Embed Size (px)
Citation preview

B R A I N R E S E A R C H 1 3 7 6 ( 2 0 1 1 ) 9 4 – 1 0 0
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Research Report
Distribution patterns of cannabinoid CB1 receptors in thehippocampus of APPswe/PS1ΔE9 double transgenic mice
Sara Kalifaa,b, Eva K. Polstona, Joanne S. Allarda, Kebreten F. Manayea,⁎aDepartment of Physiology and Biophysics, Howard University, Washington, DC, 20059, USAbDepartment of Biology, College of Science, George Mason University, Fairfax, VA, 22030, USA
A R T I C L E I N F O
⁎ Corresponding author. Fax: +1 202 806 4479.E-mail address: [email protected] (KAbbreviations: AD, Alzheimer's Disease; A
arachidonylglycerol; CB1, cannabinoid receptglial fibrillary acidic protein; PS1, presenilin 1
0006-8993/$ – see front matter © 2011 Elsevdoi:10.1016/j.brainres.2010.12.061
A B S T R A C T
Article history:Accepted 19 December 2010Available online 28 December 2010
Cannabinoids have neuroprotective effects that are exerted primarily through cannabinoidCB1 receptors in the brain. This study characterized CB1 receptor distribution in the doubletransgenic (dtg) APPswe/PS1ΔE9mousemodel for Alzheimer's disease. Immunohistochemicallabeling of CB1 protein in non-transgenic mice revealed that CB1 was highly expressed inthe hippocampus, with the greatest density of CB1 protein observed in the combinedhippocampal subregions CA2 and CA3 (CA2/3). CB1 immunoreactivity in the CA1 and CA2/3hippocampal regions was significantly decreased in the dtg APPswe/PS1ΔE9 mice compared tonon-transgenic littermates. Reduced CB1 expression in dtg APPswe/PS1ΔE9 mice wasassociated with astroglial proliferation and elevated expression of the cytokines induciblenitric oxide synthase and tumor necrosis factor alpha. This finding suggests an anti-inflammatory effect of cannabinoids that is mediated by CB1 receptor, particularly in theCA2/3 region of the hippocampus. Furthermore, the study suggests a decreased CB1 receptorexpression may result in diminished anti-inflammatory processes, exacerbating theneuropathology associated with Alzheimer's disease.
© 2011 Elsevier B.V. All rights reserved.
Keywords:Alzheimer's diseaseInflammationAstrocytesiNOSTNFα
1. Introduction
Alzheimer's disease (AD) is a devastating neurodegenerativedisease characterized by progressive memory loss, cognitivedecline and widespread loss of neurons and their synapses inthe cerebral cortex, entorhinal area, hippocampus, ventralstriatum and basal forebrain (Selkoe, 2001 and Wisniewskiand Terry, 1973). At the molecular level, the known abnormal-ities include abnormal processing of amyloid precursor protein
.F. Manaye).β, amyloid beta; APP, amor 1; CB2, cannabinoid rec; iNOS, inducible nitric o
ier B.V. All rights reserve
(APP), hyperphosphorylation of tau protein, and apoptotic-likecell death (Troncoso et al., 1996). Based on studies of transgenicmice that express mutant APP and or presenilin-1 (PS1), it ishypothesized that amyloid beta 42 (Aβ42) accumulation anddiffuse plaque formation is associated with local microglialactivation, cytokine release, reactive astrocytosis and a multi-protein inflammatory response (Eikelenboom et al, 1994;McGeer and McGeer, 1995 and Rogers et al., 1996). Generationof cytokines by glia can potentiate excitotoxicitywhich can lead
yloid precursor protein; AEA, Arachidonylethanolamide; 2-AG, 2-eptor 2; CNS, central nervous system; dtg, double transgenic; GFAP,xide synthase; TNFα, tumor necrotic factor alpha; tg, transgenic
d.

95B R A I N R E S E A R C H 1 3 7 6 ( 2 0 1 1 ) 9 4 – 1 0 0
to neuronal dysfunction and death. Therefore down-regulationof glial activation may have a favorable effect on the inductionand progression of neurodegenerative diseases. One type ofagent that may have potential in this regard is cannabinoids(Ramírez et al., 2005). Cannabinoids are a group of terpeno-phenolic compounds that are present in cannabis sativaplant (marijuana). In 1997, two receptors, CB1 and CB2, thatare responsible for mediating the effects of cannabinoids,were characterized, localized and cloned (Matsuda, 1997).Whereas CB1 receptor is most abundant in the centralnervous system (CNS), CB2 cannabinoid receptor is foundboth in peripheral immune tissues (Galiegue et al., 1995) andin CNS (Onaivi et al, 2006). The discovery of the cannabinoidreceptor and the availability of highly selective and potentcannabinoid mimetics led to the identification of a familyof lipid neurotransmitters that serve as natural ligands forthe CB1 receptor. Arachidonylethanolamide (AEA), namedanandamide from the Sanskrit word meaning ‘internal bliss’(Devane et al., 1992) and 2-arachidonylglycerol (2-AG)(Mechoulam et al., 1995; Sugiura et al., 1995) are the twoextensively characterized endocannabinoids. Both ananda-mide and 2-AG, act as true ‘endogenous cannabinoids' bybinding and functionally activating one or both cannabinoidreceptor subtypes.
Cannabinoids act as neuroprotective agents against exci-totoxicity in vitro and acute brain damage in vivo (Ramírezet al., 2005). Cannabinoids are also being investigated aspotential therapeutic agents for neurological and neurode-generative disorders (Baker et al., 2000 and Mechoulam etal., 2002). Neuroprotection by cannabinoids is thought tooccur through CB1-mediated inhibition of voltage-sensitiveCa2+ channels to reduce Ca2+ influx, glutamate release andexcitotoxicity (Shen and Thayer, 1998 and Piomelli et al.,2000). In addition, cannabinoids have been shown to haveantioxidative and anti-inflammatory effects (Hampson et al.,1998 and Marsicano et al., 2002). The endocannabinoidsystem appears to be a relevant modulator of physiologicalfunctions in the central nervous system (Di Marzo andDeutsch, 1998). However, little is known about the role of theendocannabinoid system in neurodegenerative diseasesinvolving neuroinflammation such as Alzheimer's disease.
Transgenic mouse models that express human genesassociated with familial AD, including APP and PS1, haveprovided important tools for understanding neural reactionsto the deposition of mutant Aβ proteins, and for developingnovel approaches for the therapeutic management of AD inhumans (Games et al., 1995; Hsiao et al., 1995; Malherbe et al.,1996; Hardy, 1997; Johnson-Wood et al., 1997; Sturchler-Pierratet al., 1997; Morgan et al., 2000; Wang et al., 2003). In thepresent study, we employed the double transgenic (dtg)APPswe/PS1ΔE9 mouse to investigate the relationship betweenCB1 receptor expression, neuroinflammation, and AD-likeneuropathology. Stereological approaches were used to quan-tify numbers of CB1-immunoreactive (CB1-IR) neurons andglial fibrillary acidic protein-immunoreactive (GFAP-IR) cellsin the hippocampus of middle aged dtg APPswe/PS1ΔE9 malemice and age-matched non-transgenic (non-tg) littermates.Western blot was then used to compare expression levels oftwo pro-inflammatory cytokines, iNOS and TNFα, between thedtg APPswe/PS1ΔE9 and non-tg littermate mice.
2. Results
2.1. CA2/3 contains higher numbers of CB1-immunoreactive(CB1-IR) cells than CA1 region of the hippocampus. In addition,dtg APPswe/PS1ΔE9 mice showed a significant reduction in thenumber of CB1-IR hippocampal neurons compared to non-tglittermates
Low and high magnification photomicrographs showingpatterns of CB1 labeling in the mouse hippocampus arepresented in Figs. 1(A and B). CB1 receptor was abundantlyexpressed in CA1 andCA2/3 fields of the hippocampal neuronsand, specifically, in large pyramidal cells. Stereological estima-tions of the numbers of CB1-IR cells in the CA1 and CA2/3regions of the hippocampus in middle-aged (10–12 month old)mice revealed that the numbers of CB1-IR cells were signifi-cantly higher in the CA2/3 region of the hippocampusthan in the CA1 region in both dtg and non-tg mice (p=0.001).As shown in Fig. 1C, numbers of CB1-IR neurons in dtgAPPswe/PS1ΔE9 mice were significantly lower in both the CA1(27% fewer cells) and CA2/3 (23% fewer cells) hippocampalregions compared to non-tg littermates (p=0.013).
2.2. There was a significant increase in the numbers ofGFAP-immunoreactive cells, particularly in the CA1 region, inthe dtg APPswe/PS1ΔE9 mice compared to the non-dtg group
Photomicrographs presented in Figs. 2(A and B) show GFAPimmunoreactivity in the hippocampus of dtg APPswe/PS1ΔE9mice and non-tg litter mates. GFAP-IR astrocytes were distrib-uted throughout the hippocampus of both the dtg APPswe/PS1ΔE9mice and the non-tg littermates. As shown, hippocampalGFAP-IR cells were noticeably denser in the dtg APPswe/PS1ΔE9mice than in the non-tg litter mates. Statistical analysis of thestereological results revealed a significant 90% increase inGFAPpositive cells in the CA1 hippocampal region of dtg APPswe/PS1ΔE9 animals compared to non-tg littermates (p=0.002;Fig. 2C). Regional analyses depicted in Fig. 2C showed that innon-tg control animals the distribution of GFAP positive cellswas uniform throughout the hippocampus. In contrast, numb-ers of GFAP-IR cells in the hippocampus of dtg APPswe/PS1ΔE9mice were significantly higher in the CA1 region than in CA2/3(p=0.006), with the number of GFAP-IR cell in the CA2/3region of dtg APPswe/PS1ΔE9 mice being similar to that of thenon-tg littermates.
2.3. Genotype-related increase in iNOS and TNF-α proteinexpression in dtg APPswe/PS1ΔE9 mice compared to non-dtg mice
Results from Western blot analyses of iNOS and TNF-αprotein expression levels are presented in Fig. 3. Hippocampalexpression levels of both iNOS (Fig. 3A, p≤0.001) and TNF-α(Fig. 3B, p≤0.001) were significantly elevated in the dtgAPPswe/PS1ΔE9mice compared to non-tg age-matched controls.
3. Discussion
Cannabinoid receptors are abundant in discrete areas of thebrain. In the hippocampus, a region that plays a crucial role in
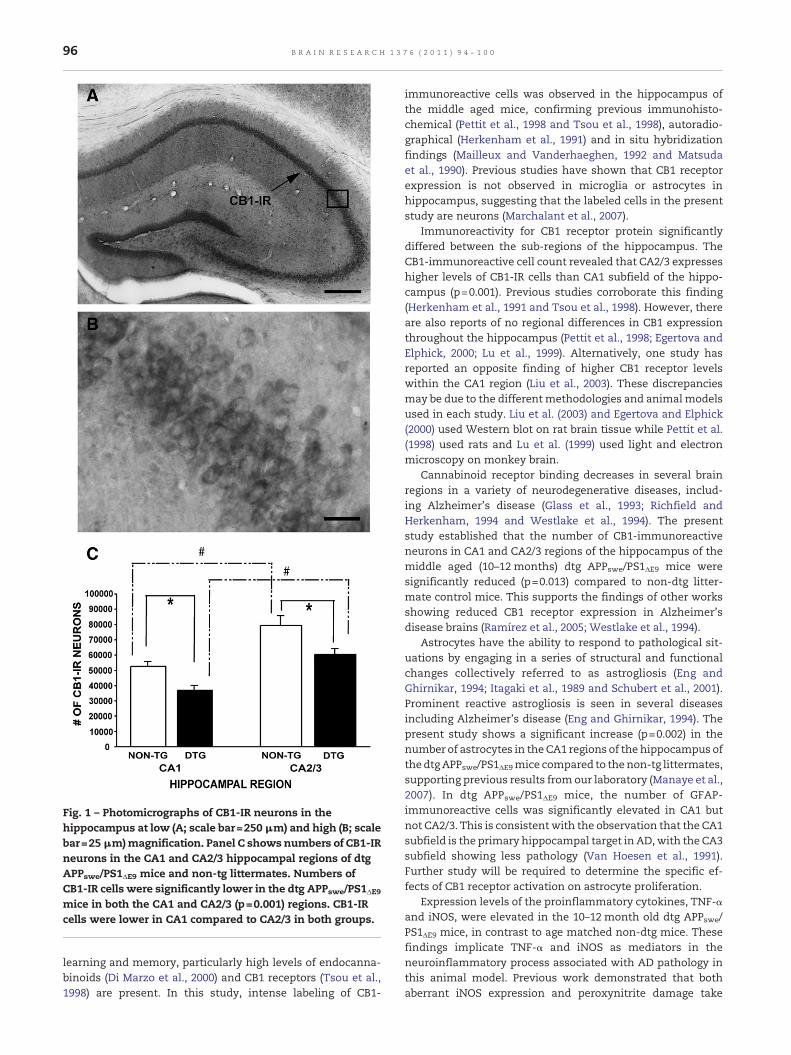
Fig. 1 – Photomicrographs of CB1-IR neurons in thehippocampus at low (A; scale bar=250 μm) and high (B; scalebar=25 μm)magnification. Panel C shows numbers of CB1-IRneurons in the CA1 and CA2/3 hippocampal regions of dtgAPPswe/PS1ΔE9 mice and non-tg littermates. Numbers ofCB1-IR cells were significantly lower in the dtg APPswe/PS1ΔE9mice in both the CA1 and CA2/3 (p=0.001) regions. CB1-IRcells were lower in CA1 compared to CA2/3 in both groups.
96 B R A I N R E S E A R C H 1 3 7 6 ( 2 0 1 1 ) 9 4 – 1 0 0
learning and memory, particularly high levels of endocanna-binoids (Di Marzo et al., 2000) and CB1 receptors (Tsou et al.,1998) are present. In this study, intense labeling of CB1-
immunoreactive cells was observed in the hippocampus ofthe middle aged mice, confirming previous immunohisto-chemical (Pettit et al., 1998 and Tsou et al., 1998), autoradio-graphical (Herkenham et al., 1991) and in situ hybridizationfindings (Mailleux and Vanderhaeghen, 1992 and Matsudaet al., 1990). Previous studies have shown that CB1 receptorexpression is not observed in microglia or astrocytes inhippocampus, suggesting that the labeled cells in the presentstudy are neurons (Marchalant et al., 2007).
Immunoreactivity for CB1 receptor protein significantlydiffered between the sub-regions of the hippocampus. TheCB1-immunoreactive cell count revealed that CA2/3 expresseshigher levels of CB1-IR cells than CA1 subfield of the hippo-campus (p=0.001). Previous studies corroborate this finding(Herkenham et al., 1991 and Tsou et al., 1998). However, thereare also reports of no regional differences in CB1 expressionthroughout the hippocampus (Pettit et al., 1998; Egertova andElphick, 2000; Lu et al., 1999). Alternatively, one study hasreported an opposite finding of higher CB1 receptor levelswithin the CA1 region (Liu et al., 2003). These discrepanciesmay be due to the different methodologies and animalmodelsused in each study. Liu et al. (2003) and Egertova and Elphick(2000) used Western blot on rat brain tissue while Pettit et al.(1998) used rats and Lu et al. (1999) used light and electronmicroscopy on monkey brain.
Cannabinoid receptor binding decreases in several brainregions in a variety of neurodegenerative diseases, includ-ing Alzheimer's disease (Glass et al., 1993; Richfield andHerkenham, 1994 and Westlake et al., 1994). The presentstudy established that the number of CB1-immunoreactiveneurons in CA1 and CA2/3 regions of the hippocampus of themiddle aged (10–12 months) dtg APPswe/PS1ΔE9 mice weresignificantly reduced (p=0.013) compared to non-dtg litter-mate control mice. This supports the findings of other worksshowing reduced CB1 receptor expression in Alzheimer'sdisease brains (Ramírez et al., 2005; Westlake et al., 1994).
Astrocytes have the ability to respond to pathological sit-uations by engaging in a series of structural and functionalchanges collectively referred to as astrogliosis (Eng andGhirnikar, 1994; Itagaki et al., 1989 and Schubert et al., 2001).Prominent reactive astrogliosis is seen in several diseasesincluding Alzheimer's disease (Eng and Ghirnikar, 1994). Thepresent study shows a significant increase (p=0.002) in thenumber of astrocytes in the CA1 regions of the hippocampus ofthedtgAPPswe/PS1ΔE9mice compared to thenon-tg littermates,supporting previous results from our laboratory (Manaye et al.,2007). In dtg APPswe/PS1ΔE9 mice, the number of GFAP-immunoreactive cells was significantly elevated in CA1 butnot CA2/3. This is consistentwith the observation that the CA1subfield is the primary hippocampal target in AD, with the CA3subfield showing less pathology (Van Hoesen et al., 1991).Further study will be required to determine the specific ef-fects of CB1 receptor activation on astrocyte proliferation.
Expression levels of the proinflammatory cytokines, TNF-αand iNOS, were elevated in the 10–12 month old dtg APPswe/PS1ΔE9 mice, in contrast to age matched non-dtg mice. Thesefindings implicate TNF-α and iNOS as mediators in theneuroinflammatory process associated with AD pathology inthis animal model. Previous work demonstrated that bothaberrant iNOS expression and peroxynitrite damage take
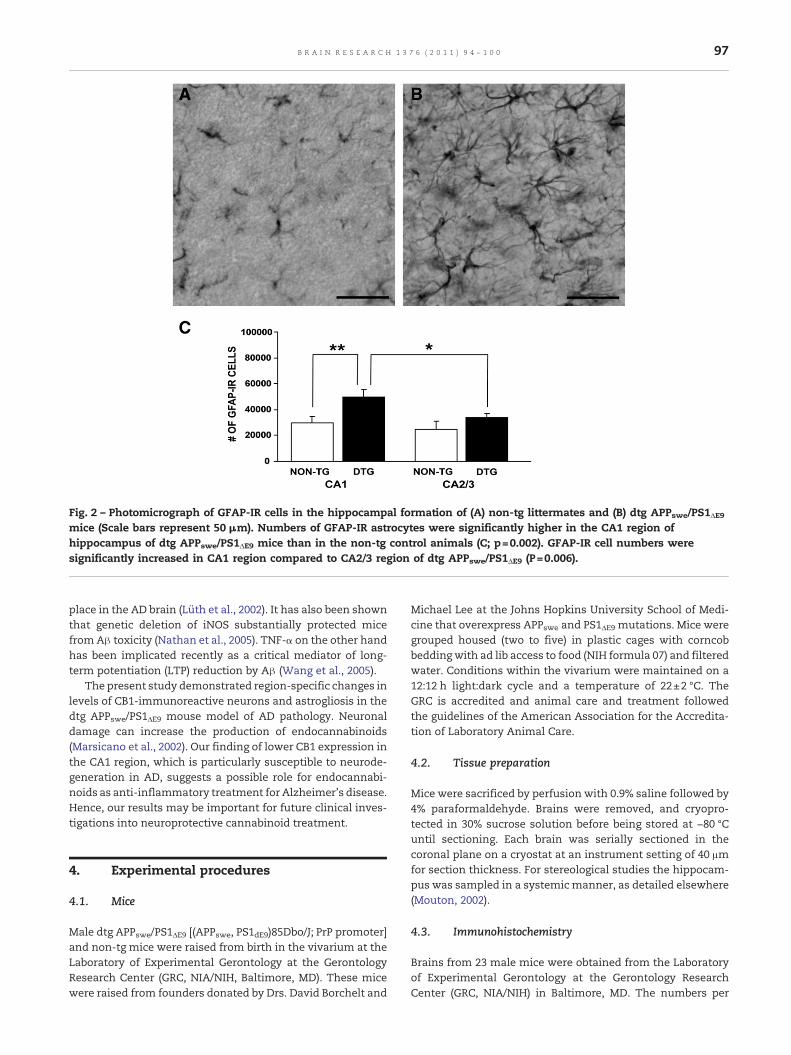
Fig. 2 – Photomicrograph of GFAP-IR cells in the hippocampal formation of (A) non-tg littermates and (B) dtg APPswe/PS1ΔE9mice (Scale bars represent 50 μm). Numbers of GFAP-IR astrocytes were significantly higher in the CA1 region ofhippocampus of dtg APPswe/PS1ΔE9 mice than in the non-tg control animals (C; p=0.002). GFAP-IR cell numbers weresignificantly increased in CA1 region compared to CA2/3 region of dtg APPswe/PS1ΔE9 (P=0.006).
97B R A I N R E S E A R C H 1 3 7 6 ( 2 0 1 1 ) 9 4 – 1 0 0
place in the AD brain (Lüth et al., 2002). It has also been shownthat genetic deletion of iNOS substantially protected micefromAβ toxicity (Nathan et al., 2005). TNF-α on the other handhas been implicated recently as a critical mediator of long-term potentiation (LTP) reduction by Aβ (Wang et al., 2005).
The present study demonstrated region-specific changes inlevels of CB1-immunoreactive neurons and astrogliosis in thedtg APPswe/PS1ΔE9 mouse model of AD pathology. Neuronaldamage can increase the production of endocannabinoids(Marsicano et al., 2002). Our finding of lower CB1 expression inthe CA1 region, which is particularly susceptible to neurode-generation in AD, suggests a possible role for endocannabi-noids as anti-inflammatory treatment for Alzheimer's disease.Hence, our results may be important for future clinical inves-tigations into neuroprotective cannabinoid treatment.
4. Experimental procedures
4.1. Mice
Male dtg APPswe/PS1ΔE9 [(APPswe, PS1dE9)85Dbo/J; PrP promoter]and non-tg mice were raised from birth in the vivarium at theLaboratory of Experimental Gerontology at the GerontologyResearch Center (GRC, NIA/NIH, Baltimore, MD). These micewere raised from founders donated by Drs. David Borchelt and
Michael Lee at the Johns Hopkins University School of Medi-cine that overexpress APPswe and PS1ΔE9 mutations. Mice weregrouped housed (two to five) in plastic cages with corncobbeddingwith ad lib access to food (NIH formula 07) and filteredwater. Conditions within the vivarium were maintained on a12:12 h light:dark cycle and a temperature of 22±2 °C. TheGRC is accredited and animal care and treatment followedthe guidelines of the American Association for the Accredita-tion of Laboratory Animal Care.
4.2. Tissue preparation
Mice were sacrificed by perfusion with 0.9% saline followed by4% paraformaldehyde. Brains were removed, and cryopro-tected in 30% sucrose solution before being stored at −80 °Cuntil sectioning. Each brain was serially sectioned in thecoronal plane on a cryostat at an instrument setting of 40 μmfor section thickness. For stereological studies the hippocam-pus was sampled in a systemic manner, as detailed elsewhere(Mouton, 2002).
4.3. Immunohistochemistry
Brains from 23 male mice were obtained from the Laboratoryof Experimental Gerontology at the Gerontology ResearchCenter (GRC, NIA/NIH) in Baltimore, MD. The numbers per
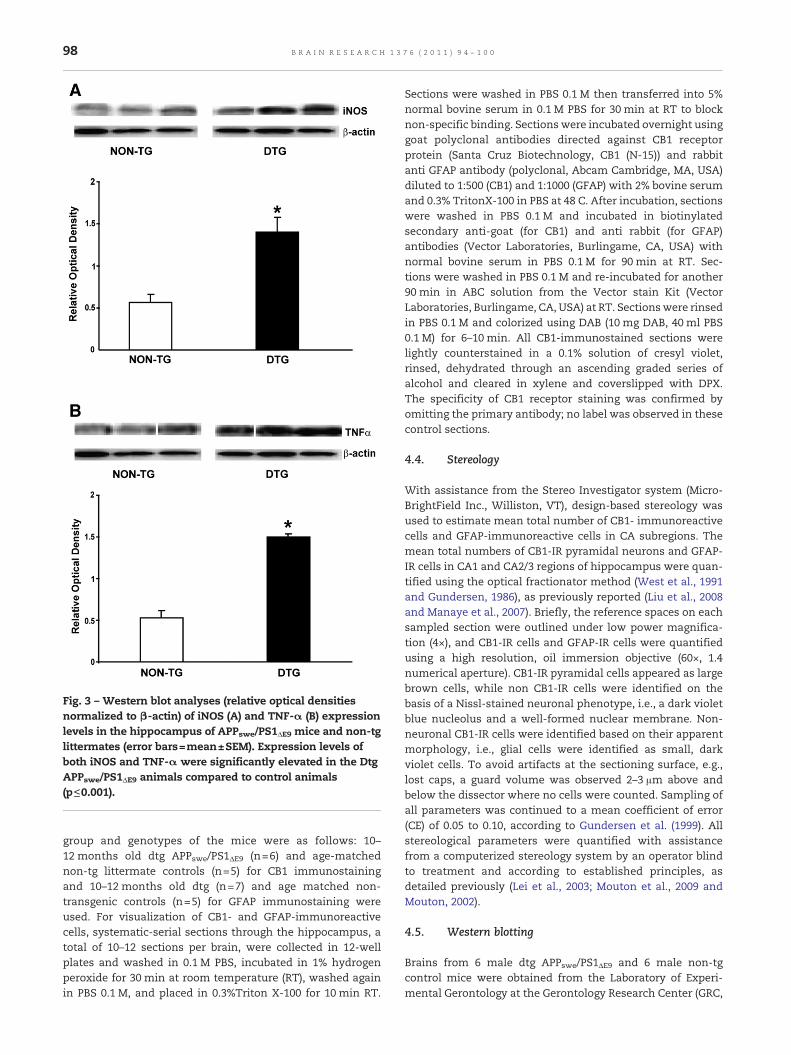
Fig. 3 – Western blot analyses (relative optical densitiesnormalized to β-actin) of iNOS (A) and TNF-α (B) expressionlevels in the hippocampus of APPswe/PS1ΔE9 mice and non-tglittermates (error bars=mean±SEM). Expression levels ofboth iNOS and TNF-α were significantly elevated in the DtgAPPswe/PS1ΔE9 animals compared to control animals(p≤0.001).
98 B R A I N R E S E A R C H 1 3 7 6 ( 2 0 1 1 ) 9 4 – 1 0 0
group and genotypes of the mice were as follows: 10–12 months old dtg APPswe/PS1ΔE9 (n=6) and age-matchednon-tg littermate controls (n=5) for CB1 immunostainingand 10–12 months old dtg (n=7) and age matched non-transgenic controls (n=5) for GFAP immunostaining wereused. For visualization of CB1- and GFAP-immunoreactivecells, systematic-serial sections through the hippocampus, atotal of 10–12 sections per brain, were collected in 12-wellplates and washed in 0.1 M PBS, incubated in 1% hydrogenperoxide for 30 min at room temperature (RT), washed againin PBS 0.1 M, and placed in 0.3%Triton X-100 for 10 min RT.
Sections were washed in PBS 0.1 M then transferred into 5%normal bovine serum in 0.1 M PBS for 30 min at RT to blocknon-specific binding. Sections were incubated overnight usinggoat polyclonal antibodies directed against CB1 receptorprotein (Santa Cruz Biotechnology, CB1 (N-15)) and rabbitanti GFAP antibody (polyclonal, Abcam Cambridge, MA, USA)diluted to 1:500 (CB1) and 1:1000 (GFAP) with 2% bovine serumand 0.3% TritonX-100 in PBS at 48 C. After incubation, sectionswere washed in PBS 0.1 M and incubated in biotinylatedsecondary anti-goat (for CB1) and anti rabbit (for GFAP)antibodies (Vector Laboratories, Burlingame, CA, USA) withnormal bovine serum in PBS 0.1 M for 90 min at RT. Sec-tions were washed in PBS 0.1 M and re-incubated for another90 min in ABC solution from the Vector stain Kit (VectorLaboratories, Burlingame, CA, USA) at RT. Sectionswere rinsedin PBS 0.1 M and colorized using DAB (10 mg DAB, 40 ml PBS0.1 M) for 6–10 min. All CB1-immunostained sections werelightly counterstained in a 0.1% solution of cresyl violet,rinsed, dehydrated through an ascending graded series ofalcohol and cleared in xylene and coverslipped with DPX.The specificity of CB1 receptor staining was confirmed byomitting the primary antibody; no label was observed in thesecontrol sections.
4.4. Stereology
With assistance from the Stereo Investigator system (Micro-BrightField Inc., Williston, VT), design-based stereology wasused to estimate mean total number of CB1- immunoreactivecells and GFAP-immunoreactive cells in CA subregions. Themean total numbers of CB1-IR pyramidal neurons and GFAP-IR cells in CA1 and CA2/3 regions of hippocampus were quan-tified using the optical fractionator method (West et al., 1991and Gundersen, 1986), as previously reported (Liu et al., 2008and Manaye et al., 2007). Briefly, the reference spaces on eachsampled section were outlined under low power magnifica-tion (4×), and CB1-IR cells and GFAP-IR cells were quantifiedusing a high resolution, oil immersion objective (60×, 1.4numerical aperture). CB1-IR pyramidal cells appeared as largebrown cells, while non CB1-IR cells were identified on thebasis of a Nissl-stained neuronal phenotype, i.e., a dark violetblue nucleolus and a well-formed nuclear membrane. Non-neuronal CB1-IR cells were identified based on their apparentmorphology, i.e., glial cells were identified as small, darkviolet cells. To avoid artifacts at the sectioning surface, e.g.,lost caps, a guard volume was observed 2–3 μm above andbelow the dissector where no cells were counted. Sampling ofall parameters was continued to a mean coefficient of error(CE) of 0.05 to 0.10, according to Gundersen et al. (1999). Allstereological parameters were quantified with assistancefrom a computerized stereology system by an operator blindto treatment and according to established principles, asdetailed previously (Lei et al., 2003; Mouton et al., 2009 andMouton, 2002).
4.5. Western blotting
Brains from 6 male dtg APPswe/PS1ΔE9 and 6 male non-tgcontrol mice were obtained from the Laboratory of Experi-mental Gerontology at the Gerontology Research Center (GRC,

99B R A I N R E S E A R C H 1 3 7 6 ( 2 0 1 1 ) 9 4 – 1 0 0
NIA/NIH) in Baltimore, MD. The hippocampi of the left hemi-sphere from all the mice were dissected out. The hippocampiof the dtg APPswe/PS1ΔE9 and that of the control mice werehomogenized separately in an ice-chilled solution (50 mMTris–HCl at pH 7.4) with protease inhibitor (cocktail tabletcomplete, from Roche). The homogenate from each mousewas centrifuged (Beckman Avanti J-25 I) at 32,000×g for30 min. The supernatant was collected and protein concen-tration was measured using the BCA Protein assay withbovine serum albumin as a standard. Equal amounts ofprotein (20 μg) were loaded onto a 4–15% SDS-PAGE thentransferred to a polyvinylidenedifluoride (PVDF) membrane.The membrane was probed with primary antibody (mouseanti TNF-α and anti-iNOS antibodies 1:1000) followed byhorseradish peroxidase-conjugated secondary antibody (Goatanti-mouse IgG, 1:10,000). Antibody detection was performedusing enhanced Chemoluminescence reagents Super SignalWest Dura. The membrane was exposed to Hyperfilm MP(Amersham Biosciences, U.S.A.) and developed using KodakGBX developer and fixer. Densitometric analysis was con-ducted using Quantity One imaging software (BioRad, CA) toquantify the intensity of bands from five independentWestern blots.
4.6. Statistical analysis
Two-way analysis of variance followed by Bonferroni post hoctest was used to compare the effects of transgene and hippo-campal subregions for measures of GFAP and CB-1 immu-noreactivity. Student's t test was used for statistical analysisof iNOS and TNFα levels determined by Western blot.
Acknowledgments
The authors wish to acknowledge support from the NIH/NINDS Grant number: U54 NS42867, Drs. Donald Ingram andPeter Mouton for their donation of the brains, and the NIAIntramural Program in Baltimore, Maryland. We would alsolike to acknowledge Dr. Jharna Das for her assistance with theWestern blot analysis.
R E F E R E N C E S
Baker, D., Pryce, G., Croxford, J.L., Brown, P., Pertwee, R.G.,Huffman, J.W., Layward, L., 2000. Cannabinoids controlspasticity and tremor in a multiple sclerosis model. Nature404, 84–87.
Devane, W.A., Hanus, L., Breuer, A., Pertwee, R.G., Stevenson, L.A.,Griffin, G., Gibson, D., Mandelbaum, A., Etinger, A., Mechoulam,R., 1992. Isolation and structure of a brain constituent thatbinds to the cannabinoid receptor. Science 258 (5090),1946–1949.
Di Marzo, V., Deutsch, D.G., 1998. Endocannabinoids: endogenouscannabinoid receptor ligands with neuromodulatory action.Trends Neurosci. 21 (12), 521–528 Review. Erratum in: TrendsNeurosci. 22 (2), 80 Feb.
Di Marzo, V., Breivogel, C.S., Tao, Q., Bridgen, D.T., Razdan, R.K.,Zimmer, A.M., Zimmer, A., Martin, B.R., 2000. Levels,metabolism, and pharmacological activity of anandamide in
CB(1) cannabinoid receptor knockout mice: evidence fornon-CB(1), non-CB(2) receptor-mediated actions ofanandamide in mouse brain. J. Neurochem. 75, 2434–2444.
Egertova, M., Elphick, M.R., 2000. Localisation of cannabinoidreceptors in the rat brain using antibodies to the intracellularC-terminal tail of CB. J. Comp. Neurol. 422, 159–171.
Eikelenboom, P., Zhan, S.S., van Gool, W.A., Allsop, D., 1994.Inflammatory mechanisms in Alzheimer's disease. TrendsPharmacol. Sci. 15, 447–450.
Eng, L.F., Ghirnikar, R.S., 1994. GFAP and astrogliosis. Brain Pathol.4, 229–237.
Galiegue, S., Mary, S., Marchand, J., Dussossory, D., Carriere, D.,Carayon, P., Bouaboula, M., Shire, D., Le Fur, G., Casellas, P.,1995. Expression of central and peripheral cannabinoidreceptors in human immune tissues and leukocytesubpopulations. Eur. J. Biochem. 232, 54–61.
Games, D., Adams, D., Alessandrini, R., Barbour, R., Berthelette, P.,Blackwell, C., Carr, T., Clemens, J., Donaldson, T., Gillespie, F.,1995. Alzheimer-type neuropathology in transgenic miceoverexpressing V717F beta-amyloid precursor protein. Nature373, 523–527.
Glass, M., Faull, R.L., Dragunow, M., 1993. Loss of cannabinoidreceptors in the substantia nigra in Huntington's disease.Neuroscience 56, 523–527.
Gundersen, H.J., Jensen, E.B., Kiêu, K., Nielsen, J., 1999. Theefficiency of systematic sampling in stereology-reconsidered.J. Microsc. 193, 199–211.
Gundersen, H.J., 1986. Stereology of arbitrary particles. A reviewof unbiased number and size estimators and the presentationof some new ones, in memory of William R. Thompson. J.Microsc. 143, 3–45.
Hampson, A.J., Grimaldi, M., Axelrod, J., Wink, D., 1998.Cannabidiol and (–)Δ9-tetrahydrocannabinol areneuroprotective antioxidants. Med. Sci. 95, 8268–8273.
Hardy, J., 1997. Amyloid, the presenilins and Alzheimer's disease.Trends Neurosci. 20, 154–159.
Herkenham, M., Lynn, A.B., Johnson, M.R., Melvin, L.S., de Costa,B.R., Rice, K.C., 1991. Characterization and localization ofcannabinoid receptors in rat brain: a quantitative in vitroautoradiographic study. J. Neurosci. 11, 563–583.
Hsiao, K.K., Borchelt, D.R., Olson, K., Johannsdottir, R., Kitt, C.,Yunis, W., Xu, S., Eckman, C., Younkin, S., Price, D., 1995.Age-related CNS disorder and early death in transgenicFVB/N mice overexpressing Alzheimer amyloid precursorproteins. Neuron 15, 1203–1218.
Itagaki, S., McGeer, P.L., Akiyama, H., Zhu, S., Selkoe, D., 1989.Relationship of microglia and astrocytes to amyloid depositsof Alzheimer disease. J. Neuroimmunol. 24, 173–182.
Johnson-Wood, K., Lee, M., Motter, R., Hu, K., Gordon, G., Barbour,R., Khan, K., Gordon, M., Tan, H., Games, D., Lieberburg, I.,Schenk, D., Seubert, P., McConlogue, L., 1997. Amyloidprecursor protein processing and AB42 deposition in atransgenic mouse model of Alzheimers disease. Proc. NatlAcad. Sci. 94, 1550–1555.
Lei, D.L., Long, J.M., Hengemihle, J., O'Neill, J., Manaye, K.F.,Ingram, D.K., Mouton, P.R., 2003. Effects of estrogen andraloxifene on neuroglia number and morphology in thehippocampus of aged female mice. Neuroscience 121,659–666.
Liu, Y., Yoo, M.J., Savonenko, A., Stirling,W., Price, D.L., Borchelt, D.R., Mamounas, L., Lyons, W.E., Blue, M.E., Lee, M.K., 2008.Amyloid pathology is associated with progressivemonoaminergic neurodegeneration in a transgenic mousemodel of Alzheimer's disease. J. Neurosci. 28, 13805–13814.
Liu, P., Bilkey, D.K., Darlington, C.L., Smith, P.F., 2003. CannabinoidCB1 receptor protein expression in the rat hippocampus andentorhinal, perirhinal, postrhinal and temporal cortices:regional variations and age-related changes. Brain Res. 979,235–239.

100 B R A I N R E S E A R C H 1 3 7 6 ( 2 0 1 1 ) 9 4 – 1 0 0
Lüth, H.J., Münch, G., Arendt, T., 2002. V Aberrant expression ofNOS isoforms in Alzheimer's disease is structurally related tonitrotyrosine formation. Brain Res. 953 (1-2), 135–143 Oct 25.
Lu, X.R., Ong, W.Y., Mackie, K., 1999. A light and electronmicroscopic study of the CB1 cannabinoid receptor in primatebrain. Neuroscience 92, 1177–1191.
Mailleux, P., Vanderhaeghen, J.J., 1992. Distribution of neuronalcannabinoid receptor in the adult rat brain: a comparativereceptor binding radioautography and in situ hybridizationhistochemistry. Neuroscience 48, 655–668.
Malherbe, P., Richards, J.G., Martin, J.R., Bluethmann, H., Maqqio, J.,Huber, G., 1996. Lack of beta-amyloidosis in transgenic miceexpressing low levels of familial Alzheimer's disease missensemutations. Neurobiol. Aging 17, 205–214.
Manaye, K.F., Wang, P.C., O'Neil, J.N., Huang, S.Y., Xu, T., Lei, D.L.,Tizabi, Y., Ottinger, M.A., Ingram, D.K., Mouton, P.R., 2007.Neuropathological quantification of dtg APPswe/PS1ΔE9neuroimaging, stereology, and biochemistry. AGE 29, 87–96.
Marchalant, Y., Cerbai, F., Brothers, H.M., Wenk, G.L., 2007.Anti-inflammatory property of the cannabinoid agonistWIN-55212-2 in a rodent model of chronic brain inflammationFeb 23 Neuroscience 144 (4), 1516–1522 Epub 2006 Dec 18.
Marsicano, G., Moosmann, B., Hermann, H., Lutz, B., Behl, C., 2002.Neuroprotective properties of cannabinoids against oxidativestress: role of the cannabinoid receptor CB1. J. Neurochem. 80,448–456.
Matsuda, L.A., 1997. Molecular aspects of cannabinoid receptors.Crit. Rev. Neurobiol. 11, 143–166.
Matsuda, L.A., Loliat, S.J., Brownstein, M.J., Young, A.C., Bonner,T.I., 1990. Structure of a cannabinoid receptor and functionalexpression of the cloned cDNA. Nature 346, 561–564.
McGeer, P.L., McGeer, E.G., 1995. The inflammatory responsesystem of brain: implications for therapy of Alzheimer andother neurodegenerative diseases. Brain Res. Rev. 21, 195–218.
Mechoulam, R., Ben-Shabat, S., Hanus, L., Ligumsky, M., Kaminski,N.E., Schatz, A.R., Gopher, A., Almog, S., Martin, B.R., Compton,D.R., 1995. Identification of an endogenous 2-monoglyceride,present in canine gut, that binds to cannabinoid receptors.Biochem. Pharmacol. 50 (1), 83–90.
Mechoulam, R., Panikashvili, D., Shohami, E., 2002. Cannabinoidsand brain injury: therapeutic implications. Trends Mol. Med. 8,58–61.
Morgan, D., Diamond, D.M., Gottschall, P.E., Ugen, K.E., Dickey, C.,Hardy, J., Duff, K., Jantzen, P., DiCarlo, G., Wilcock, D., Connor,K., Hatcher, J., Hope, C., Gordon, M., Arendash, G.W., 2000. Abeta peptide vaccination prevents memory loss in an animalmodel of Alzheimer's disease. Nature 408, 982–985.
Mouton, P.R., 2002. Principles and Practices of UnbiasedStereology: An Introduction for Bioscientists. The JohnsHopkins University Press, Baltimore, Maryland.
Mouton, P.R., Chachich, M.E., Quigley, C., Spangler, E., Ingram, D.K., 2009. Caloric restriction attenuates cortical amyloidosis in adouble transgenic mouse model of Alzheimer's disease.Neurosci. Lett. 464, 184–187.
Nathan, C., Calingasan, N., Nezezon, J., Ding, A., Lucia, M.S.,La Perle, K., Fuortes, M., Lin, M., Ehrt, S., Kwon, N.S., Chen, J.,Vodovotz, Y., Kipiani, K., Beal, M.F., 2005. Protection fromAlzheimer's-like disease in the mouse by genetic ablation ofinducible nitric oxide synthase Nov 7 J. Exp. Med. 202 (9),1163–1169 Epub 2005 Oct 31.
Onaivi, E.S., Ishiguro, H., Gong, J.P., Patel, S., Perchuk, A., Meozzi, P.A., Myers, L., Mora, Z., Tagliaferro, P., Gardner, E., Brusco, A.,Akinshola, B.E., Liu, Q.R., Hope, B., Iwasaki, S., Arinami, T.,Teasenfitz, L., Uhl, G.R., 2006. Discovery of the presence andfunctional expression of cannabinoid CB2 receptors in thebrain. Ann. NY Acad. Sci. 1074, 514–536.
Pettit, D.A., Harrison, M.P., Olson, J.M., Spencer, R.F., Cabral, G.A.,1998. Immunohistochemical localization of the neuralcannabinoid receptor in rat brain. J. Neurosci. Res. 51,391–402.
Piomelli, D., Giuffrida, A., Calignano, A., Rodríguez de Fonseca, F.,2000. The endocannabinoid system as a target for therapeuticdrugs. Trends Pharmacol. Sci. 21, 218–224.
Ramírez, B.G., Blázquez, C., Gómez del Pulgar, T., Guzmán, M.,de Ceballos, M.L., 2005. Prevention of Alzheimer's diseasepathology by cannabinoids: neuroprotection mediated byblockade of microglial activation. J. Neurosci. 25, 1904–1913.
Richfield and Herkenham, 1994. Selective vulnerability inHuntington's disease: preferential loss of cannabinoidreceptors in lateral globus-pallidus. Ann. Neurol. 36,577–584.
Rogers, J., Webster, S., Lue, L.F., Brachova, L., Civin, W.H.,Emmerling, M., Shivers, B., Walker, D., McGeer, P., 1996.Inflammation and Alzheimer's disease pathogenesisNeurobiol. Aging 17, 681–686.
Schubert, P., Ogata, T., Marchini, C., Ferroni, S., 2001. Glia-relatedpathomechanisms in Alzheimer's disease: a therapeutictarget? Mech. Ageing Dev. 123, 47–57.
Selkoe, D.J., 2001. Alzheimer's disease: genes, proteins, andtherapy. Physiol. Rev. 81, 741–766.
Shen and Thayer, 1998. Cannabinoid receptor agonists protectcultured rat hippocampal neurons from excitotoxicity. Mol.Pharmacol. 54, 459–462.
Sturchler-Pierrat, C., Abramowski, D., Duke, M., Wiederhold, K.H.,Mistl, C., Rothacher, S., Ledermann, B., Bürki, K., Frey, P.,Paganetti, P.A., Waridel, C., Calhoun, M.E., Jucker, M., Probst, A.,Staufenbiel, M., Sommer, B., 1997. Two amyloid precursorprotein transgenic mouse models with Alzheimer disease-likepathology. Proc. Natl Acad. Sci. 94, 13287–13292.
Sugiura, T., Kondo, S., Sukagawa, A., Nakane, S., Shinoda, A., Itoh,K., Yamashita, A., Waku, K., 1995. 2-Arachidonoylglycerol: apossible endogenous cannabinoid receptor ligand in brain.Biochem. Biophys. Res. Commun. 215 (1), 89–97.
Troncoso, J.C., Sukhov, R.R., Kawas, C.H., Koliatsos, V.E., 1996. Insitu labeling of dying cortical neurons in normal aging and inAlzheimer's disease: correlations with senile plaques anddisease progression. J. Neuropathol. Exp. Neurol. 55, 1134–1142.
Tsou, K., Brown, S., Sañudo-Peña, M.C., Mackie, K., Walker, J.M.,1998. Immunohistochemical distribution of cannabinoid CB1receptors in the rat central nervous system. Neuroscience 83,393–411.
Van Hoesen, G.W., Hyman, B.T., Damasio, A.R., 1991. Entorhinalcortex pathology in Alzheimer's disease. Hippocampus 1, 1–8.
Wang, Tanila H., Puolivali, J., Kadish, I., Van Groen, T., 2003.Gender differences in the amount and deposition ofamyloid-beta in APPswe and PS1 double transgenic mice.Neurobiol. Dis. 14, 318–327.
Wang, Q., Wu, J., Rowan, M.J., Anwyl, R., 2005. Beta-amyloidinhibition of long-term potentiation is mediated via tumornecrosis factor. Eur. J. Neurosci. 22 (11), 2827–2832 Dec.
West, M.J., Slomianka, L., Gunderson, H.J., 1991. Unbiasedstereological estimation of the total number of neurons inthe subdivisions of the rat hippocampus using the opticalfractionator. Anat. Rec. 231, 482–497.
Westlake, T.M., Howlett, A.C., Bonner, T.I., Matsuda, L.A.,Herkenham, M., 1994. Cannabinoid receptor binding andmessenger RNA expression in human brain: an in vitroreceptor autoradiography and in situ hybridizationhistochemistry study of normal aged and Alzheimer's brains.Neuroscience 63, 637–652.
Wisniewski, H.M., Terry, R.D., 1973. Morphology of the aging brain,human and animal. Prog. Brain Res. 40, 167–186.