Embed Size (px)
Citation preview

YONARA DE GOUVEIA CORDEIRO
Cultivo primário de células oriundas de carcinomas mamários de
cadelas e caracterização de possíveis populações de Células
Iniciadoras de Tumor
São Paulo
2015

YONARA DE GOUVEIA CORDEIRO
Cultivo primário de células oriundas de carcinomas mamários de
cadelas e caracterização de possíveis populações de Células
Iniciadoras de Tumor
Dissertação apresentada ao Programa de Pós-Graduação em Patologia Experimental e Comparada da Faculdade de Medicina Veterinária da Universidade de São Paulo para a obtenção do título de Mestre em Ciências
Departamento: Patologia
Área de Concentração: Patologia Veterinária
Orientador: Prof. Dr. Heidge Fukumasu
São Paulo
2015

Autorizo a reprodução parcial ou total desta obra, para fins acadêmicos, desde que citada a fonte.
DADOS INTERNACIONAIS DE CATALOGAÇÃO-NA-PUBLICAÇÃO
(Biblioteca Virginie Buff D’Ápice da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo)
T.3110 Cordeiro, Yonara de Gouveia FMVZ Cultivo primário de células oriundas de carcinomas mamários de cadelas e
caracterização de possíveis populações de Células Iniciadoras de Tumor / Yonara de Gouveia Cordeiro. -- 2015.
130 f. :il.
Dissertação (Mestrado) - Universidade de São Paulo. Faculdade de Medicina Veterinária e Zootecnia. Departamento de Patologia, São Paulo, 2015.
Programa de Pós-Graduação: Patologia Experimental e Comparada. Área de concentração: Patologia Experimental e Comparada. Orientador: Prof. Dr. Heidge Fukumasu.
1. Cães. 2. Células iniciadoras de tumor. 3. Câncer de mama. 4. Oncologia comparada. I. Título.

ERRATA CORDEIRO, Y. G. Cultivo primário de células oriundas de carcinomas mamários de cadelas e caracterização de possíveis p opulações de Células Iniciadoras de Tumor. 2015. 130 f. Dissertação (Mestrado em Ciências) - Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2015.
Página Onde se lê Leia-se
Capa: Título Células Iniciadoras de Tumor células iniciadoras de tumor
Página de rosto Células Iniciadoras de Tumor células iniciadoras de tumor
Folha de Avaliação Células Iniciadoras de Tumor células iniciadoras de tumor
Ficha catalográfica Células Iniciadoras de Tumor células iniciadoras de tumor
RESUMO Células Iniciadoras de Tumor células iniciadoras de tumor
ABSTRACT Células Iniciadoras de Tumor células iniciadoras de tumor


FOLHA DE AVALIAÇÃO
Nome: CORDEIRO, Yonara de Gouveia
Título: Cultivo primário de células oriundas de carcinomas mamários de cadelas e
caracterização de possíveis populações de Células Iniciadoras de Tumor
Dissertação apresentada ao Programa de
Pós-Graduação em Patologia Experimental e
Comparada da Faculdade de Medicina
Veterinária e Zootecnia da Universidade de
São Paulo para obtenção do título de Mestre
em Ciências
Data: ___/___/___
Banca Examinadora
Prof. Dr.: ____________________________________________________________
Instituição:______________________________________Julgamento:___________
Prof. Dr.: ____________________________________________________________
Instituição:______________________________________Julgamento:___________
Prof. Dr.: ____________________________________________________________
Instituição:______________________________________Julgamento:___________

À ela, meu exemplo de ser humano, meu exemplo de caráter e de coragem.
Meu porto seguro, minha conselheira, minha amiga, minha mãe.
Nada nessa minha humilde vida teria sentido sem você.
E tudo que eu conquistei, conquisto e conquistarei daqui pra frente,
foi, é e será, sempre graças à você.
Te amo muito, Ana Maria!

AGRADECIMENTOS
À Deus, sempre, por ter ouvido as minhas preces desesperadas quando já não tinha mais
pra onde correr. Por me fazer jamais perder a fé e a esperança. E por sempre me conduzir
ao caminho certo, na maioria das vezes não no momento em que pedi, mas no momento
que Ele sabia que era o certo;
À minha mãe Ana Maria, que sempre acreditou em mim. A pessoa a quem devo meu
caráter, minha força, minha independência. Obrigada por não ter desistido todas as vezes
em que eu pensei em desistir. Por ter me mantido aqui. Por ter me ensinado desde cedo
que quem faz nossa vida é a gente, basta corrermos atrás. Agradeço todos os dias por ter
vindo ao mundo através de você.
À minha família. Ao meu avô, meu exemplo de força e superação. Ao meu anjo da guarda,
minha avó, que de outro plano olha por mim em todos os momentos, e está ali me dando
proteção e acalmando meu coração. Às minhas tias, Iolanda, Nice e Tereza, pelos mimos,
carinhos e momentos de alegria. Aos tios, primos e primas, por me mostrarem o sentido da
palavra família. Nada me faz mais feliz do que voltar pra casa e sentir a alegria que é estar
com vocês.
Ao meu orientador, Prof. Dr. Heidge Fukumasu, que me recebeu desde o primeiro dia de
estágio obrigatório, e me deu a oportunidade de estar aqui hoje, anos depois. Sem dúvidas
é um exemplo de inteligência e sucesso e que, de muitas formas, contribuiu para o meu
crescimento profissional e pessoal aqui dentro.
Ao Prof. Dr. Ricardo de Francisco Strefezzi, pela contribuição com este projeto, pela
transmissão de conhecimento, e por sempre estar disposto a ajudar, foi um professor e
tanto; e ao Prof. Dr. José Bento. S. Ferraz pelos conselhos, pelas histórias, pelo carinho;
Aos clínicos veterinários que cederam amostras para este trabalho. Alessandra, Marcus,
Mariney, Juliana, Milena, Sílvio, Tânia, Beatriz, Bruno, Paulo, Nilson, Valmir, Mariana, João
Paulo. Sem vocês não teria sido possível;
Às amigas, Pâmela (sobrevivemos amiga!), Camila e Thayla, que estão comigo desde
sempre. Não teria a menor graça sem vocês aqui. Obrigada por tudo, pela amizade, por
cada lembrança compartilhada, por dividir tristezas e alegrias;
À todos os colegas e a amigos do LOCT. Principalmente ao Pedro, pelo companheirismo e
por sempre ter me ajudado quando precisei, sou muito feliz por ter você trabalhando comigo
e mais ainda por te ter como amigo; Arina, por ter me iniciado nesta vida louca de cultivo de

células, lá nos primórdios do estágio, e por ainda me ajudar nos momentos de dificuldades,
além de, claro, pelos momentos de amizade e descontração; Lídia, pela amizade, pelos
ótimos momentos aqui e nas viagens malucas, pelos conselhos, pelas dicas e ajuda com
protocolos e dissertação; Francisco, que vai entrar junto nos agradecimentos do LOCT sim!
Obrigada por tantos bons momentos, e por ter aguentado as crises de desespero. Enfim, a
todos os colegas deste novo laboratório que eu me orgulho em fazer parte. Só quem estava
aqui no antes e agora está no depois, sabe como é gratificante a sensação de estarmos tão
longe;
Aos alunos de Iniciação Científica, Fábio, Lucas, Sirlene, Erika e Fernanda, que
contribuíram para minha formação profissional aqui dentro, sempre tive muito gosto de estar
com vocês. Além de pseudo-orientados, ganhei amigos;
À Mariane, Jean e Leonardo, queridos amigos de departamento. São Paulo foi muito mais
divertida com vocês. Não teria como começar o mestrado de forma mais agradável. O
diamante de Homer ficará para sempre na memória;
Ao Prof. Dr. Flávio Meirelles, por compartilhar o espaço físico para o desenvolvimento deste
trabalho. E por todos os amigos que ali fiz durante este período, que estiveram presentes
em muitos momentos de alegria. Ana Mançanares, obrigada pela amizade e disposição
sempre. E a todos os demais colegas do LMMD;
À todos os amigos, principalmente paranaenses, por compreenderem cada vez que não
pude estar presente. Me fizeram muita falta. Câmi, Tassi e Aninha, para sempre na minha
vida e no meu coração;
À Faculdade de Zootecnia e Engenharia de Alimentos, pelo acolhimento. À Faculdade de
Medicina Veterinária e Zootecnia, e ao Departamento de Patologia Experimental e
Comparada, por proporcionar a formação acadêmica; e à CAPES, FAPESP e CNPq pelo
financiamento deste trabalho;
E à Becca, minha fiel companheira, que me dirige aquele olhar de amor que só os cães tem,
mesmo nos piores momentos de frustração.
Muito obrigada.

“E até lá, vamos viver Temos muito ainda por fazer
Não olhe pra trás
O mundo começa agora Apenas começamos”
- Renato Russo

RESUMO
CORDEIRO, Y. G. Cultivo primário de células oriundas de carcinomas mamários de cadelas e caracterização de possíveis populações de Células Iniciadoras de Tumor. [Primary cell culture of canine mammary carcinoma and characterization of possible populations of Tumor Initiation Cells]. 2015. 130 f. Dissertação (Mestrado em Ciências) - Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2015.
Em animais, a prevalência do câncer tem aumentado de forma significativa com o
passar dos anos. As neoplasias mamárias representam o tipo mais frequente de
câncer em cadelas, chegando a 52% da população de fêmeas, e entre os animais
afetados, 50% das neoplasias se apresentam sob a forma maligna. O
desenvolvimento e caracterização de modelos animais para o estudo de neoplasias
humanas é de extrema relevância para a melhoria no diagnóstico e tratamento do
câncer. Os tumores sólidos apresentam uma hierarquia entre as células que
determina o desenvolvimento e o comportamento da neoplasia. Recentemente, tem-
se estudado um pequeno grupo de células que apresentam diversas características
das células-tronco normais encontradas nos tecidos. Estas células, denominadas
Células Iniciadoras de Tumor (CITs), são descritas como sendo as principais
responsáveis pelas falhas na quimioterapia e no aparecimento de recidivas tumorais,
devido ao grande potencial de renovação e diferenciação que elas possuem. Desta
maneira, nosso objetivo foi caracterizar linhagens celulares provenientes de
neoplasia de glândula mamária de cadelas que pudessem ser utilizadas futuramente
na pesquisa básica e aplicada em oncologia comparada. Além das alterações
citogenéticas e imunocitoquímicas, verificamos que os cultivos celulares
apresentaram quantidades distintas de populações positivas e negativas quanto à
expressão de CD24 e CD44 bem como da atividade de aldeído-desidrogenase,
porém, todos exibiram potencial tumorigênico in vitro através do ensaio de formação
de tumoresferas.
Palavras-chave: Cães. Células iniciadoras de tumor. Câncer de mama. Oncologia
comparada.

ABSTRACT
CORDEIRO, Y. G. Primary cell culture of canine mammary carcinoma and characterization of possible populations of Tumor Initiation Cells. [Cultivo primário de células oriundas de carcinomas mamários de cadelas e caracterização de possíveis populações de Células Iniciadoras de Tumor]. 2015. 130 f. Dissertação (Mestrado em Ciências) - Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2015.
The prevalence of cancer in animals has increased significantly over the years.
Mammary tumors are the most common cancer in dogs, reaching 52% of female
population, and among affected animals, 50% of tumors are presented in the
malignant form. The development and characterization of animal models for the
study of human cancers is extremely important for improving the diagnosis and
treatment of cancer. Solid tumors have a hierarchy of cells that determines the
development and behavior of cancer. Recently, there has been studied a small group
of cells with different characteristics from those normal stem cells found in tissues.
These cells, known as tumor initiating cells (TICs), are described as being primarily
responsible for the failures in chemotherapy and the appearance of recurrences,
because of their potential for renewal and differentiation. Thus, our objective was to
characterize cell lines derived from mammary gland neoplasia of dogs that could be
further used for basic and applied research in comparative oncology. In addition to
the cytogenetic and immunocytochemical changes, we found that cell cultures had
different amounts of positive and negative populations of CD24 and CD44 expression
and aldehyde dehydrogenase activity, however, all exhibited tumorigenic potential in
vitro through tumorspheres formation.
Keywords: Canine. Comparative oncology. Mammary cancer. Tumor initiating cell.

LISTA DE FIGURAS
Figura 1 - Incidência e mortalidade de câncer no âmbito mundial ....................... 21
Figura 2 - Representação das taxas brutas de incidência por 100 mil indivíduos, em homens e mulheres, estimadas para o ano de 2014 .................................................................................................... 24
Figura 3 - Taxas de incidência do câncer de mama em mulheres na população mundial .............................................................................. 26
Figura 4 - Taxas brutas de incidência, em 100 mil mulheres, de neoplasias malignas de mama estimadas para o ano de 2014 ........... 26
Figura 5 - Modelo de desenvolvimento de tumores sólidos a partir da expansão clonal proposta por Nowell. ................................................. 31
Figura 6 - Modelos de desenvolvimento neoplásico. ........................................... 33
Figura 7 - Transição epitélio-mesenquimal (EMT) e transição mesenquima-epitelial (MET) para formação de tumor secundário .......................................................................................... 36
Figura 8 - Células provenientes da dissociação de neoplasias mamárias de cadelas, em condições de aderência ............................................. 64
Figura 9 - Lâmina histológica da amostra 005/13 ................................................ 66
Figura 10 - Morfologia do cultivo celular, amostra 005/13 ..................................... 67
Figura 11 - Lâmina histológica da amostra 020/14 ................................................ 68
Figura 12 - Morfologia do cultivo primário, amostra 020/14 ................................... 69
Figura 13 - Lâmina histológica da amostra 025/14 ................................................ 70
Figura 14 - Morfologia do cultivo celular, amostra 025/14 ..................................... 71
Figura 15 - Lâmina histológica, amostra 028/14 .................................................... 73
Figura 16 - Morfologia do cultivo celular, amostra 028/14 ..................................... 74
Figura 17 - Avaliação do número de cromossomos por coloração convencional, amostra 005/13 ............................................................. 77
Figura 18 - Avaliação do número de cromossomos por coloração convencional, amostra 005/13 ............................................................. 78
Figura 19 - Avaliação do número de cromossomos por coloração convencional, amostra 025/14 ............................................................. 79

Figura 20 - Avaliação do número de cromossomos por coloração convencional, amostra 025/14 ............................................................. 80
Figura 21 - Avaliação dos cromossomos por coloração convencional, amostra 028/14 ................................................................................... 81
Figura 22 - Avaliação dos cromossomos por coloração convencional, amostra 028/14 ................................................................................... 82
Figura 23 - Marcação imunocitoquímica, amostra 005/13 ..................................... 84
Figura 24 - Marcação imunocitoquímica, amostra 020/14 ..................................... 85
Figura 25 - Marcação imunocitoquímica, amostra 025/14 ..................................... 86
Figura 26 - Marcação imunocitoquímica, amostra 028/14 ..................................... 87
Figura 27 - Marcação imunocitoquímica para expressão de vimentina em diferentes passagens, amostra 020/14 ................................................ 88
Figura 28 - Marcação imunocitoquímica para expressão de vimentina em diferentes passagens, amostra 028/14 ................................................ 88
Figura 29 - Análise das populações de células com maior atividade de aldeído-desidrogenase ........................................................................ 90
Figura 30 - Formação de tumoresferas, amostra 020/14 ....................................... 97
Figura 31 - Agregados celulares formados a partir do sétimo dia, amostra 028/14 ................................................................................................. 97
Figura 32 - Formação de esferas em cultivo de baixa aderência após as separações por beads magnéticas .................................................... 102
Figura 33 - Aglomerado de células em proliferação, amostra 028/14 .................. 103

LISTA DE GRÁFICOS
Gráfico 1 - Incidência registrada de câncer na população mundial, homens e mulheres - 2012 ................................................................................. 21
Gráfico 2 - Mortalidade registrada na população mundial, homens e mulheres - 2012 .................................................................................... 22
Gráfico 3 - Frequência de neoplasias mamárias em fêmeas e distribuição dos casos por idade .............................................................................. 44
Gráfico 4 - Distribuição dos casos nos trinta e cinco cães coletados no estudo, baseado no tecido de origem e número de lesões por animal ................................................................................................... 60
Gráfico 5 - Distribuição dos casos quanto à idade (anos) e raça ............................ 61
Gráfico 6 - Análise do número de cromossomos em vinte metáfases por cultura celular ........................................................................................ 76
Gráfico 7 - Ensaio de formação de esferas em cultivo de baixa aderência em populações ALDHlow e ALDHhi, amostra 005/13 .............................. 94
Gráfico 8 - Ensaio de formação de esferas em cultivo de baixa aderência em populações ALDHlow e ALDHhi, amostra 020/14 .............................. 95
Gráfico 9 - Ensaio de formação de esferas em cultivo de baixa aderência em populações ALDHlow e ALDHhi, amostra 025/14 .............................. 95
Gráfico 10 - Ensaio de formação de esferas em cultivo de baixa aderência em populações ALDHlow e ALDHhi, amostra 028/14 .............................. 96
Gráfico 11 - Avaliação do crescimento de tumoresferas após a separação para os fenótipos CD24- e CD24+ ........................................................ 100
Gráfico 12 - Avaliação do crescimento de tumoresferas após a separação para os fenótipos CD44- e CD44+ ........................................................ 101

LISTA DE TABELAS
Tabela 1 - Distribuição das lesões segundo classificação histopatológica ............. 61
Tabela 2 - Culturas primárias de neoplasias de glândula mamária em cães, selecionadas para caracterização celular .................................... 64
Tabela 3 - Tempo de duplicação celular das quatro amostras cultivadas. .............. 75
Tabela 4 - Porcentagens de células ALDH+ nas quatro amostras de carcinomas mamários caninos .............................................................. 89
Tabela 5 - Quantificação das populações CD24 e CD44 através da separação por beads magnéticas, em neoplasia mamária de cães ...................................................................................................... 98

LISTA DE QUADROS
Quadro 1 - Incidência e mortalidade do câncer no mundo ...................................... 20
Quadro 2 - Estimativa dos dez tipos de câncer mais incidentes no Brasil, exceto pele não-melanoma - 2014 ........................................................ 23
Quadro 3 - Marcadores de células-tronco cancerosas em tumores sólidos ............. 39
Quadro 4 - Prospecção do isolamento de células-tronco de tumores mamários humanos de tecidos frescos dissociados .............................. 40
Quadro 5 - Principais características relacionadas ao estudo da espécie Canis familiaris na oncologia comparada. ............................................. 42
Quadro 6 - Clínicas parceiras que contribuíram com amostras para o presente estudo .................................................................................... 51
Quadro 7 - Anticorpos utilizados para avaliação imunocitoquímica ......................... 55

SUMÁRIO
1 INTRODUÇÃO ............................................................................................... 18
2 REVISÃO DE LITERATURA.......................................................................... 19
2.1 CONCEITOS GERAIS SOBRE CÂNCER ...................................................... 19
2.2 NEOPLASIAS MAMÁRIAS ............................................................................. 24
2.3 MODELOS DE DESENVOLVIMENTO NEOPLÁSICO ................................... 29
2.3.1 Modelo estocástico ou expansão clonal .................................................... 29
2.3.2 Modelo hierárquico ou teoria das Células-tronco Cancerosas ................. 32
2.4 CÉLULAS-TRONCO CANCEROSAS NO CÂNCER DE MAMA ..................... 37
2.5 ONCOLOGIA COMPARADA .......................................................................... 40
2.5.1 Tumores espontâneos em cães – neoplasias mamárias ........................... 40
2.5.2 Células-tronco cancerosas em neoplasias mamárias em cães ................ 46
3. OBJETIVO GERAL ........................................................................................ 49
4. OBJETIVOS ESPECÍFICOS .......................................................................... 50
5. MATERIAL E MÉTODOS .............................................................................. 51
5.1 MATERIAL UTILIZADO NO ESTUDO ............................................................ 51
5.2 CULTIVO PRIMÁRIO DE CÉLULAS ORIUNDAS DE NEOPLASIAS
MAMÁRIAS .................................................................................................... 52
4.3 TEMPO DE DUPLICAÇÃO CELULAR ........................................................... 53
5.4 AVALIAÇÃO CITOGENÉTICA POR CARIOTIPAGEM ................................... 54
5.5 IMUNOCITOQUÍMICA.................................................................................... 55
5.6 SEPARAÇÃO DE POPULAÇÕES CELULARES POR BEADS
MAGNÉTICAS ATRAVÉS DO PERFIL DE EXPRESSÃO DE CD24/CD44 .... 56
5.7 SEPARAÇÃO DE POPULAÇÕES POSITIVAS PARA A ATIVIDADE DE
ALDEÍDO-DESIDROGENASE (ALDH) ATRAVÉS DE CITOMETRIA DE
FLUXO ........................................................................................................... 57
5.8 CULTIVO CELULAR PARA FORMAÇÃO DE TUMORESFERAS .................. 58
5.9 ANÁLISE ESTATÍSTICA ................................................................................ 59
6. RESULTADOS............................................................................................... 60
6.1 DISTRIBUIÇÃO DOS CASOS CLÍNICOS ...................................................... 60
6.2 CARACTERIZAÇÃO DOS CULTIVOS CELULARES ..................................... 62
6.2.1 Dissociação e cultivo primário em monocamada ...................................... 62
6.2.2 Classificação histopatológica e morfologia da cultura primária .............. 65

6.2.3 Tempo de duplicação celular ...................................................................... 74
6.2.4 Cariotipagem ................................................................................................ 75
6.2.5 Imunocitoquímica ......................................................................................... 83
6.2.6 Separação de populações positivas para a atividade de aldeído-
desidrogenase (ALDH) através de citometria de fluxo .............................. 89
6.2.7 Separação de populações celulares por beads magnéticas através
do perfil de expressão de CD24/CD44 ........................................................ 98
7 DISCUSSÃO ................................................................................................ 104
8 CONCLUSÃO .............................................................................................. 116
9 REFERÊNCIAS............................................................................................ 117

18
1 INTRODUÇÃO
O câncer está entre as principais causas de morbidade e mortalidade no
mundo, sendo a segunda maior causa de morte, atrás apenas das doenças
cardiovasculares. Para 2030, estima-se que a carga global será de 21,4 milhões de
casos novos de câncer e 13,2 milhões de mortes por câncer, em consequência do
crescimento e do envelhecimento da população, bem como da redução na
mortalidade infantil e nas mortes por doenças infecciosas em países em
desenvolvimento. Durante décadas, a pesquisa minuciosa das bases moleculares do
câncer vem tentando esclarecer as causas da transformação oncogênica e o
aparecimento de neoplasias ainda não detectáveis.
Principalmente para tumores sólidos, como é o caso do câncer de mama, a
heterogeneidade e a complexidade molecular impõem muitos desafios para o
desenvolvimento de estratégias efetivas para a prevenção e a cura desta doença.
Neste contexto, há um grande investimento nas pesquisas que buscam entender e
desvendar a origem dos tumores, e entre elas, se encontra a teoria das células-
tronco cancerosas, onde apenas um grupo de células seria capaz de dar origem a
um novo tumor, além da mediação das vias de metástase e falhas na quimioterapia.
Se correta, pode fornecer informações valiosas que explicariam muitas das
limitações encontradas nos modelos atuais, sugerindo novas estratégias para o
controle das neoplasias mamárias. Por causa da estreita relação entre CITs com a
iniciação, progressão, metástase e resistência às terapias, técnicas de isolamento,
identificação e caracterização destas células para compreensão da biologia celular e
molecular, devem ser cada vez mais precisas para continuar os avanços na
pesquisa em câncer. Assim, temos de levar em conta os benefícios que a
investigação na oncologia comparada pode oferecer não só para a saúde animal,
mas também à medicina humana.

19
2 REVISÃO DE LITERATURA
2.1 CONCEITOS GERAIS SOBRE CÂNCER
O termo “câncer” pode ser considerado como um termo genérico, o qual
abrange um grupo de doenças de diferentes etiologias, características e implicações
que acometem qualquer parte do organismo. É definido pela proliferação
descontrolada de células que ultrapassam seus limites usuais e podem interferir não
só na arquitetura e função do próprio tecido de origem, mas também em tecidos
adjacentes a ele, seja por compressão física ou invasão de células tumorais
(KUMAR, 2011). Neste último processo, as células neoplásicas podem alcançar e
invadir vasos sanguíneos e linfáticos, disseminando-se para outras regiões e
implantando-se em um segundo tecido, dando origem a uma neoplasia secundária.
Esta evolução é conhecida como processo de invasão e formação de metástases.
Neste processo, segundo a Sociedade Americana do Câncer (ACS – American
Cancer Society), que é responsável por quase 90% das mortes pela doença no
mundo, as células do tumor primário adquirem algumas habilidades que permitem
com que estas células invadam a matriz extracelular. Para isso, é preciso que haja
uma ação concomitante entre estes dois componentes: as células primárias
necessitam selecionar as características vantajosas a elas, e a matriz extracelular
deve apresentar componentes que permitam a adaptação e acomodamento destas
células para que aconteça o desenvolvimento da nova formação (GUPTA;
MASSAGUÉ, 2006).
O câncer é uma das doenças que mais desafiam a medicina não só pela
complexidade no seu desenvolvimento, mas também pela dificuldade do tratamento,
raramente eficaz. Apesar dos grandes avanços realizados pela medicina quanto ao
diagnóstico e tratamento, seja na oncologia da pesquisa básica, ou na oncologia
clínica aplicada, ainda nos confrontamos com um alto índice de incidência e
mortalidade causado pelo câncer em todo o mundo, sendo que estes números
alarmantes não ficam restritos a uma determinada raça, idade, sexo ou condição
social. Segundo estatísticas da IARC (International Agency for Research on Cancer
– pertencente à Organização Mundial da Saúde), em 2012 foram registrados 14,1

20
milhões de novos casos, 8,2 milhões de mortes pela doença, e mais de 32 milhões
de pessoas vivendo com a doença no mundo (Quadro 1), (GLOBOCAN, 2012). Para
2025, a tendência é um aumento do número de casos: levando em consideração
somente as projeções das mudanças demográficas mundiais, estima-se que 19
milhões de casos sejam diagnosticados em homens e mulheres. As mudanças na
fertilidade e o aumento na expectativa de vida acabam por levar a uma rápida
expansão e envelhecimento da população mundial, atingindo uma escala
indesejável de problemas relacionados ao câncer, principalmente em países em
desenvolvimento (Figura 1) (ACS; IARC, 2014).
Quadro 1 - Incidência e mortalidade do câncer no mundo
Fonte: International Agency for Research on Cancer. Organização Mundial da Saúde Legenda: Em números, 57% dos casos novos, 65% das mortes pela doença e 48% casos prevalentes ocorreram em regiões menos desenvolvidas – 2012
É sabido que alguns órgãos são mais propensos a desenvolver uma
neoplasia do que outros, apresentando uma maior incidência e prevalência no
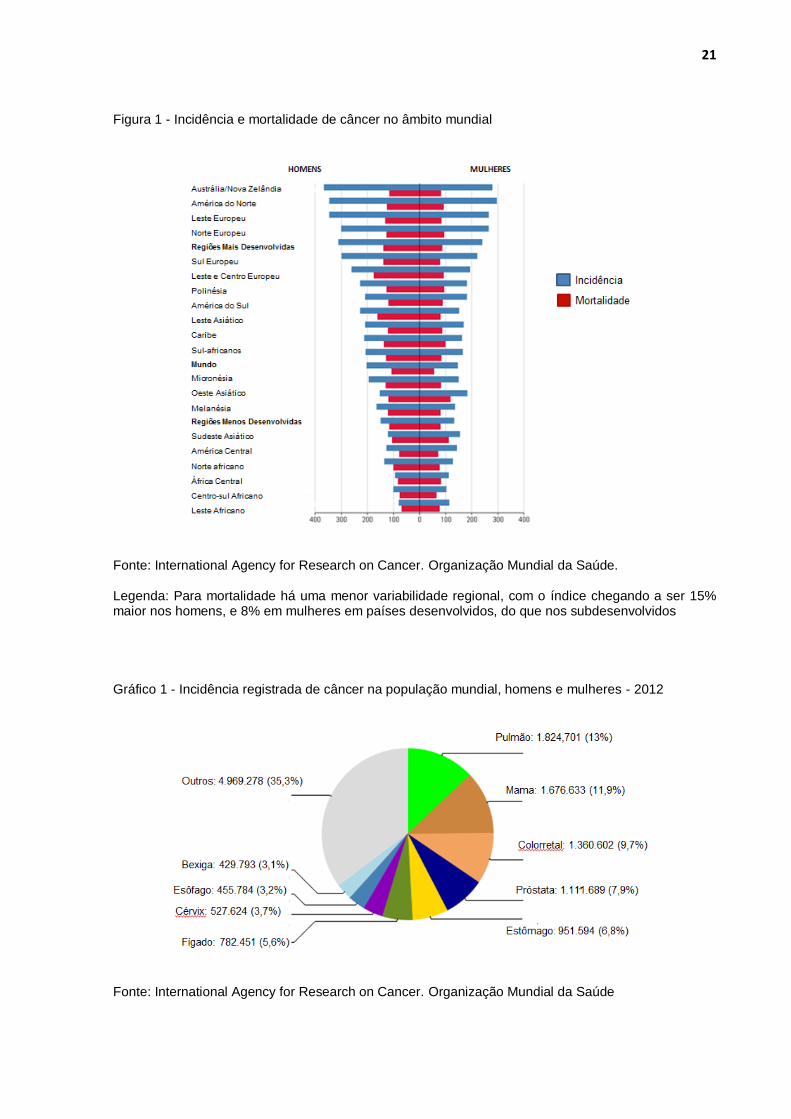
cenário atual. Dos 14 milhões de casos diagnosticados, a maior incidência é
neoplasias pulmonares, seguido de por neoplasias mamárias, colorretais, de
próstata, estômago e fígado. Este quadro não difere muito entre ambos os sexos.
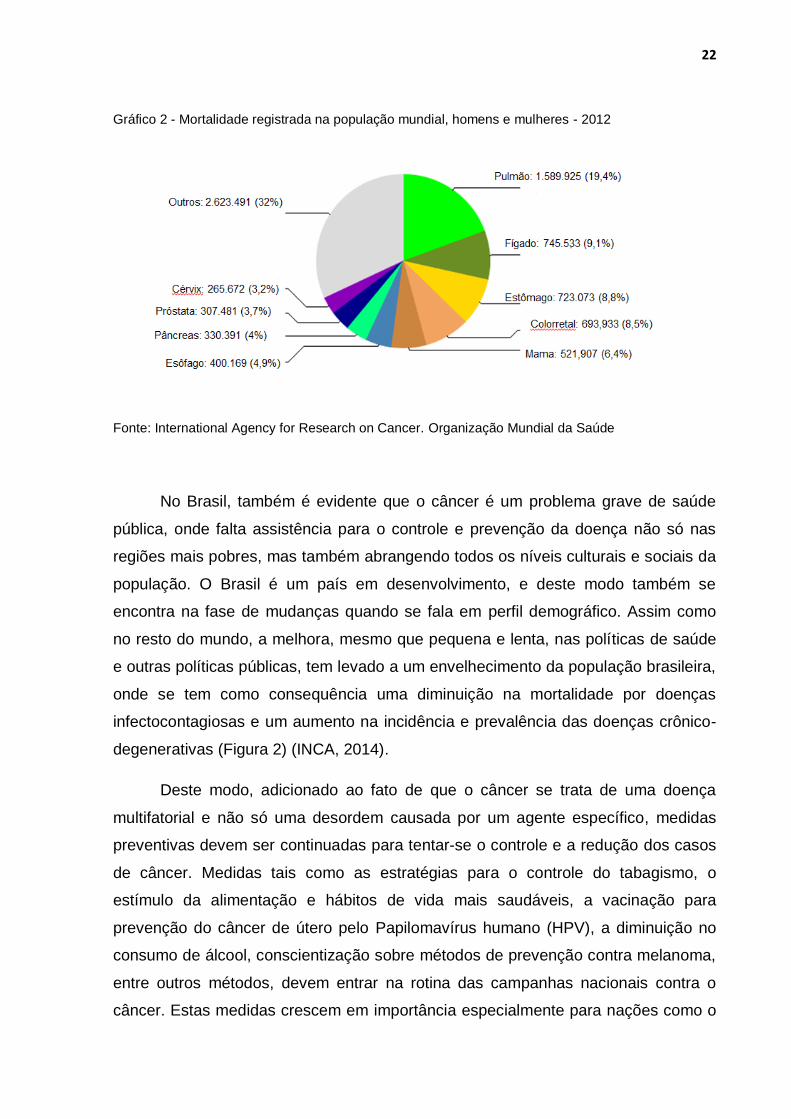
Nas mulheres, a neoplasia responsável pelo maior número de mortes ainda é o
câncer de mama, seguido das neoplasias de pulmão, colorretal, cérvix, estômago e
fígado; em homens, o principal órgão é o pulmão, seguido de fígado, estômago,
câncer colorretal e câncer de próstata (Gráficos 1 e 2).
21
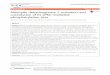
Figura 1 - Incidência e mortalidade de câncer no âmbito mundial
Fonte: International Agency for Research on Cancer. Organização Mundial da Saúde. Legenda: Para mortalidade há uma menor variabilidade regional, com o índice chegando a ser 15% maior nos homens, e 8% em mulheres em países desenvolvidos, do que nos subdesenvolvidos
Gráfico 1 - Incidência registrada de câncer na população mundial, homens e mulheres - 2012
Fonte: International Agency for Research on Cancer. Organização Mundial da Saúde
22
Gráfico 2 - Mortalidade registrada na população mundial, homens e mulheres - 2012
Fonte: International Agency for Research on Cancer. Organização Mundial da Saúde
No Brasil, também é evidente que o câncer é um problema grave de saúde
pública, onde falta assistência para o controle e prevenção da doença não só nas
regiões mais pobres, mas também abrangendo todos os níveis culturais e sociais da
população. O Brasil é um país em desenvolvimento, e deste modo também se
encontra na fase de mudanças quando se fala em perfil demográfico. Assim como
no resto do mundo, a melhora, mesmo que pequena e lenta, nas políticas de saúde
e outras políticas públicas, tem levado a um envelhecimento da população brasileira,
onde se tem como consequência uma diminuição na mortalidade por doenças
infectocontagiosas e um aumento na incidência e prevalência das doenças crônico-
degenerativas (Figura 2) (INCA, 2014).
Deste modo, adicionado ao fato de que o câncer se trata de uma doença
multifatorial e não só uma desordem causada por um agente específico, medidas
preventivas devem ser continuadas para tentar-se o controle e a redução dos casos
de câncer. Medidas tais como as estratégias para o controle do tabagismo, o
estímulo da alimentação e hábitos de vida mais saudáveis, a vacinação para
prevenção do câncer de útero pelo Papilomavírus humano (HPV), a diminuição no
consumo de álcool, conscientização sobre métodos de prevenção contra melanoma,
entre outros métodos, devem entrar na rotina das campanhas nacionais contra o
câncer. Estas medidas crescem em importância especialmente para nações como o
23
Brasil, que se encontram não só neste processo de transição demográfica, mas
também em uma condição de transição econômica, o que consequentemente leva
ao ônus global do câncer observado em países economicamente mais
desenvolvidos (INCA, 2014).
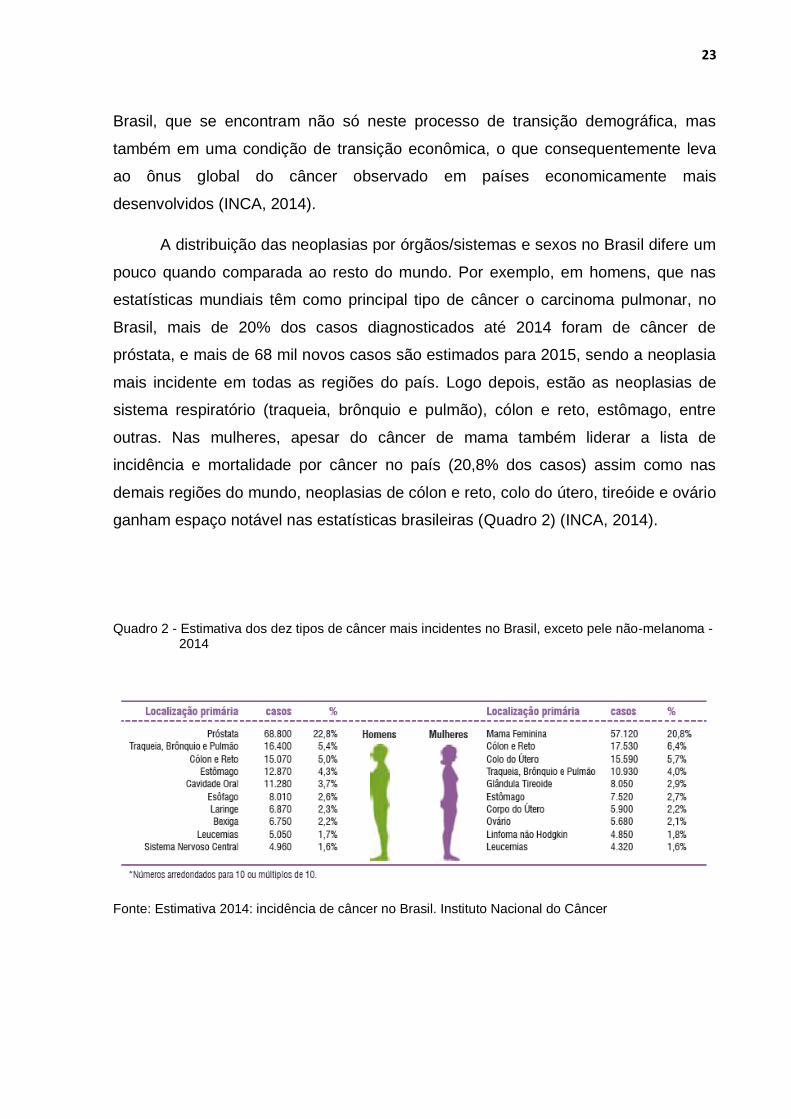
A distribuição das neoplasias por órgãos/sistemas e sexos no Brasil difere um
pouco quando comparada ao resto do mundo. Por exemplo, em homens, que nas
estatísticas mundiais têm como principal tipo de câncer o carcinoma pulmonar, no
Brasil, mais de 20% dos casos diagnosticados até 2014 foram de câncer de
próstata, e mais de 68 mil novos casos são estimados para 2015, sendo a neoplasia
mais incidente em todas as regiões do país. Logo depois, estão as neoplasias de
sistema respiratório (traqueia, brônquio e pulmão), cólon e reto, estômago, entre
outras. Nas mulheres, apesar do câncer de mama também liderar a lista de
incidência e mortalidade por câncer no país (20,8% dos casos) assim como nas
demais regiões do mundo, neoplasias de cólon e reto, colo do útero, tireóide e ovário
ganham espaço notável nas estatísticas brasileiras (Quadro 2) (INCA, 2014).
Quadro 2 - Estimativa dos dez tipos de câncer mais incidentes no Brasil, exceto pele não-melanoma - 2014
Fonte: Estimativa 2014: incidência de câncer no Brasil. Instituto Nacional do Câncer

24
Figura 2 - Representação das taxas brutas de incidência por 100 mil indivíduos, em homens e
mulheres, estimadas para o ano de 2014
Fonte: Estimativa 2014: incidência de câncer no Brasil. Instituto Nacional do Câncer Legenda: estão representadas as taxas por estados da Federação, de todas as neoplasias malignas exceto as de pele não-melanoma - 2014
Todos os dados de taxas de incidência e estimativas de novos casos de
câncer no Brasil para o biênio 2014/2015 foram retirados das publicações realizadas
pelo Instituto Nacional Do Câncer, Rio de Janeiro, Brasil. Sabe-se que estimativas e
projeções são importantes para a descrição e entendimento dos padrões atuais de
incidência de câncer, e, apesar das limitações, permitem o dimensionamento do
impacto desta doença no Brasil e no mundo, assessorando na tomada de decisões a
fim de reduzir cada vez mais os números dos próximos levantamentos.
2.2 NEOPLASIAS MAMÁRIAS
O diagnóstico, prognóstico e tratamento para o câncer de mama é uma das
áreas que mais progride em pesquisas e resultados na oncologia, principalmente
porque esta neoplasia apresenta a maior taxa de incidência e prevalência, e
representa a maior causa de mortalidade nas mulheres em todo o mundo. Segundo
a IARC, a estimativa para o ano de 2012 era de cerca de 1,67 milhões de novos
casos diagnosticados, representando 25% de todas as neoplasias (Figura 3)

25
(GLOBOCAN, 2012). No Brasil a situação não é diferente. Para 2014/2015, são
esperados 57.120 novos casos de câncer de mama, sendo o risco estimado de
56,09 casos a cada 100 mil mulheres, somando-se os dados de todos os estados
(Figura 4). As taxas de mortalidade são desiguais entre as regiões, sendo que as
áreas de melhor situação socioeconômica têm uma leve tendência à diminuição e
estabilidade, enquanto que aquelas com uma pior condição socioeconômica
apresentam uma maior probabilidade de aumento dos índices de mortalidade com o
passar do tempo. No entanto, a sobrevida de mulheres diagnosticadas com câncer
de mama no Brasil compara-se com a de países desenvolvidos, como a Inglaterra. A
sobrevida em um, cinco e vinte anos publicada para as britânicas é de 95%, 85% e
64%, respectivamente, enquanto que no Brasil, a sobrevida estimada é de
aproximadamente de 80% em cinco anos (INCA, 2014).
É a neoplasia mais comum tanto em países desenvolvidos quanto
subdesenvolvidos, com uma leve tendência a um aumento nos países de baixa e
média renda, novamente explicado pelo envelhecimento da população, urbanização,
hábitos de vida mais saudáveis e melhoria nas práticas de diagnóstico. Recursos
limitados com sistemas de saúde ineficientes, onde a incidência de câncer de mama
é relativamente baixa e a maioria das mulheres são diagnosticadas em estágios
avançados, têm a opção de implementar programas de diagnóstico precoce com
base no conhecimento dos sinais e sintomas pelas próprias mulheres, e
encaminhamento imediato para diagnóstico e tratamento. A detecção precoce, a fim
de melhorar os resultados do câncer de mama e de sobrevida após o diagnóstico
continua a ser a base do controle desta neoplasia.
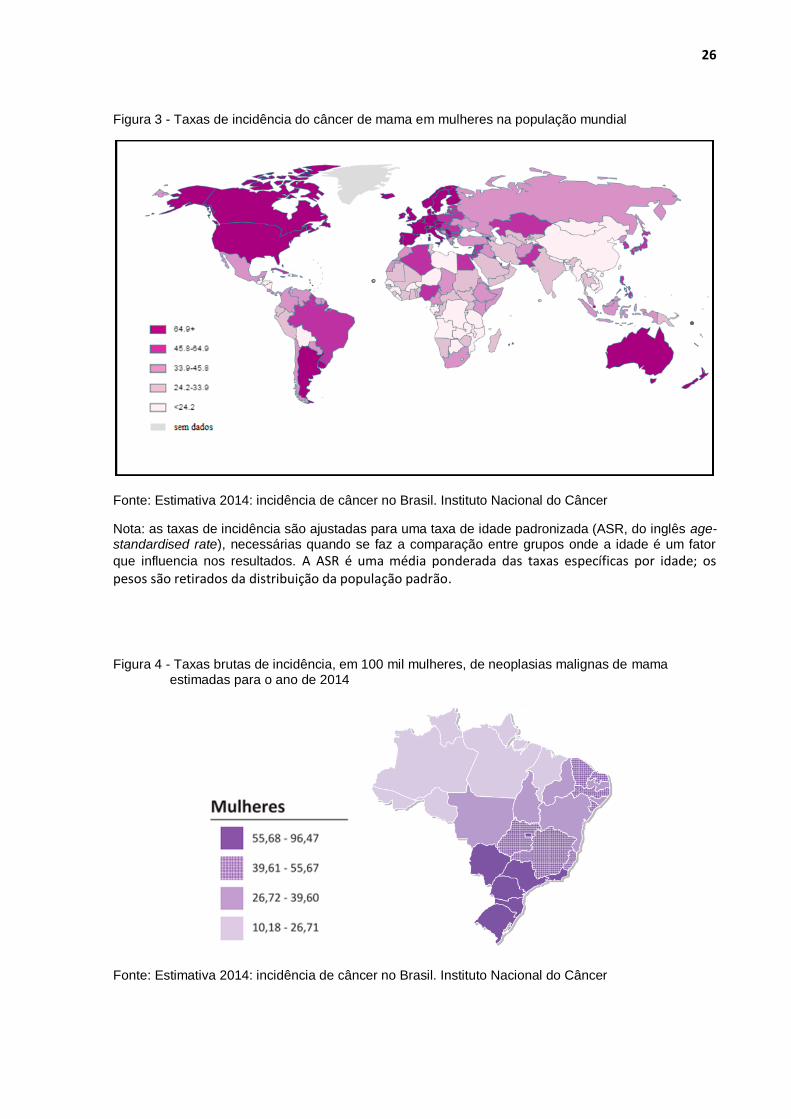
26
Figura 3 - Taxas de incidência do câncer de mama em mulheres na população mundial
Fonte: Estimativa 2014: incidência de câncer no Brasil. Instituto Nacional do Câncer
Nota: as taxas de incidência são ajustadas para uma taxa de idade padronizada (ASR, do inglês age-standardised rate), necessárias quando se faz a comparação entre grupos onde a idade é um fator
que influencia nos resultados. A ASR é uma média ponderada das taxas específicas por idade; os pesos são retirados da distribuição da população padrão.
Figura 4 - Taxas brutas de incidência, em 100 mil mulheres, de neoplasias malignas de mama estimadas para o ano de 2014
Fonte: Estimativa 2014: incidência de câncer no Brasil. Instituto Nacional do Câncer

27
Já se sabe que alguns fatores são considerados de risco para o
desenvolvimento de um tumor mamário primário. Elementos como: mutações
genéticas herdáveis bem definidas, como nos genes BRCA1 e BRCA2 (KING;
MARKS; MANDELL, 2003; WALSH; KING, 2007); número de gestações e idade ao
primeiro parto (LORD et al., 2008); obesidade e o risco associado de câncer de
mama em mulheres na pós-menopausa (CLEARY; GROSSMANN, 2009), podem
estar relacionados com uma maior ou menor probabilidade de um indivíduo, no caso
uma mulher, vir a apresentar um nódulo em que se confirme o diagnóstico de
neoplasia mamária. Mas, mesmo com algumas vias destes elementos já bem
definidas pela ciência, ainda não podemos predizer com certeza quem um dia
desenvolverá um câncer, e quem nunca apresentará a doença.
A soma dos diversos fatores de risco, estes sujeitos a modificações e sem
considerar os fatores reprodutivos, foi avaliada por Danaei et.al. (2005). Depois de
correlacionar diversos tipos de câncer a nove fatores de risco diferentes, chegaram à
conclusão de que mais de 20% de todas as mortes causadas por neoplasias
mamárias no mundo estão relacionadas ao consumo excessivo de álcool e da
obesidade, consequência do excesso de peso, falta de atividades físicas, ou de
ambos os fatores associados (DANAEI et al., 2005). Isto demonstra que a redução
da exposição aos principais fatores de risco comportamentais e ambientais impediria
uma substancial proporção de mortes por câncer no mundo.
Clinicamente, é uma doença tipicamente heterogênea, que pode ser
categorizada de diversas formas. O que normalmente é feito, é a classificação
destas neoplasias de acordo com a apresentação histológica, e posteriormente,
através da expressão ou não de marcadores tumorais, divididos em três principais
grupos: o das neoplasias que apresentam receptores de estrógeno e/ou
progesterona positivos (ER/PR), sendo o grupo mais variado e numeroso; aqueles
que são positivos para receptores da proteína HER2, o qual representa o grupo de
maior sucesso terapêutico devido à eficácia da terapia direcionada a este gene; e o
grupo dos cânceres triplo-negativos (Triple-negative Breast Cancers – TNBCs), onde
a célula cancerosa não responde a nenhum dos três receptores anteriores, sendo
considerado o mais maligno pela ineficiência terapêutica quando tratados com
medicamentos utilizados no tratamento dos demais grupos (THE CANCER
GENOME ATLAS NETWORK, 2012). Este último subtipo compreende em torno de

28
20% dos tumores mamários, ressaltando a deficiência na compreensão das vias que
regem a progressão tumoral, desfavorecendo assim o prognóstico das pacientes
dada a propensão para recidivas e metástases. (HURVITZ; FINN, 2009).
Histologicamente, os dois tipos mais frequentes são o carcinoma ductal e o
carcinoma lobular, representando cerca de 75% e 15% de todas as neoplasias
mamárias em mulheres, respectivamente (LI; URIBE; DALING, 2005; KORHONEN
et al., 2013). Os carcinomas lobulares apresentam parâmetros mais favoráveis,
como a positividade de receptores hormonais, baixos graus histológicos, baixos
índices proliferativos, apesar de com frequência apresentarem mais negatividade
para receptores HER2 quando comparados com os carcinomas ductais (DIXON;
ANDERSON; PAGE, 1996; ARPINO et al., 2004; RAKHA et al., 2008). Há um
interesse especial no estudo dos carcinomas lobulares, pois mesmo mostrando
características mais brandas do que os ductais, dados americanos salientam para o
rápido aumento nas taxas de incidência deste tumor em um intervalo de tempo
relativamente pequeno: de 1987 a 1999, enquanto a incidência de carcinoma ductal
subiu apenas 3%, as taxas de carcinoma lobular tiveram um aumento de 65% no
mesmo período, provavelmente relacionado ao uso de hormônios para reposição
pós-menopausa, e às melhorias nas técnicas de diagnóstico (LI et al., 2003).
Vários estudos vêm se dedicando a entender a diferença epidemiológica e na
progressão entre estes, que consistem nos dois principais subtipos. Muito menos se
sabe sobre os tipos histológicos mais raros, que inclui os carcinomas tubulares,
mucinosos, comedos, inflamatório, medulares, e carcinomas papilíferos, que juntos
respondem a cerca de 10% de todos os casos (LI; URIBE; DALING, 2005). Mesmo
com tantos avanços científicos, ainda há muito que se entender sobre o
desenvolvimento e progressão dos tumores sólidos, principalmente mamários,
visando sempre a prevenção, diagnóstico precoce, eficiência no tratamento e
qualidade de vida das pacientes.

29
2.3. MODELOS DE DESENVOLVIMENTO NEOPLÁSICO
A origem e a organização do câncer vêm sendo muito discutida atualmente,
uma vez que os pesquisadores ainda não conseguiram explicar como se desenvolve
a heterogeneidade característica das neoplasias, principalmente em tumores
sólidos. Hoje, teorias demonstram que células progenitoras ou mesmo células-tronco
teciduais seriam passíveis de uma transformação genética, podendo assim dar início
ao desenvolvimento neoplásico (WANG, 2010). Na tentativa de entender como a
fase de iniciação do câncer acontece, e qual seria a forma com que estas células
evoluem até os primeiros indícios de malignidade, dois modelos de desenvolvimento
tumoral foram sugeridos: o primeiro, chamado de modelo estocástico ou modelo de
expansão clonal, diz que a origem do câncer seria a partir de mutações nas próprias
células somáticas, e o consequente descontrole na proliferação permitiria a
aquisição de clones que sofreriam transformações que levariam à malignidade; e o
segundo, o modelo hierárquico, onde o desenvolvimento de um tumor se daria
através da propagação de células menos diferenciadas, como as células-tronco, que
teriam a capacidade de se diferenciar em células neoplásicas mais especializadas,
bem como se utilizaria de mecanismos para manter este potencial sempre ativo,
evitando a senescência e escapando da morte celular (Figura 5). Há ainda modelos
recentes que englobam ambos tipos de desenvolvimento, onde novas mutações
somáticas poderiam gerar diversidade clonal, aumentando ainda mais a
heterogeneidade tumoral (GREAVES; MALEY, 2012; BECK; BLANPAIN, 2013)
2.3.1 Modelo estocástico de desenvolvimento neoplásico
O modelo de desenvolvimento estocástico (ou de expansão clonal) foi
proposto por Peter Nowell em 1976, e sugere uma teoria para a evolução das
populações de células tumorais, a qual teria como principal característica uma
variação genética por etapas, podendo ser válido para a maioria das neoplasias de
mamíferos. Assim, uma célula, ou um pequeno grupo de células normais sofreriam

30
uma transformação pela ação de um agente carcinogênico, promovendo uma
proliferação desregulada, e as células-filhas geradas acabariam por obter alguma
vantagem sobre as demais células adjacentes, dando origem a uma expansão clonal
(Figura 5). A intensa multiplicação causaria uma instabilidade genômica celular,
permitindo uma série de mutações e reestruturações nos cromossomos. Este
acúmulo de mutações, então, daria origem a um novo fenótipo de células, mais
resistente e mais apto à sobrevivência, enquanto que, aquelas que não adquiriram
estas características, morreriam pela ação da imunidade própria do organismo
hospedeiro ou alterações metabólicas que desfavoreceriam a sobrevivência das
mesmas (NOWELL, 1976). O modelo estocástico então, se baseia em etapas de
transformações celulares, onde em cada fase se tem uma população dominante
sobre as outras. Este conjunto de ideias se assemelha ao conceito de seleção
natural descrito por Charles Darwin, em que o sistema de evolução é a variabilidade
genética ao acaso dos indivíduos unidos por um ancestral comum, juntamente com
a seleção natural dos mais aptos (GREAVES; MALEY, 2012).
Vários indícios podem dar suporte à teoria da expansão clonal. O primeiro
deles está relacionado ao tempo que um tumor leva até o aumento visível de
tamanho, sugerindo que as células tumorais passariam por um período de estase.
Em cada estágio do desenvolvimento neoplásico, segundo o modelo estocástico,
haveriam fatores, principalmente envolvidos com o microambiente tumoral
(arquitetura, recursos, fatores de crescimento, etc) ou mesmo fatores externos como
a quimio e radioterapia, que limitariam o crescimento de todas as células em geral,
conduzindo a uma competição inerente ao ambiente que seleciona apenas as mais
resistentes para darem continuidade à progressão. Assim, muitas, se não a maioria,
das células tumorais morreriam antes mesmo de concluir a multiplicação. Este fato é
evidenciado pelas grandes taxas de lesões pré-malígnas como carcinomas in situ,
fase de dormência do câncer, aumento lento e gradativo no tamanho macroscópico
da lesão, e reconstituição genética das semelhanças entre os clones (MALAISE;
CHAVAUDRA; TUBIANA, 1973; AGUIRRE-GHISO, 2007; REID et al., 2010).

31
Figura 5 - Modelo de desenvolvimento de tumores sólidos a partir da expansão clonal proposta por Nowell.
Fonte: adaptado de Greaves & Maley. Nature Reviews. v. 481. 2012 Legenda: subpopulações de células neoplásicas são selecionadas pelos fatores extrínsecos e intrínsecos a elas, as mais aptas sobrevivem, as outras morrem ou continuam dormentes. Os quadros em cinza representam os ecossistemas, enquanto que as linhas demonstram o momento da pressão seletiva.
Um exemplo prático seria a progressão que ocorre de forma comum em
neoplasias mamárias: uma neoplasia benigna, bem diferenciada, provavelmente
causada por um reduzido número de mutações, com o tempo começa a apresentar
um fenótipo mais maligno, como os carcinomas in situ, que apesar de menos
diferenciados ainda são de baixo grau invasivo; e por fim, as células tumorais
invadem tecidos adjacentes e vasos, causando a metástase - progressão de
hiperplasia glandular, para adenoma, carcinoma e invasividade. Mais do que simples
mutações e transformações genéticas a cada estágio da evolução clonal, as
populações celulares que estariam no controle do desenvolvimento tumoral estão
sujeitas a alterações fenotípicas desencadeadas por mudanças epigenéticas
(GREAVES; MALEY, 2012). Estas mudanças incluem alterações globais, como
PRESSÃO SELETIVA
Ecossistema 1 Ecossistema 2
Ecossistema 3
Metástase
Recidiva
Tratamento
Subclones – ‘driver mutation’

32
hipometilações do DNA e hipoacetilações da cromatina levando a uma instabilidade
genômica e a desregulação gênica do retinoblastoma e p16, por exemplo, bem
como mudanças mais específicas, assim como hipometilação e hipermetilação de
oncogenes e genes supressores tumorais, respectivamente (HODGSON et al., 2003;
FEINBERG; OHLSSON; HENIKOFF, 2006; JONES; BAYLIN, 2007).
Desde modo, propõe-se que a maioria dos neoplasmas surgem a partir de
uma única célula de origem, e os resultados de progressão tumoral e a variabilidade
genética adquirida dentro do clone original permite a seleção sequencial de
subclones mais agressivos. Populações de células tumorais são, aparentemente,
geneticamente mais instáveis do que as células normais, talvez pela ativação de um
loci específico na neoplasia, pela presença contínua do carcinógeno, ou mesmo
deficiências de recursos intratumoral.
2.3.2 Modelo hierárquico ou teoria das Células-tronco Cancerosas
Por outro lado, o modelo hierárquico sustenta a hipótese que o câncer se
desenvolve a partir de uma única subpopulação presente na massa tumoral. A teoria
das células-tronco cancerosas (CSCs, do inglês Cancer Stem Cells) propõe um
grupo de células contidas nos tumores sólidos as quais compartilham de algumas
similaridades com as células-tronco encontradas nos tecidos embrionários e tecidos
adultos saudáveis, como por exemplo, o potencial de auto-renovação, e a
capacidade de diferenciação em células mais especializadas, contribuindo para a
manutenção da neoplasia (Figura 6) (BECK; BLANPAIN, 2013). É preciso esclarecer
que o termo “célula-tronco cancerosa”, ainda gera confusão, uma vez que muitos
interpretam o termo no sentido de que tais células derivam das células-tronco do
tecido correspondente. As células-tronco no câncer podem, de fato, surgirem a partir
de células-tronco normais através de mutações em genes que tornariam as células-
tronco cancerosas, mas isto pode não ser o caso em todos os tumores. Por isso,
adotou-se também a expressão Células Iniciadoras de Tumor (CITs), que seriam
populações celulares capazes de dar início a uma nova neoplasia, mas não é
necessariamente uma célula-tronco.

33
Figura 6 - Modelos de desenvolvimento neoplásico.
Fonte: adaptado de Dick. Stem cell concepts renew cancer research. Blood, v. 112. 2008 Legenda: O tumor é composto por grupos distintos de células, biologicamente e funcionalmente. No modelo estocástico, as populações mais resistentes sobreviveriam à pressão seletiva, mas células de fenótipos variados poderiam dar origem a um novo tumor. Ao contrário, no modelo hierárquico, apenas um grupo de células teria a capacidade de se auto-renovar e dar origem aos demais grupos, e esta população poderia ser selecionada através de um fenótipo único.
O primeiro relato de isolamento destas células foi realizado na década de 90
em casos de leucemia mielóide aguda, onde Dominique Bonnet e John Dick
identificaram células-tronco normais e cancerosas, e caracterizaram ambos os
grupos quanto ao fenótipo e à capacidade de cada um deles no crescimento
neoplásico em camundongos imunodeficientes (BONNET; DICK, 1997). Neste
trabalho, os autores notaram que as leucemias mielóides poderiam responder ao
modelo hierárquico, onde uma única subpopulação celular, no caso as células-
tronco leucêmicas, se encontra no topo desta hierarquia. Quando transplantadas, as
células-tronco tumorais seriam biologicamente distintas, e se manteriam por si só,
gerando clones que se diferenciam ou geram progênies que irão adquirir fenótipo e

34
propriedades parecidas com as de células-tronco normais levando a progressão do
tumor (DICK, 2008).
A origem das CSCs depende da localização em que irá desenvolver o tumor
inicial. Assim como em tecidos normais, as CSCs estão dispostas em nichos, que
são localizações anatômicas específicas responsáveis pela proteção, manutenção
dentro do microambiente tumoral e sustentação das condições ideais para que elas
mantenham suas características (LI; XIE, 2005; SCADDEN, 2006). Acredita-se
também que as CSCs seriam as responsáveis não só por conduzir o crescimento do
tumor, mas também por sua capacidade superior de invasão, através de um
mecanismo já definido chamado de transição epitélio-mesenquimal (EMT), além de
proporcionar uma maior resistência contra a quimioterapia (HUNTLY; GILLILAND,
2005; FILLMORE; KUPERWASSER, 2008; VISVADER; LINDEMAN, 2008).
As células tumorais associam-se a mecanismos fisiológicos para se proteger
de agentes tóxicos, o que corresponde a uma significativa barreira na realização de
um tratamento quimioterápico efetivo (GOTTESMAN, 2002). Este mecanismo é
mediado principalmente por uma família de proteínas transportadoras ATP-
dependentes conhecida como transportadores ABC (ATP-binding Cassette), que
formam um sistema de efluxo de substâncias altamente eficientes, conhecido como
mecanismo de resistência a múltiplas drogas, ou MDR – do inglês Multi-drug
Resistance. O avanço da biologia molecular levou à identificação dos principais
genes MDRs, através da suspeita de que esta resistência estaria associada a
produtos de genes capazes de alterar a membrana plasmática (RONINSON et al.,
1984). E descobriu-se que os principais genes, e por consequência, as principais
proteínas responsáveis pelo efluxo de substâncias quimioterápicas para o meio
extracelular eram a ABCB1 e ABCG2, membros de uma família de 48 proteínas
conhecidas, responsáveis por uma série de processos essenciais para a
sobrevivência da célula tumoral (TWENTYMAN; FOX; BLEEHEN, 1986; DOYLE et
al., 1998; DONNENBERG; DONNENBERG, 2005).
Tanto o ABCB1 quanto o ABCG2 também são expressos em células-tronco,
não só auxiliando no efluxo de agentes tóxicos, mas também protegendo o tecido
contra a morte celular independentemente da administração de qualquer tipo de
agente externo (SIMS-MOURTADA et al., 2007). O conhecimento do papel central

35
de que os transportadores MDR atuam na proteção das células-tronco normais nos
permite refinar a hipótese das células-tronco cancerosas, e adicionar novos pontos
de vista que podem ser relevantes para explicar o fracasso dos tratamentos
quimioterápicos, recidivas tardias, metástases, e as diferentes respostas que os
tipos histológicos apresentam na apresentação geral do câncer. Tal conhecimento
também é de grande valia para a projeção de terapias inovadoras, que levem em
conta as semelhanças e diferenças entre o câncer e as células-tronco normais
(DONNENBERG; DONNENBERG, 2005).
Do mesmo modo, a transição epitélio-mesenquimal é fundamental para o
sucesso das neoplasias invasivas e malignas. Consiste na remodelação do
citoesqueleto celular, passando de uma estrutura epitelial e estável para um fenótipo
mesenquimal e de alta motilidade, capaz de passar por entre a matriz extracelular e
chegar até os vasos sanguíneos e linfáticos. Uma vez nos vasos, as células
invasoras alcançam um sítio metastático, onde novamente ultrapassam a barreira de
células endoteliais, e se instalam no tecido-alvo, retornando à forma epitelial e dando
origem a um tumor secundário (Figura 7). Entre todos os eventos de EMT
associados com metástases, um dos mais importantes é a perda de proteínas
responsáveis pela adesão célula-célula, como por exemplo, a E-caderina e
citoqueratina, e o aumento da expressão de vimentina e N-caderina. A E-caderina e
N-caderina são proteínas que desempenham um papel importante no
estabelecimento e manutenção da arquitetura celular, polaridade celular e
diferenciação (VAN ROY; BERX, 2008), enquanto que a citoqueratina, assim como a
vimentina, compõe um dos filamentos intermediários estruturais que mantêm a
morfologia da célula (BERNAL; STAHEL; BAYLIN, 1985). Nos seres humanos,
estudos já demonstraram que a perda de E-caderina e citoqueratina desencadeia
uma disseminação metastática em diversos tipos de tumores, bem como um
aumento do crescimento, invasão e resistência a drogas de células de câncer
colorretal com metástases hepáticas associadas (ONDER et al., 2008). Estudos
recentes também indicam que a EMT está fortemente associada à MDR (JIANG et
al., 2015), portanto, o aumento da motilidade celular permitiria a resistência a
agentes citotóxicos, senescência, bem como o escape do sistema imunológico e
fármacos quimioterápicos, desenvolvendo e induzindo metástases (KAJIYAMA et al.,
2007; ZHANG; MA, 2012).

36
Figura 7 - Transição epitélio-mesenquimal (EMT) e transição mesenquima-epitelial (MET) para formação de tumor secundário
Fonte: adaptado de Thompson & Haviv.The social aspects of EMT-MET plasticity. Nature Medicine, v 11. 2011.
Legenda: transição epitélio-mesenquimal (EMT) seguida de extravasamento e transição mesenquima-epitelial (MET) para formação de tumor secundário em tecido distante
Embora as células-tronco dos vários tecidos vivos apresentem propriedades
fundamentais de auto-renovação e diferenciação, o local de origem pode influenciar
a atividade de cada uma (CLARKE et al., 2006). Por isso, marcadores celulares que
são úteis na caracterização de células iniciadoras em um determinado tipo de tumor,
podem não ter a mesma eficiência em outro. Além disso, poucos marcadores que
são utilizados para o isolamento de células-tronco cancerosas são expressos
exclusivamente por elas. Por exemplo, o marcador de membrana CD133+ é
amplamente usado para isolar populações enriquecidas com CSCs em tumores
cerebrais, hepáticos, prostáticos, pancreáticos e colorretais (SINGH et al., 2003;
COLLINS et al., 2005; SUETSUGU et al., 2006; HERMANN et al., 2007; O’BRIEN et
al., 2007), mas também está presente no cérebro saudável e em células
diferenciadas de diversos tecidos do corpo. Também, já foi verificado que tanto

37
populações CD133+ quanto CD133- foram capazes de gerar tumores em
camundongos imunodeficientes (SHMELKOV et al., 2008).
O mesmo acontece com marcadores comumente utilizados no câncer de
mama, como o CD44 e o CD24. O primeiro é uma glicoproteína transmembrana
expressa em diversos tipos celulares, e está envolvido nas interações célula-célula
bem como em uma série de interações com a matriz extracelular em diversos
tecidos. O segundo, também é um marcador de membrana expresso em neutrófilos,
pré-linfócitos B e em alguns tipos de células tumorais (FOGEL et. al., 1999). O
fenótipo mais utilizado para o isolamento de populações enriquecidas com CSCs em
neoplasias mamárias corresponde ao grupo CD24-/low/CD44+, mas alguns trabalhos
já demonstraram que apenas estes dois marcadores podem não ser suficientes para
a separação desta população (AL-HAJJ et al., 2003; FILLMORE; KUPERWASSER,
2008). Além disso, em tumores altamente agressivos, como é o caso do
comedocarcinoma mamário, o fenótipo CD24+/CD44+ também foi caracterizado
como tumorigênico (AL-HAJJ; CLARKE, 2004). Isso mostra a dificuldade na
caracterização de uma molécula que seja expressa de forma semelhante e possa
ser utilizada fielmente em todos os tipos de neoplasias. Então, considerando que as
CSCs seriam as únicas células capazes de metastatizar e colonizar tecidos
distantes, e que, também são as células que apresentam uma maior resistência a
fármacos e um alto metabolismo detoxificante, o desenvolvimento de terapias-alvo
de inibidores de EMT pode proporcionar novas estratégias para a prevenção,
diagnóstico e tratamento de câncer.
2.4 CÉLULAS-TRONCO CANCEROSAS NO CÂNCER DE MAMA
Uma grande variedade de células compõe o tecido glandular mamário, como
as células epiteliais, estromais, hematopoiéticas e endoteliais. Também, a mama
representa um importante nicho de células-tronco sadias, que estão presentes em
um número constante (DONTU et al., 2003). Além do potencial de auto-renovação,
elas têm a capacidade de gerar os três grupos celulares que compõe a estrutura
lóbulo-alveolar de um tecido mamário adulto: células mioepiteliais que formam o

38
estrato basal dos ductos e alvéolos; células epiteliais ductais, que limitam o lúmen
dos ductos; e células epiteliais alveolares que sintetizam as proteínas do leite (LI;
XIE, 2005). Apresentam-se quiescentes no tecido a menos que haja algum estímulo,
como por exemplo, a chegada da puberdade ou a ocorrência da gravidez. Ao
contrário dessas alterações, que são breves e pontuais, o ciclo reprodutivo feminino
está presente em boa parte da vida destes indivíduos, expondo a mulher a uma
constante taxa de hormônios que por sua vez possuem uma grande influência na
glândula mamária, alterando a homeostasia das células-tronco mamárias (MaSC) e
consequentemente aumentando a predisposição a incidência de neoplasias
malignas (SHACKLETON et al., 2006).
Em 2003, baseando-se nos métodos utilizados nos estudos envolvendo
leucemias murinas, o grupo de Michael Clarke demonstrou a existência de uma
população celular tumorigênica peculiar no câncer de mama, que apresentaria uma
marcação similar às células-tronco teciduais normais (fenótipo CD44+/CD24low/-/Lin-).
Esta subpopulação exibia propriedades semelhantes às células-tronco encontradas
nos tecidos saudáveis: teriam potencial de auto-renovação, levariam à
tumorigenicidade, recidivas e metástases, além de apresentarem a capacidade de
se diferenciar em outros tecidos, embora aberrantes, dando origem à heterogênea
população de células tumorais (AL-HAJJ et al., 2003). Outros trabalhos alcançaram
resultados semelhantes desde então, se utilizando de diversas ferramentas e
marcadores para separação das populações (Quadro 3), mostrando que a grande
parte das células que constituem a massa tumoral, não seriam tumorigênicas devido
a falta da capacidade de auto-renovação e limitado potencial de proliferação
(Quadro 4) (PONTI et al., 2005; FILLMORE; KUPERWASSER, 2008). Além de
marcadores de membrana, outros métodos descrevem potenciais formas de
isolamento das populações de CSCs. A ALDH por exemplo, é uma enzima
responsável pela conversão de aldeídos em ácidos carboxílicos. É necessária na
produção de ácidos retinóicos e pode desempenhar um papel na regulação da
diferenciação de células-tronco hematopoiéticas, neurais e mamárias e, em seres
humanos, o elevado nível de atividade de ALDH é uma característica comum de
CSCs em tumores de mama, pulmão e pâncreas (CHUTE et al., 2006; GINESTIER
et al., 2007; JIANG et al., 2009; RASHEED et al., 2010).

39
Quadro 3 - Marcadores de células-tronco cancerosas em tumores sólidos
ABCG2 Membro da família ATP, envolvida no transporte de esteróides e outros lipídeos. ABCG2 (também conhecida como proteína de resistência do câncer de mama) é um transportador de múltiplas drogas (ver abaixo Hoechst SP). ABCG5 confere resistência à doxorrubicina.
ALDH1 A aldeído desidrogenase (ALDH) de família de enzimas catalisam a oxidação de aldeídos alifáticos e aromáticos de ácidos carboxílicos. ALDH1 tem um papel na conversão de retinol para o ácido retinóico, que é importante para a proliferação, diferenciação e sobrevivência.
CD24 (HSA) Molécula de adesão glicosilfosfatidilinositol pesadamente glicosilada, que tem um papel co-estimulador nas células B e T. O único ligante conhecido para a P-selectina. Embora não seja um marcador específico de células-tronco cancerosas, níveis baixos podem caracterizar células iniciadoras de tumor na mama
CD44 (PGP1) Molécula de adesão com múltiplas isoformas que tem papéis pleiotrópicos em sinalização, migração e homing. A forma CD44H padrão exibe uma elevada afinidade para o hialuronato; CD44V confere propriedades metastáticas.
CD133 (prominin 1) Glicoproteína de domínio transmembranar com um papel na organização da topologia da membrana plasmática. Expressa em células-tronco e progenitoras CD34+ no fígado fetal, precursores endoteliais, células-tronco neuronais fetais e do desenvolvimento de epitélio, CD133 foi detectada pelo epítopo glicosilado na maioria dos estudos. Assim, pode ser um marcador de células-tronco cancerosas mais confiável
EpCAM (molécula de adesão das células epiteliais; ESA, TROP1) Molécula de adesão celular homofílica Ca2+ independente expressa nas superfícies basolaterais da maioria das células epiteliais
Side Population (SP) Fenótipo devido ao efluxo de Hoechst33342 por bombas presentes na membrana plasmática em diferentes tipos de células. Atividade é conferida pelo transportador ABCG2. Inibida por verapamil. Fonte: adaptado de Visvader & Lindeman. Nature Review, v 8. 2008
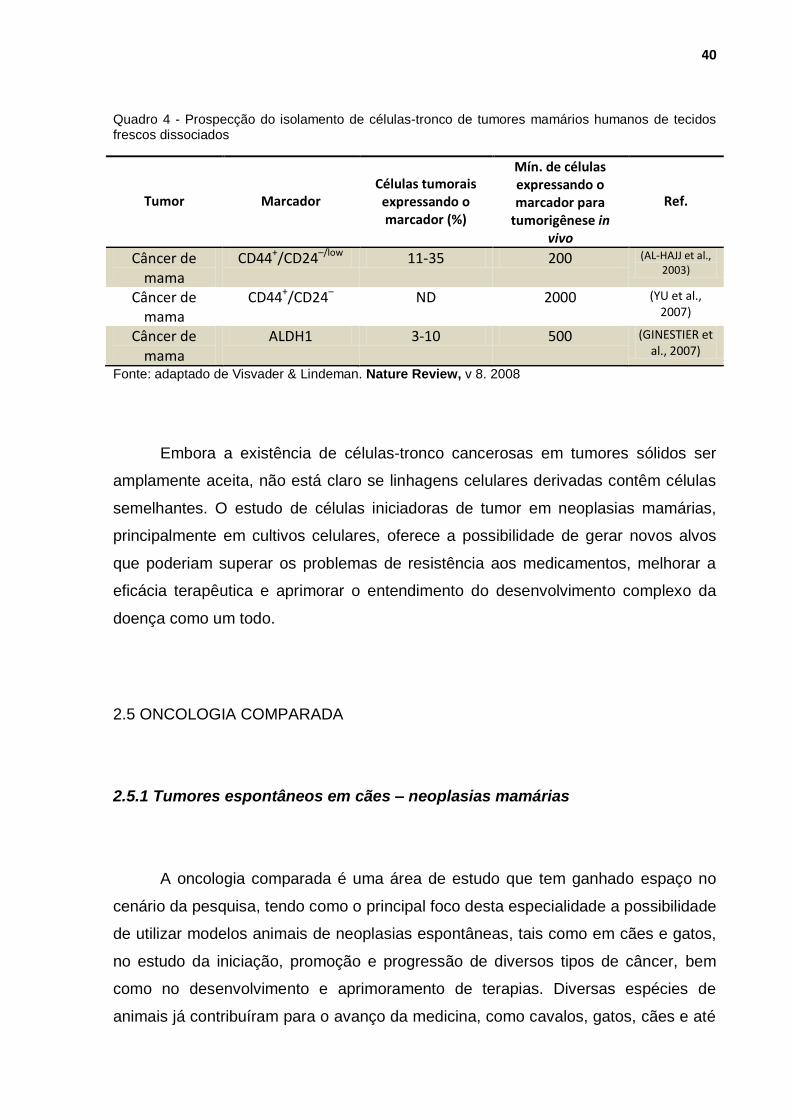
40
Quadro 4 - Prospecção do isolamento de células-tronco de tumores mamários humanos de tecidos frescos dissociados
Tumor
Marcador
Células tumorais
expressando o marcador (%)
Mín. de células expressando o marcador para
tumorigênese in vivo
Ref.
Câncer de mama
CD44+/CD24–/low 11-35 200 (AL-HAJJ et al., 2003)
Câncer de mama
CD44+/CD24– ND 2000 (YU et al., 2007)
Câncer de mama
ALDH1 3-10 500 (GINESTIER et al., 2007)
Fonte: adaptado de Visvader & Lindeman. Nature Review, v 8. 2008
Embora a existência de células-tronco cancerosas em tumores sólidos ser
amplamente aceita, não está claro se linhagens celulares derivadas contêm células
semelhantes. O estudo de células iniciadoras de tumor em neoplasias mamárias,
principalmente em cultivos celulares, oferece a possibilidade de gerar novos alvos
que poderiam superar os problemas de resistência aos medicamentos, melhorar a
eficácia terapêutica e aprimorar o entendimento do desenvolvimento complexo da
doença como um todo.
2.5 ONCOLOGIA COMPARADA
2.5.1 Tumores espontâneos em cães – neoplasias mamárias
A oncologia comparada é uma área de estudo que tem ganhado espaço no
cenário da pesquisa, tendo como o principal foco desta especialidade a possibilidade
de utilizar modelos animais de neoplasias espontâneas, tais como em cães e gatos,
no estudo da iniciação, promoção e progressão de diversos tipos de câncer, bem
como no desenvolvimento e aprimoramento de terapias. Diversas espécies de
animais já contribuíram para o avanço da medicina, como cavalos, gatos, cães e até

41
animais menos comuns, como os ferrets (KLEIN et al., 1986; ANTINOFF; HAHN,
2004; SELTENHAMMER et al., 2004; HERSHEY; DUBIELZIG, 2005; SMITH et al.,
2014).
O uso de animais de laboratório para identificar o potencial carcinogênico de
diversos produtos químicos e outros agentes ligados à iniciação e desenvolvimento
de neoplasias teve início há muitas décadas. Dentre as principais vantagens de se
ter camundongos, ratos e hamsters como principais meios de estudo in vivo em
oncologia, estão: semelhanças com os humanos nas respostas a alguns agentes
carcinogênicos em diversos aspectos; menores custos de manutenção do biotério,
visto que um pequeno espaço físico pode alojar um grande número de animais,
aumentando assim a amostragem de cada estudo; o controle do ambiente em que
eles vivem, diminuindo a interferência de fatores externos nos resultados de uma
avaliação; a possibilidade da criação de linhagens isogênicas, manipulando
geneticamente o alvo do estudo para determinadas características biológicas e
moleculares, e também diminuindo a variabilidade genética entre indivíduos,
minimizando ou até mesmo excluindo os riscos de efeitos não desejáveis em uma
determinada experimentação; e o limitado ciclo de vida destes animais, ideal para
estudos que exigem resultados rápidos e onde a idade não é um fator determinante,
ou muito determinante, nas características e na eficácia do experimento (FRESE;
TUVESON, 2007).
Mas apesar disso, muito ainda se discute sobre a extrapolação dos dados
obtidos em roedores para o que acontece com os humanos. Há um crescente grupo
de evidências experimentais indicando diferenças importantes (genéticas,
metabólicas, ontogenéticas, entre outras) entre as espécies de mamíferos na
maneira em que o câncer se desenvolve (ANISIMOV; UKRAINTSEVA; YASHIN,
2005). As células murinas requerem menos alterações genéticas e/ou epigenéticas
para que haja um descontrole na proliferação que resulte em neoplasias. Então, por
necessidade, a progressão tumoral, que em humanos necessita de várias etapas,
em roedores acaba sendo muito mais simples, onde estas diferenças na
organização das vias regulatórias celulares ditam fortemente diferentes cursos de
um tumor (RANGARAJAN; WEINBERG, 2003). Este maior número de vias
necessárias para a transformação das células humanas, ou mecanismos de
proteção contra a transformação, provavelmente deve ter sido desenvolvido, ou

42
aperfeiçoado, durante a evolução das espécies. Mas vale lembrar que, apesar de
todas as divergências entre iniciação e progressão tumoral, não se pode diminuir a
importância da modelagem em roedores para o estudo do câncer em seres
humanos; no entanto, os resultados exigem uma interpretação mais cuidadosa antes
de confirmar qualquer hipótese.
O cão recebe um destaque especial quando o assunto é a oncologia
comparada. Sabe-se que são próximos geneticamente dos seres humanos, fato
cientificamente comprovado pela publicação do genoma canino, em 2005
(LINDBLAD-TOH et al., 2005), e ocupa um espaço importante na genômica atual. A
posição do cão dentro da árvore evolutiva dos mamíferos faz com que o animal se
torne um guia importante para a análise comparativa do genoma humano, gerando
uma grande oportunidade de explorar os efeitos da genética quanto à suscetibilidade
a doenças, incluindo o câncer, como demonstrado no quadro 5 (LINDBLAD-TOH et
al., 2005). Um exemplo da evolução e organização da oncologia comparada foi a
criação do Canine Comparative Oncology e Genomics Consortium. O grupo foi
lançado por pesquisadores clínicos e científicos para proporcionar um ambiente de
discussão sobre o desenvolvimento de novas tecnologias, que permitiria o estudo
adequado dos casos de câncer canino na biologia e na terapia do câncer humano
(KHANNA et al., 2006).
Quadro 5 - Principais características relacionadas ao estudo da espécie Canis familiaris na oncologia comparada.
(Continua)
Cada vez mais os proprietários de cães se preocupam com os cuidados veterinários direcionados aos animais de estimação. Segundo a Associação Americana de Medicina Veterinária (AVMA – American Veterinary Medical Association), em 2007, o gasto em saúde com cães nos EUA foi de aproximadamente 20 bilhões de dólares. Em 2012, os números chegaram a 26 bilhões (AMERICAN VETERINARY MEDICAL ASSOCIATION, 2012). Nos últimos anos, há um interesse maior por parte dos proprietários na participação de seus animais em trials, estudos clínicos e laboratoriais, quando o sucesso do tratamento convencional não é alcançado, fortalecendo a pesquisa em oncologia veterinária comparada e translacional e dando uma nova chance ao animal de estimação.

43
(Conclusão)
• Raças de cães são semelhantes às populações humanas geograficamente isoladas, como os da
Finlândia, na Islândia, exceto que o isolamento é mais extremo. Isto oferece uma grande
vantagem na busca de genes associados a doenças complexas, que, em teoria, pode ser mais
facilmente mapeados utilizando famílias de cães do que famílias humanas (SUTTER; OSTRANDER,
2004).
• Um exemplo do sucesso do mapeamento genético do cão trazendo benefícios à saúde humana,
foi o descobrimento de uma mutação pontual causadora da RCND (sigla em inglês para
Cistoadenocarcinoma Renal e Dermatofibrose Nodular), um câncer hereditário bem definido em
Pastores Alemães, e com forte semelhança a uma desordem multissistêmica em humanos,
chamada BHD. O grupo de Lingaas et. al. constatou que o mesmo gene pode ser responsável
tanto pela doença canina quanto humana, provendo agora um modelo animal de ocorrência
natural para o estudo de uma desordem humana rara (LINGAAS et al., 2003).
• Os cães apresentam uma vida útil mais curta que os humanos, porém bem mais longa que os
roedores, sendo que a intervenção clínica pode ser estudada ao longo de um período
condensado. As taxas de sobrevida em cães são avaliadas em um ano, ao invés de cinco como na
oncologia humana. Com isso é possível obter resultados relativamente rápidos na realização de
ensaios clínicos e no monitoramento da progressão da doença.
• O estudo epidemiológico dos fatores de risco relacionados a doenças complexas humanas, como
o câncer, pode também ser acessado através do estudo com cães, uma vez que eles
compartilham do mesmo ambiente que seus proprietários e estão expostos a boa parte dos
mesmos carcinógenos (KIMURA et al., 2013; SCHALIE et al., 1999). Também, apresentam os
mesmos fatores de risco do desenvolvimento de neoplasias, por exemplo, as mamárias, que
estão associadas a fatores hormonais, obesidade, dieta, entre outros (LINDBLAD-TOH et al.,
2005).
• Semelhanças entre as características clínicas e bases moleculares análogas às doenças humanas
e caninas, permitem um teste eficaz de novos tratamentos e aprimoramento dos já existentes
para doenças humanas, em cães.
Legenda: com o avanço das pesquisas, muitas vantagens são adicionadas nesta área, aumentando os benefícios gerados na saúde humana através dos resultados obtidos principalmente pela genômica e transcriptômica do câncer.
Assim como em seres humanos, os cães desenvolvem tumores de forma
espontânea e imprevisível, ganhando destaque na oncologia comparada não só
pelas semelhanças anatomofisiológicas com os seres humanos, mas também pelas
altas taxas de incidência de câncer nesta espécie: nos Estados Unidos, são
estimados mais de um milhão de casos de neoplasias em cães de companhia por
ano (PAOLONI; KHANNA, 2008). Dentre os tipos de neoplasia, os tumores
mamários são a segunda maior causa de morte, ficando atrás apenas das
neoplasias de pele. Em 2012, Gupta e colaboradores compilaram diversas
publicações, de 1965 a 2010, em um estudo epidemiológico de neoplasias de mama
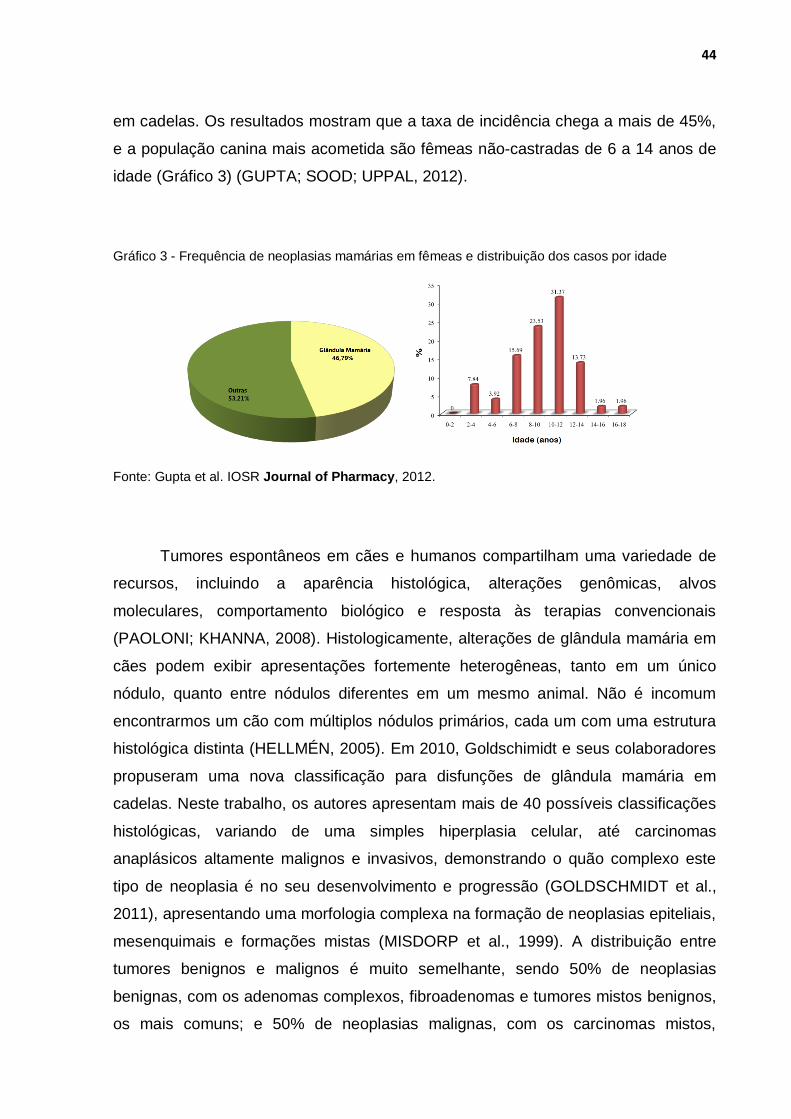
44
em cadelas. Os resultados mostram que a taxa de incidência chega a mais de 45%,
e a população canina mais acometida são fêmeas não-castradas de 6 a 14 anos de
idade (Gráfico 3) (GUPTA; SOOD; UPPAL, 2012).
Gráfico 3 - Frequência de neoplasias mamárias em fêmeas e distribuição dos casos por idade
Fonte: Gupta et al. IOSR Journal of Pharmacy, 2012.
Tumores espontâneos em cães e humanos compartilham uma variedade de
recursos, incluindo a aparência histológica, alterações genômicas, alvos
moleculares, comportamento biológico e resposta às terapias convencionais
(PAOLONI; KHANNA, 2008). Histologicamente, alterações de glândula mamária em
cães podem exibir apresentações fortemente heterogêneas, tanto em um único
nódulo, quanto entre nódulos diferentes em um mesmo animal. Não é incomum
encontrarmos um cão com múltiplos nódulos primários, cada um com uma estrutura
histológica distinta (HELLMÉN, 2005). Em 2010, Goldschimidt e seus colaboradores
propuseram uma nova classificação para disfunções de glândula mamária em
cadelas. Neste trabalho, os autores apresentam mais de 40 possíveis classificações
histológicas, variando de uma simples hiperplasia celular, até carcinomas
anaplásicos altamente malignos e invasivos, demonstrando o quão complexo este
tipo de neoplasia é no seu desenvolvimento e progressão (GOLDSCHMIDT et al.,
2011), apresentando uma morfologia complexa na formação de neoplasias epiteliais,
mesenquimais e formações mistas (MISDORP et al., 1999). A distribuição entre
tumores benignos e malignos é muito semelhante, sendo 50% de neoplasias
benignas, com os adenomas complexos, fibroadenomas e tumores mistos benignos,
os mais comuns; e 50% de neoplasias malignas, com os carcinomas mistos,

45
carcinomas complexos e carcinomas anaplásicos no topo da lista dos mais
incidentes (HELLMÉN, 2005). A maioria dos tumores mamários mistos são
caracterizados por uma mistura complexa de células epiteliais e mioepiteliais, com
presença ocasional de metaplasias cartilaginosas, ósseas, ou metaplasias
escamosas, e a análise histológica dessas neoplasias geralmente deve incluir
também um componente de células fusiformes (CASSALI et al., 2007).
Características como tamanho dos nódulos, tipo histológico, graduação
histológica e análise de linfonodos são reconhecidos como fatores prognósticos
confiáveis (MISDORP et al., 1999; KARAYANNOPOULOU et al., 2005). Mas, apesar
das neoplasias mamárias malignas representarem metade dos casos
diagnosticados, poucos marcadores prognósticos a nível molecular foram
caracterizados de forma eficiente e adicionados na rotina clínica e cirúrgica em
medicina veterinária, ao contrário do que acontece na oncologia humana. A
identificação de fatores prognósticos de confiança é essencial, a fim de estimar o
risco individual de uma evolução clínica desfavorável. Na literatura, encontramos um
número crescente de investigações em busca de marcadores adequados para o
câncer de mama canino, incluindo receptores hormonais, marcadores de
proliferação, receptor de fator de crescimento epidérmico humano (HER2), p53,
entre outros (MILL et al., 2003 LEE et al., 2004; ART, 2005). Em um trabalho de
avaliação imunohistoquímica em 102 casos de neoplasias malignas, os autores
encontraram fenótipos moleculares distintos, assim como em humanos. Foi
identificado um subtipo de carcinoma basal-like, que representou cerca de 30% dos
casos, significantemente associado com um comportamento clínico mais agressivo
(GAMA; ALVES; SCHMITT, 2008). No entanto, a experiência clínica ainda é muito
limitada, e resultados fidedignos em estudos prospectivos nem sempre estão
disponíveis.
Como comentado anteriormente, não é só na análise histopatológica que a
oncologia comparada se baseia. Estudos moleculares, principalmente ligados à
genômica, transcriptômica e proteômica, também têm sido de grande valia na
pesquisa básica em câncer de mama entre cães e humanos. Um estudo realizado
por Uva e colegas (2009), fez uma análise comparativa da expressão gênica de vias
moleculares em tecidos saudáveis e neoplásicos, de caninos e humanos, através da
técnica de microarranjo. Os resultados mostram que um número significativo de

46
genes diferencialmente expressos em neoplasias de glândula mamária em
humanos, em comparação com amostras de humanos normais, também foi
encontrado desregulado no modelo canino, incluindo as vias PI3K/AKT, RAS, PTEN,
WNT-β catenina e também a cascata MAPK (UVA et al., 2009). Em outro estudo
com carcinomas caninos, pesquisadores identificaram 21 proteínas expressas de
maneira diferente em neoplasias metastáticas e não-metastáticas. A maioria,
envolvidas em funções celulares relevantes para a propagação metastática como a
adesão celular, a modulação da matriz extracelular e a resistência à hipóxia.
Curiosamente, 19 entre as 21 proteínas já haviam sido descritas anteriormente na
medicina, sendo a expressão associada à malignidade do câncer de mama e outros
tipos de câncer humanos (KLOPFLEISCH et al., 2010).
Mas assim como em roedores, deve-se ter cuidado na interpretação dos
dados obtidos em modelos caninos quando da extrapolação para o que acontece no
câncer humano. Mesmo com diversos estudos mostrando que o cão pode ser útil
para o entendimento de neoplasias humanas, ainda pouco se sabe sobre de que
forma os tumores em cães se desenvolvem, e qual o comportamento das vias
regulatórias que regem a progressão tumoral na espécie. O entendimento das bases
biológicas e moleculares da evolução neoplásica em cães deve ser prioridade antes
da aplicação na medicina comparada e translacional.
2.5.2 Células-tronco cancerosas em neoplasias mamárias em cães
Há um grande interesse dos pesquisadores do isolamento e caracterização
de CSCs em neoplasias espontâneas de cães e gatos, por razões já comentadas
anteriormente. Em cães, são poucos os trabalhos que descrevem informações
referentes às CSCs em tumores mamários, e todos até o momento trabalharam com
cultivos de células. As linhagens celulares sem dúvidas fornecem um recurso valioso
para a investigação das características associadas, mesmo que transitoriamente, à
expressão de fenótipos. No entanto, é importante avaliar esses dados com
parcimônia, a fim de evitar uma interpretação errônea, inferindo uma característica

47
de “células-tronco tumorais” onde razões biológicas mais simples poderiam explicar
as observações.
O primeiro relato de isolamento de MaSC (Mammary Stem Cell, ou Célula-
tronco mamária) e CSCs em tumores de glândula mamária em cão foi realizado por
Cocola et al., que investigou a propagação de ambos os grupos, mamoesferas e
tumoresferas, durante cinco passagens em culturas de baixa aderência. Estas
células exibiram propriedades de auto-renovação, potencial de diferenciação em
múltiplas linhagens, formaram estruturas tubulares in vitro e tumores in vivo em
camundongos não-obesos e imunocomprometidos NOD/SCID (COCOLA et al.,
2009). A partir daí, alguns trabalhos já foram publicados, demonstrando que as
características mais eficientes para a separação de populações enriquecidas com
células que talvez correspondessem às CSCs, assim como em seres humanos,
poderiam estar ligadas principalmente à expressão dos fenótipos CD44, CD24,
CD49f, CD133, p21, CK14 (COCOLA et al., 2009; BLACKING; WATERFALL;
ARGYLE, 2011; MICHISHITA et al., 2011), e também à maior atividade de enzimas
responsáveis pelo metabolismo intracelular. Nesta, os principais métodos de
isolamento incluem a atividade intracelular de aldeído-desidrogenase (ALDH), e o
método Side Population (SP), que consiste na avaliação da célula quanto ao efluxo
de Hoechst 33342, ambos através da separação por citometria de fluxo.
Em cães, a alta expressão de aldeído-desidrogenase foi detectada em quatro
linhagens de carcinomas mamários caninos, e estas células se mostraram
enriquecidas pelo fenótipo CD44+/CD24-, mais uma vez apresentando a capacidade
de auto-renovação e tumorigenicidade em animais imunodeficientes (MICHISHITA et
al., 2012), indicando que a atividade de ALDH pode ser um marcador para CSCs em
neoplasias mamárias caninas. Já a técnica SP tem sido questionada não só devido
à variação nos resultados (SMALLEY; CLARKE, 2005), mas também por talvez
afetar a diferenciação celular (ADAMSKI et al., 2007). No entanto, afirma-se que o
perfil transcricional de células SP positivas em tumores de mama tem um
enriquecimento de expressão em genes da regulação do ciclo celular e pontos de
controle, transportadores MDRs, organogênese e vasculogênese (BEHBOD et al.,
2006), podendo ser uma análise importante no isolamento e caracterização das
populações superiores na cadeia hierárquica dos tumores sólidos.

48
Finalmente, no que diz respeito à investigação do câncer de mama e do papel
das células-tronco mamárias no desenvolvimento de neoplasias, o modelo de tumor
espontâneo canino ainda é um recurso subutilizado. No entanto, vários grupos de
pesquisa já foram capazes de isolar e caracterizar possíveis CSCs com sucesso
nesta espécie, usando protocolos semelhantes àqueles utilizados nos modelos
humanos e de roedores, indicando o potencial promissor dessas células na pesquisa
básica, bem como pesquisa oncológica translacional.

49
3 OBJETIVO GERAL
Caracterização de cultivos de células neoplásicas oriundas de carcinomas mamários
de cadelas e avaliação do potencial tumorigênico de populações celulares
selecionadas pelos marcadores ALDH, CD44 e CD24.

50
4 OBJETIVOS ESPECÍFICOS
Padronização e caracterização do cultivo de células neoplásicas de
carcinomas mamários de cadelas quanto à morfologia, proliferação celular,
citogenética e expressão de marcadores de células epiteliais e
mesenquimais;
Avaliação e quantificação da presença de populações CD24-, CD44+ e ALDHhi
em culturas primárias de células neoplásicas oriundas de carcinomas
mamários de cadelas;
Validação do potencial tumorigênico das populações acima descritas em
ensaios in vitro.
51
5 MATERIAL E MÉTODOS
5.1 MATERIAL UTILIZADO NO ESTUDO
Todas as amostras de tecidos neoplásicos foram coletadas a partir de
mastectomias unilaterais ou bilaterais realizadas no Hospital Veterinário da
Faculdade de Zootecnia e Engenharia de Alimentos/USP, bem como em clínicas
veterinárias particulares localizadas em São Paulo/SP, Pirassununga/SP e região,
que concordaram em colaborar com o presente projeto (Quadro 6). Ao todo, foram
coletadas amostras de tecido de trinta e cinco (35) cães, todas avaliadas e
classificadas pela análise histopatológica. Aquelas que tinham origem em glândula
mamária foram categorizadas seguindo a classificação de Goldschmidt
(GOLDSCHMIDT et al., 2011).
Quadro 6 - Clínicas parceiras que contribuíram com amostras para o presente estudo
CLÍNICA
MÉDICO VETERINÁRIO
CIDADE
Cia do bicho Alessandra Pirassununga/SP
CEPAC é o bicho Marcus Vazan Pirassununga/SP
Pet’s Stop Mariney Pirassununga/SP
- Juliana Casals Pirassununga/SP
Clínica Santa Rita Milena Adami Leme/SP
Hamvet Sílvio Ramos e Tânia Ávila Leme/SP
- Beatriz Martins São João da Boa Vista/SP
PetVip Bruno Monteiro São João da Boa Vista/SP
Animal Planet Paulo Côrte Neto Araras/SP
Central Nilson J. Bianchim Araras/SP
LinVet Valmir B. do Nascimento Araras/SP
- Mariana V. dos Santos Osasco/SP
- João Paulo Boccia São Paulo/SP

52
Logo após a excisão cirúrgica de cada amostra, foram removidas todas as áreas
de tecido adiposo, e possíveis áreas de necrose. O restante do material foi então
dividido em três fragmentos: o primeiro, colocado em formol tamponado a 10% para
avaliação histopatológica; o segundo, transportado em solução de RNA-later e
posteriormente conservado à temperatura de -80°C; e por fim, do terceiro fragmento
foi retirada toda a pele e tecido subcutâneo adjacente, sendo então lavado em PBS
a 5% de antibiótico e antimicótico (PenStep, Gibco®) e colocado em meio de
transporte estéril, composto por meio de cultivo DMEM-F12 (Life Technologies®) a
5% de antibiótico, à 4°C, até a chegada ao laboratório.
5.2 CULTIVO PRIMÁRIO DE CÉLULAS ORIUNDAS DE NEOPLASIAS MAMÁRIAS
O material coletado para o estabelecimento do cultivo primário foi picado
manualmente com o uso de tesoura, pinça e lâminas estéreis, em fragmentos de 1 a
2 mm3. Posteriormente, foram dissociados enzimaticamente em rotação constante
de 100 rpm por duas horas a 37°C, em meio DMEM-F12 suplementado com 200
U/mL de Colagenase tipo I (Sigma Aldrich®), 100 U/ml de Hialuronidase (Sigma
Aldrich®) e 2% de antibiótico, até que houvesse dissociação do tecido tumoral.
Depois, adicionou-se 1 mL de tripsina/EDTA, e o tubo foi levado ao shaker por mais
uma hora. O material foi passado primeiramente em um cell Strainer de 100 µm, e
logo depois filtrado em 40 µm a fim de remover grumos de células. O meio foi então
centrifugado a 350G durante 5 minutos, o sobrenadante descartado, e o pellet
ressuspendido em PBS previamente aquecido em banho-maria a 37ºC. As células
foram lavadas em PBS até que o pellet se apresentasse limpo, sendo ressuspendido
finalmente em meio de cultivo suplementado.
Após a etapa de dissociação do tecido neoplásico, as células foram avaliadas
quanto à viabilidade celular pelo método de exclusão por azul de Trypan, onde 10 μL
de meio contendo células foram adicionados a 10μL de azul de Trypan, e contados
em câmara de Neubauer. O meio contendo as células dissociadas foi levado ao
cultivo celular em garrafas estéreis de 25cm3 a 37ºC e saturação de 5% de CO₂, em
meio de cultivo DMEM-F12 suplementado com 1% de Mammary Epithelial Growth

53
Supplement (MEGS, Gibco®, Life Technologies), 5% de Soro Fetal Bovino e
adicionado de 1% de antibiótico/antimicótico.
A evolução foi acompanhada diariamente através de microscopia óptica.
Foram adicionados 2 mL de meio de cultivo novo em cada garrafa, a cada 48 horas,
para manutenção da cultura. Quando atingiam 70% de confluência, todas as
garrafas foram submetidas à atividade de Tripsina/EDTA diluída para 0,25%
(Gibco®, Life Technologies), e então subdivididas em novas garrafas estéreis para
multiplicação das colônias celulares. Brevemente, após o descarte do meio de
cultura adicionou-se 5 mL de PBS à 37ºC em cada garrafa, para remoção de debris
celulares e demais contaminantes que pudessem estar presentes à cultura. O PBS
foi prontamente descartado para a adição de 2 mL de solução Tripsina/EDTA 0,25%.
O material permaneceu em estufa à 37ºC até que todas as células fossem soltas das
garrafas, e formassem uma suspensão celular. Para inativação da enzima, usou-se
10 mL de meio de cultivo a 10% de Soro Fetal Bovino. A suspensão foi centrifugada
a 1200 rpm por cinco minutos, o sobrenadante descartado, e o pellet ressuspendido
em DMEM-F12 completo. O volume foi divido igualmente, e completado para 10 mL
por garrafa de 75cm3.
4.3 TEMPO DE DUPLICAÇÃO CELULAR
Para avaliação da velocidade de duplicação celular de cada amostra, foram
cultivadas 5 x 104 células por poço, em placa de 6 poços, com meio de cultivo
suplementado conforme descrito anteriormente. O número de células foi contado em
câmara de Neubauer em intervalos de 24 horas, por um período total de 120 horas.
Todas as amostras foram contadas no mesmo horário, e da mesma forma durante
todo o experimento. Os resultados foram avaliados através do programa Doubling-
Time (disponível em (14/01/2015): http://doubling-time.com/compute.php). O tempo
de duplicação foi realizado em dois momentos: o primeiro, antes da 10ª passagem, e
o segundo, quando todos os cultivos ultrapassaram a 20ª passagem.

54
5.4. AVALIAÇÃO CITOGENÉTICA POR CARIOTIPAGEM
A contagem do número de cromossomos foi realizada pelo método de
coloração de Giemsa. Brevemente, as células foram cultivadas em placas de Petri
durante o tempo necessário para alcançarem a fase M do ciclo celular, com base
nos resultados obtidos pelo tempo de duplicação. Decorrido o tempo da cultura
celular, adicionou-se 50 μL de solução de Colchicina (16mg/mL) para suspensão do
ciclo celular em metáfase, e a placa incubada em estufa a 37ºC durante 3 horas. O
meio de cultivo então foi coletado em um tubo Falcon de 15mL, e as células
aderidas passaram pelo processo de tripsinização para retirada do material de
cultivo, sendo colocadas juntamente com o meio armazenado na etapa anterior. O
tubo foi centrifugado a 1200 rpm durante 10 minutos, o sobrenadante descartado
cuidadosamente com uma pipeta, e o pellet ressuspendido em 5 mL de solução
hipotônica (KCl 0,075M) a 37ºC. O material foi incubado por 30 minutos a 37ºC,
sendo homogeneizado suavemente com uma pipeta em intervalos de 5 minutos.
Logo depois, adicionou-se 2 mL de fixador gelado (metanol/ácido acético 3:1) e a
solução homogeneizada novamente durante 1 minuto. O tubo foi levado a
centrifugação em 1200 rpm por 10 minutos, o sobrenadante descartado
cuidadosamente com uma pipeta, e o pellet ressuspendido em 0,5-1 mL de fixador
gelado, acertando a diluição dependendo do volume de células.
As lâminas foram lavadas e secas com papel macio para receber a
preparação, e permaneceram durante 24 horas imersas em etanol 70%, a -20ºC.
Para a confecção das lâminas, foram retiradas do etanol e secas novamente com
papel limpo e macio. Então, duas a quatro gotas da suspensão de células foram
dispensadas da pipeta a uma altura superior a 30 cm, para possibilitar o rompimento
das células e permitir o espalhamento dos cromossomos. As lâminas foram secas ao
ar, e então coradas com solução de Giemsa durante 10 minutos, em temperatura
ambiente. Depois deste processo, foram lavadas com água destilada, secas ao ar, e
montadas com lamínulas limpas e secas. Os cromossomos foram fotografados em
microscópio óptico, em imersão, no aumento de 1000x. Foram contadas vinte
metáfases por amostra.

55
5.5. IMUNOCITOQUÍMICA
As quatro amostras selecionadas para caracterização do cultivo primário
foram submetidas à técnica imunocitoquímica para avaliação da expressão dos
filamentos intermediários de citoesqueleto: citoqueratina, vimentina e α-actina de
músculo liso (detalhes descritos no quadro 7). Cerca de 105 células foram cultivadas
em lamínulas esterilizadas, em placas de 6 poços, até atingirem confluência
suficiente para a realização do experimento. Os poços foram lavados por três vezes,
com PBS, para então serem fixados através da adição de 2 mL de metanol absoluto
em cada poço, durante 10 minutos, em temperatura ambiente. As células foram
lavadas por mais duas vezes e armazenadas em PBS a 4ºC até o momento do
teste.
Quadro 7 - Anticorpos utilizados para avaliação imunocitoquímica
Anticorpo Diluição Incubação Clone Marca
Anti-citoqueratina 1:100 1 h A1/A3 Dako, Carpinteria, CA Anti-vimentina 1:100 1 h V9 Dako, Carpinteria, CA
Anti-α-sma 1:100 1 h HHF35 Dako, Carpinteria, CA
Para a marcação imunocitoquímica, as lamínulas foram lavadas previamente
por duas vezes de cinco minutos com PBS/Triton x100, para uma melhor distribuição
dos reagentes. Então, foi realizado o bloqueio da peroxidase endógena, através da
incubação com solução de peróxido de hidrogênio a 20% diluída em metanol
absoluto durante uma hora. As lamínulas foram lavadas com PBS/Triton por mais
três vezes de cinco minutos, e adicionadas de solução de leite em pó desnatado a
5%, e incubadas em estufa aquecida a 60ºC, durante uma hora, para bloqueio de
possíveis marcações inespecíficas. O anticorpo primário foi diluído em PBS/Triton
adicionado de BSA a 0,1% para todas as marcações realizadas, sendo preparados
50 μL de solução para cada lamínula cultivada. As células foram incubadas com o
anticorpo primário durante 1 hora em temperatura ambiente e então lavadas por

56
mais três vezes com PBS/Triton. Depois, utilizou-se o kit LSAB+ System-HRP (Dako,
Carpinteria, CA), sendo que o anticorpo secundário foi incubado durante 20 minutos
em temperatura ambiente, e a Streptavidina incubada por mais 20 minutos, também
em temperatura ambiente.
A marcação foi visualizada através da revelação por DAB, as lamínulas
posteriormente contracoradas com hematoxilina e montadas em lâminas para serem
observadas e fotografadas em microscópio óptico.
5.6 SEPARAÇÃO DE POPULAÇÕES CELULARES POR BEADS MAGNÉTICAS
ATRAVÉS DO PERFIL DE EXPRESSÃO DE CD24/CD44
Depois de estabilizado o cultivo celular em monocamada, ou seja, com todas
as colônias de células em condições de aderência e em proliferação, as amostras
oriundas da dissociação tumoral da glândula mamária foram submetidas à
tripsinização semelhante ao procedimento adotado para o repique convencional.
Após a centrifugação realizada posterior à inativação enzimática da Tripsina, a
suspensão celular foi avaliada quanto à viabilidade, e o número de células ajustado
para uma concentração aproximada de 2x106 células. O método escolhido para
separação das populações positivas e negativas quanto à expressão destas
proteínas foi a marcação com anticorpos biotinilados específicos para CD24 e CD44,
conjugação com as beads magnéticas (CELLection Kit, Life Technologies®) e
posterior isolamento da população positiva através do auxílio de atração magnética
promovida por um imã, conforme protocolo descrito a seguir.
Antes da realização do procedimento, se fez necessário a lavagem e
purificação das beads magnéticas antes de serem conjugadas com o anticorpo
desejado. Sendo assim, 25 μL da solução de beads foram adicionados a 1 mL de
PBS a 0,1% de Soro Fetal Bovino (consistindo do Buffer 1, CELLection Kit), e o tubo
colocado no imã durante 1 minuto. O sobrenadante foi descartado, e as beads então
estariam prontas para a realização das técnicas de separação. Tanto a técnica de
forma direta quanto a de forma indireta foram testadas para marcação de CD44:

57
● Técnica direta: em tubo estéril, 1 μg de anticorpo específico para CD44 (BD
PharmygenTM, biotin rat anti-mouse CD44) e CD24 (BD PharmygenTM, biotin rat anti-
mouse CD24) foi conjugado às beads magnéticas em temperatura ambiente durante
30 minutos. Após este período, o tubo foi colocado no imã por 1 minuto, o
sobrenadante descartado, e este procedimento repetido por mais três vezes para
remoção de anticorpos que porventura não foram conjugados. Então, as beads
ligadas ao respectivo anticorpo foram adicionadas à amostra de células em
suspensão, na concentração desejada. O material foi incubado em temperatura de
4ºC por 20 minutos, quando então foi recolocado no imã para separação da
população positiva, que no caso, ficariam aderidas na parede do tubo por força da
atração magnética promovida pelo imã. O conjunto beads/anticorpo/células foi
ressuspendido em Buffer 1 e recolocado no imã por mais duas vezes, sendo que o
sobrenadante de todas as lavagens foi retirado e colocado em tubo estéril para
verificação de células não-marcadas. Para interrupção da ligação entre as beads
magnéticas e as células, utilizou-se 4 μL de Release Buffer (solução de DNAse I) por
um período de 15 minutos em incubação e agitação leve à temperatura ambiente.
Após a liberação das células das beads magnéticas, a população marcada para
CD44 foi avaliada pelo método de exclusão de azul de Trypan e posteriormente
levada ao cultivo celular.
5.7 SEPARAÇÃO DE POPULAÇÕES POSITIVAS PARA A ATIVIDADE DE
ALDEÍDO-DESIDROGENASE (ALDH) ATRAVÉS DE CITOMETRIA DE FLUXO
A atividade celular de aldeído-desidrogenase (ALDH) foi analisada com o
auxílio do kit Aldefluor® (StemCell Technologies, EUA) conforme instruções do
fabricante. Foram preparados dois tubos para cada amostra, sendo um tubo teste e
um tubo controle. Brevemente, 106 células/mL foram ressuspendidas em 1,5 mL de
tampão e dispensadas em um tubo estéril, para então serem adicionadas de 5
µL/mL de reagente ativado Aldefluor®. Rapidamente, 500 µL desta suspensão foram
transferidas a outro tubo (controle da reação) contendo 5 µL de DEAB (inibidor da
reação entre a enzima e o reagente). As amostras foram incubadas por 40 minutos a

58
37°C, e então centrifugadas a 4°C a uma velocidade de 1000 rpm durante 5 minutos.
O sobrenadante foi descartado e o pellet ressuspendido em 0,5 mL do tampão
específico do kit. Os tubos foram mantidos no gelo até a análise. Para exclusão de
células mortas utilizou-se Hoechst (Hoechst 33342, Life Technologies ®, USA) cinco
minutos antes da análise.
5.8 CULTIVO CELULAR PARA FORMAÇÃO DE TUMORESFERAS
A capacidade de formação de colônias em condições de baixa aderência das
células avaliadas quanto ao fenótipo CD24, CD44 e atividade de ALDH foi realizada
através da cultura em condições de baixa aderência, com meio de cultivo livre de
soro fetal bovino suplementado para permitir a multiplicação celular, método este já
bem definido quanto à caracterização de Células Iniciadoras de Tumor e o potencial
de auto-renovação e formação de esferas que apresentam.
Para tal, o número de células em cada população foi ajustado para uma
concentração aproximada de 750 células/mL, no volume de 0,5 mL/poço a fim de
evitar a adesão das células e possível diferenciação celular. As amostras foram
plaqueadas em placas de baixa aderência (24-well low-adherent plates, Corning®,
USA) em meio DMEM-F12 suplementado com 20 ng/mL de EGF (PrepoTech, USA),
10 ng/mL de FGF (PrepoTech, USA), 5 μL/mL de insulina bovina (Sigma Aldrich®,
MO, USA) e 1% de antibiótico/antimicótico (PenStep, Gibco, Life Technologies®,
USA). Para manutenção do cultivo celular, 50 µL/poço de meio fresco foi adicionado
a cada 48 horas, e diariamente a placa era avaliada através de microscopia óptica. A
cultura foi analisada por 14 dias consecutivos.

59
5.9 ANÁLISE ESTATÍSTICA
A análise estatística utilizada para avaliação de significância na validação do
ensaio de tumorigênese in vitro para os grupos ALDHlow e ALDHhi, foi a análise de
variância (ANOVA) em uma via para verificação das diferenças amostrais entre os
fenótipos, seguida do pos-hoc Bonferroni. Para análise estatística quanto ao número
de esferas formadas a partir da separação dos fenótipos CD24 e CD44, foi adotado
o teste t de Student. As análises foram geradas pelo programa GraphPad Prism®
5.0, San Diego, CA, USA).

60
6 RESULTADOS
6.1 DISTRIBUIÇÃO DOS CASOS CLÍNICOS
Ao todo, trinta e cinco animais contribuíram neste trabalho. Todas as lesões
foram coletadas por excisão cirúrgica através de mastectomia eletiva, e todas foram
submetidas à avaliação anatomopatológica para definição do tecido de origem. Vinte
e nove animais (82,9%) foram considerados no presente estudo, pois apresentavam
lesão neoplásica em glândula mamária; outros dois (5,7%) foram diagnosticados
com cistos de queratina ou lesões papilomatosas, e em quatro (11,4%), o material
era insuficiente para estabelecer o diagnóstico, devido à qualidade e integridade do
tecido fixado (Gráfico 4). Portanto, estes seis animais foram excluídos das demais
análises.
Gráfico 4 - Distribuição dos casos nos trinta e cinco cães coletados no estudo, baseado no tecido de origem e número de lesões por animal
Legenda: à esquerda, distribuição dos casos segundo origem da lesão, nos trinta e cinco animais submetidos à mastectomia eletiva. À direita, número de lesões distribuídas nos vinte e nove animais com alteração em glândula mamária
A maioria dos casos (19/29) apresentava múltiplas lesões, enquanto que dez
casos tinham um único nódulo (34,5%) (Gráfico 4). Quanto à idade dos animais no
momento da cirurgia, onze (37,9%) tinham de 6 a 10 anos de idade, doze (41,4%)
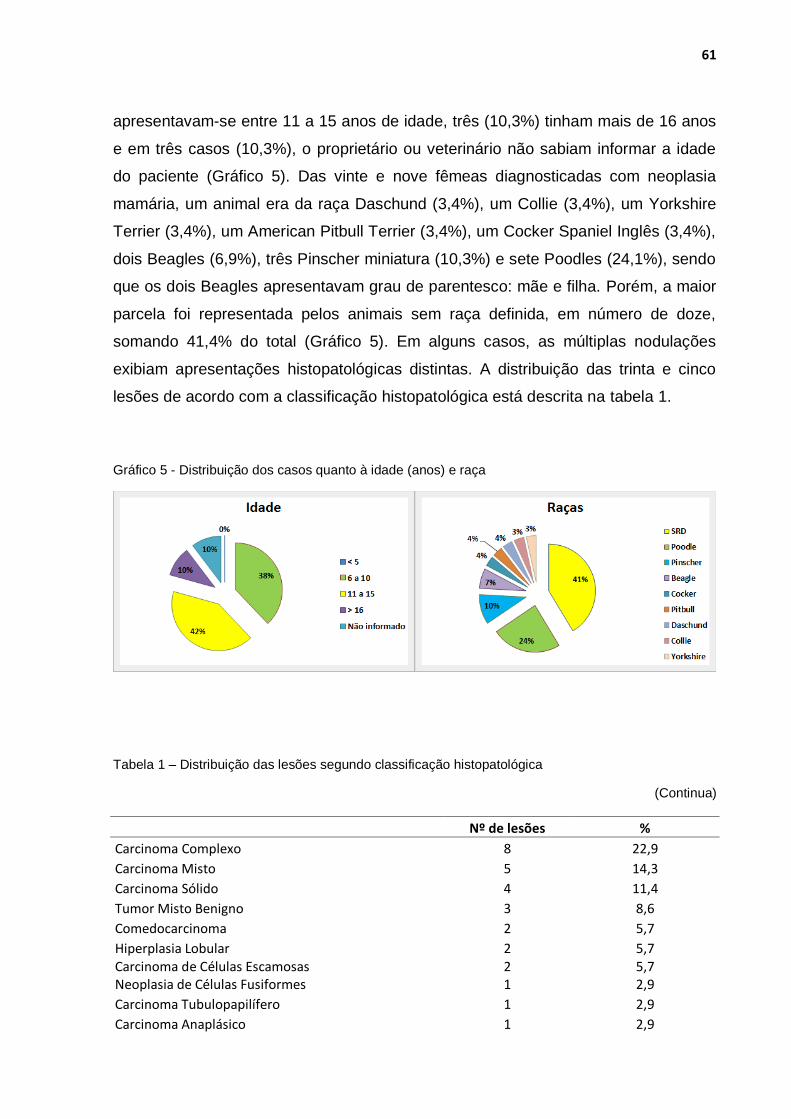
61
apresentavam-se entre 11 a 15 anos de idade, três (10,3%) tinham mais de 16 anos
e em três casos (10,3%), o proprietário ou veterinário não sabiam informar a idade
do paciente (Gráfico 5). Das vinte e nove fêmeas diagnosticadas com neoplasia
mamária, um animal era da raça Daschund (3,4%), um Collie (3,4%), um Yorkshire
Terrier (3,4%), um American Pitbull Terrier (3,4%), um Cocker Spaniel Inglês (3,4%),
dois Beagles (6,9%), três Pinscher miniatura (10,3%) e sete Poodles (24,1%), sendo
que os dois Beagles apresentavam grau de parentesco: mãe e filha. Porém, a maior
parcela foi representada pelos animais sem raça definida, em número de doze,
somando 41,4% do total (Gráfico 5). Em alguns casos, as múltiplas nodulações
exibiam apresentações histopatológicas distintas. A distribuição das trinta e cinco
lesões de acordo com a classificação histopatológica está descrita na tabela 1.
Gráfico 5 - Distribuição dos casos quanto à idade (anos) e raça
Tabela 1 – Distribuição das lesões segundo classificação histopatológica
(Continua)
Nº de lesões %
Carcinoma Complexo 8 22,9
Carcinoma Misto 5 14,3
Carcinoma Sólido 4 11,4
Tumor Misto Benigno 3 8,6
Comedocarcinoma 2 5,7
Hiperplasia Lobular 2 5,7 Carcinoma de Células Escamosas Neoplasia de Células Fusiformes
2 1
5,7 2,9
Carcinoma Tubulopapilífero 1 2,9
Carcinoma Anaplásico 1 2,9

62
(Conclusão)
Carcinoma adenoescamoso 1 2,9
Fibrossarcoma 1 2,9
Adenoma Basalóide 1 2,9
Carcinoma Tubular 1 2,9
Adenoma Tubular Simples 1 2,9
Carcinoma Ductal 1 2,9
Total 35 100%
Apenas uma das vinte e nove fêmeas era castrada antes do desenvolvimento
tumoral, porém, foi castrada aos nove anos de idade por conta de piometra. Nenhum
animal recebeu tratamento hormonal exógeno para controle do ciclo estral, ou
apresentou outro tipo de neoplasia anterior ao tumor mamário. Com exceção da
Beagle citada anteriormente, cuja mãe também apresentava câncer em glândula
mamária, não eram conhecidos os ascendentes das demais fêmeas. Não houve
informações suficientes sobre número de acasalamentos, número de gestações ou
abortos para a compilação dos dados, provavelmente dado ao desconhecimento
pelo proprietário ou clínico veterinário.
6.2. CARACTERIZAÇÃO DOS CULTIVOS CELULARES
6.2.1. Dissociação e cultivo primário em monocamada
Das trinta e cinco amostras de glândula mamária, vinte e seis foram
encaminhadas para o cultivo primário, de acordo com o descrito em material e
métodos. Foi definido um protocolo padrão para a dissociação e para o cultivo
primário das células obtidas a partir da excisão cirúrgica de neoplasias da glândula
mamária em cães. Após a dissociação e incubação, dezesseis apresentaram
quantidade considerável de células grandes, bem delimitadas, que em cerca de uma
semana já estavam completamente aderidas, formando grandes populações

63
celulares ao longo de toda a extensão da garrafa. Em todas as garrafas cultivadas,
as amostras apresentaram células bem conservadas e vivas, visualizadas através
de diluição em Azul de Trypan. Também, notou-se a presença de dois grupos
distintos de células aderidos à garrafa, que variavam em morfologia. O primeiro, com
características morfológicas de células mesenquimais, em formato fusiforme, núcleo
e nucléolos bem definidos. Algumas se apresentaram binucleadas ou
multinucleadas; já o segundo grupo, apresentava morfologia de células ovaladas ou
triangulares, com citoplasma extenso, muitas apresentando dois ou mais núcleos
(também bem definidos), dispostas em monocamada pavimentosa de formato
poligonal (Figura 8). Este protocolo permitiu o sucesso no cultivo de dezesseis das
vinte e seis amostras colocadas em cultivo, possibilitando a escolha de quatro
culturas para a caracterização quanto à origem tumoral das células que proliferaram
in vitro (Tabela 2).
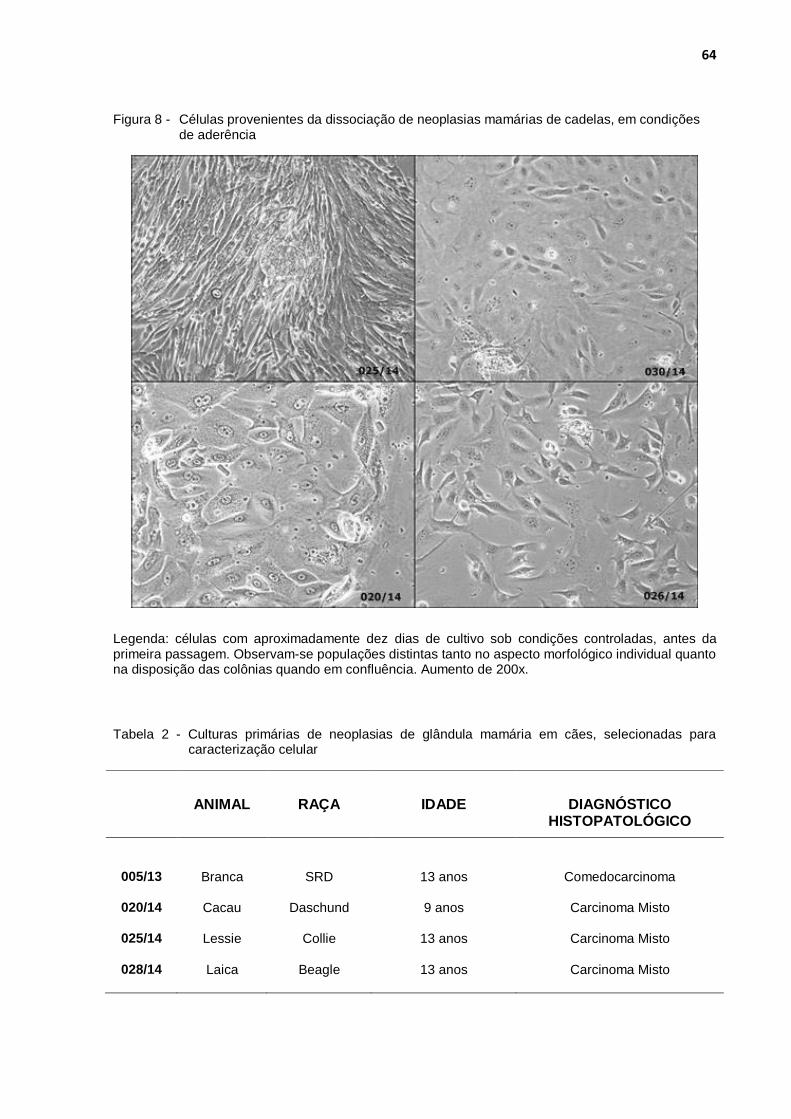
64
Figura 8 - Células provenientes da dissociação de neoplasias mamárias de cadelas, em condições de aderência
Legenda: células com aproximadamente dez dias de cultivo sob condições controladas, antes da primeira passagem. Observam-se populações distintas tanto no aspecto morfológico individual quanto na disposição das colônias quando em confluência. Aumento de 200x.
Tabela 2 - Culturas primárias de neoplasias de glândula mamária em cães, selecionadas para caracterização celular
ANIMAL
RAÇA
IDADE
DIAGNÓSTICO HISTOPATOLÓGICO
005/13
Branca
SRD
13 anos
Comedocarcinoma
020/14 Cacau Daschund 9 anos Carcinoma Misto
025/14 Lessie Collie 13 anos Carcinoma Misto
028/14 Laica Beagle 13 anos Carcinoma Misto

65
As quatro amostras foram selecionadas para caracterização quanto a: morfologia
celular do cultivo em monocamada; expressão imunocitoquímica de citoqueratina,
vimentina e alfa-actina muscular; avaliação citogenética por cariotipagem; avaliação
do tempo de duplicação celular; quantificação das populações positivas e negativas
quanto ao fenótipo CD24/CD44; e avaliação das populações quanto à expressão de
aldeído-desidrogenase.
6.2.2. Classificação histopatológica e morfologia da cultura primária
O material fixado em formol foi processado para obtenção de lâminas
histológicas, e estas coradas com hematoxilina e eosina (HE) para avaliação e
classificação histopatológica das lesões que foram encaminhadas ao cultivo de
células. Do mesmo modo, as culturas primárias foram cultivadas em lamínulas,
fixadas e coradas em HE para caracterização quanto à morfologia celular.
Amostra 005/13: Macroscopicamente, anterior à mastectomia, a lesão se
apresentava de forma inflamatória, parcialmente ulcerada, com eritema, perda de
pelos e pústulas na região abdominal. A formação neoplásica era extensa no
comprimento longitudinal, chegando a 10 cm de extensão. Histologicamente, foi
diagnosticada como comedocarcinoma, com a lâmina histológica caracterizada pela
presença de áreas necróticas em meio a agregados celulares neoplásicos em
consequência de células apoptóticas e cariorréxis subsequente. Nas áreas de
necrose, presença de grande quantidade de material eosinofílico, amorfo,
entremeado com debris celulares, neutrófilos e alguns macrófagos. O tecido
adjacente aos focos de necrose consiste em agregados celulares organizados de
maneira sólida, tubulares e em cordões, apoiados por estroma fibrovascular (Figura
9). Na cultura primária, as células aderidas em monocamada se apresentaram em
formato alongado, com tamanho variado apesar de ter em sua maioria células
pequenas, e algumas células binucleadas. Em relação ao núcleo, formato oval,
presença de anisocariose, nucléolos evidentes e múltiplos em muitas células, alguns
em formato angular. Quando em grande confluência, as células tendem a formar
redes, deixando áreas livres entre os grupos celulares. No início, era comum o
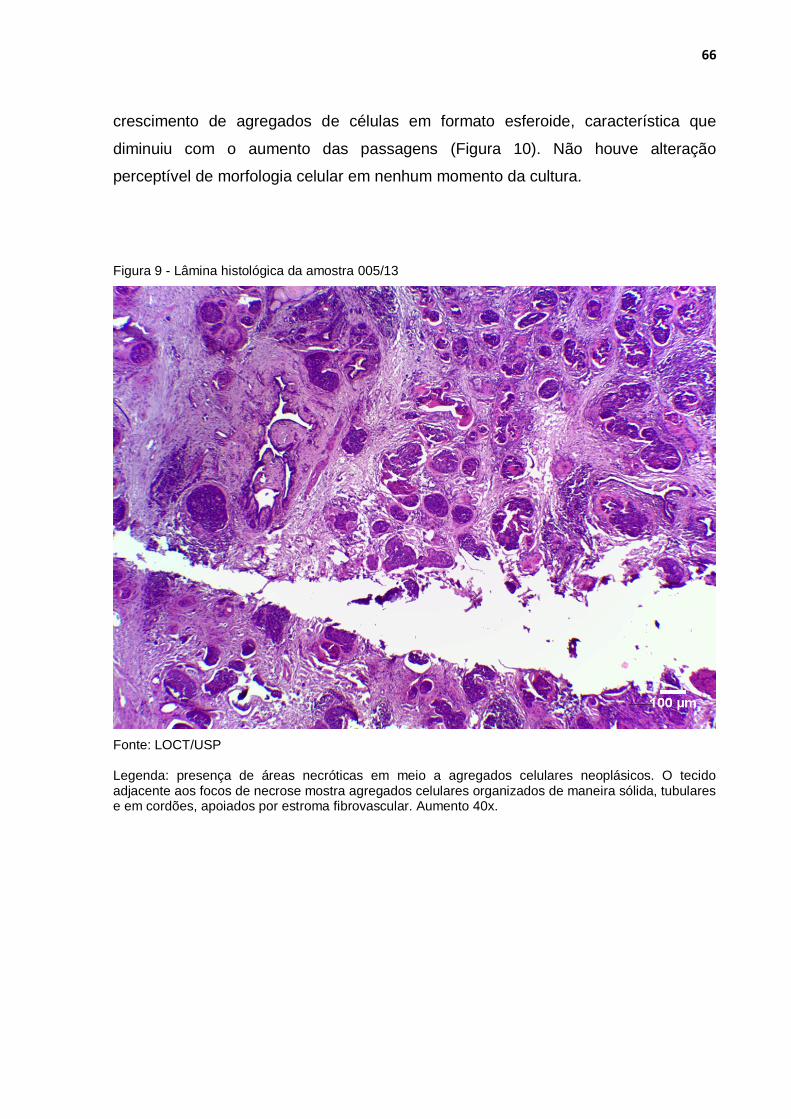
66
crescimento de agregados de células em formato esferoide, característica que
diminuiu com o aumento das passagens (Figura 10). Não houve alteração
perceptível de morfologia celular em nenhum momento da cultura.
Figura 9 - Lâmina histológica da amostra 005/13
Fonte: LOCT/USP Legenda: presença de áreas necróticas em meio a agregados celulares neoplásicos. O tecido adjacente aos focos de necrose mostra agregados celulares organizados de maneira sólida, tubulares e em cordões, apoiados por estroma fibrovascular. Aumento 40x.
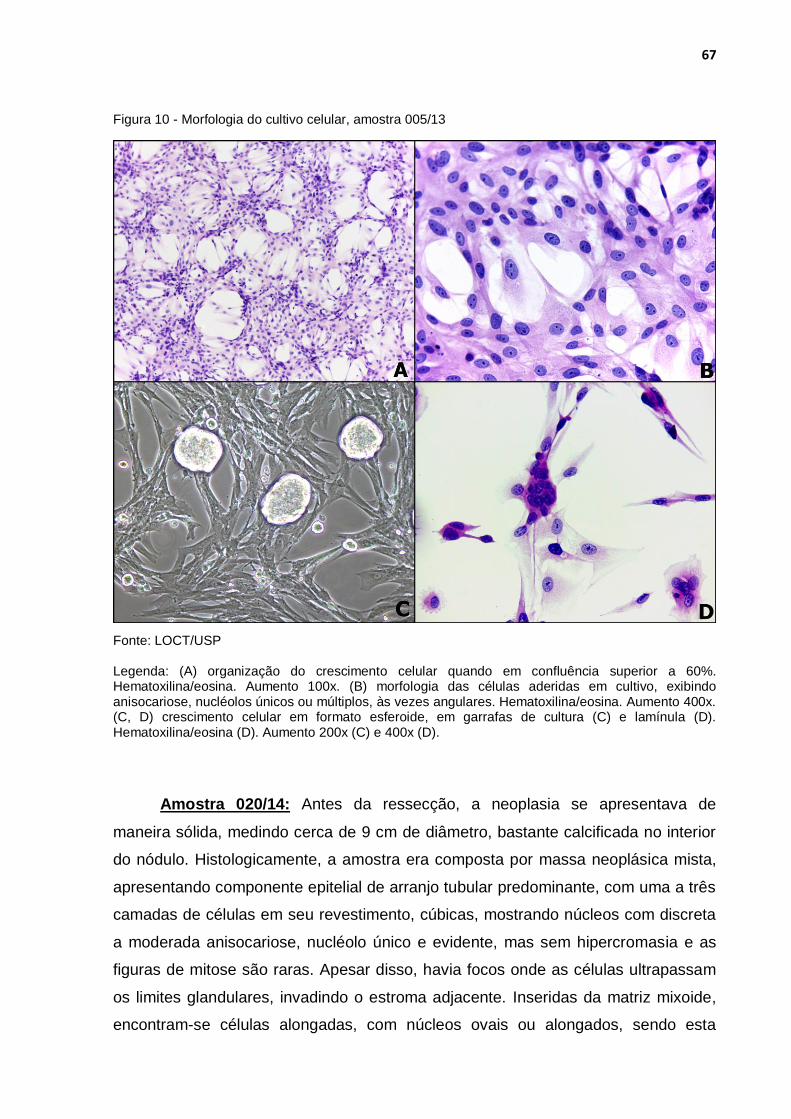
67
Figura 10 - Morfologia do cultivo celular, amostra 005/13
Fonte: LOCT/USP Legenda: (A) organização do crescimento celular quando em confluência superior a 60%. Hematoxilina/eosina. Aumento 100x. (B) morfologia das células aderidas em cultivo, exibindo anisocariose, nucléolos únicos ou múltiplos, às vezes angulares. Hematoxilina/eosina. Aumento 400x. (C, D) crescimento celular em formato esferoide, em garrafas de cultura (C) e lamínula (D). Hematoxilina/eosina (D). Aumento 200x (C) e 400x (D).
Amostra 020/14: Antes da ressecção, a neoplasia se apresentava de
maneira sólida, medindo cerca de 9 cm de diâmetro, bastante calcificada no interior
do nódulo. Histologicamente, a amostra era composta por massa neoplásica mista,
apresentando componente epitelial de arranjo tubular predominante, com uma a três
camadas de células em seu revestimento, cúbicas, mostrando núcleos com discreta
a moderada anisocariose, nucléolo único e evidente, mas sem hipercromasia e as
figuras de mitose são raras. Apesar disso, havia focos onde as células ultrapassam
os limites glandulares, invadindo o estroma adjacente. Inseridas da matriz mixoide,
encontram-se células alongadas, com núcleos ovais ou alongados, sendo esta
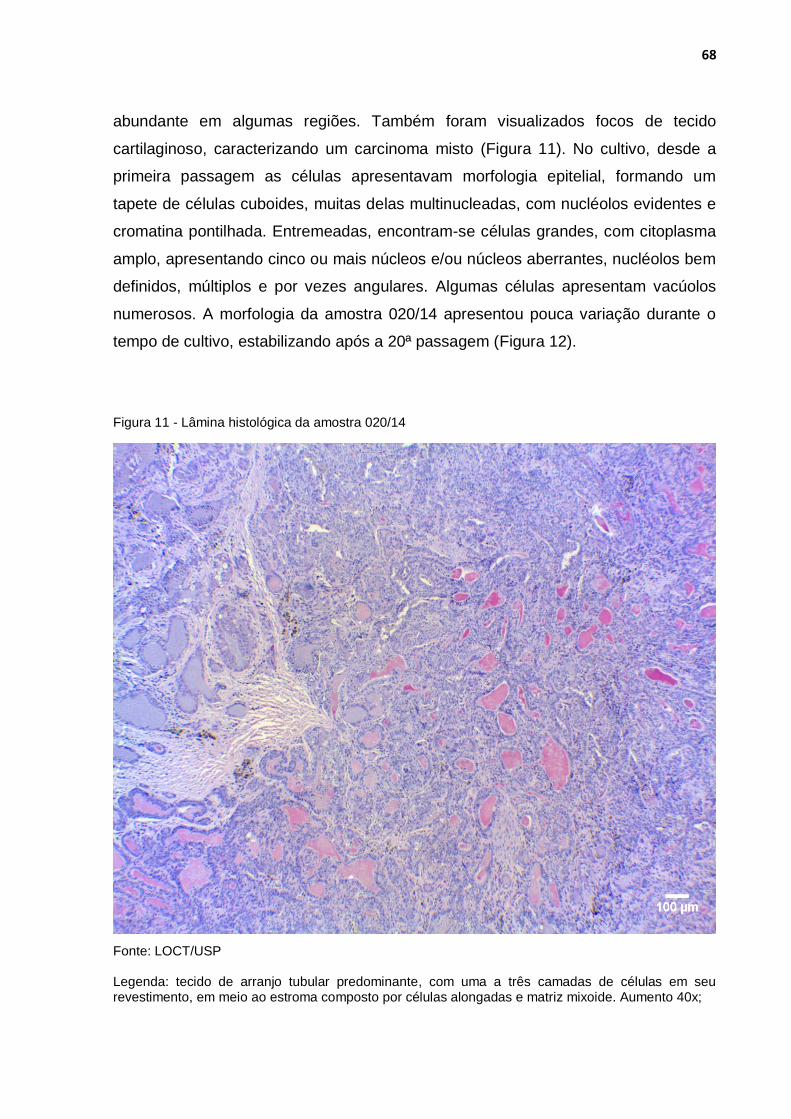
68
abundante em algumas regiões. Também foram visualizados focos de tecido
cartilaginoso, caracterizando um carcinoma misto (Figura 11). No cultivo, desde a
primeira passagem as células apresentavam morfologia epitelial, formando um
tapete de células cuboides, muitas delas multinucleadas, com nucléolos evidentes e
cromatina pontilhada. Entremeadas, encontram-se células grandes, com citoplasma
amplo, apresentando cinco ou mais núcleos e/ou núcleos aberrantes, nucléolos bem
definidos, múltiplos e por vezes angulares. Algumas células apresentam vacúolos
numerosos. A morfologia da amostra 020/14 apresentou pouca variação durante o
tempo de cultivo, estabilizando após a 20ª passagem (Figura 12).
Figura 11 - Lâmina histológica da amostra 020/14
Fonte: LOCT/USP Legenda: tecido de arranjo tubular predominante, com uma a três camadas de células em seu revestimento, em meio ao estroma composto por células alongadas e matriz mixoide. Aumento 40x;
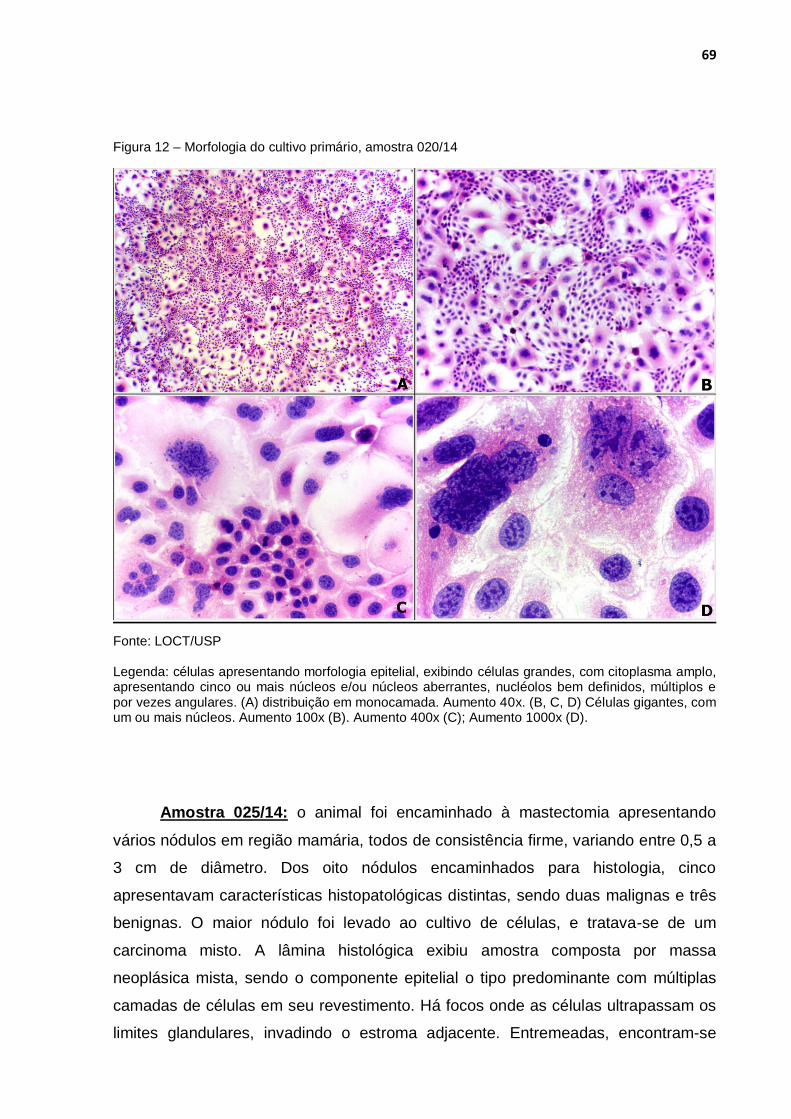
69
Figura 12 – Morfologia do cultivo primário, amostra 020/14
Fonte: LOCT/USP Legenda: células apresentando morfologia epitelial, exibindo células grandes, com citoplasma amplo, apresentando cinco ou mais núcleos e/ou núcleos aberrantes, nucléolos bem definidos, múltiplos e por vezes angulares. (A) distribuição em monocamada. Aumento 40x. (B, C, D) Células gigantes, com um ou mais núcleos. Aumento 100x (B). Aumento 400x (C); Aumento 1000x (D).
Amostra 025/14: o animal foi encaminhado à mastectomia apresentando
vários nódulos em região mamária, todos de consistência firme, variando entre 0,5 a
3 cm de diâmetro. Dos oito nódulos encaminhados para histologia, cinco
apresentavam características histopatológicas distintas, sendo duas malignas e três
benignas. O maior nódulo foi levado ao cultivo de células, e tratava-se de um
carcinoma misto. A lâmina histológica exibiu amostra composta por massa
neoplásica mista, sendo o componente epitelial o tipo predominante com múltiplas
camadas de células em seu revestimento. Há focos onde as células ultrapassam os
limites glandulares, invadindo o estroma adjacente. Entremeadas, encontram-se
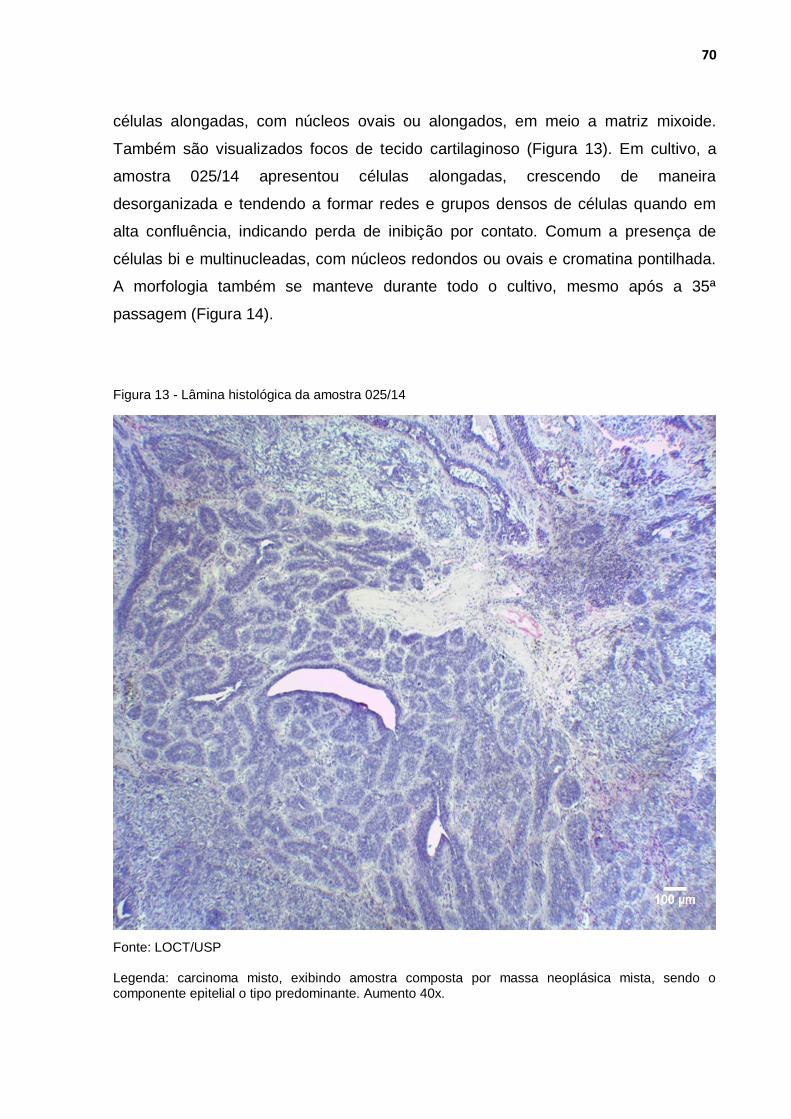
70
células alongadas, com núcleos ovais ou alongados, em meio a matriz mixoide.
Também são visualizados focos de tecido cartilaginoso (Figura 13). Em cultivo, a
amostra 025/14 apresentou células alongadas, crescendo de maneira
desorganizada e tendendo a formar redes e grupos densos de células quando em
alta confluência, indicando perda de inibição por contato. Comum a presença de
células bi e multinucleadas, com núcleos redondos ou ovais e cromatina pontilhada.
A morfologia também se manteve durante todo o cultivo, mesmo após a 35ª
passagem (Figura 14).
Figura 13 - Lâmina histológica da amostra 025/14
Fonte: LOCT/USP Legenda: carcinoma misto, exibindo amostra composta por massa neoplásica mista, sendo o componente epitelial o tipo predominante. Aumento 40x.
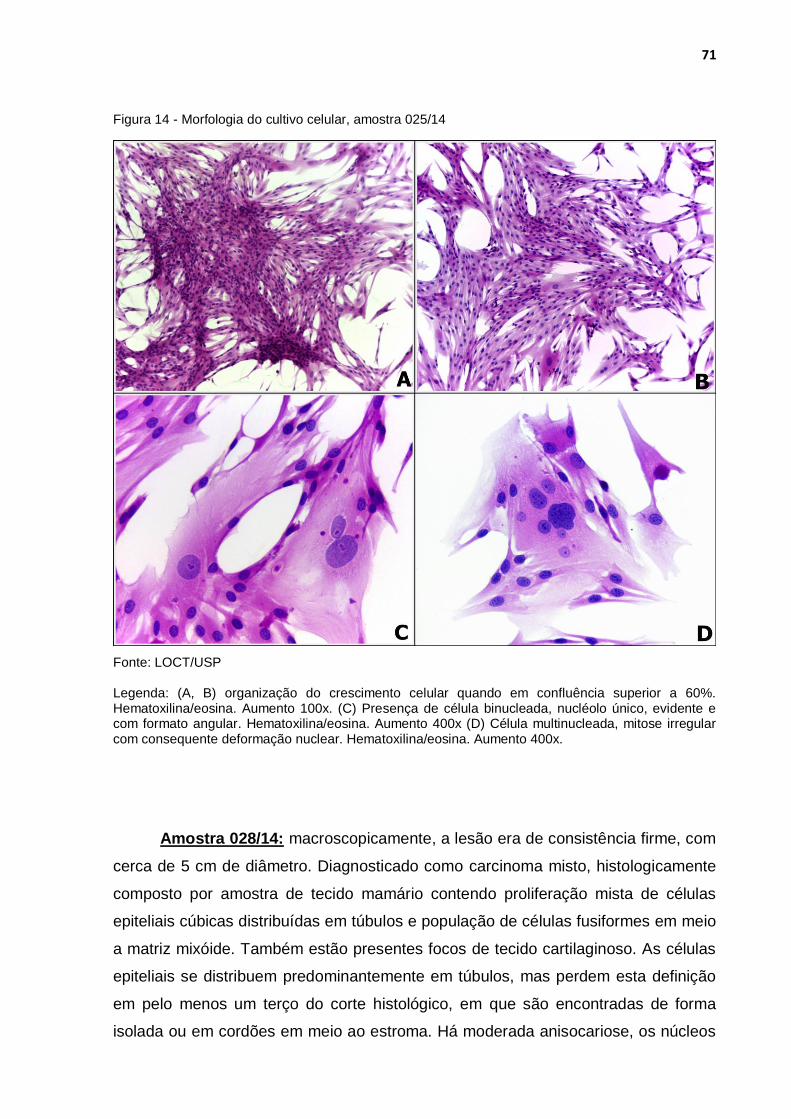
71
Figura 14 - Morfologia do cultivo celular, amostra 025/14
Fonte: LOCT/USP Legenda: (A, B) organização do crescimento celular quando em confluência superior a 60%. Hematoxilina/eosina. Aumento 100x. (C) Presença de célula binucleada, nucléolo único, evidente e com formato angular. Hematoxilina/eosina. Aumento 400x (D) Célula multinucleada, mitose irregular com consequente deformação nuclear. Hematoxilina/eosina. Aumento 400x.
Amostra 028/14: macroscopicamente, a lesão era de consistência firme, com
cerca de 5 cm de diâmetro. Diagnosticado como carcinoma misto, histologicamente
composto por amostra de tecido mamário contendo proliferação mista de células
epiteliais cúbicas distribuídas em túbulos e população de células fusiformes em meio
a matriz mixóide. Também estão presentes focos de tecido cartilaginoso. As células
epiteliais se distribuem predominantemente em túbulos, mas perdem esta definição
em pelo menos um terço do corte histológico, em que são encontradas de forma
isolada ou em cordões em meio ao estroma. Há moderada anisocariose, os núcleos

72
contém cromatina pontilhada e com nucléolo evidente na maioria das células. Há
figuras de mitose típicas em pequeno número (Figura 15). Na cultura primária, a
amostra 028/14 foi a que mais apresentou mudança no fenótipo com o aumento das
passagens. Inicialmente, a monocamada era composta por células de formato mais
alongado, com citoplasma amplo, um ou dois núcleos com nucléolo evidente,
geralmente único, redondo ou angular. Entre a 15ª e 16ª passagens, o cultivo in vitro
apresentou ilhas de células epiteliais pequenas e cúbicas cercadas por células de
aspecto mesenquimal simulando estroma tecidual, formando populações bem
definidas nas garrafas de cultura. Porém, devido a proliferação mais rápida das
células com morfologia epitelial, nas passagens seguintes elas acabaram por
dominar o cultivo primário, e a cultura estabilizou a partir da 18ª passagem. As
células se apresentam com relação núcleo/citoplasma diminuída, muitas células
multinucleadas, grande vacuolização do citoplasma, e perda evidente da inibição do
crescimento por contato, sendo possível a visualização de células sobrepostas. Os
núcleos são grandes, com nucléolos aumentados, múltiplos, muitas vezes
angulares, cromatina densa e irregular. Mitoses aberrantes também são comuns
(Figura 16).
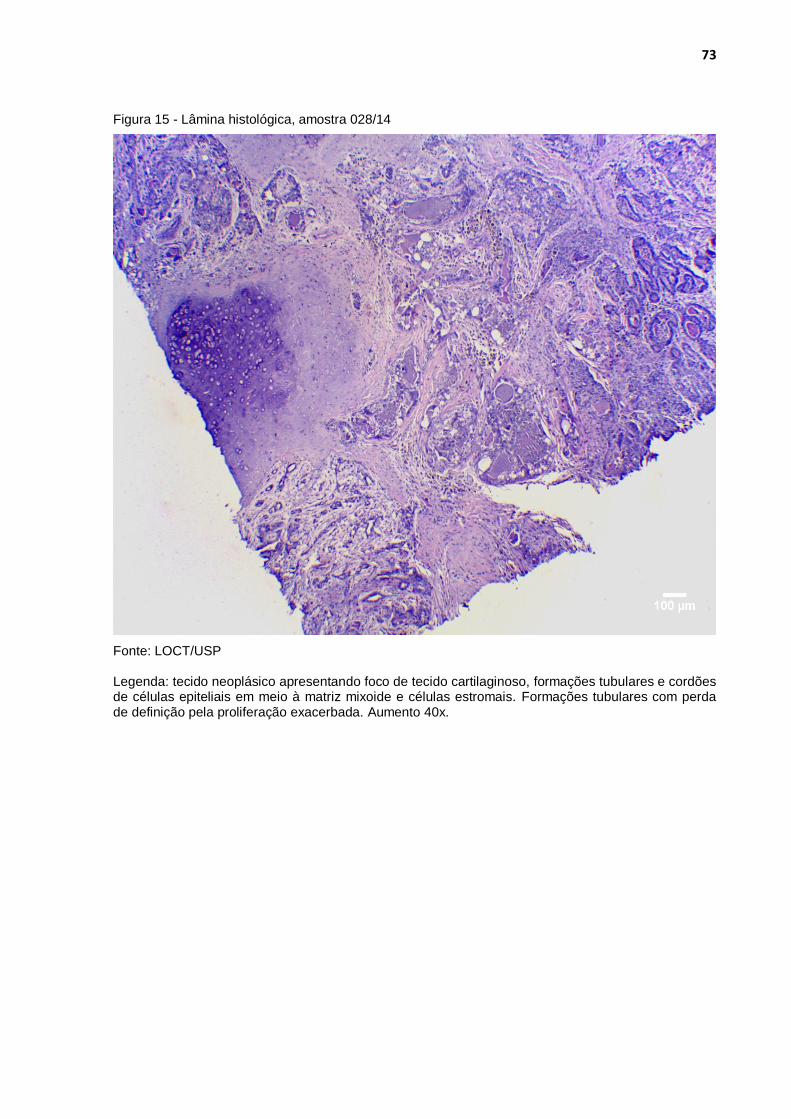
73
Figura 15 - Lâmina histológica, amostra 028/14
Fonte: LOCT/USP Legenda: tecido neoplásico apresentando foco de tecido cartilaginoso, formações tubulares e cordões de células epiteliais em meio à matriz mixoide e células estromais. Formações tubulares com perda de definição pela proliferação exacerbada. Aumento 40x.
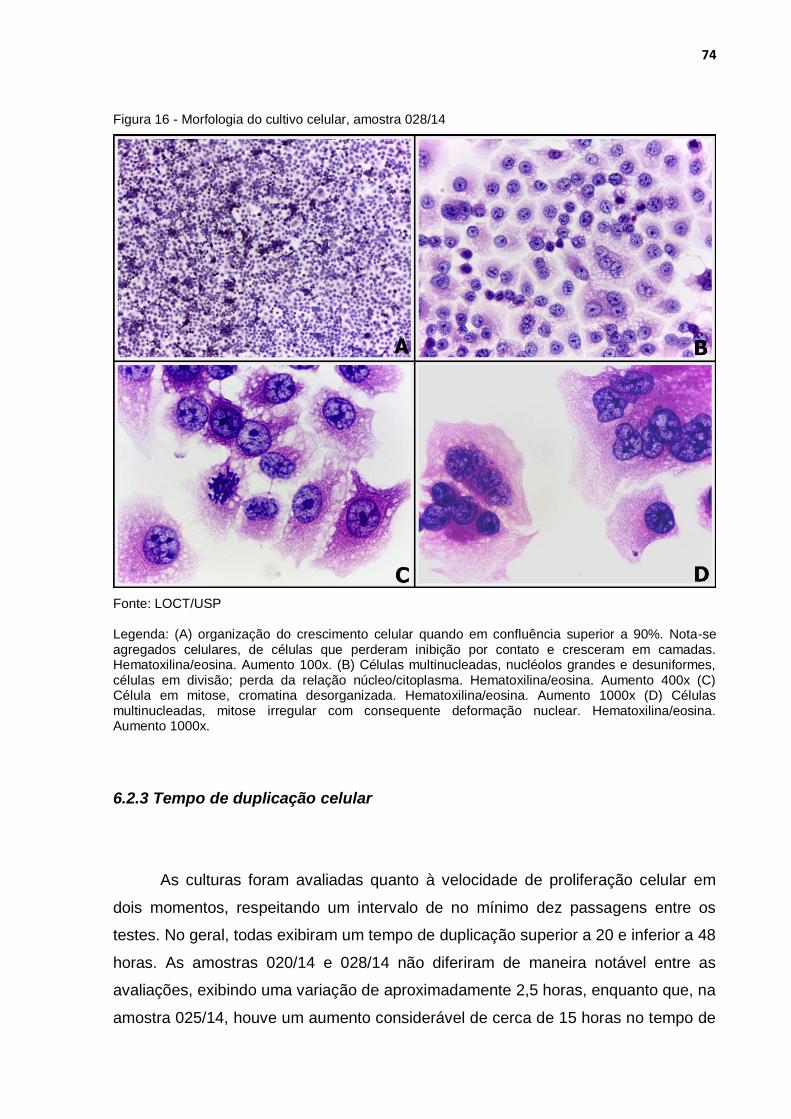
74
Figura 16 - Morfologia do cultivo celular, amostra 028/14
Fonte: LOCT/USP Legenda: (A) organização do crescimento celular quando em confluência superior a 90%. Nota-se agregados celulares, de células que perderam inibição por contato e cresceram em camadas. Hematoxilina/eosina. Aumento 100x. (B) Células multinucleadas, nucléolos grandes e desuniformes, células em divisão; perda da relação núcleo/citoplasma. Hematoxilina/eosina. Aumento 400x (C) Célula em mitose, cromatina desorganizada. Hematoxilina/eosina. Aumento 1000x (D) Células multinucleadas, mitose irregular com consequente deformação nuclear. Hematoxilina/eosina. Aumento 1000x.
6.2.3 Tempo de duplicação celular
As culturas foram avaliadas quanto à velocidade de proliferação celular em
dois momentos, respeitando um intervalo de no mínimo dez passagens entre os
testes. No geral, todas exibiram um tempo de duplicação superior a 20 e inferior a 48
horas. As amostras 020/14 e 028/14 não diferiram de maneira notável entre as
avaliações, exibindo uma variação de aproximadamente 2,5 horas, enquanto que, na
amostra 025/14, houve um aumento considerável de cerca de 15 horas no tempo de
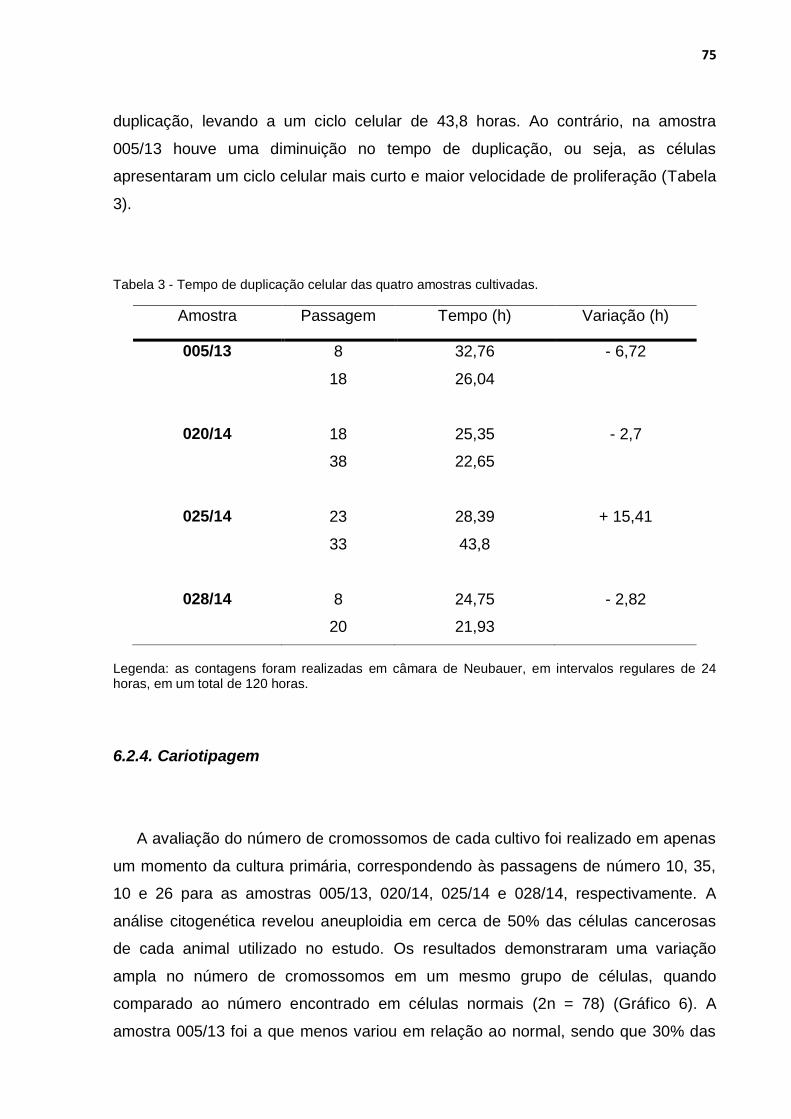
75
duplicação, levando a um ciclo celular de 43,8 horas. Ao contrário, na amostra
005/13 houve uma diminuição no tempo de duplicação, ou seja, as células
apresentaram um ciclo celular mais curto e maior velocidade de proliferação (Tabela
3).
Tabela 3 - Tempo de duplicação celular das quatro amostras cultivadas.
Amostra Passagem Tempo (h) Variação (h)
005/13 8
18
32,76
26,04
- 6,72
020/14 18
38
25,35
22,65
- 2,7
025/14 23
33
28,39
43,8
+ 15,41
028/14 8
20
24,75
21,93
- 2,82
Legenda: as contagens foram realizadas em câmara de Neubauer, em intervalos regulares de 24 horas, em um total de 120 horas.
6.2.4. Cariotipagem
A avaliação do número de cromossomos de cada cultivo foi realizado em apenas
um momento da cultura primária, correspondendo às passagens de número 10, 35,
10 e 26 para as amostras 005/13, 020/14, 025/14 e 028/14, respectivamente. A
análise citogenética revelou aneuploidia em cerca de 50% das células cancerosas
de cada animal utilizado no estudo. Os resultados demonstraram uma variação
ampla no número de cromossomos em um mesmo grupo de células, quando
comparado ao número encontrado em células normais (2n = 78) (Gráfico 6). A
amostra 005/13 foi a que menos variou em relação ao normal, sendo que 30% das
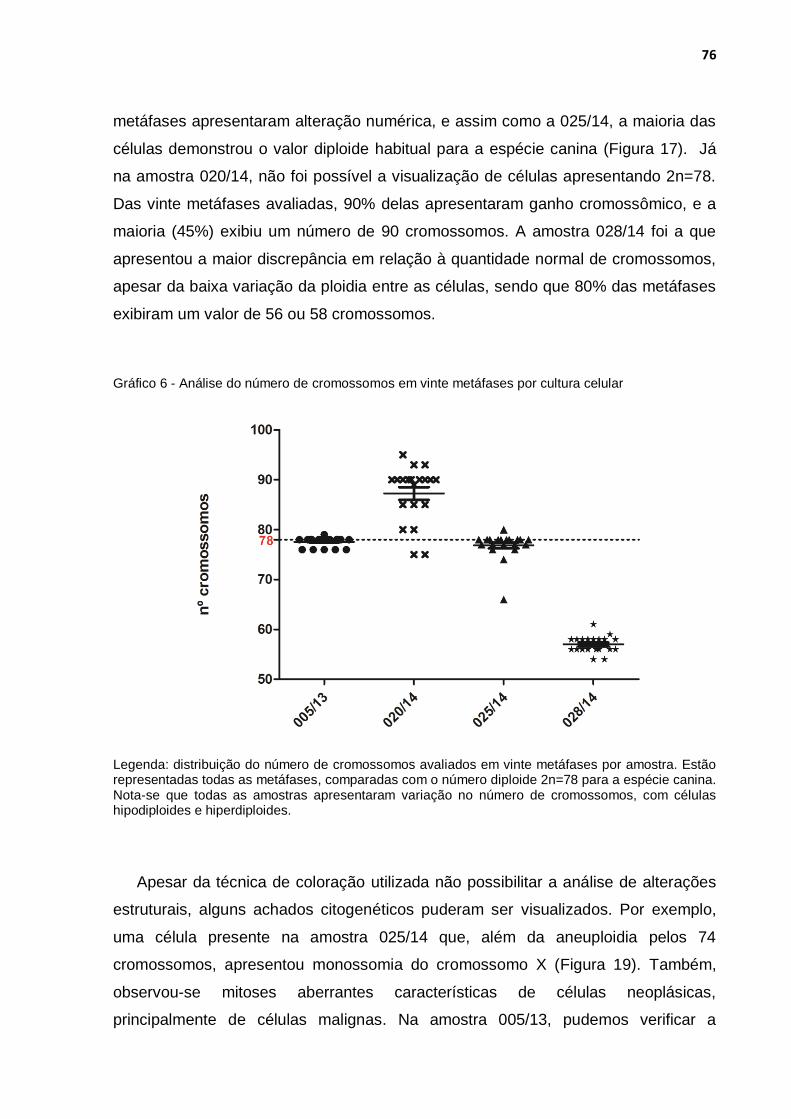
76
metáfases apresentaram alteração numérica, e assim como a 025/14, a maioria das
células demonstrou o valor diploide habitual para a espécie canina (Figura 17). Já
na amostra 020/14, não foi possível a visualização de células apresentando 2n=78.
Das vinte metáfases avaliadas, 90% delas apresentaram ganho cromossômico, e a
maioria (45%) exibiu um número de 90 cromossomos. A amostra 028/14 foi a que
apresentou a maior discrepância em relação à quantidade normal de cromossomos,
apesar da baixa variação da ploidia entre as células, sendo que 80% das metáfases
exibiram um valor de 56 ou 58 cromossomos.
Gráfico 6 - Análise do número de cromossomos em vinte metáfases por cultura celular
Legenda: distribuição do número de cromossomos avaliados em vinte metáfases por amostra. Estão representadas todas as metáfases, comparadas com o número diploide 2n=78 para a espécie canina. Nota-se que todas as amostras apresentaram variação no número de cromossomos, com células hipodiploides e hiperdiploides.
Apesar da técnica de coloração utilizada não possibilitar a análise de alterações
estruturais, alguns achados citogenéticos puderam ser visualizados. Por exemplo,
uma célula presente na amostra 025/14 que, além da aneuploidia pelos 74
cromossomos, apresentou monossomia do cromossomo X (Figura 19). Também,
observou-se mitoses aberrantes características de células neoplásicas,
principalmente de células malignas. Na amostra 005/13, pudemos verificar a
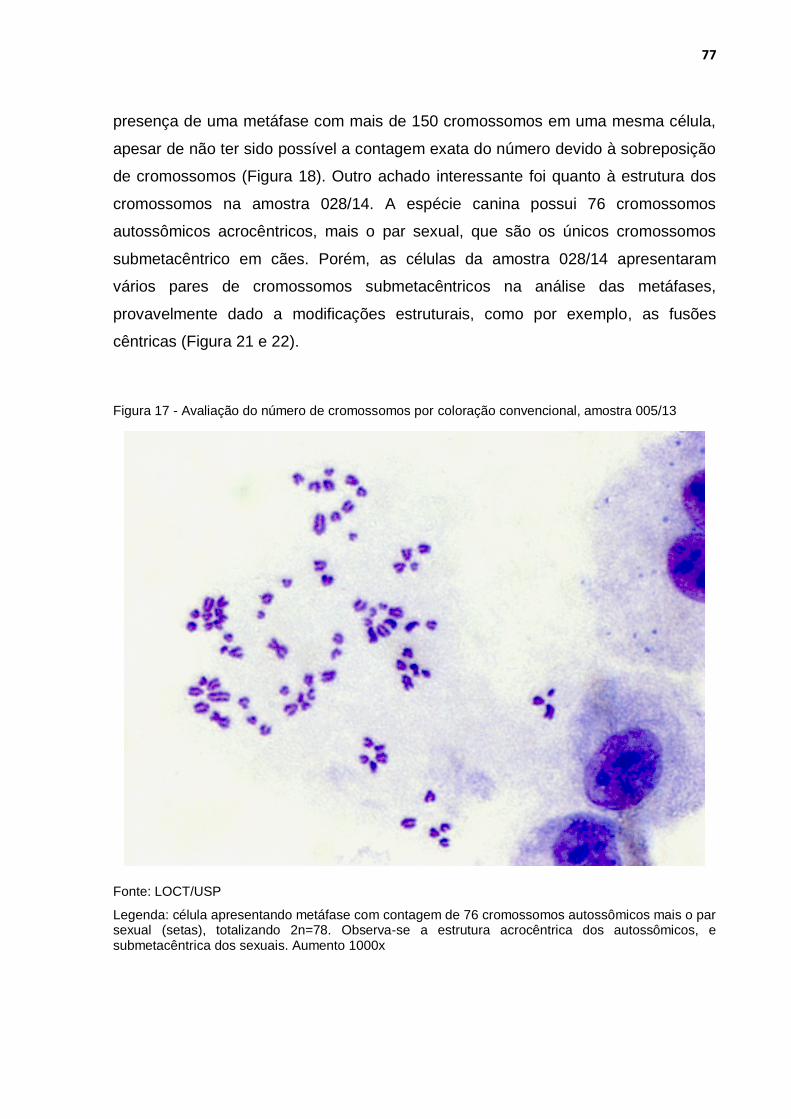
77
presença de uma metáfase com mais de 150 cromossomos em uma mesma célula,
apesar de não ter sido possível a contagem exata do número devido à sobreposição
de cromossomos (Figura 18). Outro achado interessante foi quanto à estrutura dos
cromossomos na amostra 028/14. A espécie canina possui 76 cromossomos
autossômicos acrocêntricos, mais o par sexual, que são os únicos cromossomos
submetacêntrico em cães. Porém, as células da amostra 028/14 apresentaram
vários pares de cromossomos submetacêntricos na análise das metáfases,
provavelmente dado a modificações estruturais, como por exemplo, as fusões
cêntricas (Figura 21 e 22).
Figura 17 - Avaliação do número de cromossomos por coloração convencional, amostra 005/13
Fonte: LOCT/USP
Legenda: célula apresentando metáfase com contagem de 76 cromossomos autossômicos mais o par sexual (setas), totalizando 2n=78. Observa-se a estrutura acrocêntrica dos autossômicos, e submetacêntrica dos sexuais. Aumento 1000x
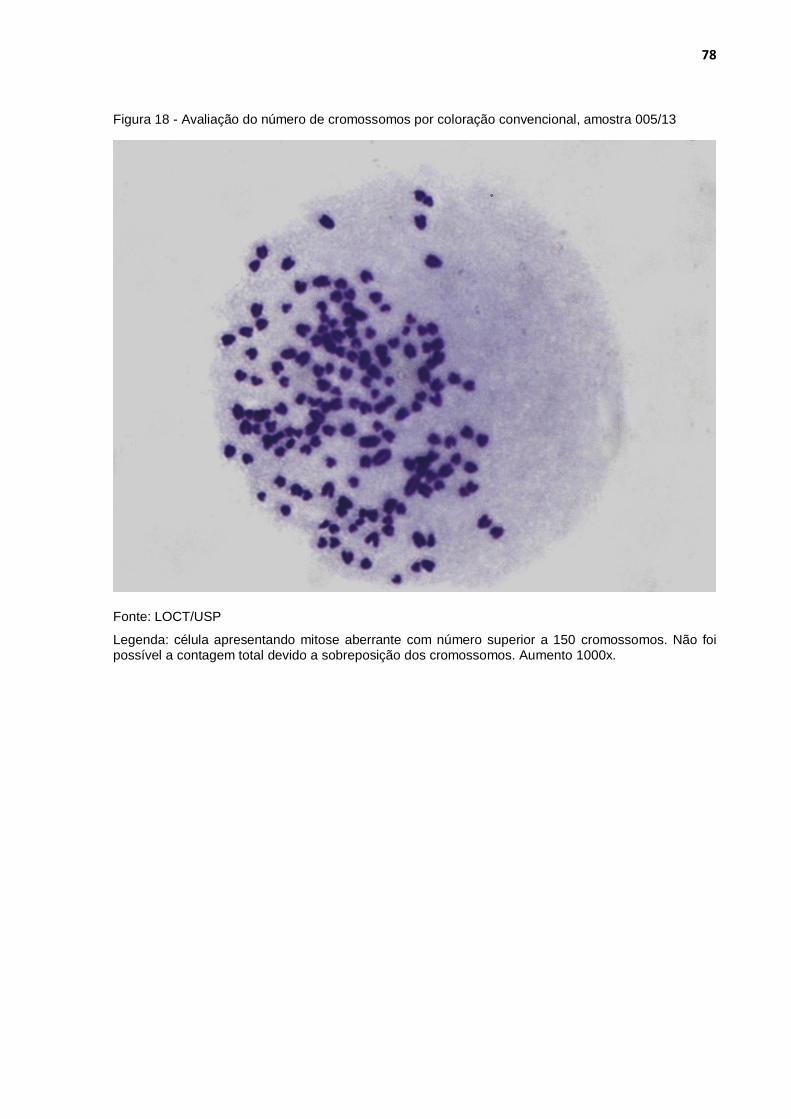
78
Figura 18 - Avaliação do número de cromossomos por coloração convencional, amostra 005/13
Fonte: LOCT/USP
Legenda: célula apresentando mitose aberrante com número superior a 150 cromossomos. Não foi possível a contagem total devido a sobreposição dos cromossomos. Aumento 1000x.

79
Figura 19 – Avaliação do número de cromossomos por coloração convencional, amostra 025/14
Fonte: LOCT/USP
Legenda: célula neoplásica com metáfase de 73 cromossomos, apresentando monossomia do cromossomo X (seta). Aumento 1000x.
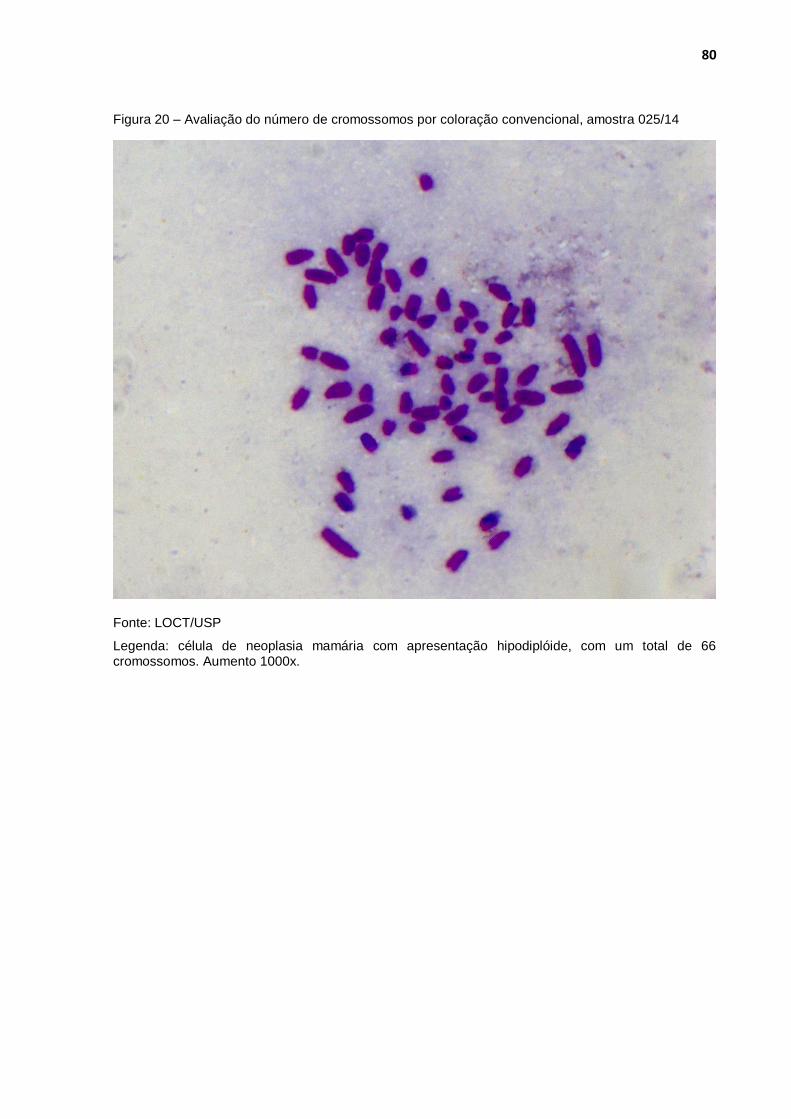
80
Figura 20 – Avaliação do número de cromossomos por coloração convencional, amostra 025/14
Fonte: LOCT/USP
Legenda: célula de neoplasia mamária com apresentação hipodiplóide, com um total de 66 cromossomos. Aumento 1000x.
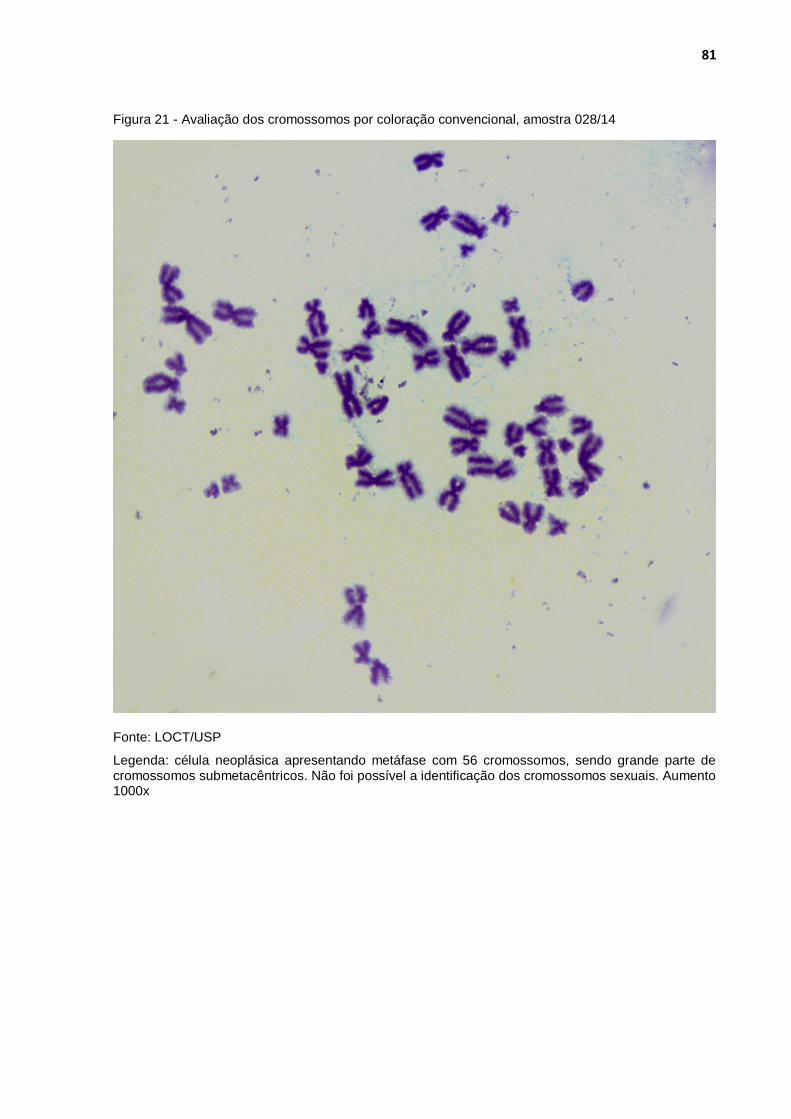
81
Figura 21 - Avaliação dos cromossomos por coloração convencional, amostra 028/14
Fonte: LOCT/USP
Legenda: célula neoplásica apresentando metáfase com 56 cromossomos, sendo grande parte de cromossomos submetacêntricos. Não foi possível a identificação dos cromossomos sexuais. Aumento 1000x
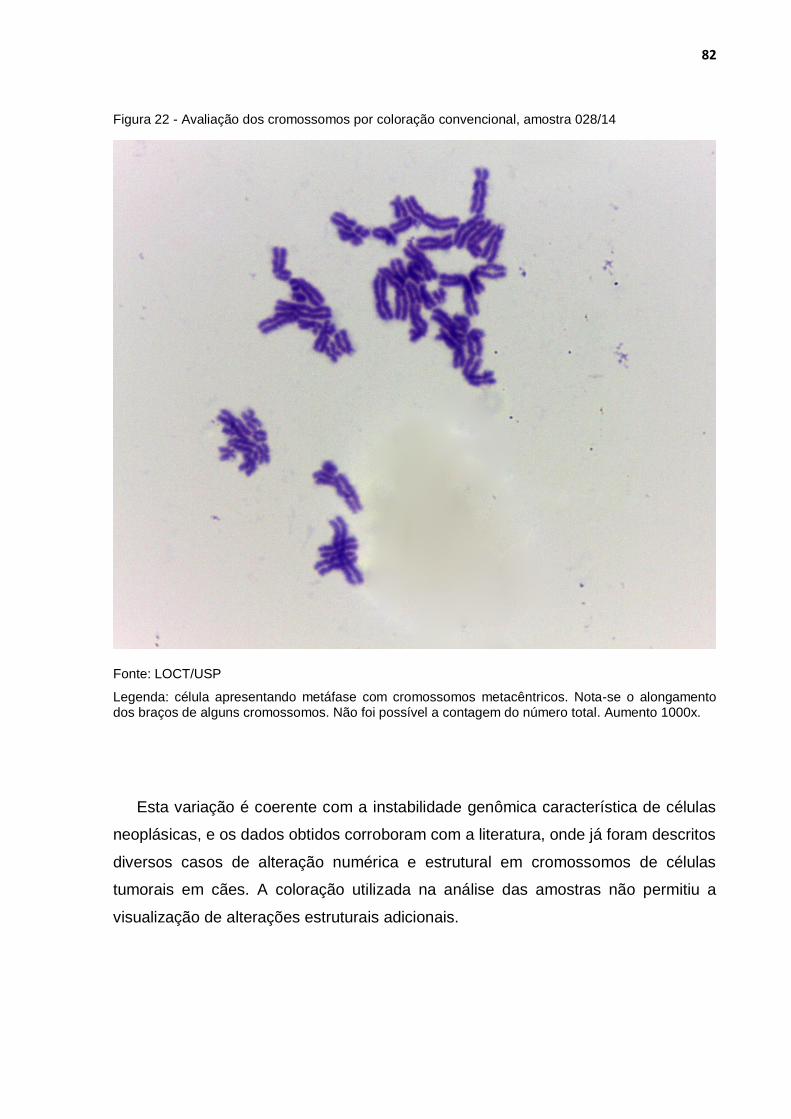
82
Figura 22 - Avaliação dos cromossomos por coloração convencional, amostra 028/14
Fonte: LOCT/USP
Legenda: célula apresentando metáfase com cromossomos metacêntricos. Nota-se o alongamento dos braços de alguns cromossomos. Não foi possível a contagem do número total. Aumento 1000x.
Esta variação é coerente com a instabilidade genômica característica de células
neoplásicas, e os dados obtidos corroboram com a literatura, onde já foram descritos
diversos casos de alteração numérica e estrutural em cromossomos de células
tumorais em cães. A coloração utilizada na análise das amostras não permitiu a
visualização de alterações estruturais adicionais.

83
6.2.5. Imunocitoquímica
Uma das principais formas de caracterização de células em cultivo primário é
a avaliação quanto à expressão de marcadores próprios de filamentos intermediários
(FI), que podem sugerir a origem tecidual, ou a morfologia que estas células
adquiriram quando colocadas em crescimento in vitro. As marcações comumente
utilizadas nestes casos são feitas pela expressão de: citoqueratina, filamentos
característicos de células de origem epitelial; vimentina, expressa por células de
origem mesenquimal; e α-actina de músculo liso, marcador de células mioepiteliais.
Neste trabalho, foi realizada a marcação imunocitoquímica para avaliação da
expressão dos três interfilamentos estruturais nas quatro amostras selecionadas
para a caracterização, bem como em fibroblastos embrionários utilizados como
controle negativo quando da marcação de citoqueratina, e controle positivo quando
da marcação de vimentina. Os resultados mostraram a expressão de citoqueratina
em todas as amostras avaliadas, sendo este FI expresso de forma heterogênea nas
amostras 005/13 e 025/14, e de maneira homogênea nas células do cultivo 020/14 e
028/14, sempre apresentando um uma marcação intensa nas células positivas.
Quanto a vimentina, foi expressa de maneira intensa em todas as células das
amostras 005/13 e 025/14, e sem mudanças no perfil de marcação entre as
diferentes passagens analisadas (Figuras 23 a 26). Já para as amostras 020/14 e
028/14, que exibiam morfologia celular mais característica de células epiteliais ao
final do experimento, a expressão de vimentina ocorreu de forma heterogênea para
as células do cultivo 028/14, e negativa para a amostra 020/14; curiosamente, o
padrão de marcação destas amostras variou com o aumento das passagens; nos
dois casos, a expressão de vimentina era positiva e de maneira homogênea em
passagens mais baixas (Figuras 27 e 28). Assim que o cultivo modificou levemente o
fenótipo e assumiu a morfologia cúbica característica de camadas epiteliais, houve
uma diminuição na expressão deste interfilamento, até chegar ao ponto de não ser
expressa, no caso da amostra 020/14. Por último, a α-actina de músculo liso foi
expressa de forma heterogênea nas amostras 005/13 e 025/14, semelhante à
citoqueratina; no cultivo 028/14, apenas algumas células isoladas expressaram este
FI, e de maneira discreta; já para a amostra 020/14, não houve marcação em
nenhuma célula avaliada (Figuras 23 a 26).
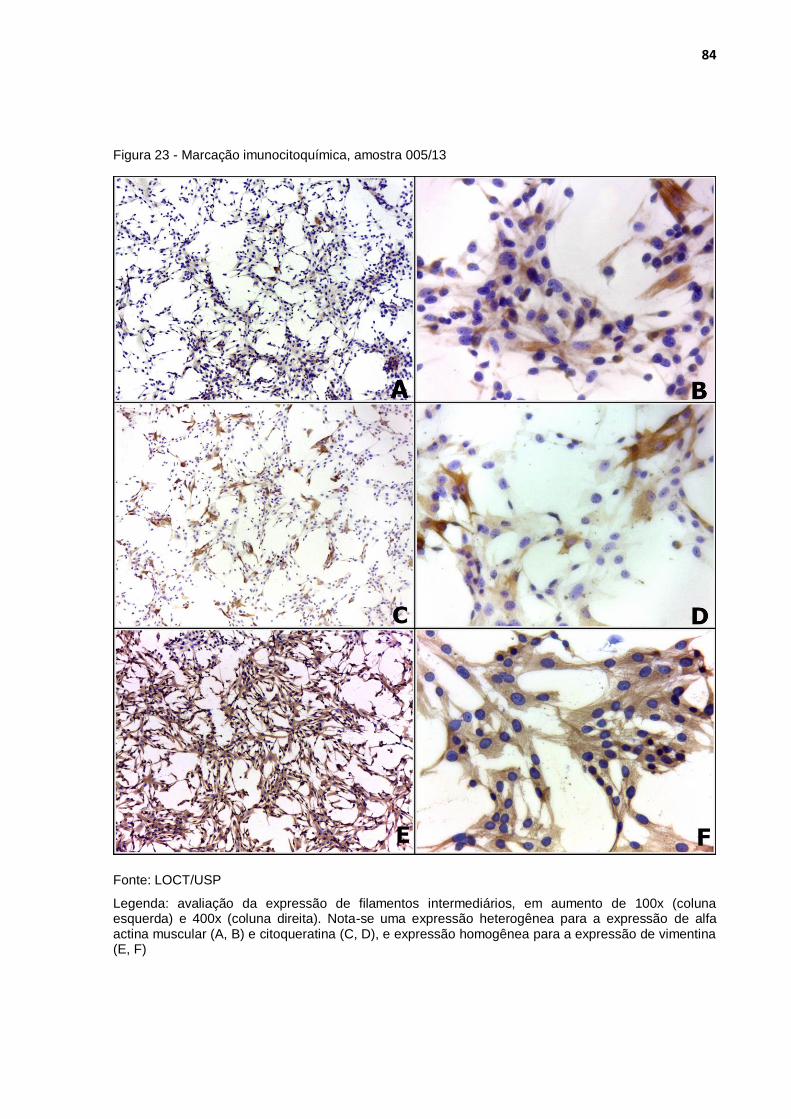
84
Figura 23 - Marcação imunocitoquímica, amostra 005/13
Fonte: LOCT/USP
Legenda: avaliação da expressão de filamentos intermediários, em aumento de 100x (coluna esquerda) e 400x (coluna direita). Nota-se uma expressão heterogênea para a expressão de alfa actina muscular (A, B) e citoqueratina (C, D), e expressão homogênea para a expressão de vimentina (E, F)
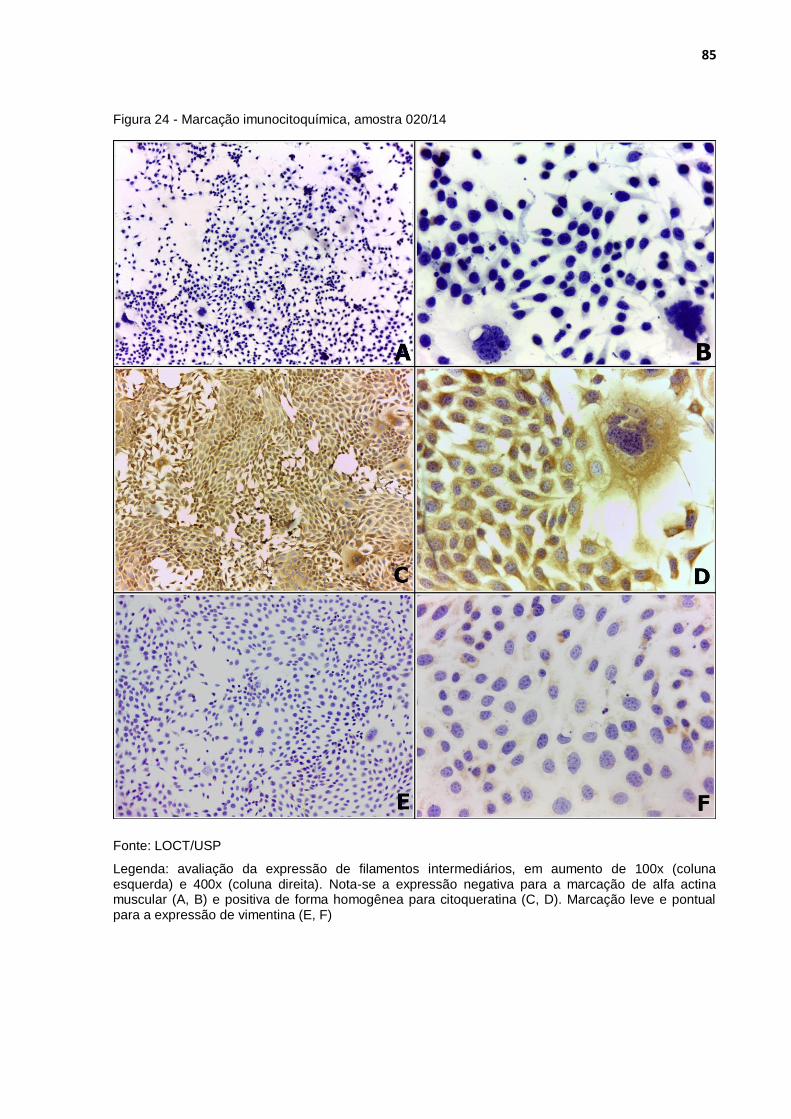
85
Figura 24 - Marcação imunocitoquímica, amostra 020/14
Fonte: LOCT/USP
Legenda: avaliação da expressão de filamentos intermediários, em aumento de 100x (coluna esquerda) e 400x (coluna direita). Nota-se a expressão negativa para a marcação de alfa actina muscular (A, B) e positiva de forma homogênea para citoqueratina (C, D). Marcação leve e pontual para a expressão de vimentina (E, F)
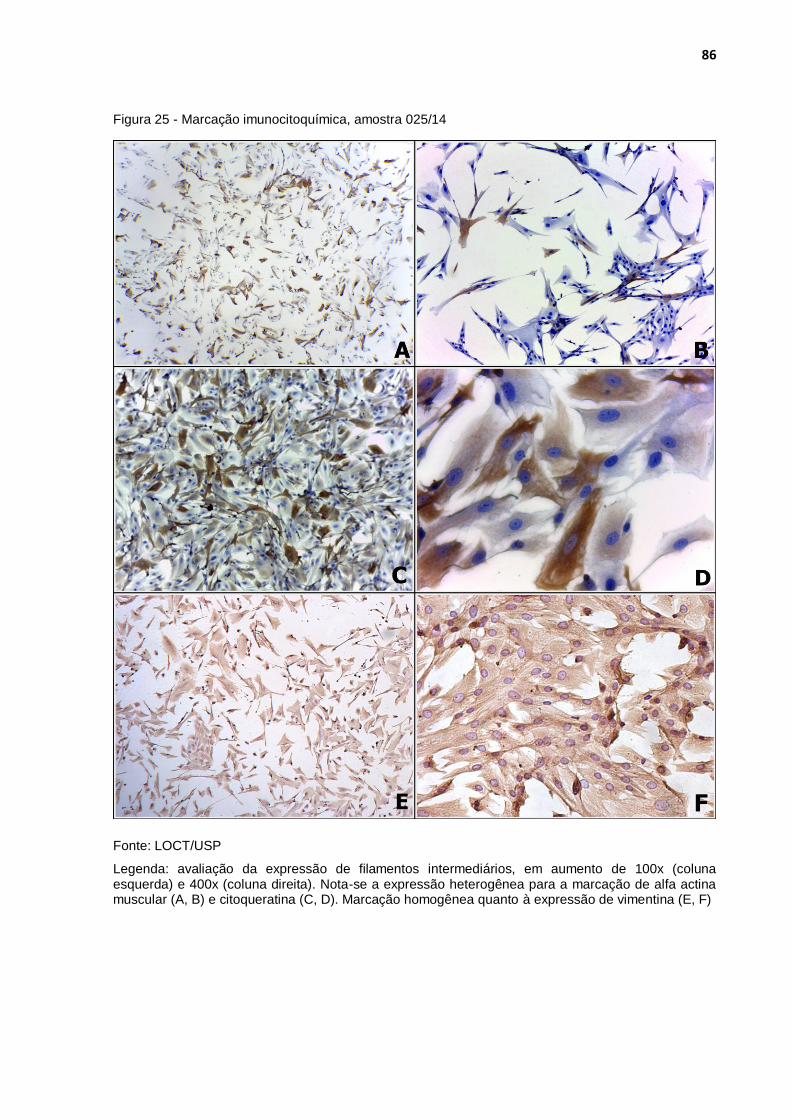
86
Figura 25 - Marcação imunocitoquímica, amostra 025/14
Fonte: LOCT/USP
Legenda: avaliação da expressão de filamentos intermediários, em aumento de 100x (coluna esquerda) e 400x (coluna direita). Nota-se a expressão heterogênea para a marcação de alfa actina muscular (A, B) e citoqueratina (C, D). Marcação homogênea quanto à expressão de vimentina (E, F)
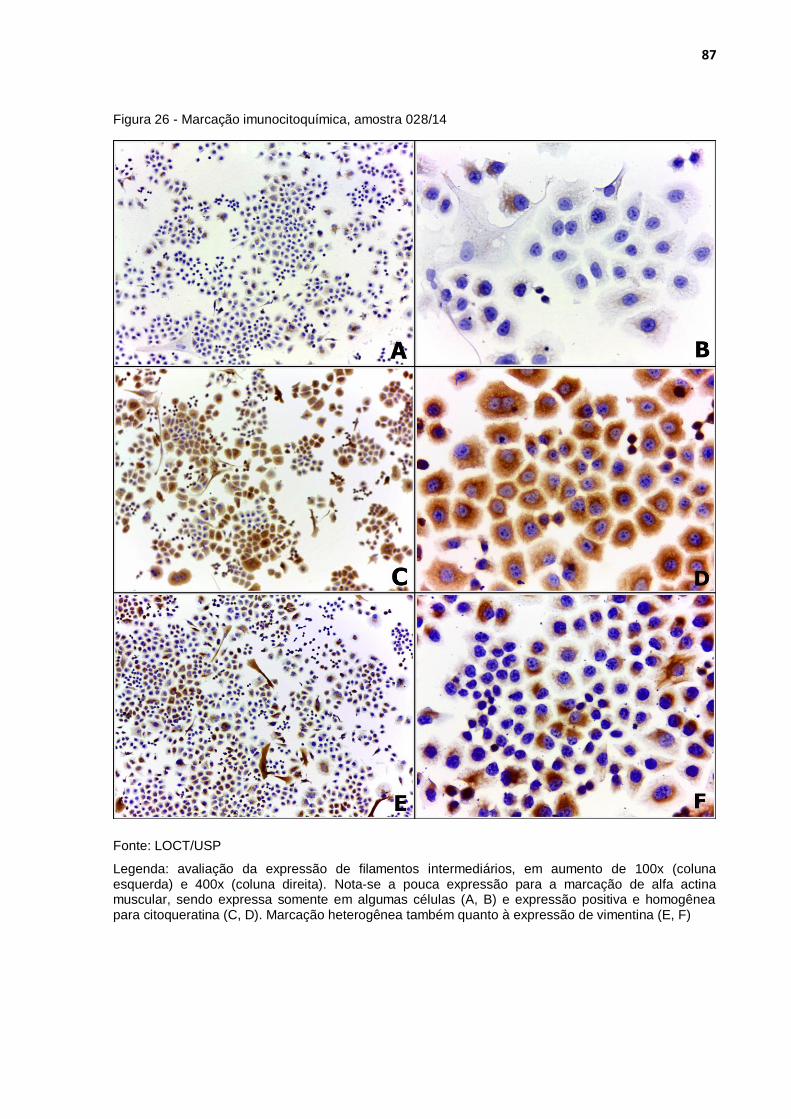
87
Figura 26 - Marcação imunocitoquímica, amostra 028/14
Fonte: LOCT/USP
Legenda: avaliação da expressão de filamentos intermediários, em aumento de 100x (coluna esquerda) e 400x (coluna direita). Nota-se a pouca expressão para a marcação de alfa actina muscular, sendo expressa somente em algumas células (A, B) e expressão positiva e homogênea para citoqueratina (C, D). Marcação heterogênea também quanto à expressão de vimentina (E, F)
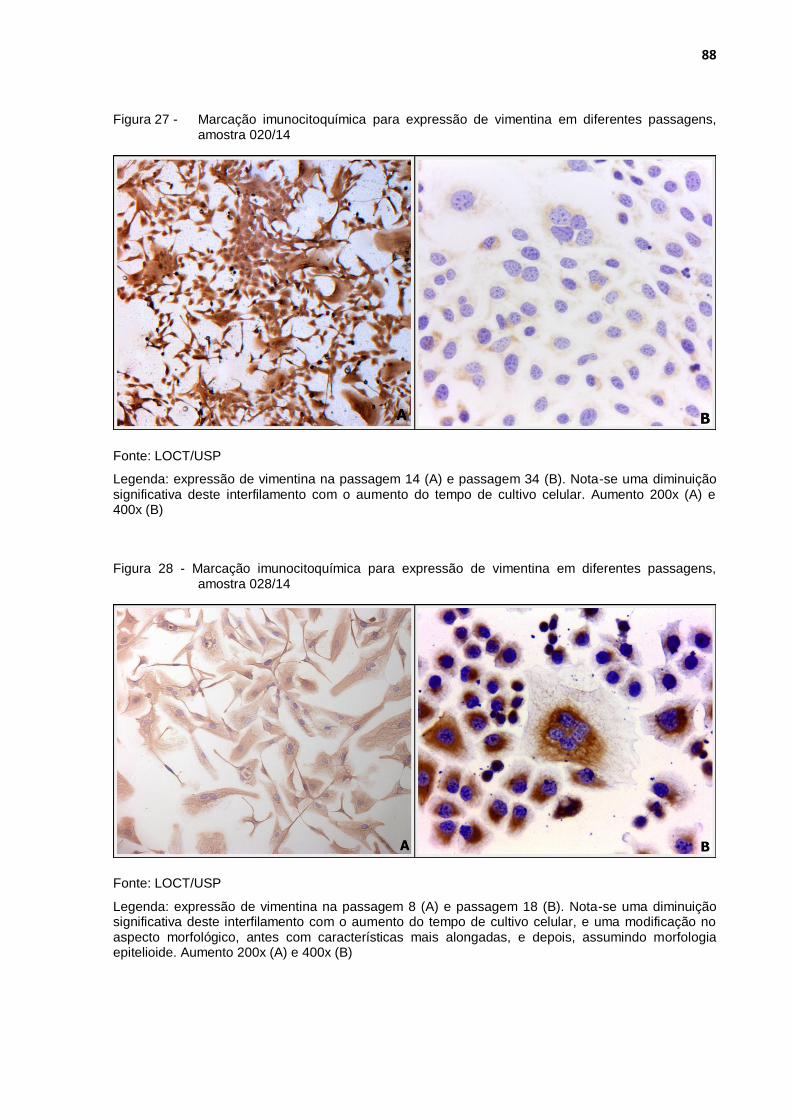
88
Figura 27 - Marcação imunocitoquímica para expressão de vimentina em diferentes passagens, amostra 020/14
Fonte: LOCT/USP
Legenda: expressão de vimentina na passagem 14 (A) e passagem 34 (B). Nota-se uma diminuição significativa deste interfilamento com o aumento do tempo de cultivo celular. Aumento 200x (A) e 400x (B)
Figura 28 - Marcação imunocitoquímica para expressão de vimentina em diferentes passagens, amostra 028/14
Fonte: LOCT/USP
Legenda: expressão de vimentina na passagem 8 (A) e passagem 18 (B). Nota-se uma diminuição significativa deste interfilamento com o aumento do tempo de cultivo celular, e uma modificação no aspecto morfológico, antes com características mais alongadas, e depois, assumindo morfologia epitelioide. Aumento 200x (A) e 400x (B)

89
Nossos dados não permitem a confirmação da origem luminal ou basal das
células obtidas no cultivo primário, uma vez que o clone utilizado para a marcação
de citoqueratina reage tanto para aquelas expressas na camada basal quanto
luminal (CK 5 e 14; e CK 7, 8, 18 e 19, respectivamente). Apenas a expressão
associada de α-sma auxilia na sugestão de que estas células teriam uma origem
basal. Também não foi possível a avaliação da coexpressão de dois ou mais
marcadores em uma mesma célula, uma vez que a técnica utilizada não permite a
dupla marcação. No entanto, pela morfologia de cada cultivo, pressupõe-se que não
se trata de populações distintas em uma mesma cultura, e sim, de uma
heterogeneidade quanto à expressão dos interfilamentos estudados em um mesmo
tipo de população celular.
6.2.6. Separação de populações positivas para a atividade de aldeído-
desidrogenase (ALDH) através de citometria de fluxo
A análise da atividade de ALDH baseada na intensidade de fluorescência de
cada célula exibiu um resultado semelhante entre três das quatro amostras
avaliadas (Figura 29). As populações que apresentaram uma maior atividade de
aldeído-desidrogenase corresponderam a 1,0%, 0,23% e 1,5% para as amostras
005/13, 025/14 e 028/14, respectivamente. Apenas a amostra 020/14 mostrou uma
maior quantidade de células ALDHhi, que corresponderam a 7,2% do total de células
analisadas (Tabela 4).
Tabela 4 - Porcentagens de células ALDH+ nas quatro amostras de carcinomas mamários caninos
005/13 020/14 025/14 028/14
Nº eventos 10000 10000 10000 10000
ALDH+ 102 723 23 148
% total 1,0 7,2 0,23 1,5
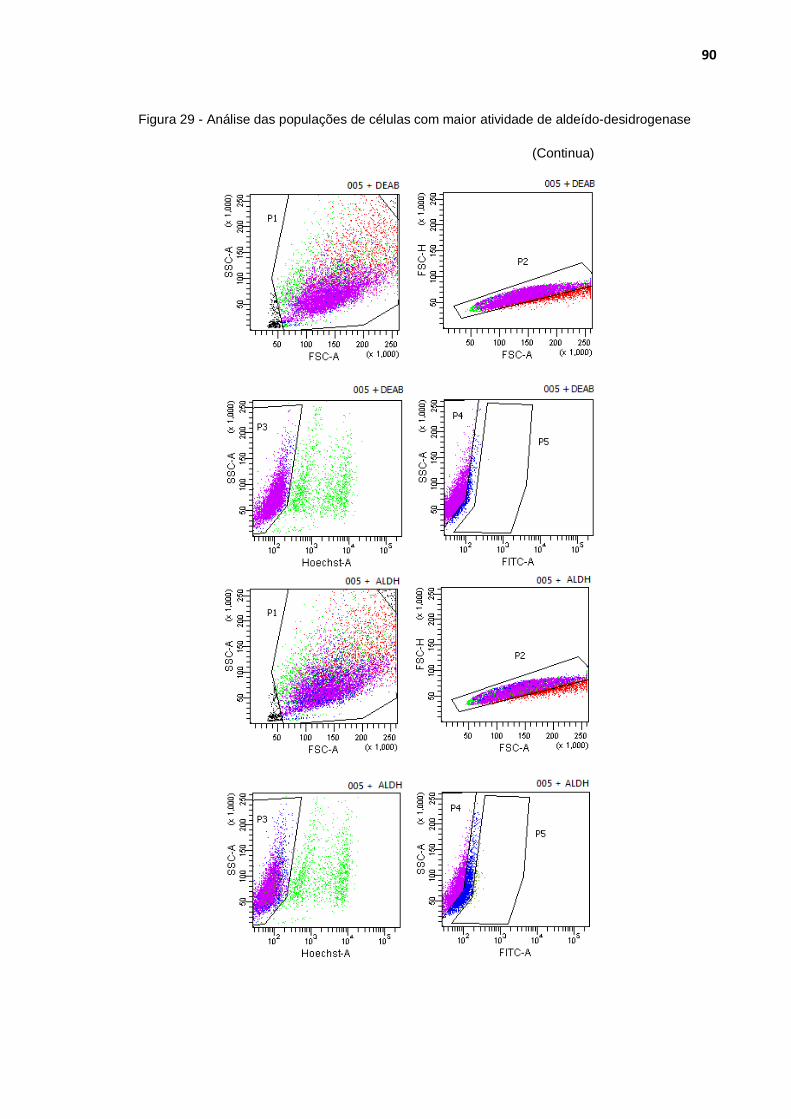
90
Figura 29 - Análise das populações de células com maior atividade de aldeído-desidrogenase
(Continua)
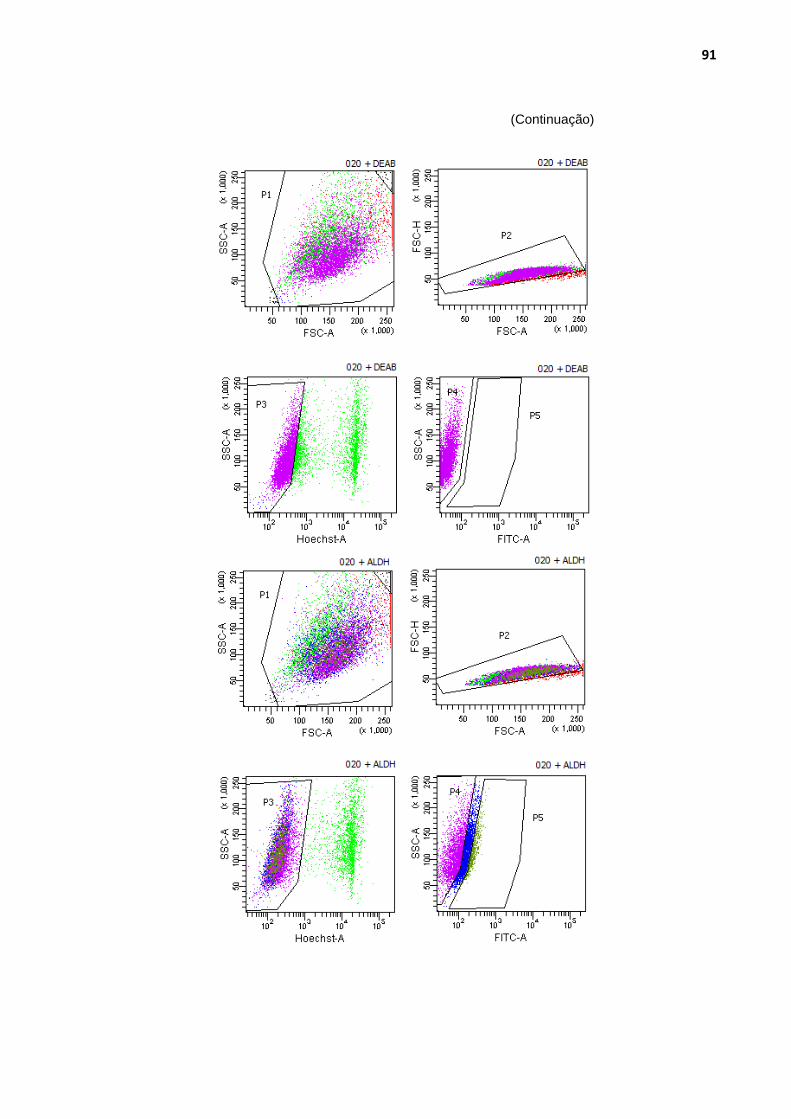
91
(Continuação)
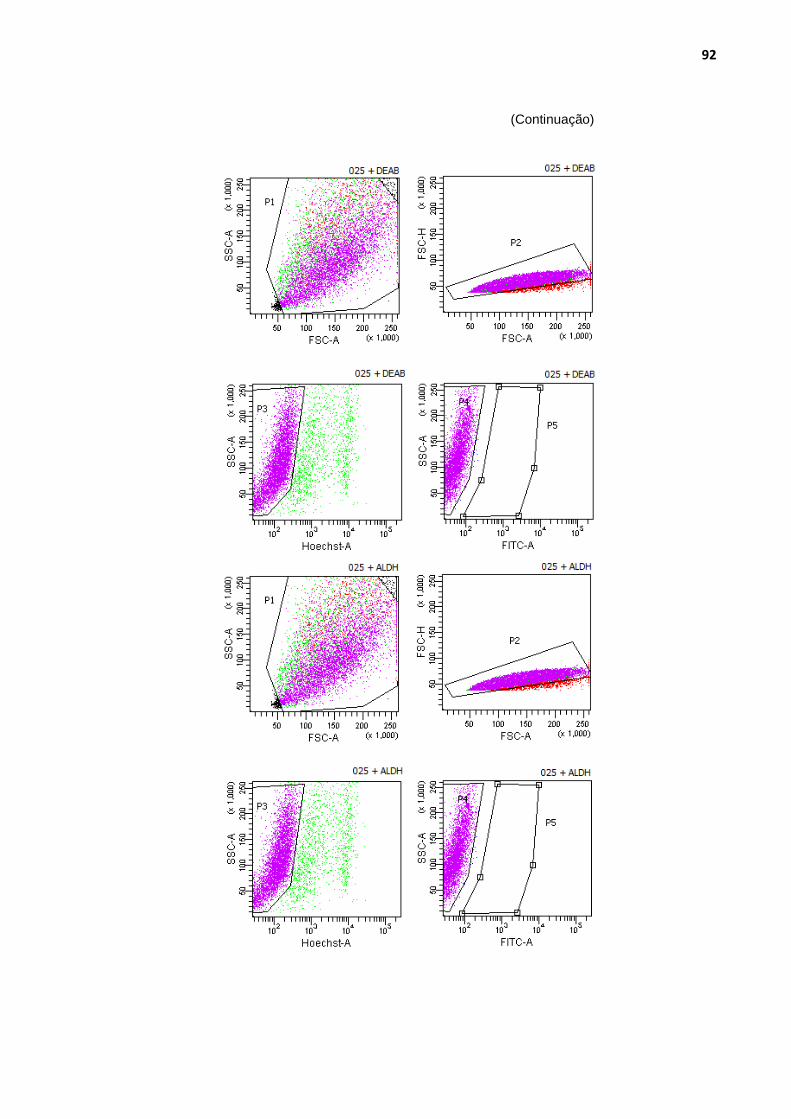
92
(Continuação)
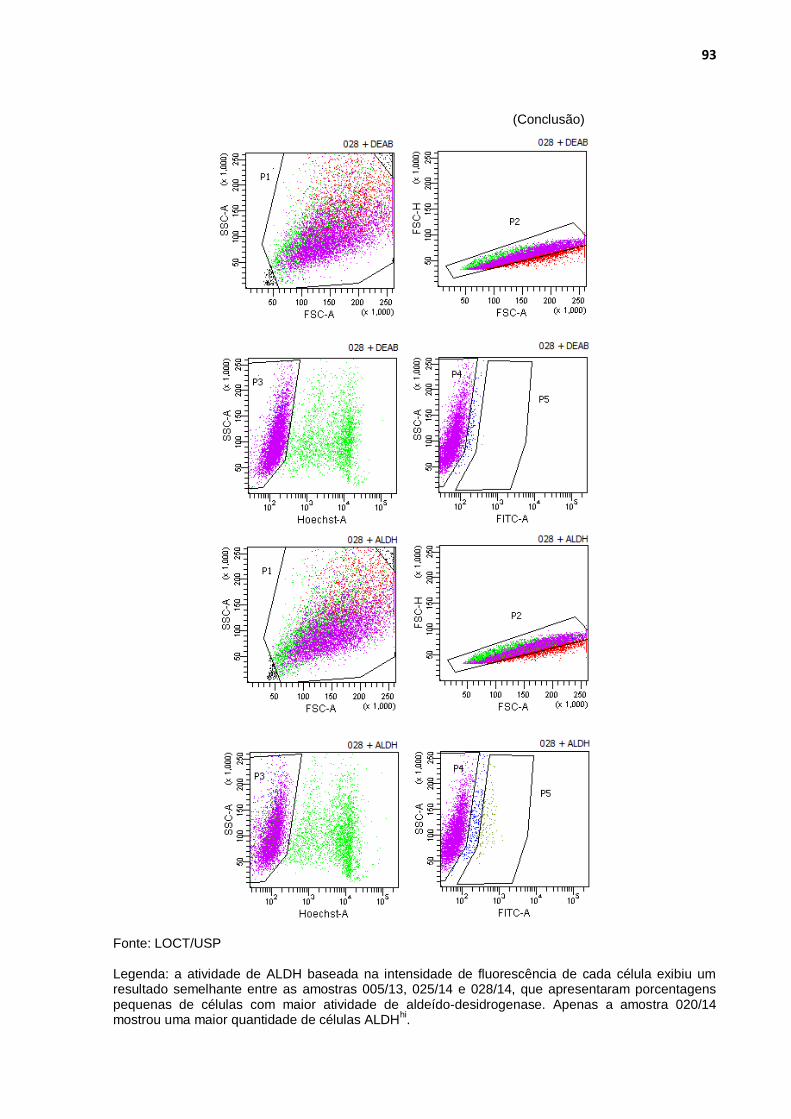
93
(Conclusão)
Fonte: LOCT/USP Legenda: a atividade de ALDH baseada na intensidade de fluorescência de cada célula exibiu um resultado semelhante entre as amostras 005/13, 025/14 e 028/14, que apresentaram porcentagens pequenas de células com maior atividade de aldeído-desidrogenase. Apenas a amostra 020/14 mostrou uma maior quantidade de células ALDH
hi.
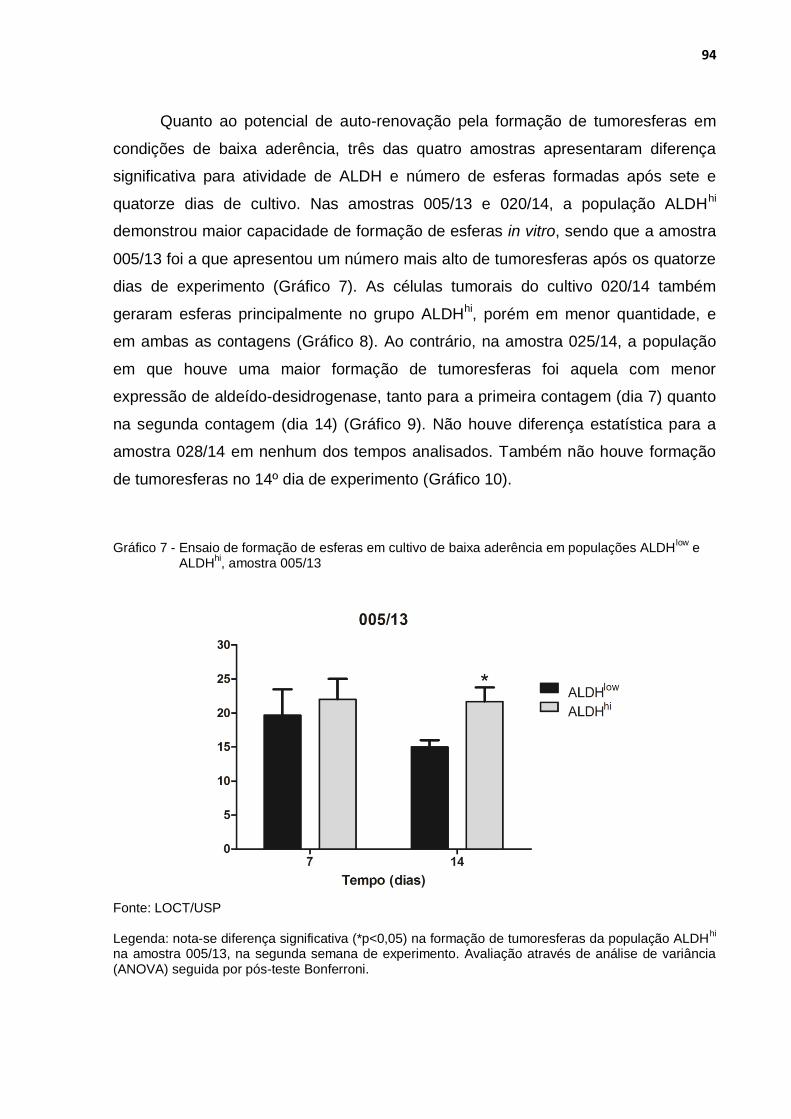
94
Quanto ao potencial de auto-renovação pela formação de tumoresferas em
condições de baixa aderência, três das quatro amostras apresentaram diferença
significativa para atividade de ALDH e número de esferas formadas após sete e
quatorze dias de cultivo. Nas amostras 005/13 e 020/14, a população ALDHhi
demonstrou maior capacidade de formação de esferas in vitro, sendo que a amostra
005/13 foi a que apresentou um número mais alto de tumoresferas após os quatorze
dias de experimento (Gráfico 7). As células tumorais do cultivo 020/14 também
geraram esferas principalmente no grupo ALDHhi, porém em menor quantidade, e
em ambas as contagens (Gráfico 8). Ao contrário, na amostra 025/14, a população
em que houve uma maior formação de tumoresferas foi aquela com menor
expressão de aldeído-desidrogenase, tanto para a primeira contagem (dia 7) quanto
na segunda contagem (dia 14) (Gráfico 9). Não houve diferença estatística para a
amostra 028/14 em nenhum dos tempos analisados. Também não houve formação
de tumoresferas no 14º dia de experimento (Gráfico 10).
Gráfico 7 - Ensaio de formação de esferas em cultivo de baixa aderência em populações ALDHlow
e ALDH
hi, amostra 005/13
Fonte: LOCT/USP Legenda: nota-se diferença significativa (*p<0,05) na formação de tumoresferas da população ALDH
hi
na amostra 005/13, na segunda semana de experimento. Avaliação através de análise de variância (ANOVA) seguida por pós-teste Bonferroni.
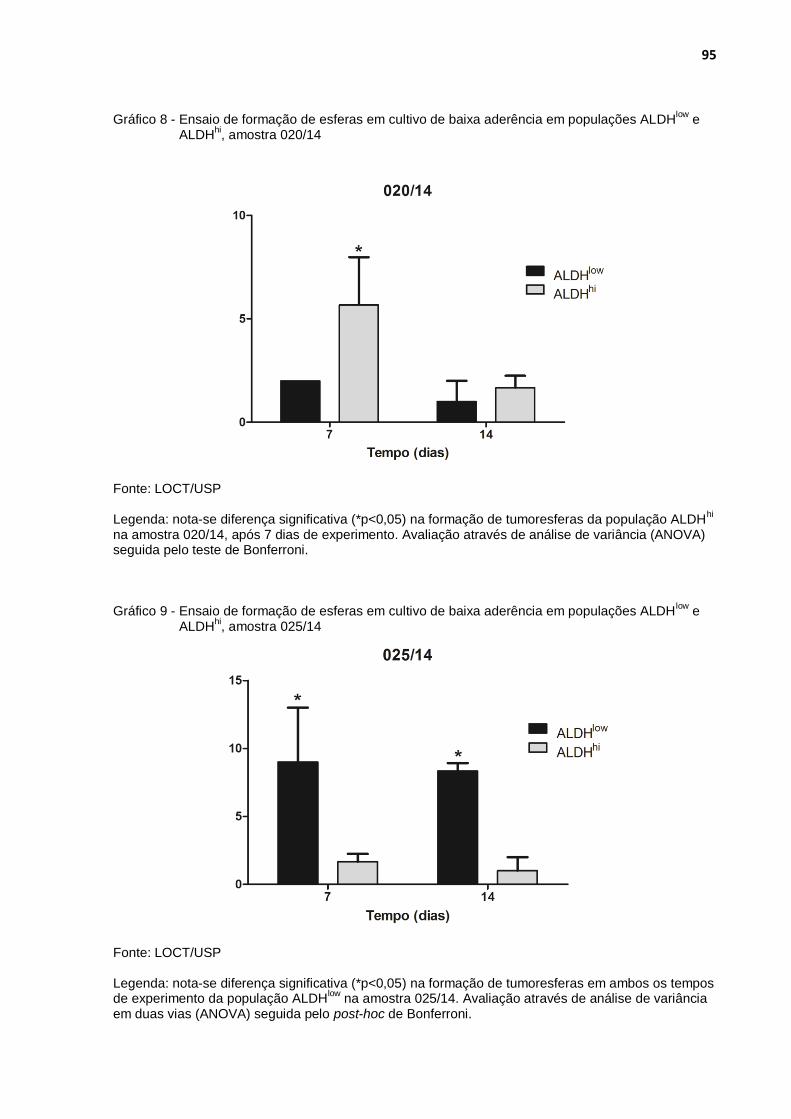
95
Gráfico 8 - Ensaio de formação de esferas em cultivo de baixa aderência em populações ALDHlow
e ALDH
hi, amostra 020/14
Fonte: LOCT/USP Legenda: nota-se diferença significativa (*p<0,05) na formação de tumoresferas da população ALDH
hi
na amostra 020/14, após 7 dias de experimento. Avaliação através de análise de variância (ANOVA) seguida pelo teste de Bonferroni. Gráfico 9 - Ensaio de formação de esferas em cultivo de baixa aderência em populações ALDH
low e
ALDHhi, amostra 025/14
Fonte: LOCT/USP Legenda: nota-se diferença significativa (*p<0,05) na formação de tumoresferas em ambos os tempos de experimento da população ALDH
low na amostra 025/14. Avaliação através de análise de variância
em duas vias (ANOVA) seguida pelo post-hoc de Bonferroni.
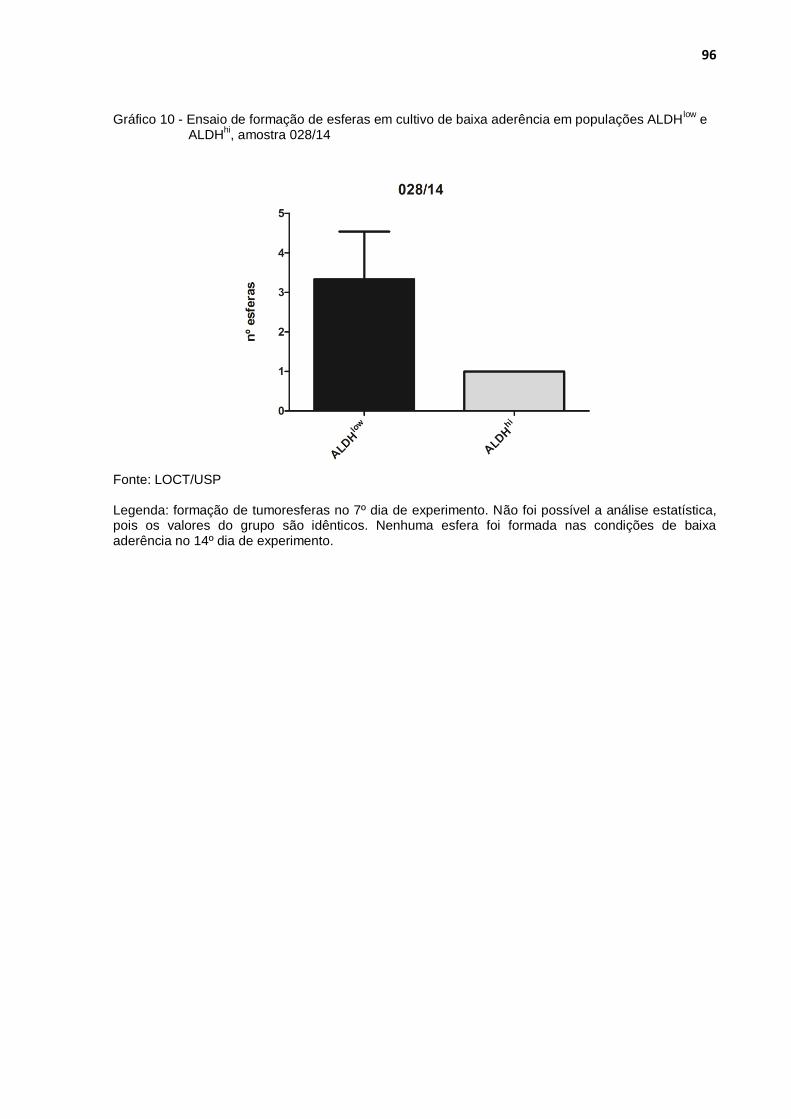
96
Gráfico 10 - Ensaio de formação de esferas em cultivo de baixa aderência em populações ALDHlow
e ALDH
hi, amostra 028/14
Fonte: LOCT/USP Legenda: formação de tumoresferas no 7º dia de experimento. Não foi possível a análise estatística, pois os valores do grupo são idênticos. Nenhuma esfera foi formada nas condições de baixa aderência no 14º dia de experimento.

97
Figura 30 - Formação de tumoresferas, amostra 020/14
Fonte: LOCT/USP Legenda: formação de esferas em condições de baixa aderência para os grupos ALDH
low (-) e ALDH
hi
(+) aos sete e quatorze dias de experimento. Aumento 100x. Escala = 100μm. Figura 31 - Agregados celulares formados a partir do sétimo dia, amostra 028/14
Fonte: LOCT/USP Legenda: amostra 028/14 exibindo aglomerados de células formados nas mesmas condições de crescimento de tumoresferas. Estes grupos não entraram na contagem de esferas ou análise estatística. Aumento 100x. Escala = 100μm

98
A análise da validação para potencial de auto-renovação e formação de
tumoresferas in vitro foi realizada somente pela contagem do número de esferas nos
poços destinados a cultura de cada população, ALDHlow e ALDHhi. Não foram
considerados o tamanho das esferas formadas.
6.2.7. Separação de populações celulares por beads magnéticas através do
perfil de expressão de CD24/CD44
Depois de estabilizado o cultivo celular em monocamada, ou seja, todas as
colônias de células em condições de aderência e em proliferação, as quatro
amostras de carcinomas mamários oriundas da dissociação tumoral foram
submetidas à separação de populações quanto à expressão dos marcadores de
membrana CD24 e CD44. A técnica demonstrou ser eficaz na separação entre
células marcadas e não marcadas para o anticorpo proposto. Todos os testes feitos
obtiveram resultados satisfatórios: porcentagem reduzida de células mortas, e,
aquelas que foram separadas positivamente, permaneceram viáveis quando
colocadas em cultivo novamente. Os resultados mostram que, as quatro amostras
analisadas apresentaram um maior número de células positivas para CD44 quando
comparado à população CD24+. A média de células CD24+ variou entre 0,04 e
0,35% enquanto que as células CD44+ variaram entre 1,75 a 8,56% do total
analisado, correspondendo a mais de seis vezes o número de CD24+. Os dados
estão representados na tabela 5.
Tabela 5 - Quantificação das populações CD24 e CD44 através da separação por beads magnéticas, em neoplasia mamária de cães
(Continua)
CD24 CD44
+ - + -
005/13
Média
0,11
99,88
1,75
98,25

99
(Conclusão)
D.P. 0,07 0,07 1,02 1,02
020/14
Média
0,35
99,65
8,56
91,44
D.P. 0,06 0,06 4,73 4,73
025/14
Média
0,12
99,87
2,42
97,58
D.P. 0,10 0,10 1,60 1,60
028/14
Média
0,04
99,96
2,03
97,97
D.P. 0,04 0,04 1,40 1,40
Fonte: LOCT/USP Legenda: na tabela estão representados a média e o desvio padrão (D.P.) das replicatas analisadas na separação de populações enriquecidas para os fenótipos CD24 e CD44. Os valores apresentados foram ajustados conforme a população de células viáveis.
Ainda, as células separadas após cada experimento foram levadas ao cultivo
celular, para validação quanto à formação de tumoresferas em condições de baixa
aderência (Figura 32). O número de esferas utilizado para análise estatística foi
aquele obtido apenas na contagem realizada no sétimo dia desde o início do
experimento, uma vez que a variação no tempo entre as contagens não teve valor
significativo entre os grupos, portanto, não considerado na presente avaliação.
A análise estatística apresentou uma diferença significativa no crescimento de
tumoresferas para o marcador CD44 nas amostras 020/14 e 025/14, onde a
população CD44+ exibiu um maior número de esferoides no cultivo de baixa
aderência. Para a amostra 005/13, o grupo CD44- foi o que mostrou mais
tumoresferas ao sétimo dia de experimento (Gráfico 12). Também, houve variação
significativa nas populações positivas e negativas para a marcação de CD24, em
que o fenótipo CD24- exibiu maior crescimento de tumoresferas nas amostras
005/13 e 025/14, e o fenótipo CD24+ apresentou crescimento considerável no cultivo
028/14 (Gráfico 11). Não houve diferença estatística entre o número de esferas nas
populações CD24 positiva e negativa para a amostra 020/14, e CD44 positiva e
negativa para a amostra 028/14. Nesta última, o fenótipo CD44+ exibiu um aumento
no crescimento de esferas em uma das replicatas do experimento.
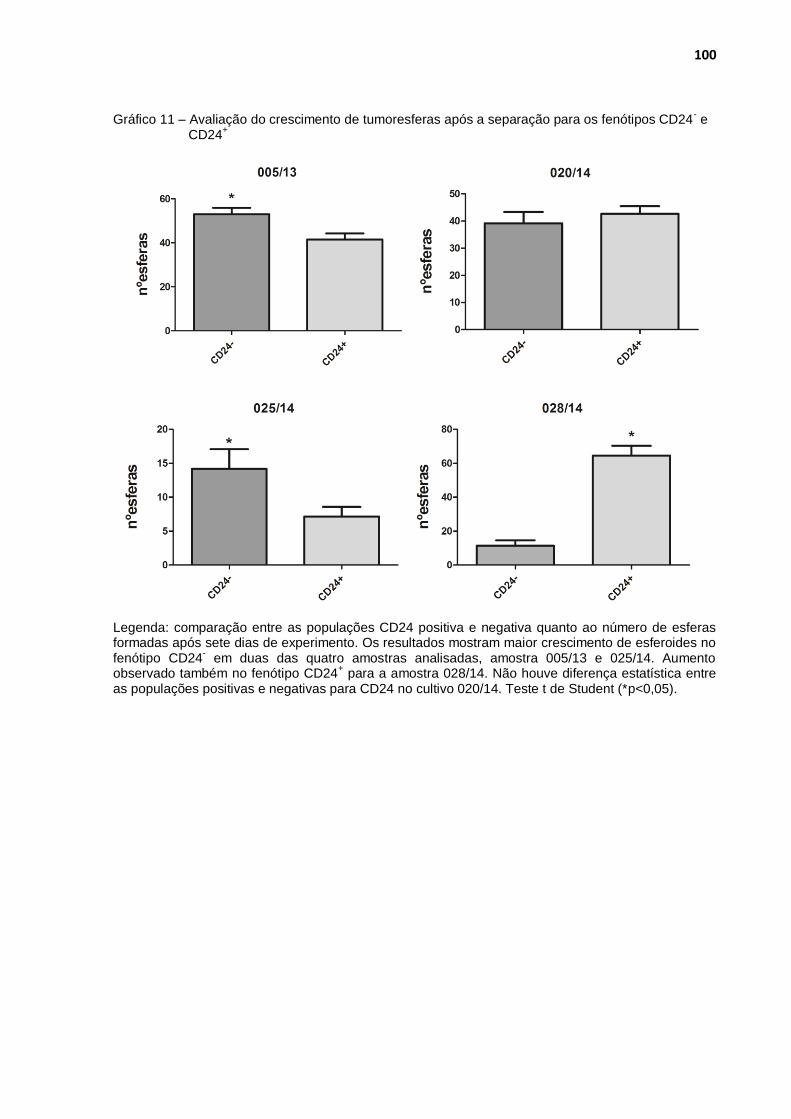
100
Gráfico 11 – Avaliação do crescimento de tumoresferas após a separação para os fenótipos CD24- e
CD24+
Legenda: comparação entre as populações CD24 positiva e negativa quanto ao número de esferas formadas após sete dias de experimento. Os resultados mostram maior crescimento de esferoides no fenótipo CD24
- em duas das quatro amostras analisadas, amostra 005/13 e 025/14. Aumento
observado também no fenótipo CD24+ para a amostra 028/14. Não houve diferença estatística entre
as populações positivas e negativas para CD24 no cultivo 020/14. Teste t de Student (*p<0,05).
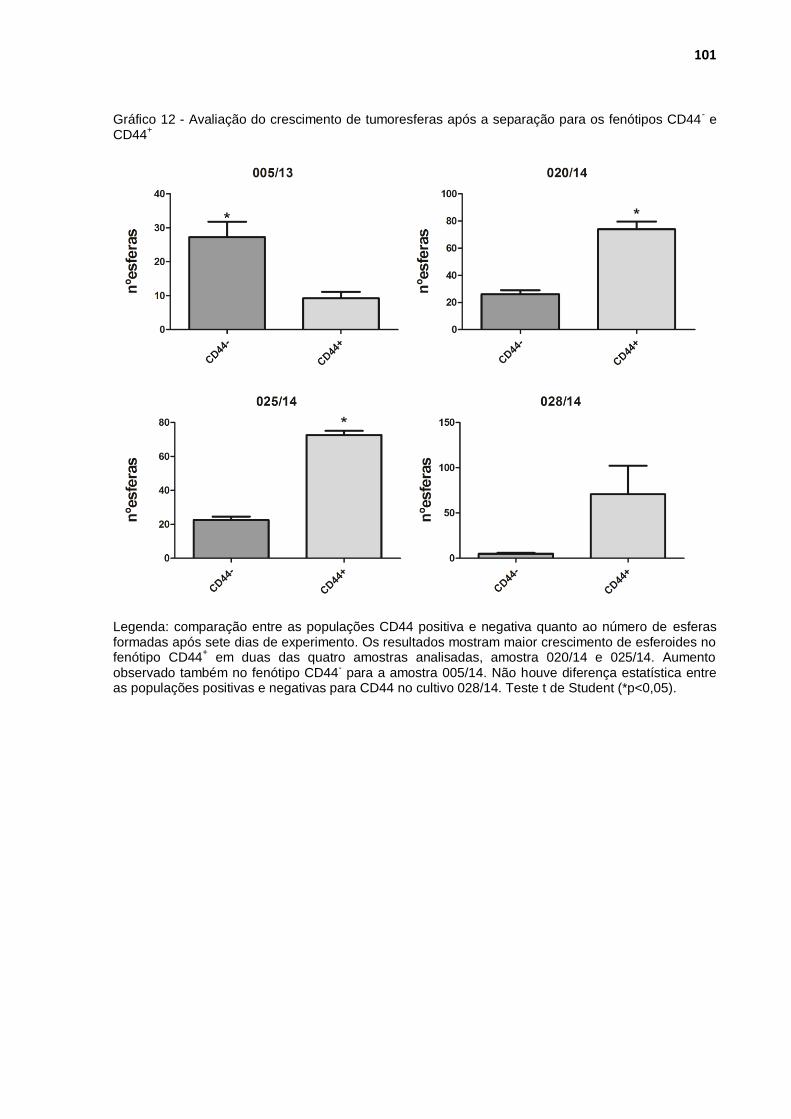
101
Gráfico 12 - Avaliação do crescimento de tumoresferas após a separação para os fenótipos CD44- e
CD44+
Legenda: comparação entre as populações CD44 positiva e negativa quanto ao número de esferas formadas após sete dias de experimento. Os resultados mostram maior crescimento de esferoides no fenótipo CD44
+ em duas das quatro amostras analisadas, amostra 020/14 e 025/14. Aumento
observado também no fenótipo CD44- para a amostra 005/14. Não houve diferença estatística entre
as populações positivas e negativas para CD44 no cultivo 028/14. Teste t de Student (*p<0,05).
102
Figura 32 - Formação de esferas em cultivo de baixa aderência após as separações por beads magnéticas
Fonte: LOCT/USP Legenda: tumoresferas formadas no cultivo de baixa aderência após o sorting magnético para separação de fenótipos CD24 e CD44 positivos e negativos. Não estão especificados os grupos em cada foto, uma vez que todos apresentaram um crescimento semelhante. Não foram considerados os tamanhos das esferas formadas em cada grupo. Aumento 100x. Escala = 100μm

103
Figura 33 - Aglomerado de células em proliferação, amostra 028/14
Fonte: LOCT/USP
Legenda: nota-se um grande agrupamento de células, possivelmente se tratando de células isoladas, e não de um crescimento em esfera propriamente dito. Estes agrupamentos não foram considerados na contagem, e foram observados somente na amostra 028/14. Aumento 100x. Escala = 100μm.
A comparação entre os tamanhos das esferas formadas durante os dias de
experimento não foi realizada na presente análise. Os cultivos foram mantidos
durante quatorze dias ao total, mas a variação entre o número de esferas formadas
entre a primeira (dia 7) e última (dia 14) contagem não foi significativa.

104
7 DISCUSSÃO
As neoplasias mamárias em cadelas ainda é um problema emergente em
países como o Brasil, onde a carência no tratamento, e principalmente, na
prevenção da doença faz com que os índices de incidência e mortalidade ainda
sejam números surpreendentes, ano após ano. Neste estudo, verificamos que as
neoplasias malignas mamárias, em especial os carcinomas, foram as mais
frequentes entre os animais com amostras coletadas no presente trabalho. Assim
como em mulheres, os níveis hormonais exercem grande influência na iniciação,
promoção e progressão tumoral de neoplasias mamárias em cadelas, principalmente
em neoplasias de origem epitelial, onde quase 80% destes tumores são responsivos
ao estrógeno (SORENMO; SHOFER; GOLDSCHMIDT, 2000). Deste modo, animais
de meia idade a idosos, não castrados, e/ou que receberam algum tipo de hormônio
exógeno para controle reprodutivo, são candidatos em potencial ao diagnóstico de
câncer de mama. Baseado neste fato, a castração em animais jovens é considerada
o método mais eficaz para evitar o desenvolvimento deste tipo de neoplasia: se
castrados até o primeiro cio, o risco destas fêmeas virem a desenvolver a doença é
de apenas 0,5%. Porém, se a cirurgia for realizada tardiamente (após o 2º cio), as
chances aumentam para 26% (SCHNEIDER; DORN; TAYLOR, 1969), evidenciando
a necessidade da implementação de programas de conscientização voltados aos
proprietários de fêmeas para a castração precoce de seus animais.
Sendo assim, este acúmulo de fatores de risco durante a vida de cada animal,
não só por fatores hormonais, mas também dieta, meio ambiente, predisposição
genética e acúmulos de mutações, faz com que raramente sejam diagnosticados
tumores de glândula mamária em cães abaixo de 2 anos de idade. A curva de
incidência torna-se ascendente a partir dos 5 anos, atingindo picos entre os 8 a 12
anos de idade, para então entrar em declínio, como evidenciado em estudos
epidemiológicos realizados nos últimos 40 anos (MULLIGAN, 1975; ZATLOUKAL;
KOHOUT; REPUBLIC, 2005; MERLO et al., 2008; FILHO et al., 2010). Resultados
que são congruentes com os verificados neste trabalho. Com relação à
predisposição racial, ainda não se pode afirmar que determinadas raças sejam mais
propensas a desenvolver tumores mamários do que outras, uma vez que este índice

105
pode estar relacionado com a preferência e distribuição racial de cada região
geográfica; porém, há relatos de tendência do aumento de casos de neoplasias de
glândula mamária em raças como Poodle, Pinscher, Cocker Spaniel e Daschund,
frequentes em trabalhos realizados tanto no Brasil como no exterior (SORENMO,
2003; FILHO et al., 2010). Neste estudo, os animais sem raça definida (SRD) foram
os mais acometidos, o que pode ser explicado pelo predomínio de cães mestiços da
região onde foram coletadas as amostras. Estes resultados estão de acordo com
outros estudos brasileiros, que também apresentaram SRDs como a população
dominante dos casos avaliados (FONSECA, 1999; HATAKA, 2004; FILHO et al.,
2010).
Neste estudo, a maior parte dos animais apresentava mais de um nódulo no
momento da cirurgia, corroborando com a literatura, que mostra que fêmeas caninas
podem apresentar múltiplas nodulações, e em alguns casos, estas nodulações
pertencem a tipos histológicos distintos (MOULTON, 1990). A maioria dos casos se
tratava de carcinomas, sendo que os tipos histológicos mais frequentes foram os
carcinomas complexos, mistos e sólidos. Também, a procedência dos cultivos
utilizados no trabalho teve quase que a totalidade de carcinomas mistos, exceto por
um comedocarcinoma, correspondente a amostra 005/13. Os carcinomas mistos
caracterizam-se pela presença de três ou mais populações (epitelial, mioepitelial e
metaplasias cartilaginosas ou ósseas) de células suportadas por um estroma
fibrovascular. Histologicamente, ainda há uma controvérsia sobre a origem do tecido
cartilaginoso e/ou ósseo formado durante o desenvolvimento dos tumores mistos: se
este seria proveniente do componente epitelial (ALLEN, 1940), da metaplasia do
tecido conectivo (PALMER; MONLUX, 1979), ou ainda, das células mioepiteliais que
compõe a camada basal da glândula mamária (TATEYAMA; COTCHIN, 1978).
Porém, evidências reforçam o papel das células mioepiteliais nas metaplasias
mesenquimais, que por algum mecanismo desconhecido, promoveriam a
transformação em cartilagem e osso, resultando numa aparência histológica
heterogênea (GÄRTNER et al., 1999). Já os comedocarcinomas, são neoplasias
extremamente agressivas, que apresentam focos de necrose em meio ao tecido
tumoral e estroma adjacente (GOLDSCHMIDT et al., 2011). Estes focos de necrose
nos tecidos tumorais podem caracterizar uma grande taxa de proliferação das
células neoplásicas, significando clinicamente tumores menos diferenciados e com

106
maior potencial de metastatização, apresentando um pior prognóstico (FRANCO,
1997).
A análise citogenética dos tumores é importante para o diagnóstico,
prognóstico e tratamento de neoplasias humanas e veterinárias (HAHN et al., 1994;
FRIEDLANDER; HEDLEY; TAYLOR, 2006; GRIMWADE et al., 2015). No presente
estudo, as quatro amostras selecionadas para caracterização do cultivo celular in
vitro apresentaram aspectos diferentes quanto ao perfil citogenético, relacionado à
variação do número e aparência dos cromossomos das células avaliadas. O
cariótipo da espécie Canis lupus familiaris é considerado um dos mais complexos e
difíceis de serem estudados, principalmente devido ao tamanho e estrutura de todos
os cromossomos autossômicos, que consistem em 76 cromossomos acrocêntricos
muito semelhantes em comprimento, com exceção do primeiro par, que apresenta
um comprimento parecido com o cromossomo X, sendo facilmente identificado
(REIMANN et al., 1999). Somente em 1996 chegou-se a um consenso sobre a
nomenclatura dos cromossomos caninos, entretanto, as recomendações exatas
sobre o uso de uma nomenclatura específica para a descrição de aberrações
cromossômicas no cariótipo canino ainda não foram alcançadas (REIMANN et al.,
1996).
Alterações numéricas em células tumorais já foram relatadas na espécie
canina. Mayr et. al. realizaram estudos citogenéticos em uma série de neoplasias em
cães. Em hemangiopericitoma, hemangiotelioma e sarcoma espinocelular,
encontraram aneuploidia em grande parte das metáfases analisadas, em
consequência de polissomias, no caso de células hiperdiploides, e fusões cêntricas
ou isocromossomos, reduzindo o número final de cromossomos em células
hipodiploides (MAYR et al., 1994). As fusões cêntricas consistem na união dos
centrômeros de dois cromossomos acrocêntricos distintos dando origem a um
metacêntrico único, levando a uma diminuição de cromossomos no número final da
célula. Mas, apesar disso, indivíduos que apresentam fusões cêntricas geralmente
têm genoma completo e fenótipo normal. Já os isocromossomos, menos frequentes,
surgem a partir de um rompimento longitudinal do centrômero e paralelo às
cromátides, fazendo com que os dois braços idênticos se liguem a um mesmo
centrômero, de modo que cada braço contenha a mesma ordem de genes
(NICHOLAS, 2012). Em um estudo com osteossarcomas, a fusão cêntrica foi um

107
achado comum entre todos os animais estudados, e nenhuma célula apresentou
metáfase com 2n=78. No mesmo trabalho, a alteração do número de cromossomos
entre os pacientes diagnosticados com o mesmo tipo de tumor foi diferente, onde um
animal exibiu células hiperdiploides, e no outro, as células eram todas hipodiploides
(MAYR et al., 1991a). Achados semelhantes foram encontrados em melanomas
orofaríngeos e em um caso de melanoma cutâneo canino. Todos apresentaram
aneuploidias com alterações estruturais características de fusões cêntricas e um
isocromossomo isolado i(12), estando de acordo com os dados também descritos
para melanomas em humanos (MAYR; SCHLEGER; LOUPALT, 1992; HÖRSTING
et al., 1999).
Em neoplasias de glândula mamária em cães, nossos dados mostraram
resultados semelhantes àqueles publicados em literatura. Todas as amostras
analisadas por coloração convencional (Giemsa) exibiram aneuploidas, mas sem um
padrão definido. Com exceção da amostra 028/14, todas apresentaram células
hipodiploides e hiperdiploides, sendo que, na 005/13 e 025/14 a maior parte
correspondia ao valor diploide normal (2n=78). Em relação a alterações estruturais,
a amostra 028/14 foi a que apresentou modificações muito evidentes, como diversos
pares de cromossomos metacêntricos e uma consequente redução no número total
de cromossomos em todas as metáfases analisadas. Os resultados corroboram com
os de Mayr et.al. em sarcomas e adenocarcinomas mamários caninos, realizados
em tumores recém-dissociados e cultivos primários. Entre os achados, estão a fusão
cêntrica ou isocromossomo no par 13, resultando em cromossomos metacêntricos e
simétricos; monossomia dos pares 4 e 6; trissomia no par 3; monossomia do
cromossomo X, entre outros (MAYR et al., 1991b, 1994; MAYR; SCHLEGER;
LOUPALT, 1992). Talvez, o cariótipo do cão suporte a perda de cromossomos
durante o processo de transformação e progressão tumoral sem a perda da
viabilidade celular com mais frequência do que o cariótipo no homem. Segundo
Rutteman et. al., que realizaram análise de ploidia em neoplasias mamárias em
cadelas, os genes que codificam para as funções celulares vitais em cães são
distribuídos em um maior número de cromossomos, permitindo assim que as células
caninas superem a perda cromossômica de uma forma menos significativa
(RUTTEMAN et al., 1988). Mesmo assim, esta perda de cromossomos é um fator

108
importante para a geração de um fenótipo maligno de células, uma vez que podem
implicar a perda de genes supressores de tumor e reparadores de danos no DNA.
Uma vez iniciado, o desenvolvimento e a progressão do tumor mamário será
influenciado por diversos fatores, especialmente de qual local e a que grupo
pertencem as células que deram origem à neoplasia. Normalmente, se originam a
partir de lesões benignas, sendo muito comum um carcinoma ou um sarcoma se
desenvolverem a partir de um tumor misto benigno ou mesmo de lesões pré-
neoplásicas, como as hiperplasias ductais ou lobulares. A possibilidade de se
estabelecer um diagnóstico específico, ou mesmo um marcador prognóstico que
seja expresso nesta fase de transição, seria extremamente bem vinda, visto que
cerca de 70% dos sarcomas e 30% dos carcinomas dão origem a metástases e
formação de tumores secundários (HELLMÉN, 2005).
Os filamentos intermediários (FI) representam um dos três principais
polímeros fibrosos da célula e atuam como suportes do citoesqueleto no núcleo e no
citoplasma (ALBERTS, 2002). São expressos nas células em maior ou menor
quantidade, dependendo do tecido de origem. Até pouco tempo atrás, acreditava-se
que um tumor expressava apenas um tipo de FI; os carcinomas expressariam
apenas citoqueratinas, enquanto que neoplasias mesenquimais teriam a expressão
somente de vimentina. Entretanto, apesar deste padrão de expressão ainda ser uma
tendência, atualmente se sabe que um mesmo tumor pode coexpressar dois ou mais
tipos de filamentos intermediários ao mesmo tempo. Frequentemente, quando há
coexpressão de FIs, a vimentina está presente na maioria dos casos (BROERS et
al., 1988). Neste estudo, verificamos diferentes padrões de expressão dos três
principais FIs utilizados na caracterização de células tumorais: citoqueratinas (CK)
de diferentes pesos moleculares, identificados pelo mesmo clone A1/A3
(especificamente CKs 1, 2, 3, 4, 5, 6, 7, 8, 10, 14, 15, 16 e 19); vimentina; e por fim,
actina de músculo liso, representada pela α-sma. Desde o início, as amostras 005/13
e 025/14, com células de morfologia mais alongada, apresentaram coexpressão de
citoqueratina e vimentina no cultivo celular, sendo a vimentina expressa em todas as
células homogeneamente, enquanto que a citoqueratina expressava de forma
heterogênea. Também, notamos a marcação para α-sma, da mesma forma que a
citoqueratina: distribuição heterogênea. No caso das amostras 020/14 e 028,
verificamos uma diminuição na expressão de vimentina com o aumento das

109
passagens, sendo que nas células da amostra 020/14, assim que o cultivo assumiu
uma morfologia epitelial, não apresentou marcação para este interfilamento. Esta
diminuição na marcação de vimentina pode ser consequência da seleção de uma
população celular durante o cultivo de células, ou do decréscimo da expressão do
interfilamento sem mudanças na natureza do cultivo celular.
As queratinas são uma família de filamentos intermediários expressos
diferencialmente no epitélio, e fornecem marcadores únicos e específicos para
estudos sobre diferenciação epitelial. As células basais e luminais dentro epitélio
mamário expressam queratinas bem definidas, em fenótipos in vitro e in vivo
(TAYLOR; LANE, 1981; O’HARE et al., 1991). A vimentina é um marcador estável de
células mesenquimais, como os fibroblastos, e também já foi descrita como presente
nas células mioepiteliais, que formam a camada basal do epitélio glandular. Além da
vimentina, a expressão de α-actina de músculo liso e a miosina já foram
caracterizadas na expressão de células mioepiteliais tanto em tecido mamário
adulto, como no infantil (GUSTERSON et al., 1982; GUELSTEIN et al., 1988;
GOULD et al., 1990; ANBAZHAGAN et al., 1991). Em humanos, a expressão de FI
na glândula mamária está associada com uma variedade de aspectos, no
desenvolvimento fetal da mama, diferenciação celular pela interferência hormonal, e
a progressão de uma neoplasia (ANBAZHAGAN et al., 1991; OSIN et al., 1998).
Nos últimos anos, tem-se relacionado a coexpressão de vimentina e
citoqueratina em carcinomas mamários com uma maior malignidade tumoral,
principalmente pela associação com a transição epitélio-mesenquimal (EMT)
responsável pela invasão do tecido adjacente e disseminação das células
neoplásicas. O subtipo tumoral que corresponderia a este grau de agressividade
devido às altas taxas de invasão e metástase seria aquele referido como triplo-
negativo, ou basal-like. Desde modo, a EMT a nível celular nestes tumores seria
caracterizada pela perda de adesão intercelular (E-caderina e ocludinas); baixa
regulação dos marcadores epiteliais (citoqueratinas); aumento da regulação de
marcadores mesenquimais (vimentina e α-actina de músculo liso); aquisição de uma
morfologia celular fusiforme, semelhantes a fibroblastos, através da reorganização
do citoesqueleto; e aumento da mobilidade, invasão e capacidade metastática das
células. Boa parte destas características foram verificadas nos estudos
imunocitoquímicos realizados nos cultivos primários deste trabalho, mesmo para

110
aquelas amostras que perderam a expressão de vimentina após o aumento no
número de passagens, como foi o caso da amostra 020/14. Em 2008, Sarrió et. al.
em um estudo com a linhagem celular MCF10, apresentou dados que corroboram
com a hipótese de que as modificações EMT-like acontecem preferencialmente no
subtipo basal do câncer de mama (SARRIÓ et al., 2008). Entretanto, nossos dados
não permitiram a confirmação da origem luminal ou basal das células obtidas no
cultivo primário pela marcação da citoqueratina específica de cada subtipo celular.
Apenas a expressão de alfa-actina muscular presente nas amostras 005/13 e 025/14
sugere que pertençam à camada basal do epitélio.
Grande parte dos estudos que envolvem associação da expressão de IF em
neoplasias mamárias são realizados através de análise imunohistoquímica. Uma vez
em cultivo, as células tendem a modificações no citoesqueleto em resposta às
condições do meio, e de confluência nas placas de cultura. Portanto, os modelos in
vitro que são utilizados para o estudo de EMT sofrem de várias limitações. Em
alguns modelos, a EMT é realizada de forma incompleta ou lenta, levando vários
dias para a conclusão. Poucos tipos celulares de carcinoma com um fenótipo
epitelial bem definido podem realizar a EMT in vitro, talvez porque o processo de
transição seja muito sensível às condições de cultura, incluindo os substratos do
meio e a presença ou ausência de soro fetal bovino (THIERY, 2002). Mesmo assim,
o cultivo de células que apresentam características semelhantes às descritas no
processo de transição epitélio-mesenquimal, naturalmente ou de forma induzida,
ainda é um método de extrema importância para o entendimento de como este
processo estaria envolvido na invasão e metástase de neoplasias agressivas, uma
vez que somente com estudos histológicos não é possível avaliar o comportamento
tumoral acessado através das células vivas, e as modificações as quais estas
células são submetidas, mimetizando a progressão do tumor in vivo.
A heterogeneidade e a complexidade biológica e molecular do câncer,
principalmente de tumores sólidos, como o câncer de mama, apresentam muitos
desafios para o desenvolvimento de estratégias eficazes para prevenir e tratar esta
doença. Por este motivo, estudos vêm dando cada vez mais crédito para a hipótese
das células-tronco cancerosas (CSCs), que, se estiver correta, fornecerá uma
explicação para muitos fatores ainda limitantes no conhecimento do
desenvolvimento e progressão tumoral. O modelo de desenvolvimento baseado em

111
CSCs consiste fundamentalmente na ideia de que o câncer seria originado a partir
de uma desregulação nas vias de auto-renovação de células-tronco e/ou
progenitoras no tecido normal (REYA et al., 2001). Como consequência, as
neoplasias teriam um componente celular chave, que manteria as suas propriedades
de células-tronco, incluindo a auto-renovação, dando início e conduzindo a
carcinogênese e diferenciação, embora aberrante, contribuindo assim para a
heterogeneidade tumoral (GUPTA; CHAFFER; WEINBERG, 2009). A principal
dificuldade encontrada no estudo e caracterização das CSCs, admitindo que esta
teoria seja de fato a origem do câncer, é a identificação de um marcador único e
exclusivo desta população tumoral que permita o isolamento somente das CSCs.
Até hoje, nenhum marcador utilizado para o isolamento das células-tronco normais e
neoplásicas de diversos tecidos são expressos somente pelas células-tronco; isto é
válido não só para marcadores de membrana, conhecidos como CDs (do inglês
Cluster of Differentiation), mas também para ensaios de funcionalidade, como os
níveis de metabolismo de enzimas ou transportadores envolvidos no processo de
detoxificação, como é o caso do ensaio Aldefluor® e Side Population.
O ensaio de formação de esferas, uma técnica in vitro onde a proliferação
celular resulta em um produto de arquitetura esferoide, juntamente com a avaliação
de tumorigênese in vivo é um dos principais métodos de validação da presença de
CSCs em cultivo primário ou linhagens estabelecidas. É muito utilizado em trabalhos
realizados com diversos tipos de tumores sólidos, para a validação de marcadores,
testes de quimioresistência, estudo das vias de transição epitélio-mesenquimal,
entre outros (SINGH et al., 2003; FILLMORE; PONTI et al., 2005; KUPERWASSER,
2008; TROPEPE et al., 2000; WELLNER et al., 2009). No caso de neoplasias
mamárias, as células normais ou tumorais que iniciam as tumoresferas quando
colocadas em condições independentes de ancoragem, têm propriedades de
células-tronco e são capazes de auto-renovação in vitro, bem como de se diferenciar
em todos os subtipos celulares encontrados na glândula mamária (PONTI et al.,
2005; FILLMORE; KUPERWASSER, 2008; KAKARALA; WICHA, 2008;).
No presente estudo, as quatro amostras de células obtidas a partir do cultivo
primário de carcinomas mamários em cães apresentaram formações em
tumoresferas no cultivo de baixa aderência. Existem poucos trabalhos publicados na
literatura que descrevem o isolamento e caracterização de CSCs em tumores

112
mamários caninos, sendo a maior parte realizada em linhagens pré-estabelecidas e
não em cultivos primários, sendo que o primeiro deles a isolar esta subpopulação foi
realizado em 2009. Neste trabalho, Cocola et. al. demonstraram que tanto as células
dissociadas de tecidos tumorais quanto saudáveis foram capazes de gerar esferas
sob as condições adequadas, sem observar diferença morfológica entre os dois
grupos (COCOLA et al., 2009). A definição de um marcador que otimize a separação
da população de CSCs em neoplasias mamárias em cães, entretanto, ainda
permanece indefinida. Mesmo para aqueles marcadores que já foram bem
documentados em estudos humanos, como é o caso do fenótipo CD24-
/low/CD44+/high, a marcação CD133+, a maior expressão de aldeído-desidrogenase e o
isolamento pelo método Side Population (AL-HAJJ et al., 2003; PATRAWALA et al.,
2005; PONTI et al., 2005; GINESTIER et al., 2007; CHARAFE-JAUFFRET et al.,
2009), nos tumores caninos, apresentou uma diferença nos resultados não só
quando comparados à literatura humana, mas também entre os animais de um
mesmo estudo.
Por exemplo, nossos dados mostram que, duas em quatro amostras
apresentaram um maior crescimento de esferas no grupo ALDHhi, como foi o caso
das amostras 005/13 e 020/14; já no cultivo 025/14, a maior expressão de aldeído-
desidrogenase não foi um fator determinante no ensaio em baixa aderência. Estes
resultados corroboram com o estudo de Michishita et. al. realizado em 2012, que
avaliaram o potencial de auto-renovação em linhagens de carcinomas mamários
caninos. Três das quatro amostras analisadas apresentaram uma maior formação de
esferas na população ALDHhi, enquanto que uma linhagem não apresentou
diferenças significativas entre os grupos ALDHlow e ALDHhi. Também, não houve
alteração na expressão gênica de ALDH1A1 entre a população positiva e negativa
da linhagem que apresentou uma maior porcentagem de expressão no ensaio
Aldefluor® (MICHISHITA et al., 2012). Em outro estudo, realizado em linhagens de
melanoma humano isoladas de sítios metastáticos, com elevado grau de
agressividade, a maior atividade de ALDH foi encontrada em quase 90% das células
avaliadas, porém sem diferenças na tumorigenicidade entre as populações ALDHlow
e ALDHhi quando da injeção em camundongos nude (PRASMICKAITE et al., 2010).
Em 2006, um Workshop realizado pela American Association of Cancer
Research, e publicado por Michael Clarke et. al. sobre o tema “Cancer Stem Cells —

113
Perspectives on Current Status and Future Directions” afirmava que nesta teoria de
desenvolvimento tumoral, as CSCs seriam uma subpopulação muito pequena,
apenas um reservatório de células que teriam como função a manutenção do tumor
(CLARKE et al., 2006). No entanto, demonstrou-se através dos resultados obtidos
nos experimentos publicados até 2009 sobre as CSCs, que a proporção de CSCs
pode diferir entre os vários tipos de tumores (GUPTA; CHAFFER; WEINBERG,
2009). Esta variação também pôde ser encontrada na comparação entre os
resultados do presente trabalho, e o estudo de Michishita et. al. comentado
anteriormente. Nossos dados apresentaram uma variação na expressão de ALDH
partindo de 0,23%, na amostra 025/14, a 7,2% na amostra 020/14, enquanto que
nos resultados publicados na literatura, a porcentagem de células com alta
expressão de ALDH variou de 5,1% a 49,1% entre as amostras analisadas
(MICHISHITA et al., 2012).
Além da expressão de ALDH, a maioria dos estudos atuais tem enfatizado a
identificação de subpopulações de células-tronco cancerosas em câncer de mama a
partir da expressão de marcadores de membrana como CD44 e CD24. O primeiro
relato de isolamento de uma subpopulação com alto potencial tumorigênico verificou
que células com expressão do fenótipo CD24- ou CD44+ apresentavam um maior
crescimento de tumores em camundongos NOD/SCID do que aquelas CD24+ e
CD24-. Ainda, quando associadas as duas marcações, o grupo CD24-/CD44+ era
capaz de formar nódulos com uma menor quantidade de células injetadas, do que
quando avaliados de forma isolada (AL-HAJJ et al., 2003). A partir de então, vários
trabalhos em humanos e cães se basearam na expressão destes marcadores na
caracterização desta subpopulação como possíveis células-tronco cancerosas
mamárias (DONTU et al., 2003; PONTI et al., 2005; FILLMORE; KUPERWASSER,
2008; FERLETTA; GRAWÉ; HELLMÉN, 2011). No presente estudo, procuramos
avaliar a capacidade na formação de tumoresferas em células de cultivo primário de
carcinomas mamários caninos, não só através da atividade de ALDH, mas também a
fim de verificar se a expressão de CD24 ou CD44 poderia selecionar subpopulações
enriquecidas com CSCs no ensaio in vitro. O método escolhido foi a separação
através de beads magnéticas, que apesar de menos frequente para este tipo de
estudo, já foi utilizado em literatura no isolamento de populações CD24 e CD44

114
positivas e negativas a partir de células tumorais para análises de CSCs e
invasividade (LEELAWAT et al., 2013; SHEN et al., 2013).
Assim como os resultados obtidos a partir da expressão de ALDH, o
crescimento de esferas no cultivo de baixa aderência diferiu entre os fenótipos
avaliados. Duas em quatro amostras apresentaram crescimento significativo na
população CD44+ quando comparada às demais, e, na amostra 025/14, o grupo
CD24- também exibiu maior número de tumoresferas no cultivo de baixa aderência,
conforme o esperado. A expressão de CD44 em células tumorais caninas talvez
esteja intimamente associada à proliferação celular em cultura, sendo que essa
expressão transitória e flutuante pode interferir na sua utilidade como um marcador
de CSCs (BLACKING; WATERFALL; ARGYLE, 2011). Esta discrepância na
expressão e potencial de formação de colônias in vitro foi relatada em linhagens
tumorais de neoplasias mamárias em humanos, bem como na avaliação da
expressão de marcadores comuns de CSCs em células dissociadas a partir de
tumores primários. Verificou-se que a expressão de CD44, CD24, ALDH, ESA,
CD133, CXCR4 e PROCR tanto em in vivo quanto in vitro, diferiu substancialmente
entre as linhagens de células pré-estabelecidas de câncer de mama, bem como nos
tumores primários, sugerindo que os marcadores previamente reconhecidos para as
células-tronco do câncer de mama podem não ser os marcadores ideais ou
universais para a identificação de populações de células-tronco do câncer nos
diversos tipos histológicos reconhecidos (HWANG-VERSLUES et al., 2009).
Uma questão fundamental é se os diferentes subtipos histológicos do câncer
de mama são derivados de uma mesma origem, ou de origens distintas. Células-
tronco cancerosas com características distintas em cada subtipo poderiam explicar
porque eles diferem não só morfologicamente, mas também em graus de invasão e
metástase, bem como no prognóstico e resposta ao tratamento de cada neoplasia. A
expressão de ALDH, por exemplo, já foi relacionada a um alto grau de agressividade
do tumor, sem necessariamente implicar em um maior crescimento de tumoresferas
no grupo ALDHhi para todos os tipos de neoplasias (PRASMICKAITE et al., 2010),
provavelmente devido à função específica de cada isoforma resultante da expressão
do gene ALDH. No câncer de mama humano, já foram descritos 19 isoformas
diferentes de ALDH, em que a progressão tumoral em diferentes subtipos tumorais
era influenciada não só por uma isoforma específica; e cada uma delas teria um

115
potencial diferente para promover um aumento na atividade do teste Aldefluor®, se
expressas em níveis suficientes (MARCATO et al., 2011). Associando o fato de que
a maior atividade de ALDH teria uma grande influência principalmente em subtipos
mais agressivos, e de que diferentes isoformas agem de maneiras distintas na
progressão de cada subtipo tumoral, a literatura daria suporte aos dados
encontrados neste trabalho; a amostra 005/13, que apresentou um maior número de
tumoresferas no grupo ALDHhi, pertence a um subtipo tumoral mais agressivo, o dos
comedocarcinomas. Assim, mesmo sem expressar uma grande porcentagem de
células com alta atividade de ALDH, aquelas que de fato expressavam, promoveram
um crescimento significativo de esferas quando colocadas em condições
específicas. Ainda, sabe-se que uma das funções da enzima aldeído-desidrogenase
é a conversão do retinol em ácido retinóico, que tem papel importante na
diferenciação celular de células-tronco neurais e hematopoiéticas (MARCATO et al.,
2011). Portanto, uma baixa atividade de ALDH, seja fisiológica ou induzida, levaria a
uma diminuição nos níveis de ácido retinóico e um consequente aumento de células
indiferenciadas, tais como as células-tronco. Esta função permitiu o desenvolvimento
de um fármaco eficaz em casos de leucemia aguda, o all-trans-retinoic-acid (ATRA),
que promove diferenciação das células tumorais e induz apoptose. Em câncer de
mama, Ginestier et. al. demonstraram que a adição de ATRA em cultivos de células
tumorais mamárias leva a uma diferenciação celular in vitro, observada através da
diminuição na formação de tumoresferas (GINESTIER et al., 2009).
A variação encontrada no crescimento de esferas em populações separadas
a partir de marcadores comuns de células-tronco cancerosas mamárias em
humanos nos permite pensar que a comparação entre CSCs e não-CSCs talvez não
seja a melhor forma de finalmente caracterizarmos esta população como sendo uma
população única; talvez, estejamos diante não só de diferentes populações de
células tumorais mamárias. Mas também, de diferentes grupos de células iniciadoras
tumorais, que levariam a diferentes subtipos de câncer de mama, e que estas
células poderiam às vezes até coexistir dentro de um mesmo tumor. Neste estudo
verificamos que neoplasias de uma mesma classificação histológica, no caso três
adenocarcinomas mistos, diferiram nas características morfológicas e citogenéticas
avaliadas, bem como na expressão de marcadores normalmente utilizados no
isolamento de CSCs em câncer de mama, apesar de todos os cultivos terem sido

116
mantidos sob as mesmas condições de meio de cultura e processamento de
dissociação primária. Assim, populações celulares diferentes seriam responsáveis
pela formação e desenvolvimento dos tumores sólidos. A definição da forma com
que acontece a progressão neoplásica permitiria o aprimoramento de terapias e
estratégias de prevenção da doença.
Os resultados obtidos no presente trabalho confirmam a possibilidade do uso
da oncologia comparada na pesquisa em oncologia básica, em busca da
compreensão da origem, desenvolvimento e organização dos tumores sólidos,
especialmente em neoplasias mamárias. O câncer de mama em cães, que
apresenta uma grande variedade e complexidade assim como em humanos, se
apresenta como uma ferramenta em potencial para o estudo das vias hierárquicas
do desenvolvimento tumoral, uma vez que é uma neoplasia espontânea e
biologicamente semelhante àquelas diagnosticadas em mulheres.

117
8 CONCLUSÃO
. A partir da caracterização citogenética e imunocitoquímica, bem como das
observações realizadas pelos ensaios de formação de esferas, conclui-se que:
• Cultivos de células tumorais provenientes de neoplasias mamárias em cadelas
podem ser padronizados in vitro, em cultivos de longo prazo, sendo excelentes
métodos para o estudo citogenético envolvendo alterações a nível
cromossômico, e também, para a análise da expressão de filamentos
intermediários in vitro;
• Cultivos celulares diferentes apresentam quantidades distintas de populações
positivas e negativas quanto à expressão de marcadores comuns de células-
tronco cancerosas em câncer de mama;
• Apesar de apresentarem respostas diferentes, todos os cultivos celulares
exibiram potencial tumorigênico através da análise in vitro.

118
REFERÊNCIAS
ACS, IARC, U. Human Development Index Transitions . Disponível em: <http://canceratlas.cancer.org/the-burden/human-development-index-transitions/>. Acesso em: 24 dez. 2014.
ADAMSKI, D. MAYOL, J-F; PLATET, N; BERGER, F; HÉRODIN, F; WION, D. Effects of Hoechst 33342 on C2C12 and PC12 cell differentiation. FEBS letters , v. 581, n. 16, p. 3076–80, 26 jun. 2007.
AGUIRRE-GHISO, J. A. Models, mechanisms and clinical evidence for cancer dormancy. Nature reviews. Cancer , v. 7, n. 11, p. 834–46, nov. 2007.
ALBERTS, B. Molecular biology of the cell . 4. ed. New York: Garland Science, 2002. p. 907–982
AL-HAJJ, M. WICHA, M S; BENITO-HERNANDEZ, A; MORRISON, S. J; CLARKE, M F. Prospective identification of tumorigenic breast cancer cells. Proceedings of the National Academy of Sciences of the United Stat es of America , v. 100, n. 7, p. 3983–8, 1 abr. 2003.
AL-HAJJ, M.; CLARKE, M. F. Self-renewal and solid tumor stem cells. Oncogene , v. 23, n. 43, p. 7274–82, 20 set. 2004.
ALLEN, A. C. So-Called Mixed Tumors of the Mammary Gland of Dog and Man. With Special Reference to the General Problem of Cartilage and Bone Formation. Archives Pathology , v. 29, p. 589–624, 1940.
AMERICAN VETERINARY MEDICAL ASSOCIATION. U.S. Pet Ownership & Demographics Sourcebook . USA: 2014
ANBAZHAGAN, R; BARTEK, J; MONAGHAN, P; GUSTERSON, B A. Growth and development of the human infant breast. The American journal of anatomy , v. 192, n. 4, p. 407–17, dez. 1991.
ANISIMOV, V. N.; UKRAINTSEVA, S. V; YASHIN, A. I. Cancer in rodents: does it tell us about cancer in humans? Nature reviews. Cancer , v. 5, n. 10, p. 807–19, out. 2005.
ANTINOFF, N.; HAHN, K. Ferret oncology: diseases, diagnostics, and therapeutics. Veterinary Clinics of North America: Exotic Animal Practice , v. 7, n. 3, p. 579–625, 2004.
ARPINO, G. BARDOU, V J; CLARK, G M; ELLEDGE, R M. Infiltrating lobular carcinoma of the breast: tumor characteristics and clinical outcome. Breast cancer research : BCR , v. 6, n. 3, p. R149–56, jan. 2004.
ART, J. M. A Prospective Analysis of Immunohistochemically Determined Estrogen Receptor and Progesterone Receptor Expression and Host and Tumor Factors as Predictors of Disease-free Period in Mammary Tumors of the Dog. Veterinary Pathology Online. v. 212, p. 200–212, 2005.
BECK, B.; BLANPAIN, C. Unravelling cancer stem cell potential. Nature reviews. Cancer , v. 13, n. 10, p. 727–38, out. 2013.

119
BEHBOD, F. et al. Transcriptional profiling of mammary gland side population cells. Stem cells (Dayton, Ohio) , v. 24, n. 4, p. 1065–74, abr. 2006.
BERNAL, S. D.; STAHEL, R. A.; BAYLIN, S. B. Cytoskeleton-associated proteins: their role as cellular integrators in the neoplastic process. Critical Reviews in Oncology/Hematology , v. 3, n. 3, p. 191–204, jan. 1985.
BLACKING, T. M.; WATERFALL, M.; ARGYLE, D. J. CD44 is associated with proliferation, rather than a specific cancer stem cell population, in cultured canine cancer cells. Veterinary immunology and immunopathology , v. 141, n. 1-2, p. 46–57, 15 maio 2011.
BONNET, D.; DICK, J. E. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nature Medicine , v. 3, n. 7, p. 730–737, 1997.
BROERS, J. L. V; ROT, M. K; OOSTENDORP, T.; BEPLER, G.; LEIJE, L.; CARNEY, D. N.; VOOIJIS, P.; RAMAERKES, F. C. S. Spontaneous change in intermediate filament proteins expression patterns in lung cancer cell. Journal of Cell Science , v. 91, p. 91–108, 1988. CASSALI, G. D.; GOBBI, H; MALM, C; SCHMITT, F C. Evaluation of accuracy of fine needle aspiration cytology for diagnosis of canine mammary tumours: comparative features with human tumours. Cytopathology , v. 18, n. 3, p. 191–196, 2007. CHARAFE-JAUFFRET, E. L; GINESTIER, C; IOVINO, F; WICINSKI, J; CERVERA, N; FINETTI, P; HUR, M; DIEBEL, M; MONVILLE, F; DUTCHER, J; BROWN, M; VIENS, P; XERRI, L; BERTUCCI, F; STASSI, G; DONTU, G; BIRNBAUM, D; WICHA, M. Breast cancer cell lines contain functional cancer stem cells with metastatic capacity and a distinct molecular signature. Cancer research , v. 69, n. 4, p. 1302–13, 15 fev. 2009. CHUTE, J. P. MURAMOTO, G; WHITESIDES, J; COLVIN, M; SAFI, R; CHAO, N J; MCDONNELL, D P. Inhibition of aldehyde dehydrogenase and retinoid signaling induces the expansion of human hematopoietic stem cells. Proceedings of the National Academy of Sciences of the United States o f America , v. 103, n. 31, p. 11707–12, 1 ago. 2006. CLARKE, M. F. DICK, J E; DIRKS, P B; EAVES, C J; JAMIESON, C H M; JONES, D L; VISVADER, J; WEISSMAN, I L; WAHL, G M. Cancer stem cells--perspectives on current status and future directions: AACR Workshop on cancer stem cells. Cancer research , v. 66, n. 19, p. 9339–44, 1 out. 2006.
CLEARY, M. P.; GROSSMANN, M. E. Minireview: Obesity and breast cancer: the estrogen connection. Endocrinology , v. 150, n. 6, p. 2537–42, jun. 2009.
COCOLA, C.; ANASTASI, P; ASTIGIANO, S; PISCITELLI, E; PELUCCHI, P; VILARDO, L; BERTOLI, G; BECCAGLIA, M; VERONESI, M C; SANZONE, S; BARBIERI, O; REINBOLD, R; LUVONI, G C; ZUCCHI, I. Isolation of canine mammary cells with stem cell properties and tumour-initiating potential.

120
Reproduction in domestic animals = Zuchthygiene , v. 44, p. 214–7, jul. 2009. Supplement 2. COLLINS, A. T.; BERRY, P; HYDE, C; STOWER, M J; MAITLAND, N J. Prospective identification of tumorigenic prostate cancer stem cells. Cancer research , v. 65, n. 23, p. 10946–51, 1 dez. 2005. DANAEI, G. VANDER HOORN, S; LOPEZ, A D; MURRAY, C J L; EZZATI, M. Causes of cancer in the world: comparative risk assessment of nine behavioural and environmental risk factors. Lancet , v. 366, n. 9499, p. 1784–93, 19 nov. 2005.
DICK, J. E. Stem cell concepts renew cancer research. Blood , v. 112, n. 13, p. 4793–807, 15 dez. 2008.
DIXON, J.; ANDERSON, T.; PAGE, D. Infiltrating lobular carcinoma of the breast. Cancer , v. 77, n. 1, p. 113–120, 1996.
DONNENBERG, V. S.; DONNENBERG, A. D. Multiple drug resistance in cancer revisited: the cancer stem cell hypothesis. Journal of clinical pharmacology , v. 45, n. 8, p. 872–7, ago. 2005.
DONTU, G.; ABDALLAH, W M; FOLEY, J M; JACKSON, K W; CLARKE, M F; KAWAMURA, M J; WICHA, M S. In vitro propagation and transcriptional profiling of human mammary stem / progenitor cells. Cancer Research. p. 1253–1270, 2003. DOYLE, L. A. DOYLE, L. A.; YANG, W.; ABRUZZO, L. V.; KROGMANN, T.; GAO, Y.; RISHI, A. K.; ROSS, D. D. A multidrug resistance transporter from human MCF-7 breast cancer cells. Proceedings of the National Academy of Sciences , v. 95, n. 26, p. 15665–15670, 22 dez. 1998.
FEINBERG, A. P.; OHLSSON, R.; HENIKOFF, S. The epigenetic progenitor origin of human cancer. Nature reviews. Genetics , v. 7, n. 1, p. 21–33, jan. 2006.
FERLETTA, M.; GRAWÉ, J.; HELLMÉN, E. Canine mammary tumors contain cancer stem-like cells and form spheroids with an embryonic stem cell signature. The International journal of developmental biology , v. 55, n. 7-9, p. 791–9, jan. 2011.
FILHO, J. C. O. YUNES, R. Estudo retrospectivo de 1 . 647 tumores mamários em cães. Química Nova . v. 30, n. 2, p. 177–185, 2010.
FILLMORE, C. M.; KUPERWASSER, C. Human breast cancer cell lines contain stem-like cells that self-renew, give rise to phenotypically diverse progeny and survive chemotherapy. Breast cancer research : BCR, v. 10, n. 2, p. R25, jan. 2008.
FONSECA, C. S. Avaliação dos níveis séricos de 17, b-estradiol e p rogesterona em cadelas portadoras de neoplasias mamárias . Dissertação de Mestrado. Faculdade de Ciências Agrárias e Veterinárias - Universidade Estadual Paulista, 1999.

121
FRANCO, J. M. “Mastologia: formação do especialista.” In: Mastologia: formação do especialista . São Paulo: Atheneu. p. 167–170. 1997.
FRESE, K. K.; TUVESON, D. A. Maximizing mouse cancer models. Nature reviews. Cancer , v. 7, n. 9, p. 645–58, set. 2007.
FRIEDLANDER, M. L.; HEDLEY, D. W.; TAYLOR, I. A. N. W. Review article Clinical and biological significance of aneuploidy in human tumours. p. 961–974, 2006.
GAMA, A; ALVES, A; SCHMITT, F. Identification of molecular phenotypes in canine mammary carcinomas with clinical implications: application of the human classification. Virchows Archiv: an international journal of pathology, v. 453, n. 2, p. 123–32, ago. 2008.
GÄRTNER, F. GERALDES, M; CASSALI, G; REMA, A; SCHMITT, F. DNA measurement and immunohistochemical characterization of epithelial and mesenchymal cells in canine mixed mammary tumours: putative evidence for a common histogenesis. Veterinary journal (London, England : 1997), v. 158, n. 1, p. 39–47, jul. 1999. GINESTIER, C. HUR, M; CHARAFE-JAUFFRET, E; MONVILLE, F; DUTCHER, J; BROWN, M; JACQUEMIER, J; VIENS, P; KLEER, C; LIU, S; SCHOTT, A; HAYES, D; BIRNBAUM, D; WICHA, M; DONTU, G. ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell stem cell , v. 1, n. 5, p. 555–67, nov. 2007.
GINESTIER, C.; CHARAFE-JAUFFRET, E; MONVILLE, F; DUTCHER, J; BROWN, M; JACQUEMIER, J; VIENS, P; KLEER, C; LIU, S; SCHOTT, A; HAYES, D; BIRNBAUM, D; WICHA, M; DONTU, G. Retinoid signaling regulates breast cancer stem cell differentiation. Cell Cycle , v. 8, n. 20, p. 3297–3302, 27 out. 2009.
GLOBOCAN. Estimated Incidence, Mortality and Prevalence World wide in 2012 . Lyon: [s.n.]. 2012. Disponível em: <http://globocan.iarc.fr/Pages/fact_sheets_cancer.aspx>.
GOLDSCHMIDT, M. PEÑA, L; RASOTTO, R; ZAPPULLI, V. Classification and grading of canine mammary tumors. Veterinary pathology , v. 48, n. 1, p. 117–31, jan. 2011.
GOTTESMAN, M. M. Mechanisms of cancer drug resistance. Annual Review Medicine , v. 53, p. 615–27, 2002.
GOULD, V. E. KOUKOULIS, G K. Coexpression Patterns of Vimentin and Glial Filament Protein with Cytokeratins in the Normal, hyperplastic and Neoplastic Breast. American Journal of Pathology. v. 137, n. 5, p. 1143–1155, 1990.
GREAVES, M.; MALEY, C. C. Clonal evolution in cancer. Nature , v. 481, n. 7381, p. 306–13, 19 jan. 2012.

122
GRIMWADE, B. D. et al. The Importance of Diagnostic Cytogenetics on Outcome in AML: Analysis of 1,612 Patients Entered Into the MRC AML 10 Trial. Blood. p. 2322–2334, 2015.
GUELSTEIN, V. I.; TCHYPYSHEVA, T. A.; ERMILOVA, V. D.; LITVINOVA, L. V.; TROYANOVSKY, S. M.; BANNIKOV, G. A. Monoclonal antibody mapping of keratins 8 and 17 and of vimentin in normal human mammary gland, benign tumors, dysplasias and breast cancer. International Journal of Cancer , v. 42, n. 2, p. 147–153, 15 ago. 1988.
GUPTA, G. P.; MASSAGUÉ, J. Cancer metastasis: building a framework. Cell , v. 127, n. 4, p. 679–95, 17 nov. 2006.
GUPTA, K.; SOOD, N.; UPPAL, S. Epidemiological studies on canine mammary tumour and its relevance for breast cancer studies. International Organization of Scientific Resarch Journals. v. 2, n. 2, p. 322–333, 2012.
GUPTA, P. B.; CHAFFER, C. L.; WEINBERG, R. A. Cancer stem cells: mirage or reality? Nature medicine , v. 15, n. 9, p. 1010–2, set. 2009.
GUSTERSON, B. A. WARBURTON, M J; MITCHELL, D; ELLISON, M; NEVILLE, A; PHILIP, S. Distribution of Myoepithelial Cells and Basement Membrane Proteins in the Normal Breast and in Benign and Malignant Breast Diseases. p. Cancer Research. 4763–4770, 1982. HAHN, K. A. RICHARDSON, R. C.; HAHN, E. ; CHRISMAN, C. L. Diagnostic and Prognostic Importance of Chromosomal Aberrations Identified in 61 Dogs with Lymphosarcoma. Veterinary Pathology , v. 31, n. 5, p. 528–540, 1 set. 1994.
HATAKA, A. Citologia aspirativa com agulha fina e histopatolog ia: valor e significado para o diagnóstico e prognóstico do cân cer de mama em cadelas . [s.l.]. Tese de Doutorado. Universidade Estadual Paulista, 2004.
HELLMÉN, E. Complex mammary tumours in the female dog: a review. Journal of Dairy Research , v. 72, n. S1, p. 90, 14 jul. 2005.
HERMANN, P. C. HUBER, S L; HERRLER, T; AICHER, A; ELLWART, J W; GUBA, M; BRUNS, C J; HEESCHEN, C. Distinct populations of cancer stem cells determine tumor growth and metastatic activity in human pancreatic cancer. Cell stem cell , v. 1, n. 3, p. 313–23, 13 set. 2007.
HERSHEY, A.; DUBIELZIG, R. Aberrant p53 expression in feline vaccine-associated sarcomas and correlation with prognosis. Veterinary Pathology , v. 811, p. 805–811, 2005.
HODGSON, J. G. EDEN, A; JACKSON-GRUSBY, L; DAUSMAN, J; GRAY, J W; LEONHARDT, H; JAENISCH, R. Induction of Tumors in Mice by Genomic Hypomethylation. v. 300, n. April, p. 489–492, 2003.

123
HÖRSTING, N. et al. Cytogenetic analysis of three oropharyngeal malignant melanomas in dogs. Research in Veterinary Science. n. 1971, p. 147–149, 1999.
HUNTLY, B. J. P.; GILLILAND, D. G. Leukaemia stem cells and the evolution of cancer-stem-cell research. Nature Reviews Cancer. v. 5, n. April, 2005.
HURVITZ, S. A.; FINN, R. S. What’s positive about “triple-negative” breast cancer? Future Oncology , v. 5, n. 7, p. 1015–1025, 1 set. 2009.
HWANG-VERSLUES, W. W. et al. Multiple lineages of human breast cancer stem/progenitor cells identified by profiling with stem cell markers. PloS one , v. 4, n. 12, p. e8377, jan. 2009.
INCA. Estimativa 2014: incidência de câncer no Brasil. Rio de Janeiro: Instituto Nacional do Câncer, 2014.
JIANG, F. et al. Aldehyde dehydrogenase 1 is a tumor stem cell-associated marker in lung cancer. Molecular cancer research : MCR, v. 7, n. 3, p. 330–8, mar. 2009.
JIANG, L. HE, D; YANG, D; CHEN, Z; PAN, Q; MAO, A; CAI, Y; LI, X; XING, H; SHI, M; CHEN, Y; BRUCE, I; WANG, T; JIN, L; QI, X; HUA, D; JIN, J; MA, X. MiR-489 regulates chemoresistance in breast cancer via epithelial mesenchymal transition pathway. FEBS Letters , v. 588, n. 11, p. 2009–2015, 2015.
JONES, P. A; BAYLIN, S. B. The epigenomics of cancer. Cell , v. 128, n. 4, p. 683–92, 23 fev. 2007.
KAJIYAMA, H. SHIBATA, K; TERAUCHI, M; YAMASHITA, M; INO, K; NAWA, A; KIKKAWA, F. Chemoresistance to paclitaxel induces epithelial-mesenchymal transition and enhances metastatic potential for epithelial ovarian carcinoma cells. International Journal of Oncology , v. 31, n. 2, p. 277–283, 2007.
KAKARALA, M.; WICHA, M. S. Implications of the cancer stem-cell hypothesis for breast cancer prevention and therapy. Journal of clinical oncology : official journal of the American Society of Clinical Oncology, v. 26, n. 17, p. 2813–20, 10 jun. 2008.
KARAYANNOPOULOU, M. et al. Histological grading and prognosis in dogs with mammary carcinomas: application of a human grading method. Journal of comparative pathology , v. 133, n. 4, p. 246–52, nov. 2005.
KHANNA, C. et al. How to stay out of a BIND To the editor : Nature Biotechnology , v. 24, n. 9, p. 1065–1066, 2006.
KIMURA, K. C.; ZANINI, D; NISHIYA, A. T.; DAGLI, M L Z. Domestic animals as sentinels for environmental carcinogenic agents. BMC proceedings , v. 7, n. Suppl 2, p. K13, jan. 2013.
KING, M.; MARKS, J.; MANDELL, J. Breast and ovarian cancer risks due to inherited mutations in BRCA1 and BRCA2. Science , v. 302, n. OCTOBER, p. 643–647, 2003.

124
KLEIN, W. R. BRAS, GE; MISDORP, W. Equine sarcoid: BCG immunotherapy compared to cryosurgery in a prospective randomised clinical trial. Cancer Immunology Immunotherapy. (21) p. 133–140, 1986. KLOPFLEISCH, R. LENZE, D; HUMMEL, M; GRUBER, A D. Metastatic canine mammary carcinomas can be identified by a gene expression profile that partly overlaps with human breast cancer profiles. BMC cancer , v. 10, n. 1, p. 618, jan. 2010. KORHONEN, T. KUUKASJÄRVI, T; HUHTALA, H; ALARMO, E L; HOLLI, K; KALLIONIEMI, A; PYLKKÄNEN, L. The impact of lobular and ductal breast cancer histology on the metastatic behavior and long term survival of breast cancer patients. Breast (Edinburgh, Scotland) , v. 22, n. 6, p. 1119–24, dez. 2013.
KUMAR, V. Bases Patológicas das Doenças . [s.l.] Elsevier Brasil, 2011.
LEE, C. KIM, W; LIM, J; KANG, M; KIM, D; KWEON, O. Mutation and overexpression of p53 as a prognostic factor in canine mammary tumors. v. 5, p. 63–64, 2004. LEELAWAT, K. KEERATICHAMROEN, S; LEELAWAT, S; TOHTONG, R. CD24 induces the invasion of cholangiocarcinoma cells by upregulating CXCR4 and increasing the phosphorylation of ERK1/2. Oncology letters , v. 6, n. 5, p. 1439–1446, nov. 2013. LI, C. I. ANDERSON, B O; DALING, J R; MOE, R E. Trends in incidence rates of invasive lobular and ductal breast carcinoma. JAMA , v. 289, p. 1421 – 1424, 2003.
LI, C. I.; URIBE, D. J.; DALING, J. R. Clinical characteristics of different histologic types of breast cancer. British journal of cancer , v. 93, n. 9, p. 1046–52, 31 out. 2005.
LI, L.; XIE, T. Stem cell niche: structure and function. Annual review of cell and developmental biology , v. 21, p. 605–31, jan. 2005.
LINDBLAD-TOH, K. et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature , v. 438, n. 7069, p. 803–19, 8 dez. 2005.
LINGAAS, F. COMSTOCK, K E; KIRKNESS, E F; SORENSEN, A; AARSKAUG, T; HITTE, C; NICKERSON, ML; MOE, L; SCHMIDT, L S; THOMAS, R; BREEN, M; GALIBERT, F; ZBAR, B; OSTRANDER, E. A mutation in the canine BHD gene is associated with hereditary multifocal renal cystadenocarcinoma and nodular dermatofibrosis in the German Shepherd dog. Human molecular genetics , v. 12, n. 23, p. 3043–53, 1 dez. 2003. LORD, S. J. BERNSTEIN, L; JOHNSON, K; MALONE, K E; MCDONALD, J; MARCHBANKS, P; SIMON, M S; STROM, B; PRESS, MF; FOLGER, S G; BURKMAN, R T; DEAPEN, D; SPIRTAS, R; URSIN, G. Breast cancer risk and hormone receptor status in older women by parity, age of first birth, and breastfeeding: a case-control study. Cancer epidemiology, biomarkers & prevention. v. 17, n. 7, p. 1723–30, jul. 2008.

125
MALAISE, E. P.; CHAVAUDRA, N.; TUBIANA, M. The relationship between growth rate, labelling index and histological type of human solid tumours. European Journal of Cancer , v. 9, n. 4, p. 305–312, abr. 1973.
MARCATO, P. DEAN, C; GIACOMANTONIO, C; LEE, PW.K.. Aldehyde dehydrogenase: Its role as a cancer stem cell marker comes down to the specific isoform. Cell Cycle , v. 10, n. 9, p. 1378–1384, 4 nov. 2011. MAYR, B. ESCHBORN, U.; LOUPAL, G; SCHLEGER, W. Characterisation of complex karyotype changes in two canine bone tumours. Research in Veterinary Science , v. 51, p. 341–343, 1991a. MAYR, B. GILLI, H.; SCHLEGER, W.; REIFINGER, M.; BURTSCHER, H. CYTOGENETIC Characterization of Mammary Tumors in Two Domestic Dogs. Journal of Veterinary Medicine Series A , v. 38, n. 1-10, p. 141–147, 12 fev. 1991b. MAYR, B. BREEDING, A; REIFINGER, M; CK, H L. Cytogenetic analyses of four solid tumours in dogs. Research in Veterinary Science. p. 88–95, 1994.
MAYR, B.; SCHLEGER, W.; LOUPALT, G. Cytogenetic Studies in a Canine Melanoma. Journal of Comparative Pathilogy. v. 106, 1992.
MERLO, D. F. ROSSI, L; PELLEGRINO, C; CEPPI, M; CARDELLINO, U; CAPURRO, C; RATTO, A; SAMBUCCO, P L; SESTITO, V; TANARA, G; BOCCHINI, VL. C a n c e r I n c i d e n c e in P e t D o g s : Fi n d i n g s o f th e A n i m a l T u m o r R egi str y of Genoa, I taly. p. 976–984, 2008. MICHISHITA, M. AKIYOSHI, R; YOSHIMURA, H; KATSUMOTO, T; ICHIKAWA, H; OHKUSU-SUKADA, K; NAKAGAWA, T; SASAKI, N; TAKAHASHI, K. Characterization of spheres derived from canine mammary gland adenocarcinoma cell lines. Research in veterinary science , v. 91, n. 2, p. 254–60, out. 2011. MICHISHITA, M. AKIYOSHI, R; SUEMIZU, H; NAKAGAWA, T; SASAKI, N; TAKEMITSU, H; ARAI, T; TAKAHASHI, K. Aldehyde dehydrogenase activity in cancer stem cells from canine mammary carcinoma cell lines. Veterinary journal (London, England : 1997), v. 193, n. 2, p. 508–13, ago. 2012. MILL, Y. FERN, V; RAM, S; MART, J; MULAS, DE; ORD, J. Oncogene HER-2 in canine mammary gland carcinomas. p. 363–367, 2003. MISDORP, W. ELSE, R W; HELLME´N, E; LIPSCOMB, TP. Histological classification of mammary tumors of the dog and the cat . Washington, DC: [s.n.]. 1999.
MOULTON, J. E. Tumors in domestic animals. In: [s.l.] Berkeley: University of California, 1990. p. 518–522.
MULLIGAN, R. M. Mammary cancer in the dog: a study of 120 cases. American journal of veterinary research , v. 36, n. 9, p. 1391–6, 1 out. 1975.

126
NICHOLAS, F. W. Introduçao à Genética Veterinária . 3 ed. [S.l.] Artmed, 2012. p. 347
NOWELL, P. The clonal evolution of tumor cell populations. Science , p. 23–28, 1976.
O’BRIEN, C. A.; POLLETT, A; GALLINGER, S; DICK, J E. A human colon cancer cell capable of initiating tumour growth in immunodeficient mice. Nature , v. 445, n. 7123, p. 106–10, 4 jan. 2007. O’HARE, M. J. ORMEROD, M G.; MONAGHAN, P; LANE, E. B; GUSTERSON, B A.. Characterization in vitro of luminal and myoepithelial cells isolated from the human mammary gland by cell sorting. Differentiation , v. 46, n. 3, p. 209–221, abr. 1991. ONDER, T. T. GUPTA, P B; MANI, S; YANG, J; LANDER, E S; WEINBERG, R. Loss of E-cadherin promotes metastasis via multiple downstream transcriptional pathways. Cancer research , v. 68, n. 10, p. 3645–54, 15 maio 2008. OSIN, P. P. ANBAZHAGAN, R; BARTKOVA, J; NATHAN, B; GUSTERSON, B. Breast development gives insights into breast disease. Histopathology , v. 33, n. 3, p. 275–283, set. 1998.
PALMER, T. E.; MONLUX, A. W. Acid Mucopolysaccharides in Mammary Tumors of Dogs. Veterinary Pathology , v. 16, n. 5, p. 493–509, 1 set. 1979.
PAOLONI, M.; KHANNA, C. Translation of new cancer treatments from pet dogs to humans. Nature Reviews Cancer , v. 2946, n. 2000, p. 2943–2946, 2008.
PATRAWALA, L. CALHOUN, T; Side Population Is Enriched in Tumorigenic , Stem-Like Cancer Cells , whereas ABCG2 + and ABCG2 À Cancer Cells Are Similarly Tumorigenic. Cancer Research. n. 14, p. 6207–6220, 2005. PONTI, D. COSTA, A; ZAFFARONI, N; PRATESI, G; PETRANGOLINI, G; CORADINI, D; PILOTTI, S; PIEROTTI, M; DAIDONE, M G. Isolation and in vitro propagation of tumorigenic breast cancer cells with stem/progenitor cell properties. Cancer research , v. 65, n. 13, p. 5506–11, 1 jul. 2005. PRASMICKAITE, L. ENGESAETER, B; SKRBO, N; HELLENES, T; KRISTIAN, A; OLIVER, N K; SUO, Z; MAELANDSMO, G M. Aldehyde dehydrogenase (ALDH) activity does not select for cells with enhanced aggressive properties in malignant melanoma. PloS one , v. 5, n. 5, p. e10731, jan. 2010. RAKHA, E. A EL-SAYED, M E; POWE, D G; GREEN, A R; HABASHY, H; GRAINGE, M J; ROBERTSON, J F R; BLAMEY, R; GEE, J; NICHOLSON, R I; LEE, A H S; ELLIS, I O. Invasive lobular carcinoma of the breast: response to hormonal therapy and outcomes. European journal of cancer (Oxford, England : 1990), v. 44, n. 1, p. 73–83, jan. 2008.

127
RANGARAJAN, A.; WEINBERG, R. Comparative biology of mouse versus human cells: modelling human cancer in mice. Nature Reviews Cancer , v. 3, p. 952–959, 2003.
RASHEED, Z. A YANG, J; WANG, Q; KOWALSKI, J; FREED, I; MURTER, C; HONG, S; KOORSTRA, J; RAJESHKUMAR, N V; HE, X; GOGGINS, MI; IACOBUZIO-DONAHUE, C; BERMAN, D; LAHERU, D; JIMENO, A; HIDALGO, M; MAITRA, A; MATSUI, W. Prognostic significance of tumorigenic cells with mesenchymal features in pancreatic adenocarcinoma. Journal of the National Cancer Institute , v. 102, n. 5, p. 340–51, 3 mar. 2010. REID, B. J. LI, XI; GALIPEAU, P C; VAUGHAN, T L. Barrett’s oesophagus and oesophageal adenocarcinoma: time for a new synthesis. Nature reviews. Cancer , v. 10, n. 2, p. 87–101, fev. 2010.
REIMANN, N. et al. Report on the progress of standardization of the G-banded canine ( Canis familiaris ) karyotype Committee for the Standardized Karyotype of the Dog ( Canis familiaris ). Chromosome Research. v. 4, p. 306–309, 1996.
REIMANN, N. BOSMA, A A; LONG, S; BARTNITZKE, S; FISCHER, P. Working with Canine Chromosomes : Current Recommendations for Karyotype Description. The Journal of Heredity. p. 31–34, 1999. REYA, T. MORRISON, S J; CLARKE, MF; WEISSMAN, I L. Stem cells, cancer, and cancer stem cells. Nature , v. 414, n. November, p. 105–111, 2001. RONINSON, I. B. ABELSON, H T; HOUSMAN, D E; HOWELL, N; VARSHAVSKY, A. Amplification of specific DNA sequences correlates with multi-drug resistance in Chinese hamster cells. Nature , v. 309, n. 5969, p. 626–628, 14 jun. 1984. RUTTEMAN, G. R. CORNELISSE, C J; DIJKSHOORN, N J; POORTMAN, J; MISDORP, W. Flow Cytometric Analysis of DNA Ploidy in Canine Mammary Tumors1. Cancer Research. 1988. SARRIÓ, D. RODRIGUEZ-PINILLA, S M; HARDISSON, D; CANO, A; MORENO-BUENO, G; PALACIOS, J. Epithelial-mesenchymal transition in breast cancer relates to the basal-like phenotype. Cancer research , v. 68, n. 4, p. 989–97, 15 fev. 2008.
SCADDEN, D. T. The stem-cell niche as an entity of action. Nature , v. 441, n. 7097, p. 1075–9, 29 jun. 2006.
SCHALIE, W. H. VAN DER JR, H S G; BANTLE, J A; ROSA, CT DE; FINCH, R A; JOHN, S; REUTER, H; BACKER, L C; BURGER, J; FOLMAR, L C; STOKES, W S. Animals as Sentinels of Human Health Hazards of Environmental Chemicals. v. 107, n. 4, p. 309–315, 1999.
SCHNEIDER, R.; DORN, C. R.; TAYLOR, D. O. N. Factors Influencing Canine Mammary Cancer Development and Postsurgical Survival. Journal National Cancer Institute , v. 43, n. 6, p. 1249–1261, 1 dez. 1969.

128
SELTENHAMMER, M. H. et al. Comparative histopathology of grey-horse-melanoma and human malignant melanoma. Pigment cell research , v. 17, n. 6, p. 674–81, dez. 2004.
SHACKLETON, M. VAILLANT, F; SIMPSON, K J; STINGL, J; SMYTH, G K; ASSELIN-LABAT, M; WU, L; LINDEMAN, G J; VISVADER, J E. Generation of a functional mammary gland from a single stem cell. Nature , v. 439, n. 7072, p. 84–8, 5 jan. 2006. SHEN, C. XIANG, M; NIE, C; HU, H; MA, Y; WU, H. CD44 as a molecular marker to screen cancer stem cells in hypopharyngeal cancer. Acta oto-laryngologica , v. 133, n. 11, p. 1219–26, nov. 2013. SHMELKOV, S. V. BUTLER, J M; HOOPER, A T; HORMIGO, A; KUSHNER, J; MILDE, T; CLAIR, R S; BALJEVIC, M; WHITE, I; JIN, D K; CHADBURN, A; MURPHY, A J; VALENZUELA, D M; GALE, NICHOLAS W; THURSTON, G; YANCOPOULOS, G D; ANGELICA, M D; KEMENY, N; LYDEN, D; RAFII. CD133 expression is not restricted to metastatic colon cancer cells initiate tumors. Journal of Clinical Investigation. v. 118, n. 6, p. 2111–2120, 2008. SIMS-MOURTADA, J. IZZO, J G; AJANI, J; CHAO, K S C. Sonic Hedgehog promotes multiple drug resistance by regulation of drug transport. Oncogene , v. 26, n. 38, p. 5674–9, 16 ago. 2007. SINGH, S. K. CLARKE, I D; TERASAKI, M; BONN, V E; HAWKINS, C; SQUIRE, J; DIRKS, P. Identification of a Cancer Stem Cell in Human Brain Tumors. Cancer Research p. 5821–5828, 2003.
SMALLEY, M. J.; CLARKE, R. B. The mammary gland “side population”: a putative stem/progenitor cell marker? Journal of mammary gland biology and neoplasia , v. 10, n. 1, p. 37–47, jan. 2005.
SMITH, B. N. BURTON, L J; HENDERSON, V; RANDLE, D D; MORTON, D J; SMITH, B; TALIAFERRO-SMITH, L; NAGAPPAN, P; YATES, C; ZAYZAFOON, M; CHUNG, L W K; ODERO-MARAH, V. Snail promotes epithelial mesenchymal transition in breast cancer cells in part via activation of nuclear ERK2. PloS one , v. 9, n. 8, p. e104987, jan. 2014.
SORENMO, K. U. Canine mammary gland tumors. Veterinary Clinics of North America: Small Animal Practice , v. 33, n. 3, p. 573–596, 2003.
SORENMO, K. U.; SHOFER, F. S.; GOLDSCHMIDT, M. H. Effect of Spaying and Timing of Spaying on Survival of Dogs with Mammary Carcinoma. Journal of Veterinary Internal Medicine , v. 14, n. 3, p. 266, 2000.
SUETSUGU, A. NAGAKI, M; AOKI, H; MOTOHASHI, T; KUNISADA, T; MORIWAKI, H. Characterization of CD133+ hepatocellular carcinoma cells as cancer stem/progenitor cells. Biochemical and biophysical research communications , v. 351, n. 4, p. 820–4, 29 dez. 2006.

129
SUTTER, N. B.; OSTRANDER, E. A. Dog star rising: the canine genetic system. Nature reviews. Genetics , v. 5, n. 12, p. 900–10, dez. 2004.
TATEYAMA, S.; COTCHIN, E. Electron microscopic observations on canine mixed mammary tumors, with special reference to cytoplasmic filamentous components. American journal of veterinary research , v. 39, n. 9, p. 1494–501, 1 out. 1978.
TAYLOR, P. J.; LANE, E. B. Keratin expression in the mammary gland. In: The Mammary Gland: Development, Regulation and Function . New York: [s.n.]. p. 181.
THE CANCER GENOME ATLAS NETWORK. Comprehensive molecular portraits of human breast tumours. Nature , v. 490, n. 7418, p. 61–70, 4 out. 2012.
THIERY, J. P. Epithelial-mesenchymal transitions in tumour progression. Nature reviews. Cancer , v. 2, n. 6, p. 442–54, jun. 2002.
TROPEPE, V. et al. Retinal Stem Cells in the Adult Mammalian Eye. Science. v. 287, n. 5460, p. 2032-2036, 2000.
TWENTYMAN, P. R.; FOX, N. E.; BLEEHEN, N. M. Drug resistance in human lung cancer cell lines: Cross-resistance studies and effects of the calcium transport blocker, verapamil. International Journal of Radiation Oncology*Biology*Physics , v. 12, n. 8, p. 1355–1358, ago. 1986.
UVA, P. et al. Comparative expression pathway analysis of human and canine mammary tumors. BMC genomics , v. 10, p. 135, jan. 2009.
VAN ROY, F.; BERX, G. The cell-cell adhesion molecule E-cadherin. Cellular and molecular life sciences : CMLS, v. 65, n. 23, p. 3756–88, nov. 2008.
VISVADER, J. E.; LINDEMAN, G. J. Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nature reviews. Cancer , v. 8, n. 10, p. 755–68, out. 2008.
WALSH, T.; KING, M.-C. Ten genes for inherited breast cancer. Cancer cell , v. 11, n. 2, p. 103–5, fev. 2007.
WANG, J. C. Y. Good cells gone bad: the cellular origins of cancer. Trends in molecular medicine , v. 16, n. 3, p. 145–51, mar. 2010.
WANG, R. LV, Q; MENG, W; TAN, Q; ZHANG, S; MO, X; YANG, X. Comparison of mammosphere formation from breast cancer cell lines and primary breast tumors. Journal of thoracic disease , n. 6, p. 829–37, jun. 2014. WELLNER, U. SCHUBERT, J; BURK, U C; SCHMALHOFER, O; ZHU, F; SONNTAG, A; WALDVOGEL, B; VANNIER, C; DARLING, D; ZUR HAUSEN, A; BRUNTON, V G; MORTON, J; SANSOM, O; SCHÜLER, J; STEMMLER, M P; HERZBERGER, C; HOPT, U; KECK, T; BRABLETZ, S; BRABLETZ, T. The EMT-

130
activator ZEB1 promotes tumorigenicity by repressing stemness-inhibiting microRNAs. Nature cell biology , v. 11, n. 12, p. 1487–95, dez. 2009. YU, F. YAO, H; ZHU, P; ZHANG, X; PAN, Q; GONG, C; HUANG, Y; HU, X; SU, F; LIEBERMAN, J; SONG, E. Let-7 Regulates Self Renewal and Tumorigenicity of Breast Cancer Cells. Cell , v. 131, n. 6, p. 1109–23, 14 dez. 2007.
ZATLOUKAL, J.; KOHOUT, P.; REPUBLIC, C. Breed and Age as Risk Factors for Canine Mammary Tumours. Acta Veterinaria Bruno. v. 74. p. 103–109, 2005.
ZHANG, J.; MA, L. MicroRNA control of epithelial–mesenchymal transition and metastasis. Cancer and Metastasis Reviews , v. 31, n. 3-4, p. 653–662, 2012.