Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, JUly 1994, P. 2265-22710099-2240/94/$04.00+0Copyright © 1994, American Society for Microbiology
Degradation of Alkylphenol Ethoxylates byPseudomonas sp. Strain TROl
HIDEAKI MAKI, NAOTO MASUDA, YASUHIRO FUJIWARA,MICHIHIKO IKE, AND MASANORI FUJITA*
Department of Environmental Engineering, Faculty of Engineering, Osaka University,2-1 Yamadaoka, Suita, Osaka 565, Japan
Received 27 December 1993/Accepted 13 April 1994
An alkylphenol ethoxylate-degrading bacterium was isolated from activated sludge of a municipal sewage
treatment plant by enrichment culture. This organism was found to belong to the genus Pseudomonas; since no
corresponding species was identified, we designated it as Pseudomonas sp. strain TROl. This strain had an
optimal temperature and pH of 30°C and 7, respectively, for both growth and the degradation of Triton N-101(a nonylphenol ethoxylate in which the average number of ethylene oxide [EO] units is 9.5). The strain was
unable to mineralize Triton N-101 but was able to degrade its EO chain exclusively. The resulting dominantintermediate was identified by normal-phase high-performance liquid chromatography (HPLC) and gas
chromatography-mass spectrometry as a nonylphenol ethoxylate with 2 mol of EO units. A carboxylatedmetabolite, [(nonylphenoxy)ethoxylacetic acid, was detected by gas chromatography-mass spectrometry. Thisbacterium also metabolized alcohol ethoxylates with various numbers of EO units but not polyethylene glycolswhatever their degree of polymerization. By oxygen consumption assay, the alkyl group or arene correspondingto the hydrophobic part of alcohol ethoxylates or alkylphenol ethoxylates was shown to contribute to theinduction of the metabolic system of the EO chain of Triton N-101, instead of the EO chain itself, whichcorresponds to its hydrophilic part. Thus, the isolated pseudomonad bacterium has unique substrateassimilability: it metabolizes the EO chain only when the chain linked to bulky hydrophobic groups.
Alkylphenol ethoxylates (APEs) are polyoxyethylene-typenonionic surfactants aimed at industrial applications ratherthan household use. APEs are divided into two groups accord-ing to the number of carbon atoms in the alkyl chain: octyl-phenol ethoxylates and nonylphenol ethoxylates (NPEs). Thelatter are more widely used at present. Most of the alkyl partconstituting the hydrophobic area of APEs is composed of thehighly branched nonyl or tert-octyl(1,1,3,3,-tetramethylbutyl)group. Because so much attention is now being paid toenvironmental considerations, APEs have been recognized as
considerably refractory compounds among chemicals releasedinto surface waters (17, 30). In fact, complete degradation hasthus far not been reported, although partial degradation was
found to occur exclusively on their hydrophilic regions, i.e.,their ethylene oxide (EO) chains. It has also been revealed thatthe shorter the EO chain of an APE becomes as a result ofbiodegradation, the stronger is the toxicity of the compound(21, 34). Moreover, bioaccumulation of APE degradationproducts in some organisms has been found because of thestability of these compounds (3, 7). The use of APEs was
recently prohibited by law in Switzerland.Common intermediates of APE biodegradation have been
found in various sources such as river water (26), effluents frommunicipal sewage treatment plants (6), and digested sludges(9). Through detailed investigations (9, 18, 24), these metab-olites have been shown to be alkylphenol and APE with 1 or 2mol of EO units and their analogous forms with a carboxylatedend of the EO chain instead of a hydroxyl end (1, 23). Theabove findings, however, were obtained through interactionsbetween numerous biocenoses such as activated sludge from
* Corresponding author. Mailing address: Department of Environ-mental Engineering, Osaka University, Yamada-oka, Suita, Osaka 565,Japan. Phone: 816-879-7674. Fax: 816-877-8497.
sewage treatment plants or other microcosms in the naturalenvironment. It remains to be elucidated whether these biointer-mediates are formed during metabolism by complex microbialgroups or by a single microbial species, in contrast to our
comparatively abundant knowledge of linear alkylbenzenesulfonate (LAS) biodegradation, in which complete degrada-tion is accomplished by synergistic metabolism of the definedbacterial group (14, 27). Thus, it is worthwhile studying themicrobial contribution to the degradation of this nonionicsurfactant. In this paper we report on the isolation andidentification of a pseudomonad bacterium which is capable ofutilizing APE as the sole carbon source. This bacterium isshown to possess a unique metabolic system for APE degra-dation, in contrast to the polyethylene glycol (PEG)-degradingbacteria reported so far (4, 6, 11, 16, 32, 33) (PEG is a
structural molecule of APE).
MATERIALS AND METHODS
Screening conditions and cultivation. An APE-degradingbacterium was isolated from activated sludge from a municipalsewage treatment plant by using enrichment culture and serialtransfer, with Triton N-101 (0.01% [wt/vol]) as the growthsubstrate. A minimal salts Ri1 medium was prepared as
reported previously (5). CGY medium (0.5% Casitone, 0.5%glycerol, 0.1% yeast extract) was used for cultivating organismsfor resting-cell preparation. The cells were grown in 100 or 300ml of liquid media in 300- or 500-ml Erlenmeyer flasks,respectively, at 28°C on a rotary shaker at 120 rpm.
Chemicals. Triton N-101 was manufactured by Bohm &Haas Co. A series of alcohol ethoxylates (AEs) was purchasedfrom Sigma Chemicals. Nonylphenol, NPEs with a range ofEO units, LAS, ethylene glycol, glycolic acid, glyoxalic acid,and fatty acids were purchased from Tokyo Chemical IndustryCo., Ltd. Triton X-100, sodium dodecyl sulfate (SDS), and
2265
Vol. 60, No. 7
on August 19, 2018 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
E 100 -u.
--0-.-. A600 -0.3
U~~~~~~~~~~~~~~~~~& 50 ° -*'O 0.20 -n. b -0.1
0T ~~~~~~~00 10 20 30 40
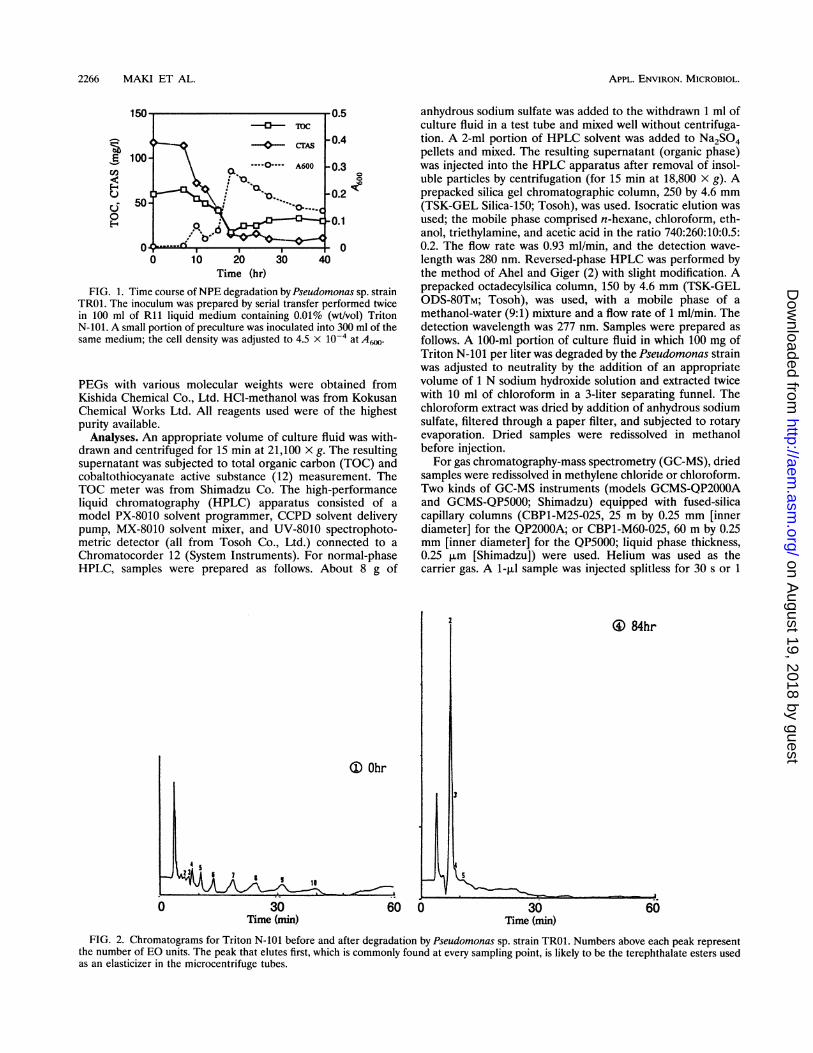
Time (hr)FIG. 1. Time course ofNPE degradation by Pseudomonas sp. strain
TRO0. The inoculum was prepared by serial transfer performed twicein 100 ml of Rll liquid medium containing 0.01% (wt/vol) TritonN-101. A small portion of preculture was inoculated into 300 ml of thesame medium; the cell density was adjusted to 4.5 x 10-4 at A6.
PEGs with various molecular weights were obtained fromKishida Chemical Co., Ltd. HCl-methanol was from KokusanChemical Works Ltd. All reagents used were of the highestpurity available.
Analyses. An appropriate volume of culture fluid was with-drawn and centrifuged for 15 min at 21,100 x g. The resultingsupernatant was subjected to total organic carbon (TOC) andcobaltothiocyanate active substance (12) measurement. TheTOC meter was from Shimadzu Co. The high-performanceliquid chromatography (HPLC) apparatus consisted of amodel PX-8010 solvent programmer, CCPD solvent deliverypump, MX-8010 solvent mixer, and UV-8010 spectrophoto-metric detector (all from Tosoh Co., Ltd.) connected to aChromatocorder 12 (System Instruments). For normal-phaseHPLC, samples were prepared as follows. About 8 g of
(D Ohr
0 30 60Time (min)
anhydrous sodium sulfate was added to the withdrawn 1 ml ofculture fluid in a test tube and mixed well without centrifuga-tion. A 2-ml portion of HPLC solvent was added to Na2SO4pellets and mixed. The resulting supernatant (organic phase)was injected into the HPLC apparatus after removal of insol-uble particles by centrifugation (for 15 min at 18,800 x g). Aprepacked silica gel chromatographic column, 250 by 4.6 mm(TSK-GEL Silica-150; Tosoh), was used. Isocratic elution wasused; the mobile phase comprised n-hexane, chloroform, eth-anol, triethylamine, and acetic acid in the ratio 740:260:10:0.5:0.2. The flow rate was 0.93 ml/min, and the detection wave-length was 280 nm. Reversed-phase HPLC was performed bythe method of Ahel and Giger (2) with slight modification. Aprepacked octadecylsilica column, 150 by 4.6 mm (TSK-GELODS-8OTM; Tosoh), was used, with a mobile phase of amethanol-water (9:1) mixture and a flow rate of 1 ml/min. Thedetection wavelength was 277 nm. Samples were prepared asfollows. A 100-ml portion of culture fluid in which 100 mg ofTriton N-101 per liter was degraded by the Pseudomonas strainwas adjusted to neutrality by the addition of an appropriatevolume of 1 N sodium hydroxide solution and extracted twicewith 10 ml of chloroform in a 3-liter separating funnel. Thechloroform extract was dried by addition of anhydrous sodiumsulfate, filtered through a paper filter, and subjected to rotaryevaporation. Dried samples were redissolved in methanolbefore injection.
For gas chromatography-mass spectrometry (GC-MS), driedsamples were redissolved in methylene chloride or chloroform.Two kinds of GC-MS instruments (models GCMS-QP2000Aand GCMS-QP5000; Shimadzu) equipped with fused-silicacapillary columns (CBP1-M25-025, 25 m by 0.25 mm [innerdiameter] for the QP2000A; or CBP1-M60-025, 60 m by 0.25mm [inner diameter] for the QP5000; liquid phase thickness,0.25 ,um [Shimadzu]) were used. Helium was used as thecarrier gas. A 1-pl1 sample was injected splitless for 30 s or 1
a) 84hr
30Time (min)
FIG. 2. Chromatograms for Triton N-101 before and after degradation by Pseudomonas sp. strain TRO. Numbers above each peak representthe number of EO units. The peak that elutes first, which is commonly found at every sampling point, is likely to be the terephthalate esters usedas an elasticizer in the microcentrifuge tubes.
2266 MAKI ET AL.
on August 19, 2018 by guest
http://aem.asm
.org/D
ownloaded from

DEGRADATION OF ALKYLPHENOL ETHOXYLATES 2267
min. The gas chromatograph temperature programs were asfollows: QP2000A, 70°C isothermal for 2 min, increasing to240°C at 4°C/min and then maintained at 240°C for 5 min;QP5000, 50°C isothermal for 5 min, 50 to 325°C at 17°C/min,and then maintained at 325°C for 30 min. The electron impactconditions were as follows: ionization energy, 70 eV; ionizertemperature, 250°C; mass ranges, 40 to 900 or 50 to 400 mlz;scan time, 1 or 1.5 s. For carboxylated metabolite detection,sample extraction was carried out in the same way as forreversed-phase HPLC, except that the culture fluid was acidi-fied to pH 2.0 by addition of an appropriate volume of 18 Nsulfuric acid before extraction. The evaporated samples werefurther dried in a desiccator. To the dried samples, ca. 1 ml of5% HCI-methanol and the same amount of dehydrated meth-anol was added, and the samples were allowed to stand in thedesiccator overnight at ambient temperature. Methanol sam-ples were diluted with 10 ml of deionized water and extractedtwice with 5 ml of methylene chloride. Methylene chloridesamples were extracted with 2 ml of deionized water to removeremaining HCI, dehydrated with Na2SO4, and concentrated bya rotary evaporator.Oxygen consumption assay with resting cells. Cells were
harvested by centrifugation for 15 min at 21,100 x g and 4°Cfrom overnight culture in CGY medium and washed once with1/15 M phosphate buffer comprising Na2HPO4 and KH2PO4(pH 7.0). Washed cells were suspended in the same buffercontaining the substance to be assayed (1 mM), the cell densitywas adjusted to 1 at A60, and the resulting suspended cellswere incubated with agitation on rotary shaker (120 rpm at28°C) for 2 h. Incubated cells were harvested, washed, andresuspended in phosphate buffer as described above. The celldensity of the suspension was adjusted to 0.5 at A6m0 and thenused for assay. The assay was carried out at 30°C by using aClark oxygen electrode connected to Flat Bed recorder(Sekonic Co., Ltd.); 6 ml of cell suspension was poured into asealed reaction vessel. After measurement of the endogenousrespiration, the assay was started by adding 60 RI of substratesolution (5% [wt/vol] Triton N-101 dissolved in phosphatebuffer). The oxygen consumption rate at the expense ofsubstrate was determined by subtracting endogenous respira-tion values from those obtained with substrate.
RESULTS
Isolation and identification of APE-degrading organism. Abacterium capable of growing on Triton N-101 as the solecarbon source was isolated by enrichment culture. Moreover, itwas shown to degrade the EO chain of Triton N-101, asconfirmed by halo formation around colonies on an Ri1 platesupplemented with 100 mg of Triton N-101 per liter in areaction with overlaid modified Dragendorffs reagent (4). Thestrain was determined to belong to the genus Pseudomonasfrom the following characteristics: gram-negative rod, motileby polar flagella, and catalase positive. However, since nospecies that completely matched all the tested properties(production of fluorescent pigment and urease; hydrolysis ofarginine, gelatin, starch, and Tween 80; reduction of nitrateand nitrite; denitrification; utilization of citrate; accumulationof polyhydroxybutyrate; meta-cleavage of protocatechuate; andgrowth at 4 and 41°C and pH 3.6) could be identified, wenamed this strain Pseudomonas sp. strain TRO0. It differs fromother reported APE-degrading bacteria-from Pseudomonassp. strain 14-1 (2) in color tone of the pigment and in nitratereduction, and from a Pseudomonas sp. nov. (20a) in pigmentproduction and cell size.
1
co0000.
0
0a:
0.5,
0 18 36 54 72 90
Time (hr)FIG. 3. Variation of the representative peak area for Triton N-101
as a function of degradation time by Pseudomonas sp. strain TRO0.Each peak area is normalized to its maximum. Symbols: O, 2 EO units;O, 7 EO units; 0, 10 EO units.
Triton N-101 degradation by Pseudomonas sp. strain TROl.A typical time course of Triton N-101 degradation by Pseudo-monas sp. strain TRO1 is shown in Fig. 1. A fluctuation inA600,which did not reflect only the cell density, was observed.Particles which could be perceived by the naked eye wereformed; these were subsequently shown by microscopic analy-sis not to be bacterial cells, judging from their great sizedifference. These particles were responsible for the fluctuationin the A600 value. Cobaltothiocyanate active substance wasefficiently removed, the final removal efficiency reaching ca.86%. Reduction in the TOC value, however, reached a plateauat 18 h, the final TOC removal efficiency being 58%. Thereaf-ter, TOC removal did not occur at all, suggesting that metab-olites which Pseudomonas sp. strain TROl could not furtherdegrade had accumulated. Structural alterations of TritonN-101 caused by biodegradation were chased by normal-phaseHPLC. In this system, the APE is separated according to thenumber of EO units composing its hydrophilic parts. At theinitial point, an impartial distribution of each peak was found,with dominance of peaks corresponding to NPEs with a higherdegree of EO polymerization (Fig. 2). At 18 h, transientincreases in peaks corresponding to NPEs with less than 8 molof EO units were observed (Fig. 3), suggesting that EO chaindegradation proceeded in a stepwise manner. Drastic increasesin the peak corresponding to an NPE with 2 mol of EO units(NP2EO) occurred at 18 and 36 h, while the peaks of thosewith higher degree of EO polymerization disappeared. At 84 h,almost no peaks except for one corresponding to NP2EO werefound (Fig. 2). NPEs with a few EO units were hardly solublein water, forming particles which had the appearance ofbacterial cells; as mentioned above, these were thought to becompounds such as NP2EO. A600 attenuation after 18 h mighthave been due to the dispersion of such insoluble compoundsby agitation in the rotary shaker. To verify the material in thepeak corresponding to NP2EO on HPLC, we collected thisfraction and subjected it to GC-MS. A sample showing a singlepeak on HPLC was further separated narrowly by high-resolution capillary-column GC (chromatogram not shown),which, on the basis of the structural heterogeneity of thebranched nonyl chain, indicated that the original compoundcomprised several isomers. Two examples of mass spectra areshown in Fig. 4; they indicate that the degradation products
VOL. 60, 1994
on August 19, 2018 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
(a)
30 e0 130 16O 230 260
(b)........ .....-----I
--- 27i'CH2. CH3j... ........................................ ......... ::
209:
:12O.H-121* J
HO-C2H4,O-C2H4±0-<(e
89H_.1O.H-O7
..........,1,-,......................... ......................................i
',CH3CH2j.C-CH3
: CH3:57
30 e0 130 0eo 230 280
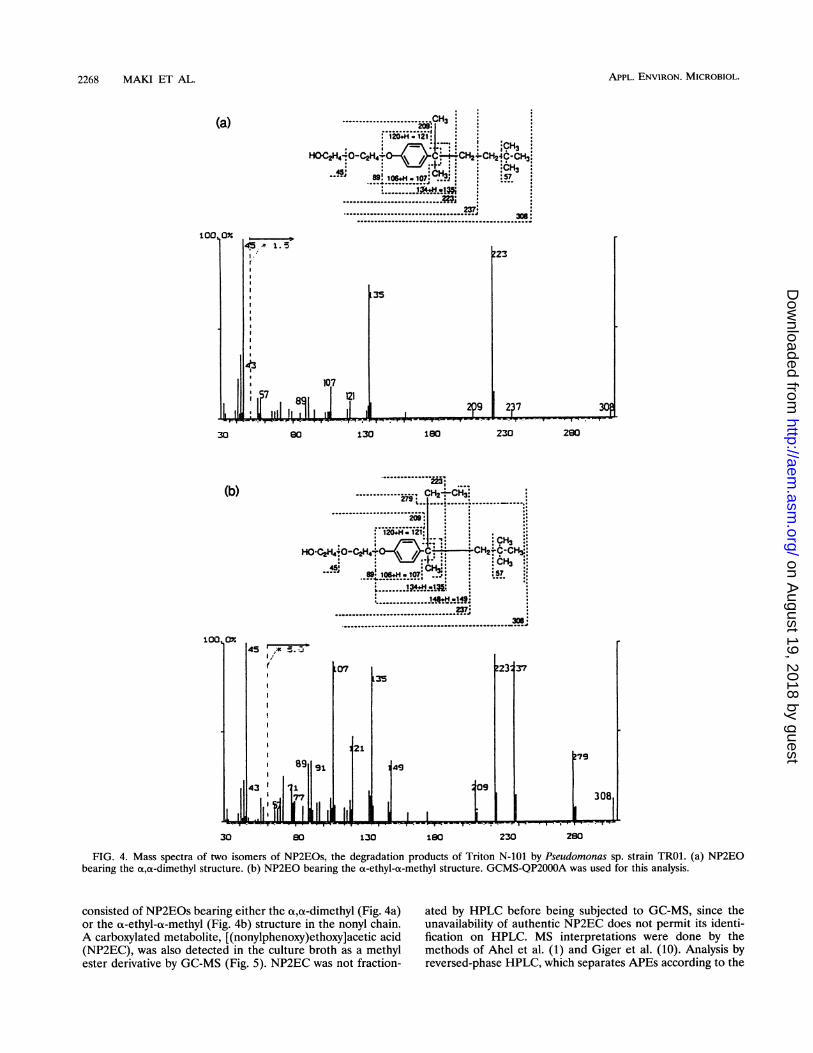
FIG. 4. Mass spectra of two isomers of NP2EOs, the degradation products of Triton N-101 by Pseudomonas sp. strain TROI. (a) NP2EObearing the a,ot-dimethyl structure. (b) NP2EO bearing the aL-ethyl-a-methyl structure. GCMS-QP2000A was used for this analysis.
consisted of NP2EOs bearing either the a,ao-dimethyl (Fig. 4a)or the ao-ethyl-a-methyl (Fig. 4b) structure in the nonyl chain.A carboxylated metabolite, [(nonylphenoxy)ethoxy]acetic acid(NP2EC), was also detected in the culture broth as a methylester derivative by GC-MS (Fig. 5). NP2EC was not fraction-
ated by HPLC before being subjected to GC-MS, since theunavailability of authentic NP2EC does not permit its identi-fication on HPLC. MS interpretations were done by themethods of Ahel et al. (1) and Giger et al. (10). Analysis byreversed-phase HPLC, which separates APEs according to the
2268 MAKI ET AL.
on August 19, 2018 by guest
http://aem.asm
.org/D
ownloaded from

V0FGRADATION OF ALKYLPHENOL ETHOXYLATES 2269
TABLE 1. Assimilability of various NPF-related substances byPseudomonas sp. strain TROI
Assimilability
AromaticsaBenzoatePhenol
Organic acidsaGlycolateGlyoxalate
Fatty acidsdFormateAcetatePropionateButylateValerateCaproateCaprylatePelargonateCaprate
AEsaC5EO3C1QEO8
C16EO20
Alcoholsa
n-NonanolEthylene glycol
Alkanean-Nonane
Anionic surfactantsSDSLAS
4h
_b
+
+
+
+
+
+
+h_h
Final TOC removalefficiency (%)
NFNT
NTNT
0
91.192.387.389.689.087.787.883.0
251;*@Z*o*e*SilCH24-CM3................... , _ _CH^,
0~~~~~~~0.......... ......-------------C-CHCH3C-CH2 0-CpH4 { CCH2!- -CH .:
H3C-0 *
''~~~ ~~~*CH3,1aHiiCHj,148+H ,1, '
0ImJz71.3
76.439.3
NTNT
_b NT
87.0NT
a Concentrations of added substances were ca. 100 mg/liter.b Judged by cell growth.NT, not tested.
d Concentrations of added substances corresponded to ca. 30 mg of TOC perliter.
number of carbon atoms in their alkyl chains (2), showed thatdegradation of the nonyl part did not occur (data not shown).
Strain TROl exhibited optimal temperature and pH of 30°Cand 7, respectively, for growth and Triton N-101 degradation.
Assimilability of substances related to Triton N-101. Com-pounds which Pseudomonas sp. strain TROl is able to metab-olize to sustain its growth were examined. The results areshown in Table 1. Triton X-100, which is the compound mostsimilar to Triton N-101 but has octylphenol as its hydrophobeinstead of nonylphenol, was assimilated in the same manner as
TABLE 2. Induction of Triton N-101-metabolizing system ofPseudomonas sp. strain TROI by compounds related to NPE
Compound used as Specific oxygen consumption Relative ratioinducer (mmol of 02/kg of cells/min)
None 3.9 1.0PEG (PEG 400) 4.1 1 1NPE (Triton N-101) 32.1 8.2AE (C,OEO8) 42.8 11.0ABS 11.9 3.1SDS 12.7 3.3
11
59
I 19 *100
265!i
7 2
135
21 149 237
200
336t
i1 16472162
3071 1279 L 321T
300
FIG. i. Mass spectrum of the methyl ester of NP2EC, a carboxy-lated degradation product of Triton N-101 by Pseudomonas sp. strainTROl. (wGCMS-QP5000 was used for this analysis.
Triton N-101. Since HPLC and GC-MS analysis showed thatthis bacterium was able to degrade the FO chain of TritonN-i(01, PFGs were also expected to be utilized. However,although a number of PEGs with various molecular weights(PFG 2(00, 400, 1000, 2000, 6000, and 20000) were tested, nonewas degraded at all. Pseudomonas sp. strain TROt coulddegrade AEs. In the degradation of some AEs, the final TOCremoval values exceeded those calculated on the basis of thecarbon content of either the EO or alkyl chain except forpolyoxyethylene 20 cetyl ether (AE with C16 and EO2,). Thefollowing final TOC removal efficiencies and the carbon con-
tent ratios of the alkyl and FO chains for each AF were
obtained: for the AE with C5 and E03, 71.3, 45.5, and 54.5%;for the AE with C1O and EO8, 76.4, 38.5, and 61.5%; and forthe AF with C16 and E020, 39.3, 28.6, and 71.4%. These resultsindicate that this strain degraded the alkyl chains as well as theEO chains of the AEs. This was supported by the fact that itassimilated n-nonanol, a series of fatty acids, and SDS. More-over, the last two compounds were found to be degradedalmost completely. LAS was not assimilated. LAS biodegrada-tion is known to be initiated by an attack on the end of its alkylchain (25). Therefore, this pseudomonad bacterium was
thought to be unable to degrade alkane, which is supported bythe disassimilability of n-nonane. With respect to aromaticcompound utilization, Pseudomonas sp. strain TROt metabo-lized benzoate but not phenol. Although the catabolites pro-duced by the bacterial degradation of PEGs or the EO chain ofAEs remain to be elucidated completely, ethylene glycol,glycolate, and glyoxalate are believed to be candidates (15, 22,28). Pseudomonas sp. strain TROl utilized the last two com-
pounds but not ethylene glycol, which suggests that the TritonN-101 degradation step by this strain might be involved ingeneration of these compouinds.
Induction of the Triton N-101-metabolizing system ofPseudomonas sp. strain TROI. An EO chain-degrading enzymeassay has not yet been established. In this study we tried toapply an oxygen consumption assay to evaluate the activities of
Compouind
Vol. 60, 1994
6a
z
on August 19, 2018 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
Triton N-101 degradation. As shown in Table 2, the specificoxygen consumption rate of the cells incubated with thesubstrate was ca. eightfold higher than that of the control. Thisresult suggested that the Triton N-101-metabolizing system ofPseudomonas sp. strain TROl was inductive rather than con-
stitutive. As mentioned above, the strain degraded the EOchain of Triton N-101 or AEs but not of PEGs. We investi-gated which moiety of Triton N-101 contributes to induction ofits metabolic system. We observed that the specific oxygen
consumption rates were amplified about threefold by incuba-tion with SDS and alkylbenzene sulfonate, indicating that theseanionic surfactants could play a role of inducer. In contrast,PEG did not exhibit an inducing effect, which accords with thelack of assimilability of PEG by this bacterium. The common
moieties that are found between nonionic surfactants such as
AEs or APEs and anionic surfactants such as SDS or ABS are
the alkyl chain or arene corresponding to their hydrophobes,and it is unlikely that the sulfate or sulfonate corresponding tothe hydrophiles of anionic surfactants contributes to induction.As a consequence, the Pseudomonas sp. strain TROl metabolicsystem of the EO chain of Triton N-101, which corresponds toits hydrophilic part, was not induced by the EO chain itself butby its hydrophobic part.
DISCUSSION
Only a few APE-degrading microorganisms have been re-
ported so far, in contrast to many that are known to degradePEG (6, 8, 13, 16, 22, 29, 32, 33). Maeda and Mikami isolatedseveral NPE-degrading pseudomonad bacteria from varioussamples and reported that these strains degraded NPE almostcompletely, the final TOC removal efficiency reaching approx.
90% (20). One of these bacteria, Pseudomonas sp. strain 14-1,was further characterized and shown to degrade AE and SDSefficiently but not phenol and PEG (19), which accords withthe characteristics of Pseudomonas sp. strain TROt. However,since Maeda and Mikami did not analyze structural changes inthe degradation products, it is unclear how NPE degradationproceeded with the strains they isolated. Nevertheless, theyspeculated that the remaining 10% of TOC was attributable toNP2EO (20), since this is known to be a ubiquitous recalcitrantmetabolite of NPE. However, judging from the value of thefinal TOC removal ratio, i.e., 90%, these pseudomonad bacte-ria also seemed to degrade NP2EO. An analysis based on thestructures of the metabolites would have provided a more
refined comparison between these two APE-degrading Pseudo-monas strains.NP2EO, a persistent compound that is generated via NPE
biodegradation and has been detected widely in surface water
environments (10, 18, 24) along with NP2EC (1, 23), was foundto be the end product of Triton N-101 degradation by Pseudo-monas sp. strain TRO1, indicating that this product can beformed by a single bacterial species. This pseudomonad bac-terium was thus assumed to play a significant role in NPEdegradation in the activated sludge. A carboxylated metabo-lite, NP2EC, was also detected as a degradation product. Thecatabolites caused by degradation of the EO chain of TritonN-101 were not identified. However, because Pseudomonas sp.
strain TROt assimilated glycolate and glyoxalate, which are
thought to be products formed by aerobic PEG degradation(15), EO chain degradation by this bacterium is likely to
proceed via the oxidative pathway, as previously proposed (15,28).
Like Pseudomonas sp. strain 14-1, strain TRO1 also degradedAEs but not PEGs. Some PEG-degrading microorganismshave displayed an ability to also assimilate AEs (6, 11, 16, 32,
33) but not APEs (4, 32). Studies on AE biodegradation haverevealed that it is initiated mainly by an attack on the methylend of the alkyl chain or by central fission which splits the AEinto the alkyl and EO chains (31). This mechanism is notapplicable to APE biodegradation, in which the EO chain ispreferentially degraded in a stepwise manner. Although APE-degrading Pseudomonas strains have also been found to de-grade AEs, the mechanism has not been studied. Furthermore,in both proposed mechanisms for AE biodegradation, freePEG which APE-degrading bacteria are not able to assimilateis released. Therefore, PEG-degrading and APE-degradingmicroorganisms are postulated to have different metabolicsystems with respect to the EO chain, although the targetmolecule is same. In other words, APE-degrading bacteriapossess fastidious EO chain assimilability compared with PEG-degrading ones, since they do not degrade the EO chain unlessit is linked to a bulky hydrophobic group such as alcohol oralkylphenol; i.e., they do degrade certain compounds as long asthey are shaped as polyoxyethylene-type nonionic surfactants.The Triton N-101-metabolizing system of Pseudomonas sp.
strain TROl was shown in an oxygen consumption assay to beinduced not only by the substrate itself but also by SDS andABS. However, the degree of induction by the last twocompounds was not comparable to that by Triton N-101 itselfor by AE. This may be due to the existence of sulfate orsulfonate in SDS or ABS, which is quite different in molecularcharacter and size from the EO chain. Also, in this assay PEGsshowed no effect as inducers. The metabolic system of the EOchain of Triton N-101 was inferred to be induced not by theEO chain itself, which is the degradation target, but by itshydrophobic part.
Again, in the case of PEG- or AE-degrading bacteria, it doesnot appear to matter whether they belong to the nonionicsurfactant or structural molecule degraders, because they showdual assimilability; i.e., they metabolize both AEs and PEGsthat are not surfactants. Judging from present data, Pseudo-monas sp. strain TROl can be regarded as an unambiguousnonionic surfactant degrader.
ACKNOWLEDGMENTS
We thank Y. Yamada (Osaka University) for providing kindly theprotocol for methylation. We are also indebted to T. Kitamura, S.Yanagida (Osaka University), J. Akino, and R. Yamaoka (KyotoInstitute of Technology) for the use of and advice on GC-MS.
REFERENCES1. Ahel, M., T. Conrad, and W. Giger. 1987. Persistent organic
chemicals in sewage effluents. 3. Determinations of nonylphenoxycarboxylic acids by high-resolution gas chromatography/mass spec-trometry and high-performance liquid chromatography. Environ.Sci. Technol. 21:697-703.
2. Ahel, M., and W. Giger. 1985. Determination of alkylphenols andalkylphenol mono- and diethoxylates in environmental samples byhigh-performance liquid chromatography. Anal. Chem. 57:1577-1583.
3. Ahel, M., J. McEvoy, and W. Giger. 1993. Bioaccumulation of thelipophilic metabolites of nonionic surfactants in freshwater organ-isms. Environ. Pollut. 79:243-248.
4. Cook, K A. 1978. A rapid method for the detection of non-ionicsurfactant-degrading micro-organisms. J. Appl. Bacteriol. 44:299-303.
5. Cripps, R. E. 1973. The microbial metabolism of thiophen-2-carboxylate. Biochem. J. 134:353-366.
6. Dwyer, D. F., and J. M. Tiedje. 1986. Metabolism of polyethyleneglycol by two anaerobic bacteria, Desulfovibrio desulfuricans and aBacteroides sp. Appl. Environ. Microbiol. 52:852-856.
7. Ekelund, R., A. Bergman, A. Granmo, and M. Berggren. 1990.Bioaccumulation of 4-nonylphenol in marine animals-a re-eval-uation. Environ. Pollut. 64:107-120.
2270 MAKI ET AL.
on August 19, 2018 by guest
http://aem.asm
.org/D
ownloaded from

DEGRADATION OF ALKYLPHENOL ETHOXYLATES 2271
8. Fincher, E. L., and W. L. Payne. 1962. Bacterial utilization of etherglycols. AppI. Microbiol. 10:542-547.
9. Giger, W., P. H. Brunner, and C. Schaffner. 1984. 4-Nonylphenolin sewage sludge: accumulation of toxic metabolites from nonionicsurfactants. Science 225:623-625.
10. Giger, W., E. Stephanou, and C. Schaffner. 1981. Persistentorganic chemicals in sewage effluents. I. Identifications of nonyl-phenols and nonylphenolethoxylates by glass capillary gas chro-matography/mass spectrometry. Chemosphere 10: 1253-1263.
11. Grant, M. A., and W. J. Payne. 1983. Anaerobic growth ofAlcaligenesfaecalis var. denitrificans at the expense of ether glycolsand nonionic detergents. Biotechnol. Bioeng. 25:627-630.
12. Greff, R. A., E. A. Setzkorn, and W. D. Leslie. 1965. A colorimetricmethod for the determination of parts/million of nonionic surfac-tants. J. Am. Oil Chem. Soc. 42:180-185.
13. Jenkins, L. D. L., K. A. Cook, and R. B. Cain. 1979. Microbialdegradation of polyethylene glycols. J. Appl. Bacteriol. 47:75-85.
14. Jimenez, L., A. Breen, N. Thomas, T. W. Federle, and G. S. Sayler.1991. Mineralization of linear alkylbenzene sulfonate by a four-member aerobic bacterial consortium. Appl. Environ. Microbiol.57:1566-1569.
15. Kawai, F. 1985. Existence of ether bond-cleaving enzyme in apolyethylene glycol-utilizing symbiotic mixed culture. FEMS Mi-crobiol. Lett. 30:273-276.
16. Kawai, F., M. Fukaya, Y. Tani, and K. Ogata. 1977. Identificationof polyethylene glycols (PEGs)-assimilable bacteria and culturecharacteristics of PEG 6000 degradation by a mixed culture. J.Ferment. Technol. 55:429-435.
17. Kravetz, L. 1981. Biodegradation of nonionic ethoxylatcs. J. Am.Oil Chem. Soc. 58:58A-65A.
18. Kravetz, L., K. F. Guin, W. T. Shebs, L. S. Smith, and H. Stupel.1982. Ultimate biodegradation of an alcohol ethoxylate and anonylphenol ethoxylate under realistic conditions. Soap Cosmet.Chem. Spec. 58:34-42, 102B.
19. Maeda, S. 1991. Degradation of TOC remained in dyeing waste-water by nonionic surfactant-degrading bacteria. Jpn. J. WaterTreat. Biol. 27:117-121.
20. Maeda, S., and I. Mikami. 1988. Degradation of the non-ionicsurfactant (polyoxyethylene-p-nonylphenyl ether) by Pseudomonasspecies. J. Water Waste 30:1056-1063.
20a.Negi, T., and K. Sugiyama. 1979. Japanese patent S54-36674.
21. Patoczka, J., and G. W. Pulliam. 1990. Biodegradation andsecondary effluent toxicity of ethoxylated surfactants. Water Res.24:965-972.
22. Pearce, B. A., and M. T. Heydeman. 1980. Metabolism of di(eth-ylene glycol) [2-(2'-hydroxyethoxy)ethanol] and other short poly-(ethylene glycol)s by gram-negative bacteria. J. Gen. Microbiol.118:21-27.
23. Reinhard, M., N. Goodman, and K. E. Mortelmans. 1982. Occur-rence of brominated alkylphenol polyethoxy carboxylates in mu-tagenic wastewater concentrates. Environ. Sci. Technol. 16:351-362.
24. Rudling, L. 1972. Swedish Water and Air Pollution Laboratoryreport B134. Swedish Water and Air Pollution Laboratory, Stock-holm.
25. Schoberl, P., and K. J. Bock. 1980. Tensidabbau und dessenMetaboliten. Tenside Deterg. 17:262-266.
26. Sheldon, L. S., and R. A. Hites. 1978. Organic compounds in theDelaware River. Environ. Sci. Technol. 12:1188-1194.
27. Sigoillot, J. C., and M. H. Nguyen. 1992. Complete oxidation ofalkylbenzene sulfonate by bacterial communities selected fromcoastal seawater. Appl. Environ. Microbiol. 58:1308-1312.
28. Steber, J., and P. Wierich. 1985. Metabolites and biodegradationpathways of fatty alcohol ethoxylates in microbial biocenoses ofsewage treatment plants. Appl. Environ. Microbiol. 49:530-537.
29. Strass, A., and B. Schink. 1986. Fermentation of polyethyleneglycol via acetaldehyde in Pelobacter venetianus. Appl. Microbiol.Biotechnol. 25:37-42.
30. Sturm, R. N. 1973. Biodegradability of nonionic surfactants:screening test for predicting rate and ultimate biodegradation. J.Am. Oil Chem. Soc. 50:159-167.
31. Swisher, R. D. 1987. Surfactant biodegradation, 2nd ed. MarcelDekker, Inc., New York.
32. Thelu, J., L. Medina, and J. Pelmont. 1980. Oxidation of poloxy-ethylene oligomers by an inducible enzyme from Pseuidomonas P400. FEMS Microbiol. Lett. 8:187-190.
33. Wagener, S., and B. Schink. 1988. Fermentative degradation ofnonionic surfactants and polyethylene glycol by enrichment cul-tures and by pure cultures of homoacetogenic and propionate-forming bacteria. Appl. Environ. Microbiol. 54:561-565.
34. Yoshimura, K. 1986. Biodegradation and fish toxicity of nonionicsurfactants. J. Am. Oil Chem. Soc. 63:1590-1596.
VOL. 60, 1994
on August 19, 2018 by guest
http://aem.asm
.org/D
ownloaded from





























