Embed Size (px)
Citation preview

ECOLOGIA
NO
CERRADOPROJETOS DE PESQUISA PRODUZIDOSNO SÉTIMO CURSO "MÉTODOS DECAMPO EM ECOLOGIA" REALIZADONA RESERVA ECOLÓGICA DO IBGE NOPERÍODO DE 1 A 15 DE MARÇO DE 1996
EDITADO PORRaimundo P. B. Henriques,
Guarino R. Colli, e
John Du Vall Hay
UNIVERSIDADE DE BRASÍLIADEPARTAMENTO DE ECOLOGIAPÓS-GRADUAÇÃO EM ECOLOGIABRASÍLIA, DF

ii
CONTEÚDO
APRESENTAÇÃO ivRaimundo P. B. Henriques
LISTA DE PROFESSORES v
LISTA DE ALUNOS v
PROJETOS EM GRUPO
DISTRIBUIÇÃO DIFERENCIAL DEINSETOS GALHADORES EMINADORES ENTRE HABITATSFlávia S. Pinto; Guilherme H. B. deMiranda; Saulo M. A. Andrade.2
VISITAÇÃO DE FORMIGAS EMNECTÁRIOS EXTRAFLORAIS DEQualea multiflora (VOCHISIACEAE) ESEU POTENCIAL COMO AGENTEANTI-HERBIVORIADelano M. S. da Silva; Rosane G. Colevatti;Everton A. dos Santos. 4
OCORRÊNCIA DE FORMIGAS EMGALHAS DE Diospyros hispida D. C. NARESERVA ECOLÓGICA DO IBGE, DFCarlos A. Bianchi; Felipe A. P. L. Costa;Terezinha A. B. Dias. 6
ESTRATÉGIAS DEFORRAGEAMENTO DE LAGARTOSFlávia S. Pinto; Saulo M. A. Andrade;Guilherme H. B. de Miranda 9
DIVERSIDADE DE ORTHOPTERARELACIONADA ÀHETEROGENEIDADE ESPACIALCarlos A. Bianchi; Felipe A. P. L. Costa;Terezinha A. B. Dias. 11
ECOLOGIA FLORAL DE QUATROESPÉCIES DO CERRADO:FREQUÊNCIA DE VISITAS E
POLINIZADORESDelano M. S. da Silva;Everton A. dos Santos; Rosane G. Colevatti.
14
APOSEMATISMO: ADVERTÊNCIAOU ATRAÇÃO?Carlos A. Bianchi; Felipe A. P. L. Costa;Terezinha A. B. Dias. 17
INFLUÊNCIA DO FOGO ANUAL NAFLORÍSTICA E DENSIDADE DEESPÉCIES LENHOSAS DECERRADÃODelano M. S. da Silva; Everton A. dosSantos; Rosane G. Colevatti. 19
EFEITO DO FOGO NAARQUITETURA DE ESPÉCIESLENHOSAS DO CERRADOFlávia S. Pinto; Guilherme H. B. deMiranda; Saulo M. A. Andrade.22
COMPARAÇÃO DECARACTERÍSTICAS DE FOLHAS DEPLANTAS DE CERRADO E MATA DEGALERIACarlos A. Bianchi; Felipe A. P. L Costa;Terezinha A. B. Dias. 25
COMPARAÇÃO DO POTENCIAL DEÁGUA DE ESPÉCIES LENHOSAS DEDIFERENTES GRUPOS FUNCIONAISDelano M. S. da Silva; Everton A. dosSantos; Rosane G. Colevatti 27
COMPARAÇÃO DE MEDIDAS DETRANSPIRAÇÃO ENTRE PLANTASHEMIPARASITAS E SUASHOSPEDEIRASFlávia S. Pinto; Saulo M. A. Andrade;Guilherme H. B. de Miranda. 29
RELAÇÃO ENTRECARACTERÍSTICAS FOLIARES E

iii
HERBIVORIA EM ESPÉCIESLENHOSAS DE MATA E DECERRADODelano M. S. da Silva; Everton A. dosSantos; Rosane G. Colevatti. 31
DISTRIBUIÇÃO DE PLANTASPARASITAS SOBRE PLANTASHOSPEDEIRAS NA RESERVAECOLÓGICA DO IBGE (BRASÍLIA,D.F)Carlos A. Bianchi; Felipe A. P. L. Costa;Terezinha A. B. Dias. 33
ESTRUTURA POPULACIONAL DEPseudobombax longifolium(BOMBACACEA). DISTRIBUIÇÃOETÁRIA E TABELA DE VIDADelano M. S. da Silva; Everton A. dosSantos; Rosane G. Colevatti. 35
TAXA DE OCORRÊNCIA DEESPÉCIES LENHOSAS COMNECTÁRIOS EXTRAFLORAIS EMCERRADOFlávia S. Pinto; Guilherme H. B. deMiranda; Saulo M. A. Andrade.37
A COEXISTÊNCIA DE ESPÉCIESCONGENÉRICAS E SEU NICHOMORFOLÓGICOFlávia S. Pinto; Guilherme H. B. deMiranda; Saulo M. A. Andrade.39
SIMILARIDADE FLORÍSTICA ENTRECOMUNIDADES DE ESPÉCIESHERBÁCEAS EM FITOFISIONOMIASEQUIVALENTESCarlos A. Bianchi; Felipe A. P. L. Costa;Terezinha A. B. Dias. 41
PROJETOS INDIVIDUAIS
PREDAÇÃO DE NINHOS ARTIFICIAISNO CERRADO: UMA COMPARAÇÃOENTRE DUAS FISIONOMIAS.Carlos Abs Bianchi 44
DISTRIBUIÇÃO ESPACIAL DE QUALEAGRANDIFLORA MART. (VOCHYSIACEAE)EM UM CERRADO DE BRASÍLIA, DF.Delano Moody Simões da Silva 49
ATIVIDADE FORRAGEIRA DE Attasexdens (LINNAEUS, 1758)(HYMENOPTERA, FORMICIDAE):RELAÇÃO COM TEMPERATURA EUMIDADE RELATIVA DO AREverton Amancio dos Santos 55
ESTRUTURA POPULACIONAL DEUMA BORBOLETA “ESTALADEIRA”,Hamadryas feronia (L.) (LEPIDOPTERA:NYMPHALIDAE)Felipe A. P. L. Costa 58
CONSIDERAÇÕES SOBRE ADISPERSÃO SECUNDÁRIA DESEMENTES DE SOLANUMLYCOCARPUM (SOLANACEAE) PORFORMIGAS.Flávia dos Santos Pinto 65
PADRÃO DE ATIVIDADE DO MICO-ESTRELA (CALLITHRIXPENICILLATA) NO CERRADO DENSOE CERRADÃO DA RESERVAECOLÓGICA DO IBGE, BRASÍLIA, DF
Guilherme H. B. de Miranda 71
ARQUITETURA E RIQUEZA DEHERBÍVOROS EM Ouratea hexaspermaBaill. (OCHNACEAE), EM UMCERRADO DA RESERVAECOLÓGICA DO IBGE, BRASÍLIA, DF

iv
Rosane G. Collevatti 75
EFEITOS DE DIFERENTES REGIMESDE QUEIMA SOBRE A VEGETAÇÃOLENHOSA DE CERRADO SENSUSTRICTO: TAXAS DIFERENCIAIS DEREBROTASSaulo Marques de Abreu Andrade 82

v
APRESENTAÇÃO
Nesta publicação são reunidas as contribuições de alunos e professores do sétimo curso"Métodos de Campo em Ecologia", desenvolvido pelo curso de Pós-graduação em Ecologia doDepartamento de Ecologia da Universidade de Brasília, na Reserva Ecológica do IBGE (DF), de1 a 15 de março de 1996.
Os trabalhos apresentados aqui são fruto de projetos diários orientados por professores,além de projetos finais realizados pelos alunos individualmente. Estes trabalhos, tratam depadrões e processos nas comunidades de Cerrado do Brasil Central. Alguns trabalhos sãobaseados em dados observacionais, porém outros foram realizados através de experimentação.
O apoio financeiro ao curso foi proporcionado por Laercio Leonel Leite, Coordenador docurso de Pós-Graduação em Ecologia da UnB. Queremos também agradecer a Maria IracemaGonzales, Diretora da Reserva Ecológica do IBGE, por sediar o curso na Reserva e Helena C.Morais pela ajuda na revisão dos projetos finais. A todos os participantes do curso, alunos eprofessores nossos sinceros agradecimentos, pela boa vontade e colaboração.
R. P. B. H.G. R. C.J. D. V. H.

vi
LISTA DE PROFESSORES ORIENTADORES
Adriana G Moreira, Dr.Reserva Ecológica do IBGEBrasília, DF
Alexandre Fernandes Bamberg de Araújo, DrDepartamento de ZoologiaUniversidade de BrasíliaBrasília, DF
Augusto César Franco, Ph.D.Departamento de BotânicaUniversidade de BrasíliaBrasília, DF
Carlos Augusto Klink, Ph.D.Departamento de EcologiaUniversidade de BrasíliaBrasília, DF
Fábio Rúbio Scarano, Ph.D.Departamento de EcologiaUniversidade Federal do Rio de JaneiroRio de Janeiro, RJ
Geraldo Wilson Fernandes, Ph.D.Departamento de Biologia GeralUniversidade Federal de Minas GeraisBelo Horizonte, MG
Guarino Rinaldi Colli, Ph.D.Departamento de ZoologiaUniversidade de BrasíliaBrasília, DF
Heloisa Sinatora Miranda, Ph.D.Departamento de EcologiaUniversidade de BrasíliaBrasília, DF
Jader Soares Marinho, DrDepartamento de ZoologiaUniversidade de BrasíliaBrasília, DF
John DuVall Hay, Ph.D.Departamento de EcologiaUniversidade de BrasíliaBrasília, DF
Mauro C. L. de Brito Ribeiro, Dr.Reserva Ecológica do IBGEBrasília, DF
Mercedes Bustamante, Ph.D.Departamento de EcologiaUniversidade de BrasíliaBrasília, DF
Paulo S. Oliveira, Dr.Departamento de ZoologiaUniversidade de CampinasCampinas, SP
Raimundo P. B. Henriques, Dr.Departamento de EcologiaUniversidade de BrasíliaBrasília, DF
Regina Macedo, Ph.D.Departamento de ZoologiaUniversidade de BrasíliaBrasília, DF
Tarcisio S. Filgueiras, Dr.Reserva Ecológica do IBGEBrasília, DF
LISTA DE ALUNOS
Alunos do Curso de Mestrado
Carlos Abs BianchiDelano Moody Simões da SilvaEverton Amancio dos SantosFlávia dos Santos PintoGuilherme H. B. de MirandaSaulo Marques de Abreu AndradeTerezinha Aparecida B. Dias
Alunos do Curso de Doutorado
Rosane Garcia Colevatti PereiraFelipe A. P. L. Costa

1
PROJETOS
DE
UM DIA

2
DISTRIBUIÇÃO DIFERENCIAL DE INSETOSGALHADORES E MINADORES ENTREHABITATS1
Flávia S. Pinto; Guilherme H. B. de Miranda; SauloM. A. Andrade.
INTRODUÇÃOGalhas são estruturas onde as larvas de
insetos se desenvolvem. Nestas estruturas, seja nafolha ou no caule, ocorre uma hiperplasia seguida deuma hipertrofia originando um tipo de câncer que éprejudicial ao crescimento e à reprodução da planta,uma vez que os nutrientes são drenados para a galha.Insetos minadores são indivíduos que similarmenteaos galhadores depositam sua larva na folha e estacome o tecido paliçado da folha, formando assim"trilhas" ou "túneis" na lâmina foliar.
As hipóteses que tentam explicar asadaptações ecológicas das galhas são (1) que estasseriam um dreno das substâncias das quais as larvasse alimentam, (2) que estas formariam um micro-ambiente para a larva, protegendo-a de alteraçõesabruptas do ambiente e, (3) também seriam umaproteção contra inimigos como parasitóides,predadores e patógenos.
Sabe-se que há uma distribuição diferencialdas galhas em ambientes xéricos (limitaçãohidronutricional) e mésicos, uma vez que o insetogalhador escolhe o melhor habitat para a exposição.Dessa forma, propôs-se verificar se a limitaçãohídrica e nutricional é importante na distribuição degalhas de insetos em habitats diferindo na qualidadehídrico-nutricional.
MATERIAL E MÉTODOSEm uma área de cerrado da Reserva
Ecológica do IBGE, tomado aqui como o ambientexérico, foram feitas 3 coletas de 20 minutos,coletando-se todas as galhas e minadores encontrados(totalizando 9 coletas, considerando cada espécie deplanta uma galha). O mesmo foi feito para a mata degaleria, tomada aqui como ambiente mésico. Todasas espécies de galhas e minas foram contadas eseparadas por habitat.
RESULTADOS E DISCUSÃOA Tabela 1 mostra que o número de galhas
foi maior no cerrado do que na mata. Enquanto quenão foi observada diferença nos minadores (Tab. 1,Figura1). Utilizou-se o teste do Wilcoxon paracomparar a mata e o cerrado: a diferença foi
1Prof. Orientador: Geraldo W. Fernandes
significativa para galhas (p=0,009), mas não foi paraminas (p=0,352). Estes resultados indicam queinsetos galhadores usam preferencialmente ambientesxéricos, provavelmente porque o parasitismo,predação e a ação de patógenos são bem menores doque no mésico. E que apesar da limitação hídrica enutricional ser bem acentuada no cerrado, o"dreno"que se forma na galha, supre a larvaplenamente (G. W. Fernandes, com. pess.)
Sugere-se também que a concentração detanino, maior em espécies de ambiente xéricos,possibilite uma maior sobrevivência, uma vez que ospatógenos são suscetíveis ao tanino, bem como aambientes secos, já que a ação de patógenos, comofungos, é potencializada em ambientes úmidos.
Um dado importante a ser levantado é que,mesmo com um maior número de espécies por metroquadrado , a mata possui menos galhadores,sugerindo que a ação de insetos galhadoresindepende da riqueza da flora, ou então que a pressãoseletiva sobre grande parte das galhas presentes namata fez com que apenas uma pequena parteresistisse à ação de parasitas, predadores e patógenose, que insetos que no passado colocaram suas larvasem espécies de mata, hoje usam espécies do cerrado.
Pode-se interpretar esta distribuiçãodiferencial como resultante da maior sobrevivência emenor mortalidade das galhas em ambientes xéricosdo em em mésicos. Por outro lado, a diferença nãosignificativa encontrada para o número de minadoresentre cerrado e mata pode ser interpretada supondo-se que as larvas dos minadores não ficamestabelecidas em galhas mas, sim, percorrem a falhapara alimentar-se. Esperaria-se então que estasbuscassem um padrão mais estável, com menos ounenhuma limitação seja hídrica, térmica ounutricional.
Dessa forma, esperaria-se encontrar ummaior número de minas na mata do que no cerrado,porém este padrão não foi observado, o que seria dese supor que talvez a pressão seletiva porparasitismo, predação ou ação de patógenos, bemcomo por estresse hídrico, não seja importante nataxa de sobrevivência e mortalidade econsequentemente não há uma distribuiçãodiferencial.

3
���������������������������������������������������������������
������������������������������������������������������������������������������������������������������������������������
NÚ
MER
O M
ÉDIO
DE
ESPÉ
CIE
S
0
5
10
15
20
GALHAS MINAS
CERRADO���� MATA DE GALERIA
Figura 1. Número médio de espécies de galhas e minas em plantas do cerrado e mata de galeria.
Tabela 1. Número de espécies de galhadores e minadores em cerrado e mata de galeria da Reserva Ecológica doIBGE.
cerrado mata de galeriaamostra galhadores minadores galhadores minadores
1 9 6 1 52 12 8 4 83 13 4 4 84 13 4 2 45 9 1 4 66 10 3 5 97 15 20 5 98 15 9 7 159 14 11 5 15
total 110 66 37 79x 12,22 7,33 4,11 8,78se 0,80 1,90 0,58 1,31
Tabela 2. Comparação das floras de mata de galeria e do cerrado (sensu lato) da Reserva Ecológica do IBGE (R. P.B. Henriques, dados não publicados).
tipo devegetação
área (ha) família gênero espécie espécie / km2 espécie /gênero
cerrado 981 83 318 742 75,6 2,3mata degaleria
104 110 290 529 508,6 1,8

4
VISITAÇÃO DE FORMIGAS EM NECTÁRIOSEXTRAFLORAIS DE Qualea multiflora(VOCHISIACEAE) E SEU POTENCIAL COMOAGENTE ANTI-HERBIVORIA2.
Delano M. S. da Silva; Rosane G. Colevatti; EvertonA. dos Santos.
INTRODUÇÃOInterações entre formigas e plantas com
nectários extraflorais (NEF) são bem descritas naliteratura. Plantas com NEF são visitadas porformigas com dietas bem amplas, as quais passeiampela planta (patrulham) atrás do néctar produzidopelos nectários. Em "troca" desse néctar as formigasprotegeriam estas plantas contra insetos herbívoros.Nectários extraflorais têm uma grande ocorrência emespécies lenhosas do cerrado, cerca de 25%,ocorrendo numa grande variedade de táxons. Paraalgumas destas espécies já foi demonstrado que apresença de formigas na planta diminui a taxa deherbivoria nas folhas e ou flores, podendo inclusiveaumentar o sucesso reprodutivo da planta.
Este trabalho tem como objetivos verificarse: (1) plantas com NEF são mais visitadas porformigas do que plantas sem NEF; (2) o padrão deforrageamento das formigas sobre as plantas estárelacionado à localização dos NEF na folhagem daplanta; (3) formigas exibem comportamentoagressivo em relação a herbívoros potenciaispresentes nas plantas.
MATERIAL E MÉTODOSO trabalho foi realizado numa área de
cerrado denso próximo à sede da reserva. A plantacom NEF utilizada foi Qualea multiflora(Vochysiaceae), uma planta com porte arbustivoarbóreo muito comum no cerrado do DF. Suas folhassão opostas e seus nectários localizam-se na inserçãodo pecíolo com o ramo. Foram utilizados 22indivíduos de Q. multiflora e 22 indivíduos de outrasespécies próximas que não possuíam NEF comocontrole. Em cada planta foram contados o númerode formigas presentes na planta. Após esta contagemutilizamos cupins para simular um herbívoro empotencial e testamos a eficiência das formigas comoprotetores. Estes cupins eram colados na face adaxialde duas folhas com cola branca. Para verificar se aeficiência das formigas como protetoras era igualpara toda a planta, pois os nectários só estão ativosem folhas jovens, utilizamos um par de folhas jovense um par de folhas velhas, tanto em Q. multifloraquanto no vizinho sem NEF.
2 Prof. Orientador: Paulo Oliveira
Após colocarmos os cupins, acompanhamoso comportamento das formigas em relação aos cupinsdurante 10 minutos. Todo evento que ocorreu nesseintervalo era anotado e ao final dos 10 minutos asformigas eram coletadas para posterior identificação.
RESULTADOS E DISCUSSÃOA tabela 1 apresenta as espécies de formigas
encontradas tanto em Qualea multiflora quanto naplanta vizinha. Dentro destas espécies, Camponotusrufipes, Camponotus crassus e Zacryptoceruspusillus foram as mais abundantes. Com relação aonúmero de formigas encontradas patrulhando asplantas observadas, encontramos que Q. multiforapossui um número significativamente maior deformigas do que as plantas vizinhas sem NEF (Tab.2). Além de visitarem mais as plantas com NEF asformigas atacam mais cupins (iscas) nas plantas com
NEF do que nas plantas sem NEF (2χ =9.95;
p=0,0016). Ambos resultados conferem com osresultados encontrados na literatura mostrando aeficiência das formigas como agentes anti-herbívoros.
Podemos concluir que as formigas visitammais Qualea multiflora do que plantas sem NEF eque além disso defendem melhor as plantas com NEF(Objetivos 1 e 3), porém quando comparamos se esteataque é maior em folhas novas (NEF ativos) que emfolhas velhas, verificamos que não existe diferença
significativa (2χ =0,11; p=0,7385). Poderíamos
pensar que apesar de não existir diferença do ataquepoderia existir uma diferença do tempo para ocorrer oataque, mas este também não mostra uma diferençasignificativa (Komogorov-Smirnov; Dmax=0,17;p=1,0; Fig. 2).
Desse modo, mais uma vez foi comprovadoque formigas que visitam NEF's atuam como agentesanti-herbívoros para estas plantas.

5
TEMPO (MINUTOS)FR
EQU
ÊNC
IA (%
)0
12
34
5
1 2 3 4 5 6 7
Folha novaFolha velha
FIGURA 1. Tempo de ataque ao cupins pelas formigas em folhas novas e velhas de Qualea multiflora.
Tabela 1. Espécies encontradas em Qualea multiflora e nas espécies vizinhas.EspécieCamponotus blandusCamponotus crassusCamponotus rufipesCamponotus spCrematogaster spPachycondyla villosaPseudomyrmex aff. flavidulusPseudomyrmex aff. gracillisPheidole spZacryptocerus pusillus
Tabela 2. Número médio de formigas encontradas em cada espécie de planta.ESPÉCIE x sd U pQualea multiflora 6,64 4,60 25,2153 <0,001Vizinho maispróximo
1,18 1,14

6
OCORRÊNCIA DE FORMIGAS EM GALHASDE Diospyros hispida D. C. NA RESERVAECOLÓGICA DO IBGE, DF3
Carlos A. Bianchi; Felipe A. P. L. Costa; TerezinhaA. B. Dias.
INTRODUÇÃOGalhas ocorrem em espécies de plantas,
provocadas pela deposição de larvas de insetos(chamados galhadores), promovendo uma hiperplasiae/ou hipertrofia no local da deposição (geralmentefolhas e caule). As galhas entre outras funções,podem servir de abrigo e fornecer nutrientes para odesenvolvimento da larva. Entretanto é possívelregistrar a utilização das galhas por outrosinvertebrados, os quais podem predar ou não a larvado galhador, ocupando o espaço disponível nointerior da mesma. Neste caso, a galha tambémfuncionaria como um abrigo. Dentre aquelesocupantes de galhas, podem ser citadas inúmerasespécies de formigas.
Este trabalho analisa a ocorrência deformigas em galhas de Diospyros hispida DC. Asquestões a serem respondidas por este estudo são: (1)quais são as espécies de formigas ocupantes degalhas de D. hispida ; (2) as galhas ocupadas sãomaiores do que as galhas não ocupadas?; (3) onúmero de formigas aumenta com o tamanho dagalha?; (4) o número de formigas varia com otamanho da galha ocupada?
MATERIAL E MÉTODOSForam coletadas 71 galhas em 18 indivíduos
de D. hispida(entre 3 a 7 galhas por indivíduo) numaárea de cerrado próxima à sede da Reserva Ecológicado IBGE, Distrito Federal. Cada galha coletada foiacondicionada em sacos de papel individuais e,posteriormente, triada no laboratório. Foi efetuada acontagem dos orifícios presentes nas galhas,estimado seu volume total, através da imersão emágua em proveta graduada (100 ml e 500 ml) ecaracterizada sua arquitetura interna (número decavidades). Após a abertura da galha, o número e aespécie de formigas presentes foram registrados,além da presença ou ausência da larva do galhador.
RESULTADOS E DISCUSSÃODentre as 71 galhas coletadas, 31 delas
estavam ocupadas por formigas, 34 não estavamocupadas por formigas e 6 foram classificadas como"podres" (quando estavam vazias e bastantequebradiças) (Fig. 1). Dos 18 indivíduos deDiospyros hispida, dos quais foram coletadas galhas, 3 Prof. Orientador: Geraldo W. Fernandes
14 apresentaram galhas ocupadas por formigas(totalizando 15 galhas) de duas ou mais espécies.Segundo os resultados apresentados na figura 1, 31galhas apresentaram formigas e, destas, 15apresentaram 2 ou mais espécies de formigas.
Foram encontradas 7 espécies de formigasem 5 gêneros, como ocupantes das galhas deDiospyros hispida. Um estudo similar realizado emárea de cerrado próximo à Belo Horizonte, MinasGerais, revelou a ocorrência de 7 espécies (6gêneros) (Tab. 3). Não houve diferença significativa(t=1,01; p>0.05; N=65) entre o volume das galhasocupadas por formigas (Tab. 1). A figura 2 mostraque existe correlação entre o número de formigaspresentes nas galhas e o volume de cada galha,apresentando gêneros como Zacryptocerus sp queocupou galhas de diferentes tamanhos com diferentesnúmeros de indivíduos.
Quando são comparadas as espécies maisabundantes, Zacryptocerus sp e Camponotus sp ,quanto ao volume das galhas que ocupavam, tambémnão se verifica diferença significativa (t=0,14;p>0.005; N=24) (Tab. 2).
Formigas de espécies diferentes ocupavam amesma galha (com entradas para cavidadesindependentes) em um dos indivíduos de D. hispida.Outro fato foi a presença de formigas do gêneroZacryptocerus sp ocupando uma cavidadeindependente em certa galha e em outra cavidade, apresença de uma larva (possivelmente do galhador)na mesma galha.
O estudo de ocorrência de formigas emgalhas de D. hispida revelou a existência de 7espécies de formiga como ocupantes das galhas.Estes resultados sugerem que as galhas são ambientesimportantes para o estabelecimento de colônias deformigas. Paralelamente, as demais plantas de D.hispida que apresentaram suas galhas ocupadas porapenas uma espécie, sugerem a possível existência decolônias destas espécies, espalhadas por 3 a 5 galhasda mesma planta.
Não houve diferença significativa entre ovolume das galhas ocupadas quando foram usadastodas as formigas ou foram usadas as duas espéciesmais abundantes (Zacryptocerus sp e Camponotussp). Pode-se concluir que não ocorre preferência portamanho de galhas, as formigas ocupam todasindependente do tamanho. Outra explicação para osresultados encontrados é que o número de formigaspoderá diferir com o tamanho da galha, dependendodo volume ocupado pelas formigas (seria necessáriocalcular o volume dos indivíduos que formam acolônia).

7
���������������������������������������������������������������
����������������������������������������������������������������������
NÚ
MER
O D
E G
ALH
AS
0
10
20
30
40
50
60
70
80
A B C D
Número total degalhas
���Número de galhasocupadas���
��� Número de galhasnão ocupadasNúmero de galhaspodres
FIGURA 1. Número de galhas coletadas e distribuição nas quatro categorias de galhas.
TAMANHO DA GALHA (ML)
NÚ
MER
O D
E FO
RM
IGAS
0
20
40
60
80
100
120
140
0 10 20 30 40
FIGURA 2. Relação entre o tamanho das galhas e o número de formigas (r=0,39; P < 0,05; N=23).

8
Tabela 1. Média e desvio padrão para o volume de galhas ocupadas e não ocupadas.valor ocupadas não-ocupadasx 9,9 11,7s 7,6 7,6t=1,01; p>0.05; N=65
Tabela 2. Comparação entre o volume das galhas ocupadas pelas espécies Camponotus sp e Zacryptocerus spCamponotus sp Zacryptocerus sp
N 7,0 17x 13,4 10,4d.p 11,9 6,4t=0,14; p>0,05; N=65
Tabela 3. Lista de gêneros e espécies na área do cerrado da reserva ecológica do IBGE e de um cerrado de BeloHorizonte.Brasília (IBGE) Belo HorizonteCamponotus pallescens Camponotus sp1Crematogaster sp. Camponotus sp2 Pseudomyrmex aff. gracilis Crematogaster brevispinosaSolenopsis sp1 P. gracilisSolenopsis sp2 Solenopsis sp1Zacryptocerus pusillus Lepthotorax spZacryptocerus sp1 Zacryptocerus pusillus

9
ESTRATÉGIAS DE FORRAGEAMENTO DELAGARTOS4
Flávia S. Pinto; Saulo M. A. Andrade; Guilherme H.B. de Miranda
INTRODUÇÃODuas estratégias de forrageamento são
conhecidas para lagartos não-herbívoros. As duasbaseiam-se nas relações de custo e benefício quevisam minimizar gastos energéticos, no caso, com aprocura de itens alimentares. Os "senta-e-espera" sãocaracterizados por apresentarem sítios definidos deforrageamento, empreendendo pequenosdeslocamentos para a obtenção de presas. São, porisso, geralmente territoriais e utilizam principalmenteo estímulo visual na procura de presas. Osforrageadores ativos, ou "procuradores", sãocaracterizados por não apresentarem sítios definidosde forrageamento, saindo em busca das presas,utilizando principalmente o estímulo olfativo nestaprocura.
Estas duas estratégias, devido à demandaenergética que está envolvida, determinariam o tipo ea quantidade das presas ingeridas. Teoricamentelagartos do tipo "senta-e-espera" apresentariammenor diversidade de itens na sua dieta, compreferência por alguns desses itens. Isto é esperadopois estes lagartos apresentam baixo gasto energéticocom a procura dos itens, podendo energeticamenteesperar presas de maior retorno energético. Osprocuradores, por sua vez, sujeitos a um déficitenergético imposto pela procura ativa do alimento,apresentariam na composição da sua dieta maiordiversidade de itens alimentares de diferentesretornos energéticos.
O objetivo principal deste trabalho éverificar a existência desses padrões para duasespécies de lagartos do cerrado, uma considerada"senta-e-espera" e um modelo aleatório por nósproduzido, através de medidas de frequência dedeslocamento e do estímulo utilizado na procura daspresas.
MATERIAL E MÉTODOSOs dados foram obtidos através de censos
realizados de minuto a minuto a partir davisualização direta dos lagartos. Foram registradosdois tipos de comportamento com relação à atividadedesenvolvida no momento da observação (parado ouandando) e caracterizado o tipo de estímulo utilizadona procura de presas, através de visualizaçãocontínua, quantificados a partir do número de
4 Prof. Orientador: Guarino R. Colli
investidas em presas, considerando estímulos visuaisou olfativos.
Os dados foram coletados na região daCachoeira de Queimados no Município de Unaí(MG). A região é caracterizada pela existência dematas de galeria associadas à afloramentos de rochaàs margens do rio.
Escolhemos para este estudo a espécieTropidurus oreadicus (Tropiduridae) bastantefrequente na área, sabidamente ”senta-e-espera”. Paratraçarmos comparações entre os dois modelospropostos criamos um modelo aleatório, que atribuiuma mesma probabilidade de, para cada observação,um lagarto imaginário estar parado (50%) ouandando (50%), com relação ao padrão de atividade,e de utilizar um estímulo visual (50%) ou olfativo(50%), com relação ao estímulo utilizado. Os dadosobservados e os dados do modelo foram confrontadosatravés do teste t pareado.
RESULTADOS E DISCUSSÃOOs dados brutos do censo realizado para
atividade e estímulo de T. oreadicus são mostradosna tabela 1. Os mesmos dados obtidos para o modeloaleatório criado estão representados na tabela 2.Foram observados ao todo 20 indivíduos de T.oreadicus. Ao todo foram 260 observações parapadrão de atividade e 64 observações para estímuloutilizado.
O modelo não apresentou nenhum padrãocomportamental segundo a teoria de forrageamentoproposta. Em aproximadamente metade dasobservações os indivíduos estavam parados (45,8%)ou andando (54,2%) e utilizaram para a visualizaçãodas presas tanto o estímulo visual (54,2%) quanto oolfativo (48,4%).
Encontramos diferença significativa nopadrão de atividade quando comparamos o númerode observações de indivíduos encontrados parados(t=-5,48 p=0,0001), o que corrobora o fato doslagartos considerados "senta-e-espera" apresentarembaixa frequência de deslocamento.
Devido à grande abundância de T. oreadicusna área vários comportamentos agonísticos foramobservados entre os indivíduos que habitavam locaispróximos mostrando a existência de territorialidadepara a população amostrada, corroborando o modeloproposto para "senta-e-espera".
A fim de identificar realmente a existênciadestes padrões, estudos considerando várias espéciesdiferentes e de diferentes grupos taxonômicos devemser realizados, a fim de descartar possíveis relaçõesfilogenéticas existentes e abrangendo maior variaçãode padrões comportamentais. Além disso outrascaracterísticas devem ser consideradas como onúmero e a quantidade de itens alimentares ingeridos.

10
Tabela 1. Padrão de atividade e estímulo utilizado por Tropidurus oreadicus. Os valores indicam o número deobservações.
atividade estímuloindivíduo parado andando visual olfativo
01 10 0 2 002 12 2 6 003 10 0 1 004 1 2 2 005 3 0 0 006 1 0 1 007 2 0 1 008 10 0 0 009 7 4 15 010 1 1 1 011 6 4 17 012 11 0 2 013 6 0 5 014 13 0 1 015 31 16 3 016 25 0 0 017 19 2 - -18 17 6 - -19 25 4 2 020 5 4 5 0
total 215 45 64 0
Tabela 2. Padrão de atividade e estímulo utilizado pelo modelo aleatório.atividade estímulo
indivíduo parado andando visual olfativo01 5 5 2 002 6 8 2 403 5 5 0 104 1 2 1 105 3 0 0 006 0 1 0 107 2 0 0 108 2 8 0 009 5 6 8 710 0 2 0 111 7 3 10 712 6 5 11 113 1 5 4 114 6 7 0 115 24 23 1 216 17 8 0 017 7 14 - -18 7 16 - -19 9 20 1 120 6 3 1 1
total 119 141 31 33

11
DIVERSIDADE DE ORTHOPTERARELACIONADA À HETEROGENEIDADEESPACIAL5
Carlos A. Bianchi; Felipe A. P. L. Costa; TerezinhaA. B. Dias.
INTRODUÇÃOA heterogeneidade espacial é importante na
determinação da diversidade de espécies. Variaçõesde tipo de solo acarretam mudanças na vegetação e,consequentemente, o mesmo efeito na fauna. Logo,ambientes mais heterogêneos tendem a suportar ummaior número de espécies em relação à ambientesmais homogêneos, isto é, um aumento naheterogeneidade espacial pode representar umaumento na diversidade de espécies de flora e fauna.A diversidade de espécies pode ser caracterizada de 3formas: a diversidade α (alfa) ou diversidadepontual, que se refere à diversidade de um pontoamostrado; a diversidade β (beta), que avalia oquanto a composição de espécies mudou de um pontoamostrado para outro e por fim a diversidade gama,que representa a diversidade regional.
O cálculo de diversidade beta é feito atravésdo Índice Whittaker (1960), dado pela fórmula:
Bw = (c / α ) −1
onde: c=riqueza total de espécies das duas áreas eα = soma do número de espécies das duas áreasdividida por 2.
Este projeto teve por objetivos verificar acomposição de espécies de Orthoptera em uma áreade campo sujo, avaliando e investigando relaçõescom a existência de um gradiente na vegetação.
MATERIAL E MÉTODOSO estudo foi realizado em uma área de
campo sujo na Reserva Ecológica do IBGE, Brasília,DF; onde foram amostrados 6 pontos. Para cadaponto efetuamos um levantamento da vegetação,classificando as espécies ocorrentes em 3 categorias:arbórea, arbustiva e herbácea; indicando o número deespécies, o número de indivíduos para cada categoria,além da altura do estrato.
Os cinco primeiros pontos estavam distantes50 m um do outro, a distância do ponto 5 ao ponto 6era de 200 m. Posteriormente 75 varreduras foramefetuados num raio de 30 m do ponto amostrado,coletando apenas os insetos Acridoidea (gafanhotos).Em seguida, foram cronometrados 15 minutos parauma nova coleta de gafanhotos, a qual era efetuada
5 Prof. Orientador: Alexandre F. B. de Araújo
com a localização visual de um exemplar econsequentemente captura com rede de varredura. Osespécimes coletados foram separados emmorfoespécies.
RESULTADOS E DISCUSSÃOForam identificadas 52 morfoespécies, num
total de 213 indivíduos coletados. A tabela 1 mostra onúmero de morfoespécies e respectivo número deindivíduos coletados para cada ponto. A figura 1apresenta uma relação entre o número de espécies e adistância percorrida. A curva apresentada na figura 1mostra uma tendência à estabilização entre 50 e 60morfoespécies, indicando que o número máximo deespécies para a área pode estar próximo destesvalores.
A diversidade beta entre os pontosamostrados é mostrada na tabela 2. Os valoresencontrados situam-se entre 0,44 (pontos 5,6) até 1,0(pontos 1,4 e 1,6). Não foi verificado um padrão demudanças na diversidade entre as áreas amostradas.Áreas próximas (dist=50 m) apresentaram tantovalores altos de dissimilaridade (Bw1,2 =0,83) comovalores baixos (Bw2,3=0,45). Do mesmo modo,áreas distantes apresentaram valores altos(Bw1,6=1,0) (dist=400 m) e valores baixos(Bw5,6=0,44; dist=200 m).
Os dados de vegetação da tabela 3 podemser agrupados em pares conforme riqueza edensidade em cada ponto. Desta forma, podemosafirmar que os pontos 1 e 3 são semelhantes emriqueza de espécies e densidade de indivíduos,porém, quanto aos Orthoptera, encontramos altovalor de dissimilaridade (Bw1,3=0,83; tabela 2).
Os pares de pontos 2 e 4, e 2 e 5 foramagrupados de acordo com riqueza e densidade,respectivamente. Ambos apresentaram valoresmédios de dissimilaridade, situados em torno de0,615 ((Bw2,4 + Bw2,5)/2). Quanto aos pares 4 e 6, e5 e 6, podemos encontrar o mesmo padrão, com valormédio de dissimilaridade em torno de 0,55 ((Bw4,6 +Bw5,6)/2).
A figura 2 apresenta os índices dedissimilaridade ao longo dos pontos amostrados, comvalor alto de P1 a P2 (Bw1,3=0,83) e valores médiosde P2 até P6 (v. figura 2). Apesar do número baixode amostras de vegetação, o padrão encontrado foiequivalente em todos os pontos, revelando umavariação bastante sutil entre pontos. Quanto àcomposição de espécies de gafanhotos de cada ponto,é possível afirmar que tais variações estavam, decerta forma, relacionadas às diferenças na vegetação,entretanto, deve-se ressaltar novamente o númeroamostral utilizado.

12
Tabela 1. Número de espécies e indivíduos coletados na área de estudo.ponto nº espécies nº indivíduos
P1 08 13P2 16 32P3 21 38P4 10 29P5 27 46P6 20 46
Tabela 2. Valores de diversidade usando o índice de Whittaker (1960) (Bw) na diagonal inferior e número deespécies comuns na diagonal superior
pontos amostradosP1 P2 P3 P4 P5 P6
P1 - 2 2 0 1 0P2 0,83 - 10 5 8 9P3 0,86 0,45 - 5 10 9P4 1,00 0,61 0,67 - 8 5P5 0,94 0,62 0,58 0,56 - 13P6 1,00 0,50 0,56 0,66 0,44 -
Tabela 3. Valores de densidade e riqueza de espécies para a vegetação nos pontos amostrados.ponto densidade (ind./m2) riqueza (nspp) agrup.dens.riq.
P1 2,0 14 (1,3)P2 3,3 24 (1,3)P3 2,0 15 (2,5)P4 2,8 23 (2,4)P5 3,3 19 (4,6)P6 2,8 20 (5,6)

13
DISTÂNCIA (m)
0
10
20
30
40
50
60
50 100 150 200 250 300 350 400 450
FIGURA 1. Relação entre o número de espécies e a distância.
P1 P2 P3 P4 P5 P6/____0,83____/____0,45___/____0,67____/____0,56____/____0,44___/
50m 50m 50m 50m 50m
FIGURA 2. Índice de dissimilaridade de Whittaker (Bw) ao longo da área de estudo.

14
ECOLOGIA FLORAL DE QUATRO ESPÉCIESDO CERRADO: FREQUÊNCIA DE VISITAS EPOLINIZADORES6
Delano M. S. da Silva; Everton A. dos Santos;Rosane G. Colevatti.
INTRODUÇÃOA polinização cruzada entre diferentes
espécies de plantas, mesmo que não resulte emfertilização ou formação de híbridos, pode diminuir ovalor adaptativo ("fitness") da planta. Isso podeocorrer tanto no componente masculino, por perda depólen, quanto no feminino, por ocupação do espaçoestigmático com pólen de outras espécies, impedindoa germinação de pólen co-específico.
As comunidades vegetais podem apresentardiversos mecanismos que impedem a polinizaçãocruzada, como: (1) florescimento em épocasdiferentes do ano; (2) ântese (cobertura da flor) emdiferentes horários do dia; (3) utilização de diferentestipos de polinizadores (abelhas, beija-flores,morcegos, etc); (4) partilha de polinizadores pelasespécies vegetais. As diferenças na morfologia floral,proporcionam uma deposição diferencial do pólen decada planta no corpo do polinizador, que correspondeà posição de recepção pelo estigma. Assim, ummesmo polinizador que visita várias espécies deplantas de uma comunidade pode apresentar pólen decada espécie de planta em diferentes partes do corpo.Além disso, espera-se que espécies com maior ofertade "recompensa" pelo "serviço" do polinizador(maior número de flores por planta, maior quantidadede néctar, pólen, óleo) apresentem uma maiorfrequência de visitas.
Foram encontradas, em uma área de cerradoquatro espécies de plantas florindo em uma mesmaépoca do ano (março), e com flores abertas durante omesmo período do dia. Dessa forma, foi levantada aquestão que foi o objetivo geral desse trabalho: hápartição do recurso polinizador por estas quatroespécies de plantas co-ocorrentes? Especificamente,este trabalho tem como objetivo responder àsseguintes questões para cada espécie vegetal: (1)quais os visitantes florais?; (2) quais os recursosoferecidos pela planta e utilizado pelos visitantes?;(3) qual a frequência das visitas?; (4) quais são osprincipais polinizadores de cada espécie?; (5) essespolinizadores são comuns às quatro espécies? Casosejam, há partilha do recurso polinizador?
MATERIAL E MÉTODOSEste trabalho foi realizado na Reserva
Ecológica do IBGE, numa área de cerrado onde co- 6 Prof. Orientador: Jader S. Marinho Filho
ocorriam quatro espécies com hábito arbustivo-herbáceo: Solanum lycocarpum (Solanaceae), Hyptissp (Labiatae), Diplusodon oblongus (Lythraceae) eBanisteriopsis campestris (Malpighiaceae).
Para observação dos visitantes foi utilizadoo método "focal contínuo" de observação, no qualcada espécie foi observada no período entre 8:10 e11:30 h por um observador. Foram registradas asseguintes observações: (1) espécie visitante; (2)horário da visita; (3) recurso utilizado; (4)comportamento de coleta do recurso. As frequênciasforam analisadas pela análise de variância não-paramétrica (Kruskal-Wallis) e teste para diferençaentre medianas. Não foi possível analisar asdistribuições de frequências de visitas por espécie deabelha e frequência total devido ao número deobservações.
RESULTADOS E DISCUSSÃOA tabela 1 apresenta os visitantes
encontrados nas quatro espécies estudadas e osprincipais polinizadores. Os recursos encontrados emcada flor e sua morfologia estão descritos abaixo:
- Solanum lycocarpum (Solanaceae): possuianteras poricidas. Embora as flores sejammorfologicamente hermafroditas, há evidências deque há indivíduos funcionalmente femininos,masculinos e hermafroditas. As flores não possuemnéctar, sendo o pólen o único recurso obtido pelasabelhas pela vibração das anteras ("buzzpollination"). A flor é actinomorfa e roxa.
- Hyptis sp (Labiatae): apresenta florzigomorfa, com as anteras dentro da corola. Essas sãoprojetadas para fora quando a abelha "força" a flor,pousando na "plataforma" que possui guias de néctarpúrpura, contrastando com a flor branca.Aparentemente há dicogamia (protandria), pois aprojeção do estigma para a parte externa da flor sófoi encontrada em flores cujas anteras estavammurchas.
- Diplusodum oblongus (Lythraceae): a floré púrpura e actinomorfa. O estigma é projetado acimada linha das anteras e, aparentemente, deve encostarna cabeça dos visitantes de tamanho corporal maiorquando estão coletando néctar.
- Banisteriopsis campestris (Malpighiaceae):A flor é branca, actinomorfa e possui 5 glândulas deóleo nas sépalas, que é o recurso principal coletadonessa flor, além do pólen. O gineceu é dialicarpelar(tricarpelar).
A figura 1 mostra a frequência de visitas porintervalos de tempo de 30 min., para cada espécie deplanta, somando todos os visitantes florais. Essasfrequências foram diferentes, pela análise devariância de Kruskall-Wallis (Kw=14,5966; p=0,0022). O teste para medianas mostra que hádiferenças no pico de visitação entre S. lycocarpum e

15
Hyptis sp (2χ =7,14; mediana=5,6; p=0,0075), S.
lycocarpum e B. campestris (2χ =7,14;
mediana=5,5; p=0,0075); Hyptis sp e D. oblongus
(2χ =5,33; mediana=60; p=0,0209), D. oblongus e
B. campestris (2χ =7,14; mediana=7,5; p=0,0075),
mas não entre S. lycocarpum e D. oblongus
(2χ =2,40; mediana=2,0; p=0,1217) e Hyptis sp e B.
campestris (2χ =2,57; mediana=11; p=1,1088).
O principal polinizador potencial de S.lycocarpum foi Epicharis sp Em Hyptis sp foi oLepidoptera Lycaenidae sp1., Epicharis sp eMyschocytharus sp (Vespidae). Para D. oblongusforam as espécies de abelha Trygona spineps,Epicharis sp e Megachilidae sp. Para B. campestris,Epicharis sp Anthophoridae sp1 e um grupo deabelhas Trygona spineps, Paratetrapedia sp1 e sp2,as quais foram reunidas na contagem de visitas, umavez que era impossível distinguí-las no campo. Afigura 2 mostra a distribuição de frequência de visitasdos principais polinizadores, para cada planta.
Como Epicharis sp foi a espécie comumentre as quatro espécies de plantas, foi analisada adiferença entre a frequência de visitas para S.lycocarpum e B. campestris, uma vez que nas outrasduas espécies a frequência de visitas foi muito baixa.A análise de Mann-Whitney mostrou que não hádiferença entre elas (U=1,4610; p=0,2268).
Apesar de não ter sido possível realizar umaanálise para diferença nas distribuições de frequênciade visita de Epicharis sp nessas duas plantas, ográfico sugere (Fig. 2) que quando há um aumento defrequência de visitas em uma das espécies há umadiminuição na outra. Além disso a frequência total devisitas e a frequência de visita de Epicharis sp em B.campestris foi bem maior que as outras espécies. Issoocorreu, provavelmente, porque B. campestrisapresentava um grande número de flores por ramo euma alta densidade de flores no que foi considerado a"mancha" da planta. Dessa forma, Epicharis sp alémdas outras abelhas, visitam várias flores dessa planta.Esse padrão de maior frequência de visitas a plantascom maior densidade de flores é bastante comum.Além disso Epicharis sp é uma espécie coletora deóleo, o que pode explicar sua maior frequência em B.campestris.
Quanto à partição do recurso polinizador,considerando principalmente Epicharis sp, não foramencontradas evidências de deposição diferencial depólen das várias espécies. Aparentemente Epicharissp coletou principalmente óleo em B. campestris,pólen em S. lycocarpum e néctar nas outras duas
espécies. Entretanto, um estudo mais detalhado, comcoleta de pólen das plantas e exame do pólen nocorpo da abelha, será necessário para conclusõesmais precisas. Além disso, foram agrupadosindivíduos machos e fêmeas dessa espécie de abelha.É possível que machos e fêmeas estivessemcoletando recursos diferentes, como é comum paraabelhas de outros grupos como Euglossini,Anthophorini, etc.
������������
������������
��������
��������
��������
�������������������� ����
����������������
������������
��������������������
���������
��������
������������������������
��������������������������������
INTERVALO DE TEMPO
0
5
10
15
20
25
1 2 3 4 5 6 7
Solanum lycocarpum���Diplusodon obolgus
������Hyptis sp.
Banisteriopsis campestris
FIGURA 1. Frequência de visitas nas flores porintervalo de tempo para cada espécie de plantaestudada. Intervalos 1: 8:10-8:39, 2: 8:40-9:09, 3:9:10-9:39, 4: 9:40-10:09, 5:10:10-1039, 6: 10:40-11:09, 7: 11:10-11:39.

16
FIGURA 2. Frequência de visitas por intervalos de tempo para os principais polinizadores de cada espécie deplanta. A. Solanum lycocarpum; B. Hyptis sp; C. Diplusodon oblongus; D. Banisteriopsis campestris.
Epicharis
02468
1012
1 2 3 4 5 6 7
A
��������
���������������
02468
1012
Myschocytharus sp�������� Lycaenidae����
Epicharis sp
B
����������
��������
��������������������������������
��������
024
68
1012
Trigona spineps�������� Epicharis sp.���� Megachilidaesp1
C
����������
���������������
02468
1012
Myschocytharus sp�������� Lycaenidae����
Epicharis sp.
D

17
APOSEMATISMO: ADVERTÊNCIA OUATRAÇÃO?7
Carlos A. Bianchi; Felipe A. P. L. Costa; TerezinhaA. B. Dias.
INTRODUÇÃOCamuflagem e aposematismo são
consequências evolutivas da predação pororganismos que se orientam visualmente. Enquanto acamuflagem diminui as chances de que uma presapotencial seja encontrada, o aposematismo diminui aschances de que um animal impalatável seja injuriadopor predadores generalistas.
Para testar a hipótese de que larvasaposemáticas de insetos são evitadas por avesinsetívoras, foi conduzido um experimento utilizandolarvas artificiais. Especificamente, foi testado se: (1)os danos e/ou a remoção seriam mais frequentes naslarvas monocromáticas (verdes ou amarelas) do quenas larvas coloridas (=aposemáticas; larvas pretascom 2 faixas transversais, uma vermelha e outraalaranjada); (2) os danos e/ou a remoção nas larvasevidentes (amarelas, contrastando com o fundo verdedas folhas) seriam mais frequentes do que nas larvascamufladas (verdes) e aposemáticas. Em resumo, nóstestamos se a frequência de danos foi decrescenteentre os 3 tipos de larvas, a saber:amarelas>verdes>coloridas.
MATERIAL E MÉTODOSLarvas artificiais (com 3,5 - 5,0 cm de
7 Profª. Orientadora: Regina Macedo
comprimento e 0,35 - 0,50 cm de diâmetro) foramfeitas com massa de modelar e colocadas sobre asfolhas de uma planta, quase sempre ao longo danervura principal.
Dentro de uma área de "cerradão", naReserva Ecológica do IBGE (Brasília, D.F.), nósestabelecemos 3 transectos paralelos e distantes 5metros entre si. Ao longo de cada um deles, foramdistribuídas, de modo alternado, 153 larvas (51amarelas, 52 verdes e 50 coloridas), cada uma sobreuma planta (alturas entre 40 - 130 cm).
Nós esperamos 3 horas para o início dasvistorias, durante a qual os danos encontrados emcada larva foram classificados de acordo com osuposto agente causador; formigas (geralmentepresentes), outros artrópodos (vespas) e aves.Especialmente nos 2 últimos casos, nós inferimos oagente dos danos de acordo com o aspecto dasmarcas encontradas. Ao final, apenas uma larva,entre as 153 colocadas, desapareceu e nós decidimosexcluí-la das análises posteriores.
RESULTADOS E DISCUSSÃODas 153 larvas distribuídas, 52 foram
reencontradas com algum tipo de dano. A ocorrênciade danos não difere das proporções relativas dos 3tipos de larvas (Tab. 1), sugerindo que as larvasforam danificadas independentemente da suacoloração. Assim, ao contrário do que era esperado,larvas "aposemáticas" foram tão frequentementedanificadas quanto larvas monocromáticas. Além
Tabela 1. Visitantes florais das quatro espécies de plantas estudadas (V) e polinizadores potenciais (P).visitante S. lycocarpum Hyptis sp D. oblongous B. campestrisBruchidae sp1 V V VChrysomelidae sp1 V VCurculionidae sp1 VMiridae sp1 VCamponotus sp1 VLycaenidae sp1 P V VBrachigaster sp1 VMyschocytharus sp P VAnthophoridae sp1 PAnthophoridae sp2 PAnthophoridae sp3 PAnthophoridae sp4 PEpicharis sp P P P PParatetrapedia sp1 PParatetrapedia sp2 PXylocopa sp PMegachilidae sp1 PNanotrigona sp VTrigona spineps P P Prazão p/(p+v) 4/4 4/6 4/6 5/14

18
disso, como as formigas foram responsáveis pelamaioria dos danos, esses resultados indicam aausência de discriminação por aqueles animais entreos 3 tipos de larvas.
Nesse ponto, nós reanalizamos os dados,comparando apenas as larvas danificas por "vespas"
contra todas as outras e o valor obtido para o 2χ foi
significativo (Tab. 2). Embora os baixos valores emalgumas células exijam uma análise mais cuidadosa,a magnitude das diferenças e sua natureza (ca. 60 %devido à célula "colorida-vespa") apontam na direçãode que as larvas aposemáticas forampreferencialmente danificadas.
Ainda que a composição da massa demodelar utilizada possa diferir com as cores (e terinfluenciado os resultados), nós imaginamos umaexplicação alternativa: larvas aposemáticas podemser mais facilmente encontradas e utilizadas poralguns artrópodos predadores. Sendo assim, étentador imaginar os benefícios que esses predadoresteriam utilizando um recurso alimentar (e.g., larvasaposemáticas) que é evitado ou pouco utilizado porpredadores vertebrados (e.g., aves).
Tabela 1. Comparação do número de larvasdanificadas de acordo com a sua coloração.cor nº larvas
coletadasnº larvas
danificadas1amarelo 51 17verde 52 17colorida 49 19total 152 531 Número esperado de larvas danificadas, de acordocom a sua abundância relativa.t=0,298 (g.l=2; p>0,05)
Tabela 2. Comparação das larvas danificadas porvespas contra os outros de diferentes cores.cor danos por
vespasoutros danos +
sem danosamarelo 2 49verde 1 51colorida 10 39total 13 139Kolmogorov-Smirnov, p<0.001)
19
INFLUÊNCIA DO FOGO ANUAL NAFLORÍSTICA E DENSIDADE DE ESPÉCIESLENHOSAS DE CERRADÃO8
Delano M. S. da Silva; Everton A. dos Santos;Rosane G. Colevatti.
INTRODUÇÃOO cerrado é uma savana tropical
caracterizada por uma vegetação rasteira, na qual hápredomínio de gramíneas, com árvores e arbustosesparsos. Este tipo de vegetação pode sercaracterizado ainda pela estacionalidade climática,com períodos secos bem marcados, onde é frequentea ocorrência de fogo. Atualmente, especula-se que aocorrência de fogo no cerrado seja um dos fatoresdeterminantes da vegetação. O objetivo deste estudofoi analisar a composição florística de duas áreas(uma afetada anualmente pelo fogo, e outraprotegida), a fim de investigar a influência do fogona composição, densidade e diversidade de espécieslenhosas.
MATERIAIS E MÉTODOSEste estudo foi realizado numa área de
cerrado denso, localizada na reserva ecológica doIBGE, e outra área adjacente, situada no JardimBotânico de Brasília. Selecionamos duas áreas, dasquais a primeira está protegida do fogo, tendo sofridoapenas uma queimada nos últimos 20 anos (em1994). A segunda área, por sua vez, sofre queimadasanuais. Estas duas áreas estão separadas entre siapenas por um aceiro de 20 m.
A densidade de plantas foi estimada usandoo método de ponto-quadrante. Em cada uma dasáreas, traçamos uma linha de 100m perpendicular aoaceiro. A cada cinco metros, medimos as distânciasdo ponto até o vizinho mais próximo de cadaquadrante, considerando a circunferência mínima dotronco de 15 cm. Ao todo, amostramos 20 pontos e80 indivíduos em cada área.
Calculamos o número de espécies e adensidade em cada área, as frequências absoluta erelativa e a densidade de cada espécie, comparamosas densidades absoluta e relativa das principaisespécies de cada área e, por fim, fizemos uma análisegráfica para comparar as curvas de diversidade decada área.
RESULTADOS E DISCUSSÃOAo compararmos o número de espécies e a
densidade total das duas áreas (Tab. 1), verificamosque a área queimada apresenta tanto um maiornúmero de espécies (32 spp. contra 25 spp. da área 8 Prof. Orientador: Fábio R. Scarana
protegida), quanto uma maior densidade total(1733,22 contra 1597,49 da área protegida).
Ao contrário do que esperávamos, o númerode espécies e a densidade observados neste estudoforam maiores na área queimada. A alta densidade naárea queimada pode ter ocorrido devido à resistêncianatural ao fogo, ou a uma alta taxa de recrutamento,ou ainda ao crescimento de estruturas vegetativas(caules) que são estimuladas pelo fogo. Porém, nãosão suficientes para uma análise conclusiva doproblema.
Quando comparamos as densidades absolutae relativa das principais espécies de ambas as áreas(Tab. 2), notamos que apenas uma espécie,Eremanthus glomerulatus, ocorre somente na áreaqueimada. É possível que esta espécie seja favorecidapelo fogo devido a adaptação especiais aindadesconhecidas. Observamos também queDidymopanax macrocarpum parece ser indiferente aofogo, uma vez que sua densidade é alta tanto na áreaqueimada quanto na área protegida. Vale notar aindaque no gênero Qualea houve inversão entre as áreas,com Q. parviflora sendo mais abundante na áreaqueimada e Q. multiflora apresentando maiordensidade na área protegida.
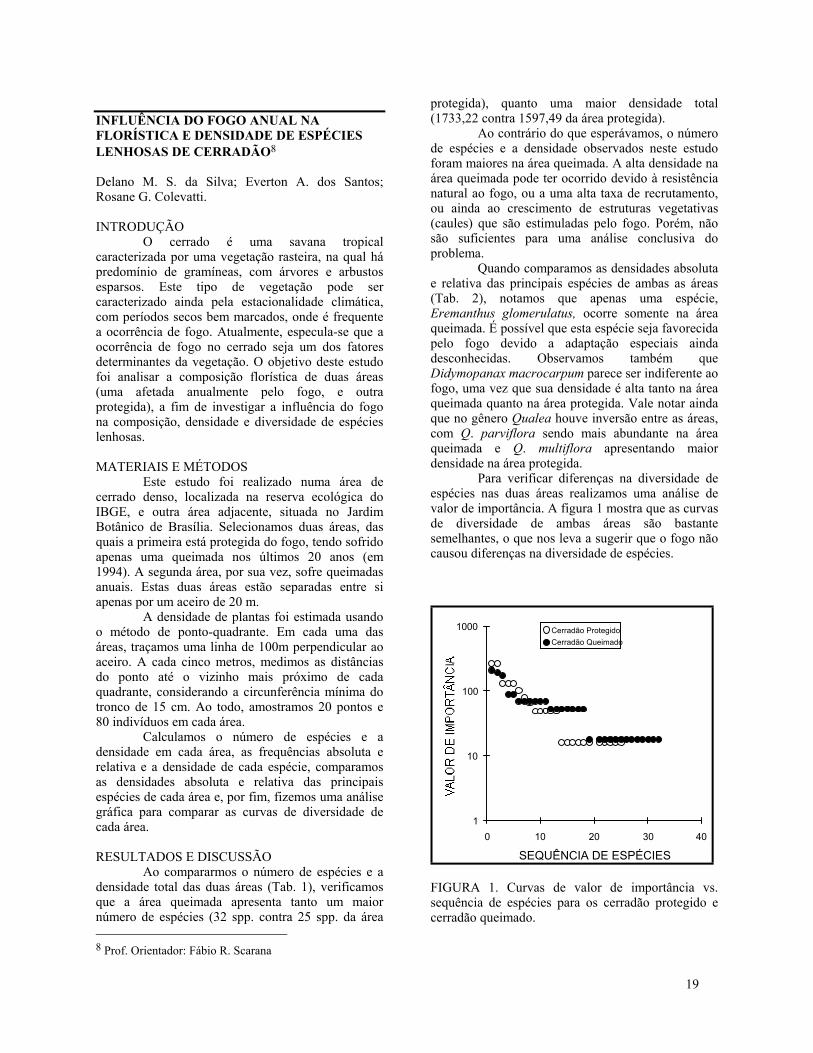
Para verificar diferenças na diversidade deespécies nas duas áreas realizamos uma análise devalor de importância. A figura 1 mostra que as curvasde diversidade de ambas áreas são bastantesemelhantes, o que nos leva a sugerir que o fogo nãocausou diferenças na diversidade de espécies.
SEQUÊNCIA DE ESPÉCIES
1
10
100
1000
0 10 20 30 40
Cerradão ProtegidoCerradão Queimado
FIGURA 1. Curvas de valor de importância vs.sequência de espécies para os cerradão protegido ecerradão queimado.

20
Tabela 1. Número de espécies e densidade de plantas nas áreas queimadas e não queimadas.área queimada área protegida
nº de espécies 32 25densidade (ind./ha) 1733 1597
Tabela 2. Densidade das principais espécies de plantas nas áreas queimadas e protegidas de cerradão.espécie área queimada área protegida
densidade % densidade %Eremanthus glomerulatus 210 12 - -Didymopanax macrocarpum 193 11 261 16Sclerolobium paniculatum 175 10 16 1Qualea multiflora 13 1 260 16
APÊNDICE 1. Densidade, frequência absoluta e relativa das espécies encontradas na área queimada.espécies frequência absoluta densidade relativa densidade absoluta
Erementhus glomerulatus 10 0,12 210,08Didymopanax macrocarpum 09 0,11 192,57Qualea parviflora 08 0,10 175,07Miconia ferruginata 04 0,05 87,53Roupala montana 04 0,05 87,53Blepharocalyx salicifolia 03 0,04 70,03Byrsonima crassa 03 0,04 70,03Guapira noxia 03 0,04 70,03Myrcinia guianensis 03 0,04 70,03Rourea induta 03 0,04 70,03Ouratea hexasperma 03 0,04 70,03Dalbergia violacea 02 0,03 52,52Erythroxylum suberosum 02 0,03 52,52Hymenaea stilbocarpa 02 0,03 52,52Lafoensia pacari 02 0,03 52,52Qualea multiflora 02 0,03 52,52Symplocus rhamnyfolia 02 0,03 52,52Vochysia thyrsoidea 02 0,03 52,52Byrsonima coccolobifolia 01 0,01 17,51Eriotheca pubescens 01 0,01 17,51Kielmeyera coriacea 01 0,01 17,51Lauracea 01 0,01 17,51Copaifera langsdorfii 01 0,01 17,51Caryocar brasiliensis 01 0,01 17,51Neea theifera 01 0,01 17,51Myrtaceae 01 0,01 17,51Qualea grandiflora 01 0,01 17,51Piptocarpha rotundifolia 01 0,01 17,51Sclerolobium paniculatum 01 0,01 17,51Stryphnodendron barbatimao 01 0,01 17,51Strychnos pseudoquina 01 0,01 17,51Palicourea rigida 01 0,01 17,51TOTAL - - 1733,22

21
APÊNDICE 2.Densidade, frequência absoluta e relativa das espécies de plantas encontradas na área protegida.espécies frequência absoluta densidade relativa densidade absolutaDidymopanax macrocarpum 13 0,16 260,15Sclerolobium paniculatum 13 0,16 260,15 Caryocar brasiliense 05 0,16 101,62Qualea multiflora 06 0,08 130,07Piptocarpha rotundifolia 06 0,08 130,07Vochysia thyrsoidea 03 0,04 65,04Stryphnodendron barbatimao 04 0,05 81,30Dimorphandra mollis 02 0,03 48,78Dalbergia violacea 02 0,03 48,78Guapira noxia 02 0,03 48,78Aspidosperma tomentosum 02 0,03 48,78Miconia ferruginata 02 0,03 48,78Blepharocalyx salicifolius 06 0,08 130,07Myrtacea 1 01 0,01 16,26Styrax ferruginea 01 0,01 16,26Palicourea rigida 01 0,01 16,26Connarus sp. 01 0,01 16,26Rapanea guianensis 01 0,01 16,26Rourea induta 01 0,01 16,26Davilla elliptica 01 0,01 16,26Qualea parviflora 01 0,01 16,26 Byrsonima verbascifolia 01 0,01 16,26B. crassa 01 0,01 16.26Eriotheca pubescens 01 0,01 16,26Pterodon pubescens 01 0,01 16,26TOTAL - - 1597,49

22
EFEITO DO FOGO NA ARQUITETURA DEESPÉCIES LENHOSAS DO CERRADO9
Flávia S. Pinto; Guilherme H. B. de Miranda; SauloM. A. Andrade.
INTRODUÇÃOO fogo ocorre comumente no cerrado,
afetando sua fauna e flora de diversas maneiras. Comrespeito à vegetação, os efeitos do fogo podem serclassificados em três níveis de intensidadedecrescente: (1) morte do indivíduo, comconsequente diminuição da densidade populacional ;(2) dano total, com a parte aérea do indivíduototalmente destruída e (3) dano parcial, com a parteaérea parcialmente destruída. A recuperação daplanta após o fogo se dá por rebrotamento que podeser: (1) subterrâneo, a partir de raízes ou outrasestruturas subterrâneas; (2) basal, da base do cauleremanescente ou (3) aéreo, das extremidades ouporção lateral do caule ou ramos remanescentes. Aarquitetura da planta que sofreu a ação do fogodepende do tipo de investimento para recuperaçãopós-queima (rebrota). Os dois primeiros tipos citadosde rebrota provocam alteração na arquitetura daplanta. A distribuição horizontal e vertical dabiomassa, por sua vez, é determinante dascaracterísticas do próximo fogo (tipo, intensidade etaxa de dispersão).
Nosso objetivo neste projeto foi comparar avegetação lenhosa de uma área sujeita a um regimeregular (bienal) de queimada com uma área protegidapor um período relativamente longo (> 20 anos). Ahipótese é que espécies arbóreas de áreas queimadaspossuem diferenças em relação à áreas nãoqueimadas, aumentando o número de brotos apósuma queimada (entouceiramento). Essas diferençassão temporárias, caso não haja recorrência do fogo.Como premissa, considerou-se que cada rebrotacorrespondia a um indivíduo.
MATERIAL E MÉTODOSA coleta de dados foi feita em duas áreas de
cerrado da Reserva Ecológica do IBGE : (1) áreaqueimada em 1992 e 1994, após um período de 16anos de proteção contra o fogo e (2) área protegidado fogo a mais de 20 anos. A primeira área localiza-se próximo à cerca de divisa com o Jardim Botânico,no limite leste da reserva e a outra área, atrás dorefeitório nas proximidades das construções da sededa reserva.
Em cada uma das áreas foram identificadose tiveram seus respectivos números de rebrotas vivase de rebrotas mortas contados todas as plantas 9 Profª. Orientadora: Heloisa S. Miranda
lenhosas que se encontravam a uma distânciaperpendicular não superior a um metro de uma linhade 50 metros estendida aleatoriamente e que tinhammais de cinco centímetros de diâmetro de caule.
RESULTADOS E DISCUSSÃONa área queimada bienalmente foram
identificadas 22 espécies lenhosas em 78 indivíduosamostradas. Ouratea hexasperma (N=11) foi aespécie mais frequente. Na área controle, 114indivíduos de 41 espécies foram amostrados. NaTabela 1 estão listadas as cinco espécies maisfrequentes em cada uma das áreas, correspondendo a55% (área queimada) e a 40% (área controle) dosindivíduos amostrados em cada área. Cinco dasespécies do cerrado queimado foram encontradosapenas uma vez. O mesmo se deu com 11 espécies daárea controle.
O índice de similaridade de Jaccard para asduas áreas foi de 31%, uma vez que apenas 15espécies eram comuns às duas áreas amostradas. Ovalor foi considerado baixo, não sendo possívelprecisar se o fogo seria o único responsável por isto,ou se seria parte do padrão de mosaico do cerrado, oualgum outro fator responsável não conhecido.
A figura 1 mostra a frequência de rebrotasvivas (20) em 78 indivíduos. A classe mais frequentefoi a de 1 rebrota (N=33). Apenas seis espécies nãotiveram representantes nesta classe, cinco dessas seisespécies ausentes na classe de uma rebrota foramjustamente as que foram encontradas uma única vezna área. Quarenta e um dos 78 indivíduos amostradosapresentaram mais de uma rebrota. O númeromáximo de rebrotas foi observado nas espécies comdistribuição por classes mais variadas, apresentandoalto grau de entouceiramento (figuras 3 e 4).
As rebrotas mortas indicando a existência dequeimadas anteriores à última também foramcontadas (130 rebrotas, variando de 1 a 6 porindivíduo) (Fig. 2). O valor mais frequente foi naclasse de uma rebrota. Na área controle foramcontados 114 indivíduos, somente dois apresentarammais de uma rebrota viva (uma Miconia fallax com 4rebrotas vivas e uma Vellozia flavicans com 3rebrotas vivas). Nenhuma rebrota morta foiencontrada.

23
Tabela 1. Espécies lenhosas mais frequêntes nas áreas amostradas.
cerrado queimado cerrado controleespécies n espécies n
Ouratea hexasperma 11 Rourea induta 18Kielmeyera coriacea 9 Rapanea guianensis 8Miconia fallax 6 Dalbergia violacea 7Davila elliptica 6 Qualea parviflora 6Byrsonima coccolobifolia 6 Roupala montana 6
NÚMERO DE REBROTOS
0
5
10
15
20
25
30
35
1 2 3 4 5 6 7 8 9 10 11 12
REBROTOS VIVOS
REBROTOS MORTOS
FIGURA 1. Número de plantas com rebrotos vivos e rebrotos mortos em um cerrado submetido a queimada bienaismodais.

24
FIGURA 2. Número de rebrotos em duas espécies de plantas em um cerrado sumetido a queimadas bienais modais.
Miconia fallax
NÚMERO DE REBROTOS
0
1
2
3
4
5
6
1 2 3 4 5 6 7 8 9 10 11 12
REBROTOS MORTOSREBROTOS VIVOS
Ouratea hexasperma
NÚMERO DE REBROTOS
0
1
2
3
4
5
6
1 2 3 4 5 6 7 8 9 10 11 12
REBROTOS VIVOSREBROTOS MORTOS
25
COMPARAÇÃO DE CARACTERÍSTICAS DEFOLHAS DE PLANTAS DE CERRADO EMATA DE GALERIA10
Carlos A. Bianchi; Felipe A. P. L Costa; TerezinhaA. B. Dias.
INTRODUÇÃOCaracterísticas foliares como tamanho,
dureza e coloração afetam a interação das plantascom fatores físicos e bióticos do ambiente. Porexemplo se a quantidade de energia luminosa variaentre dois habitats, nós deveríamos esperar que asplantas ajustassem suas características para manterum mesmo nível de eficiência. O objetivo dessetrabalho foi comparar características entre plantas deum trecho de cerrado com outro de mata ciliar. Maisespecificamente nós testamos se: 1) o tamanho, adureza e a coloração das folhas diferia entre esteshabitats, 2) de modo semelhante, a presença de pêlose látex.
MATERIAL E MÉTODOSEste trabalho foi realizado em dois tipos de
vegetação: uma área cerrado e uma área de mata degaleria, dentro da Reserva Ecológica do IBGE(Brasília, DF). Em cada área amostramos plantas de30 espécies (altura mínima de 1m), ao longo de umalinha de 50 metros. Para cada planta foramregistradas as seguintes características foliares:comprimento e largura máxima; área foliar,coloração; presença ou não de pêlos e látex. A durezarelativa de cada folha foi estimada como uso de um"penetrômetro" (3 folhas/planta e 3 medições/folha).
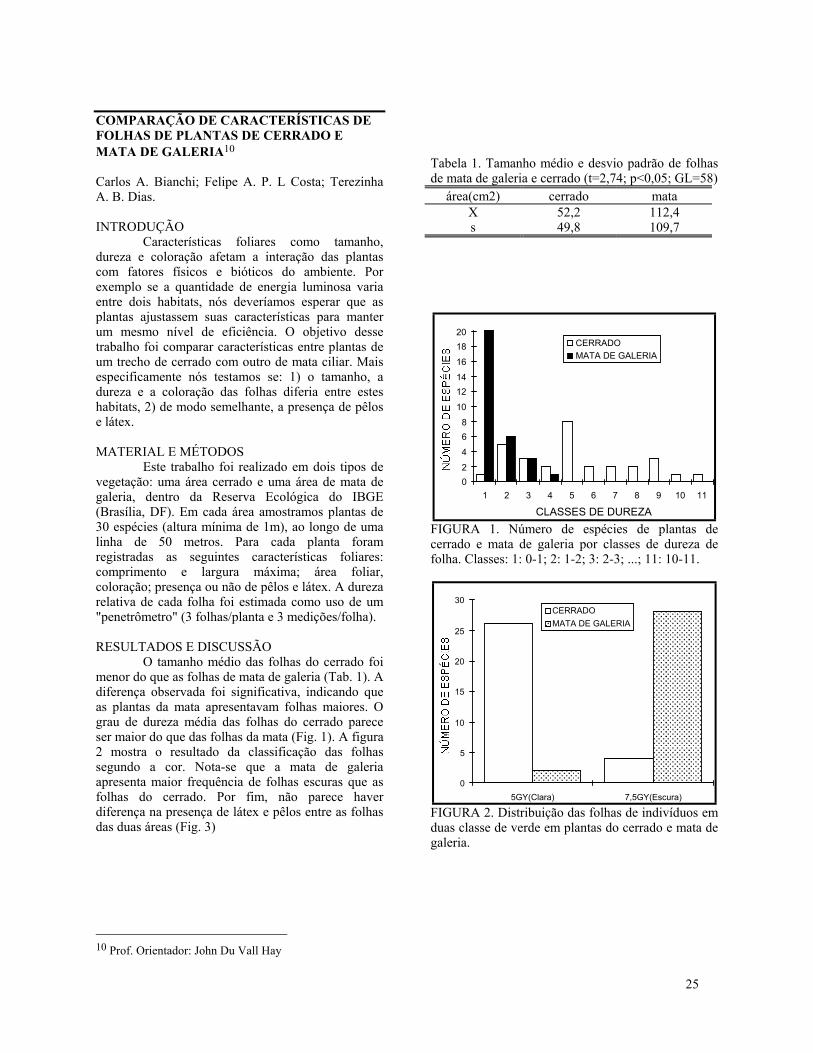
RESULTADOS E DISCUSSÃOO tamanho médio das folhas do cerrado foi
menor do que as folhas de mata de galeria (Tab. 1). Adiferença observada foi significativa, indicando queas plantas da mata apresentavam folhas maiores. Ograu de dureza média das folhas do cerrado pareceser maior do que das folhas da mata (Fig. 1). A figura2 mostra o resultado da classificação das folhassegundo a cor. Nota-se que a mata de galeriaapresenta maior frequência de folhas escuras que asfolhas do cerrado. Por fim, não parece haverdiferença na presença de látex e pêlos entre as folhasdas duas áreas (Fig. 3)
10 Prof. Orientador: John Du Vall Hay
Tabela 1. Tamanho médio e desvio padrão de folhasde mata de galeria e cerrado (t=2,74; p<0,05; GL=58)
área(cm2) cerrado mataX 52,2 112,4s 49,8 109,7
CLASSES DE DUREZA
02468
101214161820
1 2 3 4 5 6 7 8 9 10 11
CERRADOMATA DE GALERIA
FIGURA 1. Número de espécies de plantas decerrado e mata de galeria por classes de dureza defolha. Classes: 1: 0-1; 2: 1-2; 3: 2-3; ...; 11: 10-11.
����������������������������������������
������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������
0
5
10
15
20
25
30
5GY(Clara) 7,5GY(Escura)
CERRADO������MATA DE GALERIA
FIGURA 2. Distribuição das folhas de indivíduos emduas classe de verde em plantas do cerrado e mata degaleria.

26
�����������������������������������������������������������������������������
������������������������������������������������������������������������������������
�������������������������������������������������������������������������������������������������������������������������
������������������������������������������������������������������������������������������������������������������������������������
0
5
10
15
20
25
30
CERRADO MATA DEGALERIA
���COM PÊLOSEM PÊLOCOM LÁTEX���SEM LÁTEX
FIGURA 3. Número de indivíduos em relação apresença de pilosidade e presença de látex nas folhasde plantas do cerrado e mata de galeria.

27
COMPARAÇÃO DO POTENCIAL DE ÁGUADE ESPÉCIES LENHOSAS DE DIFERENTESGRUPOS FUNCIONAIS11
Delano M. S. da Silva; Everton A. dos Santos;Rosane G. Colevatti.
INTRODUÇÃOA água para as plantas além das funções já
conhecidas para outros organismos tem umimportante papel no transporte de nutrientes do solo(raízes) para as partes aéreas (folhas). Este transporteocorre devido a um gradiente do potencial de água(v) das raízes para as folhas sendo que o fluxo vai dopotencial menos negativo para o mais negativo. Issoé de grande importância para as plantas pois elaspodem controlar a direção deste gradiente da seguinteforma: os estômatos presentes nas folhas abrem-secriando um potencial negativo que deslocará o fluxode água das raízes para as folhas, pois o v das raízes émenos negativo. Quando esta água desloca-se levaconsigo os nutrientes armazenados na raiz.
Vários fatores estão influenciando estepotencial de água como podemos ver na fórmuladescrita abaixo:
v= p + m,onde v=potencial osmótico; m=potencial de matriz ep=potencial de pressão. O potencial osmótico estárelacionado à quantidade de soluto e o aumento destediminui o potencial de água. O potencial de matrizestá relacionado à permeabilidade das paredes e oaumento deste potencial diminui o potencial de água.O potencial de pressão está relacionado à pressão dosvasos e este aumenta o potencial de água. O objetivodeste trabalho é comparar o potencial de água dasfolhas de espécies lenhosas de diferentes gruposfuncionais do cerrado.
MATERIAL E MÉTODOSEste trabalho foi realizado numa área de
cerrado localizada na Reserva Ecológica doIBGE/DF, em dois horários distintos: 7:00 e 12:00. Oprimeiro funciona como nosso "ponto zero", poisimagina-se que nesse horário tanto folhas comoraízes estão no seu grau máximo de saturação. Ooutro período mostraria a variação do v.
As espécies utilizadas e o grupo funcionalde cada uma estão descritas na tabela 1. Para cadaespécie foram utilizados 3 indivíduos e de cada umforam retiradas 3 folhas. Estas folhas foramcolocadas num isopor com gelo para manutenção dov.
Para medirmos o v utilizamos uma câmarade pressão. O método consiste em colocar uma folha 11 Profª. Orientadora: Mercedes Bustamante
na câmara com o seu pecíolo de fora. Lacra-se estacâmara e introduz-se um gás inerte (N2), o qualaumenta a pressão da câmara. Quando sai uma gotade água do pecíolo fecha-se o sistema e considera-seestabelecido o equilíbrio entre as células da folha e aseiva do xilema, admitindo-se que a pressão do gáscontrabalance exatamente o potencial de água dascélulas da folha. A unidade de pressão utilizado foi obar.
RESULTADOS E DISCUSSÃOOs resultados obtidos na câmara de pressão
estão reunidos na tabela 2. Os resultadosapresentados na tabela 1 mostram que obtivemosdiferenças apenas nas espécies que são justamente asespécies coletadas antes de uma chuva.

28
Tabela 1. Lista das espécies lenhosas estudadas e seus grupos funcionais (al= acumuladora de alumínio, n-al= nãoacumuladora de alumínio).grupo funcional espécieperene
al Vochysia elliptica (Vochysiaceae)n-al Roupala montana (Proteaceae)n-al Ourateae hexasperma (Ochnaceae)n-al Styrax ferrugineus (Styracaceae)n-al Sclerolobium paniculatum (Leguminosae)
decíduaal Qualea grandiflora (Vochysiaceae)al Qualea parviflora (Vochysiaceae)
n-al Dalbergia violacea (Leguminosae)n-al Pterodon pubescens (Leguminosae)
Tabela 2. Potencial de água médio (bar) para as espécies estudadas.ESPÉCIES 7:00 14:00 ∆Vochysia elliptica* -1,7 (0,3) -5,0 (2,7) 3,3Roupala montana* -3,0 (0,5) -14,0 (8,4) 11,0Ourateae hexasperma* -2,2 (0,6) -7,0 (4,8) 4,8Styrax ferrugineus -2,0 (0,0) -2,3 (1,1) 0,3Sclerolobium paniculatum -2,7 (0,3) -2,7 (0,6) 0,0Qualea grandiflora -1,3 (0,6) -2,0 (0,0) 0,7Qualea parviflora* -1,5 (0,5) -5,0 (1,0) 3,5Dalbergia violacea -1,2 (0,3) -4,1 (0,6) 0,9Pterodon pubescens* -2,2 (0,6) -7,6 (5,8) 5,7
* Espécies medidas antes da chuva.

29
COMPARAÇÃO DE MEDIDAS DETRANSPIRAÇÃO ENTRE PLANTASHEMIPARASITAS E SUAS HOSPEDEIRAS1
Flávia S. Pinto; Saulo M. A. Andrade; Guilherme H.B. de Miranda.
INTRODUÇÃOAs plantas parasitas apresentam
especializações radiculares (haustórios) quepermitem a sua fixação em outras plantas(hospedeiras), de onde retiram nutrientes e água. Ohaustório retira seiva (bruta ou elaborada) dohospedeiro para a planta parasita. Algumas parasitasentretanto não perderam a capacidade de fazerfotossíntese e são portanto chamadas dehemiparasitas.
A condução de seiva é possível devido àrelação entre o potencial hídrico dentro dos vasos dacondução e o potencial hídrico do ar, em plantas nãosubmetidas a estresse hídrico. A diferença depotencial hídrico entre esses dois sistemas possibilitaa condução da seiva dentro dos vasos da planta. Essadiferença é obtida principalmente através da perda devapores d'água das folhas realizadas através dosestômatos. A perda d'água por transpiração produzum potencial hídrico negativo que permitiria acondução da seiva da raiz para os sítios defotossíntese. Entretanto vários fatores afetam aabertura estomática e consequentemente a taxa detranspiração, com a concentração de CO2, estressehídrico e patógeno.
Como se comportariam plantashemiparasitas neste sistema? Espera-se que plantaspara obterem seiva dos vasos condutores dahospedeira deveriam criar um potencial hídriconegativo maior do que o criado pela hospedeira,através, principalmente, do aumento da transpiraçãofoliar. Espera-se, portanto, um aumento nacondutância das espécies hemiparasitas emcomparação com suas hospedeiras.
O objetivo deste trabalho é determinarcondutâncias estomáticas: (1) se plantashemiparasitas apresentam condutância maior do quesuas hospedeiras e (2) se plantas parasitas afetam ocomportamento fisiológico das hospedeiras.
MATERIAL E MÉTODOSForam estudados dois indivíduos de
Kielmeyera coriacea e dois de Qualea multifora come sem plantas parasitas em um cerrado da Reserva doIBGE (Brasília D.F). A planta parasita encontrada em
1 Prof. Orientador: Fábio Scarano
Q. multifora foi Psittacanthus robustus e em K.coriacea. foi Phthirusa ovata.
Medidas de condutânica (c), densidade defluxo de fóton fotossintéticos (PPFD) e temperatura(T) foram feitas em cinco folhas, previamentemarcadas, para os seis indivíduos, com o auxílio deum porômetro. As medidas foram tomadas de horaem hora, no intervalo da 8:00 hs às 12:30 hs. Ao todocinco repetições foram feitas para cada plantaobservada. Nas plantas parasitas as medidas foramtomadas na porção abaxial e adaxial da folha, e paraas espécies hospedeiras apenas na porção abaxial,devido à distribuição de estômatos.
RESULTADOS E DISCUSSÃOTanto Qualea multiflora quanto Kielmeyera
coriacea não parasitadas apresentaram valores decondutância menores que plantas parasitadas damesma espécie (Fig. 1 e 2, respectivamente). Taisresultados sugerem que plantas parasitadasapresentam resposta fisiológica à presença deparasitas.
As espécies parasitas também apresentaramrespostas às variações de aumento da condutânciacom a variação da PPFD. Isto é mais marcante emPhthirusa ovata (Fig. 4 e 5). As espécies parasitasapresentaram taxas de transpiração maiores que asespécies hospedeiras parasitadas ou não (Fig. 1 e 2)indicando que tais plantas utilizariam em parte adiferença de condutância na formação de umapressão hidrostática positiva que permitiria aobtenção de seiva da planta hospedeira. Entretantonos horários da manhã em que a PPFD era baixa omesmo padrão não foi observado, indicando, maisuma vez, que a condutância destas espécies parasitasé afetada pelo grau de insolação na planta.

30
FIGURA 1. PPFD e condutância estomática em Qualea parviflora parasitada e não parasitada por Psithacanthusrobustus.
FIGURA 2. PPFD e condutância estomática de Kielmeyera coriacea parasitada e não parasitada por Phitirusaovata.
TEMPO (hora)
0
500
1000
1500
2000
2500
8:00 9:00 10:00 11:00 12:00
Psittacanthus robustusQualea parviflora(parasitada)Qualea paarviflora (nãoparasitada)
TEMPO (hora)
0
100
200
300
400
500
600
700
800
8:00 9:00 10:00 11:00 12:00
Psittacanthus robustusQualea parviflora(parasitada)Qualea paarviflora (nãoparasitada)
TEMPO (hora)
0
200
400
600
800
1000
1200
1400
8:30 9:30 10:30 11:30 12:30
Phithirusa ovataKielmeyera coriaceae(parasitada)Kielmeyera coriaceae(não parasitada)
TEMPO (hora)
0
100
200
300
400
500
600
700
8:30 9:30 10:30 11:30 12:30
Phithirusa ovataKielmeyera coriaceae(parasitada)Kielmeyera coriaceae(não parasitada)

31
RELAÇÃO ENTRE CARACTERÍSTICASFOLIARES E HERBIVORIA EM ESPÉCIESLENHOSAS DE MATA E DE CERRADO13
Delano M. S. da Silva; Everton A. dos Santos;Rosane G. Colevatti.
INTRODUÇÃOAs características fenotípicas dos
organismos servem, geralmente, como indicadores dotipo de habitat que eles ocupam, bem como podemfornecer evidências do tipo de nicho ocupado poresses mesmos organismos. Nas plantas, o tipo defolha e outras características como cor, dureza,pilosidade, presença ou ausência de látex, entreoutros, são importantes para proteção contra aherbívoros.
Neste trabalho, procuramos investigar ascaracterísticas foliares presentes em espécieslenhosas de cerrado e de mata, com o objetivo decorrelacionar algumas características das folhas coma presença de herbivoria. Analisamos principalmentea relação entre a dureza foliar e a incidência deherbivoria.
MATERIAIS E MÉTODOSEste trabalho foi realizado na reserva
ecológica do IBGE, em Brasília, em duas áreasdistintas: uma de cerrado, outra de mata. Em cadauma das áreas, traçamos uma linha de 50 m decomprimento. Ao longo de cada linha, coletamos 30espécies de plantas lenhosas com altura média de 2m.De cada espécie, amostramos 25 folhas e verificamosa ocorrência ou não de sinais de herbivoriaproduzidos por galhadores, minadores, mastigadoresou sugadores e, em seguida, anotamos o número defolhas afetadas em cada espécie. Paralelamente,retiramos 3 folhas de cada espécie, para medidas dedureza foliar. As medidas foram realizadas com umperfurômetro.
RESULTADOS E DISCUSSÃOOs dados relacionados à dureza foliar das
espécies lenhosas de cerrado e de mata estãorepresentados na figura 1. Neste gráfico, podemosobservar que as folhas da maioria das espécies demata são moles (tenras), ao passo que as das espéciesde cerrado são duras. Estes resultados sugerem que asplantas de cerrado são geralmente esclerotizadas, afim de protegê-las contra perdas de água. Por outrolado, as espécies de mata apresentam folhas maisfinas, pois nesse tipo de ambiente a umidade é maiorque a do cerrado. Portanto, o risco de perder água ébem menor nas plantas da mata do que no cerrado. 13 Prof. Orientador: John Du Vall Hay
Na figura 2, podemos observar o grau dedureza foliar das espécies de cerrado e de mata,projetados contra a percentagem de folhas afetadaspor herbívoros. Neste gráfico, notamos que asespécies de mata estão concentradas principalmentenas menores classes de dureza, enquanto as decerrado estão distribuídas em todas as classes dedureza, com a maioria das espécies colocadas nosgrupos de maior dureza. Ainda na figura 2,constatamos que existem tanto espécies de cerradocom alta dureza e baixa herbivoria, quanto espéciesde mata com baixa dureza e baixíssima taxa deherbivoria. Guapira noxia possui um grau de durezarelativamente alto, e uma taxa de herbivoriaaltíssima. Além destes dois exemplos, Vochysiaelliptica, que é uma espécie encontrada no cerrado,apresenta um grau de dureza relativamente alto, euma taxa de herbivoria quase nula. O cerradoapresentou uma dureza ( x =4,86) maior do que amata de galeria ( x =0,82), estas diferenças foramsignificativas (t=7,01; p<0,001)
A partir dos resultados obtidos, podemosconcluir que: (1) as espécies de cerrado geralmenteapresentam um grau de dureza foliar maior que o dasplantas de mata; (2) Parece que não há relaçãosignificativa entre a dureza foliar e a taxa deherbivoria, nem nas espécies de cerrado, nem nas demata.

32
CLASSES DE DUREZA
02468
101214161820
1 2 3 4 5 6 7 8 9 10 11
CERRADOMATA DE GALERIA
FIGURA 1. Distribuição das folhas por classes de durezas em plantas de cerrado e mata de galeria.
DUREZA FOLIAR
0
10
20
30
40
50
60
70
80
90
100
0 5 10 15
CERRADOMATA DE GALERIA
FIGURA 2. Relação entre dureza foliar e a porcentagem de herbivoria em espécies lenhosas de cerrado e mata degaleria.

33
DISTRIBUIÇÃO DE PLANTAS PARASITASSOBRE PLANTAS HOSPEDEIRAS NARESERVA ECOLÓGICA DO IBGE (BRASÍLIA,D.F)14
Carlos A. Bianchi; Felipe A. P. L. Costa; TerezinhaA. B. Dias.
INTRODUÇÃOPlantas hemi-parasitas ocorrem nas plantas
do Cerrado. A infecção das plantas hospedeirasocorre quando uma semente da planta hemi-parasita énela depositada e se estabelece. Como a dispersão dePhthiruza ovata e Psittacanthus sp é ornitocórica, éde se supor que plantas hospedeiras que tambémtenham este tipo de dispersão, sejam mais infectadaspor estas hemi-parasitas devido à maior visitação porpássaros. Também quando estabelecido umparâmetro mínimo de altura das plantas hospedeiras,aquelas mais abundantes em determinada área seriammais infectadas. Além disso poderia se supor que aocorrência de plantas hemi-parasitas estaria maisconcentrada em plantas hospedeiras mais altas do quebaixas, uma vez que os pássaros evitariam o solo porpressão da predação. Com o objetivo de levantardados que corroborassem as hipóteses anteriormentedescritas realizou-se um levantamento das espéciesvegetais presentes e das plantas hemi-parasitasassociadas em uma área de cerrado da ReservaEcológica do IBGE (Brasília, D.F).
MATERIAL E MÉTODOSPercorremos ao longo de uma trilha, cerca
de 180m, amostrando plantas com altura superior a1,0m que estavam até 5 m de cada lado desta trilha.Estas plantas foram identificadas botânicamente nocampo e posteriormente classificadas pelo tipo dedispersão de frutos. A altura delas foi estimada,anotando-se a presença ou ausência e o número dehemi-parasitas em cada planta hospedeira. Para asanálises estatísticas, nós utilizamos o teste do qui-quadrado e análise de regressão pelo método dosmínimos quadrados.
RESULTADOS E DISCUSSÃOForam amostradas 407 plantas de 61
espécies. Comparou-se o número de plantas com esem parasitas de dispersão ornitocórica com plantasdispersas por outros agentes (Tab. 1) e verificamosque a diferença não foi significativa. Este resultadoindica que plantas de dispersão ornitocórica não estãonecessariamente mais infectadas por plantas hemi-parasitas do que plantas que possuem outro tipo dedispersão. 14 Prof. Orientador: Raimundo P. B. Henriques
Considerando-se todas as 56 espécies(parasitadas e não-parasitadas), existe uma relaçãosignificativa entre o número de plantas encontradas eo número com hemi-parasitas. (Fig. 1). Quando asplantas não infectadas são excluídas da análise, acorrelação é maior (r=0,57; p < 0,005; N=17). Onúmero de plantas parasitadas por classe de altura éapresentado na Tabela 2. Utilizando apenas as duasprimeiras classes de altura não houve diferença
significativa (2χ =0,99; p>0,05; GL=1).

34
NÚMERO DE PLANTAS
0
2
4
6
8
10
0 10 20 30 40
FIGURA 1. Número de plantas infectadas em relação ao número total de plantas de cada espécie. Cada ponto serefere a uma espécie, existem pontos superpostos. r=0,42; p=0,001; N=56.
NÚMERO DE PLANTAS
0
2
4
6
8
10
0 10 20 30 40
FIGURA 2. Número de plantas infectadas em função do número total de plantas de cada espécie. Existem pontossuperpostos. r=0,57; p=0,02; N=17.
Tabela 1. Número de plantas com e sem parasita e tipo de dispersor da semente.tipo de dispersão com parasitas sem parasitas totalpássaros 25 (22,9) 249 (251,1) 274não pássaros 9 (11,1) 124 (121,9) 133total 34 373 407
2χ =0,062; P > 0,05; GL=1
Tabela 2. Frequência de parasitas em plantas de diferentes classes de altura.altura (m) com parasitas sem parasitas total1,00 - 3,99 308 25 3334,00 - 6,99 56 7 63
> 7,00 11 0 11total 375 32 407

35
ESTRUTURA POPULACIONAL DEPseudobombax longifolium (BOMBACACEA).DISTRIBUIÇÃO ETÁRIA E TABELA DEVIDA15
Delano M. S. da Silva; Everton A. dos Santos;Rosane G. Colevatti.
INTRODUÇÃOO cerrado apresenta duas estações bem
definidas, que podem ser caracterizadas pelaprecipitação: uma estação chuvosa, que ocorre entreos meses de outubro a abril, e uma estação seca queocorre entre maio a setembro. Muitas das espéciesdesse ambiente desenvolveram um padrão decrescimento relacionado a esse padrão sazonal dechuvas e, portanto, de disponibilidade de água: umaestação de crescimento, na época chuvosa e umaestação onde há perda das folhas e o crescimento éinterrompido.
Algumas das espécies que apresentam estetipo de estratégia de crescimento apresentam umadiferenciação nos caules e ramos, com a presença denós ou cicatrizes correspondentes à queda das folhase parada no crescimento. Cada entre-nó tem a idadede um ano (do início da estação chuvosa ao fim).Dessa forma pela contagem do número de entre-nóspodemos calcular a idade de uma planta. A partirdesses dados é possível determinar a distribuiçãoetária da população, obter uma Tabela de VidaEstática e traçar curvas de sobrevivência. Esses dadosnos dão uma idéia da estrutura da populaçãoindicando, além do padrão de mortalidade nas faixasetárias, aspectos da dinâmica populacionalrelacionados ao recrutamento de indivíduos.
Pseudobombax longifolium (Bombacaceae)é uma das espécies do cerrado que apresenta o padrãode crescimento sazonal, com diferenciação dosmódulos de crescimento de cada estação decrescimento (estação chuvosa). Ocorre,preferencialmente, próximo às matas de galeria.Assim, os objetivos do nosso trabalho foramdeterminar a distribuição etária de uma população deP. longifolium, obter a Tabela de Vida Estática etraçar a curva de sobrevivência.
MATERIAL E MÉTODOSEm uma área de 10.400 m2 (1,04 ha) foram
registrados todos os indivíduos de P. longifolium,medindo-se a circunferência na base do caule,próximo ao solo. Nos indivíduos em que era possívelcontar o número de módulos de cada estação a idadeera determinada. Foi possível determinar através das
15 Prof. Orientador: Raimundo P. B. Henriques
marcas de carvão deixadas nas plantas depois daúltima queimada na área (1994), que cada módulocresce apenas uma vez por ano.
Com os dados de circunferência e idadedesses indivíduos foi feita uma Regressão LinearSimples, para estimar a idade daqueles indivíduoscuja contagem de módulos não foi possível,utilizando o modelo: y=a + bx onde "y"= idade emanos, e "x"= circunferência, em centímetros, na basedo caule. Com a estimativa da idade dos indivíduos,estes foram agrupados em classes de idade, paraobtenção da Tabela de Vida Estática da população.
RESULTADOS E DISCUSSÃOForam encontrados 119 indivíduos na área,
resultando em uma densidade de aproximadamente114 indivíduos/ha. A Figura 1 mostra a relação entrea circunferência na base do tronco e a idade dosindivíduos, além da reta de regressão obtida. A partirda equação de regressão (y=- 6,6985 + 0,91332x) foiobtida a idade de todos indivíduos da população,cujos módulos não foi possível contar. A maioria dos indivíduos estão concentradosnas classes etárias 0 - 19 anos. Esse resultado sugereque a população está se regenerando localmente,recrutando indivíduos jovens.
A Tabela 1 apresenta a Tabela de VidaEstática da população que mostra a curva desobrevivência mostrada na Figura 3, sugere umpadrão intermediário entre as curvas do Tipo II eTipo III. Nas curvas de sobrevivência do Tipo II, ataxa de mortalidade é constante nas classes etárias(padrão comum para mortalidade de sementes), já notipo III a taxa de mortalidade é alta nas faixas etáriasmais baixas, caindo rapidamente e se estabilizandopara as faixas etárias maiores.
CIRCUNFERÊNCIA (cm)
0
10
20
30
40
50
0 10 20 30 40 50
FIGURA 1. Relação entre a circunferência na basedo tronco e idade em indivíduos de Pseudobombax

36
longiflorum em uma área de cerrado. O ponto abertono gráfico não foi incluído nas análises. Equação de
regressão: Y=0,91X - 6,69; r=0,84; p<0,001; N=25.
CLASSES DE IDADE
0
10
20
30
40
50
1 2 3 4 5 6 7 8 9 10 11
FIGURA 2. Distribuição de idades de uma população de Pseudobombax longiflorum em uma área de cerrado.Classes de idade: 1: 0-9; 2: 9-19,4; 3: 19,4-29,7; ...; 11: 101,7-112,0 (veja Tabela 1).
IDADE (ANOS)
0,001
0,01
0,1
1
0 20 40 60 80 100 120
FIGURA 3. Curva de sobrevivência para uma população de Pseudobombax longiflorum em área de cerrado.
Tabela 1. Tabela de vida estática para uma população de Pseudobombax longiflorum em uma área de cerrado.idade (anos) meio da classe
(anos)indivíduos sobreviventes taxa de sobrevivência
0-9 4,5 44 119 1,009-19,4 14,2 48 75 0,63
19,4-29,7 24,5 15 27 0,2329,7-39,9 34,8 04 12 0,1039,9-50,3 45,1 03 08 0,0750,3-60,6 55,4 01 05 0,0460,6-70,9 65,7 01 04 0,0370,9-81,1 75,9 00 03 0,0381,1-91,4 86,2 01 03 0,03
91,4-101,7 96,6 01 02 0,02101,7-112,0 106,9 01 01 0,01

37
TAXA DE OCORRÊNCIA DE ESPÉCIESLENHOSAS COM NECTÁRIOSEXTRAFLORAIS EM CERRADO16
Flávia S. Pinto; Guilherme H. B. de Miranda; SauloM. A. Andrade.
INTRODUÇÃOOs nectários extraflorais (NEF) são
estruturas presentes em diversas espécies de plantas,sendo responsáveis pela liberação de dissacarídeosexcedentes que atraem insetos nectarívoros, emespecial formigas. Estes insetos podem desempenharum papel de proteção da planta hospedeira contraseus inimigos naturais, aumentando a aptidão dasplantas com NEF o que resulta em uma interaçãomutualística "frouxa".
Diversos estudos mostram a existência deuma alta correlação negativa da frequência de NEF eformigas (diversidade, dominância e taxa depredação) e a latitude ou altitude. Neste projeto,foram obtidos, as frequências de espécies e deindivíduos lenhosos com NEF em uma área decerrado da Reserva Ecológica do IBGE ecomparando com dados publicados sobre áreas decerrado vizinhas (Jardim Botânico e Fazenda ÁguaLimpa).
MATERIAL E MÉTODOSPara o levantamento das espécies lenhosas
com mais de um metro de altura, foi percorrido umtrecho de cerrado denso no sudeste da ReservaEcológica do IBGE. Neste trecho, foramidentificados, a nível de espécie, os 500 primeirosindivíduos lenhosos ao longo de uma linha. Foramincluídas plantas a uma distância máxima da linha.Exemplares de identificação duvidosa ou ignoradativeram amostras coletadas e foram posteriormenteidentificadas em laboratório. Nas listas de espécies,foram identificadas a espécies com NEF e contadosos números de indivíduos de cada espécie a fim de seobter a frequência.
RESULTADOS E DISCUSSÃOForam identificadas 66 espécies lenhosas
para 500 indivíduos encontrados no cerrado. Cercade 21,2% das espécies (14) e 36,4% dos indivíduos(182) possuem NEF (Tab. 1). Estes valoresassemelham-se aos valores calculados em outroslevantamentos florísticos do cerrado na APA doGama-Cabeça de Veado, em particular àqueles queapresentam maior número de indivíduos. Eventuaisdiferenças metodológicas de diferenteslevantamentos de campo foram desconsiderados. 16 Prof. Orientador: Paulo S. Oliveira
Na Tabela 2, estão listadas as 14 espéciescom NEF e a posição dessas estruturas nestasespécies. A maior parte dos NEF (108ind/5spp)ocorre nas folhas. Em menor escala, ocorrem emramos (35 ind/3spp), tricomas glandulares(13ind/25spp), estípulas (12ind/1sp), sépalas(8ind/1sp) e ráquis (6ind/2spp). Duas espécies comNEF nas folhas Rapanea guianensis e Banisteriopsissp respondem por mais de 50% dos indivíduos comNEF e por quase 20% do total de indivíduosamostrados (Tab. 2).
A porcentagem das espécies com NEF emfolhas e estruturas associadas alcança cerca de 70%(127). Este valor pode ser resultante do alto valoradaptativo decorrente da presença dos NEF maisperto ou na própria folha, aumentando a proteçãodesta contra herbívoros.
Algumas das espécies arbóreas maisabundantes do cerrado (Qualea parviflora, Qualeagrandiflora, Caryocar brasiliense) possuem NEF. NaTabela 3, uma síntese dos dados de frequência deespécies com NEF e de abundância dessas espéciespara outras regiões é mostrada. Estes resultadossomados aos obtidos no presente trabalhocorroboram a hipótese do gradiente latitudinalsugerindo que os NEF são mais importantes nostrópicos.
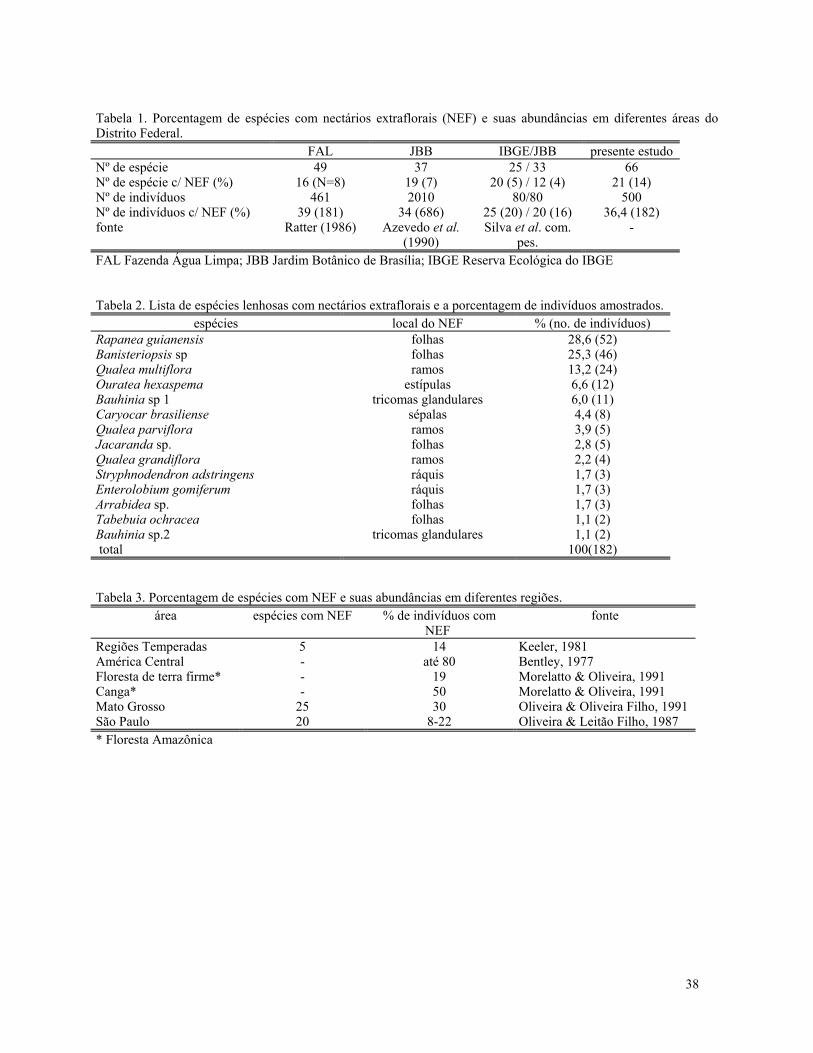
38
Tabela 1. Porcentagem de espécies com nectários extraflorais (NEF) e suas abundâncias em diferentes áreas doDistrito Federal.
FAL JBB IBGE/JBB presente estudoNº de espécie 49 37 25 / 33 66Nº de espécie c/ NEF (%) 16 (N=8) 19 (7) 20 (5) / 12 (4) 21 (14)Nº de indivíduos 461 2010 80/80 500Nº de indivíduos c/ NEF (%) 39 (181) 34 (686) 25 (20) / 20 (16) 36,4 (182)fonte Ratter (1986) Azevedo et al.
(1990)Silva et al. com.
pes.-
FAL Fazenda Água Limpa; JBB Jardim Botânico de Brasília; IBGE Reserva Ecológica do IBGE
Tabela 2. Lista de espécies lenhosas com nectários extraflorais e a porcentagem de indivíduos amostrados.espécies local do NEF % (no. de indivíduos)
Rapanea guianensis folhas 28,6 (52)Banisteriopsis sp folhas 25,3 (46)Qualea multiflora ramos 13,2 (24)Ouratea hexaspema estípulas 6,6 (12)Bauhinia sp 1 tricomas glandulares 6,0 (11)Caryocar brasiliense sépalas 4,4 (8)Qualea parviflora ramos 3,9 (5)Jacaranda sp. folhas 2,8 (5)Qualea grandiflora ramos 2,2 (4)Stryphnodendron adstringens ráquis 1,7 (3)Enterolobium gomiferum ráquis 1,7 (3)Arrabidea sp. folhas 1,7 (3)Tabebuia ochracea folhas 1,1 (2)Bauhinia sp.2 tricomas glandulares 1,1 (2) total 100(182)
Tabela 3. Porcentagem de espécies com NEF e suas abundâncias em diferentes regiões.área espécies com NEF % de indivíduos com
NEF fonte
Regiões Temperadas 5 14 Keeler, 1981América Central - até 80 Bentley, 1977Floresta de terra firme* - 19 Morelatto & Oliveira, 1991Canga* - 50 Morelatto & Oliveira, 1991Mato Grosso 25 30 Oliveira & Oliveira Filho, 1991São Paulo 20 8-22 Oliveira & Leitão Filho, 1987* Floresta Amazônica

39
A COEXISTÊNCIA DE ESPÉCIESCONGENÉRICAS E SEU NICHOMORFOLÓGICO17
Flávia S. Pinto; Guilherme H. B. de Miranda; SauloM. A. Andrade.
INTRODUÇÃOSegundo a "Teoria de Competição", espécies
com nichos semelhantes não podem coexistir umavez que pode ocorrer exclusão competitiva comextinção de uma das espécies. Espécies que sãoecologicamente parecidas, vivem juntas, logicamentehá uma diferenciação entre seus nichos. Assim pode-se assumir que entre espécies congenéricas acompetição é maior, já que são mais semelhantesentre si do que com espécies de outros gêneros. Comisso o objetivo do projeto foi verificar diferenças nasexistência do nicho morfógico de espéciescongenéricas de plantas em uma área de cerrado.
MATERIAL E MÉTODOSNuma simplificação do nicho morfológico
das espécies, já que o nicho não é bidimensional massim, multidimensional, tomou-se a relação área foliarX comprimento foliar, como uma representação donicho das espécies congenéricas de plantas. E assimcomparar as diferenças entre estes nichosmorfológicos. Para isso, foram escolhidos 10indivíduos de cada uma das espécies de Miconia,Qualea e Byrsonima (Tab. 1) e coletadas 5 folhas decada indivíduo. Mediu-se então a largura e ocomprimento de cada folha e, utilizou-se a forma daelipse para o cálculo da área foliar. Área de umaelipse (Ae)=πab/4, onde a=largura da folha eb=comprimento da folha.
RESULTADOS E DISCUSSÃOA figura 1 mostra a relação do nicho para as
3 espécies de Miconia. O porte das espécies de M.phalax e M. albicans é a mesma (Tab. 1), dessaforma esperaria-se que o nicho morfológico fossemais "distante", já que possuem o mesmo porte, istoé, para não possuírem o mesmo nicho as espéciesbuscariam mudanças na área de suas folhas para umadiferente aptidão de cada uma e consequentediferenciação de seus nichos.
A mesma relação pode ser apresentada parao gênero Qualea (Fig. 2), onde todas as espécies sãoarbóreas (Tab. 1) e esperaria-se uma diferenciação deseus nichos foliares (Fig. 2).
Para o gênero Byrsonima assumiu-se que oresultado encontrado exemplificou melhor a
17 Prof. Orientador: Raimundo P. B. Henriques
diferenciação dos nichos foliares, uma vez que suas 3espécies possuem o mesmo porte.
A separação do nicho foliar das espéciespode ser que esteja, não no nicho foliar mas sim, emoutros nichos morfológicos das plantas como,sistema radicular, arquitetura da copa, estratégia deutilização dos nutrientes, etc. Outra explicação é a deque constrangimentos filogenéticos, não permitemgrandes diferenças nos nichos foliares das espéciesestudadas.
Tabela 1. Espécies estudadas e suas formas de vida.espécie forma de vidaByrsonima coccolobifolia arbusto grossoByrsonima verbascifolia arbusto grossoByrsonima crassa arbusto grossoMiconia ferruginata arbusto grossoMiconia phalax arbusto finoMiconia albicans arbusto finoQualea parviflora arbóreaQualea grandiflora arbóreaQualea multiflora arbórea

40
COMPRIMENTO DA FOLHA (cn)
0
100
200
300
400
500
-3 2 7 12 17 22 27
Miconia albicansReta de regressãoMiconia fallaxReta de regressãoMiconiaferruginataReta de regressão
FIGURA 1. Relação entre área e comprimento de folhas de Miconia emárea de cerrado.
COMPRIMENTO DA FOLHA (cn)
0
100
200
300
0 5 10 15 20
Qualea parvifloraReta de regressãoQualea multifloraReta de regressãoQualea grandifloraReta de regressão
FIGURA 2. Relação entre área e comprimento de folhas de Qualea em área de cerrado.
COMPRIMENTO DA FOLHA (cn)
0
100
200
300
400
500
600
0 5 10 15 20 25
Byrsonima crassaReta de regressãoByrsonimacoccolobifoliaReta de regressãoByrsonimaverbascifoliaReta de regressão
FIGURA 3. Relação entre área e comprimento de folhas de três espécies de Byrsonima em área de cerrado.

41
SIMILARIDADE FLORÍSTICA ENTRECOMUNIDADES DE ESPÉCIES HERBÁCEASEM FITOFISIONOMIAS EQUIVALENTES18
Carlos A. Bianchi; Felipe A. P. L. Costa; TerezinhaA. B. Dias.
INTRODUÇÃONa vegetação dos Cerrados, muitas
formações campestres são de fisionomiassemelhantes, sugerindo um alto grau de similaridadena composição específica. O objetivo desse trabalhofoi descrever e comparar a composição em espéciesherbáceas de três fisionomias campestres numa áreade Cerrado.
MATERIAL E MÉTODOSO trabalho de campo foi desenvolvido em 3
áreas destro da Reserva Ecológica do IBGE (Brasília,DF), a saber: campo limpo (CL), brejo (BR) e"cascalheira"(CAS), (todas são de fisionomiascampestre, apresentando arbustos).
Para descrever a composição específica, nósamostramos aleatoriamente 2 parcelas (1 x 1m) emcada uma das 3 áreas. Ainda no campo, nósidentificamos e contamos o número de indivíduos decada espécie presente nas parcelas. Como índices desimilaridade entre 2 áreas, nós utilizamos osseguintes índices de similaridade: (a) Jaccard (Sj):Sj=a/a+b+c; (b) Sorensen (Ss): Ss=2a/2a+b+c; (c)Czekanowski (Sc): Sc=2 min (xi, i)/ (xi+i); onde, a=número de espécies comuns as 2 áreas; b e c =número de espécies exclusivas nas áreas 1 e 2,respectivamente; min (xi, i)=menor número deindivíduos para as espécies encontradas nas 2amostras; xi e i=número de indivíduos da espécie "i"encontrados nas 2 áreas.
Além disso, nós comparamos a diversidadeespecífica das áreas, utilizando o índice de Shannon-Wiener (H'):
H'= - ∑pi ln pi ;onde pi é a abundância relativa da espécies "i".
RESULTADOS E DISCUSSÃOConsiderando todas as 6 parcelas
amostradas, contamos 250 indivíduos, pertencentes a52 espécies de 17 famílias botânicas. O número deespécies por família é mostrado na Figura 1, onde éfácil notar a predominância das Graminae (ca. 31%do total). Essa predominância aumentatremendamente quando comparamos as abundânciasrelativas (Fig. 1). Esses resultados sustentam nossaimpressão original de que as Graminae constituem oprincipal componentes das formações campestres. 18 Prof. Orientador: Tarcisio S. Filgueiras
O número de indivíduos e espéciesencontradas variam entre as áreas, a saber: "CL"(153indivíduos de 30 espécies); "BR" (39 de 7) e"CAS"(58 de 16). Na verdade, a composiçãoespecífica de cada área foi virtualmente exclusiva,com apenas 1 espécie ocorrendo no "CL" e na"CAS".
Os valores obtidos para os índices desimilaridade entre "CL" e "CAS" são mostrados naTabela 1. O fato da única espécie em comum ter sido,ao mesmo tempo, uma das mais abundantes, explicao aumento obtido para Sc, quando comparado aosoutros índices. Deve-se notar que Sc mede asimilaridade entre amostras, levando em conta asabundâncias relativas das espécies em comum.
Os índices de diversidade para as 3 áreasestão mostrados na Tabela 2. A diferença nos valoresobtidos para "CL" e "CAS" parece resultar mais donúmero de espécies do que da equitabilidade. Poroutro lado, o baixo valor obtido par "BR" se devetanto à sua pobreza específica como também à baixaequitabilidade (mais de 50% dos indivíduospertenciam a uma única espécie).
Por fim, a Tabela 3 apresenta o número deespécies "invasoras" encontradas. Curiosamente,essas plantas só foram encontradas na "CAS", ondechegam a compor mais de 60% do total de espécies.
Concluindo, nossos resultados indicam queas Graminae dominam a paisagem das formaçõescampestres no Cerrado estudado, tanto pela riquezacomo pela abundância das espécies. Por sua vez, ariqueza específica parece ser o resultado de que asespécies se substituem ao longo de certos habitatsencontrados nos Cerrados.

42
������������������������������������������������������������������������
��������������
���������������� �������
���������������������
0
5
10
15
20
25
30
35
GRAMÍNEA LEGUMINOSAE COMPOSITAE MELASTOMATACEA OUTRAS 13 FAMÍLIAS
NÚMERO DE ESPÉCIES��
NÚMERO DE INDIVÍDUOS
FIGURA 1. Número de espécies e indivíduos por famílias.
Tabela 1. Similaridade florística entre campo limpo e cascalheira.índice de similaridade (%)
Jaccard Sorensen Czekanowski2,2 4,3 8,5
Tabela 2. Diversidade e equitabilidade em três tipos de vegetação no cerrado.tipo de vegetação
campo limpo brejo cascalheiradiversidade 2,754 1,409 2,366equitabilidade 0,810 0,724 0,874
Tabela 3. Número de espécies nativas, invasoras e espécie dominante em relação ao total de espécies na área dacascalheira.
número de espécies (%) espécie dominante % do totalnativas 6 (37,5) Mesosetum lolliforme 17invasoras 10 (62,5) Vernonia ferruginata 7total 16 (100,0)

43
PROJETOS
INDIVIDUAIS

44
PREDAÇÃO DE NINHOS ARTIFICIAIS NOCERRADO: UMA COMPARAÇÃO ENTREDUAS FISIONOMIAS.
Carlos Abs Bianchi
RESUMOA predação de ninhos em aves representa
um dos principais fatores de mortalidade para aclasse. Estudos com ninhos artificiais têm sidoutilizados para investigar padrões de predação emdiversos ambientes. Este trabalho investigou apredação de ninhos artificiais em duas fisionomias deCerrado (sensu lato): mata de galeria e cerradão, naReserva Ecológica do Roncador (IBGE), Brasília,DF; comparando taxas de predação, freqüência deretirada de ovos e efeitos de borda. Os resultadosobtidos mostraram não haver diferenças nas taxas depredação entre as duas fisionomias, entretanto, opadrão de retirada entre mata e cerradão foi diferente,na mata de galeria a taxa de predação atingiu omáximo no segundo dia enquanto que no cerradão ataxa de predação se mantave constante ao longo doperíodo de estudo. Os ovos colocados na borda damata foram estatisticamente mais predados do queaqueles colocados no interior; porém, não houvediferença entre borda e interior para o cerradão;sugerindo que o efeito de borda é mais intenso namata de galeria.
INTRODUÇÃOA predação representa uma importante
variável na determinação de diversas característicasdas comunidades; diversos estudos já foramrealizados em várias partes do mundo (Sih et al.1985), enfocando a interação predador-presa. Apredação pode ser definida como qualquer interaçãona qual ocorre fluxo de energia de um organismopara outro ou, de uma forma mais simples, como oconsumo de um indivíduo (a presa) por outro (opredador) (Begon 1990).
Predação de ninhos é a causa principal demortalidade em diversas espécies de aves (Cintra1988; Martin 1987; Angelstam 1986; Gates e Gysel1978) e certamente apresenta padrões variáveis deintensidade entre espécies e entre áreas. Aintensidade da predação sobre os ninhos pode serinfluenciada por vários fatores, tais como, o sítio denidificação (Martin 1987), a fragmentação deambientes (Santos e Tellería 1992, Angelstam 1986),ou ainda a densidade e atividade de predadores(Gates e Gysel 1978; Ruxton e Gurney 1994).
A realização de experimentos utilizandoninhos artificiais tem sido uma ferramenta bastanteimportante para testar algumas predições em
ecologia; entretanto, segundo Martin (1987), osninhos artificiais são mais conspícuos em relação aosnaturais, porém menos predados, devido à imagem deprocura desenvolvida pelos predadores associada aosninhos naturais. Tal aspecto representa uma restriçãopara comparações de taxas de predação obtidas poreste tipo de experimento com taxas de predação deninhos naturais.
Sih et al. (1985), em revisão sobre o tema,afirmam não haver diferenças marcantes nos padrõesgerais de predação quando considerada a variaçãolatitudinal, entretanto a quantidade de estudoscomparativos ainda é pequena. Os estudoscomparativos de taxas de predação entre áreas maisabertas e áreas de cobertura vegetal mais densarevelam haver diferenças entre as duas, sendo maisalta em áreas de formações florestais.
Os objetivos deste trabalho foram: (1)comparar as taxas de predação entre duas formaçõesflorestais do bioma Cerrado: mata de galeria ecerradão (Ratter, 1991); (2) comparar a freqüência deretirada e os padrões de predação para as duasfisionomias (3) comparar as taxas de predação entredois subgrupos de ninhos: ninhos de borda e ninhosde interior, em ambas as áreas, visando testar o efeitode borda sobre a predação.
MATERIAL E MÉTODOSO presente trabalho foi realizado na Reserva
Ecológica do Roncador - IBGE (15˚ 55’ 58” S e 47˚51’ 02” W), Brasília-DF; no período de 12 a 15 demarço de 1996, durante o curso “Métodos emEcologia de Campo” do Programa de Pós-graduaçãoem Ecologia da Universidade de Brasília.
Foram selecionadas para o estudo uma áreade Mata de Galeria e uma área de Cerradão (Ratter1991), distantes aproximadamente 2 quilômetrosentre si, onde foi estabelecido um gradeado para cadaárea. Cada gradeado era formado por cinco linhascompostas por sete pontos, e uma linha composta porcinco pontos, totalizando 40 pontos por gradeado. Adistância entre as linhas e os pontos foi estabelecidaem 10 metros, a mesma fixada entre os pontos decada linha (Figura 1).
O gradeado foi dividido em dois subgrupos:subgrupo “borda” e subgrupo “interior”. A primeiralinha de cada área foi estabelecida paralelamente aestrada, a 5 metros de distância desta, no sentidoborda-interior e, as três primeiras linhas (A, B, C)representaram o subgrupo “borda”, enquanto aslinhas subsequentes (D, E, F) representaram osubgrupo “interior”.
Os ninhos artificiais foram confeccionadosmanualmente com palha seca e, para cada ponto, foidepositado no chão, um ninho com dois ovos decodorna (Coturnix coturnix). Tendo em vista o

45
objetivo de apenas efetuar-se registros de predação,sem testar a capacidade dos predadores em localizaros ninhos, estes foram colocados nos pontos dosgradeados sem qualquer camuflagem. Os ninhosforam considerados predados quando o ninho e/ouum ou ambos os ovos foram removidos. As revisõesaos ninhos eram feitas no turno da manhã, numintervalo de 24 horas para as duas fisionomias.Durante a revisão, eram registrados a ocorrência ounão de ataque ao ninho, as coordenadas dentro dogradeado dos ninhos atacados, o número de ovospredados e a presença ou ausência de cascas e/ou adistância destas (quando encontradas) dos ninhospredados. Os ninhos atacados não tinham seus ovosrepostos, reduzindo desta forma, ao longo do tempo,a disponibilidade de ovos nos sítios de estudo.
Para as análises dos resultados, foramaplicados teste de qui-quadrado (χ2) para testar asignificância das taxas de predação de ovos em cadafisionomia (mata e cerradão) e as taxas de predaçãoentre borda e interior, além das representaçõesgráficas dos valores de predação do experimento.
RESULTADOSForam predados 66 (41,25%) dos 160 ovos
utilizados no experimento, sendo 37 (23,12% dototal) na mata e 29 (18,13% do total) no cerradão. AFigura 1 mostra os gradeados estabelecidos dentro decada fisionomia e o dia de predação dos ninhos.
As taxas de predação para a mata e ocerradão corresponderam a 46,25% e 36,25%,respectivamente, em relação ao total de ovos paracada fisionomia. O número de ovos predados emcada área é apresentado na Tabela 1, onde pode-severificar que não houve diferença significativa entreáreas (χ2= 1,65; P= 0,1989; g.l.=1; n= 80).
A freqüência de retirada de ovos para a matafoi bastante diferente daquela obtida para o cerradão.A Figura 2 mostra que a taxa de predação foiaproximadamente a mesma ao longo do estudo nocerradão, contrastando com os valores obtidos para amata, onde verifica-se um pico no segundo dia doexperimento, diminuindo depois.
A predação em ambas as áreas, quanto aoefeito de borda, apresentou resultados diferentes(Tabela 2). Para a mata, houve diferença significativa(χ2= 9,95; P= 0,0016; g.l.= 1; n= 40) entre os ninhospredados na borda (n= 14) e os ninhos do interior (n=5). No cerradão, o ataque aos ninhos da borda (n= 8)foi semelhante ao ataque dos ninhos do interior (n=6) (χ2= 0,80; P= 0,3702; g.l.= 1; n= 40).
Apenas um ninho na mata e três ninhos nocerradão tiverem seus ovos predados em diferentesdias do experimento. Os demais ninhos tiveram osdois ovos predados num mesmo dia.
DISCUSSÃOOs resultados obtidos neste experimento
mostram não haver diferença entre as taxas depredação na mata de galeria e no cerradão. Santos eTellería (1992), encontraram valores diferentes parapredação entre um ambiente aberto (área de plantiode fazenda) e um ambiente florestal. Entretanto,Andrén et al. (1985) e Angelstam (1986) nãoencontraram diferenças significativas entre ambientesabertos contra ambientes fechados na Escandinávia.Sugere-se que diferenças na densidade vegetalestariam associadas a diferenças nas taxas depredação, isto é, ambientes tipicamente florestaissofreriam uma pressão de predação maior em relaçãoaos ambientes mais abertos, devido, entre outrosfatores, à maior diversidade. Embora neste estudo ataxa de predação na mata de galeria tenha sido maiorque no cerradão a diferença não foi significativa. Astaxas de predação poderiam variar em um gradientede acordo com o aumento na densidade e diversidadeda vegetação de uma determinada área. A mata degaleria representa um típico ambiente florestal, comvegetação bastante densa. Dentre as fisionomias dobioma cerrado (sensu lato), o cerradão é aquela comcaracterísticas mais próximas de uma formaçãoflorestal, podendo inclusive apresentar espécies deplantas típicas de mata (Ratter 1991).
A freqüência de retirada dos ovos para cadadia foi diferente entre as duas fisionomias (Figura 2),tendo o cerradão apresentado uma taxa constante depredação, quando comparado com aquele registradopara a mata. A disposição dos ninhos nesteexperimento (gradeados com ninhos distantes 10metros entre si) foi diferente de outros estudos queutilizaram ninhos artificiais. Santos e Tellería (1992)utilizaram transectos com 30 metros de distânciamínima entre ninhos; Andrén et al. (1985) colocaramtransectos com distância mínima de 300 metros entreninhos e Angelstam (1986), utilizou transectos comdistância mínima de 500 metros. A disposição dosninhos em transectos pode diminuir as chances deataque por um mesmo predador, supondo-se que estetenha uma área de vida radial. Ninhos dispostos emgradeados, estão mais sujeitos à predação se a maiorparte do gradeado estiver dentro da área de vida deum predador potencial. Logo, distâncias maioresentre os ninhos e a disposição destes em transectos,diminuiriam relativamente as chances de ataque porum mesmo indivíduo. O pico de predação registradopara a mata no segundo dia (Figura 2) pode ou nãorepresentar a ação de um único predador, visto que osninhos atacados neste dia estão próximos entre si(Figura 1); entretanto, não existem evidênciasmaiores que comprovem este fato.
As taxas de predação (Tabela 2)correspondentes aos subgrupos “borda” e “interior”foram significativamente diferentes apenas na mata,

46
enquanto no cerradão, o ataque aos ninhos doprimeiro subgrupo não diferiram do segundo. Oresultado obtido para a mata não difere dos dadosexistentes na literatura. Santos e Tellería (1992)encontraram maior incidência de predação em ninhoscolocados na borda, assim como Angelstam (1986) eAndrén et al. (1985). Este diferente resultado obtidoentre mata e cerradão sugere que o efeito de bordapode ser mais intenso na mata do que no cerradão, epode estar relacionado a uma variação abrupta dafisionomia da mata de galeria para o cerrado, o quenão ocorre no cerradão onde a variação é maisgradual. Entretanto, outras variáveis devem serinvestigadas para melhor interpretação destesresultados.
AGRADECIMENTOSGostaria de agradecer a H. C. Morais, J. D.
Hay e R. Colevatti pelas sugestões nas análisesestatísticas do manuscrito. A D. M. Silva pelo auxílionas análises estatísticas e pela leitura e sugestões aomanuscrito. A Reserva Ecológica do Roncador(IBGE) por fornecer apoio logístico ao curso“Métodos em Ecologia de Campo”. A R. P. B.Henriques por organizar o curso, pelas sugestões aotrabalho e auxílio em algumas etapas do trabalho decampo. A C. A. Moreira pela assistência naelaboração do manuscrito. E aos colegas queparticiparam do curso de campo, pelas inúmerashoras de trabalho e descontração em conjunto.
BIBLIOGRAFIAANDRÉN, H.; ANGELSTAN, P.; LINDSTRÖM, E.
e WIDÉN, P. 1985. Differences in predationpressure in relation to habitat fragmentation: anexperiment. Oikos 45(2): 273-277.
ANGELSTAM, P. 1986. Predation on ground-nesting birds' nests in relation to predatordensities and habitat edge. Oikos 47(3): 365-373.
BEGON, M.; TOWSEND, J. L. e HARPER, C. R.1990. Ecology. Individuals, Populations andCommunities. 2nd edition, Blackwell ScienceInc. New York, USA.
CINTRA, R. 1988. Reproductive Ecology of theRuddy Ground-Dove on the Central Plateau ofBrazil. Wilson Bulletin 100(3): 443-457.
GATES, J. E. e GYSEL, L. W. 1978. Avian nestdispersion and fledging success in field-forest.Ecology 59(5): 871-883.
MARTIN, T. E. 1987. Artificial nest experiments:effects of the nest appearance and type ofpredator. Condor 89: 925-928.
RATTER, J. A. 1991. Guia para a vegetação daFazenda Água Limpa. (Brasília, DF). Editora daUniversidade de Brasília, Brasília, DF.
RUXTON, G.D. e GURNEY, W.S.C. 1994. Derivingthe functional response without assuming
homogeneity. American Naturalist 144(3): 537-541.
SANTOS, T. e TELLERÍA, J. L. 1992. Edge effectson nest predation in Mediterranean fragmentedforests. Biological Conservation 60: 1-5.
SIH, A.; CROWLEY, P.; MCPEEK, M.;PETRANKA, J. e STROHMEIER, K. 1985.Predation, competition and prey communities: areview of field experiments. Ann. Rev. Ecol.Syst. 16: 269-311.

47
CERRADÃO 7
6
5
4
3
2
1A B C D E F
MATA 7
6
5
4
3
2
1A B C D E F
LEGENDA PREDAÇÃO NO 1° DIA PREDAÇÃO NO 2° DIA PREDAÇÃO NO 3° DIA PREDAÇÃO NO 4° DIA NINHOS NÃO-PREDADOS
Figura 1. Mapa das áreas de estudo com a representação dos gradeados e data de predação dos ninhos depositadospara o estudo realizado na Reserva Ecológica do Roncador (IBGE), Brasília, DF (letras= colunas; números= linhas).
Tabela 1. Comparação do número total de ovos predados e não-predados entre a mata de galeria e o cerradão naReserva Ecológica do Roncador (IBGE), Brasília-DF.Categoria Mata de Galeria Cerradão TotalPredados 37 29 66Não-predados 43 51 94Total 80 80 160(χ2= 1.65; P= 0.1989; n= 80)
Tabela 2. Comparação de predação dos ninhos nos subgrupos “borda” e “interior” nas duas áreas de estudo naReserva Ecológica do Roncador (IBGE), Brasília, DF.Categoria Mata de Galeria Cerradão
Borda Interior Borda InteriorPredados 14 5 8 6Não-predados 5 16 11 15(mata): (χ2= 9,95; P= 0,0016; g.l.= 1; n= 40)(cerradão): (χ2= 0,80; P= 0,3702; g.l.= 1; n= 40)

48
Tempo (dias
0
5
10
15
20
25
30
35
1 2 3 4
MATA
CERRADÃO
Figura 2. Número de ovos predados na mata e no cerradão para os quatro dias do experimento na Reserva Ecológicado Roncador (IBGE), Brasília, DF.

49
DISTRIBUIÇÃO ESPACIAL DE QUALEAGRANDIFLORA MART. (VOCHYSIACEAE) EM UMCERRADO DE BRASÍLIA, DF.
Delano Moody Simões da Silva
RESUMOEste estudo tem como objetivo comparar o
padrão de distribuição espacial de Q. grandiflora emduas áreas de cerrado sensu stricto, sendo umaprotegida contra o fogo a aproximadamente 20 anos ea outra sofendo queimadas bienais desde 1989,localizadas na Reserva Ecológica do IBGE, Brasília-DF. Foram encontrados 75 indivíduos de Q.grandiflora na área de cerrado protegida e 19 naoutra área. Em ambas as áreas o padrão foi agrupado,indicando que o fogo não altera o padrão dedistribuição. A distribuição das freqüências decircunferência do fuste para as duas áreas, não difereestatisticamente, porém na área protegida indivíduoscom menos de 11 cm de circunferência foram maisfreqüentes, talvez indicando um maior recrutamentonesta área.
INTRODUÇÃOA maioria dos estudos fitossociológicos
realizados no cerrado tiveram como principaisobjetivos determinar a riqueza e a densidade dasespécies vegetais de uma determinada área. Ospoucos estudos realizados sobre o padrão dedistribuição espacial de árvores do cerradogeralmente dão maior ênfase ao padrão do conjuntode árvores de uma comunidade e não ao padrão decada população presente na comunidade (Meirelles &Luiz, 1995). Poucos estudos foram realizados comênfase na população (Oliveira et al., 1989).
Indivíduos de populações vegetais podemestar distribuídos no espaço de três maneiras: regular,aleatório e agregado. O padrão de distribuiçãoregular é o menos encontrado na natureza (Kershaw,1958), sendo o agregado o mais comum (Kershaw,1958; Hubbell, 1979; Gerig-Smith, 1979; San Jose etal., 1991; Silberbauer-Gottsberger & Eiten, 1987,Oliveira et al., !989). Segundo Kershaw (1959) opadrão de distribuição de uma dada população podeser atribuído à vários fatores, como:
1.Características da espécie: crescimento vegetativoou o modo de dispersão de suas sementes;2.Sociológicos: interação animal-planta ou planta-planta;3.Fisiográficos: ambientes em mosaico(microhabitat).
O fogo é um fator de perturbação importantepara o cerrado (Coutinho, 1979), sendo que o modocomo ele afeta o cerrado tanto ao nível de populaçõescomo de comunidades vegetais ainda não é bemconhecido. Dessa forma pouco se sabe sobre ainfluência do fogo na distribuição espacial deespécies vegetais no cerrado.
Os objetivos deste trabalho são (1)identificar e comparar o padrão de distribuiçãoespacial de Qualea grandiflora em duas áreas decerrado com diferentes regimes de queimada; (2) ecomparar a estrutura destas duas populações.
MATERIAL E MÉTODOSEste trabalho foi realizado na Reserva
Ecológica do IBGE (15˚ 55’ 58” S e 47˚ 51’ 02” W),Brasília-DF, durante a disciplina “Métodos emEcologia de Campo” pelo curso de Pós-graduação emEcologia da Universidade de Brasília no período de11 a 14 de março de 1996. Foram utilizadas duas áreaadjacentes de cerrado sensu stricto (Coutinho, 1978;Ratter, 1991) com dois históricos de queimadadiferentes. Uma das áreas é queimada de dois emdois anos desde 1989, enquanto a outra foi queimadauma única vez através de uma queimada acidental emoutubro de 1994, depois de 20 anos de proteçãocontra o fogo. Ao longo deste trabalho nóschamaremos a primeira área de cerrado bienal e asegunda de cerrado controle. Qualea grandifloraMart. (Vochysiaceae) é uma espécie de hábitoarbóreo-arbustivo comum nos cerrados de Brasília(Ratter, 1991), possuindo uma ampla distribuiçãogeográfica (Ratter & Dargie, 1990). Perde suas folhasuma vez por ano no final da estação seca (Morais etal., 1995), iniciando em seguida a brotação foliar efloral (Barbosa, 1983).
Dentre os diversos métodos de amostrageme índices que podem ser usados para detectarmos opadrões de distribuição espacial e a densidade deplantas (Brower et al., 1989), optou-se por utilizar aamostragem por parcelas, com diferentes dimensões,pois no caso de uma espécies de padrão agregado otamanho da unidade amostral que conseguirá detectaro tipo de padrão espacial é influenciado pelo tamanhodo grupo de indivíduos (Meirelles & Luiz, 1995). Foidemarcada uma área de 80x80m, tanto no cerradobienal quanto no controle, dividida em 64 parcelas de10x10m. Dentro de cada parcela foi contado onúmero total de indivíduos de Q. grandiflora emedida a circunferência do fuste ao nível do solo decada indivíduo.
Para detectarmos o grau de agrupamento oua distribuição espacial de Q. grandiflora utilizamosdois índices (Brower et al., 1989): Morisita(Id) e ocoeficiente de dispersão (CD). Para o índice deMorisita, se Id= 1,0 a distribuição é aleatória; se Id=

50
0 a distribuição é uniforme e se Id >1,0 a distribuiçãotende a ser agrupada. O coeficiente de dispersão écalculado através da razão da variância sobre amédia. Se CD < 1 então a população é uniforme; seCD > 1 a população é agrupada e se CD= 1 apopulação é aleatória. A significância de ambos ostestes é testada através de um teste qui-quadrado (χ2)(Brower et al., 1989). Além do índice de Morisita edo coeficiente de dispersão, analisamos graficamenteos padrões de agrupamento pelo método descrito porGoldsmith & Harrisson (1976), que possibilitavisualizar melhor em que escala e em qualintensidade está ocorrendo o agrupamento, através daanálise da variância do número de indivíduos queocorrem em diferentes tamanhos de parcelas. Autilização de mais de um índice ou método indicacom melhor segurança o padrão a ser aceito (Usher,1975).
RESULTADOSForam encontrados 75 indivíduos de Q.
grandiflora no cerrado controle e 19 no cerradobienal. A figura 1 mostra um croqui das áreasmarcadas com o número de indivíduos encontradosem cada parcela para as duas áreas. Analisando destaforma parece que no cerrado controle a distribuiçãotende a ser homogênea, enquanto no cerrado bienalparece ser agrupada, porém é necessário se testar seisto é verdade e em que tamanho de parcela isto estáocorrendo.
A Tabela 1 resume os resultadosencontrados para o índice de Morisita (Id) para trêstamanhos de parcelas (10x10, 20x20 e 40x40) com osseus respectivos χ2. No caso do cerrado controle sóno primeiro tamanho não detectou-se o padrãoagrupado, enquanto no cerrado bienal em todos ostamanhos foi detectado este padrão. Este mesmoresultado foi encontrado para o coeficiente dedispersão (Tab. 2).
A figura 2 mostra através da variânciaencontrada em cada unidade de amostragem ondeocorrem os agrupamentos e em que intensidade elesocorrem. Em ambas as áreas os indivíduos tendem aser agrupados, sendo que no cerrado controle ospicos ocorrem nos tamanhos de parcelas 4 e 16(20x20m e 40x40m) e no cerrado bienal ocorrempicos nos tamanhos 8 e 32 (40x20m e 80x40m).
As distribuições de circunferência de fusteencontradas nas duas áreas estão resumidas na figura3. Não existe uma diferença significativa entre essasduas distribuições (Komogorov-Smirnov= 0,09; p=1,0). É importante salientar que a divisão das classesfoi feita de maneira arbitrária e não biológica. Isso sedeve ao fato de que no cerrado controle a maioria dosindivíduos estavam rebrotando com vários ramos dabase (observ. pess.) e isto mascara a circunferência
real de indivíduos com mais de 10 cm decircunferência (< 10 cm= plântula ou jovem).Levando este fato em consideração podemos notarque no cerrado controle indivíduos comcircunferência inferior a 11 cm são mais freqüentes.Isto pode indicar um maior recrutamento nesta áreaquando comparada ao cerrado bienal (Fig. 3). Alémdisso, se as rebrotas estão ocorrendo principalmentena base, como foi dito acima, uma maiorcircunferência pode indicar um maior númerorebrotas na base no cerrado controle, mas este dadonão foi quantificado. Porém se isso for uma verdadepode explicar as maiores circunferências encontradasnesta área.
DISCUSSÃOOs dados aqui apresentados sobre a
distribuição espacial de Q. grandiflora, não diferemdos dados encontrados na literatura. Meirelles & Luiz(1995) estudando várias espécies de árvores docerrado encontraram 65 indivíduos de Q. grandifloraem uma área de 100x100m com um padrão dedistribuição agregado. Gottsberger & Silberbauer-Gottsberger (1983), também em um hectare,encontraram 183 indivíduos de Q. grandiflora comum padrão com alguns agrupamentos. Este padrãoagrupado segundo alguns autores não tem relaçãocom o modo de dispersão ou pelo menos este não étão determinante para o padrão (Gottsberger &Silberbauer-Gottsberger, 1983; San Jose et al., 1991).Talvez este padrão no caso de Q. grandiflora estejamais associado ao microhabitat. Como o cerrado éum ambiente em mosaico talvez isto explique melhora formação de agrupamentos, porém não temos dadospara fazer este tipo de afirmação.
O cerrado bienal apresentou uma menordensidade total de indivíduos e um menor número deindivíduos com menos de 11 cm de circunferênciaquando comparado com o cerrado controle (Fig. 3).Ramos (1990) sugere que o fogo pode levar a umadiminuição da densidade quando este ocorre em altasfreqüências, apesar de seus dados não mostrarem istopelo pouco tempo de estudo. A autora sugere quequeimadas frequentes levam a uma diminuição norecrutamento, uma alta mortalidade de plântulas e umgrande impacto em indivíduos com menor porte.Outra questão que a autora levanta é que plantas quesão freqüentemente queimadas sempre estão repondosuas folhas, tornando-se cada vez mais susceptíveisao ataque de patógenos, parasitas e herbívoros, sendoque estes poderiam aumentar a taxa de mortalidadedestas. Kellman (1986) também sugere que plântulastêm maior mortalidade e que plantas com rebrotabasal também teriam maior mortalidade. O autorainda sugere que regimes freqüentes de queimadaspoderiam levar algumas populações à extinçãonaquele local. Q. grandiflora tem um crescimento

51
inicial muito lento (Paulilo et al., 1993), além de umabaixa taxa fotossintética nesse período (Felippe &Dale, 1990). Sendo assim, talvez isso indique que elademore um tempo relativamente longo para atingirum tamanho suficiente para que o fogo ao passar,mate apenas a parte aérea e não o indivíduo como umtodo, explicando o menor número de indivíduos comcircunferência inferior a 11 cm no cerrado bienal.Outro fato que reforça esta hipótese é que maisimportante que o tamanho seria a circunferência doindivíduo para que este sobreviva após umaqueimada (Ramos, 1990). No caso de Q. grandifloraalém de suas plântulas com tamanho e circunferênciareduzido, suas rebrotas da base, apesar de serem deum indivíduos já estabelecido, também têm estascaracterísticas. Desse modo além das rebrotasestarem na base, mais susceptíveis à morte (Kellman,1986), elas têm uma circunferência muito pequena,talvez assim explicando uma possível maiormortalidade destas.
Desse modo com relação ao padrão dedistribuição encontrado para Q. grandiflora pareceque a freqüência de queimadas não estáinfluenciando este, pois nas duas áreas o padrão éagregado. Por outro lado, ocorreu uma redução dadensidade de Q. grandiflora na área bienal em todasas classes de tamanho, provavelmente devido àfreqüente destruição da parte aérea e da mortalidadede plântulas e rebrotas a cada queimada.
AGRADECIMENTOSGostaria de agradecer a J. D. Hay pelas
críticas e sugestões ao manuscrito. A F. S. Pinto e L.C. Baumgartem pelo leitura e críticas do manuscrito.À Reserva Ecológica do IBGE por ceder o local epelo apoio logístico ao curso “Métodos em ecologiade Campo”. A R. P. B. Henriques por organizar ocurso e pelas sugestões ao trabalho. A S. Marquespelo apoio e descontração no campo. E a todos oscompanheiros do curso de campo de 1996 pelosmomentos de trabalho e diversão em conjunto.
BIBLIOGRAFIABARBOSA, A. A. A. 1983. Aspectos da ecologia
reprodutiva de três espécies de Qualea(Vochysiaceae) num cerrado de Brasília, DF.Tese de Mestrado, UnB, Brasília, DF.
BROWER, J. E., J. H. ZAR & C. N. VON ENDE.Field and Laboratory methods for generalEcology, 3rd edition, Wm. C. Brown Publishers,Dubuque, Iowa.
COUTINHO, L. M. 1978. O conceito de cerrado.Revta. brasil. Bot. 1: 17-23.
COUTINHO, L. M. 1979. Aspectos ecológicos dofogo no cerrado. III. A preciptação de nutrientesminerais. Revta. brasil. Bot. 2: 97-102.
FELIPPE, G. M. & J. E. DALE. 1990. The effects ofphosphate supply on growth of plants fromBrazilian cerrado: Experiments with seedlings ofthe annual weed, Bidens gardneri Baker(Compositae) and the tree, Qualea grandifloraMart. (Vochysiaceae). Oecologia 82: 81-86.
GREIG-SMITH, P. 1979. Pattern in vegetation. J.Ecol. 67: 755-779.
GOLDSMITH, F. B. & C. N. M. HARRISSON.1976. Description and analysis of vegetation,116-118. In Methods in Plant Ecology, S. B.Chapman, editor, Osney Mead, Oxford.
GOTTSBERGER, G & I. SILBERBAUER-GOTTSBERGER. 1983. Dispersal anddistribution in a cerrado vegetation of Brazil,315-351. In Dispersal and Distribution. K.Kubizki, editor. Parey, Hamburg.
HENRRIQUES, R. P. B. & E. C. E. G. SOUSA.1986. Population structure , dispersion andmicrohabitat regeneration of Carapa guianensisin Northeastern Brazil. Biotropica 21(3): 204-209.
HUBBELL, S. P. 1959. Tree dispersion, abundanceand diversity in a tropical dry forest. Science203: 1299-1309.
KELLMAN, M. 1986. Fire sensivity of Casuariatorulosa in North Queensland, Australia.Biotropica 18(2): 107-110.
KERSHAW, K. A. 1958. An investigation of thestructure of a grassland community. II. Thepattern of Agrostis temus. J. Ecol. 46: 571-592.
KERSHAW, K. A. 1959. An investigation osstructure of a grassland community. II. Thepattern of Dactylis glomerata, Lolium perenne eTrifolium repens. III Discution and Conclusions.J. Ecol. 47: 31-53.
MEIRELLES, M. L. & A. J. B. LUIZ. 1995. Padrõesespaciais de árvores de um cerrado em Brasília,DF. Revta. brasil. Bot. 18(2): 185-189.
MORAIS, H. C., I. R. DINIZ & L. C.BAUMGARTEN. 1995. Padrões de produção defolhas e sua utilização por larvas de Lepidopteraem um cerrado de Brasília. Revta. brasil. Bot.18(2): 163-170.
OLIVEIRA, P. E. A. M.; J. F. RIBEIRO & M. I.GONZALES. 1989. Estrutura e distribuiçãoespacial de uma população de Kielmeyeracoriacea Mart. de cerrados de Brasília. Revta.brasil. Bot. 12: 39-47.
PAULILO, M. T. S.; G. M. FELIPPE & J. E. DALE.1993. Crescimento inicial de Qualeagrandiflora. Revta. brasil. Bot. 16(1): 37-46.
RAMOS, A. B. 1990. Efeitos da queima sobre avegetação lenhosa do cerrado. Tese de Mestrado,UnB, Brasília, DF.
RATTER, J. A. & T. C. D. DARGIE. 1990. Ananalysis of floristic composition of 26 cerrado

52
areas in Brazil. VIII Congresso da SociedadeBotânica de São Paulo.
RATTER, J. A. 1991. Guia para a vegetação daFazenda Água Limpa (Brasília, DF). Ed.Universidade de Brasília, DF.
SAN JOSE, J. J.; M. R. FARIÑAS & J. ROSALES.1991. Spatial pattern of trees and structuring
factors in a Trachypogon savanna of OrinocoLianos. Biotropica 23(2): 114-123.
SILBERBAUER-GOTTSBERGER, I & G. EITEN.1987. A hectare of cerrado. I. General aspects ofthe trees and thick stemmed shrubs. Phyton(Austria) 27: 55-91.
USHER, M. B. 1975. Analysis of patterns in real andartificial plant populations. J. Ecol. 63: 569-589.
Tabela 1 - Resultados obtidos para o índice de Morisita em três tamanhos de parcelas, para Q. grandiflora em duasáreas de cerrado na Reserva do IBGE.
Tamanho Cerrado controle Cerrado BienalId χ2 Id χ2
10x10 1,18 76,04 2,62 92,16*20x20 1,13 36,15* 2,43 40,79*40x40 1,13 12,74* 1,57 13,21** p < 0,001
Tabela 2 - Resultados obtidos para o coeficiente de dispersão (CD) em três tamanhos de parcelas, para Q.grandiflora em duas áreas de cerrado na Reserva do IBGE.Tamanho Cerrado controle Cerrado Bienal
CD χ2 CD χ2
10x10 1,21 76,04 1,46 92,16*20x20 2,41 34,14* 2,72 40,79*40x40 4,24 12,74* 5,12 15,34** p < 0,001

53
13 4 1 3 13 3 2 3 33 2 1 1 1
2 11 1 1 1 3 1 1 80 m1 2 1 4 31 3 1 3 1 21 1 1 1 2 1
80 MCerrado Controle
1 22 1 2
1 2 12 1 2 80 m
2
80 mCERRADO BIENAL
Figura 1- Esquema da área marcada nos dois tratamentos com o número de indivíduos de Q. grandifloraencontrados em cada parcela.

54
0
1
2
3
4
5
6
7
8
0 1 2 4 8 16 32
Tamanho das parcelas
Variâ
CC
CB
Figura 2- Gráfico demonstrando a intensidade do agrupamento e em que tamanho de parcela ele ocorre, através davariância do número de indivíduos em cada tamanho de parcela, para Q. grandilfora em duas áreas de cerrado naReserva do IBGE. CC= Cerrado controle; CB= Cerrado Bienal. Tamanhos correspondem: 1= 10x10 (64 par.); 2=20x10 (32 par.); 4= 20x20 (16 par.); 8= 40x20 (8 par.); 16= 40x40 (4 par.) e 32= 80x40 m (2 par.).
��������������������������������������������������������������������������������
����������������������������
�������������������������������������������������
����������������������������������������������������������������������������������������
����������������������������
����������������������������
0
5
10
15
20
25
0-10 11-20 21-30 31-40 41-50 >50
Classes de circunferência
���CCCB
Figura 3- Distribuição das freqüências de circunferências do fuste para Q. grandiflora em duas áreas de cerrado naReserva do IBGE nas duas áreas. CC= Cerrado controle; CB= Cerrado bienal.

55
ATIVIDADE FORRAGEIRA DE Atta sexdens(LINNAEUS, 1758) (HYMENOPTERA,FORMICIDAE): RELAÇÃO COMTEMPERATURA E UMIDADE RELATIVA DOAR
Everton Amancio dos Santos
RESUMOO ritmo de atividade diária da formiga
cortadeira Atta sexdens foi estudado durante três diasnuma área de cerrado situada na Reserva Ecológicado Instituto Brasileiro de Geografia e Estatística(IBGE), em Brasília. As observações foram feitas emum ninho, com intervalos de uma hora. Em cadaobservação, registrou-se o número de formigas queentravam e saíam do formigueiro, durante o intervalode um minuto. No minuto seguinte, coletou-se comuma pinça todo o material vegetal que estava sendoforrageado pelas formigas. Medidas de temperatura eumidade relativa do ar foram tomadas em cadaobservação, utilizando-se um psicrômetro. Osresultados obtidos mostram que o forrageamentoocorre tanto à noite quanto durante o dia, desde quenão seja interrompido por chuvas. No início damanhã, entre 6 e 7h, foi observado o retorno intensode formigas ao ninho, com subsequente reinício dasatividades por volta das 8h, sugerindo uma espécie de"mudança de turno". Aparentemente, a chuva tem umpapel mais importante do que a temperatura e aumidade sobre o forrageamento de Atta sexdens.
INTRODUÇÃOEntre as mais de 800 espécies de formigas,
estão as cortadeiras dos gêneros Atta (saúvas) eAcromyrmex (quenquéns), que possuem o hábito decortar e transportar fragmentos vegetais diversos paraseus ninhos subterrâneos (Della Lucia & Oliveira1993), os quais são usados como substrato para ocultivo de fungos que lhes servem de alimento(Weber 1966). Tais hábitos fazem dessas formigasuma das principais pragas da agricultura das regiõestropicais e subtropicais (Mariconi 1970).
Devido aos prejuízos causados à agricultura,essas formigas têm despertado a atenção das pessoasdesde o século XVI e várias pesquisas têm tratadodesses insetos. Contudo, embora a quantidade deestudos sobre as saúvas e quenquéns seja imensa, amaioria dos trabalhos trata de aspectos ligados aocontrole das formigas, restando muito ainda por fazerno campo da ecologia e do comportamento dessesorganismos.
Entre os trabalhos relacionados à ecologia eao comportamento dessas formigas, alguns tratam docomportamento forrageiro (e.g. Hodgson 1955;
Cherrett 1968; Lewis et al. 1974a; Gamboa 1975;Mintzer 1979; Fowler & Robinson 1979; Brener &Protomastro 1992; Brener 1993), outros tratam deaspectos como ritmo diário e sazonal de atividade(e.g. Wetterer 1990), influência dos fatoresambientais no padrão de forrageamento (e.g. Lewis etal. 1974b; Gamboa 1976; Maciel et al. 1995),distribuição de espécies de plantas atacadas ao longodas trilhas e estratégias territoriais (e.g. Brener &Sierra 1993), padrão de movimentação ao longo dastrilhas (e.g. Forti et al. 1984) e impacto da atividadeforrageira no fluxo de energia dos ecosistemas (e.g.Lugo et al. 1973).
Neste trabalho, procurou-se investigar arelação entre o padrão diário de forrageamento daformiga Atta sexdens e os fatores ambientaistemperatura e umidade relativa do ar.
MATERIAL E MÉTODOSEste estudo foi realizado na Reserva
Ecológica do IBGE Instituto Brasileiro de Geografiae Estatística, também conhecida como RECOR. Aárea constitui-se numa das Unidades de ConservaçãoPermanentes do Bioma Cerrado e está situada a35Km ao sul do centro de Brasília, entre 15€ 56'41"Se 47o53'07''W.
A RECOR está localizada na ChapadaBrasília, com uma altitude média de 1100 metros. Oclima local é estacional, com duas estações bemdefinidas: uma seca, de abril a setembro; outrachuvosa, que vai de outubro a março. A vegetação édividida em ceradão, cerrado, campos, brejos everedas e matas ciliares. O presente estudo foirealizado numa antiga área de cerrado, atualmentebastante modificada por espécies exóticas.
O ritmo de atividade diária deforrageamento de Atta sexdens foi monitoradodurante um período de três dias (12, 13 e 14 demarço de 1996). Nos dias 12 e 13, não foi possívelcobrir 24 horas de observação, devido a fortes chuvasque impediram a atividade das formigas. As medidasforam tomadas em intervalos de uma hora. A cadaobservação, registrou-se, durante um minuto, onúmero de formigas que entrava e saía do ninho ecoletou-se com uma pinça todo o material vegetalque estava sendo forrageado pelas formigas.Paralelamente, foram tomadas medidas de umidaderelativa do ar e de temperatura, utilizando-se umpsicrômetro. Nas observações noturnas, utilizou-seuma lanterna de testa, o que pode ter alterado ocomportamento das formigas, segundo Hodgson1955 e Maciel et al. 1995. No laboratório, todo omaterial vegetal coletado foi seco em estufa a 60oC,durante 24 horas, e o peso seco (biomassa) medidocom precisão de 0,01g.

56
RESULTADOSDurante o primeiro dia de observação, a
atividade das formigas teve sequência ao longo detodo o dia, tendo sido interrompida por uma fortechuva a partir das 18h. Embora as formigasestivessem ativas durante todo o dia, o forrageamentoocorreu apenas das 7 às 14h. A temperatura oscilouentre 18 e 27 oC, com atividade forrageira em todo ointervalo de temperatura, sendo que os picosocorreram entre 23 e 27oC. A umidade do ar variouentre 61 e 90% ao longo do dia, com pico deforrageamento entre 61 e 74%. Os picos deforrageamento aconteceram entre as 12 e 14h.
No segundo dia de observação, a atividadedas formigas foi monitorada das 6 às 19h. Atemperatura variou entre 18 e 28oC, com máxima às15h e mínima entre 6 e 7h. A umidade relativa do arvariou entre 60 e 95%, com màxima às 7h e mínimaàs 16h. O pico máximo de forrageamento ocorreu às16h, sob temperatura de 26oC e umidade do ar de60%.
O último dia de observação compreendeuum período de 24 horas de atividade. A temperaturavariou entre 16 e 27oC, com máxima às 13h emínima à zero hora. A umidade do ar oscilou entre64 e 90%, com máximas entre zero hora e 6h emínimas entre 12 e 14h. O forrageamento teve seupico máximo entre 21 e 23h, mas apresentou um picomenor das 2 às 4h.
Ao longo dos três dias de observação,verificou-se que no início da manhã houve umainterrupção quase total e, por vezes, total nasatividades das formigas. Este fato pode sercomparado a uma espécie de "mudança de turno", emque um grupo de formigas substitui outro em suasatividades.
DISCUSSÃOOs efeitos da temperatura, luminosidade e
umidade sobre o comportamento forrageiro deformigas dos gêneros Atta e Acromyrmex tém sidoamplamente estudado tanto nos trópicos quanto naszonas subtropicais e temperadas, porém asconclusões ainda são muito diversas. Hodgson(1955), por exemplo, afirma que o aumento daintensidade luminosa e da temperatura determina oritmo de atividade da espécie tropical Attacephalotes. Cherrett (1968), também trabalhandocom Atta cephalotes, verificou que as mudançasdiárias e sazonais no padrão de atividade forrageiraestão relacionadas com a temperatura e a umidade, aomenos em espécies tropicais. Lugo et al. (1973)observaram que Atta colombica tem seu pico deforrageio ao meio dia, quando as temperaturas sãomais elevadas.
Em Acromyrmex versicolor versicolor,Gamboa (1976) constatou que a temperatura do solo,mais que a temperatura do ar, determina a presençaou ausência de atividade de formigas na superfície,assim como o término do forrageio. Porém, eleobservou que outros fatores como umidade eintensidade luminosa podem estar agindoconjuntamente para provocar tais respostas. Mintzer(1979) verificou que em Atta mexicana o forrageio seinicia após temperaturas de 12oC e que éinterrompido quando a temperatura chega a 14oC.Fowler & Robinson (1979) trabalharam com Attasexdens no Paraguai e notaram que fatoresambientais como temperatura podem determinar osperíodos de forrageamento em formigas do gêneroAtta, mas outros fatores como disponibilidade derecursos podem estar envolvidos.
Hölldobler & Wilson (1990) dizem que asformigas são extremamente termofílicas; por isso, asvariações climáticas são consideradas determinantesde seus ritmos forrageiros, especialmente nos climassubtropicais e temperados. Segundo Hölldobler &Wilson (1990), cada espécie de formiga opera numafaixa de temperatura e umidade que funciona comoum envelope. Brener (1993) sugere que os padrões deatividades forrageiras de Atta laevigata não sãodeterminados por fatores ambientais, e sim pornecessidades nutricionais. Contudo, ele ressalta quefatores climáticos extremos poderiam tambémocasionar mudanças nos padrões de forrageio ealguns desses fatores, especialmente temperatura,possuem nos ambientes tropicais um papel maisimportante como limitante e/ou disparador do quecomo regulador da atividade das formigas.
Contrariamente, Maciel et al. (1995)atribuem um papel muito importante à temperaturacomo reguladora da atividade forrageira deAcromyrmex subterraneus subterraneus. Vale notarque a correlação de Spearman realizada por Maciel etal. (1995) mostrou-se positiva e significativa apenasquando relacionava temperatura mínima e número deoperárias em atividade. Este resultado pode ser umaevidência de que, conforme sugerido por Brener(1993), a temperatura atua mais como um fatorlimitante e/ou disparador do que como um regulador.A partir dessas informações, é possível notar oquanto as opiniões relativas ao comportamentoforrageiro dos Attini são divergentes.
Os resultados obtidos neste trabalhomostram que Atta sexdens forrageia tanto durante odia quanto à noite, exceto quando chuvas fortesinterrompem o forrageamento. A atuação da chuvacomo bloqueadora da atividade das formigas já haviasido notada por vários pesquisadores (e.g. Hodgson1955; Lugo et al. 1973; Lewis et al. 1974a, 1974b;Holldobler & Wilson 1990). As medidas relativas à

57
temperatura e à umidade mostram que esta espécieforrageia sob temperaturas que variam de 16 a 28oC,e umidade relativa do ar de 58 a 95%.
AGRADECIMENTOSSou muito grato ao Saulo Marques, por ter
cedido o psicrômetro; à Rosane Collevatti, peloempréstimo do relógio e pela ajuda na análise dosdados; ao Felipe Costa, pelas críticas e sugestões; aoRaimundo Henriques, pela orientação e auxílio combibliografia; à June Springer de Freitas, por terpermitido o uso do seu computador e, por fim, aopessoal do IBGE, por ter viabilizado a realização doCurso de Campo em sua Reserva Ecológica..
REFERÊNCIASBRENER, A. J. F. 1993. Influencia de la
estacionalidad sobre los ritmos forrajeros de Attalaevigata (Hymenoptera: Formicidae) en unasabana tropical. Revista de Biologia Tropical 41:897-899.
BRENER, A. J. F., AND PROTOMASTRO, J. 1992.Patrones forrajeros de dos especies simpatricasde hormigas cortadoras de hojas (Attini,Acromyrmex) en un bosque subtropical.Ecotropicos 5: 32-43
BRENER, A. J. F., AND SIERRA, C. 1993.Distribution of attacked plants along trails inleaf-cutting ants (Hymenoptera: Formicidae):consequences in territorial strategies. Revista deBiologia Tropical 41: 891-896.
CHERRETT, J. M. 1968. The foraging behaviour ofAtta cephalotes L. (Hymenoptera: Formicidae):I. Foraging pattern and plant species attacked intropical rain forest. Journal of Animal Ecology37: 387-403.
DELLA LUCIA, T. M. C., AND OLIVEIRA, M. A.1993. Forrageamento. In T. M. C. Della Lucia(Ed.). As formigas cortadeiras, pp.84-105. Folhade Viçosa, Viçosa, Minas Gerais.
FORTI, L. C., SILVEIRA-NETO, S., ANDPEREIRA-DA-SILVA, V. 1984. Atividadeforrageira de Atta sexdens rubropilosa Forel,1908 (Hymenoptera, Formicidae): fluxo evelocidade dos indivíduos na trilha,caracterização dos indivíduos forrageiros eduração e número de jornadas de coleta devegetais. Revista Brasileira de Entomologia 28:275-284.
FOWLER, H. G., AND ROBINSON, S. W. 1979.Foraging by Atta sexdens (Formicidae: Attini):seasonal patterns, caste and efficiency.Ecological Entomology 4: 239-247.
GAMBOA, G. J. 1975. Foraging and leaf-cutting ofthe desert gardening ant Acromyrmex versicolorversicolor (Pergande) (Hymenoptera:Formicidae). Oecologia (Berlin) 20: 103-110.
GAMBOA, G. J. 1976. Effects of temperature on thesurface activity of the desert leaf-cutter ant,Acromyrmex versicolor versicolor (Pergande)(Hymenoptera: Formicidae). American MidlandNaturalist 95: 485-491.
HODGSON, E. S. 1955. An ecological study of thebehaviour of the leaf-cutting ant Atta cephalotes.Ecology 36: 293-304.
HÖLLDOBLER, B., AND WILSON, E. O. 1990.The ants. Harvard University Press, Cambridge.
LEWIS, T., POLLARD, G. V., AND DIBLEY, G. C.1974a. Rhythmic foraging in the leaf-cutting antAtta cephalotes (L.) (Formicidae: Attini). Journalof Animal Ecology 43: 129-141.
LEWIS, T., POLLARD, G. V., AND DIBLEY, G. C.1974b. Micro-environmental factors affectingdiel patterns of foraging in the leaf-cutting antAtta cephalotes (L.) (Formicidae: Attini). Journalof Animal Ecology 43: 143-153.
LUGO, A. E., FARNWORTH, E. G., POOL, D.,JEREZ, P., AND KAUFMAN, G. 1973. Theimpact of the leaf cutter ant Atta colombica onthe energy flow of a tropical wet forest. Ecology54: 1292-1301.
MACIEL, M. A. F., DELLA LUCIA, T. M. C.,ARAÚJO, M. S., AND OLIVEIRA, M. A. 1995.Ritmo diário de atividade forrageadora daformiga cortadeira Acromyrmex subterraneussubterraneus Forel. Anais da SociedadeEntomológica do Brasil 24: 371-378.
MARICONI, F. A. M. 1970. As saúvas. São Paulo,Editora Agronômica CERES.
MINTZER, A. 1979. Foraging activity of theMexican leafcutting ant Atta mexicana(F.Smith), in a Sonoran Desert habitat(Hymenoptera: Formicidae). Insectes Sociaux26: 364-372.
WEBER, N. A. 1966. Fungus-growing ants. Science153: 587-604.
WETTERER, J. K. 1990. Diel changes in foragersize, activity, and load selectivity in a tropicalleaf-cutting ant, Atta cephalotes. EcologicalEntomology 15: 97-104.

58
ESTRUTURA POPULACIONAL DE UMABORBOLETA “ESTALADEIRA”, Hamadryasferonia (L.) (LEPIDOPTERA: NYMPHALIDAE)
Felipe A. P. L. Costa
INTRODUÇÃOHamadryas Hübner (Nymphalidae:
Nymphalinae) é um gênero composto por 20 espéciesde borboletas restritas ao Novo Mundo,especialmente os Neotrópicos (Jenkins, 1983).“Assenta-pau” e “estaladeira” são dois dos váriosnomes populares atribuídos a esses insetos emdiferentes regiões brasileiras. Os nomes fazem alusãoao hábito dos adultos pousarem sobre o tronco deárvores e aos ruídos que produzem em vôo (não emtodas as espécies), respectivamente.
Hamadryas feronia L. é uma espécie deampla distribuição, encontrada desde o sul dos EUA(Ehrlich & Ehrlich, 1961), passando pela AméricaCentral (DeVries, 1987) e chegando ao sul do Brasil(Brown, 1992). Não há dimorfismo sexual e adistinção entre machos e fêmeas depende do exameda genitália. Os adultos se alimentam de frutos eseiva fermentados (Brown, 1992) e as larvas utilizamcomo hospedeiras plantas da família Euphorbiaceae,notadamente espécies dos gêneros DalechampiaPlum. e Tragia Plum. (DeVries, 1987; Brown, 1992).
Como de resto acontece com todas asespécies dos gênero, machos e fêmeas de H. feroniapousam de cabeça para baixo e com as asas abertassobre o tronco de árvores. Nessas circunstâncias, umindivíduo pode facilmente passar desapercebido.Quando perturbada, uma borboleta em repouso podeexibir um vôo breve, retornando ao mesmo tronco,ou, se a perturbação for brusca ou persistente, elapode abandonar seu sítio original e, depois de umvôo errático, pousar em outro substrato mais oumenos próximo.
Nesse trabalho, eu apresento os resultadosde um programa de captura-marcação-recaptura (C-M-R) com uma população (sensu Brown & Ehrlich,1980) de H. feronia. Os resultados obtidos mostramque (1) pelo menos a curto prazo, machos e fêmeaspodem permanecer fiéis e restritos a uma área de vidareduzida; (2) entre os machos, o grau de dano nasasas influencia o acesso a uma fonte potencial dealimentação e, talvez indiretamente, a suaprobabilidade de acasalamento.
MATERIAL E MÉTODOSO trabalho de campo foi realizado, entre os
dias 11 e 14 de março de 1996, na Reserva Ecológicado IBGE (15˚50’ S e 47˚50’ W), Brasília, DF. Umsítio de estudo foi estabelecido em torno de uma
árvore de “ipê-roxo”, Tabebuia impetiginosa (Mart.ex DC) Standl. (Bignoniacae), plantada à margem deuma estrada asfaltada dentro da Reserva. Essa árvore,com ca. 3,5 m de altura, estava com uma ferida nabase do tronco que atraia muitos visitantes: abelhas,vespas, drosófilas, mariposas, um besouroCurculionidae e, além de H. feronia, pelo menos 7espécies de borboletas, com destaque para umaborboleta “80”, Callicore (= Catagramma) sorana(Godart), e uma “canoa-azul”, Prepona laertes(Hübner).
Aproveitando a descontinuidade davegetação, eu delimitei uma “ilha”, com ca. 1950 m2,em torno da árvore de T. impetiginosa (doravantereferida apenas como “ipê”), contendo 85 “troncos”(arbustos e árvores, as mais altas com 8-9 m dealtura, além de 3 postes de iluminação) e afastada deilhas vizinhas por trechos de vegetaçãopredominantemente herbácea. Além de um certoisolamento, o local foi escolhido pela aparentefacilidade de se encontrar indivíduos de H. feronia.Há registros de que outras espécies de Hamadryasocorrem dentro da Reserva (Ferreira, 1982), masnenhuma delas foi vista durante a realização dessetrabalho.
Outras duas agregações de H. feronia foramnotadas nas proximidades do sítio de estudo: aprimeira (A-50), distante ca. 50 m na direção NE, e asegunda (A-240), distante ca. 240 m e em torno doalojamento da Reserva; além disso, apenas mais 2indivíduos foram observados a ca. 70 m na direçãoSW. Depois de iniciado esse trabalho, ca. 15borboletas foram capturadas e/ou observadas nessasagregações, mas nenhum indivíduo havia sidoanteriormente capturado no sítio de estudo. A únicaevidência direta de movimentação entre essas áreasfoi a recaptura de um macho (#22), no dia 14, nasproximidades de A-50. Contudo, menos de 2 horasdepois, ele foi novamente capturado no limitenordeste do sítio de estudo.
No último dia de trabalho de campo, eumedi a posição dos troncos em relação à estrada e fizum mapa detalhado do local. Sobre o mapa, econsiderando apenas os 24 troncos nos quais houvepelo menos uma captura, foram calculadas asdistâncias entre troncos vizinhos (os lados de umpolígono ligando todos os 24 troncos) e a distância decada tronco em relação ao ipê.
Para a captura das borboletas dentro do sítiode estudo, foi estabelecida uma rotina de inspeção,tomando-se o ipê como ponto de partida. No início decada inspeção, eu registrei a hora do dia, as espéciese o número de borboletas pousadas no ipê. Ao final,foram executadas 55 rotinas de inspeção pela manhãe à tarde (exceto no dia 12, quando o turno da tardefoi suspenso com a chegada das chuvas). Na noite dodia 11, foi feita uma rápida visita ao local.

59
Durante as inspeções, eu evitei capturarborboletas que não estivessem pousadas, comexceção daquelas capturadas voando em torno do ipê(nesse caso, eu evitei capturar as borboletas exibindo“vôos de perseguição”, muitas vezes envolvendo 3ou 4 indivíduos). Borboletas em fuga, depois de umaprimeira tentativa de captura malsucedida, foramperseguidas e eventualmente capturadas até umadistância de ca. 10 m do tronco original.
Para cada borboleta capturada, eu registrei otronco de pouso, o estado das asas (numa escala de 1a 5, de acordo com a quantidade de danos) e o sexo.A avaliação do estado das asas visou, basicamente,estimar a proporção da superfície perdida e não o seuestado de descamação ou senescência. Emborahouvesse um componente temporal no acúmulo dedanos, alguns fatores podem ter reduzidobruscamente a superfície útil das asas. Por exemplo,em duas ocasiões, eu flagrei lagartos sobre árvorescom borboletas pousadas e desconfio que certosdanos (particularmente na base das asas anteriores)tenham sido produzidos durante investidas dessespredadores. Machos e fêmeas começaram a serreconhecidos com segurança apenas na manhã do dia12 e os indivíduos capturados antes disso foramclassificados quanto ao sexo na primeira recaptura.
Na primeira captura, as borboletasreceberam uma marca e um número nos dois ladosdas asas anteriores (escritos com uma caneta Pilotvermelha de ponta fina), tomando-se o cuidado denão atrapalhar o padrão de camuflagem. O tempo deretenção na primeira captura de cada borboleta foicronometrado e elas foram liberadas no ponto exatoem que foram encontradas ou defronte do tronco emque estavam pousadas. Eu não registrei ocomportamento das borboletas após a liberação, masem algumas ocasiões elas voltaram a pousar sobre otronco original.
Utilizando o mapa feito para o sítio deestudo, foram calculados dois parâmetros damovimentação dos indivíduos recapturados:(1) di: a distância percorrida pelo indivíduo i entre aprimeira e a segunda capturas (di1), entre a segunda ea terceira (di2), e assim por diante. Para as recapturasno mesmo tronco, d= 0;(2) D1: a distância total percorrida pelo indivíduo i,que foi igual à soma dos seus valores de d. Para asborboletas recapturadas apenas uma vez, Di= di1.
Como um teste preliminar, eu investiguei seo tempo de retenção (tR) afetou os parâmetros demovimentação das borboletas recapturadas. Para isso,foi feita uma análise de regressão linear (mínimosquadrados) entre tR vs. di1 e tR vs. Di. Nenhuma dasanálises produziu índices significativos (maior valorabsoluto de t= 0,95; P > 0,3; mantendo-se H0: b= 0),
mesmo quando os valores iguais a zero (i.e., di1 ouDi= 0) foram ignorados (maior valor absoluto de t=0,79; P > 0,4).
Esses resultados mostram que a variaçãoregistrada em tR (amplitude: 51-282 sec; média= 118sec; N= 61) não foi suficiente para afetar amovimentação das borboletas posteriormenterecapturadas. Em outras palavras, o “trauma” dacaptura parece não ter alterado a probabilidade derecaptura dos indivíduos (mas veja Morton, 1984) e,sendo assim, o tempo de retenção foi excluído dasanálises posteriores. Como um exemplo,circunstancial e drástico dessa conclusão, valeregistrar sobre um casal capturado em cópula no dia13. Após a captura (16:15 h), a fêmea (#58) e omacho (#59) permaneceram retidos por ca. 90 min(na sombra, dentro de um saco de papel). Depoisdisso, eles foram marcados e liberados e, embora omacho não tenha sido reencontrado, a fêmea foirecapturada no dia seguinte, próxima ao ponto deliberação.
Os testes estatísticos empregados nesseartigo seguem Siegel (1971) e Sokal & Rohlf (1981).Os valores críticos obtidos foram comparados comvalores tabelados fornecidos por Fisher & Yates(1971) e Rohlf & Sokal (1981). O valor crítico obtidopara o teste-G foi submetido, quando apropriado(veja Sokal & Rohlf, 1981, p. 704 e 727), a umacorreção pelo método de Williams. No que se segue,“g.l.” indica os respectivos graus de liberdade.
RESULTADOSEsforço de Amostragem
Durante os 4 dias de trabalho de campo,foram capturados 64 indivíduos de H. feronia dentrodo sítio de estudo: 28 fêmeas (F), 27 machos (M) e 9cujo sexo permaneceu desconhecido (?) (2 na TabelaI). O número de capturas aumentou com o esforço deamostragem, de tal modo que o “tempo de espera”para uma captura foi semelhante em 3 dos 4 dias (4na Tabela I). No dia 13, a redução no tempo deespera provavelmente refletiu um aumento no nívelde atividades das borboletas, depois de um dia(12/III) parcialmente chuvoso. Nos períodos semchuvas, o nível de atividade, como medido pelonúmero de borboletas encontradas no ipê, se manteveao longo do dia, com um aumento acentuado apenasno final da tarde (Fig. 2).
Levando em conta todas as recapturas, 53%das borboletas (34/64: 16F + 14M + 4?) foramreencontradas pelo menos uma vez, em 82 episódiosde recaptura (3 na Tabela I). Deixando de lado asrecapturas no mesmo dia, 51% das borboletasmarcadas nos 3 primeiros dias (29/57: 13F + 12M +4?) foram reencontradas pelo menos uma vez nosdias seguintes, em 43 episódios de recaptura (Tabela

60
II). Com esses resultados, e admitindo uma razãosexual de 1:1 (veja adiante), foi possível estimar otamanho da população dentro do sítio de estudo.Usando o Índice de Lincoln-Petersen, o tamanhofinal estimado foi de 79 borboletas (Tabela II). Essaestimativa está muito próxima do número de troncosno local (85) e levanta a suspeita de que um limitesuperior ao número de indivíduos residentes foiimposto pelo número de troncos disponíveis. Umasuspeita que é reforçada se comparamos o número decapturas em diferentes setores dentro do sítio deestudo (Fig. 1).
Razão SexualAs proporções finais de machos (27/55) e
fêmeas (28/55) capturados suportam facilmente ahipótese de uma razão sexual de 1:1. Além disso, aproporção de machos e fêmeas recapturados aomenos uma vez foram semelhantes, não importandose a comparação incluía (14/27 M e 16/28 F; G=0,15; P > 0,5) ou não (12/25 M; 13/23 F; G= 0,35; P> 0,5) recapturas no mesmo dia. Em outras palavras,machos e fêmeas foram igualmente abundantes eigualmente prováveis de serem recapturados,indicando semelhanças comportamentais entreindivíduos dos dois sexos.
Nos 3 dias com amostragens pela manhã e àtarde (o que exclui o dia 12), a proporção de capturasenvolvendo machos e fêmeas oscilou (2 e 3 naTabela I), mas sem nenhum desvio significativo deuma razão sexual de 1:1. Houve, entretanto, umadiferença consistente entre os sexos quanto aoprincipal turno de amostragem (manhã vs. tarde):71% (40/56) das capturas de machos acontecerampela manhã, enquanto, no mesmo período, foramfeitas apenas 44% (27/61) das capturas envolvendofêmeas. Para os machos, mas não para as fêmeas,esse resultado é significativamente diferente doesperado sob a hipótese de igualdade no número decaptura nos dois turnos (Tabela III). Isso mostra quea “disponibilidade” dos machos efetivamente diminuidurante o dia, talvez por eles terem reduzido seunível de atividades, permanecendo mais tempo em“refúgios” (dentro ou fora do sítio de estudo).
MovimentaçãoA movimentação de machos e fêmeas dentro
do sítio de estudo, medida pelos parâmetros d e D,foram semelhantes. A distância total percorrida pelosmachos (DM= 43,94 m; s= 29,88 m; N= 14) foi umpouco maior que a das fêmeas (DF= 36,22 m; s= 21,9m; N= 16), mas a diferença não foi significativa (t=0,814; P > 0,4). A distância média percorrida entrerecapturas foi virtualmente idêntica entre os doissexos: dM= 16,64 m (N= 37 recapturas) e dF= 16,56m (N= 35). Esses resultados sustentam a observação
feita anteriormente sobre semelhançascomportamentais entre machos e fêmeas.
Embora a distância percorrida entrerecaptura tenha variado inversamente com o tempode espera (4 e 5 na Tabela I), sugerindo um efeito dadensidade na movimentação diária das borboletas,uma análise de regressão mostrou que essa relaçãonão foi significativa (H0: b= 0; t= -2,64; gl= 2; P >0,1). Ainda que o reduzido tamanho amostral tenhaimpedido a descoberta de uma eventual relação entrea densidade e a movimentação das borboletas, deve-se notar que os resultados dos dois primeiros diasprovavelmente representaram subestimativas. Assim,enquanto as médias para os dias 13 e 14 incluíramtanto a distância entre capturas sucessivas no mesmodia como também a distância percorrida desde aúltima captura no dia anterior, a média para o dia 11envolveu apenas a distância entre recapturas naqueledia. Por fim, como já foi notado, no dia 12 o esforçode amostragem esteve restrito ao turno da manhã.
Em função dos resultados acima, todos osvalores obtidos foram agrupados (machos + fêmeas +desconhecidos) e a distribuição de frequências dasdistâncias percorridas foi comparada com a distânciaentre os troncos dentro do sítio de estudo. Se amovimentação das borboletas foi predominantementeuma movimentação entre troncos vizinhos seria de seesperar que as respectivas distribuições mostrassemuma certa concordância. Alternativamente, foi feitauma comparação entre a distribuição de frequênciasdas distâncias percorridas e as distâncias entre ostroncos e o ipê.
A Figura 3 mostra a distribuição defrequências das distâncias percorridas pelosindivíduos recapturados ao lado das frequênciasesperadas de acordo com (1) as distâncias entretroncos vizinhos; (2) as distâncias dos troncos emrelação ao ipê. Um teste-G para bondade de ajustemostrou que a distância percorrida pelas borboletasfoi muito semelhante à distância dos troncos emrelação ao ipê (G= 1,004; gl=2; P > 0,5), mas não emrelação à distância entre troncos vizinhos (G=26,304; gl= 1; P < 0,001), sustentando a interpretaçãode que a movimentação dentro do sítio de estudo foiprincipalmente pelo acesso ao ipê. Esse resultado émais facilmente interpretado se levamos em contaque o ipê foi a única fonte evidente de alimentaçãopara os adultos dentro do sítio de estudo.
Danos nas Asa e o Acesso ao IpêA classificação das borboletas capturadas de
acordo com o estado das asas é mostrado na Figura 4.Uma comparação entre machos e fêmeas (teste-U deMann-Whitney), mostrou que ambos estavamigualmente danificados (t= 0,164; g.l.= 52; P > 0,8).Apesar disso, deve-se notar a presença exclusiva defêmeas na última classe, supostamente as mais velhas

61
entre todas as borboletas capturadas. Uma dessasfêmeas já tinha perdido metade das asas posterioresquando foi capturada.
Agrupando os resultados de machos efêmeas, uma comparação entre os indivíduoscapturados ao menos uma vez no ipê (15/27 M; 18/28F) contra aqueles que foram capturados apenas nosoutros troncos (12/27 M; 10/28 F), produziuresultados distintos. Para as fêmeas, o resultadosobtido (U= 83; P > 0,1) indicou que o grau de danosnão diferiu entre as borboletas dos dois grupos,enquanto, entre os machos, a diferença encontrada foisignificativa (U= 126; P < 0,05) e mostrou que osmachos capturados no ipê tinham asas menosdanificadas.
Aparentemente, os machos com as asas maisdanificadas tinham chances reduzidas de acesso e/oupermaneciam menos tempo no ipê. Evidênciascircunstanciais sugerem que essas borboletas podempermanecer pousadas por períodos prolongados detempo. Assim, no dia 11, eu cheguei a registrar apermanência de um macho (#15, classe 4 de dano)por quase 6 horas (11:45 até 17:06 h, voando emseguida) num mesmo sítio de pouso. A diferença noestado das asas entre os indivíduos dos dois sexoscapturados no ipê levanta a suspeita de que osmachos foram mais tolerantes com as fêmeasdanificadas, talvez por elas ainda representaremparceiras potenciais.
DISCUSSÃOA população estudada de H. feronia pode ser
descrita como um conjunto de machos e fêmeasigualmente abundantes e gravitando em torno de umafonte potencial de alimentação (a seiva do ipê). Apresença de indivíduos de H. feronia no ipê durantetodo o dia (i.e., entre 8:30 e 17:30 h), ao lado deoutras espécies de borboletas e de outros insetos,sugere que a disponibilidade de seiva foi mais oumenos permanente. Além do fluxo de seiva, umlimite superior ao número de indivíduos residentesparece ter sido imposto pela disponibilidade detroncos nas proximidades do ipê.
Por qualquer dos critérios utilizados (i.e.,incluindo capturas no mesmo dia ou não), a taxa derecaptura foi elevada, principalmente secompararmos com os valores normalmente obtidosem estudos com populações de borboletas (e.g.,Ehrlich, 1984) e, mais importante, com outrosestudos envolvendo espécies de Hamadryas (e.g.,Ross, 1963; DeVries, 1983; veja ainda Young, 1974).Como os recursos alimentares normalmenteexplorados pelos adultos são relativamente efêmeros(i.e., frutos em decomposição) seria de se esperaruma alta vagilidade e, assim, uma probabilidadereduzida de recaptura. No entanto, recursos quepermanecem disponíveis por mais tempo podem
permitir o estabelecimento de populações commembros mais sedentários, como parece ter sido ocaso do presente estudo.
A diferença observada na captura demachos, mas não na de fêmeas, de acordo com oturno de amostragem, sugere, num primeiromomento, a existência de padrões distintos naexploração de uma mesma fonte de alimentação. Poroutro lado, é possível que essas diferenças tenhamsido reflexo de uma hierarquia mais acentuada entreos machos pelo acesso ao ipê (veja Marini-Filho,1996). Machos com as asas mais danificadas podiamestar reduzindo seu nível de atividades,permanecendo mais tempo empoleirados em"refúgios", dentro ou fora do próprio sítio de estudo.
O acesso dos machos ao ipê foi influenciadopelo estado das suas asas, talvez porque os machoscom as asas danificadas (i.e., com um tamanhoefetivo reduzido) fossem mais frequentementeafastados pelos outros. O acúmulo de danos, além dereduzir o tamanho efetivo das asas, pode alterar suafuncionalidade em determinados contextos, como,por exemplo, em disputas entre machos (veja Otero,1986; Marini-Filho, 1996). Existem evidências deque o tamanho das asas em Hamadryas afeta oresultado de disputas intra- e interespecíficas peloacesso a uma fonte potencial de alimento (vejaMarini-Filho, 1996).
O presente trabalho foi realizado no final daestação chuvosa de Brasília (dezembro a março) e oencontro do casal #58 e #59 (um macho da classe 2com uma fêmea da classe 5 de danos) indica que asfêmeas ainda estavam receptivas, ao mesmo tempoque sugere que elas são capazes de acasalamentosmúltiplos (veja Ehrlich & Ehrlich, 1978). A presençade fêmeas no ipê, a despeito do estado das suas asas,pode ter tido um componente sexual. Sendo maistolerantes com as fêmeas danificadas, os machos comacesso ao ipê podiam aumentar o número deparceiras potenciais. Por outro lado, se o ipê, além deum sítio de alimentação foi usado como um localpara encontro de parceiros, os machos com as asasdanificadas tiveram chances reduzidas deacasalamento.
BIBLIOGRAFIABROWN, K.S., JR. 1992. Borboletas da Serra do
Japi: diversidade, habitats, recursos alimentares evariação temporal. In: L.P.C. Morellato (Ed.).História Natural da Serra do Japi, p. 142-87.Unicamp, Campinas.
BROWN, I.R. & P.R. EHRLICH. 1980. Populationbiology of the checkerspot butterfly, Euphydryaschalcedona structure of the Jasper Ridge Colony.Oecologia 47: 239-51.

62
DEVRIES, P.J. 1983. Hamadryas februa. In: D.H.Janzen (Ed.). Costa Rican Natural History, p.729-30. Chicago University Press, Chicago.
_____. 1987. The Butterflies of Costa Rica and theirNatural History. Princeton University Press,Princeton.
EHRLICH, A.H. & P.R. EHRLICH. 1978.Reproductive strategies in the butterflies: I.Mating frequency, plugging, and egg number. J.Kans. Entomol. Soc. 51: 666-97.
EHRLICH, P.R. 1984. The structure and dynamics ofbutterfly populations. In: R.I. Vane-Wright &P.R. Ackery (Eds.). The Biology of Butterflies,p. 25-40. Academic Press, London.
_____ & A.H. Ehrlich. 1961. How to know theButterflies. W.C.Brown, Dubuque.
FERREIRA, L.M. 1982. Comparações entre Riqueza,Diversidade e Equitabilidade de Borboletas emTrês Areas com Diferentes Graus dePerturbação, Próximas à Brasília. UnB, Brasília.Tese de Mestrado.
FISHER, R.A. & F. YATES. 1971. TabelasEstatísticas para Biologia, Medicina eAgricultura. Polígono, São Paulo.
JENKINS, D.W. 1983. Neotropical Nymphalidae. I.Revision of Hamadryas. Bull. Allyn Mus. 81: 1-146.
LENKO, K. & N. PAPAVERO. 1979. Insetos noFolclore. Conselho Estadual de Artes e CiênciasHumanas, São Paulo.
MARINI-FILHO, O.J. 1996. Defesa de RecursosAlimentares e Interações Aéreas entreBorboletas Simpátricas do Gênero Hamadryas.Unicamp, Campinas. Tese de Mestrado.
MORTON, A.C. 1984. The effects of marking andhandling on recapture frequencies of butterflies.In: R.I. Vane-Wright & P.R. Ackery (Eds.). TheBiology of Butterflies, p. 56-68. AcademicPress, London
OTERO, L.D. 1986. Contribuición a la HistoriaNatural del Genero Hamadryas (Lepidoptera:Nymphalidae). Universidad Central deVenezuela, Caracas. Tese de Doutoramento.
ROHLF, F.J. & R.R. SOKAL. 1981. StatisticalTables. W.H. Freeman, San Francisco.
ROSS, G.N. 1963. Evidence for the lack ofterritoriality in two species of Hamadryas(Nymphalidae). J. Res. Lep. 2: 241-6.
SOKAL, R.R. & F.J. ROHLF. 1981. Biometry. W.H.Freeman, San Francisco.
SIEGEL, S.R. 1971. Estatística Não-Paramétrica.McGraw-Hill, São Paulo.
SOUTHWOOD, T.R.E. 1978. Ecological Methods,with Particular Reference to the Study of InsectPopulations. Chapman, London.
YOUNG, A.M. 1974. On the biology of Hamadryasfebrua (Lepidoptera: Nymphalidae) in
Guanacaste, Costa Rica. Zeitschr. Ang. Ent. 76:380-93.

63
Tabela I. Amostragem de indivíduos de H. feronia: (1) esforço de amostragem (minutos); (2) capturas e (3)recapturas por turno de amostragem (entre parêntesis, o número de indivíduos recapturados); (4) tempo de esperapara efetuar uma captura; (5) distância média percorrida pelos indivíduos recapturados.
11/MAR 12/MAR 13/MAR 14/MAR Total1.Tempo(min) 427 215 482 3152.Capturas 22 7 28 7 64a. manhã
macho 8 4 4 1fêmea 3 0 6 3? 4 3 - -
b. tardemacho 2 - 7 1fêmea 3 - 11 2? 2 - - -
3. Recapturas 10 (9) 12 (9) 38 (27) 22 (18) 82 (34)a. manhã
macho 5 5 15 7fêmea 2 3 5 8? 0 4 2 0
b. tardemacho 1 - 4 1fêmea 1 - 11 6? 1 - 1 0
4.1/2+3 (min) 12,20 11,32 7,30 10,865. Distância (m) 14,80 14,47 18,52 17,75
Tabela II. Estimativas do tamanho da população (Ni) de H. feronia(a).11/MAR 12/MAR(b) 13/MAR 14/MAR
Marcados (Mi) 0 8 18 17Não Marcados 22 7 28 7Soma (Ci) 22 15 46 24Ni - 39,11 71,74 79,17(a) usando-se o Índice de Lincoln-Petersen, com a correção de Bailey (veja Southwood, 1978), Ni= Ri(Ci + 1) / (Mi+ 1); onde Ni é a estimativa para o tamanho da população no dia i; Ri é o total de indivíduos marcados antes do diai; Ci é o total de indivíduos capturados no dia i; Mi é o número de indivíduos capturados no dia i que já estavammarcados; (b) capturas apenas pela manhã.

64
Tabela III. Capturas envolvendo machos e fêmeas de H. feronia, com teste-G para heterogeneidade sob a hipótesede que metade das capturas diárias aconteceram em cada turno de amostragem.a. Machos manhã tarde g.l. G11/MAR 13 3 1 6,738 P < 0,0113/MAR 19 11 1 2,159 P > 0,114/MAR 8 2 1 3,855 P < 0,05
40 16Total 3 12,752 P < 0,01Agrupado 1 10,626 P < 0,01Heterogeneidade 2 2,126 P > 0,3
b. Fêmeas manhã tarde11/MAR 5 4 1 0,111 P > 0,713/MAR 11 22 1 3,738 P > 0,114/MAR 11 8 1 0,476 P > 0,3
27 34Total 3 4,325 P > 0,2Agrupado 1 0,805 P > 0,3Heterogeneidade 2 3,520 P > 0,1

65
CONSIDERAÇÕES SOBRE A DISPERSÃOSECUNDÁRIA DE SEMENTES DE SOLANUMLYCOCARPUM (SOLANACEAE) PORFORMIGAS.
Flávia dos Santos Pinto
RESUMOEste trabalho sugere mais um caso de
benefício do grupo Attine sobre plantas do cerrado,indicando formigas do gênero Atta como possíveisdispersoras secundárias de Solanum lycocarpum. Nareserva Ecológica do IBGE (Brasília - D.F.) S.lycocarpum (Solanaceae) ocorre associada a ninhosde formigas do gênero Atta (sauveiros). Um censorealizado em cerrado sensu stricto mostrou que dos23 sauveiros amostrados, 65,2 % possuíam um oumais indivíduos de lobeira. Apenas 23% dosindivíduos de Solanum estavam sobre sauveiros, e44,6% estavam associados com outros grupos deformigas. Atta cf. goiana foi observada coletandosementes desta espécie localizadas nas fezes deChrysocyon brachyurus (Mammalia - Canidae), seuprincipal dispersor, assim como sementes no fruto.As sementes foram carregadas para dentro do ninho,o que poderia estar explicando a alta ocorrência destaespécie associada à sauveiros.
INTRODUÇÃOA dispersão de sementes e o estabelecimento
das plântulas representam os estágios mais sensíveisna história de vida de uma planta (Terbogh 1990);sendo essencial para essas duas fases a qualidade domicrohabitat onde estas sementes são depositadas(Hladik & Miquel 1990). O comportamento dosdispersores pode determinar o padrão de distribuiçãodessas sementes e, consequentemente, definir ascondições nas quais as plântulas se estabelecerão(Howe 1990; Howe & Smalwood 1982).
O consumo de frutos por vertebrados, queingerem ou não as sementes, tende a dispersá-las emgrupos, e em vários locais, levando a uma grandevariação de microambientes onde elas sãodepositadas (Hartgerink & Bazzard 1984). Estelocais, entretanto, podem ser propícios ou não para oseu estabelecimento.
As sementes descartadas nas fezes são, porvezes, utilizadas por formigas que atuam como pós-dispersores (Roberts & Heithaus 1986; Kaspari 1993;Levey & Byrne 1993; Loiselle 1990), causando umrearranjamento delas no ambiente. Esta relocaçãopermite maior variação de microambientes que,muitas vezes, representam sítios seguros para oestalecimento das plântulas (Culver & Beattie 1983;Heithaus 1981; Horvitz & Schemske 1986; Andersen1988; Levey & Byrne 1993).
A dispersão de sementes por formigas,denominada mirmecocoria, não é uma síndromecomum (van der Pijl 1969), tendo sido assumidacomo uma interação mutualística ainda poucodocumentada (Beattie 1985). A mirmecocoriageralmente está relacionada à presença de elaiosomasnas sementes, sendo escassos os estudos sobre adispersão de sementes de plantas não mirmecócoras,por formigas.
Estudos conduzidos por Gottsberger &Silbebauer-Gottsberger (1983) confirmam a baixaocorrência de mirmecocoria também para o cerrado.Das 271 espécies estudadas apenas 10% tinhamformigas como seu dispersor principal. Entretanto52% apresentaram dispersão zoocórica (realizada poranimais, principalmente vertebrados) mostrando aimportância da possível utilização das sementesdepositadas nas fezes. Essas relações entretanto sãopouco conhecidas para o cerrado.
Solanum lycocarpum St. Hil. (Solanaceae) éuma espécie arbustiva comum em ambientesperturbados (Sacco et al 1985; Oliveira-Filho &Oliveira 1988), estando entre as espécies dedistribuição mais ampla nos Cerrados (Sarmiento1983). Esta espécie possue frutos grandes, de cascagrossa e resistente, com grande quantidade desementes (de 600 a 800 segundo Lombardi & MottaJunior 1993). Quando maduros caem ao chão sementretanto romper a casca, estando sujeitos à ação depatógenos. É conhecida comumente como lobeira oufruta-de-lobo por ser amplamente utilizada na dietade Chrysocyon brachyurus Illiger (Canidae), o lobo-guará (Dietz 1984), que é o maior consumidor eprovavelmente seu principal dispersor, sendo comumencontrar grande quantidade de sementes nas fezes.
Na Reserva Ecológica do IBGE (Brasília -D.F.) é comum encontrar indivíduos de S.lycocarpum associados à ninhos de formigas dogênero Atta (sauveiros), tendo sido observado ocarregamento de sementes de fezes de lobo-guará poressas formigas (R. P. B. Henriques, comunicaçãopessoal).
O objetivo deste trabalho é determinar asrelações existentes entre S. lycocarpum e formigascomo possíveis pós-dispersores de suas sementes.
MATERIAL E MÉTODOSA coleta de dados foi realizada na Reserva
Ecológica do IBGE (RECOR) (15˚ 56’ 41’’ S e 47˚53’ 07’’ W) localizada no Distrito Federal à 35 kmao Sul de Brasília. A RECOR faz parte da Área deProteção Ambiental Gama-Cabeça do Veado e possuiuma área de 1350 hectares com a predominância decerrados e campos. O estudo foi conduzido noperíodo de 11 a 14 de março de 1996.
Foi realizado um censo de indivíduos de S.lycocarpum localizados ao longo de estrada de terra

66
(3km), que cerca uma área de cerrado sensu strictosujeito a queimada bienais.
Foram amostrados todos os indivíduos delobeira, bem como os sauveiros encontrados eregistrados a altura da planta, tamanho do sauveiro,ocorrência de associação de um com o outro, eespécies de formigas que possuíam formigueirolocalizado embaixo das plantas. Alguns exemplaresde cada morfotipo de formiga encontrado foramcoletados para posterior identificação.
Uma avaliação preliminar das espéciesvegetais existentes sobre os formigueiros foirealizada. Um gradeado com dez amostras de fezesfrescas de Chrysocyon brachyurus contendosementes de lobeira foi montado dentro de uma áreade cerrado sensu stricto, e cinco ao longo da estradaadjacente, sendo vistoriados a cada uma hora, para acaptura de formigas. Porções de fezes de lobo-guarácontendo sementes de lobeira também foramoferecidas a Atta cf. goiana, assim como polpa defruto maduro com sementes a fim de determinar apossibilidade de utilização das sementes nas duascondições.
RESULTADOSCenso de Solanum e Formigueiros
Foram registrados 65 indivíduos de lobeira e23 ninhos de saúvas nos 3km de trilha percorridos. S.lycocarpum apareceu 44,6% das vezes associado aformigas. Ocorrendo 23,1% das vezes sobresauveiros e 21,5% sobre ninhos de outras espécies deformiga não pertencentes ao gênero Atta (ApêndiceI), apenas 8,69% possuíam indícios de antigosformigueiros. Em 65,2% dos 23 sauveirosamostrados foram registrados um ou mais indivíduosde lobeira.
A altura das lobeiras variou de 30 cm até 4m, estando a maioria entre 1-2 m. Foram registradasde 1 a 5 plantas nos formigueiros, tendo sidoencontrada correlação positiva entre o número deplantas sobre o formigueiro e seu tamanho (r2 =0,82p<0,05).
O tamanho dos formigueiros variou desdepequenos buracos no chão até montes de terra de 5m. Não foi encontrada correlação entre o tamanhodestes formigueiros e o tamanho das plantas sobreeles.
Espécies de Plantas Encontradas SobreFormigueiros
Sete espécies de plantas foram encontradassobre os sauveiros, além da lobeira. A Tabela 1mostra a relação das espécies encontradas, suaporcentagem de ocorrência associado a sauveiros, eseu modo de dispersão. S. lycocarpum foi oito vezesmais frequente que a segunda espécie de maiorocorrência. Quatro das espécies apresentaram
dispersão endozoocórica realizada por pássaros e/oumamíferos, duas apresentaram dispersão pelo vento(anemocoria) e uma dispersão autocórica. Nenhumadelas apresentou formigas como seu dispersorprincipal segundo Gottsberger & Silbebauer-Gottsberger (1983).
Guilda de Formigas Associadas a FezesDuas subfamílias, quatro tribos, dez gêneros
e treze morfotipos de formigas foram observadosutilizando fezes de C. brachyurus durante as quatrohoras de observação (Apêndice II). Nenhuma espéciede Atta foi coletada durante as observações. Tambémnão foi observado deslocamento das sementesrealizado pelas formigas.
Espécies de Formigas Associadas a S. lycocarpumTrês espécies de formigas foram encontradas
nos formigueiros sob S. lycocarpum (Apêndice I):Atta cf. sexdens, Camponotus rufipes eMegalomyrmex sp; tendo sido encontradas utilizandofezes apenas as duas últimas.
Oferta de Frutos e Fezes a Atta goianaForam oferecidas, à Atta cf. goiana, no
campo, porções de frutos de lobeira e fezes de lobo-guará, ambas contendo sementes. As amostras foramcolocadas a aproximadamente um metro da entradado formigueiro. Nos dois casos A. cf. goiana foicapaz de cortar pedaços de fruto e da polpa fibrosadas fezes de C. brachyurus, e carregar as sementespara dentro do formigueiro. Tanto a polpa do fruto,sementes da polpa, massa das fezes quanto assemente das fezes foram utilizados.
DISCUSSÃOO censo realizado mostrou uma maior
ocorrência de S. lycocarpum sobre sauveiros emcomparação com outras plantas do cerrado, nessaárea de estudo. Acredita-se que esta relação ocorradevido ao carregamento das sementes de lobeira porestas formigas, entretanto não obtivemos dadosconclusivos a este respeito. Observações mostraramque saúvas carregam sementes de S. lycocarpumprovenientes de fezes de lobo-guará, o principaldispersor desta espécie e de frutos de Solanum para ointerior do ninho. Atta cf. goiana foi a espécieobservada carregando as sementes nestas duascondições. Outras espécies do cerrado foramobservadas associadas a sauveiros mas nenhuma comfrequência comparável à da lobeira. Quais seriam asvantagens para S. lycocarpum ter suas sementescarregadas para um sauveiro?
Nutrientes e Mudanças Físicas do SoloSegundo a hipótese do nutriente (Beattie
1985), o solo de ninhos de formigas é diferente

67
química e físicamente do solo à sua volta, podendodiferir em temperatura, porosidade, umidade, pH,conteúdo orgânico e mineral; representando assim,local propício para a germinação e estabelecimentode plântulas. Espécies de Atta e Acromyrmex sãoconhecidas como promovedoras de acúmulo dematéria orgânica e pelo aumento de nutrientes nosolo dos ninhos (Haines 1978; Alvaredo et al. 1981,Brener & Silva 1995), indicando sua importância emambientes savânicos, uma vez que savanas tropicaisestão geralmente associadas a solos pobres(Montgomery & Askewe 1983). Alguns trabalhostentaram relacionar esta riqueza no solo deformigueiros à facilidade de estabelecimento deespécies de plantas mirmecócoras (Westoby et al.1991;Culver & Beattie. 1980, 1983; Horvitz &Schemske 1986; Davidson & Morton 1981),entretanto dados conclusivos sobre esta hipóteseainda não foram obtidos.
Outra característica dos sauveiros é deprovocar mudança físicas além de químicas no solo,o que facilitaria o crescimento de estruturas vegetais,tendo sido observado um aumento na biomassa deraízes encontradas nos ninhos destas formigas emcomparação com o solo adjacente (Moutinho 1995;Nepstad et al. 1995; Brener and Silva 1995). Estasmudanças físicas e químicas poderiam estarinfluenciando a germinabilidade e o estabelecimentode S. lycocarpum sobre formigueiros. Trabalhosconduzidos por Lombardi & Motta Junior (1993)mostram um aumento na porcentagem de sementesgerminadas em solo retirado de áreas perturbadas, emcomparação com solos do cerrado, entretanto ascaracterísticas do solo destas áreas perturbadas nãosão mencionadas no artigo.
Fuga Contra Predação e PatógenosDurante o censo de formigueiros foi comum
encontrar sementes localizadas na entrada dosninhos, entretanto nenhuma semente de S.lycocarpum foi encontrada nesta condição. Sementesnesta condição desprotegida estariam maissuceptíveis ao ataque por predadores (O’Dowd &Hay 1980, Heithaus 1981), e por fungos patógenos(Augspurger 1990). Os frutos de S. lycocarpumquando não consumidos são utilizados por uma sériede insetos que facilitam a entrada de patógenos,sendo bastante comum encontrar frutoscompletamente atacados por fungos. Testes deviabilidade das sementes (teste de tetrazólio) defrutos coletados no chão em alto grau dedecomposição mostraram que apenas 42% dassementes se mostraram viáveis, enquanto quesementes retiradas de fezes frescas de C. brachyurusmostraram 82% de viabilidade. O mesmo testerealizado com frutos maduros recém coletadosmostrou aproximadamente 100% de viabilidade das
sementes (F. Borghetti comunicação pessoal),sugerindo que o ataque deste patógenos poderiamestar afetando a viabilidade das sementes.
As observações de saúvas carregandosementes de S. lycocarpum para o interior do ninho, aausência de sementes dessa planta na entrada dosninhos e o grande número de plantas sobre sauveirossugere que as sementes permaneçam protegidas epodem germinar dentro ou sobre o monte de terrasobre os sauveiros.
A ocorrência de sementes de espécies docerrado na abertura de formigueiros é bastante alta.Durante o período de realização deste trabalho foicomum encontrar várias sementes de Emmotun nitens(Icacinaceae) na abertura de formigueiros localizadospróximo à planta, assim como de outras espécies nãoidentificadas.
Vantagens da Dispersão Secundária para S.lycocarpum
A dispersão secundária de sementes de S.lycocarpum por formigas traz uma série de vantagensà espécie. Duas delas já foram citadas: a fuga contrapredadores e patógenos, e a deposição das sementesem ambiente favorável à germinação e aoestabelecimento das plântulas.
C. brachyurus, o principal dispersor de S.lycocarpum, deposita suas fezes preferencialmentesobre rochas, cupinzeiros e estradas ou trilhas(34,7%, 27,8% e 27,3%, respectivamente, segundoDietz 1984). Depositando, portanto, as sementesdescartadas nas fezes sobre substratos poucopropícios para a germinação. A dispersão secundáriapor formigas possibilitaria o deslocamento dessassementes para ambientes mais seguros, além depossibilitar o rearranjo das sementes, que sãodepositadas em grupos nas fezes.
Algumas desvantagens poderiam surgir coma associação de formigas com S. lycocarpum como,por exemplo, herbivoria. As formigas do gênero Attasão particularmente conhecidas como formigascortadeiras, representando uma ameaça aoestabeleciemento de plantas sobre formigueiros.Entretanto S. lycocarpum possue grande quantidadede pêlos nas folhas, mesmo nas mais jovens, além dapresença de alcalóides nas folhas (Ribeiro &Machado 1950), dificultando ou mesmo impedindosua utilização pelas formigas, o que poderia estarexplicando sua alta ocorrência sobre formigueiros.
Associações Benéficas Entre Attini e PlantasA existência de relações frouxas entre
formigas e sementes começa a ser investigada para ogrupo Attini. Oliveira et al. (1995) apresenta umexemplo de interação entre Micocepurus goeldii esementes de Hymenaea courbaril (Caesalpiniaceae),no sudeste do Brasil, como coadjuvantes na

68
sobrevivência das sementes encontradas em mata degaleria, onde estão sujeitas à infestação por fungosdependendo da quantidade de polpa deixada pelosdispersores primários. Os autores mostraram oprimeiro caso benéfico causado por este grupo comofacilitadora da germinação de sementes. Levantamtambém a hipótese de que outras espécies do grupoAttini promovam benefícios similares a outrasplantas.
Os resultados mostrados neste trabalhosugerem uma relação benéfica entre Atta cf. goiana, eprovavelmente outras espécies do grupo Attine, sobreo estabelecimento e possivelmente sobre agerminabilidade de S. lycocarpum.
Seleção de Substratos Pelo Gênero AttaNovas considerações surgiram com este
trabalho sobre a seleção de substratos para ocrescimento de fungos para o gênero Atta que éconsiderado como utilizador exclusivo de flores efolhas (Holldobler & Wilson 1990). Roberts &Heithaus (1986) mostram a utilização de frutos porAtta sexdens. Este trabalho é o primeiro a mostrar autilização de fezes por Atta sp e confirma a utilizaçãode frutos para este gênero.
AGRADECIMENTOSEste trabalho foi realizado durante o curso
“Métodos em Ecologia de Campo” do Curso de Pós-graduação em Ecologia da Universidade de Brasília.Meu muito obrigado ao Prof. Raimundo P. B.Henriques pela organização do curso, e pela idéiainicial que resultou neste trabalho. Ao Prof. Paulo S.Oliveira pelo esclarecimento quanto à metodologia eestímulo. À Profa. Helena C. de Moraes pelas aulassobre formigas e críticas ao manuscrito. Ao Prof.Fabian Borghetti pelas aulas sobre sementes e apoioaos meus “excessos” de criatividade. À Inara R. Lealpela identificação de Atta cf. goiana. Ao Carlos A.Bianchi pela ajuda no campo e sugestõesmetodológicas. Ao Delano M. S. Silva e Fernanda P.Pinheiro pelas críticas ao manuscrito.
BIBLIOGRAFIAALVAREDO, A., C. W. BERISH, & F. PERALTA.
1981. Leaf-cut ant (Atta cephalotes) influence onthe morphology of andepts in Costa Rica. SoilSci. Soc. Am. J. 45: 790-794.
ANDERSEN, A. N. 1988. Dispersal distance as abenefit of myrmecochory. Oecologia 75: 507-511.
AUGSPURGER, C. K. 1990. The potencial impactof fungal pathogens on tropical plantreproductive biology. In: Reproductive ecologyof tropical forest plants. Bawa, K.S. and Hadley,M. (Eds.). Man and the biosphere series
(UNESCO), vol. 7; 237-245pp. ParthenonPublishing, NewJersey.
BEATTIE, A. J. 1985. The evolutionary ecology ofant-plant mutualisms. Cambridge UniversityPress, Cambridge. 182pp.
BRENER, A. G. F. & J. F. SILVA. 1995. Leaf-cutting ant nest and soil fertility in a well-drainedsavanna in western Venezuela. Biotropica 27(2):250-253.
CULVER, D. C. & A. J. BEATTIE. 1980. The fateof Viola seeds dispersed by ants. Am. J. Bot. 67:710-714.
CULVER, D. C. & A. J. BEATTIE. 1983. Effects ofant mounds on soil chemistry and vegetationpatterns in a Colorado montane meadow.Ecology 64(3): 485-492.
DAVIDSON, D.W. & S. R MORTON. 1981.Myrmecochory in some plants (F.chenopodiaceae) of the autralian arid zone.Oecologia 50: 357-366.
DIETZ, J. M. 1984. Ecology and social organizationof the maned wolf (Chrysocyon brachyurus).Smithsonian Contribution of Zoology. n° 392.51pp.
GOTTSBERGER, G. & I. SILBERBAUER-GOTTSBERGER. 1983. Dispersal anddistribution in cerrado vegetation of Brazil,. In:Dispersal and distribution. K. Kubizki (Ed.).315-351pp. Parey, Hamburg.
HAINES, B. L. 1978. Element and energy flowsthrough colonies of the leaf-cutting ant, Attacolombica, in Panama. Biotropica 10: 270-277.
HARTGERINK, A. P. & F. A. BAZZAZ. 1984.Seedling-scale environmental heterogeneityinfluences individual fitness and populationstructure. Ecology 65:198-206.
HEITHAUS, E. R. 1981. Seed predation by rodentson three ant-dispersed plants. Ecology 62(1):136-145.
HLADIK, A. & S. MIQUEL. 1990. Seedling typesand plant establishment in a african rain forest.In: Reproductive ecology of tropical forestplants. Bawa, K. S. & Hadley, M. (Eds.). Manand the biosphere series (UNESCO), vol. 7; 261-282pp. Parthenon Publishing, New Jersey.
HÖLLDOBLER, B. & E. O. WILSON. 1990. Theants. Harvard University Press, Cambridge,Massachusetts.
HORVITZ, C. C. & D. W SCHEMSKE. 1986. Ant-nest soil and seedling growth in a neotropicalant-disperser herb. Oecologia 70: 318-320.
HOWE, H. F. & J. SMALLWOOD. 1982. Ecologyof seed dispersal. Ann. Rev. Ecol. Syst. 13: 201-228.
HOWE, H. F. 1990. Seed dispersal by birds andmammals implications for seedling demography.In: Reproductive ecology of tropical forest

69
plants. Bawa, K.S. and Hadley, M. (Eds.). Manand the biosphere series (UNESCO), vol. 7; 191-218pp. Parthenon Publishing, New Jersey.
KASPARI, M. 1993. Removal of seeds fromneotropical frugivore droppings. Oecologia 95:81-88.
LEVEY, D. J. & M. M. BYRNE. 1993. Complexant-plant interactions: rain forest ants assecondary dispersers and post-dispersal seedpredators. Ecology 74(6): 1802-1812.
LOISELLE, B. A. 1990. Seeds in droppings oftropical fruit-eating birds: importance ofconsidering seed composition. Oecologia 82:494-500.
LOMBARDI, J. A. & J. C. MOTTA JUNIOR. 1993.Seed dispersal of Solanum lycocarpum St. Hil.(Solanaceae) by the maned wolf, Chrysocyonbrachyurus Illiger (Mammalia, Canidae).Ciência e Cultura 45(2): 126-127.
MONTGOMERY, R. F. & G. P. ASKEWE. 1983.Soils of tropical savannas. In: Ecosystems of theworld 13: Tropical savannas. Bourliere, F. (Ed.).Elsevier Scientific Publishing Company. NewYork.
MOUTINHO, P. 1995. Acabar com a saúva, masnem tanto. Ciência Hoje 18(106): 10-11.
NEPSTAD, D. C., P. JIPP, P. MOUTINHO, G.NEGREIROS & S. VIEIRA. 1995. Forestrecovery following pasture abandonment inAmazonia: canopy seasonality, fire resistenceand ants. In: Evaluating and monitoring thehealth of large-scale ecosystems. D. J. Rapport,C. L. Gaudet & P. Calow. (Eds.) 333-349 pp.NATO ASI Series vol. 128.
O’DOWD, D. J. & M. E. HAY. 1980. Mutualismbetween harvester ants and desert ephemeral:seed escape from rodents. Ecology 61(3): 531-540.
OLIVEIRA, P. S., M. GALETTI, F. PEDRONI & L.P. C MORELLATO. 1995. Seed cleaning byMycocepurus goeldii ants (Attini) facilitesgermination in Hymenaea courbail(Caesalpiniaceae). Biotropica 27(2): 518-522.
OLIVEIRA-FILHO, A. T. & L. C. A. OLIVEIRA.1988. Biologia floral de uma população deSolanum lycocarpum St. Hil. (Solanaceae) emLavras. Revista brasileira de Botânica. 11: 23-32.
RIBEIRO, O. & MACHADO, A., 1950. O alcalóideda fruta de lobo. Boletim do Instituto de QuímicaAgrícola do Rio de Janeiro 16: 11-13.
ROBERTS, J. T. & E. R HEITHAUS. 1986. Antsrearrange the vertebrate-generated seed shadowof a neotropical fig tree. Ecology 67(4): 1046-1051.
SACCO, J. C. ; E. SANTOS; E. FROMM-TRINTA,N. L. M. COSTA & M. C. S. CUNHA. 1985.
Ervas daninhas do Brasil - Solanaceae- gêneroSolanum L. Documentos EMBRAPA - CNPDA,Brasília.
SARMIENTO, G. 1983. The savannas of tropicalamerica. In: Ecosystems of the world 13:Tropical savannas. Bourliere, F. (Ed.). ElsevierScientific Publishing Company. New York.
TERBORGH, J. 1990. Seed and fruit dispersal -Commentary. In: Reproductive ecology oftropical forest plants. Bawa, K. S. & Hadley, M.(Eds.). Man and the biosphere series(UNESCO), vol. 7: 181-190. ParthenonPublishing, New Jersey. 421pp.
VAN DER PIJL, L. 1969. Principle of dispersal inhigher plants. Spring-Verlag, Berlin, Germany.
WESTOBY, M., L HUGHES & B.L RICE. 1991Seed dispersal by ants: comparing infertile whitfertile soils. In: Ant-plant Interactions. Huxley,C.R. & Cutler, D.F. (Eds). Oxford UniversityPress, Oxford. 601pp.
Tabela 1. Lista de espécies vegetais encontradassobre sauveiros, suas frequências de ocorrência em23 sauveiros.
espécie %
Calliandra dysantha 4Stryphnodendron adstringens 4Erythroxylum tortuosum 4Qualea grandiflora 4Vochysia sp. 8Styrax ferrugineus 4Rapanea guianense 8Solanum lycocarpum 65
APÊNDICE I. Lista de espécies de formigas cujosninhos foram encontrados sob Solanum lycocarpum.SUBFAMÍLIA FORMICINAETribo Camponotini
Camponotus rufipesSUBFAMÍLIA MYRMICINAE
Megalomyrmex sp 1.Tribo Attini
Atta sexdens
APÊNDICE II. Lista de formigas encontradasutilizando fezes de C. brachyurus.SUBFAMÍLIA FORMICINAETribo Camponotini
Camponotus rufipesCamponotus sp.1Camponotus sp.2
SUBFAMÍLIA MYRMICINAE Allomerus sp.Megalomyrmex sp.

70
Tribo CephalotiniCephalotes attratus
Tribo AttiniAcromyrmex sp.Trachymyrmex sp.
Tribo CrematogastriniCrematogaster sp.
Tribo MyrmiciniPheidole sp.Pheidole sp. 1.Pheidole sp. 2.

71
PADRÃO DE ATIVIDADE DO MICO-ESTRELA (CALLITHRIX PENICILLATA) NOCERRADO DENSO E CERRADÃO DARESERVA ECOLÓGICA DO IBGE, BRASÍLIA,DF
Guilherme H. B. de Miranda
RESUMOA família Callitrichidae de primatas
Platyrrhini possui adaptações (porte reduzido,dentição especializada, presença de garras, etc.)ligadas à ocupação pioneira de nichos, ocorrendo emquase todos os hábitats arbóreos neotropicais. Oúnico primata encontrado no cerrado (excluindo-se asmatas ciliares) é o Callithrix penicillata (mico-estrela). O objetivo deste trabalho foi comparar asatividades desempenhadas por esta espécie singularem diferentes feições de cerrado. Durante quatrodias, na Reserva Ecológica do IBGE foram realizadasobservações (ecologia e comportamento) de doisgrupos naturais de C. penicillata (um grupocomposto por oito indivíduos habitando uma área decerrado denso e o outro, com quatro membros,ocupando uma mancha de cerradão). Foram feitosregistros individuais por varredura a cada cincominutos e registros oportunísticos sempre quepossível. No cerrado denso, foi usado como alimentopelos sagüis o exsudato proveniente de quatroespécies: Vochysia thyrsoidea, Didymopanaxmacrocarpum, Qualea grandiflora e Qualeaparviflora. No cerradão, foi observado apenas o usoda goma de V. thyrsoidea. Em ambos os ambientes, amaior parte da atividade registrada foi locomoção(57% para o cerrado denso e 50% no cerradão). Osanimais passaram a maior parte do tempo no estratomédio (altura entre 2 e 5 metros). A atividade de C.penicillata não diferiu muito nos dois hábitats, excetopela maior porcentagem de locomoção na área decerrado denso.
INTRODUÇÃOOs calitriquídeos dos gêneros Callithrix e
Cebuella possuem a dentição adaptada para aexploração alimentar de exsudatos vegetais (seiva egoma). Esses dois gêneros de calitriquídeos são osúnicos primatas que danificam ativamente árvoresgumíferas a fim de estimular a liberação dosexsudatos (Lacher et al, 1984).
São encontrados em quase todos os hábitatsarbóreos neotropicais. O gênero Callithrix é o únicoencontrado tanto na Mata Atlântica quanto naAmazônia, bem como no cerrado e caatinga, queseparam os dois grandes biomas florestais brasileiros.
A comunidade de primatas das matas docerrado do Planalto Central é constituída por três
espécies: Callithrix penicillata, Cebus apella eAlouatta caraya. Destas espécies, somente C.penicillata tem sua presença registrada em formasfisionômicas mais abertas do cerrado (cerrado s.s. ecerradão) o que foi atribuído a seu alto potencialadaptativo resultante da gumivoria (Fonseca eLacher, 1984). O cerradão é a forma fisionômicamais alta de cerrado, fisionomicamente florestal, comdossel fechado de 7 m ou mais de altura e 30% oumais de cobertura, ou arvoredo, com dossel aberto damesma altura (Eiten, 1993). Na região dos cerrados,o cerradão está presente em manchas com solosmesotróficos, podendo ocorrer também em solosdistróficos caso seu estabelecimento tenha ocorridoanteriormente ao empobrecimento do solo(Haridasan, 1992) ou como sucessão arbórea (poradensamento) a um cerrado sensu stricto sujeito alongos períodos (vários anos) sem fogo.
Callithrix penicillata é uma espécie dehábitos diurnos, que vive em grupos familiares de 5 a15 membros, insetívoro/gumívoro, com alto índice desucesso ecológico, sendo considerado umectoparasita de árvores (Lacher et al, 1984). A dietade exsudatos, de baixo valor nutritivo, mas muitoabundantes, permite suprir a escassez local e/ousazonal de recursos de maior qualidade nutritiva(insetos, outras presas animais e frutos).
MATERIAL E MÉTODOSO presente estudo foi conduzido, entre os
dias 11 e 14/03/96, na Reserva Ecológica do IBGE -RECOR (15˚56’41” S e 047˚53’07” W), que contacom cerca de 1350 ha de área, situada no km 0 daBR-251, que liga Brasília a Unaí, 35 km ao sul docentro da capital federal. O relevo local é suave e aaltitude média é de 1100 metros. As observações decampo restringiram-se a duas áreas: uma mancha decerradão distrófico (cerca de 10 ha) que ocorre noextremo sudeste da reserva e parte ( 3 ha) de umtrecho de cerrado denso (protegido do fogo por maisde 22 anos) ao sul das construções da sede da reserva(Figura 1). Durante todo o período iluminado de cadaum dos quatro dias de observação, um grupo de C.penicillata foi acompanhado em uma das áreas, desdeo momento em que foi avistado (a partir das 6 hs. e22 min.) até o fim das atividades diárias (por volta de18 hs.), sendo determinados o tamanho do grupo e onúmero de membros em cada faixa etária (adulto,juvenil e infantil). Foram também feitos registroscomportamentais por varredura de todos osindivíduos do grupo, a cada 5 minutos e registrosoportunísticos sempre que possível (Altmann, 1974).As categorias comportamentais consideradas nosregistros de varredura foram: locomoção(deslocamento individual vertical ou horizontal),descanso (qualquer posição em que o indivíduopermaneça sem deslocamento), escarificação

72
(produzir ou ampliar ferida na casca/câmbio deárvore gumífera, com o uso dos dentes, lamber eingerir o exsudato resultante) e alimentação (ingestãode outros itens da dieta, como insetos, frutos, folhas,etc.). A altura do animal observado em relação aosolo foi estimada em um de três níveis pré-definidos(baixo, até dois metros de altura; médio, de 2 a 5metros e alto, mais de 5 metros de altura). Destaforma, foram feitos 90 registros de varredura do nívelde altura dos micos-estrela. Os itens alimentaresconsumidos foram anotados.
Considerando em conjunto os quatro dias deobservação e ambos os grupos observados, osanimais foram vistos durante 9 horas e 8 minutos,totalizando 27% do tempo no campo (548/2029minutos). O grupo do cerrado denso foiacompanhado por mais tempo (6 h e 53 min),correspondendo a 28% (413/1474 minutos) doesforço de observação para este grupo. Já o grupo docerradão foi observado por apenas 2 h e 14 min, ouseja, cerca de 24,3% do tempo de observação dogrupo (134/555 minutos).
Como os esforços de observação em cadalocal foram bastante distintos (totalizando 1474minutos para a área de cerrado denso e apenas 555minutos para a área de cerradão), a fim de permitiruma comparação equilibrada entre os resultadosobtidos, os registros do cerradão foram multiplicadospor um fator de equiparação de esforço amostral (f),que consiste na razão entre os tempos de observaçãoem cada área de estudo (f= 1474/555= 2.656).
RESULTADOSNo cerrado denso, o grupo de C. penicillata
observado contava com oito indivíduos: cincoadultos, dois juvenis e um infantil. Um dos machosadultos possuía apenas metade da cauda.
No cerradão, apenas quatro indivíduoscompunham o grupo: um casal adulto, um filhotejuvenil macho e um infantil de sexo indeterminado.
Os sagüis utilizaram exsudatos de pelomenos quatro espécies arbóreas de duas famíliasdurante este estudo (Tabela 1). Na área de cerradodenso, Didymopanax macrocarpum e Vochysiathyrsoidea foram as espécies observadas com sinaisde escarificação dos troncos e ramos, ao passo que,no cerradão, a escarificação e o consumo de gomaforam vistos apenas em V. thyrsoidea, espéciebastante abundante nesta forma fisionômica.
Mesmo considerando a ponderação dosvalores obtidos para o cerradão de acordo com ofator de equiparação de esforço amostral, nas quatrocategorias comportamentais observadas, o número deregistros do grupo do cerrado denso foi maior que odo grupo do cerradão (Fig. 2).
Para o grupo do cerrado denso, o número deregistros de varredura obtidos na categoria
locomoção correspondeu a 57% (193 registros) dototal. No caso do cerradão, exatamente 50% (82) dosregistros foram locomoção, ou seja, de uma maneirageral, em ambos os grupos, os sagüis locomoviam-sea maior parte do tempo observado. É possível notarainda que a participação da categoria descanso nogrupo que habita o fragmento de cerradão (66registros) foi maior (40% das atividades do grupo)em relação ao grupo do cerrado denso (33% paradescanso).
Devido ao pequeno número obtido deregistros de altura, estes foram analisados emconjunto, independentemente do hábitat. Na maiorparte das vezes (60,0%), os animais foram avistadosno nível médio, ou seja, numa altura entre 2 e 5metros. O segundo nível mais freqüentado foi o nívelalto (altura igual ou superior a 5 metros),correspondendo a 26,7% dos registros obtidos. Porúltimo, o nível baixo (menos de 2 metros de altura)com apenas 13,3% dos registros. Esta tendência deocupação espacial ocorreu nas diversas categoriascomportamentais observadas. (Tabela 2).
DISCUSSÃOFonseca e Lacher (1984) indicaram para C.
penicillata no cerradão, que mais de 70% do tempototal gasto com alimentação relacionou-se comconsumo de exsudato. No presente trabalho, o valorencontrado foi 83% para o cerradão e 78% para ocerrado denso.
O grupo que ocupa a área de cerradão pareceser recente, o que é evidenciado pelo seu pequenotamanho (somente quatro indivíduos) e pelacomposição (com apenas dois indivíduos adultos).Esta possibilidade se encaixa com o fato do local tersido submetido à grande queimada do final de 1994.O menor tamanho deste grupo explica a menorquantidade de registros de varredura obtida mesmocom a utilização do fator de equiparação. Uma explicação para o maior deslocamentodo grupo do cerrado denso em relação aos animais docerradão é uma menor concentração espacial dosrecursos alimentares (goma/insetos) no primeirohábitat em relação ao segundo. De acordo com a literatura (Queiroz, 1991;Ferrari, 1995), C. penicillata é um primata que ocupapreferencialmente o extrato inferior da vegetação,onde há mais fontes alimentares (insetos, pequenosfrutos e árvores gumíferas) e onde estão maisprotegidos dos seus predadores mais comuns (aves derapina). Todavia, os resultados do presente estudonão estão de acordo com os estudos prévios. Aausência de outras espécies de primatas no local, oque permitiria a ocupação de um nicho espacial maisamplo; a não-habituação dos animais com a presençado observador no estrato inferior, o que os levaria aum “recuo” para os níveis mais altos e o menor porte

73
e maior espaçamento do estrato arbóreo do cerradoem relação ao das matas mesofíticas seriam osprincipais fatores responsáveis por esse resultadoaparentemente contraditório.
Foram feitos alguns registros oportunísticosde deslocamento ao nível do solo, especialmentequando o grupo passava por áreas mais abertas(tendendo a cerrado s. s.) ou cruzava uma estrada.Devido ao alto risco de exposição à predação (e àpresença do observador), estes deslocamentos foramfeitos com extrema atenção e rapidez, em geral, apósalguma hesitação.
Possivelmente, existe uma certaconvergência fisionômica entre o cerradão e ocerrado s.s. estudados, como conseqüência doimpacto do fogo (relativamente freqüente no primeirohábitat e ausente no segundo, nos últimos vinte e doisanos). De maneira que, atualmente, localmente, ocerrado apresenta um estrato arbóreo mais fechadoque o cerradão.
Em síntese, apesar de haver distinção detamanho e composição dos grupos estudados, nãoforam constatadas grandes diferenças na atividadecomportamental desses grupos que pudessem seratribuídas ao tamanho do grupo ou hábitat.
AGRADECIMENTOSO autor gostaria de agradecer ao Instituto
Brasileiro de Geografia e Estatística (IBGE) pelapermissão para usar a reserva e suas instalações. AoRaimundo P. B. Henriques e à Dóris S. de Faria,pelas sugestões e esclarecimentos prestados emdiferentes etapas do trabalho. À Regina Macedo, pelaleitura crítica de uma versão preliminar do artigo. AoDepartamento de Ecologia da Universidade deBrasília pelo apoio logístico. À CAPES pelo apoiofinanceiro, através da bolsa de mestrado.
BIBLIOGRAFIAALTMANN, J. 1974. Observational study of
behavior: sampling methods. Behavior (49):226-267.
EITEN, G. 1993. Vegetação. In: M. N. Pinto (Org.).Cerrado: caracterização, ocupação eperspectivas, 2a Edição, pp. 17-73. EditoraUniversidade de Brasília, Brasília.
FERRARI, S. F. 1995. Ecologia de primatasneotropicais: comunidades e estratégias deforrageio. Apostila do mini-curso ministradodurante o VII Congresso da Sociedade Brasileirade Primatologia. Natal, 23 a 27 de julho de 1995.
FONSECA, G. A. B. DA AND T. E. LACHER, JR.Exudate-feeding by Callithrix jacchuspenicillata in semideciduous woodland(Cerradão) in Central Brazil. Primates25(4):441-449.
HARIDASAN, M. 1992. Observations on soils, foliarnutrient concentrations and floristic compositionof cerrado sensu stricto and cerradãocommunities in central Brazil. In: P. A. Furley,J. Proctor and J. A. Ratter (Eds.). Nature andDynamics of Forest-Savanna Boundaries, pp.171-184. Chapman-Hall, London.
LACHER, T. E. , JR., G. A. B. DA FONSECA, C.ALVES, JR. AND B. MAGALHÃES-CASTRO.1984. Parasitism of trees by marmosets in acentral brazilian gallery forest.Biotropica16(3):202-209.
QUEIROZ, H. L. DE. 1991. Levantamento daspopulações de três espécies de primatas naFazenda Sucupira, Brasília, D.F. In: A. B.Rylands & A. T. Bernardes (Eds.), APrimatologia no Brasil - 3. FundaçãoBiodiversitas, Belo Horizonte, pp.369-374.

74
TABELA 1. Espécies arbóreas escarificadas por C. penicillata em duas fitofisionomias de cerrado da ReservaEcológica do IBGEcerrado densoEspécie (Família)
cerradãoEspécie (Família)
Vochysia thyrsoidea (Vochysiacea) Vochysia thyrsoidea (Vochysiacea)Didymopanax macrocarpum (Araliacea)Qualea grandiflora (Vochysiacea)Qualea parviflora (Vochysiacea)
TABELA 2. Distribuição vertical do mico-estrela, C. penicillata, no cerrado (cerradão e cerrado denso juntos) daReserva Ecológica do IBGENível Descanso Locomoção Escarificação Totalbaixo (menos de 2m de altura) 8(8,9%) 4(4,4%) 12(13,3%)médio (entre 2 e 5m de altura) 20(22,2%) 31(34,4%) 3(3,3%) 54(60,0%)alto (mais de 5m de altura) 12(13,3%) 12(13,3%) 24(26,7%)Total 40(44,4%) 47(52,2%) 3(3,3%) 90(100,0%)
Reserva Ecológica do IBGE
1 km
DrenagensLimitesEstradas
Sede da Reserva
Cerrado DensoCerradão
Fig. 1. Mapa da Reserva Ecológica do IBGE, mostrando as áreas de estudo utilizadas.
0
20
40
60
80
100
120
140
160
180
200
locomoção descanso escarificação alimentação
Categoria comportamental
cerrado denso
cerradão (equiparado)
Fig. 2. Partição da atividade de C. penicillata na Reserva Ecológica do IBGE, de acordo com os registroscomportamentais obtidos pelo método de varredura.

75
ARQUITETURA E RIQUEZA DEHERBÍVOROS EM Ouratea hexasperma Baill.(OCHNACEAE), EM UM CERRADO DARESERVA ECOLÓGICA DO IBGE, BRASÍLIA,DF
Rosane G. Collevatti
RESUMOFoi testada a hipótese de que indivíduos de
Ouratea hexasperma com arquitetura mais complexatêm maior riqueza de espécies de herbívoros, em umaregião de cerrado, na reserva do IBGE, Brasília, DF.Foi investigado, também, se a herbivoria estárelacionada à arquitetura, dureza e área foliar; como aárea foliar está relacionada ao comprimento danervura principal e largura da folha; se a dureza foliaraumenta com a área foliar; se os elementos dearquitetura (diâmetro no eixo maior e menor da copa,altura da planta e da copa, número de folhas e demódulos e volume de copa) estão correlacionados.Foram encontrados 29 morfoespécies de herbívoros.A área foliar foi explicada pelo comprimento danervura e largura da folha (89,4%) e a dureza foliartendeu a diminuir com o aumento da área foliar. Oselementos da arquitetura foram correlacionados entresi, sendo que o número de folhas e de módulos forammais correlacionados com o diâmetro. Plantasmenores tiveram tendência a ter menor herbivoria, oque pode estar correlacionado à menor dureza epalatabilidade. Plantas com menor quantidade defolhas e módulos apresentaram maior número deespécies. Porém, o número de folhas foi relacionadopositivamente para os grupos taxonômicos,separadamente. Os resultados indicam que a riquezade espécies de herbívoros em O. hexasperma, éafetada pela diversidade de recursos, expressa pela“densidade de folhas” na copa.
INTRODUÇÃOA riqueza de espécies de herbívoros em um
determinado hospedeiro pode ser explicada pordiversos fatores. A nível regional, plantas comdistribuição geográfica mais ampla tendem a termaior diversidade de herbívoros que plantas comdistribuição mais restrita, uma relação existente emespécies taxonomicamente distintas, comoZingiberales, Rosaceae, e outras (Lawton & Schröder1977, Strong 1977, Strong et al. 1977, Strong &Levin 1979, Leather 1986). A nível local, a riquezade espécies de herbívoros pode estar ligada àabundância local, ao tempo de colonização, aoisolamento taxonômico e à complexidade estruturaldo hospedeiro (Strong et al. 1977, Strong & Levin1979, Neuvonen & Niemelä 1981, Southwood et al
1982, Lawton 1983, Leather 1986, Lewinsohn 1991,Marquis 1991).
Em um gradiente de aumento decomplexidade estrutural desde plantas herbáceas atéárvores, espera-se encontrar um aumento dediversidade de herbívoros no sentido ervas, arbustose árvores. Da mesma forma, dentro de uma mesmaespécie, haveria uma tendência a um aumento dadiversidade de herbívoros com o aumento dacomplexidade estrutural da planta, ao longo do seudesenvolvimento (plântula, jovem e adulto) e demodificações sazonais (Strong & Levin 1979,Lawton 1983). A arquitetura da planta estárelacionada aos componentes: (1) tamanho da planta;(2) forma de crescimento; (3) desenvolvimentosazonal (mudanças sazonais no tamanho e forma decrescimento); (4) variedade e longevidade demódulos (Lawton & Schröder 1977, Harper 1981,Lawton 1983), que descrevem aspetos gerais dotamanho da planta, forma de crescimento ecomplexidade estrutural.
Basicamente duas hipóteses são utilizadaspara explicar essa relação na qual o número deespécies aumenta com o aumento da complexidadeestrutural: (1) área per se, na qual plantas maioresseriam mais “aparentes” (sensu Feeny 1976) eportanto seriam mais facilmente “descobertas” porherbívoros, além de suportarem populações maioresque teriam taxas de extinção menor; (2) diversidadede recursos, na qual plantas mais complexas teriamuma maior diversidade de microhabitats (alimento,local para nidificar, espaço livre de inimigos naturais)e possibilidade de estratificação vertical ou horizontalna ocupação hospedeiro, dando oportunidade para acolonização por uma maior variedade de espécies,com diferentes necessidades de recursos (Lawton &Schöder 1977, Strong & Levin 1979, Neuvonen &Niemelä 1981).
Essa tendência tem sido verificada,principalmente em plantas de região temperada,havendo uma carência de trabalhos que comprovemessa relação em regiões tropicais, principalmente noCerrado (mas veja Cytrynowicz 1991). Esse trabalhoteve como objetivo testar a hipótese de queindivíduos de Ouratea hexasperma com arquiteturamais complexa têm maior riqueza de espécies deherbívoros e relacionar herbivoria com arquitetura,dureza e área foliar. Além disso, esse trabalho tevecomo objetivos secundários verificar: (1) como a áreafoliar pode ser explicada pelas variáveiscomprimento da nervura principal e largura da folha;(2) se folhas maiores têm maior dureza; (3) se oselementos da arquitetura estão correlacionados entresi.
MATERIAL E MÉTODOS

76
Ouratea hexasperma (St.Hil.) Baill.(Ochnaceae) é uma espécie arbustiva, muito comumnos cerrados do Brasil Central. Sua arquitetura ébastante variável, podendo atingir mais de 2 m dealtura (Ratter 1986), apresentando rebrotasaparentemente induzidas após queimadas. A planta éperenifólia, apresentando folhas velhas e novas o anotodo, não ocorrendo queda acentuada e sincrônica defolhas (Morais et al. 1995). Apresenta nectáriosextraflorais nas estípulas e base do pedicelo do botãofloral, visitados principalmente por Camponotuscrassus e Zacryptoceus pusillus (Oliveira & Oliveira-Filho 1991, Oliveira et al. 1995).
O trabalho foi desenvolvido entre os dias 10e 14 de março de 1996, na Reserva Ecológica doIBGE (15˚55’58’’S, 47˚51’02’’W), situada na Áreade Preservação Ambiental (APA) Cabeça de Veado,Brasília, DF, em uma área de cerrado sensu stricto(Eiten 1990) que é submetida a queimada anual.
Foi delimitada uma parcela de 2500 m2
(50x50) onde foram registrados 93 indivíduos de O.hexasperma. Desses, 50 foram sorteados e marcadospara o estudo.
Foram medidos os seguintes componentesda arquitetura: diâmeto da copa no maior eixo(DMA) e no menor eixo (DME) perpendicular à“DMA”; altura da planta (APL); altura da copa(ACP); número total de folhas (NFL) e número totalde módulos (NMD). O volume da copa foi estimadosegundo um semi-elipsóide, ondeVCP=(π*DMA*DME*ACP)/6.
Além disso, foram coletadas folhas, aoacaso, para medir a dureza, área foliar, área foliardanificada, comprimento da nervura principal elargura maior da folha. O número de folhas coletadadependeu do número total de folhas do indivíduo (3folhas, para indivíduos com até 60 folhas; 6 folhaspara indíviduos com até 200 folhas; 9 folhas paraindivíduos com mais de 200 folhas). A dureza foimedida em folhas frescas com um penetrômetro, quemede a força necessária (g*m-2) para perfurar umcírculo de 0.5mm de diâmetro (Coley 1983). A áreafoliar foi determinada utilizando uma transparênciana qual foram desenhados quadrados de 1 cm2 deárea. A herbivoria foi calculada como % área foliardanificada/área total da folha. Foi estimada, também,a área foliar total da planta (área foliarmédia*número total de folhas) e densidade foliar(área foliar total da planta/DMA).
Em todas as plantas foram coletados ouregistrados, durante quatro dias, todos os herbívorosavistados nas plantas. As plantas foram observadasdurante o período da manhã e tarde, sequencialmente,permanecendo no máximo 5 minutos em cada planta.
Para relacionar a área foliar ao comprimentoda nervura e largura da folha foi realizada uma
regressão linear, segundo o modelo Y=a+bx1+cx2,onde Y é a área foliar, x1 é o comprimento danervura principal e x2 é a largura maior da folha.
A relação entre dureza foliar (DFL) e áreafoliar (AFL) foi analisada, também, com umaregressão linear.
Para verificar a existência de correlacãoentre os elementos de arquitetura, foi utilizado ocoeficiente de correlação de Pearson.
As variáveis herbivoria e número deespécies foram transformadas em arcoseno elogarítmo, respectivamente (Zar 1974). A relaçãoentre herbivoria e riqueza total de espécies e porgrupos taxônomicos (ordem), com os elementosarquitetônicos, dureza foliar e área foliar foianalisada por regressão múltipla por passos (Stepwisemultiple regression).
As análises estatísticas foram feitas com osoftware Statistix 4.1.
RESULTADOSForam encontradas 29 morfoespécies de
herbívoros (Tabela 1), sendo Homoptera, Orthopterae Lepidoptera os grupos com maior número demorfoespécies. A nervura principal e largura da folhaexplicaram 89,36% da variação na área foliar(r2=0,8936, F=79,77, p<0,001, N=248, Y=-50,024+2,845x1+12,5286x2, onde “x1” é ocomprimento da nervura principal e “x2” a larguramaior da folha).
A dureza foliar tendeu a aumentar com oaumento da área foliar, embora a porcentagem devariação explicada por essa variável tenha sido muitobaixa (r2=0,069, F=17,638, p<0,001, N=239,Y=5,202+0,0031*AFL).
Os elementos de arquitetura forampositivamente correlacionados entre si (Tabela 2). Onúmero de folhas (NFL) foi mais correlacionado como número de módulos (NMD), diâmetro no menoreixo (DME) e volume da copa (VCP). Uma baixaporcentagem de variação da herbivoria foi explicadapelo diâmetro no maior eixo e área foliar (Tabelas 3 e4). O número de folhas explicou a maior parte davariação da riqueza de espécies, mas com umarelação negativa (Tabelas 3 e 4). O número de folhase a altura da planta foram as variáveis responsáveispelas maiores porcentagens de explicação da variaçãode riqueza de espécies de Lepidoptera, isoladamente,com uma relação positiva (Tabelas 3 e 4).
Para Orthoptera, somente o volume de copafoi significativo, porém, a variação explicada pelomodelo foi muito baixa (Tabelas 3 e 4). ParaColeoptera, a altura de copa explicou a maior parteda variação na riqueza de espécies, mas com umarelação negativa (Tabelas 3 e 4). Para Homoptera, o

77
número de folhas, que explicou a maior parte davariação, apresentou uma relação positiva, mas ovolume de copa apresentou relação negativa (Tabelas3 e 4). Para Hemiptera, o resultado foi similar,embora somente o número de folhas e o volume decopa tenha sido significativo (Tabelas 3 e 4).
DISCUSSÃOO comprimento e a largura explicaram a
área foliar, assim, essa pode ser determinada a priori,medindo-se somente essas variáveis.
A dureza aumentou com ao aumento da áreafoliar. Folhas mais velhas acumulam mais fibras(Coley 1983, 1988, Coley et al. 1985,), sendoportanto mais duras. O. hexasperma apresentava, emum mesmo indivíduo, poucos botões foliares eindivíduos maiores e mais velhos apresentavammenor quantidade de folhas jovens que indivíduosmais jovens (observação pessoal). Entretanto, aporcentagem da variação na dureza foliar explicadapela variação na área foliar foi baixa. O. hexaspermaapresenta botões foliares durante todo o ano,principalmente na estação chuvosa (Morais et al1995). Assim, os indivíduos apresentavam folhasmais velhas e mais novas e como não houvediferenciação de estágios de desenvolvimento paracoleta e medição da dureza isso se refletiu nessavariação.
Plantas menores, com menor área foliarapresentaram maior herbivoria. Isso pode ter ocorridopor: (1) efeito amostral; (2) as folhas jovens sãoatacadas por espécies de Coleoptera que perfuram afolha antes da sua expansão, à medida que a folhaesclerifica-se e expande-se lateralmente essasperfurações são cicatrizadas, provocando umasubestimativa da porcentagem de área foliar comida;(3) folhas jovens e menores têm menor dureza, sendoportanto mais palatáveis e mais comidas (Coley &Aide 1991, Loyola, Jr. & Fernandes 1993, mas vejaCooke et al. 1984). Além disso, a porcentagem deárea foliar comida foi maior em plantas menores(diâmetro menor) que eram as plantas com maiorfrequência de folhas mais novas. Seyffarth et al.(1996) observaram que a porcentagem de ataque defolhas de O. hexasperma por lagartas de Lepidopteranão é afetada pelo comprimento foliar (que prevê aárea foliar). Entretanto, áreas queimadas apresentammaior porcentagem de ataque que áreas nãoqueimadas (17,5% contra 9,5%), indicando que oestresse causado pelo fogo pode resultar em umaumento do número de folhas jovens, maissusceptíveis ao ataque.
O número de indivíduos de herbívoros porplanta foi igual a um, praticamente para todas asespécies. Entretanto, algumas espécies foram maisfreqüentes na área (encontrados em um maior númerode plantas) que outras (Tabela 1). Embora o presente
trabalho não tenha sido realizado com o objetivo dedeterminar a abundância, aparentemente o padrão dealta riqueza e baixa abundância encontrado parece sercomum para o cerrado. Esse resultado é semelhanteao encontrado por Price et al. (1995) para lagartas deLepidoptera em Erythroxilum em um cerrado deBrasília, próximo à área do presente estudo. Ariqueza e composição de espécies de herbívoros devevariar entre as estações (Cytrynowicz, 1991), mas aabundância de insetos no verão (estação chuvosa),quando foi realizado esse trabalho, pode ser mais queo dobro que no inverno (Morais & Diniz, nãopublicado).
A relação entre complexidade estrutural eriqueza de espécies de herbívoros não apresentou umpadrão marcado e esperado pela teoria. Por um lado,plantas com maior diâmetro e altura de copaapresentaram maior número de herbívoros. Por outrolado, plantas com maior número de folhas,responsável por 15,1% da variação do número totalde espécies, tenderam a ter menor número deespécies. Esse resultado pode ter ocorrido por efeitode amostragem. Como as plantas eram observadasdurante um mesmo tempo, plantas com maior númerode folhas podem ter tido menor esforço amostral,relativamente a plantas menores. Além disso, O.hexasperma apresenta arquitetura tortuosa e bastanteramificada. Plantas maiores, com maior diâmetro ealtura apresentavam maior número de folhas, mastambém um maior espaço “aberto” no interior dacopa. Plantas menores, com menor quantidade defolhas eram mais compactas, tendo uma maior“densidade” foliar, podendo proporcionar melhorrefúgio, microclima e diversidade de recursos para osherbívoros. Para testar essa hipótese, a posteriori, foifeita uma análise de regressão linear múltipla porpassos (stepwise) considerando os elementos dearquitetura, dureza foliar e a variável densidadefoliar. O modelo foi ajustado após um passo, somentecom a entrada da variável “densidade foliar”, eexplicou 36,5% da variação da riqueza de espécies(N=50, F=29,542, p<0,001, r2=0,368,Y=1,154+0,001*DFL). Resultados diferentes podemser encontrados quando a riqueza de espécies deherbívoros é estudada a nível de comunidade deplantas. Considerando 260 espécies de plantas emuma região de cerrado no Sudeste do Brasil,Cytrynowicz (1991) encontrou 36% de explicação davariação do número de espécies de herbívoros devidoao número de folhas.
Os resultados de riqueza de espéciesseparados por ordem apresentaram a mesmatendência, sendo maior em plantas com maiordensidade de folhas, mas com uma relação positivacom o número de folhas e negativa com o volume decopa.

78
A altura da planta pode ser uma variávelarquitetônica importante para Lepidoptera (Neuvonen& Niemelä 1981) e explicou 71% da variação nariqueza de espécies de macrolepidopteras em árvorese arbustos na Finlândia. A riqueza de Lepidoptera foimaior em plantas mais altas, com maior número defolhas, porém, com menor volume de copa.Novamente aqui, a associação entre número de folhase volume mostra a preferência por plantas mais“compactas”, mas, nesse caso, em plantas maiores.Isso pode estar relacionado ao volume de recursoutilizado por lagartas. Em plantas maiores com maiorquantidade de folhas, a lagarta pode permanecer maistempo em um mesmo indivíduo, sem necessidade dedeslocamentos.
Orthoptera foi mais diverso em plantasmenores (volume de copa). Plantas menoresapresentavam maior freqüência de folhas jovens, quesão menos duras e, portanto, mais palatáveis, o quepoderia estar influenciando na escolha de plantas porespécies de Orthoptera que são raspadores. Isso podeexplicar, também, os resultados encontrados paraColeoptera, que fazem furos em folhas novas.
Riqueza de espécies de homoptera foi maiorem plantas com maior número de folhas, porém, commenor volume de copa, ou seja, em plantas commaior “densidade” de folhas. Murdoch et al. (1972)explicou 71% da variação do número de espécies deHomoptera, em uma comunidade de árvores, devidoà altura da folhagem
A relação entre riqueza de espécies deherbívoros e complexidade estrutural em O.hexasperma mostrou-se complexa. Os elementos daarquitetura, como a altura da planta, altura e diâmetrode copa, não explicaram, isoladamente, a riqueza deespécies. Isso pode estar relacionado ao desenhoarquitetônico dessa planta: plantas maiores, apesar depossuírem maior número de folhas e módulos,apresentaram copas mais abertas, com maior espaçointerno. Assim, a riqueza de herbívoros,aparentemente, foi melhor explicada pela “densidadede folhas” na copa.
Nesse trabalho, a dureza foliar não explicoua variação na riqueza de espécies de herbívoros, ouna herbivoria. Folhas novas e velhas não foramseparadas para a medição de dureza e o efeito da altafreqüência de folhas novas em plantas mais novas foirepresentado pelo efeito do tamanho da planta naherbivoria.
AGRADECIMENTOSAo IBGE, pela apoio logístico, ao Prof Dr.
Raimundo P.B. Henriques, Delano Moody e ProfaHelena C. Morais pelas valiosas sugestões. AoDepartamento de Ecologia, UnB, pela oportunidadede realizar este trabalho durante o curso deDoutorado. Ao Prof. Dr. John DuVall Hay e Prof. Dr.
Guarino Colli pelas sugestões e leitura criteriosa domanuscrito.
BIBLIOGRAFIACOLEY, P.D. 1983. Herbivory and defensive
characterisitcs of tree species in a lowlandtropical forest. Ecol. Monographs 53: 209-233.
COLEY, P.D. 1988. Effects of plant growth rate andleaf lifetime on the amount and type of anti-herbivore defense. Oecologia 74: 531-536.
COLEY, P.D., BRYANT, J.P., AND CHAPIN, III,F.S. 1985. Resource availability and plantantiherbivore defense. Science 230: 895-899.
COLEY, P.D., AND AIDE, T.M. 1991. Comparisonof herbivory and plant defenses in temperate andtropical broad-leave forest. In. P.W. Price, T.M.Lewinsohn, G.W. Fernandes, and W.W. Benson(eds). Plant-animal interactions: evolutionaryecology in tropical and temperate regions. pp.25-49, John Wiley & Sons, Inc., New York.
COOKE, F.P., BROWN, J.P., AND MOLE, S. 1984.Herbivory, foliar enzyme inhibitors, nitrogen andleaf structure of young and mature leaves in atropical forest. Biotropica 16: 257-263.
CYTRYNOWICZ, M. 1991. Resource size andpredictability, and local herbivore richness in asubtropical brazilian cerrado community. In.P.W. Price, T.M. Lewinsohn, G.W. Fernandes,and W.W. Benson (eds). Plant-animalinteractions: evolutionary ecology in tropical andtemperate regions. pp. 561-589, John Wiley &Sons, Inc., New York.
EITEN, G. 1990. Vegetação. In. M.N. Pinto (org.)Cerrado, caracterização, ocupação eperspectivas. pp. 9-66. Editora da Universidadede Brasília, Brasília.
FEENY, P. 1976. Plant apparency and chemicaldefence. Recent Adv. Phytochem. 10: 1-40.
HARPER, J.L. 1981. The concept of population inmodular organisms. In R.M. May (ed.).Theoretical ecology. pp.53-77. BlackwellScience Publ., Oxford, 2nd ed.
LAWTON, J.H. 1983. Plant architeture and thediversity of phytophagous insects. Ann. Rev.Entomol. 28: 23-39.
LAWTON, J.H., AND SCHRÖDER, D. 1977.Effects of plant type, size of geographical rangeand taxonomic isolation on number of insectspecies associated with British plants. Nature265: 137-140.
LEATHER, S.R. 1986. Insect species richness of thebritish Rosaceae: the importance of host range,plant architeture, age of establishment,taxonomic isolation and species-arearelationships. J.Anim.Ecol 55: 841-860.
LEWINSOHN, T.M. 1991. Insects in flower heads ofAsteraceae in Southeast Brazil: a case study on

79
tropical species richness. In. P.W. Price, T.M.Lewinsohn, G.W. Fernandes, and W.W. Benson(eds). Plant-animal interactions: evolutionaryecology in tropical and temperate regions. pp.525-559, John Wiley & Sons, Inc., New York.
LOYOLA, JR., R., AND FERNANDES, G.W. 1993.Herbivoria em Kielmeyera coriacea (Guttiferae):efeitos da idade da planta, desenvolvimento easpectos qualitativos de folhas. Rev. Brasil. Biol.53: 295-304.
MARQUIS, R.J. 1991. Herbivore fauna of Piper(Piperaceae) in a Costa Rican wet forest:diversity, specificity, and impact. In. P.W. Price,T.M. Lewinsohn, G.W. Fernandes, and W.W.Benson (eds). Plant-animal interactions:evolutionary ecology in tropical and temperateregions. pp. 179-208, John Wiley & Sons, Inc.,New York.
MORAIS, H.C., DINIZ, I.R., ANDBAUMGARTEN, L. 1995. Padrões de produçãode folhas e sua utilização por larvas deLepidoptera em um cerrado de Brasília. Revtabrasil. Bot. 18: 163-170.
MURDOCH, W.W., EVANS, F.C., ANDPETERSON, C.H. 1972. Diversity and pattern inplants and insects. Ecology 53: 819-829.
NEUVONEN, S., AND NIEMELÄ, P. 1981. Speciesrichness of macrolepidoptera on finnishdeciduous trees and shrubs. Oecologia 51: 364-370.
OLIVEIRA, P.S., AND OLIVEIRA-FILHO, A.T.1991. Distribution of extrafloral nectaries in thewoody flora of tropical communities in westernBrazil. In. P.W. Price, T.M. Lewinsohn, G.W.Fernandes, and W.W. Benson (eds). Plant-animal interactions: evolutionary ecology intropical and temperate regions. pp. 163-175,John Wiley & Sons, Inc., New York.
OLIVEIRA, P.S., KLITZKE, C., AND VIEIRA, E.1995. The ant fauna associated with theextrafloral nectaries of Ouratea hexasperma(Ochnaceae) in an area of cerrado vegetation inCentral Brazil. Entomologist’s MonthlyMagazine 131: 77-82.
PRICE, P.W., DINIZ, I.R., MORAIS, H.C., ANDMARQUES, E.S.A. 1995. The abundance ofinsect herbivore species in the tropics: the highlocal richness of rare species. Biotropica 27:468-478.
RATTER, J.A. 1986. Notas sobre a vegetação daFazenda Água Limpa (Brasília, DF). Editora daUniversidade de Brasília, Brasília.
SEYFFARTH, J.A.S., CALOURO, A.M., ANDPRICE, P.W. 1996. Leaf rollers in Ourateahexasperma (Ochnaceae): fire effect and theplant vigor hypothesis. Rev. Brasil. Biol. 56:135-137.
SOUTHWOOD, T.R.E., MORAN, V.C., ANDKENNEDY, C.E. 1982. The richness, abundanceand biomass of the arthropod communities ontrees. J. Anim. Ecol. 51: 635-649.
STRONG, JR., D.R. 1977. Rolled-leaf hyspinebeetles (Chrysomelidae) and their Zingiberaleshost plant in middle America. Biotropica 9: 156-169.
STRONG, JR., D.R., AND LEVIN, D.A. 1979.Species richness of plant parasites and growthform of their hosts. Am. Nat. 114: 1-22.
STRONG, JR., D.R., MCCODY, E.D., AND REY,J.R. 1977. Time and the number of herbivoresspecies: the pests of sugarcane. Ecology 58: 167-175.
ZAR, J.H. 1974. Bioestatistical analysis. Prentice-Hall, Inc. Englewood Cliffs.

80
Tabela 1. Morfoespécies de herbívoros encontradasem Ouratea hexasperma e número de plantas em queforam encontradas, em um cerrado do IBGE.Ordem/Família Espécie PlantasLepidopteraMegalopygidae Megalopyge albicollis 07Geometridae Cyclomia ocana 01Arctiidae Fregela semiluna 08Indeterminado Lepidoptera sp1 05
Lepidoptera sp2 07Lepidoptera sp3 07
OrthopteraAcrididae Acridinae sp1 15
Acridinae sp2 18Acridinae sp3 04Cyrtacanthacridina sp1 13Cyrtacanthacridinae sp2 06Cyrtacanthacridinae sp3 05
Tetigonidae Copiphorinae sp1 12Phaneropterinae sp1 08
ColeopteraBuprestidae Buprestidae sp1 01Cantharidae Cantharidae sp1 01Bruchidae Bruchidae sp1 10Scarabaeidae Melolonthinae sp1 01HemipteraCoriscidae Coriscidae sp1 05Corizidae Corizidae sp1 11HomopteraCicadellidae Cicadellidae sp1 10
Cicadellidae sp2 10Cicadellidae sp3 10Cicadellidae sp4 12Cicadellidae sp5 07Cicadellidae sp6 01Cicadellidae sp7 01
Membracidae Membracidae sp1 09Fulgoroidae Fulgoroidae 01
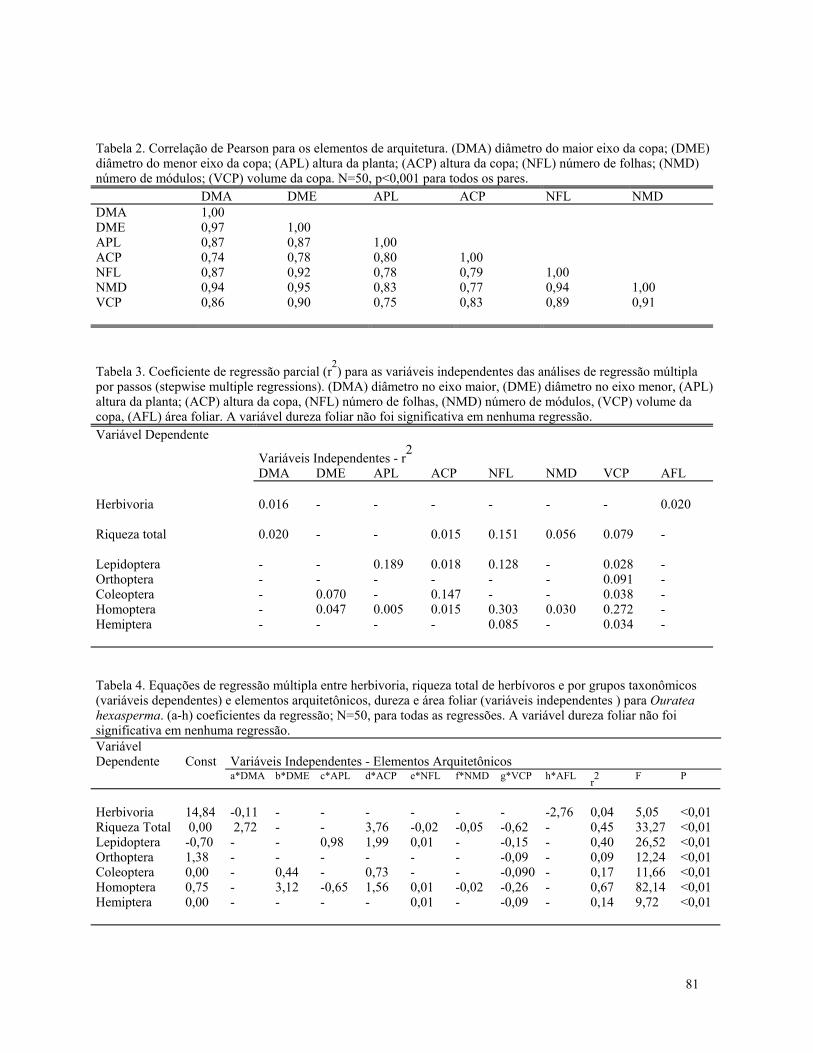
81
Tabela 2. Correlação de Pearson para os elementos de arquitetura. (DMA) diâmetro do maior eixo da copa; (DME)diâmetro do menor eixo da copa; (APL) altura da planta; (ACP) altura da copa; (NFL) número de folhas; (NMD)número de módulos; (VCP) volume da copa. N=50, p<0,001 para todos os pares.
DMA DME APL ACP NFL NMDDMA 1,00DME 0,97 1,00APL 0,87 0,87 1,00ACP 0,74 0,78 0,80 1,00NFL 0,87 0,92 0,78 0,79 1,00NMD 0,94 0,95 0,83 0,77 0,94 1,00VCP 0,86 0,90 0,75 0,83 0,89 0,91
Tabela 3. Coeficiente de regressão parcial (r2) para as variáveis independentes das análises de regressão múltiplapor passos (stepwise multiple regressions). (DMA) diâmetro no eixo maior, (DME) diâmetro no eixo menor, (APL)altura da planta; (ACP) altura da copa, (NFL) número de folhas, (NMD) número de módulos, (VCP) volume dacopa, (AFL) área foliar. A variável dureza foliar não foi significativa em nenhuma regressão.Variável Dependente
Variáveis Independentes - r2
DMA DME APL ACP NFL NMD VCP AFL
Herbivoria 0.016 - - - - - - 0.020
Riqueza total 0.020 - - 0.015 0.151 0.056 0.079 -
Lepidoptera - - 0.189 0.018 0.128 - 0.028 -Orthoptera - - - - - - 0.091 -Coleoptera - 0.070 - 0.147 - - 0.038 -Homoptera - 0.047 0.005 0.015 0.303 0.030 0.272 -Hemiptera - - - - 0.085 - 0.034 -
Tabela 4. Equações de regressão múltipla entre herbivoria, riqueza total de herbívoros e por grupos taxonômicos(variáveis dependentes) e elementos arquitetônicos, dureza e área foliar (variáveis independentes ) para Ourateahexasperma. (a-h) coeficientes da regressão; N=50, para todas as regressões. A variável dureza foliar não foisignificativa em nenhuma regressão.VariávelDependente Const Variáveis Independentes - Elementos Arquitetônicos
a*DMA b*DME c*APL d*ACP e*NFL f*NMD g*VCP h*AFL r2 F P
Herbivoria 14,84 -0,11 - - - - - - -2,76 0,04 5,05 <0,01Riqueza Total 0,00 2,72 - - 3,76 -0,02 -0,05 -0,62 - 0,45 33,27 <0,01Lepidoptera -0,70 - - 0,98 1,99 0,01 - -0,15 - 0,40 26,52 <0,01Orthoptera 1,38 - - - - - - -0,09 - 0,09 12,24 <0,01Coleoptera 0,00 - 0,44 - 0,73 - - -0,090 - 0,17 11,66 <0,01Homoptera 0,75 - 3,12 -0,65 1,56 0,01 -0,02 -0,26 - 0,67 82,14 <0,01Hemiptera 0,00 - - - - 0,01 - -0,09 - 0,14 9,72 <0,01

82
EFEITOS DE DIFERENTES REGIMES DEQUEIMA SOBRE A VEGETAÇÃO LENHOSADE CERRADO SENSU STRICTO: TAXASDIFERENCIAIS DE REBROTAS
Saulo Marques de Abreu Andrade
RESUMOAs rebrotas são eficientes mecanismos de
sobrevivência das plantas adaptadas ao fogo. Emduas áreas de 10 ha de cerrado sensu stricto,submetidas a diferentes regimes de queimas, umaqueimada bianualmente e outra vinte anos semqueima, foram amostrados todos os indivíduos, auma distância de 1 metro de quatro transectos de 50metros cada, com diâmetro de 5 centímetros a 30 cmdo solo. Todas as rebrotas foram contadas eclassificadas em aéreas, basais e subterrâneas. Aporcentagem de rebrotas aéreas foi de 30 e 8 %respectivamente, de 51 e 67 % de rebrotas basais e,19 e 25 % de rebrotas subterrâneas. Estes valorescomparados com o do controle mostram que ocorreuma mudança temporária na arquitetura davegetação.
INTRODUÇÃOO fogo seleciona aquelas espécies de
cerrado que apresentam estratégias de resistência aqueimada (Guedes 1993). Cada espécie possuicapacidade de recuperar-se frente um distúrbioconforme suas reservas de energia para rebrota,existência de bancos de semente no solo, capacidadede recolonização e habilidade de estabelecimento emcondições extremas (Frost et al. 1986). Nacomunidade lenhosa as estratégias frente ao fogo sãodistintas das do estrato rasteiro, visto que estes doiscomponentes do Cerrado possuem formas de vidasdiferentes e se localizam diferentemente com respeitoàs zonas de maior impacto de calor.
Sambuichi (1991), afirma que o fogo alémde ralear a camada lenhosa, diminui sua diversidade.Segundo Ramos (1990), áreas protegidas contraqueima, apresentam densidade arbórea, área basalviva e cobertura arbórea aproximadamente o dobrodaquelas submetidas a queimas bienais. Proteçãocontra fogo em savanas aumenta a densidade deárvores, diminuindo a camada de gramíneas (Frost etal. 1986). Com relação às plantas lenhosas jáestabelecidas, a mortalidade devida ao fogo é muitobaixa, sendo a destruição parcial da copa o danoprincipal, exceto para plântulas. Este dano varia coma intensidade e regime das queimadas, podendo levarà exclusão de espécies sensíveis, econsequentemente, a uma mudança na composiçãoflorística (Ramos 1990).
Ramos (1990), cita o dano estrutural nulo,isto é, aquele em que os indivíduos, principalmentearbóreos, com altura superior a três metros,apresentam apenas murchamento e queda das folhas.Este seria o menor efeito do fogo sobre as lenhosas.O que ocorre é que após a queima as folhas murchame caem devido ao calor. Após esta queda, aquelasgemas que conseguiram resistir ao calor emitemnovos brotos.
Armando (1994), mediu o efeito do fogo nasobrevivência e no crescimento de espécies arbóreasnas fases iniciais de crescimento (até 30 cm de alturae 5 cm de circunferência), em cerrado. Seusresultados evidenciam aumento de mortalidade ediminuição da altura atingida por rebrota. O intervalode um ano não foi suficiente para a recuperação daaltura existente antes do fogo, em juvenis e plântulas.Deste modo, queimas anuais poderão levar à reduçãoe eliminação das populações de certas espécies.
Segundo Guedes (1993), temperaturas acimade 60 0C (inclusive), causam morte do câmbiovascular. Um incêndio que apresente 380 0C por 0,7min. de duração (situação próxima à que ocorre nocerrado) alcança tal temperatura em árvores queapresentam casca de espessura menor que 6 mm.
A capacidade de rebrota está relacionada aotamanho da plântula (Armando 1994), ocorrendocom grande rapidez na época seca e chuvosa (Souza& Soares 1983; Warming 1973). A reserva em suasestruturas faz com que estas espécies mesmo após ofogo possam retomar seu crescimento, ultrapassandoa altura crítica onde o fogo causa danos diretos. Oinício, a duração e o pico de brotamento variamconforme as adaptações de cada espécie e tambémcom a disponibilidade de recursos. Água e nutrientesestão disponíveis no sistema subterrâneo da planta(xilopódios) ou no solo, permitindo o brotamento naestação seca (Souza & Soares 1983).
O estrato rasteiro ganha grande importânciana intensidade do fogo, pois quanto maior o acúmulode combustível mais intenso será o fogo (Ramos eRosa 1992), afetando, então o tipo de rebrota.Dependendo do dano ocorrido, as plantas poderãosimplesmente repor suas folhas, regenerar a partir degemas aéreas protegidas pela casca, ou a partir degemas situadas abaixo do solo, quando todo o caulefor destruído (Chandler et al. 1983). Quanto maior otamanho da parte morta, maior o entouceiramento(Ramos 1990).
Existem diversas vantagens ecológicasassociadas com a persistência por brotamentos apósdistúrbios como o fogo, principalmente em ambientesquentes e com um estresse hídrico, uma vez que estascondições são desfavoráveis para o estabelecimentode sementes. Desta forma, a utilização de estruturasde armazenamento, que possuem água e nutrientes, é

83
extremamente benéfica ao indivíduo mesmo emépocas secas (Souza & Soares 1983).
O objetivo deste estudo é documentar equantificar o número de rebrotas basais (i.e. rebrotaslogo na base do indivíduo), subterrâneas (rebrotas deestruturas vegetativas a uma distância máxima de 25cm, para serem consideradas do mesmo indivíduo) eaéreas (rebrotas que saem de debaixo da casca dopróprio caule) e, verificar se em diferentes regimesde queima há uma taxa diferencial entre esses doistipos de rebrotas, isto é, como a frequência e aintensidade do fogo afetam o número e o crescimentodas rebrotas.
MATERIAL E MÉTODOSO experimento foi realizado na Reserva
Ecológica do IBGE (RECOR) localizada a 35 km aosul de Brasília. Foram marcados 4 transectos de 50 mcada, em três áreas de 10 ha de cerrado sensu strictodo Projeto Fogo submetidas a diferentes regimes dequeima. 1.Controle- vinte anos sem queima;2.Bianual modal- com queima de 2 em 2 dois anos eque foi queimada pela primeira vez em 1992 após 20anos sem queima e depois em 1994;3.Cerrado de 1994- que era uma área de cerradocontrole 20 anos sem queima mas, que durante umincêndio em outubro de 1994 foi queimado,simulando uma queimada 20 anos sem queimaporém, não prescrita. Tomada neste trabalho comoexemplo de um incêndio natural, onde pelo grandeacúmulo de combustível espera-se uma maiorintensidade do fogo. Todos os indivíduos à distância de um metro aolongo dos transectos e com diâmetro mínimo de 5 cma 30 cm acima do solo foram anotados, contando-se onúmero de rebrotas e diferenciando-as em basais,subterrâneas e áreas (contando-se o indivíduo todocomo uma rebrota já que continua vivo).
RESULTADOS E DISCUSSÃOUtilizando-se o teste estatístico ANOVA,
para comparar a diferença entre os valores médios deespécies e indivíduos entre as áreas, verificou-se queo número de espécies nas áreas controle e cerrado1994 (Tabela 1) não se apresentaramsignificativamente diferentes (P<0,05), mesmo apósuma queimada de 20 anos sem queima na área docerrado 1994, sugerindo que apenas uma queimadaem uma área de cerrado sensu stricto protegida, aindaque por 20 anos, não é tão prejudicial. Porém onúmero de espécies e de indivíduos na bienal modal ésignificativamente diferente (P<0,05) e menor quedas áreas controle e cerrado 1994 (Tabela 1),indicando um raleamento da vegetação emconseqüência da freqüência de queima.
Na Tabela 2 observa-se que o número derebrotas basais na área do cerrado 1994 ésignificativamente diferente (P<0,05) e maior que nabienal controle assim como nesta o número derebrotas aéreas é bem maior que na do cerrado 1994,estes valores são significativamente diferentes(P<0,05).
É também nítido que no cerrado controlenão há rebrotas, de todos os 337 indivíduosamostrados nenhum apresentou rebrotas.
Porém, o tipo de rebrota está ligadodiretamente à gravidade do dano ocorrido(Chandler et al. 1983), quanto maior o dano menor acapacidade de recuperação por gemas apicais debaixoda casca (Kauffman 1991), isto é, menor número derebrotas aéreas.
Ramos (1990) mostra que quanto maior odano da parte aérea maior o entouceiramento. Istosignifica um maior número de rebrotas basais esubterrâneas. Quando a parte aérea é toda destruída aplanta não tem a capacidade de formar rebrotasaéreas, pois todas a gemas apicais foram destruídas.
O grande número de rebrotas aéreasencontradas na bienal modal (Tabela 2) ésignificativamente diferente (P<0,05), segundo testedo X2, da área do cerrado 1994 sugerindo que aintensidade do fogo foi menor, não danificando aplanta a ponto desta perder duas gemas apicais, istoem decorrência de um menor acúmulo de biomassa jáque a área foi queimada em 1992. Entretanto, nocerrado 1994 o acúmulo de biomassa era grande,fazendo com que a intensidade do fogo fosse maior,pois este acúmulo tem grande importância naintensidade do fogo (Ramos e Rosa 1992).
Cada espécie possui capacidade derecuperar-se frente um distúrbio conforme suasreservas de energia para rebrota (Frost et al.1986).Dessa forma pode-se especular que no cerrado 1994estas reservas eram maiores uma vez que não eraqueimado há vinte anos, mostrando assim um númerode rebrotas tanto basais quanto subterrâneassignificativamente diferente (P<0,05) e maior que ada bienal modal, uma vez que esta já foi queimadaduas vezes, esgotando assim a reservas para rebrotas.O número de fustes mortos indicam que a queimadado cerrado 1994 foi mais prejudicial às plantas.
Diferentes regimes de queima afetamdiferencialmente as taxas de rebrotas, sendo quequanto mais intenso, menor o número de rebrotasaéreas, uma vez que as gemas apicais são mortas. Istodetermina um maior número de rebrotas basais esubterrâneas já que estas gemas, por se encontraremprotegidas abaixo do solo, não sofrem os efeitos dasaltas temperaturas da queimadas.
As plantas que se reproduzemvegetativamente, se restabelecem predominantementepor rebrotas do tipo basal, após a queimadas.

84
O grande número de rebrotas, após asqueimadas, causam uma mudança temporária naarquitetura da vegetação, decorrente doentouceiramento. Uma vez que no cerrado protegidodo fogo por 20 anos, não se observam o grandenúmero de rebrotas encontrado nas áreas queimadas.
AGRADECIMENTOSAo professor Raimundo Paulo Henriques
pela organização do curso e grande ajuda naidentificação das espécies. A Reserva Ecológica doIBGE pelo cessão do alojamento e toda a infra-estrutura. A todos os professores que com o Rodiasolde suas inteligências exterminaram os pernilongos denossas dúvidas. Aos queridos Fátima, Formigão ecolaboradores pelas deliciosas refeições sem a quaisnão teríamos sobrevivido. A todos os meus queridosamigos que comigo desfrutaram de um grande cursoe partilharam dias de muita alegria. Aos meusqueridos parceiros de grupo que me ensinaram muitopartilhando seus conhecimentos. E finalmente aogrande e incomparável The King pela inspiração emtodos os momentos deste curso.
BIBLIOGRAFIAARMANDO, M. S. 1994. O impacto do fogo na
rebrota de algumas espécies de árvores docerrado. Dissertação de mestrado. Universidadede Brasília. 75 pp
CHANDLER, C.; CHENEY, P.; THOMAS, P.;-TRUSAND, L. & WILLIANS, S. 1983. Fireeffects on vegetation. In: Fire in forestry: forestfire behavior and effects. John Willey and Sons.1: 255-289.
FROST, P.; MEDINA, E.; MENAUT, J. C.;SOLBRIG, O.; SWIFT. M & WALKER, B. H..1986. Responses of savannas to stress anddisturbance: a proposal for a collaborativeprogramme of research. IUBS. Special Issue 10.82 pp.
GUEDES, D. M. 1993. Resistência das árvores doCerrado ao fogo: Papel da casca como isolantetérmico. Dissertação de Mestrado. Universidadede Brasília. 113 pp.
KAUFFMAN, J. B. 1991. Survival by sproutingfollowing fire in tropical forests of the EasternAmazon. Biotropica 23(3): 219-224.
RAMOS, A. E. & ROSA, C. M. M. 1992. Impactodas queimadas. In: Alternativas dedesenvolvimento dos Cerrados manejo econservação dos recursos naturais renováveis.Dias, B. F. S. (coord.). IBAMA/FUNATURA,Brasília, 34-38.
RAMOS, A. E. 1990. Efeitos da queima na vegetaçãolenhosa do cerrado. Dissertação de mestrado.Universidade de Brasília. 142 pp.
ROSA, C. M. M. 1990. Recuperação pós-fogo doestrato rasteiro de um campo sujo de cerrado.Dissertação de mestrado. Universidade deBrasília. 162 pp.
SAMBUICHI, R. H. R. 1991. Efeitos de longo prazodo fogo periódico sobre a fitossociologia dacamada lenhosa de um cerrado em Brasília, DF.Dissertação de mestrado. Universidade deBrasília. 130 pp.
SOUZA, M. H. A. O. & SOARES, J. J. 1983.Brotamento de espécies arbustivas e arbóreas,posteriormente a uma queimada, num cerradão.An. Sem. Reg. Ecol. III: 263-275.
WARMING, E. 1973. Lagoa Santa, contribuiçãopara a geographia phitobiológica. LivrariaItatiaia, Belo Horizonte & EDUSP, São Paulo,279 pp.
Tabela 1. Número médio de espécies e indivíduos portransecto encontrados nas três áreas.Área espécies
x (s)indivíduosx (s)
Bianual modal 18 (2) 61 (3)Cerrado 1994 23 (1) 73 (4)Controle 26 (2) 84 (8)
Tabela 2. Porcentagem de rebrotas nas três áreas,com o número absoluto entre parênteses, e número defustes mortos.
Rebrotas (%)Área basais aéreas subterr. fustes
mortosBienal modal 51(244) 30(144) 19(89) 179Cerrado 1994 67(506) 8(59) 25(185) 238Controle 0 0 0 20
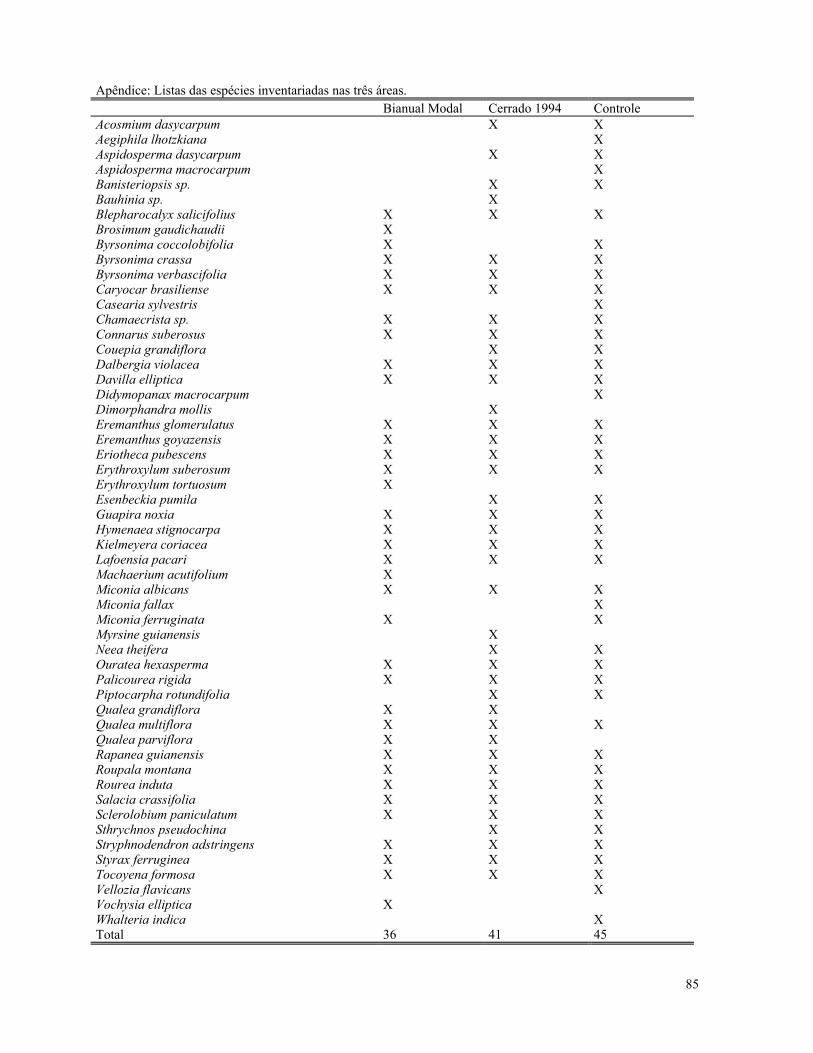
85
Apêndice: Listas das espécies inventariadas nas três áreas.Bianual Modal Cerrado 1994 Controle
Acosmium dasycarpum X XAegiphila lhotzkiana XAspidosperma dasycarpum X XAspidosperma macrocarpum XBanisteriopsis sp. X XBauhinia sp. XBlepharocalyx salicifolius X X XBrosimum gaudichaudii XByrsonima coccolobifolia X XByrsonima crassa X X XByrsonima verbascifolia X X XCaryocar brasiliense X X XCasearia sylvestris XChamaecrista sp. X X XConnarus suberosus X X XCouepia grandiflora X XDalbergia violacea X X XDavilla elliptica X X XDidymopanax macrocarpum XDimorphandra mollis XEremanthus glomerulatus X X XEremanthus goyazensis X X XEriotheca pubescens X X XErythroxylum suberosum X X XErythroxylum tortuosum XEsenbeckia pumila X XGuapira noxia X X XHymenaea stignocarpa X X XKielmeyera coriacea X X XLafoensia pacari X X XMachaerium acutifolium XMiconia albicans X X XMiconia fallax XMiconia ferruginata X XMyrsine guianensis XNeea theifera X XOuratea hexasperma X X XPalicourea rigida X X XPiptocarpha rotundifolia X XQualea grandiflora X XQualea multiflora X X XQualea parviflora X XRapanea guianensis X X XRoupala montana X X XRourea induta X X XSalacia crassifolia X X XSclerolobium paniculatum X X XSthrychnos pseudochina X XStryphnodendron adstringens X X XStyrax ferruginea X X XTocoyena formosa X X XVellozia flavicans XVochysia elliptica XWhalteria indica XTotal 36 41 45





























