Embed Size (px)
Citation preview

P a l a e o n t o g r a p h i c a A b t . В 142 Lfg. 4 - 6 105—116 S t u t t g a r t , J u n i 1 9 7 3
CUTICULAR STRUCTURE OF CRETACEOUS ANGIOSPERMS FROM THE FAR EAST OF THE
USSR BY
VALENTIN KRASSILOV, Vladivostok*)
With Plates 18—26
Abstract Ten species from the Senonian of Sakhalin and Danian of the Amur province (Tsagajan) are described. They are Platanus ray-
noldsii, Araliaephyllum polevoi, Debeya tikhonovichii, Lacuniderma meyenii, Laurophyllum sakhalinense, Limnobiophyllum scutatum, Myriciphyllum yokoyamae, Papilionaceophyllum kryshtofovichii, Protophyllum ignatianum and Trochodendroides arctica. The empha-sis is put on their cuticular structure and some comments on their natural affinities are made.
Contents Introduction 105 Material and methods 106 Descriptions 106
Genus Platanus LINNAEUS 106 Platanus raynoldsii NEWBERRY emend. BROWN 106
Genus Araliaephyllum FONTAINE 107 Araliaephyllum polevoi (KRYSHTOFOVICH) KRASSILOV 1 0 7
Genus Debeya MIQUEL emend. KNOBLOCH 108 Debeya tikhonovichii (KRYSHTOFOVICH) KRASSILOV 1 0 8
Genus Lacuniderma gen. nov 109 Lacuniderma meyenii sp. nov 109
Genus Laurophyllum GOEPPERT 109 Laurophyllum sakhalinense sp. nov 109
Genus Limnobiophyllum KRASSILOV 110 Limnobiophyllum scutatum (DAWSON) comb, nov 110
Genus Myriciphyllum CONWENTZ Ill Myriciphyllum yokoyamae (KRYSHTOFOVICH) KRASSILOV . . . . 1 1 1
Genus Papilionaceophyllum KRAUSEL & WEYLAND I l l
Papilionaceophyllum kryshtofovichii sp. nov Ill Genus Protophyllum LESQUEREUX 112
Protophyllum ignatianum KRYSHTOFOVICH & BAIKOVSKAYA 1 1 2
Genus Trochodendroides BERRY 112 Trochodendroides arctica (HEER) BERRY 112
Conclusions 113
Introduction The revision of the Cretaceous angiosperm taxonomy is badly needed. The cuticular structure is expected to
give some evidence on their natural affinities. However an amount of cuticle studies in the Late Cretaceous floras
* Address of the author: Institute of Biology and Pedology, Far-Eastern Scientific Centre, Vladivostok, USSR.
Palaeontographica Bd. 1^2. Abt. В 14

is negligible. One of the reasons is a scarcity of structurally preserved material in these floras. Only a few frag-ments of leaf cuticles have been obtained as a result of prolonged search through the extensive collections from the Late Cretaceous of Sakhalin and the Amur province. The taphofloras of both regions were described for the first time by O S W A L D H E E R ( 1 8 6 8 — 1 8 8 3 ) and reinvestigated by KRYSHTOFOVICH, who established their Cretaceous age.
Material and Methods The Senonian angiosperms are collected from deposits of chiefly deltaic origin outcropping along the western
coast of Northern Sakhalin. The largest localities (flora of Mgachi) are confined to the lower horizons. Their age is estimated as Coniacian on the evidence of marine invertebrates.
The Maastrichtian—Danian nonmarine deposits crop out along the Amur and its tributaries — Zea, Bureja and Arkhara. They consist of alluvial sands, gravels and shales with abundant plant fossils which constitute the Tsagajan flora. The dinosaur remains are reported from underlying beds.
Leaf compressions from Mgachi were treated with fluoric acid, oxidized with SCHULZE'S mixture ( H N O 3 +
КСЮз, 24 hours), then with natrium hypochlorite (12 hours). Alkali threatment was used in some cases. The cuticles from Tsagajan were macerated with HNOS(l hour) folowed by delute alkali.
Since the natural affinities of the leaf-genera remain uncertain their descriptions are arranged in alphabetical order.
Descriptions
Genus PI at an и s LINNAEUS
Plat anus raynoldsii N E W B E R R Y emend. BROWN
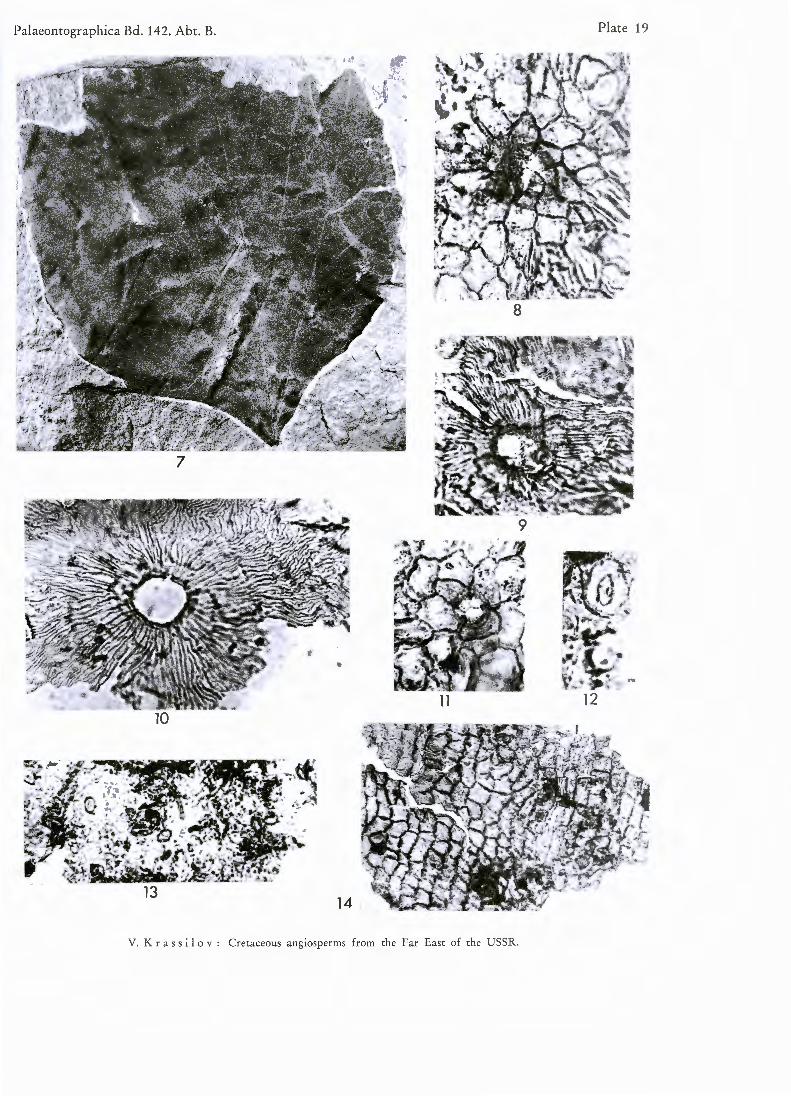
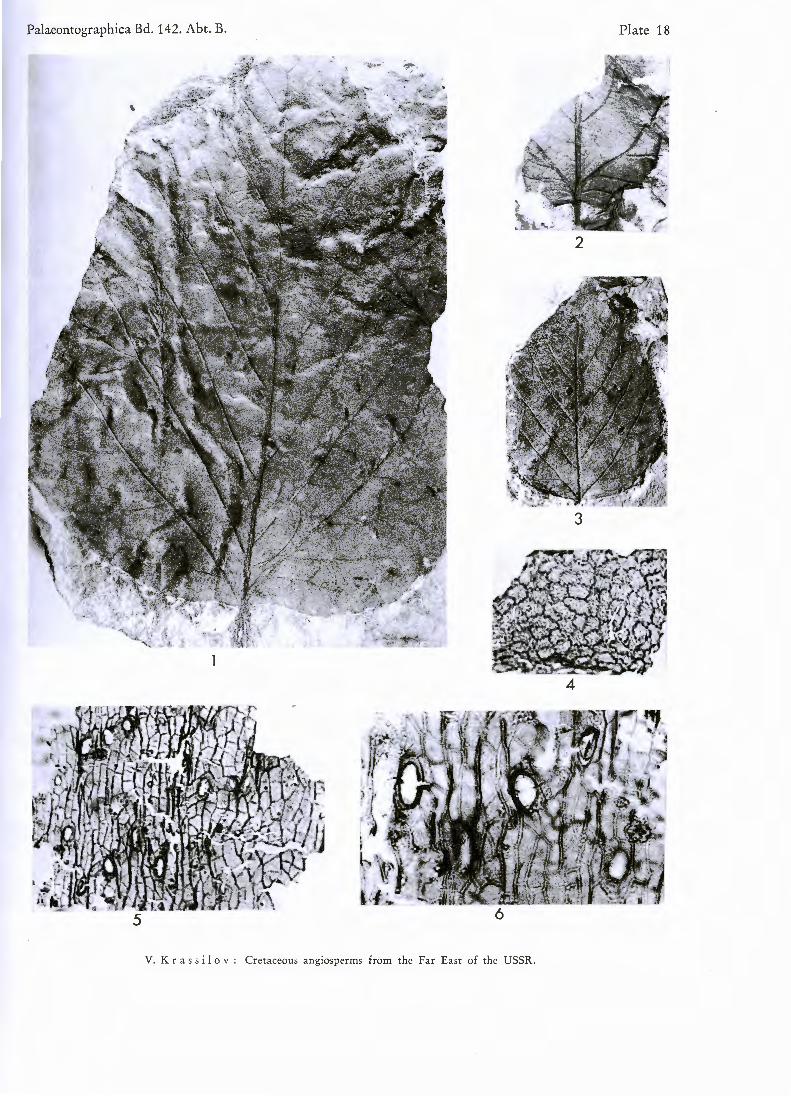
PL 18, figs. 1—6; PL 19, figs. 7—14
1962 Platanus raynoldsii — BROWN, p. 64, PL 30, figs. 1—4; PL 31, figs. 1—6; PL 66, fig. 8. The names applied to the Tsagajan leaves are:
1914 Alnus kefersteinii — KONSTANTOW, p. 9, PL 1, figs. 1—2. 1966 Quercus c f . platania — KRYSHTOFOVICH & BAIKOVSKAYA, p. 246, PL 17, f ig . 5. 1966 Platanus schmidtii — Ibid., p. 286, PI. 8, fig. 3; PL 13, fig. 1; PL 14, fig. 2; PL 16, figs. 2—3; PL 17, fig. 3. 1966 Platanus cf. platanoides — Ibid., p. 288. 1966 Platanus cf. Guillelmae — Ibid., p. 286, PL 6, fig. 1. 1966 Acer arcticum — Ibid., p. 298, Text-fig. 15. 1966 Grewiopsis yukonensis — Ibid., p. 307, PL 21, fig. 1; PL 22. 1966 Cissus amurensis — Ibid., p. 301, PL 13, fig. 2. 1966 Actinidia burejensis — Ibid., p. 308, PL 8, fig. 5. 1966 Pterospermites amurensis — Ibid., p. 309, PI. 7, fig. 2; PL 16, fig. 1; PL 20, fig. 1.
D e s c r i p t i o n : Leaf-blade typically subrhombic or broadly obovate, trilobate or obsoletely lobate, up to 30 cm long. Base peltate, cuneate or deccurrent. Margin provided with glandular teeth or only with inconspicuous glands, entire near the base. Venation palmatopinnate. Supra-basal pair of primary lateral veins vigorously branching on abaxial side and as a rule provided with strong adaxial branch. Next secondaries with one or more distal abaxial branches. Tertiaries scalariform, distant, straight or bowed, simple or once divided. Infrabasal veins of varying number, short, downwardly bending.
Underdeveloped leaves broadly ovate, entire, base rounded or truncate, margin provided with glands. Venation pinnate, basal veins abaxially branching. One pair of thin infrabasal veins generally present or only one of them is developed.
Leaf hypostomatic. Upper cuticle of moderate thickness. Costal cells rectangular, often pointed or wedge-shaped, forming files, 36—40и long, 11—\b tu wide. Cell walls conspicuous, straight. Small dark cells are scatter-ed in costal zones. Intercostal cells polygonal, about 27ju long, 20ju wide, cell walls curved or wavy, occasionally sinuous. Costal trichomes (over veins) rather frequent, trichome bases oval, longitudinally orientated, adjacent cells unspecialized. Intercostal trichomes (or druses) scattered, about 5—6 per 1 mm2. Trichome bases, oval, 27— 32,u long, surrounded by thickened rim of cuticle, disposed over a junction of 5 or 7 epidermal cells which are continuous under trichome or interrupted by small pore (which evidently corresponds to trichome foot) about

12и in diameter. Prominent cutin folds radiate from trichome bases. Away from trichomes they are thin and wavy. Lower cuticle delicate, costal zones similar to those of upper cuticle, trichomes more frequent. Intercostal cell outlines obscure. Stomata irregularly scattered, anomocytic. Guard cells exposed, about 40/.« long, outer stomatal ledges conspicuous.
C o m p a r i s o n : This species belongs to the "orientalis — occidentalis — aceroides" group of shallowly trilobate leaves according to classification, proposed by O I S H I and H U Z I O K A ( 1 9 4 3 ) . Members of "aceroides" group are hardly distinguishable and not clearly differentiated from those of "basilobata" and "guillelmae" groups of this highly conventional classification. Leaves of modern plane-trees are hypostomatic, provided with thick cuticle. Cutin ridges have been observed in all species except P. racemosa which differs from the rest in its fairly frequent stellate trichomes. Anticlinal walls of intercostal cells are straight or slightly wavy (P. orientalis, P. digi-tatay P. acerifolia) undulating or sinuous (P. lindeniana, P. racemosa). Stomata scattered in vein-meshes, anomo-cytic. Exceptional stomata with a pair of narrow paracytic subsidiary cells. Guard cells large, exposed. Our fossil species resembles P. lindeniana in cell outlines, cutin fold pattern and trichomes (trichome base surrounded by thickened rim and radiated cutin folds, adjacent cells reduced in size). Closely similar trichome structures were depicted by KRAUSEL and W E Y L A N D ( 1 9 5 9 , fig. 8 , "kopfartige Druse") in the fossil leaves of "Kadsura" breddini WLD. which are also comparable in their cutin fold pattern.
L o c a l i t y : Amur province near the mouth of Bureja River, Tsagajan. A g e : Maastrichtian-Danian.
Genus Ar ali a e p h у Hum F O N T A I N E
T y p e s p e c i e s : Araliaephyllum obtusifolium FONTAINE 1889.
Araliaephyllum polevoi (KRYSHTOFOVICH) KRASSILOV
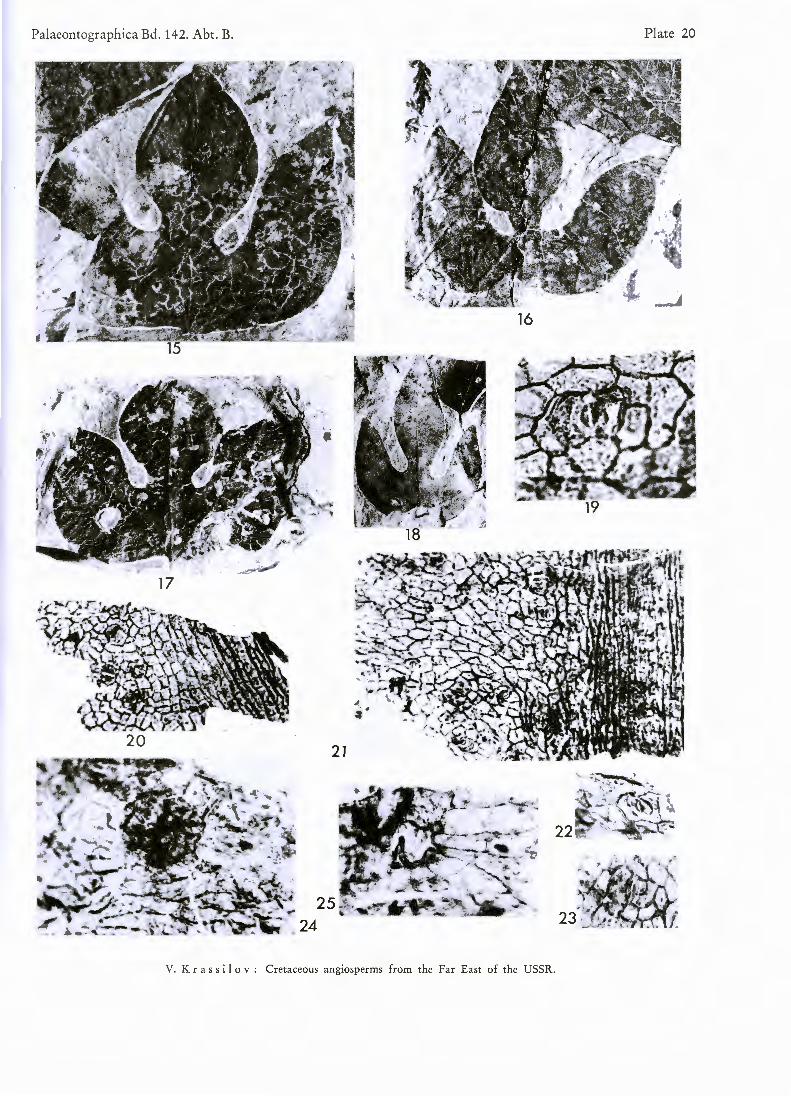
PI. 20, figs. 15—25
1918 Aralia polevoi — KRYSHTOFOVICH, p. 55, Text-fig. 12. 1937 KRYSHTOFOVICH, p. 90, PL 13, figs. 1—9; PL 14, figs. 1—3. 1962 Sassafras polevoii — KRYSHTOFOVICH, p. 276, PL 8, figs. 4—7; PL 10, fig. 6; PL 11, figs. 1, 2, 7; PL 12, figs. 1, 4. 1968 Sachalinella polevoi — IMKHANITZKAYA, p. 642, PL 1, figs. 1, 6—7; PL 2, figs. 1, 8—9; PL 3, figs. 4—5. 1969 Araliaephyllum polevoi— KRASSILOV, p. 77, Text-fig. 1—2.
D e s c r i p t i o n : Leaves typically trilobate, of perfect trident shape, up to 80 mm long, 86 mm wide (average 55 mm long, 40 mm wide). Central lobe narrow elliptical to rounded elliptical, occasionally overlapping lateral lobes. Both sinuses semicircular, on the same level, or occasionally one above the other and of unequal width. Lateral lobes inequilateral with long or shortly pointed tips. Margin entire. Base nearly horizontally trun-cated, slightly concave and deccurent on petiole. Petiole long. Exceptional leaves with reduced lobes, almost en-tire. Venation trinerved, primary veins diverge from the point of petiole insertion. Lateral primary veins are provided on abaxial side with 5 to 6 strong camptodromous branches. Intercalated veins present. Acroscopical branches are relatively short, the proximal one connects with an equaly strong branch of central primary, form-ing angular arch. Upwardly directed vein passes from this arch to the sinus. Distal acroscopic branches of lateral primaries are connected with marginal veins which run along the inner margins of the lobes. Marginal vein moves occasionally away from the margin and then approximates it again. Tertiaries rather distant, bowed, once divided.
Leaf amphistomatic. Upper cuticle of moderate thickness, lower cuticle thin. Costal cells elongated, arranged in rows. Intercostal cells irregularly polygonal or rectangular. Anticlinal walls strongly marked, straight, peri-clinal wall smooth. Stomatal frequency of upper cuticle about 20 per 1 mm2, of lower — twice as much. Sto-mata irregularly orientated, separated by several (in rare cases by only one) ordinary cells. Stomata mostly para-cyctic, some of them with only one subsidiary cell parallel to aperture. Stomatal apparatus, including subsidiary cell pair, nearly rectangular, about 45 ju wide. Subsidiary cells with somewhat thickened distal anticlinal walls, otherwise not specialized. Guard cells slightly sunken, 23—27jli long, outer stomatal ledges strongly developed. Apertures often filled with dark matter. One-celled trichomes scattered, short, conical. Druses rather frequent in lower as well as upper epidermis, rounded, about 58/i in diameter, leaving white round spots when lost.

C o m p a r i s o n : Comparable Cretaceous leaves are known under the names Araliay Cissus, Sassafras, Ben-zion, Araliophyllum, Araliphyllum, Araliaephyllumy Araliopsoides, Cissites, PlatanophyHum, Lauraceaephyllum, Sachalinella and others ( F R I T E L 1 9 1 4 ; BERRY 1 9 1 6 ; SEWARD 1 9 2 6 ; K O C H 1 9 6 3 ; IMKHANITZKAJA 1 9 6 8 ) . Details of their morphology as well as cuticular structure (which was previously unknown) invalidate an attribution to any of modern genera. Form-genus Platanophyllum is rejected since it is based on undeterminable fragments (FON-TAINE 1 8 8 9 ) . Cissites is generally used for a different type of fossil leaves. The name Araliophyllum is originally proposed for compound leaves (ETTINGSHAUSEN 1 8 6 8 ) and Araliphyllum is based on the incomplete entire leaf ( N A T H O R S T 1 8 8 8 ) . Among others, Araliaephyllum has priority. FRITEL ( 1 9 1 4 ) treated the primary vein pattern (i. e. basal versus supra-basal) as criterion for separation Aralia from Araliaephyllum. However, taxonomical significance of this feature is not certain, and FRITEL'S emendation of Araliaephyllum is not adopted here.
IMKHANITZKAJA (1968) advocated the Araliaceous affinity of Sakhalin species, but this cannot be confirmed on cuticular evidence. Cuticular structure resembles those of Sassafras and Lindera, but differs in distribution of stomata (leaves of A. polevoi are ampistomatic) and outlines of stomatal apparatus. It is worth mentioning that Liquidambar formosana has closely similar structure of stomata and trichomes. This species is similar to Araliae-phyllum groenlandicum ( H E E R ) FRITEL, however venation is slightly different. Cuticular structure of Greenland and similar North American leaves (see FRITEL 1914) is not known.
L o c a l i t y : Western coast of Northern Sakhalin, Mgachi. A g e : Senonian.
Genus Debeya M I Q U E L emend. K N O B L O C H
T y p e s p e c i e s : Debeya serrata MIQUEL 1853.
R e m a r k s : The genus Debeya is adopted here in the sense of K N O B L O C H ( 1 9 6 4 ) , i. e. including Dewalquea SAPORTA et M A R I O N and Araliophyllum sensu SCHIMPER. Systematic position of Debeya is obscure. Morphological characters suggest affinity with Proteaceae or Araliaceae. Cuticle was described up to now only for the Irish Ter-tiary species of Debeya (Dewalquea) by J O H N S O N and G I L M O R E ( 1 9 2 1 ) , who referred it to Juglandaceae. K O C H
( 1 9 6 3 ) pointed out the possibility of Picrodendraceae (Juglandaceae) affinities.
Debeya tikhonovichii (KRYSHTOFOVICH) KRASSILOV
PI. 21, figs. 26—34
1918 Aralia tikhonovichii — KRYSHTOFOVICH, p. 57, Text-fig. 13. 1972 Debeya tikhonovichii — KRASSILOV, p. 14 (without description).
D e s c r i p t i o n : Leaf compound, typically trifoliate, exceptional (probably underdeveloped) leaves trilobate. Trifoliate leaf up to 130 mm long, 90 mm wide. Leaflets coriaceous, oblanceolate, acuminate, sessile, diverge from the end of long and stout common petiole. Leaflet base cuneate. Lateral leaflets inequilateral, with midribs acro-scopically naked at base. Margin entire for short distance from the base, serrate above. Serrations delicate, at inter-vals of 2,6—3 mm, forwardly directed. Midrib strong, secondaries obscure (possibly immersed), fairly numerous, at intervals of about 3 mm, about 45° to midrib, straight proximally, slightly flexuous and curved distally, anas-tomosing with intercalated veins, looping along margin and sending short branches to the teeth. Intercalated veins between each pair of secondaries, thin, flexuous,, not reaching the margin. Secondaries and intercalated veins forming irregular network of elongated meshes.
Trilobate leaves with lobes of equal length, or middle lobe shorter than the rest. Base cuneate, asymmetrical. Margin with triangular, acute, forwardly directed teeth which are more prominent than in typical trifoliate leaves. Basal pair of secondaries arising at a short distance above the base. Details of venation as for trifoliate leaves.
Leaf hypostomatic. Upper cuticle thick. Cells polygonal, about 54ju long, irregularly disposed or forming indistinct groups of relatively isodiametric central cell and somewhat elongated cells radiating from it. Costal files of rectangular cells only above the secondaries. Minor veins of higher orders are not reflected in the cell arrangement. Cell walls straight, strongly marked. Periclinal walls with occasional round spots, possibly left by oil droplets. Trichome bases (or druses) rare, rectangular to oval, of variable size. Lower cuticle slightly thinner,

cell outlines same as in upper cuticle. Stomata scattered, about 20 per 1 mm2, evenly spaced or forming indistinct groups of 2—3 approximated complexes, with generally more than one cell between the neighbouring stomas. Stomata anomocytic, subsidiary cells, typically 5 or 6. Lateral subsidiary cells distinguishable from ordinary cells by lighter colour. Encircling cells sometimes present, forming incomplete ring. Guard cells thinly cutinized, about 31—36/л long. Stomatal ledges strongly developed, forming an oval not less than 8;u wide.
R e m a r k s : Trifoliate and trilobate leaves have been thought at first to belong to different species, but their cuticular structures, as well as details of venation, are identical. So far I am inclined to consider them as forms of one species with polymorphous leaves. I found above 30 more or less complete trifoliate leaves and only two trilobate leaves at the same locality.
C o m p a r i s o n : Debeya tikhonovichii differs from the rest of Debeya species (see KNOBLOCH 1964) in sessile, oblanceolate leaflets and fairly frequent secondary veins. The resemblance of cuticular structure to that of Araliaceae or Juglandaceae is rather remote.
L o c a l i t y : Western coast of Northern Sakhalin, Mgachi. A g e : Senonian.
Genus Lac uniderma gen. nov.
D i a g n o s i s : Cuticle with pores protected by the dome-shaped elevations.
T y p e s p e c i e s : Lacuniderma meyenii sp. nov.
R e m a r k s : Dispersed cuticles have recently been classified by MEYEN, SCHNEIDER and other authors. The name Lacuniderma is proposed for dispersed cuticles showing characteristic protected pores which are not related to stomata.
Lacuniderma meyenii sp. nov.
Pl. 22, figs 41—48
D i a g n o s i s : Fragments of stomatiferous (lower) and nonstomatiferous (upper) cuticle. Both cuticles thick, with uniform polygonal cells about 5 Oju long. Anticlinal walls straight, strongly marked. Upper cuticle with numerous irregular craks. Stomata arranged in groups, sunken in stomatal pits. Stomatal pit elliptical or angular, about 70u long, bordered with cutin folds. Guard cells with prominent stomatal ledges, about 45ju long. Subsidiary cells thinly cutinized, narrow, mostly overlapped by stomatal pit border. Upper as well as lower cuticle with scattered elliptical pores (hydathodes?), provided with dome-shaped protection. Orifice of the dome-shaped protection polygonal to irregular, about 36// in diameter, somewhat eccentric. There are occasional paired pores.
H o i о t y p e : Slide 308—78; PL 22, figs. 41—48. N A M E : After Dr. S . V . M E Y E N .
L o c a l i t y : Western coast of Northern Sakhalin, Mgachi. A g e : Senonian.
Genus Laur о p by Hum GOEPPERT
T y p e s p e c i e s : Laurophyllum beilschmiedioides G O E P P E R T , 1 8 5 4 .
R e m a r k s : The leaves described below belong to Lauroceophyllum according to classification of fossil Lauraceae proposed recently by STURM ( 1 9 7 2 ) . However the name Lauroceophyllum STURM is probably homony-mous to Lauraceaephyllum KOCH ( 1 9 6 3 ) . STURM have rejected Laurophyllum, Daphnogene and Cinnamomophyl-lum since the venation characters used as distinguishing features of these genera vary within a single species, e. g. Cinnamomum camphora. My opinion is that more familiar name Laurophyllum may be retained for the Laura-ceous leaves insufficiently preserved for more refined classification.
Laurophyllum sakhalinense sp. nov.
PL 22, figs. 35—40
D i a g n o s i s : Leaf petiolulate, lanceolate, gradually tapering to acute apex and more abruptly to the base, 70 mm long, 10 mm wide. Margin entire. Midrib prominent. Secondaries immersed, obscure, alternate, campto-dromous, arising at acute angle to midrib, at intervals of 2 to 3 mm, slightly flexuous.

Leaf hypostomatic. Cuticles of both sides delicate with similar cells and trichomes. Cells more clearly marked on lower cuticle, irregularly rectangular to polygonal, about 45/* long. Costal zones hardly distinguishable, costal cells somewhat longer than intercostal ones. Cell walls with jagged thickenings, occasionally undulating. Trichomes delicate, acute, curved, 30 to 50// long, fairly frequent, especially along the veins. Oil-cells sparse, rounded, dia-meters 45—50//, some of them with hair-like projections. Stomatal complexes including subsidiary cell pair, broad-ly elliptical, 55 to 68// wide, thinly cutinized, proximal walls of subsidiary cells not thickened. Apertures relati-vely short (about 18//), stomatal ledges inconspicuous.
H o i t y p e : 312—243, Pi. 22, figs. 36—40.
C o m p a r i s o n : Externally similar to " Proteoides" lancifolius H E E R and Laurophyllum flexuosum ( N E W B . )
BELL from the Cretaceous of Canada (BELL 1 9 6 3 ) . Their cuticle structure is not known. Comparable leaves are described by STURM ( 1 9 7 2 ) as Lauraceophylloderma vestibulibrevis STURM. However the Sakhalin leaves are smal-ler, with more frequent trichomes and somewhat larger stomata.
L o c a l i t y : Western coast of Northern Sakhalin, near the mouth of Sartunaj Creek. A g e : Senonian.
Genus Limnobiophyllum KRASSILOV
1970 Limnobiophyllum — KRASSILOV, p. 99 (nom. nud.).
D i a g n o s i s : Leaf reniforme about 12 mm wide, leaf surface tuberous. Leaf blade epistomatic, stomata rare, guard cells and lateral subsidiary cells thinly cutinized, sunken in a pit formed by polar subsidiary cells and encircling cells. Trichomes scattered, simple, short, conical. Druses frequent on the lower epidermis.
T y p e s p e c i e s : Lemna (Spirodela) scutata DAWSON 1875.
R e m a r k s : New generic name is proposed for the leaves described previously from the Cretaceous-Paleo-cene of Nor th America as LEMNA (Spirodela)y Nymphaeites, Carpolithes and Hydromystria (see W A R D 1 8 8 5 ;
D O R F 1 9 4 2 ; BROWN 1 9 6 2 ) . Externally this plant resembles Hydrocharis and Limnobium or Hydromystria, but Hydrocharitaceous (as well as Nymphaeaceous or Lemnaceous) affinity is not supported by cuticular structure. Rather similar cuticles can be found among Araceae. According to STEBBINS 6C K U S H ( 1 9 6 1 ) , two Araceous genera, Scindapsis and Dieffenbachia have 6—7 subsidiary cells grouped in a hap-hazard manner, while other genera have 2—4 subsidiaries. Stomatal complexes sunken in a pit formed by thickened encircling cells are found in several Araceous genera ( W E Y L A N D 1 9 5 7 ) . Comparable modern genus is Pistia which is thought to be related to Araceae.
Similar European Miocene fossils have been described by H E E R ( 1 8 5 9 ) , ETTINGSHAUSEN ( 1 8 5 8 ) and other authors as Hiraea fruits. H A N T K E ( 1 9 5 4 ) proved them to be the floating leaves of Hydromystria type. Their spe-cific and generic identity with Cretaceous-Paleocene plant, advocated by BROWN ( 1 9 6 2 ) , seems to me rather doubtful.
Limnobiophyllum scutatum ( D A W S O N ) comb. nov.
PL 23, figs. 49—61
1875 Lemna (Spirodela) scutata — DAWSON, p. 329, PI. 16, figs. 5, 6, 7a. 1942 Nymphaeites browni — DORF, p. 142, PL 10, fig. 9. 1962 Hydromystria expansa — BROWN, p. 52, PL 16, figs. 1, 3, 8—11. 1965 Spirodela scutata — BELL, PL 16, fig. 6.
D e s c r i p t i o n : Leaf reniforme, about 12 mm wide, 9 mm high, not less than 1,5 mm thick (judged by in-crustation thickness), acroscopic margin cordate, basiscopic margin straight or slightly concave, petiole insertion connate, somewhat removed from the centre of the base. Lamina differentiated into slightly concave middle part and marginal flange 1 mm wide with a narrow groove between them. One surface, presumably lower, densely covered with protuberances. Another surface (upper) smooth or indistinctly pitted (pits are supposed to be im-prints of protuberances through leaf blade). Protuberances cylindrical, 0,2 mm in diameter, 0,3 mm high, notched at the apex. BROWN ( 1 9 6 2 ) interpreted protuberances as fillings of air chambers, but it seems to me that they re-

fleet originally tuberous surface. Venation generally obscure but where visible consists of numerous veins radiat-ing from petiole insertion point, anastomosing and producing elongated polygonal meshes.
Leaf presumably epistomatic. Upper cuticle of moderate thickness, costal and intercostal areas indistinguish-able. Cells polygonal, about 30—35/x long. Anticlinal walls straight, strongly marked. Outer wall flat, evenly thickened. Stomata about 10 per 1 mm2, irregularly orientated. Stomatal pit oval, up to 50// long, bordered by 8 to 12 cells from which 2 are polar subsidiary cells and the rest—encircling cells. Cells of pit border somewhat smaller than ordinary epidermal cells. Polar cells bulging over stomata, sometimes modified in a hair-like struc-tures. Guard cells exposed along with 1 thin-walled lateral subsidiary cell parallel to the aperture. Exceptional stomata with 2 or none (at least none exposed) lateral subsidiary cells. Outer stomatal ledges conspicuous. Several stomatal pits are empty, stomata probably replaced by druses. Trichomes scattered, trichome bases irregularly rectangular, thickened, about 20/« long, surrounded by several rings of ordinary cells.
Lower cuticle delicate. Cells elongated, pointed, forming files, intercostal cells broader than costal. Stomata absent. Druses fairly frequent, spheroid, diameters 29—35/<, filled with dark granular contents, leaving round spots when lost.
L o c a l i t y : Amur province, near the mouth of Bureja River, Tsagajan. A g e : Maastrichtian—Danian.
Genus Myriciphyllum CONWENTZ
T y p e s p e c i e s : Myriciphyllum oligocenicum C O N W E N T Z 1886.
Myriciphyllum yokoyamae (KRYSHTOFOVICH) KRASSILOV
PL 24, figs. 66—72
1918 Celastrophyllum yokoyamae — K R Y S H T O F O V I C H , p. 54, Text-fig. 11. 1972 Myriciphyllum yokoyamae — K R A S S I L O V , p. 50, P L 22, figs. 1—8.
D e s c r i p t i o n : Leaf elongate-lanceolate, 10—30 mm wide, gradually tapering towards acute apex. Base cuneate. Margin entire below, crenulate or serrate above. Midrib prominant, secondaries obscure, arising 45 degrees to midrib at interval of 8 to 10 mm, alternate, forking and arching near the margin, giving a short basiscopic branch which enters the tooth. There are several leaves which are distally forked.
Leaf hypostomatic. Lower cuticle of moderate thickness. Cells polygonal, anticlinal cell walls straight. Sto-mata represented by rounded gaps about 25и in diameter. Trichome bases frequent, diameters 45 to 5 0 T r i c h o m e s presumably stellate, isolated branch fragments are scattered all over the surface.
C o m p a r i s o n : This leaf-type resembles Celastrophyllum grandifolium (NEWBERRY 1 8 9 5 ) as well as Myrica hollickii Ward (BERRY 1 9 1 1 ) , M. torreyi (KNOWLTON 1 9 0 0 ) and M. trifoliata (HOLLICK 1 9 3 0 ) . Forked leaves are described in Myrica conifer a from the Paleogene of Zajsan (ILJINSKAJA 1 9 6 3 ) . The venation and trichomes of Sakhalin species suggest rather Myricaceous than Celastraceous affinities.
L o c a l i t y : Western coast of Northern Sakhalin, Mgachi.
A g e : Senonian.
Genus Papilionaceophy Hum KRAUSEL & WEYLAND
T y p e s p e c i e s : Papilionaceophyllum liblarense K R A U S E L be W E Y L A N D 1 9 5 9 .
Papilionaceophyllum kryshtofovichii sp. nov.
PL 25, figs. 73—82
D i a g n o s i s : Leaflets of compound leaf. Leaf blade elliptical, inequilateral, apex obtusely pointed, margins entire, 45—56 mm long, 18—27 mm wide. Petiole short (up to 6 mm long), slightly curved. Venation camptodromous; midrib stout, secondaries 7 pairs, thin, arched, alternated; tertiaries obscure.
Leaf hypostomatic. Upper cuticle thin. Cells rectangular to polygonal or irregular. Cell walls straight, not strongly marked. Trichomes sparse, trichome base oval or rounded, about 20/г in diameter, adjacent cells unspecia-lized. Lower cuticle of the same thickness, clearly differentiated into costal bands over the veins and intercostal

stomatiferous meshes. Costal cells rectangular, straight walled, about 45/* long, 8// wide, forming files. Intercostal cells polygonal or irregular, about 16/* long, 12/* wide, walls curved but not sinuous. Stomata monocyclic, fairly frequent, irregularly orientated, closely packed, neighbouring stomata have common subsidiary cells (communo-cytic) or not more than one ordinary cell between them. Subsidiary cells typically 8, smaller than ordinary cells, not thickened, forming irregular ring 1 0/* wide bordering stomatal pit. Stomatal pit rounded, 22—23/* in dia-meter. Guard cells slightly sunken, thinly cutinized, aperture with inconspicuous outer ledges. Some subsidiary as well as ordinary cells with papilla-like projections which probably represent basal cells of two-celled tricho-mes. Cuticle of petiole of moderate thickness. Cells narrow, forming files. Trichomes frequent, two-celled, distal cell conical, with dark central strand.
H o l o t y p e : 567—716, PL 25, figs. 73, 75—82. N a m e : After Dr. A. N . KRYSHTOFOVICH.
C o m p a r i s o n : These leaflets agree with Papilionaceophyllum liblarense (KRAUSEL & W E Y L A N D 1 9 5 9 ) in details of stomata and trichomes. Druses were not seen in Tsagajan material. Leaflets of P. liblarense have much longer petioles and differ from P. kryshtofovichii in venation ( W E Y L A N D et al. 1 9 5 7 ) .
L o c a l i t y : Amur province, near the mouth of Bureja River, Tsagajan. A g e : Maastrichtian—Danian.
Genus Protophyllum LESQUEREUX
T y p e s p e c i e s : Protophyllum sternhergii LESQUEREUX 1874.
Protophyllum ignatianum KRYSHTOFOVICH & BAIKOVSKAJA
PL 24, figs. 62—65
1960 Protophyllum ignatianum — KRYSHTOFOVICH & BAIKOVSKAJA, p. 89, T e x t - f i g s . 3 1 — 3 4 .
D e s c r i p t i o n : This species comprises broadly ovate middle sized leaves. Base more or less distinctly pel-tate. Venation pinnate, secondaries, about 12 pairs, most of them branched. Cells of upper epidermis rectangular, about 36/* long. Anticlinal cell walls with jagged thickenings. Druses frequent, about 68/* in diameter. Thick cutin ridges radiate from the druse opening in star-like fashion.
R e m a r k s : R U F F L E (1958) suggested Menispermaceous affinity for the genus Gredneria, which is only conventionally separated from Protophyllum. Our species, P. ignatianum is hardly comparable to any member of the Menispermaceae. I t agrees with Liquidambar in leaf morphology and some cuticular features.
L o c a l i t y : Western coast of Northern Sakhalin, Mgachi.
A g e : Senonian.
Genus Trochodendroides BERRY
T y p e s p e c i e s : Trochodendroides rhomhoideus (LESQUEREUX) BERRY 1922.
R e m a r k s : Affinities of this genus were much discussed. BERRY coined the name Trochodendroides for the leaf-type, related not to the genus Trochodendron itself, but to the Trochodendraceae in old sense, i. e. in-cluding Tetracentron and Cercidiphyllum. Many authors placed this leaf-type in the genus Cercidiphyllum. Few anatomical features are distinguished in our material. They are not consistent with attribution either to Cerci-diphyllum or to any other extant genus. Trochodendroides was the most successfull genus among the early angio-sperms. Its abundance in Uppermost Cretaceous taphofloras justifies the name "Trochodendroides floras" ad-opted for them. Trochodendroides possessed aspen — like leaves — mechanically ideal type of deciduous leaf, which was retained in many recent angiosperms of different families.
Trochodendroides arctica ( H E E R ) BERRY
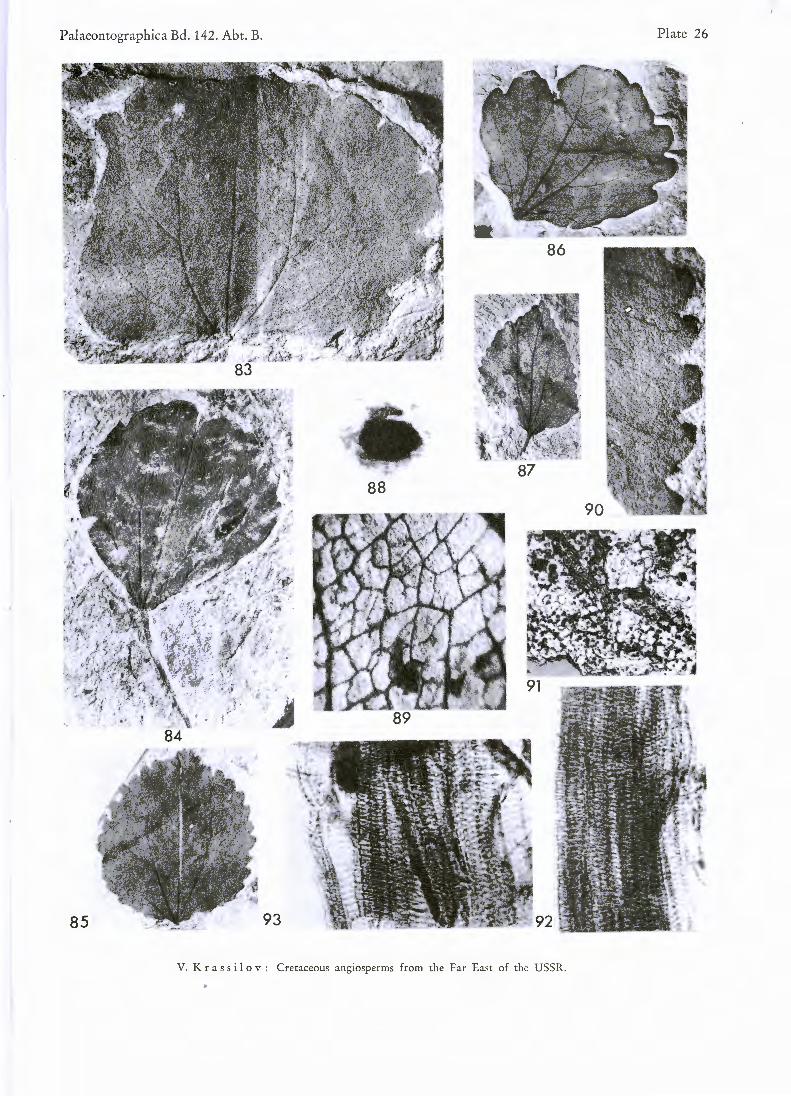
PL 26, figs. 83—92
The names applied to Tsagajan leaves of this type are:
1878 Populus arctica — HEER, p. 49, PL 15, figs. 3—5. 1914 Grewia ohovata — KONSTANTOV, p. 15, PL 5, figs. 5—8.

1914 Populus arctic а — Ibid., p. 16, PL 3, figs. 1, 2, 6. 1914 Populus zaddachii — Ibid., p. 19, PL 3, fig. 7; Pi. 4, fig. 4. 1914 Zizyphus hyperboreus — Ibid., p. 24, PL 5, fig. 2; PL 4, fig. 3. 1966 Ficus praetrinervis — K R Y S H T O F O V I C H & B A I K O V S K A J A , p. 248. 1966 Trochodendroides arctica — Ibid., p. 254, PL 7, figs. 3, 4; PL 8, figs. 1, 2, 7; PL 9, fig. 7; PL 10. figs. 4, 5, 7; PL 13, fig. 6;
PL 14, fig. 5; PL 15, fig. 2; Pl. 17, fig. 4; PL 20, fig. 4; PL 21, fig. 5. 1966 Trochodendroides cf. genetrix — Ibid., p. 261, PL 19, fig. 1. 1966 Trochodendroides smilacifolia — Ibid., p. 265, PL 9, fig. 3; PL 11, figs. 3, 4; PL 12, fig. 3; PL 13, fig. 5; PL 21. fig. 4. 1966 Trochodendroides speciosa — Ibid., p. 269, PL 7, fig. 1; PL 11, figs. 1, 2, 5, 6; PL 12, fig. 1, 2; PL 17, fig. 2, Text-fig. 9. 1966 Trochodendroides elliptica — Ibid., p. 272, PL 8, fig. 8; PL 10, fig. 8; PL 13, fig. 4. 1966 Cercidiphyllum crenatum — Ibid., p. 273. 1966 Tetrazentron amurense — Ibid., p. 281, PL 17, fig. 1; PL 18, fig. 2. 1966 Zizyphus fibrillosa — Ibid., p. 292. 1966 Zizyphus hyperborea — Ibid., Text-fig. 14a. 1966 Zizyphus phosphoria — Ibid., p. 294, PL 17, fig. 7. 1966 Paliurus cf. Colombii — Ibid., p. 297, PL 13, fig. 3, Text-fig. 14b. 1966 Dicotylophyllum tsagajanicum f. В — Ibid., p. 314, PL 14, fig. 3, Text-fig. 19b.
D e s c r i p t i o n : Four types of leaf blade are distinguishable in the Tsagajan material: 1) broadly ovate, apex triangular, base cordate; 2) nearly circular, apex rounded or truncate, base broadly cuneate; 3) obovate, apex nearly horizontally truncate or notched, base cuneate; 4) narrow ovate or elliptical, apex pointed, base cuneate and deccurent on petiole. There are wide range of transitional forms between them. Margin entire to crenulate or serrate. This characteristic is not correlated with leaf shape. Marginal teeth constantly provided with glands. Even leaves with entire margin show marginal glands. Petiole at least three times longer than leaf blade. Venation palmate-trinerved. Primary lateral veins subopposite, in exceptional leaves disposed one above the other. Original monochasial branching is apparent in such cases. Area between primary laterals is filled with arching tertiaries. Peripheral areas show series of loops, diminishing towards the margin. Network of minor veins, with polygonal meshes. Blind endings not seen.
Cuticle is extremely delicate. Cellulose film replicas show polygonal cells with straight walls and few ano-mocytic stomata. Carefull maceration revealed conducting elements of primary vein with spiral thickening. Oval bodies up to 300fi long, representing a contents of secretorgans have been extracted from the sheath of primary vein.
R e m a r k s : Long petiole, marginal glands and sympodial branching of primary veins are characteristic features of Tsagajan leaves. It is not certain, whether these features would help to distinguish among the "species" of Trochodendroides. There were several attempts to define the natural affinities of Trochodendroides complex (e. g. WOLFE 1 9 6 6 ) . Some leaf types were attributed to Cocculus, Cercidiphyllum and other living genera. How-ever the Trochodendrocarpus fructifications constantly associated with Trochodendroides leaves suggest an extinct genus of Hamamelidaceous affinities (KRASSILOV 1 9 7 3 , in press).
L o c a l i t y : Amur province, near the mouth of Bureja River, Tsagajan. A g e : Maastrichian—Danian.
Conclusions
Only one out of ten leaf types from the Upper Cretaceous of Northern Sakhalin and Amur province is attri-buted to modern genus Platanus on the evidence of cuticle structure. The rest of them belong to extinct genera. This contradicts the opinion that a majority of modern angiosperm genera have appeared in Mesozoic time and that they constituted the bulk of Late Cretaceous floras.
Prevailing cuticular characters of Cretaceous angiosperm leaves have been straight anticlinal cell walls, simple few-celled hairs, head-shaped or peltate druses and anomocytic stomata. The cuticular structure have been less diversified than in modern or even Eocene angiosperms. This suggests comparatively low taxonomical as well as ecological diversity of Cretaceous angiosperms.
Palaeontographica Bd. 142. A b t . В 15

References
BELL, W. A (1963): Upper Cretaceous floras of the Dunvegan, Bad Heart and Milk River formations of Western Canada. — Geol. Surv. Canada Bull., 94: 1—76.
—,— (1965): Illustrations of Canadian Fossils. Upper Cretaceous and Paleocene plants of Western Canada. — Geol. Surv. Canada, Pap. 63—65: 1—46.
BERRY, E. W. (1911): The flora of the Raritan formation. — Bull. Geol. Surv. N e w Jersey, 3. —,— (1916): The Upper Cretaceous deposits of Maryland. — Maryland Geol. Surv., 757—901. —,— (1922): The flora of the Woodbine sand at Arthurs Bluff, Tex. — U. S. Geol. Surv. Prof. Paper, 129: 153—180. BROWN, R. W. (1962): Paleocene flora of the Rocky Mountains and Great Plains. — U . S . Geol. Surv. Prof. Paper, 375: 1 — 119. DAWSON, J. W. (1875): Report on the geology and resources in the vicinity of the Forty-Ninth Parallel. Appendix A. Notes on
the plants collected by G. M. DAWSON from the lignite Tertiary deposits. — British N . A. Boundary Commission, 327—331. DORF, E. (1942): Flora of the Lance formation in its type locality, Niobrara County, Wyoming. — Carnegie Inst. Wash. Publ., 850:
79—159. ETTINGSHAUSEN, C. (1858): Beitrage zur Kenntnis der fossilen Flora von Sotzka in Untersteiermark. — K. Akad. Wiss. Sitzungsber.,
28 (6): 471—567. FONTAINE, W. M. (1889): The Potomac or Younger Mesozoic flora. — U. S. Geol. Surv. Monogr., 15: 1—377. FRITEL, P. H . (1914): Note sur les Aralias des flores cretaciques de l'Amerique du Nord et du Groenland. — Bull. Soc. Geol. France,
14 (1): 1—22. GOEPPERT, H. R. (1854): Die Tertiarflora auf der Insel Java. — Elberfeld. HANTKE, R. (1954): Die fossile Flora der obermiozanen Oehinger—Fundstel le Schrotzburg. — Denkschr. Schweiz Naturforsch. Ges.
Mem. 50 (2): 27—118. HEER, O. (1859): Flora Tertiaria Helvetiae, 3. Winterhur. —,— (1868—1883): Flora fossilis arctica, 1—7. Zurich. —,— (1878): Beitrage zur fossilen Flora Sibiriens und des Amurlandes. — Mem. FAcad. Imp. Sci. St.-Petersbourg, 7 ser., 25 (6):
1—58. HOLLICK, A. (1930): The Upper Cretaceous floras of Alaska. — U. S. Geol. Surv. Prof. Paper, 159: 1—116. ILYINSKAIA, I. A. (1963): The fossil f lora of the mountain Kiin-Kerish, Zaisan basin, pt. 2. — Acta Inst. Bot. Acad. Sci. URSS, ser. 8,
Paleobotanica, 4: 143—187 [in Russian]. IMKHANITZKAYA, N . N . (1968): On the problem of authenticity of Sassafras specimens found in the Cretaceous deposits of the USSR.
— Bot. J., 53 (5): 639—651 [in Russian]. JOHNSON, T. & GILMORE, J. G. (1921): The occurrence of Dewalquea in the coal-bore at Washington Bay. — Sci. Proc. Royal Soc.
Dublin, 16. KNOBLOCH, Е. (1964): Neue Pflanzenfunde aus dem siidbohmischen Senon. — Jb. Staatl. Mus. Miner. Geol., 133—201. KNOWLTON, F. H . (1900): Flora of the Montana formation. — U. S. Geol. Surv., Washington. KOCH, В. Е. (1963): Fossil plants from the Lower Paleocene of the Agatdalen (Angmartussut) area, Central Nugssuaq peninsula, North-
west Greenland. — Medd. om Gronland, 172 (5): 1—120. KONSTANTOW, S. W. (1914): Tertiary flora from the White Mountain outcrop near the mouth of the Bureja. — Travaux Geol. Com.,
new ser. 113: 1—27 [in Russian]. KRASSILOV, V. A. (1969): Some problems of investigation of the Cretaceous floras using the cuticular analysis. — In L . N . VASSILJEVA
(ed.) "Problems of botany in the Far East". Vladivostok, 73—84. —,— (1970): N e w data on the Tsagajan beds and their taphoflora. — Proc. USSR Acad. Sci., Geol. Ser., (12): 97—101. —,— (1972): The simultaneity of Cenozoic and Cenophytic lower limits. — Proc. USSR Acad. Sci., Geol. Ser., (3): 9—15. —,— (1973): Mesozoic plants and the problem of angiosperm ancestry. — Lethaia (in press). KRAUSEL, R. & WEYLAND, H . (1959): Kritische Untersuchungen zur Kutikularanalyse tertiarer Blatter (4). — Palaeontographica,
105 B , 5 — 6 : 1 0 1 — 1 2 4 .
KRYSHTOFOVICH, A . N . (1918): On the Cretaceous flora of Russian Sakhalin. — J. Coll. Sci. Imp. Univ. Tokyo, 40 (8). —,— 1937): Cretaceous flora of Sakhalin. Mgach and Polovinka. — Trans. Far-Eastern Branch of Acad. Sci. USSR, ser. geol.,
2: 1—103 [in Russian]. —,— (1962): Cretaceous flora of Sakhalin. Mgach and Polovinka. — In A . N . KRYSHTOFOVICH "Selected Works, 2". Lenin-
grad, 211—289 [in Russian]. KRYSHTOFOVICH, A . N . & BAIKOVSKAJA, T . N . (1960): Cretaceous flora of the Sakhalin. Acad. Sci. USSR. Leningrad [in Russian]. —,—, —,— (1966): The Late Cretaceous flora of the Tsagajan in the Amur district. — In A . N . KRYSHTOFOVICH, "Selected Works,
3". Leningrad, 184—320 [in Russian]. LESQUEREUX, L. (1874): Contributions to the fossil flora of Western Territories (1). The Cretaceous flora. — U . S . Geol. Surv. Terr.
Rept., 6: 1 — 136. MIQUEL, F. A. W. (1863): De fossiele plantem van het Krijt in het Hertogdom. — Geol. Kaart Nederlandsche Verh., 35—56. NATHORST, A. G. (1888): Zur fossilen Flora Japan's. — Palaeont. Abh., 4: 195—250. NEWBERRY, J. S. (1895): The flora of the Amboy clays. — U. S. Geol. Surv. Monogr., 26: 1—137. OISHI, S. & HUZIOKA, K. (1943): Studies on the Cenozoic plants of Hokkaido and Karahuto (6). On the Tertiary Platanus from
Hokkaido and Karahuto. — J. Fac. Sci. Hok . Imp. Univ. , 7 (1): 103—114.

RUFFLE, L . ( 1 9 6 8 ) : Merkmalskomplexe bei alteren Angiospermen-Blattern und die Kutikula von Credneria ZENKER (Menispermaceae). — P a l a e o n t o g r a p h i c a , 1 2 3 B : 1 3 2 — 1 4 3 .
SEWARD, A. C. (1926): The Cretaceous plant-bearing rocks of western Greenland. — Royal Soc. London. — Philos. Trans., 215 B: 57—174.
STEBBINS, G. L. & KHUSH, G. S. (1961): Variation in the organization of the stomatal complex in the leaf epidermis of monocoty-ledons and its bearing on their phylogeny. — Amer. J. Bot., 48 (1): 51—59.
STURM, M. (1971): D ie Eozane Flora von Messel bei Darmstadt. 1. Lauraceae. — Palaeontographica, 134 В: 1—60. WARD, L. (1885): Synopsis of the flora of the Laramie group. — U . S. Geol. Surv. Ann. Rept., 6: 405—570. WEYLAND, H . (1957): Kritische Untersuchungen zur Kutikularanalyse tertiarer Blatter (3). — Palaeontographica, 103 В (1—3):
34—74. WEYLAND, M.; KILPPER, K. & BERENDT, W. (1967): Kritische untersuchungen zur Kutikularanalyse tertiarer Blatter (7). — Palaeon-
tographica, 120 В (5—6): 151—168. WOLFE, J. A. (1966): Tertiary plants from the Cook Inlet region, Alaska. — Geol. Surv. Prof. Paper, 398 В: 1—32.
Explanation of Plates
Plate 18
Figs. 1—6. Platanus raynoldsii N E W B . emend. B R O W N
Figs. 1—3 Fig. 4. Fig. 5. Fig. 6.
Leaves of different shape and dimensions. X 1. Upper cuticle, cells with sinuous walls. X 146. Trichome bases of costal zone. X 146. Trichome bases of costal zone. X 395.
Plate 19
Figs. 7 — 1 4 . Platanus raynoldsii N E W B . emend. B R O W N
Fig. 7 .
Figs. 8—11 Fig. 1 2 .
Fig. 1 3 .
Fig. 1 4 .
Trilobate leaf. X 1. Trichome bases and cutin folds of upper cuticle. X 395. Stoma. X 395. Lower cuticle showing stomata and trichomes. X 146. Upper cuticle. X 146.
Plate 20
Figs. 1 5 — 2 5 . Araliaephyllum polevoi ( K R Y S H T . ) K R A S S I L O V
Figs. 15—18 Fig. 1 9 .
Figs. 2 0 — 2 1
Figs. 2 2 — 2 3
Figs. 2 4 — 2 5
. Leaf shape variation. X 1. Stoma of lower epidermis. X 562.
. Upper cuticle showing stomata and hairs. X 146.
. Stomata of upper epidermis. X 395.
. Peltate druse and druse opening. X 395.
Plate 21
Figs. 2 6 — 3 4 . Debeya tikhonovichii ( K R Y S H T . ) K R A S S I L .
Figs. 2 6 — 2 7
Fig. 2 8 .
Fig. 2 9 .
Fig. 3 0 .
Fig. 3 1 .
Figs. 3 2 — 3 4
. Trifoliate leaves. X 1. Trilobate leaf. X 1. Distribution of stomata. X 175. Hair base of upper epidermis. X 146. Lower cuticle showing stomata. X 146.
. Stomata. X 395.
Plate 22
Figs. 3 5 — 4 0 . Laurophyllum sakhalinense sp. nov.
Figs. 3 5 — 3 6
Fig. 3 7 .
Figs. 3 8 — 3 9
Fig. 4 0 .
. Leaves. X 1. Lower cuticle showing stomata. X 146.
. Stomata. X 395. Oil cells. X 395.

Figs. 41—48. Lacuniderma meyenii sp. nov.
Figs. 41—44. Stomata. X 395. Fig. 45. Paired pores. X 395. Figs. 46—47. Pores with dome-shaped protections. X 395. Fig. 48. Pore devoid of protection. X 395.
Plate 23
Figs. 49—61. Limnobiophyllum scutatum (DAWSON) comb. nov.
Figs. 49—50. Both sides of the same leaf showing tubercles. X 3. Fig. 51. Asymmetrical leaf. X 3. Figs. 52—53. Both sides of the same leaf. X 3. Fig. 54. Laef showing venation. X 3. Fig. 55. Upper cuticle showing stomata. X 146. Figs. 56—59. Stomata with different degree of exposure of guard cells and lateral subsidiary cells. The hair-like projections of
polar cells are seen in Figs. 56 and 58. X 395. Fig. 60. Lower cuticle with oil cells and oil droplets. X 146. Fig. 61. Druse opening surrounded by hairs. X 146.
Plate 24
Figs. 6 2 — 6 5 . Protophyllum ignatianum KRYSHT. & BAIK.
Figs. 62—63. Leaf apex and base. X 1. Figs. 64—65. Upper cuticle with druse openings. X 146.
Figs. 66—72. Myriciphyllum yokoyamae (KRYSHT.) KRASSIL.
Figs. 66—70. Leaf fragments, one of them (fig. 68) forked. X 1. Fig. 71. Cuticle with the fragments of hair brandies. X 146. Fig. 72. Hair bases. X 146.
Plate 25
Figs. 73—82. Papilionaceophyllum kryshtofovichii sp. nov.
Figs. 73—74. Leaflets. X 1. Fig. 75. Upper cuticle with hair bases. X 146. Fig. 76. Lower cuticle showing closely packed stomata; phasc-contrast illumination. X 146. Fig. 77. Stomata between the brandies of diverging costal zone. X 395. Figs. 78—82. Distribution of stomata. X 562 (figs. 78, 80), 395 (figs. 79, 81), 146 (fig. 82).
Plate 26
Figs. 83—92. Trochodendroides arctica (HEER) BERRY
Figs. 83—87 Fig. 88. Fig. 89. Fig. 90. Fig. 91. Figs. 92—93
Leaf shape variation. X 1. Resin body from the primary vein sheath. X 58. Details of venation. X 7. Glandular teeth. X 3. Cellulose film replica showing cell outlines and stomata. X 146. Conducting elements of primary vein. X 395.
Palaeontographica Bd. 142. Abt. B. Plate 26
V. K r as s i I o v : Cretaceous angiosperms from the Far East of the USSR.

V. K r ass i 1 o Y : giosperms Cretaceous an h Far East o from t e
Plate 25
f the USSR.

PalaeontographicaBd. 142. Abt. B. Plate 24
62
64
63
66 68 69
71
67
72 70
V. Krass i 1 o v : Cretaceous angiosperms from the Far East of the USSR.

Palaeontographica Bd. 14 2. Abt. B. Plate 23
54 60
61
V. Krass i I o v : Cretaceous angiosperms from the Far East of the USSR.

Palaeontographica Bd. 142. Abt. B. Plate 22
V. K r a s s i I o v : Cretaceous angiosperms from the Far East of the USSR.

Palaeontographica Bd. 142. Abt. B. Plate 21
V. K r a s s i 1 o v : Cretaceous angiosperms from the Far East of the USSR.
Palaeontographica Bd. 142. Abt. B. Plate 20
16
17
20
V. K r as s i I o v : Cretaceous angiosperms from the Far East of the USSR.
Palaeontographica Bd. 14 2. Abt. B. Plate 19
8
7
9
12 10
13 14
V. K r as s i I o v : Cretaceous angiosperms from the Far East of the USSR.
Palaeontographica Bd. 14 2. Abt. B. Plate 18
2
3
4
5
V. K r a s s i 1 o v : Cretaceous angiosperms from the Far East of the USSR.



![Rice OsGL1-6Is Involved in Leaf Cuticular Wax Accumulation and … · 2018. 4. 3. · quired for thetransport of wax components fromthe epidermal cells to the cuticle [23]. LTPG in](https://img.pdfslide.tips/doc/110x75/60d8d47d0935ae24ea171ab6/rice-osgl1-6is-involved-in-leaf-cuticular-wax-accumulation-and-2018-4-3-quired.jpg)

![BENVENUTI A OSLO - mytravelife.com · $ 0- " ; *0/ & & 416/ 5*/ 0 3hu od frod]lrqh r xqr vsxqwlqr sxrl vfhjolhuh xq ghol]lrvr edu vx vwudgd rssxuh xq hfrqrplfr fklrvfr ghooh fdwhqh](https://img.pdfslide.tips/doc/110x75/5e32c10a038c123f03475efb/benvenuti-a-oslo-0-0-416-5-0-3hu-od-frodlrqh-r-xqr.jpg)







![vuelos herrera de pisuerga · e 9lhqwr fux]dgr (v ho vljxlhqwh odgr gho uhfwiqjxor dg\dfhqwh d ylhqwr hq fdud \ xqd wud\hfwruld shushqglfxodu d od slvwd f 9lhqwr hq frod 7udpr sdudohor](https://img.pdfslide.tips/doc/110x75/5fbc5eea0b748e3cc8468cdc/vuelos-herrera-de-pisuerga-e-9lhqwr-fuxdgr-v-ho-vljxlhqwh-odgr-gho-uhfwiqjxor.jpg)





![BEFANA A MATERA 2020 · 2019-10-24 · sduwhq]d shu lo ulhqwur lq krwho fhqd h shuqrwwdphqwr *,2512 3ulpd frod]lrqh lq krwho h sduwhq]d shu %dul h ylvlwd jxlgdwd gho fdsroxrjr sxjolhvh](https://img.pdfslide.tips/doc/110x75/5ebffc368ad5475862755ee2/befana-a-matera-2020-2019-10-24-sduwhqd-shu-lo-ulhqwur-lq-krwho-fhqd-h-shuqrwwdphqwr.jpg)


![FROD DALAM URUS NIAGA TANAH: SATU KAJIAN … · 16 Per Romilly MR in Green v Nixon (1857) 23 Beav 530 dan 535, 53 ER 208 17 [1926] AC 101. 18 Ibid,. 138. Lihat juga Datuk Jaginder](https://img.pdfslide.tips/doc/110x75/607f599a03b01f62c10f825f/frod-dalam-urus-niaga-tanah-satu-kajian-16-per-romilly-mr-in-green-v-nixon-1857.jpg)
![villach velden klagenfurt 2019 - Yellow Rabbit...3ulpd frod]lrqh lq +rwho h sduwhq]d shu ./$*(1)857 fdsroxrjr ghood &dulq]ld ulffd gl wudgl]lrql h prqxphqwl 7hpsr d glvsrvl]lrqh shu](https://img.pdfslide.tips/doc/110x75/5f4cdffe917bd52cb2220e10/villach-velden-klagenfurt-2019-yellow-3ulpd-frodlrqh-lq-rwho-h-sduwhqd.jpg)

![5+ (55’ (55 ,1*( %$1’ ,,n1other.hjfile.cn/st/2016/07/27/1308a1ed604605f3fb534b98d99d77fb.pdf · ‰ 5hl$qjho n # kwws˛ frod herrn] iel gh ˘](https://img.pdfslide.tips/doc/110x75/5e5a1e5e241eb13eec30a730/5-55a-55-1-1a-a-5hlqjho-n-kwws-frod-herrn-iel-gh-.jpg)



![BRETAGNA NORMANDIA E CASTELLLI APRILE 2020...$JHQ]LD GL 9LDJJL 6$17,1, 9,$**, 785,602 9LD 2OLQWR 'LQL &DVWHOQXRYR GL *DUIDJQDQD HPDLO VDQWLQLYLDJJL#\DKRR LW *,2512 3ULPD FROD]LRQH](https://img.pdfslide.tips/doc/110x75/6014297e59c3f822762e854c/bretagna-normandia-e-castellli-aprile-2020-jhqld-gl-9ldjjl-6171-9.jpg)