Embed Size (px)
Citation preview

J Huazhong Univ Sci Technol[Med Sci] 30(6):2010
734
Discovery of Chrysoeriol, a PI3K-AKT-mTOR Pathway Inhibitor with Potent Antitumor Activity against Human Multiple Myeloma Cells in vitro*
Yang YANG (杨 漾), Xiaoxi ZHOU (周晓曦), Min XIAO (肖 敏), Zhenya HONG (洪振亚), Quan GONG (龚 泉),
Lijun JIANG (姜立军), Jianfeng ZHOU (周剑峰)#
Department of Hematology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, China
© Huazhong University of Science and Technology and Springer-Verlag Berlin Heidelberg 2010
Summary: This study was designed to determine the impact of chrysoeriol on proliferation and cell cy-cle progression in the human multiple myeloma cell lines RPMI 8226 and KM3, and its related molecu-lar mechanisms. Chryseoriol was identified by using the phosphorylated AKT-specific cytoblot high throughput assay. CCK-8 assay was employed to examine the growth inhibition rate and IC50 (48 h) in peripheral blood mononuclear cells (PBMNCs), RPMI 8226 and KM3 cells treated with chrysoeriol at various concentrations. Cells were labeled with 5-6-carboxyfluorescein diacetate succinimidyl ester (CFSE), and the proliferation dynamics was detected by flow cytometry and analyzed with ModFit software. The cell cycles of RPMI 8226 and KM3 cells were measured by flow cytometry when the IC50 concentration of chrysoeriol was adopted. The alterations in cell-cycle related proteins (Cyclin B1, Cy-clin D1, p21) and proteins in PI3K-AKT-mTOR pathway were determined by Western blot analysis. The results showed the proliferation of multiple myeloma cells was significantly inhibited by chrysoeriol, resulting in cell cycle arrest in G2/M phase. Chrysoeriol could significantly reduce the expression of p-AKT (s473) and p-4eBP1 (t37/46) protein, meanwhile enhanced Cyclin B1 and p21 protein expres-sion. Similar effects were not observed in PBMNCs from normal donors. It was concluded that chryso-eriol was a selective PI3K-AKT-mTOR pathway inhibitor. It restrained the proliferation of human mul-tiple myeloma cells, but didn’t affect proliferation of PBMNCs from normal donors. It might exhibit the cell cycle regulatory effect via the inhibition of PI3K-AKT-mTOR signal pathway. Key words: chrysoeriol; multiple myeloma; proliferation inhibition; G2/M arrest; PI3K-AKT-mTOR signal pathway
PI3K-AKT-mTOR signal pathway plays an impor-tant role in cell growth regulation, proliferation and me-tabolism in response to environmental cues[1, 2]. We have extended these studies by developing phosphorylated AKT-specific cytoblot high throughput assay to screen additional small-molecule inhibitors of this signal path-way. Chrysoeriol is identified from a collection of about 2000 monomeric compounds as a potent inhibitor of AKT in many kinds of cell lines. Chrysoeriol is a major flavanoid extracted from Rooibos tea (Aspalathus lin-earis)[3]. It has been shown to have various functions and bioactivities such as anti-inflammatory, anti-obesity and anti-mutagenic properties[4–8]. Studies on antitumor ac-tivity of chrysoeriol were only demonstrated in breast cancer[9], and there is no report about the influence of this compound on hematological neoplasms. We have Yang YANG, E-mail: [email protected] #Corresponding author, E-mail: [email protected] *This project was supported by grants from the National Natu-ral Sciences Foundation of China (No. 30770914; No. 30901587) and China State Key Basic Research Program (No. 2002CB513100).
demonstrated that the inhibitory effects of chrysoeriol on cell proliferation varied among blood cell types in screening assays. The proliferation inhibition in multiple myeloma cells was remarkable, but no effect was shown in peripheral blood mononuclear cells (PBMNCs) from normal donors, even though their p-AKT activities were both suppressed. Multiple myeloma is an incurable he-matological disease caused by malignant transformation of plasma cells, whose pathogenesis is still unknown[10]. The aim of this study was to examine the effect of chry-soeriol on in vitro proliferation of multiple myeloma cell lines RPMI 8226 and KM3 and the possible mechanism.
1 MATERIALS AND METHODS 1.1 Reagents
Chrysoeriol was purchased from Chromadex (USA), dissolved with DMSO, diluted to a concentration of 20 mmol/L and stored at 4°C. CCK-8 kit was purchased from Dojindo (Japan). Propidium iodide (PI) was ob-tained from Sigma–Aldrich (USA). 5-6-carboxyflu- orescein diacetate succinimidyl ester (CFSE) was pur-chased from Introvigen (USA). BCA kit and ECL detec-
30(6):734-740,2010J Huazhong Univ Sci Technol[Med Sci] DOI 10.1007/s11596-010-0649-4

J Huazhong Univ Sci Technol[Med Sci] 30(6):2010
735
tion system were from Pierce (USA). Antibodies against AKT, p-AKT (s473), p-4eBP1 (t37/46) and Cyclin D1 were from Cell Signaling Technology, Inc. (USA). The antibody against p21 was from Boster Co. (China). The antibody against Cyclin B1 was from BioLegend (USA). The antibody against β-actin was from Santa Cruz (USA). Horseradish peroxidase-conjugated goat anti-rabbit IgG was from Jackson ImmunoResearch (USA). Fi-coll-Hypaque solution was from TBD (China). Erythro-cyte lysing solution was from Tiangen (China). 1.2 Cell Lines and Cell Culture
RPMI 8226 was obtained from the American Type Culture Collection (USA), and KM3 was subcultured by our laboratory. Cells were grown in RPMI 1640 medium (Gibco, USA) containing 10% fetal calf serum in a hu-midified atmosphere (37°C, 5% CO2), and seeded at logarithmic growth phase every 1 to 2 days. 1.3 Phosphorylated AKT-specific Cytoblot High Throughput Assays
Cytoblot high throughput assays for phospho-AKT inhibition were performed as previously described[11]. In brief, BEAS-2B cells were plated into 96-well tissue culture plates at a density of 25000 cells/well. After overnight growth at 37°C, the cells were treated for 5 h in the presence of compounds (8 concentration gradients, 3 times between the adjacent concentrations, 4 alignment holes) offered by NCPC New Drug Research Center (Shijiazhuang, China). Controls were set as follows: holes without adding HRP-secondary antibody (blank); holes treated with DMSO (negative control); holes treated with 100 nmol/L of rapamycin (agonist control); holes treated with 700 nmol/L PI-103 (inhibitor control). After treatment, cells were washed in 100 μL of cold TBS and then fixed for 1 h at 4°C with cold 3.7% for-maldehyde in TBS (190 μL/well). Membranes were permeabilized during 5-min incubation in ice-cold methanol at 4°C. Cells were blocked for 30 min with 3% milk in TBS (180 μL/well) and then shaken overnight at 4°C with 3% milk in TBS (50 μL/well) containing a 1:1000 dilution of anti-phospho-AKT (s473) antibody. Antibodies were aspirated, then plates were washed twice with TBS (180 μL/well), and the cells were incu-bated with horseradish peroxidase-conjugated goat anti-rabbit IgG (dilution 1:2000) for 30 min at room tem-perature. The above washing process was repeated. Re-sults were visualized by adding Western blot chemilu-minescence reagent directly to the wells of the plates and incubating cells at room temperature for 5 min. Quanti-fication of results was done using a GS-700 scanning densitometer (Bio-Rad Laboratories, USA). 1.4 Isolation and Culture of PBMNCs
PBMNCs were isolated from 5 mL of heparinized venous blood of healthy donors by using a Fi-coll-Hypaque gradient, followed by erythrocyte lysis if necessary. Then cells were washed and cultured in IMDM medium supplemented with 10% FBS and 5 μg/mL PHA at a suitable density. 1.5 Cell Growth Inhibition Rate Assay
Cells at logarithmic growth phase were seeded in 96-well plates at a density of 1×105/mL in RPMI 1640 medium with different concentrations of chrysoeriol (200 μL/well, 8 alignment holes). Blank holes, negative holes
and solvent holes were set at the same time. After ap-proximately 48 h, 20 μL/well cell counting kit (CCK-8) was added in the dark and cells were cultured for another 2 h. After shaking gently, the absorbance (A) at 450 nm was measured by using a microplate reader. The drug concentration-cell growth inhibition rate (GIR) curve was established, on which X-axis represented the dose of the drug, and Y-axis represented the growth inhibition rate of cells. GIR was calculated using the following equation: GIR=1 - (Mean A450 of experimental groups/Mean A450 of negative control groups). 1.6 CFSE Cell Proliferation Analysis by Flow Cy-tometry
Cells were suspended in RPMI 1640 medium (without FBS) and diluted to l×107/mL, then 5 μL of CFSE stock solution was added to 1 mL of cell solution (CFSE final concentration was 5 µmoL/L). The cell so-lution was then incubated for 15 min at 37°C, shaken gently every 5 min, suspended by adding equivoluminal FBS and washed twice with PBS afterward. The cells were resuspended in complete RPMI 1640 medium to a density of l×106/mL. These processes were performed in dark. Then the stained cells were seeded in uncoated 24-well plates at a density of 1×106/mL in RPMI 1640 medium with 1 mL/hole. RPMI 8226 and KM3 cells were treated with chrysoeriol at concentrations of 2.5 and 5 µmol/L, respectively, and cultured for 96 h. The cells in negative control group were cultured for 96 h without chryseoriol. Meanwhile, optimal amount of stained cells were fixed at 4°C in dark with cold 4% paraformalde-hyde, as original parent cells. Ninety-six h later, the staining was assayed by FACS analysis in FL1 channel for green fluorescent intensity, and the data were proc-essed with Modfit software. The proliferation index was defined as the sum of the cells in all generations divided by the calculated number of original parent cells. This was useful for comparing the quantity of all cell divi-sions among cultures of the same kinds of cells under-going different treatments. 1.7 Cell Cycle Analysis by Flow Cytometry
Multiple myeloma cells in experimental groups (24 h, 48 h) and control group were harvested and washed once with ice-cold phosphate buffered saline (PBS). Af-ter that, cells were resuspended in 1 mL of 70% (w/v) ice-cold ethanol and fixed overnight at -20°C. Fixed cells were washed once with ice-cold PBS and resus-pended in 500 μL ice-cold PBS, and then 10 µL of 10 mg/mL RNase A and 50 µL of 500 μg/mL PI were added to the cells. After an incubation period of 30 min at room temperature in the dark, the cells were analyzed by flow cytometry. Similar results were observed in three inde-pendent experiments. 1.8 Protein Expression Analysis by Western Blot
Cells were harvested and lysed with M-PERTM ly-sis buffer and protein concentrations were determined using the BCA Protein Assay Kit. A total of 50 μg of each protein sample was separated by sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS- PAGE). After electrophoresis, protein was transferred to PVDF membranes, blocked in 5% non-fat dry milk for 1 h at 37°C, and subsequently washed and incubated over-night at 4°C with the corresponding primary antibodies

J Huazhong Univ Sci Technol[Med Sci] 30(6):2010
736
(1:1000). After washing with TBS-T, membranes were incubated with a peroxidase-conjugated secondary anti-body for 1 h at 37°C. Protein expression levels were de-tected by chemoluminescence using the enhanced chemiluminescence (ECL) detection system. 1.9 Statistical Analysis
The SPSS 12.0 statistical software program was used for statistical comparison. Data were presented as ±s. Student’s t test was used to determine the statistical
significance of treatment results against control group. The comparison among multiple groups was performed according to one-way ANOVA. A P<0.05 was consid-ered to be statistically significant.
2 RESULTS 2.1 Development of Phosphorylated AKT-specific Cytoblot High Throughput Assay and Identification of Chrysoeriol
Adenovirus-40 transformed, immortalized human bronchial epithelial cells, strain BEAS-2B, have been shown to highly express p-AKT protein[12], so we took BEAS-2B as model cells, and developed a 96-well plate high-throughput cytoblot assay in which the levels of phosphorylated AKT in the whole cells were detected (as described in “Materials and Methods”). We evaluated a 1893 monomeric compound library from NCPC New Drug Research Center to identify the compounds that can inhibit the activity of p-AKT in BEAS-2B, and the re-sults indicated that several compounds including could inhibit activation of p-AKT to various degrees, and chry-soeriol was one of them (CAS identifier: 497-71-4, mo-lecular weight: 300.26). Fig. 1A showed the structure of chrysoeriol, which belongs to flavanoid. Fig. 1B showed the effects of chrysoeriol on p-AKT levels. Different concentrations of chrysoeriol at holes on the 96-well plate reduced p-AKT to barely detectable levels, which were obviously lower than those in negative control group and approached the level of inhibitor control group. The curve of chrysoeriol concentration against relative luminescence values in BEAS-2B cells (fig. 1C) showed that the reduced relative luminescence value of BEAS-2B accompanied the increased concentrations of chrysoeriol. Chrysoeriol inhibited the p-AKT relative luminescence value of BEAS-2B with an IC50 of 20 µmol/L. Furthermore, we analyzed the expression of p-AKT (s473) protein in BEAS-2B cells after treated with different compounds by Western blot. Because ra-pamycin, an inhibitor of mTOR complex 1, could acti-vate p-AKT (s473) by negative regulatory loop mecha-nism, we took rapamycin as agonist control, and screened agonist by cytoblot high throughput assay. Meanwhile, for the reason that PI-103 is an inhibitor of PI3K, which is upstream protein of AKT[13], we took PI-103 as inhibitor control. Fig. 1D showed that treat-ment with rapamycin resulted in an increase in p-AKT levels in BEAS-2B cells. In contrast, PI-103 reduced p-AKT to barely detectable levels; treatment with chry-soeriol had analogous effect on p-AKT, and showed a dose-dependent depression effect.
To determine whether chrysoeriol suppresses p-AKT activity in other human cells, we evaluated sev-
eral human normal cells such as HEK293, ECV-304, hPBMNCs, several human solid tumor cell lines such as Hela, MAD-MB-231, MCF-7, OV2008 and several hu-man hematological tumor cell lines such as HL-60, Ka-sumi-1, Raji, Jurkat, RPMI 8226, KM3, SKM-1. The results showed that chrysoeriol suppressed p-AKT activ-ity in these cells to varying degrees (data not shown). Meanwhile, we examined the effects of chrysoeriol in proliferation of these cells. Interestingly, while the p-AKT level was suppressed obviously by chrysoeriol, there was no significant change in the proliferation of human normal cells. In contrast, human cancer cells ap-peared much more sensitively to the presence of chryso-eriol, especially multiple myeloma RPMI 8226 and KM3 cells. 2.2 Effects of Chrysoeriol on the Expression of p-AKT and Proliferation in hPBMNCs
To study how human multiple myeloma RPMI 8226 and KM3 cells were affected by chrysoeriol, we investi-gated whether human normal hematological cells were able to endure this compound. As shown in fig. 2A, treatment of PBMNCs with 30 µmol/L chrysoeriol led p-AKT to barely detectable levels, and it showed a good dose-dependent property, having no effect on total AKT level. Proliferation curve of PBMNCs treated with chry-soeriol (fig. 2B) suggested that this kind of cells held a high tolerance against chrysoeriol. At the dose of 30 µmol/L, chrysoeriol inhibited the growth of PBMNCs by less than 10%, with an IC50 of 175 µmol/L which was much higher than the concentration for inhibiting p-AKT. 2.3 Chrysoeriol Inhibited Proliferation of MM Cells
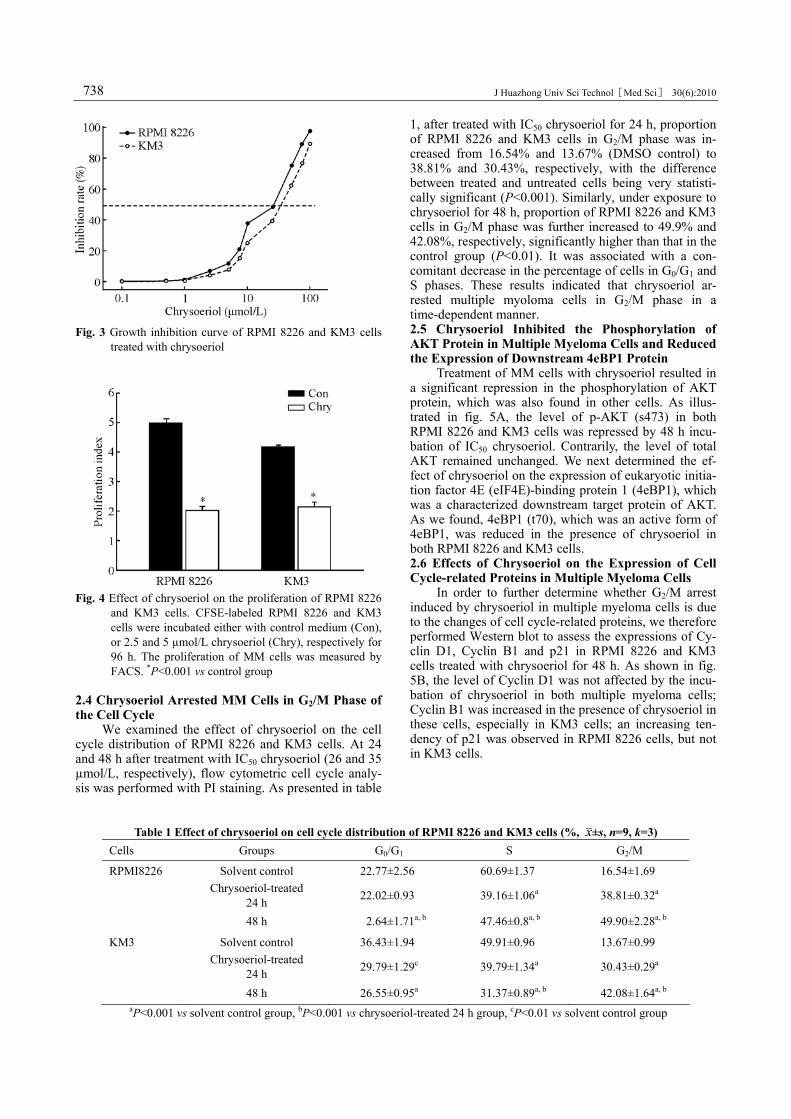
Furthermore, we showed the data which reflected the effects of chrysoeriol on multiple myeloma cells. CCK-8 assay showed that after treatment with chryso-eriol for 48 h, A450 of RPMI 8226 cells in 2.5 µmol/L group was (0.626±0.028), which was significantly dif-ferent from that in the control group (0.672±0.009) (P<0.001). In KM3 cells, the concentrations which yielded statistical significance were 5 µmol/L and more. DMSO which was set as solvent control had no inhibi-tory effect on the proliferation of MM cells.
Next, from the proliferation-inhibitory curve of chrysoeriol on RPMI 8226 and KM3 cells (fig. 3), we assessed that chrysoeriol displayed a significant growth-inhibitory effect on the multiple myeloma cells in a concentration-dependent manner. The IC50 values (in µmol/L at 48 h) of RPMI 8226 and KM3 cells were 26 and 35, respectively.
From another standpoint, RPMI 8226 and KM3 cells were stained with CFSE and cultured with 2.5 and 5 µmol/L chrysoeriol for 96 h, respectively.After incuba-tion, the staining was assayed by FACS. FACS analysis revealed that treatment of these cells with chrysoeriol resulted in significant repressed proliferation as com-pared with untreated cells, and this tendency was con-cretely marked by significantly reduced mean fluorescent intensity of untreated cells and higher green fluorescent intensity of treated cells. Taking proliferation index (PI) as parameter, PI values in both multiple myeloma cells treated with chrysoeriol were significantly lower than in untreated cells (P<0.001; fig. 4).

J Huazhong Univ Sci Technol[Med Sci] 30(6):2010
737
Fig. 1 Identification of chrysoeriol using a phosphorylated AKT-specific cytoblot high throughput assay
A: Chemical structure of chrysoeriol; B: Schematic diagram of whole-cell immunodetection assay using chemiluminescent de-tection (cytoblot). There were 5 groups: blank group, negative control group (Con), inhibitor control group (PI-103), agonist control group (Rap) and experimental groups (BEAS-2B cells treated with chrysoeriol at 0.032 and 0.011 mmol/L, respectively, Chry1 and Chry2) (four alignment holes for each group); C: Obtained relative luminescence values of BEAS-2B cells treated with chrysoeriol of different concentrations were normalized to values from corresponding DMSO-treated cells. IC50=20 µmol/L; D: Chrysoeriol suppressed phosphorylated AKT levels of BEAS-2B cells. Lane 1: Cells treated with DMSO as nega-tive control; lanes 2 and 3: Cells treated with 100 nmol/L Rap and 700 nmol/L PI-103, respectively; Lanes 4—6: Cells treated with chrysoeriol of 20, 30 and 50 μmol/L, respectively.
Fig. 2 Effects of chrysoeriol on phosphorylated AKT levels and proliferation of PBMNCs from healthy donors
A: Effect of Chrysoeriol on p-AKT (s473) and AKT protein levels; B: the curve of inhibition rate of PBMNCs against chryso-eriol. IC50 concentration of PBMNCs was about 175 µmol/L.
J Huazhong Univ Sci Technol[Med Sci] 30(6):2010
738
Fig. 3 Growth inhibition curve of RPMI 8226 and KM3 cells
treated with chrysoeriol
Fig. 4 Effect of chrysoeriol on the proliferation of RPMI 8226
and KM3 cells. CFSE-labeled RPMI 8226 and KM3 cells were incubated either with control medium (Con), or 2.5 and 5 µmol/L chrysoeriol (Chry), respectively for 96 h. The proliferation of MM cells was measured by FACS. *P<0.001 vs control group
2.4 Chrysoeriol Arrested MM Cells in G2/M Phase of the Cell Cycle
We examined the effect of chrysoeriol on the cell cycle distribution of RPMI 8226 and KM3 cells. At 24 and 48 h after treatment with IC50 chrysoeriol (26 and 35 µmol/L, respectively), flow cytometric cell cycle analy-sis was performed with PI staining. As presented in table
1, after treated with IC50 chrysoeriol for 24 h, proportion of RPMI 8226 and KM3 cells in G2/M phase was in-creased from 16.54% and 13.67% (DMSO control) to 38.81% and 30.43%, respectively, with the difference between treated and untreated cells being very statisti-cally significant (P<0.001). Similarly, under exposure to chrysoeriol for 48 h, proportion of RPMI 8226 and KM3 cells in G2/M phase was further increased to 49.9% and 42.08%, respectively, significantly higher than that in the control group (P<0.01). It was associated with a con-comitant decrease in the percentage of cells in G0/G1 and S phases. These results indicated that chrysoeriol ar-rested multiple myoloma cells in G2/M phase in a time-dependent manner. 2.5 Chrysoeriol Inhibited the Phosphorylation of AKT Protein in Multiple Myeloma Cells and Reduced the Expression of Downstream 4eBP1 Protein
Treatment of MM cells with chrysoeriol resulted in a significant repression in the phosphorylation of AKT protein, which was also found in other cells. As illus-trated in fig. 5A, the level of p-AKT (s473) in both RPMI 8226 and KM3 cells was repressed by 48 h incu-bation of IC50 chrysoeriol. Contrarily, the level of total AKT remained unchanged. We next determined the ef-fect of chrysoeriol on the expression of eukaryotic initia-tion factor 4E (eIF4E)-binding protein 1 (4eBP1), which was a characterized downstream target protein of AKT. As we found, 4eBP1 (t70), which was an active form of 4eBP1, was reduced in the presence of chrysoeriol in both RPMI 8226 and KM3 cells. 2.6 Effects of Chrysoeriol on the Expression of Cell Cycle-related Proteins in Multiple Myeloma Cells
In order to further determine whether G2/M arrest induced by chrysoeriol in multiple myeloma cells is due to the changes of cell cycle-related proteins, we therefore performed Western blot to assess the expressions of Cy-clin D1, Cyclin B1 and p21 in RPMI 8226 and KM3 cells treated with chrysoeriol for 48 h. As shown in fig. 5B, the level of Cyclin D1 was not affected by the incu-bation of chrysoeriol in both multiple myeloma cells; Cyclin B1 was increased in the presence of chrysoeriol in these cells, especially in KM3 cells; an increasing ten-dency of p21 was observed in RPMI 8226 cells, but not in KM3 cells.
Table 1 Effect of chrysoeriol on cell cycle distribution of RPMI 8226 and KM3 cells (%, ±s, n=9, k=3) Cells Groups G0/G1 S G2/M
RPMI8226 Solvent control 22.77±2.56 60.69±1.37 16.54±1.69
Chrysoeriol-treated 24 h 22.02±0.93 39.16±1.06a 38.81±0.32a
48 h 2.64±1.71a, b 47.46±0.8a, b 49.90±2.28a, b
KM3 Solvent control 36.43±1.94 49.91±0.96 13.67±0.99
Chrysoeriol-treated 24 h 29.79±1.29c 39.79±1.34a 30.43±0.29a
48 h 26.55±0.95a 31.37±0.89a, b 42.08±1.64a, b aP<0.001 vs solvent control group, bP<0.001 vs chrysoeriol-treated 24 h group, cP<0.01 vs solvent control group

J Huazhong Univ Sci Technol[Med Sci] 30(6):2010
739
Fig. 5 Effects of chrysoeriol on the expressions of some proteins in RPMI 8226 and KM3 cells
RPMI 8226 and KM3 cells were treated either with control medium, or IC50 concentration of chrysoeriol for 48 h. Protein lev-els were detected by Western blotting using antibodies against AKT, p-AKT (s473), p-4eBP1 (t37/46) (A), Cyclin B1, Cyclin D1, p21 and actin (B). Actin blotting shows equal loading.
3 DISCUSSION
PI3K-AKT-mTOR pathway plays a pivotal role in most fundamental aspects of cells, including cell growth, proliferation and metabolism[1, 2]. This signaling pathway places the normal cells in balance under the negative regulation of PTEN which is known as an anti-oncogene, contributes to the diverse cellular roles which include cell growth, proliferation, survival and apoptosis. Meanwhile, it deals with various stress status such as hypoxia and hyperthermia, adapts to environmental variations[14]. However, the equilibrium is disturbed in tumor cells, and inactivating mutations or deletions of the tumor suppressor PTEN occur, leading to constitutive abnormal pathway activation, which have been associ-ated with malignant transformation and resistance to chemotherapy[15–17]. Therefore, identification of specific inhibitors that target PI3K-AKT-mTOR pathway be-comes hot spots of targeted tumor therapy. Especially after the appearance of rapamycin which is the first mTORC1 inhibitor approved by the Food and Drug Ad-ministration (FDA), research in this field has been taken seriously more and more by people. Unfortunately, up to now, no specific pathway inhibitor has been found for a variety of reasons, for instance, the inhibition of mTORC1 by rapamycin triggers a negative feedback mechanism by activating upstream bypass, promotes the activation of p-S6K1, a molecular downstream of mTORC1, such a mechanism may attenuate the rapamy-cin effects and result in treatment failure.
In this situation, we developed an assay for phos-phorylated AKT-specific cytoblot high-throughput ac-cording to references published, evaluated nearly 2000 kinds of monomeric compounds from NCPC New Drug Research Center for their abilities to inhibit the activity of p-AKT, and obtained several compounds, e.g. chryso-eriol. Our finding is concordant with previous studies showing that chrysoeriol inhibits p-AKT in human aortic smooth muscle cells[18]. Chrysoeriol is a plant natural product identified as a bioactive flavonoid that is isolated from several plant families such as Rooibos tea and Digitalis purpurea, and it has been shown to have anti-inflammatory, anti-obesity, anti-oxidant and anti-mutagenic effects. Furthermore, in anti-tumor, chrysoeriol has been only demonstrated in breast cancer. However, little has been known about the influence of this compound on hematologic neoplasms until now.
In further study, we demonstrated that although this
compound could inhibit p-AKT activities in human nor-mal cells such as BEAS-2B, HEK293 and PBMNCs, there was no obvious proliferation inhibitory effect of chrysoeriol in these cells. In contrast, it had proliferation inhibitory abilities in many tumor cell lines, and the strongest proliferation inhibitory activity was observed in multiple myeloma RPMI 8226 and KM3 cells. The abil-ity of chrysoeriol to selectively suppress tumor cells but not normal cells suggests that a PI3K-AKT-mTOR pathway deficiency may be involved in it. Once losing constant activated AKT signal, the survivals of tumor cells are affected greatly. However, the proliferation of normal cells is not influenced by pathway inhibitor, which is due to its normal anti-oncogene and signal by-pass. Indeed, once the signal is decreased by the inhibi-tion of PI3K-AKT-mTOR pathway, the reaction of anti-oncogene subsides, then bypass signal enhances, in no time the signals in normal cells reach state of equilib-rium[19, 20]. In this report, the results of CCK-8 showed that chrysoeriol inhibits the proliferation of RPMI 8226 cells and KM3 cells at various concentrations, with 48 h IC50 concentration at 26 and 35 μmol/L respectively. The inhibition rate was increased with chrysoeriol concentra-tion. On the other hand, the results of CFSE assay dem-onstrated that PI of RPMI 8226 and KM3 cells treated with 2.5 and 5 μmol/L chrysoeriol was decreased to 2.03 and 2.15 (P<0.001) compared to 4.97 and 4.18 of control groups. These results suggest the proliferation of multi-ple myeloma cells was significantly inhibited by chryso-eriol. Flow cytometry analysis showed that the propor-tion of RPMI 8226 and KM3 cells in G2/M phase treated with chrysoeriol at an IC50 concentration was 3 times higher than the controls at 48 h. However, the effect of G2/M arrest by chrysoeriol hasn’t been identified in other tumor cell lines. These data indicates chrysoeriol might suppress the proliferation of MM cells by G2/M arrest, leading to the high sensitivity of multiple myeloma cells to chrysoeriol.
In this study, we move further to analyze the ex-pression pattern of cell cycle-related protein. G/M checkpoint, one of the cell-cycle monitoring mechanisms, can ensure the faithful replication of cells[21]. G2/M phase transition is related to cell division, including division of cytoplasm and nuclear DNA as well as transformation from one cell into the two cells. For tumor cells which grow at an exponential rate, G2/M checkpoint is an im-portant target for inhibition of cell growth. Cyclin B is produced in late S phase and G2 phase, interacts with

J Huazhong Univ Sci Technol[Med Sci] 30(6):2010
740
MPF and is degraded in M phase. It helps the transition of G2/M, making cell enter mitosis. It was reported that Cyclin B1 was enriched when cells were arrested in G/M checkpoint[22]. When RPMI 8226 cells and KM3 cells were treated with chrysoeriol at IC50 concentration, the expression of Cyclin B1 was increased as compared with controls, especially for KM3 cells. These results sug-gested that the increase of Cyclin B arrested cells in G/M checkpoint rather made cells escape the monitoring of the G/M checkpoint in multiple cells. Meanwhile, we found out that the expression of cyclin D1, which con-trols the G1/S checkpoint, didn’t change when cells were treated with chrysoeriol.
To further explore the underlying mechanism, the expression of p21 was also detected. P21 inhibits the activation of Cyclin-CDK and relates with the control of both G2 and M phases. After treatment with chrysoeriol for 48 h, the expression of p21 was up-regulated in RPMI 8226 cells but not in KM3 cells, suggesting p21 was involved in G/M arrest and inhibition of growth.
In both KM3 and RPMI 8226 cells, chrysoeriol suppressed the activity of AKT and down-regulated the phosphorylation level as well as inhibited the expression of 4eBP1, an important downstream molecule of AKT pathway. So chrysoeriol might suppress the growth of MM cells by inhibiting the PI3K-AKT-mTOR-4eBP1 pathway. High activation of PI3K-AKT-mTOR-4eBP1 signaling pathway occurs in a majority of malignant cancers, which contributes to proliferation, malignant transformation and metastasis of tumor cells[23, 24].
In conclusion, the tumor is a proliferative disease characterized by uncoordinated cell growth and it corre-lates closely with malfunction of cell-cycle control as well as abnormal activation of certain signaling pathways. Chrysoeriol significantly inhibited proliferation of MM cells and might exert its effect by regulation of cell cycle and inhibition of the PI3K-AKT-mTOR pathway. Inves-tigation on chrysoeriol’s molecular mechanism will not only lay the theoretical basis for its application in tumor therapy but also provide primary data to further explore its role as a PI3K-AKT-mTOR pathway inhibitor.
REFERENCES 1 Jacinto E, Loewith R, Schmidt A, et al. Mammalian TOR
complex 2 controls the actin cytoskeleton and is rapamy-cin insensitive. Nat Cell Biol, 2004,6(11):1122-1128
2 Sarbassov DD, Guertin DA, Ali SM, et al. Phosphoryla-tion and regulation of Akt/PKB by the rictor-mTOR com-plex. Science, 2005,307(5712):1098-1101
3 Duke JA, Bogenschutz MJ, Du CJ, et al. Handbook of Medicinal Herbs. 2nd ed. USA: CRC Press, 2002,612-613
4 Schinella GR, Giner RM, Recio MC, et al. Anti-inflammatory effects of South American Tanacetum vulgare. J Pharm Pharmacol, 1998,50(9):1069-1074
5 Han LK, Sumiyoshi M, Zheng YN, et al. Anti-obesity action of Salix matsudana leaves (Part 2). Isolation of anti-obesity effectors from polyphenol fractions of Salix matsudana. Phytother Res, 2003,17(10):1195-1198
6 Kim JH, Cho YH, Park SM, et al. Antioxidants and in-
hibitor of matrix metalloproteinase-1 expression from leaves of Zostera marina L. Arch Pharm Res, 2004,27(2):177-183
7 Choi DY, Lee JY, Kim MR, et al. Chrysoeriol potently inhibits the induction of nitric oxide synthase by blocking AP-1 activation. J Biomed Sci, 2005,12(6):949-959
8 Snijman PW, Swanevelder S, Joubert E, et al. The an-timutagenic activity of the major flavonoids of rooibos (Aspalathus linearis): some dose-response effects on mutagen activation-flavonoid interactions. Mutat Res, 2007,631(2):111-123
9 Takemura H, Uchiyama H, Ohura T, et al. A methoxyfla-vonoid, chrysoeriol, selectively inhibits the formation of a carcinogenic estrogen metabolite in MCF-7 breast cancer cells. J Steroid Biochem Mol Biol, 2010,118(1-2):70-76
10 Seidl S, Kaufmann H, Drach J, et al. New insights into the pathophysiology of multiple myeloma. Lancet Oncol, 2003,4(9):557-564
11 Yang J, Shamji A, Matchacheep S, et al. Identification of a small-molecule inhibitor of class Ia PI3Ks with cell-based screening. Chem Biol, 2007,14(4):371-377
12 Chun KH, Kosmeder JW 2nd, Sun S, et al. Effects of de-guelin on the phosphatidylinositol 3-kinase/Akt pathway and apoptosis in premalignant human bronchial epithelial cells. J Natl Cancer Inst, 2003,95(4):291-302
13 Cully M, You H, Levine AJ, et al. Beyond PTEN muta-tions: the PI3K pathway as an integrator of multiple inputs during tumorigenesis. Nat Rev Cancer, 2006,6(3):184-192
14 Hay N. The Akt-mTOR tango and its relevance to cancer. Cancer Cell, 2005,8(3):179-183
15 Grunwald V, DeGraffenried L, Russel D, et al. Inhibitors of mTOR reverse doxorubicin resistance conferred by PTEN status in prostate cancer cells. Cancer Res, 2002,62(21):6141-6145
16 Memmott RM, Dennis PA. Akt-dependent and -independent mechanisms of mTOR regulation in cancer. Cell Signal, 2009,21(5):656-664
17 Strimpakos AS, Karapanagiotou EM, Saif MW, et al. The role of mTOR in the management of solid tumors: an overview. Cancer Treat Rev, 2009,35(2):148-159
18 Cha BY, Shi WL, Yonezawa T, et al. An inhibitory effect of chrysoeriol on platelet-derived growth factor (PDGF)-induced proliferation and PDGF receptor signal-ing in human aortic smooth muscle cells. J Pharmacol Sci, 2009,110(1):105-110
19 Wendel HG, De Stanchina E, Fridman JS, et al. Survival signaling by Akt and eIF4E in oncogenesis and cancer therapy. Nature, 2004,428(6980):332-337
20 Garcia Echeverria C, Sellers WR. Drug discovery ap-proaches targeting the PI3K/Akt pathway in cancer. On-cogene, 2008,27(41):5511-5526
21 Nasmyth K. Viewpoint: putting the cell cycle in order. Science, 1996,274(5293):1643-1645
22 Cappelletti V, Fioravanti L, Miodini P, et al. Genistein blocks breast cancer cells in the G (2) M phase of the cell cycle. J Cell Biochem, 2000,79(4):594-600
23 Bjornsti MA, Houghton PJ. The TOR pathway: a target for cancer therapy. Nat Rev Cancer, 2004,4(5):335-348
24 Hoeffer CA, Klann E. mTOR signaling: at the crossroads of plasticity, memory and disease. Trends Neurosci, 2010,33(2):67-75
(Received Aug. 20, 2010)





























