Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
ÉRIKA DE ARAUJO VALONI
Envolvimento das proteínas fosfatases no acúmulo de lipídios em Saccharomyces cerevisiae e síntese em biorreator utilizando glicerina loira
como fonte de carbono
RIO DE JANEIRO
2014

ii
ÉRIKA DE ARAUJO VALONI
Envolvimento das proteínas fosfatases no acúmulo de lipídios em Saccharomyces cerevisiae e síntese em biorreator utilizando glicerina loira
como fonte de carbono
RIO DE JANEIRO 2014
Dissertação de Mestrado apresentada ao Programa de Pós-Graduação em Tecnologia de Processos Químicos e Bioquímicos, Escola de Química, Universidade Federal do Rio de Janeiro, como parte dos requisitos para obtenção do título de Mestre em Ciências.
Orientadora s: Profa. Dra Maria Alice Zar ur Coelho Profa. Dra Mônica Montero-Lomeli

iii
ENVOLVIMENTO DAS PROTEÍNAS FOSFATASES NO ACÚMULO D E LIPÍDIOS EM SACCHAROMYCES CEREVISIAE E SÍNTESE EM BIORREATOR
UTILIZANDO GLICERINA LOIRA COMO FONTE DE CARBONO
ÉRIKA DE ARAUJO VALONI
Dissertação de Mestrado submetida ao Programa Pós-G raduação em Tecnologia de Processos Químicos e Bioquímicos, Esc ola de Química, Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Mestre em Ciênc ias.
Aprovada por:
____________________________________ Maria Alice Zarur Coelho, D.Sc.
____________________________________ Mônica Montero-Lomeli, D.Sc.
____________________________________ Priscilla Filomena Fonseca Amaral, D.Sc.
____________________________________ Eleonora Kurtenbach, D.Sc.
____________________________________ Aline Machado de Castro, D.Sc.
RIO DE JANEIRO 2014

iv
FICHA CATALOGRÁFICA
Valoni, Érika de Araujo
Envolvimento das proteínas fosfatases no acúmulo de lipídios em Saccharomyces cerevisiae e síntese em biorreator utilizando glicerina loira como fonte de carbono/Érika de Araujo Valoni. Rio de Janeiro: UFRJ/EQ, 2014
(Dissertação) – Universidade Federal do Rio de Janeiro, Escola de Química, 2014.
Orientador (es): Maria Alice Zarur Coelho e Mônica Montero-Lomeli
1- Lipídios 2. Saccharomyces cerevisiae 3- Glicerina 4- Fosfatases 5- (Mestrado – UFRJ/EQ) 6- Maria Alice Zarur Coelho e Mônica Montero-Lomeli
I - Título

v
"É fazendo que se aprende a fazer aquilo que se dev e aprender a fazer."
Aristóteles

vi
DEDICATÓRIA
Dedico esta dissertação à minha querida mãe, Maria Aparecida. Obrigada por me proporcionar esse momento.

vii
AGRADECIMENTOS
Agradeço às minhas orientadoras Maria Alice Coelho e Mônica Montero pela
contribuição científica para minha carreira profissional.
Um agradecimento especial e carinhoso à minha gerente Gina Vazquez
Sebastian pelo apoio na realização desses estudos e por sempre acreditar na
capacitação de sua equipe.
Aos amigos do Cenpes/Bio me apoiaram ao longo desses meses. À querida
amiga Danielle Altomari pela amizade ao longo desses quase 11 anos. Pelos
conselhos e incentivos que me deram força para seguir em frente, mesmo quando o
cansaço da jornada dupla de trabalho e estudos era grande. À amiga Vanessa
Vólaro, por todos os anos de amizade e pela ajuda crucial na formatação da
dissertação (você é fera!). Ao amigo Bruno Sardinha, pelo companheirismo e por
escutar meus desabafos em nossos almoços no restaurante do Cenpes. Ao querido
amigo Renato Valério (Renatinho), obrigada por poder contar sempre com sua ajuda
e ideias no laboratório. À amiga Adriana Soriano pelo exemplo de profissional e
amiga (você é inspiradora!). Aos amigos que estiveram mais próximos nesses
últimos meses, César, Felipe Matias, Bruno Lacava, vocês não têm ideia da
importância que tiveram nessa reta final. Ao amigo Absai pelas análises de glicerol
no HPLC e pela acolhida tão especial no grupo de trabalho. Às amigas Ana Paula
Torres e Lídia Santa Anna, obrigada por tudo que fizeram por mim nesses últimos
meses, por mostrar que o amor que sentimos por algo que acreditamos fica dentro
da gente (e ninguém vai nos tirar).
Gostaria de agradecer também aos colegas do laboratório Biose,
especialmente a Mariana Miguez e Roseli pelos experimentos no bioreator e por
todo apoio essencial no laboratório. À Fernanda Martins pelas análises no HPLC e
aos amigos que tive o prazer de conviver nas minhas idas e vindas entre Cenpes e
EQ/UFRJ, Gizeli, Caê, Etel, Bernardo e toda a equipe.
Um agradecimento a amiga Tatiana Félix por disponibilizar o bioreator nos
momentos de aperto (que foram vários) e pela torcida para que tudo desse certo.

viii
À amiga Roberta Ribeiro, um presente que recebi do mestrado. Sempre que
precisei estava a um whatsapp de distância, me ajudando no que fosse preciso.
Obrigada pelos momentos de café, vinho e Madonna.
Aos amigos do Laboratório de Biologia Molecular de Leveduras, em especial
ao Bruno Bozaquel-Morais e Juliana Madeira que me acompanharam nos
experimentos de laboratório e contribuiram na discussão dos resultados. Vocês são
uma referência para mim, obrigada. Ao Gabriel, pelas dosagens de glicerol por via
enzimática nos minutos finais dos experimentos de bancada. Ao Cláudio Masuda
pelas trocas de ideias e por sempre estar disposto a ajudar (muito obrigada!).
Não podia deixar de agradecer à responsável por toda minha formação na
área da microbiologia, minha grande mestre Lucy Seldin. Seus ensinamentos foram
essenciais para que eu chegasse até aqui.
Aos amigos que não são do “universo da pesquisa”, mas que se envolveram e
torceram muito para que essa etapa fosse concluída. A amiga e irmã Bianca Noleto,
não tenho palavras para agradecer todos esses anos que estivemos juntas. A amiga
Beatriz Peixoto que nos seus whatsapps, quase que diários, me perguntava como
estavam os preparativos e se tinha algo que poderia ajudar.
Um agradecimento a corrida. Por ter me preparado fisicamente a enfrentar
essa jornada. O corpo e a mente precisam andar juntas para obtermos qualquer
sucesso na vida.
E por fim, mas não menos importante, a minha família. Pelo apoio incodicional
da minha mãe, que esteve sempre ao meu lado. Por ser a pessoa mais importante
em todos os aspectos da minha vida, e principalmente por ter me apontado o valor
da educação.

ix
Resumo da dissertação de mestrado apresentada ao curso de Pós-Graduação em
Tecnologia de Processos Químicos e Bioquímicos da Escola de Química/UFRJ,
como parte dos requisitos necessários para obtenção do grau de Mestre em
Ciências (M.Sc)
ENVOLVIMENTO DAS PROTEÍNAS FOSFATASES NO ACÚMULO DE LIPÍDIOS EM Saccharomyces cerevisiae E SÍNTESE EM BIORREATOR UTILIZANDO
GLICERINA LOIRA COMO FONTE DE CARBONO
ÉRIKA DE ARAUJO VALONI
A utilização de micro-organismos oleaginosos tem se mostrado como uma
tecnologia promissora, alternativa aos óleos vegetais amplamente utilizados para a
produção de biodiesel. Dentre os micro-organismos oleaginosos (aqueles que
acumulam lipídios em mais de 20% p/p, em massa seca de célula), as leveduras
apresentam vantagens sobre os outros micro-organismos em termos de taxa de
crescimento e conteúdo de óleo intracelular. Os principais ácidos graxos
encontrados nas leveduras são principalmente, o ácido mirístico, palmítico,
esteárico, linoleico e linolênico. De acordo com perfil apresentado, estes óleos
poderiam ser empregados como matéria prima para a produção de biodiesel.
Somado à isso, a utilização do glicerol, em substituição à glicose, como substrato no
meio de cultivo, apresenta vantagens ambientais, como o aproveitamento de
resíduos, e vantagens econômicas, reduzindo o custo do processo de obtenção do
óleo microbiano. No presente trabalho, foi possível avaliar o acúmulo de
triacilgliceróis (TAG) nos corpúsculos lipídicos (CL) em cepa selvagem (WT) e cepas
mutantes para os genes que codificam as proteínas fosfatases em levedura
Saccharomyces cerevisiae, cultivadas em meio respiratório, utilizando glicerina loira
como matéria prima. Inicialmente, foi possível determinar a utilização de 5% (p/v) de
glicerina loira como matéria prima no meio de cultivo de produção de corpúsculos
lipídicos. Foram realizados experimentos para avaliação da melhor relação entre o
índice de CL e
crescimento celular. A partir de uma varredura inicial para as 66 mutantes com
deleções para os genes das proteínas fosfatases, foram selecionadas oito cepas

x
para serem avaliadas em frascos com 50 mL de volume de meio de cultivo. Desse
ensaio, a cepa mutante selecionada junto com a cepa WT (cepa controle) foram
avaliadas em biorreator com volume útil de 700 mL. Os resultados obtidos sugerem
que a via de acúmulo de lipídios utilizada pela levedura foi a via ex novo. Também
foi observado nos experimentos em biorreator, que os processos de acúmulo de
lipídios e crescimento celular alcançaram seus valores máximos em 24h, tempo mais
curto que os observados nos ensaios com menor volume (48h). Além disso, não
foram observadas diferenças, no ensaio em biorreator, no acúmulo de lipídeos e
crescimento celular da cepa mutante comparada a cepa controle.

xi
Abstract of a dissertation presented to Curso de Pós-Graduação em Tecnologia de Processos Químicos e Bioquímicos – EQ/UFRJ as partial fulfillment of the requiremments for the degree of Master of Science
INVOLVMENT OF PHOSPHATASES PROTEINS ON LIPID ACCUMU LATION IN Saccharomyces cerevisiae AND SYNTHESIS IN BIOREACTOR UTILIZING CRUDE
GLYCEROL AS CARBON SOURCE
ÉRIKA DE ARAUJO VALONI
The use of oleaginous microorganisms has been considered a promising technology,
as an alternative to the plant oils which are widely use in the manufacture of
biodiesel. Among the oleaginous microorganisms (these that can accumulate more
than 20% w/w, in their dry cell mass), the yeast shows advantages over the others
microorganisms in terms of growth and intracellular lipid content. The main fatty acids
in yeast oils are myristic, palmitic, stearic, oleic, linoleic and linolenic acids. It has
been reported that such oils can be used as oil feedstock for biodiesel production.
Added to this, the use of glycerol, in substitution of glucose, takes environmental
advantage, as it utilizes an industry residue and economic advantages, reducing the
cost of the microbial oil process. In the present work, it was possible to observe the
accumulation of triacylglicerides (TAG) inside the lipid bodies (LB) in both wild strain
and mutants for the protein-phosphatase genes in Saccharomyces cerevisiae,
cultivated under respiration metabolism, using glycerol as substrate. In an initial
experiment, was possible to determine the use of 5% (w/v) of crude glycerol as
carbon source to be use in the assays. Experiments have been made to evaluate the
best relation between the level of lipid bodies and cellular growth. From the first
screening analyzing the 66 mutant strains with deletion for the protein-phosphatase
genes, eight strains have been selected to be evaluate in flask with 50 mL of
medium. From this, it was selected a mutant strain that together with the wild strain (a
control) were evaluate in a bioreactor using 700 mL of medium. The results suggest
that the pathway utilized for the accumulation of lipids by the strains was the ex novo
pathway. It was also was observed, in the bioreactor experiments, that the processes
of lipid accumulation and cellular growth reached their maximum results in shorter
times than the observed in experiments in lower volumes. Besides this, were not

xii
observed differences, in the bioreactor experiment, in the accumulation and cellular
growth between the mutant and wild strains.

xiii
ÍNDICE DE FIGURAS
Figura 3.1: Anatomia de um corpúsculo lipídico: Imagem superior - Uma micrografia eletrônica de um
corpúsculo lipídico (Lipid Droplet) em células cultivadas de hepatoma. Imagem inferior – As
características estruturais de um corpúsculo lipídico. Retirado de: FARESE, JR. e WALTHER, 2009 ...... 9
Figura 3.2: Possível relação entre os corpúsculos lipídicos (aqui representados por LD) e o retículo
endoplasmático (representado por ER). (A) Os LDs e o ER estão frequentemente localizados próximos
e podem fazer contato entre as suas membranas. Mitocôndrias (M) também estão próximas aos LDs.
(B) A figura mostra um grande LD intercalado com a membrana do retículo. As setas indicam os
pontos onde a membrana do retículo endoplasmático é contínua com o corpúsculo lipídico. (C)
Conexão por uma haste da membrana. O brotamento pode ser constituído por ésteres lipídicos entre
o LD e a membrana do retículo. Retirado de: Fujimoto e Parton, 2011. .............................................. 10
Figura 3.3: Visão geral das principais vias metabólicas da síntese de lipídios em Y. lipolytica. A glicose,
através da glicólise entra na mitocôndria como piruvato para ser utilizado no ciclo do TCA; no
entanto, o excesso de acetil-CoA é transportado da mitocôndria para o citosol através do transporte
de citrato. O acetil-CoA no citosol é convertido em malonil-CoA pela acetil-CoA carboxilase (ACC)
sendo a primeira etapa da síntese dos ácidos graxos. Após isso, a síntese de triacilglicerol (TAG) segue
a via de Kennedy, a qual ocorre no retículo endoplasmático (RE) e corpúsculos lipídicos. O acil-CoA é
precursor utilizado para a acilação do esqueleto glicerol-3-fosfato para formar o ácido lisofosfatídico
(LPA), que é então acilado para formar o ácido fosfatídico (PA). PA é desfosforilado para formar o
diacilglicerol (DAG) e então acilado pela última vez através da diacilglicerol aciltransferase (DGA)
para produzir o TAG. OAA oxaloacetato, a-KG alfa-cetoglutarato, PEP fosfoenolpiruvato, G3P
gliceraldeído-3- fosfato, DHAP dihidroxiacetona fosfato, ACL ATP citrato liase, PC piruvato
carboxilase, MDH malato desidrogenase, MAE enzima málica. Retirado de: Tai e Stephanopoulos,
2013. ...................................................................................................................................................... 14
Figura 3.4: Síntese de ésteres de esterol (EEs) e triacilglicerol (TAG). O acetil-CoA gerado a partir de
diferentes substratos é direcionado para a síntese de ácidos graxos pela enzima acetil-CoA-
carboxilase (ACCase) ou para a síntese de esteróis pela 3-hidroxi-3-metil-glutaril-CoA redutase
(HMGR), ambas reguladas pela AMP quinase (AMPK). Os ácidos graxos são convertidos a lipídios
neutros, os TAG e EEs que são estocados nos corpúsculos lipídicos. Os principais reguladores desta
via são: AMPK (AMP quinase), FAS (ácido graxo sintase), DGAT (acil-CoA: diacilglicerol
aciltransferease, LCAT (lecitina:colesterol aciltransferase e ACAT (acil-CoA;colesterol aciltransferase.
Fonte:Bozaquel-Morais (2010).............................................................................................................. 18
Figura 3.5: A fosfatase Sit4 associada à sua subunidade Sap190p regula a atividade da Snf1p,
controlando sua fosforilação, e consequentemente modulando a atividade da ACC. Além de Sit4p-
Sap190p, Snf1p é desfosforilada por Glc7p-Reg1. Fonte: Bozaquel-Morais (2010) ............................. 20
Figura 4.1: Imagem de microscopia ótica da levedura Saccharomyces cerevisiae em aumento de
1000x utilizando microscópio ótico Nikon, modelo: Eclipse E200. ....................................................... 29
Figura 4.2: Reações enzimáticas envolvidas na quantificação dos triglicerídeos. (Fonte: kit enzimático
para dosagem de triacilglicerídeos. Doles, Brasil)................................................................................. 38
Figura 5.1: Avaliação do glicerol puro (P.A.) (A) e da glicerina loira (B) como fonte de carbono. ........ 40

xiv
Figura 5.2: Curvas de crescimento da cepa selvagem (WT) BY4741 de S. cerevisiae nos meios MM –
2% e MM – 5% (p/v) de glicerol puro (A) e glicerina loira (B) e MM – 5% (p/v) de glicose. ................. 41
Figura 5.3: Acúmulo de lipídios utilizando meios contendo 2% e 5% (p/v) de glicerol puro (A) e
glicerina loira (B). Índice de corpúsculos lipídicos (CL) obtidos pelo ensaio de recuperação de
fluorescência na cepa selvagem BY47421 (WT) de S. cerevisiae com 36h e 108h de ensaio. .............. 43
Figura 5.4: Distribuição das cepas mutantes em síntese de lipídio (A) e mutantes em proteínas
fosfatases (B), de acordo com a relação crescimento celular (medida por absorvância em 600 nm) e
índice de CL no tempo de 48 horas. ...................................................................................................... 46
Figura 5.5: Acúmulo de lipídios pelas cepas mutantes selecionadas em meio com 5% de glicerol em
glicerina loira. ........................................................................................................................................ 53
Figura 5.6: Média do crescimento das cepas mutantes selecionadas. O crescimento das mutantes de
S. cerevisiae foi medido por absorvância no comprimento de onda de 600 nm ao longo da
fermentação (n=3). ............................................................................................................................... 54
Figura 5.7: Acúmulo de lipídios pelas cepa mutante rrd2 e selvagem WT (n=1) .................................. 56
Figura 5.8: Acúmulo de lipídios pelas cepa mutante rrd2 e selvagem WT (n=4). ................................. 56
Figura 5.9: Acúmulo triacilgliceróis (TAG). O percentual de TAG nas cepas de S. cerevisiae selvagem
(WT) mutante RRD2, foram medidos no tempo final de reação em biorreator em 48 horas (n=4). ... 57
Figura 5.10: Crescimento celular das cepa mutante RRD2 e cepa selvagem WT. O crescimento celular
foi medido por absrovância no comprimento de onda de 600 nm (n=4). ............................................ 58
Figura 5.11: Consumo de glicerol no meio de cultivo. O glicerol foi medido por HPLC em amostras do
meio de cultivo contendo 5% de glicerina loira. ................................................................................... 58
Figura 5.12: Consumo de glicerol no meio de cultivo. O glicerol foi medido por kit enzimático (Doles,
Brasil) em amostras do meio de cultivo contendo 5% de glicerina loira. ............................................. 59

xv
ÍNDICE DE TABELAS
Tabela 3.1: Conteúdo lipídico e perfil de ácidos graxos de leveduras e fungos filamentosos
selecionadas. A porcentagem lipídica é dada em termos de biomassa seca. ........................................ 8
Tabela 4.1: Reagentes utilizados no preparo dos meios de cultivo e os respectivos fabricantes. ....... 25
Tabela 4.2: Reagentes utilizados nos ensaios analíticos e os respectivos fabricantes ......................... 26
Tabela 4.3: Experimentos preliminares para avaliação da fonte de carbono utilizada. ....................... 32
Tabela 5.1: Cepas de S. cerevisiae mutantes deletadas para genes de síntese de lipídios e para
subunidades catalíticas e regulatórias não essenciais de fosfatases submetidos a screening pelo
método de recuperação de fluorescência e densidade ótica (DO600 nm) que não apresentaram
crescimento celular satisfatório. ........................................................................................................... 47
Tabela 5.2: Cepas de S. cerevisiae mutantes deletadas para genes de síntese de lipídios e para
subunidades catalíticas e regulatórias não essenciais de fosfatases submetidos a screening pelo
método de recuperação de fluorescência e densidade ótica (DO600 nm). As cepas destacadas em verde
foram selecionadas como hlc (alto conteúdo lipídico). ........................................................................ 48
Tabela 5.3: Cepas de S. cerevisiae mutantes deletadas para genes de síntese de lipídios submetidos a
screening pelo método de recuperação de fluorescência e densidade ótica (DO600 nm). ................. 51
Tabela 5.4: Cepas selecionadas de acordo com a média do índice de CL e a média de crescimento
celular, medida por densidade ótica em 600 nm (DO600 nm). ............................................................ 52

xvi
LISTA DE ABREVIATURAS
ACAT Acil-CoA aciltransferase
ACCase Acetil-CoA carboxilase
ACL ATP citrato liase
AMP Adenosina monofosfato
AMPK Adenosina monofosfato quinase
ATP Adenosina trifosfato
Aox Acil-CoA oxidases
BSA Bovine Serum Albumin - Albumina do soro bovino
CL Corpúsculos lipídicos
DAG Diacilglicerol
DGA Diacilglicerol aciltranferases
DGAT2
D.O. Densidade ótica
D.O.600nm Densidade ótica no comprimento de onda 600 nm
EE Éster de esterol
ERF Ensaio de recuperação de fluorescência
FAME Ésteres metílicos de ácidos graxos
FAS Ácido graxo sintase
G3P Glicerol 3-fosfato
hlc High lipid content
HMGR 3-hidroxi-3-metil-glutaril-CoA redutase
HPLC High-Performance Liquid Cromatography – Cromatografia líquida de alta eficiência
ICL Índice de corpúsculos lipídicos
IMP Inosina monofosfato
KAN Kanamicina
LCAT lecitina colesterol aciltransferase

xvii
llc Low lipid content
LPA Ácido lisofosfatídico
MM Meio mínimo
MONG Máteria orgânica não glicerinada
PA Ácido fosfatídico
P.A. Pró-análise
p/v Peso por volume
PP Proteínas fosfatases
PPM Proteínas fosfatases dependente de magnésio
PP2A Proteína fosfatase do tipo 2A
PPP Fosfoproteína fosfatase
PTP Fosfoproteína tirosina fosfatase
RE Retículo endoplasmático
RMN Ressonância magnética nuclear
rpm Rotações por minuto
SCO Single cell oil
Ser Serina
TAG Triacilglicerol
TCA Ácido tri-cloroacético
Thr Treonina
WT Wild Type - Cepa do tipo selvagem
YPD Yeast extract, peptone, dextrose
µ Taxa específica de crescimento (h-1)

xviii
SUMÁRIO
1. Introdução ....................................................................................................................................... 1
2. Justificativas e Objetivos ................................................................................................................. 4
3. Revisão Bibliográfica ....................................................................................................................... 6
3.1 Micro-organismos oleaginosos ............................................................................................... 6
3.2 Conteúdo lipídico e composição dos ácidos graxos presentes nas leveduras oleaginosas .... 7
3.3 Corpúsculos lipídicos ............................................................................................................... 8
3.4 Vias de acúmulo de lipídios em leveduras ............................................................................ 11
3.5 Síntese e acúmulo de lipídios utilizando substratos hidrofóbicos no meio de cultivo ......... 15
3.5.1 A bioquímica do acúmulo de lipídios através da via ex novo ........................................ 16
3.6 Regulação do metabolismo de lipídios ................................................................................. 17
3.6.1 As proteínas fosfatases na regulação do metabolismo lipídico .................................... 19
3.7 Manipulação genética de leveduras ..................................................................................... 20
3.8 Escalonamento para o desenvolvimento de bioprocessos ................................................... 22
4. Materiais e Métodos ..................................................................................................................... 25
4.1 Materiais ............................................................................................................................... 25
4.2 Equipamentos ....................................................................................................................... 27
4.3 Meios de cultura ................................................................................................................... 28
4.4 Micro-organismos ................................................................................................................. 28
4.4.1 Preservação ................................................................................................................... 29
4.4.2 Obtenção do pré - inoculo ............................................................................................. 29
4.4.3 Curvas de crescimento e cálculo do volume de inóculo ............................................... 30
4.5 Experimentos Preliminares ................................................................................................... 31
4.5.1 Avaliação do crescimento celular e acúmulo de lipídios em concentrações crescentes
de glicerol e glicerina loira. ........................................................................................................... 31
4.5.2 Varredura das cepas de Saccharomyces cerevisiae mutantes para avaliar o índice de
corpúsculos lipídicos, quando cultivadas em glicerina loira 5% ................................................... 32
4.5.3 Avaliação das cepas mutantes para gene fosfatase em meio 5% de glicerina loira ..... 33
4.6 Avaliação do acúmulo de corpúsculos lipídicos em biorreator por Saccharomyces cerevisiae
mutante utilizando como fonte de carbono 5% de glicerina loira .................................................... 34
4.7 Métodos Analíticos................................................................................................................ 35
4.7.1 Determinação de Biomassa ........................................................................................... 35
4.7.2 Ensaio de recuperação de fluorescência (ERF) para determinação do Índice de
Corpúsculos Lipídicos (ICL) ............................................................................................................ 35

xix
4.7.3 Determinação da concentração de glicerol presente no meio de cultivo .................... 36
4.7.4 Dosagem enzimática de triacilglicerol ........................................................................... 36
4.7.5 Determinação de Proteínas Totais ................................................................................ 38
5. Resultados e Discussão ................................................................................................................. 39
5.1 Avaliação de fontes de carbono alternativas no meio de cultivo para acúmulo de lipídios . 39
5.2 Varredura das cepas de Saccharomyces cerevisiae mutantes para escolha da cepa com
maior índice de acúmulo de lipídios crescida em glicerina loira 5% ................................................. 44
5.3 Avaliação das cepas mutantes para os genes fosfatase em meio com 5% de glicerol em
glicerina loira ..................................................................................................................................... 52
5.4 Avaliação do acúmulo de corpúsculos lipídicos na cepa mutante rrd2 de Sacchromyces
cerevisiae em biorreator utilizando glicerina loira como fonte de carbono ..................................... 55
6. Conclusões..................................................................................................................................... 63
7. Perspectivas e trabalhos futuros ................................................................................................... 65
8. Referências Bibliográficas ............................................................................................................. 66
9. Apêndices ...................................................................................................................................... 77

Capítulo 1 – Introdução Valoni, E.A.
1
1. Introdução
O aumento da população mundial e da prosperidade global está causando um
acréscimo significativo na demanda energética global. Para reduzir a dependência
da energia fóssil, metas para incorporação de energia renovável estão sendo
impostas no mundo todo (HEERES et al., 2014).
Segundo dados da Administração de Informação de Energia dos Estados
Unidos (2011), o transporte é o setor que mais cresce, e o segundo maior setor
consumidor de combustíveis nos E.U.A. (28%) depois da energia elétrica (40%).
Alternativas ao petróleo, para combustíveis líquidos e seu uso em transporte, estão
sendo procuradas devido às preocupações globais sobre as mudanças climáticas,
segurança energética e ao esgotamento iminente das reservas de petróleo.
O biodiesel é altamente degradável, não tóxico, renovável e sustentável. Ele
tem propriedades de combustão similares ao diesel de petróleo, é um combustível
de substituição chamado “drop-in”, para veículos a diesel existentes e motores de
caldeiras industriais, além de ser compatível com a infraestrutura atual de
combustível (SITEPU et al., 2014). A produção mundial de biodiesel está
aumentando fortemente para atender a demanda: a produção passou de 15 mil
barris/dia, no ano 2000, para 289.000 em 2008 (ATABANI et al., 2012). O óleo
vegetal compõe a maior parte do custo de produção do biodiesel (MIAO et al., 2006)
e o debate público sobre o tema “comida x combustível”, incentivaram o
desenvolvimento da chamada “segunda geração” de biodiesel não comestível, a
partir de óleos vegetais como o pinhão manso, mamona, além de óleos residuais
domésticos. No entanto, estes óleos podem não ser suficientes para atender às
necessidades globais e apresentar características que não se enquadram nas
especificações de qualidade do biodiesel (SITEPU et al., 2014).
Os óleos microbianos, também chamados de single cell oil (SCO), são
produzidos por alguns micro-organismos oleaginosos como leveduras, fungos
filamentosos, bactérias e microalgas (MA, 2006). Comparado com os óleos vegetais,
o óleo microbiano traz muitas vantagens como, por exemplo, o biodiesel dessa
origem ser capaz de ser produzido durante o ano todo, em terras consideradas
impróprias para a agricultura (não competindo com a produção de alimentos) e com

Capítulo 1 – Introdução Valoni, E.A.
2
taxas de produção acima de 100 vezes às alcançadas pelos óleos vegetais em
litros/hectares/ano (ATABANI et al., 2012). O custo de produção de SCO é
atualmente considerado elevado, por isso, substratos para o cultivo de micro-
organismos oleaginosos com baixo custo de aquisição tem atraído o interesse de
diversos grupos de pesquisa (BELLOU et al., 2012). Nessa perspectiva, o glicerol
bruto (também conhecido como glicerina) é uma matéria prima muito promissora,
porque é um co-produto da fabricação do biodiesel que consiste numa mistura de
glicerol (65-85%, p/p), metanol, sabões e sais minerais ( DA SILVA et al., 2009), que
podem ser valorizados na produção de lipídios.
Uma compreensão dos fatores genéticos que contribuem para o acúmulo de
lipídios é crucial para o desenvolvimento de cepas com essa capacidade de acúmulo
de lipídios. A regulação das vias metabólicas e de produção de enzimas-chave pode
ser manipulada para levar o carbono aos produtos desejados. Sistemas genéticos
foram desenvolvidos para algumas poucas espécies de leveduras, inicialmente S.
cerevisiae e Y. lipolytica. Os genomas dessas espécies de levedura foram
sequenciados para facilitar o desenvolvimento de ferramentas genéticas e
melhoramento de vias metabólicas (BEOPOULOS et al., 2009; 2010). A
manipulação genética tem gerado resultados no desenvolvimento de cepas de
leveduras com aumento ou diminuição da habilidade de formar os corpúsculos
lipídicos.
No trabalho desenvolvido por Bozaquel-Morais (2010), que teve como objetivo
determinar o papel da fosforilação na regulação da dinâmica de formação dos
corpúsculos lipídicos, foram avaliadas cepas mutantes de S. cerevisiae deletadas
para os genes das proteínas-fosfatases. Foram identificadas 13 cepas, das 65 cepas
mutantes testadas, que apresentaram níveis de corpúsculos lipídicos anormais.
Esse estudo foi capaz de identificar o envolvimento das proteínas-fosfatases na
síntese de lipídios em leveduras pela primeira vez na literatura.
Portanto, os óleos microbianos podem se tornar uma das potenciais matérias-
primas para a produção de biodiesel no futuro, embora ainda existam muitos
trabalhos associados com os micro-organismos produtores de óleo que precisam ser
desenvolvidos com maior intensidade (LI et al., 2008).

Capítulo 1 – Introdução Valoni, E.A.
3
Sendo assim, de acordo com a necessidade de contribuir para o avanço das
tecnologias de produção de óleo microbiano, o presente trabalho pretende estudar o
papel das proteínas-fosfatases no metabolismo de acúmulo de lipídios em cepas
geneticamente modificadas de S. cerevisiae cultivadas em escala de biorreator de
1L, em meio contendo como única fonte de carbono a glicerina loira, proveniente da
fabricação do biodiesel.

Capítulo 2 – Justificativas e Objetivos Valoni, E.A.
4
2. Justificativas e Objetivos
O desenvolvimento de plataformas sustentáveis utilizando micro-organismos
para a produção de bioprodutos e biocombustíveis está se tornando cada vez mais
crucial, a medida que as reservas de petróleo são diminuídas e os problemas
ambientais aumentam ao redor do mundo (ABGHARI et al., 2014). Os processos de
de bioconversão baseados nos micro-organismos podem ser utilizados para produzir
uma variedade de produtos, como biocombustíveis, biopolímeros e biossurfactantes,
a partir de uma vasta oferta de biomassa.
Muitas estratégias têm sido exploradas para melhorar a produtividade e a
competitividade de processos baseados nos micro-organismos. Para isso, deve-se
incluir nos estudos a manipulação genética e metabólica, tecnologias de
fermentação, matérias-primas de menor valor e o reuso de subprodutos (KOUTINAS
et al., 2014).
De uma maneira geral, os biolipídios produzidos pelas leveduras
oleaginosas, chamados de Single Cell Oil (SCO) são matéria prima adequada para a
produção de biodiesel (TANIMURA et al., 2014). Vários substratos utilizados para a
sua produção já foram reportados como o glicerol industrial (obtido da produção de
biodiesel ou da saponificação de gorduras) (PAPANIKOLAOU et al., 2002). A
levedura Saccharomyces cerevisiae é uma plataforma popular de engenharia
metabólica, assim como um micro-organismo produtor bastante atraente na
biotecnologia industrial. Esse status é o resultado de fatores como a intensa
experiência com esse micro-organismo em bioprocessos industriais, a facilidade de
realizar manipulações genéticas e sua robustez sob condições de processos
(SWINNEN et al, 2013).
O objetivo geral deste trabalho é aumentar o acúmulo de triacilglicerol, dentro
dos corpúsculos lipídicos, em Saccharomyces cerevisiae, para a futura produção de
biodiesel e aplicação em biocombustíveis. Para isto é importante identificar o papel
das proteínas fosfatases assim como os seus reguladores, no acúmulo de
corpúsculos lipídicos na levedura cultivada em condição respiratória, utilizando
glicerina loira como matéria prima. As modificações genéticas que apresentaram os

Capítulo 2 – Justificativas e Objetivos Valoni, E.A.
5
melhores resultados na relação de acúmulo lipídico e crescimento celular na
levedura Saccharomyces cerevisiae foram alvos para modificação na levedura
Yarrowia lipolytica, que é um dos micro-organismos oleaginosos com maior potencial
de aplicação industrial. Para alcançar o objetivo geral do projeto, este foi
segmentado nas seguintes etapas:
1. Avaliar o crescimento da levedura selvagem utilizando diferentes
concentrações de glicerol, visando determinar a melhor concentração de
substrato (% p/v) para a melhor relação entre o índice de corpúsculos
lipídicos (CL) e o crescimento celular de Saccharomyces cerevisiae.
2. Realizar uma varredura das cepas mutantes com deleções simples nos
genes que codificam para proteínas-fosfatases e suas subunidades
quando não essenciais (66 mutantes) e selecionar as cepas mutantes que
apresentem as melhores relações entre o índice de CL e crescimento
celular em meio contendo glicerol como única fonte de carbono.
3. Testar as cepas mutantes para os genes das proteínas-fosfatases que
foram selecionadas na varredura, em volumes de 50 mL de meio mínimo
(MM) contendo glicerol, em frascos do tipo erlenmeyer agitados em
agitador orbital sob temperatura de 30ºC.
4. Avaliar e comparar o comportamento da cepa mutante (selecionada no
ensaio anterior) e selvagem (WT) de Saccharomyces cerevisiae em
relação ao índice de CL e crescimento celular em biorreator de bancada.
5. Medir e avaliar o consumo de glicerol no meio reacional ao longo da
fermentação em biorreator.

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
6
3. Revisão Bibliográfica
3.1 Micro-organismos oleaginosos
A utilização de micro-organismos oleaginosos (que podem acumular lipídios
em mais de 20% p/p em seu peso seco), como fonte de óleos e gorduras em
grandes escalas de operação, representa um grande interesse industrial
(PAPANIKOLAOU et al., 2011). Segundo Sitepu et al. (2014), a chamada “3ª.
geração" de biodiesel está sendo desenvolvida utilizando micro-organismos
acumuladores de óleos, tais como microalgas, bactérias, leveduras e outros fungos.
Estudos demonstram que leveduras são mais eficientes que bactérias e
microalgas em termos de acúmulo de lipídios (ENSHAEIEH et al., 2012). Além disso,
a levedura traz vantagens como, não ser influenciada pelo clima, como as
microalgas fotoautotróficas, e poder utilizar diversas fontes de açúcares, como a
xilose, em taxas de crescimento maiores do que as microalgas heterotróficas (CHI et
al., 2011). As leveduras também demonstram maior tolerância aos íons metálicos e
tolerância à baixa disponibilidade de oxigênio, quando comparadas aos fungos
filamentosos. Diferentemente da bactéria E. coli, a levedura possui compartimentos
intracelulares, maior tamanho celular (facilitando a sua separação), maior tolerância
a co-produtos com potenciais inibitórios (como ácidos orgânicos) e alto acúmulo do
produto final (BEOPOULOS et al., 2010; AGEITOS et al., 2011).
Por décadas as leveduras oleaginosas têm sido estudadas como uma fonte
microbiana de óleos para oleoquímicos, incluindo combustíveis, produtos químicos
de plataforma e ingredientes para alimentos e rações (WOODBINE, 1959). Muitas
descobertas foram feitas nas primeiras décadas de trabalho sobre o óleo de
levedura, o que impulsionaram estudos modernos, incluindo a descoberta de
espécies que apresentavam alto acúmulo de óleos, as melhores condições de
cultivo, a conversão de resíduos em lipídios e a produção em escala piloto e
comercial de produtos para o mercado (SITEPU et al., 2014). Uma empresa de
óleos microbianos, com sede na Califórnia, chamada Solazyme, solicitou diversas
patentes para o uso de óleos de leveduras para combustíveis, químicos e

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
7
ingredientes alimentares (FRANKLIN et al., 2011; 2011b), evidenciando um forte
interesse industrial em leveduras oleaginosas.
As leveduras e fungos filamentosos oleaginosos mais conhecidas incluem os
gêneros Candida, Cryptococcus, Rhodotorula, Rhizopus, Trichosporon e Yarrowia.
Em média, estas leveduras acumulam lipídios em um nível correspondente a 40%
de sua biomassa (LI et al., 2008, MENG et al., 2009). Em condições de limitação de
nutrientes, elas podem acumular lipídios para níveis que excedem 70% de sua
biomassa.
3.2 Conteúdo lipídico e composição dos ácidos graxo s presentes nas
leveduras oleaginosas
Os lipídios são moléculas biológicas estruturalmente diversas que são
relativamente insolúveis em água e solúveis em solventes orgânicos. Eles incluem
duas categorias básicas: (1) moléculas derivadas de unidades de isopreno, como
esteróis e carotenoides, e (2) ácidos graxos e as moléculas relacionadas de cadeia
longa, como álcoois graxos, alcanos e alcenos (SITEPU et al., 2014).
As leveduras contêm vários tipos de lipídios, incluindo triacilgliceróis (TAG),
diacilgliceróis (DAG), monoacilgliceróis, ácidos graxos, ésteres de esterol, esteróis
livres e outros. Os lipídios são encontrados principalmente sob a forma de lipídios
neutros, glicolipídios, fosfolipídios e ácidos graxos livres (DEY e MAITI, 2013). As
frações de lipídios neutros, em geral, somam mais de 90% dos lipídios totais,
enquanto as frações não neutras representam menos que 10% (HUANG et al.,
2013). As classes de lipídios presentes, assim como a proporção entre lipídios de
membrana e de armazenamento, variam entre as espécies e as estirpes, condições
de cultivo e fase de crescimento da cultura (SITEPU et al., 2014).
Os triacilgliceróis (TAG) e ésteres de esterol são os principais lipídios neutros
e são sintetizados a partir do esterol, glicerol-3-fosfato e acil-CoA. A composição dos
ácidos graxos que formam os TAG intracelulares é variável, e depende da espécie
de levedura, fase de crescimento, condições ambientais e substratos do meio de
cultivo (ABGHARI et al., 2014).

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
8
Os ácidos graxos mais comuns nas leveduras são os que apresentam 16 ou 18
átomos de carbono de comprimento (C16 e C18, respectivamente), saturados (sem
duplas ligações, como o C16:0) ou monoinsaturados (uma dupla ligação, como o
C18:1). Sendo que os C18:1 (ácido oleico), C16:0 (ácido palmítico) e C18:0 (ácido
esteárico) são os que aparecem em maiores concentrações, como mostra a Tabela
3.1 (BEOPOULOS et al., 2009).
Estes também são os ácidos graxos principais dos óleos vegetais utilizados
na produção do biodiesel, como o óleo de canola e girassol (AGEITOS et al., 2011).
3.3 Corpúsculos lipídicos
Os ácidos graxos e esteróis são importantes blocos construtores das
membranas celulares, e para manter os níveis celulares balanceados, certas
quantidades são armazenadas na forma de lipídios neutros, principalmente sob a
forma de triacilgliceróis e ésteres de esteróis. Porém, como ambas as moléculas são
insolúveis no citoplasma, elas precisam ser estocadas em compartimentos celulares
que são conhecidos como corpúsculos lipídicos (CL) (CZABANY et al., 2008).
Espécie Principais Ácidos Graxos (% relativa p/p)
Lipídios (%) C16:0 C16:1 C18:0 C18:1 C18:2 C18:3
Cryptococcus curvatus 58 25 Ta 10 57 7 N.D.b
Cryptococcus albidus 65 12 1 3 73 12 0
Candida sp. 107 42 44 5 8 31 9 1
Lipomyces starkeyi 63 34 6 5 51 3 0
Rhodotorula glutinis 72 37 1 3 47 8 0
Rhodotorula graminis 36 30 2 12 36 15 4
Rhizopus arrhizus 57 18 0 6 22 10 12
Trichosporon pullulans 65 15 0 2 57 24 1
Yarrowia lipolytica 36 11 6 1 28 51 1
Tabela 3.1: Conteúdo lipídico e perfil de ácidos graxos de le veduras e fungos filamentosos selecionadas. A porcentagem lipídica é dada em term os de biomassa seca.
Ta: quantidade traço N.D.b: não detectado
Fonte: Beopoulos et al., 2009.

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
9
De acordo com MLÍĈKOVÁ et al. (2004), a estrutura desses compartimentos
é similar em todos os tipos de células, sendo bastante simples. Os CL consistem de
um núcleo formado pelos lipídios neutros, que são envolvidos por uma monocamada
de fosfolipídios com algumas proteínas incorporadas (Figura 3.1)
Figura 3.1: Anatomia de um corpúsculo lipídico: Ima gem superior - Uma micrografia eletrônica
de um corpúsculo lipídico ( Lipid Droplet ) em células cultivadas de hepatoma. Imagem inferio r
– As características estruturais de um corpúsculo l ipídico. Retirado de: FARESE, JR. e
WALTHER, 2009

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
10
Diversas hipóteses para explicar o mecanismo de formação dos CL foram
propostas. O modelo mais aceito indica que a biogênese dos CL provavelmente
resulta do brotamento do retículo endoplasmático (RE) (MURPHY et al., 2001), no
qual as enzimas no retículo endoplasmático sintetizam os lipídios neutros que são
depositados entre duas camadas da membrana do RE para formar um pré-
corpúsculo lipídico. Quando essa estrutura atinge certo tamanho, ela brota para
formar um CL maduro, conforme a Figura 3.2. (ZWEYTICK et al., 2000).
Segundo Czabany et al. (2008) e Murphy et al. (2012), algumas espécies de
leveduras podem acumular somente TAG nos corpúsculos lipídicos, enquanto outras
podem acumular somente ésteres de esterol, e ainda existem outras, como S.
cerevisiae, que podem acumular os dois.
Figura 3.2: Possível relação entre os corpúsculos l ipídicos (aqui representados por LD) e o
retículo endoplasmático (representado por ER). (A) Os LDs e o ER estão frequentemente
localizados próximos e podem fazer contato entre as suas membranas. Mitocôndrias (M)
também estão próximas aos LDs. (B) A figura mostra um grande LD intercalado com a
membrana do retículo. As setas indicam os pontos on de a membrana do retículo
endoplasmático é contínua com o corpúsculo lipídico . (C) Conexão por uma haste da
membrana. O brotamento pode ser constituído por ést eres lipídicos entre o LD e a membrana
do retículo. Retirado de: Fujimoto e Parton, 2011.

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
11
Uma das principais funções atribuídas aos CL é manter a homeostase celular
de lipídios, entretanto, nos últimos anos os CL têm sido caracterizados não apenas
como armazenadores de lipídios neutros, mas também como estruturas
intracelulares dinâmicas, envolvidas em diversos processos metabólicos (LEBER et
al., 1994).
Em S. cerevisiae, cerca de 40 proteínas estão presentes na superfície dos
corpúsculos lipídicos, e a maioria delas está envolvida com o metabolismo lipídico.
Como exemplo, o CL de S. cerevisiae ancora enzimas da biossíntese do ácido
fosfatídico (ATHENSTAEDT et al., 1999), ativação de ácidos graxos (WATKINS et
al., 1998), biossíntese de esteróis (ZINSER et al. 1993), assim como enzimas para
biossíntese e degradação de TAG (ATHENSTAEDT et al., 2005) e hidrólise de
ésteres de esterol (JANDROSITZ et al. 2005).
Por essas características, os CL encontrados em micro-organismos são
também alvos de muitos estudos para fins biotecnológicos, principalmente em
leveduras oleaginosas, como Yarrowia lipolytica (BEOPOULOS et al., 2009).
3.4 Vias de acúmulo de lipídios em leveduras
Todos os micro-organismos são capazes de sintetizar lipídios, porém,
somente as estirpes oleaginosas podem acumular quantidades significantes dentro
de suas células (PAPANIKOLAOU e AGGELIS, 2011).
Segundo BEOPOLULOS et al. (2012), os lipídios podem ser acumulados
dentro da célula a partir de duas vias diferentes, conforme a seguir.
• Via de síntese de novo: envolvendo a produção de percursores de ácidos
graxos, acetil-CoA e malonil-CoA, e sua integração com a biossíntese
lipídica. Normalmente esta via é utilizada quando o crescimento do micro-
organismo é conduzido utilizando glicose ou glicerol como substrato e com a
limitação de nitrogênio.

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
12
• Via de acúmulo ex novo: envolvendo o acúmulo de ácidos graxos, óleos e
TAGs provenientes do meio de cultura e o seu armazenamento, sem
modificações, dentro da célula. Esta via necessita que os ácidos graxos livres
sejam transportados para dentro da célula. A remontagem em frações de
TAGs e ésteres de esterol e o armazenamento são realizados dentro do
corpo lipídico.
Em uma série de estudos, foi demonstrado que os micro-organismos
oleaginosos não possuem um sistema hiperativo de biossíntese de ácidos graxos,
mas eles são capazes de produzir em quantidades significativas, acetil-CoA, a
unidade básica para a biossíntese de ácidos graxos. Dessa forma, a bioquímica da
síntese de novo pode ser dividida em duas partes distintas: o metabolismo celular
intermediário e a biossíntese de TAGs (PAPANIKOLAOU e AGGELIS, 2011).
Durante a síntese pela via de novo, o início do acúmulo de lipídios é induzido
pela exaustão ou limitação de um nutriente essencial do meio de cultura.
Usualmente, o nitrogênio é utilizado com esse propósito, pois a sua adição é fácil de
controlar. Quando o nitrogênio se torna indisponível a divisão celular diminui, pois
ele é crucial pra a síntese de proteínas e ácidos nucleicos. No entanto, o organismo
continua a assimilar a fonte de carbono (açúcares ou glicerol) do meio, que agora é
direcionada para a síntese de lipídios, ocorrendo o acúmulo de TAGs no interior da
célula e a formação de CL.
O acúmulo de lipídios por micro-organismos oleaginosos está relacionado
com a produção de ácido cítrico e a atividade da enzima isocitrato desidrogenase,
pertencente ao conjunto de enzimas presentes no Ciclo de Krebs (CK). A enzima
isocitrato desidrogenase é dependente de adenosina monofosfato (AMP). A AMP
desaminase cliva AMP em IMP (inosina monofosfato), na ausência de nitrogênio
proveniente do meio de cultivo. O grande decréscimo da concentração de AMP
intracelular altera as funções do CK. Com a alteração do ciclo, a isocitrase
desidrogenase, enzima responsável pela transformação do ácido isocítrico em ácido
α-cetoglutárico, perde sua atividade, visto que a mesma é ativada alostericamente
por AMP intracelular. Então, o ácido isocítrico é acumulado dentro da mitocôndria e
encontra-se em equilíbrio com o ácido cítrico (reação catalisada por isocitrato
aconitase). Quando a concentração de ácido cítrico dentro da mitocôndria alcança

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
13
valores críticos, ele é transportado para o citosol através do transporte citrato-malato
e é clivado pela enzima ATP citrato liase (ACL) formando oxaloacetato e acetil-CoA.
O acetil-CoA, unidade básica da biossíntese de ácidos graxos, é o precursor da
síntese lipídica de novo por micro-organismos oleaginosos (PAPANIKOLAOU e
AGGELIS, 2011).
De acordo com Beopoulos et al. (2008), o TAG é sintetizado a partir do acil-
CoA no retículo endoplasmático (RE) ou na membrana do CL através da via de
Kennedy. Esta via consiste em três etapas principais, através das quais, os ácidos
graxos são esterificados no esqueleto de glicerol 3-fosfato (G3P). A primeira
acilação do G3P para ácido lisofosfatídico (LPA) é seguida pela segunda acilação
para ácido fosfatídico (PA). A etapa final da acilação ocorre pelas três classes de
enzimas diacilglicerol aciltransferases (DGA) (ex.: DGAT1, DGAT2 e PDAT) (Zhang
et al., 2012). Um esquema com as principais etapas envolvidas na síntese dos
ácidos graxos e armazenamento de lipídios é apresentada na Figura 3.3.

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
14
Figura 3.3: Visão geral das principais vias metaból icas da síntese de lipídios em Y. lipolytica .
A glicose, através da glicólise entra na mitocôndri a como piruvato para ser utilizado no ciclo
do TCA; no entanto, o excesso de acetil-CoA é tran sportado da mitocôndria para o citosol
através do transporte de citrato. O acetil-CoA no c itosol é convertido em malonil-CoA pela
acetil-CoA carboxilase (ACC) sendo a primeira etapa da síntese dos ácidos graxos. Após isso,
a síntese de triacilglicerol (TAG) segue a via de K ennedy, a qual ocorre no retículo
endoplasmático (RE) e corpúsculos lipídicos. O acil -CoA é precursor utilizado para a acilação
do esqueleto glicerol-3-fosfato para formar o ácido lisofosfatídico (LPA), que é então acilado
para formar o ácido fosfatídico (PA). PA é desfosfo rilado para formar o diacilglicerol (DAG) e
então acilado pela última vez através da diacilgli cerol aciltransferase (DGA) para produzir o
TAG. OAA oxaloacetato, a-KG alfa-cetoglutarato, PEP fosfoenolpiruvato, G3P gliceraldeído-3-
fosfato, DHAP dihidroxiacetona fosfato, ACL ATP cit rato liase, PC piruvato carboxilase, MDH
malato desidrogenase, MAE enzima málica. Retirado d e: Tai e Stephanopoulos, 2013.

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
15
O mecanismo geral do acúmulo de lipídios em micro-organismos oleaginosos
ainda não foi totalmente esclarecido (MORIN et al., 2011). O fato de que várias
etapas de síntese são realizadas em diferentes compartimentos celulares cria uma
maior complexidade, já que existe a necessidade de transporte de compostos
intermediários entre esses compartimentos.
O acúmulo de lipídios usualmente não está associado à propriedades
de crescimento (AGEITOS et al., 2011). No entanto, uma porção significativa de
lipídios pode ser obtida durante o início da fase de crescimento (ANDRÉ et al.,
2009). O acúmulo de lipídios pode ocorrer durante a fase exponencial, em resposta
a necessidade de síntese de membrana (SHA, 2013), e geralmente alcança os
níveis máximos no início da fase estacionária, quando as células esgotaram os
nutrientes, mas ainda têm excesso de carbono no meio (TAI e STEPHANOPOULOS,
2013).
3.5 Síntese e acúmulo de lipídios utilizando substr atos hidrofóbicos no meio
de cultivo
A identificação de micro-organismos capazes de crescer e produzir lipídios a
partir de resíduos ou subprodutos do setor agroindustrial (FAKAS et al., 2008;
PAPANIKOLAOU et al., 2009), valorizando esses resíduos junto com a produção de
lipídios, pode aumentar a viabilidade do processo e ser simultaneamente benéfica
para o meio ambiente.
Poucas leveduras foram descritas como capazes de crescer em substratos
hidrofóbicos e ao mesmo tempo acumular quantidades significativas de lipídios
intracelulares. No entanto, é sabido que um maior número de micro-organismos é
capaz de consumir ácidos graxos livres, uma vez que o crescimento nesse material
é realizado independentemente da capacidade lipolítica do micro-organismo
(PAPANIKOLAOU et al., 2011). Em contraste, os micro-organismos que são
capazes de quebrar TAG ou ésteres de ácidos graxos, devem, obrigatoriamente,
possuir um sistema de lipases ativos em seu arsenal enzimático (PAPANIKOLAOU
E AGGELIS, 2010; AGGELIS et al., 1997; AGGELIS et al., 1995).

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
16
Um aspecto interessante de ser observado é que quando o crescimento das
leveduras é realizado na presença de substâncias hidrofóbicas (a chamada via de
acúmulo de lipídios ex novo), o lipídio microbiano produzido contém menores
quantidades de TAG comparado com o cultivo em meios baseados em açúcar (a
chamada via de acúmulo de novo) (PAPANIKOLAOU et al., 2001; THORPE e
RATLEDGE, 1972; PAPANIKOLAOU et al., 2002b).
3.5.1 A bioquímica do acúmulo de lipídios através d a via ex novo
Os ácidos graxos livres (presentes como substratos iniciais, ou produzidos
depois da ação das lipases nos TAG ou ésteres de ácidos graxos) são incorporados
na célula através do auxílio do transporte ativo. Os ácidos graxos incorporados são
então divididos para as necessidades de crescimento ou se tornam substratos para
transformações endocelulares (síntese de “novos” perfis de ácidos graxos que não
existiam anteriormente no substrato) (GUO et al., 1999; KINOSHITA et al., 2001;
PAPANIKOLAOU et al., 2002b).
Os ácidos graxos livres vão ser degradados pelo processo da β-oxidação, em
cadeias menores de acil-CoA e acetil-CoA, em reações catalisadas por várias acil-
CoA oxidases (Aox) (FICKERS et al., 2005) fornecendo dessa forma, primeiramente
a energia necessária para o crescimento e manutenção das células e depois a
formação de substâncias orgânicas (metabólitos intermediários) que constituem os
precursores para síntese de materiais celulares.
Uma das principais diferenças entre as sínteses de novo e ex novo, é que no
último caso, o acúmulo de lipídios ocorre simultaneamente com o crescimento
celular, sendo completamente independente do esgotamento de nitrogênio no meio
de cultivo (PAPANIKOLAOU et al., 2002; AGGELIS et al., 1997).
Ainda de acordo com Papanikolaou et al. (2011), quando o material
hidrofóbico é utilizado como única fonte de carbono e energia, não é observada
qualquer síntese pela via de novo a partir do acetil-CoA. Isso ocorre, pois a enzima
ácido graxo sintase (FAS) e a ATP-citrato liase (ACL) são fortemente inibidas pela
presença de cadeias alifáticas exógenas.

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
17
3.6 Regulação do metabolismo de lipídios
A síntese dos lipídios deve ser fortemente regulada, para que sua toxicidade
seja prevenida sem que suas funções celulares sejam prejudicadas. No entanto, os
mecanismos exatos de regulação, que controlam a expressão e a atividade das
enzimas que contribuem para a formação dos CL, não estão ainda completamente
esclarecidos.
A etapa limitante da via de síntese de lipídios é a produção de malonil-CoA e
essa reação é catalisada pela enzima acetil-CoA carboxilase (ACCase). As
leveduras apresentam, assim como os mamíferos, duas isoformas de ACCase. A
deleção dos genes que as codfica, é possível, no entanto, a mutante não é capaz de
crescer em substratos não-fermentativos (HOJA et al., 2004). De acordo com
Munday et al. (1988), a atividade de ACCase é finamente controlada por fosforilação
reversível e quando o estado energético da célula é baixo, a proteína-quinase
ativada por AMP (AMPK), é a responsável por fosforilar diretamente a ACCase no
resíduo Ser e assim inibindo a enzima (Figura 3.4).
Ainda não está claro qual proteína fosfatase é a responsável pela
desfosforilação, e portanto, ativação da ACCase (BOZAQUEL-MORAIS, 2010).
Porém, experimentos da década de 80 apontam que uma proteína-fosfatase do tipo
PP2A seria capaz de desfosforilar uma ACCase in vitro, porém ainda não se sabe a
relevância fisiológica nem se essa fosfatase seria a responsável pela desfosforilação
in vivo (GAUSSIN et al., 1996).

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
18
Figura 3.4: Síntese de ésteres de esterol (EEs) e t riacilglicerol (TAG). O acetil-CoA gerado a
partir de diferentes substratos é direcionado para a síntese de ácidos graxos pela enzima
acetil-CoA-carboxilase (ACCase) ou para a síntese d e esteróis pela 3-hidroxi-3-metil-glutaril-
CoA redutase (HMGR), ambas reguladas pela AMP quina se (AMPK). Os ácidos graxos são
convertidos a lipídios neutros, os TAG e EEs que sã o estocados nos corpúsculos lipídicos. Os
principais reguladores desta via são: AMPK (AMP qui nase), FAS (ácido graxo sintase), DGAT
(acil-CoA: diacilglicerol aciltransferease, LCAT (l ecitina:colesterol aciltransferase e ACAT (acil-
CoA;colesterol aciltransferase. Fonte:Bozaquel-Mora is (2010).

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
19
3.6.1 As proteínas fosfatases na regulação do metab olismo lipídico
A fosforilação reversível das proteínas é um mecanismo muito utilizado de
regulação bioquímica e fundamental para o controle de diversas funções celulares
(COHEN, 1997). As proteínas quinases e fosfatases apresentam alto grau de
conservação em diversos organismos, e formam redes de sinalização complexas
nas quais promovem fosforilação e desfosforilação respectivamente, de resíduos de
aminoácidos em proteínas tornando possível uma rápida resposta a estímulos
(JOHNSON e BARFORD, 1993).
Através da identificação de fosfatases por técnicas bioquímicas, as Ser/Thr
fosfatases até então chamadas proteínas fosfatases (PPs), puderam ser divididas
em dois tipos (1 e 2) (INGEBRITSEN e COHEN, 1983). Mais recentemente, através
das técnicas de biologia molecular, uma nova classificação foi proposta (COHEN,
1990) e as proteínas fosfatases foram agrupadas em duas famílias, PPP
(fosfoproteína fosfatase) e PPM (fosfoproteína fosfatase dependente de magnésio).
Além dessas duas famílias, existe um terceiro grupo chamado PTP (fosfoproteína
tirosina fosfatase), constituído por proteínas fosfatases capazes de desfosforilar
resíduos de tirosina.
Em relação a regulação por fosforilação das enzimas envolvidas no
metabolismo lipídico, existem poucos exemplos que ainda não foram totalmente
compreendidos. Sabe-se que na levedura S. cerevisiae, a ACCase é regulada por
fosforilação reversível (WITTERS e WATTS, 1990). A proteína-quinase Snf1p/AMPK
é a principal proteína quinase envolvida na fosforilação e inativação da ACCase in
vivo (WOODS et al., 1994). Porém, apesar de ainda não ser conhecida a proteína-
fosfatase responsável pela fosforilação e consequentemente, ativação da ACCase,
já foi identificado em experimentos de larga escala que existe uma interação física
entre a Ser/Thr proteína fosfatase Sit4p e Acc1p (HO et al., 2002).
Bozaquel-Morais et al. (2010) em estudo envolvendo as fosfatases como
possíveis reguladores do metabolismo de lipídios, demonstrou que a proteína
fosfatase Sit4, associada à sua sub-unidade regulatória Sap190p, está envolvida na
regulação da Snf1p. Neste trabalho, foi observado que a cepa mutante, deletada

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
20
para o gene sit4, apresentava menor acúmulo de CL em comparação a cepas
selvagens. Análises posteriores revelaram que as mutantes deletadas para os genes
sit4 e sap190 apresentavam hiperfosforilação da Snf1/AMPK, controlando, assim,
indiretamente a atividade da ACC (Figura 3.5).
Figura 3.5: A fosfatase Sit4 associada à sua subuni dade Sap190p regula a atividade da Snf1p,
controlando sua fosforilação, e consequentemente mo dulando a atividade da ACC. Além de
Sit4p-Sap190p, Snf1p é desfosforilada por Glc7p-Reg 1. Fonte: Bozaquel-Morais (2010)
3.7 Manipulação genética de leveduras
O maior conhecimento sobre as rotas biossintéticas em leveduras (HENRY et
al., 2012) combinado com a expansão de ferramentas para manipulações
bioquímicas e genéticas, tem criado um excelente momento na pesquisa sobre
acúmulo de lipídios em leveduras (KOHLWEIN, 2010; KOHLWEIN et al., 2013).

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
21
Apesar disso, poucos modelos foram desenvolvidos para o estudo do
metabolismo lipídico em leveduras. Dentre eles, podemos incluir a levedura
Saccharomyces cerevisiae, que tem sido amplamente utilizada como modelo
genético em estudos que tem melhorado significativamente o entendimento sobre o
metabolismo lipídico (CZABANY et al., 2007). As enzimas envolvidas na biossíntese
de TAG, armazenamento e degradação são muito semelhantes entre as espécies,
particularmente, entre as leveduras (BEOPOULOS et al., 2009).
S. cerevisiae oferece uma série de vantagens sobre as outras espécies de
leveduras oleaginosas como um micro-organismo hospedeiro para a produção de
ácidos graxos e seus derivados (SITEPU, 2014). Ou seja, a levedura S. cerevisiae é
mais facilmente manipulável geneticamente, quando comparada as leveduras
oleaginosas, já que as ferramentas para manipulação genética são mais abundantes
para essa espécie.
Além de S. cerevisiae, a levedura oleaginosa Yarrowia lipolytica tem recebido
destaque nos últimos trabalhos publicados envolvendo abordagens genéticas. O
genoma dessas duas leveduras foi completamente sequenciado, facilitando o
desenvolvimento de ferramentas genéticas e melhoramentos metabólicos
(BEOPOULOS et al., 2009; BEOPOULOS et al., 2010). De acordo com Sitepu et al.
(2014), a manipulação genética em leveduras oleaginosas tem resultado, por
exemplo, no desenvolvimento de cepas com maior ou menor capacidade de formar
os corpúsculos lipídicos.
De uma maneira geral, as abordagens da engenharia genética foram
desenvolvidas com base no potencial das leveduras serem utilizadas como
“fábricas” celulares para a produção de grandes quantidades de óleo, possibilitando
uma composição específica de lipídios e metabólitos de maior valor agregado.
(BEOPOULOS et al., 2009).

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
22
3.8 Escalonamento para o desenvolvimento de bioproc essos
O escalonamento do processo é geralmente a etapa final de qualquer
pesquisa com objetivo de produção industrial em larga escala de produtos por
fermentação (EINSELE, 1978).
Enquanto as abordagens de engenharia metabólica e de sistemas biológicos
são promissoras no desenvolvimento de processos otimizados em escala
laboratorial, eles precisam ser transferidos para escalas maiores sem reduzir a
sensibilidade das características de desempenho. De uma maneira mais direta, as
células que trabalham bem em escala de 1 litro são simplesmente requeridas a
desempenhar igualmente bem entre 5.000 e 10.000 litros (para químicos finos) ou
mais que 100.000 L (para commodities) (TAKORS, 2012).
É importante compreender que os processos de escalonamento de um
sistema de fermentação são frequentemente governados por um número importante
de parâmetros de engenharia, não sendo simplesmente uma questão de aumento
de volume do cultivo ou do vaso fermentador. Observa-se frequentemente que o
rendimento da biomassa e qualquer produto associado ao crescimento são
geralmente diminuídos no escalonamento de processos aeróbios (ENFORS et al.,
2001).
De acordo com Torrado e Matallana (2014), os processos industriais são
tecnologicamente otimizados para o maior rendimento de biomassa, mas pouco
caracterizados, do ponto de vista da adaptação molecular da levedura a condições
adversas de crescimento. No entanto, esse aspecto é crítico para um bom
desempenho do produto final.
As três principais escalas para o desenvolvimento de bioprocessos são: a)
laboratório, b) planta piloto e c) produção (JUNKER, 2002), podendo havere uma
escala demonstrativa. A taxa de escalonamento geralmente aplicada é de cerca de
1:10 para processos biotecnológicos até 100.000 L (VOTRUBA e SOBOTKA, 1992).
Porém, razões menores que 1:5 muitas vezes têm sido utilizadas para aumentar o
nível de segurança, isto é, diminuir o risco de um desempenho inesperado no
escalonamento. A escala de produção para vários produtos provenientes de micro-

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
23
organismos geneticamente modificados geralmente é de cerca de 10.000 L, o que é
mais uma escala piloto para outros tipos de produtos (NAVEH, 1985). A metodologia
exata que deve ser utilizada para o escalonamento depende em grande parte do tipo
e condições de processo, e se existem dados preliminares que mostram que o
procedimento escolhido é aplicável (BANKS, 1979).
Na maioria dos estudos de escalonamento envolvendo micro-organismos, a
otimização passo-a-passo é realizada. Seja a partir de frascos com agitação em
incubadores rotatórios ou em placas de poços múltiplos (possibilidade de avaliar em
paralelo um grande número de condições) para biorreatores. O rendimento do
produto depende de vários fatores que são altamente interligados. As condições do
ambiente rapidamente mudam em sistema de batelada agitado devido ao aumento
exponencial da biomassa, o que provoca muitas variações entre as culturas, um
problema que é extensivamente discutido (SIURKUS et al., 2010).
As exigências de desempenho em larga escala são desafiadoras e de
extrema importância, pois decisões econômicas sobre os investimentos para novos
bioprocessos são normalmente baseadas nos resultados de escala laboratorial. A
diminuição nas quantidades de produto, qualidade e rendimento em maiores
escalas, afeta diretamente os benefícios econômicos esperados.
Consequentemente, a atratividade de novos bioprocessos pode ter sua comparação
prejudicada com alternativas competidoras, ou até mesmo levar a sua interrupção
definitiva (TAKORS, 2012).
Por esses motivos, a elucidação de mecanismos de escalonamento para
prevenção de perdas não é somente um objetivo acadêmico, mas também uma
necessidade econômica (NOORMAN, 2011). Sem dúvidas, a rotina industrial
fornece vários exemplos de sucesso. No entanto, existem outros processos que são
sensíveis a condições de escalonamento e que mesmo fornecendo excelentes
rendimentos técnicos ou econômicos, falham em maior escala simplesmente porque
as expectativas de desempenho não podem ser cumpridas.
Sem dúvida, estudos sobre os sistemas biológicos são necessários para
entender a complexidade das respostas celulares em reatores em condições de
larga escala (LIDEN, 2002). Somente com esse entendimento holístico das
respostas celulares em múltiplos níveis será possível definir critérios que não sejam

Capítulo 3 – Revisão Bibliográfica Valoni, E.A.
24
determinados por parâmetros físicos (como atualmente) mas levando em conta as
propriedades biológicas intrínsecas. Utilizando esses critérios para os projetos
futuros, pode-se antecipar melhorias para uma bem sucedida transferência
“laboratório-para-produção” sem surpresas desagradáveis em larga escala
(TAKORS, 2012).

Capítulo 4 – Materiais e Métodos Valoni, E.A.
25
4. Materiais e Métodos
Neste capítulo são apresentados os principais materiais utilizados no estudo,
assim como as respectivas metodologias aplicadas.
4.1 Materiais
Os componentes dos meios de cultivo utilizados neste trabalho encontram-se
listados na Tabela 4.1.
Tabela 4.1: Reagentes utilizados no preparo dos mei os de cultivo e os respectivos fabricantes.
Reagente Fabricante
Extrato de Levedura Fluka
(NH4)2SO4 Vetec
KH2PO4 Vetec
Peptona Vetec
Agar-agar Vetec
Geneticina
Glicose
Glicerol P.A.
Sigma-Aldrich
Merck
Vetec
Os ensaios analíticos foram realizados com o uso dos reagentes
apresentados na Tabela 4.2.

Capítulo 4 – Materiais e Métodos Valoni, E.A.
26
Tabela 4.2: Reagentes utilizados nos ensaios anal íticos e os respectivos fabricantes
Reagente Fabricante
Kit dosagem enzimática de glicerol
Clorofórmio
Metanol
Iodeto de Potássio
Doles Brasil
Vetec
Vetec
Sigma-Aldrich
BODIPY® 493/503 Invitrogen
Formaldeído
Triton-X 100
Tris-HCl
Follin
CuSo4
Glicerol P.A.
BSA
Sigma-Aldrich
Sigma-Aldrich
Sigma-Aldrich
Sigma-Aldrich
Vetec
Vetec
Sigma-Aldrich
A glicerina loira utilizada nos experimentos descritos nesse estudo foi cedida
pela Planta Piloto do CENPES, obtida do processo de produção de biodiesel
utilizando óleo de soja e metanol, através da transesterificação alcalina com KOH.
A pureza em glicerol, caracterizado por cromatografia líquida de alta eficiência
(HPLC) no laboratório BIOSE, foi de 82% (p/v).

Capítulo 4 – Materiais e Métodos Valoni, E.A.
27
4.2 Equipamentos
Os principais equipamentos utilizados no trabalho foram:
1) Autoclave vertical Prismatec, modelo: CS;
2) Balança Analítica Adventurer;
3) Biorreator Tecnal, modelo: TecBio 1,5
4) Capela de fluxo laminar equipada com luz UV BioFlux, modelo: II A1 90 AC;
5) Centrífugas Eppendorf, modelos: Centrifuge 5804R e 5415R;
6) Centrífuga Thermo, modelo: Megafuge 16R;
7) Centrífuga CELM, modelo: LS-3 Plus;
8) Espectrofotômetro Shimadzu, modelo: UV - 1800
9) Espectrofotômetro Shimadzu, modelo: UV Mini 1240;
10) Estufa Bacteriológica Fisher Scientific, modelo: Isotemp 655D;
11) Incubador com agitação Tecnal, modelo:TE-420;
12) Microscópio ótico Nikon, modelo: Eclipse E200 acoplado a câmera Evolution VF;
13) Ultrafreezer CodLab, modelo: CL120-800;
14) Vórtex IKA, modelo: MS2 Mini agitador orbital;
15) Leitora de Placas Molecular Devices, modelo: SpectraMax 5;
16) Agitador rotatório Barnstead, modelo: Labquake;
17) Capela para exaustão de gases SP Labor;
18) Agitador de placas Boekel Scientific, modelo: The Jitterbug;

Capítulo 4 – Materiais e Métodos Valoni, E.A.
28
4.3 Meios de cultura
Os meios de cultura utilizados nos experimentos estão listados a seguir:
• YPD ( “Yeast extract, Peptone, Dextrose”) – composição (em p/v):
1% extrato de levedura
2% peptona
2% glicose
• YPD ágar – composição (em p/v):
1% extrato de levedura
2% peptona
2% glicose
2% ágar-ágar
• Meio mínimo de cultivo para acúmulo de óleo (MM – glicerol/glicerina) -
composição (em p/v) (FIEDUREK et al., 2011):
1% extrato de levedura
0,3% (NH4)2SO4
0,1% KH2PO4
0,5%, 2%, 5% ou 15% glicerol P.A/ glicerina loira
4.4 Micro-organismos
As cepas da levedura Saccharomyces cerevisiae utilizadas no presente
estudo foram a BY4741 (MATa his3∆1 leu2∆0 met15∆0 ura3∆0) (Figura 4.1) e cepas
derivadas de BY4741 com deleções simples para genes do metabolismo lipídico (31
mutantes) e genes de todas as proteínas-fosfatases e suas subunidades quando
não essenciais (65 mutantes), de acordo com a anotação do Saccharomyces
cerevisiae genome database (SGD, disponível em www.yeastgenome.org).
As cepas mutantes foram obtidas da biblioteca comercial “Gene Deletion
Library” da Open Biosystems, na qual os genes foram deletados por recombinação

Capítulo 4 – Materiais e Métodos Valoni, E.A.
29
homóloga, onde o gene deletado é substituído por um gene marcador KAN,
conferindo resistência ao antibiótico geneticina.
Figura 4.1: Imagem de microscopia ótica da levedura Saccharomyces cerevisiae em aumento
de 1000x utilizando microscópio ótico Nikon, modelo : Eclipse E200.
4.4.1 Preservação
As leveduras foram mantidas em placas de meio YPD sólido com colônias
isoladas e estocadas em temperatura de geladeira. No caso das leveduras
mutantes, elas foram mantidas em meio YPD com geneticina (200 µg/mL) e também
estocadas em geladeira.
Além disso, 2 cepas da levedura S. cerevisiae (BY4741 e rrd2) foram
mantidas em 20% de glicerol e estocadas em nitrogênio líquido, em temperatura de
-196ºC.
4.4.2 Obtenção do pré - inoculo
As células, mantidas em meio sólido (conforme descrito no item 4.4.1), foram
inoculadas em meio YPD líquido, até a fase estacionária (48 h) em agitador orbital
numa temperatura média de 30ºC e agitação de 200 rpm. O crescimento das
culturas foi estimado pela leitura da absorção da amostra em espectrofotômetro no

Capítulo 4 – Materiais e Métodos Valoni, E.A.
30
comprimento de onda de 600 nm (absorvância 600 nm). A turbidez do meio foi
expressa em D.O. (densidade ótica/ 1 mL da amostra em cubeta com 1 cm de
caminho ótico).
4.4.3 Curvas de crescimento e cálculo do volume de inóculo
As leveduras foram inoculadas a baixa densidade (0,25 DO/ml). Para calcular
o volume de pré-inoculo a ser utilizado, uma alíquota deste era amostrada
assepticamente para obtenção da medida de absorvância (AbsInóculo) a 600nm.
Utilizando a Equação 1, obtinha-se o volume de inóculo necessário para obtenção
da absorvância inicial nos cultivos.
Abs Inóculo x VInóculo = Abs Cultivo x VCultivo (1)
Onde:
Abs Inóculo = Medida de absorvância do inóculo;
V Inóculo = Volume de inóculo para se obter absorbância inicial nos cultivos;
Abs Cultivo = Valor de absorbância inicial desejada para os cultivos;
V Cultivo = Volume dos cultivos a serem inoculados;

Capítulo 4 – Materiais e Métodos Valoni, E.A.
31
4.5 Experimentos Preliminares
4.5.1 Avaliação do crescimento celular e acúmulo de lipídios em
concentrações crescentes de glicerol e glicerina lo ira.
Experimentos preliminares foram realizados com o objetivo de avaliar o
crescimento celular e acúmulo de lipídios de levedura Saccharomyces cerevisiae
utilizando um meio contendo como fonte de carbono glicerol. Dessa forma, foram
selecionados para o estudo o glicerol P.A. e a glicerina loira em concentrações que
variaram entre 2, 5 e 15% de glicerol/volume (Tabela 4.3). É importante destacar
que a concentração de glicerina loira foi corrigida, de acordo com sua pureza de
82%. Os cultivos foram inoculados utilizando um pré-inóculo da cepa S. cerevisiae
BY4741(WT) crescida previamente em YPD até a fase estacionária (48 h). A
concentração inicial de células foi de 0,050 g/L e os cultivos foram realizados em
duplicata em erlenmeyer de 250 mL, contendo 50 mL de meio de cultivo a 30ºC em
agitador orbital a 200 rpm durante 108 horas.
Foram realizadas amostragens nos tempos de 12 h, 22 h, 36 h e 108 h de
fermentação, nos quais foram realizadas leituras de densidade ótica (absorvância
600 nm) para estimação do crescimento celular.
Também foram retirados volumes das amostras contendo uma suspensão
celular em torno de 5 D.O. totais em microtubos, para análise de corpúsculos
lipídicos. Para essa análise, foram adicionados 1/9 do correspondente volume da
amostra coletada de formaldeído (concentração final de aproximadamente 3,7% v/v)
e após homogeneização, os microtubos foram incubados por 15 minutos a
temperatura ambiente. Os microtubos foram centrifugados a 3000 rpm por 5
minutos, o sobrenadante descartado e o precipitado ressuspenso em 1000 uL de
água destilada estéril. Repetiu-se mais uma vez a centrifugação à 3000 rpm por 5
minutos e o precipitado com as células fixadas foi mantido sob refrigeração em torno
de 4ºC em geladeira para posterior leitura do índice de corpúsculos lipídicos.

Capítulo 4 – Materiais e Métodos Valoni, E.A.
32
Tabela 4.3: Experimentos preliminares para avaliaçã o da fonte de carbono utilizada.
Fonte de Carbono Concentração
Glicerol P.A.
(A) 2%
(1)
5%
(2)
15%
(3) Glicerina Loira
(B)
4.5.2 Varredura das cepas de Saccharomyces cerevisiae mutantes para
avaliar o índice de corpúsculos lipídicos, quando c ultivadas em glicerina loira
5%
Para os experimentos de varredura das cepas de S. cerevisiae mutantes
foram utilizadas microplacas de 96 poços, nas quais foram distribuídas cepas que
apresentavam deleções simples para o gene do metabolismo lipídico (31 mutantes)
e deleções para os genes das proteínas-fosfatases e suas subunidades quando não
essenciais (66 mutantes). Além das cepas mutantes, mais 7 clones da cepa
selvagem BY4147 foram distribuídas, como um controle para a comparação,
somando um total de 104 cepas no experimento. A relação com todas as cepas
mutantes utilizadas nesse estudo encontra-se descrita nos Apêndices A, B e C.
Da placa com repique em YPD sólido, foi feito um pré-inóculo em microplaca
de 96 poços contendo 200 µL de meio YPD líquido, utilizando-se um replicador de
96 pinos. As células foram incubadas em estufa microbiológica a 30ºC, por 48 h.
Deste pré-inóculo, novamente com auxílio do replicador, as células foram
inoculadas em meio de cultivo para acúmulo de óleo com 5% de glicerina loira. As
células foram incubadas em incubador próprio para microplacas com agitação na
velocidade máxima estabelecida pelo equipamento e 30ºC por 48 horas.

Capítulo 4 – Materiais e Métodos Valoni, E.A.
33
Após o período de 48 horas, com auxílio de uma micropipeta multicanal, as
culturas foram fixadas para determinação do índice de corpúsculos lipídicos,
adicionando-se 23 µL de formaldeído (concentração final de aproximadamente 3,7%
v/v) e incubadas em temperatura ambiente por 15 minutos. As microplacas foram
centrifugadas a 3000 rpm por 5 minutos. O sobrenadante foi descartado e as células
lavadas com água destilada e centrifugadas novamente a 3000 rpm por 5 minutos. O
sobrenadante foi descartado e ao final do procedimento, as placas contendo o
precipitado celular foram guardadas em geladeira a temperatura de 8ºC até o
momento do ensaio.
Foram feitos três experimentos em replicata para avaliar o índice de CL no
meio de cultivo contendo 5% de glicerina loira.
4.5.3 Avaliação das cepas mutantes para gene fosfat ase em meio 5% de
glicerina loira
Para essa avaliação foram selecionadas as cepas mutantes para os genes
das proteínas-fosfatases que apresentaram os melhores resultados de crescimento
celular e índice de corpúsculos lipídicos nos experimentos anteriores.
Foram realizados experimentos em triplicata utilizando pré-inóculos das cepas
mutantes selecionadas em meio YPD líquido (para as cepas mutantes foi adicionado
200 µg/mL de geneticina ao meio). Os pré-inóculos foram adicionados numa
concentração inicial de células de 0,105 g/L em 50 mL de meio de cultivo em frascos
erlenmeyers de 250 mL e incubados a 30ºC em agitador orbital em velocidade 200
rpm durante 48 horas.
Foram retiradas amostras nos tempos de 20 h, 26 h e 45 h de fermentação
para leitura do crescimento celular (densidade ótica 600 nm) e também foram
coletadas amostras contendo 5 D.O. de células e fixadas em formaldeído 3,7% (v/v),
conforme descrito anteriormente, para a determinação do índice de corpúsculos
lipídicos.

Capítulo 4 – Materiais e Métodos Valoni, E.A.
34
4.6 Avaliação do acúmulo de corpúsculos lipídicos e m biorreator por
Saccharomyces cerevisiae mutante utilizando como fonte de carbono 5% de
glicerina loira
A partir dos experimentos prévios, foi selecionada uma cepa mutante deletada
para determinado gene regulatório não essencial de fosfatase que apresentou o
melhor resultado determinado para a relação crescimento celular e índice de CL e a
cepa selvagem utilizada como controle do experimento.
Os ensaios para avaliação de corpúsculos lipídicos foram conduzidos em
biorreator instrumentado em fermentação por 48h. Foi utilizado um biorreator
Tecnal® modelo TecBio 1,5 com vaso reacional de capacidade total de 1 L,
contendo 700 mL do meio. Todo o sistema, compreendendo o biorreator foi
previamente esterilizado em autoclave a 0,5 atm por 15 minutos.
Os experimentos foram conduzidos e controlados automaticamente a uma
temperatura de 30ºC e agitação de 200 rpm. A aeração de 1VVM foi feita através da
alimentação contínua de ar comprimido filtrado em filtro estéril com porosidade 0,45
µm.
Durante o período de fermentação, foram coletadas alíquotas nos tempos de
0 hora (logo após a inoculação do meio), 4 horas, 24 horas, 28 horas e 48 horas. As
alíquotas continham aproximadamente 5mL de amostra e foram realizadas com
auxílio de uma seringa, tomando-se o cuidado de desprezar o volume morto contido
na mangueira de amostragem.
As amostras foram coletadas nos tempos determinados, conforme citado
acima, para determinação das densidades óticas (D.O. 600nm), e a determinação do
glicerol presente no meio. Nos tempos 24 horas, 28 horas e 48 horas foram
coletadas amostras para a determinação dos corpúsculos lipídicos. No tempo de 48
horas foi coletada uma amostra para dosagem de triacilglicerol (TAG) nas células
das leveduras.

Capítulo 4 – Materiais e Métodos Valoni, E.A.
35
4.7 Métodos Analíticos
4.7.1 Determinação de Biomassa
Para conversão da densidade ótica em concentração da biomassa, efetuou-
se uma curva de calibração da densidade ótica (600 nm) versus peso seco (g/L)
utilizando-se o fator de conversão obtido pela curva de peso seco. Para isso,
usando-se uma suspensão de células em solução salina (água destilada com 0,9%
(p/v) NaCl), retira-se uma amostra (20 mL), que é filtrada em papel de filtro Millipore
(0,45 µm), seca em luz de infravermelho por 30 minutos e, seca novamente até peso
conatante. Da mesma suspensão são feitas diferentes diluições de modo a se obter
concentrações celulares distintas e, então, o valor de absorvância para cada
concentração é obtida em espectrofotômetro a 600 nm.
4.7.2 Ensaio de recuperação de fluorescência (ERF) para determinação do
Índice de Corpúsculos Lipídicos (ICL)
A determinação de corpúsculos lipídicos nas células de S. cerevisiae foi
realizada pelo método de ERF descrito por Bozaquel-Morais (2010), a partir de uma
suspensão celular em torno de 5 D.O. que foi fixada com formaldeído na
concentração final de 3,7% (v/v) durante 15 minutos a temperatura ambiente. As
células fixadas foram centrifugadas a 3000 rpm durante 5 minutos e o sobrenadante
foi descartado. As células foram lavadas 1x com água destilada e centrifugadas por
mais 5 minutos a 3.000 rpm. O precipitado celular formado foi guardado a 4ºC até a
realização do ensaio.
Foi preparado um meio de leitura, que consiste em uma solução de
BODIPY®493/503 5 µM e iodeto de potássio 500 mM. Alíquotas de 200 µL do meio
de leitura foram distribuídas nos poços de uma microplaca de 96 poços (pretas com
fundo transparente) e incubadas por 5 minutos a 30º C ao abrigo da luz para
estabilização do meio.

Capítulo 4 – Materiais e Métodos Valoni, E.A.
36
Em seguida, as amostras de células foram analisadas 4 vezes, mediante
sucessivas adições de alíquotas de 5 µL da suspensão celular a cada poço. Após
cada adição, é medida a fluorescência em excitação de 485 nm com emissão a 510
nm, cutoff de 495 nm, através de um leitor de placas. Uma leitura de absorvância é
também realizada para cada poço no comprimento de onda de 600 nm. A qualidade
dos dados obtidos foi avaliada através da análise da linearidade das medidas
comparadas a densidade ótica do poço. A partir dessa curva, obtém-se o índice de
corpúsculos lipídicos, denominado ICL, que trata-se da fluorescência relativa obtida
por unidade de densidade ótica de uma cultura de leveduras. Em caso de R2 menor
que 0,9 para a curva de uma amostra, a medida deve ser descartada e repetir uma
nova medição.
4.7.3 Determinação da concentração de glicerol pres ente no meio de cultivo
As amostras coletadas durante a fermentação nos tempos 0 hora, 4 horas, 24
horas, 28 horas e 48 horas, foram centrifugadas a 3.000 rpm durante 5 minutos para
separar o meio de cultivo das células, que formaram o precipitado celular. O meio de
cultivo sem as células foi guardado a -20ºC até o momento da análise.
As amostras coletadas foram analisadas por cromatografia líquida de alta
eficiência (HPLC).
4.7.4 Dosagem enzimática de triacilglicerol
Foram retiradas amostras contendo células crescidas no meio usado para
fermentação no biorreator após 48 h de cultivo. Foi coletado um volume equivalente
a 30 D.O. de células. As células recolhidas foram centrifugadas a 3.000 rpm por 5
minutos a temperatura ambiente. O sobrenadante foi descartado e o precipitado
guardado em ultrafreezer a -80ºC até o momento da análise.
As células foram ressuspensas em 300 µL de tampão Tris-HCl 50 mM, pH 7,5
e Triton-X 100 a 0,3% v/v. O conteúdo foi transferido para um tubo de ensaio de
vidro e adicionadas pérolas de vidro para promover o rompimento das células por
agitação, com o auxílio do vórtex por 5 ciclos de 30 segundos intercalados com

Capítulo 4 – Materiais e Métodos Valoni, E.A.
37
incubações por 30 segundos em banho de gelo. O homogeneizado foi coletado entre
as pérolas de vidro e transferido para outro tubo. Em seguida, foi adicionado mais
300 µL do mesmo tampão para lavar as pérolas e coletar novamente o
homogeneizado, centrifugando por mais 10 minutos a 3.000 rpm e transferindo o
sobrenadante coletado para um tubo de vidro de fundo cônico.
Em seguida, foi realizada uma extração de lipídeos neutros, de acordo com o
protocolo adaptado de Bligh & Dyer (BLIGH & DYER, 1959), no qual, foi adicionada
ao sobrenadante uma proporção de metanol e clorofórmio de 2:1 numa relação
máxima de água na mistura de solventes de 0,8 (água:metanol:clorofórmio). Essa
mistura foi agitada com auxílio de vórtex por dois minutos e centrifugada durante 15
minutos a 3.000 rpm, observando a formação de um precipitado e de uma única fase
sobrenadante. Todo o sobrenadante foi coletado e transferido para outro tubo de
vidro de fundo cônico, onde foi adicionado clorofórmio e água na proporção de
0,5:0,5. Após rápida agitação, os tubos foram centrifugados por 15 minutos a 3.000
rpm. A fase orgânica foi coletada com auxílio de uma pipeta Pasteur e transferida
para outro tubo de vidro de fundo cônico. A fase orgânica coletada foi seca sob fluxo
de nitrogênio e auxílio de um bloco de aquecimento. Os lipídios neutros foram
ressuspensos em 200 µL de tampão Tris-HCl 50 mM, pH 7,5 e Triton X 100.
Com os extratos lipídicos obtidos, os triacilgliceróis foram dosados como
previamente descrito por Bozaquel-Morais (2010), utilizando-se um kit clínico para
dosagem de TAG de acordo com as instruções do fabricante. O kit baseia-se em
reações enzimáticas, onde o triacilglicerol é primeiramente hidrolisado por uma
lipase gerando ácidos graxos e glicerol, e este último é dosado mediante a ação da
enzima glicerolquinase acoplada a reação da glicerol-fosfato oxidase. Nesta reação
ocorre a produção de peróxido de hidrogênio o qual é dosado mediante a oxidação
do substrato 4-aminoantipirina pela ação da enzima peroxidase, conforme as
reações descritas na figura 4.2. O substrato oxidado apresenta coloração rosada,
que foi medida no espectrofotômetro no comprimento de onda de 510 nm. Como
padrão utilizou-se uma solução de glicerol padrão na concentração de 0,1 mg/mL.

Capítulo 4 – Materiais e Métodos Valoni, E.A.
38
Figura 4.2: Reações enzimáticas envolvidas na quant ificação dos triglicerídeos. (Fonte: kit
enzimático para dosagem de triacilglicerídeos. Dole s, Brasil)
4.7.5 Determinação de Proteínas Totais
Para a dosagem de proteínas totais, foi utilizado o método de quantificação
colorimétrico de Folin – Lowry (LOWRY et al., 1951) no qual foi separada uma
alíquota de 50 µl da fase única obtida da centrifugação do homogeneizado com
mistura dos solventes metanol:clorofórmio na proporção 2:1. Foram adicionados
12% de ácido tri-cloroacético (TCA) para precipitação em banho de gelo por 10
minutos. Em seguida, as amostras foram centrifugadas em velocidade máxima por
10 minutos a 4ºC e todo sobrenadante, contendo o TCA, foi retirado com auxílio de
uma pipeta. O precipitado foi ressuspendido em 100 µL de solução alcalina.
Uma solução constituída por 98% de solução alcalina, 1% de CuSO4 e 1% de
tartarato de sódio e potássio, chamada de solução D, foi adicionada as amostras e
estas incubadas em temperatura ambiente por 10 minutos. Após esse procedimento,
foi adicionado com agitação imediata o reagente de Folin Ciocateau 1N e as
amostras protegidas da luz foram incubadas por 20 minutos em bancada. A leitura
foi medida no espectrofotômetro em comprimento de onda de 660 nm. Como padrão
utilizou-se uma solução de BSA.

Capítulo 5 – Resultados e Discussão Valoni, E.A.
39
5. Resultados e Discussão
Seguindo os objetivos da dissertação em questão, neste capítulo serão
apresentados os resultados da avaliação da cepa de levedura Saccharomyces
cerevisiae selvagem (WT) e da cepa mutante para genes que codificam proteínas
fosfatases e seus reguladores, crescidas em meio contendo o glicerol como única
fonte de carbono.
5.1 Avaliação de fontes de carbono alternativas no meio de cultivo para
acúmulo de lipídios
A maioria dos estudos de produção de lipídios a partir de micro-organismos
oleaginosos, como microalgas e leveduras, é realizado utilizando glicose como única
fonte de carbono (PAPANIKOLAOU et al., 2010). O alto custo da produção de
biodiesel a partir de micro-organismos oleaginosos tem como principal vilão o alto
custo da glicose, o qual é estimado ser 80% do custo total do meio (TSIGIE et al.,
2011). No entanto, diversos esforços têm sido realizados com a intenção de
minimizar o custo da fonte de carbono, pesquisando fontes alternativas de carbono
como o uso do glicerol (PAPANIKOLAOU et al., 2003) .
Nesse estudo, foi primeiramente avaliado o crescimento em glicerol
proveniente do glicerol puro (P.A.) e da glicerina loira, subproduto do processo de
obtenção do biodiesel. O meio utilizado foi um meio mínimo (FIEDUREK et al.,
2011), de baixo custo que poderia ser utilizado na indústria, contendo 2%, 5% e
15% (p/v) de glicerol puro (P.A.). Para a glicerina loira as mesmas concentrações
foram ajustadas de acordo com a sua pureza (82% p/v) para que todos os meios
apresentassem a mesma concentração em glicerol. A cepa de S. cerevisiae usada
para avaliação do crescimento celular foi a cepa selvagem BY4741 (WT).
Conforme podemos observar na Figura 5.1, a cepa BY4741 tem um
crescimento comparável em 2 e 5% de glicerol puro ou glicerina loira (Figura 5.1),
sendo que em 15% de glicerol puro a absorvância máxima foi duas vezes menor
que em 2 e 5% de glicerol. Quando comparado com concentrações de 2% e 5% de

Capítulo 5 – Resultados e Discussão Valoni, E.A.
40
glicerina loira, podemos observar também que o crescimento não foi diferente entre
eles.
Figura 5.1: Avaliação do glicerol puro (P.A.) (A) e da glicerina loira (B) como fonte de carbono.
Depois de avaliar o crescimento em glicerol, comparou-se este com o
crescimento em glicose, como pode ser observado na Figura 5.2, a diferença
principal entre estes dois é que a levedura utiliza um metabolismo respiratório
quando crescida em glicerol e um fermentativo quando crescida em glicose. Foi
possível observar que o crescimento da cepa WT em glicerol e glicerina loira ainda
apresenta uma fase inicial de adaptação do crescimento, também chamada de fase
lag, em 10 horas de crescimento, enquanto em meio MM com glicose em 10 h já
está na fase estacionária.

Capítulo 5 – Resultados e Discussão Valoni, E.A.
41
Em glicerol puro 2% e 5% e em glicerina loira 2% a fase exponencial de
crescimento teve início somente em 12 horas de cultivo (Figura 5.2 A e B). Esses
resultados estão de acordo com estudos anteriores que revelam que em S.
cerevisiae o consumo de glicerol, comparado ao consumo apresentado pela glicose,
é menos eficiente, com tempo de duplicação da massa celular em torno de 4 horas
(BEOPOULOS et al., 2009).
Figura 5.2: Curvas de crescimento da cepa selvagem (WT) BY4741 de S. cerevisiae nos meios MM – 2% e MM – 5% (p/v) de glicerol puro (A) e glicerina loira (B) e MM – 5% (p/v) de glicose .
A
B
C

Capítulo 5 – Resultados e Discussão Valoni, E.A.
42
Em estudo realizado por Swinnen et al. (2013), o mesmo efeito descrito foi
observado quando foram testadas diversas estirpes de S. cerevisiae em meio
sintético contendo glicerol, e a fase lag foi extremamente longa para todos os
isolados testados, até mesmo naqueles onde as cepas mostraram uma taxa
relativamente alta de crescimento após entrarem na fase exponencial de
crescimento.
O efeito que foi observado em condições de crescimento em glicerol era
esperado, pois S. cerevisiae é uma levedura que preferencialmente opta pela via
fermentativa quando na presença de glicose em concentrações a partir de 2% (p/v).
Já utilizando glicerol como única fonte de carbono, nas mesmas concentrações,
essa levedura tem como única opção realizar o metabolismo respiratório, tendo
como resultado um reflexo negativo nas taxas de crescimento celular.
Foram determinados os índices de CLs para a cepa WT crescida em meio
mínimo (MM) com glicerol puro e glicerina loira a 2% ou 5%, nos tempos de 36 horas
e 108 horas (tempo final do ensaio). Os índices de CL foram determinados pelo
ensaio de recuperação de fluorescência (ERF) (Figura 5.3 A e B). Comparando o
índice de CL entre os diferentes meios, é possível notar que os maiores índices de
CL foram alcançados quando 5% de glicerina loira foi utilizada como fonte de
carbono.
Os resultados também evidenciaram outra característica conhecida no
comportamento de acúmulo de lipídios pelas leveduras, na qual a síntese de CL tem
início na fase exponencial um pouco tardia, com seu índice máximo de acúmulo ao
longo da fase de desaceleração e início da fase estacionária de crescimento
(SANDAGER et al., 2002).

Capítulo 5 – Resultados e Discussão Valoni, E.A.
43
Figura 5.3: Acúmulo de lipídios utilizando meios co ntendo 2% e 5% (p/v) de glicerol puro (A) e
glicerina loira (B). Índice de corpúsculos lipídico s (CL) obtidos pelo ensaio de recuperação de
fluorescência na cepa selvagem BY47421 (WT) de S. cerevisiae com 36h e 108h de ensaio.
Analisando os resultados de crescimento e índice de CL, optou-se por utilizar
como fonte de carbono no meio, 5% de glicerol contido na glicerina loira. Esta
concentração foi escolhida já que foi a que mostrou um índice de CL maior e
alcançou um crescimento igual ao 2% de glicerol, mantendo uma boa relação entre
o crescimento celular e o índice de CL.
Para esta escolha também foi levado em consideração que a utilização da
glicerina loira na produção de lipídios microbianos representa uma estratégia
promissora pois permite uma valiosa reutilização de um dos maiores resíduos
produzidos pela indústria do biodiesel (RAIMONDI et al., 2014).

Capítulo 5 – Resultados e Discussão Valoni, E.A.
44
5.2 Varredura das cepas de Saccharomyces cerevisiae mutantes para
escolha da cepa com maior índice de acúmulo de lipí dios crescida em glicerina
loira 5%
Em estudo anterior realizado por Bozaquel-Morais (2010) foi avaliado o
envolvimento das proteínas fosfatases no metabolismo lipídico de S. cerevisiae.
Como estratégia foram medidos os índices de CL para as cepas deletadas para
todos os genes fosfatases e seus reguladores conhecidos presentes na levedura. A
varredura foi realizada crescendo as leveduras em meio rico (YPD) contendo 2% de
glicose como fonte de carbono até a fase exponencial. As cepas com maior ou
menor índice de CL que a cepa WT foram selecionadas e classificadas como llc e
hlc (baixo e alto conteúdo de CL, respectivamente). Nesta varredura foi possível
descobrir novos reguladores da síntese de CL.
A partir destas observações avaliou-se o comportamento e envolvimento das
proteínas fosfatases e seus reguladores na síntese de CL quando crescidas em
glicerol. Avaliou-se também o envolvimento no metabolismo respiratório, já que no
presente estudo a única fonte de carbono utilizada é o glicerol.
Foram testadas 104 cepas, divididas em dois grupos. O primeiro grupo, que
atuou como um grupo referência, é composto de 30 cepas deletadas para genes que
codificam enzimas associadas a corpúsculos lipídicos e/ou enzimas envolvidas no
metabolismo de esterol, triacilgliceróis e ceramida (Apêndice C) (Natter et al., 2005).
O segundo grupo, chamado de grupo das fosfatases, é o grupo alvo de interesse
para o estudo do envolvimento no metabolismo lipídico, e contém 67 mutantes
deletadas para as proteína-fosfatases e respectivas subunidades regulatórias
conhecidas (Apêndice B) (BOZAQUEL-MORAIS, 2010), de acordo com anotação na
base de dados online do genoma de Saccharomyces cerevisiae
(www.yeastgenome.org). Além das 97 cepas mutantes, foram incluídos 7 clones
independentes da respectiva cepa selvagem (WT) BY4741 de S. cerevisiae.
As cepas tiveram seus índices de corpúsculos lipídicos medidos através do
ensaio de recuperação de fluorescência no tempo final do ensaio, em 48 horas.
Também foram feitas medidas de absorvância no comprimento de onda 600 nm

Capítulo 5 – Resultados e Discussão Valoni, E.A.
45
para avaliar o crescimento celular. Os valores obtidos foram normalizados pela
média obtida em cada experimento e classificados do maior para o menor valor para
média de ICL obtido.
Os valores da média do índice de CL normalizados variaram entre 4,70 e
0,629. No entanto, levando-se em consideração o crescimento celular, podemos
observar que houve um comportamento muito semelhante para as cepas mutantes
dos dois grupos analisados. As cepas que apresentaram maiores índices de CL
foram as que apresentaram valores menores de crescimento celular. Assim como o
contrário também foi observado, as cepas mutantes que menos acumularam lipídios
foram as que tiveram melhor crescimento no meio contendo glicerol (Figura 5.4).

Capítulo 5 – Resultados e Discussão Valoni, E.A.
46
Figura 5.4: Distribuição das cepas mutantes em sínt ese de lipídio (A) e mutantes em proteínas
fosfatases (B), de acordo com a relação crescimento celular (medida por absorvância em 600
nm) e índice de CL no tempo de 48 horas.
A
B

Capítulo 5 – Resultados e Discussão Valoni, E.A.
47
A utilização da glicerina loira traz em sua composição diversos contaminantes
e impurezas (em particular álcoois, sabões e metais) que são conhecidos por afetar
negativamente ou até mesmo inibir o crescimento microbiano (SAMUL et al., 2014).
As cepas que, apesar de apresentarem um alto índice de CL, apresentaram
um crescimento muito baixo nos meios com glicerina loira, não foram selecionadas
para os experimentos seguintes (Tabela 5.1). Considerou-se importante para o
estudo ter cepas que além de apresentar índice de CL elevado também tivessem
crescimento satisfatório no meio proposto utilizando a glicerina loira.
Considerando o grupo das fosfatases como grupo alvo do estudo, foram
classificadas as cepas que acumularam mais lipídios e selecionadas as oito cepas
mutantes com os maiores valores da média de ICL (Tabela 5.2).
Tabela 5.1: Cepas de S. cerevisiae mutantes deletadas para genes de síntese de lipídio s e para subunidades catalíticas e regulatórias não ess enciais de fosfatases submetidos a screening pelo método de recuperação de fluorescência e dens idade ótica (DO 600 nm) que não apresentaram crescimento celular satisfatório.

Capítulo 5 – Resultados e Discussão Valoni, E.A.
48
Nome Processo Biológico Média
ICL D.P.
Media DO600 nm
D.P.
pph3 Fosfatase tipo 2A 1,826 0,750 0,559 0,413
cna1 Subunidade de calcineurina, fosfatase tipo 2B 1,600 0,874 0,750 0,579
oca1 Tirosina fosfatase 1,527 0,820 0,793 0,480
bni4 Subunidade regulatória da fosfatase tipo 1 Glc7p 1,428 0,238 0,527 0,175
pph22 Fosfatase tipo 2A 1,374 0,300 0,848 0,353
oca2 Tirosina fosfatase 1,364 0,181 0,650 0,309
yvh1 Tirosina fosfatase 1,335 0,919 1,041 0,747
rrd2 Ativador da atividade tirosina fosfatase de PP2A 1,311 0,154 0,780 0,232
nem1 Provável subunidade catalítica do complexo Nem1p-Spo7p
1,310 0,039 0,444 0,320
cnb1 Subunidade de calcineurina, fosfatase tipo 2B 1,305 0,439 0,712 0,409
ptp2 Tirosina fosfatase 1,203 0,306 0,922 0,452
ptc7 Fosfatase do tipo 2C 1,179 0,085 0,870 0,167
tip41 Interage fisicamente com Tap42p, regula Sit4p 1,161 0,242 0,892 0,123
pam1 Proteína de função desconhecida 1,087 0,134 1,149 0,414
msg5 Fosfatase dual 1,084 0,208 0,969 0,043
sap4 Subunidade regulatória da fosfatase Sit4p 1,074 0,067 0,827 0,169
ppq1 Ser/Thr fosfatase 1,074 0,270 0,943 0,078
sap155 Subunidade regulatória da fosfatase Sit4p 1,069 0,359 1,346 0,574
psr2 Fosfatase envolvida em resposta geral a estresse 1,048 0,054 1,089 0,368
sap185 Subunidade regulatória da fosfatase Sit4p 1,047 0,328 1,028 0,346
ppg1 Ser/Thr fosfatase 0,996 0,326 1,610 1,034
rts1 Subunidade regullatória da fosfatase tipo 2A 0,979 0,276 1,413 0,075
gip1 Subunidade regulatória da fosfatase tipo 1 Glc7p 0,917 0,033 1,154 0,251
ptc4 Fosfatase do tipo 2C 0,896 0,265 1,252 0,459
mih1 Tirosina fosfatase 0,896 0,266 1,105 0,654
sdp1 MAP quinase fosfatase 0,885 0,169 1,159 0,354
pps1 Fosfatase com atividade para resíduos ser, thr e tyr 0,882 0,062 1,116 0,395
YGR203W Similar a Cdc25p, Arr2p e Mih1p 0,877 0,322 1,662 0,773
Tabela 5.2: Cepas de S. cerevisiae mutantes deletadas para genes de síntese de lipídio s e para subunidades catalíticas e regulatórias não essencia is de fosfatases submetidos a screening pelo método de recuperação de fluorescência e densidade ótica (DO 600 nm). As cepas destacadas em verde foram selecionadas como hlc (alto conteúdo lipídico).

Capítulo 5 – Resultados e Discussão Valoni, E.A.
49
YNL010W Similar a serina fosfatase 0,869 0,164 1,845 0,196
reg2 Subunidade regulatória da fosfatase tipo 1 Glc7p 0,866 0,240 1,203 0,247
ptc6 Fosfatase do tipo 2C 0,864 0,186 1,081 0,416
ppz2 Fosfatase do tipo 1 0,853 0,200 1,388 0,431
rts3 Subunidade regulatória da fosfatase tipo 2A 0,842 0,066 1,713 0,409
sis2 Subunidade regulatória da fosfatase tipo 1 Ppz1p 0,835 0,308 1,293 0,567
pho13 Fosfatase alcalina com atividade proteína fosfatase
0,830 0,212 1,295 0,200
psy4 Subunidade regulatória da fosfatade tipo 2A Pph3p 0,823 0,258 1,554 0,764
gip2 Subunidade regulatória da fosfatase tipo 1 Glc7p 0,817 0,044 1,490 0,420
YBR051W ORF dúbio, parcialmente sobrepõe REG2 0,815 0,247 1,793 0,722
YER121W Proteína putativa de função desconhecida 0,809 0,248 1,784 0,010
ptc2 Fosfatase do tipo 2C 0,801 0,150 1,474 0,203
ptp1 Tirosina fosfatase 0,800 0,371 1,360 0,676
ptc3 Fosfatase do tipo 2C 0,798 0,211 1,746 0,222
ptc5 Fosfatase do tipo 2C 0,793 0,175 1,422 0,330
ltp1 Tirosina fosfatase 0,785 0,253 1,772 0,681
psr1 Fosfatase envolvida em resposta geral a estresse 0,779 0,236 1,610 0,327
sap190 Subunidade reguladora da fosfatase Sit4p 0,763 0,235 1,253 0,322
ppz1 Fosfatase tipo 1 0,742 0,250 1,892 0,821
reg1 Subunidade regulatória da fosfatase tipo 1 Glc7p 0,718 0,091 1,360 0,191
pph21 Fosfatase tipo 2A 0,696 0,206 1,494 0,379
vhs3 Subunidade regulatória da fosfatase tipo 1 Ppz1p 0,693 0,210 2,294 0,821
cmp2 Subunidade de calcineurina, fosfatase tipo 2B 0,688 0,223 1,732 0,191
siw14 Tirosina fosfatase 0,688 0,206 2,221 0,379
ptp3 Tirosina fosfatase 0,664 0,210 1,542 0,287
ppt1 Ser/Thr fosfatase similar a PP5 humana 0,662 0,187 1,468 0,473
sip5 Interage com Reg1p/Glc7p 0,642 0,141 2,165 0,250
pig2 Subunidade regulatória da fosfatase tipo 1 Glc7p 0,639 0,178 1,885 0,220
pig1 Subunidade regulatória da fosfatase tipo 1 Glc7p 0,634 0,140 1,989 0,530

Capítulo 5 – Resultados e Discussão Valoni, E.A.
50
Analisando o grupo de metabolismo lipídico (Tabela 5.3), alguns resultados
corroboram com resultados anteriormente descritos na literatura para as leveduras
realizando metabolismo fermentativo. A mutante erg4, deficiente numa enzima
relacionada a via de ergosterol, já tinha sido reportada como uma cepa com alto
conteúdo de corpúsculos lipídicos (FEI et al., 2008). Nesse estudo, apesar da cepa
mutante erg4 apresentar uma média de índice de CL alta (2,037 ± 1,02), seu
crescimento em meio contendo glicerol como fonte de carbono não foi expressivo
(0,56 ± 0,39). Outros resultados obtidos para o grupo das cepas mutantes deletadas
para genes de subunidades catalíticas e regulatórias não essenciais de fosfatases
não tiveram relação com o descrito na literatura. A cepa mutante para o gene sit4
(Apêndice A), que codifica uma Ser/Thr proteína-fosfatase e que tem papel essencial
na regulação do metabolismo de ácido graxo, apresentou altos valores de índice de
CL contrário ao já descrito. Porém, também já foi descrito que esta cepa apresenta
baixo crescimento celular em meios respiratórios (COOPER et al., 2000).

Capítulo 5 – Resultados e Discussão Valoni, E.A.
51
Nome Processo Biológico Média
ICL D.P.
Média DO600 nm
D.P.
erg3 Síntese de Esteróis 3,380 2,657 0,369 0,559
yeh2 Hidrolase (EE) 1,644 0,991 0,765 0,603
slc1 Síntese de Triacilgliceróis 1,607 0,362 0,391 0,328
csg2 Síntese de Ceramida 1,459 0,647 0,811 0,580
hmg1 Síntese de Esteróis 1,416 0,285 0,775 0,174
lac1 Síntese de Ceramida 1,349 0,366 0,734 0,342
sur2 Síntese de Ceramida 1,282 0,457 0,856 0,437
ipt1 Síntese de Ceramida 1,248 0,217 0,741 0,287
dga1 Síntese de Triacilgliceróis 1,190 0,709 1,038 0,656
sur1 Síntese de Ceramida 1,180 0,067 0,736 0,176
yeh1 Hidrolase (EE) 1,163 0,136 0,885 0,296
dpp1 Síntese de Triacilgliceróis 1,144 0,164 0,721 0,111
scs7 Síntese de Ceramida 1,103 0,430 1,263 0,805
gpt2 Síntese de Triacilgliceróis 1,082 0,117 0,740 0,085
hmg2 Síntese de Esteróis 1,077 0,175 0,898 0,248
lro1 Síntese de Triacilgliceróis 1,060 0,211 1,045 0,060
tgl3 Lipase (TAG) 1,043 0,213 1,328 0,070
tgl1 Hidrolase (EE) 1,023 0,131 0,879 0,326
are1 Síntese de Ésteres de Esterol 1,018 0,287 1,197 0,243
tgl5 Lipase (TAG) 0,989 0,168 1,060 0,181
lpp1 Síntese de Triacilgliceróis 0,955 0,347 1,248 0,445
ayr1 Síntese de Triacilgliceróis 0,945 0,161 1,095 0,289
sct1 Síntese de Triacilgliceróis 0,931 0,044 1,222 0,491
erg2 Síntese de Esteróis 0,794 0,233 1,798 0,114
erg24 Síntese de Esteróis 0,769 0,284 2,069 0,252
tgl4 Lipase (TAG) e Hidrolase (EE) 0,760 0,217 1,890 0,665
erg6 Síntese de Esteróis 0,743 0,285 1,851 0,477
lag1 Síntese de Ceramida 0,680 0,218 2,092 0,900
are2 Síntese de Ésteres de Esterol 0,674 0,181 1,724 0,246
erg5 Síntese de Esteróis 0,629 0,132 1,858 0,488
Tabela 5.3: Cepas de S. cerevisiae mutantes deletad as para genes de síntese de lipídios submetidos a screening pelo método de recuperação de fluorescê ncia e densidade ótica (DO600 nm).

Capítulo 5 – Resultados e Discussão Valoni, E.A.
52
Dessa forma, utilizando os critérios descritos acima, 8 cepas de S. cerevisiae
mutantes deletadas para genes de subunidades catalíticas e regulatórias não
essenciais de fosfatases foram selecionadas para os próximos experimentos (Tabela
5.4). Dessas cepas, somente a mutante deletada para o gene bni4, relacionado a
proteína fosfatase do tipo 1A, já tinha sido reportada no estudo anterior de Bozaquel-
Morais (2010) como uma cepa mutante deletada para o gene fosfatase que também
apresentava alto conteúdo lipídico.
Cepas Região Deletada Média
ICL
Desvio
Padrão
Media
DO600 nm
Desvio
Padrão
pph3 Fosfatase tipo 2A 1,826 0,750 0,559 0,413
cna1 Subunidade de calcineurina, fosfatase tipo 2B 1,600 0,874 0,750 0,579
oca1 Tirosina fosfatase 1,527 0,820 0,793 0,480
bni4 Subunidade regulatória da fosfatase tipo 1 Glc7p 1,428 0,238 0,527 0,175
pph22 Fosfatase tipo 2A 1,374 0,300 0,848 0,353
oca2 Tirosina fosfatase 1,364 0,181 0,650 0,309
yvh1 Tirosina fosfatase 1,335 0,919 1,041 0,747
rrd2 Ativador da atividade tirosina fosfatase de PP2A 1,311 0,154 0,780 0,232
5.3 Avaliação das cepas mutantes para os genes fosf atase em meio com 5%
de glicerol em glicerina loira
As cepas mutantes deletadas para os genes de subunidades catalíticas e
regulatórias não essenciais de fosfatases, selecionadas pela varredura do ensaio
anterior, foram avaliadas frente a um volume maior de fermentação, já que os
ensaios anteriores foram realizados em 0,2 ml. Foi realizado um ensaio utilizando-se
50 mL do meio mínimo (MM), com 5% de glicerina loira em frascos do tipo
Tabela 5.4: Cepas selecionadas de acordo com a médi a do índice de CL e a média de crescimento celular, medida por densidade ótica em 600 nm (DO60 0 nm).
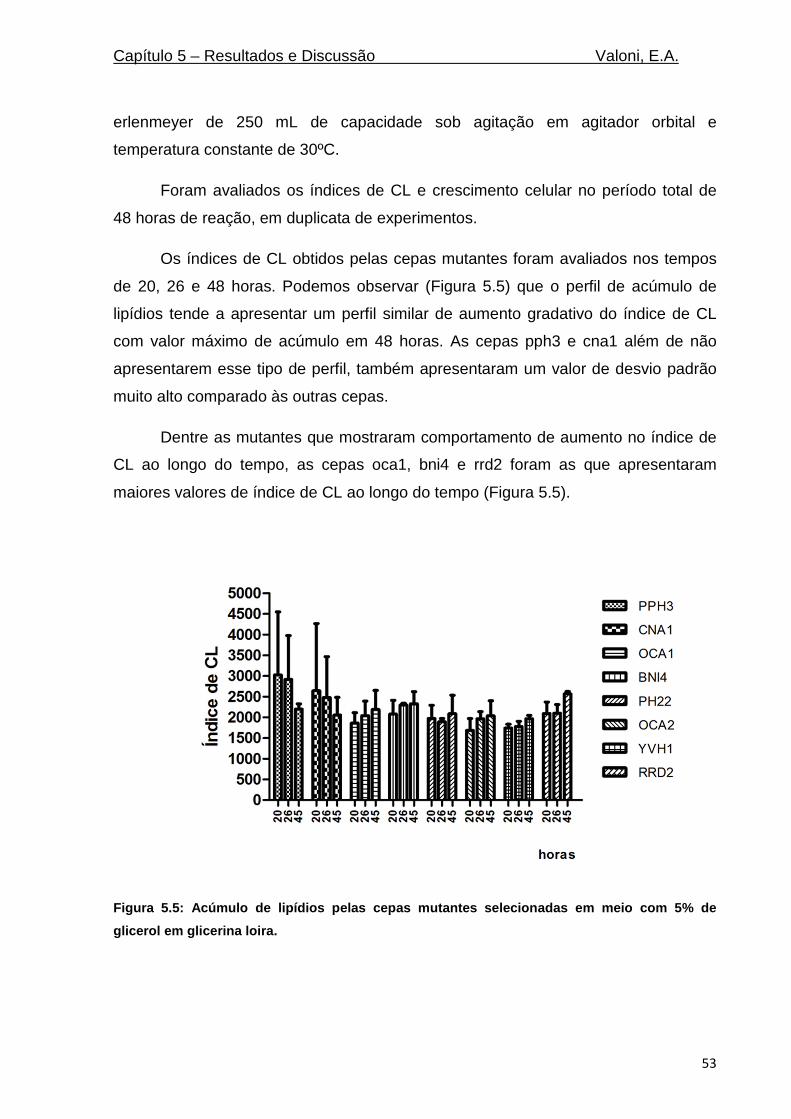
Capítulo 5 – Resultados e Discussão Valoni, E.A.
53
erlenmeyer de 250 mL de capacidade sob agitação em agitador orbital e
temperatura constante de 30ºC.
Foram avaliados os índices de CL e crescimento celular no período total de
48 horas de reação, em duplicata de experimentos.
Os índices de CL obtidos pelas cepas mutantes foram avaliados nos tempos
de 20, 26 e 48 horas. Podemos observar (Figura 5.5) que o perfil de acúmulo de
lipídios tende a apresentar um perfil similar de aumento gradativo do índice de CL
com valor máximo de acúmulo em 48 horas. As cepas pph3 e cna1 além de não
apresentarem esse tipo de perfil, também apresentaram um valor de desvio padrão
muito alto comparado às outras cepas.
Dentre as mutantes que mostraram comportamento de aumento no índice de
CL ao longo do tempo, as cepas oca1, bni4 e rrd2 foram as que apresentaram
maiores valores de índice de CL ao longo do tempo (Figura 5.5).
Figura 5.5: Acúmulo de lipídios pelas cepas mutante s selecionadas em meio com 5% de
glicerol em glicerina loira.

Capítulo 5 – Resultados e Discussão Valoni, E.A.
54
Ao avaliar o crescimento das cepas em glicerina loira (Figura 5.6), a mutante
bni4 não apresentou bom resultado de crescimento, assim como a mutante oca1.
Para esse estudo, optou-se por selecionar uma cepa mutante que apresentasse alto
valor de índice de acúmulo de CL e um bom crescimento (mesmo não sendo o maior
crescimento apresentado) no meio contendo glicerol. Sendo assim, a cepa rrd2 foi
selecionada para avaliar seu comportamento no experimento seguinte em biorreator.
Figura 5.6: Média do crescimento das cepas mutantes selecionadas. O crescimento das
mutantes de S. cerevisiae foi medido por absorvância no comprimento de onda de 600 nm ao
longo da fermentação (n=3).
O gene rrd2 codifica para um regulador de proteínas fosfatases do tipo
treonina-serina fosfatase PP2A. Ela é um ativador da proteína fosfatase sit4 que
regula o ciclo celular e também a síntese de lipídios (FELLNER et al., 2003).No
estudo anterior realizado por Bozaquel-Morais (2010), a cepa rrd2 não apresentou
alteração no metabolismo de acúmulo de lipídios quando cultivada em glicose. No
presente estudo, no entanto, foi possível observar que frente ao glicerol como fonte
de carbono, ou seja, quando a levedura utiliza a via respiratória, a deleção desse
gene apresentou uma alteração positiva para o acúmulo de lipídios.

Capítulo 5 – Resultados e Discussão Valoni, E.A.
55
5.4 Avaliação do acúmulo de corpúsculos lipídicos n a cepa mutante rrd2 de
Sacchromyces cerevisiae em biorreator utilizando glicerina loira como font e de
carbono
Para a etapa do estudo em biorreator, os resultados dos experimentos
realizados anteriormente foram cruciais para a seleção de uma cepa, dentre as 108
mutantes testadas inicialmente. Nessa etapa do estudo, a cepa poderá ser avaliada
em condições de volumes maiores, em um equipamento com controle mais preciso
de variáveis importantes para um bioprocesso, como por exemplo, temperatura,
agitação e aeração.
Os cultivos foram conduzidos em biorreator em fermentação por 48h em 4
replicatas. Além da cepa mutante rrd2, também foram realizados os mesmos
ensaios para a cepa selvagem (WT) como um controle dos estudos.
A concentração inicial de células inoculadas em 700 mL de meio foi de 0,25
D.O./mL, utilizando o fator obtido pela curva de peso seco, tem-se os valores de
0,101 g/L para a cepa WT e de 0,090 g/L para a cepa mutante rrd2.
Foram realizadas amostragens para determinação do índice CL e crescimento
celular nos tempos de 20, 24 e 48h. Também foram coletadas amostras do meio
reacional nos tempos 0, 8, 20, 24, 28 e 48 horas para verificação do consumo de
glicerol no meio. Além dessas amostras, ao final do ensaio, foi coletada uma
amostra para determinação de triacilglicerídios totais nas células crescidas por 48
horas. De acordo com os resultados obtidos no primeiro ensaio para a cepa mutante
rrd2 e para a cepa WT (Figura 5.7) pode-se observar que o índice de CL tem seu
valor máximo atingido em 24 horas de cultivo. Depois disso, observamos um
decréscimo nos tempos 28 e 48 horas.

Capítulo 5 – Resultados e Discussão Valoni, E.A.
56
Figura 5.7: Acúmulo de lipídios pelas cepa mutante rrd2 e selvagem WT (n=1)
Além disso, é possível observar para este experimento ( n=1 ) que no tempo
de 24 horas, onde ambas as cepas apresentam o índice máximo de CL, o valor
obtido para a cepa selvagem WT foi maior que o obtido para a cepa mutante rrd2.
Mas ao realizar outros três experimentos (Figura 5.8), observa-se que a média do
índice de CL obtida pela cepa mutante rrd2 é maior que a cepa selvagem, quando
determinada depois de 24 h de crescimento.
Figura 5.8: Acúmulo de lipídios pelas cepa mutante rrd2 e selvagem WT (n=4).
Alíquotas de células da cepa S. cerevisiae selvagem (WT) e mutante rrd2
foram coletadas no tempo final de fermentação no biorreator em 48 horas. Os
lipídios totais foram extraídos pelo método adaptado de Bligh & Dyer, conforme

Capítulo 5 – Resultados e Discussão Valoni, E.A.
57
descrito anteriormente. Os níveis de triacilgliceróis (TAG) foram dosados
enzimaticamente com kit clínico comercial (Doles, Brasil) (Figura 5.9), que determina
a quantidade de glicerol incorporado em TAG.
Figura 5.9: Acúmulo triacilgliceróis (TAG). O perce ntual de TAG nas cepas de S. cerevisiae
selvagem (WT) mutante RRD2, foram medidos no tempo final de reação em biorrea tor em 48
horas (n=4).
Em relação aos resultados podemos observar que a quantidade de TAG por
grama de proteína acumulada pelas cepas de S. cerevisiae WT e mutantes, foi
similar em ambas as cepas.
Os dados de crescimento celular foram obtidos ao longo do período de
fermentação em biorreator (Figura 5.10). Podemos observar que em 48 horas as
células ainda encontravam-se em fase exponencial de crescimento. Além disso, as
duas cepas tiveram crescimento similares.

Capítulo 5 – Resultados e Discussão Valoni, E.A.
58
Figura 5.10: Crescimento celular das cepa mutante RRD2 e cepa selvagem WT. O crescimento
celular foi medido por absrovância no comprimento d e onda de 600 nm (n=4).
O consumo de glicerol foi obtido através das medidas das concentrações de
glicerol nas amostras do meio reacional nos tempos inicial (0 hora), 4 horas, 24
horas e 48 horas. As medidas foram realizadas em HPLC (Figura 5.11) e pelo
método enzimático (Figura 5.12).
Figura 5.11: Consumo de glicerol no meio de cultivo . O glicerol foi medido por HPLC em
amostras do meio de cultivo contendo 5% de glicerin a loira.
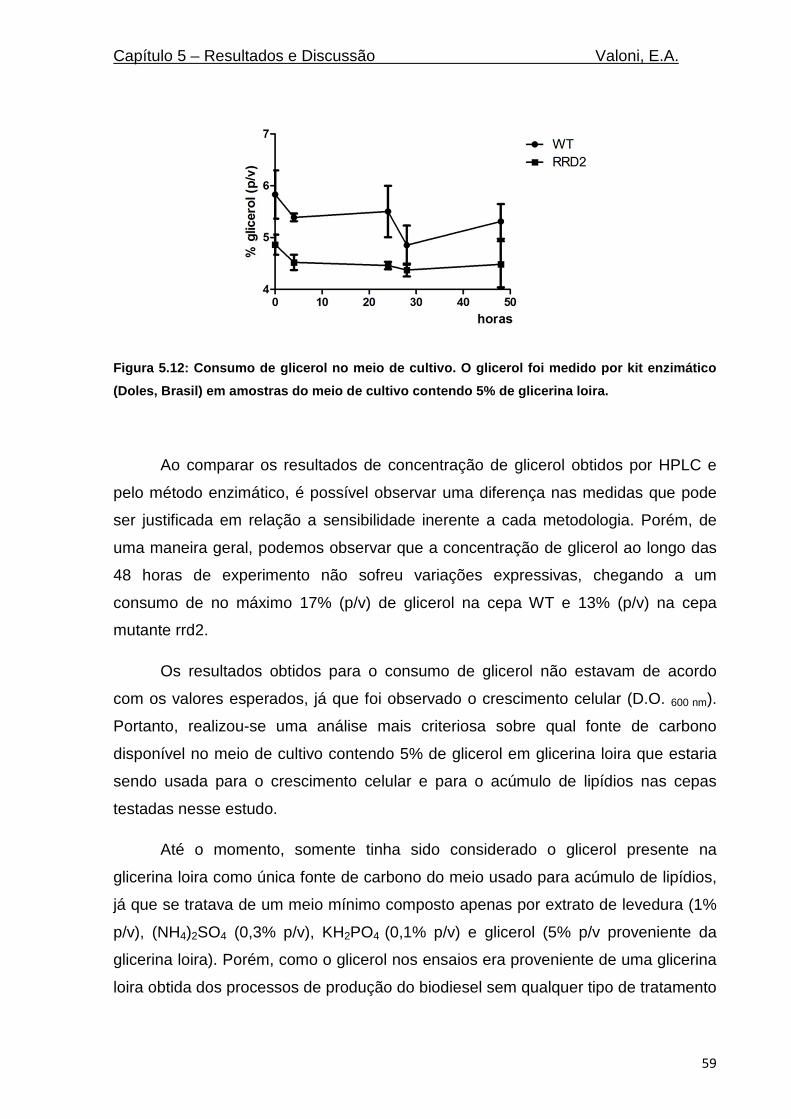
Capítulo 5 – Resultados e Discussão Valoni, E.A.
59
Figura 5.12: Consumo de glicerol no meio de cultivo . O glicerol foi medido por kit enzimático
(Doles, Brasil) em amostras do meio de cultivo cont endo 5% de glicerina loira.
Ao comparar os resultados de concentração de glicerol obtidos por HPLC e
pelo método enzimático, é possível observar uma diferença nas medidas que pode
ser justificada em relação a sensibilidade inerente a cada metodologia. Porém, de
uma maneira geral, podemos observar que a concentração de glicerol ao longo das
48 horas de experimento não sofreu variações expressivas, chegando a um
consumo de no máximo 17% (p/v) de glicerol na cepa WT e 13% (p/v) na cepa
mutante rrd2.
Os resultados obtidos para o consumo de glicerol não estavam de acordo
com os valores esperados, já que foi observado o crescimento celular (D.O. 600 nm).
Portanto, realizou-se uma análise mais criteriosa sobre qual fonte de carbono
disponível no meio de cultivo contendo 5% de glicerol em glicerina loira que estaria
sendo usada para o crescimento celular e para o acúmulo de lipídios nas cepas
testadas nesse estudo.
Até o momento, somente tinha sido considerado o glicerol presente na
glicerina loira como única fonte de carbono do meio usado para acúmulo de lipídios,
já que se tratava de um meio mínimo composto apenas por extrato de levedura (1%
p/v), (NH4)2SO4 (0,3% p/v), KH2PO4 (0,1% p/v) e glicerol (5% p/v proveniente da
glicerina loira). Porém, como o glicerol nos ensaios era proveniente de uma glicerina
loira obtida dos processos de produção do biodiesel sem qualquer tipo de tratamento

Capítulo 5 – Resultados e Discussão Valoni, E.A.
60
para remoção de impurezas ou contaminantes, analisou-se melhor os possíveis
componentes que estariam presentes em sua composição.
Uma caracterização química realizada no ano de 2013 para essa glicerina
loira, que foi cedida pelo CENPES (a íntegra das informações contidas nessa
caracterização não puderam ser divulgadas nesse estudo), constatou-se que do total
de sua composição, 3,49% (p/p) são compostos denominados matéria orgânica não
glicerinada (MONG). Através de uma caracterização qualitativa por cromatografia
líquida foram identificados entre os compostos do tipo MONG, a presença de ésteres
metílicos de ácidos graxos, ácidos graxos livres e monoacilgliceróis. Além disso,
uma análise de ressonância magnética nuclear (RMN) sugere que os ésteres
metílicos de ácidos graxos, os ácidos graxos e os monoacilgliceróis (identificados
por cromatografia líquida) possuem a distribuição típica dos constituintes do óleo de
soja. CLEMENTE et al. (2009) descrevem em seu estudo que o óleo de soja é
composto de 5 tipos de ácidos graxo: ácido palmítico (16:0), ácido esteárico (18:0),
ácido oleico (18:1), ácido linoleico (18:2) e ácido linolênico (18:3). Com percentuais
médios de 10%, 4%, 18%, 55% e 13%, respectivamente.
Sendo assim, reunindo as informações levantadas e os resultados obtidos
nos experimentos desse estudo, podemos presumir que as cepas da levedura S.
cerevisiae, tanto selvagem quanto mutante estejam utilizando como fonte de
carbono para o crescimento celular e acúmulo de lipídios os ácidos graxos presentes
na glicerina loira.
Partindo desse pressuposto, também foi possível fazer uma análise em
relação ao acúmulo de lipídios das cepas WT e mutante rrd2. Os resultados obtidos
nos 4 experimentos em replicatas não apresentaram uma diferença expressiva em
relação ao acúmulo de lipídios para a cepa selvagem e para a mutante rrd2, que foi
selecionada nos experimentos anteriores por apresentar um índice de CL maior
comparado as outra cepas testadas, inclusive a própria cepa selvagem.
O que poderia ser discutido, através dos resultados obtidos até o momento
nesse estudo, é que como o cultivo das cepas de levedura S. cerevisiae foi realizado
na presença de substâncias hidrofóbicas, estas substâncias foram utilizadas como
substratos e dessa forma, o acúmulo de lipídios observado se dá através da via ex
novo (BEOPOULOS et al., 2009). Como o consumo de glicerol no meio reacional

Capítulo 5 – Resultados e Discussão Valoni, E.A.
61
não é observado de forma expressiva nos experimentos realizados em 48 horas, e a
glicerina loira utilizada como fonte de carbono conter ácidos graxos livres, estes
compostos seriam incorporados diretamente nos CL através da via ex novo. O lipídio
microbiano produzido por essa via contém menores quantidades de TAGs
comparados com o crescimento realizado em substratos baseados em açúcares,
que utiliza a via de novo para o acúmulo de lipídios (PAPANIKOLAOU et al., 2011).
Uma grande variedade de micro-organismos é capaz de utilizar ácidos
graxos livres como única fonte de carbono e energia, independente das suas
capacidades lipolíticas (TACCARI et al., 2012).
No experimento em biorreator, ao final das 48 horas do ensaio, a
concentração de glicerol se manteve próxima das concentrações iniciais (Figura 5.11
e 5.12) e as cepas de levedura ainda se encontravam na fase exponencial de
crescimento (Figura 5.10). Com esse resultado, o estudo também pode evidenciar
que num meio contendo glicerol e lipídios neutros (ésteres metílicos de ácidos
graxos, ácidos graxos e monoacilgliceróis) a levedura S. cerevisiae
preferencialmente consumiu os ácidos graxos livres presentes. Caso o experimento
não fosse interrompido em 48 horas talvez poderia ser verificado nas horas
seguintes, o consumo do glicerol quando os ácidos graxos livres presentes na
glicerina loira tivessem sido esgotados do meio. Alguns estudos fazem referência a
fraca utilização de ácidos graxos pela levedura S cerevisiae (BEOPOULOS et al.,
2009) mas não foram encontrados estudos que fizessem referência ao consumo
preferencial de ácidos graxos frente ao glicerol como fonte de carbono.
Resumindo, foi observado nesse estudo que a levedura Saccharomyces
cerevisiae pode crescer em meio de cultivo com 5% de glicerol em glicerina loira e
acumular lipídios utilizando os ácidos graxos livres presentes na glicerina loira
através da via ex novo. Nessas condições, a deleção para o gene fosfatase rrd2 na
cepa mutante não teve influencia para o acúmulo de lipídios.
Ao comparar os resultados de índice de CL da cepa mutante rrd2 obtidos em
biorreator e nos ensaios realizados com volumes de 50 mL em frascos do tipo
erlenmeyer agitados em agitador orbital, podemos observar que o máximo para o
índice de CL que nos frascos eram obtidos em 48 horas de reação foram
antecipados para 24 horas em biorreator. Da mesma forma, observou-se que a

Capítulo 5 – Resultados e Discussão Valoni, E.A.
62
concentração média de células da cepa mutante rrd2 que era alcançada em frascos
erlenmeyer em 45 horas (4,34 D.O.) no biorreator foi alcançada entre 24 e 28 horas.
Esse é um comportamento comumente observado nas etapas de aumento de
volumes de reação, já que em biorreator é promovida uma melhor transferência de
massa, a aeração é mais eficiente e consequentemente temos os processos
acelerados. É esperado que o tempo total de batelada diminua quando se transfere
o processo para um biorreator.
Quando os resultados, tanto da cepa selvagem quanto da cepa mutante, para
cada escala de volume são comparados, observamos que os valores dos índices de
CL diminuem a medida que os volumes de reação aumentam. A cepa mutante, que
em escala de volume na ordem de µL apresenta os maiores resultados para o
acúmulo de lipídio, quando comparada a cepa selvagem sem mutação, é a que
apresenta as maiores quedas nos índices de CL quando levada para volumes de
reação próximos a 1 litro. Os processos industriais são tecnologicamente otimizados
para o maior rendimento de biomassa, porém pouco caracterizados do ponto de
vista da adaptação molecular da levedura para condições adversas de crescimento.
No entanto, esse aspecto é crítico, e deve ser estudado mais aprofundadamente,
para se obter uma boa performance do produto final (PÉREZ-TORRADO et al.,
2014).

Capítulo 6 – Conclusões Valoni, E.A.
63
6. Conclusões
Com base nos resultados obtidos neste trabalho pode-se concluir que:
� A levedura Saccharomyces cerevisiae foi capaz de crescer e acumular lipídios
quando crescida em glicerol puro (P.A.) e glicerina loira, nas concentrações de
2% ou 5%.
� A glicerina loira a 5% (p/v) foi a concentração de glicerol no meio de cultivo
mínimo que forneceu a melhor relação entre índice de CL e crescimento celular
para a levedura S. cerevisiae WT
� As cepas deletadas para o genes que codificam as proteínas fosfatases ou seus
reguladores pph3, cna1, oca1, bni4, pph22, oca2, yvh1 e rrd2 apresentaram a
melhor relação entre o índice de CL e crescimento celular e foram consideradas
para os ensaios seguintes .
� No ensaio realizado com as cepas mutantes de proteínas fosfatases e seus
reguladores, em volumes de 50 mL em frascos agitados em agitador orbital, o
melhor tempo para acúmulo de CL foi alcançado em 48 horas de reação e a
cepa mutante que apresentou a melhor relação entre maior índice de CL e
crescimento celular foi a rrd2.
� No experimento em biorreator foi possível observar que em 48 horas o consumo
de glicerol foi muito baixo, chegando ao valor máximo de 17% (p/v) para a cepa
WT e de 13% (p/v) para a cepa mutante rrd2.
� Com a caracterização da glicerina loira, foi possível concluir que o glicerol não
era a única fonte de carbono presente no meio, e que a matéria orgânica não
glicerinada (MONG) era composta por ácidos graxos livres que possivelmente
foram preferencialmente utilizados pelas cepas WT e mutante de S. cerevisiae
para o crescimento celular e acúmulo de lipídios.

Capítulo 6 – Conclusões Valoni, E.A.
64
� A via de síntese de lipídios utilizada pelas cepas de S. cerevisiae, tanto
selvagem quanto mutante, num meio contendo ácidos graxos livres
possivelmente utilizada num primeiro momento (nas primeiras 48h) foi a via ex
novo.
� Quando o experimento foi transferido para uma escala maior de volume em
biorreator (700 mL), os processos de acúmulo de lipídios e crescimento celular
foram antecipados para tempos menores de reação (índice de CL máximo
passou de 48 horas para 24 horas).
� Em biorreator a deleção do gene rrd2 não apresentou diferenças para um maior
acúmulo de lipídios, conforme o esperado pelos experimentos realizados
previamente em menores escalas de volume de reação.
Resumindo-se, foi possível concluir que a levedura Saccharomyces cerevisiae é,
muito possivelmente, capaz de utilizar os ácidos graxos livres, presentes na glicerina
loira, preferencialmente ao glicerol, acumulando lipídios nos corpúsculos lipídicos
intracelulares. Com base na utilização dos ácidos graxos livres, a via utilizada para a
síntese e acúmulo dos lipídios possivelmente foi a via ex novo, que é menos citada
na literatura. O envolvimento das proteínas fosfatases, avaliada nesse estudo pela
deleção do gene rrd2, nessa via de síntese não é muito forte, já que os resultados
de índice de CL não foram muito diferentes entre a cepa mutante e a cepa
selvagem.

Capítulo 7– Perspectivas e Trabalhos Futuros Valoni, E.A.
65
7. Perspectivas e trabalhos futuros
• Realizar um ensaio em biorreator com a cepa de Saccharomyces cerevisiae
selvagem e mutante rrd2 no mesmo meio de cultivo com 5% de glicerina loira
porém, em tempo superior a 48 horas e avaliando o consumo de ácido oleico
(C18:0) e de glicerol.
• Realizar um ensaio em biorreator com meio de cultivo contendo 5% de
glicerol puro (P.A.) com a mutante deletada para o gene das fosfatases, com
objetivo de avaliar o envolvimento da deleção no gene rrd2 acúmulo de
lipídios pela via de novo em metabolismo respiratório.
• Verificar se o ácido oleico na presença de glicerol é tóxico para as células, o
que poderia justificar o baixo crescimento celular quando os maiores índices
de CL são obtidos.
• Verificar por cromatografia em camada fina, qual o principal lipídio acumulado
nos corpúsculos lipídicos em meio contendo glicerina loira.
• Sabendo-se que os processos em biorreatores tem seus tempos de acúmulo
de CL e crescimento celular antecipados, iniciar o processo com uma
concentração de células maior do que a que foi empregada nesse estudo.

Capítulo 8 – Referências Bibliográficas Valoni, E.A.
66
8. Referências Bibliográficas
ABGHARI, A.; CHEN, S. Yarrowia lipolytica as an oleaginous cell factory platform for production of fatty acid-based biofuel and bioproducts. Front Energy Res , v. 2, p. 21, 2014.
ACHILLES, J.; HARMS, H.; MULLER, S. Analysis of living S. cerevisiae cell states – a three color approach. Cytometry , v. 69, p. 173 – 177, 2006.
AGEITOS, J.M.; VALLEJO, J.A.; VEIGA-CRESPO, P.; VILLA, T.G. Oily yeasts as oleaginous cell factories. Appl. Microbiol. Biotechnol , v. 90, p. 1219- 1227, 2011.
AGGELIS, G.; KOMAITIS, M.; PAPANIKOLAOU, S.; PAPADOPOULOS, G. A mathematical model for the study of lipid accumulation in oleaginous microorganisms I. Lipid accumulation during growth of Mucor circinelloides CBS 172 – 27 on a vegetable oil. Gracas y Aceites. , v. 46, p. 169 – 173, 1995.
AGGELIS, G.; PAPADIOTIS, G.; KOMAITIS, M. Micribial fatty acid specificity. Folia Microbiol. , v. 42, p. 117 – 120, 1997.
ANDRÉ, A.; CHATZIFRAGKOU, A.; DIAMANTOPOULOU, P.; SARRIS, D.; PHILIPPOUSSIS, A.; GALIOTOU-PANAYOTOU, M. Biotechnological conversions of biodiesel derived crude glycerol by Yarrowia lipolytica strains. Engineering in Life Sciences , v. 9, n. 6, p. 468 – 478, 2009.
ANGERBAUER, C.; SIEBENHOFER, M.; MITTELBACH, M.; GUEBITZ, G. M. Conversion of sewage sludge into lipids by Lipomyces starkeyi for biodiesel production. Bioresour. Technol. , v. 99, p. 3051 – 3056, 2008.
ATABANI, A.; SILITONGA, A., BADRUDDIN, I.; MAHLIA, T.; MASJUKI, H.; MEKHILEF, S. A comprehensive review on biodiesel as na alternative energy resource and its characteristics. Renew Sustain Energy Rev , v. 16, p. 2070-2093, 2012.
ATHENSTAED, K.; DAUM, G. Phosphatidic acid, a key intermediate in lipid metabolism. Eur J Biochem , v. 266, n. 1, p. 1 – 16, 1999.
ATHENSTAED, K.; DAUM, G. Tgl4p and Tgl5p, two triacylglycerol lipases of the yeast Saccharomyces cerevisiae are localized to lipid particles. Journal of Biological Chemistry , v. 280, p. 37301 – 37309, 2005.

Capítulo 8 – Referências Bibliográficas Valoni, E.A.
67
BANKS, G. T. Scale-up of fermentation processes. Topics in enzyme and fermentation biotechnology, New York, p. 170 – 266, 1979.
BELLOU, S.; MOUSTOGIANNI, A.; MAKRI, A.; AGGELIS, G. Lipids containing polyunsaturated fatty acids synthesized by Zigomycetes grown on glycerol. Appl Biochem Biotechnol , v. 166, n. 1, p. 146 – 158, 2012.
BEOPOULOS, A.; MROZOVA, Z.; THEVENIEAU, F.; LEDALL, M.-T.; HAPALA, I.; PAPANIKOLAOU, S. Control of lipid accumulation in the yeast Yarrowia lipolytica. Appl. Environ. Microbiol. , v. 74, p. 7779 – 7789, 2008.
BEOPOULOS, A.; CESCUT, J.; HADDOUCHE, R.; URIBELARREA, J.; MOLINA-JOUVE, C.; NICAUD, J-M. Yarrowia lipolytica as a model for bio-oil production. Progress in Lipid Research , v. 48, p. 375-387, 2009.
BEOPOULOS, A.; DESFOUGERES, T.; SABIROVA, J.; ZINJARDE, S.; NEUVÉGLISE, C.; NICAUD, J-M. The hydrocarbon-degrading oleaginous yeast Yarrowia lipolytica. Handbook of hydrocarbon and lipid microbiology , Berlin Heidelberg: Springer, p. 2111 – 2121, 2010.
BEOPOULOS, A.; NICAUD, J-M. Yeast: a new oil producer? OCL, v. 19, p. 22 – 28, 2012.
BLIGH, E. G.; DYER, W. J. A rapid method of total lipid extraction and purification. Can J Biochem Physiol. , v. 37, p. 911–917, 1959.
BOZAQUEL-MORAIS, B. L. Identificação de reguladores do metabolismo de lpídios neutros em Saccharomyces cerevisiae: desenvolvimento e aplicação de um ensaio fluorimétrico para o estudo de corpúsculos lipídicos. Tese (doutorado), Instituto de Bioquímica Médica, Universidade Federal do Rio de Janeiro, Brasil, 2010.
CHI, Z.; ZHENG, Y.; LUCKER, B.; CHEN, S. Integrated system for production of biofuel feedstock. US PATENT, 12/512, 557, 2011.
CLEMENTE, T. E.; CAHOON, E.B. Soybean oil: genetic approaches for modification of functionality and total content. Plant Physiology , v. 151, p. 1030 – 1040, 2009.
COHEN, P. T.; BREWIS, N. D.; HUGHES, V.; MANN, D. J. Protein serine/threonine phosphatases: an expanding family. FEBS Lett , v. 268, p. 355 – 359, 1990.

Capítulo 8 – Referências Bibliográficas Valoni, E.A.
68
COHEN, P. T. Novel protein serine/threonine phosphatases: variety is the spice of life. Trends Biochem Sci , v. 22, p. 245 – 251, 1997.
CZABANY, T.; ATHENSTAEDT, K.; DAUM, G. Synthesis, storage and degradation of neutral lipids in yeast. Biochim Biophys Acta , v. 1771, p. 299 – 309, 2007.
CZABANY, T.; WAGNER, A.; ZWEYTICK, D.; LOHNER, K.; LEITNER, E.; INGOLIC, E. Structural and biochemical properties of lipid particles from the yeast Sacharomyces cerevisiae. J Biol Chem , v. 283, p. 17065 – 17074, 2008.
DA SILVA, G.P.; MACK, M.; CONTIERO, J. Glycerol: A promising and abundant carbono source for industrial microbiology. Biotechnol Adv , v. 27, n.1, p. 30-39, 2009.
DAVIES, R. J.; HOLDSWORTH, J. E. Synthesis of lipids in yeasts: Biochemistry, physiology and production. Adv. Appl. Lipid Res. , v. 1, p. 119 – 159, 1992.
DEY, P.; MAITI, M.K. Molecular characterization of a novel isolate of Candida tropicalis for enhanced lipid production. J. Appl. Microbiol , v. 114, p. 1357 – 1368, 2013.
EINSELE, A. Scaling-up of bioreactors. Process Biochem , p. 13 – 14, 1978.
ENFORS, S. O.; JAHIC, M.; ROZKOV, A.; XU, B.; HECKER, M.; JURGEN, B.; KRUGER, E.; SCHWEDER, T.; HAMER, G.; O´BEIRNE, D.; NOISOMMIT-RIZZI, N.; REUSS, M. Physiological responses to mixing in large bioreactors. J. Biotechnol. , v. 85, p. 175 – 185, 2001
ENSHAEIEH, M.; ABDOLI, A.; NAHVI, I.; MADANI, M. Bioconversion of different carbon sources into microbial oil and biodiesel using oleaginous yeasts. Biol. Todays World , v.1, p. 82-92, 2012.
FAKAS, S.; PAPANIKOLAOU, S.; GALIOTOU-PANAYOTOU, M.; KOMAITIS, M.; AGGELIS, G. Organic nitrogen of tomato waste hydrolysate enhances glucose uptake and lipid accumulation in Cunninghamella echinulata. J. Appl. Microbiol. , v. 105, p. 1062 – 1070, 2008.
FARESE-JR, R.V.; WALTHER, T.C. Lipid droplets finally get a little R-E-S-P-E-C-T. Cell , v. 139, n. 5, p. 855 – 860, 2009.

Capítulo 8 – Referências Bibliográficas Valoni, E.A.
69
FARIAS, M. A. Avaliação técnico-econômica do processo integrado da produção de biodiesel via enzimática utilizando lipase de Yarrowia lipolytica. Dissertação (mestrado), Escola de Química/Universidade Federal do Rio de Janeiro, Brasil, 2012.
FEI, W.; SHUI, G.; GAETA, B.; DU, X.; KUERSCHNER, L.; LI, P.; BROWN, A.J.; WENK, M.R.; PARTON, R.G.; YANG, H. Fld1p, a functional homologue of human seipin, regulates the size of lipid droplets in yeast. J. Cell Biol. , v. 180, p. 474 – 482, 2008.
FELLNER, T.; LACKNER, D. H.; HOMBAUER, H.; PIRIBAUER, P.; MUDRAK, I.; ZARAGOZA, K.; JUNO, C.; OGRIS, E. A novel and essential mechanism determining specificity and activity of protein phosphatase 2A (PP2A) in vivo. Genes Dev , v. 17, n. 17, p. 2138 – 2150, 2003.
FICKERS, P.; BENETTI, P. H.; WACHÉ, Y.; MARTY, A. Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica, and its potential applications. FEMS Yeast Res., v. 5, p. 527 – 543, 2005.
FIEDUREK, J.; SKOWRONEK, M.; GROMADA, A. Selection and adaptation of Saccharomyces cerevisiae to increase ethanol tolerance and production. Pol. J. Microbiol , v. 60, p. 51 – 58, 2011.
FRANKLIN, S.; PIECHOCKI, J.; NORRIS, L.; DECKER, S.; WEE, J.; ZDANIS, D. Oleaginous yeast food compositions. US PATENT, 13/087, 305, 2011.
FRANKLIN, S.; DECKER, S.M.; WEE, J. Fuel and chemical production from oleaginous yeast. US PATENT, 13/087, 311, 2011b.
FUJIMOTO, T.; PARTON, R.G. Not just fat: the structure and functiom of the lipid droplet. Cold Spring Harb Perspect Biol , v. 3, n. 3, p. 1 – 17, 2011.
GASMI, N.; JACQUES, P.; KLIMOVA, N.; GUO, X.; RICCIARDI, A.; ROBERT, F.; TURCOTTE, B. The switch from fermentation in Saccharomyces cerevisiae is regulates by the Ert1 transcriptional activator/repressor. Genetics , publicado primeiro online, Disponível em: htttp://www.genetics.org/content/early/2014/08/18/genetics.114.168609.long. Acesso em: 22/09/2014.
GAUSSIN, V.; HUE, L.; STALMANS, W.; BOLLEN, M. Activation of hepatic acetyl-CoA carboxylase by glutamate and Mg2+ is mediated by protein phosphatase-2A. Biochem J , v. 316, n. Pt1, p. 217 – 224, 1996.

Capítulo 8 – Referências Bibliográficas Valoni, E.A.
70
GUO, X.; TOMONAGA, T.; YANAGIHARA, Y.; OTA, Y. Screening for yeasts incorporating the exogenous eicosapentaenoic and docosahexaenoic acids from crude fish oil. J. Biosci. Bioeng. , v. 87, p. 184 – 188, 1999.
HEERES, A. S.; PICONE, C.S.F.; VAN DER WIELEN, L.A.M.; CUNHA, R.L.; CUELLAR, M.C. Microbial advanced biofuels production: overcoming emulsification challenges for large-scale operation. Trends in Biotechnology , v. 32, n.4, p. 221 – 229, 2014.
HENRY, S. A.; KOHLWEIN, S.D.; CARMAN, G.M. Metabolism and regulation of glycerolipids in the yeast Saccharomyces cerevisiae. Genetics , v. 190, p. 317 – 349, 2012.
HO, Y.; GRUHLER, A.; HEILBUT, A.; BADER, G. D.; MOORE, L.; ADAMS, S. L.; MILLAR, A.; TAYLOR, P.; BENNET, K.; BOUTILIER, K.; YANG, L.; WOLTING, C.; DONALDSON, I.; SCHANDORFF, S.; SHEWNARANE, J.; VO, M.; TAGGART, J.; GOUDREAULT, M.; MUSKAT, B.; ALFARANO, C.; DEWAR, D.; LIN, Z.; MICHALICKOVA, K.; WILLEMS, A. R.; SASSI, H.; NIELSEN, P. A.; RASMUSSEN, K.J.; ANDERSEN, J. R.; JOHANSEN, L. E.; HANSEN, L. H.; JESPERSEN, H.; PODTELEJNIKOV, A.; NIELSEN, E.; CRAWFORD, J.; POULSEN, V.; SORENSEN, B. D.; MATTHIESEN, J.; HENDRICKSON, R. C.; GLEESON, F.; PAWSON, T.; MORAN, M. F.; DUROCHER, D.; MANN, M.; HOGUE, C. W.; FIGEYS, D.; TYERS, M. Systematic identification of protein complexes in Saccharomyces cerevisiae by mass spectrometry. Nature , v. 415, p. 180 – 183, 2002.
HOJA, U.; MARTHOL, S.; HOFMANN, J.; STEGNER, S.; SCHULZ, R.; MEIER, S.; GREINER, E.; SCHWEIZER, E. HFA1 encoding an organelle-specific acetyl-CoA carboxylase controls mithocondrial fatty acid synthesis in Saccharomyces cerevisiae. J Biol Chem , v. 279, p. 21779 – 21786, 2004.
HUANG, C.; CHEN, X.-F.; XIONG, L.; CHEN, X.-D.; MA, L.-L.; CHEN, Y. Single cell oil production from low-cost substrates: the possibility and potential of its industrialization. Biotechnol. Adv. , v. 31, p. 129 – 139, 2013.
INGEBRITSEN, T. S.; COHEN, P. The protein phosphatases involved in cellular regulation. 1. Classification and substrates specificities. Eur J Biochem , v. 132, p. 255 – 261, 1983.
JANDROSITZ, A.; PETSCHNIGG, J.; ZIMMERMANN, R.; NATTER, K.; SCHOLZE, H.; HERMETTER, A.; KOHLWEIN, S.D.; LEBER, R. The lipid droplet enzyme Tgl1p hydrolyzes both steryl esters and triglycerides in the yeast, Saccharomyces cerevisiae. Biochim Biophys Acta , v. 1735, n. 1, p. 50 – 58, 2005.
JOHNSON, L. N.; BARFORD, D. The effects of phosphorylation on the structure and function of proteins. Annu Rev Biophys Biomol Struct , v. 22, p. 199 – 232, 1993.

Capítulo 8 – Referências Bibliográficas Valoni, E.A.
71
JUNKER, B. H.; MANN, Z.; SEELEY, A.; ZHANG, J.; GREASHAM, R. Use of glucose feeding to produce concentrated yeast cells. Appl. Biochem. Biotechnol , v. 97, p. 63 – 78, 2002.
KINOSHITA, H.; OTA, Y. Concentration of docosahexaenoic acid from fish oils using Geotrichum sp. FO347-2. Biosci. Biotechnol. Biochem. , v. 65, p. 1022 – 1026, 2001.
KOHLWEIN, S.D. Triacylglycerol homeostasis: insights from yeast. J. Biol Chem , v. 285, p. 15663 – 15667, 2010.
KOHLWEIN, S.D.; VEENHUIS, M.; VAN DER KLEI, I. J. Lipid droplets and peroxisomes: key players in cellular lipid homeostasis or a matter of fat-store ‘ ‘em up or bum ‘em down. Genetics , v. 193, p. 1 – 50, 2013.
KOUTINAS, A. A.; CHATZIFRAGKOU, A.; KOPSAHELIS, N.; PAPANIKOLAOU, S.; KOOKOS, I. K. Design and techno-economic evaluation of microbial oil production as a renewable resource for biodiesel and oleochemical production. Fuel , v. 116, p. 566 – 577, 2014.
LEBER, R.; ZINSER, E.; ZELLNIG, G.; PALTAUF, F.; DAUM, G. Characterization of lipid particles of the yeast, Saccharomyces cerevisiae. Yeast , v. 10, p. 1421 – 1428, 1994.
LI, Q.; WANG, M.Y. Use food industry waste to produce microbial oil. Science and Technology of Food Industry , v. 6, p. 65 – 69, 1997.
LI, Q.; DU, W.; LIU, D. Perspectives of microbial oils for biodiesel production. Appl. Microbiol. Biotechnol ., v. 80, p. 749-756, 2008.
LIDEN, G. Understanding the bioreactor. Bioproc. Biosyst. Eng. , v. 24, p. 273 – 279, 2002
LOWRY, O. H.; ROSEBROUGH, N.J.; FARR, A.L.; RANDALL, R.J. Protein measurement with the Folin phenol reagent. The Journal of Biological Chemistry , v. 193, p. 265 – 275, 1951.
MA, Y.L. Microbial oils and its research advance. Chin J Bioprocess Eng , v.4, n.4, p. 7 – 11, 2006.

Capítulo 8 – Referências Bibliográficas Valoni, E.A.
72
MIAO,X.; WU, Q. Biodiesel production from heterotrophic microalgal oil. Bioresour Technol , v. 97, p. 841 - 84, 2006.
MENG, X.; YANG, J.; XU, X.; ZHANG, L.; NIE, Q.; XIAN, M. Biodiesel production from oleaginous microorganisms. Renewable Energy , v. 34, p. 1 – 5, 2009.
MLICKOVA, K.; ROUX, E.; ATHENSTAED, K.; D´ANDREA, S.; DAUM, G.; CHARDOT, T.; NICAUD, J-M. Lipid accumulation, lipid body formation, and acyl Coenzyme A oxidases of the yeast Yarrowia lipolytica. Appl Environ Microbiol. , v. 70, n. 7, p. 3918 – 3924, 2004.
MORIN, N.; CESCUT, J.; BEOPOULOS, A.; LELANDAIS, G.; LEBERRE, V.; URIBELARREA, J.L. Transcriptomic analyses during the transition from biomass production to lipid accumulation in the oleaginous yeast Yarrowia lipolytica. PLoS ONE, v. 6, n. 11, p. 1 – 13, 2011.
MUNDAY, M. R.; CAMPBELL, D. G.; CARLING, D.; HARDIE, D. G. Identification by amino acid sequencing of three major regulatory phosphorylation sites on rat acetyl-CoA carboxilase. Eur J Biochem , v. 175, p. 331 – 338, 1988.
MURPHY, D.J. The biogenesis and functions of lipid bodies in animals, plants and microorganisms. Prog. Lipid Res. , v. 40, p. 325 – 438, 2001.
NATTER, K.; LEITNER, P.; FASCHINGER, A.; WOLINSKI, H.; McCRAITH, S.; FIELDS, S.; KOHLWEIN, S. The spatial organization of lipid synthesis in the yeast Saccharomyces cerevisiae derived from large scale green fluorescent protein tagging and high resolution microscopy. Mol. Cell Proteomics , v. 4, p. 662 – 672, 2005.
NAVEH, D. Scale-up of fermentation for recombinant DNA products. Food Technol. , v. 39, p. 102 – 108, 1985.
NOORMAN, H. An industrial perspective on bioreactor scale-down: what can we learn from combined large-scale bioprocess and model fluid studies. Biotechnol. J. , v. 6, n. 8, p. 934 – 943, 2011.
PAPANIKOLAOU, S.; CHEVALOT, I.; KOMAITIS, M.; AGGELIS, G.; MARC, I. Kinetic profile of the cellular lipid composition in an oleaginous Yarrowia lipolytica capable of producing a cocoa-butter substitute from industrial fats. Antonie Van Leeuwenhoek , v. 80, p. 215 – 224, 2001.

Capítulo 8 – Referências Bibliográficas Valoni, E.A.
73
PAPANIKOLAOU, S., AGGELIS, G., Lipid production by Yarrowia lipolytica growing on industrial glycerol in a single-stage continuous culture. Bioresource Technology, v.82, p. 43–49, 2002.
PAPANIKOLAOU, S.; CHEVALOT, I.; KOMAITIS, M.; MARC, I.; AGGELIS, G. Single cell oil production by Yarrowia lipolytica growing on an industrial derivative of animal fat in batch cultures. Appl. Microbiol. Biotechnol. , v. 58, p. 308 – 312, 2002b.
PAPANIKOLAOU, S.; MUNIGLIA, L.; CHEVALOT, I.; AGGELIS, G.; MARC, I. Accumulation of cocoa-butter-like lipid by Yarrowia lipolytica cultivated on agro-industrial residues. Curr. Microbiol , v. 46, n. 2, p. 0124-0130, 2003.
PAPANIKOLAOU, S.; CHEVALOT, I.; GALIOTOU-PANAYOTOU,M.; KOMAITIS, M.; MARC, I.; AGGELIS, G. Industrial derivative of tallow: a promising renewable substrate for microbial lipid, single-cell protein and lipase production by Yarrowia lipolytica. Electron. J. Biotechnol. , v.10, n. 3, p. 426 – 435, 2007.
PAPANIKOLAOU, S.; CHATZIFRAGKOU, A.; FAKAS, S.; GALIOTOU-PANAYOTOU, M.; KOMAITIS, M.; NICAUD, J-M. Biosynthesis of lipids and organic acids by Yarrowia lipolytica strains cultivated on glucose. Eur. J. Lipid Sci. Technol. , v. 11, p. 1221 – 1232, 2009.
PAPANIKOLAOU, S.; AGGELIS, G. Biotechnological valorization of biodiesel derived glycerol waste through production of single cell oil and citric acid by Yarrowia lipolytica. Lipid Technol. , v. 21, p. 83 – 87, 2009b.
PAPANIKOLAOU, S.; AGGELIS, G. Yarrowia lipolytica: A model microorganism used for the production of tailor-made lipids. Eur. J. Lipid Sci. Technol , v.112, p. 639–654, 2010.
PAPANIKOLAU, S.; AGGELIS, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. European Journal of Lipid Science and Technology , v.113, n.8, p. 1031-1051, 2011.
PÉREZ-TORRADO, R.; MATALLANA, E. Enhanced fermentative capacity of yeasts engineered in storage carbohydrate metabolismo. Applied Cellular Physiology and Metabolic Engineering , doi: 10.1002/btpr.1993, aceito em setembro de 2014.
RAIMONDI, S.; ROSSI, M.; LEONARDI, A.; BIANCHI, M.; RINALDI, T.; AMARETTI, A. Getting lipids from glycerol: new perspectives on biotechnological exploitation of Candida freyschussii. Microbial Cell Factories , v. 13, p. 83, 2014.

Capítulo 8 – Referências Bibliográficas Valoni, E.A.
74
SAMUL, D.; LEJA, K.; GRAJEK, W. Impurities of crude glycerol and their effect on metabolite production. Ann. Microbiol , v. 64, p. 891 – 898, 2014.
SANDAGER, L.; GUSTAVSSON, M.; STAHL, U.; DAHLQVIST, A.; WIBERG, E.; BANAS, A.; LENMAN, M., RONNE, H.; STYMNE, S. Store lipid synthesis is non-essential in yeast. J. Biol. Chem ., v. 277, p. 6478 – 6482, 2002.
SHA, Q. A comparative study on four oleaginous yeasts on their lipid accumulating capacity. Dissertação (Mestrado), Swedish University of Agricultural Sciences, Uppsala, 2013.
SITEPU, I. R.; GARAY, L. A.; SESTRIC, R.; LEVIN, D.; BLOCK, D. E.; GERMAN, J. B.; BOUNDY-MILLS, K. L. Oleaginous yeasts for biodiesel: Current and future trends in biology and production. Biotechnology Advances , v. 32, p. 1336 – 1360, 2014.
SIURKUS, J, PANULA- PERALA, J.; HORN, U.; KRAFT, M.; RIMSELIENEZ, R.; NEUBAUER, P. Novel approach of high cell density recombinant bioprocess development: Optimisation and scale-up from microlitre to pilot scales while maintaining the fed-batch cultivation mode of E. coli cultures. Microbial Cell Factories , v. 9, p. 35, 2010.
SUBRAMANIAM, R.; DUFRECHE, S.; ZAPPI, M.; BAJPAI, R. Microbial lipids from renewable resources: Production and characterization. J. Ind. Microbiol. Biotechnol. , v. 37, p. 1271 – 1287, 2010.
SWINNEN, S.; KLEIN, M.; CARRILLO, M.; McINNES, J.; THI, H.; NGUYEN, T.; NEVOIGT, E. Re-evaluation of glycerol utilization in Saccharimyces cerevisiae: characterization of an isolate that grows on glycerol without supporting supplements. Biotechnology for Biofuels , v. 6, p. 157.
TACCARI, M.; CANONICO, L.; COMITINI, F.; MANNAZZU, I.; CIANI, M. Screeinig of yeasts for growth on crude glycerol and optimization of biomass production. Bioresource Technology , v. 110, p. 488 – 495, 2012.
TAKORS, R. Scale-up of microbial processes: Impact, tools and open questions. Journal of Biotechnology , v. 160, p. 3 – 9, 2012.
TAI, M.; STEPHANOPOULOS, G. Engineering the push and pull of lipid biosynthesis in oleaginous yeast Yarrowia lipolytica for biofuel production. Metab. Eng. , v. 15, p. 1 – 9, 2013.

Capítulo 8 – Referências Bibliográficas Valoni, E.A.
75
TANIMURA, A.; TAKASHIMA, M.; SUGITA, T.; ENDOH, R.; KIKUKAWA M.; YAMAGUCHI, S.; SAKURADANI, E.; OGAWA, J.; SHIMA, J. Selection of oleaginous yeasts with high lipid productivity for practical biodiesel production. Biosource Technology , v.153, p. 230-235, 2014.
THORPE, R. F.; RATLEDGE, C. Fatty acid distribution in triglycerides of yeasts grown on glucose and n-alkanes. J. Gen. Microbiol. , v. 75, p. 151 – 163, 1972.
TSIGIE, Y.; WANG, C.; TRUONG,C.; JU,Y. Lipid production from Yarrowia lipolytica Po1g grown in sugarcane. Bioresour. Technol , v.102, n. 19, p. 9216 – 9222, 2011.
US Energy Information Administration. Energy consumption by sector. http://www.eia.gov/totalenergy/data/annual/pdf/sec2.pdf, 2013. Acessado em: 16 de Setembro de 2014.
VOTRUBA, J.; SOBOTKA, M. Physiological similarity and bioreactor scale-up. Folia Microbiol. , v. 37, p. 331 – 345, 1992.
WATKINS, P.A.; LU, J.F.; STEINBERG, S.J.; GOULD, S.J.; SMITH, K.D.; BRAITERMAN, L.T. Disruption of the Saccharomyces cerevisiae FAT1 gene decreases very long-chain fatty acid acyl-CoA synthetase activity and elevates intracellular very long-chain fatty acid concentrations. J. Biol. Chem , v. 273, p. 18210 – 18219, 1998.
WITTERS, L. A.; WATTS, T. D. Yeast acetyl-CoA carboxylase: in vitro phosphorylation by mammalian and yeast protein kinases. Biochem Biophys Res Commum , v. 169, p. 369 - 376, 1990.
WOODS, A.; MUNDAY, M .R.; SCOTT, T.; YANG, X.; CARLSON, D. Yeast SNF1 is functionally related to mammalian AMP-activated protein kinase and regulates acetyl-CoA carboxylase in vivo. J Biol Chem , v. 269, p. 19509 – 19515, 1994.
WOODBINE, M. Microbial fat: micro-organisms as potential fat procucers. Prog Ind Microbiol , v. 1, p. 179-245, 1959.
ZHANG, H.; DAMUDE, H.G.; YADAV, N.S. Three diacylglycerol acyltransferases contribute to oil biosynthesis and normal growth in Yarrowia lipolytica. Yeast , v. 29, n. 1, p. 25 – 38, 2012.
ZINSER, E.; PALTAUF, F.; DAUM, G. Sterol composition of yeast organelle membranes and subcellular distribution of enzymes involved in sterol metabolismo. J Bacteriol. , v. 175, n. 10, p. 2853 – 2858, 1993.

Capítulo 8 – Referências Bibliográficas Valoni, E.A.
76
ZWEYTICK, D.; ATHENSTAEDT, K.; DAUM, G. Intracellular lipid particles of eukaryotic cells. Biochim. Biophys. Acta , v. 18, p. 101 – 120, 2000.

77
9. Apêndices

78
APÊNDICE A – Curvas de calibração para obtenção do fator de conversão das medidas de absorvância em comprimento de onda de 600 nm para concentração celular em massa seca [g.l-1] para cepa de Saccharomyces cerevisiae selvagem BY4741
y = 2,3701xR² = 0,9991
f= 1/a= 0,42192
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0 0,02 0,04 0,06 0,08 0,1 0,12 0,14 0,16
Ab
sorv
ân
cia
60
0 n
m
Concentração (g/L)
Curva peso seco x absorvância -
Cepa WT
y = 2,7771xR² = 0,9956
f = 1/a= 0,3601
0
0,1
0,2
0,3
0,4
0,5
0,6
0 0,05 0,1 0,15 0,2 0,25
Ab
sorv
ân
cia
60
0 n
m
Concentração (g/L)
Curva peso seco x absorvância -
Cepa WT
Figura 9.1: Curva de peso seco para quantificação do crescimento celular da cepa S. cerevisiae selvagem BY4741 através de medidas de absorvância em espectrofotômetro Shimadzu UV-1800.
Figura 9.2: Curva de peso seco para quantificação do crescimento celular da cepa S. cerevisiae selvagem BY4741 através de medidas de absorvância em espectrofotômetro Shimadzu UV Mini 1240.

79
APÊNDICE B – Curva de calibração para obtenção dos fator de conversão das medidas de absorvância em comprimento de onda de 600 nm para concentração celular em massa seca [g.l-1] para cepa de Saccharomyces cerevisiae mutante RRD2
y = 2,3303xR² = 0,9919
f=1/a=0,42913
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0 0,05 0,1 0,15 0,2
Ab
sorv
ân
cia
60
0 n
m
Concentração (g/L)
Curva peso seco x absorvância Cepa
RRD2
y = 3,4503xR² = 0,9936
f = 1/a = 0,2893
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0 0,05 0,1 0,15 0,2 0,25
Ab
sorv
ân
cia
60
0 n
m
Concentração (g/L)
Curva peso seco x absorvância
Cepa RRD2
Figura 9.3: Curva de peso seco para quantificação do crescimento celular da cepa S. cerevisiae mutante RRD2 através de medidas de absorvância em espectrofotômetro Shimadzu UV-1800.
Figura 9.4: Curva de peso seco para quantificação do crescimento celular da cepa S. cerevisiae mutante RRD2 através de medidas de absorvância em espectrofotômetro Shimadzu UV-1800.





![TRIAGEM OBSTÉTRICA: UMA REVISÃO DE LITERATURA...R114 Rabelo, Érika Marina Triagem obstétrica: uma revisão de literatura / Érika Marina Rabelo – Belo Horizonte : [s.n.], 2011](https://img.pdfslide.tips/doc/110x75/608c479c351faf058b1307a3/triagem-obsttrica-uma-revisfo-de-literatura-r114-rabelo-rika-marina-triagem.jpg)























