Embed Size (px)
Citation preview

Retina
Crystallin-b-b2-Overexpressing NPCs Support the Survivalof Injured Retinal Ganglion Cells and Photoreceptors inRats
Michael R. R. Bohm,1,2 Sarah Pfrommer,1 Carolin Chiwitt,1 Matthias Bruckner,1
Harutyun Melkonyan,1 and Solon Thanos1,2
PURPOSE. Crystallin b-b2 (crybb2) is known to support theregeneration of retinal ganglion cell (RGC) axons in culture.We investigated whether neuronal progenitor cells (NPCs)overexpressing crybb2 (crybb2-NPC) affect secondary retinaldegeneration due to optic nerve crush in vivo.
METHODS. NPCs were produced by dissociation and propaga-tion of rat embryonic neural tube and eye primordial cells atembryonic days 13.5 and 15. Retinal degeneration was inducedby injured optic nerve crush (by suture, 20 seconds). Severalgroups were built: crybb2-NPC were injected into the vitreousbody, while the Controls were comprised of recombinantcrybb2–injected and PBS-injected groups. The eyes, inparticular the retina, were analyzed by immunohistochemistryand Western blotting for different antigens at 2 and 4 weeksafter surgery.
RESULTS. At 2 and 4 weeks post surgery, crybb2-NPC residedwithin the vitreoretinal compartment, and were persistentlynestin-positive throughout the experimental period. The cellsstained positive for various neurotrophins and acted as ‘‘living’’cell factories to support the survival of injured RGCs. Thecrybb2-NPC migrated throughout the eye structures andsometimes became integrated within the tissue. Most of theocular cells responded to the appearance of crybb2-NPC withmarked changes of certain proteins, including Iba-1 (microg-lia), vimentin (glial cells), and rhodopsin (photoreceptors).Photoreceptors also displayed a better survival after crybb2-NPC injection compared to control groups.
CONCLUSIONS. Crybb2-NPC exert beneficial effects on thevitreoretinal compartment, which suggests that modifiedcrybb2-NPC could be used in a novel strategy for the treatmentof degenerative vitreoretinal diseases. However, future studies
must determine the safety of in vivo administration of crybb2-NPC. (Invest Ophthalmol Vis Sci. 2012;53:8265–8279) DOI:10.1167/iovs.12-10334
Retinal degenerative diseases affect millions of peopleworldwide1 and, hence, methods must be developed to
prevent such degeneration and the consequent visual impair-ment. One of these strategies could involve placing cells insidethe eye that produce protective substances.
Neural stem cells and neural progenitor cells (NPCs) havebeen isolated from different parts of the embryonic and adultcentral nervous system (CNS) of several mammalian species.2–5
NPCs secrete several neurotrophic factors (NTFs) and exertprotective effects on diseased neurons.6–8 NPCs can bemanipulated through pretreatment with growth factors or viagene transfection.1,2,5,9 Attempts to replace lost photorecep-tors so far has focused on injecting stem cells into the retina orthe subretinal space. Although some of the injected cellsdifferentiate into photoreceptors and integrate into the hosttissue, the quantity of cells involved generally is low, and inmost cases they fail to function.2,10–12 Alternative approachesinclude the modification of NPCs to rescue visual functionthrough the engineered expression of NTFs, such as glial cell–derived NTF (GDNF)-producing human-derived NPCs. Thesewere shown to have a positive effect after being injected intothe degenerating retinas of Royal College of Surgeons (RCS)rats.1
In addition to classical NTFs and differentiation molecules,ocular crystallins appear to have a crucial role in thedevelopment of retinal cells, including photoreceptors.13–15
Crystallins, which also are expressed in retinal neurons,16 havebeen identified in various neurodegenerative disorders, includ-ing in those affecting the retina.17 While several studies havedemonstrated that crystallin a has a role in retinal disorders,there is little information on the effects of crystallin b/c.18–21
Adult retinal ganglion cells (RGCs) have an enhanced chance ofsurvival when exposed to lens injury.16,22,23 Testing of differentclasses of crystallins has revealed that crystallins of the b/csuperfamily possess a strong neuroprotective and regenerativepotential by mimicking the effects of lens injury.16
Crystallins b and c (cryg) are expressed during thedevelopment of the mouse eye.14 Increased levels of cryghave been observed in cultured astrocytes affected byhypoxia,23 neuronal aging processes,24 and neuronal repair.25
Added crystallin b-b2 (crybb2) enhances the growth of axonsin retinal explants and in primary hippocampal neurons.26
However, to our knowledge no protective effects of crybb2 onRGCs in vivo or on photoreceptors have yet been reported.
To investigate neuroprotective features of NPCs, we usedthe optic nerve crush model to induce retinal degeneration andinjected transfected NPCs, overexpressing crybb2 intravitreally
From the 1Institute for Experimental Ophthalmology, School ofMedicine, Westfalian-Wilhelms-University Munster, Munster, Ger-many; and the 2Interdisciplinary Centre for Clinical Research (IZKF),School of Medicine, Westfalian-Wilhelms-University Munster,Munster, Germany.
Supported by a DFG Grant Th 386/18 (ST) and by IZKF grants(MRRB) and Grant THa3/002/09 (ST). The authors alone areresponsible for the content and writing of this paper.
Submitted for publication June 5, 2012; revised October 30,2012; accepted November 1, 2012.
Disclosure: M.R.R. Bohm, None; S. Pfrommer, None; C.Chiwitt, None; M. Bruckner, None; H. Melkonyan, None; S.Thanos, None
Corresponding author: Michael R. R. Bohm, Institute ofExperimental Ophthalmology, School of Medicine, Westfalian-Wilhelms-University Munster, Albert-Schweitzer-Campus 1, D15,48149 Munster, Germany; [email protected].
Investigative Ophthalmology & Visual Science, December 2012, Vol. 53, No. 13
Copyright 2012 The Association for Research in Vision and Ophthalmology, Inc. 8265
after surgery. In controls, NPCs were replaced with intravitrealinjection of recombinant crybb2 protein or injection of PBS.The aims of our study were to elucidate the survival andmigration of NPCs within the eye, the intravitreal productionof NTFs and crystallins, and the effects of overexpressedcrybb2 on degenerating RGCs and photoreceptors. Therefore,immunohistochemistry and Western blotting (WB) were usedto examine intravitreal distribution of NPCs and production ofNTFs that may affect the fate of photoreceptors, glial cells, andmicroglia.
METHODS
Animals and Drugs
Pregnant Sprague-Dawley (SD) rats at embryonic days (E)13.5 and 15
(n¼ 52) were used as a source of NPCs for these studies. Adult SD rats
at approximately 3 months of age (n¼ 64) of both sexes were used for
the surgery. All experiments were conducted in accordance with the
ARVO Statement for the Use of Animals in Ophthalmic and Vision
Research. Animals were housed in standard animal rooms under a 12-
hour light/dark cycle, with food and water provided ad libitum. All
surgical experiments were performed under general anesthesia
induced by a mixture of 10% ketamine (2 mg/kg per body weight;
Ceva Sanofi, Dusseldorf, Germany) and xylazine (1 mg/kg per body
weight; Ceva-Sanofi), administered intraperitoneally. After surgical
intervention, gentamicin (Dr. Mann Pharma, Berlin, Germany) was
applied topically to prevent infections. All surgical manipulations were
unilateral, with the contralateral eye serving as a control. Experimental
and control groups consisting of 16 animals are shown in Table 1.
Preparation and Transfection of NPCs
To retrieve and culture NPCs, embryos were removed under deep
anesthesia by cesarean section at days E13.5 and E15, and were stored
in sterile Hank’s balanced salt solution (HBSS; PAA Laboratories,
Pasching, Germany) with 1% PenStrep (Sigma-Aldrich, Hamburg,
Germany), on ice until dissection. The brains were removed and
homogenized mechanically. The homogenate then was treated with
0.1% trypsin in HBSS to obtain single cells. The suspension was
transferred into a culture medium comprising Dulbecco’s modified
Eagle’s medium–Ham’s (DMEM-F12; PAA Laboratories) supplemented
with 1% PenStrep, 1% N2 Supplement (PAA Laboratories), 10 ng/lL
recombinant human basic fibroblast growth factor (bFGF; RD Systems,
Minneapolis, MN), and 10 lL insulin (2.5 mg/mL; PromoCell,
Heidelberg, Germany). The suspension was triturated gently and 4.5
3 106 cells were plated at a density of 200,000 cells/mL on noncoated
Advanced TC Petri dishes (Greiner Bio-One, Frickenhausen, Germany)
to inhibit further differentiation. The cells were incubated at 378C in a
humidified atmosphere of 5% CO2. All cell harvesting steps were
performed under sterile conditions.
Rat crybb2 was cloned into pIRES-ACGFP (Clontech, Palo Alto, CA)
so that rat crybb2 and green fluorescent protein (GFP) could be
translated from a single bicistronic mRNA. For transfection of NPCs,
cultured cells that formed neurospheres were dissociated mechanically
into single cells and centrifuged for 5 minutes at 400 rpm (48C). The
culture medium was removed and approximately 5 3 106 cells were
resuspended in 100 lL Amaxa Rat NSC Nucleofector Kit (Lonza
Cologne, Cologne, Germany). Vector (3.5 lg DNA) was added, the cells
and DNA were mixed gently, and then the cells were electroporated
using the Nucleofector device (Lonza Cologne). The transfection
protocol used was very similar to that described by the manufacturer,
thus resulting in the highest transfection efficacies.5,27 Cells were used
for a maximum of 2 weeks after preparation. After finishing this
procedure, crybb2-overexpressing NPCs (crybb2-NPC) were placed
into the culture medium and cultured for up to 7 days, and the efficacy
of transfection was monitored using GFP fluorescence.
Transfected crybb2-NPC were prepared for injection by first
washing the cell suspensions with HBSS. Cell numbers then were
counted using the trypan-blue exclusion method, and then the cells
were transferred to the transplantation medium, HBSS. Surgery and cell
injection were performed within 1 hour of cell preparation.
Unilateral Optic Nerve Crush (ONC) andIntravitreal Treatment
To generate the primary degeneration of RGCs and a photoreceptor
transsynaptic response, we performed unilateral ONC to induce
damage of axons followed by the intravitreal injection of crybb2-
NPC. After a skin incision near the superior orbital rim, ONC was
performed according to the ‘‘mini lesion’’ technique, which involves
ligation with a suture (8.0 Ethicon; Johnson & Johnson, St-Stevens-
Woluwe, Belgium) for exactly 20 seconds.26 A 10 ll volume of crybb2-
NPC was injected through the sclera via a glass capillary, with care
being taken to avoid damage to the lens. To reveal the effects
administered by intravitreal injected crybb2-NPC, several groups in the
experimental setup were built. In the controls, 10 lL of recombinant
crybb2 protein (180 lg/lL) were injected intravitreally to reveal the
effects of recombinant crybb2 protein compared to crybb2-NPC due to
retinal degeneration. After subcloning into pQE32 (Qiagen, Hidden,
Germany), rat crybb2 was expressed and purified: bacterial cultures
were centrifuged (1500g for 20 minutes) and subsequently resus-
pended in buffer 1 (8 M urea, 50 mM NaH2PO4, 15 mM imidazole, 10
mM Tris-HCl, and 100 mM NaCl; pH 8.0). After cloning, cell lysates
were centrifuged at 20,000g for 30 minutes. The supernatant was
purified using immobilized metal affinity chromatography (Clontech);
elution was performed using buffer 1 at pH 6.0 supplemented with 100
mM imidazole. The proteins were dialyzed in aqua dest. For control
TABLE 1. Experimental Groups (In Vivo)
Experimental Group N Weights of SD Rats Experimental Manipulation Times, wk
Effects of crybb2 16 180 to 250 g Left ONC with intravitreal injection of crybb2 2, 4
IHC 4
WB 4
Effects of crybb2-NPC 16 180 to 250 g Left ONC with intravitreal injection of crybb2-NPC 2, 4
IHC 4
WB 4
Effects of PBS (control) 16 180 to 250 g Left ONC with intravitreal injection of PBS 2, 4
IHC 4
WB 4
Adult control 16 180 to 250 g Untreated 2, 4
IHC 4
WB 4
IHC, immunohistochemistry; WB, Western blotting; wk, weeks.
8266 Bohm et al. IOVS, December 2012, Vol. 53, No. 13

operations (see above), 10 lL sterile PBS were injected after ONC
instead of crybb2-NPC to exclude beneficial effects of the method of
intravitreal injection due to retinal degeneration. The experimental
protocols are listed in Table 1.
Immunohistochemistry
Cultured NPCs were examined by immunohistochemistry for selected
proteins: oncomodulin, brain-derived NTF (BDNF), calcineurin, and
cryg and crybb2. Cultures were fixed in fresh 4% paraformaldehyde for
10 minutes at room temperature (RT), washed in PBS, and then fixed
with methanol (10 minutes at�208C). After rinsing the cells with PBS,
they were incubated with blocking solution containing 10% goat serum
(2 hours at RT); the primary antibodies then were applied (Table 1).
After washing, cells were incubated with secondary antibodies (1 hour
at RT, Table 1) and then washed with PBS. Antifade mounting medium
was used to cover the cells (Mowiol; Hoechst, Frankfurt, Germany).
The expression, intraocular localization, and effects of intravitreally
injected crybb2-NPC were investigated by sacrificing operated rats
using CO2 inhalation (n ¼ 4 from each group). The eyeballs were
explanted, frozen in Tissue-Tek (Sakura Finetek Europe, Alphen aan
den Rijn, The Netherlands), and then stored at�808C until further use.
Sections were cut along the visual axis with a cryostat (10 lm thick,
CM 1500; Leica, Bremen, Germany) and mounted onto glass slides. The
same primary antibodies used to detect NTFs were used in vivo (see
above). In addition, retinal cells were stained for specific antigens to
identify potential changes caused by the NPCs. Photoreceptors were
detected with rhodopsin, while glial cells were visualized with glial
fibrillary acidic protein (GFAP) and antivimentin antibodies. The
quantitative analysis of the retinal ganglion cell axons was performed
as follows: retinal slices (10 lm) were labeled with antibodies against
NF-200 revealing the axonal cytoskeleton of retinal ganglion cells.28,29
Five microscope fields (each 0.2 mm2) of each sample were adjusted
and NF-labeled axons were counted. The mean count of axons was
calculated and expressed as a percentage of retinal ganglion cells
compared to the number of NF-stained axons in the untreated
contralateral retina. Because each ganglion cell is emitting one axon,
the axon numbers are related to the ganglion cell numbers.
ED-1 was used to identify microglial cells in different layers of the
retina, while the ionized calcium-binding adaptor molecule 1 was used
to identify activated microglia.
The procedure used to stain cell nuclei with 40,6-diamidino-2-
phenylindole is described above. The slides were viewed with the
appropriate filter on a microscope equipped with epifluorescence
(Axiophot; Carl Zeiss Meditec, Oberkochen, Germany). Negative
controls comprised sections processed without the addition of the
primary antibodies. Control and experimental sections were stained
simultaneously to avoid variations in immunohistochemical staining.
Table 2 lists the antibodies used, including their working dilutions. All
stained cells were counted within a region of 0.2 mm2 (five
randomized regions per eye) at a magnification of 3400.
Western Blotting
WB was performed to confirm the immunohistochemical findings and
evaluate retinal degeneration. Eyes taken from four animals of each
group were enucleated. The extraocular tissues were dissected, the
retina was discharged carefully from the sclera, and the vitreous was
peeled off. All isolated samples were frozen in liquid nitrogen and kept
at �808C until further use.
Samples were homogenized in SDS sample buffer (62.5 mM Tris-
HCl, 2% wt/vol SDS, 10% glycerol, 50 mM dithiothreitol, and 0.01% wt/
vol bromophenol blue). The samples were sonicated and heated, and
then the protein concentration was determined using the Bradford
method.
A total of 50 lg protein from each sample was fractionated on 8%,
10%, or 12% SDS-polyacrylamide gel electrophoresis (depending on the
protein) with a protein marker (BioRad, Hercules, CA). After
electrophoresis, the proteins were transferred onto a nitrocellulose
membrane (Schleicher & Schull BioScience, Dassel, Germany). The
blots were incubated in blocking solution (5% freshly prepared fat-free
dried milk and 0.1% Tween-20 PBS) for 1 hour, followed by incubation
overnight at 48C with a variety of antibodies (Table 3). The membrane
then was incubated with the secondary antibody conjugated with
horseradish peroxidase in blocking solution for 1 hour at RT.
Antibodies were detected with enhanced chemoluminescence (Amer-
sham, Rockville, UK) and documented on X-ray films (Amersham
Pharmacia Biotech, Amersham, UK). Loading of comparable amounts
of protein was confirmed with anticalnexin (see below). Each gel was
run three times. The relative densities of the protein spots were
analyzed using Alpha Ease (Alpha Ease FC software 4.0; Alpha
Innotech, San Leandro, CA). The protein density of a fixed area was
TABLE 2. Antibodies Used for IHC
Antibody Company Isotype Dilution Tissue
Primary antibodies
BDNF Millipore (Temecula, CA) Rabbit polyclonal 1:500 NPCs in vitro/in vivo
Calcineurin Millipore Rabbit polyclonal 1:500 NPCs in vitro/in vivo
Crybb2 Santa Cruz Biotechnology (Santa Cruz, CA) Goat polyclonal 1:100 NPCs in vitro/in vivo
Cryg Santa Cruz Biotechnology Rabbit polyclonal 1:100 NPCs in vitro/in vivo
ED-1 BMA Biomedicals (Augst, Switzerland) Mouse monoclonal 1:200 Microglia
Iba-1 Wako Pure Chemical Industries (Osaka, Japan) Rabbit 1:500 Microglia
Nestin Millipore Rabbit polyclonal 1:200 NPCs in vitro/in vivo
Nestin AbD Serotoec (Puchheim, Germany) Mouse monoclonal 1:100 NPCs in vitro/in vivo
Neurofilament 200 Sigma-Aldrich (St. Louis, MO) Mouse monoclonal 1:400 Retinal ganglion cells
Oncomodulin Swant (Marly, Switzerland) Rabbit polyclonal 1:500 NPCs in vitro/in vivo
Rhodopsin Millipore Mouse monoclonal 1:100 Photoreceptors
Vimentin Sigma-Aldrich Mouse monoclonal 1:200 Retinal glial cells
Secondary antibodies
Cy-2 Jackson ImmunoResearch Europe Ltd. (Suffolk, UK) Goat antimouse 1:200
Cy-2 Jackson ImmunoResearch Europe Ltd. Goat antirabbit 1:200
Cy-2 Abcam (Cambridge, UK) Donkey Antigoat 1:100
TRITC Sigma-Aldrich Goat antirabbit 1:400
TRITC Sigma-Aldrich Goat antimouse 1:300
TRITC Jackson ImmunoResearch Europe Ltd. Donkey antigoat 1:100
TRITC, tetramethylrhodamine isothiocyanate; Cy, cyanine.
IOVS, December 2012, Vol. 53, No. 13 Crystallin-b-b2-Overexpressing NPCs 8267

determined for each spot by subtracting the specific background
density of the particular application control. The lowest spot density
for each of the different proteins was defined as the reference mark,
and the relative values were determined and processed. The mean and
standard deviation of the relative relationships of the proteins were
generated from gels of four individual samples, each one run three
times for the individual group.
Data Evaluation
The standard deviation and optical density of the WB analysis and
several cell counts were analyzed with a test for two independent
samples (SPPS; SSPS, Chicago, IL; Statistica ver. 7; StatSoft, Tulsa, OK) to
examine the Gaussian distribution, and processed using the indepen-
dent-samples t-test (Gaussian) and Kruskal-Wallis H test. Non-Gaussian
values were prepared with image-processing software (Photoshop;
Adobe Systems, San Jose, CA). The overall brightness and contrast were
adjusted without retouching.
RESULTS
Morphologic Characterization and FactorExpression of crybb2-NPC In Vitro
As expected, NPCs harvested from embryonic brain and eyeprimordial tissue exhibited various morphologies in culture.Single cells were found along with clusters and neurospheresof NPCs (Figs. 1A, 1B). These stem cells possessed longprotrusions with a shaped cell body (Fig. 1B). NPCs stainedpositive for several NTFs, including BDNF (Figs. 1C–E),oncomodulin (Figs. 1F–H), calcineurin (Figs. 1I–K), crybb2(Figs. 1L–N), and cryg (Figs. 1O–Q). Since crybb2 is known tobe associated with regenerative retinal functions in vitro,26 itwas selected as the first-choice protein for analysis in vivo. Weobserved a transfection rate of 55 to 67%, which is comparableto previous reports.5
NPCs within the Eye
Immunohistochemistry was used to examine whether theinjected crybb2-NPC continued expression of the same NTFswithin the eye. It appeared that all proteins still were producedwithin the vitreous body (Figs. 2A–O). Particular attention waspaid to crybb2 that was stained within clusters of cells locatedwithin the vitreous (Figs. 2J–L). These data supported the viewthat non-transfected NPCs produce baseline levels of crybb2.Compared to crybb2-positive RGCs in the PBS group (Fig. 3A),crybb2-NPC–treated eyes exhibited increased and diffusestaining of RGCs after 2 weeks (Fig. 3C). No crybb2 staining
was observed 4 weeks after ONC (Figs. 3B, 3D). When theimmunohistochemistry was performed with sections fromcrybb2-NPC–treated eyes, most cells stained positive forcrybb2 (Fig. 3E).
Intraocular Localization of Intravitreally Injected
crybb2-NPC
Nestin was used as a stem-cell marker to identify thedistribution of crybb2-NPC at 2 and 4 weeks after intravitrealtransplantation. NPCs were observed as follows: arrestedcrybb2-NPC in the vitreous cavity, arrested and migratingcrybb2-NPC in the vitreoretinal compartment, and arrested andmigrating crybb2-NPC in the anterior segment, including theanterior chamber, cornea, and ciliary body (Table 4).
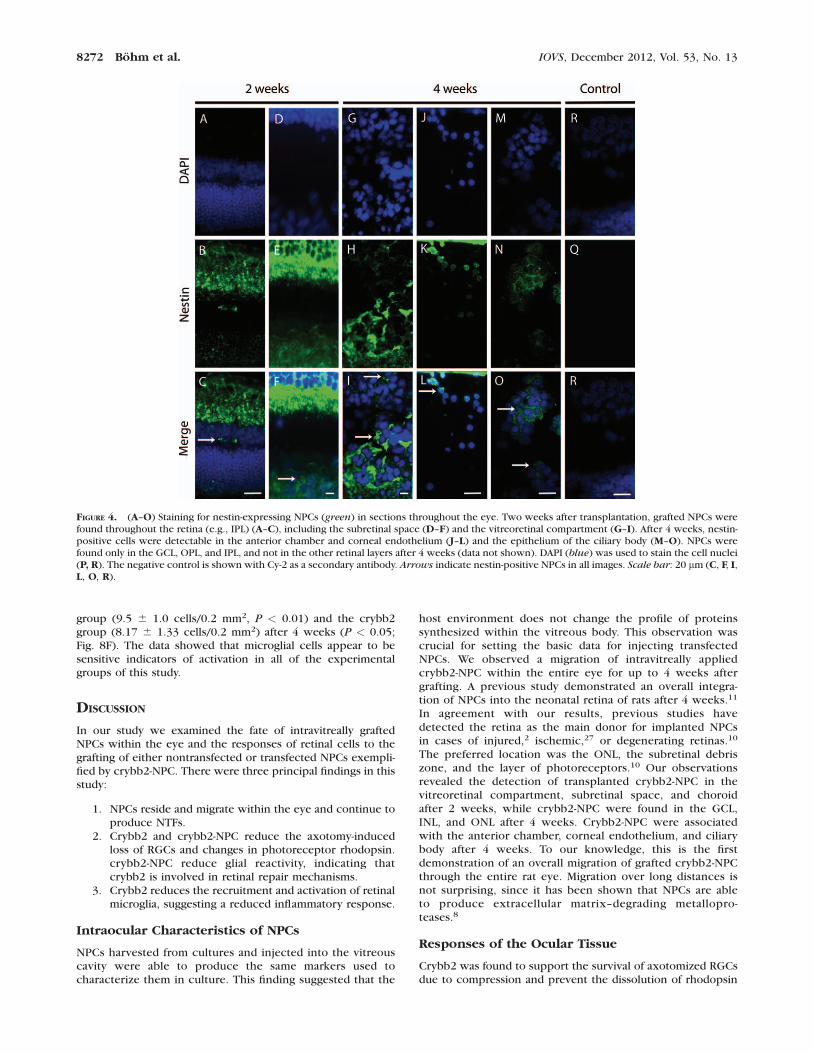
Arrested nestin-positive cells were localized in the vitrealcavity, at the vitreoretinal interface (Figs. 4G–I), in most retinallayers (Figs. 4A–C), and in the subretinal space, including thechoroidea (Figs. 4D–F) 2 weeks after transplantation. They alsowere found in the anterior chamber, associated with thecorneal endothelium (Figs. 4J–L) and epithelium of the ciliarybody (Figs. 4M–O) 4 weeks after transplantation. In the hostretina, NPCs were detected in the ganglion cell layer (GCL),inner nuclear layer (INL), and outer nuclear layer (ONL). Theseobservations show that NPCs are redistributed within the eyeduring the first 2 weeks after injection. The distribution ofNPCs was similar at 4 weeks post surgery (Fig. 4), with someNPCs having reached quite distant locations, such as thecornea (Figs. 4J–L). These data supported the view that NPCscan migrate within the eye.
Staining of Ocular Tissue after Injection of crybb2-
NPC
The marker neurofilament 200 was used to examine theresponse of RGCs to damage and to the intravitreal injection ofNPCs (Figs. 5A–D). A moderate loss of RGCs was found in thecrybb2-NPC group compared to the PBS group. Compared tothe adult control, the population of RGCs had reduced to 82.636 5.60% in the crybb2-NPC group and to 55.56 6 8.56% in thePBS group 2 weeks after implantation. After 4 weeks, thesevalues were 52.78 6 5.73% and 18.75 6 3.99%, respectively.In the crybb2 group, the population of RGCs had reduced to58.30 6 2.41% after 2 weeks and 55.56 6 3.99% after 4 weeks(Fig. 5E). After 4 weeks the population of RGCs wassignificantly higher in the crybb2 group than in the PBS group.These percentages were confirmed by WB analysis usingantibodies detecting brain-specific homeobox/POU domain
TABLE 3. Antibodies Used for WB
Antibody Company Isotype Dilution Molecular Mass
Primary antibodies
BRN-3b Covance (Princeton, NJ) Rabbit polyclonal 1:2,000 45 kD
Rhodopsin Millipore Mouse monoclonal 1:1,000 39 kD
Vimentin Sigma-Aldrich Mouse monoclonal 1:500 58 kD
Secondary antibodies
Secondary rhodopsin Cell Signaling Technology, Inc. (Danvers, MA) Horse antimouse 1:4,000
Secondary vimentin Cell Signaling Technology, Inc. Horse antimouse 1:4,000
Secondary BRN-3b Amersham Pharmacia Biotech Donkey antirabbit 1:10,000
Loading control
Calnexin Sigma-Aldrich Rabbit 1:10,000 90 kD
BRN-3b, brain-specific homeobox/POU domain protein 3b.
8268 Bohm et al. IOVS, December 2012, Vol. 53, No. 13

FIGURE 1. Morphologic and immunohistochemical characterization of NPCs derived from cultured rat fetal neural tissue. (A, B) Cultured NPCswere found in cell clusters, called neurospheres (arrows). (B) Single cells with morphologic features reminiscent of stem cells also were found(arrowheads). (C–Q) Immunohistochemical characterization of cultured NPCs (using nestin, green) and several other markers (red) in vitro.Labeling with antibodies raised against BDNF (C–E), oncomodulin (F–H), calcineurin (I–K), crybb2 (L–N), and cryg (O–Q) revealed that thesemolecules were expressed in NPCs in vitro. 40,6-Diamidino-2-phenylindole (DAPI, blue) was used to stain cell nuclei. Scale bars: 20 lm (A, B, E, H,K, N, Q).
IOVS, December 2012, Vol. 53, No. 13 Crystallin-b-b2-Overexpressing NPCs 8269

protein 3b (Fig. 5F).30 These data showed that crybb2-producing NPCs support the survival of injured RGCs.
Photoreceptor Response
The marker rhodopsin was used to examine the response ofphotoreceptors to axotomy due to optic nerve crush and to theintravitreal injection of NPCs. Immunohistochemical evalua-tion of rhodopsin revealed intense labeling of the photorecep-tor outer segments (POS) in adult retinas. A slight decrease inrhodopsin was found in all treated groups after 2 weeks (Figs.6A–D). Comparison of crybb2-NPC and crybb2 groups withthe PBS group revealed less decay of stained POS after 4 weeks(Figs. 6E–H).
Additional WB analysis revealed a massive decay ofrhodopsin in the PBS group (89.40 6 4.16% at 2 weeks and21.06 6 3.53% at 4 weeks). Comparable rhodopsin wasdetected in the crybb2-NPC group after 2 weeks (94.95 6
12.27%), while a smaller decrease in rhodopsin was observedafter 4 weeks (66.36 6 14.88%, P < 0.01; Figs. 6I, 6J).Significantly less rhodopsin was found in the crybb2-treatedanimals after 2 weeks (79.45 6 5.27%) than in the PBS group(Figs. 6I, 6J; P < 0.05). Significantly less rhodopsin was lost inthe crybb2 group (53.61 6 2.48%) compared to the PBS groupafter 4 weeks (Figs. 6I, 6J; P < 0.001). Comparable staining of
rhodopsin was found in the crybb2-NPC group after 2 weeks(94.95 6 12.27%), while a reduced loss of rhodopsin was seenafter 4 weeks (66.36 6 14.88%, P < 0.01; Figs. 6I, 6J). Thesedata showed that photoreceptors respond to the injury-induced loss of RGCs with a reduction in rhodopsin, and thatthis reduction can be neutralized at least partly by the additionof crybb2-producing NPCs.
Response of Glial Cells
Antibodies detecting GFAP and vimentin were used to detectretinal glial reactivity. A distinct labeling of both proteins in thenerve fiber layer (NFL), ONL, and OPL in the crybb2-NPCgroup was observed after 2 weeks (Figs. 7E, 7F). The intensityof the vimentin staining in the inner retinal layers wascomparable to that in the adult retinal control (data notshown). Increased labeling of vimentin (Fig. 7C) and GFAP(Fig. 7D) in all retinal layers of the crybb2 group was seen after2 weeks. Compared to the PBS group, the crybb2-NPC animalsexhibited reduced staining of vimentin and GFAP in the outerretinal layers, including the NFL, RGC layer, inner plexiformlayer (IPL), INL, and the outer plexiform layer (OPL), after 4weeks. Increased labeling of the OPL was detected. At 4 weeks,the staining profiles of both proteins were similar to thosefound at 2 weeks (Figs. 7A, 7B). WB analysis revealed lower
FIGURE 2. (A–O) Immunohistochemical characterization of intravitreally transplanted NPCs 2 weeks after propagation in culture. The sameantibodies were used for BNDF (A–C), oncomodulin (D–F), calcineurin (G–I), crybb2 (J–L), and cryg (M–O). DAPI (blue) was used to stain the cellnuclei. Scale bar: 50 lm (C, F, I, L, O).
8270 Bohm et al. IOVS, December 2012, Vol. 53, No. 13

levels of vimentin in the PBS group (122.48 6 48.17%, P <0.05) and the crybb2-NPC group (144.3 6 10.3%, P < 0.01)compared to the crybb2 group (204.84 6 17.42%) after 2weeks (Figs. 7I, 7J). In contrast, significantly decreasedvimentin was observed in the crybb2-NPC group (109.53 6
55.83%) compared to the crybb2 (205.23 6 28.46%) and PBSgroups (186.66 6 11.31%) after 4 weeks (P < 0.05; Figs. 7I,7J).
Retinal Microglia Responses to Intravitreal
Injection of crybb2-NPC and crybb2
Immunohistochemical evaluation revealed a significant in-crease in the recruitment and activation of microglia in thePBS group. Microglia were found in the GCL and NFL, while inthe crybb2 and crybb2-NPC groups those cells were observedin the IPL, INL, and OPL (Figs. 8A–D). Quantitatively, after 2weeks, 22.40 6 3.36 cells/0.2 mm2 of recruited microglia werefound in the PBS group, compared to 16.5 6 3.7 cells/0.2 mm2
in the crybb2 group (P < 0.05) and 13.86 6 1.58 cells/0.2mm2 in the crybb2-NPC group (P < 0.01; Fig. 8E). After 2weeks, increased numbers of activated microglia were found inthe PBS group (13.20 6 1.79 cells/0.2 mm2) compared to thecrybb2 group (7.50 6 2.38 cells/0.2 mm2, P < 0.05) and thecrybb2-NPC group (6.00 6 2.74 cells/0.2 mm2, P < 0.01; Fig.8F). After 4 weeks, microglia were observed in the GCL, NFL,and OPL in the PBS group, while in the crybb2 and crybb2-NPC groups they also were found in the ONL (data not shown).
In summary, fewer microglia were found in the crybb2group than in the other groups. Significantly increasednumbers of microglia were found in the PBS group (18.25 6
3.60 cells/0.2 mm2, P < 0.05) and the crybb2-NPC group(16.75 6 1.58 cells/0.2 mm2, P < 0.01) compared to thecrybb2 group (14.50 6 0.55 cells/0.2 mm2; Fig. 8E). Increasednumbers of activated cells were found in the crybb2-NPCgroup (12.13 6 1.73 cells/0.2 mm2) compared to the PBS
FIGURE 3. (A–F) Expression of retinal crybb2 (red) shown by immunohistochemical staining of slices (10 lm) of samples from animals treated withPBS (A, B), and crybb2 (C, D) after 2 and 4 weeks. (E) Expression of crybb2 by intravitreally implanted crybb2-NPC after 2 weeks. Cell nuclei aremarked blue with DAPI. (F) The negative control is shown with cyanine (Cy-3) as a secondary antibody. Scale bar: 20 lm (A–F).
TABLE 4. Localization of Grafted NPCs after Transplantation
Localization NPC – 2 weeks NPC – 4 weeks
Cornea X
AC X X
Ciliary body X
Vitreous X X
NFL
GCL X X
OPL X
ONL X X
IPL X
INL X X
RPE
Choroidea X
Grafted crybb2-NPC were found throughout the retinal layers, andin extraretinal tissue 2 and 4 weeks after grafting. AC, anteriorchamber.
IOVS, December 2012, Vol. 53, No. 13 Crystallin-b-b2-Overexpressing NPCs 8271
group (9.5 6 1.0 cells/0.2 mm2, P < 0.01) and the crybb2group (8.17 6 1.33 cells/0.2 mm2) after 4 weeks (P < 0.05;Fig. 8F). The data showed that microglial cells appear to besensitive indicators of activation in all of the experimentalgroups of this study.
DISCUSSION
In our study we examined the fate of intravitreally graftedNPCs within the eye and the responses of retinal cells to thegrafting of either nontransfected or transfected NPCs exempli-fied by crybb2-NPC. There were three principal findings in thisstudy:
1. NPCs reside and migrate within the eye and continue toproduce NTFs.
2. Crybb2 and crybb2-NPC reduce the axotomy-inducedloss of RGCs and changes in photoreceptor rhodopsin.crybb2-NPC reduce glial reactivity, indicating thatcrybb2 is involved in retinal repair mechanisms.
3. Crybb2 reduces the recruitment and activation of retinalmicroglia, suggesting a reduced inflammatory response.
Intraocular Characteristics of NPCs
NPCs harvested from cultures and injected into the vitreouscavity were able to produce the same markers used tocharacterize them in culture. This finding suggested that the
host environment does not change the profile of proteinssynthesized within the vitreous body. This observation wascrucial for setting the basic data for injecting transfectedNPCs. We observed a migration of intravitreally appliedcrybb2-NPC within the entire eye for up to 4 weeks aftergrafting. A previous study demonstrated an overall integra-tion of NPCs into the neonatal retina of rats after 4 weeks.11
In agreement with our results, previous studies havedetected the retina as the main donor for implanted NPCsin cases of injured,2 ischemic,27 or degenerating retinas.10
The preferred location was the ONL, the subretinal debriszone, and the layer of photoreceptors.10 Our observationsrevealed the detection of transplanted crybb2-NPC in thevitreoretinal compartment, subretinal space, and choroidafter 2 weeks, while crybb2-NPC were found in the GCL,INL, and ONL after 4 weeks. Crybb2-NPC were associatedwith the anterior chamber, corneal endothelium, and ciliarybody after 4 weeks. To our knowledge, this is the firstdemonstration of an overall migration of grafted crybb2-NPCthrough the entire rat eye. Migration over long distances isnot surprising, since it has been shown that NPCs are ableto produce extracellular matrix–degrading metallopro-teases.8
Responses of the Ocular Tissue
Crybb2 was found to support the survival of axotomized RGCsdue to compression and prevent the dissolution of rhodopsin
FIGURE 4. (A–O) Staining for nestin-expressing NPCs (green) in sections throughout the eye. Two weeks after transplantation, grafted NPCs werefound throughout the retina (e.g., IPL) (A–C), including the subretinal space (D–F) and the vitreoretinal compartment (G–I). After 4 weeks, nestin-positive cells were detectable in the anterior chamber and corneal endothelium (J–L) and the epithelium of the ciliary body (M–O). NPCs werefound only in the GCL, OPL, and IPL, and not in the other retinal layers after 4 weeks (data not shown). DAPI (blue) was used to stain the cell nuclei(P, R). The negative control is shown with Cy-2 as a secondary antibody. Arrows indicate nestin-positive NPCs in all images. Scale bar: 20 lm (C, F, I,L, O, R).
8272 Bohm et al. IOVS, December 2012, Vol. 53, No. 13

in photoreceptors. Crybb2 usually is produced within the
regenerating retina and facilitates the elongation of regenerat-
ing axons in retinal explants.26 Therefore, it is not surprising
that significantly fewer RGCs were lost in the crybb2 and
crybb2-NPC groups than in the PBS group. These data
confirmed the involvement of crybb2 in the repair of RGC
metabolism after injury to retinal axons, although the
mechanisms underlying the signal transduction remain to be
elucidated. Liedtke et al. have shown that GFP-labeled crybb2
is internalized by RGCs in culture.26 It is most likely that such
uptake takes place in vivo and is responsible for the survivaleffects observed in our study.
Reduction of rhodopsin and its delocalization within thePOS is an early indication of degenerative onset.31 We foundthat crybb2-NPC delayed rhodopsin decay.
It has been assumed that growth-factor failure is one reasonfor photoreceptor death in inherited retinal photoreceptordegenerations.32 The external supply of growth factors, such asbFGF, protects photoreceptors.33 Higher levels of growthfactors, such as bFGF, ciliary NTF (CNTF), and BNDF,34 werefound to be associated with photoreceptor death in RCS
FIGURE 5. (A–F) Effects of crybb2 and crybb2-NPC on RGCs in the retina. Expression of neurofilament 200 (NF-200, red) shown byimmunohistochemical staining of slices (10 lm) of samples from animals treated with PBS (A, B) and crybb2-NPC (C, D) after 2 and 4 weeks. Cellnuclei are marked blue with DAPI. Cy-3 was used for negative control as a secondary antibody (data not shown). (E) Number of persistent RGCs(per 0.2 mm2) in retinal slices (10 lm) in relation to that of adult control tissue (in %). The number of RGCs decreased in the PBS group (P < 0.001)and crybb2 group (P < 0.01) compared to crybb2-NPC–treated animals after 2 weeks. Decreased numbers of RGCs were observed in the PBS groupcompared to the crybb2 (P < 0.001) and the crybb2-NPC (P < 0.001) groups after 4 weeks. (F) WB analyses of retinas. Lysates of retinas treated asdescribed were prepared and tested for brain-specific homeobox/POU domain protein 3b (BRN-3b, 45 kiloDaltons). Calnexin was used as theloading protein control. Less protein was detected in the PBS group than in the crybb2 and crybb2-NPC groups after 2 and 4 weeks. Scale bar: 20lm (A–D). Statistical differences: **P < 0.01, ***P < 0.001.
IOVS, December 2012, Vol. 53, No. 13 Crystallin-b-b2-Overexpressing NPCs 8273

FIGURE 6. (A–D) Expression of retinal rhodopsin (green) shown by immunohistochemical staining of slices (10 lm) of samples from animals treatedwith PBS (A, E), crybb2 (B, F), and crybb2-NPC (C, G) after 2 and 4 weeks. The negative control is shown with Cy-2 (D, H) as the secondary antibody.(I) WB analyses of retinas were prepared and tested for rhodopsin expression. The expression of calnexin verified the amount of protein that loadedper lane. (J) Corresponding densitometric analyses of the WB results (n¼ 4) in PBS-, crybb2-, and crybb2-NPC–treated animals 2 and 4 weeks afterONC. The rhodopsin expression was lower in the crybb2-NPC group than in the crybb2 group after 2 weeks (P < 0.05). A significant reduction inrhodopsin expression was observed in the PBS group compared to the crybb2 (P < 0.001) and crybb2-NPC groups (P < 0.01) after 4 weeks. Therhodopsin expression was lowest in the PBS-treated group. Scale bar: 20 lm (A–H). Statistical differences: *P < 0.05, **P < 0.01, ***P < 0.001.
8274 Bohm et al. IOVS, December 2012, Vol. 53, No. 13

FIGURE 7. (A–H) Expression of retinal vimentin and GFAP, shown by immunohistochemical staining of slices (10 lm) of samples from animalstreated with PBS (A, B), crybb2 (C, D), and crybb2-NPC (E, F) 2 weeks after ONC. Glial cells are marked red with vimentin and green with GFAP.The negative control is shown with Cy-3 (G) and Cy-2 (H) as secondary antibodies. (I) WB analyses of retinas. Lysates of retinas treated as describedwere prepared and tested for vimentin expression. Calretinin expression verified the amount of protein that loaded per lane. (J) Correspondingdensitometric analyses of the WB results (n¼ 4) in PBS-, crybb2-, and crybb2-NPC–treated animals 2 and 4 weeks after ONC in relation to control.
IOVS, December 2012, Vol. 53, No. 13 Crystallin-b-b2-Overexpressing NPCs 8275

FIGURE 8. (A–D) Expression of ED-1 and Iba-1 in retinal microglial cells, as assessed by antibody staining of slices (10 lm) from animals treated withPBS (A), crybb2 (B), and NPC (C) 2 weeks after ONC. Microglial cells are marked red for ED-1 and green for Iba-1. ED-1 and IBA-1 positive cells werefound only in the NFL and GCL in the PBS-treated animals, whereas they also were found in the INL, IPL, and OPL in the crybb2- and NPC-treatedgroups. The negative control is shown with Cy-2 and Cy-3 (D) as the secondary antibodies. (E, F) Quantification of retinal microglial cells. (E) Countof ED-1/Iba-1-stained cells in the retinal layers. (F) Count of Iba-1 positive microglia cells. Scale bars: 20 lm (A–D). Statistical differences: *P < 0.05,**P < 0.01.
The vimentin expression significantly increased in the crybb2-treated animals compared to the control and the animals treated with PBS (P < 0.05)and crybb2-NPC (P > 0.01) after 2 weeks. After 4 weeks there were no significant differences in vimentin in the PBS- and crybb2-treated groups (P> 0.05), whereas there was less vimentin expression in the crybb2-NPC–treated group (P < 0.05). Scale bar: 20 lm (A–H). Statistical difference: *P< 0.05, **P < 0.01.
8276 Bohm et al. IOVS, December 2012, Vol. 53, No. 13

rats35,36 and rats with light-damaged retinas.37 It is likely thatthe factors produced by NPCs within the eye have a role inprotecting RGCs and photoreceptors from death. To this end,we have examined closely the overexpression of one of thesefactors, crybb2.
Crystallins maintain their effects on retinal proteins and arean important inhibitor of apoptosis due to toxic intermedi-ates.15 Increased levels of crystallins of the b/c superfamilywere found in animal models of retinal diseases, like retinitispigmentosa, light-induced damage, and diabetic retinopa-thy.38–40 Compared to observations of increased crybb2expression in regenerating retinas (e.g., GCL and INL) afterONC, no expression was found in unaffected eyes.26 This lineof evidence, together with our own findings, suggests thatcrybb2 exerts beneficial effects on damaged RGCs andsecondarily on photoreceptors. One of the mechanismsoperating through crybb2 may be an increase in the amountof neuroprotective CNTF released by astrocytes.16 To ourknowledge, no other studies have investigated the neuropro-tective effects of intravitreal crybb2 in vivo.
An early and significant increase in glial cell activity due tocrybb2 administration was found along with decreasedvimentin expression in the crybb2-NPC group. A complexglial network, including Muller cells and astrocytes, hasneuroprotective effects, releasing NTFs, such as bFGF, CNTF,and antioxidants after retinal injury.16,41,42
Early upregulation of vimentin and GFAP occurs in severalretinal disorders, such as retinopathia pigmentosa, maculardystrophies, or secondary retinal degenerations.43,44 and afterONC.45 The dynamic relationship between intermediatefilaments and retinal glial cells is the key mechanismunderlying the rapid modification of the Muller cell structurein response to changes in the retinal environment.46,47 It hasbeen shown that RGCs and astrocytes are donors of retinalcrystallins in vitro.23 Crystallins of the b/c superfamily arereleased during periods of critical stress in the retina.16,24
Increased retinal GFAP expression induces glial cells to switchinto a regenerative state after the intravitreal injection ofcrystallin b/c.16 Our observations confirmed the finding of adirect association between glial reactivity and crybb2. Thecrybb2-NPC–mediated effects on glial cells differ considerably.We concluded that additionally secreted NTFs by crybb2-NPCmay inhibit retinal glial activity.
NPCs and NTFs
While the exact mechanism underlying the effects of NPCswithin the retina are not understood fully, it is likely to bebased on growth-factor production.48 Retinal NTFs exertprotective effects on neighboring host neurons.49,50 SeveralNTFs delay RGC death and photoreceptor loss: fibroblastgrowth factors (e.g., bFGF), CNTF, brain-derived NTFs (e.g.,BDNF), and insulin-like growth factors. RGCs appear to bebetter protected by BDNF, while photoreceptors may be betterprotected by bFGF and CNTF.34 In our study, modified crybb2-NPC overexpressed crybb2 in addition to NTFs for up to 4weeks after transplantation in vivo.
Nestin is expressed during CNS stem-cell and progenitor-cell proliferation, and neuronal migration,51 and protectsprogenitor cells from apoptosis.33 BDNF, which is expressedby glial cells and neuronal cells,34,52 prolongs the survival ofRGCs in vitro, reduces RGC death during development, andexerts survival effects on injured adult neurons.33,53,54
Calcineurin is expressed in the RGCs of rat retinas and exertsimportant cellular effects in the immune and nervoussystems.55,56 It is involved in acute and chronic disordersresulting from glaucoma.56 Oncomodulin has a high affinity forRGCs, exerting axon-promoting effects on these cells in mature
optic nerves in vivo,57 and has a central role in inflammation-induced regeneration.58
Microglia
Microglial recruitment and activation are associated withretinal degeneration, as shown in the RCS rat.59 These cellsrespond to several proinflammatory factors, including lipo-polysaccharides and cytokines.60,61 On the other hand, theyproduce and secrete NTFs, such as CNTF, GDNF, and nervegrowth factor, to protect photoreceptors from cell death.62
The number of activated microglial cells was highest inthe PBS group after 2 weeks, suggesting an increasedresponse to dying neurons and inflammatory disorders.Inflammatory cytokines may promote the transformation oframified to active microglia. The effect is more predictablecompared to that of crybb2, which we attribute to increasedcell death within the retina. An increased activation andrecruitment of microglia in the crybb2-NPC group after 4weeks was another interesting finding. An attenuated loss ofRGCs and photoreceptors is indicative of a lower rate ofretinal cell death in this group. Furthermore, several NTFssecreted by NPCs have inhibitory effects on the cellularimmune response. We suggest a microglial reaction to thecellular response of NPCs.
CONCLUSIONS
In conclusion, to our knowledge this the first study to showthe migration of intravitreally injected NPCs through the entirerat eye. This finding implies that NPCs are able to interact withdifferent types of ocular tissue. Crybb2-NPC prevents RGC lossand decay of rhodopsin in the injury-induced model of retinaldegeneration. In addition to their survival and migration withinthe eye, crybb2-producing NPCs increase the chances ofsurvival of RGCs and photoreceptors. The expression offurther survival-promoting factors may be introduced in thesecells, thus assuring a permanent and approximately constantdelivery within the eye. This strategy of creating factor-producing ‘‘cell-factories’’ in the neighborhood of areas ofdegeneration may become a fruitful approach for thetreatment of retinal disorders by providing bioavailable growthfactors.
Acknowledgments
M. Wissing and M. Langkamp-Flock provided skillful technicalassistance, and M. Reis typed the manuscript. English ScienceEditing (ESE) provided native linguistic editing of the manuscript.
References
1. Gamm DM, Wang S, Lu B, et al. Protection of visual functionsby human neural progenitors in a rat model of retinal disease.PLoS One. 2007;2:e338.
2. Nishida A, Takahashi M, Tanihara H, et al. Incorporation anddifferentiation of hippocampus-derived neural stem cellstransplanted in injured adult rat retina. Invest Ophthalmol
Vis Sci. 2000;41:4268–4274.
3. Kim JH, Auerbach JM, Rodriguez-Gomez JA, et al. Dopamineneurons derived from embryonic stem cells function in ananimal model of Parkinson’s disease. Nature. 2002;418:50–56.
4. Wernig M, Benninger F, Schmandt T, et al. Functionalintegration of embryonic stem cell-derived neurons in vivo. J
Neurosci. 2004;24:5258–5268.
5. Richard I, Ader M, Sytnyk V, et al. Electroporation-based genetransfer for efficient transfection of neural precursor cells.Brain Res Mol Brain Res. 2005;138:182–190.
IOVS, December 2012, Vol. 53, No. 13 Crystallin-b-b2-Overexpressing NPCs 8277

6. Klein SM, Behrstock S, McHugh J, et al. GDNF delivery usinghuman neural progenitor cells in a rat model of ALS. Hum
Gene Ther. 2005;16:509–521.
7. Behrstock S, Ebert A, McHugh J, et al. Human neuralprogenitors deliver glial cell line-derived neurotrophic factorto parkinsonian rodents and aged primates. Gene Ther. 2006;13:379–388.
8. Charalambous P, Hurst LA, Thanos S. Engrafted chicken neuraltube-derived stem cells support the innate propensity foraxonal regeneration within the rat optic nerve. Invest
Ophthalmol Vis Sci. 2008;49:3513–3524.
9. Lepski G, Maciaczyk J, Jannes CE, Maciaczyk D, BischofbergerJ, Nikkhah G. Delayed functional maturation of humanneuronal progenitor cells in vitro. Mol Cell Neurosci. 2011;47:36–44.
10. Young MJ, Ray J, Whiteley SJ, Klassen H, Gage FH. Neuronaldifferentiation and morphological integration of hippocam-pal progenitor cells transplanted to the retina of immatureand mature dystrophic rats. Mol Cell Neurosci. 2000;16:197–205.
11. Takahashi M, Palmer TD, Takahashi J, Gage FH. Widespreadintegration and survival of adult-derived neural progenitorcells in the developing optic retina. Mol Cell Neurosci. 1998;12:340–348.
12. McLaren RE, Pearson RA, MacNeil A, et al. Retinal repair bytransplantation of photoreceptor precursors. Nature. 2006;444:203–207.
13. Robinson ML, Overbeek PA. Differential expression of alpha A-and alpha B-crystallin during murine ocular development.Invest Ophthalmol Vis Sci. 1996;37:2276–2284.
14. Zhang C, Gehlbach P, Gongora C, et al. A potential role forbeta- and gamma-crystallins in the vascular remodeling of theeye. Dev. Dyn. 2005;234:36–47.
15. Andley UP. Crystallins in the eye: function and pathology. Prog
Retin Eye Res. 2007;26:78–98.
16. Fischer D, Hauk TG, Muller A, Thanos S. Crystallins of the beta/gamma-superfamily mimic the effects of lens injury and promoteaxon regeneration. Mol Cell Neurosci. 2008;37:471–479.
17. Fort PE, Lampi KJ. New focus on alpha-crystallins in retinalneurodegenerative diseases. Exp Eye Res. 2010;92:98–103.
18. Sakaguchi H, Miyagi M, Darrow RM, et al. Intense lightexposure changes the crystallin content in retina. Exp Eye Res.2003;76:131–133.
19. Yoshimura N, Kikuchi T, Kuroiwa S, Gaun S. Differentialtemporal and spatial expression of immediate early genes inretinal neurons after ischemia-reperfusion injury. Invest
Ophthalmol Vis Sci. 2003;44:2211–2220.
20. Vazquez-Chona F, Song BK, Geisert EE Jr. Temporal changes ingene expression after injury in the rat retina. Invest
Ophthalmol Vis Sci. 2004;45:2737–2746.
21. Kumar PA, Haseeb A, Suryanarayana P, Ehtesham NZ, ReddyGB. Elevated expression of alphaA- and alphaB-crystallins instreptozotocin-induced diabetic rat. Arch Biochem Biophys.2005;444:77–83.
22. Fischer D, He Z, Benowitz LI. Counteracting the nogo receptorenhances optic nerve regeneration if retinal ganglion cells arein an active growth state. J Neurosci. 2004;24:1646–1651.
23. Sinha D, Esumi N, Jaworski C, Kozak CA, Pierce E, Wistow G.Cloning and mapping the mouse Crygs gene and non-lensexpression of [gamma]S-crystallin. Mol Vis. 1998;4:8.
24. Jones SE, Jomary C, Grist J, Makwana J, Neal MJ. Retinalexpression of gamma-crystallins in the mouse. Invest Oph-
thalmol Vis Sci. 1999;40:3017–3020.
25. Uchida K, Momiyama T, Okano H, et al. Potential functionalneural repair with grafted neural stem cells of early embryonicneuroepithelial origin. Neurosci Res. 2005;52:276–286.
26. Liedtke T, Schwamborn JC, Schroer U, Thanos S. Elongation ofaxons during regeneration involves retinal crystallin beta b2(crybb2). Mol Cell Proteomics. 2007;6:895–907.
27. Kurimoto Y, Shibuki H, Kaneko Y, et al. Transplantation ofadult rat hippocampus-derived neural stem cells into retinainjured by transient ischemia. Neurosci Lett. 2001;306:57–60.
28. Kong WC, Cho EYP. Antibodies against neurofilament subunitslabel retinal ganglion cells but not displaced amacrine cells ofhamsters. Life Sci. 1999;64:1773–1778.
29. Balaratnasingam C, Morgan WH, Bass L, Kang M, Cringle SJ, YuDY. Time-dependent effects of focal retinal ischemia on axonalcytoskeleton proteins. Invest Ophthalmol Vis Sci. 2010;51:3019–3028.
30. Badea TC, Nathans J. Morphologies of mouse retinal ganglioncells expressing transcription factors Brn3a, Brn3b, and Brn3c:analysis of wild type and mutant cells using genetically-directed sparse labeling. Vision Res. 2011;51:269–279.
31. Jones BW, Marc RE. Retinal remodeling during retinaldegeneration. Exp Eye Res. 2005;81:123–137.
32. Bramall AN, Wright AF, Jacobson SG, McInnes RR. The genomic,biochemical, and cellular responses of the retina in inheritedphotoreceptor degenerations and prospects for the treatmentof these disorders. Annu Rev Neurosci. 2010;33:441–472.
33. Mansour-Robaey S, Clarke DB, Wang YC, Bray GM, Aguayo AJ.Effects of ocular injury and administration of brain-derivedneurotrophic factor on survival and regrowth of axotomizedretinal ganglion cells. Proc Natl Acad Sci U S A. 1994;91:1632–1636.
34. Luthert PJ, Chong NH. Photoreceptor rescue. Eye (Lond).1998;12(Pt 3b):591–596.
35. Silverman MS, Hughes SE. Photoreceptor rescue in the RCS ratwithout pigment epithelium transplantation. Curr Eye Res.1990;9:183–191.
36. Li LX, Sheedlo HJ, Gaur V, Turner JE. Effects of macrophageand retinal pigment epithelial cell transplants on photorecep-tor cell rescue in RCS rats. Curr Eye Res. 1991;10:947–958.
37. Faktorovich EG, Steinberg RH, Yasumura D, Matthes MT, LaVailMM. Basic fibroblast growth factor and local injury protectphotoreceptors from light damage in the rat. J Neurosci. 1992;12:3554–3567.
38. Organisciak D, Darrow R, Gu X, Barsalou L, Crabb JW. Genetic,age and light mediated effects on crystallin protein expressionin the retina. Photochem Photobiol. 2006;82:1088–1096.
39. Piri N, Song M, Kwong JM, Caprioli J. Modulation of alpha andbeta crystallin expression in rat retinas with ocular hyperten-sion-induced ganglion cell degeneration. Brain Res. 2007;1141:1–9.
40. Fort PE, Freeman WM, Losiewicz MK, Singh RS, Gardner TW.The retinal proteome in experimental diabetic retinopathy:up-regulation of crystallins and reversal by systemic andperiocular insulin. Mol Cell Proteomics. 2009;8:767–779.
41. Schutte M, Werner P. Redistribution of glutathione in theischemic rat retina. Neurosci Lett. 1998;246:53–56.
42. Wilson RB, Kunchithapautham K, Rohrer B. Paradoxical roleof BDNF: BDNFþ/� retinas are protected against light damage-mediated stress. Invest Ophthalmol Vis Sci. 2007;48:2877–2886.
43. Erickson PA, Fisher SK, Guerin CJ, Anderson DH, Kaska DD.Glial fibrillary acidic protein increases in Muller cells afterretinal detachment. Exp Eye Res 1987;44:37–48.
44. Lewis GP, Matsumoto B, Fisher SK. Changes in the organiza-tion and expression of cytoskeletal proteins during retinaldegeneration induced by retinal detachment. Invest Ophthal-
mol Vis Sci. 1995;36:2404–2416.
45. Chen H, Weber AJ. Expression of glial fibrillary acidic proteinand glutamine synthetase by Muller cells after optic nerve
8278 Bohm et al. IOVS, December 2012, Vol. 53, No. 13

damage and intravitreal application of brain-derived neuro-trophic factor. Glia. 2002;38:115–125.
46. Nicholl ID, Quinlan RA. Chaperone activity of alpha-crystallinsmodulates intermediate filament assembly. EMBO J. 1994;13:945–953.
47. Fisher SK, Lewis GP, Linberg KA, Verardo MR. Cellularremodeling in mammalian retina: results from studies ofexperimental retinal detachment. Prog Retin Eye Res. 2005;24:395–431.
48. Klassen H, Ziaeian B, Kirov II, Young MJ, Schwartz PH.Isolation of retinal progenitor cells from post-mortem humantissue and comparison with autologous brain progenitors. J
Neurosci Res. 2004;77:334–343.
49. Hatakeyama J, Kageyama R. Retinal cell fate determination andbHLH factors. Semin Cell Dev Biol. 2004;15:83–89.
50. Reh TA, Fischer AJ. Retinal stem cells. Methods Enzymol.2006;419:52–73.
51. Qiu G, Seiler MJ, Thomas BB, Wu K, Radosevich M, Sadda SR.Revisiting nestin expression in retinal progenitor cells in vitroand after transplantation in vivo. Exp Eye Res 2007;84:1047–1059.
52. Jelsma TN, Aguayo AJ. Trophic factors. Curr Opin Neurobiol.1994;4:717–725.
53. Johnson JE, Barde YA, Schwab M, Thoenen H. Brain-derivedneurotrophic factor supports the survival of cultured ratretinal ganglion cells. J Neurosci. 1986;6:3031–3038.
54. Mey J, Thanos S. Intravitreal injections of neurotrophic factorssupport the survival of axotomized retinal ganglion cells inadult rats in vivo. Brain Res. 1993;602:304–317.
55. Nakazawa T, Takeda M, Lewis GP, et al. Attenuated glialreactions and photoreceptor degeneration after retinal de-tachment in mice deficient in glial fibrillary acidic protein andvimentin. Invest Ophthalmol Vis Sci. 2007;48:2760–2768.
56. Huang W, Fileta JB, Dobberfuhl A, et al. Calcineurin cleavage istriggered by elevated intraocular pressure, and calcineurininhibition blocks retinal ganglion cell death in experimentalglaucoma. Proc Natl Acad Sci U S A. 2005;102:12242–12247.
57. Yin Y, Henzl MT, Lorber B, et al. Oncomodulin is amacrophage-derived signal for axon regeneration in retinalganglion cells. Nat Neurosci. 2006;9:843–852.
58. Hauk TG, Muller A, Lee J, Schwendener R, Fischer D.Neuroprotective and axon growth promoting effects ofintraocular inflammation do not depend on oncomodulin orthe presence of large numbers of activated macrophages. Exp
Neurol. 2008;209:469–482.
59. Thanos S. Sick photoreceptors attract activated microglia fromthe ganglion cell layer: a model to study the inflammatorycascades in rats with inherited retinal dstrophy. Brain Res.1992;588:21–28.
60. Fries JE, Wheeler-Schilling TH, Guenther E, Kohler K.Expression of P2Y1, P2Y2, P2Y4, and P2Y6 receptor subtypesin the rat retina. Invest Ophthalmol Vis Sci. 2004;45:3410–3417.
61. Haynes SE, Hollopeter G, Yang G, et al. The P2Y12 receptorregulates microglial activation by extracellular nucleotides.Nat Neurosci. 2006;9:1512–1519.
62. Carwile ME, Culbert RB, Sturdivant RL, Kraft TW. Rod outersegment maintenance is enhanced in the presence of bFGF,CNTF and GDNF. Exp Eye Res 1998;66:791–805.
IOVS, December 2012, Vol. 53, No. 13 Crystallin-b-b2-Overexpressing NPCs 8279










![Research Paper HO-1 induced autophagy protects against IL ... · induce apoptosis of the nucleus pulposus cells (NPCs) in the degenerative intervertebral disc [5, 6]. Autophagy is](https://img.pdfslide.tips/doc/110x75/5e72f110b749c078843e28fa/research-paper-ho-1-induced-autophagy-protects-against-il-induce-apoptosis-of.jpg)






![Characterization of an antibody that recognizes peptides ... · in αA-crystallin (Asp 58 and Asp 151) [3], αB-crystallin (Asp 36 and Asp 62) [4], and βB2-crsytallin (Asp 4) [5]](https://img.pdfslide.tips/doc/110x75/5ff1e68e89243b57b64135f8/characterization-of-an-antibody-that-recognizes-peptides-in-a-crystallin-asp.jpg)











