Embed Size (px)
Citation preview

1819
Braz J Med Biol Res 37(12) 2004
Molecular functions and putative interactions of desmin
Desmin: molecular interactions andputative functions of the muscleintermediate filament protein
Laboratório de Diferenciação Muscular e Citoesqueleto,Departamento de Histologia e Embriologia, Instituto de Ciências Biomédicas,Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brasil
M.L. Costa, R. Escaleira,A. Cataldo, F. Oliveiraand C.S. Mermelstein
Abstract
Desmin is the intermediate filament (IF) protein occurring exclusivelyin muscle and endothelial cells. There are other IF proteins in musclesuch as nestin, peripherin, and vimentin, besides the ubiquitous lamins,but they are not unique to muscle. Desmin was purified in 1977, thedesmin gene was characterized in 1989, and knock-out animals weregenerated in 1996. Several isoforms have been described. Desmin IFsare present throughout smooth, cardiac and skeletal muscle cells, butcan be more concentrated in some particular structures, such as densebodies, around the nuclei, around the Z-line or in costameres. Desminis up-regulated in muscle-derived cellular adaptations, including con-ductive fibers in the heart, electric organs, some myopathies, andexperimental treatments with drugs that induce muscle degeneration,like phorbol esters. Many molecules have been reported to associatewith desmin, such as other IF proteins (including members of themembrane dystroglycan complex), nebulin, the actin and tubulinbinding protein plectin, the molecular motor dynein, the gene regula-tory protein MyoD, DNA, the chaperone αB-crystallin, and proteasessuch as calpain and caspase. Desmin has an important medical role,since it is used as a marker of tumors’ origin. More recently, severalmyopathies have been described, with accumulation of desmin depos-its. Yet, after almost 30 years since its identification, the function ofdesmin is still unclear. Suggested functions include myofibrillogenesis,mechanical support for the muscle, mitochondrial localization, geneexpression regulation, and intracellular signaling. This review focuseson the biochemical interactions of desmin, with a discussion of itsputative functions.
CorrespondenceM.L. Costa
Laboratório de Diferenciação
Muscular e Citoesqueleto
Departamento de Histologia e
Embriologia, ICB, UFRJ
21949-590 Rio de Janeiro, RJ
Brasil
Fax: +55-21-2562-6480
E-mail: [email protected]
Presented at the XII Brazilian
Congress of Cellular Biology,
Campinas, SP, Brazil, July 15-18,
2004.
Research supported by CNPq, FAPERJ,
PRONEX, and Fundação Universitária
José Bonifácio (FUJB).
Received July 16, 2004
Accepted September 14, 2004
Key words• Desmin• Intermediate filaments• Muscle• Myogenesis• Myopathy
Introduction
Desmin is the intermediate filament (IF)protein of muscle and endothelial cells. Thecytoskeletal category of IFs was suggestedby Holtzer’s group (1) based on the 10-nmdiameter filaments observed in electron mi-
crographs of skeletal muscle, which wereintermediate in diameter between the 5-nmmicrofilaments and the 16-nm thick filaments.Muscle IF protein was purified by Small andSobieszek (2) who called it “skeletin”, butLazarides and Hubbard (3), who also puri-fied it, named it “desmin” (desmos = link)
Brazilian Journal of Medical and Biological Research (2004) 37: 1819-1830ISSN 0100-879X Review
1820
Braz J Med Biol Res 37(12) 2004
M.L. Costa et al.
for its alleged function of linking sarco-meres. Biochemical purification led to anti-body production, which in turn led to thedetermination of its spatial distribution inseveral tissues (4). The gene has been identi-fied by Capetanaki et al. (5), and desminknockouts have been produced by the groupsof Paulin (6) and Capetanaki (7). Desminhas been shown to interact with several mol-ecules, and therefore particular functions fordesmin are attributed to each of these inter-actions. Many aspects of the cell biology ofdesmin are remarkable, like its resistance tochemical denaturation and mechanical sta-bility, and its direct interaction with the mus-cular differentiation program. It is the firstmuscle-specific structural protein and one ofthe more persistent in several differentiationconditions. The recent finding of severalcases of human diseases linked to altereddesmin expression (8) has renewed the inter-est in the physiological roles of desmin.However, despite all the curiosity it inspired,the function of desmin is still unknown.
Biochemical aspects
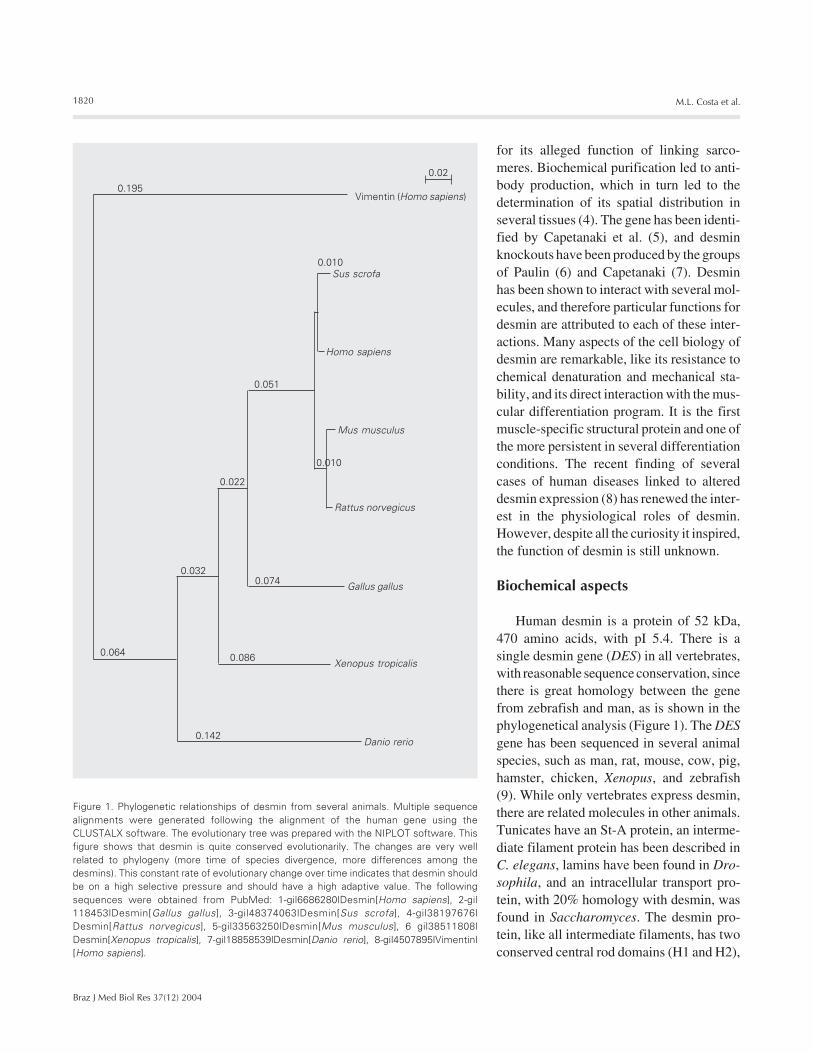
Human desmin is a protein of 52 kDa,470 amino acids, with pI 5.4. There is asingle desmin gene (DES) in all vertebrates,with reasonable sequence conservation, sincethere is great homology between the genefrom zebrafish and man, as is shown in thephylogenetical analysis (Figure 1). The DESgene has been sequenced in several animalspecies, such as man, rat, mouse, cow, pig,hamster, chicken, Xenopus, and zebrafish(9). While only vertebrates express desmin,there are related molecules in other animals.Tunicates have an St-A protein, an interme-diate filament protein has been described inC. elegans, lamins have been found in Dro-sophila, and an intracellular transport pro-tein, with 20% homology with desmin, wasfound in Saccharomyces. The desmin pro-tein, like all intermediate filaments, has twoconserved central rod domains (H1 and H2),
Figure 1. Phylogenetic relationships of desmin from several animals. Multiple sequencealignments were generated following the alignment of the human gene using theCLUSTALX software. The evolutionary tree was prepared with the NIPLOT software. Thisfigure shows that desmin is quite conserved evolutionarily. The changes are very wellrelated to phylogeny (more time of species divergence, more differences among thedesmins). This constant rate of evolutionary change over time indicates that desmin shouldbe on a high selective pressure and should have a high adaptive value. The followingsequences were obtained from PubMed: 1-gi|6686280|Desmin[Homo sapiens], 2-gi|118453|Desmin[Gallus gallus], 3-gi|48374063|Desmin[Sus scrofa], 4-gi|38197676|Desmin[Rattus norvegicus], 5-gi|33563250|Desmin[Mus musculus], 6 gi|38511808|Desmin[Xenopus tropicalis], 7-gi|18858539|Desmin[Danio rerio], 8-gi|4507895|Vimentin|[Homo sapiens].
0.02
Vimentin (Homo sapiens)0.195
0.010Sus scrofa
Homo sapiens
Mus musculus
0.010
Rattus norvegicus
Gallus gallus
Xenopus tropicalis
Danio rerio0.142
0.086
0.0740.032
0.022
0.051
0.064

1821
Braz J Med Biol Res 37(12) 2004
Molecular functions and putative interactions of desmin
4 linker regions (L1A, L1B, L2A, and L2B),and two globular end domains. The centralrod domains are responsible for polymeriza-tion by lateral association. The exact struc-ture of the filament is still unclear, but it isknown that IF molecules associate laterallyin pairs, which in turn associate with eachother. The 10-nm filament may be formed byabout 8 individual protofilaments (10). Inthis way, a truncation of the desmin carboxy-terminal domain in a few molecules leads tothe disassembly of the filament network,when a truncated desmin gene is expressedin a few nuclei in a multinuclear myotube(11).
The central IF domain is composed ofseveral heptapeptide repeats, which formcoiled-coil α-helices. This domain is presentin several protein families from spectrin toplectin. Desmin has also a less conserved IFhead domain, and the end domains are over-all quite variable among IFs. Sequence ho-mology, and therefore evolutionary relation-ships, and the ability to co-polymerize arethe main criteria for their classification into6-7 groups (10, but see Ref. 12). Thus, desminis classified as a type III IF, and shares highhomology with mesenchymal cell vimentinand glial cell GFAP (glial fibrillary acidicprotein). All type III IFs can form homopoly-mers with themselves or heteropolymers with
each other. The acid and basic cytokeratinsof epithelial cells are classified as types I andII, respectively, and they are obligatory het-eropolymers between the two classes. Thethree different neurofilament proteins com-pose the type IV IF. Type V is defined by thelamins, which are ubiquitous supporters ofnuclei in all eukaryotic cell types, and arethought to be the evolutionary more primi-tive IF protein. Lamins have a nuclear local-ization signal and an extra domain, whichinduces the formation of a lattice instead ofthe conventional filaments. While some clas-sifications propose class VI to be composedof large molecular weight proteins (synemin,paranemin), others suggest type VI to be aspecial group for lens-beaded filaments(phakinin and filensin) or as an orphan groupof unclassified proteins. Several other IFproteins, like peripherin, gelfitin, transitin,tanabin, gliarin, macrolin, etc., have beendescribed, in an ever-growing list. A total of65 IF genes have been detected in a screen-ing of a draft version of the human genome(13).
Desmin is usually phosphorylated, andthe number of isoforms correlates with phy-logeny: there is a single isoform in mamma-lian muscle, 2 in an avian muscle and 3 infish muscle (Figure 2). As is the case for allIFs, phosphorylation regulates desmin poly-
Figure 2. Two-dimensional elec-trophoresis of an extract of thecytoskeleton of 1-day-old cul-tured mouse heart cells. A, Pro-teins detected with silver stain-ing; B, corresponding autoradio-gram of cells incubated with 32Pfor 1 h, and C, A and B superim-posed, where the image of Bhas been outlined digitally. After1 h of incubation, only a fewproteins, particularly desmin, arephosphorylated, while other pro-teins known to be phosphorylat-able, such as actin and tubulin,do not incorporate phosphate.The proteins have been identi-fied by their relative positionsas: D = desmin, A = actin, T =tubulin, V = vimentin, and Tm =tropomyosin. Left is basic (OH-)and right is acid (H+). SDS = so-dium dodecyl sulfate. The arrowindicates the second dimension,from high to low molecularweight. For detailed conditions,see Ref. 17.

1822
Braz J Med Biol Res 37(12) 2004
M.L. Costa et al.
merization, so the number of isoforms maycorrelate with the structural state of eachtissue. Signaling molecules like protein ki-nase C can increase the phosphorylation ofdesmin and change the structure of the fila-ment network (14). Interestingly, some par-ticular physiological conditions or patho-logic situations can also increase the numberof desmin isoforms. Thus, there are 5-6desmin isoforms in Purkinje fibers, special-ized conducting cells in the heart of largemammalians (15). In people with familialcongenital heart disease, there are also 5-6isoforms (16). We could show that there arefive desmin isoforms in the electric organsof the electric eel Electrophorus electricusL. (17). Furthermore, our group showed thateach of the three different electric organs ofthe electric eel has a distinct desmin isoformpattern (18). All of these muscle derivativesalso have a higher IF to myofibril ratio thannormal muscle.
Myogenesis
Desmin is expressed since the early for-mation of skeletal, cardiac and smoothmuscle, and endothelial cells. In avian cells,desmin is the first muscle-specific cytoskel-etal protein or isoform to be expressed, after(or together with) the commitment of precur-sor cells to the muscle lineage. During em-bryonic development, the notochord secretesthe inductive factors SHH, the neural platesecretes Wnt and the lateral plate secretesBMP4, which will cause the somite cells tobecome committed to the muscle lineage.These proteins induce the expression of themuscle regulator factor Myf5, which in turnwill induce MyoD, which will induce mgn.The genes of most of the structural muscleproteins have a MyoD-binding regulationregion. Desmin has been reported to be ex-pressed in muscle together with Myf5 andbefore MyoD (19). The detection of desmininside the nucleus and the finding that desmincan form a dimer with MyoD are very intri-
guing (see below). The direct relationshipbetween desmin expression and muscle dif-ferentiation is difficult to establish, sincemuscle cells can develop without desmin(6,7), but muscle can also form in the ab-sence of MyoD or mgn (but not of MyoD andmgn). In chick, muscle-committed cells exitthe cell cycle, and the forced expression ofMyoD shows a coordinated beginning ofdesmin expression and inhibition of 5-bromo-2'-deoxyuridine labeling (20). In mice, repli-cating muscle cells express desmin (21). Itshould be pointed that those two modelshave quite different in vitro behavior, be-cause mammalian cells replicate more easilyand form small myotubes in culture, whileavian cells stop replicating and form enor-mous and vigorously contracting myotubes.While there are several mammalian musclecell lines, no well-differentiated avian musclelines have been established (the main avianmuscle line QM7 does not form myotubes).
The subcellular distribution of desminchanges during development from thick fila-ments widespread throughout the cell to adiffuse association with the Z-line and stria-tion. It should be noted that the mesenchy-mal precursors of muscle cells already havea vimentin IF network. Accordingly, thischange in distribution is a marker of myo-genesis, which we used to characterize theextent of MyoD conversion in transfectednon-muscle chick cultured cells (20) and topoint out the particularities of the myogen-esis in zebrafish embryos (22).
Cellular distribution
The best way to visualize the distributionof desmin in different cell types is by immu-nofluorescence (Figure 3). Desmin appearssometimes as long isolated filaments, but italso appears as a fluorescence haze, prob-ably because of its sub-resolution size (10nm, smaller than the 0.2-µm light micro-scope limit).
Desmin has been described in almost

1823
Braz J Med Biol Res 37(12) 2004
Molecular functions and putative interactions of desmin
every distinct muscular cytoskeletal struc-ture (Figure 4). In mononucleated smoothmuscle cells, desmin is spread throughoutthe cell, but is concentrated in the densebodies (Figure 4, a), where the microfila-ments also attach. Dense bodies are centers
of contraction, and have α-actinin and actinbeside desmin, being homologues to Z-linesand intercalated disks (23). Desmin can makeup to 2% of the smooth muscle mass, andchicken gizzard is commonly used for desminextraction.
Figure 3. Desmin distribution in several cell types detected by immunofluorescence. One-day-old cultured skeletalmuscle chick cell, stained for desmin (A), myosin (B) and DNA (4',6-diamidino-2-phenylindole dihydrochloride) (C).At this developmental stage, myoblasts are beginning to elongate, desmin is distributed throughout the cyto-plasm, and myosin appears first around the nucleus. Five-day-old cultured skeletal muscle chick myoblast stainedfor desmin (D), Z-line titin (E) and DNA (F). This myoblast is already striated for desmin and titin. Desmin is presenton the Z-line, but not as restricted as titin. There is also desmin around the nuclei and diffusely in the cytoplasm.Twenty-four-hour zebrafish somites stained for desmin (G), α-actinin (H) and differential interference contrast (I).These 2-hour-old myocytes in the fish embryo show a pattern similar to the 5-day chicken culture, with striationsvisible by desmin and α-actinin stain and by differential interference contrast. Three-day-old cultured heart chickcell stained for desmin (J), α-actinin (K) and DNA (L). In cardiac myocytes, desmin distribution on the Z-line is morerestricted than in skeletal muscle, but there is also more desmin in the cytoplasm. The myofibrils are not alignedas in skeletal muscle, as can be seen by the α-actinin stain. Scale bar: 10 µm for all panels. For details, see Refs.22 and 57.

1824
Braz J Med Biol Res 37(12) 2004
M.L. Costa et al.
In the multinucleated striated skeletalmuscle, again there is desmin all over thecell, but there is more desmin in the Z-line(lateral attachments of microfilaments; Fig-ures 3D,G,J and 4, b) and in the costameres(periodic structures in the membrane that areconnected to the Z-lines, Figure 4, c).Costameres are composed of several proteinclusters and link IFs and actin microfila-ments to the membrane (24). One type ofcostamere, the dystroglycan protein com-plex (DPC), connects actin through dystro-phin to the transmembrane ß-dystroglycan,which in turn binds to extracellular α-dys-troglycan, laminin 2 and collagen. Anothertype of protein complex connects actinthrough spectrin, talin, paxillin, and vinculinto trans-membrane integrins, which bind tofibronectin, laminin and collagen. Desminconnects to DPC in the costameres, throughplectin and dysferlin. The DPC alterationscause Duchenne and Becker muscular dys-trophies, frequent congenital diseases thatcan be fatal. In skeletal muscle, desmin hasbeen described in myotendinous junctions,together with nestin (Figure 4, d) (25), and in
neuromuscular junctions, associated with thecholinergic receptors (Figure 4, e) (26). Myo-tendinous junctions are specialized regionsof skeletal muscle attachment to connectivetissue, and have characteristic molecules,such as type XII and XIII collagens and α7-integrin. The neuromuscular junctions me-diate the contraction stimulus through chem-ical signaling between acetylcholine releas-ing nerve cells and the cholinergic receptorsin the synapse, a special membrane regionwith clefts (invaginations) and particular con-stituents, like type XVIII collagen and agrin.Desmin may also be present around the M-line, particularly associated with skelemin(see below). Desmin also connects myo-fibrils to the costameres, forming an extra-myofibrillar cytoskeleton (Figure 4, f).
Heart cells are mono- or binucleated andhave desmosomes, where the IFs of neigh-bor cells are connected through cadherinsand cytokeratin (Figure 4, g). Desmosomesare typical of strongly attached epithelialcells, and are composed of desmoplakin ag-glomerates that bind transmembrane cadher-ins in adjacent cells (27). In cardiac cells,desmosomes bind myocytes laterally, whilethe myofibrils of each cell end in a special Z-line that attaches to adjacent cells, the inter-calated disk (Figure 4, h). Therefore, desminand α-actinin are present in all the contrac-tion organizing centers in the three musclecell types. The distribution of both proteinsis mostly coincident, but clearly is not re-stricted to the same places. In non-musclecells, α-actinin is distributed along actin fila-ments and links them to the membrane, andin striated muscle, α-actinin aggregates onthe membrane could be nucleating centersfor myofibrillogenesis. Heart cells have thehighest proportion of desmin and also a largenumber of mitochondria.
Desmin, like all IFs, is also concentratedaround the nucleus (Figure 4, i) in the threemuscle types. In muscle cell fractionationexperiments, desmin has been found to re-main bound to the nuclear fraction (data not
Figure 4. Location of desmin in smooth, skeletal and cardiac muscle types. a, Densebodies; b, sarcomeres; c, costameres; d, myotendinous junction; e, neuromuscular junc-tion; f, extra-myofibrillar cytoskeleton; g, desmosomes; h, intercalated disk; i, perinuclearposition; j, mitochondria.

1825
Braz J Med Biol Res 37(12) 2004
Molecular functions and putative interactions of desmin
shown). Furthermore, there is a direct inter-action between cytoplasmic IFs and thenuclear lamins (28). Interestingly, the musclenuclear envelope has a particular lamin-as-sociated protein, emerin, which was identi-fied as missing in people with Emery-Dreifussmuscular dystrophy (29). It is possible tospeculate that the muscle nuclei should havea particular structure to withstand the me-chanical load of contraction, in the sameway as the desmin network could be unique.Mutations in the lamin A gene in humans arealso responsible for muscular dystrophy, andknock-out for lamin A induces the musculardystrophy phenotype (30). Because lamin Ais present in several cell types and onlymuscles are affected by its absence, musclenuclei could be subjected to a greater me-chanical stress than other cell types.
Mitochondria are somehow connected to
desmin IFs in addition to microtubules (Fig-ure 4, j).
Molecular interactions
Desmin interacts with several molecules(Table 1). First of all, it forms heteropolymerswith IF proteins of the same class, like vi-mentin, GFAP, nestin, etc., which may co-exist in some cell types, and with type VI IFproteins. The other IF proteins that may co-polymerize with desmin in vitro will not beconsidered in this review. While vimentin isco-expressed in vivo, it is down-regulatedalong myogenesis, and almost not present inadult tissues. The exact amount of vimentinprotein present in adult tissues is still un-clear, but specific silencer regulatory se-quences have been identified in the gene(31). GFAP can polymerize with desmin in
Table 1. Macromolecules that interact with desmin.
Refs. Distribution Also links to
Vimentin 28 At the periphery of Z-discs Type III or VI IFs
GFAP 32 In myofibroblasts Type III or VI IFs
Nestin 12, 33 In myotendinous junctions Type III or VI IFs
Synemin 34-36, 40 At the periphery of the Z-line and in Type III or VI IFs. α-actinin,
the sarcolemma of costameres vinculin, plectin
Paranemin 37, 38 At the periphery of the Z-line and in the sarcolemma Type III or VI IFs, plectin
and myotendinous and neuromuscular junctions
Desmuslin 39 At the periphery of the Z-line and in the sarcolemma Dystrophin, α1-syntrophin, sarcoglycans,
of costameres α-dystrobrevin, plectin
Syncoilin 41 In costameres and in neuromuscular junctions α-dystrobrevin
Lamins 28 In the nucleus αB-crystallin, emerin, nesprin, actin, DNA
Nebulin 43 On the Z-line Actin, titin, α-actinin, myosin
Skelemin 44 In the M-disc Integrin
Spectrin, ankyrin 42 At the periphery of the Z-line and in myotendinous and Ankyrin, spectrin
neuromuscular junctions
Plectin 45 At the periphery of the Z-line and in myotendinous and Myosin II, integrin ß-4, actin, spectrin, MAP,
neuromuscular junctions type III or VI IFs
Dynein 46 Along microtubules Tubulin
MyoD 47 In the nucleus Id, Mef
DNA 48 In the nucleus Histones, polymerases
αB-crystallin 49 On the Z-line Caspases, lamins
Calpain, caspase 50, 51 Desmin as substrate during apoptosis Lamins, αB-crystallin
IF = intermediate filament; GFAP = glial fibrillary acidic protein; MAP = microtubule-associated protein; Mef = muscle enhancer factor.

1826
Braz J Med Biol Res 37(12) 2004
M.L. Costa et al.
vitro, but only a few cell types actually co-express both proteins at the same time. Oneinteresting case is the dynamic and develop-mentally plastic myofibroblast, which weshowed to be positive for desmin, GFAP andvimentin (32). Nestin is typical of centralnervous system stem cells, but is also presentwith desmin in muscle ligaments (33). Sincethere is a relationship between the expres-sion of nestin and the aggressiveness of tu-mors, nestin has been an important marker incancer research. Synemin, initially purifiedas a desmin-associated molecule (34) butnow known to be a legitimate type III or VIintermediate filament, is present in striatedmuscle. Synemin may function to directlylink muscle cell intermediate filaments toboth myofibrillar Z-lines and costameres,since it has binding sites for desmin, α-actinin and vinculin (35). Curiously, syneminis expressed in all muscle tissues (althoughat higher levels in skeletal muscle), but alsoin other tissues, particularly in myoepithelialcells and glial cells (36). Paranemin was alsoinitially described as an IF-associated pro-tein (37), but later was shown to be a type VIlarge IF protein. Paranemin, but not syneminor nestin, has been shown to be essential forthe formation of an extended desmin IF net-work (38). Desmuslin is an IF protein re-cently characterized from the dystrophincomplex that associates with α-dystrobrevinin costameres and with desmin in costameresand around the Z-line (39). But sequencehomology suggests that desmuslin may bethe human homologue of synemin, morespecifically of the smaller synemin ß tran-script (40). Syncoilin is another dystrophin-associated protein that seems to concentratein neuromuscular junctions (41). Interest-ingly, nestin, desmuslin and syncoilin arenot type III filaments: while nestin may beclassified as type IV, desmuslin has beenproposed to be a type VI or VII, and syncoilincould be a type IV, since it can form fila-ments by itself. Desmin may associate withlamins, as mentioned before.
Besides IF proteins, desmin also binds toother sarcomeric and sub-membrane pro-teins. The extra-myofibrillar desmin cyto-skeleton links to the membrane adhesioncomplexes indirectly through syncoilin anddesmuslin/synemin, but there is a direct as-sociation between desmin and the proteinsspectrin and ankyrin (42). A direct interac-tion between desmin and nebulin was shownusing a yeast two-hybrid approach (43).Nebulin is a molecular ruler that controls thesize of the A-band through its binding toactin. Desmin binds to skelemin around theM-line (44). The proposed structure ofskelemin has internal myosin-binding regionsand two desmin-binding regions, one in eachextremity. In this way, desmin binds both tothe Z- and M-lines.
The three major filaments of the cyto-skeleton are linked by the protein plectin,which has been shown to bind to microtu-bules, microfilaments and IFs (45). Plectin isa member of the plakin family of cytolinkers,which include the desmosomal desmoplakin,and which also bind to integrins and cadher-ins. The distribution of plectin is not affectedin desmin knock-out mice, but in plectinknock-out mice the amounts of vinculin andspectrin are reduced. IFs seem to be trans-ported throughout the cell associated withmicrotubules and/or microfilaments via cy-toplasmic dynein (46).
Desmin can be present in the nucleus,where it has been suggested to bind to MyoDto form a heterodimeric gene regulation pro-tein (47). The interaction of desmin withMyoD establishes a new self-regulatory path-way, because the MyoD gene has E-boxes,regulatory sequences, MyoD binding sites,as does the desmin gene. Inside the nucleus,desmin can bind directly to DNA. The inter-action of type III IFs (vimentin in particular)with several DNA arrangements has beenwell characterized, and all type III filamentshave the same DNA-binding domain (48).
The desmin IF network is dependent onbinding to the small chaperone αB-crystallin

1827
Braz J Med Biol Res 37(12) 2004
Molecular functions and putative interactions of desmin
(49). First identified as a structural protein inthe eye lens, αB-crystallin is known to existin several cell types. Other crystallin iso-forms also have dual enzymatic and struc-tural roles.
The proteases calpain and caspase caninteract with desmin during differentiationand particularly in apoptosis. Caspase 6 phos-phorylates specific residues in desmin, in-duced by change in serum concentration thatcontrols differentiation (50). Desmin has beendescribed as a major substrate for proteoly-sis by a muscle-specific calpain (51).
Desminopathies
Since the IF proteins are cell type specif-ic, they have been used to type the origin ofundifferentiated cells in tumors or other cellsof unknown origin, like electrocytes (17).
The first identified involvement of desminin human diseases was a familial cardiomyop-athy that had filament deposits (15). The ad-vances in molecular diagnosis led to the iden-tification of desmin-related myopathies (52).Their symptoms include skeletal muscle weak-ness, arrhythmias, heart failure, and accumu-lation of desmin in cardiac and skeletal musclecells. The desmin accumulation can be causedby desmin mutations or mutations in αB-crys-tallin. Several mutations from different fami-lies have been characterized. This type ofdisease has been called surplus myopathy be-cause it arises not from the absence of theprotein but from its accumulation due tomisaggregation caused by the inability to poly-merize properly or by the lack of chaperones(αB-crystallin), or by its overexpression causedby problems in recycling proteins (caspase,calpain). Experimentally, a small overex-pression of desmin does not lead do anydamage, but a significant overexpressionleads to accumulation of desmin and theformation of aggregates inside the cell, typi-cal of human desminopathy patients. It isstill unclear if the accumulation is the causeor the consequence of the disease, which can
affect health to varying extents.
Putative functions
Many functions have been attributed todesmin since its identification. Some can beclassified as mechanical or structural, with arole being attributed to the filament itself,but some hypotheses propose that desminhas regulatory roles.
Desmin IFs have been proposed to berequired for myofibrillogenesis. Several linesof evidence argue against this hypothesis,including the fact that desmin knock-outmice go through their myogenesis basicallyunaffected (6,7) and that cells without the IFnetwork caused by transfection with trun-cated desmin also assemble their myofibrilsperfectly (11). By contrast, electroporationof desmin antibodies disrupts the Z-line andthe myofibril (53).
Desmin could be involved in the regula-tion of gene expression. In this case, themain argument is the ability of desmin tobind DNA and its cellular distribution fromthe cell periphery into the nucleus, where itcan interact with lamins (28,54). The nuclearlamins, once thought to be involved only inthe mechanical support of the nuclear mem-brane, now are also suspected to have regu-latory functions because of their associa-tions with DNA transcription proteins (55).
Desmin function could be the localiza-tion of mitochondria. Desmin knock-out micehave their mitochondria displaced, and thesealterations could lead to a deficiency in oxy-gen metabolism (56). It is noteworthy thatthese changes in mitochondrial distributionhappen before other disturbances.
Desmin, like most IFs, could support andposition the nucleus, together with microtu-bules. After the myoblasts fuse in skeletalmuscle, there is an important realignment ofthe nuclei. Desmin knock-out mice have mis-placed nuclei (6,7). We could show in primarychick myogenic cultures that, upon treatmentwith the phorbol ester 12-O-tetradecanoyl

1828
Braz J Med Biol Res 37(12) 2004
M.L. Costa et al.
phorbol 13-acetate, desmin is phosphorylated,the myofibrils disassemble but the IF networkpersists, and the nuclei are clustered togetherin the center (57). Our recent results show thatcholesterol withdrawal with the drug methyl-beta-cyclodextrin also induces nucleus mis-alignment (58).
Desmin could be responsible for the elas-tic properties of muscle, like resistance tostretch. This assumption originated beforethe identification of the elastic protein titin,which binds the Z- to the M-line. Further-more, desmin knock-out muscles show thesame passive elastic properties as controlmuscles (59).
While the function of desmin is still un-certain, the same is not true for other IFs andtheir proteins in other cell types. Thus, it isclear that cytokeratins have a structural rolein epithelia because there is a skin blisterphenotype both in the cytokeratin knock-outmouse and in the equivalent human disease,epidermolysis bullosa (60). On the otherhand, vimentin knock-out mice do not showany greatly altered phenotype (61) and thereare no known vimentin-related diseases. Onepossibility is that IFs perform different func-tions in specific tissues, and that some ofthem have structural roles while others haveregulatory roles. If this is true, the trendinitiated by Holtzer (1) of integrating all theintermediate filaments and their proteins in asingle cytoskeletal entity should be reversedby an approach where the differences, morethan the similarities, of each filament shouldbe emphasized.
Microfilaments and microtubules have amonotone basic structure of conserved globu-lar molecules in all cells, but they performseveral different functions because of theirgreatly varied associated proteins. IFs have adual structure, with a central conserved do-main and varied terminal domains. We couldimagine that the conserved domain is analo-gous to the basic actin and tubulin blocks,while the varied domains would function togive their cell type specific functions. Not only
may the IFs in each cell type have differentfunctions, but they could perform specific func-tions even in different places in the cell. Thereare several actin-containing compartments inone cell, each with different properties, asthere are different microtubule populations. Itis possible, for instance, that the sub-sarcolem-mal desmin has a somewhat different functionthan the nuclear desmin. But in the case ofmicrofilaments and microtubules, the associ-ated proteins are different in each sub-cellularstructure. Therefore, we can imagine that eachdesmin structure is different, maybe becauseof different associated molecules (such as nestinin myotendinous junctions) or even because offilament arrangement or isoform composition.
If there are many “sub-functions” ofdesmin, it is reasonable to assume that desminmay have (some) different roles in skeletal,cardiac and smooth muscle. Muscles are anextreme adaptation of a ubiquitous cytoskel-eton for force production, and the existenceof three main muscle types indicates quitedifferent adaptive solutions, consistent withrestricted distributions of some desmin-in-teracting molecules to each particular muscletype. However, the overall association ofdesmin with the muscular program is clear-cut, because all vertebrate muscles expressdesmin from their differentiation to all pos-sible adaptations.
The mechanical and physiological func-tions of microfilaments and microtubulesare not exclusive. Focal contacts are sites ofstrong adhesion to the substrate and of sig-naling through kinases. Likewise, desmincould have concomitant structural and regu-latory roles. In this way, it will be impossibleto look for a single function for desmin, andall the mentioned functions could be true.
Acknowledgments
We would like to thank Profs. RadovanBorojevic, Cristiano Coutinho (UFRJ) andHernandes Carvalho (UNICAMP) for helpand encouragement.

1829
Braz J Med Biol Res 37(12) 2004
Molecular functions and putative interactions of desmin
References
1. Ishikawa H, Bischoff R & Holtzer H (1968). Mitosis and intermedi-ate-sized filaments in developing skeletal muscle. Journal of CellBiology, 38: 538-555.
2. Small JV & Sobieszek A (1977). Studies on the function and compo-sition of the 10-NM(100-A) filaments of vertebrate smooth muscle.Journal of Cell Science, 23: 243-268.
3. Lazarides E & Hubbard BD (1976). Immunological characterizationof the subunit of the 100 A filaments from muscle cells. Proceed-ings of the National Academy of Sciences, USA, 73: 4344-4348.
4. Lazarides E (1980). Intermediate filaments as mechanical integra-tors of cellular space. Nature, 283: 249-256.
5. Capetanaki YG, Ngai J & Lazarides E (1984). Characterization andregulation in the expression of a gene coding for the intermediatefilament protein desmin. Proceedings of the National Academy ofSciences, USA, 81: 6909-6913.
6. Li Z, Mericskay M, Agbulut O, Butler-Borwne G, Carlsson L, ThornellL-E, Babinet C & Paulin D (1997). Desmin is essential for the tensilestrength and integrity of myofibrils but not for myogenic commit-ment, differentiation, and fusion of skeletal muscle. Journal of CellBiology, 139: 1-16.
7. Milner DJ, Waltzer G, Tran D, Bradley A & Capetanaki Y (1996).Disruption of muscle architecture and myocardial degeneration inmice lacking desmin. Journal of Cell Biology, 134: 1255-1270.
8. Carlsson L & Thornell LE (2001). Desmin-related myopathies inmice and man. Acta Physiologica Scandinavica, 171: 341-348.
9. Strelkov SV, Herrmann H & Aebi U (2003) Molecular architecture ofintermediate filaments. Bioassays, 25: 243-251.
10. Herrmann H & Aebi U (2000). Intermediate filaments and theirassociates: multi-talented structural elements specifying cytoarchi-tecture and cytodynamics. Current Opinion in Cell Biology, 12: 79-90.
11. Schultheiss T, Lin ZX, Ishikawa H, Zamir I, Stoeckert CJ & Holtzer H(1991). Desmin/vimentin intermediate filaments are dispensable formany aspects of myogenesis. Journal of Cell Biology, 114: 953-966.
12. Steinert PM, Chou YH, Prahlad V, Parry DA, Marekov LN, Wu KC,Jang SI & Goldman RD (1999). A high molecular weight intermedi-ate filament-associated protein in BHK-21 cells is nestin, a type VIintermediate filament protein. Limited co-assembly in vitro to formheteropolymers with type III vimentin and type IV alpha-internexin.Journal of Biological Chemistry, 274: 9881-9890.
13. Hesse M, Magin TM & Weber K (2001). Genes for intermediatefilament proteins and the draft sequence of the human genome:novel keratin genes and a surprisingly high number of pseudogenesrelated to keratin genes 8 and 18. Journal of Cell Science, 114:2569-2575.
14. Huang X, Li J, Foster D, Lemanski SL, Dube DK, Zhang C &Lemanski LF (2002). Protein kinase C-mediated desmin phosphory-lation is related to myofibril disarray in cardiomyopathic hamsterheart. Experimental Biology and Medicine, 227: 1039-1046.
15. Thornell LE & Eriksson A (1981). Filament systems in the Purkinjefibers of the heart. American Journal of Physiology, 241: H291-H305.
16. Rappaport L, Contard F, Samuel JL, Delcayre C, Marotte F, Tome F& Fardeau M (1988). Storage of phosphorylated desmin in a familialmyopathy. Federation of European Biochemical Societies. Letters,231: 421-425.
17. Costa ML, Moura Neto V & Chagas C (1988). Desmin heterogeneityin the main electric organ of Electrophorus electricus. Biochimie,70: 783-789.
18. Costa ML, Mermelstein CS, Froes MM, Chagas C & Moura Neto V(1988). Differences in the isodesmin pattern between the electricorgans of Electrophorus electricus L. Comparative Biochemistryand Physiology. Part B, Biochemistry and Molecular Biology, 119:715-719.
19. Li H & Capetanaki Y (1993). Regulation of the mouse desmin gene:transactivated by MyoD, myogenin, MRF4 and Myf5. Nucleic AcidsResearch, 21: 335-343.
20. Choi J, Costa ML, Mermelstein CS, Chagas C, Holtzer S & Holtzer H(1990). MyoD converts primary dermal fibroblasts, chondroblasts,smooth muscle, and retinal pigmented epithelial cells into striatedmononucleated myoblasts and multinucleated myotubes. Proceed-ings of the National Academy of Sciences, USA, 87: 7988-7992.
21. Kaufman SJ & Foster RF (1988). Replicating myoblasts express amuscle-specific phenotype. Proceedings of the National Academyof Sciences, USA, 85: 9606-9610.
22. Costa MC, Escaleira RC, Rodrigues VB, Manasfi M & MermelsteinCS (2001). Some distinctive features of zebrafish myogenesis basedon unexpected distributions of the muscle cytoskeletal proteinsactin, myosin, desmin, α-actinin, troponin and titin. Mechanisms ofDevelopment, 116: 95-104.
23. Stromer MH & Bendayan M (1988). Arrangement of desmin inter-mediate filaments in smooth muscle cells as shown by high-resolu-tion immunocytochemistry. Cell Motility and the Cytoskeleton, 11:117-125.
24. Ervasti JM (2003). Costameres: the Achilles’ heel of Herculeanmuscle. Journal of Biological Chemistry, 278: 13591-13594.
25. Tidball JG (1992). Desmin at myotendinous junctions. ExperimentalCell Research, 199: 206-212.
26. Mitsui T, Kawajiri M, Kunishige M, Endo T, Akaike M, Aki K &Matsumoto T (2000). Functional association between nicotinic ace-tylcholine receptor and sarcomeric proteins via actin and desminfilaments. Journal of Cellular Biochemistry, 77: 584-595.
27. Tokuyasu KT, Dutton AH & Singer SJ (1983). Immunoelectron mi-croscopic studies of desmin (skeletin) localization and intermediatefilament organization in chicken cardiac muscle. Journal of CellBiology, 96: 1736-1742.
28. Cartaud A, Jasmin BJ, Changeux JP & Cartaud J (1995). Directinvolvement of a lamin-B-related (54 kDa) protein in the association ofintermediate filaments with the postsynaptic membrane of the Tor-pedo marmorata electrocyte. Journal of Cell Science, 108: 153-160.
29. Holaska JM, Wilson KL & Mansharamani M (2002). The nuclearenvelope, lamins and nuclear assembly. Current Opinion in CellBiology, 14: 357-364.
30. Sullivan T, Escalante-Alcalde D, Bhatt H, Anver M, Bhat N, Naga-shima K, Stewart CL & Burke B (1999). Loss of A-type lamin expres-sion compromises nuclear envelope integrity leading to musculardystrophy. Journal of Cell Biology, 147: 913-920.
31. Farrell FX, Sax CM & Zehner ZE (1990). A negative element involvedin vimentin gene expression. Molecular and Cellular Biology, 10:2349-2358.
32. Guma FCR, Mello TG, Mermelstein CS, Fortuna VA, Wofchuk ST,Gottfried C, Guaragna RM, Costa ML & Borojevic R (2001). Interme-diate filament modulation in an in vitro model of the hepatic stellatecell activation or conversion into the lipocyte phenotype. Biochem-istry and Cell Biology, 79: 409-417.
33. Kachinsky AM, Dominov JA & Miller JB (1994). Myogenesis and theintermediate filament protein, nestin. Developmental Biology, 165:216-228.

1830
Braz J Med Biol Res 37(12) 2004
M.L. Costa et al.
34. Granger BL & Lazarides E (1980). Synemin: a new high molecularweight protein associated with desmin and vimentin filaments inmuscle. Cell, 22: 727-738.
35. Bellin RM, Huiatt TW, Critchley DR & Robson RM (2001). Syneminmay function to directly link muscle cell intermediate filaments toboth myofibrillar Z-lines and costameres. Journal of Biological Chem-istry, 276: 32330-32337.
36. Hirako Y, Yamakawa H, Tsujimura Y, Nishizawa Y, Okumura M,Usukura J, Matsumoto H, Jackson KW, Owaribe K & Ohara O(2003). Characterization of mammalian synemin, an intermediatefilament protein present in all four classes of muscle cells and someneuroglial cells: co-localization and interaction with type III interme-diate filament proteins and keratins. Cell and Tissue Research, 313:195-207.
37. Breckler J & Lazarides E (1982). Isolation of a new high molecularweight protein associated with desmin and vimentin filaments fromavian embryonic skeletal muscle. Journal of Cell Biology, 92: 795-806.
38. Schweitzer SC, Klymkowsky MW, Bellin RM, Robson RM,Capetanaki Y & Evans RM (2001). Paranemin and the organizationof desmin filament networks. Journal of Cell Science, 114: 1079-1089.
39. Mizuno Y, Thompson TG, Guyon JR, Lidov HG, Brosius M, ImamuraM, Ozawa E, Watkins SC & Kunkel LM (2001). Desmuslin, anintermediate filament protein that interacts with α-dystrobrevin anddesmin. Proceedings of the National Academy of Sciences, USA,98: 6156-6161.
40. Titeux M, Brocheriou V, Xue Z, Gao J, Pellissier JF, Guicheney P,Paulin D & Li Z (2001). Human synemin gene generates splicevariants encoding two distinct intermediate filament proteins. Euro-pean Journal of Biochemistry, 268: 6435-6449.
41. Newey SE, Howman EV, Ponting CP, Benson MA, Nawrotzki R, LohNY, Davies KE & Blake DJ (2001). Syncoilin, a novel member of theintermediate filament superfamily that interacts with α-dystrobre-vin in skeletal muscle. Journal of Biological Chemistry, 276: 6645-6655.
42. Larsen TH, Dalen H, Sommer JR, Boyle R & Lieberman M (1999).Membrane skeleton in cultured chick cardiac myocytes revealed byhigh resolution immunocytochemistry. Histochemistry and Cell Bi-ology, 112: 307-316.
43. Bang ML, Gregorio C & Labeit S (2002). Molecular dissection of theinteraction of desmin with the C-terminal region of nebulin. Journalof Structural Biology, 137: 119-127.
44. Price MG (1987). Skelemins: cytoskeletal proteins located at theperiphery of M-discs in mammalian striated muscle. Journal of CellBiology, 104: 1325-1336.
45. Wiche G (1998). Role of plectin in cytoskeleton organization anddynamics. Journal of Cell Science, 111: 2477-2486.
46. Helfand BT, Mikami A, Vallee RB & Goldman RD (2002). A require-ment for cytoplasmic dynein and dynactin in intermediate filament
network assembly and organization. Journal of Cell Biology, 157:795-806.
47. Li H, Choudhary SK, Milner DJ, Munir MI, Kuisk IR & Capetanaki Y(1994). Inhibition of desmin expression blocks myoblast fusion andinterferes with the myogenic regulators MyoD and myogenin. Jour-nal of Cell Biology, 124: 827-841.
48. Wang Q, Tolstonog GV, Shoeman R & Traub P (2001). Sites ofnucleic acid binding in type I-IV intermediate filament subunit pro-teins. Biochemistry, 40: 10342-10349.
49. Horwitz J (2003). Alpha-crystallin. Experimental Eye Research, 76:145-153.
50. Chen F, Chang R, Trivedi M, Capetanaki Y & Cryns VL (2003).Caspase proteolysis of desmin produces a dominant-negative inhib-itor of intermediate filaments and promotes apoptosis. Journal ofBiological Chemistry, 278: 6848-6853.
51. Goll DE, Thompson VF, Li H, Wei W & Cong J (2003). The calpainsystem. Physiological Reviews, 83: 731-801.
52. Goldfarb LG, Vicart P, Goebel HH & Dalakas MC (2004). Desminmyopathy. Brain, 127: 723-734.
53. Wang SM, Huang YS, Wu JC & Tseng YZ (2000). Role of desminfilaments in chicken cardiac myofibrillogenesis. Journal of CellularBiochemistry, 77: 635-644.
54. Tolstonog GV, Sabasch M & Traub P (2002). Cytoplasmic intermedi-ate filaments are stably associated with nuclear matrices and po-tentially modulate their DNA-binding function. DNA and Cell Biol-ogy, 21: 213-239.
55. Hutchison CJ (2002). Lamins: building blocks or regulators of geneexpression? Nature Reviews. Molecular Cell Biology, 3: 848-858.
56. Milner DJ, Mavroidis M, Weisleder N & Capetanaki Y (2000). Desmincytoskeleton linked to muscle mitochondrial distribution and respi-ratory function. Journal of Cell Biology, 150: 1283-1298.
57. Mermelstein CS, Costa ML, Chagas Filho C & Moura Neto V (1996).Intermediate filament proteins in TPA-treated skeletal muscle cellsin culture. Journal of Muscle Research and Cell Motility, 17: 199-206.
58. Mermelstein CS, Portilho DM, Medeiros RB, Matos AR, Einicker-Lamas M, Tortelote GG, Vieyra A & Costa ML (2004). Cholesteroldepletion by methyl-b-cyclodextrin enhances myoblast fusion andinduces the formation of myotubes with disorganized nuclei. Celland Tissue Research (in press).
59. Shah SB, Davis J, Weisleder N, Kostavassili I, McCulloch AD, RalstonE, Capetanaki Y & Lieber RL (2004). Structural and functional roles ofdesmin in mouse skeletal muscle during passive deformation. Bio-physical Journal, 86: 2993-3008.
60. Fuchs E (1996). The cytoskeleton and disease: genetic disorders ofintermediate filaments. Annual Review of Genetics, 30: 197-231.
61. Colucci-Guyon E, Portier MM, Dunia I, Paulin D, Pournin S & BabinetC (1994). Mice lacking vimentin develop and reproduce without anobvious phenotype. Cell, 79: 679-694.





























