Embed Size (px)
DESCRIPTION
Cursul 9 Bacteriologie
Citation preview

Tipuri de nutriţie
• În raport cu sursele de C şi N şi cu tipul de nutriţie, bacteriile se împart în două categorii:
– bacterii autotrofe – bacterii heterotrofe
• Bacteriile autotrofe sunt larg răspândite în sol şi ape şi sunt în totalitate saprofite
• acestea se apropie mult de regnul vegetal, fiind caracterizate prin faptul că utilizează ca surse de N şi C numai componente anorganice
• Ca surse de carbon utilizează CO2 şi carbonaţii, iar ca surse de N utilizează NH3, nitraţii şi nitriţii

• îşi sintetizează materia organică proprie şi îşi procură - total sau parţial - energia necesară
• bacteriile autotrofe îndeplinesc în natură un rol important în circulaţia C, N, S
• Unele au o contribuţie majoră la îmbogăţirea solului în compuşi azotaţi.
• bacteriile din genul Nitrosomonas oxidează NH3 în nitraţi
• cele din genul Nitrobacter transformă nitriţii în nitraţi
• Energia obţinută în cursul acestor procese chimice este utilizată la reducerea CO2, care le serveşte ca sursă de C
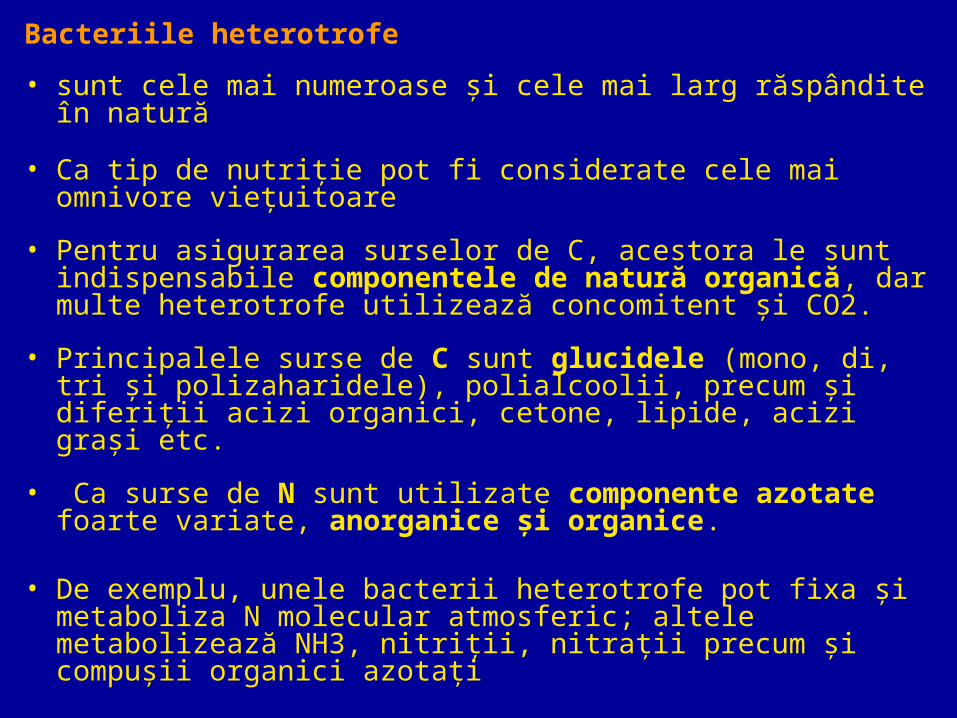
Bacteriile heterotrofe
• sunt cele mai numeroase şi cele mai larg răspândite în natură
• Ca tip de nutriţie pot fi considerate cele mai omnivore vieţuitoare
• Pentru asigurarea surselor de C, acestora le sunt indispensabile componentele de natură organică, dar multe heterotrofe utilizează concomitent şi CO2.
• Principalele surse de C sunt glucidele (mono, di, tri şi polizaharidele), polialcoolii, precum şi diferiţii acizi organici, cetone, lipide, acizi graşi etc.
• Ca surse de N sunt utilizate componente azotate foarte variate, anorganice şi organice.
• De exemplu, unele bacterii heterotrofe pot fixa şi metaboliza N molecular atmosferic; altele metabolizează NH3, nitriţii, nitraţii precum şi compuşii organici azotaţi

• între heterotrofe, in raport cu particularităţile metabolice,se individualizează:– bacteriile prototrofe– bacteriile auxotrofe -
Bacteriile prototrofe
• îşi asigură sursele de C prin metabolizarea carbohidraţilor, iar sursele de N pe seama exclusiv a unor componente anorganice, în special prin metabolizarea N molecular atmosferic.
• bacteriile din genul Azobacter, care trăiesc libere în sol şi ape
• bacteriile din genul Rhisobium care trăiesc în sol de regulă în simbioză cu plantele leguminoase, găsindu-se din abundenţă în special în nodozităţile formate pe rădăcinile acestora.

• îşi asigură cerinţele în N exclusiv prin fixarea de N molecular atmosferic - formează compuşi azotaţi care îmbogăţesc solul şi servesc plantelor ca surse nutritive.
• plantele asigură bacteriilor carbohidraţii necesari ca surse de C
Bacteriile auxotrofe
• Multiplicarea acestora este dependentă de prezenţa în mediu a unuia sau a mai multor acizi aminaţi, pe care bacteriile respective
nu-i pot sintetiza
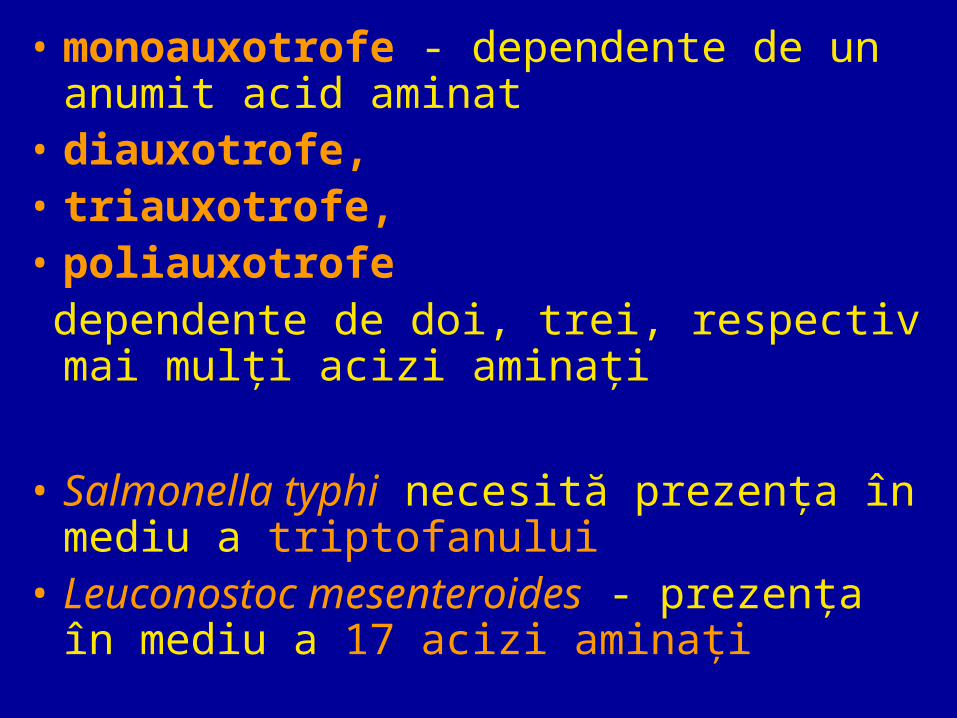
• monoauxotrofe - dependente de un anumit acid aminat
• diauxotrofe,• triauxotrofe,• poliauxotrofe dependente de doi, trei, respectiv mai mulţi
acizi aminaţi
• Salmonella typhi necesită prezenţa în mediu a triptofanului
• Leuconostoc mesenteroides - prezenţa în mediu a 17 acizi aminaţi
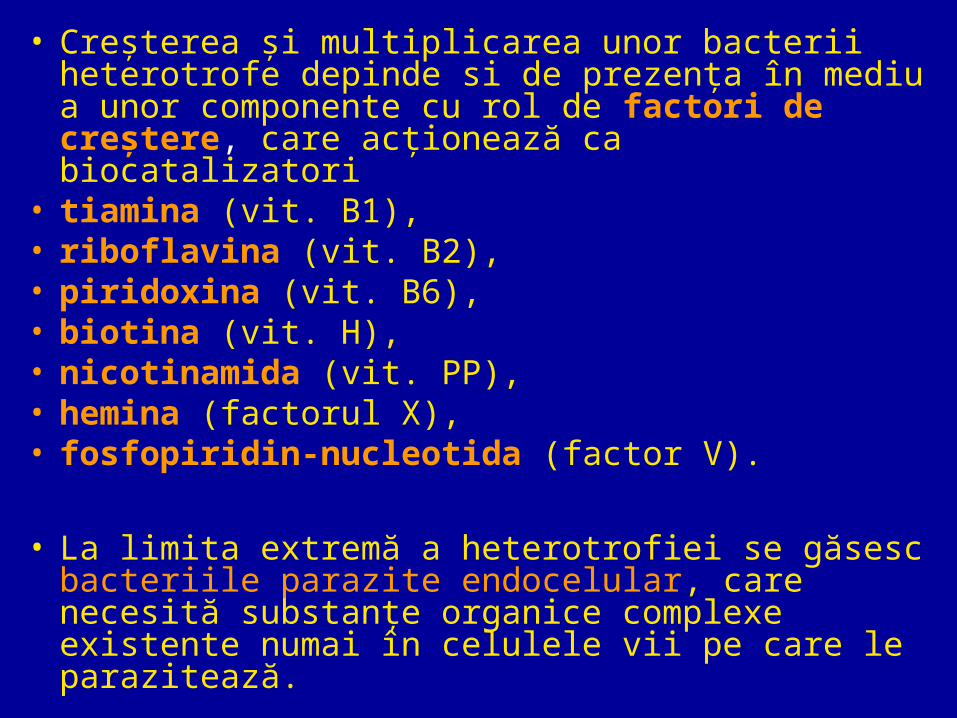
• Creşterea şi multiplicarea unor bacterii heterotrofe depinde si de prezenţa în mediu a unor componente cu rol de factori de creştere, care acţionează ca biocatalizatori
• tiamina (vit. B1),• riboflavina (vit. B2),• piridoxina (vit. B6), • biotina (vit. H), • nicotinamida (vit. PP),• hemina (factorul X),• fosfopiridin-nucleotida (factor V).
• La limita extremă a heterotrofiei se găsesc bacteriile parazite endocelular, care necesită substanţe organice complexe existente numai în celulele vii pe care le parazitează.

Pătrunderea substanţelor nutritive în celulă
• Pătrunderea substanţelor nutritive prin membrana citoplasmatică este, în general, un proces mult mai complex şi foarte selectiv, care se poate realiza prin trei mecanisme:
Mecanisme nespecificeMecanisme de difuziune Mecanismede selective - prin procese de
transport selectiv

• Mecanisme nespecifice
sunt caracteristice substanţelor nutritive solubile în lipide sau cu molecule în lanţ ce pot traversa membrana citoplasmatică strecurându-se prin porii reţelei lipoproteice;
Mecanisme de difuziune
se realizează atunci când substanţa nutritivă se găseşte în concentraţii mai mari în mediu decât în celula

METABOLISMUL GLUCIDIC• glucidele reprezintă surse energetice şi plastice deosebit de
valoroase
• căile de descompunere (catabolizare) şi de sinteză (anabolismul glucidelor) sunt deosebit de complexe şi uneori diferite de la o specie la alta,
• ac. piruvic rezultat din catabolizarea glucidelor permite realizarea unor interferări între metabolismul energetic şi plastic, în sensuri multiple
• în funcţie de calea de metabolizare (oxibiotică, anoxibiotică, fermentativă), se poate ajunge
- fie la o oxidare completă a substratului, cu formarea de metaboliţi finali (CO2, H2O),
- fie la o oxidare incompletă, cu formarea de metaboliţi intermediari care pot servi apoi ca surse pentru sinteza unor componente proteice şi lipidice proprii celulei bacteriene

Catabolizarea glucidelor
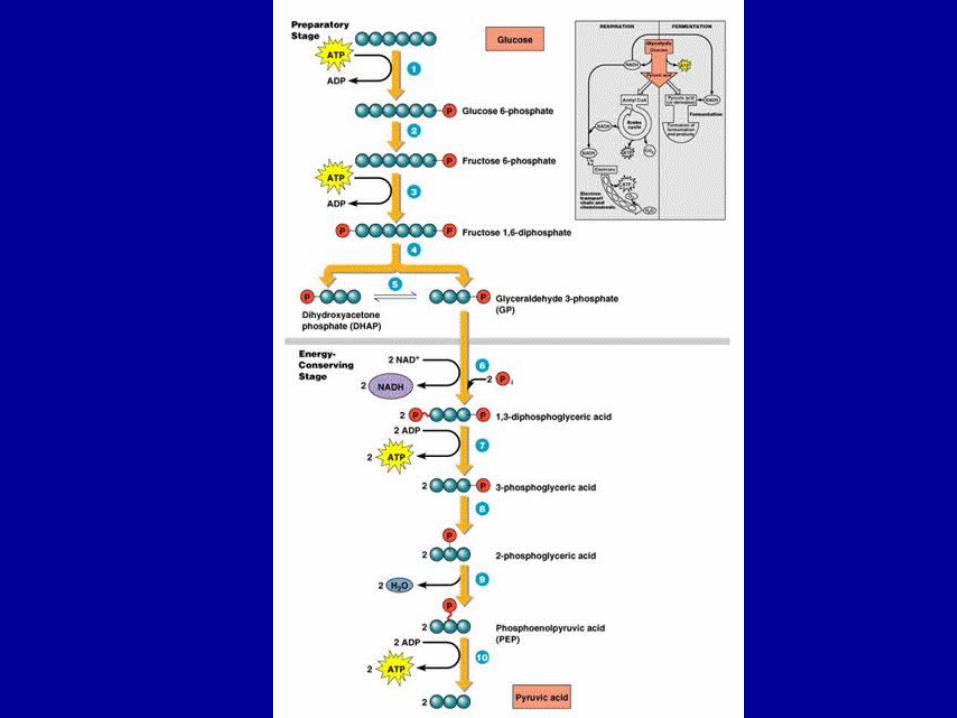
• Componentele macromoleculare glucidice sunt dezintegrate de enzimele bacteriene în componentele lor constitutive, prin procese de fosforoliză la nivelul legăturilor glicozidice, cu eliberarea de energie
• Rezultă derivaţi glucidici fosfataţi, cu structură din ce în ce mai simplă, până se atinge stadiul de ozide fosfatice, de regulă gluco-zofosfat.
• La cele mai multe microorganisme, catabolizarea glucozei urmează în continuare calea care duce la formarea de ac. piruvic

• Bacteriile pot utiliza însă alternativ şi alte căi
• Astfel, aproape toate bacteriile pot cataboliza glucoza pe calea pentozofosfaţilor obţinându-se în final acid piruvic.
• De aici, catabolizarea poate continua diferit de la o specie bacteriană la alta, pe căi – oxibiotice, – anoxibiotice – fermentative

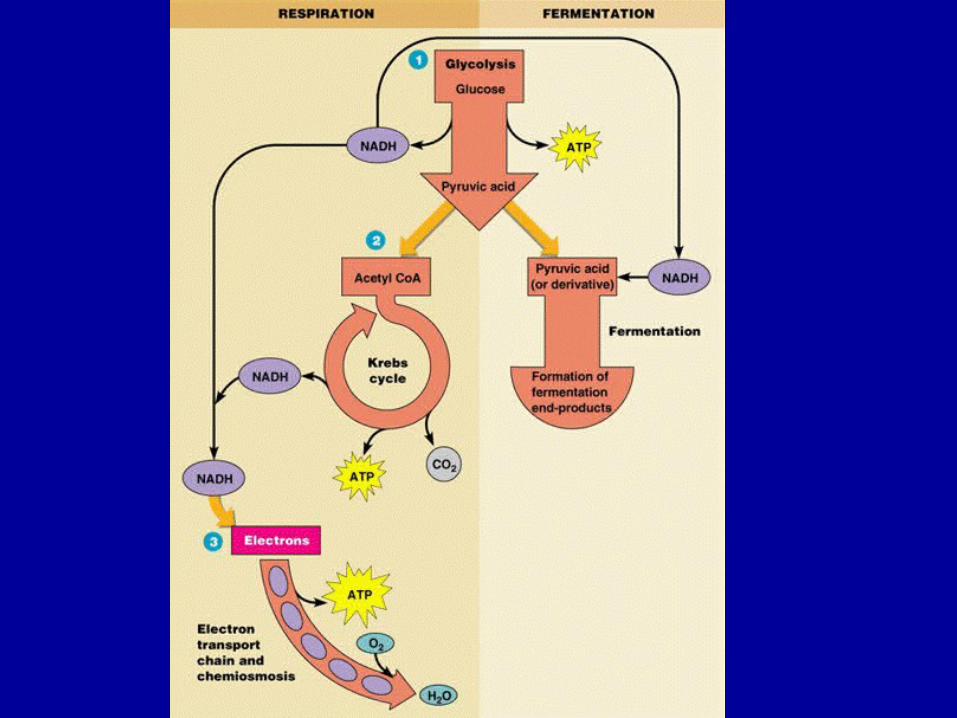
Catabolizarea oxibiotică a ac. piruvic • oxidoreduceri
• La bacteriile aerobe metabolizarea acidului piruvic pe cale oxibiotică este impusă de prezenţa în mediu a O2 (care inhibă metabolizarea pe căi fermentative)
• participa CoA
• se produce o condensare între ac. acetic şi ac. oxalo-acetic rezultând acid citric (compus cu 6 atomi C).
• Acesta este metabolizat în continuare pe calea ciclului Krebs (acizilor tricarboxilici)
• cele 10 enzime care catalizează reacţiile în ciclul Krebs sunt considerate ca enzime respiratorii
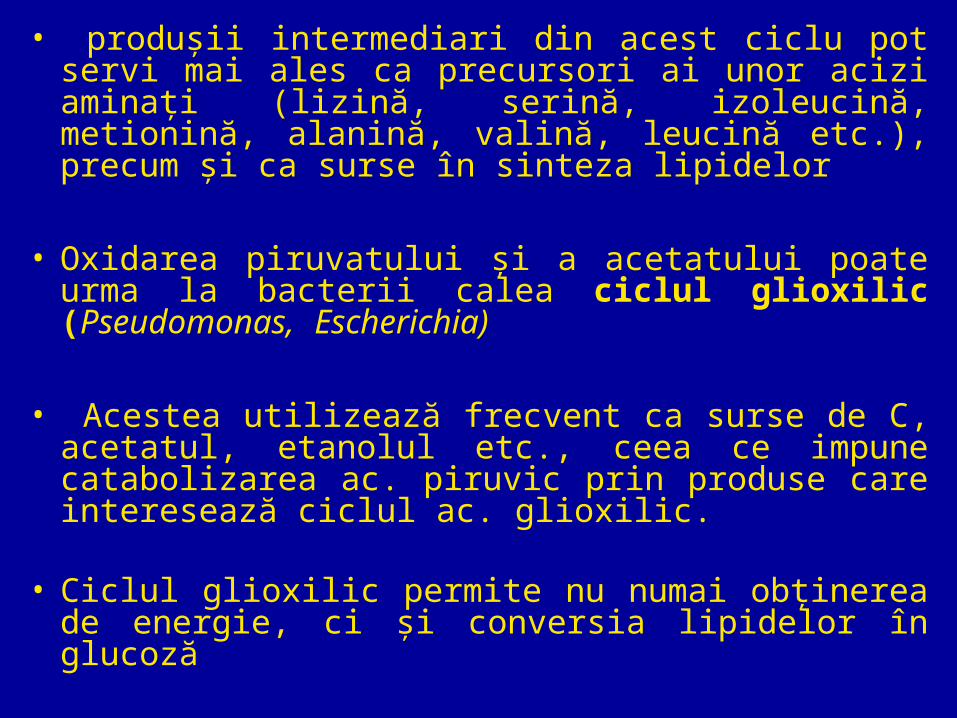
• produşii intermediari din acest ciclu pot servi mai ales ca precursori ai unor acizi aminaţi (lizină, serină, izoleucină, metionină, alanină, valină, leucină etc.), precum şi ca surse în sinteza lipidelor
• Oxidarea piruvatului şi a acetatului poate urma la bacterii calea ciclul glioxilic (Pseudomonas, Escherichia)
• Acestea utilizează frecvent ca surse de C, acetatul, etanolul etc., ceea ce impune catabolizarea ac. piruvic prin produse care interesează ciclul ac. glioxilic.
• Ciclul glioxilic permite nu numai obţinerea de energie, ci şi conversia lipidelor în glucoză

Catabolizarea anoxibiotică
• Reprezintă o altă modalitate de catabolizare a piruvatului prin decarboxilarea neoxidativă cu formarea de acetaldehidă
• poate fi metabolizată, de către bacteriile din genul Acetobacter pe calea fermentaţiei acetice;
• tot de la acetaldehidă, în condiţii de anaerobioză, metabolizarea poate decurge în sensul fermentaţiei alcoolice, proces specific unor levuri din genul Saccharomyces, precum şi unor bacterii.

• Catabolizarea anoxibiotică a ac. piruvic are obişnuit un caracter fermentativ, cu formarea în final a unor ac. graşi, în funcţie de specie şi de calea de metabolizare.
• Procesul fermentativ poate avea un caracter de: - fermentaţie lactică (caracteristică speciilor din genul
Lactobacillus şi unor coci Gram-pozitivi); - fermentaţie acetică (caracteristică bacteriilor acetice strict anaerobe);- fermentaţie citrică (caracteristică bacteriilor citrice şi
Enterobacteriaceaelor);- fermentaţie propionică (caracteristică unor bacterii
ruminale Gram-pozitive şi lactobacililor); - fermentaţie butilică sau butirică (caracteristică unor specii din genul Clostridium) etc.

Sinteza glucidelor
• Procesele anabolice urmează de regulă în sens invers căile de catabolizare a glucidelor.
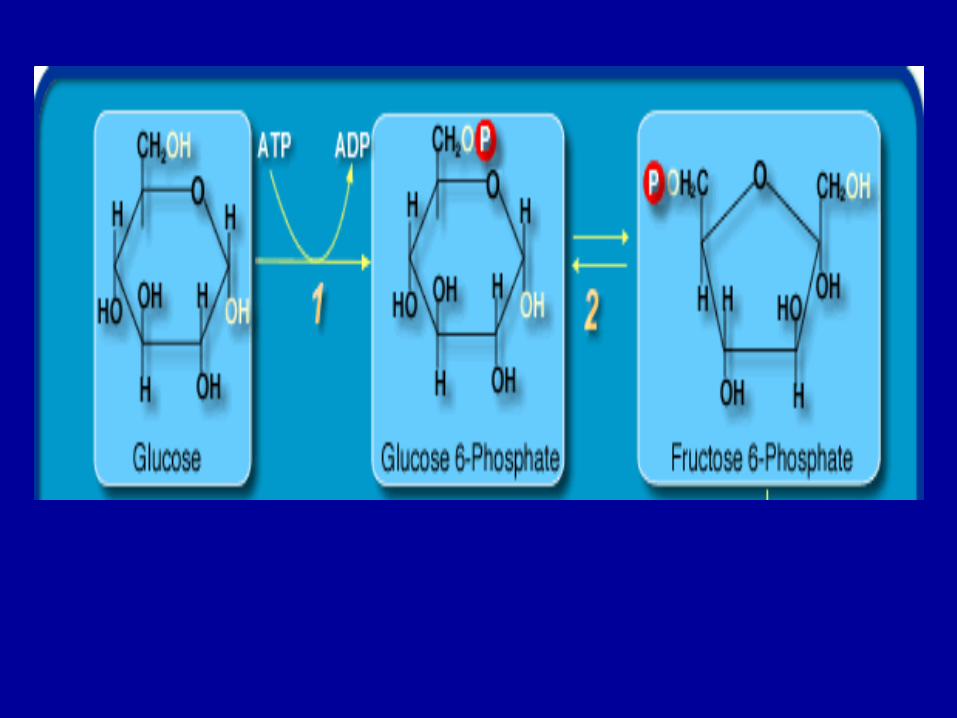
• polimerizarea glucidelor în celula bacteriană se realizează obişnuit prin procese de fosforilare a ozidelor, cu participarea ATP, ceea ce permite acumularea în structura moleculelor glucidice a unor cantităţi însemnate de energie
• polimerizarea glucozei în amidon sau glicogen este precedată de fosforilarea sa, prin participarea ATP (o grupare hidroxilică fiind înlocuită cu o grupare fosforică), obţinându-se în prima etapă glucozofosfatul; apoi, prin polimerizarea acestuia se obţine energie şi ATP.

METABOLISMUL PROTEIC• Proteinele şi acizii aminaţi sunt pentru bacterii surse
plastice şi energetice deosebit de valoroase.
• Pentru pătrunderea în celulă, proteinele sunt în prealabil hidrolizate prin enzimele proteolitice (proteaze), care le scindează în peptide;
• acestea, prin peptidaze, sunt apoi degradate la acizi aminaţi.
• transportul acestora la nivelul membranei citoplasmatice este mediat de translocaze şi permeaze

• bacteriile Gram-negative, continuă catabolizarea celor mai mulţi acizi aminaţi - pot sintetiza singure, în raport cu cerinţele celulei
• unele bacterii Gram-negative utilizează şi acumulează în celulă anumiţi acizi aminaţi, prezenţi în mediu sau rezultaţi din catabolizarea proteinelor
• bacteriile Gram-pozitive au de regulă posibilităţi mai restrânse de a sintetiza acizi aminaţi şi, de aceea, ele utilizează în măsură mult mai mare acizii aminaţi din mediu

Catabolizarea acizilor aminaţi
• Procesele de catabolizare a acizilor aminaţi sunt realizate de către bacterii prin
- dezaminare- decarboxilare
• Dezaminarea acizilor aminaţi
• duce la formarea cetoacidului corespunzător, cu liberarea de NH3, acesta putând fi utilizat apoi la sinteza altor componente azotate.
• Dezaminarea poate avea următoarele mecanisme:- dezaminare oxidativă, cu formarea de NH3 şi cetoacid
corespunzător;- dezaminare reductivă, cu formarea unui acid gras saturat;- dezaminare desaturantă, cu formarea unui acid gras
nesaturat;- dezaminare hidrolitică, cu formarea unui hidroxiacid.
• O modalitate de dezaminare frecventă la bacterii o constituie transaminarea, mediată de transaminaze
• transferul grupării NH2 de la un acid aminat la un cetoacid, acesta devenind astfel un nou acid aminat
• procesul de transaminare reprezintă, pentru cele mai multe bacterii, modalitatea cea mai frecventă de sinteză a unor acizi aminaţi proprii.
Decarboxiarea acizilor aminaţi
anaerobioză şi este mediată de decarboxilaze
procesul duce la formarea de amide, gruparea carboxilică (- COOH) fiind de asemenea scindată, cu formarea de CO2
decarboxilarea reprezintă unul din mecanismele importante de formare a CO2

Biosinteza acizilor aminaţi şi a proteinelor
Sinteza acizior aminaţi de către bacterii se poate realiza pe căi multiple, unele încă insuficient cunoscute.
Cele mai frecvente modalităţi de sinteză a acizilor aminaţi se realizează însă prin procese de aminare şi transaminare.
sinteza prin aminare - porneste de la metaboliţii intermediari din categoria ceto-acizilor, respectiv de la ac. piruvic, ac. -ceto-glutaric şi ac. oxalo-acetic, prin aminarea cărora se obţin: alanina, ac. glutamic şi ac. aspartic.

sursa de N o constituie compuşii anorganici, în special NH3
Cei mai mulţi acizi aminaţi sunt sintetizaţi însă prin transaminare
acidul glutamic are un rol important, întrucât transferul grupării sale aminice la diferiţi cetoacizi poate duce la formarea multor acizi aminaţi.
Sinteza polipeptidelor
• majoritatea proteinelor sunt sintetizate prin polimerizarea acizilor aminaţi, cu participarea de regulă a ATP-ului.
• Scindarea ATP-ului furnizează celulei bacteriene o mare cantitate de energie, utilizată în parte în procesele de polimerizare (sinteză);
• Procesul de biosinteză a proteinelor, al cărui mecanism a fost descifrat prin cercetări în sisteme acelulare de E. coli, se desfăşoară în două etape:
• - transcrierea în ARNm (transcripţie) a informaţiei genetice codificată în macromolecula de ADN;
• - traducerea informaţiei genetice purtată de ARNm la ribosomi (translaţie).

METABOLISMUL LIPIDIC
• Bacteriile posedă lipaze care le permit să degradeze componente lipidice foarte variate (fosfolipide, acizi graşi, steroli)
• lipidele au un rol redus în nutriţia bacteriilor
• prezenţa lipidelor în mediile de cultivare nu este necesară nici chiar pentru bacteriile care conţin în structura lor cantităţi mari de lipide (M. tuberculosis)
• sunt totuşi unele specii care necesită prezenţa în mediu a glicerolului, pe care îl utilizează mai ales ca sursă de C şi energie
• toate bacteriile sintetizează însă lipide, unele în cantităţi apreciabile, acestea intrând mai ales în structura componentelor parietale ale celulei, iar o parte pot fi depozitate în celulă ca materii de rezervă
Catabolizarea lipidelor
• procesele se desfăşoară pe căile metabolice clasice
• scindarea lipidelor de către lipazele bacteriene duce la formarea de acizi graşi şi glicerol.
• În funcţie de natura lipidelor catabolizate rezultă diferiţi acizi graşi, de regulă acizi saturaţi cu 14-18 atomi de C (în special ac. palmitic şi ac. stearic).
• un rol important revine acetil-CoA, care activează acidul gras.
• Fiecare etapă de degradare duce la formarea unui acid gras cu 2 atomi de C mai puţin şi cu eliberarea unui component dicarbonic cu acetil-CoA.

Sinteza lipidelor
• Se realizează probabil prin reacţii care permit obţinerea unor acizi graşi cu lanţ carbonic lung, pornind de la anumiţi acizi organici prezenţi în mediu sau rezultaţi din metabolismul celular.
• În proces intervine, de asemenea, acetil-CoA, care activează şi transportă acizii organici sub formă de derivaţi de acetil CoA
• Energia necesară sintezelor este furnizată de ATP.• Sursele utilizate de bacterii în sinteza lipidelor sunt
variate. De exemplu, M. tuberculosis îşi sintetizează lipidele mai ales pe baza surselor de C furnizate de glicocol şi acetat.

METABOLISMUL ENERGETIC
• Bacteriile - vieţuitoare chemotrofe, întrucât îşi satisfac cerinţele energetice exclusiv pe seama reacţiilor chimice exoergice
• Pe ansamblu, procesele prin care celula bacteriană îşi asigură balanţa energetică pot fi diferenţiate în două categorii:
- reacţii catabolice
- procese respiratorii.
•

Procesele catabolice şi balanţa energetică
• substratul nutritiv macromolecular este degradat prin reacţii succesive şi pe căi foarte variate, până la stadiul de metaboliţi intermediari sau produşi finali (CO2 şi H2O), fiecare etapă furnizând celulei bacteriene o anumită cantitate de energie.
• procesele decurg schematic în trei faze:
- in prima fază, macromoleculele sunt scindate în componentele moleculare constitutive (polizaharidele în monozaharide, proteinele în acizi aminaţi, lipidele în glicerol şi acizi graşi), eliberandu-se o cantitate mică de energie

• faza a doua
- catabolizarea moleculelor continuă până la stadiul de metaboliţi intermediari caracteristici căilor metabolice centrale (piruvat, acetil-CoA, trifosfaţi).
- se eliberează aproximativ 1/3 din energia conţinută în componente.
• faza a treia corespunde de fapt proceselor de respiraţie celularăc - atabolizarea şi eliberarea energiei se poate realiza pe diverse căi, dintre care cele mai importante sunt:– calea acizilor tricarboxilici - catabolizarea substratului nutritiv până la
stadiul de produşi finali (CO2 şi H2O).
– calea fermentativă - metabolizează parţial substratul nutritiv cu formarea în final a unor produşi de fermentaţie (acizi, alcooli).

RESPIRAŢIA LA BACTERII
• reacţiile biochimice de oxido-reducere, prin care celula bacteriană îşi asigură balanţa energetică
• Conceptul de respiraţie bacteriană devine astfel sinonim celui de metabolism energetic
• Procesele de oxidare biologică pun în libertate o cantitate mare de energie.
• Utilizarea raţională a acesteia de către celula bacteriană este asigurată, aşa cum s-a arătat, prin două mecanisme:
– eliberarea fracţionată, în etape, prin oxidoreduceri succesive ale substratului;
– stocarea excedentului de energie în legăturile chimice ale altor componente

• Procesele de oxido-reducere constau în dehidrogenarea (oxidarea) substratului, H fiind transportat prin mijlocirea enzimelor la un substrat acceptor care se reduce.
• În fiecare etapă se eliberează o anumită cantitate de energie, din care celula bacteriană utilizează o parte pentru satisfacerea cerinţelor imediate, iar cealaltă parte o înmagazinează în noi legături chimice de regulă fosfoergice.