Embed Size (px)
Citation preview

21
DIVERSIDADE E ECOLOGIA DE MORCEGOS, COM ÊNFASE
NA FAMÍLIA PHYLLOSTOMIDAE (CHIROPTERA: MAMMALIA),
ASSOCIADA ÀS FLORESTAS DE MANGUE E TERRA FIRME
NA FAZENDA DAS SALINAS, BRAGANÇA - PA
FERNANDA ATANAENA GONÇALVES DE ANDRADE
Prof. Dr. Marcus E. B. Fernandes
Orientador
Bragança-PA -2004-
UNIVERSIDADE FEDERAL DO PARÁ
CAMPUS UNIVERSITÁRIO DE BRAGANÇA
CURSO DE PÓS-GRADUAÇÃO EM BIOLOGIA AMBIENTAL
MESTRADO EM ECOLOGIA DE ECOSSISTEMAS COSTEIROS E ESTUARINOS

22
DIVERSIDADE E ECOLOGIA DE MORCEGOS, COM ÊNFASE NA F AMÍLIA
PHYLLOSTOMIDAE (CHIROPTERA: MAMMALIA), ASSOCIADA ÀS
FLORESTAS DE MANGUE E TERRA FIRME NA FAZENDA DAS SA LINAS,
BRAGANÇA - PA
FERNANDA ATANAENA GONÇALVES DE ANDRADE
Dissertação de Mestrado apresentada ao Curso de
Mestrado em Biologia Ambiental (Área de
Concentração: Ecologia de Ecossistemas Costeiros e
Estuarinos), da Universidade Federal do Pará,
Campus de Bragança, como parte dos requisitos
necessários à obtenção do grau de Mestre em
Biologia Ambiental.
Prof. Dr. Marcus E. B. Fernandes
Orientador
Bragança-PA - 2004-
UNIVERSIDADE FEDERAL DO PARÁ
CAMPUS UNIVERSITÁRIO DE BRAGANÇA
CURSO DE PÓS-GRADUAÇÃO EM BIOLOGIA AMBIENTAL
MESTRADO EM ECOLOGIA DE ECOSSISTEMAS COSTEIROS E ESTUARINOS
UNIVERSIDADE FEDERAL DO PARÁ
CAMPUS UNIVERSITÁRIO DE BRAGANÇA
CURSO DE PÓS-GRADUAÇÃO EM BIOLOGIA AMBIENTAL
MESTRADO EM ECOLOGIA DE ECOSSISTEMAS COSTEIROS E ESTUARINOS

23
DIVERSIDADE E ECOLOGIA DE MORCEGOS, COM ÊNFASE NA F AMÍLIA
PHYLLOSTOMIDAE (CHIROPTERA: MAMMALIA), ASSOCIADA ÀS
FLORESTAS DE MANGUE E TERRA FIRME NA FAZENDA DAS SA LINAS,
BRAGANÇA - PA
FERNANDA ATANAENA GONÇALVES DE ANDRADE
Dissertação de Mestrado apresentada ao Curso de Mestrado em Biologia
Ambiental (Área de Concentração: Ecologia de Ecossistemas Costeiros e
Estuarinos), da Universidade Federal do Pará, Campus de Bragança, como parte
dos requisitos necessários à obtenção do grau de Mestre em Biologia Ambiental.
Aprovado por:
Prof. Dr. Marcus E. B. Fernandes (UFPA – Campus de Bragança / Orientador)
Prof. Dr. José Souto Rosa Filho (UFPA – Belém)
Profa. Dra. Claudia Nunes (UFPA – Campus de Bragança)
Dr. Enrico Bernard (Conservação Internacional - CI)
Bragança-PA - 2004-

24
“Viva como se fosse morrer amanhã.
Aprenda como se você fosse viver para sempre.”
Mahatma Gandhi

25
Para a maravilhosa,
BEATRIZ
e ao encantador,
OSVALDO

26
in memorian
Aos meus amigos de Tucuruí que partiram para longe dos meus olhos, mas permanecem vivos no meu pensamento, por não termos compartilhado os últimos sorrisos. Saudades de:
Kelly, Mauro, Gilberto, Lêda e Manoel Mochida.
Em especial ao meu amigo e mestre Jairo.

27
AGRADECIMENTOS
Dos diversos encontros ocorridos na simpática Bragança, uma grande rede
de amizades foi originada, e a todos os amigos que cativei, um grande abraço de
morcego e meus sinceros agradecimentos:
Primeiramente ao Prof. Dr. Marcus Emanuel Barroncas Fernandes (UFPA-
Campus de Bragança) pela orientação, companheirismo e paciência,
contribuindo para o meu crescimento profissional, ao aceitar o grande desafio
de estudar os misteriosos morcegos;
Ao Dr. Ulf Mehling (Projeto MADAM) pela credibilidade e ajuda
proporcionando os nossos primeiros passos pela área de estudo;
À Dra. Iracilda Sampaio (UFPA – Campus de Bragança) pela confiança e
apoio durante todo o decorrer do trabalho;
À Dra. Suely Marques-Aguiar e ao Nélio Saldanha pelo auxilio na
identificação dos indivíduos;
Ao Dr. Enrico Bernard pelas dicas construtivas para o melhor desempenho
deste trabalho;
Aos aventureiros e desbravadores noturnos do manguezal: Adriana, Adriano, Brandão, Carlos André, Deivid, Dani, Geovanny, Jussara, Kelly, Magno, Natália, Renata, Stélio, Waldir e todos os demais alunos que apreciaram as mais belas estrelas cadentes das noites bragantinas;
Ao Sr. Purisso e Sra. Esmeralda por todas as madrugadas que passamos juntos acordados, e esperançosos para que cada dia trabalho fosse sempre um grande dia;
Aos amigos Priscila, Christina, César e Daniela por permitirem que a nossa
casa fosse também um laboratório;
À minha família bragantina, Elsen minha mãe-irmã Célia Serra e seus
maravilhosos filhos Roberto, Adrian e Princesa. Os quais me adotaram sem
restrições, cultivando o meu carinho e eterna gratidão;
Aos meus grandes e preciosos amigos Claudene, Helane, Elmary, Muzenilha e
Naldo;

28
Às bat-garotas Ana Paula, Elaine e Valéria por tanta dedicação e amizade.
Aos meus irmãos Socorro, Osvaldo, Denilson, Zeca, e principalmente pelo
apoio imensurável de Rosene e Atanail, pois somos uma grande família;
A colega Raquel por ter gentilmente cedido informações do seu trabalho,
contribuindo para o enriquecimento dos nossos resultados;
A todos os meus amigos do Museu Paraense Emílio Goeldi e aos dos
laboratórios da UFPA em Bragança, sintam-se abraçados também!
À Universidade Federal do Pará (Campus de Bragança), Núcleo de
Estudos Costeiros (NEC), Projeto Milênio e ao Projeto MADAM (Mangrove
Dynamics and Management) pelo apoio logístico que foi crucial para o
desenvolvimento deste trabalho;
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq) e a CAPES pela bolsa de estudo concedida.

29
FONTES DAS FIGURAS ADICIONAIS
Capa:
Buldog Bat ou Morcego Pescador (Noctilio leporinus – Noctilionidae). Bats: natural
history. HILL, J. E. & SMITH, (1986). 2nd Ed. University of Texas Press, USA,
página 64.
Dedicatória:
Símbolo chinês Wu-fu. Murcielagos de Venezuela. LINARES, O. J. (1987).
Cuadernos Lagoven, Departamento de Relaciones Públicas de Lagoven S. A.,
Caracas, página 09.
Agradecimentos:
Pteropus giganteus. Bats Biology and Behaviour. ALTRINGHAM, J. D. (1996).
Oxford University Press, Oxford, New Jersey and Tokyo, página 234.

30
SUMÁRIO
EPÍGRAFE i
DEDICATÓRIA ii
AGRADECIMENTOS iii
FONTE DAS FIGURAS ADICIONAIS v
SUMÁRIO vi
LISTA DE TABELAS viii
LISTA DE FIGURAS ix
RESUMO xi
ABSTRACT xii
1. INTRODUÇÃO
1
1.1. Biologia e ecologia de morcegos 3
2. OBJETIVOS 7
2.1. Objetivo Geral 7
2.2. Objetivos Específicos 7
3. MATERIAL E MÉTODOS 8
3.1. Área de Estudo 8
3.2. Procedimento de campo e tratamento das Espécimes 10
3.3. Análise dos Dados 14
3.3.1. Abundância de Indivíduos, Esp écies e Esforço de
Captura 14
3.3.2. Estimativas de Riqueza 15
3.3.3. Estimativas de Diversidade 16
3.3.3.1. Índice de Shannon-Wiener (H’) 17
3.3.3.2. Índice de Simpson (D) 17
3.3.4. Cálculos de Similaridade 18
3.3.5. Análise de Classificação 19
Página

31
3.3.6. Período de Atividade das Espécies mais
Abundantes 19
3.3.7. Razão Sexual 19
3.3.8. Flutuações Sazonais das Principais Guild as 19
3.3.9. Similaridade da Quiropterofauna no Estad o do
Pará 20
4. RESULTADOS 21
4.1. Abundância de Indivíduos e Espécies 21
4.1.1. Esforço de Captura de Indivíduos e Espéc ies 21
4.2. Estimativa de Riqueza em Espécies 26
4.3. Período de Atividade das Espécies mais Abundantes 30
4.4. Categorias Reprodutivas das Fêmeas e Razão Sexual 34
4.5. Flutuações Sazonais das Principais Guildas 34
4.6. Resultados das Capturas Diurnas e Noturnas Semestrais 36
4.7. Similaridade da Quiropterofauna no Estado do Pará 39
5. DISCUSSÃO 40
6. CONCLUSÕES 46
7. REFERÊNCIAS BIBLIOGRÁFICAS 47
ANEXOS 56

32
LISTA DE TABELAS
Página
Tabela 1 – Coordenadas geográficas dos sítios de trabalho nos dois ecossistemas na Fazenda das Salinas, Bragança-Pará.
13
Tabela 2 – Caracterização da quiropterofauna da Fazenda das Salinas, incluindo informações taxonômicas, nomes vulgares no Brasil (Emmons, 1997; Wilson et al.,1996; Silva, 1994), guildas (Emmons, 1997; Findley, 1993; Hill & Smith, 1986; Wilson et al., 1996), categoria de conservação (Fundação Biodiversitas, 2002; Ministério do Meio Ambiente, 2003), grau de ocorrência na área de estudo (Marques-Aguiar et al., 2002a) e total de indivíduos por espécies capturadas nos ecossistemas manguezal e terra firme. (+) = rara, de 1 a 5 indivíduos; (++) = baixa, de 6 a 10 indivíduos; (+++) = comum, acima de 10 (Marques-Aguiar et al., 2002a). 23
Tabela 3 – Proporções de capturas realizadas com redes-neblina em dois ecossistemas, manguezal e terra firme, e seus respectivos sítios de trabalho na Fazenda das Salinas, Bragança, Pará. H.R = total de horas de campo x o número de redes abertas; Ni = número total de indivíduos capturados; Ne = número total de espécies capturadas; DI = razão entre Ni e hora.rede, que representa a taxa de capturas dos espécimes; DE = razão entre Ne e hora.rede, que representa a taxa de espécies capturas; REC = razão entre Ne e Ni (espécies/captura). 25
Tabela 4 – Estimativas de riqueza em espécies para a Fazenda das Salinas. DV = Desvio Padrão; % complementar = percentual das espécies obtidas no presente trabalho, diante do esperado por cada estimador; ACE = Abundance-based Coverage Estimator; ICE = Incidence-based Coverage Estimator; MMMean = Michaelis-Menten Mean. Os cálculos foram realizados através do programa EstimateS 6.0b 1. 28 Tabela 5 – Índices de diversidade calculados para os ecossistemas separadamente e agrupados. 1 – D = Coeficiente de diversidade de Simpson; 1 / D = Nº de espécies comuns para gerar o coeficiente. 29

33
LISTA DE FIGURAS
Página
Figura 1 – Localização da área de estudo na península bragantina, destacando as quatro transecções abertas na “ilha” de terra-firme e na área de manguezal. T1 a T3 = Transecção 1; T2 e T4 = Transecção 2. Modificado de Mehlig (2001). 12
Figura 2 – Desenho esquemático da instalação da rede-neblina. Fonte: Auricchio (2002).
13
Figura 3 – Principais medidas morfométricas que foram obtidas para a identificação dos indivíduos capturados: comprimento do trago (T), da orelha interna e externa (O) e do antebraço (A). Modificado de Auricchio (2002).
13
Figura 4 – Percentual de famílias de quirópteros (A) e subfamílias de Phyllostomidae (B) capturados na Fazenda das Salinas em Bragança - Pará. 25
Figura 5 – Dendrograma mostrando a similaridade entre os quatro sítios de trabalho, com base nos seus respectivos valores de abundância. 26
Figura 6 – Curva de acumulação das espécies capturadas: (A) nos sistemas agrupados e (B) nos ecossistemas separados: M = manguezal e TF = terra firme, na Fazenda das Salinas, Bragança - Pará. 27
Figura 7 – Dendrograma mostrando a similaridade entre os quatro sítios de trabalho, com base na presença/ausência de indivíduos de cada espécie. 29
Figura 8 – Atividade noturna das cinco espécies de morcegos mais abundantes na Fazenda das Salinas, de acordo com os horários de captura, durante o período de dezembro de 2002 a outubro de 2003. Para tanto, foram utilizadas redes-neblina, abertas de 18:00 à 1:00 h,
totalizando sete horas diárias de campo.
31

34
Figura 9 – Diagrama climático da área de estudo, representando a flutuação da Temperatura (°C) e da Precipitação (mm ) ao longo do período de estudo.
32
Figura 10 – – Horário de captura nas duas estações (chuvosa e seca), das três espécies de morcegos mais abundantes na Fazenda das Salinas, Artibeus jamaicensis (A), Artibeus obscurus (B) e Carollia perspicillata (C), acompanhados ao longo de um ano (Dezembro de 2002 a outubro de 2003). Para tanto, foram utilizadas redes-neblina, abertas de 18:00 à 1:00 h, totalizando sete horas diárias de campo. 33
Figura 11 – Abundância relativa das guildas de morcegos nos ecossistemas estudados da Fazenda das Salinas, durantes o período de dezembro de 2002 a outubro de 2003. 35
Figura 12 – Variação sazonal do número de machos e fêmeas das guildas Frugívora (A) e Onívora (B), comparada com a variação sazonal das taxas de frutificação (g.m.-2.mês-1) na terra firme e precipitação (mm) na área de estudo, Fazenda das Salinas, Bragança-Pará. 37
Figura 13 – Variação sazonal do número de fêmeas reprodutivas grávidas (A) e lactantes (B) da guilda Frugívora, comparada com a variação sazonal das taxas de frutificação e de precipitação na área de estudo, Fazenda das Salinas, Bragança-Pará. 38
Figura 14 – Análise de Cluster revelando semelhanças na composição das espécies de seis estudos realizados em localidades geograficamente distintas, incluindo as amostras da Fazenda das Salinas em Bragança, Pará.
39

35
RESUMO
Este trabalho teve como objetivo estudar a diversidade e alguns aspectos
ecológicos da fauna de morcegos, com ênfase na família Phyllostomidae,
associada aos ecossistemas manguezal e terra firme da Fazenda das Salinas,
Bragança, Pará. Quatro sítios de trabalho foram escolhidos para realizar as
campanhas bimestrais de captura, durante um ciclo anual. As coletas foram
efetuadas com redes-neblina distribuídas em quatro transecções, duas na terra
firme e duas no manguezal. Durante as capturas, das 18:00 à 01:00 h, foi obtido
um total de 438 indivíduos de cinco famílias e 23 espécies, sendo 217 para a
terra firme e 221 para o manguezal. A família Phyllostomidae foi a mais
representada com 428 indivíduos. A análise comparativa entre os dois
ecossistemas mostrou não haver diferença significativa no que diz respeito a
quiropterofauna (ANOSIM, R=0,001; p>0,05) e a análise de cluster para os quatro
sítios de trabalho mostrou que há similaridade entre alguns sítios dos dois
ambientes. Os horários de atividade das espécies mais abundantes não foram
sobrepostos, bem como não foi registrada correlação significativa com a
precipitação. Provavelmente, os dois ecossistemas funcionem como fontes
diferenciadas de alimento para as espécies de morcegos. Assim, as “ilhas” de
terra firme seriam uma fonte principal de frutos, visto que normalmente há maior
disponibilidade de frutos neste sistema, ao passo que o manguezal poderia
simplesmente funcionar, para os frugívoros, como corredor de acesso a essas
“ilhas”. Artibeus jamaicensis foi uma das espécies mais capturadas e exibiu um
padrão reprodutivo poliéstrico bimodal, além do mais, este também foi o padrão
de distribuição das fêmeas lactantes ao longo do ano. O sucesso reprodutivo dos
morcegos frugívoros parece estar relacionado com a estação seca. A guilda
Frugívora foi a mais abundante com 64% dos indivíduos capturados, não havendo
contudo, diferença significativa entre o número de indivíduos das guildas
considerando os ambientes, mas somente quando as guildas foram comparadas
entre si (ANOVA, F=693; gl=2; p<0,05). Por fim, as altas taxas de captura da
subfamília Stenodermatinae e a presença de Desmodus rotundus na área de
estudo, poderiam ser possíveis indicadores biológicos da qualidade ambiental dos
bosques de terra firme da Fazenda das Salinas.

36
ABSTRACT
This work aimed to study the diversity and some ecological aspects of the
bat fauna, with emphasis on the family Phyllostomidae, associated to mangrove
and terra firme ecosystems of Fazenda das Salinas, Bragança, Pará. Four study
sites were chosen to undertake the bimonthly capture efforts, during an annual
cycle. Fieldwork were carried out by using mist net distributed in for transects, two
in terra firme and two in mangrove. During the captures, from 18:00 à 01:00 h, a
total of 438 individuals of five families and 23 species were obtained, being 217 for
terra firme and 221 for mangrove. The family Phyllostomidae was the most
represented with 428 indiviiduals. The comparative analysis between ecosystems
showed that there is non significant difference considering the bat fauna
(ANOSIM, R=0,001; p>0,05), and the cluster analysis comprising the for study
sites showed that there is similarity among some sites for both environments. The
time of activity of the most abundant species were not overlapped as well as there
was none significant correlation with precipitation. Probably, both ecosystems
work as different sources of food for the bat species. So, the “islands” of terra
firme would be the main source of fruits, since there is more availability of fruits in
this system, while mangrove could simply work for the frugivorous as a corridor of
acces to these “islands”. Artibeus jamaicensis was one of the species most
captured and showed a bimodal polyoestry reproductive pattern, furthermore this
was also the pattern of distribution of the lactant females during the year. The
reproductive success of the frugivorous bats seems to be related to the dry
season. The frugivorous guild was the most abundant with 64% of the captured
individuals, however, there was non significant difference between the number of
individuals of the guilds considering the environments, but only when the guilds
were compared with each other (ANOVA, F=693; gl=2; p<0,05). Finally, the high
rates of capture of the subfamily Stenodermatinae and the presence of Desmodus
rotundus in the study area could be feasible biological indicators of the
environmental quality of the terra firme stands at the Fazenda das Salinas.

37
1. INTRODUÇÃO
A classe Mammalia é considerada um grupo relevante por influenciar na
estrutura e composição da vegetação nas áreas em que habitam. De maneira
geral, os mamíferos são importantes em diversos acontecimentos, tais como
herbivoria (Xenarthra e Primates - Queiroz, 1995), atividades de polinização e de
dispersão de sementes (Primates – Pjil, 1982; Chiroptera – Zanzini, 2000;
Marsupialia e Chiroptera - Charles-Dominique, 1986). Algumas espécies também
podem servir como indicadores de produção primária (Queiroz, 1995) e/ou do
estado de conservação ambiental (Wilson et al., 1996; Marques-Aguiar et al.,
2002a). De modo geral, a influência no ecossistema pode ser preponderante até
mesmo quando se trata de espécies de menor tamanho corpóreo, como alguns
pequenos mamíferos terrestres (Carey & Harrington, 2001) e voadores (Richards,
1990a, b).
Entretanto para Pardini et al. (2003), certos fatores como a existência de
muitas espécies de hábitos noturnos, com áreas de vida relativamente grandes,
baixas densidades populacionais e até mesmo o tipo de habitat, podem influenciar
nos estudos ecológicos que abordem sobre estrutura, diversidade e dinâmica das
comunidades de mamíferos. Portanto, é provável que o baixo número de registros
de mamíferos em determinados ecossistemas como os manguezais, seja um
reflexo da influência de vários dos fatores citados anteriormente.
De acordo com Fernandes (2000), os mamíferos teriam 111 espécies de 14
ordens associados às áreas de mangue ao redor do mundo, o que representaria
7,5% do total de 1.467 espécies de vertebrados registrados nesse ecossistema.
Ainda para o mesmo autor, os quirópteros corresponderam a 15,5 % do total de
mamíferos com apenas 14 espécies e duas famílias (Pteropodidae e Molossidae),
constituindo o quarto grupo mais diversificado em áreas de mangue distribuídas
na Ásia, Oceania, costa oeste e leste das Américas e costa oeste e leste da
África.
Outro estudo realizado nos manguezais de Kimberley, na Austrália, por
McKenzie & Rolfe (1986), registrou a ocorrência de mais dezessete espécies de
quirópteros das seguintes famílias: Vespertilionidae, Hipposideridae, Molossidae e
Emballonuridae. No entanto, a ordem Chiroptera é, de acordo Linares (1987),
Koopman (1993), Altringham (1996) e Emmons (1997), a segunda maior ordem

38
da classe Mammalia com cerca de 950 espécies distribuídas em 186 gêneros e
duas subordens: Megachiroptera (uma família, Pteropodidae, do velho mundo) e
Microchiroptera (dezessete famílias de distribuição mundial, com exceção da
Antártida). Segundo Findley (1993), quase 90% dessas espécies seriam
exclusivamente neotropicais.
No Brasil, em outros ecossistemas florestais Amazônicos, a ordem
Chiroptera corresponderia a 40% da diversidade total de mamíferos, com
aproximadamente 124 espécies de morcegos e o maior número de endemismo
dentre os biomas brasileiros (Marinho-Filho & Sazima, 1998). Trabalhos recentes
de Bernard & Fenton (2002), Marques-Aguiar & Aguiar (2002), Marques-Aguiar et
al. (2002a); Marques-Aguiar et al. (2002b), Cáceres (2003), Fonseca (2003) e
Marques-Aguiar et. al. (2003), pontuam somente para o estado do Pará, um total
de 102 espécies, 56 gêneros e nove famílias (Emballonuridae, Noctilionidae,
Mormoopidae, Phyllostomidae, Molossidae, Thyropteridae, Vespertilionidae,
Furipteridae e Natalidae).
Na realidade, assim como as demais espécies de mamíferos, os morcegos
são pouco estudados em áreas de manguezal, o que certamente subestima a
quantidade de táxons associadas a este ecossistema. Silva Júnior et al., (2002),
relatou que para muitos mastozoólogos, os ecossistemas litorâneos brasileiros,
são extensões mais pobres em diversidade florística e faunística de áreas
continentais adjacentes. Em função desta visão equivocada, os manguezais
seriam florestas menos favoráveis para estudos dos aspectos ecológicos da
biodiversidade animal. Conseqüentemente, as relações e a importância dos
mamíferos e de muitos outros táxons animais, para com o referido ambiente
continuaria sendo negligenciada.
Contudo, na verdade os manguezais seriam importantes áreas de transição
que se encontram submetidas à interação entre os fatores continentais, marinhos
e atmosféricos, numa faixa de latitude entre 32°N e 38°S, em zonas costeiras
tropicais (Chapman, 1975; Schwamborn & Saint-Paul, 1996). O desenvolvimento
estrutural máximo dessas zonas estuarinas, aumenta na medida em que se
aproxima da linha do equador (Saenger & Snedaker, 1993; Schaeffer-Novelli,
1995).

39
Somente o litoral brasileiro, é constituído por cerca de 1,38 milhão de
hectares de manguezais ao longo de uma costa de aproximadamente 6.800 Km,
compreendendo uma das mais extensas áreas de manguezais no mundo (Kjerve
& Lacerda, 1993). Dentre toda essa dimensão litorânea do ecossistema, destaca-
se a costa norte, que representa cerca de 85% dos manguezais do Brasil
(Vanucci, 1999), dos quais 270.000 ha estão no litoral nordeste do Estado do
Pará (Senna & Mello, 1994). Mas para Proust et al. (1988) e Nittrouer et al.
(1991), os manguezais da costa norte apresentariam juntamente com os
manguezais da Guiana Francesa, níveis de destruição em larga escala.
Outras análises realizadas por Lara & Cohen (2003), também indicaram
uma perda de cobertura vegetal em áreas da zona costeira paraense de
aproximadamente 166 Km de comprimento, que inclui a planície costeira de
Bragança e áreas adjacentes. Para Souza Filho (2001), os principais problemas
ambientais da planície costeira bragantina estão relacionados à erosão e à
ocupação desordenada. No entanto, a costa norte brasileira, ainda é
caracterizada por Knoppers et al. (2002), como uma das áreas mais bem
preservadas e produtivas em termos de recursos naturais, que apresenta uma
interação dinâmica e equilibrada entre espécies vegetais e animais, abrigando
desde formas microscópicas até organismos maiores (Vanucci, 1999, Fernandes
2000).
1.1 Biologia e ecologia de morcegos
Em termos evolutivos, há 60 milhões de anos atrás durante o Eoceno, os
quirópteros, já eram semelhantes aos exemplares atuais, que estão
morfologicamente mais próximos das ordens Dermoptera (lêmures voadores),
Primates (prossímios, símios e macacos) e Scandentia (mussaranhos
arborícolas), formando um grupo supraordinal chamado Archonta, sendo os
morcegos um grupo derivado dos mamíferos eutérios (Pough et al., 1999). No
entanto, este autor aponta questões pouco esclarecidas sobre a ordem
Chiroptera, as quais envolvem o conhecimento do grupo de origem, as relações
entre os grandes grupos subordinados e também o de muitas famílias extintas e
atuais.

40
De modo geral, os representantes da ordem Chiroptera, conquistaram
diferentes regiões geográficas no decorrer de sua evolução, auxiliados por
adaptações que permitiram a utilização de uma gama bastante diferenciada de
recursos alimentares, tais como: néctar, pólen, sangue, frutos, insetos, peixes,
crustáceos e pequenos animais (Findley, 1993; Hill & Smith, 1986; Wilson et al.,
1996; Emmons, 1997).
Uma das mais interessantes estratégias adaptativas, encontrada
principalmente, entre os morcegos insetívoros e carnívoros do grupo
Microchiroptera é o chamado sistema de ecolocalização (Linares, 1987; Findley,
1993). Este consiste na transmissão de pulsos ultrassônicos devolvidos na forma
de ecos aos ouvidos, distinguindo para o animal o tamanho, tipo e velocidade de
vôo da presa, diferenciando entre os alimentos, os comestíveis ou não. Já para
outros grupos de quirópteros ocorre uma variação na eficiência visual, sendo que
determinadas espécies de morcegos frugívoros, em sua maioria pertencente à
subordem Megachiroptera, teriam visão ativa enxergando melhor do que os
insetívoros, devido aos primeiros apresentarem o sistema de ecolocalização
deficiente ou inexistente (Findley 1993; Pough et al., 1999).
Ainda sobre a biologia e ecologia de morcegos, de acordo com relatos de
Wilson (1973), haveria entre certas espécies tropicais uma estreita relação entre a
reprodução, disponibilidade alimentar e estação chuvosa, enquanto para os
quirópteros de regiões de clima temperado, esta relação seria direta e
decrescente, pois as fêmeas de muitas espécies de morcegos empregariam a
chamada diapausa embrionária durante a hibernação no inverno. De acordo com
Pough et al. (1999), este fenômeno estaria relacionado com a redução do
metabolismo, que envolve diretamente o desenvolvimento do feto, sendo essa
uma estratégia que acompanha a baixa disponibilidade de recursos alimentares
Em sendo assim, várias espécies de morcegos nas regiões tropicais
apresentariam comportamento reprodutivo do tipo sazonal, que seria muito mais
em função da dieta do que em função das atividades de forrageio (Hill & Smith,
1986). Durante a estação chuvosa, as florestas tropicais pluviais, normalmente
apresentam maior disponibilidade de frutos, que são apreciados por muitas
espécies de morcegos da família Phiylostomidae. No entanto, durante a estação
seca, para suprir a necessidade alimentar, e conseqüentemente auxiliar a
reprodução, algumas dessas espécies modificariam seus hábitos, passando a

41
uma dieta rica em pólen e flores. Outras espécies, por sua vez, passariam a
capturar insetos.
Contudo, certas espécies de quirópteros, como algumas do gênero
Artibeus de dieta alimentar a base de frutos, tanto realizam a diapausa
embrionária (Altringham, 1996), quanto à modificação da dieta em função da
atividade reprodutiva (Mikich, 2002). Dessa forma, são consideradas altamente
adaptadas a diferentes ambientes, apresentando o comportamento alimentar
categorizado como frugívoro generalista, este comportamento foi evolutivamente
favorecido por características como acessibilidade, disponibilidade sazonal e
composição nutricional variável (Fleming, 1986).
Independente da existência de sazonalidade na reprodução, cada ciclo
sexual das fêmeas de quirópteros, é dividido em quatro fases: pró-estro
(crescimento do folículo ovariano), estro (ovulação), meta-estro (regressão do
corpo lúteo) e diestro (período estacionário). Cada uma dessas fases é definida
por eventos ocorridos no ovário maduro, enquanto o sincronismo das mesmas é
controlado pela liberação de hormônios sexuais secretados no sangue pela
glândula pituitária (Pough et al., 1999). Esses hormônios pituitários são os
mesmos para ambos os sexos, os quais são produzidos de acordo com estímulos
externos (precipitação, temperatura e nutrição). Nos machos, a atuação desses
hormônios é refletida no processo de espermatogênese, com aumento e posterior
descida dos testículos para a bolsa escrotal, o que ocorre somente durante o
período de reprodução, sendo que fora deste período os testículos são
localizados na cavidade abdominal (Hill & Smitn, 1986).
Em zonas temperadas o período de diestria das fêmeas de morcegos é
prolongado por quase todo o ano, com a receptiva sexual das mesmas aos
machos ocorrendo apenas uma vez em cada ano (espécies monoéstricas). Nas
áreas tropicais as espécies são monoéstricas ou poliéstricas, mas de curto
período de diestria, tornando-se receptivas sexualmente várias vezes durante
uma estação reprodutiva, a qual, geralmente, coincide com o período chuvoso
(Findley, 1993). A maioria das fêmeas de morcegos apresenta taxa reprodutiva
mínima de uma cria a cada gestação, a qual pode variar de dois meses a um ano,
com o desenvolvimento pós-natal demorado. Esses filhotes, nos primeiros
estágios de vida, viajam agarrados à porção ventral das mães durante o vôo
(Altringham, 1996).

42
Por fim, todos as informações que envolvem biologia e ecologia de
morcegos no ecossistema manguezal, assim como dados de diversidade, são
escassas ou inexistentes, limitando-se tão somente às áreas de mangue das
regiões do Velho Mundo (Fernandes, 2000; Mckenzie & Rolfe, 1986). Dessa
forma, o presente estudo deverá contribuir para o conhecimento da dinâmica de
algumas dessas características que envolvam espécies de quirópteros
associadas às florestas de mangue e terra firme localizadas na Fazenda Salinas,
a qual pertence a grande área denominada manguezal bragantino na costa norte
brasileira.

43
2. OBJETIVOS
2.1. Objetivo Geral
Estudar a diversidade de espécies e alguns aspectos ecológicos da
quiropterofauna, com ênfase na família Phyllostomidae, associada aos
ecossistemas contíguos de manguezal e terra-firme, que compõem a paisagem
na parte central da península bragantina, no litoral paraense.
2.2. Objetivos específicos
- Fazer um inventário da quiropterofauna associada aos ecossistemas
manguezal e terra firme na Fazenda das Salinas;
- Estimar e comparar a diversidade de espécies, a abundância e a densidade da
comunidade de quirópteros entre esses ecossistemas;
- Avaliar a eficiência do esforço de captura, com base nas estimativas de
diversidade de espécies, abundância e densidade para os dois ecossistemas
estudados;
- Investigar a influência dos padrões de frutificação e precipitação sobre as
atividades alimentar e reprodutiva das guildas mais abundantes.
3. MATERIAL E MÉTODOS
3.1. Área de Estudo
O manguezal bragantino está localizado na costa leste do Estado do Pará,
a cerca de 150 Km ao sul da foz do rio Amazonas, próximo à cidade de Bragança
que está situada a 250 km da capital Belém. A área desse manguezal é de
aproximadamente 116 km2, acompanhada ao longo de sua extensão pelos 36 km
da PA-458. Essa Estrada foi construída para facilitar o acesso entre Bragança e a

44
“Vila dos Pescadores”, ou mais especificamente à praia de Ajuruteua,
aumentando assim o turismo local (Behling et al., 2001).
A composição vegetal do manguezal bragantino inclui um imenso mosaico
vegetacional, que de acordo com Mehlig (2001), apresenta extensas coberturas
por florestas de mangue divididas em ilhas e penínsulas, cortadas por canais e
“creeks” (Figura 1). Dentro desse mosaico também estão presentes dunas e
praias (na parte mais ao norte), áreas degradadas (na parte mais ao sul), algumas
ilhas de terra firme e manchas de campos salinos (na parte mais central),
dominados na sua maioria, por gramíneas. Esses dois últimos ecossistemas
juntamente com os bosques de mangue, constituem a propriedade particular,
denominada de Fazenda das Salinas, localizada entre as coordenadas
046°40’20,4”W e 0°55’27”S.
A Fazenda das Salinas, foi definida como a área para o desenvolvimento
do presente estudo (Figura 1). A mesma encontra-se a aproximadamente 20 Km
da cidade de Bragança, nas planícies mais elevadas da península bragantina,
sendo que as suas paisagens contíguas de campos salinos, florestas de mangue
e de terra-firme, são menos freqüentemente inundadas (<28 dias/ano, pelas altas
marés de sizígia), do que as extensas áreas colonizadas por manguezais (Lara &
Cohen, 2003).
As florestas de terra firme são representadas apenas por algumas “ilhas”
de composição vegetal heterogênea, com presença de espécies botânicas
arbustivas e arbóreas, que seriam de uma vegetação relíquia, originada da
transgressão dos manguezais e dos campos salinos em direção à terra firme
(Behling et al., 2001). De acordo com os estudos de Abreu et al. (submetido), a
estrutura arbórea da maior “ilha” de terra firme, dentre as presentes na Fazenda
das Salinas, possui altura máxima de 23 m (com uma média entre todos os
indivíduos de 7,9 m), extensão total de 33 ha e localização a 1,5 Km da PA-458
(Figura 1). Nessa mesma “ilha” foram fixados sítios de trabalho para o presente
estudo com quirópteros. Quanto a constituição vegetal, os autores identificaram
na mesma 71 espécies de 40 famílias, com alta dominância relativa da espécie
Simarouba amara Aubl (Marupá), pertencente à família Simaroubaceae. A maior
diversidade foi das famílias Myrtaceae, Arecaceae, Sapindaceae e
Lecythidaceae. Já o maiores valores de densidade absoluta, foram para

45
Arecaceae e Burseraceae. Através de comunicação pessoal de Fernandes,
também são incluídas palmeiras (Ericaceae) e embaúbas (Cecropiaceae), sendo
que, a presença de Cecropiacea é geralmente um forte indicativo de mata
secundária. Dados sobre o período de frutificação dessa mesma “ilha”, indicaram
dois picos de produção na estação seca (meses de agosto e dezembro), com
biomassa aproximada de 4 g.m-2.mês-1 (Nascimento, 2004).
Para representar o ecossistema manguezal neste estudo com quirópteros,
os bosques onde foram fixados os sítios de trabalho, estavam contíguos a “ilha”
de terra firme. Estes, localizaram-se nas planícies de lama cobertas por
vegetação típica de manguezais (Rhizophora mangle, Avicennia germinans e
Laguncularia), principalmente por Avicennia, que se propagam por 3 a 6 Km em
direção às planícies mais baixas, que são freqüentemente inundadas, passando
de uma área de bosques de mangues mistos, para um setor dominado por
Rhizophora (Lara & Cohen, 2003). Estes bosques de mangues aparentavam ser
bem antigos, devido à presença de muitas árvores mortas caídas, além da
estrutura vegetacional ser mais aberta permitindo a maior entrada de luz.A
caracterização climática da Fazenda das Salinas, foi esquematizada através da
construção de um diagrama climático, referente ao período de realização do
presente trabalho. Os valores de precipitação (mm) foram obtidos a partir da
Estação Meteorológica Automática, situada no Furo Grande (01o42´30´´S e
051o31´51´´W), que é monitorada pelo Departamento de Meteorologia da
Universidade Federal do Pará (UFPA). Também constam no mesmo diagrama,
as taxas de temperatura do ar (°C), coletadas utili zando-se sensor Data Logger,
programados para armazenar valores a cada uma hora em pontos na terra firme
e no manguezal da Fazenda das Salinas.
3.2. Procedimento de campo e tratamento das Espécimes
Para a garantir a captura de um maior número de espécies de quirópteros
na Fazenda das Salinas, as coletas foram realizadas através de três
procedimentos, sendo estes: a capturas diurnas, a capturas noturnas e a
capturas noturnas semestrais. O primeiro método foi fundamentado na busca
diurna em abrigos utilizando puçás, com a finalidade de adicionar táxons que são
dificilmente coletados com redes-neblina (mist nets), as quais foram utilizadas

46
nos demais procedimentos de captura. Essa técnica de coleta com redes-neblina
é altamente seletiva para determinadas espécies de morcegos, principalmente
frugívoros, da família Phyllostomidae (Voss & Emmons, 1996; Simmons & Voss,
1998; Milliken & Ratter, 1998; Sekiama et al., 2001).
Os procedimentos de coletas noturnas e noturnas semestrais, foram
desenvolvidos em períodos de lua nova ao longo de um ciclo anual. A principal
diferença entre os mesmos, inclui o tempo de trabalho de campo, o qual para as
“capturas noturnas” é constituído por seis campanhas realizadas a cada dois
meses, sendo cada uma composta por três noites de 7 hs diárias de trabalho de
campo (18:00 à 1:00 h), ao passo que o segundo procedimento das “capturas
noturnas semestrais”, foi dividido em duas campanhas, uma referente a estação
chuvosa e outra a estação seca, incluindo também três noites em campo, sendo
agora cada noite de 12 hs diárias (18:00 às 6:00 hs).
Os resultados obtidos pelo método de “captura noturna”, foram utilizados
na maioria das análises do presente estudo, enquanto os dados do terceiro
procedimento (capturas noturnas semestrais), serviram apenas na tentativa de
adicionar táxons, que apresentariam atividade noturna mais intensa ao
amanhecer. O intervalo de 30 dias entre as “capturas noturnas”, foi adotado no
intuito de minimizar a habituação dos morcegos à localização das redes, pois de
acordo com Bernard (2002), estes animais apresentam boa memória espacial e,
conseqüentemente, poderiam evitar áreas muito amostradas.
Para a exposição das redes-neblina, quatro transecções de 400 m cada
(T1, T2, T3 e T4), foram abertas e posicionadas com auxílio de GPS (Sistema de
Posicionamento Global por Satélite), modelo E-trex (Garmin), sendo as duas
primeiras na floresta de terra firme e as outras duas na floresta de mangue
(Figura 1). No meio de cada transecção foi fixado um sítio de trabalho (Tabela 1),
e para cada sítio eram destinadas quatro redes-neblina, medindo 3 m de altura x
7 m de comprimento, perfazendo um total de 112 m2 de rede por noite, que
permaneciam suspensas a três metros do solo (Figura 2). As mesmas eram
vistoriadas em intervalos de uma hora.
A disposição das redes dependeu da estrutura dos bosques em cada sítio
e da definição dos locais de maior facilidade para a passagem de quirópteros,
originando distribuições das redes em formatos “T” nos bosques mais abertos
(sítios #1 e #4) e “X” em bosques mais fechados (sítios #02 e #03).

47
Depois de capturados, os morcegos vivos foram individualmente mantidos
em sacolas de pano, obedecendo aos procedimentos citados por Auricchio
(2002), para obtenção das seguintes medidas morfométricas em campo:
comprimentos do antebraço, da folha nasal, da orelha e do trago (Figura 3).
Também foram feitas outras anotações referentes às coletas, tais como, data,
local, hora e número de captura, sexo, estado reprodutivo, faixa etária e peso. A
faixa etária dos indivíduos capturados, foi determinada através da consolidação
dos discos cartilaginosos nas articulações entre os metacarpos e as falanges
(Anthony, 1988). Para a determinação do estado reprodutivo das fêmeas, foi
seguida à caracterização de Racey (1988): jovens imaturas, grávidas, lactantes e
pós-lactantes.
A identificação dos espécimes foi realizada com base em chaves de
classificação (Handley, 1987; Linares, 1987; Marques-Aguiar, 1994; Emmons,
1997; Eisenberg & Redford, 1999; Gregorin & Taddei, 2002). Os exemplares de
identificação duvidosa, eram mortos, etiquetados e fixados utilizando-se solução
de formol 10% e glicerina 5%, para posterior conservação em solução de etanol
70%. Destes, cerca de 150 exemplares que incluíam as diversas espécies de
quirópteros capturadas, foram analisados pela Dra. Suely Marques Aguiar,
especialistas do Museu Paraense Emílio Goeldi, onde receberão o número de
tombo, representando assim, o material testemunho da Fazenda das Salinas.
Apenas um exemplar de cada espécie, permanecerá na coleção científica de
referência da península bragantina, localizada na Universidade Federal do Pará –
Campus, Bragança - Pará.

48
46°36’W
T3
T4
T1
T2
1°00’S
0°48’S
Figura 1 – Localização da área de estudo na península bragantina, destacando as quatro transecções
abertas na “ilha” de terra-firme e na área de manguezal. T1 a T3 = Transecção 1; T2 e T4 = Transecção
2. Modificado de Mehlig (2001).

49
Ecossistemas Sítios Longitude (W) Latitude (S)
Terra-firme
Manguezal
#01
#02
#03
#04
046° 40’ 11,5”
046° 40’ 14,4”
046° 40’20,4”
046° 40’ 23,2”
0° 55’ 21,3”
0° 55’ 33,4”
0° 55’ 26,7”
0° 55’ 43,3”
A
Sítio#4
Figura 2 – Desenho esquemático da instalação da rede-neblina. Fonte: Auricchio
(2002).
Figura 3 – Principais medidas morfométricas que foram obtidas para a identificação dos
indivíduos capturados: comprimento do trago (T), da orelha interna e externa (O) e do antebraço
(A). Modificado de Auricchio (2002).
Tabela 2 - Coordenadas geográficas dos sítios de trabalho nos dois ecossistemas na
Fazenda das Salinas, Bragança-Pará.

50
3.3. Análise dos Dados
3.3.1. Abundância de Indivíduos, Espécies e Esforço de Captura
Neste trabalho foi utilizada a abundância (número total de indivíduos
capturados), para a análise do grau de associação em nível de indivíduos e
espécies por ambiente amostrado (terra firme e manguezal), bem como para os
ambientes agrupados (Fazenda das Salinas). O esforço de captura foi avaliado
dos ambientes separadamente e agrupados, utilizando-se valores calculados da
Densidade por Indivíduos (DI), Densidade por Espécie (DE) e da razão entre o
total de Espécies e das Capturas (REC), obtido pelas seguintes fórmulas:
DI = N° total de indivíduos / hora. rede
O nível de associação do total dos quirópteros capturados em cada
ecossistema e nos sítios de trabalho, foi avaliado através da matriz de
similaridade gerada pelo teste estatístico ANOSIM do programa Primer v5 (Clarke
& Gorley, 2001). Este teste unifatorial utiliza a abundância dos táxons de cada
subamostra, sendo a matriz gerada com os dados transformados pela raiz
quadrada, utilizando-se o coeficiente de similaridade de Bray-Curtis, o que
segundo Clarke & Gorley (2001), são as melhores escolhas para gerar uma matriz
de abundância de espécies.
Ainda no mesmo programa, com a análise de cluster (UPGMA), foi obtido
um dendrograma a partir da matriz de similaridade dos sítios de trabalho, baseado
DE = N° total de espécies / hora. rede
REC = N° total de espécies / N° total de capturas

51
na abundância total dos indivíduos. O dendrograma foi construído com a
utilização do modo de agrupamento “Group Average”, isto é, permitindo a
maximização da correlação de similaridade entre as amostras (van Tongeren,
1995).
3.3.2. Estimativas de Riqueza
A quiropterofauna da Fazenda das Salinas foi estimada ao nível de riqueza
em espécies, sendo para isso, necessário a utilização de métodos paramétricos e
não-paramétricos, amplamente aplicados em estudos de diversidade biológica,
pois os mesmos permitem comparações entre conjuntos de dados amostrais,
obtidos com metodologias e esforços diferentes. A seguir, esses métodos são
apresentados e descritos, de acordo com as bases conceituais de Santos (2003).
O único método paramétrico utilizado foi à curva de acumulação, ou curva
do coletor destinada à avaliação do quanto o inventário se aproximou de capturar
todas as espécies do local estudado. Quando a curva torna-se estável, isto é,
atinge um ponto em que o aumento do esforço de coleta não implica em aumento
do número de espécies, significa que toda a riqueza da área foi amostrada. A
aplicação da equação de “Michaelis-Menten Mean” (MMMean), permite estimar a
assíntota a partir de qualquer ponto da curva de acumulação construída, ou seja,
prevê o ponto de estabilização desta curva, além de ser um modelo facilmente
ajustável aos diferentes conjuntos de dados.
No caso do método não-paramétrico o ACE (Abundance-based Coverage
Estimator), serviu para a avaliação dos acontecimentos através da proporção de
espécies raras, isto é, espécies representadas por apenas um único indivíduo na
amostra, enquanto o ICE (Incidence-based Coverage Estimator), trabalha com a
proporção das espécies infrequentes ou espécies que ocorrem em apenas uma
unidade amostral. Esses dois métodos (ACE e ICE), permitem determinar os
limites para que uma espécie seja considerada rara ou infrequente.
Outros dois métodos também não-paramétricos, recentemente adaptados
para a estima de riqueza são o Chao1 e Chão 2. A riqueza estimada por Chao1 é
igual à riqueza observada, somada ao quadrado do número de espécies
representadas com “singletons” (espécies representadas por um único indivíduo
nas amostras), dividido pelo dobro do número de espécies representados por

52
dois indivíduos (doubletons). O Chao1 somente atinge valor máximo quando
todas as espécies, menos uma, são “singletons”. Enquanto o método de Chao2
utiliza o número de espécies que ocorrem em uma ou duas unidades amostrais,
ditas como “uniques” e “duplicates”, respectivamente. Este método apresenta
valor máximo quando todas ás espécies, menos uma, são “uniques”.
O método Jacknife 1 foi bastante útil na estimativa da riqueza total,
esclarecendo se a riqueza estimada é igual ou não a observada. Para tanto é
somada a riqueza observada (número de espécies coletadas) a um parâmetro
calculado a partir do número de espécies raras e do número de amostras que
deve ser menor do que 20. Desse modo, o método atinge valor máximo de
estimativa, se todas as espécies, menos uma, são “uniques”.
Entretanto, o “Bootstrap”, ao contrário dos métodos apresentados acima,
utiliza informações de todas as espécies coletadas para estimar a riqueza total,
não se restringindo apenas às espécies raras. Calcula-se este índice somando a
riqueza observada ao total do inverso da proporção de amostras em que ocorre
cada táxon (Smith & van Belle, 1984). Mesmos devendo ser esse número de
amostras maior do que 20 amostras, o valor do Boostrap foi gerado durante o
cálculo no programa EstimateS 6.0b 1(Colwell, 1997), sendo assim, foi
acrescentado no resultado do presente estudo apenas ao nível de informação.
Enquanto as demais estimativas dos métodos anteriormente citados foram
realizadas através do programa EstimateS 6.0b 1, a curva de acumulação de
espécies foi produzida utilizando-se o programa Excel-2000 v.7.0.
3.3.3. Estimativas de Diversidade
Neste estudo o conceito de diversidade de espécie, é o mesmo de
Lewinsohn & Prado (2002), que refere-se à variedade de espécies dentro de
algum tipo de ambiente ou região definida, maior ou menor.
3.3.3.1. Índice de Shannon-Wiener (H’):

53
Índice baseado na teoria da informação, muito empregado em
estimativas de diversidade (Magurran, 1988). O índice de Shannon-Weiner é
representado pela seguinte fórmula:
H’ = - Σ (ni / N) ln (ni / N)
Onde:
ni = no total de indivíduos da espécie i;
N = no total de indivíduos na amostra;
ni/N = pi abundância proporcional de cada espécie na amostra total.
A Equitabilidade (E) é um componente da diversidade, que reflete o grau
de uniformidade com que os indivíduos estão distribuídos entre as várias
espécies da amostra por ambiente, podendo ser um indicativo de dominância
numérica relativa (Zanzini, 2000; Magurran, 1988). A equitabilidade possui
valores de 0 (uniformidade nula) até 1 (uniformidade máxima), que podem ser
obtidos através do seguinte cálculo:
E = H’ / In S
Onde:
S = no total de espécies na amostra;
H’ = valor obtido do índice de Shannon-Wiener.
3.3.3.2. Índice de Simpson (D):
De acordo com Zanzini (2000), assume valores de 0 a 1 e fornece a
estimativa da probabilidade de dois indivíduos, retirados aleatoriamente de uma
amostra, pertencerem à mesma espécie. Portanto, quanto maior o valor assumido
pelo índice, menor a diversidade da amostra. O índice de Simpson (D) é dado
pela seguinte fórmula:

54
D = Σ n (n – 1) / N (N – 1)
Onde:
D = estimativa da probabilidade de que dois indivíduos selecionados ao acaso
sejam da mesma espécie;
ni = nº de indivíduos da i-ésima espécie;
N = nº total de indivíduos na amostra.
A análise inversa do índice de Simpson (1/D) fornece o resultado numérico
das espécies comuns. Dessa forma, as observações das comunidades podem ser
feitas de modo mais geral (Krebs, 1989).
Os índices de diversidade foram calculados utilizando-se os programas
Excel-2000. v.7.0. e Species Diversity & Richness-v. 2.5 (Henderson & Seaby,
1997).
3.3.4. Cálculos de Similaridade
Para a comparação quanto ao número de espécies entre os sítios de
trabalho, foram calculados os índices qualitativos de Jacacard (CJ) e Sorenson
(CS). Estes índices foram desenhados para igualar a 1 (um), nos casos de
completa similaridade (onde os ambientes comparados apresentam o mesmo
número de espécies) e a 0 (zero) se estes ambientes são dissimilares (Magurran,
1988). Todos os cálculos foram feitos manualmente. As fórmulas para Jacacard e
Sorenson são as seguintes:
C(Jaccard) = c/(a+b-c) C(Sorenson) = 2c/a+b
Onde:
a = número de espécies presentes na amostra a;

55
b = número de espécies presentes na amostra b;
c = número de espécies comuns às amostras a e b.
3.3.5. Análise de Classificação
Através da análise multivariada de cluster foi gerado, dos dados totais de
todas as amostras, um dendrograma dos sítios de trabalho baseado na presença
e ausência das espécies. O pacote estatístico Primer v5 foi utilizado para este fim
(Clarke & Gorley, 2001).
3.3.6. Período de Atividade das Espécies mais Abundantes
Para investigar a relação entre os horários de intensa atividade noturna e a
flutuação no número de indivíduos ao longo do ciclo anual das espécies mais
abundantes, foi utilizada a análise de variância do tipo ANOVA (bifatorial).
Previamente, os dados brutos foram testados quanto à normalidade (método de
Lilliefors), para em seguida, de acordo com Centeno (1999), ser realizada a
transformação dos mesmos (logaritmo natural ou a raiz quadrada). O teste de
Tukey-HSD (Honestly Significant Difference), serviu para a verificação da ordem
de significância dos parâmetros avaliados. Essas análises foram feitas utilizando-
se o pacote estatístico BioEstat 3.0 (Ayres et al., 2000)
3.3.7. Razão Sexual
Para avaliar a razão sexual das espécies mais abundantes foi utilizada a
análise não-paramétrica do Qui-quadrado, no qual os valores esperados possuem
uma regência ou padrão conhecido representado pela proporção de 1:1 ou 50%
para cada acontecimento (Centeno, 1999). Os cálculos também foram realizados
no programa BioEstat 3.0.
3.3.8. Flutuações Sazonais das Principais Guildas
A definição das guildas obedeceu aos dados da literatura (Hill & Smith,
1986; Findley, 1993; Wilson et al., 1996; Emmons, 1997), sendo que as mais
abundantes no presente trabalho, foram relacionadas com os ecossistemas (terra

56
firme e manguezal) e com os quatro sítios de trabalho, utilizando-se ANOVA
(bifatorial). O teste de normalidade dos dados, foi verificado através do método de
Lilliefors no programa BioEstat 3.0 (Ayres et al., 2000). No mesmo programa
estatístico foi gerado o teste de Tukey-HSD (Honestly Significant Difference), para
verificar a ordem de significância dos parâmetros avaliados. Na busca de
prováveis padrões sazonais para os indivíduos das diferentes guildas, foi
realizada uma análise de Regressão Linear correlacionando a proporção entre
machos e fêmeas. E por fim, também foi correlacionado o número de fêmeas
grávidas e lactantes das guildas mais abundantes com as variações sazonais de
precipitação e frutificação ao longo do ciclo anual. Os valores médios mensais de
frutificação (g.m-2.mês-1) são oriundos dos estudos de Nascimento (2004).
3.3.9. Similaridade da Quiropterofauna no Estado do Pará
Apenas em nível de comparação, foram utilizadas listas de quirópteros de
outras cinco localidades no Estado do Pará, obtidas pela mesma técnica de coleta
noturna, mas em diferentes espaços de tempo: 1) Belém (Fonseca, 2003); 2)
Paragominas (Cáceres, 2003); 3) Caxiuanã: Melgaço (Marques-Aguiar & Aguiar,
2002); 4) Alter do Chão (Bernard & Fenton, 2002); 5) Xingu (Marques-Aguiar et
al., 2002a) e 6) Marajó: entre Anajás e Muaná (Marques-Aguiar et. al., 2002b). A
planilha utilizada para esta análise encontra-se no Anexo 1. O dendrograma foi
gerado pela análise de cluster no pacote estatístico Primer v.5 (Clarke & Gorley,
2001).

57
4. RESULTADOS
4.1. Abundância de Indivíduos e Espécies
As campanhas noturnas bimestrais, empreendidas das 18:00 até 01:00 h
na Fazenda das Salinas, resultaram em 438 capturas. Deste total, 217 indivíduos
de 14 espécies foram obtidos na terra firme, enquanto que no manguezal foram
registrados 221 indivíduos de 18 espécies (Tabela 2 e Anexo 2).
A família Phyllostomidae representou 98% de todas as capturas com 428
indivíduos de 18 espécies, enquanto as outras quatro famílias, individualmente,
não constituíram mais do que 2% das capturas (Figura 4A). Os filostomídeos da
área de estudo se dividiram em cinco subfamílias, encontradas tanto na floresta
de terra firme quanto no manguezal (Figura 4B). Dentre elas destacam-se a
subfamília, Stenodermatinae com 65%, seguida por Carollinae (23%),
Glossophaginae (7%), Phyllostominae (4%) e Desmodontinae com apenas 1%
das capturas.
A subfamília Stenodermatinae foi a mais representativa, principalmente,
pela proporção de indivíduos capturados das espécies Artibeus jamaicenses e
Artibeus obscurus, compreendendo 29% e 25,6% de todas as capturas, Carollia
perspicillata (subfamília Carolliinae) representou 22%, Glossophaga soricina
(subfamília Glossophaginae) totalizou 6,8%, enquanto as demais subfamílias
(Desmodontinae e Phyllostominae) representaram um total de apenas 5% (Tabela
2).
4.1.1. Esforço de Captura de Indivíduos e Espécies
As taxas de capturas foram as mesma tanto para o manguezal quanto para
a terra firme, de 0,22 ind/hora.rede, sendo também este o valor registrado na
avaliação da Fazenda das salinas somadas todas as suas capturas (Tabela 3). A
análise comparativa entre os dois ecossistemas, mostrou não haver diferença
significativa no que diz respeito as associações desses ambientes com o total de
espécimes de morcegos capturadas (ANOSIM, R=0,001; p>0,05).

58
Talvez, os sítios devam ser estruturalmente tão diferentes dentro de cada
ecossistema, que a análise utilizando o total de capturas não registrou diferenças
entre os ecossistemas. Sendo assim, foi realizada a comparação entre os valores
de DI dos quatro sítios de trabalho. Os resultados mostraram diferenças
significativas entre os sítios #01 e #02, ambos da terra firme (ANOSIM, R=0,014;
p<0,001); entre os sítios #01 da terra firme e #03 do manguezal (ANOSIM,
R=0,009; p<0,05); e entre os sítios #02 e #04, terra firme e manguezal,
respectivamente (ANOSIM, R=0,006; p<0,05). Estes resultados são corroborados
com a análise de cluster envolvendo os sítios, cujos agrupamentos mostraram os
sítios #02 e #03 são os mais similares quanto ao número de indivíduos
capturados (coeficiente de Bray-Curtis = 99,69) e os sítios #01 e #04 os mais
distantes do Sítio#02, com 99,26 e 99,52, respectivamente. Por fim, como não
foram encontradas diferenças significativas entre os ecossistemas, e os valores
de similaridade do número de espécimes capturados entre os sítios estão acima
de 99%, o mais provável é que os mesmos sejam visitados de modo semelhantes
(Figura 5).
Na densidade por espécie (DE), o esforço empreendido em toda a área de
estudo foi de 0,011 espécies/hora.rede, enquanto para cada ambiente, a
eficiência do esforço distribuiu-se em 0,014 espécies/hora.rede na terra firme e
0,018 espécies/hora.rede no manguezal (Tabela 3). O Sítio#01 foi o que mais
contribuiu para a DE da terra firme com 0,026 espécies/hora.rede, enquanto para
os sítios do manguezal (Sítios #03 e #04) os valores foram quase equivalentes
(0,026 e 0,028 espécies/hora.rede). O Sítio#02 foi o que apresentou menor
esforço.
A avaliação da razão entre o total das espécies e das capturas (REC) na
terra firme foi de 0,064 espécies/captura, destacando-se o Sítio#02 (0,13
espécies/n°.capturados). Apesar do sucesso de captu ra das espécies por
hora.rede, ter sido menos sucedido para o manguezal, este ecossistema
apresentou 0,081 espécies/captura, sendo principalmente auxiliado pelo valor de
0,13 espécies/ n°.capturados no Sítio#03 (Tabela 3) .

59
Tabela 2 – Caracterização da quiropterofauna da Fazenda das Salinas, incluindo informações taxonômicas, nomes vulgares no Brasil
(Emmons, 1997; Wilson et al.,1996; Silva, 1994), guildas (Emmons, 1997; Findley, 1993; Hill & Smith, 1986; Wilson et al., 1996),
categoria de conservação (Fundação Biodiversitas, 2002; Ministério do Meio Ambiente, 2003), grau de ocorrência na área de estudo
(Marques-Aguiar et al., 2002a) e total de indivíduos por espécies capturadas nos ecossistemas manguezal e terra firme. (+) = rara, de
1 a 5 indivíduos; (++) = baixa, de 6 a 10 indivíduos; (+++) = comum, acima de 10 (Marques-Aguiar et al., 2002a).
TÁXONS
NOME
VULGAR
GUILDAS
STATUS DE CONSERVAÇÃ
O
OCORRÊNCIA
ECOSSISTEMAS
TOTA
L
Abundânci
a relativa
(%) M TF
EMBALLONURIDAE
Saccopteryx leptura
(Schreber, 1771)
Morc. de
duas
linhas
Insetívora
Estável
(+)
-
01
01
0,2
MOLOSSIDAE
Molossus molossus (Pallas,
1766)
Morcego
das casas
Insetívora
Estável
(+)
03
-
03
0,7
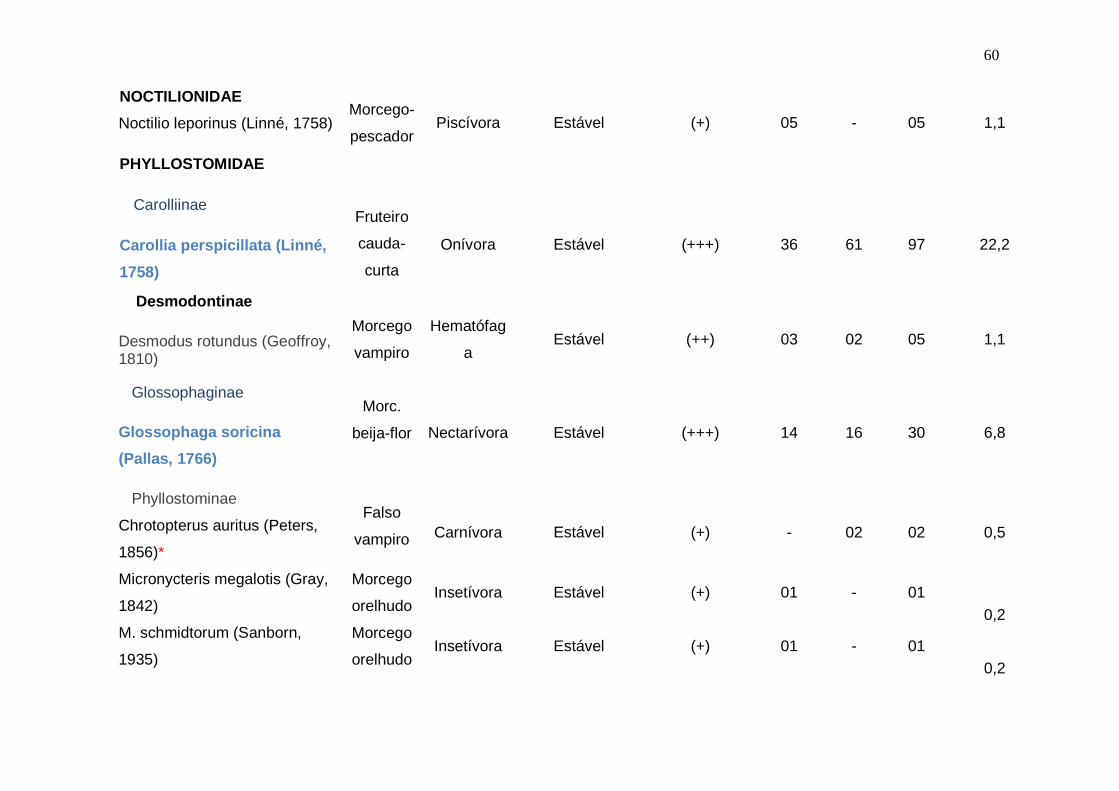
60
NOCTILIONIDAE
Noctilio leporinus (Linné, 1758)
Morcego-
pescador
Piscívora
Estável
(+)
05
-
05
1,1
PHYLLOSTOMIDAE
Carolliinae
Carollia perspicillata (Linné,
1758)
Fruteiro
cauda-
curta
Onívora
Estável
(+++)
36
61
97
22,2
Desmodontinae
Desmodus rotundus (Geoffroy, 1810)
Morcego
vampiro
Hematófag
a
Estável
(++)
03
02
05
1,1
Glossophaginae
Glossophaga soricina
(Pallas, 1766)
Morc.
beija-flor
Nectarívora
Estável
(+++)
14
16
30
6,8
Phyllostominae
Chrotopterus auritus (Peters,
1856)*
Falso
vampiro
Carnívora
Estável
(+)
-
02
02
0,5
Micronycteris megalotis (Gray,
1842)
Morcego
orelhudo Insetívora Estável (+) 01 - 01
0,2 M. schmidtorum (Sanborn,
1935)
Morcego
orelhudo Insetívora Estável (+) 01 - 01
0,2

61
Mimon crenulatum (Geoffroy,
1810)
Morc.
nariz-
peludo
Insetívora Estável (+) 02 - 02 0,5
Phylloderma stenops (Peters,
1865)
Morc.
nariz de
arpão
Onívora Estável (++) 05 02 07
1,6
Tabela 2 – Continuação...
Tonatia saurophila (Koopman
& William, 1951)
Morc.
orelha-
redonda
Insetívora Estável (+) 01 02 03 0,7
Trachops cirrohosus (Spix,
1823)
Morc.
lábio-de-
franja
Carnívora Estável (+) 01 - 01 0,2
Stenodermatinae
Artibeus obscurus (Schinz,
1821)
Morc.
fruteiro-
grande
Frugívora Estável (+++) 58 54 112 25,6
Artibeus cinereus (Gervais,
1855)
Morc.
fruteiro Frugívora Estável (+++) 04 22 26 5,9
Artibeus jamaicensis (Leach,
1821)
Morc.
fruteiro
comum
Frugívora Estável (+++) 82 45 127 29,0
23

62
Chiroderma villosum (Peters,
1860)
Morc.
Grande
de linhas
brancas
Frugívora Estável (+) 01 - 01 0,2
Platyrrhinus helleri (Peters,
1867)
Frugívoro
de linhas
brancas
Frugívora Estável (+) - 03 03 0,7
Sturnira lilium (Geoffroy, 1810) Morc.
frugívoro
pequeno
Frugívora Estável (+) - 01 01 0,2
Uroderma bilobatum (Peters, 1866)
Morcego
construtor Frugívora Estável (++) 02 05 07 1,6
Uroderma magnirostrum
(Davis, 1968)
Construto
r amarelo Frugívora Estável (+) 01 - 01 0,2
Vampyressa bidens (Dobson, 1878)
Morc.
orelha-
amarela
Frugívora Estável (+) - 01 01 0,2
VESPERTILIONIDAE
Myotis albescens (Geoffroy,
1806)
Morc.
borboleta-
prata
Insetívora Estável (+) 01 - 01 0,2

63
TOTAL
221 217 438 100
N°. DE ESPÉCIES 18 14 23
N°. DE ESPÉCIES
EXCLUSIVAS
09 05
24

64
Tabela 3 - Proporções de capturas realizadas com redes-neblina em dois
ecossistemas, manguezal e terra firme, e seus respectivos sítios de
trabalho na Fazenda das Salinas, Bragança, Pará. H.R = total de horas de
campo x o número de redes abertas; Ni = número total de indivíduos
capturados; Ne = número total de espécies capturadas; DI = razão entre Ni
e hora.rede, que representa a taxa de espécimes capturas; DE = razão
entre Ne e hora.rede, que representa a taxa de espécies capturas; REC =
razão entre Ne e Ni (espécies/captura).
Ambiente Sítios H.R Ni Ne DI DE REC
Terra firme (TF) 1008 21
7
14 0,22 0,014 0,065
Sítio#01 504 14
2
13 0,28 0,026 0,092
Sítio#02 504 75 10 0,15 0,020 0,15
Figura 4 – Percentual de famílias de quirópteros (A) e subfamílias de Phyllostomidae (B) capturados na
Fazenda das Salinas em Bragança - Pará.
98%
51%49%
0102030405060708090
100
Famílias
Terra firme (TF)Manguezal (M) M + TF
65%
23%
7%4%
1%
34.6%30.1%
0102030405060708090
100
Percentual de Indivíduos
Sufamílias
Pe
rce
ntu
al
A B
Subfamília

65
Manguezal (M) 1008 22
1
18 0,22 0,018 0,082
Sítio#03 504 99 13 0,20 0,026 0,13
Sítio#04 504 12
2
14 0,24 0,028 0,12
Fazenda
(M+TF)
2.016 43
8
23 0,22 0,011 0,053
S 1
S4
S3
S 2
99.0 99.1 99.2 99.3 99.4 99.5 99.6 99.7 99.8 99.9 100.0Coeficiente de Similaridade de Bray-Curtis
4.2. Estimativa de Riqueza em Espécies
Para a captura de 438 indivíduos pertencentes a 23 táxons da
quiropterofauna local, utilizou-se um esforço de 2.016 hora.rede. Analisando a
curva de acumulação dos três ambientes (Figura 6 A e B), observou-se que os
valores encontrados estão relativamente distantes de alcançar a assíntota da
curva, mas que a fauna de quirópteros em cada ambiente foi, razoavelmente
capturada, sendo talvez, apenas necessária à continuidade das coletas, dentro do
mesmo esforço empregado neste trabalho, para alcançar os valores estimados de
riqueza em espécies de morcegos na Fazenda das Salinas.
Os estimadores de riqueza de espécies para os dados agrupados
(Fazenda das Salinas), predizem valores entre 23 e 38 espécies esperadas para
a área de estudo, colocando as capturas num patamar entre 61 e 100%,
Figura 5 – Dendrograma mostrando a similaridade entre os quatro sítios de trabalho, com
base nos seus respectivos valores de abundância.
Sítio#02 Sítio#03 Sítio#04
Sítio#01

66
respectivamente (Tabela 4). Para cada ambiente os resultados obtidos com os
mesmos estimadores, registraram uma variação entre 78 e 100% das espécies de
morcegos esperadas para a terra firme e entre 60 e 90% para o manguezal.
Quanto a diversidade de espécies, os índices de Shannon-Wiener (H’) e
Simpson (D), resultaram em valores aproximados para os dois ecossistemas,
sendo H’ = 1,84 e 1-D = 0,80 para a terra firme, e H’ = 1,80 e 1-D = 0,76 para o
manguezal. A diversidade geral da área de estudo, foi novamente calculada pelo
agrupamento dos dados de cada ambiente, e os resultados encontrados
mostraram que H’ = 1,90 e 1-D = 0,79.
0
7
14
21
28
0 100 200 300 400 500
N°. de Espécies
N °. de Indivíduos
0
7
14
21
28
0 50 100 150 200 250
N°. de Espécies
N°. de Indivíduos
M
TF
( A )
( B )
M + TF
Figura 6 - Curva de acumulação das espécies capturadas: (A) nos sistemas agrupados e (B) nos
ecossistemas separados: M = manguezal e TF = terra firme, na Fazenda das Salinas, Bragança - Pará.

67
O dendograma da Figura 7, evidenciou para os quatro sítios estudados,
uma maior similaridade, quanto à presença/ausência de indivíduos, entre os sítios
#01 e #02 (78,26), ou seja, maior semelhança entre os sítios caracterizados por
vegetação de terra firme, sendo estes dois, em conjunto, mais similares ao
Sítio#03 (64,4) do que ao Sítio#04 (58,95). Além do mais, dos 18 táxons obtidos
no ecossistema manguezal, nove foram exclusivos desta floresta, ao passo que
na área de terra firme das quatorze espécies capturadas, nove eram comuns para
os dois ecossistemas. A análise de similaridade entre esses ecossistemas
mostrou valores bastante altos, isto é, Jaccard (Cj) = 64,28% e Sorenson (CS) =
78,26% (Tabela 5).

68
Tabela 4 - Estimativas de riqueza em espécies para a Fazenda das Salinas. DV = Desvio Padrão; % complementar =
percentual das espécies obtidas no presente trabalho, diante do esperado por cada estimador; ACE = Abundance-based
Coverage Estimator; ICE = Incidence-based Coverage Estimator; MMMean = Michaelis-Menten Mean. Os cálculos foram
realizados através do programa EstimateS 6.0b 1.
Estimativas
Dados Agrupados
(N=23)
±DV
% complementa
r
Terra firme (N=14) ±DV
% complementa
r
Manguezal (N=18) ±DV
% complementar
ACE 34 1,73 68
16
0,88 86 25 1,4 72
ICE 35 0,02 66
18
0,01 78 30 0,04 60
Chao1
35 0,20 66 15 1,77 93 25 0,13 72
Chao2 38 0,24 61
14
1,46 100 28 0,16 64
Jackknife1
33 3,8 70
17
1,64 82 26 3,7 69
Boostrap 27 0,68 85 16 0,35 86 21 0,9 86

69
MMMean 23 100
16
86 20 90
28

70
S4
S3
S1
S2
50 60 70 80 90 100Coeficiente de Similaridade de Bray-Curtis
Tabela 5 - Índices de diversidade calculados para os
ecossistemas separadamente e agrupados. 1 – D =
Coeficiente de diversidade de Simpson; 1 / D = Nº de
espécies comuns para gerar o coeficiente.
ÍNDICES Sistemas
Agrupados Terra firme Manguezal
Número de espécies
23 14 18
Shannon-Wiener (H’)
1,90 1,84 1,80
Equitabilidade (E’)
0,60 0,59 0,62
1 – D
0,79 0,80 0,76
1 / D 4,86 5,07 4,24
Figura 7 – Dendrograma mostrando a similaridade entre os quatro sítios de
trabalho, com base na presença/ausência de indivíduos de cada espécie.
Sítio#02 Sítio#01 Sítio#03 Sítio#04

71
4.3. Período de Atividade das Espécies mais Abundantes
Somente cinco espécies de quirópteros coletadas na Fazenda das Salinas,
apresentaram abundância suficiente para a análise do período de atividade
noturna (Figura 8). Todos os indivíduos dessas espécies resultam no total de 392
capturas. Da subfamília Stenodermatinae, a espécie Artibeus jamaicensis
apresentou dois picos, um mais intenso na terceira hora da noite (21:00 hs) e
outro menor, à 1:00 h da manhã (Figura 8A). Enquanto A. obscurus concentrou as
atividades às 20:00 hs, seguida por uma aparente homogeneidade na captura dos
indivíduos, ao longo da noite (Figura 8B). O pico de atividade da espécie Carollia
perspicillata, subfamília Carollinae, ocorreu ao longo das três primeiras horas
após o pôr-do-sol, seguido por um decréscimo a partir dás 21:00 hs, e
posteriormente, apresentando uma certa uniformidade para o resto da noite
(Figura 8C). Já a atividade para a espécie Glossophaga soricina (Figura 8D), da
subfamília Glossophaginae, parece ter se concentrado na primeira hora da noite
(19:00 hs). E, finalmente, a espécie Artibeus cinereus, cujas seis primeiras horas,
apresentaram duas oscilações de baixa densidade, sendo que o maior pico
ocorreu apenas à 1:00 h da manhã (Figura 8E).
Para a avaliação de variações populacionais, das espécies mais
abundantes de quirópteros da Fazenda das Salinas (Artibeus jamaicensis, A.
obscurus e C. perspicillata), tendo como base as estações do ano (chuvosa e
seca), foi construído um diagrama climático utilizando-se as taxas de temperatura
e chuvas (Figura 9). De modo geral, a área é caracterizada por um clima bi-
sazonal, com estação chuvosa no período de janeiro a junho, acompanhada de
estação mais seca de agosto a novembro, além de temperatura média anual alta
de 27.7±1.5°C.
A espécie Artibeus jamaicensis apresentou dois picos de atividade, bem
evidentes durante a estação chuvosa, sendo um às 21:00 e outro às 23:00 hs,
(Figura 10A). Também foram registrados nas duas estações do ano, para A.
obscurus e Carollia perspicillata, picos de atividade em diferentes horários (Figura
10 B e C). Sendo que, no decorrer da estação chuvosa, no primeiro horário (18:00
às 19:00 hs), a espécie C. perspicillata apresentou o maior número de capturas

72
(Figura 10C), enquanto o segundo horário (19:00 às 20:00 hs), destacou A.
obscurus como a espécie com maior número de registros (Figura 10B).
Per
cent
ual d
e C
aptu
ras
(%)
Figura 8 – Atividade noturna das cinco espécies de morcegos mais abundantes na Fazenda das
Salinas, de acordo com os horários de captura, durante o período de dezembro de 2002 a outubro de
2003. Para tanto, foram utilizadas redes-neblina, abertas de 18:00 à 1:00 h, totalizando sete horas
diárias de campo.
Artibeus obscurus
0
10
20
30
Carollia perspicillata
0
10
20
30
Glossophaga soricina
0
5
10
15
20
Artibeus cinereus
0
2
4
6
8
19:00 20:00 21:00 22:00 23:00 0:00 1:00
Artibeus jamaicensis
0
10
20
30
40
A
E
D
C
B
Horário de Captura

73
Em seguida, no terceiro horário, o maior número de indivíduos registrado
foi de A. Jamaicensis. É importante observar, que não ocorreu sobreposição dos
picos de atividade noturna entre as três espécies na duas estação. A exclusão
das espécies G. soricina e A. cinereus para essa análise, foi em função dos
baixos valores numéricos, observados após a distribuição do total de capturas nas
respectivas estações (chuvosa e seca), o que poderia não assegurar os
resultados para as espécies.
De maneira geral, as variações no número de indivíduos das espécies
avaliadas, referentes aos horários de atividade noturna (18:00 à 1:00 h) nas duas
estações (chuvosa e seca), foi significativa para a espécie A. jamaicensis, tanto
entre as estações (ANOVA, F=360; gl=1; p<0,001) quanto entre os horários de
atividade (ANOVA, F=56,6; gl=6; p<0,05). Para a espécie A. obscurus, a única
variação significativa ocorreu na avaliação da abundância entre as estações
(ANOVA, F=62,5; gl=1; p<0,05), ao passo que para C. perspicillata não houve
variação significativa (ANOVA, p<0,05) para nenhum dos acontecimentos acima
considerados.
Figura 9 – Diagrama climático da área de estudo, representando a flutuação da
Temperatura (°C) e da Precipitação (mm) ao longo do período de estudo.

74
PE
RC
EN
TU
AL
DE
CA
PT
UR
AS
(%
)
Figura 10 – Horário de captura nas duas estações (chuvosa e seca), das três espécies de
morcegos mais abundantes na Fazenda das Salinas, Artibeus jamaicensis (A), Artibeus
obscurus (B) e Carollia perspicillata (C), acompanhados ao longo de um ano (Dezembro de
2002 a outubro de 2003). Para tanto, foram utilizadas redes-neblina, abertas de 18:00 à 1:00 h,
totalizando sete horas diárias de campo.
Artibeus jamaicensis
0
5
10
15
20
25
30
Artibeus obscurus
0
5
10
15
20
25
30
Chuvoso
Seco
Carollia perspicillata
0
5
10
15
20
25
30
19:00 20:00 21:00 22:00 23:00 0:00 1:00
Hora de Captura
A
C
B

75
4.4. Categorias Reprodutivas das Fêmeas e Razão Sexual
No presente estudo, das 225 fêmeas de morcegos capturadas, 21 foram
classificadas como pós-lactantes, 11 lactantes e 34 grávidas. A maioria das
fêmeas (n=159) não apresentou características reprodutivas evidentes, sendo
assim foram definidas como não reprodutivas. As espécies que se destacaram em
cada categoria, apresentaram os seguintes valores: 32,35% das fêmeas grávidas
foram de Carollia perspicillata; 45,5% de lactantes e 71,4% de pós-lactantes
foram da espécie A. jamaicensis e 29,5% das fêmeas não reprodutivas foram da
espécie A. obscurus. Dos 213 machos, 161 não apresentavam descida testicular
evidente, este processo reprodutivo somente foi encontrado em 51 machos,
principalmente capturados em abril e agosto, do quais A. jamaicensis representou
44%. No entanto, a descida testicular pode ter outras causas, que incluem o
estresse gerado pela captura e manipulação do animal.
Do total de quirópteros do sexo masculino capturados na Fazenda das
Salinas (n=213), 52,8% foram obtidos no ecossistema manguezal e 47,4% na
terra firme. Enquanto, das fêmeas capturadas (n=225), 48,4% foram coletadas no
manguezal e 51,5% nos bosques de terra firme. A proporção sexual entre machos
e fêmeas somente das espécies A. jamaicensis, A. obscurus, G. soricina e C.
perspicillata foi de aproximadamente 1:1. A avaliação dessa proporcionalidade,
realizada no teste do Qui-quadrado, não foi significativa (p>0,05). Só houve
correlação significativa para os indivíduos da espécie A. cinereus (5:1), no
entanto, este valor é baseado em baixos números de captura (n=26).
4.5. Flutuações Sazonais das Principais Guildas
Espécies de diferentes guildas exibiram heterogeneidade de ocorrência
(Figura 11). A guilda Frugívora englobou a maioria dos indivíduos da Fazenda das
Salinas (64%). Em seguida encontra-se a guilda de Onívoros com 24% das
capturas e por fim as demais guildas, sendo estas Nectarívora (7%), Insetívora
(3%), Carnívora, Hematófaga e Piscívora, que juntas representam 3%. Os
piscívoros foram capturados, exclusivamente, nos bosques de mangue em noites
de preamar.

76
Os valores finais de indivíduos capturados de cada uma das guildas
mais representativas (Frugívora, Onívora e Nectarívora), foram comparados
por ambiente e entre ambientes (terra firme e manguezal), bem como por sítio
e entre os sítios de trabalho, utilizando-se a análise de variância ANOVA
(bifatorial). As diferenças significativas encontradas resultaram somente da
relação entre guildas, por ambiente (ANOVA, F=693; gl=2; p<0,05) e por sítio
de trabalho (ANOVA, F=654,8; gl=2; p<0,01). Sendo que em cada ambiente,
estas diferenças foram induzidas, principalmente, pelas maiores e menores
proporções do número de indivíduos frugívoros e nectarívoros,
respectivamente (Tukey=51,21; p<0,05). Enquanto, para cada sítio de trabalho,
teria sido resultado da relação do menor número de indivíduos nectarívora com
as outras duas guildas restantes (Tukey=161,60; p<0,05 – Nectarívora X
Frugívora e Tukey=88,57; p<0,05 – Nectarívora X Onívora).
A busca por prováveis variações sazonais, do número de indivíduos
de certas guildas de quirópteros nos ecossistemas manguezal e terra firme, ao
longo do período de estudo, foi limitada a utilização apenas das espécies com
valores elevados de capturas. Para tanto, os indivíduos pertencentes a guilda
Frugívora, que apresentaram tempo de atividade noturna semelhantes, foram
agrupados dentro das respectivas categorias sexuais e em seguida
Figura 11 – Abundância relativa das guildas de morcegos nos ecossistemas estudados
da Fazenda das Salinas, durantes o período de dezembro de 2002 a outubro de 2003.

77
correlacionados com fatores externos representados, pelos valores mensais de
precipitação mensal e frutificação nos dois ecossistemas. Como o mesmo
evento foi observado para as espécies que compõem a segunda maior guilda
(Onívora), todo o processo de agrupamento e análise foi também aplicado para
estas espécies.
A primeira análise a ser considerada é a relação entre as variações
do período de frutificação somente da terra firme (Nascimento, 2004) e de
precipitação do manguezal bragantino. As regressões lineares ajustadas não
foram significativas entre estes dois fatores ambientais (p>0,05). Quanto à
regressão linear, agora utilizada para relacionar a variação do número de
machos e fêmeas de cada guilda (Frugívora e Onívora), com produção de
frutos e precipitação (Figura 12), essa também não apresentou resultados
significativos (precipitação = p>0,05 e frutificação = p>0,05).
Considerando as populações de morcegos da guilda Frugívora e
utilizando os dados da categoria aqui especificada como fêmeas reprodutivas
(grávidas e lactantes), o resultado foi o mesmo, ou seja, não houve nenhuma
relação significativa dessas fêmeas reprodutivas com a variação de
precipitação (p>0,05) e nem com o período de frutificação (p>0,05) ao longo do
ciclo anual (Figura 13). Esta mesma análise não foi possível para a guilda
Onívora, devido à insuficiência de dados.
4.6. Resultados das Capturas Diurnas e Noturnas Semestrais
As buscas diurnas em abrigos utilizando puçás, resultaram apenas na
visualização dos indivíduos em ocos de árvores de mangue (Avicennia
germinans) a 10 m de altura, sem a possibilidade de captura e identificação das
espécies. As árvores mortas, caídas no chão do manguezal, não parecem ser
úteis como abrigos para a fauna de quirópteros, em função da influência das
marés.
Nas capturas noturnas semestrais (das 18:00 às 06:00 hs), foram coletados
84 indivíduos com padrões de abundância das espécies semelhantes aos
encontrados nas capturas noturnas bimestrais (das 18:00 à 01:00 h).
Apresentando também como predominantes, as espécies Artibeus jamaicenses

78
(44%), Artibeus obscurus (23%), e Carollia perspicillata (12%), de acordo com os
percentuais de indivíduos capturados. As campanhas noturnas semestrais,
também contribuíram para o registro de mais uma espécie, Myotis nigricans, com
apenas uma captura. As capturas foram, principalmente, obtidas em maior
proporção nos bosques de mangue. Outro dado relevante é a captura de 11
fêmeas na campanha de outubro (estação seca), enquadradas na categoria
reprodutiva grávidas-lactantes, das quais nove fêmeas são da espécie A.
jamaicenses, uma de A. obscurus e a outra de C. perspicillata. Esse fato pode ser
um indicativo para estas espécies de ciclo reprodutivo poliéstrico bimodal.
Frugívora
0
20
40
60
D F A J A O
2002/2003
N°.
de In
diví
duos
0
200
400
600
800
Precipitação
(mm
)
FêmeasMachosPrecipitação
Frugívora
0
20
40
60
D F A J A O
2002/2003
N°.
de In
diví
duos
0
2
4
6
Frutificação
(g.m-2.m
ês-1)
FêmeasMachosFrutificação
Onívora
0
10
20
30
D F A J A O
2002/2003
N°.
de
Indi
vídu
os
0
2
4
6
Frutificação
(g.m-2.m
ês-1)
FêmeasMachosFrutif icação
Onívora
0
10
20
30
D F A J A O
2002/2003
N°.
de
Indi
vídu
os
0
200
400
600
800
Precipitação
(mm
)
FêmeasMachosPrecipitação
A
B
Figura 12 – Variação sazonal do número de machos e fêmeas das guildas Frugívora (A) e Onívora (B),
comparada com a variação sazonal das taxas de frutificação (g.m.-2.mês-1) na terra firme e precipitação (mm) na
área de estudo, Fazenda das Salinas, Bragança-Pará.

79
Figura 13 – Variação sazonal do número de fêmeas reprodutivas grávidas (A) e lactantes (B) da
guilda Frugívora, comparada com a variação sazonal das taxas de frutificação e de precipitação
na área de estudo, Fazenda das Salinas, Bragança-Pará.
A
B
0
5
10
15
D F A J A O
2002/2003
Frut
ifica
ção
(g.m
-2.m
ês-1
)
e n°
. de
fêm
eas
gráv
idas
0
200
400
600
800
Precipitação (m
m)
GrávidasFrutificaçãoPrecipitação
0
5
10
15
D F A J A O
2002/2003
Frut
ifica
ção
(g.m
-2.m
ês-1
)
e n°
. fêm
eas
gráv
idas
0
200
400
600
800
Precipitação (m
m)
LactantesFrutificaçãoPrecipitação

80
4.7. Similaridade da Quiropterofauna no Estado do Pará
É importante ressaltar que a lista de quirópteros aqui apresentada, para
fins de comparação com outras localidades do Estado do Pará, é a compilação de
todas as espécies identificadas ao longo deste trabalho, inclusive Myotis
nigricans. Essa lista representa a quiropterofauna da península bragantina, que
no dendrograma foi denominada de Bragança, acompanhada por dois outros
grandes grupos principais (Figura 14). O primeiro grande grupo inclui as
localidades Belém e Caxiuanã com 69,4% de similaridade, juntamente com Alter
do Chão e Xingu, sendo que a maior diferença foi entre os resultados de Alter do
Chão e Belém (5,34%). Como grupo irmão apresentando 46,84% de similaridade
com Alter do Chão, está o segundo grande grupo formado por Paragominas e
Marajó (Figura 14). O terceiro grupo foi originado do isolamento da localidade de
Bragança, que representa os dados os dados obtidos nos ecossistemas da
Fazenda das Salinas. Esta apresentou apenas 42,36% de similaridade com os
grupos 2 e 1, respectivamente (Figura 14).
Xingu
Caxiuanã
Belém
Alter do Chão
Marajó
Paragominas
Bragança
40 60 80 100
Coeficiente de Similaridade de Bray-Curtis
Figura 14 – Análise de Cluster revelando semelhanças na composição das espécies de seis
estudos realizados em localidades geograficamente distintas, incluindo as amostras da Fazenda
das Salinas em Bragança, Pará.
Coeficiente de Similaridade de Bray-Curtis
40 60 80 100

81
5. DISCUSSÃO
Este trabalho constitui o primeiro estudo da quiropterofauna na península
bragantina, onde está localizada a Fazenda das Salinas e seus ecossistemas.
Provavelmente, também seja o primeiro esforço para apresentar a fauna de
morcegos associada a manguezal no Brasil.
Dos hábitats manguezal e terra firme da Fazenda das Salinas, foram
registrados 23 táxons, dentre os quais 18 espécies, na maioria insetívoras,
apresentaram menos de 10 indivíduos coletados, compreendendo a 10,5% do
total de capturas. Sendo assim, a captura de 280 indivíduos no presente estudo,
confirma a maior seletividade do método de coleta aqui empregado, para táxons
da família Phyllostomidae, a qual é principalmente composta por espécies
frugívoros (Voss & Emmons, 1996; Simmons & Voss, 1998; Kunz & Kurta, 1988).
Contudo, a quiropterofauna da Fazenda das Salinas, correspondeu a 24% do total
de espécies atualmente registradas para todo o estado do Pará, que inclui cerca
de 102 táxons, 56 gêneros e nove famílias de morcegos (Bernard & Fenton, 2002;
Cáceres, 2003; Fonseca, 2003; Marques-Aguiar & Aguiar, 2002; Marques-Aguiar
et al., 2002 a e b; Marques-Aguiar et. al., 2003). De modo mais abrangente, os
resultados do presente estudo, acrescentam 18 espécies a lista organizada por
Fernandes (2000), que apresenta o número de espécies de morcegos associadas
a áreas de mangue no mundo.
Na alta representatividade dos táxons da família Phyllostomidae, as
espécies Artibeus jamaicensis, A. obscurus e Carollia perspicillata foram as
dominantes na Fazenda das Salinas. Essas espécies também estiveram entre as
cinco mais abundantes no trabalho de Bernard & Fenton (2002), que comparou
inventários da quiropterofauna de cinco localidades da Amazônia Central. Estes
mesmos autores, ainda relataram que a espécie Carollia perspicillata prevaleceu
em três desses locais. Entretanto, é difícil comparar inventários, mesmos quando
estes utilizam métodos iguais, pois a riqueza amostral depende do esforço de
coleta.
Estimadores de riqueza total em espécies, são parâmetros úteis que
permitem comparações dos índices de diferentes inventários, devido à

82
independência dos mesmos com relação ao esforço amostral. Desse modo, estes
estão sendo amplamente utilizados em diversos estudos, como por exemplo, na
fauna de aranhas, ácaros, morcegos e outros seres vivos (Santos, 2003). De
acordo com alguns estimadores de riqueza em espécies, a fauna de morcegos
dos ecossistemas estudados na Fazenda das Salinas, não foi totalmente
inventariada. Assim, o esforço amostral não proporcionou a estabilização da curva
de acumulação, apesar dos valores estáveis de integralidade estimados
(próximos a 90%), principalmente no ambiente de terra firme.
Para Bergallo et al. (2003), é importante considerar que o aumento do
esforço de captura (hora.rede), numa dada localidade, é menos eficiente do que o
aumento do número de sítios amostrados. Sendo assim, os valores aqui
apresentados corroboram com essa hipótese, haja vista o esforço empregado,
quanto à densidade de indivíduos e espécies, ter sido mais bem sucedido no
Sítio#01(terra firme) e no Sítio#04 (manguezal). No entanto a razão entre
espécie/indivíduos capturados nesses sítios é relativamente baixa, indicando que
mesmo esses sítios apresentando um alto número de indivíduos capturados, o
número de espécies foi menor. Dessa maneira, se as campanhas noturnas
fossem mais bem distribuídas espacialmente, talvez o número de espécies
tivesse sido bem maior dentro do mesmo esforço de captura, a ponto de refletir
uma curva de acumulação estabilizada.
Estudos de Morrison (1979), indicam que A. jamaicensis é capaz de viajar
até 10 Km entre as suas áreas de dormida e de forrageio. A espécie Carollia
perspicillata apresentou em Alter do Chão, Pará, um alcance de vôo de 2,5 Km
(Bernard & Fenton, 2003). Considerando a capacidade de vôo destas espécies, é
razoável pensar que as distâncias de aproximadamente 500 m entre os sítios de
trabalho, não são grandes barreiras para estes animais voadores, podendo visitar
todos os quatro sítios várias vezes em uma mesma noite.
Há provável preferência dos indivíduos capturados, na maioria frugívoros,
por áreas mais abertas nos ecossistemas da Fazenda Salinas (Sítios #01-TF e
#04-M), o que parece coincidir com relatos de outros estudos. Fleming (1986) e
Mikich (2002), por exemplo, citam que a maioria dos morcegos frugívoros tem
uma dieta generalista, com preferência por frutos que apresentem fácil
acessibilidade na planta e disponibilidade sazonal. Geralmente, áreas mais

83
abertas são bosques apropriados para determinadas famílias de plantas como
Cecropiacae e Moraceae. De acordo com Passos et al. (2003), essas famílias
botânicas são as preferidas por muitas espécies frugívoras de morcegos como as
do gênero Artibeus, da mesma forma que as espécies da família Piperaceae são
apreciadas por Carollia perspicillata.
Para Fernandes (1997), o manguezal é uma floresta de alta produtividade
primária, mas de baixa diversidade de espécies de plantas. Além do mais, os
frutos e propágulos produzidos pelos três tipos de mangue (Rhizophora,
Avicennia e Laguncularia) não apresentam odores atraentes, não são carnosos e
possuem muitos compostos secundários. Essas características são muito
apreciadas pelos morcegos frugívoros (Tomlinson, 1986). A presença de
morcegos em maior quantidade nos bosques de mangue, pode estar relacionada
com o fato de que os ambientes manguezal e terra firme são contíguos, tornando
a diversidade semelhante entre os mesmos. Outro fato que deve ter contribuído
para o registro do maior número de taxóns (não obtidos dos valores de
diversidade) no manguezal, foi a captura de espécies representadas por menos
de cinco indivíduos (espécies raras), principalmente encontradas nas guildas de
insetívoros, carnívoros, hematófagos e piscívoros. Assim, enquanto na terra firme
foram capturadas apenas duas espécies de insetívoros, no manguezal foram
capturados seis, provavelmente atraídas pela disponibilidade de alimentos e pela
facilidade de captura, proporcionada pelo espaço físico bastante aberto.
Estudos relataram que morcegos insetívoros apresentariam maior
capacidade de executar manobras com destreza, para capturar presas voadoras
(McKenzie & Rolfe, 1986; Altringham, 1996). Essas manobras podem ser
favorecidas, não só pela presença de algumas características morfológicas que
incluem formato da asa, peso e tamanho corpóreo, mas também pelo espaço
físico.
Parece provável que os dois ecossistemas, manguezal e terra firme, são
fonte de recursos diferenciados para as diferentes espécies de morcegos. Então,
as “ilhas” de terra firme funcionariam como uma fonte principal de frutos, pois
normalmente há maior diversidade de plantas frutíferas neste sistema, ao passo
que o manguezal poderia simplesmente funcionar, para os frugívoros, como
corredor de acesso a essas “ilhas” de terra firme. Hill & Smith (1986), ressaltam

84
que muitos morcegos frugívoros também adaptam sua dieta, durante o período de
escassez de frutos passando a alimentar-se de insetos. Este relato pode servir
como explicação da alta captura de frugívoros no manguezal. Como não há
trabalhos disponíveis sobre a fauna de insetos na área de estudo, essa
explicação permanecerá apenas no plano hipotético para a área da Fazenda das
Salinas. Sendo assim, as espécies frugívoras capturadas no presente trabalho,
podem obedecer ao comportamento alimentar do tipo oportunista e generalista,
que é favorecido por uma estratégia alimentar de menor gasto energético durante
o forrageio.
Quanto às atividades noturnas dos morcegos, foram intercalados por
períodos de descanso, no qual os mesmos ou estariam retornando aos locais de
moradia ou permaneceriam em áreas próximas ao território de forrageio (Hill &
Smith, 1986). Para a Fazenda das Salinas, a única espécie que se aproximou de
um padrão bimodal de atividade noturna foi Artibeus jamaicensis, enquanto a
espécie A. obscurus, apresentou três picos de atividades. Esses resultados de A.
jamaicensis e A.obscurus, corroboram com o comportamento relatado para as
mesmas por Bernard (2002).
A atividade mais intensa da espécie Carollia perspicillata na área da
Fazenda das Salinas, foi apenas nas três primeiras horas após o pôr-do-sol,
sendo este resultado mais aproximado ao de Marques (1985), diferenciando
apenas no atraso do pico inicial (na segunda hora da noite). Outro resultado do
estudo de Marques (1985), relata que o pico inicial de atividade da espécie G.
soricina, ocorreu na segunda hora após o escurecer, seguido por atividade
continua ao longo da noite, o que se assemelha ao encontrado para os
exemplares de G. soricina, capturados na Fazenda das Salinas, diferenciando-se
apenas quanto ao início do primeiro pico.
Quanto a proporcionalidade de 1:1, entre machos e fêmeas das espécies
A. jamaicensis, A. obscurus, G. soricina e C. perspicillata, a mesma não reflete o
comportamento reprodutivo desses táxons, talvez este fato, seja uma
conseqüência do baixo número de capturas de indivíduos. Geralmente essas
espécies formam haréns, com diversas fêmeas e um macho (Altringham, 1996;
Emmons, 1997). Mesmo diante deste resultado, foi realizada uma breve avaliação

85
dos padrões de reprodução dessas espécies, baseado na condição reprodutiva
das fêmeas. Para tanto, foram utilizadas apenas as categorias de fêmeas
grávidas e lactantes, pois de acordo com Marques-Aguiar (1986), o sucesso
reprodutivo é admitido de acordo com a presença de fêmeas lactantes, as quais
garantem a manutenção da prole.
A maioria das fêmeas frugívoras grávidas e lactantes, capturadas neste
estudo, apresentaram um pico no meio do período seco (agosto a outubro). O
estudo de Bernard (2002), também descreveu um pico para fêmeas grávidas na
Amazônia brasileira no mesmo período (outubro e novembro). Segundo Hill &
Smith (1986), a produção de muitas proles por ano não é simultânea nas espécies
tropicais, para as quais alimento e clima seriam menos restritivos nos trópicos do
que em zonas temperadas, provavelmente devido à presença de variadas fontes
de recursos alimentares associada à dieta generalista de muitos frugívoros. Além
do que, espécies como Artibeus jamaicensis apresentam o recurso fisiológico
extra da diapausa embrionária, que pode prolongar a gravidez se as condições
externas mostrarem-se contrárias (Altringham, 1996).
A falta de correlação da presença de fêmeas grávidas e lactantes com o
período chuvoso, onde, conseqüentemente, deveria haver mais disponibilidade de
alimento para as mesmas, não parece ser um fator limitante para a produção de
uma nova prole também na Fazenda das Salinas. No entanto, os valores apontam
para um período de abundância de frutos durante o período seco (Nascimento,
2004), o qual coincidiu com os picos de captura do maior número de fêmeas
grávidas do que de lactantes. Este fato, fortalecido pela citação de Hill & Smith
(1986) e Altringhan (1996), indica que provavelmente no final da estação seca, a
maioria das fêmeas adultas já deveria ter cessado a amamentação dos infantes e
iniciado um novo ciclo de estro para a geração de uma nova prole, durante a
estação chuvosa.
Os dados de frutificação de Nascimento (2004), não são muito coerente
com áreas de abrangência de floresta pluvial tropical, principalmente na
Amazônia. Os mesmos são provenientes do método de coleta para serapilheira,
realizadas na área de estudo, paralelamente às coletas de morcegos. Porém, de
acordo com Stashko & Dinerstein (1988), este método usado para estimar a
disponibilidade dos frutos para morcegos frugívoros apresenta certos problemas,

86
como a possibilidade de remoção das sementes e frutos por outros animais, além
da localização das armadilhas na área de estudo, que pode gerar uma falta de
representatividade dos frutos coletados pelas mesmas em uma floresta
heterogênea.
Quanto às campanhas noturnas semestrais, estas aumentaram o número
de indivíduos da espécie A. jamaicensis, com um provável terceiro pico de
atividade na madrugada, além da captura de 13 fêmeas grávidas-lactantes, sendo
11 da espécie A. jamaicensis e as duas demais de C. perspicillata e G. soricina. A
presença desta categoria reprodutiva para estas espécies confirma o padrão
poliéstrico bimodal encontrado por Marques (1985). Alguns estudos citam que
muitas espécies de morcegos exibem uma ampla variação nos padrões
reprodutivos, com dois ou mais ciclos éstricos ao longo de um ano, que deveriam
coincidir com o período de maior abundância dos recursos alimentares (Hill &
Smith,1986; Altringham, 1996).
Por fim, os morcegos apresentam um papel ecológico bastante evidente
nas florestas tropicais. São cruciais para a disseminação de sementes de plantas
pioneiras em áreas impactadas (Charles-Dominique, 1986), bem como auxiliam
nos mecanismos de regeneração e sucessão secundária dessas florestas
(Frumhoff, 1995). De acordo com Fenton et al. (1992), a distribuição de morcegos
filostomídeos, principalmente os da subfamília Phyllostominae, reflete os níveis de
perturbação ambiental, sendo a diversidade dessa subfamília maior em locais
relativamente inalterados. No presente estudo, a subfamília Phyllostominae
apresentou baixas taxas de captura, além do que foi registrada a presença da
espécie Desmodus rotundus. Para Marques-Aguiar et al. (2002a), a espécie D.
rotundus também serve como um bom indicador de áreas impactadas, pois o
processo de ocupação humana, associado à criação de animais domésticos, pode
favorecer a densidade de hematófagos. Além do mais, os bosques de terra firme
na Fazenda das Salinas, sofreram um processo de alteração ambiental
proporcionado por acontecimentos naturais e por atividades antrópicas, no
decorrer dos anos (Behling et al., 2001; Souza Filho, 2001).

87
6. CONCLUSÕES
Abaixo estão as conclusões sobre alguns aspectos da ecologia da fauna de
quirópteros, elaboradas a partir dos resultados do monitoramento de bosques de
mangue e terra firme na península bragantina. Os resultados permitiram concluir
que:
1. O levantamento da quiropterofauna na Fazenda das Salinas registrou cinco
famílias, 19 gêneros e 24 espécies; A fauna de morcegos das “ilhas” de terra
firme não difere significativamente daquela encontrada no ecossistema
manguezal, tanto em termos de número de indivíduos quanto em diversidade;
2. A área da Fazenda das Salinas apresenta baixa similaridade com a de
outras áreas estudadas no estado do Pará.
3. As “ilhas” de terra firme apresentaram valores de riqueza em espécies mais
próximos ao esperado do que o manguezal, de acordo com os estimadores de
riquezas empregados neste estudo;
4. Os horários de atividade noturna das espécies mais abundantes, ao longo
do ciclo anual, não foram sobrepostos, indicando independência nas atividades
entre espécies da mesma guilda na área de estudo;
5. A proporcionalidade entre machos e fêmeas para as espécies mais
abundantes estudadas (1:1), não representou os padrões conhecidos para estas
espécies;
6. As fêmeas lactantes apresentaram um padrão de distribuição ao longo do
ano, que sugere um sucesso reprodutivo de algumas espécies da guilda de
frugívoros, durante a estação seca;

88
7. REFERÊNCIAS BIBLIOGRÁFICAS
ABREU, M. M. O., MEHLING, U., NASCIMENTO, R. A. & MENEZES, M. P. M.
(2004). Análise de composição florística e estrutura florestal de um
fragmento de bosque de terra firme em um manguezal adjacente na
península de Ajuruteua, Bragança, Pará. Submetido ao Boletim do Museu
Paraense Emílio Goeldi.
ALTRINGHAM, J. D. (1996). Bats Biology and Behaviour. Oxford University Press,
Oxford, New Jersey and Tokyo, ix-261p.
ANTHONY, E. L. (1988). Age determination in bats. In: Ecological and behavioral
methods for the study of bats. KUNZ, T. H. (Ed.). Smithsonian Institutuion,
Washington D. C., USA, 47-58pp.
AURICCHIO, P. (2002). Mamíferos. In: Técnicas de Coleta e Preparação –
Vertebrados. AURICCHIO, P. & M. da SALOMÃO, G. (Eds.). Instituto Pau
Brasil de História Natural, São Paulo-SP, 150-194 pp.
AYRES, M., AYRES-JR, M. & SANTOS, A.S. (2000). BioEstat 3.0 Aplicações
Estatísticas nas áreas das Ciências Biológicas e Médicas. Sociedade Civil
Mamirauá, Belém, 290p.
BEHLING, H., COHEN, M. C. L. & LARA, R. J. (2001). Studies on Holocene
mangrove ecosystem dynamics of the Bragança Peninsula in north-eastern
Pará, Brazil. Paleogeography, Paleoclimatology, Palaeoecology , 167:225-
242.
BERGALLO, H. G., ESBÉRARD, C. E. L., MELLO, M. A. R., LINS, V.,
MANGOLIN, R., MELO, G. S. & BAPTISTA, M. (2003). Bat species richness
in Atlantic Forest: What is the minimum sampling effort?. Biotropica, 35
(2):278-288.
BERNARD, E. (2002). Diet, activity and reproduction of bats species (Mammalia,
Chiroptera) in Central Amazonia, Brazil. Revta. Bras. Zool., 19(1):173-188.
BERNARD, E. & FENTON. M. B. (2002). Species diversity of bats (Mammalia:
Chiroptera) in forest fragments, primary forest, and savannas in central
Amazonia, Brazil. Can. J. Zool., 80:1124-1140.
BERNARD, E. & FENTON, M. B. (2003). Bat mobility and roosts in a fragmented
landscape in Central Amazonia, Brazil. Biotropica, 35 (2):262-277.

89
CÁCERES, H. J. (2003). Levantamento da quiropterofauna (Mammalia:
Chiroptera) na região de Paragominas (Pa), com observações sobre sua
ecologia. Monografia não publicada, Graduação em Ciências Biológicas,
Universidade Federal do Pará. Belém, 63p.
CAREY, A. B. & HARRINGTON, C. A. (2001). Small mammals in young forests:
implications for management for sustainability. Forest Ecology and
Management, 154:289-309.
CENTENO, A. J. (1999). Curso de Estatística Aplicada à Biologia. 2°. Ed. UFG,
Goiânia, 234p.
CHAPMAN, V.J. (1975). Mangrove biogeography. In: Biology and Management of
Mangroves. Proced Int. Symp. Hawaii. Gerald Walsh et al., (eds.), University
of Florida, Gainsville. Vol. I:3-22.
CHARLES-DOMINIQUE, P. (1986). Inter-relations between frugivorous
vertebrates and pioneer plants: cecropia, birds and bats in Franch Guyana.
In: Frugivorous ad seed dispersal. ESTRADA, A. FLEMING, J. H. (Eds). Dr.
W. Junk Publishers, USA, 105-135pp.
CLARKE, K.R. & GORLEY, R.N. (2001). Primer v.5: User manual / Tutorial.
Primer-E: Plymouth. 91p.
COLWELL, R. K. (1997). EstimateS: Statistical estimation of species richness and shared species from samples. Version 6.0b1.
EISENBERG, J. F. & REDFORD, K. H. (1999). Mammals of the Neotropics. The
Central Neotropicaics. Vol. 3. Ecuador, Peru, Bolívia, Brazil. The University of
Chicago Press, Chicago and London.
EMMONS, L.H. & FEER, F. (1997). Neotropical Rainforest Mammals: a Field
Guide. 2nd ed. The University of Chicago Press, Chicago & London, 307p.
FENTON, M. B., ACHARYA, L., AUDET, D., HICKEY, M. B. C., MERRIMAN, C.,
OBRIST, M. K., SYMES, D. M. & ADKINS, B. (1992). Phyllostomid bats
(Chiroptera: Phyllostomidae) as Indicators of habitat disruption in the
neotropics. Biotropica, 24 (3):440-446.
FERNANDES, M. E. B. (1997). The ecology and productive of mangroves in the
Amazon region, Brazil. Tese de doutorado apresentada ao departamento de
biologia da Universidade de York, York, Inglaterra, 214p.

90
FERNANDES, M. E. B. (2000). Association of mammals with mangrove forests: a
world wide review. Boletim do Lab. Hidrobiologia, 13:83-108.
FINDLEY, J. S. (1993). Bats: a Community Perspective. Cambridge studies in
ecology. Cambridge University Press. Cambridge, UK, xi-167p.
FLEMING, T. H. (1986). Oportunism versus specialization: the evolution of feeding
strategies in frugivorous bats. In: Frugivorous ad seed dispersal. ESTRADA,
A. FLEMING, T. H. (Eds). Dr. W. Junk Publishers, USA, 105-135pp.
FONSECA, R. T. D. (2003). Diversidade de quirópteros de Belém (Pará, Brasil) e
suas implicações ambientais. Trabalho de Conclusão de Curso (Graduação
em Ciências Biológicas) - Universidade Federal do Pará, 57 pp.
FRUMHOFF, P. C. (1995). Conserving wildlife in tropical forest managed for
timber to provide a more viable complement to protected areas. Bioscience,
45 (7):456-464.
FUNDAÇÃO BIODIVERSITAS. (2002). Lista das espécies terrestres da fauna
brasileira ameaçadas de extinção segundo o Workshop da Fundação
Biodiversitas (com categorias UICN). (http: //www.biodiversitas.org.br/f
ameaçada/form fauna.asp)
GREGORIN, R. & TADDEI, V. A. (2002). Chave artificial para a identificação de
molossídeos Brasileiros (Mammalia, Chiropteros). Mastozoologia
Neotropical/ J. Neotrop. Mammal., 9(1):13-32.
HANDLEY, C.O. (1987). New species of bats from Northen South America: fruit-
eating bats, genus Artibeus Leach. In: Studies Neotropical Mammalogy:
Essays in honor of Philip Hershkovitz Fieldiana Zoologgy (New series).
PATTERSON, B. D. & TIMM, R. M. (Eds). 1382 (39):163-172.
HENDERSON, P. A. & SEABY, R. M. H. (1997). Species diversity & Richness.
Pisces Conservation Ltda, Pennington & Lymington, UK.
HILL, J. E. & SMITH, J. D. (1986). Bats: natural history. 2nd Ed. University of Texas
Press, USA, 243p.
KJERVE, B. & LACERDA, L. D. (1993). Mangroves of Brazil. In: Conservation and
Sustainable Utilizacion of Mangrove Forest in Latin America and Africa

91
Regions. Part 1: Latin America. ITTO/ISME Project PD114/90 (F), 245-
272pp.
KNOPPERS, B., EKAU, W., FIGUEIREDO JÚNIOR, A. G. & SOARES-GOMES,
A. (2002). Zona Costeira e Plataforma Continental do Brasil. In: Biologia
Marinha. PEREIRA, R. C. & SOARES-GOMES, A. (Eds.). Interciência, Rio
de Janeiro-RJ, 353-361pp.
KOOPMAN, K. F. (1993). Order Chiroptera. In: Mammal species of the World: a
taxonomic and geographic reference. WILSON, D. E. & REEDER, D. M.
(Eds). 2nd Ed. Smithsonian Institution Press. Washington and London, 137-
241pp.
KREBS, C. J. (1989). Ecological Methodology. Harper Collins Publishers, New
York, 652p.
KUNZ, T. H. & KURTA, A. (1988). Capture methods and holding devices. In:
Ecological and behavioral methods for the study of bats. KUNZ, T. H. (Ed.).
Smithsonian Institutuion, Washington D. C., USA, 1-28pp.
LARA, R. J. & COHEN, M. C. L. (2003). Sensoriamento remoto. Em: Os
manguezais da costa norte brasileira. FERNANDES, M. E. B. (Org).
Fundação Rio Bacanga, São Luís, Maranhão, 11-28pp.
LEWINSOHN, T. M. & PRADO, P. I. (2002). Biodiversidade brasileira: Sínteses
do estado atual do conhecimento. Editora Contexto, São Paulo, 176p.
LINARES, O. J. (1987). Murcielagos de Venezuela. Cuadernos Lagoven,
Departamento de Relaciones Públicas de Lagoven S. A., Caracas, 122p.
MAGURRAN, A. E. (1988). Ecological Diversity and Its Measurement. Princeton
Uiversity Press., New Jersey, 179p.
MARINHO-FILHO, J. & SAZIMA, I. (1998). Brazilian bats and conservation
biology: a first survey. In: Bat biology and conservation. KUNZ, T. H. &
RACEY, P. A. (Eds.). Smithsonian Institution Press. Washington and London,
282-294pp.
MARQUES, S. A. (1985). Novos registros de morcegos no Parque Nacional da
Amazônia (Tapajós), com observações do período de atividade noturna e
reprodução. Boletim do Museu Paraense Emílio Goeldi (Zoologia). Vol 2
(1):71-83.

92
MARQUES-AGUIAR, S. A. (1986). Activity cycle, feeding and reproction of an
population of Molossus ater from Brazil (Chiroptera: Molossidae). Boletim do
Museu Paraense Emílio Goeldi 2 (2): 159-179.
MARQUES-AGUIAR, S. A. (1994). A systematic review of the large species of
Artibeus Leach, 1821 (Mammalia: Chiroptera) with some phylogenetic
inferences. Boletim do Museu Paraense Emílio Goeldi (Zoologia), 10 (1):3-
83.
MARQUES-AGUIAR, S. A. & AGUIAR, G. F. S. (2002). Interações de quirópteros
em ecossistemas tropicais: perspectivas de estudo para Caxiuanã. In:
Caxiuanã: Populações Tradicionais, Meio Físico e Diversidade Biológica.
LISBOA, P.L.B. (Org.). Belém, p. 651-668.
MARQUES-AGUIAR, S. A, REIS-FILHO, V. O.R., SALDANHA, N., ROCHA, M.
M. B., AGUIAR, G. F. S., QUEIRÓZ, J. A. L., ROCHA, A. L. & BRÍGIDA, J.
N. S. (2002a). Levantamento de quirópteros da região de implementação do
CHE Belo Monte, Pará. Relatório Final, Belém, Pará. 51p.
MARQUES-AGUIAR, S. A., MELO, C. C. S. & AGUIAR, G. F. S. & QUEIRÓZ, J.
A. L. (2002b). Levantamento preliminar da mastofauna da região de Anajás-
Muaná, Ilha de Marajó, Pará, Brasil. Revista Brasileira de Zoologia, Curitiba,
PR, 19 (3): 841-854.
MARQUES-AGUIAR, S. A., AGUIAR, G. F. S., SALDANHA, N., SILVA JÚNIOR,
J. S. E. & ROCHA, M. M. B. (2003). Caracterização e perspectivas de
estudo os quirópteros da Estação Científica Ferreira Penna - Município de
Melgaço - PA. Em: Estação Científica Ferreira Penna - Dez Anos De
Pesquisa Na Amazônia,. Idéias e Debates volume 6. Belém: Museu
Paraense Emílio Goeldi/MCT, Belém. 66p.
McKENZIE, N. L. & ROLFE, J. K. (1986). Structure of bat guilds in the Kimberley
mangroves, Australia. Journal of Animal Ecology, 55:401-420.
MEHLIG, U. (2001). Aspects of tree primary production in an equatorial mangrove
forest in Brazil. ZMT contribution. Ph.D. Thesis, Bremen, xiii-155p.
MIKICH, S. B. (2002). A dieta dos morcegos frugívoros (Mammalia, Chiroptera,
Phyllostomidae) de um pequeno remanescente de Floresta Estacional
Semidecidual do sul do Brasil. Revta. Brás. Zool. 19 (1):239-249.

93
MILLIKEN, W. & RATTER, J.A. (1998). The vegetation of the Ilha de Maraca. In
Maraca: the biodiversity and environment of an Amazonian Rain forest.
MILLIKEN W. & RATTER J.A. (Eds). John Wiley & Sons, New York, 71- 112
pp.
MINISTÉRIO DO MEIO AMBIENTE. (2003). Nova lista da fauna silvestre
brasileira ameaçadas de extinção (http://www.ibama.gov.br/). Anexo da
Instrução Normativa n°.3, de 27 de maio de 2003
(http://www.ibama.gov.br/fauna/in 03MMA.doc)
MORRISON, D. W. (1979). Apparent male defense of tree hollows in the fruit bat,
Artibeus jamaicensis. J. Mammal. 60:11-15.
NASCIMENTO, R. E. S. A. (2004). Produção de serapilheira em dois
ecossistemas: terra firme e manguezal na península de Ajuruteua,
Bragança, Pará, Brasil. Monografia apresentada ao curso de Licenciatura
em Biologia, Universidade Federal do Pará, Campus de Bragança. 55p.
NITTROUER, C. A., KUEHL, S. A., RHINE, J. M., FIGUEIREDO, A. G., FARIA, L.
E. C., DIAS, G. T. M., SILVA, M. A. M., UNDERKOFFLER, E. C. &
BORGES, H. V. (1991). Sedimentology and stratigraphy of the Amazon
continetal shelf. Oceanography, 25:33-38.
PARDINI, R., DITT, E.H., CULLEN, JR., BASSI, C. & RUDRAN, R. (2003).
Levantamento rápido de mamíferos de médio e grande porte. Em: Métodos
de Estudo em Biologia da Conservação & Manejo da Vida Silestre. CULLEN
Jr., RUDRAN, R. & VALLADARES-PADUA (Orgs.). Curitiba, Ed. UFPR;
Fundação O Boticário de Proteção a Natureza, 181-201pp.
PASSOS, C. F., SILVA, W. R., PEDRO, W. A. & BONIN, M. R. (2003). Frugivoria
em morcegos (Mammalia, Chiroptera) no parque estadual Intervales,
sudeste do Brasil. Revista Brasileira de Zoologia, 20 (3):511-517.
PIJL, L. v. d. (1982). Principles of dispersalin higher plants. 3°. Edition. Springer-
Verlag, Berlin, IX-147p.
POUGH, F. H., JANIS, C. M. & HEISER, J. B. (1999). Vertebrates Life. 5th ed.
Prentice Hall, Upper Saddle River, New Jersey, xviii-733p.

94
PROUST, M. T., LINTIER, M. & BARTHES, B. (1988). Evolution cotière en
Guyane Française: la zone de Sinnamary. Congresso Brasileiro de
Geologia, Abstracts, Belém, Brazil: 406-407pp.
QUEIROZ, H. L. (1995). Preguiças e Guaribas: os Mamíferos folívoros
arborícolas do Mamirauá, vol. II. CNPq (DF), Sociedade Civil Mamirauá
(AM), Rio de Janeiro, 124p.
RACEY, P. A. (1988). Reproductive assesment in bats. In: Ecological and
behavioral methods for the study of bats. KUNZ, T. H. (Ed.). Smithsonian
Institutuion, Washington D. C., USA, 31-45pp.
RICHARDS, G. C. (1990a). The spectacled Flying-Fox, Pteropus conspicillatus
(Chiroptera: Pteropodidae), in north Queesland, 2. Roost sites and
distribution patterns. Australian Mammalogy, 13:17-24.
RICHARDS, G. C. (1990b). The spectacled Flying-Fox, Pteropus conspicillatus
(Chiroptera: Pteropodidae), in north Queesland, 2. Diety, seed dispersal and
feeding ecology. Australian Mammalogy 13:25-31.
SAENGER, P. & SNEDAKER, S. C. (1993). Pantropical trends in mangrove
above-ground biomass and annual litterfall. Oecologia, 96: 293-299.
SANTOS, A. J. (2003). Estimativas de riqueza em espécies. Em: Métodos de
Estudo em Biologia da Conservação & Manejo da Vida Silestre. CULLEN Jr.,
RUDRAN, R. & VALLADARES-PADUA (Orgs.). Curitiba, Ed. UFPR;
Fundação O Boticário de Proteção a Natureza, 19-41pp.
SCHAEFFER-NOVELLI, Y. (1995). Manguezal: Ecossistema entre a Terra e o
Mar. Caribbean Ecological Research., São Paulo, 64p.
SCHWAMBORN, R. & SAINT-PAUL, U. (1996).Mangroves, forgotten forests? Nat.
Resourc and Develp., 43/44:13-36.
SEKIAMA, M. L., REIS, N.R., PERACCHI, A.L. & ROCHA, V.J. (2001). Morcegos
do Parque Nacional do Iguaçu, Paraná (Chiroptera, Mammalia). Revista
Brasileira de Zoologia, Curitiba, 18 (3):749-754.
SENNA, C. & MELLO, C. (1994). Impactos naturais e antrópicos em manguezais
do litoral do nordeste do Pará. II Workshop ECOLAB. Anais. Amapá, 9-11pp.
SILVA, F. (1994). Mamíferos Silvestres – Rio Grande do Sul. 2a ed. – Porto
Alegre, Fundação Zoobotânica do Rio Grande do Sul, publicações Avulsas
FZB, n° 7, 246p.

95
SILVA JÚNIOR, J. S., AVELAR Á. A., SILVA, L. S. (2002): Diversidade de
mamíferos no litoral da Amazônia brasileira. Coordenação de zoologia –
Museu Paraense Emílio Goeldi. VI Workshop ECOLAB (CDROM), Belém-
Pará.
SIMMONS, N. B. & VOSS, R. S. (1998). The mammals of Paracou, French
Guiana: a Neotropical lowland rainforest fauna. Part 1. Bats. Bulletin of the
American Museum of Natural History, 237:1-219.
SMITH, E. P. & vAN BELLE, G. (1984). Nonparametric estimation of species
richness. Biometrics 40: 119-129.
SOUZA FILHO, P. W. M. (2001). Impactos naturais e antrópicos na planície
costeira de Bragança (NE do Pará). Em: Ecossistemas Costeiros: Impactos e
Gestão ambiental. PROST, M. T. R. C. & MENDES, A. (Eds). Museu
Paraense Emílio Goeldi, Belém, 133-144pp.
STASHKO, E. R. & DINERSTEIN, E. (1988). Methods estimating fruit availability
to frugivorous bats. In: Ecological and behavioral methods for the study of
bats. KUNZ, T. H. (Ed.). Smithsonian Institutuion, Washington D. C., USA,
221-230pp.
TOMLINSON, P. B. (1986). The botany of mangrove. Cambridge University Press.
413p.
van TONGEREN, O S. R. (1995). Cluster analyse. In: Data Analyse in community
and landscape ecology. JOUNGMAN, R. H. G., TRE BRAAK, C. J. S. & van
TONGEREN, O. S. R. (Eds.). Cambridge University Press, United Kingdon.
174-212pp.
VANUCCI, M. (1999).Os Manguezais e Nós/ Uma Síntese de Percepções.
EDUSP, São Paulo, 233p.
VOSS, R. S. & EMMONS, L. H. (1996). Mammalian diversity in Neotropical
lowland rainforests: a preliminary assessment. Bulletin of the American
Museum of Natural History, 230:1-115.
WILSON, D. E. (1973). Reproduction in neotropical bats. Period. Biol., 75:215-
217.

96
WILSON, D. E., ASCORRA, C. F. & SOLARI, S. (1996). Bats a Indicators of
Habitat Disturbance. In: Biodiversity of Southeastern Peru. WILSON, D. E. &
SANDOVAL, A. (Eds.). Smithsonian Institution Press., Washington, 679p.
ZANZINI, A. C. S. (2000). Fauna Silvestre - Introdução ao estudo de gestão
ambiental. Curso de Pós-graduação “Lato Sensu” (Especialização) à
distância – Gestão e Manejo Ambiental em Sistemas Agrícolas. Lavras:
UFLA/FAEPE, 80p.

97
ANEXOS

98
Anexo 1 – Espécies de morcegos registradas para sete localidades no Estado do Pará. Localidades
Alter do Chão Bragança Caxiuanã Xingu Paragominas
Belém Marajó
Ametrida anderseni 1 0 0 1 0 0 0 Ametrida centurio 1 0 0 1 1 1 1 Anoura geoffroy 0 0 0 1 0 0 0 Artibeus cinereus 1 1 0 0 0 1 1 Artibeus concolor 1 0 0 0 1 1 1 Artibeus gnomus 1 0 1 0 0 1 1 Artibeus jamaicensis 1 1 1 1 0 1 1 Artibeus lituratus 1 0 1 1 1 1 1 Artibeus obscurus 1 1 1 1 1 1 1 Carollia brevicauda 1 0 1 1 0 1 0 Carollia perspicillata 1 1 1 1 1 1 1 Chiroderma trinitatum 1 0 0 0 0 1 0 Chiroderma villosum 1 1 0 0 0 1 0 Choeroniscus minor 1 0 1 0 1 1 1 Chrotopterus auritus 1 1 1 0 0 1 0 Cormura brevirostris 1 0 0 1 0 1 0 Desmodus rotundus 1 1 1 1 1 1 1 Diaemus youngi 0 0 1 1 0 1 0 Diclidurus albus 0 0 0 0 0 1 0 Diclidurus ingens 1 0 0 0 0 0 0 Diclidurus scutatus 0 0 0 0 0 1 0 Ectophylla macconnelli 1 0 0 0 0 0 0 Eptesicus brasiliensis 1 0 1 0 0 1 0 Eptesicus chiriquinus 1 0 0 0 0 0 0 Eptesicus furinalis 0 0 0 0 1 1 0 Eumops bonariensis 1 0 0 0 0 0 0 Eumops perotis 0 0 0 0 0 1 0 Eumops trumbulli 0 0 0 0 0 1 0 Furipterus horrens 0 0 0 1 0 0 0 Glossophaga soricina 1 1 1 1 1 1 1 Glyphonycteris daviesi 1 0 0 0 0 0 0

99
Glyphonycteris sylvestris 1 0 0 0 0 0 0 Lasiurus blossevilli 1 0 0 0 0 0 0 Lasiurus brachyotis 1 0 0 0 0 0 0 Lasiurus ega 0 0 1 0 0 0 0 Lichonycteris obscura 1 0 0 0 1 1 0 Lionycteris spurrelli 0 0 1 0 1 1 0 Lonchophylla thomasi 1 0 1 1 1 1 1 Lonchorhina aurita 0 0 0 1 0 0 0 Macrophyllum macrophyllum 0 0 0 1 0 1 0 Mesophylla macconnelli 0 0 1 0 0 1 0 Micronycteris brachyotis 0 0 0 0 0 1 0 Micronycteris daviesi 0 0 1 0 0 1 0 Micronycteris hirsuta 1 0 1 1 0 0 0 Micronycteris homezi 1 0 0 1 0 0 0 Micronycteris megalotis 1 1 1 0 0 1 0 Micronycteris minuta 1 0 1 0 0 1 0 Micronycteris nicefori 0 0 1 1 0 1 0 Micronycteris schmidtorum 1 1 0 0 0 1 0 Micronycteris sylvestris 0 0 1 0 0 1 0 Mimon crenulatum 1 1 1 1 0 1 0 Molossops matogrossensis 0 0 0 1 0 0 0 Molossops neglectus 0 0 0 0 0 1 0 Molossops paranus 1 0 0 0 0 0 0 Molossops planirostris 1 0 0 1 0 0 0 Molossops temminckii 1 0 0 0 0 0 0 Molossus ater 0 0 0 0 0 1 0 Molossus molossus 1 1 0 0 0 1 0 Myotis albescens 1 1 0 1 0 1 0 Myotis nigricans 0 1 1 1 0 1 0 Myotis riparius 1 0 0 0 0 1 0 Myotis simus 0 0 0 0 0 1 0

100
Neonycteris pusilla 0 0 0 1 0 0 0 Noctilio albiventris 1 0 1 1 0 0 0 Noctilio leporinus 0 1 0 0 0 1 1 Peropteryx kappleri 0 0 0 0 0 1 0 Anexo 1 – Continuação... Peropteryx leucoptera 1 0 0 1 0 0 0 Peropteryx macrotis 1 0 1 1 0 1 0 Phylloderma stenops 1 1 1 1 1 1 0 Phyllostomus discolor 1 0 0 1 1 1 0 Phyllostomus elongatus 1 0 1 1 1 1 0 Phyllostomus hastatus 1 0 1 1 0 1 0 Platyrrhinus brachycephalus 1 0 0 0 0 0 1 Platyrrhinus helleri 1 1 1 1 1 1 1 Pteronotus gymnonotus 1 0 0 0 0 0 0 Pteronotus parnellii 1 0 0 1 0 0 0 Rhinophylla fischerae 1 0 1 1 1 1 0 Rhinophylla pumilio 1 0 1 1 1 1 1 Rhynchonycteris naso 1 0 1 1 0 1 0 Saccopteryx bilineata 1 0 1 1 0 1 0 Saccopteryx canescens 1 0 1 0 0 1 0 Saccopteryx gymnura 0 0 0 0 0 1 0 Saccopteryx leptura 0 1 1 1 0 1 0 Scleronycteris ega 1 0 0 0 0 0 0 Sturnira lilium 1 1 1 1 1 1 1 Sturnira tildae 1 0 1 1 1 1 1 Thyroptera lavali 1 0 1 0 0 0 0 Thyroptera tricolor 1 0 1 1 0 1 0 Tonatia bidens 0 0 1 0 0 0 0 Tonatia brasiliense 1 0 1 1 0 1 0 Tonatia carriekeri 1 0 0 0 0 1 0 Tonatia saurophila 1 1 0 1 0 1 0 Tonatia schulzi 1 0 0 0 0 0 0 Tonatia silvicola 1 0 1 1 1 1 0 Trachops cirrhosus 1 1 1 1 0 1 1 Trinycteris nicefori 1 0 0 0 0 0 0
101
Uroderma bilobatum 1 1 1 1 1 1 1 Uroderma magnirostrum 1 1 0 1 1 1 1 Vampyressa bidens 1 1 1 1 0 1 1 Vampyressa pusilla 0 0 1 1 0 0 0 Vampyrodes caraccioli 0 0 0 1 1 1 0 Vampyrum spectrum 1 0 0 0 1 1 0

102
Anexo 2 – Informações gerais e as principais características morfológicas
utilizadas para a identificação dos táxons capturados no manguezal e na terra
firme da Fazenda das Salinas, município de Bragança – Pará.
_________________________________________________________________
1. Emballonuridae
Esta família é de distribuição mundial, tropical e subtropical, e é constituída
por 48 espécies, das quais 18 estão no Novo Mundo. São morcegos pequenos
que pesam de 3 a 15 g, sendo as fêmeas geralmente maiores que os machos em
muitas das espécies. Possuem como característica principal a porção terminal da
cauda se projetando livremente sobre o uropatágio (Esquema 1-A). As orelhas
são bem separadas e de tamanho moderado, largamente triangular e com a
margem interna começando desde a parte frontal, logo acima dos olhos. O rosto
apresenta, na maioria das vezes, um focinho alongado e nos lábios superiores
uma divisão (Esquema 1-B). As asas são largas e estreitas, devido à redução
progressiva dos metacarpos do terceiro ao quinto dedo, sendo assim, o segundo
dedo possui um metacarpo e os demais dois. Em muitas espécies, os machos ou
ambos os sexos possuem sacos glandulares, na membrana da asa acima do
cotovelo (Esquema 1-C), destinados à produção de odores para demarcação de
território e atividades reprodutivas (Hill & Smith, 1986; Linares, 1987; Altringham,
1996). No presente trabalho, foi capturada apenas um exemplar da espécie
Saccopteryx leptura (Esquema 1), utilizando redes-neblina a 3 m de altura em
área de terra firme.
C
B
A
Esquema 1 - Principais características morfológicas distintivas de morcegos da família Emballonuridae,
utilizando como modelo a espécie Saccopteryx leptura, único representante da família capturado no
presente trabalho: (A) cauda livre e (B) focinho alongado e (C) saco glandular.

103
2. Família Molossidae
É constituída por 90 espécies e 14 gêneros de distribuição ampla em
regiões tropical e subtropical, sendo que 10 destes gêneros estão presentes no
Novo Mundo. São morcegos de tamanho que vai do médio ao grande, cauda livre
e grossa que se projeta para além da margem posterior do uropatágio (Esquema
2-A). Esta cauda auxilia na direção durante o vôo. O rosto é obtuso, com narinas
elevadas, boca larga, presença de pregas grossas verticais nos lábios superiores,
que estão associadas à captura de insetos, além da presença de rugas. As
orelhas grandes e compridas são dobradas para frente, unidas frontalmente por
uma prega grossa de pele (Esquema 2-B). A forma precisa do anti-trago e da
extremidade superior da orelha, são importantes características para a
identificação de gênero. As asas são pontiagudas, estreitas e compridas. Todas
as espécies voam alto, sendo assim, são raramente capturadas com redes-
neblina (Hill & Smith, 1986; Linares, 1987; Altringham, 1996). No presente
trabalho, apenas três exemplares da espécie Molossus molossus foram
capturados com redes-neblina a 3 m de altura na área de manguezal.
B A
Esquema 2 - Detalhes morfológicos da espécie Molossus molossus, único representante desta
família capturado na Fazenda das Salinas: (A) cauda livre, adaptado de Hill & Smith (1986) e
(B) orelhas dobradas para frente.

104
3. Família Noctilionidae
Constituída de um gênero e duas espécies, que são restritas às regiões
neotropicais e subtropicais do Novo Mundo. Apresentam orelhas pontiagudas,
compridas e estreitas (Esquema 3-A). A pelagem é curta e de cor laranja
brilhante. Os membros posteriores incluem patas grandes com garras fortes e
afiladas (Esquema 3-B), adaptadas para o forrageio sobre a superfície d’água,
principalmente para a captura de peixes, sendo que também se alimentam de
pequenos crustáceos e insetos aquáticos (Hill & Smith, 1986; Emmons, 1997). A
cauda tem as mesmas características dos embalonurídeos (Esquema 3-C). No
entanto, serão principalmente reconhecidos pela forma peculiar de seus lábios
superiores, do tipo leporino (Esquema 3-D). Devido a esta característica, em
outros países são conhecidos popularmente como “Bulldog Bats”. No presente
trabalho foram capturados cinco exemplares da espécie Noctilio leporinus no
ecossistema manguezal, somente quando a maré estava cheia.
A
B D
C
Esquema 3 - Detalhes morfológicos da espécie Noctilio leporinus: (A) coloração típica, laranja
brilhante e orelhas longa e estreita; (B) patas grandes com garras fortes e afiladas; (C) cauda
projetando-se a partir da membrana interfemural (adaptado de Hill & Smith, 1986) e (D) detalhe do
rosto mostrando o lábio superior fendido.

105
4. Família Phyllostomidae
Os filostomídeos incluem cerca de 6 subfamílias descritas, 51 gêneros e
147 espécies, o que pode variar de acordo com o autor (Altringham, 1996,
Emmon, 1997). Possuem uma variedade de formas adaptadas às regiões
tropicais e subtropicais do Novo Mundo. Evoluíram de um ancestral insetívoro,
mas atualmente possuem uma variedade de preferências alimentares, sendo
incluídas nesta família todas as guildas neotropicais.
As espécies componentes desta família apresentam, como característica
principal, uma estrutura tecidual denominada folha nasal, que tem a forma de uma
lança e está localizada sobre o nariz (Esquema 4-A). Em algumas subfamílias ela
pode ser ausente. Essa folha varia de acordo com a espécie, podendo ser
pequena e sensível, até grande e complexa. Há uma considerável variação
relativa para o comprimento da cauda e forma do uropatágio (Esquema 4-B). A
seguir serão apresentadas as subfamílias e as espécies capturadas no presente
trabalho, realizado na Fazenda das Salinas, Bragança-Pará.
A B
Esquema 4 - Características típicas dos filostomídeos: (A) Folha nasal apresentada sobre a
esquematização da espécie Mimon crenulatum, adaptado de Emmons (1997) e (B) comprimentos e
formas do uropatágio e da cauda, adaptado de Hill & Smith (1986).

106
4.1. Subfamília Carollinae
São morcegos que se alimentam de frutos e insetos, sendo a dieta
complementada com néctar durante a estação seca. Geralmente são os mais
numerosos morcegos em florestas tropicais e parecem ser muito comuns em
hábitats perturbados. Também, devido à abundância do grupo, são
provavelmente um dos mais importantes dispersores de sementes das
pimenteiras do gênero Piper (Fleming 1988, Altringham 1996, Emmons, 1997),
além de muitas outras plantas com pequenos frutos. As colônias consistem de
haréns, com um macho e várias fêmeas. No presente estudo foram capturados
107 indivíduos da espécie Carollia perspicillata (Fotografia 1).
4.2. Subfamília Desmodontinae
Esta subfamília é constituída por três espécies que se alimentam de
sangue de mamíferos, mais raramente de pássaros e em cativeiro até mesmo de
anfíbios e répteis (Emmons, 1997). Eles possuem sensores no nariz que
detectam áreas ricas em capilares, caninos modificados para cortar, incisivos
afiados para abrir feridas, língua estriada para transportar o sangue rapidamente
para a boca e anticoagulante na saliva (Altringham, 1996). Estão sendo
amplamente estudados no auxílio de doenças do aparelho circulatório. Possuem
Fotografia 1 - Carollia perspicillata, a única
espécie da subfamília Carollinae capturada
nos dois ecossistemas, manguezal e terra
firme, na Fazenda das Salinas, Bragança,
Pará.

107
estômago e rim especializados na remoção rápida do plasma, sendo na verdade,
as células vermelhas, as mais importantes para a sua alimentação. Na Fazenda
das Salinas foram capturados seis exemplares da espécie Desmodus rotundus
(Fotografia 2). A mesma, de acordo com Emmons (1997), possui uma
organização social complexa com as fêmeas formando grupos estáveis de 8 a 12
indivíduos, que se alimentam a longas distâncias, depois retornam para as
colônias e compartilham o alimento com os outros do grupo por regurgitação. Os
machos formam associações temporárias e competem entre si por grupos de
fêmeas. Estes morcegos são naturalmente raros em florestas, mas tornaram-se
comuns principalmente, em áreas onde se pratica bovinocultura, dessa forma está
espécie deveria ser protegida em florestas, devendo ser o controle empreendido
nas populações que atacam bovinos em criações extensivas, as quais podem
estar contaminadas pelo vírus da raiva (Marques-Aguiar et al., 2002).
Fotografia 2 - Desmodus rotundus, subfamília
Desmodontinae, capturados, tanto no manguezal
quanto na terra firme.

108
4.3. Subfamília Glossophaginae
São morcegos polinizadores de muitas plantas, que se alimentam do
néctar das flores, mas também podem comer frutos e insetos. A alimentação é
auxiliada por dentes reduzidos, uma longa língua e uma coleção de papilas em
forma de escova no final da mesma, a qual aumenta a quantidade de néctar
transportado (Altringham 1996, Emmons, 1997). Foram capturados neste trabalho
36 indivíduos da espécie Glossophaga soricina (Fotografia 3).
.
4.4. Subfamília Phyllostominae
São morcegos, em sua maioria generalistas, que consomem desde
vertebrados, insetos até frutos. Apresentam focinho estreito, orelhas bem
desenvolvidas, folha nasal em quase todas as espécies e cauda sobressaindo ou
não do uropatágio. A ecolocalização é presente em todos os táxons, auxiliando na
captura de presas (Altringham 1996, Emmons, 1997). Nos ecossistemas da
Fazenda das Salinas, foram capturados 18 indivíduos pertencentes à sete
espécies da família Phyllostominae: Chrotopterus auritus, Micronycteris megalotis,
Micronycteris schmidtorum, Mimon crenulatum, Phylloderma stenops, Tonatia
saurophila re Trachops cirrohosus). Destes, quatro estão representados na
Fotografia 4.
Fotografia 3 - Glossophaga soricina,
subfamília Glossophaginae.

109
A
C D
B
Fotografia 4 – Indivíduos da subfamília Phyllostominae capturados
na Fazenda das Salinas: (A) Micronicteris megalotis, (B)
Micronycteris schimidtorum, (C) Trachops chirrosus e (D)
Chrotopterus auritus.

110
4.5. Subfamília Stenodermatinae
Suas características incluem ausência de cauda no uropatágio, que se
estende até a metade das extremidades posteriores formando um “V”, quando os
calcâneos são abertos. No rosto curto e alargado podem apresentar duas linhas
esbranquiçadas, que passam entre as orelhas e outras duas abaixo da orelha,
além de uma listra mais comprida no dorso. Possuem hábito alimentar à base de
frutos, mas algumas espécies podem alimentar-se de insetos (Hill & Smith, 1984).
Foram capturados neste trabalho nove espécies da família Stenodermatinae:
Artibeus obscurus, Artibeus cinereus, Artibeus jamaicensis, Chrioderma villosum,
Platyrrhinus helleri, Sturnira lillium, Uroderma bilobatum, Uroderma magnirostrum
e Vampyressa bidens. Destes, cinco estão representados na Fotografia 5.
B
C
E D
A
Fotografia 5 – Indivíduos da subfamília Stenodermatinae
capturados na Fazenda das Salinas: (A) Artibeus obscurus, (B)
Artibeus cinereus, (C) Artibeus jamaicensis, (D) Vampyressa
bidens e (E) Sturnira lillium.

111
5. Família Vespertilionidae
É uma das famílias de morcegos insetívoros mais numerosas, com
distribuição mundial. Morfologicamente são pequenos, com rosto sem apêndices
ou protuberâncias, de orelhas bem separadas, que por muitas vezes são
extremamente desproporcionais ao rosto, devido às longas dimensões. O trago é
alargado e às vezes curvo, a cauda também é larga e ocupa toda a extensão do
uropatágio, já as asas são longas e estreitas (Linares, 1987).





























