luminosa. Este é um mecanismo que tem sido relatado
como freqüente em ambientes áridos (Larcher 2000). Apesar da redução das perdas de

86
água na evapotranspiração, é possível que mesmo concentrando as trocas gasosas à noite
ainda possam ser atuantes fatores que afetem a condutância estomática, tal como a
umidade relativa do ar. A velocidade de difusão de água entre a folha e o ambiente é
diretamente relacionada com a diferença entre a umidade relativa do ar na cavidade
subestomática e o ar circundante (Nobel 1980).
O ambiente de restinga é um ambiente com certo grau de aridez. Embora, em
grande parte do litoral do estado do Rio de Janeiro a precipitação anual esteja próximo aos
valores de 2000-2200mm, o solo arenoso que apresenta caracteristicamente baixa retenção
hídrica, limita o fornecimento de água ao curto período de tempo após as chuvas. Mas,
mais ainda, a demanda evaporativa do ar e a incidência de ventos podem ser os fatores
decisivos no balanço hídrico da vegetação crescendo na restinga (Barbiere 1984). O
ambiente costeiro é certamente um local propício a incidência de ventos (Dau 1960, Martin
et al 1993). No Parque Nacional da Restinga de Jurubatiba, o vento SSE é o de maior
intensidade e o segundo em freqüência, marcando a entrada das frentes frias (Correia,
1998).
As diferenças transpiracionais observadas nas rosetas de N. cruenta entre as
localidades refletem provavelmente diferenças nas características atmosféricas do ar retido
sobre a moita de vegetação e o exterior. As movimentações de ar entre o oceano e as
planícies costeiras podem contribuir para a formação dos gradientes de umidade entre o
interior das moitas de vegetação e o exterior, contudo estudos que tenham avaliado a
proximidade com o mar como um fator que afeta a vegetação de restinga não encontraram
resultados significativos (Reinert 1995, Pimentel 2002).
A remoção da camada limítrofe entre a folha e o ar circundante pode aumentar
significativamente a taxa de transpiração nas plantas (Nobel 1991). Em especial no caso de
plantas CAM, não há influência da quantidade de radiação interceptada por áreas foliares

87
maiores e, assim, podemos reduzir a influência da área foliar ao espessamento da camada
limítrofe. Se considerarmos que a proteção do dossel da moita reduz a circulação de ar
local, é possível prever as diferenças transpiracionais encontradas.

88
Capítulo 5
Conclusões gerais

89
5. Discussão
Neoregelia cruenta é uma bromélia CAM tanque, que possui tricomas
especializados recobrindo a epiderme e que foram relatados com tendo função de absorção
de água e nutrientes (Martin 1994, Benzing, 2000). Isto implica que a fisiologia desta
espécie além de permitir o uso eficiente (Metabolismo CAM) e o acúmulo de água
(suculência foliar), pode ser independente das raízes para a absorção mineral e para o
balanço hídrico. Assim, como em outras espécies da família, N. cruenta apresenta
resistência na colonização de ambientes áridos, tais como no hábito epífitico e no
estabelecimento em substratos arenosos e rochosos. Estas características provavelmente
levaram à noção de que esta pode ser uma espécie focal nas vegetações abertas de restingas
(Lacerda & Hay, 1977; 1978; Hay & Lacerda, 1980, Lacerda, Andrade 1992, Mantovani
2002)
Entretanto, não apenas os aspectos fisiológicos conferem robustez a N. cruenta,
mas também aspectos arquiteturais, dentre eles, a capacidade de reprodução clonal e
forrageamento lateral do ambiente. Neste sentido um indivíduo clonal amplia as suas
chances de encontrar uma mancha favorável de recurso no ambiente (Stuefer 1997).
Estudos experimentais mostram que a especialização funcional de partes do indivíduo
clonal a situações locais de ambientes heterogêneos são favoráveis ao crescimento do
indivíduo como um todo.
Esta tese mostrou que N. cruenta apresenta características plásticas (abertura da
roseta, comprimento foliar, espessura do parênquima aqüífero, densidade de tricomas na
epiderme adaxial e condutância estomática) e não plásticas (largura do limbo foliar,
comprimento do estolão, espessura do parênquima clorofiliano, razão clorofila a/b).
Entretanto, por vezes as características mensuradas embora tenham variado entre rosetas
conectadas estiveram mais relacionadas a síndromes de estresse luminoso (teor de clorofila

90
e carotenóides).
A especialização intraclonal somente pode favorecer a produtividade em ambientes
heterogêneos, em face à possibilidade de translocação de recursos entre os rametes. Os
resultados encontrados neste estudo mostram que, de fato, as rosetas de N. cruenta são
mais freqüentes no micro-hábitat heterogêneo da borda. O crescimento populacional foi
negativo apenas para a situação completamente exposto ao sol pleno, o que juntamente
com a incapacidade de ajuste plástico do parênquima clorofiliano e a forte clorose
encontrada, corrobora a idéia de que o ambiente luminoso da restinga é excessivo,
característico de ambientes costeiros.
As características relacionadas ao balanço hídrico, de outra forma, mostraram que
são passíveis de alguns ajustes. O número de tricomas foi maior nas folhas expostas do que
nas folhas protegidas nos segmentos clonais crescendo no micro-hábitat borda.
Provavelmente ajudam a compensar o tempo de retenção hídrica da água no copo com a
velocidade de absorção. O estoque de água em estratos do parênquima aqüífero foi maior
também nos rametes expostos quando comparados aos seus pares no micro-hábitat borda,
permitindo resistência por mais tempo à dessecação. Estes resultados sugerem mais
independência das rosetas de N. cruenta do ambiente hídrico do que do ambiente
luminoso.
Neste estudo foi proposto que ocorra uma integração do fluxo do xilema entre os
rametes parentais e filiais, que seria direcionado por diferenças nas taxas de transpiração
estomática. O dissimilaridade das taxas transpiracionais entre os rametes conectados ocorre
em função das distintas condições abióticas entre o interior e exterior das moitas de
vegetação nas restingas abertas. Distinção que pode ser “percebida” por conexões de
rametes crescendo nas bordas das moitas de vegetação, isto é, parcialmente no interior e
parcialmente no exterior. Embora, N. cruenta seja CAM, e abra os estômatos à noite, a

91
proteção contra o vento e o tamponamento das amplitudes térmicas e da umidade relativa
são capazes de criar ambientes de trocas gasosas distintos, como mensurado no capítulo 4.
O desacoplamento das taxas de transpiração e o fluxo direcional do xilema como
mecanismos que induzem o crescimento clonal são consonantes com os resultados de
outros estudos sobre a biologia de bromélias das restingas fluminenses e sobre as
condições ambientais em restingas abertas. Dau (1960) mostrou que o ambiente do interior
e do exterior das moitas de vegetação são distintos, tanto durante o dia quanto à noite.
Reinert (1995) encontrou diferenças transpiracionais em N. cruenta relacionadas as faixas
de cordões arenosos, que em última análise apresentaram diferenças nas médias de
Umidade Relativa. Mantovani (2002) sugere que N. cruenta pode ter papel importante na
expansão lateral das moitas de vegetação por ser muito freqüente nas bordas. Assim, como
Sampaio (2004) apresenta resultados do crescimento direcional de A. nudicaulis do interior
para a borda. Ambos podem ser subsidiados pelo mecanismo de fluxo diferencial do
xilema, que implica no direcionamento da região de menor taxa de transpiração (protegido)
para a região de maior condutância estomática (borda).
Durante toda a observação, o crescimento de N. cruenta seguiu necessariamente a
direção acropétala. Nestes sentido, é possível que a integração fisiológica ocorra apenas
unidirecionalmente, isto é, apenas nos vasos xilemáticos, mas a retranslocação de
fotossintatos não ocorra no sentido inverso. Isto permitiria a independência do substrato,
mas não do ambiente luminoso.
Os resultados encontrados apontam para a possibilidade de um trade-off entre a
sobrevivência da roseta, o tamanho e a reprodução clonal. A sobrevivência das rosetas
variou significativamente entre os micro-hábitats e pareceu ser inversamente proporcional
a produção de novos rametes. Em situações mésicas (interior das moitas) o investimento no
maior tamanho da roseta retardou a produção de novos rametes. Muito embora a baixa taxa

92
de mortalidade tenha mantido a taxa de crescimento populacional positiva e similar a
situação borda, diferente do encontrado por Sampaio (2004) para A. nudicaulis. A
reprodução clonal em N. cruenta não á apenas uma extensão do crescimento vegetativo. O
estresse ambiental parece estimular o investimento em crescimento clonal, que dependendo
da intensidade e de fatores intrínsecos do status fisiológico da planta parental pode
provocar a sua senescência.
Os resultados permitem concluir que a variação espacial e temporal das condições
ambientais pode influenciar o crescimento, a estabilidade e a manutenção das sub-
populações de N. cruenta. É possível que mudanças ambientais que diminuam a estação
chuvosa e, portanto, o período de nebulosidade (aumentando a radiação total), afetem a
sub-população crescendo exposta ao sol pleno. Porém, uma extinção local é improvável
porque os micro-hábitats protegido e semi-exposto poderiam fornecer novos indivíduos.
Embora o risco de extinções locais seja alto para a família Bromeliaceae (Siqueira Filho &
Tabarelli 2006). Contudo, o papel de iniciadora de sucessão apresentado por Andrade
(1992) e de espécie-chave no melhoramento do solo apresentado por Hay & Lacerda
(1981) ficaria comprometido.

93
Referências Bibliográficas

94
Referências Bibliográficas
Alpert, P. & Mooney, H.A. 1986. Resource sharing among ramets in the clonal herb, fragarai chiloensis. Oecologia, 70: 227-233.
Alpert, P.& Stuefer, J.F. 1997. Division of labour in clonal plants. In de Kroon, H. & van
Groenendael, J. (eds.) The ecology and evolution of clonal plants, Academic Publishing , Amsteram.
Andrade, L.R. 1992. Análise da estrutura e composição específica das comunidades
vegetais de moitas de restinga, em Maricá, RJ. Dissertação de M.Sc., Departamento de Ecologia /UNB, Brasília, Brasil.
Araujo, D.S.D. & Henriques, R.P.B. 1984. Análise florística das restingas do estado do Rio
de Janeiro. In: Lacerda, L.D., Araujo, D.S.D., Cerqueira, R. & Turcq, B. (eds). Restingas: Origem, Estrutura e Processos. Niterói, CEUFF, pp. 159-194.
Araujo, D.S.D., Scarano, F.R., Sá, C.F.C., Kurtz, B.C., Zaluar, H.L.T., Montezuma,
R.C.M. & Oliveira, R.C. 1998. Comunidades vegetais do Parque Nacional da Restinga de Jurubatiba. In: Esteves, F.A., (ed.) Ecologia das lagoas costeiras do Parque Nacional da Restinga de Jurubatiba e do município de Macaé, Macaé, NUPEM-UFRJ, pp. 39-63.
Araujo, D.S.D. 2000. Análise florística e fitogeográfica das restingas do Estado do Rio de
Janeiro. Tese de D.Sc., PPGE/UFRJ, Rio de Janeiro, RJ, Brasil. Archer, S. 1989. Have Southern Texas savannas been converted to woodlands in recent
history? The American Naturalist, 134(4): 545-561. Archer, S., Scifres, C., Bassham, C.R. & Maggio, R. 1988. Autogenic sucession in a
subtropical savanna: conversion of grassland to thorn woodland. Ecological Monographs, 58(2):111-127.
Arens, K. 1958. Considerações sobre as causas do xeromorfismo foliar. Bol. Fac. Fil.
Ciênc. Letr. Univ. São Paulo, 305 Botânica, 22: 25-56. Arruda, R.C.O. 2003. Características foliares em espécies de cerrado e restinga. Tese de
Doutorado, USP, São Paulo. Assunção, J & Nascimento, M.T. 2000. Estrutura e composição florística de quatro
formações vegetais de restinga no complexo lagunar Grussaí/Iquipari, São João da Barra, RJ, Brasil. Acta Botanica Brasílica, 14(3): 301-315.
Barbiére, E.B. 1984. Cabo frio e Iguaba grande, dois microclimas distintos a um curto
intervalo espacial. In Lacerda, L.D., Araujo, D.S.D., Cerqueira, R. & Turcq, B. (Orgs.) Restingas: origem, estrutura e processos, Niterói, CEUFF, 3-14..
Barnes, P.W. & Archer, S. 1996. Influence of an overstorey tree (Prosopis glandulosa) on
associated shrubs in a savanna parkland: implications for patch dynamics.

95
Oecologia, 105: 493-500. Benzing, D.H., Seemann, J. & Renfrow, A. 1978. The foliar epidermis in Tillandsioideae
(Bromeliaceae) and its role in habitat selection. American Journal of Botany, 65(3): 359-365.
Benzing, D.H. & Burt, K.M. 1970. Foliar permeability among twenty species of th Bromeliaceae. Bulletin of the Torrey Botanical Club, 97(5): 269-279.
Benzing, D.H. 2000. Bromeliaceae: profile of an adaptative radiation. Cambridge,
Cambridge Univ. Press. Berg, A. Birgit, O.Mattos, E.A., Duarte, H.M. & Lüttge, U. Expresssion of crassulacean
acid metabolism in Clusia hilariana Schlechtendal in diferent stages of development in the field. Trees, Structure and Function, 18(5): 553-558.
Bertness, M.D. & Callaway, R.M. 1994. Positive interactions in communities. Trends in
Ecology and Evolution, 9:191-193. Björkman O. 1981. Responses to different quantum-flux density. In: Lange OL, Nobel PS,
Osmond CB, Ziegler H, eds. Encyclopaedia of Plant Physiology 12 A. Berlin: Springer, 57-107.
Boardman, N.K. 1977. Comparative photosynthesis of sun and shade plants. Annu. Rev.
Plant Physiology, 28:355-377. Bradshaw, A.D. 1965. Evolutionary significance of phenotypic plasticity in plants.
Advances in Genetics, 13: 115-155. Braz, M.I.G. 2002. Comparação dos atributos foliares em espécies de Cerrado e Restinga.
Dissertação de M.Sc. PPGE/UFRJ, Rio de Janeiro, RJ, Brasil. Brouwer, R. 1963. Some aspects of the equilibrium between overground and aboveground
plant parts. Meded. Inst. Biol. Scheik. Onderz., 213; 31-39. Brouwer, R. 1983. Functional equilibrium: sense or non-sense? Netherlands Journal of
Agricultural Science, 31: 335-348. Callaway, R.M. 1992. Effect of shrubs on recruitment of Quercus douglasii and Quercus
lobata in California. Ecology, 73:2118-2128. Callaway, R.M. 1995. Positive interactions among plants. Botanical Review, 61: 306-349. Chapin III, F.S. 1980. The mineral nutrition of wild plants. Annual Review Ecology
Systematic, 11: 233-260. Chapin III, F.S. 1991. Integrated responses of plants to stress. BioScience, 41: 29-36. Chazdon, R.L. & Kaufmann, S. 1993. Plasticity of leaf anatomy of two rain forest shrubs
in relation to photosynthetic light acclimation. Functional Ecology, 7: 385-394.

96
Chazdon, R.L. 1992. Photosynthetic plasticity of two rains forest shrubs across natural gap transects. Oecologia, 92: 586-595.
Cogliatti-Carvalho, L. Freitas, A.F.N., Rocha, C.F.D. & Sluys, M.V. 2001. Variação na
estrutura e na composição de Bromeliaceae em cinco zonas de restinga no Parque Nacional da Restinga de Jurubatiba, Macaé, RJ. Revista Brasileira de Botânica, 24(1): 1-9.
Cordeiro, S.Z. 2005. Composição e distribuição da vegetação herbácea em três áreas com
fisionomias distintas na Praia do Peró, Cabo Frio, RJ, Brasil. Acta Botanica Brasilica, 19(4): 679-693.
Correia, C.M.B. 1998. Estrutura de populações, associação entre quatro espécies-chaves e
formação de moitas em restinga de Clusia no norte fluminense. Dissertação de M.Sc. PPGE/UFRJ, Rio de Janeiro, RJ, Brasil.
Crawford, R.M.M. 1989. Studies in plant survival. Oxford, Blackwell Science, 296pp. Dau, L. 1960. Microclimas das restingas do sudeste do Brasil. Arquivos do Museu
Nacional, vol. L: 79-133. Davies, P.J. 1987. Plant hormones and their role in plant growth and development.
Amsterdan, KluwerAcademic Publishers. de Kroon, H. & Hutchings, M.J. 1995. Morphological plasticity in clonal plants: the
foraging concept reconsidered. Journal of Ecology, 83: 143-152. Demmig-Adams, B. & Adams, W.W. 1992. Photoprotection and other responses of plants
to high light estress. Annual Raview of Plant Physiology and Plant Molecular Biology, 43: 599-626.
Demmig-Adams, B. & Adams, W.W. 1996a. Chlorophyll and carotenoid composition in
leaves of Euonymus kiautschovicus acclimated to different degrees of light stress in the field. Australian Journal of Plant Physiology, 23:649-659.
Demmig-Adams, B. & Adams, W.W. 1996b. The role of xanthophyll cycle carotenoids in
the protection of photosynthesis. Trends in Plant Science, 1:21-26. Fernandes, J. Chaloub, R.M. & Reinert, F. 2002. Influence of nitrogen supply on the
protective response of Neoregelia cruenta (Bromeliaceae) under high and low light intensity. Functional Plant Biology, 39: 757-762.
Ferri, M.G. 1955. Contribuições ao conhecimento do Cerrado e da Caatinga. Estudo
comparativo da economia de água de sua vegetação. Boletim da Faculdade de Filosofia, Ciência e Letras da Universidade de São Paulo, 12 (195): 1-170.
Fialho, R.F. & Furtado, A.L.S. 1993. Germination of Erythroxylum ovalifolium
(Erythroxylaceae) seeds within terrestrial bromeliad Neoregelia cruenta. Biotropica, 25(3): 359-362.

97
Flores-Martínez, A., Ezcurra, E. & Sánchez-Colón, S. 1998. Water availability and the competitive effect of a columnar cactus on its nurse plant. Acta Oecologica, 19(1): 1-8.
Franco, A.C. & Nobel, P.S. 1988. Interactions between seedlings of Agave deserti and the
nurse plant Hilaria rigida. Ecology, 69: 1731-1740. Franco, A.C. & Nobel, P.S. 1989. Effect of nurse plants on the microhabitat and growth of
cacti. Journal of Ecology, 77: 870-886. Franco, A.C. & Nobel, P.S. 1990. Influences of root distribution and growth on predicted
water uptake and interspecific competition. Oecologia, 82:151-157. Franco, A.C., Valeriano, D.M., Santos, F.M., Hay, J.D.; Henriques, R.P.B. & Medeiros,
R.A. 1984. Os microclimas das zonas de vegetação da praia da restinga de Barra de Maricá, Rio de Janeiro. In: Lacerda, L.D.; Araújo, D.S.D.; Cerqueira, R. & Turc, B. (Eds.) Restingas: origem, estrutura e processos. CEUFF, Niterói, pp. 413-423.
Franco-Pizaña, J., Fullbright, T.E., Gardiner, D.T. & Tipton, A.R. 1996. Shrub emergence
and seedling growth in microenvironments created by Prosopis glandulosa. Journal Vegetation Science, 7: 257-264.
Fulbright, T.E., Kuti, J.O. & Tipton, A.R. 1995. Effects of nurse-plant canopy
temperatures on shrub seed germination and seedling growth. Acta Oecologica, 16(5): 621-632.
Gass, L. & Barnes, P. 1998. Microclimate and understory structure of live oak Quercus
fusiformis clusters in central texas, USA. The Southwest Naturalist, 43(2): 183-194. Garcia-Franco, J.G. & Rico-Gray, V. 1995. Population structure and clonal reproduction in
Bromelia pinguin L. (Bromeliaceae) in dry forests of coastal Veracruz, Mexico. Tulane Studies of Zoology and Botany, 30: 27-37.
Garnier, E., Shipley, B., Roumet, C. & Laurent, G. 2001. A standardized protocol for the
determination of specific leaf area and leaf dry matter content. Functional Ecology, 15: 688-695.
Griffiths, H. 1988. Carbom balance during CAM: an assesment of respiratory CO2
recycling in the epiphytic bromeliads Aechmea nudicaulis and Aechmea fendleri. Plant, Cell and Environment, 11: 603-611.
Hay, J.D. & Lacerda, L.D. 1980. Alterações nas características do solo após a fixação de
Neoregelia cruenta (R. Grahan L. Smith (Bromeliaceae), em um ecossistema de restinga. Ciência e cultura, 32(7): 863-867
Hay, J.D., Lacerda, L.D. & Tan, A.L. 1981. Soil cation increase in a tropical sand dune
ecosystem due to a terrestrial bromeliad. Ecology, 65(5): 1392-1395. Hay, J.D. & Lacerda, L.D. 1984. Ciclagem de nutrientes do ecossistema de restinga. In:
Lacerda, L.D., Araujo, D.S.D., Cerqueira, R. & Turcq, B. (eds). Restingas: Origem,

98
Estrutura e Processos. Niterói, CEUFF, pp. 459-475. Henriques, R.P.B., Araujo, D.S.D. & Hay, J.D. 1986. Descrição e classificação dos tipos
de vegetação da restinga de Carapebus, Rio de Janeiro. Revista Brasileira de Botânica, 9: 173-189.
Hutchings, M.J. & Wijesinghe, D.K. 1997. Patchy habitats, division of labour and growth
dividends in clonal plants. Trends in Ecology & Evolution, 12: 390-394. Jackson, L.W.R. 1967. Effect of shade on leaf structure of deciduous tree species. Ecology,
48(3): 498-499. Jordan, P.W. & Nobel, P.S. 1979. Infrequent establisment of seedlings of Agave deserti
(Agavaceae) in the Northwestern Sonoran desert. American Journal of Botany, 66(9): 1079-1084.
Köppen, W. 1948. Climatologia: Com un estudio de los climas de la tierra. Fondo de
Cultura Economica, México. Krause, G.H., Koroleva, O.Y., Dalling, J.W. & Winter, K. 2001. Acclimation of tropical
tree seedlings to excessive light in simulated tree-fall gaps. Plant, Cell and Environment, 24: 1345-1352.
Lacerda, L.D., Araujo, D.S.D. & Maciel, N.C. 1993. Dry coastal ecosystems of tropical Brazilian coast.. In: E. Van de Maarel (ed.). Dry coastal ecosystems: Africa, America, Asia, Oceania. Elsevier, Amsterdam, pp. 477-493.
Lacerda, L.D. & Hay, J.D. 1982. Habitat of Neoregelia cruenta (Bromeliaceae) in coastal
sand dunes of Maricá, Brazil. Revista de Biología Tropical,30(2): 171-173. Lambers, H. Chapin III, F.S. & Pons, T.L. 1998. Plant physiological ecology. New York,
Spinger-Verlag Inc. Larcher, W. 2000. Ecofisiologia Vegetal. 1ed. (Trad. C.H. Prado), São Carlos, RiMA. Levin, S. 1992. The problem of pattern and scale in ecology. Ecology, 73: 1943-1967. Levitt, J. 1989. Responses of plants to environmental stress. Academic Press, New York. Lichtenthaler, H.K. 1987. Chlorophylls and carotenoids: pigments of photosynthetic
biomembranes. Methods in enzymology, 148: 350-382. Lovett Dous, L. 1981. Intraclonal variation and competition in Ranunculus repens. New
Phytologist, 89(3): 495-502. Lovett Doust, L & Lovett Doust, J. 1982. The battle strategies of plants. New Scientist, 95:
81-84. MacKinney, G. 1941. Absorption of light by chlorophyll solutions. Journal of Biological
Chemistry, 140: 315-322.

99
Mantovani, A. 1999. A method to improve leaf succulence quantification. Brazilian Archives of Biology and Technology, 42(1): 9-14.
Mantovani, A. Iglesias, R.R. 2001. Bromélias terrestres na restinga de Barra de Maricá,
Rio de Janeiro: influência sobre o microclima, o solo e a estocagem de nutrientes em ambientes de borda de moitas. Leandra, 16: 17-37.
Mantovani, A. 2002. Bromélias terrestres na Restinga de Barra de Maricá: alocação de
recursos na floração, germinação de sementes, estabelecimento e papel facilitador.Tese de Doutorado, PPGE/UFRJ, Rio de Janeiro.
Mantuano, D.G. 2003. Distribuição espacial e respostas ecofisiológicas do sub-bosque da
formação arbustiva aberta de Clusia à diminuição da cobertura da espécie-focal Clusia hilariana no PARNA da Restinga de Jurubatiba. Dissertação de Mestrado, PPGE/UFRJ, Rio de Janeiro.
Mantuano, D.G., Barros, C.F. & Scarano, F.R. 2006. Leaf anatomy within and between
three “restinga” populations of Erythroxylum ovalifolium Peyr. (Erythroxylaceae) in Southeast Brazil. Revista Brasileira de Botânica, 29(2): 209-215.
Martin, L., Suguio, K. & Flexor, J.M. 1993. As flutuações de nível do mar durante o
quaternário superior e a evolução geológica de “deltas” brasileiros. Boletim – IG-USP Publ. Esp., 15:1-186.
Martin, C. 1994. Physiological ecology of the Bromeliaceae. The Botanical Review, 60(1):
1-82. Mattos, E.A. 1998. Perspectives on comparative ecophysioloy of some Brazilian
Vegetation Types: leaf CO2 and H20, gas exchange, chlorophyll a fluorescence and carbon isotope discrimination. In F.R. Scarano & Franco, A.C. (Orgs) Ecophysiological strategies of xerophytic and amphybious plants in the neotropics, Computer & Publish ed., Rio de Janeiro, IV: 1-13.
Mattos, E.A., Braz, M.I.G., Cavalin, P.O., Rosado, B.H.P., Gomes, J.M., Martins, L.S.T. &
Arruda, R.C. 2004. Variação espacial e temporal em parâmetros fisioecológicos de plantas. In Rocha, C.F.D., Esteves, F.R. & Scarano, F.R. (Orgs.) Pesquisa de Longa Duração na Restinga de Jurubatiba: ecologia, história natural e conservação. RiMA, São Carlos, 99-116.
McGill, R., Tukey, J.W. & Larsen, W.A. 1978. Variations of Box Plots. The American
Statistician, 32(1): 12-16. Medina, E., Delgado, M., Troughton, J.H. & Medina, J.D. 1977. Physiological ecology of
CO2 fixation in Bromeliaceae. Flora, 166: 137-152. Munzbergova, Z. & Ward, D. 2002. Acacia trees as keytone species in Negev desert
ecossystems. Journal of Vegetation Science, 13:227-236. Murchie, E.H. & Horton, P. 1997. Acclimation of photosynthesis to irradiance and spectral
quality in British plant species: chlorophyll content, photosynthetic capacity and

100
habitat preference. Plant, Cell and Environment, 20: 438-448. Negrelle, R.R.B. & Muraro, D. Aspectos fenológicos e reprodutivos de Vriesea incurvata
Gaudich (Bromeliaceae). Acta Scientiarum (UEM), 28(2):95-102. Niinemets, Ü. 1996. Plant growht form alters the relationship between foliar morphology
and species shade tolerance ranking in temperate woody taxa. Plant Ecology, 124(2): 145-156.
Niinemets, Ü. 2001. Global-scale climatic controls of leaf dry mass per area, density, and
thickness in trees and shrubs. Ecology, 82: 453-469. Nobel, P.S. 1991. Physiochemical Plant Physiology. Academic Press, San Diego. Nunes-Freitas, A.F., Rocha-Pessôa, T.C., Cogliatti-Carvalho, L. & Rocha, C.F.D. 2006.
Bromeliaceae da restinga da Reserva Biológica Estadual da Praia do Sul: composição, abudância e similaridade da comunidade. Acta Botanica Brasilica, 20(3); 709-717.
Oliveira, M.G.N. & Rocha, C.F.D. 1997. O efeito da complexidade da bromélia-tanque
Neoregelia cruenta (R.Graham) L.B. Smith sobre a comunidade animal associada. Bromélia, 4(2):13-22.
Oliveira, M.G.N., Rocha, C.F.D. & Bagnall, T. 1994. A comunidade animal associada à
bromélia-tanque Neoregelia cruenta (R. Graham) L.B. Smith. Bromélia, 1(1): 22-29.
Ormond, W.T. 1960. Ecologia das restingas do sudeste do Brasil. Arquivos do Museu
Nacional, vol. L.:185-236. Osmond, C.B. 1994. What is Photoinhibition? Some insights from comparisons of shade
and sun plants. In: Baker, NR, Bowyer, JR, (eds.) Photoinhibition of photosynthesis – from molecular mechanism to the field, Oxford, Bios Scientific, pp.1-24.
Pereira, M.C.A., Araujo, D.S.D. & Pereira, O.J. 2001. Estrutura de uma comunidade
arbustiva da restinga de Barra de Maricá – RJ. Revista Brasileira de Botânica, 24(3): 273-281.
Pereira, M.C.A., Cordeiro, S.Z. & Araujo, D.S.D. 2004. Estrutura do estrato herbáceo na
formação aberta de Clusia do Parque Nacional da Restinga de Jurubatiba, RJ, Brasil. Acta Botanica Brasilica, 18(3): 677-687.
Pierce, S., Maxwell, K., Griffiths, H. & Winter, K. 2001. Hydrophobic trichome layers and
epicuticular wax powders in Bromeliaceae. American Journal of Botany, 88(8): 1371-1389.
Pimentel, M. 2002. Variação espacial na estrutura de comunidades vegetais da formação
arbustiva aberta de Clúsia no Parque nacional da Restinga de Jurubatiba, RJ. Tese de Doutorado, PPGE/UFRJ, Rio de Janeiro.

101
Pittendrigh, C.S. 1948. The bromeliad-Anopheles-Malaria complex in trinidad. I. – The bromeliad flora. Evolution, 2: 58-89.
Pons, T.L., Jordi, W. & Kuiper, D. 2001. Acclimation of plants to light gradients in leaf
canopies: evidence for a possible role in cytokinins transported in the transpiration stream. Journal of Experimental Botany, 52(360): 1563-1574.
Reich, P.B. & Ellsworth, D.S. 1992. Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems. Ecological monographs, 62: 365-392.
Reich, P.B., Ellsworth, D.S., Walters, M.B. Vose, J.M., Gresham, C., Volin, J.C.,
Bowman, W.D. 1999. Generality of leaf trait relationships: a test across six biomes. Ecology, 80:1955-1969.
Reich, P.B., Uhl, C., Walters, M.B. & Ellsworth, D.S. 1991. Leaf lifespan as a determinant
of leaf structure and function among 23 amazonian tree species. Oecologia, 86: 16-24.
Reich, P.B., Walters, M.B. & Ellsworth, D.S. 1991. Leaf age and season influence the
relationships between leaf nitrogen, leaf mass per area and photosynthesis in maple and oak trees. Plant, Cell and Environment, 14: 251-259.
Reinert, F. 1995. On the Bromeliaceae of the restinga of Barra de Maricá in Brazil:
environmental influences on the expression of crassulacean acid metabolism. Phd Thesis, Agricultural and Environmental Science, New CastleUniversity, UK.
Rizzini, C.T. 1979. Tratado de Fitogeografia do Brasil. Aspectos sociológicos e florísticos.
São Paulo, Hucitec. Rôças, G. Barros, C.F. & Scarano, F.R. Leaf anatomy plasticity of Alchornea triplinervea
(Euphorbiaceae) under distinct light regimes in a Brazilian montane Atlantic rain forest. Trees Structure and Function, 11: 469-473.
Rôças, G., Scarano, F.R. & Barros, C.F. 2001. Leaf anatomical variation in Alchornea
triplinervea (Spreng.) Mull. Arg. (Euphorbiaceae) under distinct light and soil water regimes. Botanical Journal of the Linnean Society, 136: 231-238.
Rosevear, M.J., Young, A.J. & Johnson, G.N. 2001. Growth conditions are more important
than species origin in determining leaf pigment content of British plant species. Fuctional Ecology, 15: 474-480.
Ruzin, S. 1999. Plant microthechnique and microscopy. Oxford University Press, New
York. Sampaio, M.C., Perissé, L.E., Oliveira, G.A. & Rios, R.I. 2002. The contrasting clonal
architecture of two bromeliads from sandy coastal plains in Brazil. Flora, 197: 443-451.
Sampaio, M.C., Picó, F.X. & Scarano, F.R. 2005. Ramet demography of a nurse bromeliad
in Brazilian restingas. American Journal of Botany, 92: 676-683.

102
Sampaio, M.C. 2004. Respostas da bromélia clonal Aechmea nudicaulis à heterogeneidade ambiental da restinga aberta de Clusia. Tese de Doutorado, PPGE/UFRJ, Rio de Janeiro.
Sass, J.E. 1951. Elements of botanical microtechnique. McGraw-Hill, New York. Scarano, F.R., Ribeiro, K.T., Moraes, L.F.D. & Lima, H.C. 1997. Plant establishment on
flooded and unflooded patches of a freshwater swamp forest in southeastern Brazil. Journal of Tropical Ecology, 14:793-803.
Scarano, F.R., Duarte, H.M., Ribeiro, K.T., Rodrigues, P.J.F.P., Barcellos, E.M.B., Franco,
A.C., Brulfert, J., Deléens, E. & Lüttge, U. 2001. Four sites with contrasting environmental stress in southeastern Brazil: relations of species, life form diversity, and geographical distribution to ecophysiological parameters. Botanical Journal of the Linnean Society, 136: 345-364.
Scarano, F.R., Duarte, H.M., Rôças, G., Barreto, .M.B. Amado., E.F., Reinert, F., Wendt,
T. Mantovani, A., Lima, H.R.P. & Barros, C.F. 2002. Acclimation or stress sympton? An integrated study of intraspecific variation in the clonal plant Aechmea bromelifolia, a widespread CAM tank-bromeliad. Botanical Journal of the Linnean Society, 140: 391-401.
Scarano, F.R., Duarte, H.M., Franco, A.C., Gessler, A., Mattos, E.A., Rennenberg, H. &
Lüttge,, U. 2005a. Physiological synecology of tree species in relation to geographic distribution and ecophysiological parameters at the Atlantic forest periphery in Brazil: an overview. Trees, Structure and Function, 19: 497-509.
Scarano, F.R., Duarte, H.M., Franco, A.C., Gessler, A., Mattos, E.A., Nahm, M.,
Rennenberg, H. Zaluar, H.L.T. & Lüttge,, U. 2005b. Ecophysiology of selected tree species in different plant communities at the periphery of the Atlantic Forest o SE – Brazil. I. Behaviour of three different species of Clusia in an array of plant communities. Trees, Structure and Function, 19: 497-509.
Scarano, F.R. 2002. Structure, function and floristic relationships of plant communities in
stressful habitats marginal to the Brazilian Atlantic Rainforest. Annals of Botany, 90; 517-524.
Schlichting, C.D. 1986. The evolution of phenotypic plasticity in plants”. Annual Review
of Ecology and Systematics, 17:667-693. Siqueira Filho, J.A. & Machado, I.C.S. 2001. Biologia reprodutiva de Canistrum
aurantiacum E. Morren (Bromeliaceae). Acta Botanica Brasilica, 14: 427-443. Siqueira Filho, J.A. & Tabarelli, M. 2006. Bromeliad species of the Atlantic forest of noth-
east Brazil: losses of critical populations of endemic species. Oryx, 40(2): 218- Soave, R.C.F. & Silva, O.A. 1993. Aspectos fenológicos e variação nos conteúdos das
clorofilas a e b em Caryocar brasiliensis Camb. Arq. Biol. Tecnol., 36(1): 57-69. Souza, A.F. 2004. Aspectos da dinâmica populacional de uma palmeira clonal na floresta

103
paludícola da Reserva Municipal de Santa Genebra (Campinas, SP). Tese de Doutorado, Instituto de Biologia, UNICAMP.
Stuefer, J.F. 1997. Division of Labour in clonal plants? On the response of stoloniferous
herbs to environmental heterogeneity. PhD Thesis, Department of Plant Ecology & Evolutionary Biology, Utrecht University, The Netherlands.
Stuefer, J.F., During, H.J. & de Kroon, H. 1994. High benefits of clonal integration in two
stoloniferous species, in response to heterogeneous light environments. Journal of Ecology, 82: 511-518.
Stuefer, J.F., de Kroon, H. & During, H.J. 1996. Exploitation of environmental
heterogeneity by spatial division of labour in a clonal plant. Functional Ecology, 10: 328-334.
Stuefer, J.F. 1996. Potential and limitations of current concepts regarding the response of
clonal plants to environmental heterogeneity. Vegetatio, 127: 55-70. Suguio, K. & Tessler, M.G. 1984. Planícies de cordões litorâneos quaternários do Brasil:
origem e nomeclatura. In Lacerda, L.D., Araujo, D.S.D., Cerqueira, R. & Turcq, B. (Orgs.) Restingas: origem, estrutura e processos, Niterói, CEUFF, 15-26..
Taiz, L. & Zeiger, E. 1998. Plant Physiology. Sinauer Associates, Sunderland. Valiente-Banuet, A. & Ezcurra, E. 1991. Shade as a cause of the association between the
cactus Neobuxbaumia tetetzo and the nurse plant Mimosa luisana. Journal of Ecology, 76:961-971.
Valiente-Banuet, A., Vite, F. & Zavala-Hurtado, A.J. 1991a. Interactions between the
cactus Neobuxbaumia tetetzo and the nurse plant Mimosa luisana. Journal Vegetation Science, 2:11-14.
Valiente-Banuet, A., Bolongaro-Crevenna, A., Briones, O., Ezcurra, E., Rosas, M., Nuñez,
H., Bernard, G. & E. Vazquez. 1991b. Spatial relationships between cacti and nurse shrubs in a semiarid environment in central Mexico. Journal of Vegetation Science, 2:15-20.
Valladares, F. & Pearcy, R.W. 1997. Interactions between water stress, sun-shade
acclimation, heat tolerance and photoinhibition in the sclerophyll Heteromeles arbutifolia. Plant, Cell and Environment, 20:25-36.
Vernon, L.P. 1960. Spectrophotometric determination of chlorophylls and pheophytins in
plant extracts. Analytical Chemistry, 32(9): 1144-1150. Vetaas, O.R. 1992. Micro-site effects of threes and shrubs in dry savannas. Journal of
Vegetation Science, 3: 337-344. Vieira, R.C. 1995. Anatomia foliar de Bauhinia radiata Vell. Em diferentes ambientes.
Brazilian Archives of Biology and Technology, 38: 63-107.

104
Villegas, A.C. 2001. Spatial and temporal variability in cloanl reproduction of Aechmea magdalenae, a tropical understory herb. Biotropica, 33(1): 48-59.
Wylie, R.B. 1951. Principles of foliar organization shown by sun-shade leaves from ten
species of deciduous dicotyledonous trees. American Journal of Botany, 38: 355-361.
Winter, K. & Smith, J.A.C. 1996. Crassulacean acid metabolism: current status and
perspectives. In: K.Winter, J.A.C. Smith (eds.) Crassulacean acid metabolism: biochemistry, ecophysiology and evolution. Ecological Studies Analysis and Synthesis, 114: 389-426.
Witkowski, E.T.F. & Lamont, B.B. 1991. Leaf specific mass confounds leaf density and
thickness. Oecologia, 88: 486-493. Yarranton, G.A. & Morrison, R.G. 1974. Spatial dynamics of primary succession:
nucleation. Journal of Ecology, 62: 417-428. Yeaton, R.I. 1978. A cyclical relationship between Larrea tridentata and Opuntia
leptocaulis in the Northern Chihuahuan Desert. Journal of Ecology, 66:651-656. Zaluar, H.L.T. 1997. Espécies focais e formação de moitas na restinga aberta de Clusia,
Carapebus, RJ. Dissertação de M.Sc., Programa de Pós-graduação em Ecologia – UFRJ.
Zaluar, H.L.T. & Scarano, F.R. 2000. Facilitação em restingas de moitas: um século de
buscas por espécies focais. In: Esteves FA & Lacerda LD (eds.). Ecologia de Restingas e Lagoas Costeiras. Rio de Janeiro: NUPEM-UFRJ, pp. 3-23.
Zaluar, H.L.T. 2002. Dinâmica da vegetação em restingas abertas fluminense: uma
aproximação através das interações entre plantas. Tese de D.Sc., PPGE/UFRJ, Rio de Janeiro, RJ, Brasil.
Zar, J.H. 1996. Biostatistical analysis. 3 ed. Prentice Hall. New Jersey, USA.





























