Embed Size (px)
Citation preview

7/27/2019 edificando la célula
http://slidepdf.com/reader/full/edificando-la-celula 1/10
Clls highly complx stuctus1. This complxity ffcts biologicl fuctios t my siz scls2. But how dos this complxity is? Wht mchisms pttth chitctu of th cll d wh dos th ifom-tio to spcify distict chitctu fo difft clltyps com fom?
Th mout of ifomtio tht is dd to dictth ctio of th cllul ssmbly pthwys could b mii-ml bcus cllul mophogsis is sl-organizing
procss3. Subjct to simpl iputs tht c bis how thsystm dlops, slf-ogizig pocsss c gtcomplx, pttd sposs. I picipl, thfo, thcll chitctu could b spcifid by smll umb of distict gs, which would s to modult th ih-t slf-ogizig pocsss tht ld to th physiclshp of clls d oglls. Udstdig th oigisof cll ogiztio thus quis us to sw th qus-tio of how th slf-ogizig physicl d biochmiclpoptis of cllul costituts c cot simpltmpol chgs i g-xpssio lls ito complxchgs i physicl, th-dimsiol stuctus. Thisiw ddsss this qustio t th stuctul ll
of th ogll by skig how th gomty (th siz,umb, positio d mophology) of oglls isspcifid d gultd.
I ll css, w will ttmpt to idtify dsig piciplsby which simpl iput cotiig miiml ifomtioc poduc complx output i tms of mophologiclchgs. I giig tmiology dsig piciplis simpl ul tht, wh followd i th dsig of mchi, sus o t lst icss th liklihoodof pop ssmbly d fuctio. W ot tht ou us of th tm dos ot imply th xistc of ‘dsig’, butgi th cut itst i sythtic biology w suggsttht th dsig picipls usd i olutio could ow b
usd to pogmm clls i tiol wy. Fo xmpl,w gu tht commo sttgy usd by clls to gultogll siz is to limit ith th ssmbly o disssm-bly t of ogll by pocss tht is ihtly dpdt o ogll siz. Oth spcific piciplstht w discuss th cotol of ogll umb by th blc of ogll fomtio d ptitioig; thstblishmt of cllul polity by slf-ogiztio thtis bisd by xtl iputs; th cotol of ogll posi-tio by dict motio o polizd cytoskltl ys;d th cotol of ogll shp by mmb-bidigpotis tht di locl cutu chgs.
How cells measure size and length
Lgth d siz fudmtl pmts fo spci-fyig gomty. Ogll siz must b ppopitly scld s fuctio of cll siz4 d physiologicldmds. Mitochodio siz i yst, fo xmpl, ispopotiol to cll olum (G. Psc, Th MolculScics Istitut, Bkly, psol commuictio).Ogll siz is tightly cotolld: fo xmpl, thcofficit of itio i th lgths of ukyotic cilia
d laglla (th two tms will b usd itchgbly i this iw) is lss th 10% (Ref. 5). But how thsizs of subcllul stuctus dtmid? Gticscs h idtifid gs tht iold i sizcotol fo ious oglls, icludig cili d flg-ll6, poxisoms7 d lipid doplts8. How, i mostcss th mchistic bsis fo siz lttios i thsmutts mis ukow.
Measuring leng th using molecular rulers. O wy to cotol th siz of li stuctus is fo th cll tomsu th lgth of th stuctu with molculul poti (fIG. 1a) which hs physicl lgth tht
Department of Biochemistry
and Biophysics,
University of California,
San Francisco,
600 16th Street,
San Francisco,
California 94158, USA.
e‑mails:
doi:10.1038/nrm2460
Self-organizing process
A procss by which a st
o componnts that can, in
principl, b connctd in
various possibl pattrns
will spontanously associat
into a limitd subst o
pattrns, without any xtrnal
input o inormation. ‘Sl-
organization’ is to b
contrastd with ‘sl-assmbly’,in which componnts can
only it togthr such that only
on pattrn is possibl.
Cilium
A microtubul-basd motil
and snsory organll
that projcts rom th surac
o many ukaryotic clls.
Flagellum
An altrnativ trm or
cilia whn applid to
ukaryotic clls.
Building the cell: design principles of cellular architectureSusanne M. Rafelski and Wallace F. Marshall
Abstract | The astounding structural complexity of a cell arises from the action of a
relatively small number of genes, raising the question of how this complexity is achieved.
Self-organizing processes combined with simple physical constraints seem to have key
roles in controlling organelle size, number, shape and position, and these factors then
combine to produce the overall cell architecture. By examining how these parametersare controlled in specific cell biological examples we can identify a handful of simple design
principles that seem to underlie cellular architecture and assembly.
R E V I E W S
naTUre revIeWS | molecular cell biology vOLUMe 9 | aUGUST 2008 | 593
7/27/2019 edificando la célula
http://slidepdf.com/reader/full/edificando-la-celula 2/10
L e n g t h
2 L
L e
n g t h
1 . 5
L
L e n g t h
L
Molecular rulera
Organelle
OrganellePrecursor materialb
Cell
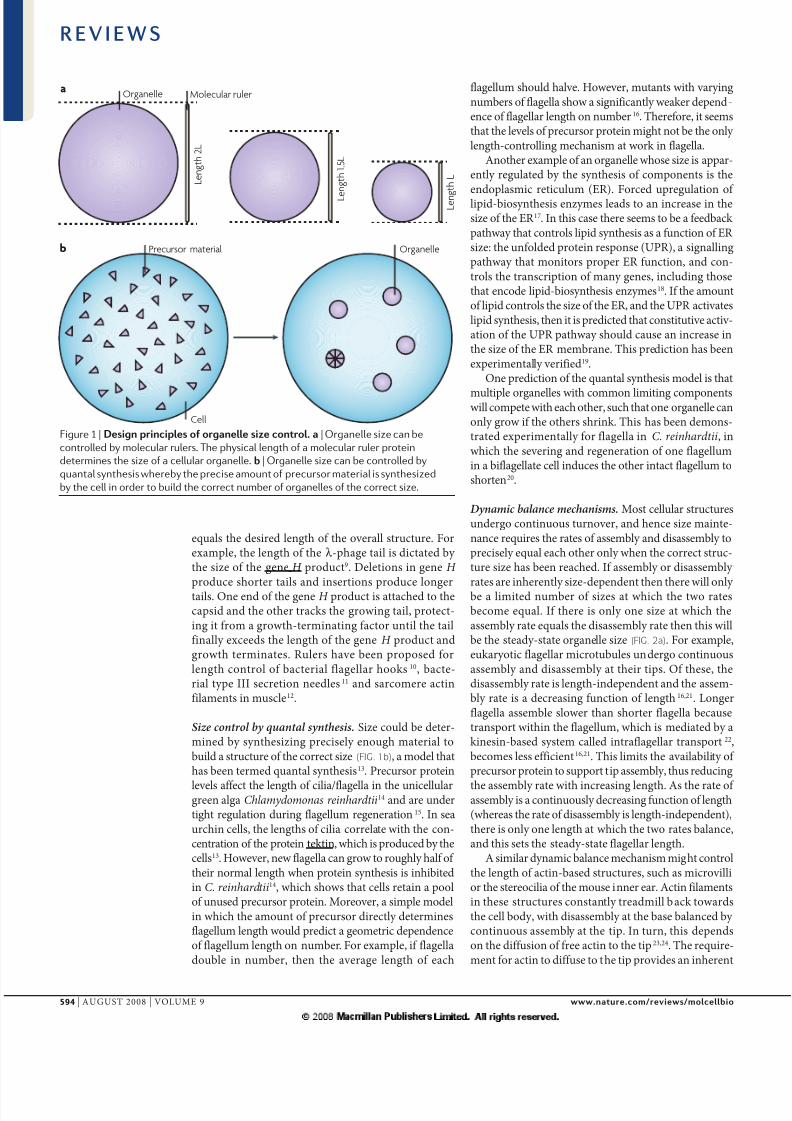
quls th dsid lgth of th oll stuctu. Foxmpl, th lgth of th λ -phg til is dicttd by th siz of th g H poduct9. Dltios i g H poduc shot tils d istios poduc logtils. O d of th g H poduct is ttchd to thcpsid d th oth tcks th gowig til, potct-ig it fom gowth-tmitig fcto util th tilfilly xcds th lgth of th g H poduct dgowth tmits. ruls h b poposd folgth cotol of bctil flgll hooks10, bct-il typ III sctio dls11 d scom ctifilmts i muscl12.
Size control by quantal synthesis. Siz could b dt-mid by sythsizig pcisly ough mtil tobuild stuctu of th coct siz (fIG. 1b), modl thths b tmd qutl sythsis13. Pcuso poti
lls ffct th lgth of cili/flgll i th uicllulg lg Chlamydomonas reinhardtii14 d udtight gultio duig flgllum gtio15. I suchi clls, th lgths of cili colt with th co-cttio of th poti tkti, which is poducd by thclls13. How, w flgll c gow to oughly hlf of thi oml lgth wh poti sythsis is ihibitdi C. reinhardtii14, which shows tht clls ti poolof uusd pcuso poti. Moo, simpl modli which th mout of pcuso dictly dtmisflgllum lgth would pdict gomtic dpdcof flgllum lgth o umb. Fo xmpl, if flglldoubl i umb, th th g lgth of ch
flgllum should hl. How, mutts with yigumbs of flgll show sigifictly wk dpd-c of flgll lgth o umb16. Thfo, it smstht th lls of pcuso poti might ot b th oly lgth-cotollig mchism t wok i flgll.
aoth xmpl of ogll whos siz is pp-tly gultd by th sythsis of compots is thdoplsmic ticulum (er). Focd upgultio of lipid-biosythsis zyms lds to ics i thsiz of th er 17. I this cs th sms to b fdbck pthwy tht cotols lipid sythsis s fuctio of er siz: th ufoldd poti spos (UPr), siglligpthwy tht moitos pop er fuctio, d co-tols th tsciptio of my gs, icludig thostht cod lipid-biosythsis zyms18. If th moutof lipid cotols th siz of th er, d th UPr ctitslipid sythsis, th it is pdictd tht costituti cti-tio of th UPr pthwy should cus ics ith siz of th er mmb. This pdictio hs bxpimtlly ifid19.
O pdictio of th qutl sythsis modl is tht
multipl oglls with commo limitig compotswill compt with ch oth, such tht o ogll coly gow if th oths shik. This hs b dmos-ttd xpimtlly fo flgll i C. reinhardtii, iwhich th sig d gtio of o flgllumi biflgllt cll iducs th oth itct flgllum toshot20.
Dynamic balance mechanisms. Most cllul stuctusudgo cotiuous tuo, d hc siz mit-c quis th ts of ssmbly d disssmbly topcisly qul ch oth oly wh th coct stuc-tu siz hs b chd. If ssmbly o disssmbly ts ihtly siz-dpdt th th will oly b limitd umb of sizs t which th two tsbcom qul. If th is oly o siz t which thssmbly t quls th disssmbly t th this willb th stdy-stt ogll siz (fIG. 2a). Fo xmpl,ukyotic flgll micotubuls udgo cotiuousssmbly d disssmbly t thi tips. Of ths, thdisssmbly t is lgth-idpdt d th ssm-bly t is dcsig fuctio of lgth16,21. Logflgll ssmbl slow th shot flgll bcustspot withi th flgllum, which is mditd by kisi-bsd systm clld itflgll tspot22,bcoms lss fficit16,21. This limits th ilbility of pcuso poti to suppot tip ssmbly, thus ducig
th ssmbly t with icsig lgth. as th t of ssmbly is cotiuously dcsig fuctio of lgth(whs th t of disssmbly is lgth-idpdt),th is oly o lgth t which th two ts blc,d this sts th stdy-stt flgll lgth.
a simil dymic blc mchism might cotolth lgth of cti-bsd stuctus, such s micoillio th stocili of th mous i . acti filmtsi ths stuctus costtly tdmill bck towdsth cll body, with disssmbly t th bs blcd by cotiuous ssmbly t th tip. I tu, this dpdso th diffusio of f cti to th tip23,24. Th qui-mt fo cti to diffus to th tip poids iht
Figure 1 | D pp z t. | Organelle size can be
controlled by molecular rulers. The physical length of a molecular ruler protein
determines the size of a cellular organelle. | Organelle size can be controlled by
quantal synthesis whereby the precise amount of precursor material is synthesized
by the cell in order to build the correct number of organelles of the correct size.
R E V I E W S
594 | aUGUST 2008 | vOLUMe 9 www.t./w/

7/27/2019 edificando la célula
http://slidepdf.com/reader/full/edificando-la-celula 3/10
Number decrease
Number increase
Disassembly rate
b
a
Assembly rate
Disassembly rate
Assembly rate
Number decrease
Number increase
Organelle
Number increase anddecrease alter organellenumber
Number increase and/ordecrease depends onorganelle number
Cell
Assembly and/or disassemblyrate depends on size
Assembly and disassemblyalter size
Mitotic spindle
A highly dynamic array
o microtubuls that orms
during mitosis and srvs
to mov th duplicatd
chromosoms apart.
Centriole
A short, barrl-lik array
o microtubuls that
organizs th cntrosom
and contributs to cytokinsis
and cll-cycl progrssion.
Autophagy
A pathway or th rcycling
o cllular contnts, in which
matrials insid th cll ar
packagd into vsicls and ar
thn targtd to th vacuol
or lysosom or bulk turnovr.
dpdc of th ssmbly t o th lgth of thfilmt. Diffusio-iducd lgth dpdc of th ssmbly t hs lso b suggstd fo thcotol of bctil flgll lgth25.
Yst kisi-8 bids log th lgth of mico-tubuls, mos pocssily to thi tips d thctlyss disssmbly. Log micotubuls cuitmo kisi-8, which sults i lgth-dpdtdisssmbly pocss tht, wh combid with costt t of ssmbly, yilds uiqu stdy-sttmicotubul lgth26. Ulik flgll d micoilli, itsms to b th disssmbly of th micotubuls, thth th ssmbly, tht is gultd s fuctio of lgth. I complx cytoskltl ssmblis, such sth mitotic spindl, lgth cotol might iol multi-pl opposig lgth-dpdt pocsss i mocomplictd dymic blc27.
Th sm typs of dymic blc mchisms tht usd to cotol lgth could lso b usd to co-tol th olum o sufc of th ogll. If ogll such s th Golgi pptus xchgs sicls
with th er, sicl fusio o th ogll occus t t tht is dicttd by th t of sicl buddig fomth er, th th by th ogll siz. By cotst,
sicl buddig fom th ogll could occu t ttht is popotiol to th sufc of th ogll(ssumig costt dsity of buddig sits). Th tsof mmb dditio d mmb mol wouldoly blc ch oth fo sigl sufc lu,thby dicttig th sufc of th ogll. If th ogll bcm lg th this th buddig twould ics, ldig to ductio i sufc :if th ogll bcm too smll th buddig twould dcs lti to th fusio t d hcit would lg to its stdy-stt sufc .
How cells control organelle number
a typicl ukyotic cll hs o uclus, two cntriols,o er, multipl mitochodi, scos of dosomsd my ibosoms. How is th umb of ogllscotolld?
Number control as a dynamic balance. Th umb of oglls flcts th blc btw two compt-ig pocsss: th ics i umb du to de novo sythsis o fissio of p-xistig oglls; dth dcs i umb du to fusio with othoglls, dgdtio pocsss such s autophagy, o
th ptitioig of oglls ito dught clls du-ig cll diisio. To cotol th umb of oglls,o o mo of ths pocsss must b gultd.It hs b poposd, fo xmpl, tht th sythsis of chlooplsts is gultd s fuctio of th umbof chlooplsts i lgl clls28. Clls tht ihit toofw chlooplsts will mk mo, whs clls thtihit too my will stop chlooplst sythsis.a simil schm hs b xpimtlly dmo-sttd fo ctiols29. Thfo, th cotol of ogll umb shs cocptully simil sttgy to th dymic blc mchisms of ogll sizcotol (fIG. 2).
Controlling the increase in organelle number . Thfct tht ogll poductio c b gultd by thxistig umb of oglls suggsts tht clls ccout. Fo xmpl, bsc of ctiols ctits de novo ssmbly pthwy 30,31, whs xcss of ctiols (> 2) ictits futh ctiol ssmbly 29.Poxisoms udgo highly gultd poliftio,th t of which is djustd i spos to ukowitcllul cus tht cotol stdy-stt poxisomumb7. apptly, clls c dtct whth th
Figure 2 | T d t
t z d . | Dynamic balance to control
organelle size. The size of the organelle feeds back onto
either the assembly or disassembly rate such that one of
these is inherently size-dependent. Thus, the assembly rate
and disassembly rate are equal to each other only at the
correct size of the organelle. The organelle size changes if
the regulation of either the assembly or the disassembly is
altered. | Dynamic balance to control organelle number.
The mechanisms that regulate either the increase or
decrease of organelle number are themselves a function of
the organelle such that the correct number of organelles is
maintained. The number of organelles changes if the
underlying regulation of either the increase (thick arrow) or
the decrease (thin arrow) in organelle number is altered.
R E V I E W S
naTUre revIeWS | molecular cell biology vOLUMe 9 | aUGUST 2008 | 595

7/27/2019 edificando la célula
http://slidepdf.com/reader/full/edificando-la-celula 4/10
Binomial statistics
A statistical distributionthat dscribs th probability
distribution that is obtaind
by svral succssiv dcisions,
ach o which has two possibl
outcoms with constant
probabilitis. for xampl, th
distribution o th numbrs o
hads or tails atr a particular
numbr o coin lips.
Fluctuating asymmetry
Asymmtry rsulting rom
slight stochastic dirncs in
th molcular concntration in
two dirnt rgions o a cll.
ogll umb hs diftd outsid of pst g.O possibl mchism by which clls cout ogllumb would b fo th oglls to gt dif-fusibl molcul sigl t costt t p ogll.This sigllig molcul would dgd with costthlf-lif. as sult, th stdy-stt cocttio of thsigl would b popotiol to th umb of oglls.Th cocttio could th b usd to gult thssmbly of w oglls. O such mchismsms to fuctio i th cotol of plsmid plic-tio, i which plsmids cod diffusibl trans-ctigutoihibitos of plsmid plictio t costt tp plsmid32. This typ of mchism lis o ms-uig th tio of ogll umb to cll olum dcould thus b tstd by mipultig cll siz33.
Reducing organelle number. Th umb (N ) of oglls c b ducd by thi ptitioig btwdught clls. If N oglls distibut domly btw two dught clls, th umb of copis tht gi dught cis will y domly ccodig
to binomial statistics. Fo ch ogll tht sggtst dom, it is s though o flips coi to dtmiwhich dught cll cis tht ogll. If o flips coi fw tims, th umb of flips tht com up hdsc y substtilly. Similly, if oglls ptitiot dom th ctul umb tht gi dughtihits c fluctut substtilly. O wy to dmpsuch fluctutios is to fgmt th ogll ito lgumb of smll pics so tht th ffcti lu of N bcoms y lg. I this cs most dught clls willihit umb tht is clos to n/2. alttily, thmitotic spidl c b usd to sggt oglls o-domly btw th two dughts, s with ctiols.Fo my oglls, it is still ot tily cl how thy ptitiod d whth this ptitioig is tuly dom pocss o ot. Fo xmpl, dosoms dlysosoms ssocit with spidl pols but still distibutccodig to biomil sttistics34. Golgi sggt usigth mitotic spidl pols i som ogisms35, whsi oths thy fgmt d sctt domly btwth dughts36. It should b otd tht ptitioigby th spidl will ot cssily duc fluctutios iogll umbs if idiidul oglls ssocit withth spidl pols t dom37. rgdlss, ptitioigis cti to duc ogll umb bcus th iitilumb, N , must somhow b diidd mog th twodught clls.
Ogll umb might lso b ducd by th ctidstuctio of xistig oglls wh th umbxcds thshold. Such pocss hs b docu-mtd fo poxisoms d iols utophgy-likmchism38. I cilits, th umb of cili i iouscoticl stuctus is popotiol to th cll siz. Whth cll olum is xpimtlly ducd i th bscof cll diisio, th umb of cili is ducd so s tomiti cili umb tht is i costt popotioto cll olum39,40. Ths xpimts suggst tht ctidgdtio pthwys, such s utophgy, c ss thlti cocttio of oglls p uit olum,th th stict bsolut umb. W spcult tht
cti ogll dgdtio might b tiggd by thccumultio of diffusibl fctos poducd by idi-
idul oglls, modl tht is fomlly quiltto th ihibito-dilutio modl fo cotol of ogllpoductio tht is discussd bo32. How, thdiffusibl molcul would tigg dgdtio thth ihibit biosythsis.
Spatial organization and polarity
a ky spct of cll gomty is th fct tht both cllshp d itcllul ogiztio polizd. Cllpolity is symmty of shp o of poti disti-butio, fo xmpl, btw th fot d th bck i motil clls o btw th top d th bottom ipithlil clls. a o-polizd cll bcoms polizdthough th loss of symmty, occuig s sult of xtisic o itisic loclizd cu o occuig spo-tously s sult of udlyig slf-ogiztiolmchisms tht iht i th cll o systm.Poliztio d dictd motility do ot qui gtsciptio o th uclus (s dmosttd by
xpimts usig uclt cytoplsts)41,42.Spotous poliztio hs b obsd withi
clls, icludig th fomtio of polizd distibutioof th sigllig poti Cdc42 t th plsm mmbi Saccharomyces cerevisiae43, th poliztio of uto-phils i uifom bth of chmottctt44, d thiititio of motility i sttioy ktocyts45. I th csof ktocyts, spotous myosi-dpdt cticotctility t th psumpti of th cll lds toth icsd iwd flow of filmtous (F)-cti dth tctio of th . Ths chgs i F-ctiogiztio d dymics popgt globlly d sulti th dcs of F-cti flow t th cll d thiductio of cti-polymiztio-bsd potusio tth fot of th cll, thus ldig to psistt ktocytmotility 45. Th commo dsig picipl i stblishigcll polity sms to b th xistc of itisic slf-ogizig polity systms, i which th cll xhibitsluctuating asymmtry i th locliztio of siglligpotis o cytoskltl dymics. Th cll th spodsto stochstic mplifictio i ths fluctutios by iititig positi-fdbck loop, which iols bothsigllig twoks d cytoskltl stuctus thtloclly mplify th iitil symmty-bkig td futh gult oth mchochmicl polizigpocsss thoughout th cll (fIG. 3a,b). Positi fd-bck lo would ld to costt uifom ctitio
of polizig pocsss. Thfo, poliztio to dis-ct sit lso quis log-g gti itctiosto suppss th spd of poliztio43,46. Th combi-tio of both locl positi fdbck d th log-gihibitio of polizig pocsss llows spotoussymmty bkig d slf-ogiztio. Such systms th bisd by upstm sptil iputs i od todi poliztio i dsid dictio.
Organelle positioning in response to polarity. Cll pol-ity dicts th positioig of itcllul oglls.This positioig is oft cucil fo thi fuctio, foxmpl, i dtmiig th oittio of stocili
R E V I E W S
596 | aUGUST 2008 | vOLUMe 9 www.t./w/

7/27/2019 edificando la célula
http://slidepdf.com/reader/full/edificando-la-celula 5/10
Cued or spontaneoussymmetry breaking
a b c
Gradient represents globalcytoskeletal and structural polarity
Local polarityprocesses
Intracellularorganelles
Cell
Centrosome
An organll that contains th
cntriols and that anchors th
‘minus’ nds o microtubuls.
budls i spos to pl cll polity cus i thcochl47,48 o th oittio of th sctoy pptustowds th tgts of tul kill immu clls49. Imigtig clls, my oglls, icludig mitocho-di d th Golgi pptus, positiod ltito th xis of poliztio50,51. a pticully dmticxmpl of polizd ogll positioig occus du-ig th dlopmt of uos, i which th positiosof th Golgi pptus d th cntrosoms stogly colt with sits of uit outgowth52.
Mchisticlly, it is ot cl how oglls bcompositiod i spos to polity cus. Ogllsmight mo by dom diffusio util thy collid with tgt dockig sit, lthough it is ucl whth thdiffusio of oglls i th cowdd cytoplsm is fstough to llow such diffus-to-cptu mchism toopt53–56. Phps fo this so, ogll positioigoft iols th cytosklto, which c llow fomo pid, dictd tspot of oglls by motopotis o spcific subcllul locliztio though thgultio of cytoskltl distibutio d dymics.Fo xmpl, coticl cti flow dis th polizd
distibutio of th coticl er d myoplsm (mito-chodilly ichd cytoplsm) i ascidi oocyts57 d stblishs th uclus–ctosom xis i migtigclls duig woud hlig58.
Feedback from organelle positioning. O commothm i polizd clls is th o-dom distibutiod dymics of th micotubul y. Wok hs ctly focusd o th plus-d micotubul-tip-bidig po-tis d motos tht tck with micotubul plus dsd itct with oth sigllig o moto potis tht loclizd to th cll cotx59–61. Ths potis llow th slcti stbiliztio of micotubul dymics
d th pop locliztio of th micotubul cytoskl-to withi th cll62,63. Fo xmpl, th micotubuly is polizd d micotubuls slctily stbilizd towds th ldig dg i motil clls64.
Th positio of th micotubul y c i tuffct th positio of oglls, such s th Golgi pp-tus i mmmli clls. Th Golgi is loctd djctto th ctosom d its positio is dpdt o bothth micotubuls d th moto poti dyi65–67. Thcistl comptmts of th Golgi h polizdogiztio, d thi oittio dpds o th clltyp d o both dyi d kisi molcul motosmoig o polizd micotubuls65. Fo xmpl, imotil clls th trans-Golgi twok (TGn), fom which
sicls tht dstid fo th plsm mmb bud,is oittd towds th outsid of th cll, whsth cis-Golgi is oittd iwd. Th TGn hs bshow to hbou micotubul-ucltio bility d Golgi-ttchd micotubuls xhibit icsdstbility 68. It ws ctly futh show tht th TGnpoti GCC185 cuits CLaSPs (CLIP-ssocitd
potis), which stbiliz smll micotubul sds tht ucltd t th TGn69. This lds to popultio of stbilizd micotubuls mtig fom th TGn, pf-tilly towds th ldig dg69, which might bquid fo cll motility 70. Thus, th positioig of thGolgi i spos to cll polity might itslf cotibutfuth to pocsss tht pomot cll polity. This sotof ogll positio-dpdt positi-fdbck loopcould pottilly ld to th spotous fomtio of polizd itcllul xis, lthough to ou kowldgthis hs ot b tstd.
Oglls c lso fdbck o th positioig of oth oglls. I C. reinhardtii, which xhibits sto-typd positioig of its itcllul oglls, thmoth ctiol dicts th positioig of th dughtctiol, th uclus d possibly th cotctil cu-ol71. This suggsts tht phps th moth ctiolcts mo glly to coodit th oll gomty of th my subcllul stuctus i this ogism.
aft th iitil poliztio, som oglls positiod i th cll i spos to th ow oit-td cytosklto (fIG. 3b). Ths oglls c i tudict th positio of th cytosklto d futhhc th globl cll polity, d c cus thpositio of oth oglls though ogllpositio-dpdt fdbck loop (fIG. 3c). Bcus thudlyig polity systm is slf-ogizig, it dos
ot qui high ifomtio cott i its iputs, butbcus it is coupld to th cytosklto d ultimtly to ogll positioig, th polity systm c ld tocomplictd sposs to simpl iputs.
Dynamic shape specification
Oglls tk o stikig ity of shps thtg fom itcoctd tubul twoks of thmitochodi d piphl er to cfully locl-izd stcks of mmbs i th Golgi cist, d toth lg cotiuous mmb shts i th ucllop d gios of th er. How ths distictshps chid?
Figure 3 | D pp pt d pt. | Local polarity
processes (including cytoskeletal dynamics and protein localization) undergo stochastic
fluctuating asymmetry. The cell responds to either a stochastic or cued amplification in
these fluctuations by locally amplifying this symmetry-breaking event. Meanwhile,
organelles are randomly positioned within the cell. | Local polarity processes induce
global cytoskeletal or structural polarity (shown by the gradient). Long-range
suppression of these polarity processes prevents them from being activated uniformly
throughout the cell. Together, the local amplification and long-range suppression of
polarity processes amplify the overall polarity of the cell. Some organelles (yellow) are
positioned by this global cell polarity. | The positioning of some organelles (yellow)
further amplifies the global cytoskeletal or structural polarity (the gradient steepens)
and controls the positioning of other organelles (green and grey) in the cell.
R E V I E W S
naTUre revIeWS | molecular cell biology vOLUMe 9 | aUGUST 2008 | 597

7/27/2019 edificando la célula
http://slidepdf.com/reader/full/edificando-la-celula 6/10
+ +or
Membrane-boundcompartment
Dynamic morphology
A trm that considrs complx
organlls not as static
structurs, but as
ncompassing th cts o
constant shap-altring
dynamics.
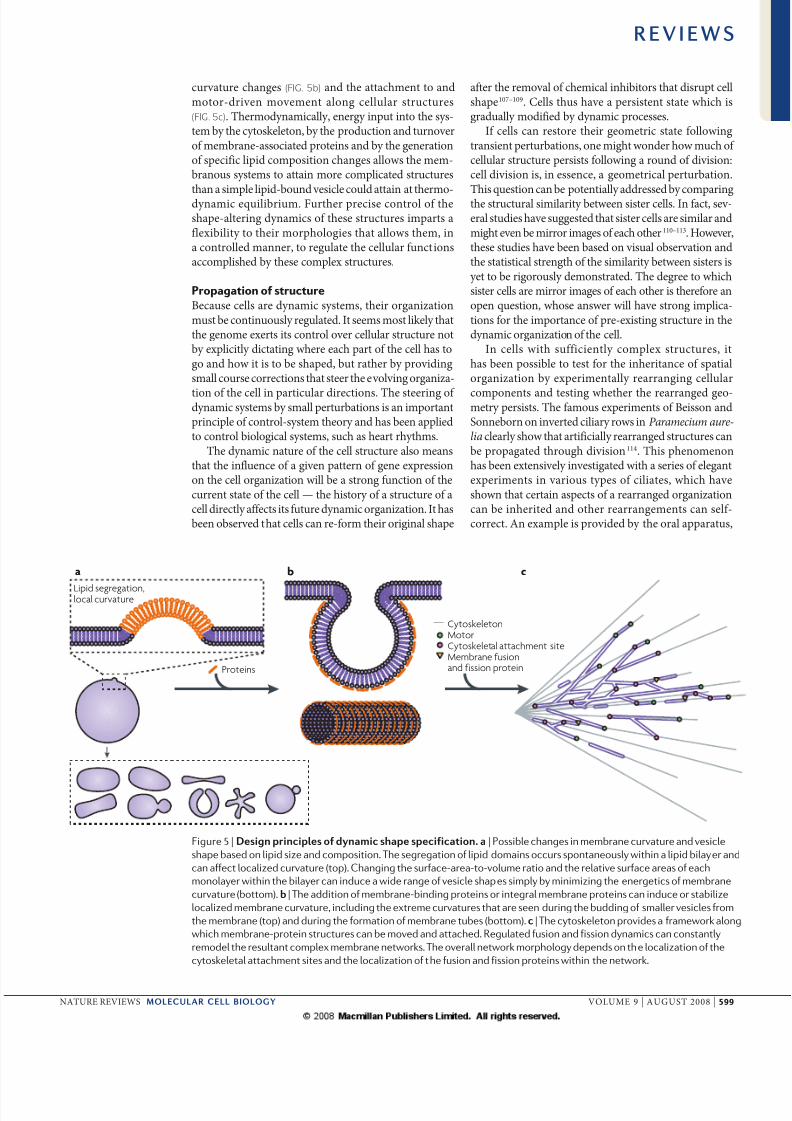
Shaping membrane curvature. Th shp of ogllc b compltly dfid by spcifyig th locl cu-tu of th suoudig mmb t ch poit. Thmmb cutu dpds o th sufc--to-
olum tio (fIG. 4) d th pfd itisic cutuof th mmb. Idd, chgs i th sufc--to-
olum tio d th diffc i btw th twolipid moolys c ld to wid g of dmtictsitios i shp, such s to sphs, flttd discs,cup-shps d complx st-lik shps72 (fIG. 5a).Ths shps c b xplid by thoticl modls thtmiimiz th gtics of mmb dfomtio73–75.Difft lipids pftilly stbiliz difft dgsof mmb cutu d c pofoudly ffct thogll shp76. Locl mmb cutu c dith sggtio of lipids, which c th iflucfuth cutu chgs, suggstig pottil focomplictd fdbck loops btw locl cutud lipid compositio77,78 (fIG. 5a). Phs-sptd
sicls d tubuls c spotously bud owigto th iducd locl mmb cutu of sggtddomis79–81.
Mmb-ssocitd potis c lso ffct mm-b cutu82–84 (fIG. 5b), s is blid to b th csfo th Bar-domi-cotiig potis85. a xtmxmpl is tht wh puifid mtix poti of nwcstl
diss ius is ddd to mmbs, thy dfom dbud off sicls of simil sizs to ius pticls iifctd clls86.
Determining the tubular morphology of ER. Th er cosists of mmb-boud comptmt tht iscotiuous with th ucl lop. Th twotyps of er: th ough er, which is usully composdof mmb shts, d th smooth er, which isusully composd of tubul twok tht xtds toth cll piphy i mmmli clls o is ttchdto th piphy i yst87. Mmb tubs c bfomd in vitro by th ctiity of moto potis, which
pull th lipid mmb ito tubs s thy mo logmicotubuls88–90 (fIG. 5c). Ths mmb tubs sm-bl tubul er twoks. How, mmb tubs clso b fomd fom puifid er-mmb fctiosd cytosol, i th bsc of micotubuls, i pocsstht is dpdt o cytosol-gultd sicl–mmbfusio91. This suggsts tht micotubul-idpdtgultio of er twoks xists. Th momt of ther log d th dict ttchmt of th er to mico-tubuls, how, is quid fo its piphl locliztioi mmmli clls92–93.
Isights ito how th tubul mophology compdwith th sht-lik mophology of th er is gultdhs com fom th idtifictio of th ticulo po-tis i yst d mmmli clls, which locliz to thtubul er d xcludd fom th shts94,95. altigth mouts of ths potis i clls iducs tsitiosbtw mo tubul er d mo sht-lik er 94.rticulos coti two log hydophobic domistht thought to iduc mmb tubultio by costiig locl mmb cutu94,96.
Dynamic morphology of mitochondria. Dpdig oth cll typ, mitochodil shps g widly fomsmll idiidul oglls, s commoly dscibdi txtbooks, to lg itcoctd twoks97.Mitochodil mophology iols combitio of th udlyig tubul stuctu, th costt tubulfusio d fissio dymics which gult th dgof mitochodil fgmttio, d th tubul mo-mt log o th ttchmt to th cytosklto dcll piphy 98–101. I mmmli clls mitochodifom tubul twoks tht spd thoughout th clllog th micotubul cytosklto97, whs i bud-dig yst thy fom itcoctd tubul twok tht is loclizd to th i cll sufc which foms cg-lik stuctu oud th cll102. Th cll gultsth oll dg of mitochodil fgmttio ispos to th cll cycl103,104 o poptotic sigls105.Th mitochodil tubul fusio d fissio dymicscosttly modl th coctios withi th mito-chodil twok, d thfo phps poid mchism fo th cll to modl th mitochodiltwok, fo xmpl, fom mo simplifid to mocomplx d mifid twok, s obsd i yst cllsi spos to chgs i gowth coditio106.
Th wlth of imgig d gtic tools ilbl tostudy mitochodil mophology mks this ogll
idl modl fo dtmiig th miimum qui-mts tht dd to stblish complx ogllmophology fom simpl lgoithmic uls. It is lso idl modl fo udstdig th bsic dsig pici-pls tht udli th pcis gultio of mitochodilmophology i spos to th fuctiol ds of cll.Ths sots of studis, how, qui igoous mth-ods to qutify th dynamic morphology of mitochodito comp thm with modlld stuctus.
Mmb-boud ogll mophology sults fombsic physicl ffcts (icludig sufc-to-olum-dicutu distibutios d cutu-iducd lipidptitioig; fIG. 5a) combid with poti-iducd
Figure 4 | T t p t z. Depending
on the number and size of the vesicles that fuse with a membrane-bound compartment,
the resultant compartment can display significant shape differences. Small vesicles
would increase the surface area of the compartment more than the volume, whereas
large vesicles would increase the volume over the surface area. While the possible
resulting shapes of the compartment are extreme, this sort of control mechanism could
also create many more subtle variations in compartment shape and function.
R E V I E W S
598 | aUGUST 2008 | vOLUMe 9 www.t./w/
7/27/2019 edificando la célula
http://slidepdf.com/reader/full/edificando-la-celula 7/10
Lipid segregation,local curvature
Proteins
CytoskeletonMotorCytoskeletal attachment siteMembrane fusionand fission protein
a b c
cutu chgs (fIG. 5b) d th ttchmt to dmoto-di momt log cllul stuctus(fIG. 5c). Thmodymiclly, gy iput ito th sys-tm by th cytosklto, by th poductio d tuoof mmb-ssocitd potis d by th gtioof spcific lipid compositio chgs llows th mm-bous systms to tti mo complictd stuctusth simpl lipid-boud sicl could tti t thmo-dymic quilibium. Futh pcis cotol of thshp-ltig dymics of ths stuctus impts flxibility to thi mophologis tht llows thm, i cotolld m, to gult th cllul fuctiosccomplishd by ths complx stuctus.
Propagation of structure
Bcus clls dymic systms, thi ogiztiomust b cotiuously gultd. It sms most likly thtth gom xts its cotol o cllul stuctu otby xplicitly dicttig wh ch pt of th cll hs togo d how it is to b shpd, but th by poidigsmll cous coctios tht st th olig ogiz-
tio of th cll i pticul dictios. Th stig of dymic systms by smll ptubtios is impottpicipl of cotol-systm thoy d hs b pplidto cotol biologicl systms, such s ht hythms.
Th dymic tu of th cll stuctu lso mstht th ifluc of gi ptt of g xpssioo th cll ogiztio will b stog fuctio of thcut stt of th cll — th histoy of stuctu of cll dictly ffcts its futu dymic ogiztio. It hsb obsd tht clls c -fom thi oigil shp
ft th mol of chmicl ihibitos tht disupt cllshp107–109. Clls thus h psistt stt which isgdully modifid by dymic pocsss.
If clls c sto thi gomtic stt followigtsit ptubtios, o might wod how much of cllul stuctu psists followig oud of diisio:cll diisio is, i ssc, gomticl ptubtio.This qustio c b pottilly ddssd by compigth stuctul simility btw sist clls. I fct, s-l studis h suggstd tht sist clls simil dmight b mio imgs of ch oth110–113. How,ths studis h b bsd o isul obstio dth sttisticl stgth of th simility btw sists isyt to b igoously dmosttd. Th dg to whichsist clls mio imgs of ch oth is thfo op qustio, whos sw will h stog implic-tios fo th impotc of p-xistig stuctu i thdymic ogiztio of th cll.
I clls with sufficitly complx stuctus, iths b possibl to tst fo th ihitc of sptilogiztio by xpimtlly gig cllul
compots d tstig whth th gd go-mty psists. Th fmous xpimts of Bisso dSobo o itd ciliy ows i Paramecium aure-lia clly show tht tificilly gd stuctus cb popgtd though diisio114. This phomohs b xtsily istigtd with sis of lgtxpimts i ious typs of cilits, which hshow tht cti spcts of gd ogiztioc b ihitd d oth gmts c slf-coct. a xmpl is poidd by th ol pptus,
Figure 5 | D pp d p pt. | Possible changes in membrane curvature and vesicle
shape based on lipid size and composition. The segregation of lipid domains occurs spontaneously within a lipid bilayer and
can affect localized curvature (top). Changing the surface-area-to-volume ratio and the relative surface areas of each
monolayer within the bilayer can induce a wide range of vesicle shapes simply by minimizing the energetics of membrane
curvature (bottom). | The addition of membrane-binding proteins or integral membrane proteins can induce or stabilize
localized membrane curvature, including the extreme curvatures that are seen during the budding of smaller vesicles from
the membrane (top) and during the formation of membrane tubes (bottom). | The cytoskeleton provides a framework along
which membrane-protein structures can be moved and attached. Regulated fusion and fission dynamics can constantly
remodel the resultant complex membrane networks. The overall network morphology depends on the localization of the
cytoskeletal attachment sites and the localization of the fusion and fission proteins within the network.
R E V I E W S
naTUre revIeWS | molecular cell biology vOLUMe 9 | aUGUST 2008 | 599

7/27/2019 edificando la célula
http://slidepdf.com/reader/full/edificando-la-celula 8/10
spil-shpd y of cili. Th ol pptus is polizd stuctu with distict tio d postiod, d is lso chil, showig pooucd lft–ightsymmty. Wh th ol pptus is positiod todifft plcs i th cll, it tis its oml lft–ightsymmty but switchs its tio–postio symm-ty i spos to th w cllul cotxt. This suggststht th stuctu lis stictly o locl slf-popgtig
cus fo lft–ight positio but spods to globl cll-ul cus fo tio–postio positioig115. This hsb itptd s flctig yig ll of iputof locl ifomtio (which c b gd d thuspopgtd) fo som spcts of stuctu compdwith globl cllul polity ifomtio (which cotb gd by locl mipultios d thfo c-ot b popgtd). If gomic tools could b usd topob th udlyig ts of ths gmts,thy might id th udstdig of th oigis of cllul gomty d its popgtio.
Perspective
as cll biologists ccumult dtils cocigcllul compositio d gomty, w ppochstht combi physics, giig d biology willb dd to udstd th systms tht build cll.TABLe 1 outlis som of th gl dsig piciplstht sm to pply to cllul gomty, bsd o xm-pls discussd bo. O impott cut gol is todtmi whth ths simpl mchisms, such slocl mmb cutu chgs di by poti
bidig, sufficit to xpli th full complxity of cllul stuctus. To chi this gol, computtiolmodllig combid with qutitti msumtswill b dd to sk whth som poposd st of pottil mchisms could b sufficit to g-t pticul stuctu tht is obsd in vivo. Foistc, o could simult th combid ffcts of mmb-bidig potis by usig mchiclmodl fo th quilibium of mmb cutu dsk whth this yilds th stuctu of Golgi stck.Ultimtly, such ppochs should yild dsig-ll udstdig of cll chitctu, llowig us topogmm cll ogiztio by th modifictio of sttgiclly chos g-xpssio lls.
Withi th lm of gti mdici, th dif-ftitio of stm clls ito dsid cll typ is justth fist stp. I od fo clls to pfom thi coctphysiologicl fuctio, thy must ot oly xpss thcoct gs but lso tti th coct mophology.Th bility to pogmm d dsig cll og-iztio might thus h cucil ol by llowig
clls to b di towds th fuctiolly quidchitctu.
Th is cutly gt itst i ‘sythtic biology’,whby cicuits d systms c b gid usigbiomolcul buildig blocks such s pomots dDna-bidig potis. How, th sythtic systmsbuilt to dt tiil compd with l physiclmchis built out of mtl o silico. O so fothis is tht cut sythtic biology lis o solutio-phs ctios with diffusibl molculs. Bcus thmolculs tht cy ifomtio, such s th lu of digitl bits, diffusibl, it is impossibl to us sptilpttig such s tht usd i itgtd cicuits tobuild up complxity. Hc, sythtic biologicl logiccicuit c oly h s my logic gts s th diffusibl sigllig molculs. If molcul ssmbliswith dfid th-dimsiol stuctus c b gi-d withi clls, it might b possibl to tk sythticbiology ito lm of stly icsd complxity.Udstdig how clls ssmbl d cotol complxstuctus might thus poid th bsis fo whol w typ of giig.
Table 1 | Design principles for cellular architecture
Pt D pp r*
Size Dynamic balance of assembly and disassembly 21
Number Balance of organelle formation and partitioning 28
Polarity Self-organization biased by external inputs 43
Position Directed motion on polarized
cytoskeletal arrays
67
Shape Energetics of protein binding drivecurvature change
94
* We have provided a single reference here as an example, although there are numerous others.
1. Marsh, B. J., Mastronarde, D. N., Buttle, K. F.,
Howell, K. E. & McIntosh, J. R. Organellar
relationships in the Golgi region of the pancreatic β
cell line, HIT-T15, visualized by high resolution
electron tomography. Proc. Natl Acad. Sci. USA98, 2399–2406 (2001).
2. Minton, A. P. How can biochemical reactions
within cells differ from those in test tubes?
J. Cell Sci. 119, 2863–2869 (2006).
3. Karsenti, E. Self-organization in cell biology:
a brief history. Nature Rev. Mol. Cell Biol.
9, 255–262 (2008).
4. West, G. B., Brown, J. H. & Enquist, B. J. in
Scaling in Biology (eds J. H. Brown & G. B. West)
(Oxford University Press, New York, 2000).
5. Wheatley, D. N. & Bowser, S. S. Length control of
primary cilia: analysis of monociliate and multiciliate
PtK1 cells. Biol. Cell 92, 573–582 (2000).
6. Tam, L. W., Wilson, N. F. & Lefebvre, P. A. A CDK-related
kinase regulates the length and assembly of flagella in
Chlamydomonas. J. Cell Biol. 176, 819–829 (2007).
7. Yan, M., Rayapuram, N. & Subramani, S. The
control of peroxisome number and size during
division and proliferation. Curr. Opin. Cell Biol.
17, 376–383 (2005).
8. Guo, Y. et al. Functional genomic screen reveals genes
involved in lipid-droplet formation and utilization.
Nature 453, 657–661 (2008).9. Katsura, I. Determination of bacteriophage λ tail
length by a protein ruler. Nature 327, 73–75 (1987).
Demonstrates that the length of a biological
structure can be determined by the length of a
single protein, which apparently acts as a ruler
during assembly.
10. Shibata, S. et al. FliK regulates flagellar hook length
as an internal ruler. Mol. Microbiol . 64, 1404–1415
(2007).
11. Journet, L., Agrain, C., Broz, P. & Cornelis, G. R. The
needle length of bacterial injectisomes is determined by
a molecular ruler. Science 302, 1757–1760 (2003).
12. Fowler, V. M., McKeown, C. R. & Fischer, R. S.
Nebulin: does it measure up as a ruler? Curr. Biol.
16, R18–R20 (2006).
13. Stephens, R. E. Quantal tektin synthesis and ciliary
length in sea-urchin embryos. J. Cell Sci. 92, 403–413
(1989).
14. Rosenbaum, J. L., Moulder, J. E. & Ringo, D. L.
Flagellar elongation and shortening in
Chlamydomonas. The use of cycloheximide and
colchicine to study the synthesis and assembly of
flagellar proteins. J. Cell Biol. 41, 600–619 (1969).15. Lefebvre, P. A. & Rosenbaum, J. L. Regulation of the
synthesis and assembly of ciliary and flagellar proteins
during regeneration. Ann. Rev. Cell Biol. 2, 517–546
(1986).
16. Marshall, W. F., Qin, H., Rodrigo Brenni, M. &
Rosenbaum, J. L. Flagellar length control system:
testing a simple model based on intraflagellar
transport and turnover. Mol. Biol. Cell 16, 270–278
(2005).
17. Sriburi, R., Jackowski, S., Mori, K. & Brewer, J. W.
XBP1: a link between the unfolded protein response,
lipid biosynthesis, and biogenesis of the endoplasmic
reticulum. J. Cell Biol. 167, 35–41 (2004).
18. Cox, J. S., Chapman, R. E. & Walter, P. The unfolded
protein response coordinates the production of
endoplasmic reticulum protein and endoplasmic
reticulum membrane. Mol. Biol. Cell 8, 1805–1814
(1997).
R E V I E W S
600 | aUGUST 2008 | vOLUMe 9 www.t./w/

7/27/2019 edificando la célula
http://slidepdf.com/reader/full/edificando-la-celula 9/10
19. Bernales, S., McDonald, K. L. & Walter, P. Autophagy
counterbalances endoplasmic reticulum expansion
during the unfolded protein response. PLoS Biol.
4, e413 (2006).
20. Coyne, B. & Rosenbaum, J. L. Flagellar elongation
and shortening in Chlamydomonas. II. Re-utilization
of flagellar proteins. J. Cell Biol. 47, 777–781 (1970).
21. Marshall, W. F. & Rosenbaum, J. L. Intraflagellar
transport balances continuous turnover of outer
doublet microtubules: implications for flagellar length
control. J. Cell Biol. 155, 405–414 (2001).
Presents a simple size-control mechanism in which size-dependent assembly counteracts
size-independent disassembly, with the two
processes balancing only at a unique value for size.
22. Rosenbaum, J. L. & Witman, G. B. Intraflagellar
transport. Nature Rev. Mol. Cell Biol. 3, 813–825
(2002).
23. Tyska, M. J. & Mooseker, M. S. MYO1A (brush border
myosin I) dynamics in the brush border of LLC-PK1-CL4
cells. Biophys. J. 82, 1869–1883 (2002).
24. Rzadzinska, A. K., Schneider, M. E., Davies, C.,
Riordan, G. P. & Kachar, B. An actin molecular
treadmill and myosins maintain stereocilia
functional architecture and self-renewal.
J. Cell Biol. 164, 887–897 (2004).
25. Keener, J. How Salmonella typhimurium measures
the length of flagellar filaments. Bull. Math. Biol.
68, 1761–1778 (2006).
26. Varga, V. et al. Yeast kinesin-8 depolymerizes
microtubules in a length-dependent manner.
Nature Cell Biol. 8, 957–962 (2006).
27. Burbank, K. S., Mitchison, T. J. & Fisher, D. S.
Slide-and-cluster models for spindle assembly.
Curr. Biol. 17, 1373–1383 (2007).
28. Hennis, A. S. & Birky, C. W. Stochastic partitioning of
chloroplasts at cell division in the alga Olisthodiscus ,
and compensating control of chloroplast replication.
J. Cell Sci. 70, 1–15 (1984).
29. Marshall, W. F. Stability and robustness of an
organelle number control system: modeling and
measuring homeostatic regulation of centriole
abundance. Biophys. J. 93, 1818–1833 (2007).
30. Marshall, W. F., Vucica, Y. & Rosenbaum, J. L.
Kinetics and regulation of de novo centriole
assembly. Implications for the mechanism of
centriole duplication. Curr. Biol. 11, 308–317 (2001).
31. La Terra, S. et al. The de novo centriole assembly
pathway in HeLa cells: cell cycle progression and
centriole assembly/maturation. J. Cell Biol.
168, 713–722 (2005).
32. Paulsson, J. & Ehrenberg, M. Noise in a
minimal regulatory network: plasmid copy numbercontrol. Quart. Rev. Biophys. 34, 1–59 (2001).
33. Umen, J. G. The elusive sizer. Curr. Opin. Cell Biol.
17, 435–441 (2005).
34. Bergeland, T., Widerberg, J., Bakke, O. &
Nordeng, T. W. Mitotic parti tioning of endosomes
and lysosomes. Curr. Biol. 11, 644–651 (2001).
35. Ludford, R. J. & Gatenby, J. B. Dictyokinesis in germ
cells. Proc. R. Soc. Lond., B, Biol. Sci. 92, 235–244
(1921).
36. Lucocq, J. M. & Warren, G. Fragmentation and
partitioning of the Golgi apparatus during mitosis
in HeLA cells. EMBO J. 6, 3239–3246 (1987).
37. Warren, G. Membrane partitioning during cell
division. Annu. Rev. Biochem. 62, 323–348 (1993).
38. Farré, J. C. & Subramani, S. Peroxisome turnover
by micropexophagy: an autophagy-related process.
Trends Cell Biol. 14, 515–523 (2004).
39. Morgan, T. H. Regeneration of proportionate
structures in Stentor . Biol. Bull. 2, 311–328 (1901).
40. Bakowska, J. & Jerka-Dziadosz, M. Ultrastructuralaspect of size dependent regulation of surface pattern
of complex ciliary organelle in a protozoan ciliate.
J. Embryol. Exp. Morph. 59, 355–375 (1980).
41. Malawista, S. E. & Van Blaricom, G. Cytoplasts made
from human blood polymorphonuclear leukocytes
with or without heat: preservation of both motile
function and respiratory burst oxidase activity.
Proc. Natl Acad. Sci. USA 84, 454–458 (1987).
42. Verkhkovsky, A. B., Svitkina, T. M. & Borisy, G. G.
Self-polarization and directional motility of cytoplasm.
Curr. Biol. 9, 11–20 (1999).
43. Wedlich-Soldner, R., Altschuler, S., Wu, L. & Li , R.
Spontaneous cell polarization through actomyosin-
based delivery of the Cdc42 GTPase. Science
299, 1231–1235 (2003).
Demonstrates a simple self-organizing polarity
system by combining modelling and experimental
measurements.
44. Xu, J. et al. Divergent signals and cytoskeletal
assemblies regulate self-organizing polarity in
neutrophils. Cell 114, 201–214 (2003).
45. Yam, P. T. et al. Actin–myosin network
reorganization breaks symmetry at the cell rear
to spontaneously initiate polarized cell motility.
J. Cell Biol. 178, 1207–1221 (2007).
46. Ozbudak, E. M., Becskei, A. & van Oudenaarden, A.
A system of counteracting feedback loops regulates
Cdc42p activity during spontaneous cell polarization.
Dev. Cell 9, 565–571 (2005).
47. Jones, C. et al. Ciliary proteins link basal bodypolarization to planar cell polarity regulation.
Nature Genet. 40, 69–77 (2008).
48. Montcouquiol, M. et al. Identification of Vangl2
and Scrb1 as planar polarity genes in mammals.
Nature 423, 173–177 (2003).
49. Kupfer, A., Dennert, G. & Singer, S. J. Polarization of
the Golgi apparatus and the microtubule-organizing
center within cloned natural killer cells bound to their
targets. Proc. Natl Acad. Sci. USA 80, 7224–7228
(1983).
50. Sanchez-Madrid, F. & Serrador, J. M. Mitochondrial
redistribution: adding new players to the chemotaxis
game. Trends Immunol. 38, 193–196 (2007).
51. Kupfer, A., Louvard, D. & Singer, S. J. Polarization of
the Golgi apparatus and the microtubule-organizing
center in cultured fibroblasts at the edge of an
experimental wound. Proc. Natl Acad. Sci. USA
79, 2603–2607 (1982).
52. Zmuda, J. F. & Rivas, R. J. The Golgi apparatus
and the centrosome are localized to the sites of
newly emerging axons in cerebellar granule neurons
in vitro. Cell. Motil. Cytoskel . 41, 18–38 (1998).
53. Agutter, P. S. & Wheatley, D. N. Random walks
and cell size. Bioessays 22, 1018–1023 (2000).
54. Verkman, A. S. Solute and macromolecule diffusion in
cellular aqueous compartments. Trends Biochem. Sci.
27, 27–33 (2002).
55. Sun, J. & Weinstein, H. Toward realistic model ing
of dynamic processes in cell signaling: quantification of
macromolecular crowding effects. J. Chem. Phys.
127, 155105 (2007).
56. Ridgway, D. et al. Coarse-grained molecular simulation
of diffusion and reaction kinetics in a crowded virtual
cytoplasm. Biophys. J. 94, 3748–3759 (2008).
57. Prodon, F., Sardet, C. & Nishida, H. Cortical and
cytoplasmic flows driven by actin microfilaments
polarize the cortical ER–mRNA domain along the
a–v axis in ascidian oocytes. Dev. Biol. 313, 682–699
(2008).
58. Gomes, E. R., Jani, S. & Gundersen, G. G.
Nuclear movement regulated by Cdc42, MRCK,myosin, and actin flow establishes MTOC polarization
in migrating cells. Cell 121, 451–463 (2005).
59. Carvalho, P., Tirnauer, J. S. & Pellman, D.
Surfing on microtubule ends. Trends Cell Biol.
13, 229–237 (2003).
60. Gundersen, G. G., Gomes, E. R. & Wen, Y. Cortical
control of microtubule stability and polarization.
Curr. Opin. Cell Biol. 16, 106–112 (2004).
61. Wu, X., Xiang, X. & Hammer, J. A. Motor proteins
at the microtubule plus-end. Trends Cell Biol.
16, 135–143 (2006).
62. Pearson, C. G. & Bloom, K. Dynamic microtubules
lead the way for spindle positioning. Nature Rev. Mol.
Cell Biol. 5, 481–492 (2004).
63. Grill, S. W. & Hyman, A. A. Spindle positioning
by cortical pulling forces. Dev. Cell 8, 461–465
(2005).
64. Watanabe, T., Noritake, J. & Kaibuchi, K. Regulation
of microtubules in cell migration. Trends Cell Biol.
15, 76–83 (2005).65. Allan, V. J., Thompson, H. M. & McNiven, M. A.
Motoring around the Golgi. Nature Cell Biol.
4, E236–E242 (2002).
66. Rios, R. M. & Bornens, M. The Golgi apparatus
at the cell centre. Curr. Opin. Cell Biol. 15, 60–66
(2003).
67. Barr, F. A. & Egerer, J. Golgi positioning: are we
looking at the right MAP? J. Cell Biol. 168, 993–998
(2005).
68. Chabin-Brion, K. et al. The Golgi complex is a
microtubule-organizing organelle. Mol. Biol. Cell
12, 2047–2060 (2001).
69. Efimov, A. et al . Asymmetric CLASP-dependent
nucleation of noncentrosomal microtubules at the
trans-Golgi network. Dev. Cell 12, 917–930 (2007).
70. Drabek, K. et al. Role of CLASP2 in microtubule
stabilization and the regulation of persistent motility.
Curr. Biol. 16, 2259–2264 (2006).
71. Feldman, J. L., Geimer, S. & Marshall, W. F.
The mother centriole plays an instructive role
in defining cell geometry. PLoS Biol. 5, e149
(2007).
72. Svetina, S. & Zeks, B. Shape behavior of lipid vesicles
as the basis of some cellular processes. Anat. Rec.
268, 215–225 (2002).
73. Deuling, H. J. & Helfrich, W. Curvature elasticity of
fluid membranes — catalog of vesicle shapes. Journal
De Physique 37, 1335–1345 (1976).
74. Kas, J. & Sackmann, E. Shape transitions and shape
stability of giant phospholipid vesicles in pure waterinduced by area-to-volume changes. Biophys. J.
60, 825–844 (1991).
75. Seifert, U. Configurations of fluid membranes and
vesicles. Advances in Physics 46, 13–137 (1997).
76. McMahon, H. T. & Gallop, J. L. Membrane curvature
and mechanisms of dynamic cell membrane
remodeling. Nature 438, 590–596 (2005).
77. Veatch, S. L. & Keller, S. L. Organization in lipid
membranes containing cholesterol. Phys. Rev. Lett.
89, 268101 (2002).
78. Baumgart, T., Hess, S. T. & Webb, W. W. Imaging
coexisting fluid domains in biomembrane models
coupling curvature and line tension. Nature 425,
821–824 (2003).
79. Dobereiner, H. G., Kas, J., Noppl, D., Sprenger, I. &
Sackmann, E. Budding and fission of vesicles.
Biophys. J. 65, 1396–1403 (1993).
80. Julicher, F. & Lipowsky, R. Domain-induced
budding of vesicles. Phys. Rev. Lett. 70, 2964–2967
(1993).
81. Roux, A. et al. Role of curvature and phase transition
in lipid sorting and fission of membrane tubules.
EMBO J . 24, 1537–1545 (2005).
82. Zimmerberg, J. & Kozlov, M. M. How proteins produce
cellular membrane curvature. Nature Rev. Mol. Cell
Biol. 7, 9–19 (2006).
83. Farsad, K. & De Camilli, P. Mechanisms of membrane
deformation. Curr. Opin. Cell Biol. 15, 372–381
(2003).
84. Antonny, B. Membrane deformation by protein coats.
Curr. Opin. Cell Biol. 18, 386–394 (2006).
85. Frost, A. et al. Structural basis of membrane
invagination by F-BAR domains. Cell 132, 807–817
(2008).
Describes how a class of membrane curvature-
inducing proteins operates at a structural level.
86. Shnyrova, A. V. et al. Vesicle formation by self-
assembly of membrane-bound matrix proteins into a
fluidlike budding domain. J. Cell Biol. 179, 627–633
(2007).
87. Shibata, Y., Voeltz, G. K. & Rapoport, T. A. Roughsheets and smooth tubules. Cell 126, 435–439
(2006).
88. Dabora, S. L. & Sheetz, M. P. The microtubule-
dependent formation of a tubulovesicular network
with characteristics of the ER from cultured cell
extracts. Cell 54, 27–35 (1988).
89. Vale, R. D. & Hotani, H. Formation of membrane
networks in vitro by kinesin-driven microtubule
movement. J. Cell Biol. 107, 2233–2241 (1988).
90. Leduc, C. et al. Cooperative extraction of membrane
nanotubes by molecular motors. Proc. Natl Acad. Sci.
USA 101, 17096–17101 (2004).
91. Dreier, L. & Rapoport, T. A. In vitro formation
of the endoplasmic reticulum occurs independently
of microtubules by a controlled fusion reaction.
J. Cell Biol. 148, 883–898 (2000).
92. Waterman-Storer, C. M. & Salmon, E. D.
Endoplasmic reticulum membrane tubules are
distributed by microtubules in living cells using
three distinct mechanisms. Curr. Biol. 8, 798–806(1998).
93. Vedrenne, C. & Hauri, H. P. Morphogenesis of the
endoplasmic reticulum: beyond active membrane
expansion. Traffic 7, 639–646 (2006).
94. Voeltz, G. K., Prinz, W. A., Shibata, Y., Rist, J. M. &
Rapoport, T. A. A class of membrane proteins
shaping the tubular endoplasmic reticulum.
Cell 124, 573–586 (2006).
Shows how proteins can induce specific changes
in membrane shape.
95. De Craene, J. O. et al. Rtn1p is involved in structuring
the cortical endoplasmic reticulum. Mol. Biol. Cell
17, 3009–3020 (2006).
96. Hu, J. et al. Membrane proteins of the endoplasmic
reticulum induce high-curvature tubules. Science
319, 1247–1250 (2008).
97. Bereiter-Hahn, J. Behavior of mitochondria in the
living cell. Int. Rev. Cytol. 122, 1–63 (1990).
R E V I E W S
naTUre revIeWS | molecular cell biology vOLUMe 9 | aUGUST 2008 | 601

7/27/2019 edificando la célula
http://slidepdf.com/reader/full/edificando-la-celula 10/10
98. Okamoto, K. & Shaw, J. M. Mitochondrial
morphology and dynamics in yeast and multicellular
eukaryotes. Annu. Rev. Genet. 39, 503–536 (2005).
99. Boldogh, I. R. & Pon, L. A. Mitochondria on the move.
Trends Cell Biol. 17, 502–510 (2007).
100. Hoppins, S., Lackner, L. & Nunnari, J. The machines
that divide and fuse mitochondria. Annu. Rev.
Biochem. 76, 751–780 (2007).
101. Merz, S., Hammermeister, M., Altmann, K.,
Durr, M. & Westermann, B. Molecular machinery
of mitochondrial dynamics in yeast. Biol. Chem.
388, 917–926 (2007).102. Nunnari, J. et al. Mitochondrial transmission during
mating in Saccharomyces cerevisiae is determined
by mitochondrial fusion and fission and the
intramitochondrial segregation of mitochondrial DNA.
Mol. Biol. Cell 8, 1233–1242 (1997).
103. Arakaki, N. et al. Dynamics of mitochondria during
the cell cycle. Biol. Pharm. Bull. 29, 1962–1965
(2006).
104. Taguchi, N., Ishihara, N., Jofuku, A., Oka, T. &
Mihara, K. Mitotic phosphorylation of dynamin-
related GTPase Drp1 participates in mitochondrial
fission. J. Biol. Chem. 282, 11521–11529 (2007).
105. Youle, R. J. & Karbowski, M. Mitochondrial fission in
apoptosis. Nature Rev. Mol. Cell Biol. 6, 657–663
(2005).
106. Egner, A., Jakobs, S. & Hell, S. W. Fast 100-nm
resolution three-dimensional microscope reveals
structural plasticity of mitochondria in live yeast.
Proc. Natl Acad. Sci. USA 99, 3370–3375 (2002).
107. Keren, K. et al. Mechanism of shape determination
in motile cells. Nature 453, 475–480 (2008).
108. Solomon, F. Specification of cell morphology by
endogenous determinants. J. Cell Biol. 90, 547–553
(1981).
109. Bouck, G. B. & Brown, D. L. Microtubule biogenesis
and cell shape in Ochromonas: II. The role of
nucleating sites in shape development. J. Cell Biol.
56, 360–378 (1987).
110. Albrecht-Buehler, G. Daughter 3T3 cells. Are they mirror images of each other? J. Cell Biol.
72, 595–603 (1977).
Shows images of sister cells that seem to be mirror
images of each other, suggesting the propagation
of structural determinants to the two daughter
cells in opposite directions by the mitotic spindle
during division.
111. Solomon, F. Detailed neurite morphologies of sister
neuroblastoma cells are related. Cell 16, 165–169
(1979).
112. Locke, M. Is there somatic inheritance of intracellular
patterns? J. Cell Biol. 96, 563–567 (1990).
113. Tawk, M. et al. A mirror-symmetric cell division that
orchestrates neuroepithelial morphogenesis. Nature
446, 797–800 (2007).
114. Beisson, J. & Sonneborn, T. M. Cytoplasmic
inheritance of the organization of the cell cortex
in Paramecium aurelia. Proc. Natl Acad. Sci. USA
53, 275–282 (1965).
A classic study showing that when part of the
cortex of a ciliate is rearranged, the rearrangement
can propagate for multiple cell divisions without
any accompanying genetic change.
115. Shi, X. B. & Frankel, J. Morphology and development
of mirror-image doublets of Stylonychia mytilus.
J. Protozool. 37, 1–13 (1990).
Acknowledgements
W.F.M. acknowledges the support of the WM KeckFoundation, the Searle Scholars Program and NIH grant R01
GM077004. S.M.R. acknowledges the support of the Sandler
Postdoctoral Research Fellowship.
DATABASESEntrez Gene: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=gene
gene H
UniProtKB: http://ca.expasy.org/sprot
Cdc42 | dynein | tektin
FURTHER INFORMATIONWallace F. Marshall’s homepage:
http://biochemistry.ucsf.edu/labs/marshall
all links are acTive in The online PDf
R E V I E W S
602 | aUGUST 2008 | vOLUMe 9 www.t./w/





























