Embed Size (px)
Citation preview


Efectos del estrés abiótico y factores
bióticos en las interacciones planta-
planta: implicaciones para el
funcionamiento y la restauración de los
ecosistemas semiáridos
Autor: Santiago Soliveres Codina1,2
Directores: Fernando T. Maestre Gil1, Adrián Escudero Alcántara1 y Fernando
Valladares Ros2
1Área de Biodiversidad y Conservación. Departamento de Biología y Geología, Escuela
Superior de Ciencias Experimentales y Tecnología, Universidad Rey Juan Carlos.
2 Instituto de Recursos Naturales, Centro de Ciencias Medioambientales, Consejo Superior de
Investigaciones Científicas.
Madrid, 2010


Dr. Fernando T. Maestre Gil y Dr. Adrián Escudero Alcántara, Profesor Titular y Catedrático
de Universidad del Departamento de Biología y Geología de la Universidad Rey Juan Carlos,
respectivamente, y Fernando Valladares Ros, Profesor de Investigación del Instituto de
Recursos Naturales (Centro de Ciencias Medioambientales) del Consejo Superior de
Investigaciones Científicas,
CERTIFICAN:
Que los trabajos de investigación desarrollados en la memoria de tesis doctoral: “Efectos del
estrés abiótico y factores bióticos en las interacciones planta-planta: implicaciones para
el funcionamiento y la restauración de los ecosistemas semiáridos”, son aptos para ser
presentados por el Ldo. Santiago Soliveres Codina ante el Tribunal que en su día se consigne,
para aspirar al Grado de Doctor en Ciencias Ambientales por la Universidad Rey Juan Carlos
de Madrid.
VºBº Director Tesis VºBº Director de Tesis
Dr. Fernando T. Maestre Gil Dr. Adrián Escudero Alcántara
VºBº Director Tesis
Dr. Fernando Valladares Ros

A mi madre, por ser mi inspiración
A Soraya, por ser mi Todo.
La transición de los datos a la teoría requiere imaginación
creativa. Las hipótesis y teorías científicas no se “derivan” de
los hechos observados, sino que se “inventan” para dar cuenta
de ellos.
Carl Hempel

AGRADECIMIENTOS
“Joer, que cuatro años más largos” es lo primero que me viene a la mente al empezar a
escribir estos agradecimientos. Pero después echo la vista atrás y me doy cuenta que he vivido
un montón de experiencias, muchas más buenas que malas, y que he aprendido mucho en este
proceso que es la tesis, he disfrutado muchísimo y me he formado como científico, y aún más
importante, como persona (nunca es tarde para esto último, jeje). Todo esto se lo tengo que
agradecer a la gente que me ha rodeado, apoyado y ayudado durante toda esta importante
etapa de mi vida, a ellos va dedicada esta sección.
En primer lugar les quiero agradecer a mis directores de tesis el haberme brindado esta
oportunidad, el haber trabajado mucho y muy duro para que pudiera colaborar con un montón
de gente distinta, aprender cuanto necesitara en el camino y no tener que preocuparme por
problemas de financiación, algo básico para poder hacer ciencia. Muy especialmente te quería
agradecer a ti, Fernando (Maestre), por tu apoyo continuo y por ser un director inmejorable.
Sin tu ayuda y apoyo, tus innumerables ideas y la inmensa cantidad de horas que has
invertido, mirando y revisando cada uno de los capítulos de esta tesis en cada una de sus
fases, esto no habría sido posible. Pero te quiero agradecer mucho más, el haberme enseñado
una forma de trabajar que no tiene precio, y el haberme dado un ejemplo a seguir como
profesional y como persona. A ti, Adrián, te tengo que agradecer tu infinita paciencia y todos
los comentarios y aportaciones que has hecho a muchos de los trabajos que aquí se presentan.
Me has ayudado muchísimo a entender la ecología de comunidades y muchas otras ecologías.
Las discusiones científicas que hemos tenido y todos tus comentarios al respecto, han sido
todo un placer y un aprendizaje estupendo, sin mencionar la estadística! Y sobre todo me has
enseñado a ser crítico con todo lo que leo y escribo, y a aprender a formarme un criterio
propio, lo cual sin duda me ayudará a ser mejor científico. Fernando (Valladares) es sin duda
el responsable de que yo pudiera tener una beca para poder hacer la tesis. Te agradezco
también que siempre hayas conseguido sacar un rato para ayudarme en muchas etapas de esta
tesis, tus continuas aportaciones a muchos de estos trabajos sin duda han mejorado su calidad.
Dudo que pocas personas sepan más del efecto de la sombra y de ecofisiología en general que
tú, sólo espero haber sido un buen alumno.

A Pablo decirte que has sido un compañero de tesis excepcional, no tengo palabras para
agradecerte tu continuo apoyo a lo largo de toda esta etapa de mi vida. He aprendido
muchísimo de ti y, para mí, eres todo un ejemplo a seguir por muchas cosas. Por tu inmensa
capacidad de trabajo, por no dejarte caer nunca y tirar siempre adelante, fueran cuales fueran
las circunstancias y porque tienes unos coj… como no los tiene nadie. Muchísimas gracias
también por todos los buenos momentos que he pasado contigo, en la uni, en los taludes, en
las fieldtrips, en las roadtrips, y en las trips callejeras y camperas nocturnas y diurnas, y por
acogerme en tu casa siempre que lo he necesitado. También por ser sin duda el maestro de
ceremonias del grupo, por las Aranjuez summer festival, las fiestas en tu casa, por enseñarme
las jam-sessions madrileñas, por las cenas y por todo lo demás, mi experiencia madrileña
hubiera sido mucho peor si no hubieras estado. Espero que nos encontremos muchas veces
más en el camino. Andrea, a ti también te quería agradecer el ser una compañera estupenda, y
tu ayuda en muchas fases de esta tesis. No te creas que me olvidé de quien se quedó conmigo
aquellos primeros días hasta las 10 de la noche cortando los p… protectores para los conejos.
Espero que te vaya muy bien en lo que emprendas en el futuro. Los tres hemos vivido muchos
momentos juntos que recordaré siempre con cariño (eso de subir garrafas monte arriba es que
une mucho, jeje), y que espero que se repitan en el futuro. Me gustaría agradecer a Jorgito
(alias Piruan) por ser un compañero estupendo durante mis primeros pasos en el mundillo de
la ecología, por tu incontenible curiosidad y tus ganas de aprender siempre cosas nuevas.
Tampoco me olvido de que me has apoyado siempre, y has confiado en mí como pocos.
Espero que te esté yendo genial allá por la Puna.
Tengo que decir que estoy tremendamente orgulloso de pertenecer y haber trabajado en el
departamento de Biología y Geología de la URJC. El nivel científico de los Biodiversos es
incuestionable, pero aún es mucho mayor el nivel humano, el buen ambiente que se respira y
lo fácil que es trabajar con todos vosotros. Desde luego hacéis que ir a trabajar cada día sea
más un placer que una obligación. Muchísimas gracias por haberme aceptado y acogido, y
sobre todo, por haberme soportado. Recuerdo cómo Rubén y yo hablábamos en susurros el
primer día, cuando me estaba enseñando los despachos y presentándome a la gente…aquella
fue la única vez que hablé flojo en ese departamento, y sin embargo, todos los biodiversos han
sabido perdonarme (o eso espero)… También espero que sepáis perdonarme mi sentido del
humor algo “brusco” y no siempre acertado, espero que sepáis que siempre fue sin mala
intención. En especial quería agradecerles a mis compis de despacho todos los buenos
momentos que he vivido con vosotros durante estos años. A Cris Escolar, la pequeña del

grupo fernandiano, por tu eterna alegría contagiosa y por ser una compañera genial. A Edu,
aquel señor bajito, con el pelo de dudosa procedencia, y oriundo de “un país chiquito, al ladito
del cielo”. Simplemente, personas como tú hacen que el mundo sea un lugar mejor. Muchas
gracias por preocuparte por todos nosotros siempre, por ser el alma de Biodispersos (Úbeda
queda como testigo) y por no parar de organizar cosas para que, aquellos que estuviéramos
fuera y añoráramos a nuestra gente, nos sintiéramos siempre como en casa. A todos los
demás, Ares, Sonia, Mari Carmen, Samu, Pablo, Alberto, Rubén Milla y Alf (si, si…os
escapasteis, pero para mí seguís siendo de mi despacho), muchas gracias por vuestra paciencia
y ayuda, por todo lo que nos hemos reído y por los buenos recuerdos que me llevo de todos
vosotros. Gracias a todos por hacer de ese despacho un lugar cálido donde diera gusto
trabajar. También quería agradecer a toda la gente que me ayudó en algún momento con el
trabajo de esta tesis, a Kike, Cristina Alcalá, Vicky, Patri, Dolo, Becky, Yoli, Chele, los
Rubenes (Milla y Torices), Mariajo, José Margalet, Luis Giménez, Ozeluí y muchos otros que
seguro que se me olvidan, pido perdón por ello. A Luisiño, por su optimismo y simpatía, y
por ser un gran amigo y compañero. También porque sin él los eventos lúdico-deportivos de
este departamento serían inexistentes. Me gustaría también agradecerles a todos mis otros
compañeros biodispersos el que me hayan ayudado cada vez que los he necesitado, y por
todos los buenos momentos que hemos vivido juntos. A Rocío, María y Mónica, que junto
con Andrea (Javi, Samu y Rubén Torices eran, supuestamente, la parte sensata) han sido el
terror de vigilantes de metro y porteros de discoteca, me lo he pasado genial con todos
vosotros cada vez que nos hemos juntado y espero que nos sigamos juntando muchas veces
más…sea en el país que sea. También a Isa, Ana Millanes, Cris Fernández, Raúl, Javi, Sonja,
Luis Giménez y Julián porque, sin duda, habéis ayudado a pasar estos años en la distancia de
una forma mucho más agradable. No quiero olvidar a los nuevos, Peska, Carlos, Gema, Sonia
(la otra) y los demás, me voy bien tranquilo sabiendo que las nuevas generaciones seguirán
formando un departamento cojonudo donde trabajar y que pasarán muchos años antes de que
haya un cochinillo aburrido en Navidad, espero eso sí, que mejoréis el nivel futbolístico
actual.
Resulta que a las pocas horas de llegar a Móstoles me vi metido en un coche con un tipo
asturiano con barba y pelo largo que me llevaba a ver un piso en la c/ Camino de Humanes,
12. Allí esperaba una andaluza, fumando como un carretero, y con un tembleque extraño en la
pierna. Resulta que Rubén y Mariajo eran esa gente, y que fueron mi familia durante más de 2
años. No puedo dejar de agradeceros que hicierais de esa casa un hogar en el que poder

desahogarme y estar completamente a gusto, vuestro continuo apoyo y el que hayáis sido mis
“hermanos mayores”, lo bien que lo hemos pasado y todo lo que he aprendido y disfrutado
con vosotros. Me alegro mucho de poder teneros como amigos. A la familia tengo que sumar
a Lucia y Horta, ha sido un gustazo conoceros y he disfrutado de cada momento que hemos
pasado juntos.
A toda la gente que ha colaborado en algún capítulo de esta tesis, a Lucia por ayudarme con
toda la parte de dendro y carbohidratos, y por enseñarme lo poco que se, sin tu ayuda el
capítulo 2 no habría sido posible. A Chemi Olano, que con sus ideas y participación hizo que
éste capítulo fuera mucho mejor. David Eldridge (alias Mr. Fantastic), the aussie guy with
italian accent, thanks for all your help and your extraordinary sense of humour; and especially
for completing the aussie gradient!!!. It has been a pleasure to meet and work with you. Matt
Bowker, thanks for a lot of things, it has been wonderful everytime I worked with you and
also the time we spent outside the job. Thanks also for an excellent feedback in our scientific
discussions, for your patience and for introducing me in the causal correlation world. I´m very
grateful because it was you who showed me that stop and think is the first thing you should do
to start doing good science. To the aussie team, Nick Reid and Matt Tighe, for hosting me in
Armidale and helping me with all the logistics during my stay. I especially thank Nick Schultz
for helping me during all the fieldwork in Australia. A Rubén, porque sin él el capítulo 5 no
existiría, por ser tan rápido y eficiente currando y porque ha sido un gustazo descubrir que,
además de un buen amigo, eres muy buen científico. A Estrella, por iniciarme con el mundillo
R y por ayudarme a buscar parcelas en Alicante, y a Bea Amat por echarme un cable con los
árboles de regresión.
A mi gente de Alicante. Pero sobretodo al “Kanutet Vilero”, sin Jorge, Curro, Juande, Joanmi
(gràcies per lo de la portada, rei), Raúl y Juanito mi vida en general, y los cuatro años de la
tesis en particular, no serían lo mismo (no sé si mejor o peor, pero desde luego, mucho más
aburrida). Sólo decir que tengo mucha suerte de teneros como amigos. Muchas gracias por
estar siempre ahí, por perdonarme mis repetidas ausencias, y por estar siempre dispuestos a
pasar conmigo el poco tiempo libre que he podido sacar, por las risas que nos hemos echado,
y por ayudarme en todo cuanto os he pedido. Lo de ayudarme a plantar 2300 árboles el plena
navidad no se me olvidará nunca…aunque con todo el alcohol que me habéis hecho filtrar,
cabrones, ni siquiera puedo entender cómo es que aún recuerdo mi propio nombre! Tampoco
me quiero olvidar de los alicantinos expatriados, Jorgito, JM (ex-expatriados), el Cuñao,

Kiket, Vicen, Carla, Paula y Aitana, gracias por hacer que me sintiera en Madrid como en
casa, o mejor dicho, que me sintiera acompañado fuera de ella.
Sin mi madre nada de esto (ni muchísimas otras cosas) nunca hubiera sido posible, tinc que
agraírte moltes coses, pero sobretot que sempre hages estat ahí, que sigues una lluitadora i tot
un exemple a seguir, i que ho hages donat tot per uns fills que no et mereixen, gràcies mare
PER TOT. También me gustaría agradecerle al resto de mi familia, A Benilde, Juan Antonio,
Jorge y Katia, porque sin su apoyo uno muchas veces no podría seguir adelante. Y como no, a
mi recua de mascotas, las que siguen y las que se fueron, a Stick, Peluso, Lluna, Mac, y a mis
últimos “compañeros de despacho”, a Acho y Legaña. Por estar siempre dispuestos a hacerme
compañía y darme cariño sin pedir nada a cambio, por lo que me habéis hecho reír y disfrutar,
y por enseñarme cosas tan útiles como que, si te echas al suelo en verano, se está más
fresquito que en el sofá. No sabéis leer (creo), pero valéis un imperio.
Y uno siempre se deja lo mejor para el final. A Soraya, por ser mi continuo apoyo y por estar
siempre junto a mí. Por soportar 3 años y medio de distancia y por acompañarme al final del
mundo cuando te lo pedí (11 vuelos en 13 días, y eso que te da miedo volar!). Por dejarte
robar horas por ese infame ordenador, unas horas que me gustaría, y debería, haber pasado
contigo, y nunca reprochármelo (yo sí que lo hago, créeme). Por ayudarme en todo, siempre.
Porque sin tí nada tiene mucho sentido y contigo y tu eterna sonrisa los problemas parecen
unos puntitos lejanos en el horizonte. Y porque no recuerdo haber vivido mejores momentos
en mi vida que los que paso junto a ti, es imposible pensar que haya alguien mejor con quien
uno podría estar.
Seguro que me he dejado a mucha gente, y espero que sepan perdonarme…
A TODOS, GRACIAS POR TODO


ÍNDICE
� Resumen
Antecedentes 3
Objetivos 26
Metodología general y área de estudio 29
Estructura general de la tesis 35
� Capítulo 1
Predicted climate change effects in rainfall regime modulate the outcome of grass-
shrub interactions in two semiarid communities. 39
� Capítulo 2
Spatio-temporal heterogeneity in abiotic factors modulate multiple ontogenetic shifts
between competition and facilitation. 65
� Capítulo 3
Temporal dynamics of herbivory and water availability interactively modulate the
outcome of a grass-shrub interaction in a semiarid ecosystem. 89
� Capítulo 4
On the relative importance of climate and biotic non-trophic interactions as drivers of
local plant species richness 111
� Capítulo 5
On the relative importance of environmental conditions, biotic interactions and
evolutionary relationships as drivers of the structure of semiarid communities. 153
� Discusión y conclusiones generales 185
� Bibliografía y afiliación de los coautores 218


RESUMEN

RESUMEN
2

3
ANTECEDENTES
Las llamadas “tierras secas” (drylands) engloban todos aquellos ecosistemas de ambientes
desde hiperáridos a secos-subhúmedas; representando en total un 41% de la superficie
emergida del planeta (Millenium Ecosystem Assessment [en adelante MEA] 2005, Reynolds
et al. 2007a,b; Fig. A1). Las tierras secas se caracterizan generalmente por tener
precipitaciones escasas y variables, temperaturas aéreas extremas y una evapotranspiración
potencial elevada (Noy-Meir et al. 1973, Whitford 2002, Reynolds et al. 2007b). Estas
características ambientales hacen que estos sistemas tengan una baja productividad, que es
altamente variable dependiendo de las condiciones de cada año y de una alta heterogeneidad
espacial en la disponibilidad de nutrientes y la productividad vegetal (Whitford 2002). Pese a
ello, las tierras secas representan una parte importante de la biodiversidad global (Convención
para la Lucha contra la Desertificación 2005; en adelante CLD), y son el hogar y la fuente de
sustento del 38% de la población mundial (Reynolds et al. 2007a,b). Los impactos
antropogénicos (i.e. cambios en el uso del suelo, sobre-explotación de recursos, aumento de
las infraestructuras) y el aumento de la aridez provocado por el cambio climático son algunas
de las causas más importantes de la degradación de las tierras secas, comúnmente llamada
desertificación (MEA 2005, Reynolds et al. 2005, 2007b). Un aumento del nivel de
degradación implica una pérdida del funcionamiento y de los servicios ecosistémicos,
afectando directamente al bienestar de una parte importante de la población humana (MEA
2005, Reynolds et al. 2007a). Una vez que una zona ha sido degradada, revertir estos cambios
es difícil, ya que se requieren profundas transformaciones socio-económicas que afecten al
desarrollo y manejo de estas áreas, inversiones sustanciales de recursos externos, y un
profundo conocimiento de los factores que condujeron a la merma de la productividad y de
funcionamiento ecosistémico (MEA 2005, Reynolds et al. 2007a,b). Un alto porcentaje de las
tierras secas, hasta el 70% según la CLD, están amenazadas de degradación, mientras que un
10-20% de ellas ya están degradadas en mayor o menor grado (MEA 2005).
Más de las dos terceras partes del territorio español pertenecen a lo que se define como
tierras secas, y hasta el 36% de su territorio está amenazado por la desertificación,
concentrándose la mayoría de este área en la mitad sur Peninsular (Ministerio de Medio
Ambiente, Rural y Marino; Fig A1). Aunque no es la única condición, un mejor conocimiento
sobre cómo funcionan los ecosistemas en las tierras secas, sobre los factores que afectan a su

RESUMEN
4
biodiversidad, y sobre cómo están respondiendo estos ecosistemas al cambio climático o a
incrementos en las perturbaciones que les afectan, y que están asociados con motores de
cambio global (i.e. herbivoría, incendios), nos permitirá desarrollar herramientas para
prevenir y combatir la desertificación (MEA 2005, Reynolds et al. 2005, 2007a). Los
ecosistemas naturales presentan un cierto nivel de resiliencia, de forma que pueden resistir
cierto nivel de perturbación o incremento de estrés sin verse severamente afectados, y
pudiendo recuperarse bajo condiciones ambientales promedio (Noy-Meir 1975, Westoby et al.
1989, Briske et al. 2003); el problema es que, una vez alcanzados los umbrales de resistencia
de estos ecosistemas, éstos pueden sufrir cambios repentinos a estados severamente
degradados, desde donde el retorno puede ser imposible pese a que las condiciones
ambientales que provocaron el tránsito del umbral se modifiquen (Briske et al. 2003, Cortina
et al. 2005, Kefi et al. 2007, Scheffer et al. 2009). Por tanto, es fundamental establecer cuales
son los diferentes umbrales (i.e. niveles de perturbación, aridez) que hacen colapsar los
mecanismos de resiliencia de estos ecosistemas a estados más degradados; como por ejemplo
cambios en la disponibilidad de hábitats (Bascompte y Rodríguez 2001), el colapso de la
expansión de nicho promovida por las plantas nodriza (Michalet et al. 2006), o cambios en el
patrón espacial y tamaño de los parches de vegetación que afecten a la captura y
redistribución de los recursos (Schlesinger et al. 1990, Tongway y Hindley 1995, Kefi et al.
2007). Esta tesis doctoral se centra en los sistemas semiáridos, y por tanto, es sobre ellos en
concreto sobre los que hablaremos a partir de ahora.
LA DINÁMICA Y EL FUNCIONAMIENTO DE LOS SISTEMAS SEMIÁRIDOS
La principal característica de los ambientes áridos y semiáridos es que llueve poco (índices de
aridez [precipitación anual/evapotranspiración potencial] de entre 0,5-0,05), que esta lluvia es
generalmente impredecible, y que normalmente se produce en pulsos más o menos extensos
seguidos de temporadas secas prolongadas (Noy-Meir 1973, Whitford 2002). Por
consiguiente, el reclutamiento de nuevas plántulas sólo se produce durante pulsos de elevada
disponibilidad de agua, que son poco frecuentes e irregulares a lo largo del tiempo; estos
pulsos suelen corresponder con años particularmente benignos desde el punto de vista
climático (p. ej. Eldridge et al. 1991, Escudero et al. 1999, Kitzberger et al. 2000, Whitford
2002, Holmgren et al. 2006). Aunque los pulsos de agua más abundantes y continuos existen,
son los pulsos de agua cortos y de escasa intensidad (<5mm) los que predominan (Whitford
2002, Huxman et al. 2004). La diferente frecuencia entre los distintos tipos de pulsos, y cómo

5
los distintos componentes del ecosistema (i.e. microorganismos del suelo, costra biológica y
distintos tipos funcionales de plantas) aprovechan estos pulsos de agua, son la razón de que
los ecosistemas áridos y semiáridos sustenten niveles de diversidad relativamente altos pese a
su escasa productividad (Sala y Lauenroth 1982, Fowler 1986, Ogle y Reynolds 2004,
Schwinning et al. 2004).
Figura A1 Distribución global de las tierras secas (arriba; Fuente: MEA 2005), y áreas amenazadas de desertificación en España (de verde a amarillo representa de menor a mayor grado de amenaza, imagen de abajo. Fuente: Ministerio de Medio Ambiente, Rural y Marino: http://www.mma.es/portal/secciones/biodiversidad/desertificacion/desertificacion_espnia/index.htm).
La cobertura vegetal discontinua, organizada en manchas discretas de vegetación
embebidas en una matriz de suelo desnudo, con escasa cobertura vegetal (Fig. A2),

RESUMEN
6
característica de muchos ambientes áridos y semiáridos es el origen de la dinámica fuente-
sumidero que define el funcionamiento de estos ecosistemas (Ludwig y Tongway 1995,
Aguiar y Sala 1999, Puigdefábregas et al. 1999). La dinámica fuente-sumidero consiste en el
arrastre de agua, nutrientes y semillas durante los eventos lluviosos de cierta magnitud desde
la matriz de suelo desnudo o de muy escasa cobertura (fuente) hasta las manchas de
vegetación existente (sumideros). Aunque la costra biológica del suelo y los microorganismos
presentes en la matriz de suelo desnudo tienen un papel importante en la captura de carbono y
el reciclado de nutrientes de estos ecosistemas (Belnap y Lange 2003, Huxman et al. 2004,
Castillo-Monroy et al. 2010); la mayor parte de la productividad, reciclado de nutrientes y
captura de carbono ocurre en las manchas de vegetación, dominadas en su mayoría por
arbustos y herbáceas graminoides perennes, que actúan como islas de recursos en un ambiente
mucho más pobre (Franco y Nobel 1987, Schlesinger y Pilmanis 1998, Aguiar y Sala 1999).
La concentración de los recursos en dichas manchas conlleva generalmente un aumento de la
productividad, heterogeneidad ambiental y diversidad a nivel local (Noy-Meir 1973, Huxman
et al. 2004, Schwinning et al. 2004), algo que sería más difícil de alcanzar si los recursos se
repartieran de forma homogénea en el ecosistema (Noy-Meir 1973, Aguiar y Sala 1999). A su
vez, es precisamente esta concentración de los recursos la que mantiene la estructura
discontinua, con la presencia de manchas discretas de vegetación, a largo plazo, ya sea en
áreas donde esta estructura existía de por sí (Ludwig y Tongway 1995, Puigdefábregas et al.
1999, Rietkerk y Van der Koppel 2008), o bien en lugares donde cambios en la composición
vegetal han generado una distribución heterogénea de los recursos (Schlesinger et al 1990,
Archer 1994). La dinámica fuente-sumidero mantiene esta estructura heterogénea mediante
procesos de retro-alimentación positivos a escala de mancha (más recursos, más
productividad en las manchas de vegetación, más captura de recursos), y negativos a mayores
escalas (concentración de recursos y semillas en las manchas de vegetación dificulta el
reclutamiento en sitios libres de vegetación, y por tanto, el desarrollo de patrones espaciales
uniformes a nivel del ecosistema; Schlesinger et al. 1990, Archer 1994, Rietkerk y Van de
Koppel 2008).
Las diferencias en las condiciones ambientales (sombra, fertilidad del suelo,
infiltración), junto con una mayor cantidad de semillas concentradas al quedar atrapadas en
las manchas de vegetación por el efecto de la escorrentía o por la deposición de aves y
mamíferos, hace que sea en estos lugares donde se concentra una gran parte del reclutamiento
de nuevas plántulas (Aguiar y Sala 1999), aunque esto dependerá de los requerimientos
ecológicos de cada especie (p. ej. Miriti et al. 1998, 2007, Caballero et al. 2008). El efecto

7
positivo que tienen unas plantas sobre otras mediante la mejora de las condiciones
ambientales (p. ej. reducción de temperatura y viento, mejora de las condiciones hídricas
mediante sombreo, mayor fertilidad del suelo) y/o la defensa frente a la herbivoría recibe el
nombre de “facilitación” (Callaway 1995; Figura A2), siendo uno de los factores
fundamentales que interviene en el reclutamiento de nuevas plántulas en estas manchas de
vegetación (Niering et al. 1963, McAuliffe 1988, Eccles et al. 1999). Sin embargo, la
intercepción del agua de lluvia por los doseles de las plantas adultas y la mayor competencia
por agua, luz y nutrientes debajo de estos manchas de vegetación, junto con los efectos
negativos de la caída hojarasca, tanto físicos como químicos, relacionados con la liberación
de compuestos alelopáticos, producen efectos negativos que pueden llegar a superar a los
efectos beneficiosos según las especies implicadas y las condiciones ambientales reinantes
(revisado en Fowler 1986, Callaway 2007). Además de esto, las mismas condiciones
ambientales que son beneficiosas para las plántulas (i.e. poca radiación incidente) pueden
resultar negativas para plantas más adultas (Schupp 1995), lo que hace que las interacciones
positivas o facilitativas puedan volverse competitivas a medida que las plantas facilitadas
avanzan en su desarrollo (Fowler 1986, Callaway y Walker 1007, Miriti 2006). Además, lo
que puede ser beneficioso para una plántula en un determinado momento y condiciones puede
ser pernicioso en otros, sin necesidad de cambios ontogenéticos (De la Cruz et al. 2008). El
equilibrio entre facilitación/neutralidad/competencia es un determinante fundamental de la
dinámica de las comunidades vegetales en los ecosistemas semiáridos (Fowler 1986, Aguiar y
Sala 1999). Por tanto, un mayor conocimiento sobre la importancia relativa de las
interacciones positivas frente a las negativas en distintos procesos y atributos ecosistémicos
(i.e. composición, estructura), así como un entendimiento de los condicionantes para que se
de uno u otro signo en la interacción, son fundamentales para poder entender el ensamblaje de
las especies, la dinámica y funcionamiento de estos ecosistemas y mejorar su restauración
(Fowler 1986, Cortina et al. 2005, Callaway 2007, Brooker et al. 2008, Gómez-Aparicio
2009).

RESUMEN
8
Figura A2 Estructura espacial heterogénea con parches de vegetación herbácea y arbustiva en un espartal de Stipa tenacissima en Zorita, España (arriba). Individuos de Austrostipa scabra reclutando bajo un arbusto adulto en un bosque abierto de Eucalyptus populnea en Nyngan, Australia (abajo).

9
LA HIPÓTESIS DEL GRADIENTE DE ESTRÉS Y LA INTRODUCCIÓN DE LA FACILITACIÓN EN LA TEORÍA
ECOLÓGICA
Aunque las interacciones positivas entre plantas son conocidas desde hace tiempo (Clements
1916, Shreve 1931, Niering et al. 1963), han sido ampliamente ignoradas en ecología hasta
hace 25 años (Callaway 1995). Durante los años previos enfatizó el papel de procesos como la
competencia entre plantas, el efecto de las perturbaciones o el estrés abiótico en el ensamblaje
de comunidades (p. ej. Grime 1973, Huston 1979, 1999, Tilman 1988), ignorando la
importancia de la facilitación en este proceso (ver revisión en Callaway 2007). Prueba de ello
es que sólo 27 artículos científicos versaron sobre facilitación en el período 1900-1989,
mientras que el número de artículos sobre este tema en los últimos 20 años asciende hasta
1252 (Pakeman et al. 2009). Además, la facilitación ha sido introducida como un proceso
importante en el ensamblaje de especies en comunidades, especialmente en medios
“estresantes” (i.e. medios áridos y semiáridos, alta montaña, ecosistemas salobres,
comunidades intermareales), ganándose un sitio en el marco de la teoría general ecológica
(Callaway 1997, Hacker y Gaines 1997, Stachowicz 2001, Bruno et al 2003, Lortie et al.
2004, Michalet et al. 2006, Callaway 2007).
Gran parte de este “viraje” en la atención prestada a la facilitación en ecología se la
debemos a Mark Bertness y Ragan Callaway, que en 1994 publicaron su Hipótesis del
Gradiente de Estrés (en adelante SGH, del inglés “Stress-Gradient hipótesis”). Estos autores
propusieron un modelo teórico sencillo y biológicamente muy plausible, en el cual predecían
un aumento de la importancia y la frecuencia de las interacciones facilitativas frente a las
competitivas a medida que el estrés abiótico o las perturbaciones aumentan. A partir de ese
momento, las interacciones positivas pasaron a ser un mecanismo a tener en cuenta a la hora
de estudiar la dinámica de las poblaciones, y luego las comunidades vegetales en todo el
mundo (Callaway 2007). Bajo el paraguas de la SGH, numerosos ecólogos han fijado su
atención en estas interacciones positivas y su efecto en la dinámica de ciertos pares de
especies en particular (p. ej. Valiente-Banuet et al. 1991, Maestre et al. 2001, Gómez-Aparicio
et al. 2004, Sthultz et al. 2007), en el efecto de ciertas especies clave sobre las demás especies
(p. ej. Holzapfel y Mahall 1999, Pugnaire y Luque 2001, Maestre y Cortina 2005, Badano y
Cavieres 2006), o, más raramente, en la importancia de la facilitación a nivel de comunidades
enteras (p. ej. Hacker y Bertness 1999, Kikvidze et al. 2005, Maestre et al. 2010). Además,
esta hipótesis no solo ha sido evaluada en plantas, sino que existen estudios sobre su
aplicabilidad a comunidades intermareales (p. ej. Stachowicz 2001, Kawai y Tokeshi 2007,

RESUMEN
10
Daleo y Iribarne 2009), costra biológica del suelo (Mulder et al. 2001, Maestre et al. 2008,
Bowker et al. 2010) e incluso entre herbívoros de diferente tamaño (Arsenault y Owen-Smith
2002). Prueba de su importancia son las más de 830 citas que ha recibido el artículo de
Bertness y Callaway (1994) hasta la fecha.
A pesar de constituir un punto de partida inmejorable para las investigaciones sobre
facilitación, la SGH ha encontrado numerosas excepciones que hacen poner en duda la
generalidad de sus predicciones. Así, resultados obtenidos cuando se evalúa la actividad
vegetal con diferentes parámetros (Goldberg y Novoplansky 1997, Maestre et al. 2005,
Gómez-Aparicio 2009), se estudian niveles extremadamente altos de estrés (Kitzberger et al.
2000, Tielbörger y Kadmon 2000a, Maestre y Cortina 2004), o se consideran de forma
simultánea distintos factores de estrés (Ibañez y Schupp 2001, Kaway y Tokeshi 2007,
LeRoux and McGeoch 2010) contradicen las predicciones de la SGH. Además, las
interacciones planta-planta son altamente específicas, independientemente del nivel de estrés
en el que ocurran (Callaway 2007). Esto hace pensar que los diferentes rasgos ecológicos de
las especies que interactúan (Choler et al. 2001, Liancourt et al. 2005, Gross et al. 2009,
Maestre et al. 2009a, Gómez-Aparicio 2009) o sus relaciones evolutivas (Valiente-Banuet et
al. 2006, Valiente-Banuet y Verdú 2007, Castillo et al. 2010) son factores fundamentales que
afectan al resultado de estas interacciones. Estos resultados contradictorios han sido el punto
de partida de numerosos debates y redefiniciones de la SGH, que han tratado de incorporar el
efecto de la identidad y características ecológicas de las especies implicadas, los distintos
niveles y tipos de estrés o las distintas medidas de rendimiento vegetal utilizadas (Holmgren
et al 1997, Maestre et al. 2005, 2006, 2009a, Lortie y Callaway 2006, Callaway 2007,
Malkinson y Tielbörger 2010, Holmgren y Scheffer 2010). De todas estas aportaciones se
puede concluir que: a) la facilitación será especialmente intensa y frecuente a niveles
intermedios de estrés; esto puede deberse a que, bajo niveles elevados de estrés, el efecto
negativo de la competencia supera al efecto positivo; o bien a que las plantas nodriza no son
capaces de mejorar suficientemente las condiciones ambientales bajo condiciones tan
extremas (Maestre y Cortina 2004, Michalet et al. 2006, Holmgren y Scheffer 2010), b) el
resultado de las interacciones dependerá de las características ecológicas de las especies
implicadas (competidoras o tolerantes al estrés, sensu Grime 2001) y de si el tipo de estrés
está directamente relacionado con un recurso (p. ej. agua o luz), o no (p. ej. temperatura o
salinidad; Maestre et al. 2009a), c) el resultado de las interacciones dependerá del efecto de la
nodriza sobre la disponibilidad de recursos (p. ej. luz, agua o temperatura), y las tolerancias
relativas de las especies facilitadas (p. ej. tolerancia a la sombra, a la sequía o al frío;

11
Holmgren et al. 1997, Choler et al. 2001, Liancourt et al. 2005, Malkinson y Tielbörger 2010,
Holmgren y Scheffer 2010), d) el resultado de las interacciones bajo distintos tipos de estrés
puede ser de muy diferente naturaleza, pudiendo darse jerarquías entre los distintos tipos de
estrés presentes (Baumeister y Callaway 2006), sinergias o anulaciones de sus efectos (Kawai
y Tokeshi 2007) o bien, simplemente que éstos sean aditivos (Riginos et al. 2005), y e) las
interacciones entre plantas tenderán a ser más positivas cuanto menos relacionadas (i.e. más
distantes en la evolución) estén las especies que interactúan (Valiente-Banuet et al. 2006,
Valiente-Banuet y Verdú 2007, Castillo et al. 2010).
Además de estas complicaciones añadidas a la simplicidad excesiva de la SGH,
hemos de tener en cuenta que, en medios áridos y semiáridos la disponibilidad hídrica y de
nutrientes se produce en pulsos, seguidos de largos períodos de tiempo con una disponibilidad
de agua o nutrientes muy limitada (inter-pulsos; Goldberg y Novoplansky 1997, Whitford
2002), lo que complica aún más la predicción sobre el resultado de las interacciones planta-
planta. Por ejemplo, Goldberg y Novoplansky (1997) desarrollaron un modelo en el que
predecían un efecto negativo de las nodrizas sobre el crecimiento durante los pulsos o épocas
benignas, mientras que este efecto cambiaría a positivo, aumentando la supervivencia, durante
los inter-pulsos o épocas más secas. El resultado final de la interacción dependería de: 1) el
efecto relativo de la competencia con la nodriza frente a los factores abióticos en el
agotamiento de los nutrientes durante los inter-pulsos, y 2) del efecto de la reducción en el
crecimiento durante los pulsos sobre la supervivencia posterior en los inter-pulsos (Goldberg
y Novoplansky 1997). A pesar de su importancia, esta variabilidad intra-anual en los recursos
y su importancia relativa sobre el resultado final de las interacciones planta-planta ha sido
poco estudiada (Barchuk et al. 2005, Kikvidze et al. 2006, Sthultz et al. 2007, de la Cruz et al.
2008).
Los diversos modelos existentes sobre los efectos del cambio climático para la región
Mediterránea semiárida predicen, además de una reducción de la cantidad anual en las
precipitaciones, un cambio sustancial en su patrón temporal (IPCC 2007). En el futuro es
esperable que las épocas secas o inter-pulsos sean más largos, las lluvias durante los pulsos
menos abundantes, y los eventos de lluvias torrenciales más frecuentes (IPCC 2007, Knapp et
al. 2008, Miranda et al. 2009). Es muy probable que estos cambios tengan efectos profundos
sobre la vegetación de los ecosistemas semiáridos (Ogle y Reynolds 2004, Holmgren et al.
2006, López et al. 2008, Heisler-White et al. 2009, Miranda et al. 2009, Pías et al. 2010). Por
consiguiente, es fundamental estudiar cómo las interacciones planta-planta van a verse
afectadas por estos cambios, o cómo pueden mitigar estos cambios aumentando la resiliencia

RESUMEN
12
del ecosistema ante el incremento de aridez y el cambio en el patrón temporal de las
precipitaciones (Brooker et al. 2008). Sin embargo, son muy pocos los estudios que han
evaluado el efecto de estos cambios en la frecuencia y abundancia de los eventos de lluvia
sobre las interacciones planta-planta (Zavaleta 2006, Knapp et al. 2008, Matías 2010).
Ya se ha comentado anteriormente que los cambios en los requerimientos ecológicos
de las distintas especies durante la ontogenia pueden modificar los resultados de sus
interacciones con otras plantas (Callaway y Walker 1997, Miriti 2006). Estos cambios han
sido relacionados con la existencia de interacciones positivas durante la germinación y el
desarrollo de plántula, debido a una mayor vulnerabilidad a la sequía o a la herbivoría
(Schupp 1995, Cavender-Bares y Bazzaz 2000, Ibañez y Schupp 2001) y con el incremento de
la competencia por la luz y el agua a medida que las plántulas crecen y se convierten en
individuos reproductivos (Miriti 2006, 2007, Schiffers y Tielbörger 2006, Armas y Pugnaire
2009). Sin embargo, esta relación no siempre es tan evidente, y parece depender del nivel de
estrés ambiental reinante en cada estado ontogenético y del estado fisiológico (Ibañez y
Schupp 2001, Sthultz et al. 2007, Butterfield et al. 2010), de las diferencias en la forma de
crecimiento de las especies que interactúan (Gómez-Aparicio 2009), o de las relaciones
evolutivas que definen las diferencias entre los rasgos ecológicos entre estas especies
(Valiente-Banuet y Verdú 2008). La inmensa mayoría de los estudios que evalúan cambios
ontogenéticos en las interacciones planta-planta se centran en ventanas temporales concretas a
lo largo del desarrollo de las especies facilitadas (Armas y Pugnaire 2005, 2009, Miriti 2006,
2007, Valiente-Banuet y Verdú 2008). Sin embargo, estas aproximaciones no dan una visión
global del tema, ya que no tienen en cuenta el efecto de arrastre que las condiciones
ambientales de años anteriores pueden tener en el presente. El efecto de las condiciones del
año anterior sobre el crecimiento presente es fundamental en medios semiáridos (Whitford
2002). Por consiguiente, el no tener en cuenta dicho efecto puede confundir la relación entre
el resultado de una interacción planta-planta dada y las condiciones ambientales reinantes en
un año concreto. Estudios que sigan el desarrollo de estas interacciones durante el tiempo
necesario para que estas especies de vida larga pasen de plántula a adulto son logísticamente
prohibitivos, por lo que el uso de medidas indirectas (i.e. estudios dendrocronológicos)
podrían ser una solución adecuada para evaluar la interacción clima-ontogenia en estas
especies (véase Armas y Pugnaire 2005, Sthultz et al. 2007, Aragón et al. 2008, Pías et al.
2010 para otras aproximaciones).
Además, es fundamental que estos estudios a lo largo de distintos estados
ontogenéticos se realicen con distintas combinaciones de formas de crecimiento que

13
interactúen entre ellas (Gómez-Aparicio 2009). Aunque muchas plántulas tienen escasa
tolerancia a la sequía y pueden estar menos estresadas bajo el dosel de una planta adulta
(Schupp 1995 pero ver Caballero et al. 2008, Matías 2010), las diferencias en los
requerimientos ecológicos de individuos adultos entre tipos funcionales o formas de
crecimiento diferentes pueden ser muy importantes. Por ejemplo, distintas tolerancias a la
sombra o la sequía (Holmgren et al. 1997, Hastwell y Facelli 2003) o diferentes
profundidades de enraizamiento (Sala y Lauenroth 1982, Schwinning et al. 2001, Ogle y
Reynolds 2004) pueden ser clave a la hora de definir el resultado de las interacciones entre
adultos de distintas especies. Los estudios sobre cambios de facilitación a competencia a lo
largo de la ontogenia se centran en interacciones arbusto-arbusto, con ambas especies
teniendo características morfológicas y funcionales parecidas, por lo que es esperable que
compartan sus nichos ecológicos de explotación de agua, luz y nutrientes (p. ej. Miriti 2006,
Armas y Pugnaire 2009). Sin embargo, estudios centrados en otro tipo de interacciones (i.e.
herbácea-arbusto o entre arbustos con estrategias y formas contrastadas), muy abundantes por
otro lado en medios semiáridos (Aguiar y Sala 1999), pueden arrojar resultados muy
diferentes. Ello es así debido a que ambas formas de crecimiento difieren en la toma de agua y
nutrientes por sus distintas profundidades de enraizamiento (Sala y Lauenroth 1982, Fowler
1986) y a que tienen alturas contrastadas, lo que puede reducir la competencia entre ambos
grupos por la luz. Por ejemplo, Armas y Pugnaire (2005) encontraron que los individuos
juveniles de la gramínea perenne Stipa tenacissima tenían efectos neutros o negativos sobre el
arbusto Cistus clusii, dependiendo del clima; pero que este efecto negativo se volvía neutro (o
incluso positivo, en condiciones más secas) cuando individuos adultos de ambas especies
interactuaban. En otro estudio, Gasque y García-Fayos (2004) encontraron que tanto las
plántulas como los adultos de Pinus halepensis se desarrollaban mejor cuando crecían cerca
de una macolla de S. tenacissima, sin encontrar cambios en estas interacciones positivas en
distintos momentos del desarrollo de P. halepensis, aunque los efectos positivos de S.
tenacissima sobre las plántulas de P. halepensis desaparecieron durante la sequía estival, ya
que ninguna plántula sobrevivió. Resultados que contrastan con una relación monotónica
entre el paso de facilitación a competencia con la edad (p. ej. Miriti 2006, Valiente-Banuet y
Verdú 2008) no se han encontrado sólo en interacciones leñosa-herbácea, si no que también
son comunes en otro tipo de interacciones. Tirado y Pugnaire (2003) encontraron efectos
facilitativos, independientemente del momento ontogenético, al estudiar la interacción entre
dos arbustos (Asparagus albus y Ziziphus lotus). Sthultz et al. (2007) también encontraron
que el efecto del arbusto Fallugia paradoxa sobre el árbol Pinus edulis era positivo en sitios

RESUMEN
14
de elevado estrés, independientemente de la edad de P. edulis, aunque el efecto positivo de F.
paradoxa se tornaba negativo a medida que P. edulis crecía en sitios menos estresantes. Las
interacciones entre especies herbáceas parecen estar normalmente dominadas por la
competencia (Gómez-Aparicio 2009), aunque esto depende en un grado importante de las
condiciones ambientales reinantes (Graff et al. 2007, Veblen 2008), y con toda probabilidad
de los rasgos funcionales de las herbáceas que interactúen (Cahill et al. 2008, Gómez-
Aparicio 2009). En consecuencia, la relación entre el signo de las interacciones planta-planta,
la ontogenia y las condiciones climáticas parece ser altamente específica de cada especie, o
por lo menos para cada forma de crecimiento (Gómez-Aparicio 2009), por lo que no sirve un
modelo simple para predecir la evolución de las interacciones. Sin duda, estudios destinados a
entender mejor estas relaciones son fundamentales para entender la dinámica de las
comunidades vegetales en los escenarios climáticos presentes y futuros (Fowler 1986,
Butterfield 2009).
EL PAPEL DE LA HERBIVORÍA Y SU INTERACCIÓN CON EL CLIMA EN LAS INTERACCIONES PLANTA-
PLANTA
La herbivoría es un factor fundamental que afecta al desarrollo de las plantas en condiciones
semiáridas (McNaughton 1978, Milchunas et al. 1989, Fowler 2002, Kefi et al. 2007).
Sabemos que la presencia de herbívoros y su preferencia por ciertas especies pueden influir en
la dinámica competitiva entre las plantas (Gurevitch et al. 2000, Fowler 2002), con profundos
efectos en la diversidad y abundancia relativa de las especies vegetales (Westoby et al. 1989,
Fuhlendorf et al. 2001, Briske et al. 2003). También se conoce el efecto que algunas especies
no palatables pueden ejercer en el mantenimiento de la diversidad en las comunidades
vegetales mediante su papel protector/facilitador sobre otras (Hay et al. 1986, Callaway et al.
2000, Rebollo et al. 2002, Baraza et al. 2006, Veblen 2008). De hecho, este efecto protector
puede compensar el efecto negativo derivado de la competencia por agua o nutrientes, dando
como resultado neto una asociación positiva entre pares de especies (Graff et al. 2007). La
herbivoría es, por tanto, un factor determinante de las interacciones planta-planta en
ambientes semiáridos (Rebollo et al. 2002, Baraza et al. 2006, Graff et al. 2007, Veblen
2008).
Sin embargo, al igual que pasa con la mejora microclimática, el efecto protector de
estas plantas nodriza tiene límites, y bajo niveles extremadamente altos de herbivoría u otras
perturbaciones, este efecto positivo puede llegar a desaparecer (Brooker et al. 2006, Smit et

15
al. 2007, 2009, Graff et al. 2007, Forey et al. 2009). En medios semiáridos, la presión de los
herbívoros puede aumentar a medida que la disponibilidad de forraje se reduce con menores
disponibilidades hídricas (Illius y O´Connor 1999, Chase et al. 2000) o por aumento de cargas
de estos herbívoros por dinámicas endógenas; a su vez, las condiciones abióticas pueden
acelerar o retrasar la recuperación de las comunidades vegetales después de la perturbación
(Fuhlendorf et al. 2001). Es lógico pensar que ambos factores, herbivoría y aridez, interactúan
a la hora de definir el resultado de las interacciones planta-planta y la dinámica de las
comunidades vegetales (Van Auken 2000, Roques et al. 2001, Smit et al. 2009). Sin embargo,
a pesar de los numerosos estudios realizados para evaluar las interacciones planta-planta bajo
niveles diferentes de estrés abiótico o herbivoría de forma separada (revisados en Callaway
2007), muy pocos trabajos han evaluado el efecto conjunto que la interacción entre ambos
tipos de estrés produce sobre estas interacciones (Ibañez y Schupp 2001, Veblen 2008,
Anthelme et al. 2009). La co-ocurrencia de ambos tipos de estrés puede afectar de forma
importante a las interacciones planta-planta, provocando profundos efectos en la dinámica de
las comunidades, que son difíciles de predecir si sólo consideramos ambos factores de estrés
por separado (Smit et al. 2009).
Además del efecto protector descrito anteriormente, las plantas nodriza pueden
incrementar la tolerancia de las plantas facilitadas a la herbivoría (Rand 2004, Acuña-
Rodríguez et al. 2006). Las mejores condiciones hídricas y la mayor concentración de
nutrientes presentes frecuentemente bajo el dosel de las plantas nodriza pueden ayudar a la
recuperación de ciertas especies después de la pérdida de biomasa producida por la herbivoría
(Crawley et al. 1998). Sin embargo esto no es tan sencillo, ya que el efecto que unas mejores
condiciones hídricas o nutricionales, pero una menor radiación solar, tengan sobre la
tolerancia a la herbivoría dependerá de cual es el recurso limitante en cada caso, y de cómo la
pérdida de biomasa producida por la herbivoría afecte a la toma de este recurso. Wise y
Abrahamson (2005, 2007) propusieron el Modelo de Recursos Limitados (“Limited Resource
Model”), que predice correctamente la inmensa mayoría de relaciones entre tolerancia a la
herbivoría y disponibilidad de recursos. Este modelo sugiere que la tolerancia a la herbivoría
será mayor en condiciones más húmedas cuando el agua es el factor más limitante (algo
común en sistemas semiáridos), ya que estas condiciones más húmedas favorecen mayores
tasas de fotosíntesis, permitiendo compensar las pérdidas de biomasa ocasionadas por los
herbívoros (Crawley et al. 1998). Sin embargo, en los casos en los que la luz es también un
factor limitante (p. ej. Marañón y Bartolomé 1993, Seifan et al. 2010a), la tolerancia a la
herbivoría será más baja conforme aumente la humedad (Baraza et al. 2004, Wise y

RESUMEN
16
Abrahamson 2005, 2007; Fig. A3). Esto se debe a que la fotosíntesis de estas plantas es
potencialmente más alta bajo condiciones más húmedas, lo que hace que la pérdida relativa de
rendimiento sea mayor cuando la fotosíntesis se ve limitada por la baja disponibilidad de luz
producida por la pérdida de biomasa causada por la herbivoría. En cambio, en condiciones
más secas, el crecimiento vegetal ya está limitado por la baja disponibilidad hídrica, por lo
que el efecto relativo que tiene una menor disponibilidad lumínica en la fotosíntesis es mucho
menor y la tolerancia es más alta (Fig. A3). Por consiguiente, tanto el efecto que las plantas
nodriza tendrán sobre la tolerancia de las plantas facilitadas a la herbivoría, como las
variaciones de este efecto bajo distintos niveles de disponibilidad hídrica son difíciles de
predecir.
Figura A3 Predicciones del Modelo de Recursos Limitados en el caso de que la herbivoría afecte a la toma del recurso principal (Hrp; en nuestro caso el recurso principal sería agua) o a la toma de un recurso alternativo (Hra; en nuestro caso luz). Estas predicciones varían si el nivel inicial del recurso principal es bajo (panel de la izquierda) o alto (panel de la derecha). Modificado de Wise y Abrahamson 2007.
Debido a la gran importancia de ambos factores (herbivoría y agua) en ambientes
semiáridos, parece prioritario llevar a cabo estudios destinados a resolver como las
interacciones planta-planta afectan a la incidencia de los herbívoros sobre las plantas
facilitadas, y a la tolerancia a la herbivoría de estas plantas bajo distintas disponibilidades
hídricas. Esto trabajos son fundamentales para entender la dinámica de las comunidades
vegetales semiáridas (Smit et al. 2009). Este conocimiento puede ayudar a desarrollar
estrategias de manejo adecuadas, que mantengan unos niveles razonables de pastoreo
dependiendo de las condiciones de cada año, permitiendo un desarrollo sostenible en
ambientes semiáridos (el 55% del área ocupada por estos ambientes se destina a la ganadería;

17
MEA 2005). Particularmente, aquellos estudios enfocados en las interacciones leñosa-
herbácea pueden ayudar a predecir el efecto de la herbivoría y el incremento de la aridez en
ecosistemas afectados por procesos de matorralización.
EL PAPEL DE LA FACILITACIÓN EN EL ENSAMBLAJE DE COMUNIDADES
Uno de los debates que más ha fascinado a los ecólogos a lo largo de su historia (ver Gotelli y
Graves 1996, Callaway 1997, 2007, Hubbell 2001, Alonso et al. 2006 para revisiones sobre
este debate) es el iniciado a partir de los trabajos de Clements (1916) y Gleason (1926), los
cuales proponen modelos opuestos sobre la concepción de las comunidades vegetales: un
superorganismo en el cual las interacciones bióticas juegan un papel fundamental (Clements
1916), o bien una coincidencia de especies que coexisten debido a sus adaptaciones
independientes a las condiciones ambientales de cada lugar (Gleason 1926). Resolver este
debate no sólo tiene un atractivo teórico, también tiene implicaciones fundamentales para la
conservación de los ecosistemas naturales. Comprender la naturaleza de una comunidad y ver
las distintas especies como entidades reemplazables y sustituibles, o como organismos con
fuertes relaciones de interdependencia puede conllevar cambios drásticos en como
entendemos y gestionamos la biodiversidad (discutido en Callaway 2007). Existen numerosos
estudios que apoyan ambas teorías, siendo clave la escala temporal, y sobretodo espacial, a la
que estos estudios se realizan (Hubbell 2001, Stokes y Archer 2010).
Estudios conducidos en áreas extensas, a niveles biogeográficos o regionales, parecen
indicar que las interacciones bióticas no son importantes, y que las comunidades por tanto, no
son estables, sino que cambian con el tiempo debido a procesos estocásticos de especiación,
dispersión y extinción (Hubbell 2001, pero ver Gotelli et al. 2010). Sin embargo, estudios a
escalas locales advierten sobre la suma importancia que las interacciones bióticas tienen en el
mantenimiento de estas especies en un lugar dado, así, procesos como la exclusión
competitiva, la segregación de nicho o los mutualismos entre numerosos organismos (p. ej.
insectos polinizadores y plantas) han sido demostrados como clave para mantener el conjunto
concreto de especies que coexisten en una comunidad dada (Levin 1970, Diamond 1975,
Huston 1979, 1999, Rezende et al. 2007), especialmente en aquellas más consolidadas o
menos pioneras (Stokes y Archer 2010). Por otro lado, sabemos que las interacciones
facilitativas son claves para mantener la diversidad en las comunidades naturales (Bruno et al.
2003, Callaway 2007). Así, muchas de las especies nodriza han sido catalogadas como
ingenieros del ecosistema (sensu Jones et al. 1996) en numerosos ecosistemas de todo el

RESUMEN
18
mundo (Hacker y Bertness 1999, Stachowicz 2001, Maestre y Cortina 2005, Badano y
Cavieres 2006). Como ya se ha comentado anteriormente, estas especies modifican las
condiciones microambientales en sus cercanías, lo que permite la existencia de especies
menos adaptadas a las condiciones ambientales particulares de ese sitio, ya que sin la
presencia de estas especies nodriza no podrían sobrevivir (expansión de nicho; Bruno et al.
2003).
Así pues, parece claro que ambas visiones de lo que es una comunidad son
complementarias, y que la escala espacial que consideremos (no tanto la temporal, como se
discutirá en la siguiente sección) es fundamental para entender la importancia de los procesos
estocásticos frente a las interacciones bióticas para el ensamblaje de las comunidades. De
hecho, Hubbell (2001) reconoce que “aunque la asunción de neutralidad completa es, sin
duda, falsa, pocos ecólogos negaran que las poblaciones y comunidades reales no están
sujetas sólo a los factores físicos y las interacciones bióticas, sino también a la estocasticidad
demográfica…Las comunidades ecológicas están indudablemente gobernadas por reglas de
ensamblaje de nicho y dispersión, junto con la estocasticidad demográfica, pero la pregunta
importante es: ¿Cuál es la importancia relativa y cuantitativa de estos procesos?”. Bien, han
pasado casi diez años desde que se hizo esta pregunta, y algo hemos aprendido en el camino.
Tanto las aproximaciones teóricas como empíricas parecen apuntar al mismo sitio: mientras
que son los procesos estocásticos a gran escala (especiación y dispersión) los que determinan
que especies de plantas “aparecen” en una comunidad dada, son los factores físicos (también
estocásticos en algún grado) y las interacciones bióticas los que determinan en mayor parte
cuales de estas especies permanecen (Huston 1999, Lortie et al. 2004a, Rajaniemi et al. 2006,
Rezende et al. 2007, pero ver Gotelli et al. 2010).
Si bien se presupone que tanto los factores ambientales (i.e. clima, perturbaciones)
como las interacciones bióticas condicionan el ensamblaje de comunidades a escalas locales,
poco se sabe sobre la importancia relativa de ambos factores como determinantes de dicho
ensamblaje (Butterfield et al. 2010). Callaway (2007) advierte sobre el efecto tampón que
algunas plantas nodriza pueden tener sobre la variabilidad inter-anual en las precipitaciones
(que podemos considerar estocástica) de algunos desiertos y zonas semiáridas, promoviendo
ambientes más estables en estos ecosistemas. Esta revisión sugiere, por tanto, que las posibles
interacciones entre las condiciones abióticas y las interacciones planta-planta son clave a la
hora de definir la dinámica de las comunidades naturales, ya que pueden modificar los efectos
de la estocasticidad climática en estas poblaciones.

19
Algunos estudios evaluaron la hipótesis de que las interacciones bióticas
(fundamentalmente la competencia) eran clave en los sistemas más productivos, mientras que
las condiciones ambientales, y la falta de adaptaciones fisiológicas de la mayoría de especies a
estas condiciones, determinaban la caída en la riqueza de especies observada en los ambientes
menos productivos (Grime 1973, Huston 1979). De estos estudios se deriva una relación
unimodal entre la riqueza y la productividad de las comunidades naturales, donde en ambos
extremos se vería una caída de la diversidad de especies respecto a niveles intermedios de
productividad, donde los niveles de diversidad serían máximos (Grime 1973). Tanto Hacker y
Gaines (1997) como Michalet et al. (2006) discutieron el papel que podrían jugar las
interacciones facilitativas en esta relación unimodal entre productividad y diversidad. Ambos
trabajos afirman que las plantas nodriza, mediante modificación del microambiente bajo su
dosel, pueden reducir el filtro abiótico que se da en las condiciones menos productivas,
aumentando así la diversidad en niveles medios-altos de “estrés”. Estos trabajos difieren en
que Hacker y Gaines (1997) asumen que la relación entre las interacciones positivas y el
incremento de estrés es positiva y monotónica, mientras que Michalet et al. (2006) apuntan a
un colapso de estos efectos facilitativos bajo niveles extremadamente altos de estrés, donde
incluso el crecimiento de las plantas nodriza, y por tanto su capacidad de modificación del
microambiente, estaría limitado.
Aunque estos modelos sencillos fueron un punto de partida excelente para empezar a
evaluar el papel relativo de los factores físicos y las interacciones bióticas en el ensamblaje de
las especies a nivel local, son insuficientes. Por un lado, se basan en una relación diversidad-
productividad unimodal, la cual no es, ni mucho menos, tan general como se esperaba (Grace
1999, Waide et al. 1999, Gillman y Wright 2006, sólo en 2010 se han publicado en la revista
Ecology cinco estudios discutiendo la generalidad de esta relación). Por otro lado, presuponen
la existencia de un gradiente de “estrés” que afecta de forma general todas las especies de una
comunidad, y que este nivel de estrés aumenta a medida que se reduce la productividad, lo
cual tampoco tiene por qué ser cierto (ver discusión en Körner 2003, 2004, Lortie et al.
2004b). Cada especie presenta unas características propias que le permitirán tener un óptimo
ambiental en unas condiciones particulares (sean estas más o menos productivas); por tanto, a
medida que nos alejamos de estas condiciones ambientales óptimas, esta especie en particular
verá aumentado su nivel de estrés (Chapin et al. 1987, Körner 2003). Sin embargo, las
distintas especies que coexisten en una comunidad difieren en mayor o menor grado en sus
óptimos ambientales y, por tanto, es incorrecto considerar que todas ellas se verán afectadas
de la misma manera a medida que cambien las condiciones ambientales (Chapin et al. 1987,

RESUMEN
20
Lortie et al. 2004b). Por lo tanto, el uso de aproximaciones basadas en el papel de las especies
“tolerantes al estrés” facilitando la existencia de las especies más “competidoras” a medida
que el estrés aumenta (p. ej. Travis et al. 2005) es inadecuado porque estas estrategias
ecológicas cambian con las condiciones ambientales (una especie tolerante a un determinado
factor de estrés no tiene porque ser tolerante a otros tipos de estrés; y una especie competidora
verá modificadas sus habilidades competitivas dependiendo de las condiciones ambientales en
las que se desarrolle). La revisión y cuestionamiento de estos tres supuestos (la existencia de
una relación unimodal entre riqueza y productividad, de un nivel de estrés único que afecte a
comunidades naturales enteras y de estrategias ecológicas que permanecen estables a lo largo
de gradientes ambientales amplios), permitirá explorar alternativas más realistas sobre el
papel que juega la expansión de nicho y las modificaciones microclimáticas promovidas por
las plantas nodriza a lo largo de gradientes ambientales. Sería de esperar, entonces, que el
efecto positivo de las plantas nodriza sobre la diversidad local se extienda con igual
importancia a lo largo de gradientes ambientales amplios, ya que este efecto positivo afectará
al mismo número de especies, aunque su identidad (tanto de las nodrizas como de las
facilitadas) vaya cambiando a medida que cambien las condiciones ambientales y unas
especies u otras se alejen de su óptimo ambiental (Greiner la Peyre 2001, Choler et al. 2001,
Liancourt et al. 2005, Chu et al. 2008, Holmgren y Scheffer 2010). La superación de los
supuestos aludidos debería tener profundas implicaciones en nuestra forma de ver la
importancia relativa de las interacciones positivas a lo largo de gradientes ambientales,
haciendo innecesario hablar de un nivel de estrés único a nivel de comunidad, ya que este
nivel cambiará con cada especie y condición ambiental. Esta es quizás, la razón fundamental
de los resultados contradictorios sobre los cambios en el signo y la intensidad de las
interacciones planta-planta a lo largo de gradientes ambientales (Maestre et al. 2005, 2006,
Lortie y Callaway 2006). Asimismo, la frecuencia e importancia de las interacciones positivas
a nivel de comunidad deberían mantenerse estables a lo largo de gradientes ambientales
amplios, ya que la identidad, pero no la cantidad, de las especies facilitadas es lo que
cambiará a lo largo de estos gradientes. Sin duda, estudios diseñados para evaluar estas
predicciones serán de ayuda para finalmente entender tanto el papel relativo de los factores
físicos y las interacciones bióticas en el mantenimiento de la diversidad local, como la
relación de esta diversidad con la productividad (Mulder et al. 2001, Callaway 2007).
La inmensa mayoría de las aproximaciones que versan sobre el efecto de la
facilitación en las comunidades naturales, tanto teóricas como empíricas, se centran en los
niveles medios-altos de “estrés” (Hacker y Gaines 1997, Lortie et al. 2004a, Travis et al.

21
2005, Michalet et al. 2006, Callaway 2007), otros estudios revelan que los efectos de ciertas
plantas sobre sus vecinas pueden extenderse en condiciones muy productivas (Levine 1999,
Laird y Schwamp 2006, 2009, Brooker et al. 2008), donde cabría esperar que la exclusión
competitiva jugara un papel fundamental (Grime 1973, 2001). Uno de los mecanismos
propuestos (la complementariedad de nicho, descrito a continuación) no puede considerase
como facilitación, ya que no implica que una determinada especie se beneficie por la
presencia de otra. En ambientes heterogéneos, una mayor diversidad de especies, o grupos
funcionales distintos, puede llevar a una mayor y más eficiente explotación de los recursos,
aumentando la productividad de la comunidad a mayores niveles de diversidad debido a la
complementariedad de nicho (p. ej. Hector et al. 1999).
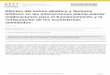
Figura A4 Quizás el ejemplo más conocido de redes de competencia intransitiva es el juego de “Piedra, papel o tijeras”. Podemos observar como la complejidad de las redes intransitivas aumenta a medida que añadimos más actores (especies) al juego. Este tipo de redes de competencia intransitiva es más probable que ocurran en las manchas de vegetación cuanto mayor sean la riqueza de especies y la heterogeneidad en los recursos por los que compiten. Esto puede encontrarse a medida que nos movemos desde áreas de suelo desnudo hacia las manchas de vegetación de mayor complejidad (imágenes de la parte de debajo de la figura).
Sin embargo, otros mecanismos sí que están directamente relacionados con las
interacciones positivas entre plantas. Entre ellos destacan la facilitación indirecta, es decir el
efecto positivo de una especie determinada sobre otra, mediado por el efecto negativo de la
primera sobre una tercera especie (Levine 1999, Callaway 2007, Brooker et al. 2008, Cuesta

RESUMEN
22
et al. 2010). Si imaginamos una sencilla comunidad de tres especies (A, B y C), donde hay
una jerarquía competitiva marcada (A>B>C), entonces, es fácil de imaginar que A puede
facilitar a C mediante su efecto negativo sobre B. Otro mecanismo que atañe a las
interacciones positivas de una manera indirecta es la competencia intransitiva, esto es, la
inexistencia de una jerarquía marcada en las habilidades competitivas de las especies que
coexisten (Gilpin 1975). Si volvemos a nuestra comunidad de tres especies, será fácil de
entender que si A>B>C>A entonces el balance competitivo está más equilibrado y se pueden
mantener mayores niveles de diversidad (Laird y Schwamp 2006, 2009, Bowker et al. 2010;
Fig. A4). Esta competencia intransitiva sólo puede existir cuando la heterogeneidad en los
recursos y en los grupos funcionales que coexisten permite un equilibrio en las habilidades
competitivas de las especies en una comunidad (Grace 1993, Huston 1999). Ya hemos dicho
anteriormente que las plantas nodriza, y los parches que estas forman, son una de las mayores
fuentes de heterogeneidad en ecosistemas semiáridos (p. ej. Pugnaire et al. 1996a, Tracol et al.
2010). Bajo el dosel de estas plantas se dan condiciones heterogéneas de luz, agua, nutrientes
o redes micorrícicas (Pugnaire et al. 1996a, Holmgren et al. 1997, Wolfe et al. 2009 y
referencias en ese texto) que pueden generar las condiciones necesarias de heterogeneidad
para que se de competencia intransitiva o, alternativamente, segregación de nicho. Ambos
mecanismos pueden promover un aumento de la diversidad de especies que coexisten bajo su
dosel (Grace 1993, Pugnaire et al. 1996a, Hastwell y Facelli 2003, Silvertown 2004, Maestre
y Cortina 2005, Badano y Cavieres 2006, Laird y Schwamp 2006). Así pues, es probable que
se den procesos de retroalimentación positiva entre ambos procesos (más heterogeneidad y
más diversidad generan competencia intransitiva o segregación de nicho, que a su vez
aumentan la diversidad) que aumenten de forma desproporcionada la diversidad local de las
comunidades vegetales. No obstante, hasta la fecha sólo hay un estudio que evalúe el efecto
de las plantas nodriza sobre la dinámica competitiva de sus especies facilitadas (Tielbörger y
Kadmon 2000b), y no se ha evaluado este efecto conjuntamente con otros mecanismos como
la mejora microambiental y expansión de nicho. Estudios que evalúen los efectos de las
plantas nodriza sobre la riqueza de especies local, teniendo en cuenta en un mismo marco
general todos los posibles mecanismos por los que estas plantas pueden aumentar la
diversidad (expansión de nicho, competencia intransitiva o segregación de nicho), son
necesarios para entender finalmente el papel de las interacciones planta-planta en la
diversidad local y, por tanto, en la productividad y el funcionamiento ecosistémico (Mulder et
al. 2001, Hooper et al. 2005) a lo largo de gradientes ambientales (Callaway 2007).

23
EL PAPEL DE LA FACILITACIÓN EN LA EVOLUCIÓN DE LAS COMUNIDADES VEGETALES
Aunque se ha sugerido que las interacciones bióticas no son importantes en comparación con
los procesos estocásticos de especiación y dispersión a lo largo de escalas de tiempo
evolutivas (Hubbell 2001), numerosos estudios indican lo contrario (p. ej. Bascompte 2009).
Ejemplos clásicos de ello son los procesos de coevolución que pueden existir entre diferentes
especies de plantas y sus animales asociados, ya sean polinizadores o herbívoros (Darwin
1859). Rezende et al. (2007) encontraron una señal filogenética clara en redes de
interacciones animal-planta. Sus resultados indican que las interdependencias entre ambos
tipos de organismos pueden llevar a procesos de coextinción cuando una de las especies
desaparece, mostrando un alto grado de dependencia interespecífica que parece extenderse
durante toda la historia evolutiva de las especies que forman las comunidades naturales. Estos
procesos de coevolución han sido demostrados también en las interacciones entre plantas. Por
ejemplo, Callaway y Aschehoug (2000) evaluaron las interacciones entre Centaurea diffusa,
una planta nativa de Asia, e invasora en Estados Unidos, y sus vecinas en ambas regiones. En
este estudio vieron como, al añadir carbón activo para secuestrar los compuestos alelopáticos,
no se encontró ninguna diferencia entre las vecinas asiáticas (que por otro lado eran menos
sensibles a la competencia con C. diffusa). Sin embargo, las vecinas americanas
experimentaron un menor efecto competitivo de C. diffusa al añadir carbón activo, lo que
demuestra que estas especies estaban menos adaptadas a los compuestos alelopáticos. Estos
resultados sugieren que las vecinas asiáticas habían experimentado algún grado de adaptación
a estos compuestos debido a la coexistencia con C. diffusa. Pero sin duda, el mejor ejemplo de
interdependencia entre plantas a lo largo de escalas de tiempo evolutivas lo encontramos en
Valiente-Banuet et al. (2006). Estos autores encontraron que las especies de origen Terciario
(condiciones más húmedas que las actuales) dependen de la presencia de especies originadas
durante el Cuaternario (condiciones más áridas) para mantener su nicho de regeneración en
diversos ecosistemas Mediterráneos. La conclusión de estos autores fue que la mejora
microclimática promovida por las especies del Cuaternario ha sido clave para mantener a las
especies del Terciario en ambientes a los que no estaban adaptadas, lo que indica que las
interacciones planta-planta son fundamentales para mantener la diversidad de las
comunidades naturales a lo largo de escalas de tiempo evolutivas.
El reciente desarrollo de las filogenias moleculares ha permitido a los ecólogos evaluar
el efecto de distintos mecanismos (i.e. interacciones bióticas, factores físicos) en el
ensamblaje de las comunidades a lo largo de estas escalas de tiempo evolutivas (Webb et al.

RESUMEN
24
2002, Cavender-Bares et al. 2009). Si asumimos que muchos rasgos ecológicos están bien
conservados a lo largo de la evolución (Herrera 1984, 1992, Ackerly 2003, Valiente-Banuet et
al. 2006), una dominancia de las interacciones competitivas producirá la coexistencia de
especies con rasgos marcadamente diferentes, que permitan cierta segregación de nichos
ecológicos para su coexistencia; por tanto, el patrón filogenético de esa comunidad será más
disperso o equitativo de lo que sería esperable por azar (Webb et al. 2002). En cambio, si los
filtros abióticos son de mayor importancia para esa comunidad, el conjunto de especies que la
formen se caracterizará por una cierta homogeneidad en sus rasgos ecológicos, que
corresponde con sus adaptaciones morfológicas y fisiológicas a esas condiciones ambientales;
por tanto, el patrón filogenético de esa comunidad será más agregado de lo que sería esperable
por azar (Webb et al. 2002, Pausas y Verdú 2007). Esta asunción sencilla fue el punto de
partida de la explosión de estudios que durante los últimos diez años han tratado de inferir los
mecanismos dominantes en el ensamblaje de una comunidad dada a partir su patrón
filogenético (revisado en Cavender-Bares et al. 2009, Vamosi et al. 2009).
Estudios recientes advierten sobre otros posibles mecanismos que pueden afectar al
patrón filogenético de una comunidad y que antes no habían sido considerados. Por citar
algunos ejemplos, la preferencia de herbívoros o polinizadores por taxones filogenéticamente
relacionados (Webb et al. 2006, Sargent y Ackerly 2008), la escala a la que se realice el
estudio (Cavender-Bares et al. 2006, Kraft et al. 2007, Kraft y Ackerly 2010), diferencias en
el nicho de regeneración o las habilidades competitivas entre las especies que coexisten
(Myfield y Levine 2010), o las interacciones positivas entre plantas (Valiente-Banuet y Verdú
2007, Verdú et al. 2009), son algunos de los mecanismos que pueden afectar a la estructura
filogenética de las comunidades. Es por ello que, para inferir los mecanismos de ensamblaje a
partir de patrones filogenéticos se recomiendan medidas complementarias de otros procesos,
como patrones de co-ocurrencia (indicador de interacciones bióticas positivas y negativas;
Tirado y Pugnaire 2005), variables físicas (filtros abióticos) o la conservación de rasgos
ecológicos importantes a lo largo de la evolución (Cavender-Bares et al. 2009, Pausas y
Verdú 2010). Sin embargo, estudios que incluyan las medidas de estos otros mecanismos y las
posibles interacciones entre ellos, tanto a nivel de comunidad como a nivel de especie, son
aún muy escasos pese a que las interacciones entre algunos de esos procesos son clave para el
ensamblaje de las comunidades semiáridas (Holmgren y Scheffer 2010, Butterfield et al.
2010).
La idea de Darwin (1859) en relación a que las especies más parecidas necesariamente
tenían que competir de una forma más intensa ha permeado en la teoría ecológica durante 150

25
años (Webb et al. 2002, Cahill et al. 2008). Sin embargo, a pesar de sus profundas
implicaciones para la diversidad local y el ensamblaje de las comunidades naturales, esta idea
ha sido pobremente estudiada experimentalmente (Valiente-Banuet et al. 2006, Valiente-
Banuet y Verdú 2007, 2008, Cahill et al. 2008, Castillo et al. 2010). Cahill et al. (2008) no
encontraron ninguna relación entre la distancia filogenética y el efecto de la competencia al
evaluar una base de datos amplia que incluía la relación entre 50 especies objetivo y 92
especies competidoras distintas. Ellos atribuyeron esta falta de relación a la diferencia entre
las interacciones entre mono- y dicotelodóneas, ya que la intensidad de la competencia
aumentaba con la distancia filogenética para las monocotiledóneas, ocurriendo lo contrario
con las dicotiledóneas. Los trabajos de Valiente-Banuet y colaboradores (2006, 2007, 2008) y
Castillo et al. (2010), conducidos en su mayoría con especies dicotiledóneas, concluyen que la
competencia disminuye también con la distancia filogenética entre dos especies, siendo más
probable que se den interacciones positivas entre especies distanciadas en la evolución. Por
tanto, la idea de Darwin parece confirmarse en la mayoría de casos estudiados, al menos para
plantas dicotiledóneas. De estos estudios se concluye, por tanto, que la relación evolutiva es
clave para decidir el resultado de la interacción entre dos especies. Sin embargo, se ha
discutido con anterioridad en este texto, y durante 20 años en la literatura ecológica en
general, que las condiciones ambientales son fundamentales para definir el resultado de estas
interacciones. Entonces, ¿cuál es la importancia relativa de las condiciones ambientales frente
a las relaciones evolutivas a la hora de definir el resultado de las interacciones planta-planta?,
¿interactúan ambos factores a la hora de definir estos resultados? Hasta la fecha ningún
estudio se ha planteado responder a estas preguntas, las cuales son clave para establecer la
importancia de las relaciones planta-planta en el ensamblaje de las comunidades bajo distintas
condiciones ambientales y a lo largo de escalas de tiempo amplias. Asimismo, estudios
enfocados en la interacción entre la distancia filogenética entre las especies implicadas y el
clima en el que se dan estas interacciones nos pueden ayudar a mejorar nuestras inferencias
sobre los procesos reinantes en el ensamblaje de una comunidad a partir del estudio de los
patrones filogenéticos.

RESUMEN
26

27
OBJETIVOS
El objetivo general de esta tesis es evaluar el efecto de distintos niveles de estrés, tanto biótico
como abiótico, en el resultado de las interacciones entre pares de especies vegetales,
estudiando también cómo estas interacciones y los factores climáticos afectan a la diversidad
local de especies y a la estructura filogenética de las comunidades vegetales en medios
semiáridos. Para poder conseguir este objetivo general se han desarrollado siete objetivos
específicos, que se describen a continuación y se abordarán en los cinco capítulos que
conforman el cuerpo de esta tesis doctoral.
OBJETIVOS A NIVEL DE PAR DE ESPECIES
• Evaluar el efecto del cambio en el patrón temporal de las precipitaciones predicho por
diversos modelos de cambio climático en el resultado de la interacción entre Retama
sphaerocarpa (planta facilitada) y distintas especies herbáceas (plantas nodriza) en un
ecosistema natural (espartal dominado por Stipa tenacissima) y uno emergente
(herbazal de talud de carretera; Capítulo 1).
• Evaluar el efecto de la variabilidad espacio-temporal en la disponibilidad de agua
sobre la interacción entre el arbusto gipsófilo Lepidium subulatum (planta facilitada) y
la herbácea perenne S. tenacissima (planta nodriza) a lo largo de diferentes estados
ontogenéticos de L. subulatum (Capítulo 2).
• Determinar el efecto simultáneo de dos factores distintos de estrés (herbivoría y
aridez), así como de su dinámica temporal, en el resultado de la interacción entre R.
sphaerocarpa (planta facilitada) y la herbácea S. tenacissima (planta nodriza; Capítulo
3).
• Evaluar la generalidad de los modelos teóricos existentes para predecir el signo de las
interacciones planta-planta a lo largo de gradientes ambientales en dos regiones
semiáridas contrastadas (Capítulo 4).

RESUMEN
28
• Definir la importancia relativa de las relaciones filogenéticas y las condiciones
ambientales a la hora de definir el signo de las interacciones entre pares de especies
vegetales presentes en espartales de S. tenacissima a lo largo de un gradiente
ambiental amplio (Capítulo 5).
OBJETIVOS A NIVEL DE COMUNIDAD
• Estudiar la importancia relativa de distintos mecanismos de facilitación/competencia
(expansión de nicho, mejora microambiental, competencia intransitiva y segregación
de nicho) y de los factores climáticos, así como la interacción entre ambos, a la hora
de determinar la riqueza local de especies en dos comunidades semiáridas de
características contrastadas a lo largo de gradientes ambientales amplios (Capítulo 4).
• Evaluar la extensión del efecto de las interacciones bióticas, los factores climáticos, y
su interacción, sobre el patrón filogenético en espartales de Stipa tenacissima a lo
largo de un gradiente ambiental amplio (Capítulo 5).

29
METODOLOGÍA GENERAL
Y ÁREA DE ESTUDIO
REA DE ESTUDIO
Salvo dos excepciones (parte de los capítulos 1 y 4), esta tesis doctoral se centra en su
totalidad en los espartales de Stipa tenacissima situados en el centro y sudeste Peninsular.
Este ecosistema es uno de los más representativos de las zonas semiáridas de España y el
Norte de África (LeHoureu 2001). Los espartales se extienden sobre suelos pedregosos,
limosos, arcillosos, calizos o yesosos, en zonas desde el nivel del mar hasta 2000 m de altitud,
y con precipitaciones que pueden llegar hasta los 700 mm, aunque preferentemente se dan en
la franja entre 200-400 mm (revisado en Maestre et al. 2007). Los espartales son formaciones
vegetales abiertas, con coberturas que oscilan entre el 18% y el 60% (Maestre 2002, Ramírez
2006), de estructura y composición heterogéneas (Puigdefábregas y Sánchez 1996,
Puigdefábregas et al. 1999, Maestre et al. 2007). Al igual que otros sistemas semiáridos, la
estructura espacial de la vegetación en los espartales, caracterizada por la presencia de
manchas de vegetación discreta embebidos en una matriz de suelo desprovisto de plantas
vasculares, generan una dinámica fuente-sumidero que resulta clave en la dinámica hídrica y
ecológica de estas comunidades (Puigdefábregas et al. 1999, Maestre y Cortina 2004c,
Ramírez y Bellot 2009).
Los espartales han sido intensamente manejados por el hombre desde hace no menos
de 4000 años, principalmente para la explotación de fibras vegetales (Barber et al. 1997). Sin
embargo, la llegada de las fibras sintéticas y el abandono general del campo que ocurrió en
España a partir de los 1960s, promovió el cese del manejo humano de estos ecosistemas
(Maestre et al. 2007). Este cese ha provocado la recolonización, aunque muy lenta y poco
abundante, de los arbustos rebrotadores típicos de estos climas, que anteriormente eran
eliminados por su posible efecto negativo sobre el crecimiento del esparto (Cortina y Maestre
2005, Maestre et al. 2007, Maestre et al. 2009b). Así, arbustos como Pistacia lentiscus,
Quercus coccifera, Rhamnus lycioides o Ephedra fragilis, entre otros, han aumentado
levemente su cobertura en estos ecosistemas desde el abandono de su explotación (Maestre et

RESUMEN
30
al. 2007). Estos arbustos, a pesar de representar una parte pequeña en cuanto a cobertura en
estos ecosistemas, juegan un papel fundamental, ya que incrementan la heterogeneidad y
diversidad local (Cortina y Maestre 2005, Maestre y Cortina 2005), y afectan positivamente a
la fertilidad y el funcionamiento ecosistémico (Pugnaire et al. 1996a, Caravaca et al. 2003,
Maestre et al. 2009b). A diferencia de estos arbustos, que extienden su sistema radicular no
sólo bajo su dosel, si no también en las áreas de suelo desnudo circundantes, el esparto centra
sus raíces exclusivamente bajo su dosel, y a profundidades inferiores a 40 cm de profundidad,
dependiendo su rendimiento en gran parte de su capacidad para recoger el agua de escorrentía
generada durante los eventos de lluvia (Puigdefábregas et al. 1999; pero véase Ramírez et al.
2007 para una visión alternativa). Esta capacidad de capturar el agua de escorrentía, junto con
los efectos que su dosel produce sobre la reducción de la radiación incidente y la demanda
evaporativa, son claves para entender el efecto positivo que el esparto produce sobre otras
especies y su papel como “islas de recursos” (Maestre et al. 2001, 2003, Gasque y García-
Fayos 2004, Armas y Pugnaire 2005, Barberá et al. 2006, Navarro et al. 2008).
Los capítulos 1, 2 y 3 de esta tesis se centran en espartales del centro de la Península
Ibérica que crecen en suelos gipsícolas. Estos suelos presentan características químicas
(exceso de iones de sulfato o Calcio, baja retención de agua; Meyer 1986, Escudero et al.
1999, 2000) y físicas (costra superficial dura; Romao y Escudero 2005) que hacen que la
colonización vegetal difícil para muchas especies, siendo su composición florística
particularmente abundante en especialistas de estos sustratos (revisado en Caballero 2006,
Matesanz 2008). Esto, junto con la combinación de características climáticas adversas (estos
suelos se desarrollan sobretodo en medios semiáridos) hace que estos suelos yesosos
presenten una gran cantidad de endemismos, adaptados a las condiciones particulares de estos
suelos y climas. Estas características hacen de los ecosistemas Mediterráneos yesíferos
hábitats de interés para su conservación por la particularidad de las especies que los
conforman (Caballero 2006, Matesanz 2008 y referencias en esos textos). Por tanto, en los
espartales escogidos para el desarrollo de los capítulos 1, 2 y 3 pueden encontrarse especies
propias de los matorrales gipsófilos (especialistas de suelos yesíferos; Helianthemum
squamatum, Lepidium subulatum, Centaurea hyssopifolia) y “gipsovags” (generalistas que
pueden vivir en suelos yesíferos; Retama sphaerocarpa, Rosmarinus officinalis o Thymus
vulgaris). En cambio, los capítulos 4 y 5 se centran en los espartales presentes en los suelos
calcáreos desde Guadalajara a Murcia (ver Fig. A5). La amplia zona de distribución de estos
espartales sobre un suelo relativamente homogéneo hace posible la realización de
experimentos observacionales a lo largo de un gradiente climático amplio, que oscila entre

31
13-17 ºC de temperatura media, y 273-488 mm de precipitación media anual, con mucha
mayor cantidad y frecuencia de heladas en el extremo occidental (Guadalajara-Madrid) que en
el oriental (Alicante-Murcia) del gradiente.
Alternativamente, parte de los experimentos de esta tesis doctoral (Capítulo 1) se han
desarrollado sobre herbazales de talud de carretera, un tipo de ecosistema emergente que está
aumentando su importancia a nivel global año a año (García-Palacios et al. 2010). Estos
sistemas se caracterizan por una cobertura herbácea dominada por especies anuales,
generalmente ruderales que presentarán mayor o menor cobertura dependiendo del tipo de
talud (desmonte o terraplén; Matesanz et al. 2006) o de la disponibilidad de agua durante la
germinación y desarrollo de las plántulas (Bochet y García-Fayos 2004; ver García-Palacios
2010 para una revisión extensa sobre la dinámica de estos sistemas emergentes). En estos
herbazales, es común que la sucesión secundaria se vea ralentizada, bien por la escasa
disponibilidad de agua o nutrientes (Bolling y Walker 2000), o bien porque coberturas
herbáceas muy desarrolladas no dejan huecos libres para la colonización de nuevas especies
(Burke y Grime 1996). La introducción de especies leñosas en estos sistemas ha sido
recomendada para acelerar la sucesión secundaría (Booth et al. 1999); sin embargo, sabemos
muy poco sobre como las herbáceas dominantes en este tipo de sistemas afectan al éxito de
estas plantaciones, especialmente bajo diferentes niveles de disponibilidad hídrica. Un mejor
entendimiento de la interacción entre ambos grupos vegetales (herbáceas y leñosas) nos
permitirá conciliar los dos mayores retos en la restauración de estos taludes de carretera: el
establecimiento de una cobertura herbácea suficientemente densa como para prevenir
procesos de erosión (Andrés y Jorba 2000), y el establecimiento de especies leñosas para
acelerar su sucesión secundaria (Jorba y Vallejo 2008).
Por otro lado, en el Capítulo 4 se evalúa el efecto de las interacciones bióticas y el
clima en la diversidad local, no sólo de espartales, si no de ecosistemas semiáridos
australianos. Concretamente se han muestreado comunidades pertenecientes a las alianza de
Eucalyptus populnea y Callitris glaucophylla y de Casuarina pauper y Alectyron oleifolius
(Beadle 1948), comúnmente encontradas en las llamadas “tierras rojas” de este continente.
Estas tierras rojas se caracterizan por tener textura arenosa, suelos profundos y con contenidos
bajos en nutrientes (Isbell 1996). Estos ecosistemas se caracterizaban originalmente por
presentar coberturas herbáceas continuas, con individuos dispersos de Eucalyptus spp. y
algunos arbustos. Sin embargo, la elevada presión ganadera a la que han sido sometidas estas
áreas (Keith 1998) ha incrementado notablemente el reclutamiento de estos arbustos (p. ej.
Geijera parviflora, Eremophylla spp., Callitris glaucophylla, etc.) y ha reducido la cobertura

RESUMEN
32
herbácea. Lo que finalmente ha generado una estructura discontinua, de manchas discretas de
vegetación embebidas en una matriz de suelo desnudo, equivalentes a las de los espartales
anteriormente descritos (Tongway y Hindley 1995). Estas comunidades ocupan una amplia
superficie en el este de Australia, que permitió seleccionar 10 parcelas con vegetación
perteneciente a estas comunidades a lo largo de un gradiente climático amplio (16º–19º C y
280–630 mm) con el objetivo de complementar el muestreo realizado en España para el
Capítulo 4 (descrito con más detalle en la siguiente sección).
Figura A5. Distribución aproximada de las parcelas utilizadas a lo largo del gradiente ambiental referido en los capítulos 4 y 5. El diagrama ombroclimático (Fuente: www.globalbioclimatics.org) y una imagen general de los dos extremos del gradiente junto a una posición intermedia aparecen mostrados en la figura.
El clima de todas las áreas de estudio escogidas para esta tesis es Mediterráneo
semiárido, difiriendo en su grado de continentalidad y en sus precipitaciones medias. En las
parcelas situadas en el centro Peninsular, las temperaturas en invierno son mucho más frías y
las precipitaciones anuales son algo mayores. La lluvia sigue una distribución bimodal, con
máximos marcados en primavera (Abril-Mayo) y otoño (Septiembre-Octubre). La
variabilidad interanual de la precipitación es muy grande, siendo impredecibles tanto la

33
cantidad como el patrón temporal de las lluvias de cada año (revisado en Puigdefábregas et al.
1999; ver figuras con precipitaciones registradas durante el período de estudio en los capítulos
1 y 3). En las parcelas situadas en el sudeste peninsular, las temperaturas son más suaves en
invierno y las precipitaciones medias algo menores. La distribución temporal de los eventos
de lluvia en estas parcelas es más unimodal, centrándose las precipitaciones hacia finales del
verano y comienzos del otoño (Fig. A5). Las precipitaciones en el gradiente australiano
siguen una distribución relativamente homogénea a lo largo del año, con una ligera
predominancia de eventos de lluvia durante el verano (el 60% de estos eventos se concentra
en verano).
METODOLOGÍA GENERAL
En esta tesis doctoral se han realizado experimentos tanto manipulativos como
observacionales, en los que se incluyen distintos niveles de estrés abiótico (generalmente
aridez) y biótico (herbivoría). Todos ellos han sido llevados a cabo bajo condiciones
naturales. Los diferentes niveles de aridez se han conseguido mediante riegos en los
experimentos manipulativos, o mediante el uso de gradientes ambientales amplios en las
aproximaciones observacionales. En éstas últimas se han homogeneizado, tanto como ha sido
posible, la pendiente, orientación, tipo de suelo e historia de manejo previo de las parcelas
seleccionadas, con la intención de evitar la influencia de otros factores que no estuvieran
considerados en el experimento y que pudieran confundir la interpretación de sus resultados.
Los únicos herbívoros considerados han sido los conejos, que eran especialmente abundantes
en las parcelas seleccionadas (véanse los tres primeros capítulos). Su nivel de herbivoría ha
sido controlado mediante mallas de exclusión, combinado con el seguimiento de plantones no
protegidos a lo largo de un año. Los experimentos manipulativos se centran en interacciones a
nivel de par de especies. Las aproximaciones observacionales, en cambio, han sido utilizadas
para evaluar el efecto de estas interacciones tanto a nivel de especie como a nivel de
comunidad. La complejidad de las interacciones bióticas se ha evaluado mediante análisis de
co-ocurrencia (Gotelli 2000) o utilizando distintos indicadores del desarrollo vegetal
(crecimiento, supervivencia y eficiencia fotosintética en los manipulativos, crecimiento o
cobertura en los observacionales) en individuos que crecían bajo el dosel de la planta nodriza
escogida y en áreas libres de vegetación para cada estudio. La metodología utilizada en cada
caso se detalla en la sección dedicada a ese fin en los distintos capítulos de esta tesis doctoral
(ver siguiente sección).

RESUMEN
34

35
ESTRUCTURA GENERAL DE LA TESIS
Los cinco capítulos que conforman el cuerpo de esta tesis han sido escritos en inglés, para su
publicación en revistas científicas de ámbito internacional. A continuación se da una breve
descripción de estos capítulos y de la metodología utilizada en cada uno de ellos.
Capítulo 1. Las modificaciones en el régimen de lluvias predichas con el cambio
climático modulan las interacciones herbácea-arbusto en dos comunidades
semiáridas.
Pese al gran interés que ha suscitado en los últimos 20 años la relación entre las interacciones
planta-planta y el nivel de estrés, muy pocos estudios han evaluado cómo los cambios en el
patrón temporal de las precipitaciones dentro del mismo año afectan al resultado de las
interacciones planta-planta. Este patrón temporal es incluso más importante que la cantidad de
lluvia que cae durante un año determinado para el funcionamiento de los medios semiáridos.
El objetivo de este capítulo es evaluar el efecto de las modificaciones en la abundancia y
frecuencia de los eventos de lluvia predichos con el cambio climático en la interacción entre
plantones de R. sphaerocarpa y diversas especies herbáceas. Para ello aumentamos
experimentalmente la cantidad de agua disponible entre abril y julio, imitando a la inversa la
reducción de las precipitaciones durante este período (el tratamiento control, sin riego, sería el
futuro escenario de cambio climático y los tratamientos de riego serían los escenarios
actuales). El riego fue distribuido en dos o cuatro pulsos de lluvia, imitando el incremento en
la frecuencia de eventos torrenciales. El estudio se llevó a cabo durante tres años en dos
sistemas marcadamente diferentes situados sobre sustratos ricos en yeso: un espartal y un
herbazal de terraplén de carretera, dominados por S. tenacissima y diversas anuales nitrófilas,
respectivamente. Para evaluar el efecto de las herbáceas sobre R. sphaerocarpa, se midió la
supervivencia, crecimiento y eficiencia fotosintética durante estos tres años. Los objetivos de
este capítulo eran evaluar las diferencias entre 1) diferentes vecinas herbáceas, 2) el efecto de
las vecinas a lo largo de pulsos e interpulsos y 3) el cambio en el efecto de las vecinas bajo
diferentes abundancias o frecuencias de eventos lluviosos.

RESUMEN
36
Capítulo 2. La heterogeneidad espacio-temporal en los factores abióticos modula los
cambios entre competencia y facilitación que ocurren a lo largo de la ontogenia.
Las interacciones planta-planta están determinadas, en parte, por las condiciones ambientales
y la ontogenia de las especies implicadas. Pese a que el efecto de ambos factores en el
resultado de estas interacciones ha sido evaluado de forma separada, muy pocos estudios han
investigado su efecto conjunto. El objetivo de este capítulo era testar este efecto, así como su
variabilidad espacio-temporal. Para ello se evaluó la interacción entre el arbusto gipsófilo
Lepidium subulatum y la herbácea perenne S. tenacissima en tres zonas del centro Peninsular.
Dentro de estas tres zonas evaluamos el resultado neto de la interacción entre ambas especies
usando análisis de co-ocurrencia en laderas con orientación norte (menos estrés) y sur (más
estrés). En una de las tres zonas (Aranjuez) se evaluaron cambios en el signo de la interacción
a lo largo de distintas etapas ontogenéticas de L. subulatum en las dos orientaciones mediante
una combinación de experimentos de siembra y medidas dendrocronológicas, de floración y
de acumulación de carbohidratos. Este conjunto de técnicas nos permitió estudiar en detalle el
efecto de la ontogenia y de la variabilidad espacial en la disponibilidad hídrica, así como su
interacción en la relación entre L. subulatum y S. tenacissima. Las hipótesis principales
fueron: 1) la interacción entre ambas especies pasará de fuertemente positiva a fuertemente
negativa a lo largo del desarrollo de L. subulatum; 2) dado que ambas especies son tolerantes
al estrés, es de esperar que las interacciones positivas dominen a niveles intermedios de estrés
hídrico; y 3) un mayor nivel de estrés hídrico debería reducir el efecto negativo de S.
tenacissima en individuos adultos de L. subulatum.
Capítulo 3. Las dinámicas temporales de la herbivoría y la disponibilidad hídrica
interactúan modulando el resultado de una interacción herbácea-arbusto en un ecosistema
semiárido.
La herbivoría y la aridez son dos factores de estrés que comúnmente coinciden en los medios
semiáridos, jugando ambos un papel fundamental en el resultado de las interacciones planta-
planta. Sin embargo, su efecto conjunto ha sido pobremente estudiado. En este capítulo se
estudia el efecto conjunto de ambos tipos de estrés, y su variabilidad temporal a lo largo del
año, en el resultado de la interacción entre S. tenacissima y plantones del arbusto R.
sphaerocarpa. Para ello se utilizó una combinación de aproximaciones observacionales y

37
experimentales. Estas aproximaciones incluyen el seguimiento de la supervivencia y el nivel
de daño provocado por conejo en plantones sin protección a lo largo de un año, y el efecto de
la manipulación del nivel de aridez y el daño por herbivoría en el signo de la interacción entre
ambas especies. Las hipótesis iniciales de este estudio fueron: 1) S. tenacissima reducirá el
daño por conejo sobre R. sphaerocarpa mediante su papel de ocultadora frente a los
herbívoros; 2) las mejores condiciones hídricas, pero menores niveles de luz, reducirán la
tolerancia de R. sphaerocarpa bajo el dosel de S. tenacissima; y 3) el efecto conjunto de
niveles altos de ambos tipos de estrés (herbivoría y aridez) anularán el efecto positivo
derivado de la protección y la mejora microclimática de S. tenacissima.
Capitulo 4. Sobre la importancia relativa del clima y las interacciones bióticas no tróficas
como determinantes de la riqueza local de especies vegetales.
En este capítulo se pretende evaluar el efecto relativo de las condiciones climáticas y distintos
componentes de las interacciones bióticas (expansión de nicho y efecto sobre la dinámica
competitiva de las especies vecinas –intransitividad en la competencia o segregación de
nicho), así como la interacción entre ambos factores, a la hora de definir la riqueza local de
especies vegetales en comunidades semiáridas a lo largo de gradientes ambientales amplios.
Para ello se llevo a cabo un estudio observacional a diversas escalas en comunidades
semiáridas de España y Australia. Se evaluó el efecto de ocho variables climáticas, resumidas
mediante análisis de componentes principales, sobre la riqueza específica de cada localidad,
así como sobre diversos indicadores del signo, intensidad e importancia de las interacciones
bióticas a nivel de especie y de comunidad. Además, se estudió el efecto de estas
interacciones sobre la riqueza específica, y la variación de dicho efecto a lo largo del
gradiente climático escogido en cada región. Se midió la intensidad y la importancia de estas
interacciones, tanto a nivel de par de especies como de comunidad, utilizando índices
disponibles en la literatura. La frecuencia de las interacciones facilitativas a nivel de
comunidad fue cuantificada calculando el porcentaje de especies, con respecto al número total
de especies en la localidad, que se desarrollaron mejor (beneficiarias), o dependían
directamente (obligadas) de la presencia de dos especies nodrizas distintas en cada región. La
expansión de nicho fue evaluada mediante el número de especies obligadas, y también
utilizando un índice de similaridad entre las poblaciones de los microambientes nodriza (bajo
una de las dos especies nodriza seleccionadas en cada región) y claro (en suelo desnudo). Los
cambios en la dinámica competitiva de las vecinas fueron analizados mediante una

RESUMEN
38
aproximación observacional a escala de mancha, y utilizando modelos nulos de estructura
agrupada (“guild-structure null models”) para medir los patrones de co-ocurrencia. Nuestras
hipótesis iniciales predecían una igual importancia de las interacciones bióticas para la
diversidad a lo largo de los gradientes ambientales, provocada porque la expansión de nicho
afecta al mismo número de especies, aunque su identidad cambie, a lo largo de estos
gradientes. Alternativamente, proponemos que los cambios en la dinámica competitiva
(segregación de nicho o competencia intransitiva), junto con el aumento de la riqueza de
especies debido a la expansión de nicho, conducen a efectos positivos desproporcionados
sobre la riqueza local de especies debido a procesos de retroalimentación positiva.
Capitulo 5. Sobre la importancia relativa de las condiciones ambientales, las interacciones
bióticas y las relaciones evolutivas como determinantes de la estructura de las comunidades
semiáridas.
En este capítulo se evalúa el efecto relativo de las condiciones ambientales y las interacciones
bióticas (competencia/facilitación a nivel de comunidad) sobre el patrón filogenético de
espartales semiáridos dominados por S. tenacissima. En el mismo se estudia también cómo
interactúan estos dos factores a la hora de determinar dicho patrón, así como la variación en
sus efectos relativos a lo largo de un gradiente ambiental amplio. Para ello se utilizan parte de
las zonas de estudio e indicadores de interacciones bióticas descritos para el capítulo 4, junto
con medidas de co-ocurrencia a nivel de parcela. También se determinó el patrón filogenético
de cada una de las localidades muestreadas y se evaluó el efecto sobre este patrón de los
distintos mecanismos derivados de las condiciones climáticas, las interacciones bióticas, y la
interacción entre ambos factores, mediante regresiones lineares y correlaciones parciales.
Asimismo, se analizó el efecto de la distancia filogenética entre la planta facilitada y su
nodriza, de las condiciones climáticas de cada lugar, y de la interacción entre ambos factores,
como moduladores de la relación entre un total de 200 pares de especies a lo largo de este
gradiente ambiental mediante árboles de regresión. Las hipótesis principales de este capítulo
fueron: 1) la importancia relativa de las condiciones climáticas y las interacciones bióticas
para el ensamblaje de las comunidades, y por tanto para su patrón filogenético, varía a lo
largo del gradiente ambiental, y 2) la distancia filogenética y las condiciones climáticas
interactúan a la hora de definir el signo de las interacciones planta-planta.

Santiago Soliveres, Fernando T. Maestre, Pablo García-Palacios,
Adrián Escudero, and Fernando Valladares.
Manuscrito inédito
1
Predicted climate change effects in rainfall regime modulate the
outcome of grass-shrub interactions in two semi-ari d communities

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
40

CHAPTER 1
41
ABSTRACT
Much research has been devoted to understand how plant-plant interactions behave along water stress gradients in drylands. However, few studies have evaluated how changes in the magnitude and frequency of rainfall events, which are an important component of ongoing climate change, modulate the outcome of such interactions. We evaluated the response of the interaction between seedlings of the shrub Retama sphaerocarpa (L.) Boiss., our target plant, and different herbaceous neighbours to those changes in rainfall availability during three years. The experiment was conducted in natural and anthropogenic grasslands dominated by a perennial stress-tolerator and ruderal annual species, respectively. Competition between herbaceous plants and Retama seedlings prevailed, and increased with water stress. These negative effects were reduced through time, suggesting niche segregation between the interacting plants. Less frequent, but more intense rainfall events, accelerated this niche segregation in the natural grassland, where the stress-tolerator grass took more advantage of light rainfall events than Retama and competition was stronger. Thus, increases in the frequency of heavy rains could counteract the negative effects of the increased competition between grasses and shrubs expected under higher water stress conditions. However, in the anthropogenic grassland the phenology of the annuals made more frequent and lighter rainfall events more useful to avoid competition by water, being heavy rains uneffective in this case. Our findings suggest the existence of a trade-off between the shade tolerance of protégée plants and the effects of nurses on light and water availability that defines the outcome of a given plant-plant interaction. Our results challenge current predictions on the outcome of these interactions under climate change, and could be used to further refine our forecasts on how plant-plant interactions, and therefore plant communities, will respond to such change in ecosystems where grass-shrubs interactions are prevalent.

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
42
INTRODUCTION
he study of plant-plant interaction
dynamics along water stress
gradients has been a major topic
in dryland ecology during the last decade
(e.g. Pugnaire and Luque 2001, Maestre
and Cortina 2004a). The seminal “stress
gradient hypothesis” (SGH), which predicts
an increase in the frequency of facilitative
interactions as abiotic stress increases
(Bertness and Callaway 1994), constitutes a
paradigmatic framework for these studies
(see Callaway 2007 and Brooker et al. 2008
for reviews). However, the generality of its
predictions has been recently debated
(Maestre et al. 2005, 2006, Lortie and
Callaway 2006, Callaway 2007, Maestre et
al. 2009a, Smit et al. 2009, Malkinson and
Tielbörger 2010). Furthermore, the
relationship between plant-plant
interactions and environmental gradients is
especially complex in arid and semiarid
environments, where water availability is
highly pulsed, with erratic and typically
short periods of enough water availability
triggering ecosystem processes
(Schwinning and Sala 2004). Indeed, the
size and frequency of individual rain events
registered in a given period may have more
importance for the functioning of semiarid
ecosystems than the total rainfall
accumulated (Whitford 2002, Ogle and
Reynolds 2004). These temporal dynamics
should be considered when studying plant-
plant interactions and community dynamics
in water-limited ecosystems (Goldberg and
Novoplansky 1997, De la Cruz et al. 2008).
Understanding community responses to
water pulses in water limited ecosystems is
particularly timing because current rainfall
events are likely to be extremer as a
consequence of the ongoing climate change
(Knapp et al. 2008). For example, forecasts
for the Mediterranean Basin predict a
decrease in the amount of annual rainfall,
the lengthening of drought periods and the
increase of the frequency of heavy storms
(IPCC 2007). Grass-shrub interactions are
particularly instructive for studying the
effects of these climatic changes on the
outcome of plant-plant interactions because
of their contrasted water acquisition
strategies and their abundance in natural
and anthropogenic ecosystems worldwide
(e.g. Sala et al. 1989, Scholes and Archer
1997). While grasses tend to use more
efficiently the water derived from light and
sparse rainfall events (Sala et al. 1989,
Reynolds et al. 2004), shrubs generally
perform better after continuous rains, which
recharge deeper soil profiles (Sala et al.
1989, Schwinning and Ehleringer 2001,
Schwinning and Sala 2004). These grass-
shrub interactions are likely to be
particularly sensitive to the changes in
T

CHAPTER 1
43
overall water availability and frequency of
rainfall events predicted under future
climate change scenarios. For example,
shrub encroachment may be promoted by
the increase in the frequency of heavy
storms (Ogle and Reynolds 2004), or
reduced by the enlargement of summer
drought (López et al. 2008), two features of
the ongoing climate change. Alternatively,
grasses may foster shrub survival and
recruitment under moderate drought
conditions, but these positive effects may
be overcome by the increase in competition
registered under periods of very low water
availability (Kitzberger et al. 2000, Maestre
and Cortina 2004a, 2004b). The possible
responses of these interactions to changes
in rainfall amount and frequency seem very
complex and at times counter-intuitive, and
are further complicated by the fact that the
multiple responses described above are
likely to occur simultaneously.
A particular case of grass-shrub
interaction, and also farily common in
semiarid environments worldwide, occurs
when annuals act as nurse plants for shrubs
(e.g. Holzapfel and Mahall 1999 and
references therein). These annual-shrub
interactions are likely to behave
differentially along environmental gradients
because of the different life-strategy of the
former. However, they have been largely
ignored in the facilitation literature, as the
bulk of studies on grass-shrub interactions
have evaluated the effects of shrubs on
annuals (see Callaway 2007 for a review).
Thus, studies directed to clarify the effects
of grasses (both perennials and annuals) on
shrubs across realistic water stress gradients
are of crucial importance to understand
plant community responses to ongoing
climate change in those ecosystems where
grasses and shrubs coexist. In this study we
aimed to test the response of grass-shrub
interactions to changes in the degree of
abiotic stress. The study was conducted in
two different semiarid Mediterranean
communities: a natural Stipa tenacissima L.
steppe (hereafter called “natural grassland”)
and an annual-dominated grassland located
in a motorway embankment (hereafter
called “anthropogenic grassland”).
Degraded landscapes such as the latter
represent good examples of novel
ecosystems, which are increasing in
importance worldwide because of the rise
in anthropogenic disturbances (Hobbs et al.
2006). Given that the structure and
functioning of these novel ecosystems often
differs from that found in natural ones
(Hobbs et al. 2006), studies focusing on
both natural and novel ecosystems may
help to further refine our predictions of the
response of plant communities to climate
change (Brooker 2006). Our focal species
were the leguminous shrub Retama
sphaerocarpa (L.) Boiss. (hereafter
Retama) and different grass species (the

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
44
stress-tolerator tussock grass Stipa
tenacissima –hereafter Stipa– and several
ruderal annual species), the latter acting as
potential nurse plants for Retama.
The outcome of grass-shrub
interactions in semiarid ecosystems, either
natural or anthropogenic, depends strongly
on soil water availability (Eliason and Allen
1997, Maestre and Cortina 2004a), and
therefore, both grassland-types are excellent
study systems to test changes in the
outcome of grass-shrub interactions across
abiotic stress gradients. We obtained a
realistic water stress gradient by modifying
the amount and timing of water availability
separately, according to the most likely
climate change scenarios for the study area.
We tested the following hypotheses: i) both
Stipa and annual grasses will reduce
Retama growth because of competition by
water during wet seasons, when most of
vegetation growth concentrates in semiarid
environments (Goldberg and Novoplansky
1997, Escudero et al. 1999); ii) Stipa and
annuals will differ in their effect on Retama
survival. Although both Stipa and annuals
will increase Retama survival during
summer drought, mainly via microclimate
amelioration and improvement of soil
properties (Goldberg and Novoplansky
1997, Maestre et al. 2003), these effects
will change with the lengthening of summer
drought. Under these conditions, the
positive effects of annuals will be more
intense because they will be mainly derived
from the shade produced by their dry
tissues (annuals die during summer), but
Stipa will reduce Retama survival because
of the increased competition by water will
outweight the positive environmental
buffering promoted by shade (Maestre and
Cortina 2004a); and iii) the increase in the
frequency of heavy storms will reduce
competition between Retama and both Stipa
and annual grasses by recharging deeper
soil layers and promoting niche segregation
(Sala et al. 1989, Ogle and Reynolds 2004).
METHODS
STUDY AREA
Both the natural and anthropogenic
grasslands selected for this study are
located in the center of the Iberian
Peninsula (natural grassland: 40º03´60´´N,
3º54´91´´W, 545 m.a.s.l.; anthropogenic
grassland: 52º16´00´´N, 3º43´13´´W, 604
m.a.s.l.) and share the same climate and soil
type. The climate is semiarid
Mediterranean, with average annual
precipitation and temperature of 388 mm
and 15 ºC, respectively, and with a strong
summer drought (Aranjuez weather station;
1994-2005 period; Marqués et al. 2008).
Both communities are located on gypsum-
rich soils, classified as Xeric Haplogypsid
(Marqués et al. 2008), although in the
anthropogenic grassland the original

CHAPTER 1
45
substrate has been altered by the mixture
with gravels and components from external
sources during the construction of the
motorway. Vegetation in the natural
grassland is an open steppe dominated by
Stipa tenacissima, with a perennial plant
cover of 24%. Vegetation in the
anthropogenic grassland is dominated by
annuals, with a mean cover of 75% and
with Bromus rubens L., B. diandrus Roth.,
and Medicago sativa L. as the most
abundant species (16, 14 and 14% of the
total cover, respectively; García-Palacios et
al. 2010). Hereafter we refer to those
herbaceous annuals as annual grasses for
simplicity. Both study sites hold a high
density of rabbits (Oryctolagus cunniculus
L.), as suggested by visual contacts, and by
the high number of warrens and latrines
found (S. Soliveres, pers. obs.).
EXPERIMENTAL DESIGN
In December 2006, 176 two-year old
Retama seedlings, with a mean height of of
27 ± 2 cm, were planted in each site by
using manually-dug holes of 20×20×20 cm.
These seedlings came from a nursery in
central Spain (viveros Bárbol, Madrid). We
randomly assigned these seedlings to two
different microsites: “Nurse” and “Open”.
Because of the heterogeneous patch-
interpatch structure of the natural steppe
system, and the homogenous herbaceous
cover of the anthropogenic site, these
microsite types were defined differently in
the two ecosystems. Nurse microsites were:
1) located upslope and adjacent to Stipa
tussocks (stress tolerator nurse) of ca. 1 m
width (< 15 cm from the edge of the north
face of the tussock, where facilitative
effects of this species on target shrubs have
been found, e.g. Maestre et al. 2003) in the
natural grassland, and 2) located in the
center of a multi-specific 50-cm diameter
grass patch (ruderal nurse) of ca. 40 cm
height and 75-100% cover (in spring) in the
anthropogenic grassland. The rest of the
seedlings were assigned to randomly
selected Open microsites. These were either
located in bare ground areas at least 80 cm
away from any perennial plant (natural
grassland site), or placed in sites where all
aboveground vegetation within 80-cm
diameter circles surrounding the target
seedling was monthly clipped
(anthropogenic grassland site).
The seedlings were randomly allocated
to establish two full factorial experiments
with two factors each, which were run in
parallel in the two grasslands studied. In the
first experiment (hereafter Experiment 1)
the two factors were Microsite (Nurse vs.
Open) and Irrigation. This irrigation
treatment consisted in three different
amounts of water applied during eight
watering events (once every month between
April and July in both 2007 and 2008). In
each watering event, the 0%, 25% and 50%

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
46
of the median from the last 30 years for that
particular month was applied to the control
(hereafter +0%), amount 1 (hereafter
+25%), and amount 2 (hereafter +50%)
treatments, respectively. In the second
experiment (hereafter Experiment 2) the
factors were i) Microsite (Nurse vs. Open),
and ii) Irrigation frequency. In this case we
applied the same amount of water
employed in the +50% treatment, but with
two different frequencies: two and four
pulses (hereafter named as 2x and 4x
treatments, respectively). The first
frequency treatment (4x) was applied in
four monthly pulses, from April to July, as
explained above; the second frequency
treatment (2x) was applied every two
months, in May and July. In each of the
watering events applied in the 2x treatment,
the amounts of water added equaled the
sum of April and May, and June and July
irrigations applied in the 4x treatment,
respectively (Fig. 1.1, Table 1.1). Both
irrigation treatments were applied
irrespectively of the rainfall registered in
each month (Fig. 1.1). With these two
experiments we aimed to evaluate the
effects on the outcome of the interaction
studied of: i) an increase in summer drought
(summer drought was longer in less-
watered plants [+0% > +25% > +50%]
because of the low rainfall levels typically
registered during June and July in the study
area), ii) a reduction of rainfall during the
wet season (spring rainfall was less
abundant in non-watered plants [+0% <
+25% < +50%]), and iii) changes in the
frequency of heavy showers (magnitude of
individual events was higher in the 2x than
in the 4x treatment, despite both treatments
received the same amount of water, Fig.
1.1). All these effects, i.e. the increase in
summer drought, the reduction in the total
rainfall amount and the increase in the
frequency of heavy showers are predicted
by future climate change scenarios for the
Mediterranean Basin (IPCC, 2007; see
Table 1.1). Because of the high density of
rabbits observed, and to avoid seedling
predation, the seedlings were protected
from browsing by using a thin-wire mesh.
This mesh did not shade the seedlings, and
thus did not confound the effects of any of
the factors studied in the experiment.

CHAPTER 1
47
Figure 1.1 Climatic data (mean monthly temperature, black circles; and monthly rainfall, black bars) obtained from a meteorological station (Onset, Pocasset, MA, USA) located in the natural grassland. The increment in monthly rainfall promoted by the irrigation treatments applied during 2007 and 2008 is represented by different colors: +25% (dark grey) = irrigation of 25% of the median of April-July rainfall in four pulses, +50 -4x (light grey) = irrigation of 50% of the median of April-July rainfall in four pulses, and +50 -2x (white) = irrigation of 50% of the median of April-July rainfall in two pulses).
Table 1.1 Details of the water amount added (l · m-2) in each irrigation treatment (Experiments 1 and 2) and the periods when these pulses took place. Last row explains the specific climate change effect that each irrigation treatment emulated.
Irrigation
Pulses
EXPERIMENT 1:
Amount treatment
EXPERIMENT 2:
Frequency treatment
+0% +25% +50% 4x 2x
April 0 6 12 12
May 0 9.25 18.5 18.5 30.7
June 0 10.85 21.7 21.7
July 0 5.65 11.3 11.3 33 Total water
applied 0 31.75 63.5 63.5 63.5
Simulated
climate change
effect

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
48
MONITORING OF SOIL MOSITURE
AND PLANT PERFORMANCE
Soil moisture was measured by time-
domain reflectometry (TDR; Topp and
Davis 1985) using a Campbell TDR100
system (Campbell Scientific Ltd,
Loughborough, UK). In 10 randomly
selected planting holes per each of the eight
possible treatments combinations, 10 cm
long TDR probes were vertically installed
(n = 80). This soil depth was chosen
because we expected the concentration of
most of the roots of the interacting species
to be in the upper part of the soil during the
study period (herbs and young woody
seedlings should concentrate their roots in
the upper soil layers; Scholes and Archer,
1997). A strong relationship between TDR
values and soil gravimetric moisture has
been found in the natural grassland (R2 =
0.84; P < 0.0001; Soliveres et al. unpubl.
data); thus, this measurement can be
considered as a good proxy for soil
moisture availability. Soil moisture was
measured every two months during the
study period, starting and ending in April
2007 and September 2009, respectively. As
summer drought is considered the
bottleneck for plant recruitment in semiarid
Mediterranean environments (Escudero et
al. 1999), this sampling was conducted
monthly during the summer (June-
September).
As the magnitude of the effect of Stipa
on woody seedlings follows seasonal
dynamics of water availability (Maestre et
al. 2003), the effects of herbaceous
neighbours on soil moisture availability
were tested considering different dry/wet
periods. To evaluate how the irrigation
treatments and the presence of grasses
affected soil moisture, the relative
interaction index (RII; Armas et al. 2004)
was calculated for each sampling date as:
(TDRNu – TDROp)( (TDRNu + TDROp),
where TDRNu and TDROp are soil moisture
data obtained in Nurse and Open
microsites, respectively. To aid interpreting
our results, RII data were grouped in
intervals with percentages in soil moisture
above and below 10% (wet and dry
seasons, respectively). This limit
corresponds to the natural seasonal
dynamics in water availability; periods
below and above 10% moisture occur
mainly during summer drought and when
plant activity concentrates, respectively
(Schwinning and Sala 2004, Reynolds et al.
2004). We randomly paired the samples by
microsite (Nurse vs. Open) and between the
four irrigation treatments obtained with the
two experiments (+0%, +25%, +50%-4x,
and +50%-2x; 10 pairs for each microsite ×
irrigation combination). Then, RII data of
these paired samples were grouped for wet
and dry seasons separately. With these data,
the average RII for all wet and dry seasons
was obtained for each of the four irrigation

CHAPTER 1
49
treatments.
The outcome of plant-plant interactions
varies widely depending on the
performance measure used (Goldberg and
Novoplansky 1997, Maestre et al. 2005).
Thus, as recommended when studying these
interactions along stress gradients (Brooker
et al. 2008), we used several performance
measurements to test the effect of
neighbour grasses on Retama seedlings.
Seedling height, root collar diameter and
survival were measured after each summer,
in September 2007, 2008 and 2009. A
strong relationship between standing
biomass and root collar diameter has been
previously observed for Retama seedlings
(R2 = 0.823, P < 0.0001; Soliveres et al.
unpubl. data), so this measure was used as
our surrogate of seedling biomass in the
field. The relative growth rate (hereafter
RGR) of Retama seedlings for each year
was obtained as: ln RCD1 – ln RCD0)/(T1 –
To); being RCD1 and RCD0 the root collar
diameter at the precedent (T0) and current
(T1) sampling date, respectively. The
slenderness coefficient, which is considered
a good indicator of light competition
(Kurashige and Agrawal 2005), was also
calculated as the ratio between height and
root collar diameter lnv H/ln RCD. Light
competition can be an important factor
affecting plant-plant interactions, even in
water-limited environments (Seifan et al.
2010a, Soliveres et al. 2010). Therefore,
measuring it is important to understand its
relative role in the interactions studied.
Both plant growth and survival are
integrative measurements that include the
effects of microsite or irrigation on plant
performance during the whole year.
Therefore, they are not appropriate to detect
seasonal differences in the effect of the
assayed treatments on seedling
performance. To assess for such
differences, we estimated the potential
photochemical efficiency of Retama
seedlings by measuring the maximum
quantum yield of PSII (Fv/Fm) of dark-
adapted leaves (30 min, at midday) with a
pulse-modulated fluorometer (FMS2,
Hansatech Instruments, Norfolk, UK). This
parameter has been widely used as an
indicator of plant stress in semiarid regions
(e.g. Pugnaire et al. 1996b, Maestre et al.
2003, Aragón et al. 2008); small changes in
photochemical efficiency have been
associated with water limitations during
important stages of the lifecycle of woody
plants in these environments (Aragón et al.
2008), and have been found to match
results with other performance measures
(e.g. survival) when evaluating the outcome
of plant-plant interactions (e.g. Maestre et
al. 2003, 2004). Six seedlings per
combination of treatments and grassland
type were randomly selected for these
measurements (n = 48 per grassland).
Different randomly selected plants were

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
50
measured in each sampling period, which
took place on the same dates than TDR
surveys (see above). To test for differences
in the intensity of biotic interactions in
response to the treatments evaluated within
dry or wet seasons, the RII index was
calculated using Fv/Fm data as described
above for the TDR data.
STATISTICAL ANALYSES
RII data (calculated from both TDR and
Fv/Fm measurements) were organized by
dry/wet seasons to test treatment effects
during seasons with contrasted rainfall
availability during the year. We calculated
95% confidence intervals of this index for
each level of irrigation amount or
frequency, and per each wet/dry period to
test for significant differences from zero
(which means neutral effect of the nurse on
the response variable). If 95% confidence
intervals did not overlap with zero or with
each other, we took as significant the
differences of the RII from zero or between
treatments, respectively. These differences
were interpreted as increases (if
significantly higher) or decreases (if
significantly lower) of soil moisture or
Fv/Fm under the canopy of the herbaceous
neighbours in comparison with open areas.
Relative growth rate data obtained for
root collar diameter and slenderness
coefficient were correlated from one year to
another (r > 0.375, P < 0.0001 in all cases).
Thus, we used multivariate analysis of
variance (MANOVA) to test the effects of
microsite and irrigation on this variable at
each study site. These analyses were
conducted separately for Experiments 1 and
2. Slenderness coefficient data were
squared-root transformed to meet
MANOVA assumptions.
Damage derived from rabbit activity
(warrens and territory coverage) was an
important source of seedling death despite
of the grazing protection provided (see
results below). Thus, we separated the
survival status of Retama seedlings into
three levels: alive, death by drought or
death by rabbit. We analyzed survival
percentages of these seedlings separately
for each year and study site by using a
hierarchical log-linear analysis, with
microsite and irrigation as fixed factors. To
assess the effects of the factors assayed
during each year, only those seedlings that
survived the previous summer were taken
into account (for example, to analyze
survival of 2008, we only considered those
seedlings alive after the summer of 2007).
With this approach, we were able to assess
the consistency of the effect of the
treatments evaluated over the years. This
approach also avoids the potential
“dragging” that an extremely strong effect
of a given treatment during a given year
may have on the overall net results (e.g. if
the +50% treatment would had strong

CHAPTER 1
51
effects in 2007, but not in the rest of years
we could detect it with our approach; in
contrast, taking into account survival
percentage from the beginning may produce
an overall net significant effect of this
treatment over the study period). Survival
data from one year to another were not
correlated (Pearson r < 0.4; P > 0.2 in all
the cases); thus, independency is expected.
As with growth data, separate analyses
were performed for each experiment.
Statistical analyses were conducted using
SPSS 13.0 for Windows (Chicago, Illinois,
USA).
RESULTS
SOIL MOISTURE
The different irrigation treatments produced
contrasted results depending on the
grassland considered. In the natural
grassland, the frequency, but not the
amount, of water added modified the effect
of Stipa on soil moisture; the opposite
response was found in the anthopogenic
grassland. In the natural grassland, Stipa
had a mostly neutral effect on soil moisture
in both dry and wet periods, regardless of
the amount of water added (Fig. 1.2A). This
neutral effect shifted to slightly positive
when the +50% treatment was applied in
two pulses (2x treatment), a response not
observed in the 4x treatment (Fig. 1.2B). In
the anthopogenic grassland, the increase in
water amount reduced monotonically the
negative effect of annuals on soil moisture
(Fig. 1.2C). Annuals reduced soil moisture
in the +0% treatment, but did not affect, or
even increased, water availability
comparing to open microsites in the +25%
and the +50% treatments, respectively. The
latter had the same effect regardless of the
frequency of watering pulses (Fig. 1.2D).
These effects were consistent in both dry
and wet periods.
PLANT PERFORMANCE
In Experiment 1, the irrigation treatments
modified the effect of grasses on the Fv/Fm
of Retama seedlings in a way that
mimicked the effects of the former on soil
moisture in the natural grassland (Fig. 1.3).
However, the amount of water applied did
not affect the studied interaction in the
natural grassland, with the exception that
Stipa effect shifted from neutral to slightly
positive as water availability increased
during wet seasons (neutral in the +0%
treatment and slightly positive in the rest;
Fig. 1.3A). Although the amount of water
was not important affecting the outcome of

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
52
Figure 1.2 Relative effects of herbaceous nurse plants, as measured with the Relative Interaction Index (RII), on soil water availability during dry/wet seasons (periods with soil moisture above and below 10%, respectively). RII was calculated for the mean values of ten Nurse/Open pairs for each of the irrigation treatments assayed. Legend of the Experiment 1 (panels A and C) and Experiment 2 (panels B and D) treatments as follows: +0% = no irrigation, +25% = irrigation of 25% of the median of April-July, +50% = irrigation of 50% of the median of April-July in four pulses, 4x = irrigation of 50% of the median of April-July rainfall in four pulses, and 2x = irrigation of 50% of the median of April-July rainfall in two pulses. Data represent means ± 95% confidence interval (n = 10) in the natural (A and B) and the anthropogenic (C and D) grassland, respectively.
the studied interaction during dry seasons,
less frequent but heavier water inputs (2x
treatment) neutralized the negative effect of
Stipa on Retama found in dry periods (Fig.
1.3B). Alternatively, nurse annuals reduced
monotonically their negative effect on
Retama seedlings during dry seasons as the
water amount added increased (Fig 1.3C),
but this treatment did not affect the studied
interaction during wet periods. Contrary to
the results found in the natural grassland,
the 2x treatment did not affect the
interaction outcome neither in dry nor in
wet seasons in the anthropogenic grassland
(Fig. 1.3D).

CHAPTER 1
53
Figure 1.3. Relative effects of herbaceous nurse plants, as measured with the Relative Interaction Index (RII), on seedling photochemical efficiency during dry/wet seasons (periods with soil moisture above and below 10%, respectively). RII was calculated for the mean values of six Nurse/Open pairs for each of the irrigation treatments assayed. Data represent means ± 95% confidence interval (n = 6) in the natural (A and B) and the anthropogenic (C and D) grassland, respectively. Rest of legend as in Fig. 1.2.
Neither microsite nor irrigation
treatments affected the growth rate of
Retama in the natural grassland, which was
almost nill in all the cases (Table 1.2; see
detailed statistics in Appendix A in
Supplementary Material). The 2x treatment,
but not the rest, slightly decreased the
slenderness coefficient of Retama seedlings
(Table 1.3). This trend was constant for the
three years of study, but was significant
only in 2007 (F3,53 = 3.63; P = 0.019). The
growth rate of Retama was much higher in
the anthropogenic grassland than in the
natural grassland, and was negatively
affected by the presence of annuals
(MANOVA Pillai´s Trace: F2,53 = 4.6; P =
0.014). According to the results found with
the Fv/Fm measurements, increases in
water availability (+50% treatment)
neutralized this negative effect of annuals,
regardless of the frequency of its
application. The slenderness coefficient was
no affected neither by microsite nor by any
of the irrigation treatments in the
anthropogenic grassland.

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
54
Table 1.2. Relative growth rate (cm · cm-1 · year-1) of root collar diameter (RGR) of Retama sphaerocarpa seedlings in the different experiments and study sites. Values are means ± SE (n = 4-16, depending on the treatment and the sampling period). Open = areas without perennial vegetation, Nurse = areas under the canopy of annual plants (anthropogenic grassland) or Stipa tenacissima (natural grassland), +0% = unwatered seedlings, +25% = irrigation of 25% of the median of April-July rainfall in four pulses, 4X = irrigation of 50% of the median of April-July rainfall in four pulses, and 2X = irrigation of 50% of the median of April-July rainfall in two pulses. Different letters or asterisks indicate significant differences (MANOVA test; P < 0.05) between microsites or among irrigation treatments, respectively. See Appendix A in Supplementary Information for detailed statistical results.
Experiment 1: Amount treatment
Microsite Irrigation Natural grassland Anthropogenic grassland
2007-2008 2008-2009 2007-2008 2008-2009
Open
+0% 0.00 ± 0.02 0.02 ± 0.01 1.37 ± 0.13 1.66 ± 0.13a
+25% 0.06 ± 0.01 0.00 ± 0.01 1.54 ± 0.08 1.83 ± 0.06a
+50% 0.05 ± 0.01 0.00 ± 0.01 1.40 ± 0.08 1.63 ± 0.08a
Nurse
+0% 0.00 ± 0.02 0.01 ± 0.01 1.40 ± 0.09 1.70 ± 0.11b
+25% 0.02 ± 0.01 0.00 ± 0.01 1.59 ± 0.13 1.56 ± 0.12b
+50% 0.03 ± 0.01 0.00 ± 0.01 1.56 ± 0.22 1.75 ± 0.13b
Experiment 2: Frequency treatment
Open 4X 0.05 ± 0.01a 0.00 ± 0.01 1.40 ± 0.08 1.63 ± 0.08
2X 0.04 ± 0.01a 0.01 ± 0.02 1.41 ± 0.11 1.66 ± 0.11
Nurse 4X 0.03 ± 0.01b 0.00 ± 0.01 1.56 ± 0.22 1.75 ± 0.13
2X 0.00 ± 0.01b 0.00 ± 0.02 1.46 ± 0.09 1.60 ± 0.08
The presence of grasses reduced the
survival of Retama in the two grasslands
studied during 2007 and 2008, a negative
effect that disappeared in 2009 (Table 1.4,
see also Appendix B in Supplementary
Material for detailed statistical results). In
the anthropogenic grassland, we found a
negative effect of grasses on the ability of
Retama seedlings to resist summer drought
in 2007, but a positive effect was detected
by the protection against rabbit damage
during this year, resulting in a net neutral
effect (see Retama seedlings death by
drought or by rabbits in Table 1.4).

CHAPTER 1
55
Table 1.3. Slenderness coefficient (unitless) of Retama sphaerocarpa seedlings in the different experiments and study sites. Values are means ± SE (n = 4-16, depending on the treatment and the sampling period). Rest of legend as in Table 1.2
Experiment 1: Amount treatment
Microsite Irrigation Natural grassland Anthropogenic grassland
2007 2008 2009 2007 2008 2009
Open
+0% 2.4 ± 0.3 2.2 ± 0.2 1.8 ± 0.1 2.7 ± 0.2 2.2 ± 0.1 1.7 ± 0.1
+25% 3.2 ± 0.2 2.2 ± 0.2 2 ± 0.2 3.5 ± 0.7 2.1 ± 0.2 1.8 ± 0.1
+50% 3.4 ± 0.4 2 ± 0.2 2 ± 0.1 4 ± 0.5 2.5 ± 0.2 1.6 ± 0.1
Nurse
+0% 2.9 ± 0.6 3 ± 0.5 2.5 ± 0.5 2.7 ± 0.2 2.2 ± 0.1 1.7 ± 0.1
+25% 2.9 ± 0.2 2.4 ± 0.2 2.3 ± 0.2 3.1 ± 0.3 2.2 ± 0.2 2 ± 0.1
+50% 3.7 ± 0.5 2.7 ± 0.4 2.4 ± 0.2 3.4 ± 0.4 2.4 ± 0.2 1.9 ± 0.1
Experiment 2: Frequency treatment
Open 4X 3.4 ± 0.4 2 ± 0.2 2 ± 0.1 4 ± 0.5 2.5 ± 0.2 1.6 ± 0.1
2X 2.4 ± 0.2* 1.9 ± 0.1 1.9 ± 0.2 4.1 ± 0.5 2.6 ± 0.2 1.7 ± 0.1
Nurse 4X 3.7 ± 0.5 2.7 ± 0.4 2.4 ± 0.2 3.4 ± 0.4 2.4 ± 0.2 1.9 ± 0.1
2X 2.3 ± 0* 2.2 ± 0.4 2.1 ± 0.2 3.7 ± 0.3 2.7 ± 0.1 2.4 ± 0.2
These negative effects were independent of
the irrigation treatments, with the exception
of Experiment 1 in the anthropogenic
grassland in 2008. During this year, the
survival of Retama monotonically increased
with water availability (+0% < +25% <
+50%), and the negative effects of annuals
on the survival of this species disappeared
in the +50% treatment (Table 1.4,
Appendix B). Different irrigation
frequencies did not affect the survival of
Retama seedlings neither in the natural nor
in the anthropogenic grassland.

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
56
Table 1.4. Percentage of Retama sphaerocarpa seedlings death by drought or by rabbit (separated by /) during the three years of study in the different experiments and study sites. Percentage were calculated taking as 100% those seedlings which had survived after the summer of the previous year. Initial n was 22 for each combination of treatments. Different letters or asterisks indicate significant differences (Log-linear test; P < 0.05) between microsites or among irrigation levels within each experiment, respectively. See Appendix B in Supplementary Information for detailed statistical results. Rest of legend as in Table 1.2.
Amount treatment
Microsite Irrigation Natural grassland Anthropogenic grassland
2007 2008 2009 2007 2008 2009
Open
+0% 4 / 0 10 / 3a 33 / 3 0 / 12a 12 / 12a 33 / 15
+25% 4 / 8 18 / 0a 33 / 0 0 / 9a 10 / 19a 38 / 8
+50% 0 / 4 21 / 0a 44 / 0 0 / 13a 4 / 13a* 13 / 13
Nurse
+0% 13 / 0 60 / 0b 75 / 0 8 / 0b 30 / 13b 52 / 11
+25% 8 / 0 50 / 0b 75 / 0 9 / 4b 30 / 22b 40 / 20
+50% 0 / 0 53 / 11b 63 / 11 10 / 0b 19 / 0b* 38 / 5
Frequency treatment
Open 4X 0 / 4 21 / 0a 44 / 0 0 / 13a 4 / 13 13 / 13
2X 0 / 0 19 / 5a 40 / 15 0 / 16a 0 / 21 20 / 24
Nurse 4X 0 / 0 53 / 11b 63 / 11 10/ 0b 19 / 0 38 / 5
2X 17 / 4 53 / 5b 74 / 5 12 / 4b 21 / 4 55 / 3
DISCUSSION
WATER STRESS INCREASES THE
INTENSITY OF GRASS-SHRUB
COMPETITION
According to our first hypothesis, we found
a negative effect of both Stipa and annuals
on the growth of Retama seedlings
(Goldberg and Novoplansky 1997).
However, this effect was almost nill in the
natural grassland, maybe due to the low
growth rates found in this ecosystem
regardless of the treatment applied. The
negative effect of annuals found in the
anthropogenic grassland seems to be mainly
caused by competition by water, an effect
that increased with water stress. According
to previous studies (Knoop and Walker
1985, Davis et al. 1999, Maestre and

CHAPTER 1
57
Cortina 2004a), the highest competition
intensities were found under the higher
water stress levels (both summer drought
and non-watered treatments in the
Experiment 1), a response that contradicts
predictions of the SGH (Bertness and
Callaway 1994). Both Stipa and the annuals
negatively affected the growth, stress level
and survival of Retama seedlings. These
results partially reject our second
hypothesis, based on the model proposed by
Goldberg and Novoplansky (1997), because
herbaceous neighbours reduced shrub
survival and did not show any positive
effect during dry periods (Figs. 1.2 and
1.3). Also, and in contrast with our second
hypothesis, this effect remained equal
regardless of the water amount added in the
natural grassland, but was more negative in
the anthropogenic grassland as water stress
increased (see results of Experiment 1). The
most plausible explanation for these results
is that the negative effects of both Stipa and
annuals on the stress experienced by
Retama seedlings during wet periods
reduced growth and resource capture,
compromising survival during summer
regardless of the environmental stress
experienced during this season.
Ruderal plants have been suggested as
possible nurse plants in degraded areas,
where other nurse plants are absent, and its
potential use as a restoration tool has been
suggested (Brooker et al. 2008). However,
our results do not fully support this
recommendation because of the inexistence
of positive effects of these herbaceous
plants during the entire study period (see
also McDonald 1986, Eliason and Allen
1997). Interestingly, when considering the
impact of rabbit activity, which was
considerable in the anthropogenic
grassland, the negative effect of annuals on
the ability of seedlings to overcome
summer drought was compensated by the
protective role of these annuals (see Retama
seedlings death by drought/rabbit in Table
1.4). This result illustrates how the effects
that multiple stressors have on a particular
nurse-protégée interaction is a crucial factor
affecting the potential of this particular
nurse to have positive, neutral or negative
effects on the protégée depending on the
particular environment where this
interaction occur (Smit et al. 2009,
Soliveres et al. in press).
DIFFERENT NURSE GRASSES HAVE
UNEVEN EFFECTS ON RETAMA
SEEDLINGS
Competition between Retama and annuals
was clearly driven by water competition,
and therefore increases in soil moisture
reduced the negative effect of these species.
However, the effects of Stipa on soil
moisture (neutral or positive) did not fully
match those on seedling performance
(survival reduction and increased stress),

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
58
suggesting that competition by factors other
than water were also important drivers of
the final outcome of the interaction studied.
The shade intolerance of Retama
(Valladares and Pugnaire 1999, Valladares
et al. 2003), together with the trend to
increase slenderness coefficient underneath
Stipa canopy, seems to point to light
competition as a key factor defining the
outcome of the studied interaction in the
natural, but not in the anthropogenic,
grassland, where Stipa canopies could
shade Retama seedlings enough to limit
carbon gain (Davis et al. 1999, Soliveres et
al. 2010, Seifan et al. 2010a). Light
competition could explain the contrasting
results found between both grassland types,
where a lack of influence of water amount
in the natural, but not in the anthropogenic
grassland was found. Our findings, and
those from previous studies (Prider and
Facelli 2004, Seifan et al. 2010a), point to
the existence of a trade-off between the
shade and drought tolerances of the
protégée plant and how nurse plants affect
water and light supply for their protégées.
For example, although nurses may exert a
positive effect on the availability of one
resource (i.e. water), the overall effect of a
nurse on a particular target will be negative
if shade provided by this nurse is deeper
than the physiological limits of this
particular protégée plant can withstand,
regardless of its positive effect on water
availability (Malkinson and Tielbörger
2010). This trade-off should be taken into
account when studying specific plant-plant
interactions along stress gradients because
it will define the final outcome of a given
interaction and how it changes across
environmental gradients (Holmgren et al.
1997, Prider and Facelli 2004, Malkinson
and Tielbörger 2010).
CHANGES IN RAINFALL
FREQUENCY, HERBACEOUS
PHENOLOGY AND POTENTIAL FOR
NICHE SEGREGATION
Niche segregation is among the most
important factors fostering coexistence in
semiarid environments (Fowler 1986, Sala
et al. 1989, Scholes and Archer 1997).
Despite that our study only lasted three
years, it seems enough time for Retama
seedlings to avoid competition by grasses,
as demonstrated by the lack of effect of
nurse herbs on Retama survival during
2009 at both study sites. Retama seedlings
are able to reach deep soil layers soon after
its establishment (Padilla and Pugnaire
2007), and thus it is likely that they were
able to reach deeper soil profiles than those
achieved by its herbaceous neighbours in
three years. These results agree with
previous studies with other species (Brown
and Archer 1990), which found a high
ability of woody seedlings to reach soil

CHAPTER 1
59
resource partitioning with grasses very
early in their life cycle.
Contrary to our expectations (Ogle and
Reynolds 2004, Knapp et al. 2008) and
despite of the existence of niche
segregation described above, the assayed
differences in the frequency of heavy
storms (2x vs. 4x treatment in Experiment
2) did not increase seedling survival when
growing with grasses in any of the studied
sites. However, taking into account other
measurements (soil moisture and Fv/Fm),
the two assayed frequencies had differential
effects depending on the site considered.
While more frequent water inputs (4x
treatment) promoted higher soil moisture
and shrub performance in the anthropogenic
grassland, less frequent but heavier water
inputs (2x treatment) had the same effects
in the natural one. Following the inverse
soil texture hypothesis proposed by Noy-
Meir (1973), this could be caused by
differences in soil depth between both
grasslands, with deeper soils (i.e. the
natural grassland) allowing niche
segregation and shallow soils (i.e.
anthropogenic grassland) preventing it.
However, both soil types had similar
geological basis and thus it is likely that
both had similar textures and depths. Thus,
the most plausible explanation for this
differential response between grassland
types is that differences on the life-strategy
(annuals vs. perennials) of the herbaceous
nurses caused it (Gómez-Aparicio 2009). In
the anthropogenic grassland more water
pulses (those of spring and early summer)
in the 4x than in the 2x treatment increased
the amount of water available for seedlings
when the annual nurses were active. This
increase in water availability might
compensate competition by water between
annuals and Retama seedlings, as
demonstrated by the significant reduction of
negative effects on survival, growth or
stress found. On the other hand, in the 2x
treatment one of the two irrigation pulses
was applied during summer, when most
ruderal nurses were death. Therefore,
negative effects of annuals on Retama
seedlings were less important when the
plant received this irrigation, resulting in a
less effective compensation of water
competition when compared to the same
amount of water applied more frequently.
In contrast, Stipa is not only perennial and
therefore competitive during the entire year,
but also highly efficient taking water from
short and light rainfall inputs, even during
summer (Pugnaire et al. 1996b, Balaguer et
al. 2002). Thus, the 4x treatment more
likely benefited Stipa than reduced
competition between this species and
Retama seedlings in the natural grassland.
However, the less frequent but heavier
water pulses applied in the 2x treatment
could reach deeper soil profiles, where
Stipa is not able to take water, and therefore

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
60
may enhance niche segregation, increasing
shrub performance (Fv/Fm results; see also
Sala et al. 1989, Schwinning and Ehleringer
2001, Ogle and Reynolds 2004). In spite of
this competition was importantly affected
by light (see discussion above), an
improvement in water status could increase
the ability of Retama to compete for light
(Fahey et al. 1998), as demonstrated by the
significant increase in the slenderness
coefficient and Fv/Fm observed in the 2x
treatment. Unfortunately, we did not
measure root structural traits, but the
extremely low shoot growth rates of
Retama seedlings found in this ecosystem,
compared with those observed in the
anthropogenic grassland (Table 1.2),
suggests a major investment in root growth
to promote this niche segregation. The
differential responses to different water
availabilities and frequencies found
depending on the life-strategy of the
herbaceous plants could help to reconcile
contrasting results observed in the
literature, which support or reject niche
segregation between woody plants and
grasses.
CONCLUDING REMARKS
Despite the plethora of studies devoted to
test facilitation/competition shifts along
stress gradients (see Callaway 2007 for a
review), to our knowledge none of them
have experimentally tested these shifts
within the same site, avoiding site-to-site
confounding factors, and with more than
two points along stress gradients driven by
water availability. We did so in two
contrasting ecosystems with different
herbaceous potential nurses, and following
a four-point realistic water stress gradient,
derived from predictions for future climate
change scenarios for the study areas, within
each of the three study years. Our results
suggest that the expected increase in water
stress under climate change will reduce
shrub recruitment and performance in
semiarid grasslands, but these effects will
depend on the specific species involved and
the suggested trade-off between
shade/drought tolerances of protégée and
nurse effects on light and water availability.
However, the increase in the frequency of
heavy rains may counteract this effect by
enhancing niche segregation among
coexisting plants (Knapp et al. 2008), an
effect that will be mediated by the life-
strategy of the nurses involved. The results
presented here challenge current predictions
of plant-plant interactions in response to the
ongoing climate change in Mediterranean
ecosystems (Brooker 2006), and therefore
raise caution on current generalizations on
how grass-shrub interactions will respond
to climate change.
ACKNOWLEDGEMENTS We thank Matthew Bowker for revising the English of this manuscript. A.P. Castillo-Monroy, M.

CHAPTER 1
61
Carpio, E. Pigem, C. Alcalá, P. Alonso, R. Milla, L. Gimenez and J. Margalet for their help during fieldwork. We thank the Instituto Madrileño de Investigación y Desarrollo Rural, Agrario y Alimentario (IMIDRA) and CINTRA S.A. for allowing us to work in their sites. SS and PGP hold PhD fellowships from the EXPERTAL project, funded by Fundación Biodiversidad and CINTRA S.A. This work was supported by the EXPERTAL,
REMEDINAL2, INTERCAMBIO and EFITAL [B007/2007/3-10.2] projects, the latter funded by the Ministerio de Ciencia e Innovación (MICINN). FTM acknowledges support from the European Research Council under the European Community's Seventh Framework Programme [(FP7/2007-2013)/ERC Grant agreement n° 242658].

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
62
Supplementary Material for Chapter 1.
Appendix A. Detailed statistical results of the MANOVA perfomed with growth rates and
slenderness coefficients.
Statistical results of the MANOVA perfomed with growth rate and slenderness coefficient
data in the natural (A, B; separate analyses for each variable) and the anthropogenic (C, D;
separate analyses for each variable) grasslands. Significant (P < 0.05) or marginally
significant (0.05 < P < 0.1) are highlighted in bold and italics, respectively. RGR = relative
annual growth of root collar diameter; SC = slenderness coefficient.
Table S1. MANOVA results showing the overall effect of the factors assayed (microsite and irrigation) on the 5 variables tested (RGR 2007-08 and 2008-09, and SC for 2007, 2008 and 2009) in the natural grassland. This analysis was performed separately for Experiments 1 (Amount treatment) and 2 (Frequency treatment).
Experiment 1: Amount treatment
Factor Pillai´s trace F df P-value Microsite (M) 0.574 4.859 5,18 0.005
Irrigation (I) 0.394 0.933 10,38 0.515 M x I 0.192 0.404 10,38 0.937
Experiment 2: Frecuency treatment
Microsite (M) 0.386 2.518 5,20 0.063
Irrigation (I) 0.402 2.686 5,20 0.052
M x I 0.115 0.520 5,20 0.758
Table S2. Results from separate ANOVA analyses for each variable in the natural grassland. Analyses were conducted separately for Experiments 1 and 2 (Amount and Frequency treatments, respectively). Experiment 1: Amount treatment
Dependent variable
Microsite (M) Irrigation (I) M x I
F df P F df P F df P RGR 2007-08 3.9 1,42 0.06 0.4 2,42 0.66 0.1 2,42 0.91 RGR 2008-09 1.6 1,42 0.22 1.1 2,42 0.35 0.3 2,42 0.73
SC 2007 1.9 1,42 0.19 1.8 2,42 0.18 0.2 2,42 0.79 SC 2008 5.0 1,42 0.04 0.6 2,42 0.56 0.4 2,42 0.69 SC 2009 3.3 1,42 0.08 0.6 2,42 0.55 0.2 2,42 0.80
Experiment 2: Frecuency treatment RGR 2007-08 7.6 1,24 0.01 2.6 1,24 0.12 0.2 1,24 0.65 RGR 2008-09 0.5 1,24 0.48 0.9 1,24 0.77 0.0 1,24 0.93
SC 2007 0.7 1,24 0.80 9.1 1,24 0.01 0.5 1,24 0.48 SC 2008 3.4 1,24 0.08 1.8 1,24 0.20 0.5 1,24 0.49 SC 2009 2.9 1,24 0.1 0.9 1,24 0.36 0.5 1,24 0.48

CHAPTER 1
63
Table S3. MANOVA results showing the overall effect of the factors assayed (microsite and irrigation) on the 5 variables tested (RGR 2007-08 and 2008-09, and SC for 2007, 2008 and 2009) in the anthropogenic grassland. This analysis was performed separately for Experiments 1 (Amount treatment) and 2 (Frequency treatment).
Experiment 1: Amount treatment
Factor Pillai´s trace F Hypothesis/error df P-value
Microsite (M) 0.286 5.056 5,63 0.001
Irrigation (I) 0.238 1.731 10,128 0.081
M x I 0.054 0.358 10,128 0.962 Experiment 2: Frecuency treatment
Microsite (M) 0.341 4.653 5,45 0.002
Irrigation (I) 0.085 0.839 5,45 0.529 M x I 0.045 0.424 5,45 0.829
Table S4. Results from separate ANOVA analyses for each variable in the anthropogenic grassland. Analyses were conducted separately for Experiments 1 and 2 (Amount and Frequency treatments, respectively).
Experiment 1: Amount treatment Dependent
variable Microsite (M) Irrigation (I) M x I
F df P F df P F df P RGR 2007-08 3.3 1,67 0.07 0.3 2,67 0.73 0.0 2,67 0.97 RGR 2008-09 5.3 1,67 0.03 0.3 2,67 0.76 0.8 2,67 0.48
SC 2007 0.6 1,67 0.44 3.0 2,67 0.06 0.3 2,67 0.76 SC 2008 0.0 1,67 0.95 1.2 2,67 0.30 0.1 2,67 0.94 SC 2009 0.17 1,67 0.70 0.2 2,67 0.78 0.6 2,67 0.54
Experiment 2: Frecuency treatment RGR 2007-08 3.4 1,49 0.07 2.1 1,49 0.16 0.3 1,49 0.56 RGR 2008-09 3.4 1,49 0.07 3.7 1,49 0.06 1.8 1,49 0.19
SC 2007 1.0 1,49 0.31 0.4 1,49 0.55 0.1 1,49 0.80 SC 2008 0.7 1,49 0.79 1.0 1,49 0.33 0.2 1,49 0.70 SC 2009 0.3 1,49 0.62 2.7 1,49 0.10 1.6 1,49 0.22

CLIMATE CHANGE AFFECTS PLANT INTERACTIONS
Appendix B: Detailed statistical results of the log-linear analyses performed with survival frequencies.
Hierarchical log-linear results for the different models for both study sites and for each year of study (2007-2009). Table S5 shows the results of
the analyses conducted with the three amount treatments evaluated (+0%, +25% and +50%; Experiment 1), without considering the frequency
levels. Table S6 shows the results of the analyses conducted with the two frequency treatments evaluated (2x and 4x; Experiment 2), without
introducing the amount levels. P-values < 0.05 are shown in bold
Table S5
Factor df
Anthropogenic grassland Natural grassland
2007 2008 2009 2007 2008 2009
G P G P G P G P G P G P
Microsite 2 14.5 0.0007 6.9 0.031 0.54 0.763 5.6 0.062 23.6 >0.001 2 0.365
Irrigation 6 0.06 0.999 11.9 0.018 2.4 0.655 8.8 0.065 1.6 0.813 0.1 0.999
Table S6
Factor df
Anthropogenic grassland Natural grassland
2007 2008 2009 2007 2008 2009
G P G P G P G P G P G P
Microsite 2 12 0.0024 3.5 0.174 1.8 0.410 5.9 0.052 12.9 0.002 2.3 0.314
Irrigation 6 0.5 0.778 1.7 0.434 2.7 0.253 5.7 0.057 0.4 0.816 3.4 0.186

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
Santiago Soliveres, Lucía DeSoto, Fernando.T. Maestre and José Miguel Olano
Manuscrito publicado en:
Perspectives in Plant Ecology, Evolution and Systematics 12: 227-234.
2
Spatio-temporal heterogeneity in abiotic factors mo dulate multiple
ontogenetic shifts between competition and facilita tion

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
66

CHAPTER 2
67
ABSTRACT
Plant-plant interactions are largely influenced by both environmental stress and ontogeny. Despite the effects of each of these factors on the overall outcome of these interactions has received considerable attention during the last years, the joint effects of both factors as drivers of such outcome are poorly understood. We used the combination of spatial pattern analysis, fruit production surveys, carbohydrate assays, sowing experiments and dendrochronological techniques to explore the interaction between Stipa tenacissima (nurse) and Lepidium
subulatum (protégée) in two different slope aspects. This battery of techniques allows us to study the effects of the nurse plant during the whole life cycle of the protégée, and to assess the role of spatio-temporal variability in abiotic stress as a modulator of ontogenetic shifts in plant-plant interactions. Spatial pattern analyses suggested a net facilitative effect of S. tenacissima on L. subulatum. This effect was particularly important during the germination, shifting to competition (growth reduction) early after establishment. Competition was gradually reduced as the shrub aged, suggesting niche differentiation. The magnitude of competition was reduced under low rainfall levels in south-facing slopes, whereas this response was observed due to other abiotic factors in north-facing slopes. Our results highlight the crucial effect that positive interactions at early life-stages have to determine the long-term outcome of a given plant-plant interaction, and the existence of multiple shifts between facilitation and competition along different life-stages of the protégée. They also show how these ontogenetic shifts are modulated by abiotic factors, which differ among slope aspects. These findings may help to refine conceptual and theoretical models about shifts between facilitation and ontogeny under current climate change scenarios.

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
68
INTRODUCTION
he analysis of the spatio-temporal
variation of facilitative and
competitive interactions along
abiotic stress gradients has become a major
research topic in community ecology
during the last two decades (Kikvidze
1996, Maestre and Cortina 2004a,
Kikvidze et al. 2006, Brooker et al. 2008).
In arid and semiarid areas, such variations
are particularly important, as water, which
is the most limiting factor, shows a strong
spatio-temporal variability (Whitford 2002,
Holmgren et al. 2006). These environments
are characterized by a high inter-annual
variability in rainfall distribution, and as a
result, plant recruitment is limited to
particularly rainy years (Holmgren et al.
2006). Furthermore, water availability also
experiences a strong spatial variation
between slope aspects: radiation and
temperatures, and thus water stress, are
higher in south-facing slopes, while in
north-facing slopes water stress and
evapotranspiration are lower (Friedman et
al. 1977, Bellot et al. 2004, Aragón et al.
2007, Pueyo and Alados 2007). These
spatio-temporal changes in water
availability have been pointed as a major
factor defining the final outcome of plant-
plant interactions in drylands (Tielbörger
and Kadmon 2000a, Pugnaire and Luque
2001, Gómez-Aparicio et al. 2004, Miriti
2007).
The relationship between abiotic
stress and the final outcome of plant-plant
interactions is further complicated by the
ontogenetic changes that plants experience
throughout their life cycle, which can
strongly modulate facilitation/competition
shifts (Miriti 2006, Schiffers and
Tielbörger 2006, Armas and Pugnaire
2009). Short-term studies, which form the
core of facilitation/competition research
(see Callaway 2007 for a review), are
insufficient to fully understand the
magnitude of ontogenetic shifts in plant-
plant interactions, but long-term studies are
often logistically prohibitive because of
economic and temporal constraints. Some
studies have overcome these limitations by
using annual plants (Schiffers and
Tielbörger 2006), or by sampling specific
temporal windows of the plant life cycle
(Armas and Pugnaire 2005, 2009, Miriti
2006, Valiente-Banuet and Verdú 2008).
These approaches in isolation are
insufficient to test ontogenetically-driven
facilitation/competition shifts along the
whole plant life, particularly in long-lived
perennial plants, and to assess the effects
of spatio-temporal changes in abiotic stress
on such ontogenetic shifts. These problems
can be circumvented using
dendrochronological techniques, assigning
annual rings to calendar years
(Schweingruber 1988). Since xylem acts as
T

CHAPTER 2
69
conductive area for water and nutrients
within a plant (e.g. Dyer and Bailey 1987,
Bascietto and Scarascia-Mugnozza 2004),
this technique can reconstruct investment
in secondary growth along plant life, and
therefore, act as a measurement of plant
performance in each year during its whole
life.
Although it is known that the
outcome of plant-plant interactions may be
affected by the interaction between abiotic
stress and the ontogeny of the target
species (Goldberg et al. 2001), there is a
lack of studies evaluating the simultaneous
effects of abiotic stress, both in space and
in time, and ontogeny as drivers of on the
outcome of plant-plant interactions (but see
Schiffers and Tielbörger 2006; Sthultz et
al. 2007). Improving our understanding on
the interacting effects of these factors will
allow us to further refine current
conceptual and mathematical models
aiming to predict how plant-plant
interactions change along stress gradients
(Michalet 2007), and to increase the
precision of our estimates about how plant
individuals and communities will respond
to ongoing climate change (Brooker 2006).
In this study, we combine spatial pattern
analyses, sowing experiments,
dendrochronological and reproductive
surveys, and carbohydrate assays to
explore the relationship between the
tussock grass Stipa tenacissima L.
(Poaceae; the nurse plant) and the shrub
Lepidium subulatum L. (Brassicaceae; the
protégée plant) in two slope aspects (north
and south) with contrasting abiotic stress
environments. This combination of
approaches provides us with a continuous
set of data, suitable for testing the presence
of ontogenetic facilitation/competition
shifts throughout the entire life cycle of the
protégée. Furthermore, we aimed to assess
the role of spatio-temporal changes in
abiotic stress (differences between abiotic
factors controlling plant growth and
survival among slope aspects, and
differences between water availability
among years) as a modulator of these
shifts. We tested the following hypotheses:
(i) the germination and survival of L.
subulatum seeds and seedlings will be
higher under S. tenacissima than in
adjacent bare ground zones because of the
improvement of environmental conditions
under the canopy of this nurse plant
(Maestre et al. 2001, 2003, Barberá et al.
2006); (ii) the outcome of the interaction
will shift from facilitation to competition
with shrub age, resulting in less growth
and fruit production of L. subulatum when
growing under the canopy of S.
tenacissima (Miriti 2006); (iii) given that
both S. tenacissima and L. subulatum are
primarily stress-tolerant species (Pugnaire

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
70
et al., 1996b, Palacio et al. 2007), and that
water deficit (a resource-related stress) is
the main limiting factor, we expect to find
facilitative interactions mainly under
moderate abiotic stress levels, (Maestre et
al. 2009a). This will happen because the
positive effect of microclimatic
amelioration is limited after a threshold
under high levels of abiotic stress, where
competition, mainly for water, overcomes
the positive effect of this amelioration and
competition arises again (Maestre and
Cortina 2004a); and (iv) the magnitude of
facilitation and competition is modulated
by spatio-temporal changes in climatic
conditions.
METHODS
TARGET SPECIES
Lepidium subulatum is a dwarf summer
deciduous shrub linked to gypsum
outcrops. It is distributed along the
Western Mediterranean where it coexists
with S. tenacissima. Seeds are small,
exhibiting atelechory (Escudero et al.
2000) and forming a small permanent seed
bank (Caballero et al. 2008). Emergence
concentrates in winter, but spans to June,
with densities ranging from 19 to 700
seedlings m2 (Escudero et al. 2000, J.M.
Olano, unpubl. data). Plant recruitment
bottlenecks occur during the first summer
after germination, when survival rates
range from 0.1% to 10%, with high
survivorship linked to especially
favourable years (Escudero et al. 2000,
J.M. Olano, unpubl. data). Annual survival
rates increase sharply afterwards, reaching
71–95% for adults, depending mainly on
autumn and spring conditions.
Flowering starts at 2-4 years (M.
Eugenio, pers. com.). Primary growth
occurs in two pulses, from mid February to
June, and from September to November,
respectively (Palacio and Montserrat-Martí
2005, Palacio et al. 2007). Secondary
growth also occurs in spring. Flowering
and fruiting stages last from April to June.
A detailed description on the natural
history of S. tenacissima and the grasslands
it forms is given in Maestre et al. (2009b).
STUDY AREA
Three sites were selected in central Spain
for this study: Aranjuez (40º10´30´´N, 31º
54´09´´W; 545 m.a.s.l.); Tielmes (40º
12´40´´N, 31º25´02´´W; 595 m.a.s.l.); and
Noblejas (40º10´03´´N, 31º37´01´´W; 526
m.a.s.l.). Their climate is Mediterranean
semiarid, with average annual precipitation
of 388 mm, characterized by a high inter-
annual variability and a characteristic
strong summer drought. Mean annual
temperature is 14.6 ºC, ranking from 25 ºC
in July to 5.6 ºC in January (Data from
National Meteorological Service, 1994–
2005. Marqués et al. 2008). The three sites

CHAPTER 2
71
were located on gypsum-rich soils,
classified as Typic Gypsiorthid (Tielmes
and Noblejas) and Xeric Haplogypsid
(Aranjuez; Soil Survey Staff 1994).
Vegetation was in all cases an open steppe
dominated by S. tenacissima, and
contained shrub species like L. subulatum,
Retama sphaerocarpa (L.) Boiss. and
Helianthemum squamatum (L.) Dum.
Cours. Perennial plant cover is below 45%
in all cases.
EXPERIMENTAL DESIGN
At each site, experimental plots in north-
and south-aspect slopes were established,
with slopes varying between 13º and 22º.
Perennial cover was different depending on
the slope aspect in all the studied areas:
31% vs. 33%; 24% vs. 42%; and 32% vs.
35% for south- vs. north-aspect slopes in
Tielmes, Aranjuez and Noblejas,
respectively. Since cover can be a good
surrogate of productivity in ecosystems
such as those studied (Flombaum and Sala
2009), and productivity is a good proxy for
abiotic stress at the level of entire plant
communities (Lortie and Callaway 2006),
we assume that these differences are
related to higher levels of abiotic stress in
the south- than in the north-facing slopes.
This agrees with many studies conducted
in arid and semiarid areas showing higher
abiotic stress in south- vs. north-aspect
slopes (e.g. Friedman et al. 1977,
Sternberg and Shoshany 2001, Bellot et al.
2004; Aragón et al. 2008, Pueyo and
Alados 2007). Differences in perennial
cover between slope aspects were
particularly evident in the Aranjuez site
(Appendix C in Supplementary Material),
so this site was selected to carry out the
bulk of the fieldwork in this study.
OBSERVATIONAL MEASUREMENTS
In April 2008, five 25 m × 4 m transects
were randomly established in each of the
experimental plots (30 transects in total).
Every L. subulatum individual found along
the transect band was registered. Those
individuals located at distances shorter
than 20 cm and larger than 50 cm from the
edge of a S. tenacissima tussock were
considered as growing in association with
S. tenacissima and in isolation,
respectively. These situations are hereafter
called Tussock and Open microsites,
respectively. This distance has been used
as separation between microsites in other
studies with S. tenacissima, detecting
significant differences in both biotic and
abiotic features between Tussock and Open
microsites (e.g. Maestre et al. 2001, 2003,
2009b). L. subulatum individuals growing
at distances among 20–50 cm from the
edge of a S. tenacissima tussock were not
considered for further analyses.

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
72
SOWING EXPERIMENT
In October 2007, a seed germination
experiment was conducted in the Aranjuez
site. It was designed as a fully factorial
experiment with two treatments: slope
aspect (north vs. south) and microsite
(open areas devoid of vascular vegetation,
north and south-face of S. tenacissima
tussocks). Ten replicates were established
per treatment combination (each consisting
in a 25 cm × 25 cm plot), and 75
commercial seeds of L. subulatum were
seeded in each replicate (25 holes, 3 seeds
per hole). We chose this approach over the
alternative neighbor removal approach,
because neighbor removal does not erase
the facilitative legacy effects of a nurse
plant upon soil infiltration and fertility.
Seeds were buried at 0.5 cm to avoid ant
depredation, irrigated with 40 ml of water
and protected from rabbits (Oryctolagus
cuniculus L.) by using a metallic mesh that
did not shade the seeding site. A
germination test conducted under
controlled conditions revealed that the total
germination rate of the pool of seeds
employed was 89% after one month in a
growth chamber (16 light hours at 20ºC
and 8 dark hours at 10ºC). Seed emergence
and seedling survival were monitored
monthly until July 2008, when all
germinated seedlings died during the
summer drought. Because of this extreme
mortality event and the lack of germination
in Open sites, sapling survival data
analyses cannot be provided.
DENDROCHRONOLOGICAL
SURVEYS
In June 2007, adult individuals of L.
subulatum were randomly selected in the
Aranjuez site for dendrochronological
measurements. These individuals were
chosen among those naturally growing
under four different conditions, resulting
from the combination of two microsites
(Tussock vs. Open) and slope aspects
(north vs. south); 16 individuals were
selected for each combination (64 in total).
After harvesting, a section of the stem
including the root collar was selected to
measure the annual growth ring widths as
an indicator of plant growth over the
course of its life. Annual rings were dated
and measured following standard
denchronological techniques as detailed in
Appendix D in Supplementary Material. A
section of the main root of the same plants
was also collected to measure the content
of non-structural carbohydrates using the
anthrone method (see Olano et al. 2006 for
a full account of the methodology). Two
different fractions of non-structural
carbohydrates were measured in this study:
nonsoluble and soluble carbohydrates. In
L. subulatum, non-soluble carbohydrates
are used to overcome respiration rates in
the leafless plant during summer drought

CHAPTER 2
73
(Palacio et al. 2007). Therefore, low
contents in non-soluble carbohydrates may
compromise the ability of this species to
survive this critical moment. We interpret
differences in this variable between
microsites or slope aspects as differences
in the ability of L. subulatum to respond to
summer drought stress. On the other hand,
soluble carbohydrates provide a surplus of
sugars that can be stored for use under
favourable conditions (Chapin et al. 1990),
and that are susceptible to be immediately
allocated to functions such as growth.
Thus, higher contents of soluble
carbohydrates in a given combination of
microsite/slope aspect may indicate that
the plant experiences more benign
conditions there. Both soluble and non-
soluble carbohydrates inform us about the
status of the reserves of the plant, and thus
are an integrative measurement of plant
performance during the whole year. These
variables were measured in June, just
before summer drought, and in the main
root because this organ and date match
with the maximum starch content organ
and period of the year for L. subulatum,
respectively (Palacio et al. 2007).
FRUIT/INFRUCTESCENCE RATIO
SURVEYS
In June 2008, ten reproductive L.
subulatum individuals in each combination
of two slope aspects (north vs. south) and
microsites (Tussock vs. Open) were
randomly selected in the Aranjuez site. Ten
infructescences per plant were randomly
chosen, and the number of fruits in each
infructescence was registered. The canopy
area of each sampled individual was
calculated using the ellipse formula with
the diameters parallel and perpendicular to
slope. This measurement was introduced in
the analysis as covariate to control for
plant size.
STATISTICAL ANALYSES
The frequency of naturally occurring L.
subulatum individuals in the surveyed plots
was analyzed by using a Chi-square
goodness of fit test. Our null hypothesis
was that L. subulatum individuals have a
random spatial pattern (depending directly
on the cover of each microsite). Data were
tested for independence in 6 separate one-
way tables (resulting from each
combination of site and slope aspect)
including only the microsite factor
(Tussock vs. Open). A joint analysis that
would permit testing the interaction
between the factors included in the model
was not possible because the relative
Tussock/Open microsites cover (and
therefore the expected frequencies) in the
different site × slope aspect combinations
were not equiprobable, a general

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
74
assumption of the null hypothesis used
when analyzing contingency tables with
multiple factors. To calculate the expected
frequencies, the number of total plants
found in each transect were multiplied per
the percentage of cover of each microsite.
The sum of predicted/observed frequencies
of the five transects per each site × slope
aspect combination was used to run the
Chi-square test. To adjust for the increase
in Type I error because of multiple testing,
the Bonferroni correction was used
(corrected a: 0.05/6 = 0.0083).
The effects of slope aspect (north
vs. south) and microsite (open areas, north
face of S. tenacissima tussocks and south
face of S. tenacissima tussocks) on the
cumulative number of germinated seeds
were tested by using generalized linear
models (GLMs). GLMs were run with a
Poisson error distribution combined with a
log link function; Type I log-likelihood
ratios were used to analyze main effects.
As recommended to counteract data over-
dispersion and to adjust the statistics
properly, the scale parameter was
estimated by dividing the square root of the
Pearson’s Chi-square statistic by the
degrees of freedom (McCullagh and
Nelder, 1989). Differences between
microsites were tested using a post-hoc test
based on least-square means. Growth data
obtained from dendrochronological
measurements were analyzed using two
complementary approaches. First, the ring
width data of all individuals measured in a
given combination of microsite and slope
aspect were grouped according to L.
subulatum age, independently of
recruitment year, and averaged. The
average ring width of each of these four
groups was used to estimate the effect of S.
tenacissima on the growth of L. subulatum
throughout the ontogeny of the later by
using the relative interaction index (RII;
Armas et al. 2004). This index was
calculated as (Gst – Go)/(Gst + Go), where
Gst and Go are the average ring widths of
L. subulatum individuals growing in
Tussock and Open microsites, respectively.
RII values were obtained from 1- to 12-
year old individuals to maintain enough
sample size along the whole age range (n =
12–16 individuals in all the groups for all
the ages analyzed; sample size
dramatically decreased for older
individuals). This 12-year period recovers
different ontogenetic stages of the protégée
plant: seedling (first year), juvenile (from 2
to 4 years) and adult stage (from 5 years
onward). Therefore, it is sufficient to test
possible ontogenetic shifts in the sign of
the interaction studied. To remove
potential autocorrelation derived from
repeated measures of ring width, a Prais–
Winsten autoregression was performed to
test the relationship between the values of
the RII index and L. subulatum age. This

CHAPTER 2
75
analysis takes into account the non-
independence of the samples by estimating
a regression equation whose errors follow
a first-order autoregressive process (SPSS,
2004). Second, ring width data were
averaged for each calendar year, without
taking into account the age of each L.
subulatum individual, to evaluate the effect
of climate inter-annual variability on
annual growth rates. The RII index was
calculated with these data as described
above, evaluating the relationship between
this index and the rainfall registered in
March by using linear regression. March
rainfall was selected as the best possible
rainfall predictor of abiotic stress because
it was highly correlated with the
standardized ring width series of L.
subulatum (Pearson’s correlation index:
0.78 and 0.62, P < 0.05 for ring widths
growth of L. subulatum in Open microsites
in south- and north-aspect slopes,
respectively, see Appendix E in
Supplementary Material). To ensure that
no other important rainfall variable was
ruled out without the proper test, we
evaluated the bivariate correlations
between the width growth of L. subulatum
rings and the rest of possible rainfall
indicators of abiotic stress: rainfall of the
rest of the months, total annual rainfall,
monthly rainfall of September–December
of the previous year, cumulative rainfall of
spring (March-May), and the cumulative
rainfall of the two annual pulses of L.
subulatum growth (February-June and
September-November). Of all these
correlations, only March rainfall, which is
prior to the main pulse of L. subulatum
primary growth (Palacio et al. 2008), was
statistically correlated with the width
growth of L. subulatum rings (Appendix E
in Supplementary Material). Therefore, we
selected rainfall in this month as our
surrogate of abiotic stress. Separate
regression and autoregression analyses
were conducted for north and south slopes.
This approach allowed us to test
differences in the effects of rainfall on the
final outcome of the interaction depending
on the variability in the abiotic conditions
among the two slope aspects considered.
The relative importance of both ontogeny
and abiotic stress as drivers of the outcome
of the interaction studied was also
explored. For doing this, three sequential
analyses were run. First, the effects of
microsite (Tussock vs. Open) and slope
aspect (north vs. south) on the growth of L.
subulatum were evaluated using repeated
measures ANOVA. For doing this, ring
width data were grouped according to L.
subulatum age, independently of the
calendar year when they were formed. As
abiotic conditions are different in north and
south slopes due to different irradiation,

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
76
temperature and water stress levels
(Friedman et al. 1977, Bellot et al. 2004,
Aragón et al. 2008), separated analyses
were run for each slope aspect. This
procedure allowed us to test potential
differences in the nature of abiotic factors
controlling the final outcome of the
interaction at each slope aspect. The
second step was to introduce the
recruitment year as covariate in this
analysis to assess the specific weight of
climatic variability. L. subulatum
individuals were of contrasting ages
(recruitment dates of individuals analyzed
vary from 1981 to 2000), so the ring width
corresponding to each individual age may
be influenced by the particular abiotic
conditions in the period they grew, and
therefore a high intra-group variability was
expected to be found. As abiotic factors
can produce non-random differences
between individuals, and these differences
could mask the effect of an experimental
treatment, its inclusion as a covariate has
been recommended to gain power when
testing the effects of the factors of interest,
particularly when a high intra-group
variability is found (see Engqvist 2005 and
references therein). If the effect of this
covariate is significant, and changes that of
S. tenacissima, this would suggest that
abiotic stress is modulating the net effects
of S. tenacissima on the growth of L.
subulatum during ontogeny. Lastly, the
same analysis was performed but changing
the covariate to the median of March
rainfall during the lifetime of each
individual included in the analysis. March
rainfall is a key driver of L. subulatum
growth (Palacio and Montserrat-Martí
2005, Palacio et al. 2007; Appendix E in
Supplementary Material), and its median
would be a good estimator of the water
stress level suffered by each individual
during its entire life. With this analysis we
aimed to differentiate the effects of rainfall
from those of unmeasured abiotic factors
characterizing each year as modulators of
ontogenetic shifts in plant–plant
interactions. The effects of microsite
(Tussock vs. Open) and slope aspect (north
vs. south) on the mean
fruits/infructescence ratio and on soluble
and non-soluble carbohydrates content
were evaluated with a two-way ANCOVA,
where the size of L. subulatum individuals
was used as a covariate. GLM analyses
were carried out using the GENMOD
procedure of SAS 9.0 (SAS Institute, Cary,
NC, USA). The remaining statistical
analyses were conducted using SPSS 13.0
for Windows (Chicago, IL, USA). The
non-soluble carbohydrates content did not
meet the ANOVA assumptions (normality
and homocedasticity), and was transformed
by using the arcsin transformation. The rest
of the data met these assumptions, and
were not transformed.

CHAPTER 2
77
RESULTS
Lepidium subulatum was more frequent
than expected under S. tenacissima
canopies than in open microsites in five of
the six site × slope aspect combinations
studied (Table 2.1). The overall cumulative
germination rate was very low due to the
drought conditions of the study year (Fig.
2.1). However, it was higher under the
canopy of S. tenacissima than in Open
microsites. A significant microsite × slope
aspect interaction was also found,
suggesting that the magnitude of the
differences between microsites varied with
the slope aspect considered. More
germination events occurred in the north-
than in the south-face of S. tenacissima
canopies in the south slope (χ2 = 9.2; P =
0.002), but no differences between the
north- and south-face of the canopy were
found in the north slope (χ2 = 1.2; P =
0.277).
Table 2.1. Expected and observed frequency of Lepidium subulatum individuals found under the canopy of Stipa tenacissima (Stipa) and in bare ground areas (Open) in north- and south-facing slope aspects at the three studied sites. Data represent the sum of the number of individuals of the five transects measured in each site × slope aspect combination. Chi-square test results evaluate the effects of microsite (χ2 and P-value) in the various site and slope aspect combinations. Significant results (Bonferroni-corrected α) are in bold.
Site Slope
Aspect
Expected
frequencies
Observed
frequencies χ2
P-value
Open Stipa Open Stipa
Tielmes South 75.5 29.5 40 26 17.1 <0.0001
North 63.62 23.38 52 9 10.97 0.009
Aranjuez South 176.95 28.05 137 29 9.05 0.002
North 63.56 33.44 24 34 24.63 <0.0001
Noblejas South 99.44 33.56 89 17 9.27 0.002
North 146.42 63.58 119 49 8.48 0.004

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
78
Figure 2.1. Cumulative germinations registered in bare ground areas (Open), north face of Stipa tenacissima tussocks (Stipa north) and south face of S. tenacissima tussocks (Stipa south). Data represent mean + SE; n = 10. Different letters indicate significant differences between microsites in each slope aspect (post-hoc test based on the differences of least-square means).
A significant positive relationship between
the RII values and L. subulatum age was
found, suggesting that the negative effect
of S. tenacissima on the growth of L.
subulatum decreased as individuals aged
(Fig. 2.2A). This relationship was found in
both slope aspects (R2 = 0.83 and 0.51 for
south and north slopes, respectively). A
negative linear relationship between the
RII values and March rainfall was found in
the south slope, suggesting that negative
interactions dominated in years of high
March rainfall (Fig. 2.2B). No significant
relationships were found in the north slope.
Repeated-measures ANOVA showed a
negative effect of S. tenacissima on the
growth of L. subulatum individuals
(Appendix D in Supplementary Material),
which was particularly evident in the north
slope (north: F1,32 = 5.54, P = 0.025; south:
F1,27 = 3.88, P = 0.059). When recruitment
year was introduced as a covariate in this
analysis, the effects of microsite became
non-significant in both the south
(recruitment year: F1,26 = 6.71, P = 0.015;
microsite: F1,26 = 0.46, P = 0.503) and
north (recruitment year: F1,31 = 4.24, P =
0.048; microsite: F1,31 = 3.22, P = 0.082)
slopes. When the median of March
precipitation was used as a covariate, it did
not change substantially the effects of S.
tenacissima on the growth of L. subulatum
in the north slope (median March
precipitation: F1,31 = 0.89, P = 0.35;
microsite F1,31 = 4.66, P = 0.039), but it did
so in the south slope (median March

CHAPTER 2
79
precipitation: F1,26 = 3.63, P = 0.068;
microsite: F1,26 = 0.58, P = 0.454). Non-
structural carbohydrate content was higher
in Open than in Tussock microsites (F1,58 =
4.44; P = 0.039 and F1,58 = 4.71; P = 0.034,
for soluble and non-soluble carbohydrates,
respectively; Fig. 2.3).
Figure 2.2. (A) Relationships between the effect size of Stipa tenacissima on the growth of Lepidium subulatum, as measured by the RII index, and the age of L. subulatum. (B) Relationships between values of this index and the median of rainfall registered in March during the period 1995–2007. Results of significant autoregressions (A) and linear regressions (B) are shown. Each RII value is obtained by averaging growth data from 12 to 16 L. subulatum individuals.
B
A

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
80
Figure 2.3. Soluble and non-soluble root carbohydrates content (black and grey bars, respectively) of Lepidium subulatum individuals harvested into two different slope aspects (north vs. south) and growing underneath Stipa tenacissima canopy (Stipa) or in areas without perennial vegetation (Open). Data represent means ± SE; n = 16. Asterisks mark significant differences in root carbohydrates content between microsites within each slope aspect. Different letters mark significant differences among slope aspects for soluble (normal letters) and non-soluble carbohydrates (capital letters).
Figure 2.4. Number of fruits per infructescence (mean ± SE; n = 10) of Lepidium subulatum individuals growing into two different slope aspects (north vs. south) and microsites (underneath Stipa tenacissima canopy, Stipa, or in areas without perennial vegetation, Open).

CHAPTER 2
81
A higher content of soluble carbohydrates
was found in north- than in south-aspect
slopes (F1,58 = 28.82; P < 0.001), but non-
significant effects of slope aspect were
found when analyzing non-soluble
carbohydrates. The average number of
fruits/infrutescence ratio was affected by
the size of L. subulatum individuals (F1,35 =
10.70, P = 0.002), as larger plants had
more fruits. However, no significant
effects of slope aspect and microsite were
found (slope aspect: F1,35 = 0.91, P=0.346;
microsite: F1,35 = 0.04, P = 0.845. Fig. 2.4).
DISCUSSION
In contrast to studies showing a single
facilitation/competition shift throughout
the ontogeny of perennial plants (Miriti
2006, Valiente-Banuet and Verdú 2008,
Armas and Pugnaire 2009), our results
provide evidence of multiple ontogenetic
shifts between facilitation/competition
during the life cycle of the protégée. We
also found that spatio-temporal changes in
abiotic stress modulated these ontogenetic
shifts. To our knowledge, these responses
have not been described before. While the
importance of studying different abiotic
stressors and their interaction has been
highlighted (Holmgren et al. 1997, Riginos
et al. 2005, Baumeister and Callaway
2006), our study illustrates how the spatial
variability of these stressors, even at small
spatial scales, and their interaction with
ontogeny determine the final outcome of
plant–plant interactions.
As expected, we found higher
germination rates under the canopy of S.
tenacissima than in bare ground areas in
both slope aspects (Barberá et al. 2006,
Schiffers and Tielbörger 2006). This effect
seems crucial to define the final outcome
of the interaction studied, as indicated by
the strong net positive effect of S.
tenacissima on the abundance of L.
subulatum individuals in most of the
situations and sites studied. It is interesting
to note, however, that if only two
microsites (e.g. open vs. north face of S.
tenacissima tussocks) would had been
considered, as it has been done by most
facilitation studies (see Callaway 2007),
the higher positive effect of S. tenacissima
on the germination of L. subulatum found
in the south-aspect slope would point to a
higher facilitative effect with increases in
abiotic stress (Bertness and Callaway
1994). However, the contrasting results
found between slope aspects when
evaluating the germination of L. subulatum
in the north- and south-faces of S.
tenacissima tussocks suggest a complex
interplay between abiotic stress and
facilitation, which is strongly influenced
by spatial variability in these abiotic
stressors (e.g. light availability; Parker and

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
82
Muller 1982, Marañón and Bartolome
1993; S. Soliveres, unpubl. data). One
possible explanation for these results is
that in south-aspect slopes, where water
stress is higher than in north-aspect slopes
(Bellot et al. 2004, Aragón et al. 2008),
seeds predominantly germinate under the
north face of tussocks, where the
improvement in microclimate by S.
tenacissima is maximal due to water inputs
coming from run-off and to the shadow
provided by the canopy of this species
(Maestre et al. 2001, 2003). This microsite
preference is not so evident in the north-
aspect slope, as seed germination in both
faces of S. tenacissima tussocks was
similar. These results seems to suggest a
trade-off between microclimatic
amelioration in the north face of S.
tenacissima tussocks (Maestre et al. 2003)
and the increase in interference
competition with other neighboring plants
(Goldberg et al. 2001, Miriti 2006), as they
are less abundant under the south-face of
the tussocks (S. Soliveres, pers. obs.).
Stipa tenacissima had a negative
effect on the growth and carbohydrate
content of L. subulatum, suggesting that
not only sink activity (growth), but also
resource levels (carbohydrate content) are
lower in this microsite. Although we
cannot statistically differentiate the effect
found on L. subulatum growth from a
neutral one (we do not have error bars as
we grouped our data by the four possible
treatments combinations and averaged
them to calculate RII), there is a clear
reduction trend on the strength of the
negative effect on growth as L. subulatum
individuals aged. This effect, together with
the positive effect found on germination,
points to multiple facilitation/competition
shifts along the life cycle of L. subulatum.
Differences between our results and those
from previous studies (Miriti 2006,
Valiente-Banuet and Verdú 2008, Armas
and Pugnaire 2009) can be explained
because our study focuses on a grass–
woody plant interaction, while previous
studies have focused on woody–woody
plant interactions. In a grass–woody
interaction, the growth of the woody
individuals as they age helps to avoid
water and light competition from the
grasses (Fowler 1986, Van Auken 2000),
whereas the benefits of shade and
increased soil resources under the canopy
of the later still exist (Maestre et al. 2003).
This may render competition less
important, as effective nicheseparation is
likely to occur with increasing age (Fowler
1986, Van Auken 2000, Armas and
Pugnaire 2005). This is less likely to occur
when the nurse and protégée share the
same ecological traits (e.g. annuals:
Goldberg et al. 2001; Schiffers and
Tielbörger 2006; or shrubs: Miriti 2006),
as they are likely exhibiting greater niche

CHAPTER 2
83
overlap (Maestre et al. 2009a) and the
increase of standing biomass can lead to an
increase in resource competition (Schiffers
and Tielbörger 2006). This difference
suggests that differentiation in the traits of
the plants involved are likely to play an
important role in determining ontogenetic
facilitation/competition shifts (Armas and
Pugnaire 2009), and thus should be
explicitly taken into account when trying
to generalize the results of particular
studies.
Apart from the life story traits of
the interacting species, differences in the
tolerance to abiotic stress and the
competitive ability of the interacting
species are crucial when studying plant–
plant interactions (Liancourt et al. 2005,
Maestre et al. 2009a, Gross et al. 2010). In
our study, two stress-tolerant species
coexist along a temporal stress gradient
driven by water availability of each year.
As both species are stress tolerant, and
water is a resource-related factor, positive
interactions are expected to be dominant at
intermediate levels of abiotic stress
because of the existence of thresholds in
both sides of the gradient (Maestre et al.
2009a). This prediction was not met, as we
found less competition under more
stressful (i.e. lower rainfall) conditions in
the south slope, and no relationships
between abiotic stress and the outcome of
the interaction were found in the north
slope. The relationship with rainfall
observed in the south slope can be
explained by the compensation of the
negative effect of shade on growth with the
reduction in the water stress experienced
by L. subulatum, particularly in dry years
(Fig. 2.2B; Holmgren et al. 1997, Hastwell
and Facelli 2003). In our case, it seems that
rainfall modulated the trend toward
escaping light competition with age, being
shade less negative under dry years. This is
suggested by the reduction observed in the
negative effects of S. tenacissima on the
growth of L. subulatum when rainfall was
introduced as a covariate. In the north
slope, the ontogenetic trend was the same,
but abiotic factors other than rainfall
modulated this trend, as indicated by the
change in microsite effect along ontogeny
when recruitment year, but not rainfall,
was introduced in the analysis. Our study
shows how the same nurse effect, and its
interaction with ontogeny, depends on the
spatio-temporal changes in the overall
amount of abiotic stress experienced by the
interacting individuals, and on the
resources driving such stress (Holmgren et
al. 1997, Hastwell and Facelli 2003).
Moreover, we show how a longitudinal
track of competitive or facilitative
interactions and its relationship with
climatic conditions can be easily obtained

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
84
using secondary growth data present in
annual rings.
CONCLUDING REMARKS
In contrast with previous studies, which
found an increase in competition as
prote´ge´e plants grew (Miriti 2006,
Schiffers and Tielbörger 2006, Valiente-
Banuet and Verdú 2008), we found a
reduction of competition as plant aged.
This result may be influenced by
differences in ecological traits of species
involved in our study comparing to the
previous studies, which can result in an
effective niche separation between grasses
and shrubs, and therefore a reduction of
competition (e.g. Armas and Pugnaire
2005; our case) or in a niche overlap,
which together with the increase in
biomass can lead to higher competition
(e.g. Miriti 2006 for shrubs; Schiffers and
Tielbörger 2006 for annuals). Because
perennial grasses conform an important
component of many vegetation formations
worldwide (Zimmermann et al. 2010), our
results provide useful information to refine
current theoretical models about
facilitation/competition shifts along
ontogeny of perennial species. Our results
also provide important insights on how
spatio-temporal changes in abiotic stress
can modulate multiple
facilitation/competition shifts. As rainfall
increased competition escape in south-
aspect slopes, other factors were more
important in north-aspect slopes. Despite
the mostly negative effect of the nurse
plant on growth and reserve accumulation,
the positive effect found on early stages of
the life cycle of the protégée (germination)
may be driving the net positive sign of the
interaction, as demonstrated by the spatial
aggregation found between studied species.
Given the implications of understanding
how plant–plant interactions change along
stress gradients for accurately predicting
global change impacts on communities and
ecosystems (Brooker 2006), future studies
should pay special attention to the
interplay between abiotic stress and
ontogeny as joint drivers of
facilitation/competition shifts, and more
specifically, on the effect of nurse plants in
key stages of the life cycle under different
environmental conditions (Goldberg et al.
2001). This is particularly true when
working with long-lived species in
stressful environments.
ACKNOWLEDGMENTS We thank M. Bowker for an early revision on a previous version of this manuscript. E. Marcos, P. García-Palacios, A.P. Castillo, E. Pigem, C. Alcalá, J.C. Rubio (CESEFOR) and M. Méndez for their help during field and laboratory work. SS was supported by a fellowship from Fundación Biodiversidad-CINTRA (EXPERTAL project). LDS was supported by a fellowship from Junta de Castilla y León. FTM was supported by a ‘‘Ramón y Cajal’’ contract from the Spanish Ministerio de Ciencia e Innovación (MICINN), cofunded by the European Social Fund, by the British Ecological Society (Studentship 231/1975), and by the MICCIN project CGL2008-00986-E/BOS. JMO was supported by Junta de Castilla y León

CHAPTER 2
85
VA069A07 project. This research was supported by the EXPERTAL and INTERCAMBIO (BIOCON06/105) projects, funded by Fundación
Biodiversidad-CINTRA and Fundación BBVA, respectively.

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
86
Supplementary material for Chapter 2
Appendix C. Pictures showing differences among perennial cover in north (picture above) vs.
south (picture below) aspect slopes in the Aranjuez study site.

CHAPTER 2
87
Appendix D. Detailed description of growth measurements and ring-width growth data
used
After manually polishing Lepidium disc surfaces until the xylem cellular structure was clearly
visible, ring widths were visually dated by assigning calendar years (Stokes and Smiley 1968)
and measured to the nearest 0.001 mm using a sliding-stage micrometer (Velmex, Bloomfield
INC., NY, US) interfaced with a computer. The COFECHA software (Grissino-Mayer 2001)
was used to quantitatively check for cross-dating errors. The synchronised and highly inter-
correlated ring-width chronologies for each combination of microsite and slope were selected
from the pool of raw chronologies and used to build a master chronology. The master
chronologies represent the common ring-width growth for each combination of factors
(Figure S1). Master chronologies were standardized with the ARSTAN computer program
(Cook and Holmes 1996) by fitting to a spline function with a 50% frequency response of 32
yr, which was flexible enough to reduce the non-climatic variance by preserving high-
frequency climatic information.
Figure S1. Ring-width growth data in north and south slopes as Lepidium subulatum individuals aged. Stipa and Open indicate individuals located less than 20 cm and further away than 50 cm from the canopy of a Stipa tenacissima tussock. Most individuals from the Stipa microsite were located under its canopy. These data were used to calculate Relative Interaction Indexes (RII) shown in Figure 2.2 of the main text. Data represent means ± SE (n = 12-16).

MULTIPLE ONTOGENETIC SHIFTS IN PLANT INTERACTIONS
88
Appendix E. Climate Response of Lepidium subulatum growth
The obtained residual chronologies were correlated against monthly rainfall during the
growing season (September of the previous year to June) to assess which months determined
plant growth and can be used as indicators of stress level. Precipitation data were obtained
from the nearest meteorological station (period 1995–2007; Dosbarrios Meteorological
Station, located 11 km SE from Noblejas site).
Table S7. Pearson two-tailed correlations (r) between the standardized ring width growth data of Lepidium subulatum for the four slope aspect × microsite combinations and rainfall registered at the nearest weather station (39º53’04’’N, 3º28’33’’W. 714 m.a.s.l.). We consider each month separately and also the sum of the rainfall registered during February-June, September-November and March-May, which match with the growth pulses of L. subulatum. In all cases, n = 12. Stipa and Open indicate individuals located less than 20 cm and further away than 50 cm from the canopy of a Stipa tenacissima tussock, respectively. Most individuals from the Stipa microsite were located under its canopy. Correlations with P < 0.05 are in bold.
North -aspect slope
South -aspect slope
Open Stipa Open Stipa
r P r P r P r P Sep prev. 0.12 0.71 0.23 0.47 0.12 0.71 0.01 0.97 Oct. prev. 0.59 0.04 0.09 0.78 0.41 0.19 0.01 0.97 Nov. prev. -0.46 0.13 -0.09 0.78 -0.08 0.81 0.33 0.30 Dec. prev. -0.48 0.15 -0.19 0.56 -0.12 0.7 0.42 0.17 January -0.42 0.17 -0.21 0.51 0.06 0.85 0.33 0.29 February 0.22 0.49 -0.3 0.34 -0.1 0.82 -0.12 0.70 March 0.77 <0.01 -0.24 0.45 0.62 0.03 0.43 0.17 April -0.02 0.94 -0.03 0.92 -0.01 0.99 -0.04 0.91 May -0.31 0.34 -0.04 0.91 0.09 0.78 0.5 0.10 June -0.13 0.69 -0.27 0.39 0.18 0.58 -0.21 0.51 September -0.22 0.48 0.31 0.33 -0.06 0.86 0.46 0.13 October 0.33 0.29 0.18 0.58 -0.1 0.76 -0.4 0.20 November -0.25 0.44 -0.19 0.56 -0.07 0.83 0.06 0.86 December -0.23 0.48 -0.09 0.78 0.02 0.96 0.41 0.19 Annual -0.15 0.64 -0.34 0.29 0.14 0.67 0.46 0.13 Feb-June 0.17 0.61 -0.29 0.37 0.32 0.32 0.37 0.24 Sep-Nov. 0.26 0.42 0.14 0.67 0.39 0.21 0.28 0.38 March-May 0.15 0.65 -0.16 0.63 0.36 0.25 0.54 0.07

Santiago Soliveres, Pablo García-Palacios, Andrea P. Castillo-Monroy, Fernando T. Maestre,
Adrián Escudero and Fernando Valladares
Manuscrito publicado en: Oikos (en prensa). D.O.I. 10.1111/j.1600-0706.2010.18993.x
3
Temporal dynamics of herbivory and water availabili ty
interactively modulate the outcome of a grass-shrub interaction
in a semiarid ecosystem

BIOTIC AND ABIOTIC STRESSORS AFFECT PLANT-PLANT INTERACTIONS
90

CHAPTER 3
91
ABSTRACT
The study of plant-plant interactions along grazing and abiotic stress gradients is a major research topic in plant ecology, but the joint effects of both stressors on the outcome of plant-plant interactions remains poorly understood. We used two different factorial experiments conducted in a semiarid Mediterranean steppe to assess: 1) the role of the perennial grass Stipa tenacissima, a low-palatability species, providing protection from rabbit herbivory to the shrub Retama sphaerocarpa (Experiment 1), and 2) the effects of environmental amelioration provided by Stipa on the recovery of Retama after rabbit damage under two contrasted levels of water availability (Experiment 2). In the Experiment 1, water stress worked as an indirect modulator of herbivore protection by Stipa. This species protected Retama seedlings from rabbit herbivory during the wetter conditions of spring and winter, but this effect dissapeared when rabbit pressure on Retama increased during summer drought due to the decrease in alternative food resources. In the Experiment 2, Stipa exerted a negative effect on the survival of Retama seedlings during the three years of the experiment, regardless of inter-annual differences in rainfall or the watering level applied. This negative effect was mainly due to excessive shading. However, Stipa increased Retama recovery after initial rabbit impact, overriding in part this negative shade effect. Conversely, Stipa impact on the Fv/Fm of Retama seedlings depended on the intra-annual water dynamics and its experimental manipulation, overall contradicting predictions from the Stress-Gradient Hypothesis. The complex interactions found between herbivory, microclimatic amelioration from Stipa, and water availability as drivers of Retama performance illustrate the importance of considering the temporal dynamics of both biotic and abiotic stressors to fully understand the outcome of plant-plant interactions.

BIOTIC AND ABIOTIC STRESSORS AFFECT PLANT-PLANT INTERACTIONS
92
INTRODUCTION
nteractions among plants can be
negative, neutral, or positive,
depending their direction and
magnitude on the level and type of the
stressors governing a given community (see
Callaway 2007 for a review). Important
efforts have been devoted during the past
two decades to understand the dynamics of
plant-plant interactions along abiotic stress
or herbivory gradients, highlighting how
the presence of multiple abiotic stressors or
different levels of consumer pressure
importantly affect the outcome of plant-
plant interactions (e.g. Baumeister and
Callaway 2006, Graff et al. 2007).
However, and despite their common co-
occurrence in nature, the joint effects of
both herbivory and water stress on the
outcome of these interactions remain
largely ignored (Ibañez and Schupp 2001,
Veblen 2008, Smit et al. 2009). Considering
both abiotic stress and herbivory together is
crucial to understand the role of plant-plant
interactions in dryland ecosystems, where
these stressors are major factors influencing
plant community dynamics (Fischer and
Turner 1976, Whitford 2002).
The Stress-Gradient Hypothesis
(SGH; Bertness and Callaway 1994), a
framework in which most studies focused
on plant-plant interactions rely on, predicts
a continuous increase in the frequency of
positive interactions with increases in either
consumer pressure or abiotic stress.
However, several studies suggest that
positive plant-plant interactions may
collapse under extremely high levels of
both consumer pressure and abiotic
stressors directly related to resources, such
as water or light (e.g. Graff et al. 2007,
Maestre et al. 2009a). These studies caused
the generality of SGH predictions to be
challenged (Maestre et al. 2005, 2006,
Lortie and Callaway 2006). As a result of
this debate, predictions from the SGH have
been refined to consider the effect of
different abiotic stressors and the ecological
strategy of the species involved, and to
introduce consumer pressure as a major
factor affecting plant-plant interactions
along abiotic stress gradients (Maestre et al.
2009a, Smit et al. 2009). Furthermore, in
arid and semiarid areas water availability is
characterized by a strong inter- and intra-
annual variability, with marked temporal
dynamics that profoundly affect ecosystem
functioning (Whitford 2002). These
temporal dynamics add complexity to the
response of plant-plant interactions to
abiotic stress (Goldberg and Novoplansky
1997, Pugnaire and Luque 2001, de la Cruz
et al. 2008), and may also modulate the
I

CHAPTER 3
93
effect of herbivores on vegetation (Veblen
2008). Thus, they must be specifically
considered when studying the relationship
between plant-plant interactions and
different environmental biotic and abiotic
stressors in arid ecosystems.
Herbivory is also a major driver of
plant-plant interactions in these regions
(Baraza et al. 2006, Graff et al. 2007,
Veblen 2008). Unpalatable nurse plants
may protect their understorey vegetation
from herbivory damage by hiding more
palatable plants under their canopies, or by
sharing their defenses against herbivores
(Baraza et al. 2006, Veblen 2008), a process
commonly refered to as “associational
resistance” (sensu Hay 1986). This
protection against herbivory can strongly
affect vegetation dynamics and biodiversity
in grazed systems (Veblen 2008).
Alternatively, the improvement on water
status that nurse plants usually provide to
their target plants may also positively affect
target plant recovery from herbivory
damage (Rand 2004, Acuña-Rodriguez et
al. 2006). However, even in dry
environments, shade casted by nurse plants
can lead to light limitation for the protégée
plants (Seifan et al. 2010a, Soliveres et al.
2010). The joint effect that both shade and
an improved water status provided by nurse
plants have on the recovery from herbivory
of the protégée plants will depend on the
relative importance of water and light as
limiting factors for plant performance and
how herbivory affects their uptake (Wise
and Abrahamson 2005, 2007). Hence, the
final outcome on the protégée plants
response to herbivory is difficult to
generalize and predict, so more studies
along these lines are needed to refine
predictions on how plant communities
respond to different levels of abiotic and
biotic stressors (Graff et al. 2007, Smit et
al. 2009).
We conducted two simultaneous field
experiments to test the effects of rabbit
herbivory and water availability on the
interaction between the tussock grass Stipa
tenacissima L. (the nurse) and seedlings of
the leguminous resprouter shrub Retama
sphaerocarpa (L.) Boiss. (the protégée) in a
semiarid Mediterranean steppe. Stipa has an
overall low palatability (Ben Salem et al.
1994), and therefore could provide
herbivory protection for shrub seedlings by
associational resistance. Furthermore, the
positive effect that microclimatic
amelioration provided by Stipa has on the
survival of Mediterranean shrub seedlings
is well known (e.g. Maestre et al. 2001,
2003). Since water stress and the impact of
herbivores can prevent Retama
establishment in these environments
(Espigares et al. 2004, Rueda et al. 2008), it
is likely that the protection from herbivores

BIOTIC AND ABIOTIC STRESSORS AFFECT PLANT-PLANT INTERACTIONS
94
and the microclimatic amelioration
provided by Stipa canopies can play a key
role improving Retama colonization in
semiarid grasslands. We tested the
following hypotheses: i) Stipa provides
protection from rabbit herbivory to Retama,
enhancing its survival when rabbits are
present; ii) Shade provided by the Stipa
canopy enhances Retama water status
increasing its survival, but this positive
effect wane during extremely dry years due
to the overwhelming effect of competition
for water, iii) Regardless of plant water
status, light reduction produced by Stipa
will decrease Retama seedling recovery
from rabbit herbivory; and iv) Facilitation
of Retama seedlings by Stipa will collapse
under extreme levels of stress produced by
the joint action of herbivore damage and
drought stress.
METHODS
STUDY AREA
We conducted the study in the Aranjuez
Experimental Station, located in the center
of the Iberian Peninsula (40º03´60´´N,
3º54´91´´W). The climate is semiarid
Mediterranean, characterized by cold
winters and a strong summer drought, with
average annual precipitation and
temperature of 388 mm and 15 ºC,
respectively (1994-2005; Marqués et al.
2008, see also Appendix F in
Supplementary Material). The soil is
classified as Xeric Haplogypsid (Marqués
et al. 2008). Vegetation is an open steppe
dominated by Stipa (this species accounts
up to 90% of the total perennial cover),
with a perennial plant cover of 24%. Sparse
adult individuals of Retama and small
shrubs such as Lepidium subulatum L. and
Helianthemum squamatum (L.) Dum. Cours
are also present. The study site has a high
diversity of annual plants, which reach their
production peak in spring and constitute an
important part of plant productivity during
this period (Peco et al. 2009).
The study area harbours a high
density of rabbits (Oryctolagus Cunniculus
L.), as suggested by the high number of
visual contacts and the number of warrens
and latrines found (S. Soliveres, pers. obs.).
Domestic livestock or other large
herbivores are absent, and thus rabbits are
the only herbivores affecting vegetation
there. Rabbit activity tracks seasonal
variation in vegetation productivity. These
animals feed near their burrows to avoid
predation during spring and winter, when
their prefered food –mainly annuals– is
abundant; however, during summer drought
−when annuals dry out and food is scarcer−,
rabbits increase their exploration to obtain
enough food to survive (Rueda et al. 2008).
The selection of woody seedlings as a food
resource by rabbits increases during
summer (Maestre et al. 2001). Rabbit

CHAPTER 3
95
browsing, in the case of Retama seedlings,
may suppose the virtual removal of all the
aboveground biomass.
EXPERIMENTAL DESIGN
In December 2006, we planted 371 two-
year old Retama seedlings, with a mean
height of 27 ± 2 cm (mean ± SE, n = 20), by
using manually-dug holes of 20×20×20 cm.
The planted seedlings came from central
Spain (viveros Bárbol, Madrid, Spain), and
were maintained in the original nursery
until the week prior to planting. In the
nursery, seedlings were grown under
optimal conditions (full sunlight, fertilized
1:1 peat:coco-peat substrate, watering to
field capacity each week). We selected
these two-year old seedlings to ensure they
had enough size and reserves to resist rabbit
damage at least once (see Experiment 2
below). The selection of these two-year old
seedlings does not underestimate potential
facilitative effects of Stipa because Retama
seedlings were grown under optimal
conditions prior to their plantation, and thus
they were still sensitive to water stress and
to the environmental amelioration provided
by Stipa. We randomly planted these
seedlings on two different microsites:
“Tussock” and “Open”. The former
microsite was located < 15 cm from the
upslope face of an Stipa tussock (ca. 1 m
width and 80 cm height). Open microsites
were located in bare ground areas, > 50 cm
away from any perennial plant. One month
after planting, rabbits browsed some of the
seedlings, and we set up two parallel
experiments then. In the first experiment
(hereafter Experiment 1) we did not give
the seedlings any protection from further
grazing. This allowed us to test the
protection against herbivores provided by
Stipa canopy. In the second experiment
(hereafter Experiment 2) we evaluated the
joint effects of contrasted levels of water
availability and Stipa environmental
amelioration on the recovery of Retama
seedlings after rabbit browsing.
-Experiment 1: Nurse plant protection
against herbivores
From the 371 Retama seedlings planted, we
left a total of 195 seedlings without any
protection from herbivores (the remaining
plants after Experiment 2 was set up, see
below). From these seedlings, 103 and 92
were located in Tussock (refuge) and Open
(control) microsites, respectively. We
scored which of these plants had been
browsed by rabbits and which of them were
able to resprout and survive after the virtual
removal of their aboveground biomass (i.e.
the effect of rabbit damage) during January,
March and September 2007. Because of the
high number of rabbits present in the study
area and their repeated browsing, no plants

BIOTIC AND ABIOTIC STRESSORS AFFECT PLANT-PLANT INTERACTIONS
96
remained alive after September 2007, so we
stopped monitoring then.
-Experiment 2: Joint effects of herbivory
and abiotic stress on the outcome of the
interaction
This experiment had three factors: i)
Microsite (Tussock vs. Open), ii) Herbivory
(Retama seedlings whose aboveground
biomass was completely eaten by rabbits
during the first month in the field vs.
unbrowsed seedlings), and iii) Irrigation
(watered vs. non watered seedlings). Since
inter-annual variability is of crucial
importance in dry environments, and it may
strongly affect the results found and our
conclusions (Ibañez and Schupp 2001), we
conducted this experiment during a three
year-period (and therefore under three
contrasted environmental conditions) to add
confidence to the results found. We set up
22 replicates per each combination of
treatments (176 seedlings in total). All the
seedlings from this experiment were
protected from further herbivory after the
first month in the field using a metallic
mesh. The diameter of the openings in the
mesh was 5 cm, which casted no detectable
shade to the seedlings, and did not
confound the effects of any of the factors in
the experiment. In this experiment we
monitored the variables described below
(see Field monitoring section below).
We conducted a spatial analysis of
browsed seedlings using the Spatial
Analysis by Distance Indices (SADIE)
methodology (see Perry 1998 for details).
The spatial pattern of herbivory damage by
rabbits was random (SADIE’s Aggregation
Index [Ia] = 0.95; P = 0.56; n = 176). Thus,
we do not expect unmeasured variables
with spatial structure (e.g. soil depth,
distance to a rabbit burrow or slope
position) to influence seedling response to
the assayed treatments. We did not measure
seedling attributes that could influence
rabbit behaviour (e.g. plant C/N ratio, initial
plant height). However, the large number of
seedlings randomly assigned to each
treatment, and the fact that rabbit damage
was equally intense (i.e. complete removal
of aboveground biomass) regardless of the
microsite considered, should control for the
experimental noise that any unmeasured
factors potentially affecting rabbit behavior
could have on the results of this
experiment.
The irrigation treatment consisted in
eight supplementary pulses of water, once
every month, between April and July in
both 2007 and 2008. The wettest and driest
periods of the study area are spring (from
March to May) and summer (from June to
September), respectively. Thus, the
irrigation treatment affected both wet and
dry periods. In each monthly watering, we
applied an amount of water equivalent to

CHAPTER 3
97
the 50% of the median rainfall registered
during the past 30 years for this month,
irrespectively of the rainfall registered (i.e.
6, 9.3, 10.9 and 5.7 mm for April, May,
June and July, respectively; see Appendix 1
in Supplementary material). With this
irrigation scheme we aimed to evaluate the
joint effects of reducing the summer
drought (summer drought was longer in
unwatered plants because of the lack of
rainfall in June and July) and increasing
precipitation during the wet season (spring
rainfall was less abundant in unwatered
plants) on the outcome of the interaction
studied.
Field monitoring of Experiment 2
-Plant performance measurements
We monitored seedling height and survival
after each summer, which is the most
critical season for seedling survival in
Mediterranean semiarid regions (e.g.,
Maestre et al. 2001, 2003). We measured
these variables in September 2007, 2008
and 2009. Height was well correlated with
aboveground biomass, as demonstrated by
an allometric relationship performed with
seedlings of contrasted sizes and ages
(Spearman correlation: ρ = 0.65, P <
0.0001; n = 45), and thus was used as our
surrogate for seedling biomass in the field.
We measured photochemical
efficiency by using the in situ chlorophyll
fluorescence parameter Fv/Fm; it is
calculated from photosystem II (PSII)
fluorescence signals as the ratio between
the variable (Fv) and the maximum (Fm)
fluorescence signal, which are obtained
from a short light pulse after 20 minutes of
dark adaptation. Fv is the difference
between Fm and the minimal fluorescence
signal right before the saturating light pulse,
being Fm the light that plant cannot absorb
when its absortion capability has been
collapsed by a previous pulse. Fv/Fm was
determined with a pulse-modulated
fluorometer (FMS2, Hansatech Instruments,
Norfolk, UK). This variable is an estimator
of the overall plant stress (Maxwell and
Johnson 2000), and has been widely used as
an indicator of plant stress in numerous
studies in semiarid areas (e.g. Pugnaire et
al. 1996b, Maestre et al. 2001, 2003,
Aragón et al. 2008). Furthermore, small
changes in the concentration of chlorophyll
in leaf tissues associated with water
limitation, which can be tracked by
measuring Fv/Fm, can be crucial during
important stages of plant life (Aragón et al.
2008). Although the use of Fv/Fm can be
problematic for the detection of water stress
in some species (Resco et al. 2008), it is a
good proxy for plant stress in our case
because the canopy structure, lack of leaves
and the high tolerance of Retama to solar
radiation minimizes photoinactivation and

BIOTIC AND ABIOTIC STRESSORS AFFECT PLANT-PLANT INTERACTIONS
98
downregulation in this species (Valladares
and Pugnaire 1999), a confounding factor
that could mask the relationship between
Fv/Fm and water stress. We randomly
selected eight seedlings per treatment (n =
64) for Fv/Fm measurements. We measured
Fv/Fm in four key moments for seedling
performance in Mediterranean semiarid
environments: during the wettest period of
spring (April-May), in the middle and at the
end of the summer drought (July and
September, respectively) and during the
coldest month (December-January). Fv/Fm
was measured in the same plants from May
2007 to September 2009.
-Soil moisture measurements
We measured soil moisture by time-domain
reflectometry (TDR; Topp and Davis 1985)
using a Campbell TDR100 system
(Campbell Scientific Ltd, Loughborough,
UK). In eight randomly selected planting
holes per treatment, we vertically installed
10 cm long probes (n = 64). We chose this
soil depth because the vast majority of root
biomass of the planted seedlings
concentrates near the soil surface (the pots
where Retama seedlings were grown had
ca. 15 cm. depth), and because Stipa
tussocks also concentrate the majority of
their roots in the upper layers of the soil
(Puigdefábregas et al. 1999). We conducted
a site-specific calibration between Time
Domain Reflectometry measurements and
gravimetric soil moisture (R2 = 0.84; P <
0.0001; n = 68) to assess for the validity of
these measurements in gypsum soils and to
convert them in soil gravimetric moisture
data. We measured soil moisture in the
same plants in all the samplings, and in the
same sampling periods as Fv/Fm surveys
(see above). When these measurements
coincided with irrigation pulses, soil
moisture was measured at least one week
after the irrigation. With this approach we
avoided giving too much importance to the
occasional influence of irrigation in our
measurements, and assessed more
realistically the soil moisture available for
Retama seedlings during a given period. In
addition to these measurements, and to
further evaluate the effects of irrigation on
soil water availability, we measured soil
moisture one day after the irrigation pulse
was applied on June 2007 in the planting
holes of 10 undamaged plants (the eight
replicates selected for the previously
described soil moisture measurements plus
two extra replicates) of each of the four
combinations between microsite and
irrigation (n = 40). We measured soil
moisture only in unbrowsed plants because
we were only interested in assessing for
differences in soil moisture between the
irrigation treatments applied and the
microsites tested.

CHAPTER 3
99
STATISTICAL ANALYSES
As all Retama seedlings used in Experiment
1 died at the end of the monitoring, we did
not perform any statistical comparison with
the survival data at this period. However,
we used the log-rank statistic of the Kaplan-
Meyer procedure to compare the shape of
the survivorship and number of browsed
plants curves between Tussock and Open
microsites. We analyzed the survival of
Retama seedlings from Experiment 2
separately for each year by using a
hierarchical log-linear analysis, with
microsite, irrigation and herbivory as fixed
factors. To assess for the effects of the
factors assayed during the different
environmental conditions characterizing
each year, only those seedlings that
survived the previous summer were taken
into account for this analysis (for example,
to analyze survival of 2008, we only
considered those seedlings alive after the
summer of 2007). With this approach, we
were able to assess the consistency of
treatment effects over the years. It also
avoids the potential “dragging” that an
extremely strong effect of a particular
treatment during a given year may have on
the overall net results (i.e. we could detect
if herbivory had strong effects in 2007, but
not in the rest of years). Survival data from
one year to another were not correlated
(Spearman correlation: ρ < 0.4; P > 0.2 in
all the cases) and thus independency is
expected. Since browsing by rabbits
removed most aerial biomass of planted
seedlings, and initial seedling height after
rabbit impact was not measured, we did not
consider such height as a covariate in
further statistical analysis.
Months after planting0 2 4 6 8 10
unbr
owse
d se
edlin
gs (
%)
0
20
40
60
80
100
OPENTUSSOCK
Months after planting0 2 4 6 8 10
seed
ling
surv
ival
(%
)
0
20
40
60
80
100
OPENTUSSOCK
Figure 3.1. Dynamics of herbivore damage (A) and overall survivorship (B) of unprotected Retama sphaerocarpa seedlings growing under the canopy of Stipa tenacissima (Tussock) and in bare ground areas (Open) from December 2006 to September 2007.
We analyzed gravimetric soil
moisture (obtained from TDR data), Fv/Fm
A
B

BIOTIC AND ABIOTIC STRESSORS AFFECT PLANT-PLANT INTERACTIONS
100
and seedling height data by using repeated
measures ANOVA, with microsite,
irrigation and herbivory as fixed factors.
Data were square-root transformed to reach
normality and homocedasticity assumptions
when necessary. We found significant
interactions between time and the
treatments evaluated when analyzing soil
moisture and Fv/Fm data (data not shown).
These interactions can lead to the
misinterpretation of the effects of the fixed
factors (Quinn and Keough 2002), and are
of biological importance, as intra-annual
dynamics in water availability can strongly
affect the effects of nurse plants (Goldberg
and Novoplansky 1997, De la Cruz et al.
2008). To properly assess the effect of the
assayed treatments, and to explore how the
intra-annual dynamics in water availability
modulates them, we grouped both soil
moisture and Fv/Fm data for the three study
years in wet/dry periods (periods with soil
moisture values above and below 10%,
respectively) and analyzed them separately
using repeated measures ANOVA. We
established this 10% value to separate
wet/dry periods because it corresponds to a
biological threshold that separates periods
when most plant activity concentrates
(those with soil moisture > 10%) in
semiarid environments (Noy-Meir 1973,
Valladares et al. 2005). These analyses
could lack independency because results
obtained in dry periods were not completely
independent from those coming from wet
periods. However, by pooling the data from
the three study years together (we
conducted two separate RM ANOVA, one
with dry and another for wet periods data
from the three study-years) only consistent
results for the three years may result
significant and this lack of independency
disappears. Furthermore, with this approach
we removed the interactions with the
assayed treatments and time, avoiding the
confounding effect that the strong temporal
variability in water availability could have
on the interpretation of the main treatment
effects (Quinn and Keough 2002).
We evaluated differences in soil
moisture between watered vs. unwatered
plants (soil moisture measures after the
irrigation pulse of June 2007) using a two
factor (microsite and irrigation) ANOVA.
These data followed the assumptions of this
analysis, and thus were not transformed.
We conducted all statistical analyses using
SPSS 13.0 for Windows (Chicago, Illinois,
USA).
RESULTS
EXPERIMENT 1: NURSE PLANT
PROTECTION AGAINST HERBIVORES
-Survival and herbivore damage of
unprotected plants
Fewer plants were browsed by rabbits when
growing under Stipa canopies than in Open

CHAPTER 3
101
microsites during spring (Fig. 3.1A; Log-
rank test, P < 0.0001). Rabbit predation
intensity on Retama seedlings increased
during summer drought. This, together with
the removal of most aboveground biomass
produced after each browsing impact,
caused the depletion of the resprouting
ability of Retama seedlings, and all the
plants died during summer regardless of the
microsite where they were planted (Fig.
3.1B). Thus, we did not find differences in
survival among microsites during the study
period (Log-rank test, P = 0.466).
EXPERIMENT 2: JOINT EFFECTS OF
HERBIVORY AND ABIOTIC STRESS
-Plant performance
Stipa tenacissima reduced significantly the
survival of Retama seedlings during the
three years of study (2007: G2 = 12.7, df =
1, P = 0.005; 2008: G2 = 9.3, df = 1, P =
0.002 and 2009: G2 = 3.6, df = 1, P < 0.06,
Fig. 3.3. 2). Survival was 17%, 27% and
15% lower in Tussock than in Open
microsites for 2007, 2008 and 2009,
respectively. Irrigation increased the
survival of Retama seedlings during the
first summer by 27% (G2 = 10.29, df = 1, P
= 0.001), but did not affect the negative
effect of Stipa. Browsing damage did not
affect mortality rates per se in this
experiment, but reduced the negative effect
of Stipa on Retama (Microsite × Herbivory:
0
20
40
60
80
100
2009
Sur
viva
l (%
)
0
20
40
60
80
100
OPENTUSSOCK
2008
0
20
40
60
80
100
2007
H- H+
I-
H- H+
I+
I ; P < 0.0005M ; P < 0.0005M x H ; P = 0.005
M ; P < 0.005
M ; P = 0.06I x H ; P < 0.005
Figure 3.2. Survival of Retama sphaerocarpa seedlings during the three years of study in the eight combinations of treatments evaluated. Open = bare ground areas, Tussock = Stipa tenacissima canopies, I- = no irrigation, I+ = irrigation of 50% of the median of April-July period rainfall in four pulses, H- = no herbivore damage, and H+ = seedlings partially eaten by rabbits. Initial n = 22. G
2 = 7.7, df = 1, P = 0.005, Fig. 3.2). We
found a significant interaction between
prior herbivory damage and water
availability in 2009 (G2 = 8.85, df = 1, P =
0.003), being the survival of unbrowsed
seedlings higher than that of browsed

BIOTIC AND ABIOTIC STRESSORS AFFECT PLANT-PLANT INTERACTIONS
102
seedlings when these were unwatered.
When watered, seedlings that were browsed
showed a higher survival rate than those
that were not browsed (Fig. 3.2).
Herbivory decreased seedling height
during the three-years of study (RM
ANOVA: F1,171 = 4.6; P = 0.036), an effect
that was especially evident in 2008 (Fig.
3.3). We did not detect any microsite or
watering effect, neither any interaction
between the treatments evaluated or with
time, when analyzing seedling height. Stipa
tenacissima reduced the Fv/Fm ratio of
Retama seedlings during dry seasons (RM
ANOVA: F1,56 = 9.6; P = 0.003), but this
effect decreased when plants were watered
(Microsite x Irrigation: F1,56 = 3.9; P =
0.05). This Microsite x Irrigation
interaction was also found in wet periods,
when irrigation reduced the positive effects
of Stipa on Retama Fv/Fm (Fig. 3.4;
Appendix G.A).
-Soil moisture
2007 was the wettest year of the studied
period, with soil moisture levels well above
20% during spring (Appendix G.2B).
Conversely, 2009 was the driest year, with
soil moisture levels below 10% in three of
the four periods sampled (Appendix G.2B).
Stipa tenacissima slightly (< 2%) reduced
soil water availability during dry periods
(RM ANOVA: F 1,56 = 5.02; P = 0.029).
During wet periods, a significant microsite
× herbivory interaction was found (F 1,56 =
5.35; P = 0.024), with more water available
under Stipa canopies and Open microsites
for browsed and unbrowsed plants,
respectively (Fig. 3.5). Although irrigation
increased soil moisture values by an
average of 35% after watering (Two-way
ANOVA, F1,36 = 8.05; P < 0.001), it did not
affect soil moisture at the long-term, as this
treatment had no significant effects on this
variable when analyzing the data gathered
during the whole year (Repeated-Measures
ANOVA; P = 0.929).
DISCUSSION
The results of our study highlight the
importance of herbivory as a major factor
affecting the relationship between plant-
plant interactions and abiotic stress. The
increase in rabbit pressure during summer
drought, indirectly caused by the lack of
alternative food resources during this
season, overrided the herbivory protection
provided by Stipa during wetter periods,
when rabbit pressure upon Retama
seedlings was lower. Conversely, the initial
loss of biomass produced by rabbit
browsing shifted the interaction between
Stipa and Retama from negative to neutral.

CHAPTER 3
103
2009
10
20
30
40
2008
10
20
30
40
2007
Mea
n se
edlin
g he
ight
(cm
)
10
20
30
40
OPENTUSSOCK
H- H+
I-
H- H+
I+
RM ANOVA: H ; P < 0.005
Figure 3.3. Height of Retama sphaerocarpa seedlings during the three years of study in the eight combinations of treatments evaluated (mean ± SE; n depended on survival). Rest of Legend as in Figure 3.2.
Our results suggest that the negative effect
of Stipa on the performance (Fv/Fm) of
Retama was driven by water availability,
but that competition for other resources
rather than water modulated the effect of
Stipa on Retama survival. The complex
interactions between herbivory, abiotic
stress and their temporal dynamics as
drivers of the outcome of plant-plant
interactions highlight the importance of
considering these stressors together to fully
understand the outcome of plant-plant
interactions along environmental gradients
(Goldberg and Novoplansky 1997, de la
Cruz et al. 2008, Anthelme and Michalet
2009).
Pho
toch
emic
al e
ffici
ency
(F
v/F
m)
0.0
0.6
0.8
OPENTUSSOCK
DRY SEASON
0.0
0.6
0.8
WET SEASON
H- H+
I-
H- H+
I+
M ; P < 0.005M x I ; P = 0.005
M x I ; P = 0.05
Figure 3.4. Photochemical efficiency (Fv/Fm) of Retama sphaerocarpa seedlings during wet/dry periods (periods above/below 10% gravimetric soil moisture, respectively). Data are means ± SE of the three years-study period data pooled by wet/dry seasons; n = 8). Rest of Legend as in Figure 3.2.

BIOTIC AND ABIOTIC STRESSORS AFFECT PLANT-PLANT INTERACTIONS
104
Gra
vim
etric
soi
l moi
stur
e (%
)
0
2
4
6
8
10
OPENTUSSOCK
DRY SEASON
WET SEASON
H- H+
I-
H- H+
I+
0
5
10
15
20
25
30
M ; P < 0.05
M x H ; P < 0.05
Figure 3.5. Gravimetric soil moisture (inferred from time-domain reflectometry measurements) during wet/dry periods. Rest of Legend as in Figure 3.4.
PROTECTION AGAINST HERBIVORES
BY STIPA TENACISSIMA
Although Retama seedlings were protected
from herbivore damage when Stipa was
present, this effect dissapeared during
summer drought, when the higher rabbit
impact upon Retama seedlings
overshadowed the refuge effect of Stipa
(Fig. 3.1). Annual plants, which provide an
important fraction of plant productivity in
arid and semiarid systems (Fischer and
Turner 1978), have completed their life
cycle before the onset of summer drought in
our study area (Peco et al. 2009). Thus, the
corresponding increase in rabbit predation
upon perennials due to changes in diet
produced by the lack of annuals during
summer (Rueda et al. 2008) can explain the
suppression of this facilitative effect during
this season. Similar reductions of
facilitative effects under high herbivory
pressure have been previously reported
(Graff et al. 2007, Smit et al. 2007), and
should be common when food resources are
less abundant and the same number of
herbivores may exert higher pressure on the
remaining plants (but see Veblen 2008,
Anthelme and Michalet 2009).
DOES SHADE INTOLERANCE OF THE
PROTÉGÉE EXPLAIN THE NEGATIVE
EFFECT OF THE NURSE PLANT?
In contrast with previous studies using the
same nurse plant (e.g. Maestre et al. 2001,
2003), we found a net negative effect of
Stipa on survival of Retama seedlings.
Plant-plant interactions depend up to a great
degree on the identity of the species
involved (Callaway 2007), and thus these
contrasting results are not fully surprising.
Plant competition in drylands is generally
attributed to water or nutrients (Whitford
2002). Interestingly, most of the negative
effects of Stipa on Retama were not
influenced by increases in water
availability, neither were explained by the
effect of Stipa on this variable. The reduced
light availability under the canopy of Stipa

CHAPTER 3
105
tussocks (which suppose >80% of incident
PAR reduction; see Maestre et al. 2003),
has been identified as the major driver of
the positive effect of Stipa on shrub
seedlings (Maestre et al. 2003). However,
this same shade could be also a key factor
affecting the negative outcome of the
studied interaction (Seifan et al. 2010a,
Soliveres et al. 2010). The lack of leaves of
Retama and the concentration of the total
photosynthetic area on its cladodes, is
associated with the high light requirements
of this species (Valladares and Pugnaire
1999, Valladares et al. 2003, Espigares et
al. 2004), and suggests that a reduction in
available light might underlie the outcome
observed.
Most plant species adapted to
drought are not able to cope with deep
shade (Niinemets and Valladares 2006), and
therefore, it is likely that species that are
more adapted to full sunlight and drought,
which are abundant in dry environments, do
not benefit from the presence of a nurse
plant unless the positive effects of the nurse
on the water status of the protégée
overcome negative effects promoted by
light reduction (Holmgren et al. 1997). The
more drought-tolerant the protégée plant is,
the less positive the shade effect is expected
to be, according to the general ideas
discussed in recent revisions of the SGH
(Maestre et al. 2009a, Malkinson and
Tielbörger 2010). The same may happen
with different life stages of a given species,
as plants are often more shade tolerant early
during their ontogeny than in later stages of
development, and therefore are more likely
to benefit from nurse’s shade (Callaway and
Walker 1997, Miriti 2006). Our results
highlight the species-specific nature of such
effects (Callaway 2007), since the same
shade that is beneficial for some species
(Maestre et al. 2001, 2003) could
conceivably be negative for Retama or
other shade-intolerant plants (Marañón and
Bartolomé 1993, Seifan et al. 2010a). Thus,
more studies involving species with
different ecological strategies and drought
and shade relative tolerances are needed to
improve our understanding on the responses
of plant-plant interactions to abiotic stress
and herbivory at the entire community
level.
THE ROLE OF FACILITATION AND
RESOURCE AVAILABILITY ON THE
RECOVERY OF BROWSED PLANTS
Irrigation increased survival during part of
the studied period (Fig. 3.2) and also the
degree of stress experienced by Retama
seedlings was lower during wet periods.
Both results indicate that water was limiting
the performance of this species, regardless
of the microsite tested. However, light was
also an alternative limiting factor for plant

BIOTIC AND ABIOTIC STRESSORS AFFECT PLANT-PLANT INTERACTIONS
106
performance in Tussock but not in Open
microsites (discussed above). The Limited
Resource Model (LRM), which correctly
explains most of the relationships between
different resource levels and tolerance to
herbivory, predicts a differential recovery
from tolerance depending on the nature of
the limiting resources and the way
herbivory damage affect their uptake (Wise
and Abrahamson 2005, 2007). Particularly,
this model foresees a higher tolerance to
herbivory under wetter conditions when
water is the only limiting factor (such as in
Open microsites), but a lower tolerance
under these same wet conditions when
herbivory exacerbates the limitation of an
alternative resource, light in our case (such
as in Tussock microsites). This latter
outcome is explained because plants
growing under drought are already at their
optimum light conditions, and are mainly
limited by water availability, while plants
growing under Stipa canopies are closer to
their optimum moisture conditions, but also
limited by light. Thus, the latter plants will
be much more sensitive to the reduced
uptake of light (their alternative limiting
resource), and therefore to the loss of
biomass produced by herbivory, than the
former (Baraza et al. 2004, Wise and
Abrahamson 2005, 2007).
Following these predictions, we may
expect lower tolerance to herbivory in
Tussock than in Open microsites, especially
in watered plants, since plants growing
beneath the canopy of Stipa should be more
limited by light, while plants growing in
Open microsites should be limited only by
water. However, our results did not fully
match predictions from the LRM, maybe
because our target plant was a resprouter,
and therefore its tolerance to herbivory
might be affected not only by the amount of
resources available, but also by the reserves
of each seedling might have (Vesk et al.
2004). In contrast, a higher survival in
browsed plants was found when they were
watered, regardless of the microsite where
they were planted. This finding could
indicate an overcompensation of browsing
damage by plants when environmental
conditions were more benign (Crawley et
al. 1998). Why the LRM predictions did not
correctly explain the effects of shade
provided by Stipa on Retama recovery after
herbivory damage? Shade provided by
Stipa might increase the water status of
Retama seedlings by a reduction in
transpiration (Holmgren et al. 1997). Thus,
it is likely that this improvement in the
water status of Retama seedlings increased
their tolerance to herbivory in a similar way
that watering did it in Open microsites. This
positive effect in the recovery from
herbivory shifted the negative effect that
this same shade exerted on Retama survival
on unbrowsed plants (we found a
significant Microsite × Herbivory

CHAPTER 3
107
interaction when analyzing seedling
survival). The compensation of resource
competition due to herbivory protection has
been observed in previous studies in
semiarid environments (Graff et al. 2007).
Our results suggest that this compensation
also occurs when the nurse plant improves
recovery from herbivory, and highlight the
complex interaction between water stress
and herbivory as drivers of the outcome of
an interaction strongly influenced by shade.
TEMPORAL DYNAMICS IN WATER
AVAILABILITY MODULATE THE
EFFECT OF STIPA ON RETAMA
In contrast with our initial hypothesis, Stipa
negatively affected Retama survival during
the three years of study, regardless of the
inter-annual variation in rainfall availability
among years. This may be explained
because the studied interaction was mainly
driven by light competition, and therefore
water availability only played a secondary
role in its outcome. However, intra-annual
dynamics in water availability and our
irrigation treatment modulated the effect of
Stipa on the stress level experienced by
Retama seedlings. Goldberg and
Novoplansky (1997) proposed a conceptual
model to introduce the effect of intra-annual
resource dynamics on plant-plant
interactions. In their model, nurse plants
affected negatively protégée growth during
pulses (our wet seasons) due to competition
by resources, while increased survival
during periods with low nutrient availability
(our dry seasons). The final outcome would
depend on how much the negative effect on
growth during wet seasons affects survival
during dry periods, and on the relative
importance of plant uptake or abiotic
factors affecting resource availability
during these dry periods (Goldberg and
Novoplansky 1997, Hastwell and Facelli
2003). However, the effects of Stipa on the
Fv/Fm (our surrogate of plant stress) of
Retama seedlings during wet/dry seasons
found differed from the expected responses
arisen from the predictions of Goldberg and
Novoplansky (1997). Specifically, we
detected a trend towards facilitation and
competition during wet and dry seasons,
respectively (although it must be considered
that we only measured the degree of stress
experienced by Retama and not its growth
or survival seasonally, which could be a
better test for this model). Differences
produced in the outcome of the interaction
studied within these seasonal dynamics
varied with irrigation, which overall
suggest a reduction of competition intensity
at intermediate levels of abiotic stress
(watered plants during summer or
unwatered plant during spring), but a
prevalence of competition in the rest of
assayed situations, as suggested by the

BIOTIC AND ABIOTIC STRESSORS AFFECT PLANT-PLANT INTERACTIONS
108
refined SGH when the two species involved
are stress tolerators (Maestre et al. 2009a).
CONCLUDING REMARKS
Collectively, our experiments indicate the
existence of a complex hierarchy of
mechanisms affecting the interaction
studied (Baumeister and Callaway 2006). In
our case, water shortage per se was
irrelevant under extremely high herbivory
impacts (all seedlings died in Experiment 1
but none of them due to drought), but
indirectly modulated herbivory impacts by
affecting alternative rabbit food resources.
Conversely, the initial loss of biomass
produced by herbivory was a major factor
affecting the outcome of the interaction
studied. Stipa exerted a negative effect on
Retama seedlings mainly by light
competition, but this shade improved
seedlings recovery after herbivory,
overriding the negative effects found on
unbrowsed plants. Our findings illustrate
how the complex interactions between
herbivory and water stress jointly influence
the outcome of plant-plant interactions.
They provide insights to fully understand
the interplay between facilitation and
competition, and they can be used to further
refine conceptual models aiming to predict
the outcome of plant-plant interactions
along composite stress gradients.
ACKNOWLEDGEMENTS We wish to thank E. Chaneton, D. Eldridge, M. Seifan and three anonymous referees for their useful comments and corrections on a previous version of this manuscript. E. Pigem, C. Alcalá, S. Constán-Nava, J. Papadopoulos and E. Barahona helped during the fieldwork. We thank the Instituto Madrileño de Investigación y Desarrollo Rural, Agrario y Alimentario (IMIDRA) for allowing us to work in the Finca de Sotomayor (Aranjuez). SS and PGP hold PhD fellowships from the EXPERTAL grant, funded by Fundación Biodiversidad and CINTRA S.A. APC was supported by a PhD fellowship from the INTERCAMBIO (BIOCON06/105) grant, funded by Fundación BBVA. FTM acknowledges support from the European Research Council under the European Community's Seventh Framework Programme (FP7/2007-2013)/ERC Grant agreement n° 242658. This research was funded by the EXPERTAL grant, and with additional funds from INTERCAMBIO and REMEDINAL2 grants.

CHAPTER 3 .
109
Supplementary Material for Chapter 3
Appendix F. Climatic data (mean monthly temperature, black circles; and monthly rainfall,
grey bars) obtained from a meteorological station (Onset, Pocasset, MA, USA) located at the
study site. White bars represent the increment in monthly rainfall by the irrigation treatments
applied during 2007 and 2008.
Mc A
My Jn Jl Au S O N D Ja F
Mc A
My
Jn Jl
Au S O N D Ja
F M
c A
My
Jn
Jl
Au
S
Rai
nfal
l (m
m)
0
20
40
60
80
100
120
140monthly rainfallwatering
2007 2008 2009
Mea
n te
mpe
ratu
re (
ºC)
0
5
10
15
20
25
30

BIOTIC AND ABIOTIC STRESSORS AFFECT PLANT-PLANT INTERACTIONS
110
Appendix G. Photochemical efficiency of Retama sphaerocarpa seedlings (A) and
gravimetric soil moisture (A) in the different sampling dates for the eight possible
combinations of treatments. Open = bare ground areas, Stipa = Stipa tenacissima canopies, I-
= no irrigation, I+ = irrigation of 50% of the median of April-July period rainfall in four
pulses, H- =no herbivore damage, and H+ = seedlings partially eaten by rabbits. Data
represent means ± SE (n = 8).
Pho
toch
emic
al e
ffici
ency
(F
v/F
m)
2007 2008 2009
MayJuly
Sept.
January
May
July Sept.
January
April
July
Sept.
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Open I- H- Open I- H+ Open I+ H- Open I+ H+Stipa I- H-Stipa I- H+Stipa I+ H- Stipa I+ H+
MayJu
lySept.
January M
ay Ju
ly Sept.
JanuaryApril
July
Sept.
0
10
20
40Open I- H- Open I- H+ Open I+ H-Open I+ H+Stipa I- H-Stipa I- H+Stipa I+ H-Stipa I+ H+
Gra
vim
etric
soi
l moi
stur
e (%
)
2007 2008 2009
A
B

Santiago Soliveres, David J. Eldridge, Fernando T. Maestre, Matthew A. Bowker,
Matthew Tighe and Adrián Escudero.
Manuscrito en revisión en Ecological Monographs
4
On the relative importance of climate and biotic no n-trophic
interactions as drivers of local plant species rich ness in semiarid
communities

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
112

CHAPTER 4 .
113
ABSTRACT
Most studies including the role of positive plant-plant interactions as drivers of plant richness along environmental gradients assume an unimodal richness-productivity relationship, which is not as general as previously thought, and the existence of an overarching stress gradient which affects equally the different species forming a community. We aimed to evaluate the relative roles of facilitation/competition and environmental conditions as drivers of local species richness without these assumptions, and to clarify their contribution to the richness-productivity relationship. We conducted an observational experiment across wide environmental gradients in semiarid regions from Spain and Australia, assessing how the intensity, importance and frequency of positive interactions, and the degree of niche expansion provided by the nurse plants changed along these gradients. We also tested the particular mechanism (niche segregation, competitive exclusion or intransitivity) underlying the effects of nurses on their understorey vegetation. Nurse plants increased local richness by expanding the niche of the less adapted species both in Spain and Australia. The high variability of niches often found underneath their canopies may be the main reason why higher niche segregation and species coexistence was found under nurse than in open microsites. The outcome of the competition-facilitation continuum changed depending on the type of stress gradient considered. When it was driven by both rainfall and temperature (Spanish sites), the community-wide importance of nurse plants remained constant along the gradients. When the stress gradient was driven only by rainfall (Australian sites), the importance of nurses showed a unimodal relationship with the gradient, indicating a collapse of facilitation under both extremes of rainfall availability. Particular pairwise interactions outcomes were poorly predicted using abiotic measurements as an overarching stress level, and we propose to use each species distance to its environmental optimum as a better approach for this purpose. Our study provides a complete mechanistic understanding of the relative roles of plant-plant interactions and environmental conditions shaping local species richness in semiarid environments. These results can also be used to refine our predictions of the response of plant communities to environmental gradients, and clarify the relative importance of biotic interactions as a driver of such responses.

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
114
INTRODUCTION
he study of the mechanisms
controlling the composition of
communities has been a major
topic in ecology since the early days of this
science (see Gotelli and Graves 1996 for a
review). Recent research has highlighted
the fact that local-scale non-random
processes such as abiotic constraints and
biotic interactions determine the species
that are able to successfully colonize a
given environment (Huston 1999, Lortie et
al. 2004a, Rajaniemi et al. 2006).
Pioneering studies have suggested that,
among local-scale processes, competition
regulates richness at high levels of
productivity, while limited physiological
tolerances to abiotic stress or disturbance
reduce species recruitment, and thus
richness, at low levels of productivity
(Grime 1973, Huston 1979). An inherent
assumption of this observation is that
competition is less important at the lower
productivity end of the gradient. Overall,
the joint effects of abiotic constraints and
competition should generate a hump-shaped
relationship between species richness and
local productivity, which is reputed to be
ubiquituous in nature (Grime 1973, 2001,
Huston 1979). However, plant-plant
interactions are key drivers of community
structure in both high and low productivity
environments (Tilman 1988, Callaway
2007). Over the past decade, ecologists
have revisited the hump-shaped richness-
productivity relationship to explore the
potential effects of positive, non-trophic
interactions (hereafter ‘facilitation’; Hacker
and Gaines 1997, Michalet et al. 2006).
However, the relative effects of plant-plant
interactions and abiotic conditions on
changes in species richness along
environmental gradients, and therefore their
influence on the richness-productivity
relationship, remain uncertain (Rajaniemi et
al. 2006). Studies aimed at clarifying the
roles of these factors can help to explain
why several studies cast doubt on the
generality of the hump-shaped richness-
productivity relationship, particularly at
both local and regional scales (Waide et al
1999, Gillman and Wright 2006).
Many empirical studies (e.g. Hacker
and Bertness 1999, Kikvidze et al. 2005,
Valiente-Banuet et al. 2006, Cavieres and
Badano 2009) and theoretical models
(Bertness and Callaway 1994, Bruno et al.
2003, Lortie et al. 2004a) developed over
the past two decades have emphasized the
importance of facilitation for maintaining
community richness at low to moderate
levels of productivity (Hacker and Gaines
1997, Michalet et al. 2006). Environmental
buffering (both microclimatic amelioration
and protection from herbivory) by nurse
T

CHAPTER 4 .
115
species can increase the realized niche of
less adapted species, and therefore increase
local richness (niche expansion; Bruno et
al. 2003). Although the relationship
between the importance and frequency of
positive plant-plant interactions and
abiotic/biotic stress was originally thought
to be monotonic and positive (Bertness and
Callaway 1994), some studies have
suggested a waning of positive interactions
under either extreme abiotic stress levels
(e.g. Kitzberger et al. 2000, Ibañez and
Schupp 2001, Maestre and Cortina 2004a)
or intense disturbance (Ibañez and Schupp
2001, Smit et al. 2007, Forey et al. 2009),
thus obviating niche expansion (Michalet et
al. 2006). Explanations invoked to explain
this lack of niche expansion under such
conditions are: 1) the competitive effects of
nurse plants may outweight the positive
effects of environmental amelioration,
particularly when abiotic stress is promoted
by a resource such as water (Maestre and
Cortina 2004a, Maestre et al. 2009a), and 2)
nurse plants may not be able to attain a size
large enough to ameliorate harsh abiotic
conditions in extremely stressful
environments (Michalet et al. 2006). We
henceforth refer to both of these potential
mechanisms as facilitation waning models
(see Fig. 4.1).
Most studies describing facilitatory
mechanisms in relation to the diversity-
productivity curve have tended to focus on
unproductive to moderately-unproductive
environments (Hacker and Gaines 1997,
Michalet et al. 2006). Recent studies,
however, have revealed that ecological
processes such as niche segregation (Hector
et al. 1999, Silvertown 2004), competition
intransitivity (lack of hierarchy in
competition networks: Gilpin 1975, Laird
and Schwamp 2006, Bowker et al. 2010),
and indirect facilitation (Levine 1999,
Brooker et al. 2008) may be key
mechanisms enhancing species richness in
more productive conditions. More benign
conditions often found under nurse plants
(Franco and Nobel 1989) may increase the
local species pool and the heterogeneity of
available niches, also allowing finer
partitioning of variable resources. Although
largely ignored in facilitation research
(Brooker et al. 2008, but see Tielbörger and
Kadmon 2000b), these localized effects
may potentially increase niche segregation
(Pugnaire et al 1996,a Maestre and Cortina
2005) and competition intransitivity
beneath nurse plant canopies, enhancing
overall local richness and productivity.
Despite the interest generated in the joint
effects of nurse plants on both niche
expansion and changes in competition

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
116
outcomes, these mechanisms have largely
been explored separately in the literature.
There is a clear need, therefore, to develop
a unifying framework that integrates our
understanding of the roles of niche
expansion and competition in explaining
the role of plant-plant interactions on local
richness (Brooker et al. 2008).
Most conceptual models of
facilitation developed to date are based on
the notion that stress-tolerant plants
increase the performance of competitive
plants under “high levels of stress”. Thus
the underlying assumption of an
overarching stress gradient affecting plant
communities has permeated the facilitation
literature (Lortie et al. 2004a, Travis et al.
2005, Michalet et al. 2006). However,
stress is a complex concept (see Körner
2003, Lortie et al. 2004b, Körner 2004, for
a recent discussion) and is more
appropriately applied at the level of
individual species than at the community
level. Individual species are adapted to
tolerate particular environmental
conditions, and under these particular
conditions, a given species will experience
little or no stress (Körner 2003). However,
the morphological and physiological
adaptations to particular environmental
conditions constitute trade-offs in a species´
ability to cope with different stressors
(Tilman 1988, Niinemets and Valladares
2006). When a given species has to cope
with environmental conditions which it is
maladapted therefore, intuitively it could be
considered to be far from its environmental
optimum, and therefore limited or stressed
(Lortie et al. 2004b). Since species forming
a community do not need to be adapted to
exactly the same environmental conditions,
we can find, within a given community,
species differing in their optima and
therefore differences in the degree of stress
that they experience across environmental
gradients (Chapin et al. 1987, Greiner La
Peyre et al. 2001). We refer to this concept
henceforth as the individual-based stress
concept (Fig. 4.1). This may have profound
implications for our understanding on how
plant-plant interactions affect local richness
and its relationship with productivity, since
niche expansion does not necessarily need
to increase in the “moderate to high stress
direction” (Lortie et al. 2004a, Travis et al.
2005, Michalet et al. 2006). Rather, a
general mechanism operating across the
entire environmental gradient should be that
better adapted plants increase the realized
niche of less adapted species to a given set
of environmental conditions (Bruno et al.
2003). Thus when dealing with natural
plant communities, there is a clear need to
question the existence of an overarching
stress gradient for the whole community, or
of ecological strategies that remain constant
along these gradients (Greiner La Peyre et
al. 2001, Prider and Facelli 2004, Holmgren

CHAPTER 4 .
117
and Scheffer 2010). A reassessment of the
assumptions underlying these notions will
pave the way for an improved
understanding of the relative roles of biotic
interactions and abiotic conditions as
drivers of species diversity along
productivity gradients.
In this study we aimed to develop a
unifying framework that explains the
relative effects of plant-plant interactions
and environmental conditions on local
species richness across environmental
gradients. Our framework includes the two
main processes by which plant-plant
interactions may affect local diversity: 1)
niche expansion due to facilitation, and 2)
changes in competitive outcomes beneath
nurse canopies. We have included in this
framework an assessment of changes in the
intensity, importance and frequency of
positive interactions on community-level
plant richness and productivity along an
environmental gradient. We have also
evaluated changes in the intensity and
importance of a large number of pairwise
interactions along such gradients. To
develop our framework we established plots
across wide environmental gradients in
Spain and Australia, and assessed the
relative roles of plant-plant interactions and
abiotic constraints as drivers of the local
species richness at each plot. We chose
these two regions because they have
contrasting vegetation communities and
management histories, and both exhibit a
relatively wide diversity in species richness
across their respective environmental
gradients. Our main hypotheses were that:
1) nurse plants will enhance local species
richness via niche expansion and changes in
the competitive networks (niche segregation
and competition intransitivity) of their
understorey vegetation; 2) positive pairwise
interactions will wane at environmental
conditions corresponding to levels of
extreme stress for involved plants (Michalet
et al. 2006, Maestre et al. 2009a) and 3)
since different species co-ocurring in a
community differ in their relative tolerances
to given environmental conditions, the
intensity, importance and frequency of
facilitation at the community level, and
therefore niche expansion, will remain
constant across stress gradients as the
identity, but not the amount, of facilitated
species changes along such gradients
(Greiner La Peyre et al. 2001).
METHODS
STUDY AREA
Two semiarid regions were selected for this
study, one located in the Stipa tenacissima
steppes of central and south-eastern Spain,
and the other in the semiarid eucalypt

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
118
woodlands of eastern Australia. We
surveyed 10 (Australia) and 11 (Spain) sites
along climatic gradients ranging from 13º–
17º C mean annual temperature and 273–
488 mm average annual rainfall in Spain,
and from 16º–19º C and 280–630 mm in
Australia. Plot selection included the full
range of environmental conditions existing
within the distribution of both vegetation
communities, as recommended to test the
relationship between plant-plant
interactions, community processes and
environmental gradients (Brooker et al.
2008, Lortie 2010). Within each country the
selected plots shared a common soil type,
management style and vegetation
community type, and were selected with
similar orientation and slopes to minimize
any experimental noise that could
potentially influence the effect of climatic
conditions on the stress experienced by the
plant community. Stipa sites were located
on limestone soils. Vegetation was in all
cases an open steppe (mean cover 35 to
68%) dominated by the perennial tussock
grass Stipa, with sparse resprouting shrubs
such as Quercus coccifera, Pistacia
lentiscus and Rhamnus lycioides. Sites in
south-eastern Australia were open
woodlands located on clay loam soils.
Canopy cover, which ranged from 18-70%,
was dominated by Eucaliptus populnea, E.
intertexta, E. microtheca, Geijera
parviflora and several shrub species
(Eremophila mitchelii, E. sturtii, Dodonaea
viscosa, Acacia spp. and Senna spp.).
Details of the study sites and images of the
communities are given in Appendix H.
VEGETATION SURVEY
At each site we established a 30 m × 30 m
plot, containing the representative
vegetation of the surrounding area. This
plot size permitted the inclusion of several
shrub and tree patches within this area,
enough to conduct the facilitation surveys
described below. Within each plot we
centrally aligned three 30 m long transects,
8 m apart, down the slope for the vegetation
survey. Along each transect we placed 20
contiguous 1.5 m × 1.5 m quadrats, and
recorded the cover and abundance of all
perennial plant species within the quadrat.
These data provided us with a
presence/absence matrix of 80 columns
(four transects by 20 quadrats) for each
plot. The total cover of each plot, which has
been shown to be a good surrogate of
productivity in semiarid environments
(Flombaum and Sala 2009), was derived
from the average cover of perennial plants
across the 80 quadrats. This survey was
used to examine differences in community
composition derived from contrasting
environmental optima of the different
species forming each community across the

CHAPTER 4
119
Figure 4.1. Conceptual diagram synthesizing current facilitation models (upper panel) and our newly proposed model (lower panel). Current facilitation models are the Stress-Gradient Hypothesis (SGH), as originally presented by Bertness and Callaway (1994) and the Facilitation waning models, as a synthesis of new empirical data and several proposed modifications of the SGH (see main text). The SGH predicts an increase in positive interactions either under high consumer pressure or abiotic stress level (parts A, and E-F of the upper panel), being competition more important under moderate conditions (part C of the upper panel). Facilitation waning models propose that positive interactions dominate under high, but not extreme, levels of either abiotic stress or consumer pressure (parts B, D and E of the panel). In contrast with the SGH, these positive interactions collapse when consumer pressure reaches extreme high levels (e.g. Smit et al. 2007; part A in the upper panel) or when abiotic stress reaches this extreme levels (part F in the upper panel). Among facilitation waning models, some of them propose that negative interactions dominate again under such stressful conditions (e.g. Maestre et al. 2009a; discontinuous line), while others suggest that the sign of the interaction becomes neutral (e.g. Michalet et al. 2006). This differentiation is caused by the different explanations invoked to explain this facilitation waning (see main text). Our newly proposed model (Individual-based stress concept; lower panel) does not take into account if the “abiotic stress” or “consumer pressure” are extremely high, moderate or low, because their effects may differ depending on the physiological tolerances of the species involved, and thus are difficult to predict. Instead, we use the distance to the environmental optimum of each target species to predict the effects of nurse plants. As a given

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
120
particular species moves further away from its optimum, the effects of a nurse plant will become more positive (Greyner La Peyre et al. 2001, Holmgren and Scheffer 2010). This interaction will become more negative as the target species gets closer to its environmental optimum. This model also predicts a facilitation collapse when the given environmental conditions are so severe for a given target plants that the recruitment is impossible, even under the safe sites provided by the nurse plants (Kitzberger et al. 2000, Ibañez and Schupp 2001., Soliveres et al. in press)
gradient in each region (see Appendix I for
a detailed description of this analysis).
FACILITATION SURVEY
Because of differences in vegetation
physiognomy, we conducted facilitation
surveys using slightly different protocols in
Spain and Australia. In each plot located in
Spain, ten Stipa tussocks (hereafter Stipa
microsite) were randomly selected, and the
total area under their canopy sampled with
0.5 m × 0.5 m quadrats. The abundance and
cover of all perennial plant species were
recorded within each quadrat. Since most of
the species in the studied areas have
canopies smaller than the 0.5 m × 0.5 m
area, we believe that this is an appropriate
size to evaluate species co-ocurrence on a
distance closer enough to ensure that the
species found were interacting someway.
Ten paired open areas, located at least 1 m
from any Stipa tussock or resprouting shrub
(hereafter Open microsite), were randomly
selected adjacent to these tussocks. We
balanced our sampling effort among
microsites by sampling the same area (i.e
the same number of 0.25 m2 quadrats) of
Open microsites as that sampled under
Stipa. Finally, we sampled the same area
under the canopies of five Quercus
coccifera (or another resprouting shrub
species when Quercus was absent from the
plot; hereafter Shrub microsite).
For the Australian plots, we sampled
three different microsites; Open, Shrub and
Tree. Shrub microsites were represented as
inverse cone-shaped (sensu Whitford 2002)
shrubs such as Eremophila mitchellii,
Dodonaea viscosa, Senna artemisioides or
juvenile Callitris glaucophylla, depending
on the species present in each plot. Our
sampling protocol changed slightly
depending on the canopy size of shrubs.
Where the canopy was sufficiently large,
we sampled six 0.5 m × 0.5 m quadrats
under each of five shrubs. Where shrubs
were smaller, a larger number of shrubs
were sampled in order to sample a total of
30 quadrats. Paired Open microsites (> 1 m
from any shrub or tree) were also sampled
adjacent to these shrubs to yield the same
sampling area. Finally, we sampled the
same number of quadrats under the Tree
microsites, which were represented by
different species of eucalypts (E. populnea,
E. intertexta, E. microtheca), Casuarina
pauper or Geijera parviflora. Because of
the large area occupied by these tree species

CHAPTER 4 .
121
(up to 200 m2 for some Eucalyptus trees),
we sampled only three trees in each plot.
The canopy area of all shrubs and trees was
calculated based on the area of a circle. For
both Spain and Australia, plot-level
richness was calculated by summing up the
total number of different species found in
the 30 0.5 m × 0.5 m, and eighty 1.5 m ×1.5
m quadrats sampled.
STATISTICAL ANALYSES
-Reduction of climatic data
Eight climatic variables (annual radiation,
minimum, maximum and mean
temperature, and rainfall, temperature range
[maximum-minimum temperature], and
minimum and maximum temperatures for
the coldest and warmest month,
respectively) were collected for each site
using available climatic models (Ninyerola
et al. 2005) and data from the Bureau of
Meteorology (www.bom.gov.au) in Spain
and Australia, respectively. We reduced
these climatic variables to a single synthetic
variable for each country using PCA.
Summarizing environmental variables in a
PCA allowed us to obtain a more general
assessment of the influence of all of our
environmental variables at both community
and species-specific levels. This approach is
strongly recommended for testing
relationships between plant-plant
interactions and abiotic stress along
environmental gradients (Lortie 2010). We
used the first PCA axis as our surrogate for
the climatic gradient present at our sites in
both countries (hereafter referred to as
Climate). This axis explained 88.6%
(Eigenvalue = 8.08·103) and 86.2%
(Eigenvalue = 1.07·104) of the variance in
the climatic data for Spain and Australia,
respectively. This axis was highly
correlated with rainfall and radiation in
Spain (Eigenvectors = -0.864 and 0.502 for
rainfall and radiation, respectively; the
remainder of the eigenvectors were < 0.03
in all cases) but only with rainfall in
Australia (Eigenvector = 0.996; the rest of
eigenvectors were < 0.1 in all cases).
Principal Component Analyses were carried
out in Primer v. 6 statistical package for
Windows (PRIMER-E Ltd., Plymouth
Marine Laboratory, UK).
We evaluated the relationships
between Climate and both cover and
species richness at the plot level, and the
relationship between cover and richness,
using both linear and quadratic regressions
because either linear or unimodal
relationships between these variables are
expected from previous studies (e.g. Grime
1973, Whitford 2002). Regression analyses
were carried out using SPSS 13.0 for
Windows (Chicago, Illinois, USA).

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
122
-Assessing biotic interactions
Because plant-plant interactions cannot be
measured in a simple way (Maestre et al.
2005, Brooker et al. 2005), we applied three
complementary indices to quantify their 1)
intensity, 2) importance and 3) frequency.
The intensity and importance of plant-plant
interactions were assessed using both plant
richness and cover. To measure the
intensity of the interactions, i.e., the effect
that neighbours have on species richness
and cover regardless of other environmental
factors (Brooker et al. 2005), we used the
Relative Interaction Index (RII; Armas et
al. 2004). This index is calculated for each
microsite pair as (PNurse – POpen)/(PNurse +
POpen), where PNurse is either mean cover or
mean species richness under the canopy of
a nurse plant (Stipa, Shrub or Tree
microsites) and POpen is either mean cover
or mean species richness in the Open
microsite. This index has good statistical
properties, which make it suitable for
comparing the intensity of plant-plant
interactions across environmental gradients;
it has defined limits (-1,+1), is symmetrical
around zero, and has identical absolute
values for competition and facilitation. It is
also linear, unbiased at low intensity
interactions, and has no discontinuities in
its range (Armas et al. 2004). For each plot
we calculated the mean index obtained from
all the Nurse-Open microsite pairs sampled.
To assess the importance of plant-
plant interactions, i.e., the relative effect
that Stipa, shrubs and trees had on richness
and cover compared to that of other
environmental factors (Brooker et al. 2005),
we used the Interaction Importance Index
(Iimp; Seifan et al. 2010b), which has similar
statistical properties to RII and is therefore
comparable among sites located across the
environmental gradient sampled. This index
is calculated as Iimp= Nimp/│Nimp│+│Eimp│,
where Nimp and Eimp are the nurse plant and
environmental contributions to species
richness or total cover, respectively. Nimp is
calculated as PNurse – POpen, and Eimp as POpen
– MPOpen/Nurse, where MPOpen/Nurse is the
maximum value of species richness or
mean cover found in the entire gradient,
irrespective of the microsite sampled.
Finally, the frequency of positive
interactions was measured as the percentage
of either facilitation obligates and
facilitation beneficiaries (sensu Butterfield
2009), as a percentage of the total species
pool of each plot. We considered as
facilitation obligates those species found
only under the canopy of a given nurse
plant but not in the Open microsites
(regardless of the identity of the nurse
plant), while facilitation beneficiaries were
species with more individuals growing
under the canopy of a nurse than in the
Open microsites. We used the number of
recruited individuals because seedling

CHAPTER 4 .
123
germination and establishment, particularly
during the first year, are known to be the
principal bottlenecks in plant recruitment in
semiarid environments, such as those
sampled (Eldridge et al. 1991, Escudero et
al. 1999, Maestre et al. 2001). Thus, we
believe that a higher number of individuals
recruiting in a given microsite is indicative
of superior environmental conditions
experienced in this microsite for a given
species, an approach followed by previous
studies on the topic (Valiente-Banuet et al.
2006).
The three attributes used to quantify
plant-plant indicators (intensity, importance
and frequency) were plotted against
Climate and mean nurse size for each plot
(i.e. the area occupied by the nurse plants).
These relationships were tested with both
linear and quadratic regressions, following
predictions from previous facilitation
models (Bertness and Callaway 1994,
Michalet et al. 2006, Maestre et al. 2009a).
In the case of frequency, we calculated the
percentage of obligate and beneficiary
species for each microsite and country
separately, and then for the whole
community. For the whole community we
added together data for both nurse
microsites (Stipa and Shrub, Spain; or
Shrub and Tree, Australia) and also tested
them separately for each country. This
battery of approaches provided us with a
complete assessment of the relationships
between plant-plant interactions and abiotic
stress. Furthermore, it allowed us to
evaluate the importance of nurse area as a
driver of this relationship. This area
influences the effects of nurses on
microclimatic amelioration and niche
availability, and thus affects the richness
and cover of understorey plants (Pugnaire et
al. 1996a, Maestre and Cortina 2005,
Michalet et al. 2006).
Furthermore, to assess the relative
role of abiotic stress and nurse size at the
pairwise level, we selected for each country
separately, those species present in at least
three different sites across the sampled
environmental gradient, which ensured that
we had at least three points with which to
test the relationship between plant-plant
interactions and abiotic stress using climatic
features, as recommended (Lortie 2010).
With the selected species (16 and 13 for
Spain and Australia, respectively), we
calculated Iimp and RII for all the species
using cover of the target species as our
proxy of plant performance. If the intensity
or importance of facilitation increase with
abiotic stress, as predicted by the “stress-
gradient hypothesis” (SGH; Bertness and
Callaway 1994, Callaway 2007), we would
expect to detect an increasing trend of

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
124
positive interaction outcomes with
increasing stress. Alternatively, if
facilitative interactions collapse at
extremely high stress levels (i.e. low
rainfall), we would expect to find a
unimodal relationship between facilitation
indicators and climate, which may or may
not be related to nurse size (facilitation
waning; Maestre and Cortina 2004a,
Michalet et al. 2006, Maestre et al. 2009a).
To test the generality and validity of these
three models (SGH, facilitation waning
because of smaller nurses, and facilitation
waning because of increasing competition),
we evaluated the relationships between RII,
Iimp obtained with all the pairwise
interactions tested in each country, and the
climatic PCA axis or the nurse size using
Spearman correlations. To assist us in the
interpretation of these analyses, the
percentage of pairwise interactions that
followed each model prediction was also
calculated for each country and nurse plant.
If the Spearman correlation with Climate is
significant and negative, this will give
support to the SGH, if the relationship
between the interaction indices and Climate
is unimodal, this will give support to
facilitation waning models. If the latter is
also related with nurse size, this will
support the facilitation collapse derived
from nurse plant growth limitation. The
percentage of cases explained for a given
model is another measurement of the fitness
of each model to our data.
-Measuring changes in plant-plant
interaction outcomes depending on
microsite
Nurse plants may affect the competitive
outcomes of their understorey plants by
increasing competitive intransitivity or by
niche segregation (Brooker et al. 2008).
Both mechanisms are related to the
maintenance of higher species richness than
if competitive exclusion alone dominates
interactions among understorey plants
(Silvertown 2004, Laird and Schwamp
2006). Recent models have highlighted the
importance of competition intransitivity as
a key modulator of species richness (Laird
and Schamp 2006, 2008, Bowker et al.
2010). The degree of intransitivity can be
defined as the absence of a competitive
hierarchy among the species coexisting in a
community (Gilpin 1975). However, this
concept assumes that the competitive ability
of those species making up a given
community are constant along the whole set
of possible environmental conditions. To
adequately test intransitivity it is necessary
to measure the competitive ability of every
species against every other one (Grace et al.
1993), making it exceptionally difficult to
test empirically. Alternatively, niche
segregation has been identified as a crucial
mechanism increasing species richness (e.g.

CHAPTER 4 .
125
Silvertown 2004), and is more likely to
occur under the most heterogeneous
conditions found beneath nurse plants,
rather than in Open microsites (Pugnaire et
al. 1996a, Maestre and Cortina 2005).
However, niche segregation may change
along environmental gradients (Huston
1999), and is also difficult to measure
empirically when considering the whole
plant community.
We attempted to measure changes in
competition intransitivity and niche
segregation, and their relationship with the
plot-scale diversity of species found in each
microsite, by using null models of guild
structure based on patterns of species co-
occurrence (Gotelli and Graves 1996,
Gotelli et al. 2010). These null models are
organized a priori by groups of ecological
significance, such as different functional
groups or trophic levels (i.e. species guilds),
and allow testing of the role of competition
in structuring the community within each
guild separately (Gotelli and Graves 1996).
This analysis is not limited to grouping by
species guild. In reality, any type of a priori
group could be examined.
For co-occurrence analyses, we
organized our presence/absence data
(obtained from the 0.5 m × 0.5 m quadrats)
by microsite guilds, that is, we calculated
species co-occurrence (C-score index,
explained below) independently for each of
the Stipa, Shrub and Tree microsites by
pooling all the sampled quadrats of these
microsites within each plot (n = 30),
obtaining a unique value per microsite and
per plot. Most of the species sampled in
Spain and Australia are small shrubs or
grasses, and therefore the quadrat size used
is particularly suitable to include
interactions among them without including
random co-ocurrence or exclusions not
related to competition among them. We
estimated species co-occurrence with the C-
score index, a metric commonly used in this
kind of analyses (e.g. Dullinger et al. 2007,
Maestre et al. 2008, Rooney et al. 2008).
This index is calculated for each pair of
species as (Ri - S)(Rj - S), where Ri and Rj
are the number of total occurrences for
species i and j, and S is the number of
quadrats in which both species occur. This
score is then averaged over all possible
pairs of species in the matrix (Gotelli
2000). The C-score is related to the
competitive exclusion concept of
“checkerboardness” i.e., how many of the
possible species pairs in a given community
never appear in the same quadrat together.
Thus, positive and large values of this index
indicate that competition may be the
prevalent mechanism determining the co-
occurrence patterns observed (Gotelli

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
126
2000). To determine the strength of co-
occurrence in a sample, the observed C-
score value is compared against a set of null
models which serve as a baseline for what a
community unstructured by species
interactions would look like (Connor and
Simberloff 1979). As the values of the C-
score are dependent on the number of
species and co-occurrences observed within
each plot, we obtained a standardized effect
size (SES) as (Iobs - Isim) ⁄ Ssim, where Iobs is
the observed value of the C-score, and Isim
and Ssim are the mean and standard
deviation, respectively, of this index
obtained from the n simulations performed
(Gotelli and Entsminger 2006).
We used ‘fixed rows–equiprobable
columns’ null models and 5000
simulations. With this approach, each
species conserved its own abundance (rare
species remained rare and common species
remained common) and each quadrat was
assumed to have the same probability of
being colonized as the remainder,
regardless of the number of species found
in each quadrat, during the simulations.
This null model has been recommended for
standardized samples collected in
homogenous habitats (Gotelli 2000), such
as the ones gathered in this study. We also
used the “fixed rows-fixed columns”
algorithm (both species and quadrats
conserved their relative abundance and
richness, respectively) to add confidence to
our conclusions. The results obtained with
this analysis were similar to those obtained
with the ‘fixed rows–equiprobable
columns’ null model, and thus are not
shown.
Standardized Effect Size (SES)
values of the C-score less than or greater
than zero indicate prevailing spatial
segregation and aggregation among the
species within a community, respectively.
To assess the extent to which changes in
competitive outcomes affect local diversity,
we compared the SES obtained with the
plot-level richness found in each microsite
(hereafter ‘plot richness’). The logic
underlying the use of SES and plot richness
values to measure competition intransitivity
or niche segregation is that we assume that
SES will be higher when competitive
exclusion is more important at the quadrat-
scale. High SES values can lead to two
different outcomes: 1) a reduction in plot
richness because a few dominant species
occupy all of the available space (i.e. when
the differences in competitive ability among
co-existing species are high, competitive
transitivity leads to competitive exclusion
and low diversity), or 2) an increase in plot
richness, if there is a lack of a competitive
hierarchy and competitive dominants in
each quadrat, depending on the
microenvironmental conditions existing in
each different quadrat. In this case, a high
quadrat-scale competition will generate

CHAPTER 4 .
127
high turnover/heterogeneity in the
dominance of a given species, ultimately
leading to a high plot richness
(intransitivity increases plot richness; Laird
and Schwamp 2006, 2008). Alternatively,
SES values will be lower if 1) the
competitive ability of co-existing species is
more equilibrated at the quadrat-scale, and
thus niche segregation prevails, or 2) spatial
aggregation, and therefore positive effects
of the nurse on their understorey species,
prevail (Tirado and Pugnaire 2005). If we
analyze the effects of nurse plants on SES
and plot richness separately, there are four
possible responses: 1) Nurse plants have a
joint effect reducing SES and increasing
plot richness compared to Open microsites:
these plants promote the development of
understorey/neighbour species via niche
segregation and this has positive effects on
the overall plot-scale richness, 2) Nurse
plants increase both SES and plot richness:
these plants increase quadrat-scale
competition, but species with competitive
advantage vary among quadrats, generating
a high species turnover, and therefore
increasing plot-scale richness
(intransitivity), 3) Nurse plants increase
SES and reduce plot richness: competitive
exclusion is the dominant interaction
between understorey species and a smaller
set of competition winners dominate all
quadrats, 4) Nurse plants do not affect SES,
regardless of their effects on plot richness:
changes in the competitive outcomes are
not an important factor modulating the
effect of nurses on plot-scale diversity.
Differences in SES and plot
richness values obtained among microsites
were compared using separate one-way
ANCOVAs for each variable. In these
models, microsite (Open, Stipa and Shrub –
Spain; Open, Shrub and Tree –Australia)
was introduced as a fixed factor, and mean
plot cover (our surrogate of overall site
productivity) was used as a covariate.
Standardized Effect Size data were √(x+1)
transformed to meet assumption of
ANCOVA analyse (normal distribution of
residuals and homoscedasticity). Tukey’s
HSD post-hoc tests were used to assess for
differences among the three microsites of
each country. We tested for relationships
among the residuals of the ANCOVA and
nurse plant canopy area using Spearman
correlations. This was necessary in order to
assess the importance of nurse size, as we
could not use nurse size as a covariate in
our model because Open microsites do not
have a size and the relationships between
nurse area and their effect on SE and plot
richness might not necessarily be linear.
To add confidence to our results,
we also developed an alternative approach

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
128
to detect the relative importance of
competitive intransitivity and niche
segregation based on a modification of
Whittaker´s beta-diversity index (Whittaker
1972). Results from this approach were
very similar, and only suggested that it was
more sensible to detect niche segregation
(see Appendix J for methodological details
and results). Null model analyses were
conducted with Ecosim 7.22 (Gotelli and
Entsminger 2006). ANCOVA analyses
were carried out using SPSS 13.0 for
Windows (Chicago, Illinois, USA).
SPAIN AUSTRALIA
mean plot cover (%)
30 40 50 60
spec
ies
richn
ess
0
10
20
30
40
50
A
R2 = 0.66; P = 0.002
mean plot cover (%)
10 20 30 40 50 60 70 80
spec
ies
richn
ess
10
20
30
40
50
60
B
-200 -150 -100 -50 0 50 100
spec
ies
richn
ess
0
10
20
30
40
50
sp richness: R2 = 0.64; P = 0.017
cover: R2 = 0.77; P = 0.003
- RAINFALL +
+ MINIMUM TEMPERATURE/RADIATION -
Cov
er (
%)
30
35
40
45
50
55
60
65
C
- RAINFALL +
-200 -100 0 100 200 300
spec
ies
richn
ess
10
20
30
40
50
60
Cover: R2 = 0.4; P = 0.047
D
Cov
er (
%)
10
20
30
40
50
60
70
80
Figure 4.2. Relationships between cover, our surrogate of standing biomass, and species richness at the community level in Spain (A) and Australia (B), respectively. The relationship between both cover (open dots, continuous line) and richness (black dots, dashed line) and the first axis of a PCA derived from climatic variables is shown for both Spain (C) and Australia (D), respectively. Significant relationships (P < 0.05) are shown as bold lines.

CHAPTER 4
129
-Assessment of niche expansion
We calculated the Chao-Jaccard abundance-
based similarity index (hereafter Chao
index; Chao et al. 2005) to assess the
relative role of niche expansion by nurse
plants on community species richness. This
index is based on the probability that two
randomly chosen individuals, one from
each of two samples (referred to as a
“pair”), belong to species shared by both
samples. It takes into account not only the
number of shared species among different
microsites, but also differences in their
relative abundances (Chao et al. 2005). We
assume that, as the influence of niche
expansion increases, more species should
be present, or more abundant, under a given
nurse plant than in Open microsites. We
calculated the Chao index at the community
level by summing over the number of
individuals and species recorded in all
quadrats for a given microsite within each
plot. Thus, the higher the dissimilarity
among nurse microsites and open areas in a
given plot, the higher the effect of niche
expansion provided by nurse canopies on
the overall community richness in this plot.
Differences in the Chao index between
microsite pairs (Stipa/Shrub vs. Open,
Shrub/Tree vs. Open, Stipa/Shrub vs.
Shrub/Tree for Spain and Australia,
respectively) were compared with one-way
ANOVA, with microsite pair as fixed
factor. Tukey’s post-hoc HSD tests were
used to assess significant differences among
pairs. After conducting the analysis, and to
assess the influence of climate in niche
segregation, we evaluated the relationship
between Climate and Chao index using
both linear and quadratic regressions. With
both approaches we can correctly evaluate
the differences in the understorey
populations between each microsite (each
nurse-type may have different effects on a
given target plant, and this may translate in
a high dissimilarity not only between
Nurse/Open microsites, but also between
different nurses), and to account for the
possible non-linear relationships between
Climate and the effects of the different
nurse plants tested on their understorey
vegetation. The Chao index was calculated
using EstimateS 8.2.0 for Windows
(Colwell 2000;
http://viceroy.eeb.uconn.edu/estimates).
ANOVA and correlation analyses were
carried out using SPSS 13.0 for Windows
(Chicago, Illinois, USA).

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
130
Table 4.1. Relationship between the indicators of interaction intensity (Relative Interaction Index, RII) and importance (Interaction Importance Index, Iimp) calculated for particular pairwise interactions and Climate. Particular pairwise interactions for both studied regions and the two different nurse microsites tested in each region are included (Stipa/Shrub and Shrub/Tree in Spain and Australia, respectively). The relationship between RII/Iimp and Climate can be nil (0), monotonically positive or negative (+ and -, respectively) or hump-shaped (hs). The last column indicates the number of sites along the gradient in which each species was present.
Spain RII
Stipa
RII
Shrub
Iimp
Stipa
Iimp
Shrub Sites
Asparragus horridus + + 0 - 3
Asphodelus ramosus hs Hs hs hs 4
Brachypodium retusum 0 0 0 + 6
Cistus clusii - - 0 - 6
Fumana ericoides 0 - + 0 7
Fumana thymifolia 0 0 0 0 6
Helianthemum cinereum 0 - - 0 6
Helianthemum violaceum 0 - 0 - 5
Rosmarinus officinalis 0 Hs 0 0 8
Sedum sediforme 0 - 0 - 6
Stahelina dubia - + hs hs 4
Stipa offneri 0 + 0 hs 5
Teucrium capitatum 0 0 0 0 6 Teucrium
pseudochamaepytis hs Hs - 0 9
Thymus vulgaris 0 0 0 hs 11
Thymus zygis hs + 0 0 3
Australia RII
Shrub
RII
Tree
Iimp
Shrub
Iimp
Tree Sites
Austrodanthonia caespitosa 0 - - - 4
Austrostipa scabra 0 - 0 0 7
Boerhavia dominii 0 0 hs hs 4
Cenchrus ciliaris - - - 0 4
Chenopodium desertorum - - hs 0 4
Einadia nutans 0 0 0 + 6
Enteropogon acicularis 0 - 0 0 6
Maireana enchylaenoides hs 0 0 0 3
Maireana sclerolaenoides hs - hs + 4
Sclerolaena muricata - - + + 5
Sida corrugata - 0 0 + 5
Sida cunninghamii - - 0 0 4
Vittadinia cuneata 0 + 0 + 4

CHAPTER 4
131
Table 4.2. Percentage of pairwise interaction outcomes, as measured with the Relative Interaction Index (RII) and the Interaction importance index (Iimp), explained by the two major current models tested in this work, the Stress-Gradient Hypothesis and the waning of facilitation under extremely high stress (Waning). As a special case in the facilitation waning model, the percentage of cases that are positively related with nurse size (compared with the total number of cases tested) are indicated below. st = Stipa tenacissima, sh = resprouting shrubs in Spain (SP) or inverse cone-shaped shrubs in Australia (AU), and tr = Eucalyptus or Geijera parviflora trees.
Model
RII Iimp
SP
(st)
SP
(sh)
AU
(sh)
AU
(tr)
SP
(st)
SP
(sh)
AU
(sh)
AU
(tr)
Stress-Gradient Hypothesis 23.5 29.4 38.5 61.5 17.6 29.4 15.4 7.7
Facilitation
Waning
23.5 17.7 23 0 11.8 23.5 15.4 7.7
Nurse size 19 0 23 0 11.8 0 15.4 7.7
RESULTS
DIVERSITY-BIOMASS RELATIONSHIP
AND THE EFFECT OF CLIMATE
A total of 96 and 131 perennial species
were found in Spain and Australia,
respectively, with a plot-level richness
ranging from 9 to 47 in Spain, and from 16
to 51 in Australia, respectively. The number
of species found in each plot was linearly
and negatively related to mean plot cover in
Spain (Fig. 4.2A). This was particularly
evident near the centre of the climatic
gradient, under conditions of both moderate
drought and moderately low temperatures
(Fig. 4.2C). In contrast, richness was
largely independent of either cover or
climate in Australia (Figs. 4.2B and 4.2D).
Mean cover at the plot level showed a
monotonic and positive increase with water
availability in both Spain and Australia,
reaching its maximum value in the more
mesic plots.
PLANT-PLANT INTERACTIONS AND
ABIOTIC STRESS
Contrasting results were found between the
studied regions. While neither intensity nor
importance of plant-plant interactions were
related to abiotic stress in Spain, we
detected a hump-shaped relationship
between most RII and Iimp values and
rainfall for the Australian sites (Fig. 4.3).
Both the percentage of facilitation
beneficiaries and obligates tended to
decrease with rainfall. This negative trend
was found to be significant for beneficiaries
in Spain and for obligates in Australia (Fig.
4.4). The percentage of plants with more
individuals under the canopy of any nurse
plant than in Open microsites (facilitation
beneficiaries) decreased from about 50% in
the drier and warmer sites to about 30% in

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
132
SPAIN AUSTRALIA
-200 -100 0 100 200 300-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
Shrub (richness) Tree (richness) Shrub (cover)Tree (cover)
-200 -100 0 100 200 300-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
B
D
Tree (cover): R2= 0.7; P = 0.005
Tree (richness): R2= 0.49; P < 0.05
Shrub (cover): R2= 0.56; P = 0.055
Tree (cover): R2= 0.57; P = 0.053
Tree (richness): R2
= 0.7; P < 0.05
-200 -150 -100 -50 0 50 100
Inte
ract
ion
inte
nsity
(R
II)
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
Stipa (richness)Shrub (richness)Stipa (cover)Shrub (cover)
-200 -150 -100 -50 0 50 100
Inte
ract
ion
impo
rtan
ce (
Iimp)
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
A
C
Climatic PCA axis values
Figure 4.3. Relationships between the indicators of interaction intensity (Relative Interaction Index, RII) and importance (Interaction importance index, Iimp), calculated for both community richness and cover, and the first axis of a PCA derived from climatic variables. Values are means ± SE per plot and microsite. Significant (P < 0.05) relationships are shown with a bold line; marginally significant ones (0.05 < P < 0.10) are also showed in the figure.
the wetter sites for Spain. This relationship
was different, however, for Australia, where
facilitation obligate plants followed a
unimodal relationship with climate. Only
10% of the species at the community level
required a nurse plant to occur at the wettest
sites, but this percentage increased up to
40% in the driest sites, and showed a
maximum (ca. 60%) at the middle of the
environmental gradient (Fig. 4.4). When we
tested the relationship between frequency of
positive interactions of each nurse plant and
climate separately, we found different
results depending on the nurse plant
analyzed at both studied regions. We
detected a similar relationship with climate
at the community level for Spanish Shrub
microsites and Australian Tree microsites,
but there were no significant relationships
for Stipa (Spain) or Shrub (Australia)
microsites. We found a marginally
significant linear relationship between nurse

CHAPTER 4
133
SPAIN AUSTRALIA
Shrub: R2 = 0.58; P < 0.005
Total: R2 = 0.61; P < 0.05
B
D
-200 -100 0 100 200 300
% fa
cilit
atio
n ob
ligat
es
0
10
20
30
40
50
60
70
ShrubTreeTotal
Climatic PCA axis values
-200 -100 0 100 200 300
% fa
cilit
atio
n be
nefic
iarie
s
0
20
40
60
80
100-200 -150 -100 -50 0 50 100
0
10
20
30
40
StipaShrubTotal
-200 -150 -100 -50 0 50 1000
10
20
30
40
50
60
C
ATree: R2 = 0.51; P = 0.02
Total: R2 = 0.61; P = 0.036
+ RAINFALL - Figure 4.4. Relationships between the percentage of facilitation beneficiaries (species with more individuals recruiting under nurse plants than in Open microsites) and facilitation obligates (species that only recruit under the canopy of nurse plants), regarding total species richness in each plot, and the first axis of a PCA derived from climatic variables. Significant (P < 0.05) relationships are shown with a bold line.
area and the percentage of the species
growing under Stipa (R2 = 0.34; P =
0.061); the same relationship for Shrub
microsites was not significant.
Relationships among nurse size and
interaction indices were not significant for
any microsites in Australia.
When testing the validity of the Stress-
Gradient hypothesis or alternative models
to predict pairwise interaction outcomes,
the Spearman correlations between
interaction indicators (RII and Iimp) and
Climate showed that facilitation tended to
decrease monotonically with rainfall in
three of the four cases in Australia (ρ
ranged from -0.27 to -0.46, P < 0.05 in all
the cases except Tree Iimp; see Fig. 4.5).
However, these interaction indicators were

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
134
not significantly correlated with Climate in
Spain (P > 0.6 in all cases). When we
visually assessed each pairwise interaction,
RII values appeared to decline with rainfall
in 38% of the total cases studied, according
to the SGH (Table 4.2). The relationship
between RII and Climate was more
common in Australia (Shrub: 38.5%, Tree:
61.5%) than in Spain (Stipa: 23.5%;
Shrub: 29.4%), which adds confidence to
the results obtained with the overall
Spearman correlations. In contrast, the
relationship between Iimp and Climate was
lower, in general. Iimp showed a
decreasing trend with rainfall in 18%
(Stipa) and 29% (Shrub) of cases for
Spain, and in 15% (Shrub) and 8% (Tree)
of cases for Australia, respectively. The
intensity and importance of the interactions
showed a hump-shaped relationship when
plotted against Climate in 20% and 14% of
cases for Spain and Australia, respectively
(Table 4.2). These cases provided support
for the facilitation waning model.
However, when we tested the relationships
between RII, Iimp and nurse size with
Spearman correlations, none of these
indices was correlated with nurse size in
any of the nurse microsites tested in both
countries (ρ < 0.2 in all cases).
Competitive exclusion, as indicated by
SES values, was lower under the canopy of
both Stipa and Shrub than in Open
microsites along the entire Spanish
gradient (Open: 1.03 ± 0.35; Stipa: 0.12 ±
0.14, Shrub: 0.03 ± 0.16; means ± SE; F2,29
= 3.01; P = 0.008). Tukey’s HSD post-hoc
tests revealed differences between Stipa
and Open (P = 0.028) and Shrub and Open
(P = 0.015) microsites, but not between
Stipa and Shrub microsites (P = 0.961).
Mean plot cover did not affect SES results,
but when analyzing the effects of microsite
on plot-level richness, we found a
significant effect of cover (F1,29 = 53.9; P <
0.0001) and a marginal positive effect of
microsite (F2,29 = 3.01; P = 0.065). Overall,
these results suggest an effective niche
expansion by nurse plants, indicating an
increase in local richness under both
nurses. However, this positive effect
decreased substantially with mean plot
cover (Spearman correlation between
residuals of the ANOVA and mean plot
cover = -0.78; P < 0.0001). Nurse size
showed no relationship with the residuals
of the ANCOVA models fitted with both
SES and plot richness in Spain or in
Australia. Differences in competitive
exclusion among microsites, as measured
with SES, were not found for the
Australian sites (F2,27 = 2.4; P = 0.101).

CHAPTER 4
135
SPAIN AUSTRALIA
-200 -100 0 100 200 300
-1.0
-0.5
0.0
0.5
1.0
Climatic PCA axis values
-200 -150 -100 -50 0 50 100
Inte
ract
ion
inte
nsity
(R
II)
-1.0
-0.5
0.0
0.5
1.0
StipaShrub
-200 -150 -100 -50 0 50 100
Inte
ract
ion
impo
rtan
ce (
Iimp)
-1.0
-0.5
0.0
0.5
1.0-200 -100 0 100 200 300
-1.0
-0.5
0.0
0.5
1.0
ShrubTree
- RAINFALL +
A
C
B
D
Shrub: ρ = -0.46; P < 0.0001Tree: ρ = -0.34; P = 0.003
Shrub: ρ = -0.27; P = 0.02
Figure 4.5. Scatter plot showing the relationship between pairwise facilitation indicators (intensity [RII, panels A and B for Spain and Australia, respectively] and importance [Iimp, panels C and D for Spain and Australia, respectively]) and Climate. Spearman correlation coefficients and P values are shown in each case. However, microsite significantly affected
plot richness in this country (F2,26 = 4.05; P
= 0.03). While the highest richness was
found in Shrub microsites (plot richness =
17±1.7 and 11 ± 1.5 for Shrub and Open
microsites, respectively; mean ± SE; Tukey
HSD: P = 0.029), this effect was less
marked in Tree microsites (plot richness =
15.8 ± 1.5; Tukey HSD: P = 0.093 for Tree
vs. Open microsites). In contrast to the
results found in Spain, mean plot cover did
not modify the effect of microsite on SES
or plot richness.
NICHE EXPANSION
Although the similarity index was slightly
lower for Shrub vs. Open microsites (0.64 ±
0.08, mean ± SE, n = 11) than for Stipa vs.
Open microsites (0.74 ± 0.08), we did not
find significant differences in the similarity
index among nurse microsites in Spain
(Stipa vs. Shrub, 0.68 ± 0.06, n = 11;
ANOVA: F2,29 = 0.622; P = 0.54). Shrub
and Stipa microsites shared about 70% of
their understorey populations with Open
sites. Significant differences in similarity
among microsite pairs were found,

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
136
however, in Australia (F2,26 = 4.57; P =
0.02), suggesting some degree of niche
expansion. Populations shared between
Shrub-Open or Shrub-Tree pairs were close
to 60%, though the similarity index for
Tree-Open microsites showed that they
shared about 40% of the individuals (Tukey
HSD post-hoc test; P = 0.034, 0.710 and
0.171 for Open-Shrub vs. Open-Tree,
Open-Shrub vs Shrub-Tree, Open-Tree vs
Shrub-Tree pairs, respectively).
-200 -150 -100 -50 0 50 100
Cha
o-Ja
ccar
d si
mila
rity
inde
x
0.0
0.2
0.4
0.6
0.8
1.0
Open vs Stipa Open vs Shrub Stipa vs Shrub
SPAIN
-200 -100 0 100 200 3000.0
0.2
0.4
0.6
0.8
1.0
Open vs ShrubOpen vs TreeShrub vs Tree
AUSTRALIA
Climatic PCA axis values
- RAINFALL +
Open vs Tree: R2 = 0.64; P = 0.028Shrub vs Tree: R2 = 0.63; P = 0.032
Figure 4.6. Relationships between Chao-Jaccard similarity index, our surrogate of niche expansion, and Climate in Spain and Australia. Data from the three different microsite pairs for each country (Open vs Stipa, Open vs Shrub, and Stipa vs Shrub for Spain; Open vs Shrub, Open vs Tree, and Tree vs Shrub for Australia) are shown. Significant relationships (P < 0.05) are shown as bold lines.
Overall, these results indicate that, while
Shrub microsites shared 60% of the
populations beneath their canopies with
both Tree and Open microsites, the
similarities between Tree and Open
populations were lower (ca. 40% shared).
Similarity among the microsites tested did
not show any relationship with Climate in
Spain, but two of the three indices
calculated showed a quadratic relationship
with this variable in Australia (Fig. 4.6),
suggesting a trend of increasing similarity
at both ends of the climatic gradient.
DISCUSSION
SPECIES RICHNESS-PRODUCTIVITY
RELATIONSHIP AND THE CONCEPT
OF STRESS
The results of our studies from both Spain
and Australia did not conform with the
hump-shaped relationship between richness
and productivity predicted by pioneering
studies (Grime 1973, Huston 1979). Several
reviews and meta-analyses have questioned
the universality of this hump-shaped
relationship (Grace 1999, Waide et al.
1999, Gillman and Wright 2006). Although
the unimodal richness-productivity
relationship is rarely proven empirically, it
continues to be used to invoke the role of
facilitative interactions on increasing plant
community diversity (Hacker and Gaines
1997, Michalet et al. 2006). Previous

CHAPTER 4 .
137
studies have described the roles of stress-
tolerant plants on niche expansion of more
competitive species (Lortie et al. 2004a,
Travis et al. 2005, Michalet et al. 2006).
However, we argue that, as different species
are adapted to different environmental
conditions, their environmental optima will
occur at different points along any given
productivity gradient (Chapin et al. 1987,
Ibañez et al. 2007, Holmgren and Scheffer
2010; see also Appendix I). A more general
framework should consider the species-
specific nature of ‘stress’ (Körner 2003)
and the ‘distance’ of each species from its
ecological optimum, rather than the
ecological strategy that it employs. Given
the absence of a clear overarching stress
level affecting whole plant communities
along environmental gradients (Chapin et
al. 1987), such a framework is clearly
needed. Our results suggest that plant-plant
interactions not only enhance diversity at
mid to low productivity levels (sensu
Hacker and Gaines 1997, Michalet et al.
2006), but that this effect extends to the
entire productivity gradient via
environmental amelioration by nurse plants
of the less adapted species to a given set of
environmental conditions (Holmgren and
Scheffer 2010). This positive effect is
reinforced because nurse plants not only
increase the available species pool in a
given plot, but also promote the existence
of a richer community because of greater
environmental heterogeneity (Pugnaire et
al. 1996a, Maestre and Cortina 2005).
PLANT-PLANT INTERACTIONS
ALONG ENVIRONMENTAL
GRADIENTS
Community productivity in both Spain and
Australia was limited mainly by water
availability, consistent with the
expectations for arid and semiarid
environments worldwide (Noy-Meir 1973,
Whitford 2002; Fig. 4.2). Thus, the
negative trend found in the frequency of
positive interactions with increasing rainfall
in both areas provides strong support for the
original predictions of the SGH (Bertness
and Callaway 1994). However, at the
Australian sites, where environmental stress
seemed to reach extremely high levels (plot
cover declined to 17% in some cases; see
Appendix H), positive effects of nurses on
community richness decreased at the
highest stress levels, consistent with results
from other semiarid environments
(Kitzberger et al. 2000, Maestre and Cortina
2004a, Anthelme et al. 2007). We found
some evidence that increased nurse size
could explain increased facilitative effects
of Stipa in the Spanish sites. Percentage of
facilitation obligate species and the

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
138
intensity of the positive effects on
community diversity increased with Stipa
size, suggesting an increased capacity to
ameliorate harsh environmental conditions
by larger tussocks. However, in contrast to
theoretical predictions (Michalet et al.
2006), nurse area was unrelated to either
importance, intensity or frequency of
positive interactions at the community level
in the other microsites tested. The model
developed by Michalet et al. (2006) was
based on empirical data using mostly
tussock-like nurses, plants similar to Stipa,
or cushion-like plants (e.g. Choler et al.
2001, Callaway et al. 2002, Liancourt et al
2005, Anthelme et al. 2007). Small
increases in the size of these nurses are
likely to have relatively large facilitatory
effects on understorey species (Michalet et
al. 2006, Anthelme et al. 2007). However,
relatively small changes in large nurse
plants, such as the Australian eucalypt trees
(with canopy areas up to 200 m2), would be
less likely to be influential. We suggest
caution, therefore, in generalizing the
effects of nurse plants in semiarid
environments without a consideration of
their size. Even when the smallest nurse
plants almost always exceed an area of 1.5
m2, such as the microsites tested in this
studies (excepting Stipa), we would not
expect a strong relationship between nurse
size and their tendency to alleviate
environmental stress. An alternative
explanation for the positive effects of nurse
size found in pairwise interactions (Table
4.2) could result from increased
heterogeneity in microclimate (e.g. shade,
temperature, light) at different parts of the
canopy, which would be expected to
increase niche segregation (Pugnaire et al.
1996a, Maestre and Cortina 2005). Thus,
the higher availability of different niches
and the associated decreases in inter-
specific competition via niche segregation
(Huston 1979, see discussion below) could
enhance the performance of particular
species more than any increase in the ability
of the nurse to buffer environmental
stressors per se.
Our results demonstrated
inconsistent relationships between
environmental stress and both interaction
intensity and importance, contrary to
prevailing facilitation theory (Brooker et al.
2005, Callaway 2007). In Spain, neither
interaction intensity nor importance were
related to the environmental gradient we
evaluated, but a hump-shaped relationship
for both facilitation/competition indicators
was found in Australia. How can we
account for this difference? Environmental
stress in the Spanish gradient was driven by
two negatively correlated and distinct
stressors; water and radiation/temperature.
Thus, it is likely that in the coldest or
warmest extremes of this gradient, less
cold- or drought-adapted plants would

CHAPTER 4 .
139
benefit from nurse canopies, respectively
(Choler et al. 2001, Liancourt et al. 2005).
This may explain why the net positive
effects of nurse plants were equally intense
or important at the community level along
the entire gradient (Tilman 1988). Indeed,
this is suggested by the fact that the
proportion of facilitation obligate species
was not significantly related to climate in
this region. The higher number of
facilitation beneficiary species under drier
conditions could be related to the fact that,
regardless of their physiological adaptions
or environmental optima, germination and
recruitment of most semiarid species are
limited by periods of adequate soil moisture
(Westoby 1978/79). These periods are more
easily achieved under the more shaded
onditions beneath nurse plants than in Open
microsites (Franco and Nobel 1989).
Therefore, under drier conditions we would
likely record more individuals of most of
species under the canopy of nurse plants
than in Open microsites (Kitzberger et al.
2000). The environmental stress in the
eastern Australian gradient was
predominantly driven by a single stressor,
rainfall, through its influence on soil
moisture availability. Consequently, nurse
plants could conceivably have an important
role in allowing recruitment and persistence
of taxa less adapted to low soil moisture
(Kitzberger et al. 2000, Soliveres et al.
2010). This facilitative role collapsed at
extremely high stress levels. The most
parsimonious explanation for this is that
facilitation collapsed because species with
low tolerances to drought and/or herbivory
were unable to overcome the environmental
filters controlling their recruitment,
regardless of the presence of nurse plants
(Kitzberger et al. 2000, Ibañez and Schupp
2001, Michalet et al. 2006, Soliveres et al.
in press), and therefore could not be
included in our pairwise analyses. Thus, the
positive effects on richness and productivity
at the community level collapsed under
these extremely stressful conditions (Forey
et al. 2009). This is consistent with
observations of higher percentage of
facilitation obligate species and lower
similarity at moderate levels of drought
stress, and with the breakdown of these
effects under extremely high levels of
drought (Fig. 4.4).
THE EFFECT OF PLANT-PLANT
INTERACTIONS ON DIVERSITY:
NICHE EXPANSION AND
SEGREGATION
The results from the Chao index of
similarity, and the lack of relationship
between this index and Climate (Fig. 4.6),
suggest that facilitation from nurse plants

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
140
promotes niche expansion for the less
adapted species to the given environmental
conditions present in a particular site across
the gradient, rather than increased niche
expansion of competitive species under
harsh conditions. Our results partially
contrast with previous studies, which have
suggested that stress-tolerant species
increase the realized niche of competitive
species on harsh (i.e. drier) environments
(Travis et al. 2005, Michalet et al. 2006).
We argue that this is not caused by the
stress-tolerator or competitor strategy of the
species involved, but rather the distance of
each target species to its ecophysiological
optimum. It should be realized that about
20-30% of the sampled species recruited
only under the canopies of nurse plants
(Fig. 4.4), regardless of the environmental
conditions present in each plot. The total
percentage of facilitation obligate species
(when considering both microsites together)
was greater than those for separate
microsites in both the Spanish and
Australian gradients surveyed. These results
suggest that the identity of facilitation
obligate species changed according to the
particular nurse plant examined. These
changes in the identity of facilitated species
depending on the nurse plants were,
perhaps, due to their different phylogenetic
relationships (Valiente-Banuet and Verdú
2007, 2008) or to differences in their
ecological strategy (Prider and Facelli 2004,
Maestre et al. 2009a).
In contrast with previous studies
(Tielbörger and Kadmon 2000b), our
results showed that nurse plants affected the
competition outcomes of their understorey
vegetation in comparison with Open areas
through increases in niche segregation, but
not by increasing competition intransitivity.
While this was apparent in Spain, it was not
in Australia. On the Spanish gradient,
competitive exclusion was significantly
lower under nurse canopies than in Open
microsites. This could be explained because
the more productive conditions found under
the nurse plants allowed more species to
recruit. Since nurse plants provide some
degree of microclimatic heterogeneity
(Pugnaire et al. 1996a), the joint effect of
both processes (increase in species pool and
variability in the resources that these
species compete for) might increase niche
segregation, and therefore local diversity
(Huston 1979, 1999, Silvertown 2004). We
found, however, that despite the relatively
constant effect of nurse plants upon the
competitive outcome of their understorey
vegetation, the effect of nurses on local
diversity decreased with productivity. It is
conceivable that the relative differences in
the microenvironmental conditions between
nurse and Open microsites that allowed
more species to recruit under nurses than in
the unvegetated interspaces declined under

CHAPTER 4 .
141
wetter conditions. In Australia, the lack of
nurse effects on competitive outcomes, but
their positive effect on local diversity, may
have resulted from environmental buffering
(both microclimatic amelioration and
grazing protection), allowing fewer
drought- and herbivory-tolerant species to
recruit, and therefore increased diversity by
direct facilitation. However, their relative
homogeneous microenvironmental
conditions did not allow niche segregation
to occur (Huston 1979). In contrast with
previous studies (Bowker et al. 2010), we
did not find an important contribution of
competition intransitivity to local species
richness in any of the studied regions, even
in the more productive and heterogeneous
environments beneath nurse plants. This
could be a matter of scale, since the
availability of slightly different niches (i.e.
differences in shade or nutrients), even at
the 0.5 m × 0.5 m scale, might prevent the
changes in competitive dominance of
different species depending on the different
conditions in each quadrat, causing the
niche segregation, rather than competitive
exclusion at this scale.
PAIRWISE INTERACTIONS AND THE
ABSENCE OF AN APPROPRIATE
EXPLANATORY MODEL
None of the models we tested i.e. SGH
(Bertness and Callaway 1994) or
Facilitation waning model (Michalet et al.
2006, Maestre et al. 2009a; see Fig. 4.1)
predicted more than 60% of the pairwise
interactions studied (see Tables 4.1 and
4.2). The percentage of cases explained,
however, varied strongly among microsites
and regions, as well as with the identity of
the particular nurse plant (Callaway 2007,
Table 4.2). These results highlight the
difficulties in establishing generalities when
predicting how the outcome of plant-plant
interactions change along environmental
gradients. Our results are not completely
unexpected, as plant-plant interactions are
driven by a complex set of factors including
the number and type of stressors considered
(Baumeister and Callaway 2006, Maestre et
al. 2009a, le Roux and McGeoch 2010), the
particular adaptations of species to cope
with current environmental conditions
(Choler et al. 2001, Liancourt et al. 2005),
the relative effect of nurse shade on target
plants (Holmgren et al. 1997, Prider and
Facelli 2004, Soliveres et al. 2010),
phylogenetic relationships (Valiente-Banuet
and Verdú 2007), and the ontogenetic stage
of interacting plants (Callaway and Walker

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
142
1997, Miriti 2006, Soliveres et al. 2010).
This complex array of mechanisms
involved in the outcome of particular
pairwise interactions makes it extremely
difficult to develop general predictions
about the response of a given pairwise
interaction to particular environmental
conditions without a detailed knowledge of
many of the attributes described above.
Extrapolation from a few pairwise
comparisons to the broader community of
species should only be made with extreme
caution. Future general models aimed at
predicting the outcomes of particular
pairwise interactions should therefore
consider the complexities and multiplicity
of mechanisms shaping such outcomes. We
suggest here that this complexity could be
better integrated by using a surrogate of the
distance of each target species from its
optimum, either from detailed knowledge
of its physiological tolerances (i.e Choler et
al. 2001, Liancourt et al. 2005) or by
phylogenetic derived assumptions
(Valiente-Banuet and Verdu 2007). The
outcomes of pairwise interactions could
therefore be better predicted by using a
distance to optimum approach rather than
an approach that considers all species in a
given community equally, by invoking the
stress gradient model.
CONCLUDING REMARKS
Our study has highlighted the fact that
nurse plants increase local richness of plant
communities across broad environmental
gradients. Nurses increase niche
segregation and species coexistence by
providing a range of available niches
beneath their canopies, thereby allowing
species that are less adapted to a particular
position within the gradient to survive and
recruit (niche expansion). Our results show
that the importance of nurse plants is
relatively constant along environmental
gradients when different independent
stressors (e.g. low temperature and rainfall,
herbivory) have differential effects on the
stress experienced by different species
within the community. However, the
importance of niche expansion increases
along environmental gradients when a
given stressor, or a combination of
positively correlated stressors (e.g. high
temperatures and salinity or drought),
affects the stress level of different species
within the community (e.g. Callaway et al.
2002). In the latter case, this positive effect
is likely to collapse under extremely high
levels of stress through several mechanisms
previously discussed in the literature
(Michalet et al. 2006, Smit et al. 2007,
Maestre et al. 2009a).
Given the clumped nature of
vegetation in arid and semiarid areas, and
thus the tendency for plants to interact, it is

CHAPTER 4 .
143
not surprising to find that plant-plant
interactions play a major role in shaping the
relationships between richness and
productivity in these environments.
However, our results indicate that the effect
of these interactions on increasing or
decreasing richness along environmental
gradients is more complex than previously
thought, and depends on the number of
stressors involved and their
interrelationships. Our findings also help to
explain why the unimodal relationship
between diversity and productivity is rarely
found in arid and semiarid environments
(Waide et al. 1999). We maintain that the
outcome of particular pairwise interactions
is best predicted by the distance of a given
plant to its environmental optimum rather
than by an overarching stress gradient. This
has profound implications for interpreting
previous studies evaluating the interplay
between facilitation and competition along
stress gradients, and should be considered
by future research on this important theme
of community ecology. We highlight the
fact that the complex array of mechanisms
shaping the outcomes of pairwise plant-
plant interactions makes it difficult to
develop a universal model that is able to
successfully predict their outcome along
environmental/productivity gradients. We
propose that approaches considering
multiple models, such as that followed in
this study, may provide important insights
into the mechanisms driving such
outcomes, and on the community and
ecosystem-level consequences of plant-
plant interactions. By using a multiplicity of
conceptual and analytical approaches, and
an appropriate dataset collected in two
contrasted semiarid regions, our study
provides a more complete mechanistic
understanding of the relative role of biotic,
non-trophic interactions and environmental
conditions shaping local richness. It also
helps to refine our predictions of the
response of plant communities to
environmental gradients, and clarifies the
relative importance of biotic interactions as
a driver of such responses.
ACKNOWLEDGMENTS We thank David Tongway and Nick Reid for their help during plot selection and fieldwork in Australia. Nick Reid also hosted SS during a research stay in his lab. Nick Schultz, Megan Good, María D. Puche, Pablo García-Palacios, Erin Roger, Ian Telford, James Val and Madeleine Rankin assisted with fieldwork and/or plant identification. Peter Weston, Anthony Gibson, Kevin Mitchell, Andrew Mosely and Patty Byrne allowed us access to their properties and gave us valuable information on land management issues in semiarid Australian woodlands. SS was supported by a PhD fellowship from the EXPERTAL project, funded by Fundación Biodiversidad and CINTRA S.A. This research was funded by the CEFEMED, INTERCAMBIO (BIOCON 06/105) and REMEDINAL projects, funded by the Universidad Rey Juan Carlos-Comunidad de Madrid, Fundación BBVA and Comunidad de Madrid, respectively. FTM acknowledges support from the European Research Council under the European Community's Seventh Framework Programme (FP7/2007-2013)/ERC

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
144
Grant agreement n° 242658. DJE is supported by grant LP0882630 from the Australian Research Co

CHAPTER 4 .
145
Supplementary material for Chapter 4.
Appendix H. Characteristics of vegetation types and sites selected
Stipa tenacissima grasslands, the vegetation type selected in Spain, constitute one of the more
important European/North African ecosystem types, as they can be found from the semi-
desert steppes of Ukraine to the western Mediterranean (Izco 1984), with the greatest
extensions being found in the Maghreb and the Iberian Peninsula (Le Houeróu 1986, 2001).
In the semiarid parts of the Mediterranean Basin, these grasslands are distributed over 32,000
km2 in a thin latitudinal fringe in North Africa, from Libya to Morocco, and in the
southeastern Iberian Peninsula (Le Houérou 2001). See Maestre et al. (2009b) for a detailed
account of the natural history of this ecosystem. Soils in the Spanish plots are Lithic
calciorthid (Soil Survey Staff 1994) and characterized by a high CaCO3 content, high pH
values, low depth, and a stony surface.
Sites in the semiarid woodlands in eastern Australia were located in the Bimble box
(Eucalyptus populnea)–White cypress pine (Callitris glaucophylla) alliance on the Cobar
Pediplain, the Belah (Casuarina pauper)–Rosewood (Alectryon oleifolius) woodlands in far
western New South Wales (NSW), and woodlands dominated by White box (Eucalyptus
albens) in central western NSW. While these communities have slightly different community
dominants, physiognomically they are similar and characterized by an open woodland on clay
loam soils with canopy cover ranging from 18-70% (Keith 1998). The midstorey shrub cover
at all sites varied depending on grazing intensity and rainfall (Beadle 1948). Soils in the
Australian plots were non-sodic Kandosols to Demosols under the Australian classification
(Isbell 1996), or a mixture of Luvisols, Yermosols and Ferrasols (FAO 1998) and are
commonly grouped as Red earths (Stace et al 1968). They are characterized by deep clay
loam to loamy surface textures with a gradual increase in clay content with depth. They have
relatively low available nutrient and water holding capability (Isbell 1996).

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
146
Table S7. Main charactertistics of the study sites. TSR = travelling stock reserve.
Country Site
Latitude
Longitude
Land use
Annual
rainfall
(mm)
Mean
projected
Cover
(%)
Australia Cowra 33º50’41”S 148º36’12”E TSR 630 70
Australia Quandialla 33º53’19”S 147º51’27”E TSR 570 51
Australia Nevertire 31º51'27"S 147º42´28”E TSR 490 70
Australia Condobolin 33º07’15”S 142º24’44”E State Forest 455 18
Australia Nyngan1 31º10’00”S 142º24’44”E Grazing 406 31
Australia Nyngan2 31º10’00”S 142º24’44”E Grazing 406 43
Australia Florida 31º33’22”S 146º18’35”E Grazing 398 44
Australia Truganini 32º07’00”S 146º39’50”E property 375 44
Australia Etiwanda 32º09’40”S 145º53’40”E Grazing 360 21
Australia Buronga 34º07’35’’S 141º05’09”E Grazing 280 35
Spain Barrax 39º02’91’’N 2º13’82’’W Hunting area 433 46
Spain
Camporreal 40º19’72’’N 3º25’36’’W Hunting area 457 54
Spain
Carrascoy 37º48’02’’N 1º18’32’’W Hunting area 282 37
Spain
Crevillente 38º14’15’’N 0º55’49’’W Hunting area 273 35
Spain
El Ventós 38º28’14’’N 0º37’03’’W Hunting area 319 36
Spain
Morata 40º27’62’’N 3º05’31’’W Hunting area 455 58
Spain
Sierra Espuña 37º49’27’’N 1º40’41’’W Hunting area 364 49
Spain
Titulcia 40º11’28’’N 3º30’13’’W Hunting area 440 57
Spain
Villarrobledo 39º12’64’’N 2º30’77’’W Hunting area 446 63
Spain
Yecla 38º35’40’’N 1º12’15’’W Hunting area 350 36
Spain
Zorita 40º21’30’’N 2º52’62’’W Hunting area 434 45

CHAPTER 4 .
147
SPAIN AUSTRALIA
Figure S2. General appearance of the vegetation types studied in Spain and Australia.

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
148
Appendix I. Measuring community composition changes across environmental
gradients
To test for differences in community composition across the environmental gradient, we
constructed for each country a data matrix comprising the relative frequency of each species
in the 80 1.5 m by 1.5 m quadrats within each plot. Thus, a species occurring in all 80
quadrats at a given site received a relative frequency of 100%. We examined differences in
community structure between the sites with Multi-Dimensional Scaling (MDS) after applying
a fourth-root transformation to the data to downweight the influence of the most dominant
species (Clarke and Warwick 2001). Extremely rare species (those appearing in less than the
1% of the sampled quadrats), were removed prior to analysis as recommended to improve the
clarity in the analyses (Anderson et al. 2008). These rare species accounted for about half of
the sampled species (46 of 96 for Spain, 57 of 131 for Australia). We used the Bray-Curtis
distance measure to construct the similarity matrix. This distance measure controls for the
relative abundance of each species in a given matrix, and helps to avoid the undue influence
of extremely abundant or extremely rare species (Clarke and Warwick 2001). We carried out
these analyses using PRIMER v6 statistical package for Windows (PRIMER-E Ltd.,
Plymouth Marine Laboratory, UK). We analysed the data in both two and three dimensions,
and then chose the 2-D ordination, because it provided satisfactory results (i.e. low stress
values) for both studied regions (Stress = 0.10 and 0.12 for Spain and Australia, respectively).
The stress value reflects how well the data can be represented in any given number of
dimensions. Stress values < 0.05 indicate that the n axes provide an excellent representation
of the relationships among samples, while values of > 0.20 are regarded as a poor
representation. We performed the analyses with 25 random starts to reduce the risk of finding
a local instead of the global minimum of this stress function.
To identify the species or abiotic factors responsible for the ordination patterns found,
the first two axes of the MDS biplot were correlated with the PCA axis obtained with climatic
data (Climate) and with the abundance of each species in each plot using Spearman
correlation coefficients. For the latter analyses, only the species present in at least three of the
80 plots were considered. Variables with a correlation coefficient > 0.5 are represented in the
MDS plot (Fig. B1).
The ordination (see Figure below) showed a significant effect of rainfall on
community composition in Spain, where different species were related both positively or

CHAPTER 4 .
149
negatively with this factor, depending on their ecophysiological tolerances. However, this
effect was less clear in Australia, suggesting the existence of unmeasured factors affecting
community composition. The relatively high density of domestic and native herbivores in the
Australian semiarid zone (Noble and Tongway 1986) suggests that different levels of grazing
pressure among plots could be one such factor.
MDS 1
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
MD
S 2
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
Barrax
Camporreal
El Ventós
Crevillente
Morata
Zorita
Villarrobledo
Sierra Espuña
Titulcia
Carrascoy
Yecla
Rainfall: 0.79 Hh: 0.70Gs: 0.67
Ac: 0.69 Ar: 0.64 Br: 0.61 Ss: 0.67Ft: 0.77 Hc: 0.87 Hv: 0.62
Ch:
0.8
5 T
z: 0
.70
A
MDS 1
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5 2.0
MD
S 2
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
CowraQuandialla
Gibson1
Gibson2
Condobolin
Nevertire
Buronga
Florida
TruganiniEtiwanda
Ed: 0.64 Des: 0.75 Cs: 0.65Pd: 0.75 Scl: 0.66
Rai
nfal
l: 0.
59
M
e: 0
.81
Cal
: 0.8
0
Al:
0.75
B
Fig. S3 . MDS plot showing the dissimilarity distance among communities of the plots sampled along environmental gradients in Spain (A) and Australia (B). Spearman correlation coefficients >0.5 are showed in a box for each axis. Legends are: Rainfall = Climatic PCA, highly correlated with rainfall in both countries. Spain: Tz = Thymus zygis, Gs = Genista scorpius; Ch = Carex humilis; Ac = Anthyllis citisoides; Ar = Asphodelus ramosus; Br = Brachypodium retusum; Ss = Sedum sediforme; Ft = Fumana thymifolia; Hh, Hc and Hv = Helianthemu hirtum, H. cinereum and H. violaceum, respectively. Australia: Al = Atriplex leptocarpa; Me = Maireana enchylaenoides; Cal = Calotis sp.; Ed = Eragrostys dielsii; Des = Desmodium sp.; Cs = Cheilantes sieberi; Scl = Sclerolaena sp.; Pd = Panicum decompositum.

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
150
Appendix J. An alternative approach to measure changes in competitive outcomes
depending on microsite
As stated in the main text, three principal mechanisms may determine competitive outcomes
in the vegetation in Open microsites and under nurse plants: competitive exclusion,
intransitivity and niche segregation. While competitive exclusion is likely to cause low
diversity at the plot-scale, both intransitivity and niche segregation are likely to increase it.
We aimed to to test for differences in the competitive outcomes among the three microsites
tested in each country. As an alternative approach to that presented in the main text, we
developed an index based on Whittaker´s beta-diversity index (Whittaker 1972): A/(B-1),
where A is the global species richness, and B the local species richness. In our case, A will be
the species richness found under each microsite at the whole plot level, while B will be the
species richness found in each one of the quadrats sampled for each microsite (n = 30). To
calculate B, those quadrats with no species occurring on them where not considered.
The rationale behind this approach is similar to that described in the main text: 1) if
competitive exclusion dominates, we will find a reduction of B causing a decrease in A (few
dominant species will control all the space available; low B, low A = competitive exclusion);
2) if intransitivity dominates, we will find a low B but a high A because of few dominant
species will dominate at the small-scale (quadrat): however, the identity of these dominant
species will change depending on the particular environmental conditions of each patch and
the original species mixture that colonized it. This will generate a high turnover of species
that will increase the plot-scale richness (low B, high A = competition intransitivity); 3) if
niche segregation dominates, both A and B will be high. At small spatial scales, the
differential exploitation of resources (niche segregation) will enhance species coexistence,
and therefore increase the chances of several species to recruit and coexist under a given
microsite (high B, high A = niche segregation). We analyzed separately both A and B with
univariate ANCOVA models, with microsite (three levels) as fixed factor and mean plot cover
(a surrogate of productivity) as a covariate (Table S7).

CHAPTER 4 .
151
Table S8. ANCOVA results for both small-scale (B) and plot-scale (B) richness for studied region.
Variable Factor df F P-value Effect Underlying
mechanism
Sp
ain
A Cover 1,29 53.9 <0.0001 Reduction of
microsite effect Higher niche segregation under nurse plants, this difference
reduces with higher
productivity
Microsite 2,29 3.0 0.65 Nurses increase A
B Cover 1,29 8.1 0.008 Reduction of
microsite effect
Microsite 2,29 4.7 0.017 Nurses increase B
Au
stra
lia
A Cover 1,26 0.2 0.628 None Higher niche
segregation under nurse
plants, regardless of ecosystem productivity
Microsite 2,26 4.0 0.030 Nurses increase A
B Cover 1,26 2.7 0.113 None
Microsite 2,26 4.0 0.030 Nurses increase B

BIOTIC INTERACTIONS DRIVE LOCAL-SCALE RICHNESS
152

Santiago Soliveres, Rubén Torices, Fernando T. Maestre.
Manuscrito en revisión en Journal of Ecology
5
On the relative importance of climate and biotic no n-
trophic interactions as drivers of local plant spec ies
richness in semiarid communities

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
154

CHAPTER 5
155
ABSTRACT
Molecular phylogenies are being increasingly used to get new insights on the mechanisms structuring plant communities. However, the large number of factors potentially affecting the phylogenetic structure of plant communities cautions against the sole use of this information to properly infer the mechanisms shaping them. We jointly evaluated the effects of environmental conditions and biotic interactions on the phylogenetic structure of 11 semiarid Stipa tenacissima L. communities along an ample environmental gradient. We also assessed the relative importance of phylogenetic relatedness (PD) and abiotic conditions as drivers of pairwise interactions across such gradient. Habitat filtering and biotic interactions promoted a random phylogenetic structure in most of the communities studied. While positive biotic interactions increased phylogenetic evenness by niche expansion and habitat differentiation, more benign environmental conditions reduced this evenness indirectly by reducing the effects promoted by nurse plants. Phylogenetic relatedness was the primary factor affecting pairwise interactions. Values of this variable between 207-272.8 Myr led to competition, those outside this range led to neutral or positive interactions, depending on climate. Our study illustrates, for the first time, the relative importance of climate and biotic interactions on the phylogenetic structure of plant communities, and shows how the evolutionary relationships and environmental conditions interact to determine particular pairwise interactions. We also provide a comprehensive set of easy-to-measure and interpret tools for avoiding misleading interpretations when inferring mechanisms from phylogenetic structure data in observational studies.

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
156
INTRODUCTION
he recent development of
molecular phylogenies has
provided ecologists with a
powerful tool to get new insights on the
mechanisms structuring plant communities
(Webb et al. 2002). The phylogenetic
structure of a given community has been
extensively used to assess the relative
importance of environmental conditions
(environmental filtering) or competition as
drivers of community structure (reviewed
in Webb et al. 2002, Cavender-Bares et al.
2009, Vamosi et al. 2009). Biotic
interactions and environmental conditions
are known to interactively affect plant
community structure and dynamics
(Butterfied et al. 2010). However, the
relative importance of both factors as
drivers of the phylogenetic structure of
plant communities is still poorly
understood (Cavender-Bares et al. 2004,
Verdú et al. 2009). Furthermore, the
phylogenetic structure of a given
community may be strongly affected by
other factors, including herbivore or
pollinator preference for closely related
taxa (Webb et al. 2006), the scale
considered (Kraft and Ackerly 2010), or
differences in niche and competitive ability
among co-occurring species (Myfield and
Levine 2010). This complex array of
factors makes the use of phylogenetic
structure alone insufficient to properly
infer the mechanisms shaping plant
communities (Cavender-Bares et al. 2009,
Myfield and Levine 2010). Thus, more
comprehensive approaches, including the
study of environmental factors and co-
occurrence patterns, have been
recommended to further refine the
conclusions drawn from phylogenetic
methods (Pausas and Verdú 2010).
Positive interactions among plants
have been shown to largely influence the
structure and diversity of plant
communities in virtually all terrestrial
ecosystems (Callaway 2007, Brooker et al.
2008), and can even promote the expansion
of realized species niches over
evolutionary time frames (Valiente-Banuet
et al. 2006). These interactions have been
shown to depend up to a great degree on
the abiotic environment (Callaway 2007;
Maestre et al. 2009), and on the
phylogenetic distance (hereafter PD)
between the interacting species (Valiente-
Banuet et al. 2006, Castillo et al. 2010).
Overall, these studies suggest that positive
interactions are more likely to occur
among phylogenetically distant species
pairs, or under harsh environmental
conditions (e.g. Callaway 2007, Valiente-
Banuet and Verdú 2007). In the same way
that the different ontogenetic stages of
involved species interact with their PD in
defining the outcome of pairwise plant-
T

CHAPTER 5
157
plant interactions (Valiente-Banuet and
Verdú 2008, Castillo et al. 2010), both
environmental conditions and PD are likely
to jointly determine the outcome of these
interactions, and hence the structure of
plant communities. Although it is expected
that harsh environmental conditions cause
phylogenetic clustering (Webb et al. 2002),
the prevalence of pairwise positive
interactions at the community level might
lead to an even phylogenetic structure
(Valiente-Banuet and Verdú 2007).
Therefore, knowing how PD and
environmental conditions jointly affect the
outcome of pairwise species interactions
will provide additional insights to
understand the relative importances of
environmental filtering and biotic
interactions as drivers of the phylogenetic
structure of entire plant communities
(Cavender-Bares et al. 2009).
To our knowledge, no previous
study has evaluated the relative
importances and joint effects of biotic
interactions and environmental conditions
as determinants of the phylogenetic
structure of whole plant communities along
wide environmental gradients, nor the joint
effects and the relative roles of PD and
environmental conditions as drivers of
pairwise plant-plant interactions. We
aimed to do so by simultaneously
measuring the phylogenetic structure,
different components of biotic interactions
(co-occurrence patterns, niche expansion
promoted by nurse plants, and differences
in the environmental filtering among
facilitated/non-facilitated species guilds),
and a set of environmental conditions in 11
semiarid Stipa tenacissima communities
located along an environmental gradient in
Spain. Additionally, we assessed how the
abiotic conditions, the PD between the
involved species, and the interaction
between both factors modulated a large set
of pairwise interactions outcomes along
such gradient. We addressed the following
questions: How do environmental filtering
and biotic interactions jointly affect the
community phylogenetic structure across
environmental gradients?, and What is the
relative importance of PD and abiotic
conditions in defining the outcome of
particular pairwise interactions?

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
158
Figure 5.1 Conceptual model synthesizing the unifying approach employed in this study at both community and pairwise levels. Arrows are the different processes considered affecting community assemblage and its phylogenetic structure. Circular boxes represent the surrogates of each process measured in this study. The phylogenetic structure of a given community will be affected by several factors such as abiotic conditions (i.e. environmental filtering) or biotic non-trophic interactions (i.e. facilitation/competition shifts, niche expansion or double environmental filtering [phylogenetic clustering among facilitated/non-facilitated species guilds, or differences in similarity among microsites]). However, these processes do not affect independently the community assemblage and they are likely to interact in many ways. We tested for shifts in the biotic interactions across a wide environmental gradient and how they affect the phylogenetic structure of the studied community. At the pairwise level, interaction outcomes should be more positive when these conditions represent higher stress (i.e. low rainfall, cold temperatures; Callaway 2007). A different line of inquiry highlights that the phylogenetic distance (PD) between involved species as a crucial factor affecting these interactions; the outcome will be more positive when higher the PD among the species involved

CHAPTER 5
159
(Valiente-Banuet et al. 2006, Valiente-Banuet and Verdú 2007, Castillo et al. 2010). We hypothesize that the PD distance between a nurse plant and a given beneficiary would need to increase for their interaction to be positive as abiotic conditions become less stressful.
MATERIALS AND METHODS
STUDY AREA
We studied 11 Stipa tenacissima
communities along a climatic gradient
spanning from the center to the south-east
of Spain (see Table S9). Our sites have
annual precipitation and temperature
values ranging from 273 mm to 488 mm,
and from 13ºC to 17ºC, respectively. To
minimize the experimental noise produced
by environmental factors other than
climate, which could affect our
conclusions, all the sites shared the same
general soil type (Lithic Calciorthid, Soil
Survey Staff 1994), and had similar
orientation and slope values. Vegetation
was in all cases an open grassland
dominated by S. tenacissima, with total
cover values ranging from 35% to 68%.
Sparse resprouting shrubs like Quercus
coccifera, Pistacia lentiscus or Rhamnus
lycioides were also present in all sites.
VEGETATION SURVEY
At each site we established a 30 m × 30 m
plot containing the representative
vegetation of the surrounding area. This
plot size allowed the inclusion of a number
of shrub patches large enough to conduct
the survey described below. In each site,
we located four 30 m long transect
downslope for the vegetation survey, each
8 m apart across the slope. Along each
transect, we placed 20 contiguous 1.5 m ×
1.5 m quadrats, and recorded the
presence/absence of each perennial plant
species within each quadrat.
To evaluate particular pairwise
interactions for both dominant nurse types
in the study region (sprouting shrubs and S.
tenacissima tussocks, hereafter Shrub and
Stipa microsites, respectively), and to
assess the total number of facilitated
species at each site, we established a
complementary sampling design. We
randomly selected ten Stipa tussocks in
each site, and sampled the total area under
their canopy using 0.5 m × 0.5 m quadrats
(~30 quadrats per site). Ten paired open
areas (areas located at least 1 m away from
any Stipa tussock or resprouting shrub,
hereafter Open microsite), were randomly
selected adjacent to these tussocks. The
same number of 0.5 m × 0.5 m quadrats
sampled in each Stipa microsite was
sampled in each Open microsite selected,
to balance the sampling effort. Finally, the
same area was also sampled under the

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
160
canopies of five sprouting shrubs (mostly
Q. coccifera, Appendix K). The abundance
(number of individuals) of all perennial
plant species was recorded within each
sampled quadrat.
ASSESSMENT OF PHYLOGENETIC
STRUCTURE
We assembled a phylogenetic tree for the
86 species included in this study using
Phylomatic2 (Webb et al. 2008). All the
families in our dataset matched the family
names of the angiosperm megatree used in
Phylomatic (R20091110.new), which was
based on the APG III phylogenetic
classification of flowering plant orders and
families (Angiosperm Phylogeny Group
2009). Within-family phylogenetic
relationships were further resolved based
on data from various published molecular
phylogenies (Appendix K, Table S10).
After assembling the phylogenetic tree, we
adjusted its branch lengths with the help of
the Phylocom BLADJ algorithm, which
fixes the age of internal nodes based on
clade age estimates, whereas undated
internal nodes in the phylogeny are spaced
evenly (Webb et al. 2008). According to
Vamosi and Vamosi (2010), we used
TimeTree (Hedges et al. 2006) to fix as
many nodes in the tree as possible (see
Appendix K for methodological details).
This procedure resulted in the fixation of
48 nodes (representing more than 70% of
internal nodes of our tree).
Once we assembled the
phylogenetic tree for all the species
surveyed (Fig. S8), we measured two
different indicators of phylogenetic
relationships among co-occurring plants at
each of our sites: the mean phylogenetic
distance (hereafter MPD; Webb et al.
2002), and the pairwise PD among every
possible pair of co-occurring species in
each site. Since MPD is related to the
species pool, we avoided this confounding
factor by calculating its standardized effect
size (SES) with the Picante package for R
(Kembel et al. 2010), version 2.10.1 (R
Development Core Team 2009). It was
calculated as (MPDobs - MPDsim) ⁄
sdMPDsim, where MPDobs was the
observed value of MPD, and MPDsim and
sdMPDsim were the mean and standard
deviation, respectively, of this index
obtained from the 1000 simulations
performed under the null model. Positive
SES, and with a P-value > 0.95 indicate
significant phylogenetic evenness in the
sampled community, while negative SES
with P-values < 0.05 indicate phylogenetic
clustering.

CHAPTER 5
161
Figure 5.2 Graphical synthesis of the results obtained. Different surrogates of biotic interactions (C-E), environmental filtering (A) and their interactions (B) are separated. Blue arrows indicate an increase in community phylogenetic evenness due to a given mechanism, while red arrows mean an increase in phylogenetic clustering (dashed lines indicate an indirect effect). Crossed-red circles indicate no effect (in the case of arrows) or no interaction with climate (in the case of variables). MPD = mean phylogenetic distance among all possible species pairs of a given site; MPDfac = mean phylogenetic distance among pairs of the cluster of facilitated species; C-score = standardised effect size of the C-score; Similarity = Chao-Jaccard abundance-based similarity index calculated for Stipa vs. Open or Shrub vs. Open microsites; % Obligates = percentage of species found only under a nurse plant comparing to the total species richness found in each site; Climate = values of the first axis of a PCA performed with eight environmental variables. Detailed statistical results from each relationship shown in this figure are in Fig. S10.
EVALUATING PLANT-PLANT
INTERACTIONS AT THE
COMMUNITY LEVEL
The 80 1.5 m �1.5 m quadrats surveyed at
each site were used to examine the co-
occurrence pattern at the whole community
level by using the SES of the C-score
index (Gotelli et al. 2000). This metric is
commonly used to assess the outcome of
biotic interactions at the community level
(e.g. Rooney et al. 2008, Bowker et al.
2010), and was calculated as described

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
162
above for the MPD (see Appendix K for
methodological details).
To measure niche segregation or
the “double habitat filtering” (see below)
promoted by nurse plants we used two
different measurements: 1) the Chao-
Jaccard abundance-based similarity index
(hereafter CI; Chao et al. 2005), and 2) the
difference between MPD for the whole set
of species in a given community (described
above), and the MPD for only the
facilitated/non-facilitated species guilds
(MPDfac and MPDnf, respectively). We
refer here to “double environmental
filtering” to describe the effect of the
different environmental conditions found
in Open or nurse microsites, which
generate different understorey species
guilds for each microsite (i.e., higher
dissimilarity among microsites; Badano
and Cavieres 2006). These guilds could be
phylogenetically clumped within each
microsite because of their shared
adaptations to the same environmental
conditions, but can be phylogenetically
even at the whole community level
because of the different environments
existing at each site, i.e., MPD > MPDfac
or MPDnf. The CI is based on the
probability that two randomly chosen
individuals, one from each of two selected
microsites (Open, Stipa or Shrub), belong
to species shared by both microsites. A
higher dissimilarity (lower CI) among
Stipa/Shrub and Open microsites will
indicate higher influence of niche
differentiation provided by nurse canopies.
We calculated the CI by summing over the
number of individuals and species recorded
in all the 0.5 m × 0.5 m quadrats per
microsite and site (n ~ 30) using EstimateS
8.2.0 for Windows (Colwell 2000). We
also compared the MPDfac and MPDnf
with the MPD index (see rationale in
Assessing the effects of environmental
filtering and biotic interactions on
community phylogenetic structure below).
Finally, to evaluate the degree of
the realized niche expansion (sensu Bruno
et al. 2003) provided by nurse plants, we
calculated the percentage of facilitation
obligates (sensu Butterfield 2009), in
comparison to the total number of species
found in each site. Facilitation obligates
were those species with individuals
recruiting only under a nurse plant, and
therefore, only able to colonize a given site
under the microclimatic protection
provided by nurses.
EVALUATING PLANT-PLANT
INTERACTIONS AT THE PAIRWISE
LEVEL
We measured facilitation intensity and
importance, i.e., the effect that neighbours
have on their target species regardless of
other environmental factors and the
relative effect of nurses on their target

CHAPTER 5
163
species compared to that of other
environmental factors, respectively
(Brooker et al. 2005), of each possible
Stipa- and Shrub-target species pairs at
each site. For doing this we used the
Relative Interaction Intensity index (RII)
and the Interaction Importance index
(Iimp) provided by Armas et al. (2004) and
Seifan et al. (2010), respectively (see
Appendix K for details). Both indices
oscillate between -1 and 1 (relative units);
positive and negative values indicate
facilitation and competition, respectively.
The higher the index value, the higher the
intensity (RII) or importance (Iimp) of
such effect. For these analyses we used the
total number of individuals found in the
~30 0.5 m × 0.5 m quadrats sampled at
each site, and calculated a unique RII and
Iimp index for each species and site. We
used the number of recruited individuals as
an indicator or each species performance to
calculate these indices, because the number
of recruited individuals indicates superior
environmental conditions for a given
species in a given microsite, an approach
followed by previous studies (Valiente-
Banuet et al. 2006, Valiente-Banuet and
Verdú 2007).
Alternatively, the pairwise
phylogenetic distances among both nurses
(Stipa and Shrub) and their co-occurring
species in each site were calculated using
the “cophenetic” command of R. The mean
PD of facilitation beneficiaries (species
with more individuals recruiting under a
nurse plant) and obligates (described
above) at both Stipa and Shrub microsites
was calculated in each site using these data
(see Fig. 5.1).
ASSESSING THE EFFECTS OF
ENVIRONMENTAL FILTERING AND
BIOTIC INTERACTIONS ON
COMMUNITY PHYLOGENETIC
STRUCTURE
Prevalent climatic conditions (rainfall,
radiation and a temperatures) for each site
were collected using available climatic
models (Ninyerola et al. 2005) and reduced
to a single synthetic variable using PCA
conducted with the Primer v. 6 statistical
package for Windows (PRIMER-E Ltd.,
Plymouth Marine Laboratory, UK).We
used the first axis of this PCA (hereafter
Climate) for the analyses explained below.
This axis explained 88.6% of the variance
and was inversely and positively correlated
with radiation and rainfall, respectively
(Appendix K).
The relationships between the
surrogates of biotic interactions used (C-
score, percentage of facilitation obligates,
and CI) and both MPD and Climate were

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
164
evaluated by using linear regressions. This
set of analyses gives us an idea of the
relative importance of environmental
conditions as drivers of biotic interactions,
and of the effects of these interactions on
the phylogenetic structure of the
community. Additionally, the relationship
between Climate and MPD was assessed
by using both linear regression (direct
effect) and partial correlations (indirect
effect, mediated by the effect of Climate
on C-score, percentage of facilitation
obligates, CI and MPD-MPDfac/nf,
respectively). If C-score, percentage of
facilitation obligates or CI increase MPD,
this will mean that either competition,
niche expansion or niche differentiation
provided by nurse plants, respectively,
increase the phylogenetic evenness of the
community. The relation between these
biotic interactions indicators and Climate
will give us an idea on how climatic
conditions influence such interactions.
Finally, if both linear regression and partial
correlations between Climate and MPD are
significant, this will mean that climatic
conditions have a direct effect on the
phylogenetic structure of the studied
communities. However, if only linear
regressions, but not partial correlations, are
significant, this will mean that climatic
conditions are affecting MPD only via
their indirect effects on a particular biotic
interaction mechanism (depending on the
partial correlation that is non-significant).
Finally, as a second evaluation of
the degree of habitat differentiation, and
how abiotic conditions affected it, we
evaluated the effect of Climate on the
difference between MPD and both MPDfac
and MPDnf, respectively. If this difference
is significantly higher than 0, niche
differentiation is producing a phylogenetic
clustering in facilitated or non-facilitated
species guilds, and thus the difference in
microclimatic conditions between nurse
and Open microsites affect the
phylogenetic structure of this particular
community. We first compared the MPD –
MPDfac/nf difference from zero by using
t-tests, with sites acting as replicates. Then,
the relationship between Climate and this
difference was evaluated using linear
regressions to asses the effect of climatic
conditions on this “double habitat filtering”
(Jones et al. 1997, Badano and Cavieres
2006; Fig. 5.1)
ASSESSING THE EFFECTS OF
CLIMATIC CONDITIONS AND PD ON
THE OUTCOME OF PAIRWISE
PLANT-PLANT INTERACTIONS
To evaluate the existence of an interaction
between PD and abiotic conditions, we
assessed the relationships between Climate
and the MPD obtained from all the nurse-
facilitated species pairs in each site by

CHAPTER 5
165
using linear regressions. These species
pairs were divided according to the type of
facilitative relationship (beneficiaries or
obligates) and nurse plant (Stipa or
shrubs). We also fitted the relationships
between Climate, PD, RII and Iimp by
using Regression trees (De´ath and
Fabricius 2000), as implemented in the
Tree package of R. We used 10-fold cross-
validation to fit the most parsimonious
model to each dataset (De´ath and
Fabricius 2000). Climate and the PD
between each target species and its nurse
were used as predictor variables in the four
regression trees performed (RII and Iimp
indices for Stipa and shrub nurses). Despite
this low number of predictors, we used this
procedure because of the heavily skewed
nature of the PD values (Fig. S5), a
characteristic commonly found with these
data (Castillo et al. 2010). Regression trees
also allow detecting non-linear
relationships, and are insensible to the
distribution of either the predictor or
response variables (De´ath and Fabricius
2000).
RESULTS
EFFECTS OF ENVIRONMENTAL
FILTERING AND BIOTIC
INTERACTIONS ON COMMUNITY
PHYLOGENETIC STRUCTURE
Most of the studied sites showed a random
phylogenetic structure (Table S9). Climate
was negatively related to MPD at the
community level, but this relationship was
only marginally significant (r = -0.53; P =
0.09). When we removed the indirect
effect mediated by the relationship
between Climate and the MPD – MPDfac
difference by using partial correlations,
this relationship disappeared (ρ = 0.32; P =
0.4), suggesting the lack of a direct effect
of Climate on MPD. In contrast, the
different measures of plant-plant
interactions were all positively, but
marginally, related to MPD at the
community level (r = 0.60, 0.56 and 0.57;
P = 0.07, 0.09 and 0.07 for C-score SES,
CI between Open and Shrub microsites and
percentage of facilitation obligates,
respectively; Fig. 5.2, Fig. S6). The
relationship between CI (Open vs. Stipa
microsites) and MPD was not significant (r
= - 0.11; P = 0.44). Most of the surrogates
of biotic interactions employed were not
related to Climate (P > 0.35 in all cases;
Fig. 5.2). However, the negative
relationship between the MPD-MPDfac
difference and Climate found (r = -0.82; P
= 0.006) suggests a decrease in
phylogenetic clustering among facilitated
species with increased rainfall availability.
This difference was also significantly

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
166
-200 -150 -100 -50 0 50 100
200
300
400
500
600
700
PCA climate
-200 -150 -100 -50 0 50 100
Mea
n P
D fr
om fa
cilit
atio
n be
nefic
iarie
s
200
250
300
350
400
StipaShrub
Total mean PD for Stipa
Total mean PD for Shrub
Mea
n P
D fr
om fa
cilit
atio
n ob
ligat
es
Total mean PD for Stipa
Total mean PD for Shrub
A
B
- RAINFALL +
Figure 5.3 Relationship between the PCA axis obtained from climatic values (PCA climate) and mean phylogenetic distance (PD) between facilitation beneficiaries (A) or obligates (B) species and their nurses. No relationships were found for any of the assayed variables and climate (R2 < 0.1 and P > 0.3 in all the cases).

CHAPTER 5
167
Figure 5.4 Regression trees conducted with both interaction intensity (RII; panel A) and importance (Iimp; panel B) indices. Split values for each predictor used (PD, in million years [Myr], or climatic PCA axis values) are shown in each branch. Terminal nodes show the mean value for each group of the response variable introduced and the number of cases in each node (between parenthesis; n = 200 cases for each tree). Positive and negative values indicate facilitative and competitive interactions, respectively. The higher the values, the more intense or important the effect of the nurse upon the target species. In each panel, the general fit of the model (D2, percentage of variance explained by the model), extracted from the null deviance (Deviance root), and the deviance of the final chosen tree after 10-fold cross-validation (Deviance tree) are shown.

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
168
different from zero, but the MPD-MPDnf
difference was not (mean difference = 1.09
± 0.15 and 0.69 ± 0.20; t = 2.36 and 1.15;
P = 0.046 and 0.284 for facilitated and
non-facilitated species, respectively).
These results suggest a phylogenetic
clustering, comparing with the general
species pool of each site, for the facilitated
but not for the non-facilitated species.
EFFECTS OF CLIMATIC CONDITIONS
AND PHYLOGENETIC DISTANCE ON
THE OUTCOME OF PAIRWISE
INTERACTIONS
The PD between nurse-facilitated species
remained constant across the entire
environmental gradient sampled (Fig. 5.3).
Climate and PD were poor predictors for
the RII and Iimp data calculated with the
target species tested and Stipa as nurse
plant, respectively (Regression trees D2 =
0.02 and 0 for RII and Iimp, respectively).
Conversely, regression trees predicted 25%
and 14% of the variance of the RII and
Iimp indices for Shrub microsites and their
target species (Fig. 5.4). Values of PD
between the shrubs and their target species
between 207 and 272.8 million years (Myr)
rendered negative or neutral interactions,
while values of PD outside this values
(<207 Myr or >272.8 Myr) indicated
positive results for both RII and Iimp
indices. Climate was a modulator of
secondary importance for these pairwise
interactions. When 207 Myr < PD < 272.8
Myr, values of Climate higher than 47.7
(the wettest sites) render negative RII
values, which were neutral otherwise.
When 207 Myr < PD > 272.8 Myr, RII
values were positive in the dryer sites
(Climate values < 127.2), but neutral in the
rest of climatic conditions (Fig. 5.4A).
Values of the Iimp index rendered slightly
different results; when PD > 272.8 Myr
and Climate values < -67 (dryer sites),
shrubs were not important for the
performance of their target species; when
Climate values where higher than -67 (mid
to wet sites), shrubs exerted a negative
effect upon their target species (Fig. 5.4B).
DISCUSSION
By jointly considering information on
abiotic conditions, state-of-the-art
phylogenetic tools, and different aspects of
biotic non-trophic interactions
(competition/facilitation shifts, niche
expansion and niche differentiation), we
were able to explore the relative
importances of plant-plant interactions and
the environment as drivers of the
phylogenetic structure of the studied
communities. Our results show that these
interactions are important to determine
phylogenetic structure along a wide
environmental gradient, but that the effect
of climatic conditions on this structure is

CHAPTER 5
169
indirectly mediated by its effects upon the
“double environmental filtering” provided
by the difference in microclimatic
conditions between nurse plants and Open
microsites. The phylogenetic evenness
promoted by biotic interactions and the
phylogenetic clustering promoted by
climatic conditions, mediated by its effects
on plant-plant interactions, caused a
random phylogenetic structure in most of
the studied communities. At the pairwise
level, we found that PD was a primary
modulator of plant-plant interactions, with
Climate playing a secondary role. PD
values between 207-278.2 Myr rendered
negative outcomes, while PD values
outside this range yielded positive or
neutral outcomes, depending on
environmental conditions.
DIRECT AND INDIRECT EFFECTS OF
ENVIRONMENTAL FILTERING AND
BIOTIC INTERACTIONS ON THE
COMMUNITY PHYLOGENETIC
STRUCTURE
In sharp contrast with previous studies
(Michalet et al. 2006, Callaway 2007), we
did not find any relationship between our
surrogates of biotic interactions or niche
expansion at the community level (C-score
and percentage of facilitation obligates
species, respectively) and the climatic
variables measured. This does not mean
that plant-plant interactions were not
important at this level of organization, but
rather that these interactions, mainly
positive ones, importantly influenced
community assemblage through niche
expansion and habitat differentiation
across the entire environmental gradient
studied (Fig. 5.2; Fig. S6). How does the
equal importance of biotic interactions
across wide environmental gradients affect
the relative roles of such interactions and
environmental filtering in determining the
phylogenetic structure of plant
communities?
When evaluating the effects of
climatic conditions or biotic interactions on
the phylogenetic structure of the studied
communities separately, we found support
for the patterns found by previous studies:
while competition increased the
phylogenetic evenness of a given
community, environmental filtering did the
contrary (Webb et al. 2002, Cavender-
Bares et al. 2004). However, when
considering both processes and their
interaction together, our results showed
that phylogenetic structure is determined
by both direct and indirect effects mediated
by climatic conditions and biotic non-
trophic interactions (Fig. 5.2), which
finally caused a random phylogenetic

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
170
pattern. To our knowledge, such
interaction has not been described before.
These results are in the line of the most
recent literature, which suggests that the
phylogenetic structure alone, even with a
perfect knowledge of the trait conservatism
of co-occurring species, may not be
enough to infer the implied mechanisms in
the assemblage of a given community
(Pausas and Verdú 2010). Our study
demonstrates that this is particularly true
when dealing with more than one
important mechanism shaping natural
communities, something fairly common in
nature (Butterfield et al. 2010). In contrast
with previous studies (e.g. Valiente-Banuet
and Verdú 2007, Verdú et al. 2009), we did
not find any increase in phylogenetic
evenness with the increase of facilitation
importance, as shown by the C-score
measurements at the community level.
This could be explained because of
contrasting ontogenetic stages comparing
to those studies (more adult-adult
interactions in our sampled sites than in the
others; Valiente-Banuet and Verdú 2008),
or because of some of the patchiness
registered with the C-score may be more
related to a shared dispersion syndrome
than to facilitation processes, and therefore
this could lead to a phylogenetic clustering
(see additional discussion below).
However, the increase of the relative
importance of niche expansion (sensu
Bruno et al. 2003), another important
surrogate of facilitation at the community
level, promoted phylogenetic evenness, in
agreement with previous findings
(Valiente-Banuet and Verdú 2007, Verdú
et al. 2009). This latter result highlights the
necessity of measuring different indicators
of plant-plant interactions, and not only co-
occurrence patterns, to correctly assess the
role of such interactions on the
phylogenetic structure of plant
communities.
We were able to establish how an
increase in rainfall counter-intuitively
acted as an environmental filtering,
increasing community phylogenetic
clustering, and how its effect was mediated
by the reduction in the habitat
differentiation provided by nurse plants
(more rainfall reduced the difference
between MPD and MPDfac; Fig. 5.2). Our
results also highlight the importance of
considering not only multiple processes,
but also employing different proxies to
identify their direct and indirect effects on
community assemblage and its
phylogenetic structure. Using only co-
occurrence patterns, as previously
suggested (Pausas and Verdú 2010), would
not suffice to detect these indirect effects.
To take into account the whole set of
processes affecting phylogenetic structure
may also help to disentangle the
importance of phylogenetic diversity per se

CHAPTER 5
171
on ecosystem functioning or other
important ecosystem services (e.g. Forest
et al. 2007, Maherali and Klironomos
2007, Cavender-Bares et al. 2009),
avoiding potential confounding effects
produced by different processes causing a
particular phylogenetic pattern (Mayfield
and Levine 2010).
IS THE EVOLUTIONARY
RELATIONSHIP MORE IMPORTANT
THAN ABIOTIC CONDITIONS
DEFINING PAIRWISE INTERACTIONS
OUTCOMES?
A hierarchy between both PD and climatic
conditions modulated the outcome of the
large set of pairwise interactions tested. In
our case, PD was the primary factor
affecting such interactions, while climatic
conditions played a secondary role (Fig.
5.4). We found a double threshold in PD
values that defined the sign of the
interactions, with PD values between 207
and 272.8 Myr always rendering
competition, and values outside these
thresholds always leading to facilitation.
While results from the upper threshold (PD
> 272.8 Myr leading to facilitation) agree
with current literature (Valiente-Banuet
and Verdú 2007, 2008, Castillo et al.
2010), those from the lower threshold
(facilitation with PD < 207 Myr) do not.
The idea that a threshold in the PD
between the involved species may define
the outcome of their interaction is
appealing, and fits surprisingly well –
although the authors did not discuss the
data in that way– results from a recent
study in the Mexican scrubland (Castillo et
al. 2010). These authors found that lower
PDs always indicate negative interactions,
while higher PDs could mean either
positive or negative outcomes (see Figs 2
and 3 in Castillo et al. 2010); in our case,
this threshold would be PD < 272.8 Myr.
Conversely, the lower PD threshold could
be related not with a facilitatory effect of
the nurse shrub, but with the role of the
shrubs studied as a refuge for animals and
their perch effect (Pausas et al. 2006). This
could promote an increase in the
deposition of seeds by animals of other
Tertiary and animal-dispersed shrubs, and
therefore foster the co-occurrence of
species phylogenetically related with the
studied shrubs by nucleation processes
(Herrera 1992, Verdú and García-Fayos
1996). The fact that these results may be
related mostly to a dispersion syndrome,
rather than to the facilitatory effect of the
shrubs themselves, could be a potential
explanation of the contrasting results found
in other studies regarding this low
threshold in PD values (Castillo et al.

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
172
2010), and regarding the effect of
facilitation in the phylogenetic structure
(we found an increase in phylogenetic
clustering with lower SES C-scores, but
Valiente-Banuet and Verdú [2007] or
Verdú et al. [2009] found the contrary
using other measurements of co-
occurrence patterns). What factors, then,
determine the outcome of pairwise
interactions when PD is high enough to
allow positive interactions to occur? Our
results show that environmental conditions
play a major role once this threshold has
been reached in some cases, with dryer
sites usually yielding more positive
outcomes (Callaway 2007). However, we
wish to highlight that only 25% of the
variance of the tested interaction outcomes
were predicted by our regression trees.
Therefore, other unmeasured factors, such
as different ontogenetic stages of the
involved plants (Valiente-Banuet and
Verdú 2008), differences in herbivory
pressure among sites (Smit et al. 2009), or
ecophysiological traits labile through
evolutionary time frames and therefore not
detected with our phylogenetic approach
(Cavender-Bares et al. 2004, 2009), could
be important factors affecting such
interactions (e.g. Liancourt et al. 2005).
Such unmeasured factors could play a
major role in defining the outcomes of the
interaction between Stipa and its target
species (e.g. Soliveres et al. 2010;
Soliveres et al. in press), since nor PD
neither climatic conditions were good
predictors of such outcomes.
CONCLUDING REMARKS
Inferring the mechanisms shaping plant
communities from their phylogenetic
structure alone may drive to misleading
conclusions (Myfield and Levine 2010).
The use of manipulative experiments
including different communities under
contrasting environmental conditions, and
the measurement of the direct and indirect
effects of biotic interactions have been
recommended to overcome these
limitations (Cavender-Bares et al. 2009,
Vamosi et al. 2009). However, such
experiments are often logistically
prohibitive. The observational and
analytical approach employed here may
serve as an alternative to experimentation,
and can help to avoid misleading
conclusions when inferring the several
possible mechanisms underlying the
assemblage of natural communities. Our
study illustrates the complexity of direct
and indirect effects of environmental
filtering and biotic interactions as drivers
of the phylogenetic structure of natural
communities across environmental
gradients. It also highlights the necessity of
taking into account environmental
conditions and different components of the
biotic non-trophic interactions when

CHAPTER 5
173
studying the processes shaping plant
communities and their phylogenetic
structure.
This study also represents, to our
knowledge, the first attempt to test the
relative importance and possible
interactions between the evolutionary
relationships of two interacting species and
climate as drivers of the outcome of their
interactions. Interactions between different
factors affecting plant-plant interactions
may lead to counterintuitive or antagonistic
responses and should be studied jointly to
properly infer their relative importances as
drivers of such interactions (Baumeister
and Callaway 2006, Soliveres et al. in
press). Regression trees revealed as a
useful tool for detecting the hierarchy and
the non-linear responses in the effect of
climate and PD on such interactions. We
found that PD was of primary importance
acting as a threshold, which could explain
the species-specific nature often found in
plant-plant interactions (Callaway 2007).
Conversely, climatic conditions affected
these interactions only once PD reached
this particular threshold. In contrast, other
unmeasured factors in the case of Stipa,
such as herbivory, ontogenetic stage or
specific ecophysiological traits not
included in the PD relationships seemed
more important drivers of such outcomes,
illustrating the difficulties of predicting
pairwise interactions outcomes with simple
models involving just one or few
predictors.
ACKNOWLEDGMENTS Estrella Pastor, Beatriz Amat, Luis Cayuela, María D. Puche, Matt Bowker, and Pablo García-Palacios assisted with fieldwork and/or statistical analyses. Adrián Escudero provided help with species identification. SS was supported by a PhD fellowship from the EXPERTAL project, funded by Fundación Biodiversidad and CINTRA S.A. FTM acknowledges support from the European Research Council under the European Community's Seventh Framework Programme (FP7/2007-2013)/ERC Grant agreement n° 242658. This research was funded by the CEFEMED, and INTERCAMBIO (BIOCON 06/105) projects, funded by the Universidad Rey Juan Carlos-Comunidad de Madrid, and Fundación BBVA, respectively.

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
174
Supplementary material for Chapter 5.
Appendix K. Detailed materials and methods.
SUITABILITY OF STIPA TENACISSIMA STEPPES
Stipa tenacissima steppes are well suited ecosystems for testing the relative roles of biotic
interactions and habitat filtering in the phylogenetic structure of plant communities, and to
assess the relative importances and possible interactions between PD and climate as drivers of
plant-plant interactions outcomes for different reasons. Important ecophysiological traits are
extremely well-conserved through evolutionary time in the Mediterranean flora (Herrera
1992; Valiente-Banuet et al. 2006), which avoid potential confounding factors in the
interpretation of a given phylogenetic structure (Webb et al. 2002). Conversely, the PD
between two interacting species is a good indicator of dissimilarity in their ecological niches
and traits (Valiente-Banuet et al. 2006, Valiente-Banuet and Verdú 2007). Furthermore, S.
tenacissima steppes are one of the most extended Mediterranean semiarid community types
(Le Houreu 2001), and are strongly shaped by facilitation and its interaction with abiotic
stress (e.g., Maestre and Cortina 2004a, Armas and Pugnaire 2005). See Maestre et al.
(2009b) for additional details on the natural history of these communities.
SYNTHESIZING CLIMATIC CONDITIONS WITH PCA
Eight climatic variables (annual radiation, minimum, maximum and mean temperature,
annual rainfall, temperature range [maximum-minimum], and minimum and maximum
temperatures for the coldest and warmest month, respectively) were collected for each site
using available climatic models (Ninyerola et al. 2005). We reduced them to a single synthetic
variable using PCA to obtain a more general assessment of the influence of all of our
environmental variables at both the community and pair of species levels. We used the first
axis of this PCA (referred in the main text to as Climate) as our surrogate of the climatic
gradient present at our sites. This axis explained 88.6% (Eigenvalue = 8.08·103) of the
variance in the climatic data, and was highly correlated with both rainfall and radiation
(Eigenvectors = -0.864 and 0.502 for rainfall and radiation, respectively; the remainder of the
eigenvectors were < 0.03 in all cases). PCA was carried out using the Primer v. 6 statistical
package for Windows (PRIMER-E Ltd., Plymouth Marine Laboratory, UK).

CHAPTER 5
175
METHODOLOGICAL DETAILS OF THE PHYLOGENETIC HYPOTHESIS
The phylogenetic distances between species pairs were estimated by assembling a
phylogenetic hypothesis for all the species included in this study using Phylomatic2 (avalaible
at: http://www.phylodiversity.net/phylomatic/phylomatic.html). All the families in our data
set matched the family names of the angiosperm megatree used in Phylomatic
(R20091110.new), which was based on the Angiosperm Phylogeny Group III phylogenetic
classification of flowering plant orders and families (APG 2009). Within-family phylogenetic
relationships were further resolved based on data from various published molecular
phylogenies (Asteraceae: Funk et al. 2005; Susanna et al. 2006; Cistaceae: Guzmán and
Vargas 2009; Guzmán et al. 2009; Fabaceae: Allan and Porter 2000, Allan et al. 2004,
Wojciechowski et al. 2004; Poaceae: Bouchenak-Khelladi et al. 2008, 2010; Rubiaceae:
Bremer and Eriksson 2009).
Once we had assembled the phylogenetic hypothesis, we adjusted its branch lengths
with the help of the Phylocom BLADJ algorithm (Webb et al. 2008), which fixes the age of
internal nodes based on clade age estimates, whereas undated internal nodes in the phylogeny
are spaced evenly between dated nodes to minimize tree-wide variance in branch length
(Webb et al. 2008). Thus, BLADJ is a simple tool that fixes the root node of a phylogeny at a
specified age and fixes the other nodes for which age estimates are available. It sets all other
branch lengths by placing the nodes evenly between dated nodes, as well as between dated
nodes and terminals (of Age 0). The Phylocom manual (Webb et al. 2008) suggests using the
age estimates from Wikström et al. (2001); however, new analyses estimating divergence
times for angiosperms have been published since the publication of this seminal work (e.g.
Bremer et al. 2004, Anderson et al. 2005, Magallón and Castillo 2009, Smith et al. 2010,
Wang et al. 2010). In addition, TimeTree (Hedges et al. 2006), a public knowledge-base of
divergence times among organism is publicly available online (http://www.timetree.net). This
utility allows exploration of the thousands of divergence times among organisms in the
published literature (Hedges et al. 2006). A tree-based (hierarchical) system is used to identify
all published molecular time estimates bearing on the divergence of two chosen taxa, such as
species, compute summary statistics, and present the results. We mainly used this database to
fix the ages of internal nodes on our phylogenetic hypothesis, completing TimeTree results
with other published sources when this database did not provide any date (Cistaceae: Guzmán
and Vargas 2009, Guzmán et al. 2009; Asteraceae: Kim et al. 2005, Torices 2010; Poaceae:

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
176
Bouchenak-Khelladi et al. 2010; Fabaceae: Lavin et al. 2005, Bello et al. 2009; Brassicaceae:
Franzke et al. 2009; Caryophyllaceae: Valente et al. 2010). Eventually, we fixed the age of 48
internal nodes (see Table S2).
Apart from the data showed in the text, we used this tree to calculate the mean nearest
phylogenetic taxon index (MNTD; Webb et al. 2002), which measures the phylogenetic
distance to the most closely related neighbor. This index was calculated using the Picante
package for R (Kembel et al. 2010). Since MNTD was highly correlated with MPD
(Pearson´s correlation: r = 0.87; P < 0.0001), and its results were very similar to those
obtained with the MPD index, we only used results from the latter (presented in Table S1).
MEASURING CO-OCCURRENCE PATTERNS AND PLANT-PLANT INTERACTIONS
OUTCOMES
We measured co-occurrence patterns in each community by using null models based on
patterns of species co-occurrence found with the 80 1.5 m × 1.5 m quadrats (Gotelli and
Graves 1996). We estimated species co-occurrence with the C-score index, a metric
commonly used by studies aiming to infer species interactions at the community level from
co-occurrence data (e.g. Rooney et al. 2008, Maestre et al. 2008, Bowker et al. 2010). This
index is calculated for each pair of species as (Ri - S)(Rj - S), where Ri and Rj are the number
of total occurrences for species i and j, and S is the number of quadrats in which both species
occur. This score is then averaged over all possible pairs of species in the matrix (Gotelli
2000). The C-score is related to the competitive exclusion concept of “checkerboardness”
i.e., how many of the possible species pairs in a given community never appear in the same
quadrat together. Thus, positive and large values of this index indicate that competition may
be a prevalent mechanism determining the co-occurrence patterns observed (Gotelli 2000).
As the values of the C-score are dependent on the number of species and co-
occurrences observed within each plot, we obtained a standardized effect size (SES) as (Iobs -
Isim) ⁄ Ssim, where Iobs is the observed value of the C-score, and Isim and Ssim are the mean and
standard deviation, respectively, of this index obtained from the n simulations performed
(Gotelli and Entsminger 2006). Standardized Effect Size (SES) values of the C-score less than
or greater than zero indicate prevailing spatial segregation (competition prevalence) and
aggregation (facilitation dominance; Tirado and Pugnaire 2005) among the species within a
community, respectively. We used ‘fixed rows–equiprobable columns’ null models and 5000
simulations. With this approach, each species conserved its own abundance (rare species
remained rare and common species remained common) and each quadrat was assumed to

CHAPTER 5
177
have the same probability of being colonized as the remainder, regardless of the number of
species found in each quadrat during the simulations. This null model has been recommended
for standardized samples collected in homogenous habitats (Gotelli 2000), such as the ones
gathered in this study. We also tried the “fixed rows-fixed columns” algorithm (both species
and quadrats conserved its relative abundance and richness, respectively) to add confidence to
our conclusions. The results obtained with these analyses were similar than those obtained
with the ‘fixed rows–equiprobable columns’ null model, and thus are not shown.
We also calculated interaction intensity and importance indices (RII; Armas et al.
2004 and Iimp; Seifan et al. 2010, respectively). RII indices were calculated as (PNurse –
POpen)/(PNurse + POpen), where PNurse was the number of individuals under the canopy of a nurse
plant (Stipa or Shrub) and POpen was the number or individuals recruited in the Open
microsite. Alternatively, Iimp indices were calculated as Iimp= Nimp/│Nimp│+│Eimp│, where
Nimp and Eimp were the nurse plant and environmental contributions to the total number of
individuals recruited for each species, respectively. Nimp was calculated as PNurse – POpen, and
Eimp as POpen – MPOpen/Nurse, where MPOpen/Nurse is the maximum number of recruited
individuals for a given species found in the entire gradient, irrespective of the microsite
sampled.

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
178
Table S9. Main characteristics of the study sites. MPD = Mean phylogenetic distance and SES = Standardized effect size. The SES of the MPD was calculated as MPDobs – meanMPDnull/ sdMPDnull; where MPDobs is the observed phylogenetic distance in our sample, and meanMPDnull and sdMPDnull are the mean and standard deviation of the phylogenetic distances obtained from the n simulated communities under the null model. Positive SES, and with a P-value > 0.95 indicate significant phylogenetic evenness in the sampled community, while negative SES with P-values < 0.05 indicate phylogenetic clustering.* Indicates a significant phylogenetic clustering in this community. The dominant shrub species is Quercus coccifera in all sites excepting at Carrascoy, where it was Rhamnus lycioides.
Site
Latitude
Longitude
Annual rainfall
(mm)
Species richness
MPD (SES) P (MPD)
Barrax 39º02’91’’N 2º13’82’’W 433 24 -0.88 0.22
Camporreal 40º19’72’’N 3º25’36’’W 457 9 -0.77 0.16
Carrascoy 37º48’02’’N 1º18’32’’W 282 38 -0.09 0.42
Crevillente 38º14’15’’N 0º55’49’’W 273 36 1.60 0.91
El Ventós 38º28’14’’N 0º37’03’’W 319 35 0.37 0.65
Morata 40º27’62’’N 3º05’31’’W 455 22 -1.01 0.15
Sierra Espuña 37º49’27’’N 1º40’41’’W 364 32 -1.49 0.04*
Titulcia 40º11’28’’N 3º30’13’’W 440 21 -0.84 0.24
Villarrobledo 39º12’64’’N 2º30’77’’W 446 18 -0.83 0.18
Yecla 38º35’40’’N 1º12’15’’W 350 47 0.52 0.74
Zorita 40º21’30’’N 2º52’62’’W 434 38 1.52 0.91

CHAPTER 5
179
Table S10. Estimated age of branching events.
Node
label
Estimated
time (Myr) Reference
1 334.5 Hedges et al. (2006) 2 298.0 Hedges et al. (2006) 3 150.1 Hedges et al. (2006) 4 147.8 Hedges et al. (2006) 5 122.0 Hedges et al. (2006) 6 101.0 Hedges et al. (2006) 7 67.0 Hedges et al. (2006) 8 112.0 Hedges et al. (2006) 9 57.0 Bouchenak-Khelladi et al. (2010) 10 44.7 Bouchenak-Khelladi et al. (2010) 11 28.0 Bouchenak-Khelladi et al. (2010) 12 125.0 Hedges et al. (2006) 13 121.0 Hedges et al. (2006) 14 109.0 Hedges et al. (2006) 15 98.0 Hedges et al. (2006) 16 69.0 Hedges et al. (2006) 17 89.0 Hedges et al. (2006) 18 94.0 Hedges et al. (2006) 19 84.0 Bello et al. (2009) 20 55.0 Lavin et al. (2005) 21 24.6 Lavin et al. (2005) 22 97.5 Hedges et al. (2006) 23 84.8 Hedges et al. (2006) 24 39.0 Wikström et al. (2001) 25 19.0 Franzke et al. (2009) 26 51.0 Hedges et al. (2006) 27 14.5 Guzmán and Vargas (2009); Guzmán et al. (2009) 28 2.0 Guzmán and Vargas (2009); Guzmán et al. (2009) 29 1.6 Guzmán and Vargas (2009); Guzmán et al. (2009) 30 6.1 Guzmán and Vargas (2009); Guzmán et al. (2009) 31 122.0 Hedges et al. (2006) 32 122.0 Hedges et al. (2006) 33 55.8 Valente et al. (2010) 34 16.6 Valente et al. (2010) 35 73.5 Wikström et al. (2001) 36 113.0 Magallón and Castillo (2009); Anderson et al. (2005); Janssens et al. 2009) 37 111.9 Magallón and Castillo (2009); Anderson et al. (2005); Janssens et al. 2009) 38 96.6 Magallón and Castillo (2009); Janssens et al. (2009) 39 47.0 Kim et al. (2005) 40 35.5 Kim et al. (2005) 41 24.7 Torices (2010) 42 108.0 Hedges et al. (2006) 43 17.0 Bremer and Eriksson (2009) 44 106.0 Hedges et al. (2006) 45 90.0 Hedges et al. (2006) 46 76.0 Hedges et al. (2006) 47 23.0 Wikström et al. (2001) 48 8.5 Stevens (2010)

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
180
Figure S4. Phylogenetic tree of the regional species pool. Phylogenetic hypothesis are developed in Appendix S1. Main nurse plants are highlighted in blue. Node labels are given in Table S9.

CHAPTER 5
181
0 100 200 300 400 500 600 700 800
num
ber
of c
ases
0
20
40
60
80
100
120
140
160
180
Phylogenetic distance nurse-target species
100 200 300 400 500 600 700 800
num
ber
of c
ases
0
20
40
60
80
100
120
A
B
Figure S5. Frequency histogram of the phylogenetic distances between the different target plants and their nurses, either Stipa tenacissima (A) or shrubs (B). Notice the discontinuous distribution and the skewness of the data.

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
182
ENVIRONMENTAL CONDITIONS
Climatic PCA axis values
-200 -150 -100 -50 0 50 100
MP
D (
SE
S)
-2.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
R2= 0.30; P = 0.09
- RAINFALL +
Phylogenetic evenness
Phylogenetic clustering

CHAPTER 5
183
HABITAT × BIOTIC INTERACTIONS
-200 -150 -100 -50 0 50 100
C-s
core
-3
-2
-1
0
1
2
3
R2 = 0.13; P = 0.57
Climatic PCA axis values
-200 -150 -100 -50 0 50 100
% fa
cilit
atio
n ob
ligat
es
5
10
15
20
25
30
35
40
R2 = 0.12; P = 0.30
-200 -150 -100 -50 0 50 100
Sim
ilarit
y in
dex
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Open vs. Stipa: R2 = 0.05; P = 0.54 Open vs. Shrub: R2 = 0.01; P = 0.80
Climatic PCA axis values
-200 -150 -100 -50 0 50 100
MP
Dge
n -
MP
Dcl
ust
-2
-1
0
1
2
3
4
MPDfacilitated: R2 = 0.68; P = 0.006MPDnon-facilitated: R2 = 0.05; P = 0.56
Phylogenetic clusteringamong facilitated/non-facilitated species
Climatic PCA axis values
- RAINFALL +

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
184
BIOTIC INTERACTIONS
C-score-3 -2 -1 0 1 2 3
MP
D
-2.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
R2 = 0.36; P = 0.07
Similarity index0.4 0.5 0.6 0.7 0.8 0.9 1.0
-2.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
Open vs. Stipa: R2 = 0.01; P = 0.75Open vs. Shrub: R2 = 0.30; P = 0.09
% facilitation obligates
10 20 30 40
MP
D
-2.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
R2 = 0.01; P = 0.77
MPDgen - MPDclust
-2 -1 0 1 2 3 4
Sim
ilarit
y in
dex
0.4
0.5
0.6
0.7
0.8
0.9
1.0
MPDfac vs Open-Stipa similarity MPDfac vs Open-Shrub similarity MPDnf vs Open-Stipa similarity MPDnf vs Open-Shrub similarity
Figure S6. Detailed results of the conceptual diagram shown in Figure 5.2. Results are organized following sections provided in Figure 5.2 (effects of environmental conditions, biotic interactions and habitat × biotic interactions). In all cases, linear regressions results are shown in each plot.

CHAPTER 5
185

BIOTIC AND ABIOTIC FILTERS FOR PHYLODIVERSITY
186

DISCUSIÓN Y CONCLUSIONES GENERALES

DISCUSIÓN GENERAL
188

189
DISCUSIÓN GENERAL
Los trabajos presentados en esta tesis doctoral constituyen una de las evaluaciones más
exhaustivas sobre la evolución de las interacciones planta-planta a lo largo de gradientes
ambientales realizadas hasta la fecha. Por un lado, se ha testado la evolución de la interacción
entre Stipa tenacissima y Retama sphaerocarpa a lo largo de cuatro puntos contrastados de
disponibilidad hídrica, derivados de predicciones realistas sobre futuros escenarios de cambio
climático (IPCC 2007, capítulo 1). Por otro lado, se ha testado el efecto de la co-ocurrencia de
la aridez con distintos factores de importancia en la determinación del signo y la intensidad de
las interacciones planta-planta, como son la herbivoría (p. ej. Baraza et al. 2006), la ontogenia
(p. ej. Miriti 2006) o la relación evolutiva entre las especies implicadas (p. ej. Castillo et al.
2010). Estas aproximaciones han revelado complejas interacciones y jerarquías existentes
entre todos estos factores (capítulos 2, 3 y 5). Los experimentos realizados a nivel de
comunidad han permitido evaluar la generalidad de los distintos modelos propuestos sobre la
evolución de las interacciones planta-planta a lo largo de gradientes ambientales (p. ej.
Bertness y Callaway 1994, Michalet et al. 2006, Maestre et al. 2009a), tanto a nivel de par de
especies como al de comunidades vegetales enteras (capítulo 4).
Los resultados obtenidos apuntan a que –por orden de importancia– la herbivoría, las
características ecológicas de las especies implicadas (deducidas tanto a partir de la tolerancia
a distintos factores de estrés o fases ontogenéticas como de su historia evolutiva) y, por
último, las condiciones climáticas y sus interacciones con los factores anteriormente
mencionados, son los factores clave que definen el signo y la intensidad de las interacciones
entre pares de especies vegetales. A pesar de que la mayoría de estudios sobre la dinámica de
las interacciones planta-planta en medios semiáridos se centran en la disponibilidad hídrica
como factor abiótico clave (p. ej. Holzapfel y Mahall 1999, Tielbörger y Kadmon 2000,
Pugnaire y Luque 2001, Maestre y Cortina 2004a), los trabajos presentados en esta tesis
apuntan a la disponibilidad de luz y a la intolerancia a la sombra de las especies beneficiarias,
o de sus distintas fases ontogenéticas, como un factor de gran importancia a la hora de definir
dichas interacciones y su relación con la disponibilidad hídrica (Valladares y Pearcy 2002,
Prider y Facelli 2004, Valladares et al. 2008, Seifan et al. 2010). Por otro lado, se ha
demostrado que el patrón temporal, y no sólo la cantidad, de lluvia es algo importante a
considerar si queremos entender la evolución de estas interacciones a lo largo de gradientes

DISCUSIÓN GENERAL
190
ambientales, o su respuesta frente a futuros escenarios climáticos (Zavaleta 2006, Knapp et al.
2008). Los resultados obtenidos apuntan a que, mientras que el aumento de aridez puede
conllevar un aumento de la competencia entre las especies estudiadas, el incremento de los
eventos de lluvia torrencial puede acelerar la segregación de nicho entre las especies
implicadas, reduciendo este efecto competitivo. Los efectos de ambos patrones (aumento de
aridez y eventos torrenciales) dependerán de las tolerancias relativas a la sombra y a la sequía
de la especie beneficiaria, y de la posibilidad de que se produzca una segregación de nicho
efectiva entre la planta nodriza y su beneficiaria (Holmgren et al. 1997, Knapp et al. 2008,
capítulos 1–3).
Los múltiples factores que afectan de forma conjunta a las interacciones planta-planta
(herbivoría, aridez y diferentes características ecológicas), que comúnmente coexisten en los
ecosistemas naturales, hacen que sea difícil desarrollar modelos generales que predigan la
evolución de estas interacciones a lo largo de gradientes ambientales. Especialmente
discutibles son aquellas aproximaciones que asumen un “gradiente de estrés” que afecta
igualmente a todas las especies de una comunidad dada, ya que éstas difieren en sus
adaptaciones ecofisiológicas y, por tanto, en sus tolerancias a los distintos factores de estrés
que representan unas condiciones ambientales dadas (Chapin et al. 1987, Greiner la Peyre et
al. 2001, Körner 2003).
Los resultados presentados en el capítulo 4 apuntan a que los efectos positivos de las
plantas nodriza sobre la riqueza de especies a nivel de comunidad están promovidos tanto por
la expansión de nicho como por la reducción en la exclusión competitiva entre plantas
vecinas. Estos efectos positivos se mantienen constantes a lo largo de gradientes ambientales
donde coexisten distintos factores de estrés inversamente relacionados, o no relacionados
entre sí (p. ej. aridez y bajas temperaturas), al contrario de lo que predicen los modelos
teóricos actuales (p. ej. Lortie et al. 2004a, Michalet et al. 2006). Esta falta de variación de la
importancia de las interacciones positivas a lo largo de estos gradientes ambientales deriva de
la inexistencia de una gradiente de estrés único que afecta a todas las especies presentes en
una comunidad. Las distintas condiciones microclimáticas que se encuentran bajo el dosel de
las plantas nodriza beneficiarán a las especies menos adaptadas a las condiciones locales, sean
éstas las que sean (Greyner la Peyre et al. 2001, Liancourt et al. 2005). Sin embargo, en
gradientes ambientales gobernados por factores de estrés únicos o correlacionados entre sí (p.
ej. aridez, bajas temperaturas), las interacciones entre plantas seguirán una relación unimodal,
con predominio de las interacciones positivas a niveles medios-altos de estrés y de las de

191
competencia en ambos extremos del gradiente, tal como predicen los modelos teóricos en la
actualidad (Michalet et al. 2006, Holmgren y Scheffer 2010).
Por último, en esta tesis se muestran, por primera vez, los efectos conjuntos, directos e
indirectos, de las condiciones climáticas (filtro de hábitat) e interacciones bióticas (distintos
indicadores de competencia/facilitación) en el patrón filogenético de comunidades vegetales
semiáridas (capítulo 5). El patrón filogenético encontrado fue aleatorio en la mayoría de
casos, debido al efecto conjunto de un incremento de la dispersión filogenética promovida por
las interacciones bióticas y del aumento de la agregación en este patrón causado por los
efectos indirectos del clima sobre dichas interacciones. Los procesos de nucleación, derivados
de la dispersión zoocora concentrada en arbustos y árboles remanentes que comúnmente se
encuentra en ambientes Mediterráneos (Verdú y García-Fayos 1996, Méndez et al. 2008),
también parecen importantes en esta agregación del patrón filogenético. Los arbustos
remanentes tienden a concentrar semillas de otros arbustos dispersados por animales, un
síndrome de dispersión altamente conservado a lo largo de la evolución en la flora
Mediterránea (Herrera 1992); por tanto, estos procesos de nucleación se dan entre especies
evolutivamente próximas entre sí, causando agregación en el patrón filogenético. Se ofrecen
en este último capítulo una serie de herramientas, fáciles de medir e interpretar, que pueden
ayudar a evitar conclusiones erróneas derivadas de la inferencia de los mecanismos
implicados en el ensamblaje de especies vegetales a partir del patrón filogenético de las
comunidades en estudios observacionales (Cavender-Bares et al. 2009, Mayfield y Levine
2010, Pausas y Verdú 2010).
HACIA UN NUEVO MODELO SOBRE LA EVOLUCIÓN DE LAS INTERACCIONES ENTRE PARES DE
ESPECIES A LO LARGO DE GRADIENTES BIÓTICOS Y ABIÓTICOS
Las numerosas excepciones a la Hipótesis del Gradiente de Estrés (Bertness y Callaway 1994)
comúnmente encontradas en los sistemas naturales, han originado un debate sobre la
generalidad de sus predicciones (Maestre et al. 2005, 2006, Lortie y Callaway 2006, Callaway
2007, Lortie 2010) que ha dado lugar a nuevas interpretaciones y revisiones del modelo
inicial propuesto (Maestre et al. 2009a, Smit et al. 2009, Malkinson y Tielbörger 2010,
Holmgren y Scheffer 2010). De estas nuevas aproximaciones se concluye que la naturaleza
del estrés (si está relacionado o no con recursos directamente tomados por las plantas, p. ej.
luz o agua frente a temperatura o salinidad), la co-ocurrencia de distintos factores de estrés
abiótico o de éstos con la herbivoría, o las respuestas no lineales derivadas de las distintas

DISCUSIÓN GENERAL
192
tolerancias fisiológicas de las especies beneficiarias a los cambios microclimáticos
promovidos por las plantas nodriza, son factores clave a considerar para entender la evolución
de las interacciones planta-planta a lo largo de gradientes ambientales. Sin embargo, incluir
todos estos factores en un único modelo, que permita además inferir lo que ocurre en
comunidades vegetales enteras a partir del estudio de las interacciones entre uno o pocos
pares de especies es difícil. Incluso los modelos más recientes no son lo suficientemente
generales a la hora de arrojar predicciones sobre la evolución de la frecuencia, intensidad e
importancia de las interacciones planta-planta en la estructura y composición de las
comunidades vegetales a lo largo de distintos gradientes ambientales (pero véase Holmgren y
Scheffer 2010 para una aproximación muy meritoria). Esto ocurre porque es extremadamente
difícil que los modelos relativamente sencillos, que podrían ser extrapolables a numerosos
ecosistemas y fácilmente evaluables, incluyan la gran variedad de factores que influyen en las
interacciones planta-planta. Y modelos que incluyan todos los factores serían extremadamente
complejos y, por tanto, poco útiles.
La dicotomía entre complejidad y utilidad se ilustra con un modelo conceptual que
predice la evolución en las interacciones planta-planta a lo largo de gradientes ambientales en
zonas semiáridas a nivel de par de especies (Fig. B1). Para su elaboración se han tenido en
cuenta algunas de las aportaciones más significativas de los últimos estudios publicados, así
como los resultados presentados en esta tesis doctoral. En este modelo se evitan aspectos
problemáticos a la hora de elaborar predicciones generales, como son la existencia de un
gradiente general de estrés, que afecte a todas las especies de una comunidad por igual, o la
existencia de estrategias ecológicas que permanecen constantes a lo largo de gradientes
ambientales amplios (Travis et al. 2005, Michalet et al. 2006, Maestre et al. 2009a, Smit et al.
2009). Para ello, el modelo se basa en las tolerancias relativas de las especies implicadas a
factores ambientales concretos (derivadas bien de sus rasgos ecofisiológicos o de su historia
evolutiva), medibles y extrapolables a cualquier ambiente.
El modelo presentado en la Figura B1 toma como base el modelo teórico presentado
por Holmgren et al. (1997), que considera la eficacia biológica de las especies beneficiarias a
lo largo de un gradiente, desde el centro del dosel de la planta nodriza hasta un claro libre de
vegetación, bajo distintas condiciones de aridez. A este modelo se le ha añadido la respuesta
diferencial, y no lineal, promovida por las distintas tolerancias ecofisiológicas de las especies
implicadas o de sus distintas fases ontogenéticas, así como el efecto conjunto de distintos
niveles de aridez con el impacto de la herbivoría. Por simplicidad se asume que la carga de
herbívoros es constante a lo largo de las distintas condiciones ambientales (Smit et al. 2009).

193
Esta asunción podría ser problemática, ya que se ha demostrado que ambos factores de estrés
interactúan de diversas maneras y que, por tanto, no se puede considerar que varíen
independientemente (Illius y O´Connor 1999, Ibañez y Schupp 2001, Silliman et al. 2005,
Veblen 2008, capítulo 3). Además, el efecto de la herbivoría depende de la tolerancia a la
misma de las especies beneficiarias así como de la palatabilidad de las especies nodriza
(Baraza et al. 2006, Zamora et al. 2008). Ambos factores (tolerancia y palatabilidad) son
directamente dependientes del nivel de recursos existente (Crawley 1998, Baraza et al. 2004,
Wise y Abrahamson 2005, 2007, Smit et al. 2009). Las tolerancias ecofisiológicas han sido
organizadas en tres grupos: 1) especies con una tolerancia media a la sequía y a la sombra,
que constituyen la mayoría de especies existentes (Niinemets y Valladares 2006), 2) especies
con una tolerancia a la sombra muy alta, pero intolerantes a la sequía (p. ej., Hedera helix,
Rhagodia spinescens), y 3) especies altamente tolerantes a la sequía, pero intolerantes a la
sombra (p. ej., Retama sphaerocarpa, Helianthemum squamatum, Ambrosia dumosa). Debido
a la existencia de compromisos en las adaptaciones fisiológicas a ambos factores, no se
considera la existencia de especies con una alta tolerancia conjunta a la sombra y a la sequía,
ya que es poco probable que estas especies existan en la naturaleza (Niinemets y Valladares
2006). Por simplicidad, tampoco se pueden considerar en el modelo otras tolerancias
fisiológicas que podrían ser relevantes en zonas áridas y semiáridas, como la tolerancia a
suelos salinos o pobres en nutrientes (Pugnaire et al. 2004, Brady et al. 2005, Riginos et al.
2005, Armas y Pugnaire 2009), a bajas temperaturas (capítulo 4), o a vientos (Baumeister y
Callaway 2006). Asimismo, las tolerancias a la aridez y a la sombra no se dividen en
categorías discretas, ya que cada especie se localizará en un punto concreto dentro de un
continuo de rasgos ecológicos que les permitirán una mayor o menor tolerancia a las
condiciones ambientales (Kobe et al. 1995, Ackerly 2003, Niineemets y Valladares 2006).
En el modelo que se introduce en la Figura B1 se consideran también tres fases
ontogenéticas distintas: germinación y establecimiento temprano de las plántulas, juvenil, e
individuos adultos reproductivos. Estas fases son claves a la hora de definir 1) el
reclutamiento de nuevas especies en la comunidad (Escudero et al. 1999, Maestre et al. 2001,
Holmgren et al. 2006), y 2) la demografía de estas especies (Escudero et al. 2000, Miriti et al.
2007), contribuyendo todos ellos de forma sustancial a la dinámica de las comunidades
vegetales en las tierras secas (McAuliffe 1988, Eldridge et al. 1991, Eccles et al. 1999,
Butterfield et al. 2010). Además, se considera la posibilidad de una segregación de nicho
efectiva entre individuos adultos de las dos especies que interactúan, de forma que se predicen
diferentes cambios en las interacciones entre adultos si existe dicha segregación (Fowler

DISCUSIÓN GENERAL
194
1986, Sala et al. 1989, Stokes y Archer 2010), o si por el contrario sus nichos ecológicos se
solapan (Ludwig et al. 2004, Miriti 2006, Armas y Pugnaire 2009). Esta última separación
puede tener implicaciones importantes en la respuesta de las comunidades vegetales a los
cambios en el patrón de las precipitaciones predichos con el cambio climático (Schwinning y
Sala 2004, Knapp et al. 2008, capítulo 1). Para hacer más entendibles los resultados de las
interacciones derivados de este modelo, se han calculado también los resultados que serían
esperables en la intensidad e importancia de las interacciones (sensu Brooker et al. 2005, ver
capítulo 4) para cada estrategia ecológica, cada fase ontogenética y cada uno de los tres
niveles de aridez propuestos (Fig. B2).

195
Figura B1. Modelo teórico donde se predice la evolución de la eficacia biológica (fitness) en un continuo dosel de planta nodriza-claro libre de vegetación a lo largo de un gradiente de aridez (modificado de Holmgren et al. 1997). Las estrategias ecológicas incluidas son tolerancia media a la sequía y a la sombra (en verde), intolerancia a la sequía (en azul), e intolerancia a la sombra (en rojo). Se indican variaciones en dicha evolución a lo largo de distintas fases ontogenéticas para cada estrategia: germinación y establecimiento inicial de plántulas (panel superior), crecimiento y supervivencia de individuos juveniles (2º panel) y coexistencia entre individuos adultos sin (3er panel) o con segregación de nicho (4º panel). Las flechas en el interior de cada panel indican el efecto de una carga constante de herbívoros a lo largo del gradiente de aridez, que es indiferente a las tolerancias a la sombra o a la sequía. Este efecto consiste en un aumento del fitness más cerca del dosel de la planta nodriza, que incrementará su importancia a niveles medios de aridez. A niveles más elevados de aridez, la escasez de fuentes alternativas de forraje hará que los herbívoros incrementen su esfuerzo de búsqueda y acaben perjudicando incluso a las plantas bajo el dosel (flecha diagonal en los paneles de la derecha). Este efecto será mucho más marcado en juveniles que en adultos, como muestran las flechas en los paneles.

DISCUSIÓN GENERAL
196
Figura B2. Intensidad e importancia de las interacciones planta-planta predichas a lo largo de un gradiente de aridez y en distintas fases ontogenéticas (germinación, juvenil y adulto; de arriba a abajo) predichos a partir del modelo teórico presentado en la Figura B1. Las distintas estrategias ecológicas incluidas siguen el mismo código de colores de dicha figura. La línea discontinua en medio de cada panel corresponde con el 0 (interacción neutra) de los indicadores tanto de intensidad como de importancia de las interacciones. Valores por encima o debajo de 0 implican facilitación o competencia, respectivamente. Las líneas continuas y discontinuas del panel inferior indican interacciones entre adultos con y sin segregación de nicho, respectivamente.
TRES ESTRATEGIAS, TRES RESPUESTAS
Las especies tolerantes a la sombra se comportarán prácticamente del mismo modo a lo largo
de su ontogenia (Fig. B1). Su elevada tolerancia a la sombra hace que sean capaces de
aprovechar la mayor fertilidad en el suelo y disponibilidad hídrica comúnmente encontradas
bajo el dosel de la planta nodriza (p. ej. Franco y Nobel 1989, Callaway 2007). Por tanto, su
eficacia biológica siempre será mayor bajo una planta nodriza que en lugares libres de
vegetación, independientemente de las condiciones de aridez. Un ejemplo clásico de este tipo
de interacciones se da entre arbustos tolerantes a las altas temperaturas y los cactus
columnares asociados en el sur de Estados Unidos y norte de Méjico. Así, los cactus
columnares, como por ejemplo Carnegia gigantea (saguaro) o Neobuxbaumia tetetzo, ambos

197
intolerantes a las altas temperaturas, crecen y sobreviven únicamente asociados a la sombra de
plantas nodriza, que reducen la temperatura bajo su dosel (Shreve 1931, Niering et al. 1963,
Valiente-Banuet y Ezcurra 1991). Este comportamiento también se ha observado en la
herbácea Brachypodium retusum a lo largo del gradiente ambiental estudiado en el capítulo 4
(Fig. B3; ver también Maestre y Cortina 2004b), o en las especies Enchylaena tomentosa y
Rhagodia spinescens en el semiárido australiano (Hastwell y Facelli 2003, Prider y Facelli
2004). Especies intolerantes a altas radiaciones también se benefician de la presencia de
especies nodriza en condiciones más mésicas. Por ejemplo, Gómez-Aparicio et al. (2006)
encontraron efectos muy positivos de la reducción de la luz incidente para las especies Acer
opalus y Quercus pyrenaica en una zona seco-subhúmeda (871 mm de lluvia anual), por lo
que infirieron que el nicho de regeneración de estas especies se encontraría mayoritariamente
bajo el dosel de especies nodriza, fundamentalmente arbustos (Castro et al. 2002, Gómez-
Aparicio et al. 2004, 2005). En todos los casos mencionados, estas especies mostraron una
distribución restringida a la sombra de alguna especie nodriza bajo un amplio rango de
condiciones de aridez. Esto da lugar a interacciones positivas muy intensas (al haber una
eficacia biológica muy baja en los claros, cualquier índice que mida la intensidad de la
interacción siempre será muy positivo). Sin embargo, a medida que aumente la aridez,
algunas de estas especies se irán alejando de su óptimo ambiental, siendo más difícil su
reclutamiento en medios muy áridos (nótese la bajada en la eficacia biológica de estas
especies a medida que aumenta la aridez en la Fig. B1). Esta reducción de la eficacia
biológica puede llegar incluso a la desaparición de esta especie bajo esas condiciones en
particular, pese a la mejora microclimática promovida por la planta nodriza; esto provocaría
un colapso de las interacciones positivas para estas especies (Michalet et al. 2006, Forey et al.
2009). Si se diera esta desaparición, obviamente la intensidad de la facilitación pasaría en un
cambio brusco de muy positiva a cero (Fig. B2), ya que la especie ni siquiera estaría presente.
Al contrario que la intensidad, la importancia de la facilitación para las especies intolerantes a
la sequía se reducirá con el nivel de aridez. Esto ocurre porque la importancia es el efecto
relativo de la planta nodriza con respecto a otras condiciones ambientales (Brooker et al.
2005) y, como se aprecia en la Fig. B1, la eficacia biológica de estas especies será mucho
mayor en condiciones más húmedas, independientemente del efecto de la planta nodriza.
Además, el efecto de esta planta nodriza no será tan positivo en condiciones más áridas, ya
que el consumo de agua de esta nodriza puede llegar a superar a su mejora microclimática,
dando lugar a condiciones de “sombra seca” y a efectos netos negativos sobre las especies
vecinas (Valladares 2001, Valladares y Pearcy 2002).

DISCUSIÓN GENERAL
198
PCA climático-150 -100 -50 0 50 100
Índi
ces
de in
tera
cció
n
-1.0
-0.5
0.0
0.5
1.0
RII ST RII SH Iimp ST Iimp SH
FA
CIL
ITA
CIÓ
NC
OM
PE
TE
NC
IA
- LLUVIA + Figura B3. Índices de intensidad (RII) e importancia (Iimp) de la interacción de Brachypodium retusum con sus especies nodriza Stipa tenacissima (ST) y arbustos rebrotadores (SH) a lo largo del gradiente climático estudiado en los capítulos 4 y 5. Nótese la nula relación con el clima, y los relativamente altos valores de los índices calculados.
Las especies con una tolerancia media a la sombra y a la sequía presentarán un
comportamiento muy distinto a lo largo de su ontogenia. Por un lado, la germinación y
establecimiento temprano de estas especies será mayor bajo las plantas nodriza, debido
fundamentalmente a un suelo más fértil y a una menor radiación solar (p. ej. McAuliffe 1988,
Barberá et al. 2006). Este efecto positivo puede colapsar a medida que aumente la aridez, ya
que las plántulas de estas especies son más sensibles al estrés hídrico y pueden no germinar o
morir bajo niveles muy elevados de aridez, incluso considerando los cambios microclimáticos
de la planta nodriza (Kitzberger et al. 2000, Ibañez y Schupp 2001, Gasque y García-Fayos
2004). Esto dará lugar a una relación unimodal de la importancia e intensidad de las
interacciones a medida que aumente la aridez (Fig. B2). Los juveniles de estas especies, por
otro lado, competirán con la nodriza en condiciones más húmedas, tal como predicen todos
los modelos teóricos (p. ej. Bertness y Callaway 1994, Brooker y Callaghan 1998, Michalet et
al. 2006, Maestre et al. 2009a). Sin embargo, estas especies aumentarán su eficacia biológica
bajo el dosel de una planta nodriza en condiciones intermedias de aridez, fundamentalmente
por el efecto positivo de la sombra sobre el estado hídrico de los juveniles (Valiente-Banuet y
Ezcurra 1991, Maestre et al. 2003, Callaway 2007). A medida que aumente la aridez, la

199
intercepción de la escasa lluvia por el dosel de la planta nodriza (Tielbörger y Kadmon
2000a), o el incremento de la competencia entre ambas especies por el agua (Maestre y
Cortina 2004a), generarán condiciones de “sombra seca” que reducirán la eficacia biológica
de las especies beneficiarias bajo el dosel de la planta nodriza. Esto originará una relación
unimodal a lo largo del gradiente de aridez, tanto de la intensidad como de la importancia de
la facilitación sobre especies con una tolerancia media a la sombra y a la sequía (Tielbörger y
Kadmon 2000a, Maestre y Cortina 2004, Barchuck et al. 2005). En individuos adultos, la
competencia se extenderá hasta niveles medios de aridez cuando no haya separación de nicho,
ya que las especies competirán por el agua y perderán parte de los beneficios de la sombra
promovida por la planta nodriza al sobrepasar la altura de su dosel (Miriti 2006, Callaway
2007, Valiente-Banuet y Verdú 2008). Sin embargo, en condiciones más áridas, procesos
como el levantamiento hidráulico (hydraulic lift) o, simplemente, el efecto de la sombra sobre
la humedad del suelo puede incrementar el efecto positivo de la nodriza (Callaway 2007).
Esto generará una relación monotónica o unimodal entre la intensidad de la interacción y la
aridez, dependiendo de los efectos relativos de la sombra o el levantamiento hidráulico y la
competencia en el agua disponible para las plantas adultas facilitadas (podemos encontrar
efectos contrastados en Dawson 1993, Maestre et al. 2003 y Ludwig et al. 2004, Maestre y
Cortina 2004a). Sin embargo, si existe segregación de nicho, la intensidad (y la importancia)
de la facilitación aumentará de forma lineal con la aridez, ya que la competencia entre ambas
especies se verá reducida por la segregación de nicho a la vez que el efecto de la sombra y la
mayor fertilidad del suelo continuarán, incluso en condiciones más áridas (Armas y Pugnaire
2005, Sthultz et al. 2007).
Las especies intolerantes a la sombra, por otro lado, experimentarán mayores tasas de
germinación y supervivencia temprana lejos de plantas adultas a niveles bajos e intermedios
de aridez (p. ej. Veblen 1989, Baskin y Baskin 1998), pero pueden verse beneficiadas por
cierto nivel de sombreo en lugares más áridos (Escudero et al. 2005, Pueyo et al. 2009,
capítulo 2, pero ver Olano et al. 2005). Aunque esto puede depender de otros factores, como
el momento en que emergen estas plántulas (de la Cruz et al. 2008). La relación entre
juveniles de especies intolerantes a la sombra y una planta nodriza será casi siempre negativa,
como se ha visto en numerosos sistemas semiáridos de todo el mundo (Parker y Müller 1982,
Marañón y Bartolomé 1993, Escudero et al. 1999, Forseth et al. 2001, Prider y Facelli 2004,
Seifan et al. 2010, capítulos 1–3). Este efecto negativo se incrementará con el nivel de aridez
(Eliason y Allen 1997, Davis et al. 1999, Espigares et al. 2004, Valladares et al. 2008,

DISCUSIÓN GENERAL
200
capítulos 1 y 3, Soliveres et al. en preparación; Fig. B2), al contrario de lo que predice la
Hipótesis del Gradiente de Estrés (Bertness y Callaway 1994).
Por último, la interacción entre individuos adultos de una especie intolerante a la
sombra y una planta nodriza dependerá en buena medida de si esta especie puede “escapar” de
la competencia por la luz u otros recursos a medida que crece o no. En el primer caso se
puede reducir el efecto negativo de la nodriza de forma gradual a medida que la especie
beneficiaria crece (capítulo 2); mientras que en el segundo, incluso los individuos adultos van
a seguir compitiendo por este recurso, manteniendo así la interacción negativa a lo largo de
toda la vida del individuo (Miriti 2006, Miriti et al. 2007).
En general, la herbivoría aumentará el efecto positivo de las nodrizas en todos los
casos, sin importar las tolerancias relativas de las especies implicadas a otros factores
ambientales. Se ha visto una reducción de las interacciones competitivas cuando los
herbívoros están presentes en numerosos ecosistemas semiáridos de todo el mundo
(McNaughton 1978, Gurevitch et al. 2000, Fowler 2002, Rebollo et al. 2002, Veblen 2008).
De hecho, este cambio en las interacciones debido a la herbivoría puede llegar a compensar
los efectos negativos de la competencia por recursos, generando efectos netos positivos (Graff
et al. 2007). Sin embargo, bajo presiones muy elevadas de herbivoría, como las que se pueden
dar si aumenta la carga de herbívoros con las mismas condiciones climáticas o se reduce la
productividad vegetal con la misma carga de herbívoros (Illius y O´Connor 1999), el efecto
protector de las plantas nodriza frente a la herbivoría puede colapsar, dando como producto
interacciones ligeramente positivas o neutras (Graff et al. 2007, Smit et al. 2007, capítulo 3,
Figs. B1 y B2). No obstante, esto dependerá de la palatabilidad de la planta beneficiaria y sus
vecinas, el grado de tolerancia a la herbivoría de las especies implicadas y sus diferentes fases
ontogenéticas, y el tipo de herbívoro presente en la zona de estudio (revisado en Zamora et al.
2008).
¿EXISTE UN MODELO SENCILLO Y GENERAL QUE PREDIGA LOS RESULTADOS
DE LAS INTERACCIONES ENTRE PARES DE ESPECIES?
Incluso evitando, por simplicidad, incluir en un mismo modelo algunos de los factores clave
que modulan las interacciones planta-planta –como la presencia de factores de estrés
relacionados o no con recursos (Maestre et al. 2009a), la continuidad de rasgos ecológicos que
definen la tolerancia de las especies a dichos factores (Kobe et al. 1995, Ackerly 2003,
Liancourt et al. 2005), o la co-ocurrencia de diversos tipos de estrés a la vez (Riginos et al.
2005, Baumeister y Callaway 2006, Kawai y Tokeshi 2007), por nombrar unos cuantos– el

201
modelo presentado en la Figura B1 ha generado un total de 36 escenarios distintos,
modulados a su vez por el nivel de herbivoría presente en cada sitio. Algo que la mayoría de
ecólogos definirían quizás como un modelo demasiado complejo y poco útil. Este modelo, sin
embargo, es el único que ilustra la gran complejidad existente y los numerosos factores que
afectan al comportamiento de las interacciones planta-planta en los sistemas semiáridos a lo
largo de gradientes climáticos (en este caso de aridez). La gran cantidad de factores que
intervienen en las interacciones planta-planta haría necesario crear modelos de esta
complejidad para cada bioma o sistema de estudio (p. ej. sistemas de alta montaña o
ecosistemas salobres requerirían de modelos diferentes al presentado, pero igualmente
complejos). Además, a la hora de aplicar estos modelos a nivel de comunidad, deberíamos
incluir los efectos derivados de que las especies no están organizadas en pares de individuos
aislados, sino que están organizadas en manchas discretas donde coexisten numerosos
individuos de diversas especies (p. ej. Aguiar y Sala 1999). Esto podría generar fenómenos de
facilitación indirecta (Levine et al. 1999, Cuesta et al. 2010), segregación o
complementariedad de nicho (Hector et al. 1999, Silvertown 2004, Stokes y Archer 2010), o
competencia intransitiva (Laird y Schwamp 2006, 2009), que podrían variar todos los
resultados predichos en los 36 escenarios mostrados en el modelo.
Se sugiere, por tanto, que el único modelo sencillo, que podría resultar útil para
predecir los resultados de las interacciones entre pares de especies e inferir estos resultados a
redes de interacciones más complejas, como las encontradas en las comunidades naturales, y
en todos los sistemas de estudio de forma general, es el modelo basado en el estrés individual,
descrito en el capítulo 4. Este modelo resume todos los posibles resultados de todos los
posibles escenarios prediciendo que las interacciones planta-planta se tornarán más positivas a
medida que la especie beneficiaria se aleja de su óptimo ambiental (ya sean estas condiciones
más o menos áridas, con o sin herbivoría, frío, salinidad, etc.), llegando a un colapso cuando
las condiciones ambientales son tan duras para esta especie en concreto que ni siquiera puede
reclutar bajo las plantas nodriza. Definir el óptimo ambiental de cada especie es una tarea
ardua, pero sin embargo, es la única forma de predecir de forma general el resultado de la
interacción de una especie en concreto con sus vecinas. Para inferir la distancia de una especie
en concreto a su óptimo podría considerarse utilizar el número de individuos reclutados en
áreas libres de vegetación como un indicador de la distancia al óptimo de esa especie (más
individuos reclutados significarán mejores condiciones ambientales para esa especie
[Valiente-Banuet et al. 2006, Valiente-Banuet y Verdú 2007]). También se pueden inferir las
tolerancias relativas a cualesquiera que sean los factores de estrés dominantes en cada área de

DISCUSIÓN GENERAL
202
estudio a partir de las bases de datos y publicaciones disponibles sobre rasgos ecológicos de
las especies implicadas. Por ejemplo, podemos encontrar algunas bases de datos de libre
acceso para especies mediterráneas (i.e. BROT [Paula et al. 2009] o BASECO [Gachet et al.
2005]) o artículos científicos que incluyen rasgos ecológicos de multitud de especies de todo
el mundo (p. ej. Niinemets y Valladares 2006, Poorter et al. 2009, Wright et al. 2004). Un
ejemplo de este tipo de inferencia lo podemos encontrar en Pavoine et al. (2010).
Alternativamente, se podrían utilizar modelos de distribución potencial (p. ej. Guisan y
Zimmermann 2000, Loiselle et al. 2003) de las especies beneficiarias. Según el modelo
basado en el estrés individual, estas especies experimentarán un efecto más positivo de la
presencia de una nodriza a medida que nos alejemos del centro de su área de distribución
potencial.
FACILITACIÓN A NIVEL DE COMUNIDAD: IMPLICACIONES PARA LA ESTRUCTURA Y EVOLUCIÓN DE
LOS ECOSISTEMAS SEMIÁRIDOS
El modelo basado en el estrés individual soluciona algunos de los supuestos más
problemáticos anteriormente mencionados (existencia de un gradiente de estrés general para
todas las especies de una comunidad dada, y de estrategias ecológicas que se mantienen
estables a lo largo de gradientes ambientales amplios) mediante el establecimiento de un
gradiente de estrés único para cada especie. Este gradiente está basado en medidas de la
tolerancia ecológica de cada especie a distintos factores ambientales, cuantitativas y
extrapolables a cualquier sistema de estudio. Así pues, la ausencia de estos supuestos, junto
con la consideración de la diferente naturaleza de los gradientes ambientales que podemos
encontrar en la naturaleza (compuestos por factores de estrés independientes o
correlacionados entre sí) y las diferencias en las tolerancias relativas entre las especies que
coexisten en una comunidad dada, nos permitirá considerar finalmente la evolución de la
importancia de las interacciones planta-planta a nivel de comunidad bajo distintas condiciones
ambientales.
En la Figura B4 se muestra un modelo conceptual basado en una sencilla comunidad
de tres especies (A, B y C). Cada especie difiere en su óptimo ambiental, y por tanto, tendrá
una distribución diferente (basada en su eficacia biológica bajo distintas condiciones) a lo
largo de un gradiente ambiental. Si no consideramos las interacciones positivas entre plantas,
podemos observar un solapamiento mínimo en las distribuciones de estas tres especies, que
vendría dado por: 1) las diferencias en sus tolerancias específicas a los distintos factores de

203
estrés encontrados a lo largo de ese gradiente ambiental (Chapin et al. 1987), y 2) la exclusión
competitiva que ejerce la especie más adaptada a las condiciones locales sobre las demás
(Grime 1973; panel izquierdo de la Fig. B4). En cambio, si consideramos las interacciones
facilitativas, se puede observar como la distribución espacial de cada especie aumenta
(expansión de nicho; Bruno et al. 2003), ya que las especies más adaptadas a las condiciones
locales aumentan las eficacia biológica de las menos adaptadas (panel derecho de la Fig. B4).
Sin embargo, incluso teniendo en cuenta estas interacciones positivas, llega un momento en
que las condiciones ambientales son demasiado duras para una especie dada, y el
reclutamiento de esta especie en concreto es imposible, dando como resultado una eficacia
biológica igual a cero, pese a la mejora microambiental que pudiera ejercer la especie nodriza
(Kitzberger et al. 2000, Ibañez y Schupp 2001; flechas en el panel derecho de la Fig. B4). Sin
embargo, el que el efecto positivo de la facilitación desaparezca para esa especie en concreto
no significa que haga lo propio a nivel de la comunidad entera, ya que habrá otras especies
capaces de reclutar bajo esas condiciones locales, pero lo suficientemente poco adaptadas
como para verse beneficiadas por la presencia de una especie nodriza. Esto explicaría por qué
en diversas comunidades se observa una reducción de la facilitación a escala de mancha, pero
no un efecto significativo de esta reducción en la facilitación sobre la diversidad local
(revisado en Michalet et al. 2006).

DISCUSIÓN GENERAL
204
Figura B4. Modelo teórico donde se predice la eficacia biológica de tres especies con rasgos ecológicos contrastados a lo largo de un gradiente ambiental. En el panel izquierdo se consideran sólo las tolerancias ecofisiológicas de cada especie y la exclusión competitiva. En el panel de la derecha se considera la expansión de nicho promovida por las especies nodriza (especies más adaptadas a las condiciones locales). Bajo condiciones demasiado severas para cada especie, su reclutamiento es imposible, incluso incluyendo la expansión de nicho, y la facilitación colapsa a nivel de esa especie (flechas en el panel). Cuando dejan de existir especies adaptadas a las condiciones locales a lo largo de este gradiente ambiental, la facilitación colapsa a nivel de comunidad (línea superior en el panel derecho). Hasta llegar a ese punto, la importancia de las interacciones positivas a nivel de comunidad permanece estable, ya que la identidad, pero no la cantidad, de especies facilitadas es lo que cambia a lo largo del gradiente.
Cuando el gradiente ambiental esté formado por diversos factores de estrés,
independientes entre sí, la importancia de la facilitación a nivel de la comunidad entera se
mantendrá constante hasta que ya no quede ninguna especie adaptada a las condiciones
locales que pueda facilitar a las demás, lo que llevaría al colapso de la facilitación (Silliman et
al. 2005, Michalet et al. 2006; línea superior del panel izquierdo de la Fig. B4). Casos en que
el gradiente ambiental esté dominado por un solo factor de estrés, o varios correlacionados
entre sí (p. ej. Bertness y Shumway 1993, Callaway et al. 2002), serían simplemente una
fracción del escenario anteriormente descrito. Ambos casos están separados por la línea
discontinua en la Figura B4. Este sencillo modelo conceptual resume el por qué las
interacciones positivas son importantes en numerosos ecosistemas de todo el mundo, y no
sólo en los considerados “estresantes” y el por qué las interacciones positivas serán más
importantes bajo niveles “intermedios de estrés”, se trate del bioma que se trate (revisado en
Holmgren y Scheffer 2010). También explica el colapso de las interacciones positivas bajo

205
elevados niveles de estos factores de estrés a lo largo de gradientes ambientales, ya sea para
especies concretas (Kitzberger et al. 2000, Ibañez y Schupp 2001), o en comunidades enteras
(Silliman et al. 2005, Michalet et al. 2006). La Figura B4 ayuda a entender el papel de la
facilitación en la evolución y resiliencia de las comunidades naturales. Estas interacciones
positivas asegurarán el nicho de regeneración de las especies menos adaptadas a las
condiciones locales (Valiente-Banuet et al. 2006), sean estas las que sean, o el mantenimiento
de la diversidad y el funcionamiento ecosistémico en condiciones ambientales menos
productivas (Mulder et al. 2001, Kikvidze et al. 2005, Badano y Cavieres 2006), como las
predichas con el cambio climático para la cuenca Mediterránea (Brooker 2006). Esto ocurrirá
siempre y cuando queden especies que presenten tolerancias ecofisiológicas a las condiciones
ambientales existentes y puedan suavizar estas condiciones para las especies menos
adaptadas.
Ahora bien, al igual que la mayoría de artículos relacionados con las interacciones
entre plantas, el modelo propuesto en la Figura B4 no considera el hecho de que las especies
de una comunidad no se relacionan par a par, si no que forman ensamblajes más o menos
complejos que atañen a todas las especies que coexisten en esa comunidad (Maestre et al.
2010). Un ejemplo de ello serían las manchas de vegetación de las tierras secas, donde
coexisten un elevado número de especies, y donde es probable que se den interacciones
positivas y negativas entre multitud de ellas, dando lugar a complejas redes de interacción
(Verdú et al. 2010). Como se ha mencionado antes, esto puede dar lugar a procesos de
facilitación indirecta, competencia intransitiva, segregación o complementariedad de nicho
(Grace et al. 1993, Hector et al. 1999, Levine et al. 1999, Silvertown 2004). El incremento de
la diversidad mediante la expansión de nicho, junto con el aumento en la heterogeneidad en
los recursos por el que estas especies compiten, promovidos ambos por la presencia de
especies nodriza, puede dar lugar a aumentos desproporcionados en la diversidad local
(Bowker et al. 2010). Aunque estos efectos ya se conocían y han sido estudiados por separado
(revisado en Brooker et al. 2008), hasta ahora ningún modelo teórico ha incluido ambos
mecanismos a la hora de predecir el papel de la facilitación en la diversidad de las
comunidades naturales (Callaway 2007). Se pretende dar ese paso con el modelo mostrado en
la Figura B5. Este modelo se basa en la distribución que podemos encontrar para cualquier
especie a lo largo de un gradiente espacial o temporal en cualquier libro de ecología, esto es,
una campana de Gauss más o menos apuntada. Como hemos explicado antes, los rasgos
ecológicos de cada especie, y sus tolerancias relativas a los diferentes factores de estrés
presentes a lo largo de ese gradiente, definirán su óptimo ambiental, que variará según la

DISCUSIÓN GENERAL
206
especie. En este modelo, se utiliza un conjunto inicial de siete especies (de la A a la G). El
nicho potencial de estas especies (panel superior izquierdo de todos los escenarios de la Fig.
B5) vendrá indicado por los diversos filtros de establecimiento, esto es, llegada de propágulos
(dispersión) y tolerancia a las condiciones locales. Una vez llegados a este punto es donde las
interacciones bióticas adquieren mayor importancia (p. ej. Huston 1999, Rajaniemi et al.
2006, pero ver Mitchell et al. 2009, Gotelli et al. 2010).

207
Figura B5. Modelo teórico donde se predicen los nichos potenciales y ocupados de distintas especies a lo largo de un gradiente ambiental, y como esto se refleja en la riqueza local de especies de una comunidad dada a partir de un conjunto inicial de siete especies (A–G). Las predicciones se realizan en cuatro escenarios distintos teniendo en cuenta: A) homogeneidad en los recursos por los que compiten las plantas, la cual hace imposible la intransitividad en la competencia o la segregación de nicho, B) heterogeneidad en los recursos, haciendo posible cierto grado de segregación de nicho o competencia intransitiva entre las especies existentes, C) expansión de nicho de las especies menos adaptadas a las condiciones locales promovida por especies nodriza, lo cual genera la llegada de nuevas especies que inicialmente no estaban en la comunidad, pero condiciones homogéneas en los recursos por los que estas especies compiten, y D) el doble efecto de las plantas nodriza (incrementar la heterogeneidad de nutrientes y aumentar el conjunto de especies mediante expansión de nicho) en un sistema ya de por sí heterogéneo. En los escenarios A y C se han sombreado los nichos ocupados por cada especie, para facilitar su comprensión. En los escenarios B y D esto no ha sido posible por el alto grado de solapamiento entre nichos que se predice. Nótese como la relación entre la riqueza de especies y la productividad (panel de abajo a la derecha en cada escenario) va cambiando según consideremos unos procesos u otros.

DISCUSIÓN GENERAL
208
En un primer escenario suponemos una disposición homogénea de los recursos por los
que compiten estas siete especies (escenario A en la Fig. B5). En estas condiciones, es
imposible que exista segregación de nicho o intransitividad en la competencia (Huston 1979,
1999). También obviamos aquí el efecto facilitador que pueden ejercer unas especies sobre
otras. Por tanto, la exclusión competitiva es la única interacción planta-planta que nos queda.
En unas condiciones más o menos homogéneas, las especies más adaptadas a las condiciones
locales (competitivas, sensu Grime 1979) pueden desplazar a las menos adaptadas, haciendo
que su nicho real sea menor que el potencial. Poniendo como ejemplo las especies A, B y C,
cumpliéndose que A>B>C tenemos que A excluirá a B y C en los lugares donde esté presente,
reduciendo el nicho real de B y C. Y B hará lo mismo con C (áreas sombreadas del escenario
A).
En el escenario B se considera el efecto de la heterogeneidad ambiental, esto es, la
variabilidad espacial o temporal en los recursos por los que compiten las especies presentes en
una comunidad. Esto permitirá 1) que no haya una jerarquía marcada en la competencia
(competencia intransitiva), si no que, dependiendo de las condiciones microambientales, la
especie A pueda desplazar a las demás o pueda ser desplazada por otras (Gilpin 1975, Grace
et al. 1993), y 2) que se pueda dar segregación de nicho, esto es, los nichos reales de
diferentes especies pueden mantenerse aunque éstas compitan por el mismo recurso, ya que
difieren en la toma de este recurso en el espacio o en el tiempo (Silvertown 2004). Ambos
procesos son importantes, ya que pueden aumentar la diversidad a nivel local (Grace 1993,
Huston 1999, Laird y Schwamp 2006; ver Tilman 1994 para una aproximación alternativa).
En el escenario B de la Figura B5 podemos ver, por ejemplo, como A no excluye
competitivamente a B de su nicho potencial (aunque reduce su nicho real considerablemente)
pese a su superioridad competitiva. Esto ocurre debido al efecto indirecto de D, una especie
que supera competitivamente a A. Por tanto, A, B y D coexisten (competencia intransitiva:
A>B>D>A, que genera facilitación indirecta de D sobre B). Alternativamente, G y F pueden
coexistir a pesar de que sus nichos potenciales se solapan. Esto se debe a que ambas especies
no compiten exactamente entre ellas pese a necesitar los mismos recursos, ya que los toman
en lugares o momentos diferentes (segregación de nicho, p. ej. Sala et al. 1989, capítulo 1). En
sistemas heterogéneos y relativamente ricos en especies, estas especies pueden
complementarse, de forma que toman los recursos disponibles de forma más eficiente,
incrementando la productividad a nivel del ecosistema (Hector et al. 1999). Aunque esta
complementariedad de nicho es clave para entender la relación entre diversidad y
productividad, por lo que se ha incluido en el modelo, no afecta a la riqueza de especies, si no

209
a la relación de la riqueza con la productividad total del sistema, por lo que no se discutirá
ahora el papel de este proceso.
El escenario C, por otro lado, es exactamente idéntico al A, salvo que se ha incluido la
expansión de nicho (sensu Bruno et al. 2003). Hasta el momento, este es el único efecto de las
interacciones positivas que se había incluido en los modelos teóricos sobre el efecto de las
interacciones planta-planta en la riqueza local de especies (Hacker y Gaines 1997, Bruno et al.
2003, Lortie et al. 2004a, Michalet et al. 2006). Como predicen todos estos modelos, y se ha
matizado en esta discusión, las especies más adaptadas a las condiciones locales
incrementarán la eficacia biológica de las especies menos adaptadas, e incluso pueden
permitir la colonización de otras especies, que no habían podido cruzar los filtros ambientales
por ellas mismas (H e I, en el escenario C). Al haber ahora más especies en la comunidad, hay
mayores posibilidades de que se de segregación o complementariedad de nicho. Sin embargo,
este escenario sigue asumiendo unas condiciones homogéneas, que permiten la existencia de
especies “competitivas” que excluyan a otras especies de sus nichos potenciales. Esto daría
lugar al incremento de la riqueza, gracias a la expansión de nicho, predicho en condiciones
más improductivas debido a que las especies “tolerantes al estrés” facilitan a las especies
“competitivas” en condiciones improductivas (Hacker y Gaines 1997, Travis et al. 2005,
Michalet et al. 2006; especie I en el escenario C). Aunque también puede incrementar la
diversidad en situaciones mucho más productivas, donde la especie más adaptada a las
condiciones locales (A en este caso), facilita la entrada de una especie menos adaptada (H).
Este fenómeno ha sido observado en sistemas considerados muy productivos, como el bosque
lluvioso tropical o bosques de ribera, entre otros (revisado en Holmgren y Scheffer 2010).
Finalmente, en el escenario D se considera un medio de por sí heterogéneo, como la
mayoría de medios naturales, donde además se incluye el doble efecto positivo que ejercen las
especies nodrizas sobre la diversidad local en una comunidad dada. Estos efectos son: 1)
incrementar el conjunto de especies disponibles por medio de la expansión de nicho (Bruno et
al. 2003, Lortie et al. 2004a, Badano y Cavieres 2006), y 2) aumentar la heterogeneidad
ambiental bajo su dosel, lo que incrementa aún más la heterogeneidad en los recursos por los
que estas especies compiten (p. ej. Pugnaire et al. 1996a, Cuesta et al. 2010). Este escenario
no había sido considerado hasta el momento para evaluar el efecto de la facilitación sobre la
diversidad local (Callaway 2007). Sin embargo, es en este escenario donde las posibilidades
de que se de competencia intransitiva y la segregación o complementariedad de nicho son
máximas, lo que llevaría a un aumento de la diversidad desproporcionado provocado por
procesos de retroalimentación entre ambos procesos (más especies disponibles y más

DISCUSIÓN GENERAL
210
heterogeneidad en la competencia generan competencia intransitiva y/o segregación de nicho,
que a su vez permiten que más especies puedan coexistir). Este proceso de retroalimentación
positiva aumenta la diversidad local gracias a dos efectos fundamentales. Primero, porque
aumenta el nicho potencial de la mayoría de las especies del conjunto, y segundo, porque se
reducen las posibilidades de exclusión competitiva. Pocas especies son excluidas de sus
nichos potenciales ya que, o bien difieren en el nicho de las especies con las que coexisten (H,
D y A, en el escenario D), o bien, terceras especies evitan que estas sean excluidas (D permite
que B no sea excluida por A). Este proceso de retroalimentación positiva, que lleva a la
reducción de la exclusión competitiva para muchas especies, podría ser la causa del
incremento de diversidad a lo largo de la sucesión en algunos ecosistemas (Johnston y Odum
1956, McKindsey y Bourget 2001), o de los resultados contrastados en la relación entre
diversidad e invasibilidad de los ecosistemas (revisado en Bruno et al. 2003). Sistemas más
diversos podrán atraer nuevas especies (sean estas invasoras o no) si las características del
sistema de estudio (i.e. heterogeneidad en los recursos por los que compiten las especies)
permiten que se den estos procesos de retroalimentación, pero evitarán la entrada de nuevas
especies si estos procesos de retroalimentación no se dan y domina la exclusión competitiva.
La coexistencia o no de todos estos factores (expansión, segregación y
complementariedad de nicho, y competencia intransitiva) puede modificar de forma sustancial
la relación entre la riqueza de especies y la productividad del ecosistema, incluso con las
mismas especies, estudiadas a la misma escala y con la misma amplitud del gradiente
estudiado (panel inferior derecho de todos los escenarios; ver Whitakker 2010, Mittlebach
2010 y referencias en estos textos para una discusión detallada sobre los efectos de la escala
de estudio en la relación riqueza-productividad). Esto podría ayudar a explicar la gran
cantidad de excepciones que se han encontrado a esta relación unimodal entre riqueza y
diversidad en numerosos ecosistemas de todo el mundo a nivel local (p. ej. Grace 1999,
Waide et al. 1999, Gillman y Wright 2006). El modelo propuesto contradice no sólo la idea de
que la facilitación es sólo importante en los sistemas más improductivos y estresantes (p. ej.
Bertness y Callaway 1994), si no también que la facilitación sólo afecte a esta relación entre
diversidad y productividad a niveles medios-bajos de ésta última (Michalet et al. 2006). Se
propone en esta discusión que evaluar el papel relativo de la expansión, complementariedad y
segregación de nicho, junto con el grado de intransitividad de la competencia nos permitirá,
por un lado, evaluar de una forma completa el papel de las interacciones planta-planta en la
diversidad local (Brooker et al. 2008), y por otro, definir la relación entre esta diversidad local
y la productividad y el funcionamiento del ecosistema (Mulder et al. 2001, Callaway 2007,

211
Maestre et al. 2010). Ambas cuestiones no son sólo atractivas desde el punto de vista teórico,
ya que nos permiten arrojar algo de luz sobre los mecanismos implicados en la relación entre
la diversidad y la productividad de los ecosistemas (Whitakker 2010), sino que también son
cruciales para finalmente entender el papel de la biodiversidad en el funcionamiento de los
ecosistemas y los servicios que éstos proveen (Hooper et al. 2005, Michalet et al. 2006) y, por
tanto, en como entendemos y manejamos la naturaleza. Sin embargo, evaluar el papel de cada
uno de los procesos mencionados a nivel de comunidad requeriría experimentos muy costosos
y logísticamente inabordables, por lo que se ha sugerido el uso de estudios observacionales
para evaluar la importancia relativa de las interacciones bióticas a nivel de comunidad (p. ej.
Gotelli y Graves 1996, Brooker et al. 2008, Pausas y Verdú 2010). A pesar de que las
herramientas existentes han permitido un mejor entendimiento de los mecanismos de
ensamblaje de especies y el papel de las interacciones bióticas en este proceso (p. ej. Gotelli y
Graves 1996, Dullinger et al. 2007, Maestre et al. 2008, 2010, Rooney 2008, Bowker et al.
2010, Gotelli et al 2010), éstas son insuficientes por el momento para discernir entre los
procesos anteriormente mencionados. Aunque queda mucho por hacer en este aspecto, se
ofrecen en los trabajos desarrollados en esta tesis doctoral (capítulos 4 y 5) una serie de
herramientas para diferenciar entre los procesos de competencia intransitiva, expansión y
segregación de nicho, y su efecto sobre la riqueza de especies y el patrón filogenético local, a
partir de estudios observacionales. El refinamiento de estas técnicas ayudará a entender
finalmente el papel de las interacciones bióticas no tróficas en el ensamblaje de las especies
que forman las comunidades naturales, y la relación entre la diversidad de estas especies y la
productividad de los ecosistemas.
IMPLICACIONES DE LA FACILITACIÓN EN LA RESTAURACIÓN DE LOS SISTEMAS SEMIÁRIDOS
El papel que la facilitación puede jugar en la restauración de los ecosistemas Mediterráneos
en general, y los semiáridos en particular, ya ha sido discutido ampliamente con anterioridad
(Pugnaire et al. 1996, Maestre et al. 2001, Castro et al. 2002, Gómez-Aparicio et al. 2004,
Cortina y Maestre 2005, Padilla y Pugnaire 2006, Valladares y Gianoli 2007, Pueyo et al.
2009, Cortina et al. 2010). Sin embargo, en la inmensa mayoría de estos trabajos se trata el
papel de las interacciones facilitativas en un contexto puramente climático, asumiendo que la
importancia de estas interacciones para el éxito de la restauración aumentará con el nivel de
aridez. Muy pocos trabajos han tenido en cuenta el colapso de estas interacciones facilitativas
bajo niveles extremadamente altos de aridez o herbivoría (Valladares y Gianoli 2007, Pueyo

DISCUSIÓN GENERAL
212
et al 2009, Cortina et al. 2010), o la importancia relativa de las interacciones facilitativas
frente a otros múltiples procesos y herramientas importantes para la restauración (Méndez et
al. 2008, Pueyo et al. 2009, Cortina et al. 2010). Se discute en esta sección la idoneidad de
utilizar las interacciones facilitativas en restauración en sistemas semiáridos, con respecto a
otras técnicas disponibles. También se discute como puede adaptarse el manejo de los
ecosistemas semiáridos a las futuras condiciones ambientales que se esperan con el cambio
climático, y el papel que la facilitación puede jugar en dicho proceso.
El funcionamiento ecosistémico en la mayoría de ambientes semiáridos viene dado por
la dinámica fuente-sumidero, que permite a las manchas de vegetación retener el agua de
escorrentía, recursos y semillas procedentes de las áreas de suelo desnudo. Esto favorece la
captura y reciclaje de nutrientes y aumenta la colonización de nuevas especies en las manchas
de vegetación (Ludwig y Tongway 1995, Aguiar y Sala 1999, Puigdefábregas et al. 1999).
Como ya se ha comentado en la introducción general, la degradación de estos sistemas viene
dada por la pérdida de esta estructura de la vegetación (reducción del tamaño y
distanciamiento entre los parches [Maestre et al. 2006, Kefi et al. 2007], o reducción de la
cobertura [Maestre y Escudero 2009]). La degradación de los sistemas semiáridos no es un
proceso lineal, sino que experimenta distintos umbrales de degradación correspondientes con
cambios drásticos en la estructura, composición y funcionamiento ecosistémicos (Van de
Koppel et al. 1997; ver Fig. B6, modificado de Cortina et al. 2010). Aunque se ha visto que el
funcionamiento ecosistémico no está necesariamente unido a su “restaurabilidad” (Cortina et
al. 2005, Maestre et al. 2006), conocer en qué estado funcional se haya el ecosistema es
básico para: 1) evaluar las medidas más adecuadas para su restauración, y 2) definir áreas
prioritarias para la conservación o restauración (Cortina et al. 2005, 2010, Méndez et al.
2008). Así por ejemplo, en el caso de los espartales de Stipa tenacissima, aunque dependerá
también de objetivos sociales, políticos y económicos, dos tipos de áreas prioritarias para la
restauración de las estepas de Stipa tenacissima serían aquellas con niveles de funcionalidad
intermedios (“estepas” y “estepas empobrecidas” sensu Cortina et al. 2010). En el caso de las
estepas empobrecidas, para asegurarnos que la pérdida en la cobertura vegetal y la diversidad
no alcanza estados de degradación severos (Maestre y Escudero 2009). En el caso de las
estepas, en cambio, la prioridad se debería a que podemos ganar bastante en funcionalidad y
servicios ecosistémicos con relativamente poca inversión.

213
Figura B6. Esquema donde se resumen los cuatro posibles estados de degradación de una estepa de Stipa tenacissima: estepa con matorral (A), estepa sin matorral (B), estepa degradada o empobrecida (C) y área desertificada o “badland” (D). Se indican de forma esquemática las actuaciones prioritarias en cada etapa. La introducción de especies leñosas, prestando atención a la procedencia y calidad del material utilizado, es recomendable en todas las etapas, aunque el grosor de la flecha indica en cada caso el grado de prioridad de esta práctica. Se señalan las áreas prioritarias para la restauración con un círculo. EI = estructuras inertes para la captura y retención de escorrentía; ME = mallas de exclusión de herbívoros.
Hemos de tener en cuenta que el principal cuello de botella para el establecimiento de
las plantas en medios semiáridos es el agua disponible (Eldridge et al. 1991, Whitford 2002,
Holmgren et al. 2006). Por tanto, sea cual sea la herramienta que se utilice y el estado de
degradación de la zona donde se actúe, un año seco dará un rendimiento menor del esfuerzo

DISCUSIÓN GENERAL
214
invertido (Navarro et al. 2006 y referencias en dicho texto). Por tanto, predecir de alguna
manera las condiciones climáticas venideras nos daría una información crucial sobre cuando
actuar para maximizar el éxito de los esfuerzos de restauración (Cortina et al. 2010). La gran
importancia de la alternancia de los eventos climáticos globales (El Niño/La Niña) para el
patrón de precipitaciones de numerosas regiones de todo el mundo, incluida España (ver
Holmgren et al. 2006), nos ofrece esta herramienta de predicción (Valladares y Gianoli 2007).
Así, la alternancia entre las fases del Niño y la Niña nos ofrece ventanas temporales de vital
importancia para predecir estos años lluviosos y centrar en ellos los proyectos de restauración.
Si bien la alternancia de estos fenómenos no es predecible al 100%, presenta un tiempo de
recurrencia aproximado de entre tres y seis años (Holmgren et al. 2006 y referencias en ese
texto). Por tanto, mientras no se desarrollen mejores herramientas de predicción, cuando en un
año en concreto se de la fase del Niño (condiciones más lluviosas para España), podemos
inferir que los dos siguientes años serán igualmente benignos. Los esfuerzos destinados a la
restauración de los sistemas semiáridos deberían centrarse exclusivamente en estos años,
probablemente más lluviosos que la media, donde el éxito de los proyectos de restauración
seguramente será mayor, sean cuales sean las herramientas que apliquemos.
Una vez establecido el cuándo y el dónde es mejor actuar, podemos centrarnos en
cómo es mejor hacerlo. Se utiliza aquí como base el diagrama modificado de Cortina et al.
(2010) para ilustrar el papel relativo de las interacciones planta-planta frente a otros procesos
y herramientas útiles para asegurar el éxito de la restauración en los espartales de Stipa
tenacissima (Fig. B6). En este diagrama se establecen cuatro estados alternativos en la
degradación del funcionamiento ecosistémico en el caso de estos espartales que varían desde
el suelo desnudo a la estepa con vegetación arbustiva. En las etapas menos degradadas (estepa
con vegetación arbustiva, estepa y estepa empobrecida) interesaría incrementar la cobertura
de vegetación arbustiva, lo cual se ha demostrado muy positivo para el funcionamiento y la
diversidad de los espartales ibéricos (Maestre y Cortina 2005, Cortina y Maestre 2005,
Maestre et al. 2009b). Para ello, en el caso de la estepa con vegetación arbustiva (no en el
resto de etapas de degradación), podemos confiar en los procesos de nucleación que
comúnmente suceden en sistemas Mediterráneos, donde los animales dispersan de forma
natural propágulos de numerosas especies bajo los arbustos o árboles remanentes (efecto
percha), que a su vez se establecerán con más éxito bajo estos arbustos debido a las
condiciones de sombreo y mayor fertilidad del suelo (Verdú y García-Fayos 1996, Pausas et
al. 2006, Méndez et al. 2008). Si bien este proceso natural puede incrementar la diversidad y
la cobertura de arbustos rebrotadores de forma natural en estos espartales y en numerosos

215
ecosistemas Mediterráneos (restauración pasiva), es poco probable que se aumente de forma
significativa la colonización de estas especies en lugares donde no haya arbustos o árboles
remanentes que atraigan a los animales dispersores. En otras palabras, la colonización natural
sólo se dará en lugares donde previamente ya haya algún arbusto presente (los animales
depositarán las semillas en esos lugares), no creándose nuevos parches de vegetación
arbustiva en otros lugares (Cortina y Maestre 2005). Por tanto, incrementar la tasa de
colonización y el número de parches formados por estos arbustos es deseable, especialmente
en las estepas donde éstos no estén presentes (Cortina y Maestre 2005, Maestre y Cortina
2005). Para ello, utilizar las interacciones positivas del esparto con estas especies ha sido
recomendado (p. ej. Maestre et al. 2001, Gasque y García-Fayos 2004, Barberá et al. 2006,
Navarro et al. 2008). Sin embargo, hay que considerar el hecho de que estas interacciones
pueden colapsar bajo niveles extremos de aridez o herbivoría (Maestre y Cortina 2004a,
capítulo 3). Bajo estas condiciones, el establecimiento de pilas de ramas u otras estructuras
inertes (p. ej. microcuencas) que provean mejores condiciones hídricas y de fertilidad del
suelo, o bien que reduzcan los efectos de la herbivoría (p. ej. mallas protectoras), para los
plantones introducidos o para la colonización natural son recomendables (Tongway y Ludwig
1996, Ludwig y Tongway 1996, Holmgren et al. 2006, Soliveres et al. 2008, Pueyo et al.
2009).
El uso de estructuras inertes destinadas a aumentar la retención del agua de escorrentía
y otros recursos, también es recomendable en lugares muy degradados, independientemente
de su nivel de aridez, debido a la ausencia de plantas que puedan ejercer como nodriza (estado
más degradado en la Fig. B6). También en estas zonas más degradadas, donde con seguridad
se ha perdido parte, o todo el horizonte orgánico del suelo, puede ser conveniente incrementar
la fertilidad del suelo mediante el uso de enmiendas orgánicas. Si bien esto dependerá de las
características físico-químicas del sitio a restaurar y de las condiciones climáticas (Soliveres
et al. en prensa y referencias en ese texto). Otra alternativa para incrementar la cobertura
vegetal, especialmente en los sitios más áridos o degradados, es la introducción de especies
más heliófilas (p. ej. Retama sphaerocarpa; Moro et al. 1997, Caravaca et al 2003). Los
capítulos 1 y 3 de esta tesis doctoral han demostrado que el uso de las interacciones
facilitativas tampoco sería recomendable para la introducción de estas especies, siendo
preferible introducir los plantones en lugares libres de vegetación. Las interacciones
facilitativas, por tanto, aunque son una herramienta de restauración muy útil en determinados
escenarios de restauración, no son recomendables en condiciones de aridez extrema o
herbivoría muy intensa. En estas situaciones, otras medidas como las anteriormente

DISCUSIÓN GENERAL
216
explicadas (y las discutidas más abajo) pueden dar mejores resultados en los proyectos de
restauración.
En algunas ocasiones, especialmente en los futuros escenarios predichos con el cambio
climático, las condiciones ambientales pueden ser extremadamente adversas para la gran
mayoría de especies de interés, de forma que su reclutamiento sea imposible, incluso bajo el
dosel de una especie nodriza o con la presencia de las estructuras inertes mencionadas con
anterioridad. En este caso, una cuidadosa selección de las poblaciones de donde se obtiene el
material para la restauración, y el establecimiento de una calidad de planta adecuada para las
condiciones ambientales puede mejorar el rendimiento de algunas especies (Cortina et al.
2006, Villar et al. en prensa). Aunque esta selección y preparación del material es importante
en cualquier proyecto de restauración, cobra especial importancia en sistemas particularmente
áridos, donde la procedencia o la calidad de la planta pueden decidir el destino de las
plántulas introducidas (Navarro et al. 2006, Trubat et al. 2008). La aplicación conjunta de la
selección del material vegetal adecuado (especies y procedencias), en conjunto con una buena
calidad de planta y el uso de estructuras inertes, puede llevarnos al éxito en la restauración
bajo estas condiciones más áridas o degradadas (p. ej. proyecto FUNDIVFOR en el sureste
ibérico, http://80.24.165.149/fundivfor/).
A pesar de que se ha sugerido que los arbustos pueden ser la mejor opción posible
como planta nodriza en ecosistemas Mediterráneos en las actuales condiciones climáticas
(Gómez-Aparicio et al. 2004, Gómez-Aparicio 2009), se debería conceder un mayor crédito
como especies nodriza a las herbáceas perennes como Stipa tenacissima en el futuro. Estas
herbáceas es probable que ejerzan efectos más positivos sobre las especies leñosas de interés
en las futuras condiciones predichas con el cambio climático, ya que el incremento en la
frecuencia de eventos de lluvia torrenciales puede incrementar la segregación de nicho entre
S. tenacissima y sus arbustos vecinos, pero no así en el caso de que la nodriza sea otro
arbusto.

217
CONCLUSIONES GENERALES
De los trabajos desarrollados en la presente tesis doctoral se extraen las siguientes conclusiones generales: 1) La tolerancia a la sombra es un factor clave a la hora de entender las interacciones planta-planta en medios semiáridos. La sombra generada por Stipa tenacissima ejerció un efecto negativo sobre Retama sphaerocarpa y Lepidium subulatum, ambas especies intolerantes a la sombra. 2) Los efectos de los cambios en el patrón de las precipitaciones predichos con el cambio climático varían dependiendo de las especies implicadas. En general, la reducción en las lluvias de primavera aumentó los efectos negativos sobre las especies intolerantes a la sombra. 3) El efecto competitivo de las especies herbáceas sobre las leñosas se reduce con la edad de éstas últimas, lo que sugiere la existencia de procesos de segregación de nicho. El aumento de los eventos de lluvia torrencial, predicho con el cambio climático, puede acelerar la segregación de nicho entre especies leñosas y herbáceas. 4) Los factores climáticos que influyen en las interacciones planta-planta a lo largo de su ontogenia presentan una heterogeneidad espacial marcada. Mientras que la lluvia fue el modulador fundamental de la interacción entre Lepidium y Stipa en la ladera de solana, otros factores climáticos fueron importantes en la ladera de umbría. 5) La herbivoría es un factor clave en las interacciones planta-planta, que puede llegar a compensar los efectos negativos derivados de la competencia por nutrientes (i.e. luz o agua). La reducción del impacto de la herbivoría promovida por el efecto protector de Stipa sobre Retama redujo la competencia entre ambas especies, dando como resultado un efecto neto positivo de Stipa sobre Retama. 6) La herbivoría y el estrés hídrico presentan interacciones complejas, existiendo una jerarquía entre ambos factores. La aridez por sí misma no tuvo un efecto importante en la interacción entre Stipa y Retama bajo niveles muy altos de herbivoría. Sin embargo, la aridez moduló indirectamente la presión de herbivoría afectando a la productividad vegetal, y por tanto, a la cantidad de forraje disponible para los herbívoros y a su impacto sobre la especie estudiada. 7) Existe una jerarquía entre las condiciones climáticas y la distancia filogenética entre los arbustos rebrotadores y sus vecinas a la hora de definir la interacción entre ambas. Distancias filogenéticas entre 207 y 273 millones de años produjeron siempre interacciones competitivas. Mientras que valores fuera de este rango dieron lugar a interacciones neutras o facilitativas, dependiendo de las condiciones climáticas. Ni la distancia filogenética ni el clima fueron factores clave entre la interacción entre Stipa y sus especies vecinas.

CONCLUSIONES
218
8) El efecto positivo de Stipa sobre Lepidium durante la germinación de éste último fue clave a la hora de definir el resultado neto de la interacción entre ambas especies. Esto quedó demostrado por el elevado grado de co-ocurrencia encontrado a pesar de la dominancia de interacciones negativas entre ambas especies a lo largo de la vida de Lepidium. 9) Las condiciones climáticas, interpretadas como un gradiente general de estrés, son predictores muy pobres del resultado de las interacciones entre pares de especies, ya que estas interacciones dependen de multitud de factores además del clima. Por tanto, ninguno de los modelos conceptuales dominantes en la actualidad predice una suficiente cantidad de estas interacciones como para ser aceptado de forma universal. 10) Las plantas nodriza aumentan la riqueza de especies y la diversidad filogenética a nivel local no sólo por la expansión de nicho, promovida por la mejora de las condiciones microclimáticas bajo su copa, sino también por un aumento de la segregación de nicho entre sus especies beneficiarias, derivado de una mayor heterogeneidad ambiental bajo su dosel. 11) La evolución del efecto positivo que las plantas nodriza ejercen sobre la riqueza local de especies a lo largo de gradientes ambientales depende de la naturaleza de dichos gradientes. Mientras que en España se mantuvo constante a lo largo de todo el gradiente estudiado (definido de forma opuesta por la aridez y las bajas temperaturas), este efecto aumentó con la aridez en Australia, donde la lluvia era el factor de estrés dominante en las comunidades estudiadas. En este último caso, se detectó un colapso del efecto positivo de las nodrizas bajo niveles extremos de aridez, como apuntan algunos modelos teóricos existentes. 12) Las condiciones climáticas y las interacciones bióticas interactúan a la hora de definir el patrón filogenético de las comunidades vegetales. Mientras que la facilitación aumentó la dispersión del patrón filogenético, condiciones climáticas “más benignas” (p. ej. más lluvia) incrementaron la agregación de este patrón mediante su efecto indirecto en la reducción de la diferenciación de nicho entre especies facilitadas y no facilitadas. El efecto conjunto de ambos factores generó un patrón filogenético aleatorio en la mayoría de comunidades estudiadas. Estos resultados advierten sobre la problemática derivada de inferir los mecanismos dominantes en el ensamblaje de una comunidad dada a partir únicamente de su patrón filogenético, aunque tengamos un conocimiento detallado del grado de conservación de importantes rasgos ecofisiológicos en las especies de esa comunidad a lo largo de la evolución.

BIBLIOGRAFÍA Y AFILIACIÓN DE LOS CO AUTORES

BIBLIOGRAFÍA
220

BIBLIOGRAFÍA
Ackerly, DD. 2003. Community assembly,
niche conservatism and adaptive evolution in changing environments. International Journal of Plant Sciences 164: S165–S184.
Acuña-Rodríguez I, LA Cavieres, and E Gianoli. 2006. Nurse effect in seedling establishment: facilitation and tolerance to damage in the Andes of central Chile. Revista Chilena de Historia Natural 79: 329–336.
Aguiar MR, and OE Sala. 1999 Patch structure, dynamics and implications for the functioning of arid ecosystems. Trends in Ecology and Evolution 14: 273–277.
Allan GJ, J Francisco-Ortega, A Santos-Guerra, E Boerner, and A Zimmer. 2004. Molecular phylogenetic evidence for the geographic origin and classification of Canary Islands Lotus (Fabaceae: Loteae). Molecular Phylogenetics and Evolution 32: 123–138.
Allan GJ, and JM Porter. 2000. Tribal delimitation and phylogenetic relationships of Loteae and Coronilleae (Faboideae: Fabaceae) with special reference to Lotus: evidence from nuclear ribosomal ITS sequences. American Journal of Botany 87: 1871-1881.
Anderson CL, K Bremer, and EM Friis. 2005. Dating phylogenetically basal eudicots using rbcL sequences and multiple fossil reference points. American Journal of Botany 92: 1737-1748.
Anderson MJ, RN Gorley, and KR Clarke. 2008. PERMANOVA+ for PRIMER: Guida to software and statistical methods. PRIMER-E, Plymouth, UK.
Andrés, P, and M Jorba. 2000. Mitigation strategies in some motorway embankments (Catalonia, Spain). Restoration Ecology 8: 268-275.
Angiosperm Phylogeny Group. 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botany Journal of the Linnean Society 161: 105–121.
Anthelme F, and R Michalet. 2009. Grass-to-tree facilitation in an arid grazed environment (Aïr Mountains, Sahara). Basic and Applied Ecology 10: 437-446.
Anthelme F, R Michalet, and M Saadou. 2007. Positive associations involving the tussock grass Panicum turgidum Forssk. In the Aïr-Ténéré Reserve, Niger. Journal of Arid Environments 68: 348–362.
Aragón CF, A Escudero, F. Valladares. 2008. Stress-induced dynamic adjustments of reproduction differentially affect fitness components of a semiarid plant. Journal of Ecology 96: 222–229.
Archer SR. 1995. The role of herbivores in mediating grass-woody plant interactions. Tropical Grasslands 29:218–235.
Archer SR, TW Boutton, and KA Hibbard. 2001. Trees in grasslands. In: Biogeochemical consequences of woody plant expansion in global biogeochemical cycles in the climate system. ED Schulze, SP Harrison, M Heimann, EA Holland, J Lloyd, IC Prentice, and D Schimel (Eds.). Academic press, San diego.
Armas C, R Ordiales, and FI Pugnaire. 2004. Measuring plant interactions: A new comparative index. Ecology 85: 2682–2686.
Armas C, and FI Pugnaire. 2005. Plant interactions govern population dynamics in a semi-arid plant community. Journal of Ecology 93: 978–989.
Armas C, and FI Pugnaire. 2009. Ontogenetic shifts in interactions of two dominant shrub species in a semi-arid coastal sand dune system. Journal of Vegetation Science 20: 535–546.
Aragón CF, A Escudero, and F Valladares . 2008. Stress-induced dynamic adjustments of reproduction differentially affect fitness components of a semiarid plant. Journal of Ecology 96: 222–229.
Arsenault R, and N Owen-Smith. 2002. Facilitation versus competition in grazing herbivore assemblages. Oikos 97: 313–318.
Badano EI, and LA Cavieres. 2006. Ecosystem engineering across ecosystems: do engineer species sharing common features have generalized or idiosyncratic effects on species diversity? Journal of Biogeography 33: 304–313.
Balaguer L, FI Pugnaire, E Martínez-Ferri, C Armas, F Valladares, and E Manrique . 2002. Ecophysiological significance of chlorophyll loss and reduced

BIBLIOGRAFÍA
222
photochemical efficiency under extreme aridity in Stipa tenacissima L. Plant and Soil 240: 343–352.
Baraza E, JM Gómez, JA Hódar, and R Zamora. 2004. Herbivory has a greater impact in shade than in sun: response of Quercus pyrenaica seedlings to multifactorial environmental variation. Canadian Jorunal of Botany 82: 357–364.
Baraza E, R Zamora, and JA Hódar. 2006. Conditional outcomes in plant-herbivore interactions: neighbours matter. Oikos 113: 148–156.
Barber A, MR Cabrera, and I Guardiola. 1997. Sobre la cultura de l'espart al territori valencià. Fundació Bancaixa, Spain.
Barberá GG, JA Navarro-Cano, VM Castillo. 2006. Seedling recruitment in a semi-arid steppe: The role of microsite and post-dispersal seed predation. Journal of Arid environments 67: 701–714.
Barchuk AH, A Valiente-Banuet, MP Díaz. 2005. Effect of shrubs and seasonal variability of rainfall on the establishment of Aspidosperma quebracho-blanco in two edaphically contrasting environments. Austral Ecology 30: 695–705.
Bascietto M., and G Scarascia-Mugnozza. 2004. A collection of functions to determine annual tree carbon increment via stem-analysis. Annals of Forest Science 61: 597–602.
Bascompte J. 2009. Disentangling the Web of Life. Science 325: 416–419.
Bascompte J, and MA Rodríguez. 2001. Habitat patchiness and plant species richness. Ecology Letters 4: 417–420
Baskin CC, and JM Baskin. 1998. Seeds: ecology, biogeography, and evolution of dormancy and germination. Academic Press, San Diego, USA.
Baumeister D, and RM Callaway. 2006. Facilitative effects of Pinus flexilis during succession: a hierarchy of mechanisms benefits other plant species. Ecology 87: 1816–1930.
Beadle NCW. 1948. The Vegetation and Pastures of Western New South Wales with Special Reference to Soil Erosion. Department of Conservation of NSW, Sydney.
Bello MA, A Bruneau, F Forest, and JA Hawkins. 2009. Elusive relationships within Order Fabales : Phylogenetic Analyses using matK and rbcL sequence data. Systematics and Botany 34: 102-114.
Bellot J, FT Maestre, and N Hernández. 2004. Spatio-temporal dynamics of chlorophyll fluorescence in a semi-arid shrubland. Journal of Arid Environments 58: 295–308.
Belnap J, and OL Lange (Eds). 2003. Biological Soil Crusts: Structure, Function, and Management, Springer-Verlag: Berlin.
Ben Salem H, A Nefzaoui, and H Abdouli. 1994. Palatability of shrubs and fodder trees measured on sheep and camels. CIHEAM Options Mediterraneennes 4: 35–48.
Bertness MD, and RM Callaway. 1994. Positive interactions in communities. Trends in Ecology and Evolution 9: 191–193.
Bertness MD, and SW Shumway. 1993. Competition and facilitation in marsh plants. American Naturalist 142: 718–724.
Bochet, E, and P García-Fayos. 2004. Factors controlling vegetation establishment and water erosion on motorway slopes in Valencia, Spain. Restoration Ecology 12:166-174.
Bolling, JD, and LR Walker. 2000. Plant and soil recovery along a series of abandoned desert roads. Journal of Arid environments 46:1-24.
Booth, DT, JK Gores, GE Schuman, and RA Olson. 1999. Shrub Densities on Pre-1985 Reclaimed Mine Lands in Wyoming. Restoration Ecology 7 :24-32.
Bouchenak-Khelladi Y, N Salamin, V Savolainen, F Forest, M van der Bank, MW Chase, and TR Hodkinson. 2008. Large multi-gene phylogenetic trees of the grasses (Poaceae): Progress towards complete tribal and generic level sampling. Molecular and Phylogenetic Evolution 47: 488-505.
Bouchenak-Khelladi Y, GA Verboom, V Savolainen, and TR Hodkinson. 2010. Biogeography of the grasses (Poaceae): A phylogenetic approach to reveal evolutionary history in geographical space and geological time. Biological Journal of the Linnean Society 162: 543-557.
Bowker MA, S Soliveres, and FT Maestre. 2010. Competition increases with abiotic stress and regulates the diversity of biological soil crusts. Journal of Ecology 98: 551–560.
Brady KU, AR Kruckeberg, and HD Bradshaw Jr. 2005. Evolutionary ecology of plant adaptation to serpentine soils. Annual Review of Ecology, Evolution, and Systematics 36: 243–266.
Bremer K, EM Friis, and B Bremer. 2004. Molecular phylogenetic dating of asterid flowering plants shows Early Cretaceous diversification. Systematics and Biology 53: 496–505.
Bremer, B., and Eriksson, O. (2009). Time tree of Rubiaceae: Phylogeny and dating the

223
family, subfamilies and tribes. Int. J. Plant Sci., 170, 766-793.
Briske DD, SD Fuhlendorf, and FE Smeins. 2003. Vegetation dynamics on rangelands: a critique of the current paradigms. Journal of Applied Ecology 40: 601–614.
Brooker RW. 2006. Plant–plant interactions and environmental change. New Phytologist 171: 271–289.
Brooker RW, and TV Callaghan. 1998. The balance between positive and negative plant interactions and its relationship to environmental gradients: a model. Oikos 81: 196–207.
Brooker RW, Z Kikvidze, FI Pugnaire, RM Callaway, P Choler, CL Lortie, and R Michalet. 2005. The importance of importance. Oikos 109: 63–70.
Brooker RW, FT Maestre, RM Callaway, CL Lortie, LA Cavieres, G Kunstler, P Liancourt, K Tielbörger, JMJ Travis, F Anthelme, C Armas, L Coll, E Corcket, S Delzon, E Forey, Z Kikvidze, J Olofsson, FI Pugnaire, P Saccone, K Schiffer, M Seifan, B Touzard, and R Michalet. 2008. Facilitation in plant communities: the past, the present, and the future. Journal of Ecology 96: 18–34.
Brown JR, and SR Archer. 1990. Water relations of a perennial grass and seedling versus adult woody plants in a subtropical savanna, Texas. Oikos 57: 366–374.
Bruno JF, JJ Stachowicz, and MD Bertness. 2003. Inclusion of facilitation into ecological theory. Trends in Ecology and Evolution 18: 119–125.
Burke, MJW, and JP Grime. 1996. An Experimental Study of Plant Community Invasibility. Ecology 77: 776-790.
Butterfield BJ. 2009 Effects of facilitation on community stability and dynamics: synthesis and future directions. Journal of Ecology 97: 1192–1201.
Butterfield BJ, JL Betancourt, RM Turner, and JM Briggs. 2010. Facilitation drives 65 years of vegetation change in the Sonoran Desert. Ecology 91: 1132–1139.
Caballero I. 2006. Estructura espacio-temporal de un banco de semillas: Las comunidades gipsícolas del centro de la Península Ibérica. PhD Thesis.
Caballero I, JM Olano, J Loidi, and A Escudero. 2008. A model for small-scale seed bank and standing vegetation connection along time. Oikos 117: 1788–1795.
Caballero I, JM Olano, A Escudero, and J Loidi. 2008. Seed bank spatial structure in semi-arid environments: beyond the patch-
bare area dichotomy. Plant Ecology 195: 215–223.
Cahill JF, SW Kembel, EG Lamb, and PA Keddy. 2008. Does phylogenetic relatedness influence the strength of competition among vascular plants? Perspectives in Plant Ecology, Evolution, and Systematics 10: 41–50.
Callaway RM. 1995. Positive interactions among plants. The Botanical Review 61: 306–349.
Callaway RM. 1997. Positive interactions in plant communities and the individualistic-continuum concept. Oecologia 112: 143–149.
Callaway RM. 2007. Positive Interactions and Interdependence in Plant Communities. Springer; New York.
Callaway RM, and ET Aschehoug. 2000. Invasive plants versus their new and old neighbors: a mechanism for exotic invasion. Science 290: 521–523.
Callaway RM, RW Brooker, P Choler, Z Kikvidze, CJ Lortie, R Michalet, L Paolini, FI Pugnaire, B Newingham, ET Aschehoug, C Armas, D Kikodze, and BJ Cook. 2002. Positive interactions among alpine plants increase with stress. Nature 417: 844–888.
Callaway RM, D Kikodze, and Z Kikvidze. 2000. Facilitation by unpalatable weeds may conserve plant diversity in overgrazed meadows in the Caucasus Mountains. Oikos 89:275–282.
Callaway RM, and LR Walker. 1997. Competition and facilitation: A synthetic approach to interactions in plant communities. Ecology 78: 1958–1965.
Caravaca MF, MM Alguacil, D Barea, JM Navarro, JM Roldán, and A Garrigos. 2003. Re-establishment of Retama sphaerocarpa as a target species for reclamation of soil physical and biological properties in a semi-arid Mediterranean area. Forest Ecology and Management 182: 49–58.
Castillo JP, M Verdú, and A Valiente-Banuet. 2010. Neighborhood phylodiversity affects plant performance. Ecology (in press)
Castillo-Monroy, AP, FT Maestre, M Delgado-Baquerizo, and A Gallardo. 2010. Biological soil crusts modulate nitrogen availability in semi-arid ecosystems: Insights from a Mediterranean grassland. Plant and Soil 333: 21-34.
Castro J, R Zamora, JA Hódar, and JM Gómez. 2002. The use of shrubs as nurse plants: a new technique for reforestation in Mediterranean mountains. Restoration Ecology 10: 297-305.

BIBLIOGRAFÍA
224
Cavender-Bares J, DA Ackerly, D Baum and FA Bazzaz. 2004. Phylogenetic overdispersion in Floridian oak communities, American Naturalist, 163: 823-843.
Cavender-Bares J, and FA Bazzaz. 2000. Changes in drought response strategies with ontogeny in Quercus rubra: Implications for scaling from seedlings to mature trees. Oecologia 124: 8–18.
Cavender-Bares J, A Keen, and B Miles. 2006. Phylogenetic structure of Floridian plant communities depends on taxonomic and spatial scale. Ecology 87: S109-S122.
Cavender-Bares J, K. Kozak, P Fine, and S Kembel. 2009. The merging of community ecology and phylogenetic biology. Ecology Letters 12: 693–715.
Cavieres LA, and EI Badano. 2009. Do facilitative interactions increase species richness at the entire community level? Journal of Ecology 97: 1181–1191.
Chao A, RL Chazdon, RK Colwell, and T Shen. 2005. A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecology letters 8: 148–159.
Chapin III, FS, AJ Bloom, CB Field, and RH Waring. 1987. Plant responses to multiple environmental factors. BioScience 37: 49–57.
Chapin III, FS, ED Schulze, and HA Mooney. 1990. The ecology and economics of storage in plants. Annual Review in Ecology and Systematics 21: 423–447.
Chase JM, MA Leibold, MA, AL Downing, and JB Shurin. 2000 The effects of productivity, herbivory, and plant species turnover in grassland food webs. Ecology 81: 2485–2497.
Choler Ph, R Michalet, and RM Callaway. 2001. Facilitation and competition on gradients in alpine plant communities. Ecology 82: 3295–3308.
Chu CJ, YS Wang, FT Maestre, and G Wang. 2008. On the relevance of facilitation in alpine meadow communities: An experimental assessment with multiple species differing in their ecological optimum. Acta Oecologica 33: 108–113.
Clarke KR, and RM Warwick. 2001. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. 2nd edition: PRIMER-E, Plymouth, UK.
Convención para la Lucha contra la Desertificación (CLD) 2005. Economic opportunities in the drylands under the united nations convention to combat desertification:
http://www.unccd.int/cop/cop7/docs/ss_background-eng.pdf.
Clements FE. 1916. Plant succession: An analysis of the development of vegetation. Washington, DC: Carnegie Institute. Washington.
Crawley MJ. 1998. Herbivory. In: Physiological plant ecology, MC Press et al. (Eds.), Blackwell Scientific Publications, Germany.
Colwell RK. 2000 EstimateS: statistical estimation of species richness and shared species from samples. V 8.2.0. Dept. of Ecol. and Evol. Biol., Univ. of Connecticut, USA. http://viceroy.eeb.uconn.edu/estimates.
Connor EF, and D Simberloff. 1979. The assembly of species communities: chance or competition? Ecology 60: 1132–1140.
Cook ER, and RL Holmes. 1996. Guide for computer program ARSTAN. In: Grissino-Mayer HD, Holmes RL, Fritts HC, eds. The international treering data bank program library version 2.0 user’s manual. Tucson, AZ, USA: Laboratory of Tree-Ring Research, University of Arizona. 75–87.
Cortina J, and FT Maestre. 2005. Plant effects on soils in drylands: implications on community dynamics and ecosystem restoration. In: Tree Species effects in Soils: Implications for Global Climate Change. Binkley D, and O. Menyailo (Eds.). NATO Science Series, Kluwer Academic Publishers, Dordrecht.
Cortina J, FT Maestre, and D Ramírez. 2010. Innovations in semiarid restoration. The case of Stipa tenacissima L. grass steppes. In: Bautista S, J Aronson, and R Vallejo (Eds.). Land Restoration to Combat Desertification: Innovative Approaches, Quality Control and Project Evaluation. Fundación CEAM, Spain.
Cortina J, FT Maestre, R Vallejo, MJ Baeza, A Valdecantos, and M Pérez-Devesa. 2005. Ecosystem structure, function, and restoration success: Are they related? Journal of Nature Conservation 14: 152–160.
Cortina J, RM Navarro, and A del Campo. 2006. Evaluación del éxito de la reintroducción de especies leñosas en ambientes mediterráneos. In: Cortina J, JL Peñuelas, J Puértolas, R Savé, and A Vilagrosa (Eds.). Calidad de planta forestal para la restauración de ambientes mediterráneos. Ministerio de Medio Ambiente, Spain.
Daleo P, and O Iribarne. 2009. Beyond competition: the stress-gradient hypothesis tested in plant– herbivore interactions. Ecology 90: 2368–2374

225
Darwin C. 1859. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. John Murray, London.
Davis MA, KJ Wrage, PB Reich, MG Tjoelker, T Schaeffer, C Muermann. 1999. Survival, growth, and photosynthesis of tree seedlings competing with herbaceous vegetation along a water-light-nitrogen gradient. Plant Ecology 145: 341–350.
Dawson, TE. 1993. Hydraulic lift and water use by plants: implications for water balance, performance and plant-plant interactions. Oecologia 95: 565-574.
De´ath G, and KE Fabricius. 2000. Classification and regression trees: a powerful yet simple technique for ecological data analysis. Ecology 81: 3178-3192.
de la Cruz, M, RL Romao, A Escudero, FT Maestre. 2008. Where do seedlings go? A spatio-temporal analysis of seedling mortality in a semi-arid gypsophyte. Ecography 31: 720–730.
Diamond JM. 1975. Assembly of species communities. In: Ecology and evolution of communities, Cody ML, and JM Diamond (Eds), Harvard Press, USA.
Dullinger S, I Kleinbauer, H Pauli, M Gottfried, R Brooker, L Nagy, JP Theurillat, JI Holten, O Abdaladze, JL Benito, JL Borel, G Coldea, D Ghosn, R Kanka, A Merzouki¸ C Klettner, P Moiseev, U Molau, K Reiter, G Rossi, A Stancisi, M Tomaselli¸ P Unterlugauer, P Vittoz, and G Grabherr. 2007. Weak and variable relationships between environmental severity and small-scale co-occurrence in alpine plant communities. Journal of Ecology 95: 1284–1295.
Dyer ME, and RL Bailey. 1987. A Test of Six Methods for Estimating True Heights from Stem Analysis Data. Forest Science 33: 3–13.
Eccles N, KJ Esler, and RM Cowling. 1999. Spatial pattern analysis in Namaqualand desert plant communities: evidence for general positive interactions. Plant Ecology 142: 71–85.
Eldridge DJ, M Westoby, and KG Holbrook. 1991. Soil-surface characteristics, microtopography and proximity to mature shrubs: effects on survival of several cohorts of Atriplex vesicaria seedlings. Journal of Ecology 79: 357–364.
Eldridge DJ, and D Freudenberger. 2005. Ecosystem wicks: woodland trees enhance water flow in a fragmented agricultural landscape in eastern Australia. Austral Ecology 30: 336–347
Eliason SA, and EB Allen. 1997. Exotic grass competition in supressing native shrubland re-establishment. Restoration Ecology 5: 244–255.
Engqvist L. 2005. The mistreatment of covariate interaction terms in linear model analyses of behavioral and evolutionary ecology studies. Animal Behavior 70: 967–971.
Escudero A, JM Iriondo, JM Olano, A Rubio, and R Somolinos. 2000. Factors affecting the establishment of a gypsophyte: The case of Lepidium subulatum. American Journal of Botany 87: 861–871.
Escudero A, RC Somolinos, JM Olano, and A Rubio. 1999. Factors controlling the establishment of Helianthemum squamatum, an endemic gypsophile of semiarid Spain. Journal of Ecology 87: 290–302.
Espigares T, A López-Pintor, and JM Rey-Benayas. 2004. Is the interaction between Retama sphaerocarpa and its understorey herbaceous vegetation always reciprocally positive? Competition-facilitation shift during Retama sphaerocarpa establishment. Acta Oecologica 26: 121–128.
Fahey TJ, JJ Battles, GF Wilson. 1998. Responses of early successional northern hardwood forests to changes in nutrient availability. Ecological Monographs 68: 183–212.
Fischer RA, and NC Turner. 1978. Plant productivity in the arid and semi-arid zones. Annual Review in Plant Physiology 29: 277–317.
Flombaum, P., and O. E. Sala. 2009. Cover is a good predictor of biomass in arid systems. Journal of Arid Environments 73: 597–598.
Forest F, R Grenyer, M Rouget, TJ Davies, RM Cowling, DP Faith, A Balmford, JC Manning, S Proches, and M van der Bank. 2007. Preserving the evolutionary potential of floras in biodiversity hotspots. Nature 445: 757–760.
Forey E, B Touzard, and R Michalet. 2009. Does disturbance drive the collapse of biotic interactions at the sever end of a diversity-biomass gradient? Plant ecology 206: 287–295.
Forseth IN, DA Wait, and BB Casper. 2001. Shading by shrubs in a desert system reduces the physiological and demographic performance of an associated herbaceous perennial. Journal of Ecology 89: 670–680.
Fowler N. 1986. The role of competition in plant communities in arid and semiarid

BIBLIOGRAFÍA
226
regions. Annual Review of Ecology and Systematics 17: 89–110.
Fowler N. 2002. The joint effects of herbivory, competition, and topographic position upon six grasses. Ecology 83: 2477–2488.
Franco AC, and PS Nobel. 1989. Effect of nurse plants on the microhabitat and growth of cacti. Journal of Ecology 77: 870–886.
Franzke A, D German, IA Al-Shehbaz, and K Mummenhoff. 2009. Arabidopsis family ties: Molecular phylogeny and age estimates in Brassicaceae. Taxon 58: 425-437.
Friedman JJ, G Orshan, and Y Ziger-Cfir. 1977. Suppression of Annuals by Artemisia herba-alba in Negev Desert of Israel. Journal of Ecology 65: 413–426
Fuhlendorf SD, DD Briske, and FE Smeins. 2001. Herbaceous vegetation change in variable rangeland environments: the relative contribution of grazing and climatic variability. Applied Vegetation Science 4: 177–188.
Funk VA, R Bayer, S Keeley, R Chan, L Watson, B Gemeinholzer, E Schilling, J Panero, B Baldwin, NT García-Jacas, A Susanna, and RK Jansen. 2005. Everywhere but Antarctica: Using a supertree to understand the diversity and distribution of the Compositae. Biol. Skrifter 55: 343-374.
Gachet S, E Véla, and T Tatoni. 2005. BASECO: a floristic and ecological database of Mediterranean French flora. Biodiversity and Conservation 14: 1023–1034.
García-Palacios P. 2010. Interacciones planta-suelo en taludes de carretera: implicaciones para la restauración. PhD Thesis.
García-Palacios P, S Soliveres, FT Maestre, A Escudero, A Castillo-Monroy, and F Valladares. 2010. Dominant plant species modulate responses to hydroseeding, irrigation and fertilization during the restoration of semiarid motorway slopes. Ecological Engineering, (In press).
Gasque M, and P García-Fayos. 2004. Interaction between Stipa tenacissima and Pinus halepensis: consequences for reforestation and the dynamics of grass steppes in semiarid slopes of western Mediterranean areas. Forest Ecology and Management 189: 251–261
Gillman LN, and SD Wright. 2006. The influence of productivity on the species richness of plants: a critical assessment. Ecology 87: 1234–1243.
Gilpin ME. 1975. Limit cycles in competition communities. American Naturalist 109: 51–60.
Gleason HA. 1926. The Individualistic Concept of the Plant Association. Bulletin of the Torrey Botanical Club 53: 7–26.
Goldberg DE, and A Novoplansky. 1997. On the relative importance of competition in unproductive environments. Journal of Ecology 85: 409–418.
Goldberg DE, R Turkington, L Olsvig-Whittaker, and AR Dyer. 2001. Density dependence in an annual plant community: variation among life history stages. Ecological Monographs 71: 423–446.
Gómez-Aparicio L. 2009. The role of plant interactions in the restoration of degraded ecosystems: a meta-analysis across life-forms and ecosystems. Journal of Ecology 97: 1202–1214.
Gómez-Aparicio L, JM Gómez, and R Zamora. 2005. Microhabitats shift rank in suitability for seedling establishment depending on habitat type and climate. Journal of Ecology 93: 1194–1202.
Gómez-Aparicio L, F Valladares, and R Zamora. 2006. Differential light responses of Mediterranean tree saplings: linking ecophysiology with regeneration niche in four co-occurring species. Tree Physiology 26: 947–958.
Gómez-Aparicio L, R Zamora, JM Gómez, JA Hódar, J Castro, and E Baraza. 2004. Applying plant facilitation to forest restoration in Mediterranean ecosystems: a meta-analysis of the use of shrubs as nurse plants. Ecological Applications 14: 1128–1138.
Gotelli NJ. 2000. Null model analysis of species co-occurrence patterns. Ecology 81: 2606–2621.
Gotelli NJ, and GL Entsminger. 2006. EcoSim: Null Models Software for Ecology, Version 7. Acquired Intelligence Inc. and Kesey-Bear. Available at: http://garyentsminger.com/ecosim.htm.
Gotelli NJ, and GR Graves. 1996. Null Models in Ecology, Smithsonian Institution Press, Washington, DC.
Gotelli NJ, GR Graves, and C Rahbek. (2010). Macroecological signals of species interactions in the Danish avifauna. Proceedings of the National Academy of Sciences USA (In press).
Graff P, MR Aguiar, and EJ Chaneton. 2007. Shifts in positive and negative plant interactions along a grazing intensity gradient. Ecology 88: 188–199.

227
Grace JB. 1999. The factors controlling species density in herbaceous plant communities: an assessment. Perspectives in Plant Ecology, Evolution and Systematics 2: 1–28.
Grace JB, GR Guntenspergen, and JR Keough. 1993. The examination of a competition matrix for transitivity and intransitive loops. Oikos 68: 91–98.
Greiner La Peyre MK, JB Grace, E Hahn, and IA Mendelssohn. 2001. The importance of competition in regulating species abundance along a salinity gradient. Ecology 82: 62–69.
Grime JP. 1973. Competitive exclusion in herbaceous plant communities. Nature 242: 344–347.
Grime JP. 2001. Plant Strategies, Vegetation Processes, and Ecosystem Properties, 2nd edn.Wiley, Chichester, UK.
Grissino-Mayer HD. 2001. Evaluating crossdating accuracy: a manual and tutorial for the computer program COFECHA. Tree-Ring Research 57: 205–221.
Gross N, P Liancourt, Ph Choler, KN Suding, and S Lavorel. 2010. Strain and vegetation effects on local limiting resources explain the outcomes of biotic interactions. Perspectives in Plant Ecology, Evolution and Systematics 12: 9–19.
Guisan A, and NE Zimmermann. 2000. Predictive habitat distribution models in ecology. Ecological Modelling 135: 147-186.
Gurevitch J, JA Morrison, and LV Hedges. 2000. The interaction between competition and predation: a meta-analysis of field experiments. American Naturalist 155: 435–453.
Guzmán B, and P Vargas. 2009. Historical biogeography and character evolution of Cistaceae (Malvales) based on analysis of plastid rbcL and trnL-trnF sequences. Organisms Divers. Evolution 9: 83-99.
Guzmán B, MD Lledó, P Vargas. 2009. Adaptive radiation in Mediterranean Cistus (Cistaceae). Plos ONE, 4, e6362
Hacker SD, and SD Gaines. 1997. Some implications of direct positive interactions for community species diversity. Ecology 78: 1990–2003.
Hacker SD, and MD Bertness. 1999. The role of positive interactions in the plant species diversity of salt marsh plant communities. Ecology 80: 2064–2073.
Hastwell GT, and JM Facelli. 2003. Differing effects of shade-induced facilitation on growth and survival during the
establishment of a chenopod shrub. Journal of Ecology 91: 941–950.
Hay ME. 1986. Associational plant defenses and the maintenance of species diversity: turning competitors into accomplices. The American Naturalist 128: 617–641.
Hector A, B Schmid, C Beierkuhnlein, CMC Caldeira, M Diemer, PG Dimitrakopoulos, JA Finn, H Freitas, PS Giller, J Good, R Harris, P Högberg, K Huss-Danell, J Joshi, A Jumpponen, C Körner, PW Leadley, M Loreau, A Minns, CPH Mulder, G O'Donovan, SJ Otway, JS Pereira, A Prinz, DJ Read, M Scherer-Lorenzen, ED Schulze, ASD Siamantziouras, EM Spehn, AC Terry, AY Troumbis, FI Woodward, S Yachi, and JH Lawton. 1999. Plant diversity and productivity experiments in European grasslands. Science 286: 1123–1127.
Hedges SB, J Dudley, and S Kumar. 2006. TimeTree: a public knowledge-base of divergence times among organisms. Bioinformatics 22: 2971-2972.
Heisler-White JL, JM Blair, EF Kelly, K Harmoney, and AK Knapp. 2009 Contingent productivity responses to more extreme rainfall regimes across a grassland biome. Global Change Biology 15: 2894–2904.
Herrera CM. 1984. Tipos morfológicos y funcionales en plantas leñosas del matorral mediterráneo del sur de España. Studia Oecologica 5: 7–34.
Herrera CM. 1992. Historical effects and sorting processes as explanations of contemporary ecological patterns: character syndromes in Mediterranean woody plants. The American Naturalist 140: 421-446.
Hobbs RJ, S Arico, J Aronson, JS Baron, P Bridgewater, VA Cramer, PR Epstein, JJ Ewel, CA Klink, AE Lugo, D Norton, D Ojima, DM Richardson, EW Sanderson, F Valladares, M Vilá, R Zamora, and M Zobel. 2006. Novel ecosystems: theoretical and management aspects of the new ecological world order. Global Ecology and Biogeography 15: 1–7.
Holmgren M, and M Scheffer. 2010. Strong facilitation in mild environments: the stress gradient hypothesis revisited. Journal of Ecology (In press). doi: 10.1111/j.1365-2745.2010.01709.x
Holmgren M, M Scheffer, and MA Huston. 1997. The interplay of facilitation and competition in plant communities. Ecology 78: 1966–1975.
Holmgren M, P Stapp, CR Dickman, C Gracia, S Graham, JR Gutiérrez, C Hice, F Jaksic,

BIBLIOGRAFÍA
228
DA Kelt, M Letnic, M Lima, BC López, PL Meserve, WB Milstead, GA Polis, MA Previtali, M Richter, S Sabaté, and FA Squeo. 2006. Extreme climatic events shape arid and semiarid ecosystems. Frontiers in Ecology and the Environment 4: 87–95.
Holzapfel C, and BE Mahall. 1999. Bidirectional facilitation and interference between shrubs and annuals in the mojave desert. Ecology 80: 1747–1761.
Hooper DU, FS Chapin III, JJ Ewel, A Hector, P Inchausti, S Lavorel, JH Lawton, D Lodge, M Loreau, S Naeem, B Schmid, H Setälä, AJ Symstad, J Vandermeer, and DA Wardle. 2005. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecological Monographs 75: 3–35.
Hubbell SP. 2001. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton University Press, USA.
Huston MA. 1979. A general hypothesis of species diversity. American Naturalist 113: 81–101.
Huston MA. 1999. Local processes and regional patterns: appropriate scales for understanding variation in the diversity of plants and animals. Oikos 86: 393–401.
Huxman TE, JM Cable, DD Ignace, AJ Eilts, N English, J Weltzin, and DG Williams. 2004. Response of net ecosystem gas exchange to a simulated precipitation pulse in a semiarid grassland: the role of native versus non-native grasses and soil texture. Oecologia 141:295–305.
Ibáñez I, JS Clark, S LaDeau, and J Hille Ris Lambers. 2007. Exploiting temporal variability to understand tree recruitment response to climate change. Ecological Monographs 77: 163–177.
Ibañez I, and EW Schupp. 2001. Positive and negative interactions between environmental conditions affecting Cercocarpus ledifolius seedling survival. Oecologia 129: 543–550.
Illius AW, and TG O’Connor. 1999. On the relevance of non-equilibrium concepts to arid and semi-arid grazing systems. Ecological Applications 9: 798–813.
IPCC. 2007. Climate Change 2001: Impacts, Adaptation and Vulnerability. International Panel of Climate Change, Geneva, Switzerland
Isbell RF. 1996. The Australian soil classification. CSIRO Publishing. Collingwood, Australia.
Izco J. 1984. Madrid Verde. Ministerio de Agricultura, Pesca y Alimentación, Spain.
Janssens SB, EB Knox, S Huysmans, EF Smets, and V Merckx. 2009. Rapid radiation of Impatiens (Balsaminaceae) during Pliocene and Pleistocene: Result of a global climate change. Molecular Phylogenetics and Evolution 52: 806–824.
Jones CG, JH Lawton, and M Shachak. 1997. Positive and negative effects of organisms as physical ecosystem engineers. Ecology 78: 1946–1957.
Jorba M, and RVallejo. 2008. The ecological restoration of quarries: a case with application of organic amendment and irrigation. Ecosistemas 17: 119-132.
Kawai T, and M. Tokeshi. 2007. Testing the facilitation-competition paradigm under the stress-gradient hypothesis: decoupling multiple stress factors. Proceedings of the Royal Society London B 274: 2503–2508.
Kéfi S, M Rietkerk, CL Alados, Y Pueyo, A ElAich, V Papanastasis, and PC de Ruiter. 2007. Spatial vegetation patterns and imminent desertification in Mediterranean arid ecosystems. Nature. 449: 213–217.
Keith D. 1998. Ocean shores to desert dunes: The native vegetation of NSW and the ACT. Department of Environment and Climate Change, Australia.
Kembel SW, PD Cowan, MR Helmus, WK Cornwell, H Morlon, DD Ackerly, SP Blomberg, and CO Webb. 2010. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 26: 1463-1464.
Kikvidze Z. 1996. Neighbour interaction and stability in subalpine meadow communities. Journal of Vegetation Science 7: 41–44.
Kikvidze Z, L Khetsuriana, D Kikodze, and RM Callaway. 2006. Seasonal shifts in competition and facilitation in subalpine plant communities of the central Caucasus. Journal of Vegetation Science 17: 77–82.
Kikvidze Z, FI Pugnaire, RW Brooker, Ph Choler, CJ Lortie, R Michalet, and RM Callaway. 2005. Linking patterns and processes in alpine plant communities: a global study. Ecology 86: 1395–1400.
Kim KJ, KS Choi, and RK Jansen. 2005. Two chloroplast DNA inversions originated simultaneously during the early evolution of the sunflower family (Asteraceae). Molecular Biology and Evolution 22: 1783–1792.
Kitzberger T, DF Steinaker, and TT Veblen. 2000. Effects of climatic variability on facilitation of tree establishment in northern Patagonia. Ecology 81: 1914–1924.
Knapp AK, C Beier, DD Brisker DD, AT Classen, Y Luo, M Reichstein, MD Smith,

229
SD Smith, JE Bell, PA Fay, JL Heisler, SW Leavitt, R Sherry, B Smith, and E Weng. 2008. Consequences of more extreme precipitation regimes for terrestrial ecosystems. BioScience 58: 811–821.
Knoop WT, and BH Walker. 1985. Interactions of woody and herbaceous vegetation in a Southern African savanna. Journal of Ecology 73: 235–253.
Kobe RK, SW Pacala, JA Silander, and CD Canham. 1995. Ecological Applications 5: 517–532.
Körner C. 2003: Limitation and stress - always or never? Journal of Vegetation Science 14: 141–143.
Körner C. 2004. Individuals have limitations, not communities: A response to Marrs, Weiher and Lortie et al. Journal of Vegetation Science 15: 581–582.
Kraft NJB, and D Ackerly. 2010. Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonian forest. Ecological Monographs 80: 401–422
Kraft NJB, WK Cornwell, CO Webb, and DD Ackerly. 2007. Trait evolution, community assembly and the phylogenetic structure of ecological communities. American Naturalist 170: 271–283.
Kurashige NS, and AA Agrawal. 2005. Phenotypic plasticity to light competition and herbivory in Chenopodium album (Chenopodiaceae). American Journal of Botany 92: 21–26.
Laird RA, and BS Schamp. 2006. Competitive intransitivity promotes species coexistence. American Naturalist 168: 182–193.
Laird RA, and BS Schamp. 2008. Does local competition increase the coexistence of species in intransitive networks? Ecology 89: 237–247.
Lavin M, PS Herendeen, and MF Wojciechowski. 2005. Evolutionary rates analysis of Leguminosae implicates a rapid diversification of lineages during the Tertiary emergence. Systematics and. Biology 54: 575-594.
Le Houérou HN. 1986. The desert and arid zones of Northern Africa. In: Ecosystems of the world 12B. Hot deserts and arid shrublands. Evenary, M, I Noy-Meir, and DW Goodall (Eds). B, Elsevier, Amsterdam.
Le Houérou HN. 2001. Biogeography of the arid steppeland north of the. Sahara. Journal of Arid Environments 48: 103–128.
Le Roux PC, and MA McGeoch. 2010. Interaction intensity and importance along two stress gradients: adding shape to the
stress-gradient hypothesis. Oecologia 162: 733–745.
Levin SA. 1970. Community equilibria and stability, and an extension of the competitive exclusion principle. American Naturalist 104: 413–423
Levine JM. 1999. Indirect facilitation: evidence and predictions from a riparian community. Ecology 80: 1762–1769.
Liancourt P, RM Callaway, and R Michalet. 2005. Stress tolerance and competitive-response ability determine the outcome of biotic interactions. Ecology 86: 1611–1618.
Loiselle BA, CA Howell, CH Graham, JM Goerck, T Brooks, KG Smith, and PH Williams. 2003. Avoiding pitfalls of using species distribution models in conservation planning. Conservation Biology 17: 1591–1600.
López BC, M Holmgren, S Sabaté, CA Gracia. 2008. Estimating annual rainfall threshold for establishment of tree species in water-limited ecosystems using tree-ring data. Journal of Arid Environments 72: 602–611.
Lortie CJ. 2010. Synthetic analysis of the stress-gradient hypothesis. In: Positive Plant Interactions and Community Dynamics. Pugnaire FI (Ed.). C.R.C. Press, USA.
Lortie CJ, RW Brooker, Ph Choler, Z Kikvidze, R Michalet, FI Pugnaire, and RM Callaway. 2004a. Rethinking plant community theory. Oikos 107: 433–438.
Lortie CJ, RW Brooker, Z Kikvidze, and RM Callaway. 2004b. The value of stress and limitation in an imperfect world: a reply to Körner. Journal of Vegetation Science 15: 577–580.
Lortie CJ, and RM Callaway. 2006. Re-analysis of meta-analysis: support for the stress-gradient hypothesis. Journal of Ecology 94: 7–16.
Ludwig F, TE Dawson, HHT Prins, F Berendse, and H de Kroon. 2004. Below-ground competition between trees and grasses may overwhelm the facilitative effects of hydraulic lift. Ecology letters 7: 623–631.
Ludwig JA, and DJ Tongway. 1995. Spatial organization of landscapes and its function in semi-arid woodlands, Australia. Landscape Ecology 10: 51–63
Ludwig JA, and DJ Tongway. 1996. Rehabilitation of semi-arid landscapes in Australia II: restoring productive soil patches. Restoration Ecology 4: 398-406.
Maestre FT. 2002. La restauración de la cubierta vegetal en zonas semiáridas en función del patrón espacial de factores bióticos y abióticos. PhD Thesis.

BIBLIOGRAFÍA
230
Maestre FT, S Bautista, J Cortina, and J Bellot. 2001. Potential of using facilitation by grasses to establish shrubs on a semiarid degraded steppe. Ecological Applications 11: 1641–1655.
Maestre FT, S Bautista, and J Cortina. 2003. Positive, negative and net effects in grass-shrub interactions in semiarid Mediterranean steppes. Ecology 84: 3186–3197.
Maestre FT, MA Bowker, C Escolar, MD Puche, S Soliveres, S Mouro, P García-Palacios, AP Castillo-Monroy, I Martínez, and A Escudero. 2010. Do biotic interactions modulate ecosystem functioning along abiotic stress gradients? Insights from semi-arid Mediterranean plant and biological soil crust communities. Philosophical Transactions of the Royal Society B 365: 2057–2070.
Maestre FT, RM Callaway, F Valladares, and CJ Lortie. 2009a. Refining the stress-gradient hypothesis for competition and facilitation in plant communities. Journal of Ecology 97: 199–205.
Maestre FT, and J Cortina. 2004. Do positive interactions increase with abiotic stress? A test from a semi-arid steppe. Proceedings of the Royal Society of London, Series B (Supplement) 271: S331–S333.
Maestre FT, and J Cortina. 2004b. Are Pinus halepensis afforestations useful as a restoration tool in semiarid Mediterranean areas? Forest ecology and management 198: 303–317.
Maestre FT, and J Cortina. 2004c. Insights on ecosystem composition and function in a sequence of degraded semiarid steppes. Restoration Ecology 12: 494–502
Maestre FT, J Cortina, and S Bautista. 2004. Mechanisms underlying the interaction between Pinus halepensis and the native late-successional shrub Pistacia lentiscus in a semiarid plantation. Ecography 27: 776–786.
Maestre FT, and J Cortina. 2005. Remnant shrubs in Mediterranean semi-arid steppes: effects of shrub size, abiotic factors and species identity on understorey richness and occurrence. Acta Oecologica 27: 161–169.
Maestre FT, C Escolar, I Martínez, and A Escudero. 2008. Are soil lichen communities structured by biotic interactions? A null model analysis. Journal of Vegetation Science 19: 261–266.
Maestre FT, and A Escudero. 2009. Is the patch-size distribution of vegetation a
suitable indicator of desertification processes? Ecology 90: 1729-1735.
Maestre FT, MD Puche, MA Bowker, MB Hinojosa, I Martínez, P García-Palacios, A P Castillo, S Soliveres, AL Luzuriaga, AM Sánchez, JA Carreira, AGallardo, and A Escudero. 2009. Shrub encroachment can reverse desertification in Mediterranean semiarid grasslands. Ecology Letters 12: 930–941.
Maestre FT, DA Ramírez, and J Cortina. 2007. Ecología del esparto (Stipa tenacissima L.) y los espartales de la Península Ibérica. Ecosistemas 16: 117–136.
Maestre FT, F Valladares, and JF Reynolds. 2005. Is the change of plant-plant interactions with abiotic stress predictable? A meta-analysis of field results in arid environments. Journal of Ecology 93: 748–757.
Maestre FT, F Valladares, and JF Reynolds. 2006. The stress-gradient hypothesis does not fit all relationships between plant–plant interactions and abiotic stress: further insights from arid environments. Journal of Ecology 94: 17–22.
Magallón S, and A Castillo. 2009. Angiosperm diversification through time. American Journal of Botany 96: 349-365.
Maherali H, and JN Klironomos. 2007. Influence of phylogeny on fungal community assembly and ecosystem functioning. Science 316: 1746–1748.
Malkinson D, and K Tielbörger. 2010. What does the stress-gradient hypothesis predict? Resolving the discrepancies. Oikos 119: 1546–1552.
Marañón T, and JW Bartolome. 1993. Reciprocal transplants of herbaceous communities between Quercus agrifolia woodland and adjacent grassland. Journal of Ecology 81: 673–682.
Marqués MJ, R Bienes, R Pérez-Rodríguez, and L Jiménez. 2008. Soil degradation in Central Spain due to sheet water erosion by low-intensity rainfall events. Earth Surface Processes and Landforms 33: 414–423.
Matesanz S. 2008. Ecología funcional de plantas de yesos: respuestas y vulnerabilidad al cambio global. PhD Thesis.
Matesanz S, F Valladares, D Tena, M Costa-Tenorio, and D Bote. 2006. Early Dynamics of Plant Communities on Revegetated Motorway Slopes from Southern Spain: Is Hydroseeding Always

231
Needed?. Restoration Ecology 14: 297–307.
Matías, L. 2010. Efectos del Cambio Climático sobre la regeneración del bosque mediterráneo: una aproximación experimental. PhD Thesis.
Maxwell K, and GN Johnson. 2000. Chlorophyll fluorescence: a practical guide. Journal of Experimental Botany 51: 659–668.
Mayfield MM, and JM Levine. 2010. Opposing effects of competitive exclusion on the phylogenetic structure of communities. Ecology Letters (In press).
McAuliffe JR. 1988. Markovian dynamics of simple and complex desert plant communities. American Naturalist 131: 459–490.
McCullagh P, and J Nelder. 1989. Generalized Linear Models, Second Edition. Chapman and Hall, London.
McDonald PM. 1986. Grasses in young conifer plantations-Hindrance and help. Nortwest Science 60: 271–278.
McNaughton SJ. 1978 Serengeti Ungulates: Feeding Selectivity Influences the Effectiveness of Plant Defense Guilds. Science 17: 806–807.
Méndez M, D García, FT Maestre, and A Escudero. 2008. More ecology is needed to restore Mediterranean ecosystems. A reply to Valladares and Gianoli. Restoration Ecology 16: 210-216.
Meyer SE. 1986. The ecology of gypsophile endemism in the Eastern Mojave desert. Ecology 67: 1303–1313.
Michalet R. 2007. Highlighting the multiple drivers of change in the interactions along stress gradients. New Phytologist 173: 1–2.
Michalet R, RW Brooker, LA Cavieres, Z Kikvidze, CJ Lortie, FI Pugnaire, A Valiente-Banuet, and RM Callaway. 2006. Do biotic interactions shape both sides of the humped-back model of species richness in plant communities? Ecology Letters 9: 767–773.
Milchunas DG, WK Lauenroth, PL Chapman, and MK Kazempour. 1989. Effects of grazing, topography, and precipitation on the structure of a semiarid grassland. Vegetatio 80: 11–23.
Millennium Ecosystem Assessment. 2005. Ecosystems and Human Well-being: Desertification Synthesis. World Resources Institute, Washington, DC.
Miranda JD, FM Padilla, and FI Pugnaire. 2009. Response of a Mediterranean semiarid community to changing patterns of water
supply. Perspectives in Plant Ecology, Evolution, and Systematics, 11: 255–266.
Miriti MN. 2006. Ontogenetic shift from facilitation to competition in a desert shrub. Journal of Ecology 94: 973–979.
Miriti MN, HF Howe, and SJ Wright. 1998. Spatial patterns of mortality in a Colorado desert plant community. Plant Ecology 136: 41–51.
Miriti MN, SJ Wright, and HF Howe. 2007. Twenty years of changes in spatial association and community structure in a community of desert perennials. Ecology 88: 1177–1190.
Mitchell, MGE, JF Cahill, and DS Hik. 2009. Plant interactions are unimportant in a subarctic-alpine plant community. Ecology 90: 2360-2367.
Mittlebach GG. 2010. Understanding species richness–productivity relationships: the importance of meta-analyses. Ecology 91: 2540-2544.
Moro MJ, FI Pugnaire, P Haase, and J Puigdefábregas. 1997. Effect of the canopy of Retama sphaerocarpa on its understorey in a semiarid environment. Functional Ecology 11: 175-184.
Mulder CPH, DD Uliassi, and DF Doak. 2001. Physical stress and diversity-productivity relationships: the role of positive interactions. Proceedings of the National Academy of Sciences 98: 6704–6708.
Navarro RM, A del Campo, and J Cortina. 2006. Factores que afectan al éxito de una repoblación y su relación con la calidad de planta. In: Cortina J, JL Peñuelas, J Puértolas, R Savé, and A Vilagrosa (Eds.). Calidad de planta forestal para la restauración de ambientes mediterráneos. Ministerio de Medio Ambiente, Spain.
Navarro FB, J Lorite, E Fernández-Ondoño, MA Ripoll, and MN Jiménez. 2008. Stipa tenacissima as a nurse planto f the endemic species Haplophyllum bastetanum near Granada, SE Spain. Applied Vegetation Science 11: 63–72.
Niering WA, RH Whittaker, and CH Lowe. 1963. The saguaro: a population in relation to environment. Science 142: 15–23.
Niinemets U, and F Valladares. 2006. Tolerance to shade, drought, and waterlogging of temperate Northern Hemisphere trees and shrubs. Ecological Monographs 76: 521–547.
Ninyerola M, X Pons, and JM Roure. 2005. Atlas Climático Digital de la Península Ibérica. Metodología y aplicaciones en bioclimatología y geobotánica.

BIBLIOGRAFÍA
232
ISBN932860-8-7. Universidad Autónoma de Barcelona, Bellaterra, Spain.
Noble JC, and DJ Tongway. 1986. Herbivores in arid and semi-arid rangelands. In: Australian Soils: The Human Impact. Russell JS, and RF Isbell (Eds.). University of Queensland Press, Australia.
Noy-Meir I. 1973. Desert ecosystems: environment and producers. Annual Review in Ecology and Systematics 4: 25–51.
Noy-Meir I. 1975. Stability of grazing systems: an application of predator–prey graphs. Journal of Ecology 63: 459–481.
Olano JM, I Caballero, A Escudero, and J Loidi. 2005. Prediction of plant cover from seed bank analysis in a semi-arid plant community on gypsum. Journal of Vegetation Science 16: 215-222.
Olano JM, ES Menges, and E Martinez. 2006. Carbohydrate storage in five resprouting Florida scrub plant across a fire chronosequence. New Phytologist 170: 99−106.
Ogle K, and JF Reynolds. 2004. Plant responses to precipitation in desert ecosystems: integrating functional types, pulses, thresholds, and delays. Oecologia 141: 282–294.
Padilla FM, and FI Pugnaire. 2007. Rooting depth and soil moisture control Mediterranean woody seedling survival during drought. Functional Ecology 21: 489–495.
Pakeman RJ, FI Pugnaire, R Michalet, CJ Lortie, K Schiffers, FT Maestre, and J Travis. 2009. Is the cask of facilitation ready for bottling? A symposium on the connectivity and future directions of positive plant interactions. Biology Letters 5: 577–579.
Palacio S, A Escudero, G Montserrat-Martí, M Maestro-Martínez, R Milla, and MJ Albert. 2007. Plants living in gypsum: beyond the specialist model. Annals of Botany 99: 333–343.
Palacio S, M Maestro, and G Montserrat-Martí. 2007. Seasonal dynamics of nonstructural carbohydrates in two species of Mediterranean sub-shrubs with different leaf phenology. Environmental and Experimental Botany 59: 34–42.
Palacio S, R Milla, J Albuixech, C Pérez-Rontomé, JJ Camarero, M Maestro, and G Montserrat-Martí. 2008. Seasonal variability of dry matter content and its relationship with shoot growth and nonstructural carbohydrates. New Phytologist 180: 133–142.
Palacio S, and G Montserrat-Martí. 2005. Bud morphology and shoot growth dynamics in two species of Mediterranean sub-shrubs co-existing in gypsum outcrops. Annals of Botany 95: 949–958.
Parker VT, and CH Muller. 1982. Vegetational and environmental-changes beneath isolated live oak trees (Quercus agrifolia) in a California annual grassland. American Midland Naturalist 107: 69–81.
Paula S, M Arianoutsou, D Kazanis, Ç Tavsanoglu, F Lloret, C Buhk, F Ojeda, B Luna, JM Moreno, A Rodrigo, JM Espelta, S Palacio, B Fernández-Santos, PM Fernandes, and JG Pausas. 2009. Fire-related traits for plant species of the Mediterranean Basin. Ecology 90: 1420.
Pausas JG, A Bonet, FT Maestre, and A Climent. 2006. The role of the perch effect on the nucleation process in Mediterranean semi-arid oldfields. Acta Oecologica 29: 346–352.
Pausas JG, and M Verdú. 2010. The jungle of methods for evaluating phenotypic and phylogenetic structure of communities. BioScience 60: 614–625.
Pavoine S, E Vela, S Gachet, G de Bélair, and MB Bonsall. 2010. Linking patterns in phylogeny, traits, abiotic variables and space : a novel approach to linking environmental filtering and plant community assembly. Journal of Ecology. In press.
Peco B, L Rico, and FM Azcárate. 2009. Seed size and response to rainfall patterns in annual grasslands: 16 years of permanent plots. Journal of Vegetation Science 20: 8–16.
Perry JN. 1998. Measures of spatial pattern for counts. Ecology 79: 1008–1017.
Pias B, S Matesanz, A Escudero, TE Gimeno, and F Valladares. 2010. Effects of Global Change on early fitness components and offspring performance of an endemic Mediterranean gypsophyle. Oikos (In press) doi: 10.1111/j.1600-0706.2010.18232.x
Prider JN, and JM Facelli. 2004. Interactive effects of drought and shade on three arid zone chenopod shrubs with contrasting distributions in relation to tree canopies. Functional Ecology 18: 67–76.
Poorter H, Ü Niinemets, I Poorter, IJ Wright, and R Villar. 2009. Causes and consequences of variation in leaf mass per area (lma): a meta-analysis. New Phytologist 182: 565–588.
Pueyo Y, and CL Alados. 2007. Effects of fragmentation, abiotic factors and land use on vegetation recovery in a semi-arid

233
Mediterranean area. Basic and Applied Ecology 8: 158–170.
Pueyo Y, CL Alados, B García-Ávila, S Kéfi, M Maestro, and M Rietkerk. 2009. Comparing direct abiotic amelioration and facilitation as tools for restoration of semiarid grasslands. Restoration Ecology 17: 908–916.
Puigdefábregas J, A Sole, L Gutierrez, G del Barrio, and M Boer. 1999. Scales and processes of water and sediment redistribution in drylands: results from Rambla Honda study site in Southeast Spain. Earth-Science Reviews 48: 39–70.
Puigdefábregas J, and G Sánchez. 1996. Geomorphological implications of vegetation patchiness on semiarid slopes. In: Advances in hillslope processes, Volume 2. Anderson MG, and SM Brooks (Eds.). John Willey y Sons, London.
Pugnaire FI, C Armas, and F Valladares. 2004. Soil as a mediator in plant-plant interactions in a semi-arid community. Journal of Vegetation Science 15: 85-92.
Pugnaire FI, P Haase, J Puigdefábregas, M Cueto, LD Incoll, and SC Clark. 1996a. Facilitation and succession under the canopy of the leguminous shrub, Retama sphaerocarpa, in a semi-arid environment in south-east Spain. Oikos 76: 455–464.
Pugnaire FI, P Haase, LD Incoll, and SC Clark. 1996b. Response of the tussock grass Stipa tenacissima to watering in a semi-arid environment. Functional Ecology 10: 265–274.
Pugnaire FI, and MT Luque. 2001. Changes in plant interactions along a gradient of environmental stress. Oikos 93: 42–49.
Quinn GP, and MJ Keough. 2002. Experimental design and data analysis for biologists. Cambridge University Press.
Rajaniemi TK, DE Goldberg, R Turkington, and AR Dyer. 2006. Quantitative partitioning of regional and local processes shaping regional diversity patterns. Ecology Letters 9: 121–128.
Ramírez DA. 2006. Estudio de la transpiración del esparto (Stipa tenacissima L.) en una cuenca del semiárido alicantino: un análisis pluriescalar. PhD Thesis.
Ramírez DA, and J Bellot. 2009. Linking population density and habitat structure to ecophysiological responses in semiarid Spanish steppes. Plant Ecology 200: 191–204.
Ramírez DA, J Bellot, F Domingo, and A Blasco. 2007. Can water responses in Stipa tenacissima L. during summer season be promoted by non-rainfall water gains in soil? Plant Soil 291: 67–79.
Rand TA. 2004. Competition, facilitation, and compensation for insect herbivory in an annual salt marsh forb. Ecology 85: 2046–2052.
Rebollo S, DG Milchunas, I Noy-Meir, and PL Chapman. 2002. The role of a spiny plant refuge in structuring grazed shortgrass steppe communities. Oikos 98: 53–64.
Resco V, DD Ignace, W Sun, TE Huxman, JF Weltzin, and DG Williams. 2008. Chlorophyll fluorescence, predawn water potential and photosynthesis in precipitation pulse-driven ecosystems: implications for ecological studies. Functional Ecology 22: 479–283.
Reynolds JF, PR Kemp, K Ogle, and RJ Fernandez. 2004. Modifying the 'pulse-reserve' paradigm for deserts of North America: precipitation pulses, soil water, and plant responses. Oecologia 141:194–210.
Reynolds JF, FT Maestre, E Huber-Sannwald, JE Herrick, and PR Kemp. 2005. Aspectos socioeconómicos y biofísicos de la desertificación. Ecosistemas 14: 3–21.
Reynolds, JF, DM Stafford Smith, EF Lambin, BL Turner II, M Mortimore, SPJ Batterbury, TE Downing, H Dowlatabadi, RJ Fernández, JE Herrick, E Huber-Sannvald, R Leemans, T Lynam, FT Maestre, M Ayarza, and B Walker. 2007a. Global desertification: Building a science for dryland development. Science 316: 847–851.
Reynolds JF, FT Maestre, PR Kemp, DM Stafford-Smith, and E Lambin. 2007b. Natural and human dimensions of land degradation in drylands: causes and consequences. In: Terrestrial Ecosystems in a Changing World. Canadell J, D Pataki, and LF Pitelka (Eds.). Springer-Verlag, Berlin.
Rezende E, J Lavabre, P Guimaraes, P Jordano, and J Bascompte. 2007. Non-random coextinctions in phylogenetically structured mutualistic networks. Nature 448: 925–928.
R Development Core Team. 2009. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna.
Rietkerk MG, and J van de Koppel. 2008. Regular pattern formation in real ecosystems. Trends in ecology and evolution 23: 169–175.
Riginos C, SJ Milton, and T Wiegand. 2005. Context-dependent interactions between adult shrubs and seedlings in a semi-arid shrubland. Journal of Vegetation Science 16: 331–340.

BIBLIOGRAFÍA
234
Riginos C, and TP Young. 2007. Positive and negative effects of grass, cattle, and wild herbivores on Acacia saplings in an East African savanna. Oecologia 153: 985–995.
Romao R, and A Escudero. 2005. Gypsum physical soil surface crusts and the existence of gypsophytes in semi-arid central Spain. Plant Ecology 181: 1–11.
Roques KG, TG O'Connor, and AR Watkinson. 2001. Dynamics of shrub encroachment in an African savanna: relative influences of fire and herbivory, rainfall and density dependence. Journal of Applied Ecology 38: 268–280.
Rooney TP. 2008. Comparison of co-occurrence structure of temperate forest herb-layer communities in 1949 and 2000. Acta Oecologica 34: 354–360.
Rueda M, S Rebollo, L Gálvez-Bravo, and A Escudero. 2008. Habitat use by large and small herbivores in a fluctuating Mediterranean ecosystem: Implications of seasonal changes. Journal of Arid Environments 72: 1698–1708.
Sala OE, RA Golluscio, WK Lauenroth, and CA Soriano. 1989. Resource Partitioning Between Shrubs and Grasses in the Patagonian Steppe. Oecologia 81: 501–505.
Sala OE, and WK Lauenroth. 1982. Small rainfall events: an ecological role in semiarid regions. Oecologia 53: 301–304.
Sargent R, and DD Ackerly. 2008. Plant-pollinator interactions and community assembly. Trends in Ecology and Evolution 23: 123–130.
Scheffer M, J Bascompte, WA Brock, V Brovkin, SR Carpenter, V Dakos, H Held, EH van Nes, M Rietkerk, and G Sugihara, 2009. Early warning signals for critical transitions, Nature 461: 53–59.
Schiffers K, and K Tielbörger. 2006. Ontogenetic shifts in interactions among annual plants. Journal of Ecology 94: 336–341.
Schlesinger WH, and AM Pilmanis. 1998. Plant-soil interactions in desert. Biogeochemistry 42: 169–187.
Schlesinger WH, JF Reynolds, GL Cunningham, LF Huenneke, WM Jarrell, RA Virginia, and WG Whitford. 1990. Biological feedbacks in global desertification. Science 247: 1043–1048.
Scholes RJ, and SR Archer. 1997. Tree-grass interactions in savannas. Annual Review in Ecology and Systematics 28: 517–544.
Schupp EW. 1995. Seed-seedling conflicts, habitat choice and patterns of plant recruitment. American Journal of Botany 82: 399–409.
Schweingruber FH. 1988. Tree Rings: Basics and Applications of Dendrochronology. D. Reidel, Dordrecht; The Netherlands.
Schwinning S, and JR Ehleringer. 2001. Water use trade-offs and optimal adaptations to pulse-driven arid ecosystems. Journal of Ecology 89: 464–480.
Schwinning S, and OE Sala. 2004. Hierarchy of responses to resource pulses in and semiarid ecosystems. Oecologia 141: 211–220.
Seifan M, K Tielbörger, and R Kadmon. 2010a. Direct and indirect interactions among plants explain counterintuitive positive drought effects on an eastern Mediterranean shrub species. Oikos 119: 1601-1609.
Seifan M, T Seifan, C Ariza, and K Tielbörger. 2010b. Facilitating an importance index. Journal of Ecology 98: 356–361.
Shreve F. 1931. Physical conditions in sun and shade. Ecology 12: 96–104.
Silliman BR, J Van de Koppel, MD Bertness, L Stanton, and I Mendelssohn. 2005. Drought and herbivory lead to catastrophic salt marsh collapse. Science 310:1803–1805
Silvertown J. 2004. Plant coexistence and the niche. Trends in Ecology and Evolution 19: 605–611.
Smit C, M Rietkerk, and MJ Wassen. 2009. Inclusion of biotic stress (consumer pressure) alters predictions from the stress gradient hypothesis. Journal of Ecology 97: 1215– 1219.
Smit C, C Vanderberghe, J den Ouden, and H Müller-Schärer. 2007. Nurse plants, tree saplings and grazing pressure: changes in facilitation along a biotic environmental gradient. Oecologia 152: 265–273.
Smith SA, JM Beaulieu, and MJ Donoghue. 2010. An uncorrelated relaxed-clock analysis suggests an earlier origin for flowering plants. Proceedings of the National Academy of Sciences USA 107: 5897-5902.
Soliveres S, V Contreras, JP Fernandes, J Cortina, P García-Palacios, M Martínez, and J Fort. Ecotecnología aplicada a la restauración de infraestructuras lineales. In. F Valladares et al. (Eds.). Ecología e infraestructuras lineales: bases científicas para soluciones técnicas. Ministerio de Medio Ambiente, Spain. In press.
Soliveres S, L DeSoto, FT Maestre, and JM Olano. 2010. Spatio-temporal heterogeneity in abiotic factors modulate multiple ontogenetic shifts between competition and facilitation. Perspectives

235
in Plant Ecology, Evolution and Systematics 12: 227–234.
Soliveres S, P García-Palacios, AP Castillo-Montroy, FT Maestre, A Escudero, and F Valladares. 2010. Temporal dynamics of herbivory and water availability interactively modulate the outcome of a grass-shrub interaction in a semi-arid ecosystem. Oikos (In press) Doi: 10.1111/j.1600-0706.2010.18993.x
Soliveres S, J Monerris, and J Cortina. 2008. El uso de parches artificiales mejora el rendimiento de una repoblación de Rhamnus lycioides en medio semiárido. Cuadernos de la Sociedad Española de Ciencias Forestales 28: 125-130.
Soil Survey Staff. 1994. Keys to Soil Taxonomy, Sixth edition. USDA Soil Conservation Service, Pocahontas Press; Blacksburg.
SPSS. 2004. SPSS® Trends 13.0 . SPSS Inc.; Chicago.
Stace HCT, GD Hubble, R Brewer, KH Northcote, JR Sleeman, MJ Mulcahy, and EG Hallsworth. 1968. A Handbook of Australian Soils. Rellim Technical Publications, CSIRO and International Society of Soil Science, Glenside, South Australia.
Stachowicz JJ. 2001. Mutualisms, positive interactions, and the structure of ecological communities. BioScience 51: 235–246.
Sternberg M, and M Shoshany. 2001. Influence of slope aspect on Mediterranean woody formations: comparison of a semiarid and an arid site in Israel. Ecological Research 16: 335–345.
Stevens PF. 2010. Angiosperm Phylogeny Website: http://www.mobot.org/MOBOT/research/APweb/.
Sthultz CM, CA Gehring, and TG Whitham. 2007. Shifts from competition to facilitation between a foundation tree and a pioneer shrub across spatial and temporal scales in a semiarid woodland. New Phytologist 173: 135–145.
Stokes MA, and TL Smiley. 1968. An Introduction to Tree-ring Dating. University of Chicago Press; Chicago.
Susanna A, N Garcia-Jacas, O Hidalgo, R Vilatersana, and T Garnatje. 2006. The Cardueae (Compositae) revisited: insights from ITS, trnL-trnF, and matK nuclear and chlorpoplast DNA analysis. Annals of the Missouri Botanical Garden 93: 150-171.
Tielbörger K, and R Kadmon. 2000a. Temporal environmental variation tips the balance
between facilitation and interference in desert plants. Ecology 81: 1544–1553.
Tielbörger K, and R Kadmon. 2000b. Indirect effects in a desert plant community: is competition among annuals more intense under shrub canopies? Plant Ecology 150: 53–63.
Tilman D. 1988. Plant strategies and the dynamics and structure of plant communities. Princeton University Press, New Jersey.
Tilman D. 1994. Competition and biodiversity in spatially structured habitats. Ecology 75: 2-16.
Tirado R, and FI Pugnaire. 2003. Shrub spatial association and consequences for reproductive success. Oecologia 136: 296–301.
Tirado R, and FI Pugnaire. 2005. Community structure and positive interactions in constraining environments. Oikos 111: 437–444.
Tongway DJ, and N Hindley. 1995. Assessment of soil condition of tropical grasslands. CSIRO Ecology and Wildlife, Canberra, Australia.
Tongway DJ, and JA Ludwig. 1996. Rehabilitation of semi-arid landscapes in Australia II: restoring productive soil patches. Restoration Ecology 4: 388-397.
Tongway DJ, AD Sparrow, and MH Friedel. 2003 Degradation and recovery processes in arid grazing lands of central Australia. Part 1: soil and land resources Journal of arid environments 55: 301-326.
Topp GC, and JL Davis. 1985. Measurement of soil water control using time domain reflectometry (TDR): a field application. Journal of Soil Science 49: 9–22.
Torices R. 2010. Adding time-calibrated branch lengths to the Asteraceae supertree. Journal of Systematics and Evolution 48: 271-278.
Tracol Y, JR Gutiérrez, and FA Squeo. 2010 Plant Area Index and microclimate underneath shrub species from a Chilean semiarid community. Journal of arid environments (In press).
Travis JMJ, RW Brooker, and C Dytham. 2005. The interplay of positive and negative species interactions along an environmental gradient: insights from an individual based model. Biology Letters 1: 5–8.
Trubat R, J Cortina, and A Vilagrosa. 2008. Short-term nitrogen deprivation increases field performance in nursery seedlings of Mediterranean woody species. Journal of Arid Environments 72: 879–890.

BIBLIOGRAFÍA
236
Valente LM, V Savolainen, and P Vargas. 2010. Unparalleled rates of species diversification in Europe. Proceedings of the Royal Society of London, series B 277:1489-1496.
Valiente-Banuet A, and E ezcurra. 1991. Shade as a cause of the association between the cactus Neobuxbaumia tetetzo and the nurse plant Mimosa luisana in the Tehuacán valley, mexico. Journal of Ecology 79: 961–970.
Valiente-Banuet A, and M. Verdú. 2007. Facilitation can increase the phylogenetic diversity of plant communities. Ecology Letters 10: 1029–1036.
Valiente-Banuet A, and M. Verdú. 2008. Temporal shifts from facilitation to competition occur between closely related taxa. Journal of Ecology 96: 489–494.
Valiente-Banuet A, A Vital, M Verdú, and RM Callaway. 2006. Modern Quaternary plant lineages promote diversity through facilitation of ancient Tertiary lineages. Proceedings of the National Academy of Sciences USA 103: 16812–16817.
Valiente-Banuet A, F Vite, and A Zavala-Hurtado. 1991. Interaction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana. Journal of Vegetation Science 2: 11–14.
Valladares F. 2001. Características mediterráneas de la conversión fotosintética de la luz en biomasa: de órgano a organismo. pp: 67–94 In: Zamora R, and FI Pugnaire (Eds). Aspectos funcionales de los ecosistemas mediterráneos. CSIC-AEET, Granada.
Valladares F, I Aranda, and D Sánchez-Gómez. 2008. La luz como factor ecológico y evolutivo para las plantas y su interacción con el agua. pp 337–372 In: Valladares F. (Ed.). Ecología del bosque mediterráneo en un mundo cambiante (segunda edición). Ministerio de Medio Ambiente. Egraf, S.A., Madrid.
Valladares F, LG Hernández, I Dobarro, C García-Pérez, R Sanz, and FI Pugnaire. 2003. The ratio of leaf to total photosynthetic area influences shade survival and plastic response to light of green-stemmed leguminous shrub seedlings. Annals of Botany 91: 577–584.
Valladares F, and FI Pugnaire. 1999. Tradeoffs between irradiance capture and avoidance in semiarid environments assessed with a crown architecture model. Annals of Botany 83: 459–469.
Valladares F, I Dobarro, D Sánchez-Gómez, and RW Pearcy. 2005. Photoinhibition and drought in Mediterranean woody saplings:
scaling effects and interactions in sun and shade phenotypes. Journal of Experimental Botany 56: 483–494.
Valladares F, and E Gianoli. 2007. How much ecology do we need to know to restore Mediterranean ecosystems? Restoration Ecology 15: 363-368.
Valladares F, and RW Pearcy. 2002. Drought can be more critical in the shade than in the sun: a field study of carbon gain and photoinhibition in a Californian shrub during a dry El Niño year. Plant Cell and Environment 25: 749.
Valladares F, J Zaragoza-Castells, D Sánchez-Gómez, S Matesanz, B Alonso, A Portsmuth, A Delgado, and OK Atkin. 2008. Is shade beneficial for mediterranean shrubs experiencing periods of extreme drought and late-winter frosts? Annals of Botany 102: 923–933.
Vamosi SM, SB Heard, JC Vamosi, and CO Webb. 2009. Emerging patterns in the comparative analysis of phylogenetic community structure. Molecular Ecology 18: 572–592.
Vamosi JC, and SM Vamosi. 2010. Key innovations within a geographical context in flowering plants: towards resolving Darwin's abominable mystery. Ecology Letters 13: 1270–1279.
Van Auken OW. 2000. Shrub invasions of North American semiarid grasslands Annual Review of Ecology and Systematics 31: 197–215.
Van de Koppel J, M Rietkerk, and FJ Weissing. 1997. Catastrophic vegetation shifts and soil degradation in terrestrial grazing systems. Trends in Ecology and Evolution 12: 352-356.
Veblen KE. 2008. Season- and herbivore-dependent competition and facilitation in a semi-arid savanna. Ecology 89: 1532–1540.
Veblen T. 1989. Tree regeneration responses to gaps along a transandean gradient. Ecology 77: 541–543.
Verdú M, and P García-Fayos. 1996. Nucleation processes in a Mediterranean bird-dispersed plant. Functional Ecology 10: 275–280.
Verdú M, P Jordano, and A Valiente-Banuet. 2010. The phylogenetic structure of plant facilitation networks changes with competition. Journal of Ecology, In press.
Verdú M, and JG Pausas. 2007. Fire drives phylogenetic clustering in Mediterranean Basin woody plant communities. Journal of Ecology 95: 1316–1323.
Verdú M, PJ Rey, JM Alcántara, G Siles, and A Valiente-Banuet. 2009. Phylogenetic

237
signatures of facilitation and competition in successional communities. Journal of Ecology 97; 1171-1180.
Vesk PA, DI Warton, and M Westoby. 2004. Sprouting by semi-arid plants: testing a dichotomy and predictive traits. Oikos 107: 72–89.
Villar P, S Soliveres, and JL Quero. Introducción de especies leñosas. In. F Valladares et al. (Eds.). Ecología e infraestructuras lineales: bases científicas para soluciones técnicas. Ministerio de Medio Ambiente, Spain. In press.
Waide RB, MR Willig, CF Steiner, G Mittelbach, L Gough, SI Dodson, GP Juday, and R Parmenter. 1999. The relationship between productivity and species richness. Annual Review in Ecology and Systematics 30: 257–300.
Wang H, MJ Moore, PS Soltis, CD Bell, SF Brockington, R Alexandre, CC Davis, M Latvis, SR Manchester, and DE Soltis. 2010. Rosid radiation and the rapid rise of angiosperm-dominated forests. Proceedings of the National Academy of Sciences USA 106: 3853-3858.
Webb CO, DD Ackerly, M McPeek, and MJ Donoghue. 2002. Phylogenies and community ecology. Annual Review of Ecology and Systematics 33: 475–505.
Webb CO, DD Ackerly, and SW Kembel. 2008. Phylocom: software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics 24: 2098–2100.
Webb CO, GS Gilbert, and MJ Donoghue. 2006. Phylodiversity dependent seedling
mortality, size structure, and disease in a Bornean rain forest. Ecology 87: S123–S131.
Westoby M. 1979/80. Elements in a theory of vegetation dynamics. Israel Journal of Botany 28: 169–194.
Westoby M, BH Walker, and I Noy-Meir. 1989. Opportunistic management for rangelands not at equilibrium. Journal of Rangeland Management 42: 266–274.
Whitford WG. 2002. Ecology of desert systems. Academic Press; California.
Whittaker RJ. 2010. Meta-analyses and mega-mistakes: calling time on meta-analysis of the species richness-productivity relationship. Ecology 91: 2522-2533.
Wikström N, V Savolainen, and MW Chase. 2001. Evolution of the angiosperms: calibrating the family tree. Proceeding of
the Royal Society of London, series B 268: 2211-2219.
Wise MJ, and WG Abrahamson. 2005. Beyond the compensatory continuum: environmental resource levels and plant tolerance of herbivory. Oikos 109: 417–428.
Wise MJ, and WG Abrahamson. 2007. Effects of resource availability on tolerance of herbivory: a review and assessment of three opposing models. The American Naturalist 169: 443–454.
Wojciechowski MF, M Lavin, and MJ Sanderson. 2004. A phylogeny of legumes (Leguminosae) based on analysis of the plastid matK gene resolves many well-supported subclades within the family. American Journal of Botany 91: 1846-1862.
Wolfe BE, JL Parrent, AM Koch, BA Sikes, M Gardes, and JN Klironomos. 2009 Spatial Heterogeneity in Mycorrhizal Populations and Communities: Scales and Mechanisms. In: Mycorrhizas - Functional Processes and Ecological Impact. Azcón-Aguilar C, et al. (Eds.). Springer-Verlag, Berlin.
Wright IJ, PB Reich, M Westoby, DD Ackerly, Z Baruch, F Bongers, J Cavender-Bares, FS Chapin, JHC Cornelissen, M Diemer, J Flexas, E Garnier, PK Groom, J Gulias, K Hikosaka, BB Lamont, T Lee, W Lee, C Lusk, JJ Midgley, ML Navas, Ü Niinemets, J Oleksyn, N Osada, H Poorter, P Poot, I Prior, VI Pyankov, C Roumet, SC Thomas, MG Tjoelker, E Veneklaas, and R Villar. 2004. The world-wide leaf economics spectrum. Nature 428: 821-827.
Zamora R, P García-Fayos, and L Gómez-Aparicio. 2008. Las interacciones planta-planta y planta animal en el contexto de la sucesión ecológica. pp 337–372 In: Valladares F. (Ed.). Ecología del bosque mediterráneo en un mundo cambiante (segunda edición). Ministerio de Medio Ambiente. Egraf, S.A., Madrid.
Zavaleta ES. 2006. Shrub establishment under experimental global changes in a California grassland. Plant Ecology 184: 53–63.
Zimmermann J, S Higgins, V Grimm, J Hoffmann, and A Linstädter. 2010. Grass mortality in semi-arid savanna: The role of fire, competition and self-shading. Perspectives in Plant Ecology, Evolution and Systematics 12: 1–8.

COAUTORES
238
AFILIACIÓN DE LOS COAUTORES
Fernando T. Maestre Gil
Área de Biodiversidad y Conservación, Departamento de Biología y Geología, Escuela
Superior de Ciencias Experimentales y Tecnología, Universidad Rey Juan Carlos, 28933
Móstoles, Spain. E-mail: [email protected]
Adrián Escudero Alcántara
Área de Biodiversidad y Conservación, Departamento de Biología y Geología, Escuela
Superior de Ciencias Experimentales y Tecnología, Universidad Rey Juan Carlos, 28933
Móstoles, Spain. E-mail: [email protected]
Fernando Valladares Ros
Instituto de Recursos Naturales, Centro de Ciencias Medioambientales, C.S.I.C., Serrano 115,
E-28006 Madrid, Spain. E-mail: [email protected]
Pablo García-Palacios
1) Área de Biodiversidad y Conservación, Departamento de Biología y Geología, Escuela
Superior de Ciencias Experimentales y Tecnología, Universidad Rey Juan Carlos, 28933
Móstoles, Spain.
2) Instituto de Recursos Naturales, Centro de Ciencias Medioambientales, C.S.I.C., Serrano
115, E-28006 Madrid, Spain. E-mail: [email protected]
Andrea P. Castillo Monroy
Área de Biodiversidad y Conservación, Departamento de Biología y Geología, Escuela
Superior de Ciencias Experimentales y Tecnología, Universidad Rey Juan Carlos, 28933
Móstoles, Spain. E-mail: [email protected]

239
Lucia DeSoto Suárez
Centro de Ecogía Funcional, Departamento de Ciencias da Vida. Universidade de Coimbra
3001 – 455 Coimbra. Portugal. E-mail: [email protected]
Jose Miguel Olano
Área de Botánica, Departamento de Ciencias Agroforestales, E.U.I. Agrarias de Soria,
Universidad de Valladolid, Campus de los Pajaritos, 42004 Soria, Spain. E-mail:
David Eldridge
Department of Environment, Climate Change and Water, Evolution and Ecology Research
Centre, School of Biological, Earth and Environmental Sciences, University of New South
Wales, Sydney, NSW 2052, Australia. E-mail: [email protected]
Matthew A. Bowker
Colorado Plateau research Station, US Geological Survey, Flagstaff (USA). E-mail:
Matthew K. Tighe
Ecosystem Management, School of Environmental and Rural Science, University of New
England, Armidale, NSW 2351, Australia. E-mail: [email protected]
Rubén Torices Blanco
Área de Botánica, Departamento de Ciencias Agroforestales, E.U.I. Agrarias de Soria,
Universidad de Valladolid, Campus de los Pajaritos, 42004 Soria, Spain. E-mail:

COAUTORES
240
Diseño portada e imágenes:
Joan Miquel Fuster Mollà y Santiago Soliveres Codina
Maquetación:
Soraya Constán Nava y Santiago Soliveres Codina