Embed Size (px)
Citation preview

I SOLYMVARI All E
BY DE SOORT A&RoTis 5TOLONIFERA L.
LEO SOLDAAT
doctoraalverslag
plantenoecologie /populatiegenetica
februari — juni 1985
1111
1110
110 I
UI 0 Iii Ill I ill
•__—•I0
110
1111
11 ''I
0
11
11
I
I
''I
111
III
1111
I0
1111
101
I
Iii
IIll
III
111
llIll
I 111
101
I 00
0 1(11)
II 1101
o iiiii
I Ill
I II
[1 IIQI
00!
011 I
IIIII
IllIII
D 400

I SOLYM VARI All F
BY O .SOORT AGROTJS STOLONIFERA L.
LEO SOLDAAT
doctoraalverslag
plantenoecologie /populatiegenetica
februari — juni 1985
BEGELEI DING
plaritenoecologie:
Drs. C. Kik
populatiegenetica:
Drs. K. Wolff
Dr. R. Bijlsrna
RjksuniverSteit Groninc!Ofl
BibIothe3I( BIooIch ContrumKerklaan 30 — postbus 'I 4
9753 AA HAEN

Doktoraalverslag
Vakgroep Plantenoecologie R.U.G
Biologisch Centrum
Haren (Gn).
Doktoraalverslagen van de Vakgroep Plantenoecologie zijri internerapporten, dus geen offici1e pubflkaties.
De inhoud variert van een eenvoudige bespreking van onderzoeks-
resutaten tot een concluderende diskussie van gegevens in wijder
verband.
De konklusies, veelal sechts gesteund door kortlopend onderzoek,
zijn meestal van voorlopige aard en komen voor reken ing van de
auteur(s)
Overname en gebruik van gegevens slechts toegestaan na overleg met
auteur(s) en/of Vakgroepbestuur.

INHOUDblz.
1. SUMMARY 1
2. SAMEN VATTING 2
3. INLEIDING 3
3.1 milieuheterogeniteit en isozymvariatie 3
3.2 po1yp1odie 4
3.3 vraagstelling 5
4. POPULATIEBESCHRIJVING 6
5. MATERIAAL & METHODE 7
6. RESULTATEN 9
6.1 opbouw van de zymogrammen 9
6.2 isozymvariatie 12
6.3 temporele variatie in de ADH—expressie 16
6.4 po1yp1odie 16
7. DISCUSSIE 18
7.1 isozymvariatie 18
7.2 temporele variatie in do ADH—expressie 19
7.3 polyploTdie 20
8. LITERATUUR 25
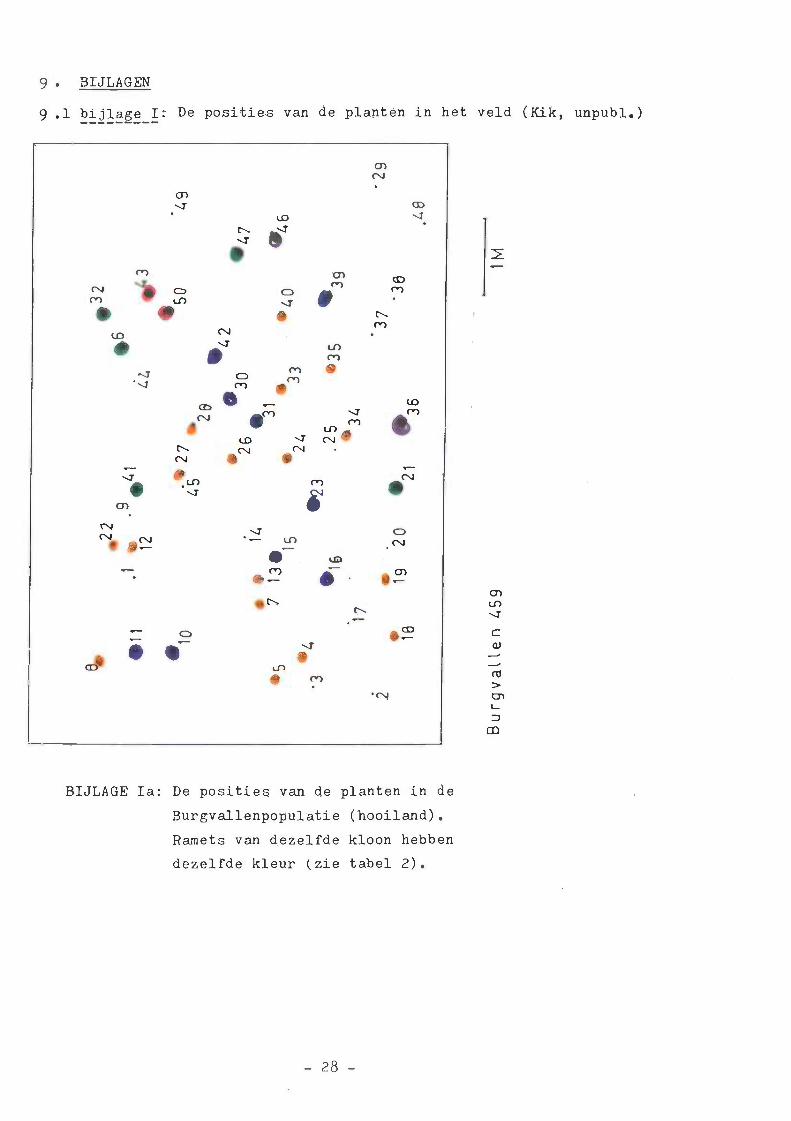
9. BIJLAGEN 28
9.1 bijiage I 28
9.2 bijiage II 32
9.3 bijiage III 34
9.4 bijiage IV 48

1. SUMMI\PY
Isozyme variation in four populations of' Agrostis stolonifera L.
in contrasting environments was determined. The enzyme systems
assayed were GPI, EST, PGM, 6-PGD, ADH, IDH.
The variation in isozyme phenotypes between the populations was
great. Only one of' 127 phenotypes occured in two populations, All
other phenotypes occured. in only one of the four populations.
No correlation was found between environmental factors and iso—
zyme variation, indicating that the enzyme—systems analysed
were selectively neutral. The polymorphy-indices in the popula-
tions were about equal.
In one population the amount of different phenotypes was much
lower than in the other three populations. This most presumable
was the result of clonal reproduction. The low level of' genetic
variation due to clonal reproduction may partly be compensated
for by genetic variation (in the form of' differences in plody—
level) within clones.
Also temporal isozyme variation was discovered, During the second
electrophoresis ADH showed other phenotypes than during the first
electrophoresis. The most likely explanation of this phenomenon
is change of ADI-I-expression caused by environmental changes in
the greenhouse the plants were grown in.
A mechanism bringing about isozyme variation within clones and
within individual plants is discussed.
-1—

2. SAMENVATTING
In vier, in hun milieu contrasterende Dopulaties van Agrostis
stolonifera L. werd de isozymvariatie voor zes enzymsystemen
(GPI, EST, PGM, 6—PGD, ADH, IDH) bepaald.
Het aantal verschillende isozymfenotypen in de vier populaties
bleek groot te zijn. Slechts n van de 127 gevonden fenotypen
kwam in twee populaties voor. Alle andere fenotypen kwamenhooguit in n van de populaties voor.Er is geen relatie gevonden tussen milieufactoren en isozymva—
riatie, wat aangeeft dat de onderzochte enzymsystemen selectief
neutraal zijn.
De polymorfie-indices waren in de vier populaties ongeveer
even groot.
Het aantal verschillende isozymfenotypen was in ên populatie
veel lager dan in de andere drie. De oorzaak hiervan is waar—
schijnlijk de sterkere kloonvorming in die populatie. Deze
geringe mate van genetische variatieals gevoig van kloonvorming
kan gedeeltelijk gecompenseerd worden door genetische variatie
(in de vorm van p1odie-verschil1eh) binnen kionen.
Ook temporele isozymvariatie kwam aan het licht. De ADH—feno—
typen bleken bij de tweede electroforese meestal anders te zijn
dan bij de eerste electroforese. De meest waarsciüjnlijke ver—
klaring hiervoor is verandering van de ADH-expressie onder in—
vloed van het milieu in de kas waarin de planten groeiden.
Een mechanisme waardoor ook isozymvariatie binnen kionen en
binnen individuele planten mogelijk is wordt besproken.
—2—

3. INLEIDING
3.1 milieuheterogeniteit en isozymvariatie
Aan het eind van de zestiger jaren kwam de enzymelectroforese in
gebruik in do populatiebiologie. Dit gebeurde nadat gebleken was
dat isozympatronen genetisch genterpreteerd konden worden (15,
19). Het bleek dat de genetische variatie in natuurlijke popula—
ties veel groter was dan men altijd gedacht had. Sindsdien is er
veel gediscussieerd over de vraag in hoeverre het milieu, via na—
tuurlijke selectie, invloed heeft op de genetische variatie in
een populatie.
Veel onderzoekers komen theoretisch tot de conclusie dat in niet—
stabiele milieus de genetische variatie hoger moet zijn dan in
stabiele milieus. Voldoende genetische variatie maakt het voor
een soort mogelijk om tolerant te zijn tegen milieuveranderingen
en om in versehillende milieus te kunnen overleven. Echt.er, in de
meeste onderzoeken met betrekking tot de relatie milieuheteroge—
niteit — genetische variatie kan deze relatie niet bewezen worden.
Er zijn vrijwel altijd alternatieve interpretaties van de onder—
zoeksresultaten mogelijk. De mate van genetische variatie in een
populatie kan bijvoorbeeld beinvloed zijn door het reproductieve
systeem van de soort, genetische drift, migratie en andere facto—
ren.
Do rol van natuurlijke selectie in het handhaven van enzympolymor—
fin en het belang van specifieke milieufactoren als selectieve
krachten, kan vastgesteld worden door de adaptieve betekenis van
individuele enzympolymorfin vast te stellen. Hierbij is het van
belang te weten dat allozymen en isozyrnen vaak verschillende func—
ties hebben, veroorzaakt door:
— verschillen in kinetisch gedrag: bindingaffiniteit substraat—
enzym, maximale reactiesneiheid, stabiliteit onder hoge tem—
peratuur, etc.
— verschillen in drie—dimensionale vormenHet potentieel voor functionele differentiatie is dus aanwezig(16). Het zoeken naar relaties tussen milieufactoren en individu—
ele enzympolymorfin is tot nu toe echter niet erg vruchtbaar ge—
weest. Metabolische processen in eon organisme worden doorgaans
ook niet door én enzym bestuurd, maar door een groot aantal ver—
schillende enzymen, elk verantwoordelijk voor eon deelreactie van

het proces. Daarom zijn multi—locus benaderingen van de relatie
milieuheterogeniteit — enzympolymorfie van belang. Een dergelijk
multi—locus onderzoek is bijvoorbeeld uitgevoerd door Allard et.
al. (2,3,8,14) aan Avena barbata.
Ret onderwerp van dit onderzoek is de interactie tussen het milieu
en de isozymvariatie bij Agrostis stolonifera L. Ret onderzoek
werd uitgevoerd in het kader van het onderzoek van Kik, waarin
gekeken wordt naar de interactie tussen het milieu en een set van
gecorreleerde levenscycluseigenschappen bij een aantal in hun mil-
ieu contrasterende populaties van Agrostis stolonifera L.
3.2 polyplodie
Eên van de factoren diede mate van genetische variatie kan bein—
vloeden is het plodie-niveau van een soort. PolyploTde planten
kunnen een groter aantal allelen voor een locus hebben dan diplo—
de planten. Hierdoor kunnen polyplode planten meer en, in het
geval van multimere enzymen, nieuwe isozymen vormen (1,11,12,23,
24,25,29). Ret grote aantal isozymen in polyplode planten maakt
de genetische interpretatie van electroforesepatronen vaak moei—
lijk. Veel onderzoek is dan ook verricht aan diplode soorten.
Onderzoek bij polyplode plantensoorten naar de relatie tussen
het milieu en iso-/allozymvariatie is door verschillende onder—
zoekers verricht:
Allard et. al. (2,3,8,14,17,21,22) bijvoorbeeld, vonden bij
Avena barbata een correlatie tussen de frequentie van bepaal—
de allozymfenotypen en temperatuur- en vochtigheidsparameters.
In koperverontreinigde milieus vonden Wu et. al. (31,32) p0—
pulaties van Agrostis stolonifera L. met een veel groter aan—
tal verschillende isozymfenotypen dan in populaties in niet-
verontreinigde milieus. Z±j concluderen dat sterke selectie-
voorwaarden (koperverontreiniging) zeker niet per sê leiden
tot reductie van het aantal genotypen in een populatie.
Al Mouemar en uasquez (4) vonden bij Chenopodium album L. het
tegenovergestelde. In een met herbicide behandeld veld (sterke
selectievoorwaarden) bleek de polymorfiegraad lager te zijn
dan in een niet behandeld veld. Zij durven echter niet te con—
cluderen dat dit het gevoig is van selectie.
— —

l3rown et. al. (7) tenslotte vonden een relatie tussen allo—
zymfrequenties en bodemvochtgehaite bij tromus mollis L.
Doze relatie werd gevonden voor drie loci, echter niet voor
het ADH—locus waarvan juist eon relatie met het bodemvocht—
gehalte verwacht word.
Agrostis stolonifera L. is een a11otetrap1od of amphidip1od
(5,6), dat wil zeggen een soort met twee, onafhankelijk van el—
kaar overervende, diplode genomen. Bj$rkman (6) vo.nd bij Agros—
tis stolonifera L. naast tetraploiden ook penta-/hexa— en aneu—
p1oden. Aston (5) vond alleen tetrap1oden en hexaploiden. De
hexaploTden vond hij vooral in natte, productierijke milieus.
Kik en Linders hebben in de vier populaties uit dit onderzoek
planten met de volgende ploTdie-niveau's gevonden (18):
tetraploden (2n = LfX = 28)
pentaploden (2n = 5x = 35)hexaploTden (2n = 6x = )2)aneusomen (planten met een per cel varirend aantal
chromosomen)
In dit onderzoek zal nagegaan worden of er een relatie bestaat
tussen doze plodie-niveau's en het aantal gevonden isozymbanden.
Hierbij wordt er vanuit gegaan dat de mate van isozymvariatie good
gecorreleerd is met de mate van genetische variatie op de onder—
zochte loci.
3.3 vraagstelling
Do vraagstelling bij dit onderzoek luidt als volgi:
1. Hoe groot is do mate van isozymvariatie tussen en binnen vier
populaties van Agrostis stolonifera L.
2. [s er een relatie tussen hot p1odie—niveau en het aantal
isozymbanden.
—5-

4. POPULATIEBESCHRIJVING
De vier populaties waarin de isozymvariatie bepaald is, en een
karakterisering van hun milieu staan in tabel 1.
POPULATIE MILIEUFACTOREN
BV NC ZC BP
BUPGVALLEN hooiland + + - +
NOOPDPOLDERZIJL kwelder ÷ + + +
LAUWEPSMEEP : polder ± ÷ ÷ ±
SCHIERMONNIKOOG : duin — - - -
BV = %bodemvocht NC = nutrintenconcentratieZC zoutconcentratie BP = biomassaproductie+ = relatief veel + intermediair — relatief weinig
TABEL 1. De vier onderzochte populaties van Agrostis stolonifeia
L. en hun milieu. (gegevens naar Kik, unpubi.)
De kwelder in Noordpolderzijl wordt voor een gedeelte van het
jaar begraasd.
Het hooiland in Burgvallen wordt ên keer per jaar, in het najaar,
gemaaid.
—6—

5. MATERIAAL & METHODE
In april 1983 heeft Kik zestig planten verzameld uit elke popu—
latie (zie bijiage I voor de posities van de planten in het veld).
Deze planten staan sindsdien in plastic potten met een rijk grond-
mengsel in een kas. De temperatuur in de kas is nauwelijks te re-
gelen, maar schommelt meestal rond de 20°C. Ongeveer Lf5 van deze
planten uit elke populatie zijn elk twee keer gelectroforeerd.
Van elke plant werd 300 mg. jong bladmateriaal gextraheerd engelectroforeerd op de wijze beschreven door van Dijk en van Del—
den (9).
De gebruikte buffers zijn:
pH 7.0 0,1 M citraat met tris op pH 7.0 gebracht
pH 8.65 (poulik) 0,76 M tris met citroenzuur op pH 8.65
gebrachtpoulik tankbuffer 0,3 N H3B03 met NaOH op pH 8.1 gebracht
(wordt onverdund in de tank gedaan)
De gebruikte gels zijn:
6 % polyacrylamide, buffer pH 7.0 (voor GPI,PGM,6PGD,IDH)
7,5 % polyacrylamide, buffer pH 7.0 (voor ADH)
7,5 % polyacrylamide, buffer pH 8.65 (voor EST)
Na de electroforese werden de gels gekleurd bij 37°C. De gebruikte
kleuringen staan in bijiage II.
CHPOMOSOOMTELLINGEN
Kik en Linders(18) hebben van de meeste gelectroforeerde planten
het p1odie-niveau bepaald.
FENOTYPISCHE POLYMORFIE & GENETISCHE AFSTANDEN
Deze beide gootheden zijn slechts te gebruiken voor onderlinge
vergelijking tussen de populaties. Vergelijking met andere onder—zoeken is niet mogelijk omdat in dit onderzoek de monomorfe sys—temen niet in de berekening zijn meegenomen.
Be fenotypische polymorfie is berekend op de manier zoals beschre—
yen is door Kahler et. al. (17):
—7—

(Pd) voor een enzymsysteem in een populatie
P p(1 - = 1
= frequentie van het i—de fenotype
n = aantal fenotypen per enzymsysteem per populatie
De gewogen gemiddelde fenotypische polymorfie (P) over alle
enzymsystemen is:
k(1/N.)p.
_j=i
(i/N.)j=i 3
N. = totaal aantal fenotypen per j—de enzymsysteem
voor k enzyrnsystemen
De genetische afstanden tussen de populaties zijn als volgtberekend:
= de kans dat twee willekeurige fenotypen voor
een enzymsysteem in populatie X identiek zijn
J idem voor populatie Yyy
= de kans dat twee fenotypen identiek zijn als
de eeri geoen wordt ult populatie X en de
ander uit populatie Y
I = de genora1iseerde identiteit voor het enzymsys—
teem:
J- xy
\/7T1V XX yy
D de genetische afstand tussen populatie X en Y
= _iogIDe genetische afstand oP basis van meerdere enzymsystemen wordt
berekend met behuip van het wiskundig gemiddelde van J enJ voor die systemen.Xy
—8—

6. RESULTATEN
6.1 opbouw van de zymogranimen
In deze paragraaf zullen de zymogrammen (bijiage III) van de
verschillende enzymsystemen besproken worden in het licht van
hun mogelijke genetische basis.
Om een betrouwbare genetische interpretatie van de zymogrammen
te kunnen geven moeten kruisingsexperimenten uitgevoerd worden.
Dit valt echter buiten het kader n dit onderzoek.
Ret aantal polypeptideketens waaruit een enzym is opgebouwd is
overgenomen van Ferguson (10), die deze aantallen geeft voor
menselijke enzymen.
GPI (glucose fosfaat isomerase, dimeer)
Waarschijnlijk gaat het hier om twee loci waarvan de zymogrammen
vlak bij elkaar liggen of elkaar overlappen.Het ene locus geeft altijd een bandje op 4.0 cm. van de originen meestal nog n of misschien meer bandjes ervoor.Ret andere locus beslaat bijna alle isozymbanden vôôr 4.0 cm.
Dit locus heeft waarschijnlijk een groot aantal allelen.
— in enkele gevallen heeft een plant slechts n allel
(bijv. L31, L4l)
- meestal zijn meerdere allelen aanwezig waardoor veel
heteromultimeren gevormd kunnen worden.
Plant S36 bijv. heeft drie allelen: S,I en F en zou
genotype SIIF kunnen hebben. Daardoor kunnen de volgende
dimeren gevormd worden:
dimeren : SS SI IT/SF IF FF
verhouding: 1 : 4 : 6 : 4 : 1
zyrnogram : I U
Plant N42 heeft twee allelen (S en F) en genotype SFFF,
zodat de volgende dimeren gevormd worden:
dimeren : SS SF FF
verhouding: 1 : 6 : 9
zymogram : 9 1De meeste zymogrammen op dit locus zijn echter moeilijk te
interpreteren.
_0_

EST (esterase , mono— en dimeren)
De opbouw van de esterase zyrnogrammen is erg onregelmatig.
Aangezien af en toe planten voorkomen met slechts n isozymband
(bijv. S20, S22, S23) zou er sprake kunnen zijn van &n locus
met een groot aantal allelen.
PGM (fosfoglucomutase,rnonomeer)
PGM gaf twee activiteitsz5nes te zien die PGM—I en PGM—II genoemd
zijn. PGM—I gaf banden te zien inhet gebied 3.2 — L-.1 cm. vanaf
de origin en PGM—II in het gebied L.Lf — 5.1 cm. vanaf de origin.
PGM—II gaf meestal erg onduidelijke banden.
De opbouw van de PGM—I zymogrammen lijkt vrij eenvoudig. Er icomen
vier allelen voor, namelijic S, I, F en P. Ret F allel komt bij
elke plant voor, dus n genoom is waarschijnlijk gefixeerd voor F.
Ret andere genoom variert De verschillende genotypen leveren
de volgende zymogrammen op:
GENOTYPE ZYMOGRAM
1 FFFF
2 IFFF to3 11FF EJO! SFFF Li
5 SSFF U U
6 PFFF [1 I
7 PPFF on8 IPFF Ill9 SIFF
10 SPFFI B I
De zymogrammen voor penta— en hexap1ofde planten zijn hieruit
gemakkelijk af te leiden.
Aangezien zymogrammen Lf, 5 en 10 niet voorkomen en zymograin 9
vrijwel niet is het S—allel waarschijnlijk erg zeldzaana. Ock
het P—allel komt slechts zelden voor (350, N3, Ni', S8).
6—PGD (6—fosfogluconaat dehydrogenase)
Waarschijnlijk gaat het om twee loci. Het linker locus bestaat
vaak uit drie bandjes met af en toe een bandje er tussenin
(bijv. L3k). Dit wijst op een dimere structuur van het enzym.
- 10 -

Andere zymogrammen lijken dit echter tegen te spreken. Plant
S13 en SlLf zouden narnelik niet het fenotype (fill 1) moetenI 13
hebben, maar (fJIlfl), dus ook een heteromultimeer tussen band
2 en 3.
Met rechter locus vertoont altijd een bandje op 3.9 cm. vanaf
de origin en vaak nog n of twee bandjes daarvoor. Blijkbaar
is ên genoom gefixeerd voor het allel dat bandje 3.9 veroor—zaakt.
ADH (alcohol dehydrogenase, dimeer)
De isozymbanden op 2.1 en 3.1 cm. zouden afkomstig kunnen zijn
van twee verschillende loci. Isozyinband 2.6 is altijd erg zwak
gekleurd en zou veroorzaakt kunnen zijn door post—translationele
modificatje. De beide veronderstelde locj vertonen meestal n
allel, waarvan de mate van expressie variert (zie paragraaf
6.3). Af en toe is er een tweede allel aanwezig (L/+, L5, L3Lf).
Dit tweede allel resulteert echter niet in drie isozymbanden,
zoals bij een dimeer enzym verwacht mag worden, behalve in plant
N25 waar dat juist wêl het geval is.
IDH (isocitraat dehydrogenase, dimeer)
De zymogranimen zouden geinterpreteerd kunnen worden als het
resultaat van twee allelen (S en F) op n locus, waarbij n
genoom gefixeerd is voor het F—allel.
ZYMOGRAN VERHOUDING GENOTYPE
TETRA- j 131] 1 : 6 : 9 SFFF
PLOTD 0 I -- 1 : 2 : 1 SSFF'
PENTA- j 131 1 : 8 : 16 SFFFF
PLOTDI 9 13 4 : 12 : 9 SSFFF
UUI 9:12: 4 SSSFF
Echter, het fenotype ( I 131) komt ook voor bij tetraplode planten.
Moeilijk te verklaren S ook het feit dat nooit homozygoot F—indi—
viduen gevond.en zijn. Alleen plant B20 was homozygoot F, maar dit
is geen Agrostis stolonifera L. rnaar Agrostis tenuis.
Er is aangenornen dat bij een niet al te sterke kleuringsintensi—
teit de dunste isozymbanden onzichtbaar blijven. Dit betekent dat
C I 131) en ( 19) dezelfde fenotypen zijn ( zie bijvoorbeeld de
duplo's van plant N39, bijiage III).
— 11 -

6.2 isozymvariatie
Voor het bepalen van de isozymvariatie zijn planten met identieke
isozymfenotypen geclusterd. Hiervoor is gebruik gemaakt van de
enzymsystemen GPI, EST, PGM-I en IDEI. PGM—II en 6-PGD werden niet
gebruikt omdat de zymogrammen voor deze enzymsystemen vaak ondul-
delijk waren. ADH werd niet gebruikt omdat er grote verschillen
waren tussen de zymogrammen van de eerste en de tweede electro—
foreseronde (paragraaf 6.3). De werkwijze voor het clusteren was
als volgt:
a. Alle planten werden ingedeeld aan de hand van het PGI"l—Ifenotype. Daarbij werden de volgende fenotypen onder-scheiden (zie ook paragraaf 6.1):
FENOTYPE
SIFR1
I
2
3 H14 IL5 II!
Er is geen rekening gehouden met verschillen in kleurings-
intensiteit tussen de isozymbanden, omdat deze verschillen
niet altijd duidelijk waren.
b. Vervolgens werden per PGM-I fenotype de planten gescheiden
op grond van de volgende 1DM fenotypen:
F ENOT YP E
1
2 jon3 001If
c. Binnen de nu ontstane clusters werden de planten onderlingvergeleken voor GPI en EST.
d. De planten waarvan de duplo's voor PGM—I en/of 1DM nietnduidig waren, werden vervolgens met alle andere planten
vergeleken voor GPI en EST.
De verdeling van de PGM—I en IDH fenotypen over alle onderzochte
planten staat in bijiage IV.
Na de bewerkingen a t/m d bleven een aantal clusters over waarvan
alle planten voor elk enzymsysteem hetzelfde fenotype bezaten.
- 12 —

Aangenomen is dat de planten uit n cluster ramets zijn van
dezelfde doon. 1)it is waarschijnlijk, omdat:
— de planten uit een cluster vaak viak bij elkaar staan
in het veld (bijlage I)
- de kans dat twee planten na sexuele voortplanting voor
alle enzyrnsystenien hetzelfde fenotype hebben erg klein
is, gezien het grote aantal rnogelijke fenotypen voor elk
enzymsysteem
De samenstelling van de kionen in de vier populaties staat in
tabel 2.
POPULATIE KLOON RAMETS
A B4 B5 37 B8 B12 B13 318 B19 B22BUPGVALLEN
B2Lf B26 B27 B28 B33 B31 B35 B1O(hooiland)
B BlO Bli B15 B16 B23 B30 B31 B39 BLf2
C B6 321 B32 BL1.l B47
D B36 B46
E BI+3 B50
A N39 NL4.L4 N47 N48 NLf9
B N5 N13N15NOORDPOLDEPZIJL
c N3 N4(kwelder)
D N30 N31
E N35 N36
F N18 N22
G N8 N9
H N27 N28
A L12 L13 L20 L39 L42 LLi7 L50LAUWEPSEEP
B L30 L36 L40(polder) L23 L26
D L19 L22
E L14 L38
SCHIEPMONNIKOOGA S7 S9 Sb
B S2 S3(duin)
C S40 SLf2
D SLfL S'45
TABEL 2. Samenstelling van de kionen in de vier populatiesDe plaats van de planten in het veld is te vinden inbijlage I.
- 13 —

Het enige isozymfenotype dat in twee populaties gevonden is,
is het fenotype van plant B38 en L21.
Als maat voor de mate van kloonvorming in de pooulaties is ge-
nomen: 100 — (% verschillende fenotypen van het totaal aantal
bepaalde fenotypen). De getallen staan in tabel 3.
De mate van kloonvorming is in de Burgvallen-populatie dus dui-
delijk sterker dan in de andere drie populaties. In de discussie
wordt hier verder op in gegaan.
De polymorfie-index van de vier populaties is berekend aan de
hand van de enzymsystemen GPI, PGM-I en IDH (tabel 4),op de ma—
nier zoals die beschreven is in het hoofdstuk Materiaal & Metho—
de (blz.7). De enzymsystemen PGM—II, 6—PGD en ADH zijn niet ge—
bruikt om eerder genoemde redenen (paragraaf 6.2). EST is niet
gebruikt om de volgende twee redenen:
— Na de eerste electroforeseronde bleken er veel verschil—
lende isozymbanden te zijn voor GPI en EST. Hierdoor Wa—
ren de fenotypen van planten die op verschillende gels
geelectroforeerd waren, moeilijk te vergelijken. Bij de
tweede electroforeseronde is er daarom voor gekozen om
planten die bij de eerste ronde ongeveer hetzelfde GPI—
fenotype hadden, op ên gel te electroforeren. Zodoende
waren deze planten voor GPI goed met elkaar te vergelijken.
VoOr EST kon dit niet gebeuren omdat dat teveel tijd zou
kosten, zodat onderlinge vergelijking voor EST minder be—
trouwbaar is.
— Nadat in de kas waarin de planten groeiden met een insec-
ticide gespoten was, was elke EST—activiteit in de planten
— 14 —
POPULATIE AANTAL GEELEC— AANTAL VERSCFJIL-TPOFOREERDE LENDEPLANTEN FENOTYPEN
BUPGVALLEN 44 14 (31,8%)
NOORDPOLDEPZIJL 47 35 (74,5%)LAUWERSMEER 46 35 (76,1%)SCHIERMONNIKOOG 47 42 (89,4%)
% KLOON-VORMING
68,225,523,9
10,6
TABEL 3. De mate van kloonvorming in de vier populaties

verdwenen. Hierdoor konden in de eerste electroforeseronde
in de Lauwersmeerpopulatie geen EST—fenotypen bepaald wor—
den, en in de tweede ronde konden in geen enkele populatie
EST—fenotypen bepaald worden.
POPULATIE POLYMORFIE-INDEX (P.) (gewogen
GPI PGM—I IDH gemiddelde)
BURGVALLEN 0,787 0,504 0,455 0,499
NOORDPOLDEPZIJL 0,950 0,462 0,593 0,542
LAUWERSMEER 0,910 0,484 0,649 0,568
SCHIEPMONNIKOOG 0,96b 0,503 0,583 0,556
TABEL 4. De polymorfie-indices in de vier populaties.
Uit tabel 4 blijkt dat, ondanks de sterke kloonvorming in de
Eurgva11enpopu1atie (tabel 3), de fenotypische polymorfie in deze
populatie nauwelijks lager is dan in de andere drie populaties.
De genetische afstanden tussen de populaties voor GPI, PGM—I en
IDH en voor deze drie enzymsystemen samen staan in tabel 5.
POPULATIES GENETISCHE AFSTAND (D) D gemiddeldGPI PGM-! IDH
B — N -400 0,094 2,112 0,750
B — L —OQ 0,191 1,737 0,794
B — S —)oc, o,487 1,485 0,968
N — L —400 0,029 0,091 0,129
N — s —>oo 0,187 0,034 0,160
L — S -4o0 0,066 0,230 0,204
TABEL 5. De genetische afstanden tussen de populaties.
t3=Burgvallen N=Noordpolderzi ji L=Lauwersmeer
S=Schiermonnikoog. —>00 = gaat naar oneindig.
De genetische afstanden voor de drie enzymsystemen verschillen
nogal. Voor GPI gaan de afstanden naar oneindig. Dit komt door
de grote fenotypische polymorfie van UPI (tabel 4).
— 15 —

6.3 temporele variatie in de ADII—expressie
Veel onderzochte planten gaven bij de tweede electroforeseronde
andere ADH—fenotypen te zien dan bij de eerste electroforeseron—
de (bijiage III). Bijvoorbeeld de ramets van de grootste kloon
in de Burgvallenpopulatie (kloon A, tabel 2) gaven bij de eerste
electroforeseronde zonder uitzondering het fenotype ( J J ) te
zien. Bij de tweede ronde bleken de ADH—fenotypen tussen de ra—
mets behoorlijk te verschillen. In het algemeen was er bij de
tweede ronde meer activiteit op het rechter bandje dan bij de
eerste ronde (tabel 6).
POPULATIE VEPSCHUIVINGSRICHTING—R L— GEEN
BURGVALLEN 86 2 12
NOOPDPOLDERZIJL 8 L. 28
LAUWERSMEEP 57 0
SCHIERMONNIKOOG 27 11 61
TABEL 6. % van de gevallen waarin de ADH-acti--
viteit bij de tweede electroforese—
ronde naar rechts (— H), links (L —)
of niet (GEEN) verschoven is t.o.v. de
eerste ronde.
6.2+ po1yplodie
Van de zes onderzochte enzymsystemen zijn alleen GPI, EST en
PGM—I gebruikt voor hot bepalen van het aantal isozymbanden per
p1odieniveau (tabel 7). De overige enzymsystemen gaven te on—
betrouwbare resultaten (PGM-II, 6—PGD) of gaven een waarschijn-.
lijk niet variérend aantal isozymbanden (ADH, IDH). Vanwege het
kleine aantal penta— en hexaploTden (resp. 9 en 1) zijn deze
samen met de aneusornen als ên p1odieniveau ( >28) vergeleken
met de tetraploden (23).
De hypothese: er bestaat tussen do twee ploIdieniveau's geen
verschil in het aantal isozymbanden per plant
werd getoetst met de Mann—Whitney—U test. De resultaten staan
in tabel 7.
— 16 —

EN ZY M-
SYSTEEM
(x)
2
GPI
gem. aantal banden
p
AANTAL PLANTEN MET
13'VALLEN N'ZIJL
28 >28 23 >28
2
1/+
3
1
x ISOZYMBANDEN
L'MEER S'OOG
28 >28 28 >28
1
1
1 5 4
8 3 8 2
3 1 4 1
5 3 3
1 2
5,8 5,20,093
Aantal isozymbanden per plant voor twee verschillende
plodieniveau's in de vier populaties.
28 = tetrap1oden>23 = pentaplolden, hexaploden en aneusomen
P = kans dat het verschil tussen de gemiddelden
toeval is.
Bij i-=0,05 (5%) bestaat voor geen enkel enzymsysteem een verschil
in het gemiddeld aantal isozymbanden per plant tussen de twee plo—
dieniveau' s.
— 17 —
AANTAL ISO-
ZYMB A NJ) EN
2
31
2
1
7 5,70,090
(x) 28 >28
1
5,3 5,30,443
28 >28
EST 3
gem. aantal banden
P
5,2 4,6
0,077
28 >28
5 2
6 6
10 3
1
2,4 2,1
0,223
2 14
3 16
1
3,6 3,60,299
28 >28
5
11 2
1
1,8 2,0
0,298
(x) 28 >28 23 >28 28 >28 28 >28
PGn-I
gem. aantal banden
p
TABEL 7.
2 25
3 7
2,6 2,2
0,088
6 2
15 4
2
1,7 2,0
0,190
11 4 17 1
10 11 14 2
1,5 1,7 1,5 1,7
0,097 0,272

7. I)LSCUSSIE
7.1 isozyrnvariatie
Net feit dat elke populatie een unieke set isozymfenotypen heeft
(op het ene fenotype na dat Nurgvallen en Lauwersmeer gemeenschap—
pelijk hebben) wordt voornamelijk veroorzaakt door de isozymvaria—
tie voor GPI en EST. De variatie voor deze beide enzymsystemen was
veel groter dan voor de andere systemen (bijiage 111). Vandaar dat
de genetische afstanden voor (iPI veel groter zijn dan voor PGM—I
en IDH (tabel 5).
Er lijkt geen verband te bestaan tussen de gevonden isozymfenoty—
pen en het milieu. Een aanwijzing voor een dergelijk verband kan
pas gevonden worden als in een bepaald milieu n of enkele feno—
typen of isozymbanden domineren, zoals bijvoorbeeld werd gevonden
bij Avena barbata in Californi (3, 8, 21, 22). Bij deze haver—
soort vonden Allard et. al. n genotype dat meestal overheerste
in droge, warme gebieden. Ook binnen ên gebied met drogere en
nattere habitats bleek dit genotype vooral in de drogere habitats
voor te komen. Hoewel niet duidelijk was of natuurlijke selectie
aangreep op de onderzochte enzymsystemen zeif, of op nauw daaraan
gekoppelde systemen, bestaat in dit Avena—onderzoek zeker een ver—
band tussen het milieu en het isozymfenotype (in dit geval ook
het genotype).
Voor een dergelijk verband zijn in dit onderzoek geen aanwijzingen
gevonden. De onderzochte enz.ymsystemen zijn waarschijnlijk selec—
tief neutraal.
De kloonvorming in de Burgvallenpopulatie is duidelijk groter dan
in de drie andere populaties (tabel 3). De verspreiding van deze
klonen vindt plaats via de stolonen van de planten. In de kwelder,
waar korte stolonen gevormd worden, staan de ramets dichter bij
elkaar dan in het hooiland, waar langere stolonen gevormd worden
(bijlage I).
De sterke kloonvorming in Burgvallen (hooiland) geeft aan dat de
voortplanting daar voornamelijk vegetatief is. Vegetatieve voort—
planting heeft in het hooiland waarschijnhijk een voordeel ten op—
zichte van sexuele voortplanting. Door de hoge vegetatie kunnen
kleine kiemplanten immers gemakkelijk gebrek aan licht, water en
nutrinten krijgen en doodgaan. Jonge ramets echter zijn minder
— 18 —

afhankelijk van hun directe abiotische omgeving dan kiemplanten,
omdat zij nog via een stolon voedlngsstoffen van do adulte plant
kunnen krljgen (20).
Door de sterke kloonvorming in het hooiland is het aantal isozym—
fenotypen per opervlakteeenheid laag. Zo'n relatie tussen het
reproductieve systeem en de fenotypefrequentie werd bijvoorbeeld
ook gevonden door Uray et. al. (13) bij Puccinellia maritima. In
begraasde kwelders vonden zij sterkere kloonvorming dan in onbe—
graasde kwelders, waarschijnlijk omdat in begraasde kwelders mm-
der ruimte beschikbaar was voor kiemplanten. Het reproductieve
systeem bleek van invloed te zijn op de fenotypefrequenties. In
de, onbegraasde kwelders (sexuele reproductie) kwamen ongeveer
twee keer zoveel isozymfenotypen per oppervlakte—eenheid voor danop de begraasde kwelders (vegetatieve reproductie).Hoewel het aantal isozymfenotypen per oppervlakte—eenheid relatief
laag is in hot hooiland, is de fenotypische polymorfie (P, tabel Lf)
slechts weinig lager dan in de andere drie milieus. De oorzaak
hiervan is dat de polymorfie-index een gewogen gemiddelde is van de
polymorfie—indices van de verschillende enzymsystemen. hierdoor
tellen minder variabele systemen als i-'GM—I en IDH zwaarder mee dan
zeer variabele systemen als UP!. Dat do fenotypische polymorfie inhet hooiland, ondanks de sterkere kloonvorming, maar lets lager is
dan in de andere milieus, geeft aan dat de isozymvariatie tussen
de kionen onderling en de overige planten groot is.
7.2 temporele variatie in de ADi-i—expressie
In tabel 6 was te zien dat de ADH—activiteit tijdens de tweede elec—
troforeseronde naar rechts verschoven is ten opzichte van de eerste
ronde, Het enige verschil tussen de beide ronden is de periode
waarin deze plaats vonden. De eerste ronde duurde van 22/3 — 2LF/4,
de tweede ronde van 21/5 — 21/6. Tijdens de eerste ronde was de ge—
middelde temperatuur in de kas 20,3°C en tijdens de tweede ronde
23,6°C. Deze temperaturen zijn de gemiddelden over alle dagen,00 00bij per dag vier keer de temperatuur bepaald is (om 6. , 12.
18.00 en 24.°°h.). De standaarddeviatie was tijdens de eerste
ronde 2,0 en tijdens de tweede rondo 5,3. Naast een hogere gemid—
delde temperatuur was de variatie in de temperatuur tijdens de
tweede rondo dus ook hoger. Af en toe kwam do temperatuur boven de
40°C. Hierdoor was hot bodemvochtgehalte in de potten tijdens de
- 19 -

tweede ronde waarschijnlijlc lager dan tijdens de eerste ronde.
De planten zeif bevatten in ieder geval duidelijk minder water,
hetgeen bleek bij de bereiding van de extracten: na centrifugatie
bleef minder supernatant over dan tijdens de eerste ronde het ge—
val was.
Deze gegevens doen verrnoeden dat er een verband bestaat tussen
het isozymfenotype en het milieu, zoals dat bij Bromus mollis het
geval is (7). Het byzondere hier is echter dat er sprake is van
temporele variatie in de ADH—expressie binnen individuele plan—
ten.
Het hier beschreven fenomeen lijkt veel op wat Puvinsky et. al.
(26) alternation genoemd hebben:
PtAlternation is a nonmutational inheritable change
in the state (active inactive) of single or
multiple genes. "
Zij vonden binnen kionen van de watervlo Daphnia pulex variatie
in de expressie van G6PD. Deze variatie trad spontaan op n was
te induceren met behuip van glucose (27). De mechanismen waarmee
deze activatie — inactivatie bewerkstelligd wordt zijn nog niet
duidelijk.
Als er werkelijk een relatie bestaat tussen het bodemvochtgehalte
en de ADH—expressie is dat van adaptieve betekenis. Een plant is
dan minder kwetsbaar door schommelingen in het bodemvochtgehalte.
Misschien is dit verschijnsel verantwoordelijk voor de grote fe—
notypische flexibiliteit die Aston (5) vond in sommige populaties
van Agrostis stolonifera L. Deze populaties bleken zowel onder
natte als onder droge omstandigheden goed te overleven.Zie ook 3.
7. polyplodieEen toename van het aantal isozymbanden voor een enzymsysteem bij
een hoger ploTdie—niveau is in geen van de drie populaties gecon—
stateerd (tabel 7). Mogelijk is dit te wijten aan het kleine aan—
tal geteste planten en het relatief kleine verschil in plodie—
niveau tussen de twee testgroepen. In onderzoeken waarin wel een
groter aantal isozymbanden bij een hoger p1odie-niveau gevonden
werd ging het meestal om diplode en tetraploTde planten (12, 23,
25, 28, 29). Roose en Gottlieb (25) bijvoorbeeld hebben onderzoek
gedaan aan drie diploTde Tragopagon soorten (T. dubius, T. porn—
f'olius, T. pratensis) en hun tetraploTde hybriden (T. mirus en
- 20 -

P. miscellus). De diploden waren voor veel loci homozygoot. De
tetraploTden echter waren voor dezelfe loci vaak heterozygoot,
waarbij ze de isozymbanden van de beide dip1ode voorouders te
zien gaven en, in het geval van multimere enzymen, sozns ook
nieuwe isozymbanden.
Interessant is het voorkomen van meerdere p1odie-niveau's binnen
ê6n k],oon. Kloon A in de Burgvallenpopulatie bijvoorbeeld (de
eerste 17 planten in bijiage III) bevat twee tetrap1ode planten,
vier pentaploTden en tien aneusomen, De p1odie—niveau's van de
planten (overgenomen vai Kik & Linders,18) staan achteraan bijiage
I. Om te begrijpen hoe deze p1odieverschi11en kunnen ontstaan
moeten we eerst weten hoe de penta— en hexap1oden en de aneusomen
gevormd worden. Dit zou als volgt voorgesteld kunnen worden:
Tijdens de meiose worden niet alleen normaal geredu—
ceerde gameten gevormd, maar in sommige gevallen ook
unilateraal gereduceerde en ongereduceerde gameten.
Na samensmelting met andere gameten kunnen zo penta—
en hexap1ode planten ontstaan. Deze planten zijn
functioneel steriel (5, 6). In de loop van de tijd
vallen deze planten terug naar het tetraplo!de (en
fertiele) niveau. Tijdens dit proces kan een vari'é—
rend aantal chromosomen in de plantencellen aanwezig
zijn; de planten zijn dan aneusoom.
Tussen de ramets van n kloon kan hierdoor variatie in het aantal
chromosomen ontstaan, en ook binnen een aneusorne rainet bestaat Va—
riatie in p1odie—niveau tussen de plantecellen.
Theoretisch kunnen er tijdens het terugvallen naar het tetrap1o—
de niveau twee dingen gebeuren, die verschillende gevolgen hebben
voor de plant. Dit wordt duidelijk in het volgende voorbeeld:
VOORBEELD
De al1otetrap1ode plant met voor een bepaald locus
het genotype A1A2B1B1 (A,B = chromosoom van genoom
A, resp. 3; 1,2 = allel 1,2) zal bij een normale mel—
ose twee typen garneten vormen, namelijk A1B1 en A2B1.
Bij samensmelting met bijv. gameet A3B3 ontstaan twee
typennakornelingeri: A1AB1B en A2A3B1B3.
Wanneer echter plant A1A2B1B1 ook unilateraal ongere—
— 21 —

duceerde gameten vormt, ontstaan de volgende gameten:
A1A2B1, A1B1B1 en A2131B1. Na samensmeiting met gameet
A3B3 worden de nakomelingen A1A2AB1B3, A1A3B1B1B3 en
A2A331B133 gevormd. Deze pentapl&fde planten vallen in
de loop van de tijd terug naar het tetrapiode niveau.
De twee manieren waarop dat in z'n werk kan gaan zijn:
1.in aile planteieilen verdwijnt hetzelfde chromo—
soom. De pentap1oden vorrnen dan uiteindeiijk de
volgende tetrapio!de pianten:
A1A2A3B1B3 vormt A1A2B1B3 Of A1A331B3 Of A2A3B1B3
A1A3B1B1B3 vormt A1A331B3 Of AA3BB1
A2A3B1B1B3 vormt A2A3B1B3 Of A2A3B1B1
Plant A1A2B1B1 heeft flu dus vijftypennakornelingen.
2.in eike plantecel kan een ander chromosoom verdwij—
neri. Bijvoorbeeld uit pentapiod A1A2A3B133 ont—
staat een tetrapiod met drie typen celien (zie 1.)
oftewel een genetisch mozalek. In elke plant zal de
verhouding tussen deze typen celien anders zijn, zo—
dat een haast oneindig aantal typen nakomelingen
ontstaat.EINDE VOORBEELD
In de literatuur is over een dergeiijk proces niets te vinden.
Het voorkomen van genetische mozaTeken zou een verkiaring kunnen
zijn voor de kleine verschillen in het isozymfenotype van planten,
die regeimatig tussin de twee electroforeseronden gevonden werden
(zie bijv. het GPI—fenotype van Li, L2, L3, Lik, L19, LLi-l en vele
andere). Tijdens de eerste electroforeseronde is immers een andere
populatie van pianteceilen geêlectroforeerd dan tijdens de tweederonde.Of penta— en hexapialde pianten inderdaad terugvailen naar het
tetraplalde niveau en op weike manier dat gaat zal onderzocht moe—ten worden. Wat zon onderzoek ook op zal leveren, n ding staat
al vast, namelijk dat er genetische variatie (in de vorm van plo—
die—verschillen) mogelijk is binnen n kioon en zelfs binnen n
plant. De pianten die dit soort genetische variatie kunnen hebben
(pianten met een verhoogd pioidie—niveau) komen voornamelijk voor
in de Burgvalienpopulatie, zoals te zien is in tabel 8.
- 22 —

Hiervoor zijn verschillende verklaringen mogelijk:
1. Het milieu in de Burgvallen veroorzaakt een sterke
kloonvorming waardoor het aantal genotypen per opper—
vlakte—eenheid verlaagd wordt (blz. 19). Via de vorming
van planten met een verhoogd p1odie-niveau kan de po—
pulatie deze verlaging van de genetische variatie jets
compenseren.
2. Het vochtige milieu in de Burgvallen is op een of andere
manier verantwoordelijk voor het voorkomen van veel plan-
ten met een verhoogd ploIdie-niveau. Dit is niet zomaar
een hypothesem maar is gebaseerd op waarnemingen van
Aston (5) die bij Agrostis stolonifera L. vond dat hexa—
ploden vooral in vochtige milieus voorkwamen. Aangezien
de penta— en hexaploTden functioneel steriel zijn (5, 6)
planten zij zich vegetatief voort. Vandaar de sterke
kloonvorming in de Burgvallenpopulatie.
Deze twee verklaringen in een schema:
— 23 —
POPULATI E MILIEU % TETRAPLOIDEN
BURGVALLEI' hooiland
NOORDPOLDERZIJL kwelder
LAUWERSf4EER polder
SCHIERMONNIKOOG duin
% PENTAPLOIDENHEXAPLOIDENA NEU SON EN
19
70
63
90
81
30
37
10
TABEL 8. % tetraploden en % (penta/hexap1oden en aneusomen)
in de vier populaties.

Behalve voor de populatie kan dit soort genetische variatie ook
betekenis hebben voor individuele planten. Als een plant een
genetisch mozaek is, kan hij daardoor een grotere kans hebben
om milieuveranderingen te overleven. Whitham (30) vond bijvoor—
beeld bij Populus angustifolia dat planten met een (fenotypische)
mozaek—structuur voor bladoppervlakte een verhoogde resistentie
bezaten tegen eon parasiet.
Tot flu toe is weinig bekend over de in deze paragraaf genoemde
vormen van gnetische variatie.
— 24 -

8• LITERATUUP
1. Adams, W.T. & P.11. Allard (1977). Effect of polyploidy onphosphoglucose isomerase diversity in Festuca micro—stachys. Proc. Nati. Acad. Sci. USA. vol. 74(4), pp.1652—1656.
2. Allard, H.W., G.IR. Babbel, M.T. Clegg & A.L. Kahler (1972).Evidence for coadaptation in Avena barbata. Proc. Nati.Acad. Sci. USA. vol. 69, pp. 3043—3048.
3. Allard, R.W., P.D. Miller & A.L. Kahler (1978). The rela-tionship between degree of environmental heterogeneityand genetic polymorphism. In: Structure and functioningof plant populations. Freysen, A.H.J. & J.W. Woldendorp.(eds.). New York: North Holland Pubi. pp. 49—73.
4. Al mouemar, A. & J. Gasquez (1983). Environmental conditionsand isozyme polymorphism in Chenopodium album L. WeedPesearch. vol. 23, pp. 141—149.
5. Aston, J.L. (1962). Evolutionary divergence in the Gramineae,with special reference to Agrostis stolonifera L.
6. Bjrkman, S.0. (1954). Chromosome studies in Agrostis. II.Hereditas (Lund). vol. 40, pp. 254—258.
7. Brown, A.H.D., D.P. Marshall & L. Albrecht (1974). The main-tenance of alcohol dehydrogenase polymorphism in Bromusmollis L. Aust. J. Biol. Sci. vol. 27, pp. 545— 559.
8. Clegg, M.T. & P.W. Allard (1972). Patterns of genetic dif-ferentiation in the slender wild oat species Avena bar—bata. Proc. Nati. Acad. Sci. USA. vol. 69, pp. 1820-1824.
9. DIjk, H. van & W. van Delden (1981). Genetic variability inPlantago species in relation to their ecology. I. geneticanalysis of the allozyme variation in P. major subspecies.Theor. Appl. Genet. vol. 60, pp. 285—290.
10. Ferguson, A. (1980). Biochemical systematics and evolution.Blackie. Glasgow and London.
11. Gottlieb, L.D. (1973). Genetic control of glutamate oxaloacetatetransaminase isozymes in the dip1od plant Stephanomeriaexigua and its a1lotetraplod derivative. Bioch. Gen. vol.9, pp. 97—107.
12. Grant, J.E., A.H.D. Brown & J.P. Grace (1984). Cytologicaland isozyme diversity in Glycine tomentella Hayata (Legumi-nosae). Aust. J. Bot. vol. 32, pp. 665—677.
13. Gray, A.J.,, H.J. Pai-sell & R. Scott (19 ). The genetic struc-ture of plant populations in relation to the developmentof salt marshes. In: Ecological processes in coastal envi-ronments. Jefferies, P.L. & A.J. Davy (eds.), pp.43—63.
14. Hamrick, J.L. & P.11. Allard (1975). Correlations between quan-titative characters and enzyme genotypes in Avena barbata.Evolution. vol. 29, pp. 438—442.
15. Harris, H. (1966). Enzyme polymorphisms in man. Proc. Roy. Soc.Ser. B. vol. 164, pp. 298—310.
- 25 —

16. Johnson, j.B. (1976). Genetic polymorphism and enzyme function.In: Molecular evolution. Ayala, i".J. (ed.). Sinauer Asso-ciates, Sunderland, Massachusetts. pp. 46—59.
17. Kahier, A.L., P.W. Allard, M. Krzakowa, C.F. Wehrhahn & E. Nevo(1980). Associations between isozyme phenotypes and environ-ment in the slender wild oat (Avena barbata) in Isral.Theor. Appi. Genet. vol. 56, pp. 31—1+7.
18. Kik, C. & T.E. Linders (unpubl.). On the occurence of aneusomatyin the species Agrostis stolonifera L. and its possibleecological significance.
19. Lewontin, P.C. & J.L. Hubby (1966). A molecular approach to thestudy of genic heterozygosity in natural populations. II.Amount of variation and degree of heterozygosity in naturalpopulations of Drosophila pseudoobscura. Genetics. vol. 54,pp. 595—609.
20. Lovett Doust, L. (1981). rntraclonal variation and competitionin Panunculus repens. New Phytol. vol. 89, pp. 495—502.
21. Marshall, D.P. & P.W. Allard (1970 a). Maintenance of isozymepolymorphisms in natural populations of Avena barbata.Genetics. vol. 66, pp. 393—399.
22. Marshall, D.P. & P. W. Allard (1970 b). Isozyme polymorphismsin natural populations of Avena fatua and A. barbata.Heredity. vol. 25, pp. 373—382.
23. Mitra, P. & C.R. Bhatia (1971). Isozymes and polyplody. I.Qualitative and quantitative isozyme studies in the Triti—cinae. Genet. Pes. Camb. vol. 18, pp. 57—69.
24. Reddy, M.M. & E.D. Garber (1971). Genetic studies of variantenzymes. III. Comparitive electrophoretic studies of estera—ses and peroxidases for species, hybrids and amphidiplodsin the genus Nicotiana. Bot. Gaz. vol. 132(2), pp. 158-166.
25. iRoose, M.L. & L.D. Gottlieb (1976). Genetic and biochemicalconsequences of polyploTdy in Tragopagon. Evolution. vol.30, pp. 818-830.
26. Ruvinsky, A.O., Yu.I. Lobkov & D.K. Belyaev (1983). Spontaneousand induced activation of genes affecting the phenotypicexpression of glucose-6phosphate dehydrogenase in Daphniapulex. I. Intraclonal variations in the electrophoreticmobility of G6PD. Mol. Gen. Genet. vol. 189, pp. 485—489.
27. Puvinsky, A.0., Yu.I. Lobkov & D.K. 3elyaev (1983). Spontaneousand induced activation of genes affecting the phenotypicexpression of glucose-6-phosphate dehydrogenase in Daphniapulex. II. Glucose—induced changes in the electrophoreticmobility of G6PD. Mol. Gen. Genet. vol. 189, pp. 490—494.
28. Sheen, S.J. (1972). Isozyme evidence bearing on the origing ofNicotiana tabacum L. Evolution. vol. 26, pp. 143—154.
29. Smith, H.H., D.E. Hamill, E.A. Weaver & K.H. Thompson (1970).Multiple molecular forms of peroxidases and esterases amongKicotiana species and amphiploTds. J. Heridity. vol. 61,pp. 203-212.
- 26 —

30. Whitham, i.G. (19 ). Host manipulation of parasites: within—plant variation as a defense against rapidly evolvingpests. In:
31. Wu, L., A.D. Bradshaw & D.A. Thurman (1975). The potential forevolution of heavy metal tolerance in plants. III. Therapid evolution of copper tolerance in Agrostis stolonifera.Heredity. vol. 3Lf, pp. 165—87.
32. Wu, L. (1976). Esterase isoenzymes in populations of Agrostisstolonifera L. Bot. Bull. Academia Sinica. vol. 17, pp.175_l8Lf.
33. kot, . C., 3. STiLcovc G. Rcz A V. stac.iVacLiOfr, yd.WLv xooiC c 0svv cud-' csi 4-ov a c2-oc £I: -obc uitov&
c.o. ti-i. CrQw'orc (edc.. Arri Arbor
— 27 —
9 • BIJLAGEN
9 .1 bijiagel: De posities van de planten in het veld (Kik, unpubi.)
0')
c—i
-4t-. —r
0)(N IC') Li)•
(N(0
• •C,-)
•C'(N C')Li)
w(N S
(-'4CJ.—
— (y)
0)N.
•Li)I >
0)I-
CD
BIJLAGE Ta: De posities van de planten in de
Burgvallenpopulatie (hooiland).Ramets van dezelfde kloon hebben
dezelfde kleur (zie tabel 2).
— 28 —

'C",
S (N (N(N
('Si
(N
— 29 —
IIJLA(.iE Ib:
De posities vai
de planten in
de Noordpolder—
zijipopulatie
(kwelder).
Pamets van de—
zelfde kloon
hebben dezeif—
de kleur (zie
tabel 2).
(N (-v)If)
U)U)
"Jo
U)It)
(0 ON-
(N-4
1p
'C')U)
0)'(V.)
(D$(V'-4
(V.)(V.)
cD0) (0
(N
It)(N
.4(N
(N CDI—0')
(N
U)
CD0)
(0
NJL.
-o
00'000z

[cD - - -T.U)
I*UD 14)
cD c3)•
NN iD U)
() (_)
(Nr) m () (N (N
'-U(N
(N Q-)(N -,
0• iJ)3(N 0) a)
• .CD
C..-. (0 a)
LI)
_______
-J
BIJLAGE Ic: De posities van de planten in de
Lauwersmeerpopulatie (polder).
Rainets van dezelfde kloon hebbendezelfde kleur (zie tabel 2).
— 30 —

(j-n3-
3o3 —
0 — —o 10
NJ. tOC)
0)NJ
C-,,
NJ C-fl(0 - £- CD
(i).(J1 0'
(Aj C-flNJ 0) 0)
(0NJ
(A) .L)01 0)
0)
(A)
Li
(0
BIJLAGE Id: De posities van de planten in de
Schiermonnikoogpopulatie (duin).
Pamets van dezelfde kloon hebben
dezelfde kleur (zie tabel 2).
— 31 —

9.2 bijiageIl: Overzicht van de gebruikte kleuringen.
(WI (glucose fosfaatdehydrogenase)
30 ml Tris—HC1—MgC12 (0,1M), met tris op
pH 7,5 gebracht+5mg NADP10 mm, in:
50 mg Fructose—6—fosfaat
6 mg MTT
0,03 ml Glucose—6-fosfaat dehydrogenase
na 10 mm.toegevoegd: 1 mg PMS
EST (esterase)
30 mm. in fosfaatbuffer (0,1M), pH 6,5
(30 ml Fosfaatbuffer (0,1M), pH 6,5na 30 mm. in:
20 mg Fast Red
5 mg — naftylacetaatI ç opgelost in 0,5 mlL 5 mg (3 — naftylacetaat S aceton
PGM (fosfoglucomutase)
(30 ml Tris—HC1—MgC12 (0,1M), pH 7,5
5 mg NADP10 mm. in: 60 mg Glucose—1-fosfaat
6mg MTT
L 0,03 ml Glucose—6—fosfaat dehydrogenase
na 10 mm.toegevoegd: 1 mg PMS
6—PGD (6—Fosfogluconaat dehydrogenase)
(30 ml Tris—HC1—MgC12 (0,1M), pH 7,5
5 mg NADP30 mm. in:
6 mg 6—Fosfogluconaat
L 6 mg
na 30 mm.toegevoegd: 1 mg PMS
ADH (alcohol dehydrogenase)
(30 ml Tris-HC1 (0,05M), p1-i 8,5
30 mm. in: 6 mg NAD1 ml Ethanol
6mg MTT
— 32 —

na 30 mm.toegevoegd: 1 tug PMS
IDH (isocitraat dehydrogenase)
(30 ml Tris—HC1-MgC12 (o,1M), pH 7,5
I 5mg NADP30 nun, in:40 mg Na-isocitraat
6 tug MTT
na 30 mm,toegevoegd: 1 mg PMS
- 33 -

9.3 bijiage Ill: Isozymfenotypen en ploTdieniveau's
isozymfenotypen
Elke plant is twee keer g&électroforeerd. De eerste keer (in de
periode 22/3 — 21-f/4) werd het bovenste fenotype gescoord, de
tweede keer (in de periode 21/5 — 21/6) het onderste fenotype
dat achter elke plant staat.
De betekenis van de tekens is als volgt:
aanwezigheid van de isozymband is niet zeker
I isozymbarid met lage kleuringsintensiteit
0 isozymband. met matige kleuringsintensiteit
I isozymband met sterke kleuringsintensiteit
U brede isozymband met matige kleuringsintensiteit
I brede isozymband met sterke kleuringsintensiteit
wel kleuring, maar het aantal betrokken isozymbanden is niet
duidelijk
kleuringszOne met plaatselijk sterkere kleuring
grens van de kleuringszOne onduidelijk
Ui.kleuringszone die waarschijnlijk uit (in dit voorbeeld) drie
isozymbanden bestaat
Bij elk enzymsysteem staat de afstand van minstens ên van de
isozymbanden tot de origin (links van de banden) aangegeven in cm.
De onderlinge afstanden tussen de banden, vermenigvuldigd met een
factor l,Lf komen overeen met de onderlinge afstanden op de gels.
Niet alle enzymsystemen gaven even goede resultaten. het volgende
tabelletje geeft een overzicht van de betrouwbaarheid van de
gescoorde isozymfenotypen:
GPI EST PGM—I PGM—II 6—PGD ADH IDH
electroforese + + + + + ÷ +
e2 electroforese + ÷ ÷ — — + +
+ = betrouwbaar÷ = rninder betrouwbaar
— = niet betrouwbaar
- 3L -

p1odie—niveau' s
Voor elke populatie staat aan het eind van de lijst met isozym-
fenotypen in die populatie de ploTdie—niveau' s (overgenomen vanKik ? Linders, 18).
De betekenis van de tekens is als volgt:
o tetrap1od0. tetrap1od of aneusoom
El pentap1od0 pentap1od of aneusoomV hexaploTdV. hexap1od of aneusoom• aneusoom
- 35 —

IV/LLI\ / HOOILFj&PI EST PGM -PGD ADH IDHI
.0 .8 '(.4
H II II ID 01 lID10 III 1011 0 11 1101
01 1 i 11 111 ii 11
Dl I I II III 1110 II 0 001
Bi II DII I ID III lIDII III lOll 11 I
138 :1! ID 01 I II0 I I III IOU II I 001
II VI. I I Il 111 lID111 III IOU DID 0111II 0111 1101 1111. I I III III 100 DI I VII
l UI UI I I ID DI DIII 10 1111 001
l0U DIII III 11111 DII 0111
1111 III I I ID II 1101 ID II 0 DVI
IDD 01 I I 10 III 10 DID VOl
1111 DI I I 10 0 I DIII ID 1111 001
Ill11 Dl I I ID III ID 0111 001
till DI I I III DI 001 lOll lID 0111
IDII DI I I Ill III ID 010 001
Dl DI I I ID III 0111 00 110 001
II Ill 11 IV! 0 111 0111
• Bzj I 11 I ill I I ID DI 1101 00 lID DIII
DI I I 10 III II OIl 001
1111 DI I I ID DI 11111 DII lID DIII
II ID 11 III! ! I 0 1101
33 IDI DI I I 00 DI 0111 iD II 11 ODI
111 I I ID III ID 1110 DIII
1111 III I I 011 DI 011! 0 1111 1101
II III 11 101 010 001
1111 III I I 011 II I 001 0 I I 0 001
tOO DI I I ID III 10 DII DIII
1111 flu I I 00 01 001 V 110 001
I I I 0 VI 000 10 I
III lID lIlfl III 1111
— III 0 III 01111 J I I
- 36 -

I I
I
I I
I
I
I
I
I
I
I
I
I I
I I
I I Ill (Oil 11 Ill 0 (III
Bis II 11 01 0011 I I
I 110 I0!I 0 DII (DI
Bi 001 11 DI I I U I I lOll 11 I 101
III 1101 1111 II Ii 11 0(1123 II II Ol DID II lilI a I 11111
3O
II
OIl11 III
0 0!
lID
IlO
I I
III
lilt0
11 ..
I Ill.0 .
...i
iiiI III 11 I 000 (0 liii Ill Ill 1111
liii 0 01 (III II III! 0 I (Ill
BIII1100III0011
I
VOl
11 II
0V0
I
lb
00
1100
I
II(0
(lUll0
I
fIll
JIll
U
ID
1.11(00
U .
1111
I
11(11
I
0 .1. 11
,
(00
...._.i
101
(01
BC
fl
B4
(II III I I
I 11 I
HI110I
I I I
I I I
...
I
(0
(11
10
(DI
III
. .
(III
100
101
1110101
1111 .
IOJ
.
.
.
.
.....
. 0111
III I
lIDIII I
HiI] .1 I
I I U .
:_..J
1101
DIII.
1101
I11L.±ii0111
DOI
B1
B3
B'i
I I I
I I I
III
II I Il IIIII l
I
IOU
110
011
III
1111
10
Ill
III
1111
UI
DI
II
.
00
IOU
(110
1111
1111
III
ID
11
ID .
111011,
.
.
;
01 I
. I I
oil.
.
lID1111
:.. $1.11
OIl..
.
DIII
Ill
IoU! 1101..Oil
III i
1111! .....i.._1
1111 11.__11
6
B
83
B
I OHl11j I0( I
IOU! II(Ill! I
OIII IIIIII
III 1
.
110111
DIIIII 11
11111! LU
.
OI I II
..
(80
IOU
ID
10
1111
110
ID
DI
.10
III
.11 I
Al
ill
III
I
IIIUI
Ill .
Ill
II
Ill
0
.
.
.
.
1
.
UI
0111.
lID.11(11
11
11 Vi.
0 I 11
.
.
l0l1/.IIOL..
.
.101;
.1111 ......J
OOI
III I I DI I II 111 III III 11 I 111 001 1
3? —
Ill
ID
ID
Ill
10
ID
10
(11
00
ID

III III 11:10: III III
H J ( ID 0 11. I 08
(Acosic . 81 11 0 11 I 8 0
0 00 ItO 0
L B 108011 III 11 0 I I 8 I 101
1 11111. U I 0 Jil( 0JII .18 OIl II8 lID Oil
.. II I I 8 81 8 11111.II . . 18:81 ID VI 0 18118 JillIIII till.
0 Oil tat/too
11011 DI II U I 10 81110Ili (1 8 11 II U 801
1(11.111 .. ill 11 II 0111
(tIll II 10 11 01 081Il II . II I 11I 0 11 111(0 tot
ii 01 lilt I 8 18(/180
PLOoe PLQo
L... B. 0. a.
B3
83 0 BBs
H.. B . .. B
.. .B •
L.. s o .
L . Bct
L . .a. I;Icc
H0
H • Bc;
H •
. .B1. 0
H
— 38 —

GPI EST/ KVJLD
AD Loll
NOO RDPOLOER7YL
P&Mi it
Ni
.o4.((III
4.04
11(11
$94, 1
11110 I 0
3.+II &"
8I fill 0 0 Ii UI 1101 0 U II (00
N2. II11 DI III 0 11 I (00H I U 11 11(0 lUll
N3 I I I (DI Ill II I I LU
Ij 0 11 • III! 01¼14 II I 1 (UI 11 (I 1110 to
j_ (0 11 III1'Is• iii II 11110 0(0 110 (110Ill ill III 11 I R IOU
N (fill IV (1100 I11 10110 U
I. 11 DII Ill
M. ii 011 110 U II (II)II t
11 11 (00
ta 101 II II ID 11 III (11 .. 1111
III VII11 11 I
N (DI II II (11 0 III 11 0 10
III H OIl11 a IDIII
11 1101 (0110 00
I1 11 I U 11 (110
Nt I1 1111 III .0 EU
II II H 1111 11 1 11.11 ot/Ntz 111111 III. I 11 (011 .111 110
I 010311 11 flU ID II 1ll
Nt3 11111111 H 1111 III (UI Ill I LII (1111lilt Di11 110 1111
N4 (UI ID I I Ill (II (00 (11 i 01 0 (011
I00 II 11 '11 1111. I 0 1 1111 ([III
N 11111111 II 811 111 1111 111 1(11 (110lIlt Dl11 111 (011
Nq. (11 II (UI 0 ID III ID I 11 III
01 Ii to! 11 11 1111
N IID 111 11 0 III ! .10 i I 0 III
DIII 10! 11 DII IOU
Mi 1(111111 III 00 110 III I 0 . IOU
011I 11(0 0 . 1 UI, IOU
fta I 8010 01 DO 11 IOU .18 I 0 III
I11 iio Ill
1
I
1
—3
— 39 —

0l
11 III
I 0 DI
Iii UI
II DI 0I
DI
10 UI
Ia II
III DI
Ill 0 DI
I U D
11
DI 0
U
U
UI
DI I 10 0
HI 11
ill U
Ill U
U DI
III ll
UI 01
11
DI DI
111
UI UI
U
ID U
11
0 0
flu
0 11
II ItO I H11011
UI •I I I
I I .101
II
IIII I I .1 I I I I
11001
11 0 01
0 1101
Ii 110
II 11110
lilt I 1101
I I Ill II
H 001
111111
110111
I I 1111
I I liii 1111111
DI
I I©Il
I I I
DI 0 II
IDOl
110 011
01@
111 DI
II DI 0 i
— Oi - DI U ID U
UI 0111 DI
01 Ht
UI
I I
UIII fill
11
0:
A
ID
.IUI U
rH
II o
U'
'ml . .. Dl
11111 I I 11 0
- 11111 0 0 D III
101 0 Iii' II
0111 1' UI 100
ID) 0 I I I 0
.
'1111 I 1111011
1101 11011 0
001 I DI U
0111 II II U
1101 I UI DI
11101
.101
11
I
101
I I
11
UI: I
.
IOU U II ' 11'
'1011 I . 01 1101
lAO (1 I I 11
IOU I ''" DI 1101
H 001 lID 0
1101 11 II Dl Ei
AOl 0 I I 0
001 0 1 ' 0 1101
1101 11 II U
H 0111 UI DI 0111
11111 1 II U
1101 Fl Dl. tIll H oo II F
1101 I lot DI l oi i tü A
UI
lOt/tOt
I
11
11
10
DI, 1111
0
r I o III
1101 0 I I 11
1111 11 DI 0111
0 UI ID
11111
U I
0
II 11110
I I
010
UI
II II I
1111
0
Ii UI UI
DI Ill
I ID lID I0
1111
0
ID
DIII
11 Ill
II
II
jO
IhN
N
h4
t
) IJ
hi N
Li
I.

4
— -E+1 -
o tt4
0'
o Ot4
o Gi
'A 9t
•A tt
o
• Qtt) •
0 N •O 'R
o
•
o
'-H
'1
0 Slj
o '14
o £ 0'
0 'N
o
o
o 8MH o w
• N
Q S'
• 9 • H • 'N H
toio1o
Oh o 9"t1
o m o ?h
VN
o ThN
0•
•
o Mj
o I
I ID
all II I'll
II VII
II Ill
a ii Ill
II 1111 I
I II Vi II
U
DI
II
0
II
U
II
a
UI III
a
0
11
V III
U
O fIll
III
aü i
U
UI
a
01
0
DI
DO I
DO!
U
Oaf
DO
ID'
OUt
DI
IV VI
I V UI
1 DI
II DI
0
H VI
I VI
II VI
VI
II VI
I VI
0!. VI
DI
ID Vl I A
I' V
Il' III
111011
UI III I' 11101
'U •
IV lID
I II Fl V
1 II IVI
I 111111
Dliii 1111111
I III ill 1 11 lUll
II 1110
I 'I DI I V I
1 U 111111
--H
:,111J
'N1 --H
VN

. LAU\JK(itt.k ./. PULU-&.PI :E.5T. P&t1 -P&D ., ID
. t .-¼.4 . r
.
,9 ' J.4
,
. 4
H tO (0 8111 tO • till]
1101 II tO [I till to i tO 11111
II 11 ill tO 0111 II
110.118 811 0111 l0 0. 1Q tO
o.oi it Ii to tot to ito too
011110 .11 (1 I [OJItI 8101 tOil
111111! tO l till tO 00 1
11181010 • C) I 1111 11 00 0 tO
II tO 111 000 tO 081 0 tOll
Ill tO . 0.11 . . I 11 lOt I tOil
It • .... to to tO 110 (1111
.001. 10 .1 III IJJ to 0! 8 lilt
Jilt!: Ill ... 11 .111 lOt (0. 118 108
tlt I , 0. Ii 11 II 11 tOt ii to at itt it 11
111110 I II ID it lOt 0 I 10 ID
II 0 tO III ill ItO tilttll DO II 11 III (00
(tIlt: II III 10 11 ItO18100 0 JO 111 11 lID tOllhIDI
tOO II 11 01 to I I II
li_li I I 11 0 I 11 tO
III tO GO III Ill ItO ID!
I 011 0 0 It 0 C)! 11 10!
00 00 110 III Ill I I till
I tOO tO 11 II 11 0 I 0 JO!
00 1111 • 10 00 111 tO I 111 lOll
1101111 II Jill [ lOt 1110 1811
11100000 111 011 In tO I II tOO
tIlt HO I 0 0 0 lID 100
0111 II 011 1110 II Ill I IOU
Ill 10 III 011 10 i 10 DII 001/101
1000 DO 11 II I Oil tO 81 tOO
1000 1111 I V 111 I I 0 100
11810 00 tO 10 F II! 810 ID
10101 l ill III 10 81 ii tO
1011 tO It UI tO ItO 1111
I 100 Ill 0 H U 010 III!
— 2 —

• H
-1
_1
0 00 II] 00 11011 III 11111 1101
H II 0 III III 111 010 00I
122. 0011IJ I 11 11 I F Ill 01 I II
0001 I 111 11! 0 11! Ii Ill
Lz HJ1011 01 l.I11 1111 ID
10110 .1 0 11! I .0 .. .110 lU
L2. - 00 - .I. Ill 000111 - .1111 1.01.
I 0 -- H I - 11 11 1.0 ;.i . -
L.z H0. 10 Dl I1ll lID III
1101111 Dl II Dl 1 11 I Ill Ill
Lzi 1111 00 I ill IF 0 DID IOU
lOt ID 11 I V DI 11l 1011
128 00 H HI I Ill DII IOU
III 111 I .1 I 11 • it II 1110
Lzo 11010 H •• 11 Dl 11111 10 I I IA
110110 I I V DI I .0 • - - lID 0
1o OOIIII lOll lOUD III ID
101 ID I II11 1111 11
13 11 011 01 Ill 110 III
I 0 II0 1111 11
L32 l II 0 1111 III ID I 101
OI JO I I 11 .0 IL3 011 III tO I ID II 10
11 0(11 ID I I 11 Il I Ill
L3 I II 011 Al 1111 IA lID 1011
I1100 1111 0011 0 lUll
L3 001 II 1111 01 ID 110 III
00) 10 H11 OlD II
137 11011111 011 I 0 10 0111 IV!
I Oil DI• 011 0 Dl I loll/till
L OUt III 11 I II ID Ill IOU
001111 II 10 0! 11 OlD lOll
133 II ID II III ID ItO IDO
O O III U I 11 1110 101 loot
L40 1101111 DI IA ID I IA
lOt 11 0 II11 Dl / Ill
L41 I I I 0 0 10 1101 I 1111
I 0 I I 0 DII tei/ooi192 0I lOlA 0111 lID Ill!
O lUll IA 0 II 0 DIll 1111
L43

III . ••. 11 00 • U • I 10 IOU
L_I a IOU
LL _JUIIIIII U ID E U 0(0 118
• [0101 jU ........ 1 0 0 II I (GO/lot
F
IDIOt IU —— tO '11 11 0 110 ID
— UI 811 DI to
0 II It 0 101 III
'JUIOII 1 I 0 0 lID :110
10 I I U 118 101
I IOU 10 11 II U 0 I 0 tillI I 0 0 111 I 0 010 lOt
• U 1 8 10 IOU 11 DI I IOU
L Lco . . .. I II I I 0 I I 0 lOt
• I. IOU [II U II 0 010 0!
2.0ioH L o L33
• L o
H b 0. . 0..0 . • • C'
• Lz. 0
U o• o• L .0
• • o. Lz3 o L, E1..
• L 0. L'i o to 0
..•. L o Lzs 0.Lo 0 LA 0
L:_ (. • o (.'H... Lr7. .• .1.28 o L4'i O•
L 0 o L'is 0.
F-—'• • .
o
F—L o • o.Li o • a a
L4j
- L4 —

-1.
---
.-
(A•
U"-
-r-
•
.Ir
.!r•I
—:.
•;•'
••i
•..•
U',•
.U
'-Lty
-N
.
C =
— C
a =
00
= =
0 =
00 a
0 C
- =
-
—-=
C =
= -
•In
.1
— * rn 0 z Zr ot 0-
a a
0 C
C —
C=
= =
= C
= =
C 0
= =
a =
= =
— —
0 0
C 0
0 =
aO
00.
-C.
0 —
= 0
= 0
= C
= .
= ..
=•
— r
-.. —
— .
— —
—0
—=
— —
a a
0 0
= =
= =
C C
C —
— =
C C
= —
—
— =
—-
——
•—=
._-C
-_•
—
---:
•-
-—-
-
—..—
--—
-•--
.•-
-.•
•
- -
- •
-
= -
— —
— =
a —
==
— —
- —
-o H
C -
= C
= =
0 0
= =
0 =
000
= C
0 0
C —
• =
•0 —
= -
—-
--
- =
-
= •
-•=
= =
.=
--=
c —
_C
= =
C 0
00
0
= 0
= 0
— =
C—
-0
•--0
-.
. --
C.
C
o=
C=
a=
—0
= =
— —
C =
= C
= =
0—
0 0
0 0
0 =
0-
.•
.•
—-
- —
—C
a —
==
-
-C
.—
I•
•
z-
=C
t O
-
— a
——
—•-
:
.- H
0 =
C =
C
— —
• •
0•0•
0 0
—C
- =
— C
o•c
•• —
= =
— C
C0
==
— •
= •
— -
C
= —
C =
C

011/101
11IJ I
01
Ill
fill
Ill
log
1111!
001
ID!
11111
Ill I
DIII
III
I00
11111
11111
III
1101
Ill I
0111
1110
1111
lOll
IC;
OIl
00!
III
II
111
III
Ill
11
a'
Ill
III
ohç
o 11 1 I O H
III!) 111111
ID
I 111111
Ill I 11101
I 0111101
II 11111
ID. 10 (I lID 011
I 11 11111
10
II 110 I 01011
1111 hIll, II I II. 11111
11 l I! Ill!
10 0111101
0 I 11111
11 101111
II III
II 101
UI f 11101 01 II
Al :. liii II
H
lIQ !
f 11fF II
101
Ill III III 1111
Ill 1110
OHI
1111
DII CII
11
DII 11 III
III III fill Ill ill' fill fill 1111
Ill II III Ito III II 0111
110
010 11
1111
Ill fill fill
0
11
11
0
II
A.
11
10
11
ill
0
ill
11
111
0
I
11
0
I
11
DI
11
11
II
F
11
'I
11
III
11
II
II
III
Dl
11
DI
DI
DI
— 9+7 —
Dl
UI Ill
III 11
Ill II
00 II 0
II III
II DI
I ill
II DI
III 11
II I
I Dl
I 110
II 11
III 11
I 11
II 11
II 0
111 11
II 11
II U
I I
II 11
1111 DI
JO UI
0
ID ill ID II
Ill U
I I
I, I
11
II 11
ill
II 00
ID DI
II UI
1 II
Ill
1111
11111
Oil
II
III
'Oil
III
II
Ill
I
III
III
III
II!
II!
00
I!!
1110
III
Ill
Iii
Ill
Ifl
II
III
DO?
lii
ocsh
I 11
Ill
II
111
I I —
LIII H
I
H J•ff
I 1 I I
loll
DI 1010 . . HI III
O lID I
I 0011 1
Ill III —
I ii
11H

11111 II 0 II UI U . lID IOU
DIII HO I II 001 V III U]
III OIl ID ID II III 0 010 00
IDO I ID DII II DVIII lID lUll
11101 I I I II U I HI! U U I I I DO
ID I I LU UI II DID IOU
I DI I 0 !D U II I 0 0. U I U 1111I0 ItO 00 V 1110
011110 U ID ID III, U . VIII ID
DOt D Ill I II II 1111 .553 0 III U II III III 0 0 I lOll
I!O DID II Ill UI II . . lID Ill
IHII ID III DI III 0 . lID Ill
10110 0 Dl 101 0. 0111 100
II II I I I U ID III U. GIll lOll
O II II U I I lOll
LoiE LOWtE
Si 0 0
57. 0 Zo 0
53 0 0 S3
Sq 0 Szz 0 SMO..0
Ss o 5z 0
S 0' 0 942 0
0 5 0
58 0' 0 0
5' o S o & .0
So 0 $18 0 0
SI' 0 0
Sa 0 53o 0 5 o
S'3 0 531 0
5' 0 5 o .0
Ss 0 5 0
S 53l
59 0• 55 0
5e 0
- 47 -

9.L. bijiageiV: verdeling van PGM-I en IDH fenotypen (zie biz. 12)
PGM-1 PGt1-2 PGM- PGM-1-i- PGM-5
N2Lf N6 N8 N9 N21 N3 N4 320 Ni?
L2 L7 Lii N39 N40 N41 NLfLf
L19 L22 L23 N1+7 NLf8 NLf9
L26 L29 LLf L8 L30 L3i
S5 S26 S28 L34 L36 LL+O LL+5
S41 S1+5 511 S23 S51
N2 N7 N12 N5 NiO N13 N15 Ni Ni1+
N33 N3k N37 N18 N22 N27 N28 N20 N26
N L3 L18 L27 N29 N1+6 N50 NLf3
L13 L4/+ Li L5 L14 L35 L16 L28
0 S2 S3 S13 L38 S29
S21 S33 S3L4. 56 S12 SlLi. S20
S36 s38 SLf1 sI s48 S19
S1+7 S55
B38 Bk5 S8 BLf 35 B6 37
N30 N3i 38 39 B12 Bi3() S4 S37 B18 B19 B21 322
B21+ B26 B27 B28
B32 B33 B31+ B35
336 31Q B/+i 31+6
B47 Bk8
S25 S3i
L32 B2 B3 B14 B1+3 350 Bi BlO
S22 N35 N36 N38 Bli B15
L12 L13 L20 L39 316 B23
L1+2 L'+7 L50 B30 B31
B39 B42
B37 Nh N25 N32 L21 L21
N23 N1+2 L6 L10 Li? L37 L41
L9 L/+6 Ll8 Si s18 S1+0 S1+2 S30 S53
S7 S9 Sb S1+6 S51+
Si5 s16 S50H
- 1+8 —





























