Embed Size (px)
Citation preview

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis Doctoral
Estructuración poblacional delEstructuración poblacional delCardenal Amarillo (GubernatrixCardenal Amarillo (Gubernatrixcristata): implicancias para sucristata): implicancias para su
conservaciónconservación
Domínguez, Marisol
2015-12-21
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:
Domínguez, Marisol. (2015-12-21). Estructuración poblacional del Cardenal Amarillo(Gubernatrix cristata): implicancias para su conservación. Facultad de Ciencias Exactas yNaturales. Universidad de Buenos Aires.
Cita tipo Chicago:
Domínguez, Marisol. "Estructuración poblacional del Cardenal Amarillo (Gubernatrix cristata):implicancias para su conservación". Facultad de Ciencias Exactas y Naturales. Universidad deBuenos Aires. 2015-12-21.

UNIVERSIDAD DE BUENOS AIRES FACULTAD DE CIENCIAS EXACTAS Y NATURALES
DEPARTAMENTO DE ECOLOGÍA, GENÉTICA Y EVOLUCIÓN & IEGEBA-CONICET
Estructuración poblacional del Cardenal Amarillo (Gubernatrix cristata): implicancias para su conservación
Tesis presentada para optar al título de Doctor de la Universidad de Buenos Aires en el área: CIENCIAS BIOLÓGICAS
MARISOL DOMÍNGUEZ
Directora
Dra. Bettina Mahler Consejero de estudios
Dr. Juan Carlos Reboreda
Buenos Aires, 2015

I
ESTRUCTURACIÓN POBLACIONAL DEL CARDENAL AMARILLO (GUBERNATRIX CRISTATA): IMPLICANCIAS PARA SU CONSERVACIÓN
RESUMEN
El Cardenal Amarillo (Gubernatrix cristata) es un passeriforme endémico de América del Sur, que está categorizado como ―En peligro‖ por la Unión Internacional para la Conservación de la Naturaleza como consecuencia de la alta tasa de captura para su comercio ilegal como ave de jaula y la modificación de su hábitat. Su distribución actual es altamente discontinua, con escasas regiones donde se registra la especie, separadas por zonas donde su presencia es muy escasa o nula.Los objetivos del trabajo fueron: 1) Estudiar los principales parámetros reproductivos del Cardenal Amarillo; 2) analizar la estructuración poblacional de la especie en Argentina y Uruguay; y 3) estudiar el efecto que tiene la hibridación con la especie emparentada Diuca diuca minor en la región sur de su distribución. A través de un seguimiento de parejas reproductivas en la provincia de Corrientes se describieron los parámetros reproductivos para el Cardenal Amarillo y se observó una disminución en la supervivencia de los nidos a medida que avanza la temporada reproductiva y como consecuencia del parasitismo por el tordo renegrido Molothrus bonariensis y por larvas de moscas del género Philornis. El impacto del parasitismo no había sido considerado previamente como una posible amenaza para esta especie. Para estudiar la estructuración poblacional se analizaron individuos provenientes de cuatro regiones que se ubican en los extremos norte (Litoral, provincia de Corrientes) y sur (Provincias de Río Negro y La Pampa) y en el centro (Provincia de San Luis) de su distribución en Argentina. Además, se estudiaron individuos de la República Oriental del Uruguay. Las poblaciones fueron analizadas a nivel genético (736 pb de la región control de ADNmt y 10 loci microsatélites), morfológico (11 medidas morfométricas) y comportamental (vocalizaciones). Se encontraron diferencias entre poblaciones en todas las características estudiadas. En base a estos resultados se pudieron definir cuatro unidades de manejo para el Cardenal Amarillo, compuestas por la población de Uruguay, la población de Corrientes, la población de San Luis y norte de La Pampa, y la población de Río Negro y sur de La Pampa. En la población austral, se estudió cómo la hibridación con la Diuca Común (Diuca diuca minor) afecta la variabilidad de estos caracteres y aunque no se encontró una introgresión genética entre especies, se observó una mayor similitud de los cantos de Diuca Común con los de la población sur del Cardenal Amarillo.La caracterización de las poblaciones existentes del Cardenal Amarillo podrá utilizarse para delimitar áreas protegidas que permitan mantener una representatividad adecuada de la variabilidad poblacional a lo largo de su distribución geográfica. Estos resultados permitirán, además, asignar la proveniencia de individuos incautados por las Direcciones de Fauna, lo cual podrá servir como base para delinear futuros programas de reintroducción.
Palabras clave: Gubernatrix cristata, Cardenal Amarillo, unidades de conservación, estructuración poblacional, biología reproductiva, hibridación.

II
POPULATION STRUCTURE OF THE YELLOW CARDINAL ( GUBERNATRIX CRISTATA): CONTRIBUTIONS TO ITS CONSERVATION
ABSTRACT
The Yellow Cardinal (Gubernatrix cristata) is an endemic South American passerine, which is categorized as "endangered" by the International Union for Conservation of Nature as a result of illegal traffic and habitat modification. Its current distribution is highly discontinuous, with few areas where the species is present separated by areas with scarce or null records. The objectives of this thesis were to: 1) study the main reproductive parameters of the Yellow Cardinal; 2) analyze the population structure of the species in Argentina and Uruguay; and 3) study the effect of the hybridization with the related species Diuca diuca minor in the southern part of its distribution. Through monitoring of breeding pairs in the province of Corrientes, reproductive parameters were described and a decrease in nest survival was observed as the breeding season advances and as a result of parasitism by the Shiny Cowbird (Molothrus bonariensis) and fly larvae of the genus Philornis. The impact of parasitism had not been previously considered as a potential threat to this species. In order to study Yellow Cardinal population structure individuals from four regions at the extremes and center of the distribution were analyzed. The southern region is located in the provinces of La Pampa and Río Negro, the central region in the province of San Luis and the northern region in the province of Corrientes. In addition, individuals from Uruguay were studied. Populations were analyzed at genetic (736 bp of mtDNA control region and 10 microsatellite loci), morphological (11 morphometric measurements) and behavioral level (vocalizations). Differences between populations were found in all the characters that were studied. Based on these results, it was possible to define four management units for the Yellow Cardinal, composed by the populations of Uruguay, Corrientes, San Luis and northern La Pampa, and Río Negro and southern La Pampa. We also studied whether hybridization with the Common Diuca (Diuca diuca minor) affects the variability of these characters. Even though a genetic introgression between species was not found, a greater similarity of the songs of Common Diucas with the southern Yellow Cardinal population was observed. The description of the remaining populations of the Yellow Cardinal can be used to delineate protected areas in order to maintain a proper representation of population variability throughout its range. These results will also help to identify the origin of confiscated individuals, which may serve as a basis for outlining future reintroduction programs.
Keywords: Gubernatrix cristata, yellow cardinal, conservation units, population structure, reproductive biology, hybridization.

III
AGRADECIMIENTOS
Durante el desarrollo de esta tesis tuve la suerte de conocer a hermosas personas con quienes establecí profundas relaciones de amistad y que resultaron muy importantes para este trabajo. A ellas les estoy infinitamente agradecida.
A mi directora de tesis, Bettina Mahler, por ser un ejemplo a seguir y guiarme en el camino hacia la obtención del título de doctorado. Camino que puedo decir fue muy grato de recorrer y que superó todas mis expectativas.
A Juan Carlos Reboreda por ser mi consejero de estudios, pero también por la permanente motivación y por estar siempre que necesité su opinión.
A la Facultad de Ciencias Exactas y Naturales, por brindarme de manera gratuita la capacitación necesaria para obtener los títulos de licenciada y de doctora.
Al Servicio Alemán de Intercambio Académico (DAAD) y al Ministerio de Ciencia, Tecnología e Innovación Productiva (MINCYT) por permitirme tener una de las experiencias más enriquecedoras y a Ralph Tiedemann por aceptarme en su laboratorio y enseñarme a perfeccionar las técnicas moleculares.
Al Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Aves Argentinas y a la Universidad de Buenos Aires por financiar el proyecto.
A las autoridades nacionales y provinciales que otorgaron los permisos necesarios y por reconocer la importancia de este trabajo.
A la ONG AOP/Aves Argentinas por todo el esfuerzo que pone en generar acciones de conservación para muchas especies de aves amenazadas. Especialmente a Rodrigo Fariña, Inés Pereda y Rocío Lapido con quienes armamos un proyecto especialmente para el Cardenal Amarillo.
A Conservation Land Trust y todo el personal de la Estancia Rincón del Socorro, a Carlos Figuereo, Fabian Tittarelli, Matias Ayarragaray y Luciano Segura, por facilitar sitios de estudio y haberme permitido estar en el campo con los cardenales.
A los asistentes de campo, Martin Hoffman, Alejandro Cinalli y Sergio Briones, por la ayuda en la toma de datos y la compañía.
A mis compañeros de laboratorio, Caro, Cyn, Naty Cossa, Nati Rebolo, Romi, Juanma, Ali, Kini, Ceci, Lucho, Diego y Vani por su compañerismo y agradables jornadas laborales. A Dario, Pablito, Luciano y Naty de la división de Ornitología del Museo. Adrian e Igor por brindarme su experiencia en la conservación de fauna silvestre.
A mi familia, mamá, papá, Sebas, Mimi, Abu, tíos y Brian y a mis amigos de la vida, Mar, Gaby, Sabri So., Ray, Sabri Sa., Flor S. y Flor H., Ale, Ema G., Lu R.P., Pablito, Rama, Gaby K., Giomara, Yiyi, Belen, Lu, Miriam y a mis amigos de Iberá Kari, Ceci, Yamil y Ema.

IV
INDICE
Resumen I
Abstract II
Agradecimientos III
Índice IV
Capítulo 1: Introducción general 1
1.1 Conservación de aves amenazadas 1
1.2 Estrategias de manejo 2
1.2.1 Técnicas de manejo 2
1.2.2 Unidad de conservación 3
1.3 Especie de estudio: el Cardenal Amarillo 4
1.3.1 Filogenia 4
1.3.2 Descripción de la especie 4
1.3.3 Ambiente 6
1.3.4 Distribución 9
1.3.5 Categoría de amenaza y marco legal 11
1.4 Amenazas para el Cardenal Amarillo 12
1.5 Objetivos 13
1.5.1 Objetivo general 13
1.5.2 Objetivos particulares 13
1.7 Referencias bibliográficas 13
Capítulo 2: Biología Reproductiva y efectos del parasitismo 21
2.1 Introducción 21

V
2.2 Métodos 22
2.2.1 Sitio de estudio 22
2.2.2 Toma de datos 23
2.2.3 Análisis de datos 25
2.2.4 Análisis estadístico 26
2.3 Resultados 28
2.3.1 Construcción del nido, puesta e incubación de huevos 28
2.3.2 Eclosión y crecimiento de pichones 29
2.3.3 Distribución de las parejas 29
2.3.4 Efectos del parasitismo 30
2.3.5 Supervivencia de nidos 31
2.4 Discusión 32
2.5 Referencias bibliográficas 35
Capítulo 3: Variación geográfica del canto 40
3.1 Introducción 40
3.1.1 El comportamiento animal en la biología de la conservación 40
3.1.2 Adquisición del canto 40
3.1.3 Variación geográfica del canto 42
3.1.4 Variaciones en las características de los dialectos 42
3.1.5 Objetivos 43
3.2 Metodología 44
3.2.1 Grabación de cantos 44
3.2.2 Análisis de los cantos 45
3.2.3 Análisis estadísticos 47
3.3 Resultados 47

VI
3.4 Discusión 53
3.5 Referencias bibliográficas 56
Capítulo 4: Variación geográfica de la morfología 63
4.1 Introducción 63
4.2 Metodología 65
4.2.1 Colección de datos 65
4.2.2 Análisis estadísticos 67
4.3 Resultados 68
4.3.1 Machos 68
4.3.1.1 Peso 68
4.3.1.2 Coloración 68
4.3.1.3 Análisis multivariados de las variables morfológicas 70
4.3.2 Hembras 73
4.3.2.1 Peso 73
4.3.2.2 Coloración 73
4.3.2.3 Análisis multivariados de las variables morfológicas 74
4.4 Discusión 75
4.5 Referencias bibliográficas 76
Capítulo 5: Variación genética 82
5.1 Introducción 82
5.1.1 Conservación de la variabilidad genética 82
5.1.2 Efecto de la fragmentación del hábitat sobre la diversidad genética 82
5.1.3 Los riesgos de las poblaciones pequeñas 83
5.1.4 Estructuración de la variabilidad genética 83

VII
5.1.5 Objetivos 85
5.2 Metodología 85
5.2.1 Colección de datos 85
5.2.2 Análisis Genéticos 85
5.2.3 Estructuración poblacional genética 87
5.3 Resultados 88
5.3.1 Estadística descriptiva 88
5.3.2 Estructuración poblacional genética 92
5.4 Discusión 97
5.5 Referencias bibliográficas 99
Capítulo 6: Hibridación con la Diuca Común 104
6.1 Introducción 104
6.2 Metodología 108
6.2.1 Colección de datos 108
6.2.2 Análisis genéticos 109
6.2.3 Cantos 110
6.3 Resultados 110
6.3.1 Genética 110
6.3.2 Cantos 111
6.4 Discusión 113
6.5 Referencias bibliográficas 115
Capítulo 7: Discusión general y consideraciones finales 119
7.1 Principales resultados 119

VIII
7.1.1 Exito reproductivo 119
7.1.2 Amenazas 120
7.1.3 Unidades de manejo 120
7.2 Apliaciones para la conservación de la especie 121
7.2.1 Conservación de la variabilidad 121
7.2.2 Manejo de los cardenales decomisados 122
7.3 Referencias bibliográficas 123

INTRODUCCIÓN GENERAL
1
CAPÍTULO 1
INTRODUCCIÓN GENERAL
“El Cardenal Amarillo es uno de los fringílidos argentinos más hermosos. Es un ave vivaz y graciosa, encantadora con su traje amarillo y negro, y con una voz fuerte y musical. Por desgracia, estas cualidades han hecho de él uno de los favoritos para estar enjaulados. Los pichones son muy buscados en las florestas del norte de Argentina y Paraguay, y sacados del nido para ser educados con destreza. Su canto se compone de 4 o 5 notas de gran poder o dulzura”
Hudson 1874
1.1. Conservación de aves amenazadas
La diversidad biológica del planeta está siendo rápidamente eliminada (Lawton & May 1995). A diferencia de las extinciones masivas prehistóricas, las tasas de extinción actuales son sensiblemente más altas y están llevando a una pérdida de especies sin precedentes (Ceballos et al. 2015). Según la Lista Roja Mundial de las Aves de 2014, elaborada por BirdLife International, el 13% del total de las especies de aves conocidas en el mundo está en peligro de extinción. Esto equivale a aproximadamente una de cada ocho especies (Birdlife International 2014). En la Argentina se encuentran 50 especies de aves globalmente amenazadas (BirdLife International 2015). A nivel global, la fragmentación de los ambientes naturales, junto a la sobreexplotación e introducción de especies exóticas, son consideradas las causas más importantes de pérdida de biodiversidad (Harris 1984, Vitousek et al. 1997, Fahrig 2003, Betts et al. 2007).
La disminución de las poblaciones por la pérdida, degradación y destrucción del hábitat natural de una especie puede verse agravada por la extracción de individuos de su ambiente. Este es el caso del Cardenal Amarillo (Gubernatrix cristata), que está catalogado como ―En peligro‖ por la Unión Internacional para la Conservación de la Naturaleza (BirdLife International 2013). El tráfico ilegal de animales es y ha sido desde hace muchos años un problema para la fauna silvestre. En Argentina, la cantidad de especies silvestres (no permitidas para la venta) que se comercializan viene aumentando año tras año (Argibay et al. 2005). Una gran cantidad de comercios en todo el país venden ilegalmente diferentes

INTRODUCCIÓN GENERAL
2
especies de aves silvestres y en Capital Federal están algunos de los principales centros de comercio, como la Feria de Pompeya y de Villa Dominico, que aprovechan el status legal de venta de especies de criadero para también vender, ilegalmente, especies silvestres, algunas de ellas amenazadas de extinción.
1.2. Estrategias de manejo
1.2.1. Técnicas de manejo
Ante las evidentes disminuciones en los tamaños poblacionales, muchos planes de manejo tienden a aumentar el número de individuos para garantizar la viabilidad de las poblaciones (Allendorf & Luikart 2007).
Un proyecto de conservación idealmente debería incluir un manejo integrado, incluyendo las técnicas más adecuadas dependiendo de las amenazas que enfrente la especie, sumado a acciones que promuevan la educación y participación de la comunidad. El proceso de recuperación de una especie amenazada requiere de varias etapas. En primer lugar, es necesario hacer una síntesis del conocimiento que se tiene de la especie y evaluar los problemas que enfrenta para diagnosticar las causas que la llevaron al declive poblacional (Norris & Pain 2002). La investigación ayudará a llenar los vacíos de conocimientos que se identifiquen. Luego se deberán encarar acciones de manejo inmediatas para aumentar el tamaño poblacional y asegurar su viabilidad. Al mismo tiempo se debe trabajar con los factores que afectan negativamente a la especie para asegurar la supervivencia a largo plazo. Finalmente, un plan de monitoreo es necesario para evaluar el éxito del programa (Sutherland et al. 2004).
Las estrategias de conservación se definen de acuerdo a las amenazas que afronta cada especie y, en general, sus resultados se ven a largo plazo. El manejo exitoso de las poblaciones amenazadas requiere un conocimiento básico de la biología de la especie, incluyendo las tasas de reclutamiento, de supervivencia, longevidad, los requerimientos de hábitat y el comportamiento social. Algunos aspectos incluidos en programas exitosos de manejo comprenden factores bióticos y abióticos que aumentan las tasas reproductivas y de supervivencia de la especie amenazada, como ser: 1) manejo del hábitat, nido, refugio (ej. Philesturnus carunculatus en Nueva Zelanda, Lovegrove 1996) o alimento (ej. Cóndor Californiano Gymnogyps californianus, Wilbur 1978, y Columba mayeri en la isla Mauritius, Jones et al. 1999); 2) manejo de parásitos y enfermedades (ej. Petroica traversi, Tisdall & Merton 1988); 3) manejo de predadores y competidores exóticos (ej. recuperación de Pomarea dimidiata en las islas Cook por el control de Rattus rattus, McCormack & Künzle 1990); 4) manejo genético (ej. Petroica traversi, Merton 1990, 1992, Ardern & Lambert 1997); 5) translocaciones (Bell 1975, 1991) o reintroducciones

INTRODUCCIÓN GENERAL
3
(ej. Tetrao urogallus en Inglaterra, Perrins 1987); y 6) cría en cautiverio (ej. Cóndor Californiano, Wallace 2005).
1.2.2. Unidades de Conservación
Identificar unidades intraespecíficas que deben ser conservadas es crítico para guiar los esfuerzos de conservación (Caughley & Gunn 1996, Allendorf & Luikart 2007). En 1974, Otto Frankel menciona por primera vez la importancia de conservar la diversidad genética que se originó a lo largo de los millones de años de evolución, ya que no podemos predecir los cambios a los deberán hacer frente las especies en el futuro. Esto quiere decir que es importante conservar la mayor parte de la variabilidad original de una especie, maximizando de esta forma la habilidad para evolucionar en respuesta al cambio ambiental. A mayor variabilidad, mayor será el potencial adaptativo de una especie.
A fines de los años 90 surge como disciplina la Genética para la Conservación. En base a las variantes genéticas se definen dentro de una especie unidades evolutivamente significativas con prioridad de conservación (UESs) (Ryder 1986). Las UES representan grupos conespecíficos con monofilia recíproca que atravesaron un periodo de aislamiento suficientemente largo que podría haber llevado a diferencias adaptativas entre las poblaciones y cada una de ellas representaría un componente importante para el potencial adaptativo de la especie.
En la actualidad, este concepto se considera demasiado estricto, ya que sólo especies que muestren evidencias de subdivisión en varias subespecies caerían dentro del concepto de UES (DeWeerdt 2002). Por este motivo, es más ampliamente utilizado el concepto de unidades de manejo (UM) definido por Moritz (1994). Las UMs se componen de poblaciones con diferencias substanciales en las frecuencias alélicas de varios loci. Estas poblaciones serían demográficamente independientes, con frecuencias alélicas distintivas, pero también con cierto grado de flujo génico entre sí. En general, dan información de historias demográficas más recientes que las ESUs. Las UMs pueden estar definidas no solamente por caracteres genéticos sino también por caracteres relacionados con la morfología y el comportamiento (Green 2005, Allendorf & Luikart 2007). De esta forma, el concepto de UM es superador ya que permite conservar, además de la variabilidad genética, la variabilidad morfológica y comportamental presente en una especie (Mortiz 1994, Crandall et al. 2000, Palsbøll et al. 2006).

INTRODUCCIÓN GENERAL
4
1.3 . Especie de estudio: el Cardenal Amarillo
1.3.1. Filogenia
El Cardenal Amarillo, Gubernatrix cristata (Vieillot 1817), es un paseriforme endémico del sur de América del Sur y es el único representante del género monotípico Gubernatrix. Está incluido dentro de la familia Thraupidae junto con otros géneros de aves granívoras como Diuca, Paroaria y Lophospingus (Campagna et al. 2011, Barker et al. 2013, Burns et al. 2014). El género Gubernatrix ha variado su posición filogenética dentro de los paseriformes (familia Emberizidae: De la Peña 1981, Bascarán 1987, Collar et al. 1994, Pessino & Tittarelli 2006; familia Fringillidae: Meyer de Schauensee 1966, Sibley & Ahlquist 1990, Sibley & Monroe 1990). Sin embargo, el género fue transferido de Emberizidae a Thraupidae de acuerdo con la Propuesta N° 512 al South American Classification Committee (SACC) en base a la morfología y datos genéticos provistos por Campagna et al. 2011. Actualmente, existe consenso en que Gubernatrix es un género integrante de la familia Thraupidae (Barker et al. 2013, Burns et al. 2014, Barker et al. 2015). La familia Thraupidae está compuesta por más de 90 géneros y casi 400 especies de aves distribuidas en el Neotrópico (Sick 1997, Isler & Isler 1999). Es la segunda familia más numerosa, después de Tyrannidae, ya que representa cerca del 4% de todas las especies de aves (Burns et al. 2014). Debido a su colorido plumajeson unas de las especies más llamativas del Neotrópico (Isler & Isler 1999), y, sumado a su adaptabilidad al cautiverio, son muy comercializadas como mascotas.
1.3.2. Descripción de la especie
El Cardenal Amarillo, también conocido como Cardenal de Copete Negro o Cardeal amarelo (Brasil), es un ave que mide aproximadamente 20 cm de longitud (Bascarán 1987) y posee un plumaje mayormente amarillo oliváceo con algunas manchas y estrías negras, especialmente en el dorso. Su copete y garganta son negros y la cola es amarilla con timoneras centrales negras (Narosky & Yzurieta 2003). El pico es cónico y grisáceo, las patas oscuras y los ojos marrones. En su estadio adulto se evidencia un notable dimorfismo sexual: el macho ostenta un plumaje más colorido que la hembra (Figura 1.1). El macho tiene líneas superciliares, malar y vientre de color amarillo (Figura 1.1A), mientras que la hembra tiene líneas superciliares y malar blancos en lugar de amarillos, con mejillas, pecho y flancos grises (Figura 1.1B; Bascarán 1987). El juvenil es más parecido a la hembra y exhibe una coloración apagada donde predomina el color gris moteado de negro en partes superiores e inferiores (Figura 1.2). Es un ave muy activa, y en particular el macho tiene un canto melodioso y defiende agresivamente su territorio durante la etapa reproductiva (Collar et al. 1992, Sick 1997). Las hembras no cantan pero producen débiles voces de contacto (Chebez 2008).

INTRODUCCIÓN GENERAL
5
Figura 1.1. Fotografías de un macho adulto (A) y una hembra adulta (B) de Cardenal Amarillo donde se puede apreciar el dimorfismo sexual.
Figura 1.2. Fotografías de juveniles de Cardenal Amarillo. A= juvenil de 35 días de edad. B= hembra (a la izquierda) y juvenil de 25 días de edad (a la derecha).
La alimentación de los adultos consiste principalmente de granos, frutos y pastos tiernos (como verdolaga, diente de león, acelga silvestre y achicoria silvestre, entre otros),
A B
A
B

INTRODUCCIÓN GENERAL
6
pero los pichones son alimentados básicamente con insectos (Bascarán 1987, Sick 1997). El tipo de granos y frutos depende de la zona donde se encuentren.
1.3.3. Ambiente
El Cardenal Amarillo habita bosques abiertos y matorrales característicos de la ecorregión del espinal, representada por la presencia de árboles y arbustos espinosos. También se lo encuentra en el ecotono con las ecorregiones chaqueña y del monte. El espinal presenta, en casi toda su extensión, parches de bosque intercalados con pastizales, un paisaje que recuerda a la sabana africana. Estos ambientes mixtos son los utilizados por el Cardenal Amarillo.
A lo largo del espinal, estos bosques van reemplazando sus especies dominantes formando tres distritos diferentes. El distrito norteño (Figura 1.3) ocupa el centro y sur de la mesopotamia argentina y el Ñandubay (Prosopis affinis) es la especie principal. En este tipo de espinal se pueden encontrar también Algarrobo Negro (Prosopis nigra), Tala (Celtis ehrenbergiana), Chañar (Geoffroea decorticans), Sombra de toro (Jodina rhombifolia) y el Molle (Schinus longifolius).
Figura 1.3. Ambiente del espinal en el distrito norte. Fotografía tomada en la provincia de Corrientes.
En el distrito oeste (Figura 1.4), que ocupa las provincias de Santa Fe y Córdoba, se destacan los Algarrobos Blanco (Prosopis alba) y Negro. Otras especies acompañantes en

INTRODUCCIÓN GENERAL
7
esta región son el Espinillo (Acacia cavens), Molle, Coronillo (Scutia buxifolia) y Quebracho Blanco (Aspidosperma quebracho-blanco).
Figura 1.4. Ambiente del espinal en el distrito oeste. Fotografía tomada en la provincia de Santa Fe.
En el distrito sur (Figura 1.5), que se ubica en el centro y sur de San Luis, La Pampa, Río Negro y Buenos Aires, la especie dominante es el Caldén (Prosopis caldenia), el cual es endémico de la región. Otras especies que se encuentran en este distrito son el chañar, Sombra de toro y Algarrobo Negro. El Cardenal Amarillo acompaña dicha formación, a lo largo de los tres distritos.
Figura 1.5. Ambiente del espinal en el distrito sur. Fotografía tomada en la provincia de Río Negro.

INTRODUCCIÓN GENERAL
8
En el pasado, el espinal constituía un anillo boscoso que rodeaba la región pampeana, conformando una transición entre el pastizal pampeano, la región chaqueña y el monte (Cabrera 1976, Figura 1.6). El espinal se extendía por más de 24.000.000 hectáreas, cubriendo más del 8 por ciento de la superficie de Argentina (Brown & Pacheco 2006).
Figura 1.6. Ecorregiones fitogeograficas según Cabrera 1976. Se observa la extensión que cubría el espinal.
Debido al avance de la frontera agropecuaria y a la extracción de leña, los bosques de espinal han disminuido considerablemente. En el año 2005 se estimó una pérdida de alrededor del cuarenta por ciento de esta superficie (Brown & Pacheco 2006) con más de 9.000.000 de hectáreas transformadas. En el distrito norteño, el desmonte se incrementó con la rápida expansión de los cultivos de soja. Esta expansión afectó tierras del espinal que habían permanecido bajo un uso ganadero extensivo en Entre Ríos y Corrientes. Además, en estas provincias, los bosques del espinal han sido y siguen siendo reemplazados por plantaciones de Eucalyptus. En el distrito sur, los caldenales, por su menor aptitud agrícola, fueron reemplazados para aprovechamiento forestal (Lell 2004). El espinal ha sufrido también la invasión de especies exóticas como la morera (Morus alba), el ligustro

INTRODUCCIÓN GENERAL
9
(Ligustrum lucidum) y el paraíso (Melia azederach). Debido a estos factores, Brown & Pacheco proponen en 2006 un nuevo mapa de ecorregiones donde se puede observar la disminución de la superficie del espinal (Figura 1.7).
Figura 1.7. Mapa de ecorregiones argentinas según Brown & Pacheco (2006). Se observa la retracción del espinal en las provincias de Santa Fe y La Pampa.
El Cardenal Amarillo depende de este ambiente para sobrevivir. El Espinal también proporciona el hábitat a una de las aves rapaces más amenazadas de la Argentina, el Águila Coronada (Harphyaeletus coronatus), brinda refugio a poblaciones del Venado de las Pampas (Ozoteceros bezoarticus), y es habitado por otros mamíferos amenazados como el Aguará Guazú, el Pecarí de Collar y el Lobo de Gargantilla.
1.3.4. Distribución
La distribución antigua del Cardenal Amarillo se extendía desde el extremo sur de Brasil hasta el centro de Argentina incluyendo Uruguay (Ridgely & Tudor 1997, Figura 1.8). Incluso llegó a considerarse una especie común y de amplia distribución en Argentina
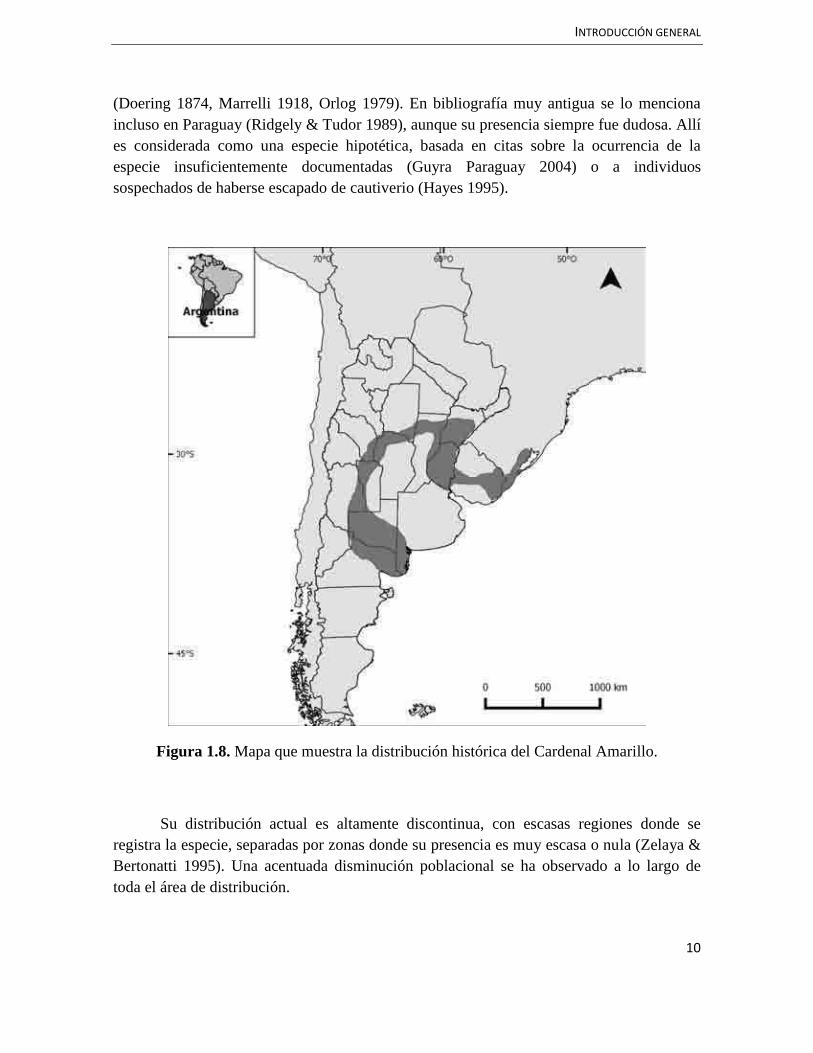
INTRODUCCIÓN GENERAL
10
(Doering 1874, Marrelli 1918, Orlog 1979). En bibliografía muy antigua se lo menciona incluso en Paraguay (Ridgely & Tudor 1989), aunque su presencia siempre fue dudosa. Allí es considerada como una especie hipotética, basada en citas sobre la ocurrencia de la especie insuficientemente documentadas (Guyra Paraguay 2004) o a individuos sospechados de haberse escapado de cautiverio (Hayes 1995).
Figura 1.8. Mapa que muestra la distribución histórica del Cardenal Amarillo.
Su distribución actual es altamente discontinua, con escasas regiones donde se registra la especie, separadas por zonas donde su presencia es muy escasa o nula (Zelaya & Bertonatti 1995). Una acentuada disminución poblacional se ha observado a lo largo de toda el área de distribución.

INTRODUCCIÓN GENERAL
11
En Brasil, la especie ha sido avistada ocasionalmente sólo en el Parque Estatal Espinilho, en Río Grande do Sul (Martins-Ferreira et al. 2010). En Uruguay existen registros actuales en Sierras del Este y en el litoral del Río Uruguay (Azpiroz et al. 2012).
Dentro de Argentina, fue inventariada para 28 AICAs (Áreas importantes para la conservación de las aves, Di Giacomo 2005c), distribuidas a lo largo de 11 provincias. En el litoral, su abundancia está centrada en Corrientes (Di Giacomo 2005a, Fraga 2005, Giraudo & Di Giacomo 2005, Giraudo et al. 2005) y Entre Ríos (Chebez & Morandeira 2005, Di Giacomo 2005b, Frucella & Bosso 2005, Veiga 2005) y algunos sectores del noreste de Santa Fe (Pautasso 2002; De la Peña 2005; Di Giacomo, Di Giacomo & Pautasso 2005; Ortiz 2008), mientras que en el sur, las poblaciones más estables se encuentran en las provincias de La Pampa (Maceda 2005a, Maceda 2005b, Maceda 2005c, Tittarelli 2005, Veiga & Tittarelli 2005) y Río Negro (González 2005, Chebez 2008). También han sido registradas poblaciones pequeñas en las provincias de San Luis (Domínguez et al. En prensa) y Mendoza (Sosa et al. 2011).
Existen pequeñas poblaciones de Cardenal Amarillo que se encuentran en áreas protegidas: en el Parque Nacional Lihué Calel (La Pampa), en los parques provinciales Quebracho de la Legua (San Luis) e Iberá (Corrientes) y en la Reserva Provincial Parque Luro (La Pampa).
1.3.5. Categoría de amenaza y marco legal
El Cardenal Amarillo es una especie categorizada globalmente como ―En Peligro‖ por la Unión Internacional para la Conservación de la Naturaleza debido a la pérdida de hábitat y a la disminución poblacional (criterios A2cd, 3cd y 4cd, Collar et al. 1992, BirdLife 2013). Si bien no es endémica de Argentina, gran parte de su distribución se centra en el país (Figura 1.9). A nivel nacional el Cardenal Amarillo también está considerado como ―En Peligro‖ por resolución Nº 348/2010 de la Secretaría de Ambiente y Desarrollo Sostenible de la Nación. Esta especie se encuentra protegida bajo la Ley Nacional de Conservación de la Fauna Silvestre Nº 22.421 y su Decreto Reglamentario Nº 666/97. El artículo 20 establece que el Poder Ejecutivo Nacional debe adoptar medidas de emergencia a fin de asegurar la repoblación y perpetuación, pudiendo disponer también la prohibición de la caza, del comercio interprovincial y de la exportación de los ejemplares y productos y subproductos de las especies silvestres autóctonas que se encuentran amenazadas, como es el caso del Cardenal Amarillo.
En el contexto del comercio internacional, Argentina adhierió en 1981 a la Convención Internacional para el Comercio de Especies Amenazadas de Fauna y Flora Silvestres (CITES) mediante la Ley 22.344. Dicha Convención posee listados que se

INTRODUCCIÓN GENERAL
12
denominan apéndices, los cuales se encuentran divididos en categorías denominadas I, II y III, donde figuran las especies según su grado de amenaza. El Cardenal Amarillo se encuentra incluido en el apéndice II, lo cual implica que su comercio debe regularse.
El Cardenal Amarillo ha recibido reconocimiento en algunas provincias que aún albergan poblaciones naturales. En Entre Ríos, fue declarado monumento natural provincial en 2002 mediante el decreto 4933/02 y en General Conesa, Río Negro, es considerada ave emblemática del Valle de Conesa por declaración 07/2012.
1.4. Amenazas para el Cardenal Amarillo
Esta especie ha sufrido históricamente una fuerte presión de captura (particularmente los ejemplares macho) para abastecer el mercado ilegal de aves de jaula. Esta explotación constante llevada a cabo por más de 100 años (Collar et al. 1992), sumada a la pérdida de hábitat por su destrucción y modificación para uso ganadero, tierras de cultivo o explotación maderera, han resultado en una rápida disminución de la población silvestre en los últimos años. Según BirdLife International (2013) su población total actual rondaría entre los 1000 y 2000 individuos con una clara tendencia decreciente. En Uruguay sobrevivirían unos 300 ejemplares (Azpiroz et al. 2012) y en Brasil, la población silvestre se estima en tan solo 50 individuos (Serafini 2013). En la mayor parte de su área de distribución es una especie rara (Collar et al. 1992), y en la zona sur, se ha registrado una disminución poblacional significativa en los últimos 40 años (Pessino & Tittarelli 2006). Esto ubica a la Argentina en un lugar clave para llevar adelante un proyecto de conservación de la especie.
El comercio ilegal de especies vivas es una de las principales amenazas para la biodiversidad. Según encuestas y relevamientos en comercios realizados en las provincias de Santa Fe, La Pampa (Pessino & Tittarelli 2006) y Buenos Aires (Loydi 2008, Argibay et al. 2005), el Cardenal Amarillo es la especie más buscada por los tramperos de pájaros silvestres. Los machos reaccionan agresivamente ante la presencia de otro ejemplar del mismo sexo. Este comportamiento facilita su captura ilegal utilizando jaulas trampa con un cardenal ―llamador‖, cuya presencia atrae a los cardenales amarillos que se hallan en las cercanías. En la Argentina, un ejemplar macho puede rondar los cien dólares (Argibay et al. 2005), un precio más de diez veces superior al de la gran mayoría de las aves vendidas en el comercio ilegal, comparable al precio que se paga por algunas especies de psitácidos. Se estima que se capturan alrededor de 500 ejemplares de Cardenal Amarillo por año en Argentina (Zelaya & Bertonatti 1995).

INTRODUCCIÓN GENERAL
13
1.5. Objetivos
1.5.1. Objetivo general
El objetivo de la tesis es estudiar de las poblaciones remanentes y el grado de conexion entre estas, para poder establecer los factores que están jugando un papel fundamental en la disminución poblacional, con el objeto de poder generar un plan informado de conservacion y manejo para la especie. La necesidad inminente de tomar acciones para la conservación del Cardenal Amarillo hace que sea de particular interés estudiar sus poblaciones. Muchos individuos son decomisados por las autoridades y depositados en centros gubernamentales de recepción. La ausencia de conocimientos básicos sobre la ecología, la historia evolutiva y el comportamiento del Cardenal Amarillo no permite delinear un plan estratégico nacional para la conservación de la especie. Como consecuencia, los individuos incautados no son liberados ni tampoco incorporados a programas de conservación ex situ. 1.5.2. Objetivos particulares
Se pueden enumerar tres objetivos específicos de esta tesis:
1) Estudiar los parámetros reproductivos básicos y los factores que afectan la supervivencia de los nidos en una población de Cardenal Amarillo.
2) Estudiar la variabilidad morfológica, comportamental y genética de las principales poblaciones remanentes, con el objetivo de delimitar las unidades de manejo.
3) Analizar el efecto que tiene la hibridación con la especie emparentada Diuca diuca minor en la región sur de su distribución.
1.7. Referencias Bibliográficas
Allendorf, F. W. & G. Luikart. 2007. Conservation and the Genetics of Populations. Pp 404. Blackwell Publishing, Malden, Massachusetts, USA.
Ardern, S. L. & D. M. Lambert. 1997. Is the black robin in genetic peril? Molecular Ecology 6: 21–28.
Argibay, H., C. Dabul & V. Diaz. 2005. Actualizaciones sobre la venta de fauna silvestre en las ferias de Nueva Pompeya y Villa Domínico. Grupo Fauna Silvestre – CT / Aves Argentinas.
Azpiroz, A. B., M. Alfaro & S. Jiménez. 2012. Lista Roja de las Aves del Uruguay. Una evaluación del estado de conservación de la avifauna nacional con base en los

INTRODUCCIÓN GENERAL
14
criterios de la Unión Internacional para la Conservación de la Naturaleza. Dirección Nacional de Medio Ambiente, Montevideo.
Barker, K., K. Burns, J. Klicka, S. Lanyon & I. Lovette. 2013. Going to extremes: Contrasting rates of diversification in a recent radiation of New World Passerines Birds. Systematic Biology 62: 298–320.
Barker, K., K. Burns, J. Klicka, S. Lanyon & I. Lovette. 2015. New insights into New World biogeography: An integrated view from the phylogeny of blackbirds, cardinals, sparrows, tanagers, warblers, and allies. The Auk 132: 333–348.
Bascarán, J. L. 1987. El Cardenal Amarillo. Rev. Asociación Civil de Criadores de Especies Silvestres, Proveedores y Afines (ACCESPA) N° 6, Buenos Aires.
Bell, B. D. 1975. The rare and endangered species of the New Zealand region and the policies that exist for their management. Bulletin of the International Council for Bird Preservation 12: 165–172.
Bell, B. D. 1991. Recent avifaunal changes and the history of ornithology in New Zealand. Acta XX Congressus Internationalis Ornithologici 2: 193–230.
Betts, M. G., G. J. Forbes & A. W. Diamond. 2007. Thresholds in songbird occurrence in relation to landscape structure. Conservation Biology 21: 1046–1058.
BirdLife International. 2013. Species factsheet: Gubernatrix cristata. Downloaded from http://www.birdlife.org on 23/6/2015.
BirdLife International. 2014. One in eight of all bird species is threatened with global extinction. Presented as part of the BirdLife State of the world's birds website. Available from: http://www.birdlife.org/datazone/sowb/casestudy/106. Checked: 24/10/2015.
BirdLife International. 2015. Country profile: Argentina. Available from: http://www.birdlife.org/datazone/country/argentina. Checked: 2015-10-23.
Brown, A. D. & S. Pacheco. 2006. Propuesta de actualización del mapa ecorregional de la Argentina. En: Brown, A.; Martínez Ortiz, U.; Acerbi, M. y J. Corcuera (eds). La situación ambiental argentina 2005. Fundación Vida Silvestre Argentina, Buenos Aires.
Burns, K. J., A. J. Shultz, P. O. Title, N. A. Mason, F. K. Barker, J. Klicka, L. Scott & I. J. Lovette. 2014. Phylogenetics and diversification of tanagers (Passeriformes: Thraupidae), the largest radiation of Neotropical songbirds. Molecular phylogenetics and evolution 75: 41–77.
Cabrera, A. L. 1976. Regiones fitogeográficas Argentinas, Enciclopedia Argentina de Agricultura y Jardinería, Segunda Edición, Vol II. Buenos Aires.

INTRODUCCIÓN GENERAL
15
Campagna, L., K. Geale, P. Handford, D. A. Lijtmaer, P. L. Tubaro & S. C. Lougheed. 2011. A molecular phylogeny of the Sierra-Finches (Phrygilus, Passeriformes): extreme polyphyly in a group of Andean specialists). Molecular Phylogenetics and Evolution 61: 521–533.
Caughley, G. & A. Gunn. 1996. Conservation Biology in Theory and Practice. Blackwell Science, Cambridge, Massachusetts, U.S.A.
Ceballos, G., P. R. Ehrlich, A. D. Barnosky, A. García, R. M. Pringle & T. M. Palmer. 2015. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Science Advances 1: e1400253.
Chebez, J. C. & N. S. Morandeira. 2005. Parque Nacional El Palmar. En A. S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 168-170. Temas de Naturaleza y Conservación 5. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires.
Chebez, J. C. 2008. Los que se van. Tomo 2: Aves. Ed Albatros. Buenos Aires.
Collar, N. J., L. P. Gonzaga., N. Krabbe., A. Madroño Nieto, L. G. Naranjo, T. A. Parker & D. G. Wege. 1992. Threatened Birds of the Americas. The Internacional Council for Bird Preservation/Internacional Union for Conservation of Nature Red Data Book, Cambridge, Reino Unido.
Collar, N. J., Crosby & A. J. Stattersfield. 1994. Birds To Watch 2. The World List of Threatened Birds. Birdlife International, Cambridge, UK.
Crandall, K. A., O. R. Bininda-Emonds, G. M. Mace & R. K. Wayne. 2000. Considering evolutionary processes in conservation biology. Trends of Ecology and Evolution 15: 290–295.
De la Peña, M. R. 1981. Notas nidológicas sobre pepiteros, cardenales, etc. (Aves: Emberizidae). Primera parte. Historia Natural. 2: 1–4.
De la Peña, M. R. 2005. Lista y distribución de las aves de Santa Fe y Entre Ríos. L.O.L.A. Monografía 15: 5–137.
DeWeerdt, S. 2002. What really is an evolutionarily significant unit? The Debate over integrating genetics and ecology in conservation biology. Conservation Biology in Practice 3: 10–17.
Di Giacomo, A. G., A. S. Di Giacomo & A. Pautasso. 2005. Dorso occidental subhúmedo de Santa Fe. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 436–437. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.
Di Giacomo, A. S. 2005a. Espinal de Mercedes. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la

INTRODUCCIÓN GENERAL
16
conservación de la biodiversidad: 160. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.
Di Giacomo, A. S. 2005b. Ñandubaysal-Gualeguaychú. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 172–173. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.
Di Giacomo, A. S. (editor). 2005c. Áreas importantes para la conservación de las aves en Argentina: sitios prioritarios para la conservación de la biodiversidad. Temas de Naturaleza y Conservación 5. Aves Argentinas/AOP. Buenos Aires.
Doering, A. 1874. Noticias ornitologías de las regiones ribereñas del río Guayquiraro. Period. Zoologico 1: 237–258.
Domínguez, M., M. Ayarragaray & R. Lapido. En Prensa. Nuevos registros de ejemplares y nidos de Cardenal Amarillo (Gubernatrix cristata) en la provincia de San Luis, Argentina. Nuestras Aves.
Fahrig, L. 2003. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution and Systematics 34: 487–515.
Fraga, R. 2005. Felipe Yofre. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 161–162. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.
Frankel, O. H. 1974. Genetic conservation: our evolutionary responsibility. Genetics. 78: 53–65.
Frucella, S. & A. Bosso. 2005. Selva de Montiel. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 178–179. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.
Giraudo, A., J. Alonso., J. Herrera., M. Almiron. & D. Baldo. 2005. Región de la Estancia Oscuro. En Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 163. Temas de Naturaleza y Conservación 5. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires.
Giraudo, A. & A. S. Di Giacomo. 2005. Rincón del Socorro – Iberá. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 156–157. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.
González, P. M. 2005. San Antonio Oeste. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación

INTRODUCCIÓN GENERAL
17
de la biodiversidad: 341–343. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.
Green, D. M., 2005. Designatable units for status assessment of endangered species. Conservation Biology 19: 1813–1820.
Guyra Paraguay, 2004. Lista comentada de las aves de Paraguay. Annotated checklist of the Birds of Paraguay. Pp 117. Asunción, Paraguay.
Harris, L. D. 1984. The fragmented forest: island biogeography theory and the preservation of biotic diversity. Chicago, Illinois, Estados Unidos.
Hayes, F. E. 1995. Status, distribution and biogeography of the birds of Paraguay. American Birding Association, Colorado Springs.
Isler, M. L. & P. R. Isler. 1999. The Tanagers: natural history, distribution, and identification. Smithsonian Institution Press, Washington, Estados Unidos.
Jones, C.G., K. Swinnerton, J. Hartley & Y. Mungroo. 1999. The restoration of the free-living populations of the Mauritius kestrel, (Falco punctatus), Pink pigeon, (Columba mayeri) and Echo parakeet, (Psittacula eques). Proceedings of The Seventh World Conference on Breeding Endangered Species, Cincinnati, Ohio, 22–26 May 1999, pp. 77–86.
Lawton, J. H. & R. M. May. 1995. Extinction Rates. Oxford University Press, Oxford.
Lell, J. D. 2004. El caldenal: una visión panorámica enfatizando en su uso, en: Arturi, M. F., J. L. Frangi y J. F. Goya (eds.), Ecología y manejo de los bosques de Argentina, La Plata, Editorial de la Universidad Nacional de La Plata.
Lovegrove, T. G. 1996. A comparison of the effects of predation by Norway (Rattus norvegicus) and Polynesian rats (R. exulans) on the Saddleback (Philesturnus carunculatus). Notornis 43: 91–112.
Loydi, A. 2008. Situación actual del comercio ilegal de aves en la ciudad de Bahía Blanca. BioScriba 1: 9–6.
Maceda, J. J. 2005a. Reserva Provincial Parque Luro. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 243–244. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.
Maceda, J. J. 2005b. Victorica. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 244–245. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.
Maceda, J. J. 2005c. Jagüel del Monte. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación

INTRODUCCIÓN GENERAL
18
de la biodiversidad: 245–247. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.
Marrelli, C. A. 1918. Aves de Curuzú Cuatiá. El Hornero. I : 74–80.
Martins-Ferreira, C., M. O. Santos, O. Hadddrath, A. J. Baker & T. R. O. Freitas. 2010. Isolation and characterization of 10 microsatellite loci in the Yellow Cardinal Gubernatrix crista En. Molecular Ecology Resources 10: 751–754.
McCormack, G. & J. Künzle. 1990. Kakerori - Rarotonga's endangered flycatcher. Rarotonga: Cook Islands Conservation Service.
Merton, D. V. 1990. The Chatham Island black robin: how the world’s most endangered bird was saved from extinction. Forest and Bird. Journal of the Royal Forest & Bird Protection Society NZ Inc. 21: 14–19.
Merton, D. V. 1992. The legacy of ―Old Blue‖. New Zealand Journal of Ecology 16: 65–68.
Meyer de Schauensee, R. 1966. The species of birds of South Arrerica and their distribution. The Academy of Natural Sciences of Philadelphia, Livingston.
Moritz, C. 1994. Defining ―evolutionarily significant units‖ for conservation. Trends in Ecology & Evolution 9: 373–375.
Narosky, T. & D. Yzurieta. 2003. Guía para la identificación de las aves de Argentina y Uruguay. Asociación Ornitológica del Plata, Vázquez-Mazzini Ed., Buenos Aires, Argentina.
Norris, K. & D. J. Pain. 2002. Conserving Bird Biodiversity: General Principles and Their Application. Cambridge University Press, Cambridge, UK.
Olrog, C. C. 1979. Nueva lista de avifauna argentina. Pp 1–324. Opera Lilloana 27. Fundación Miguel Lillo, Tucuman, Argentina.
Ortiz, D. 2008. Distribución histórica y actual del Cardenal Amarillo (Gubernatrix cristata) en el Litoral Fluvial Argentino. Pp 121–126. En: Aceñolaza F.G. (editores). Temas de la biodiversidad del litoral fluvial argentino III. INSUGEO, San Miguel de Tucumán.
Palsbøll, P. J., M. Bérubé & F. W. Allendorf. 2006. Identification of management units using population genetic data. Trends in Ecology & Evolution 22: 11–1.
Pautasso, A. A. 2002. Nuevos registros para aves poco citadas en Santa Fe, Argentina. Nuestras Aves 43: 19–21.
Perrins, C. 1987. Collins New Generation Guide to the Birds of Britain and Europe. London: Collins.

INTRODUCCIÓN GENERAL
19
Pessino, M. & R. F. Tittarelli. 2006. The Yellow Cardinal (Gubernatrix cristata): a diagnosis of its situation in the province of La Pampa, Argentina. Gestion Ambiental 12: 69–76.
Ridgely, R. S. & G. Tudor. 1989. The birds of the South America. Volume II.The Suboscine Passerines. Univ.of Texas Press, Austin, Texas.
Ridgely, R. S. & G. Tudor. 1997. The birds of South America. Volume I, The Oscine Passerines. University of Texas Press, Austin.
Ryder, O. A. 1986. Species conservation and systematics: the dilemma of subspecies. Trends in Ecology & Evolution 1: 9–10.
Serafini, P. P. 2013. Plano de Ação Nacional para a conservação dos passeriformes ameaçados dos campos sulinos e espinilho. Instituto Chico Mendes de Conservação da Biodiversidade, ICMBIO, Brasília.
Sibley, C. G. & J. E. Ahlquist. 1990. Phylogeny and Classification of Birds: A Study in Molecular Evolution. Yale University Press, New Haven, Connecticut.
Sibley, C. G. & B. L. Jr. Monroe. 1990. Distribution and Taxonomy of the Birds of the World. Yale University Press, New Haven, Connecticut.
Sick, H. 1997. Ornitologia Brasileira. Nova Fronteira, Rio de Janeiro, Brasil.
Sosa, H., S. Martin & A. Zarco. 2011. Nuevos registros del Cardenal Amarillo (Gubernatrix cristata) en la provincia de Mendoza, Argentina. BioScriba 4(1): 21–25.
Sutherland, W. J., I. Newton & R. E. Green 2004. Bird Ecology and Conservation. A Handbook of techniques. Oxford University Press, Nueva York.
Tisdall, D. J. & D. V. Merton. 1988. Disease surveillance in the Chatham Islands black robin. Surveillance 15, Journal of MAFQUAL, New Zealand Ministry of Agriculture.
Tittarelli, F. 2005. Parque Nacional Lihue Calel. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 247–248. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.
Veiga, J. O. 2005. Ceibas. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 177. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.
Veiga, J. O & F. Tittarelli. 2005. Bañados del Río Atuel. En A.S. Di Giacomo (editor), Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 248–249. Temas de Naturaleza y Conservación 5. Aves Argentinas / Asociación Ornitológica del Plata, Buenos Aires.

INTRODUCCIÓN GENERAL
20
Vitousek, P. M., H. A. Mooney, J. Lubchenko & J. M. Melillo. 1997. Human domination of Earth‘s ecosystems. Science 277: 494–499.
Wallace, M. 2005. Re-introduction of the California Condor to Baja California, Mexico. Re-introduction News 24: 27–28.
Wilbur, S. R. 1978. Supplemental feeding of California Condors. Pp. 135–140. En Endangered birds. Management techniques for preserving threatened species, ed. S.A. Temple, Wisconsin: University of Wisconsin Press.
Zelaya, D. & C. Bertonatti. 1995. Nuestro Libro Rojo: Cardenal Amarillo. Vida Silvestre 43: 21–22.

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
21
CAPÍTULO 2
BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
Domínguez, M., J.C. Reboreda & B. Mahler. 2014. Impact of Shiny Cowbird and botfly parasitism on the reproductive success of the globally endangered Yellow Cardinal Gubernatrix cristata. Bird Conservation International 25: 294-305. (DOI: 10.1017/S095927091400015X)
2.1. Introducción
Uno de los principales objetivos para el manejo de especies amenazadas es mantener o aumentar su éxito de supervivencia y su tasa de reproducción (Gosling & Sutherland 2004). Para ello, es fundamental conocer los parámetros reproductivos y los factores que pueden afectarlos (Sutherland 2000). El desconocimiento de datos básicos de la biología de las especies amenazadas limita los esfuerzos para protegerlas.
Para el Cardenal Amarillo hay muy poca información acerca de los principales parámetros reproductivos, tales como la duración de la época reproductiva, el tamaño de puesta, el éxito de eclosión y la supervivencia de pichones. La única información disponible proviene de De la Peña (1981, 1987), que describe las características del nido y de los huevos de esta especie. Además, hay un único reporte antiguo que identifica al Cardenal Amarillo como especie hospedadora del Tordo Renegrido, Molothrus bonariensis (Friedmann 1934), aunque hasta el momento no ha sido evaluado el impacto que el parasitismo de cría genera sobre el éxito reproductivo de los cardenales.
El parasitismo del Tordo Renegrido podría potencialmente ser una amenaza para los cardenales amarillos ya que el Tordo Renegrido está presente en la totalidad del área de distribución del Cardenal Amarillo y los tordos han aumentado su población como resultado de la transformación del hábitat y de las prácticas agrícolas (por ejemplo, la creación de feedlots). Para varias especies hospedadoras se ha mostrado que el parasitismo del Tordo Renegrido genera un costo reproductivo, ya que las hembras de esta especie parásita suelen perforar los huevos del hospedador cuando visitan los nidos (Massoni y Reboreda 2002, Peer 2006), disminuyendo el tamaño de la puesta y aumentando la probabilidad de abandono del nido (Massoni & Reboreda 1998, Astié & Reboreda 2006). Asimismo, el parasitismo puede disminuir el éxito de eclosión y la supervivencia de los pichones del hospedador por la competencia generada en el nido (Fraga 1978, Astié & Reboreda 2006, Tuero et al. 2007).
Por otro lado, se ha reportado que las larvas de mosca del género Philornis (Orden Diptera) pueden ser una grave amenaza para las especies de aves (Fessl & Tebbich 2002).

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
22
Estas larvas se alojan en el tejido subcutáneo de los pichones y se alimentan de fluidos serosos, de restos de tejido y de células sanguíneas (Uhazy & Arendt 1986, Texeira 1999) reduciendo el crecimiento y la supervivencia de estos pichones (Dudaniec & Kleindorfer 2006, Rabuffetti & Reboreda 2007, Quiroga & Reboreda 2012). Aunque no hay registros anteriores de parasitismo de las larvas de esta mosca en cardenales amarillos, su presencia se ha descrito para otros paseriformes en el área de distribución del cardenal (Antoniazzi et al. 2010, Quiroga et al. 2012).
El objetivo general de este capítulo es aportar nuevos datos sobre la biología reproductiva del Cardenal Amarillo. Los objetivos particulares son: 1) estimar los principales parámetros reproductivos, 2) examinar el efecto de factores temporales (fecha de inicio del nido, estadío del nido) sobre la supervivencia de nidos, y 3) investigar si el parasitismo por parte del Tordo Renegrido y por larvas de moscas del género Philornis afectan negativamente el éxito reproductivo del Cardenal Amarillo.
2.2. Métodos
2.2.1. Sitio de estudio
El estudio se realizó en dos sitios de la provincia de Corrientes (Figura 2.1) durante las temporadas reproductivas (principios de octubre - finales de diciembre) de 2011 y 2012.
Figura 2.1. Mapa que muestra los sitios de estudio en la Provincia de Corrientes: 1 = Reserva Rincón del Socorro, 2 = Estancia La Paz.
2
1

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
23
El primer sitio se encuentra en la 'Reserva Rincón del Socorro' (28°32'S, 57°10'O) al margen de los esteros del Iberá. La reserva es una mezcla de pastizales abiertos inundados temporalmente, sabanas arboladas, bosques hidrófilos y humedales. El segundo sitio se encuentra a unos 100 km del primero, en la 'Estancia La Paz' (29°20'S, 58°26'O), cerca de la ciudad de Mercedes. Este sitio se caracteriza principalmente por la presencia pastizales abiertos con parches de bosque caducifolio espinoso dominado por Prosopis affinis (Figura 2.2). Mientras que en el primer sitio no se realizan actividades productivas, en el segundo se practica ganadería extensiva.
Figura 2.2. A la izquierda se muestra el ambiente de la 'Reserva Rincón del Socorro', a la derecha de la 'Estancia La Paz'.
2.2.2. Toma de datos
Durante las temporadas reproductivas 2011 y 2012, se capturaron 16 machos y 14 hembras mediante redes de niebla. Los individuos se pesaron con una balanza de resorte de 60 g marca Pesola AG (error ± 0.5 g) y se anillaron en el tarso con un anillo de aluminio numerado (Porzna Ltd.) y dos anillos de plástico con una combinación única de colores (A. C. Hughes Ltd., Middlesex, UK). Esto facilitó la identificación de los individuos para el seguimiento de las parejas y sus intentos reproductivos.
Se realizaron búsquedas de nidos de Cardenal Amarillo en los territorios reproductivos identificados en cada temporada en las áreas de sabana arbolada dominados por árboles bajos (< 8 m de altura) de los géneros Prosopis y Acacia (Tressens et al. 2002). Los nidos fueron encontrados mediante una búsqueda sistemática de potenciales sitios de nidificación y observando el comportamiento de las parejas territoriales (por ejemplo, el acarreo de material para construir el nido o de comida para alimentar a los pichones). Para las observaciones se siguieron las técnicas estandarizadas en Martin & Geupel (1993) que minimizan las molestias ocasionadas a la pareja reproductora. La localización del territorio de cada pareja se realizó de forma auditiva o por observación directa de los adultos. Una

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
24
vez localizado el territorio, los nidos fueron buscados entre las ramas de los árboles. En los nidos de difícil acceso se utilizó una vara de 3 m de largo con un espejo a 45º en la punta para la inspección del contenido del nido. Para todos los nidos se determinó el estadío en el que se encontraba al momento de la detección (construcción, huevos o pichones). Durante el período de estudio, se encontraron un total de 46 nidos: 21 correspondientes a 16 parejas seguidas durante el año 2011, y 25 nidos correspondientes a 18 parejas seguidas durante 2012. Se encontraron 13 nidos (28%) durante la etapa de construcción del nido, 5 (11%) durante la puesta, 18 (39%) durante la incubación, y 8 (22%) después de la eclosión de los huevos.
Una vez detectados, los nidos se visitaron cada 1-4 días hasta que los pichones abandonaran el nido o el nido hubiera fracasado. Durante cada visita, se registró el número de huevos o pichones, la presencia de picaduras por hembras de Tordo Renegrido en el caso de huevos y la presencia de larvas de mosca en los pichones. Además, se removió con cuidado el contenido completo del nido (huevos y/o pichones) para realizar las mediciones correspondientes, lo cual llevó unos 10-15 minutos, dependiendo de la accesibilidad del nido y el número de huevos o pichones presentes. Los huevos fueron removidos una única vez donde fueron identificados con tinta resistente al agua (con un número si se lograba identificar el orden de puesta del huevo, o con letras si el nido ya se encontraba en incubación; Figura 2.3) y se midieron su largo y ancho con un calibre de reloj marca Measy2000 (error ± 0.1 mm) y su peso con una balanza de resorte de 10 g marca Pesola AG (error ± 0.1 g).
Figura 2.3. Nido de Cardenal Amarillo con 3 huevos marcados con tinta indeleble para su seguimiento individual.

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
25
Después de la eclosión, los pichones fueron marcados en el tarso con tinta resistente al agua y pesados con una balanza de resorte de 30 g marca Pesola AG (error ± 0.2 g). En cada visita, los pichones fueron inspeccionados cuidadosamente para detectar la presencia subcutánea de larvas del género Philornis. Los pichones fueron anillados a los 8-10 días de edad (Figura 2.4) y posteriormente no fueron manipulados para evitar el abandono prematuro del nido.
Figura 2.4. Pichón de Cardenal Amarillo marcado con dos anillos de plástico coloreados y un anillo de aluminio para su identificación individual.
2.2.3. Análisis de datos
El tamaño de puesta fue estimado a partir de una muestra de nidos encontrados durante la construcción o en puesta y que llegaron a completar la puesta (n = 17). Una puesta fue considerada completa cuando el número de huevos no se modificó entre dos visitas consecutivas. Para calcular los promedios poblacionales del largo y ancho de los huevos, primero se calculó un promedio por puesta, dado que estos huevos no son independientes. Se estimó el período de incubación como el tiempo transcurrido entre la puesta del último huevo y la eclosión del último pichón, en aquellos nidos donde todos los huevos eclosionaron. La frecuencia de parasitismo fue estimada como la proporción de

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
26
nidos parasitados con huevos de tordos renegridos y la intensidad de parasitismo se estimó como el número promedio de huevos de tordo en nidos parasitados. Para esta estimación consideramos nidos encontrados durante la construcción, puesta e incubación (n = 33) considerando que el parasitismo puede ocurrir hasta los primeros días de la incubación (Briskie & Sealy 1990, Kattan 1995, Fiorini et al. 2009). Se estimó la prevalencia de parasitismo por parte de moscas el género Philornis como la proporción de nidos en los que los pichones fueron infestados con larvas de mosca. Para esta estimación se consideraron únicamente los nidos en los que los pichones sobrevivieron al menos hasta los 5 días de edad (n = 18), ya que la mayoría de los eventos de parasitismo por larvas ocurren en la etapa temprana de pichones, cuando presentan la piel más descubierta (Rabuffetti & Reboreda 2007, Segura & Reboreda 2011). Se estimó el éxito de eclosión como el número de pichones eclosionados dividido el número de huevos al momento de la eclosión. La supervivencia de los pichones se estimó como la proporción de pichones eclosionados que lograron abandonar exitosamente el nido. Para estimar el éxito de eclosión y la supervivencia de los pichones sólo se consideraron los nidos encontrados durante la etapa de construcción, puesta o incubación en los que los pichones eclosionaron o abandonaron el nido exitosamente, respectivamente. Consideramos que un nido había sido abandonado si los huevos o pichones estaban fríos y ninguna actividad parental era observada cerca del nido durante la visita; consideramos que había sido predado si el contenido del mismo desaparecía entre visitas consecutivas y había una ausencia de actividad de los padres cerca del nido.
2.2.4. Análisis estadístico
Las tasas de supervivencia diarias de los nidos (TSD) fueron modeladas con el programa MARK 6.2 (White & Burnham 1999) el cual incorpora los efectos hipotéticos de las variables que puedan afectar a la supervivencia de nidos y evalúa el apoyo de cada modelo utilizando el enfoque de teoría de la información (Burnham & Anderson 2002). Se estimó la TSD en la etapa de los huevos (puesta e incubación) y en la etapa de pichones (después de la eclosión del primer pichón). Para nidos observados durante las dos etapas, el período de observación fue truncado en el último día de la primera etapa (ver abajo) e iniciado en ese día para la etapa siguiente (Dinsmore & Dinsmore 2007). El período de observación se truncó en la fecha de la eclosión del primer pichón del hospedador (etapa de huevos), y en la fecha de abandono de los volantones (etapa de pichones) para los nidos que completaron con éxito cada etapa. Cuando la fecha exacta de abandono del nido por parte de los pichones era desconocida, el periodo de observación se truncó en la edad promedio en la que los pichones abandonan exitosamente el nido (14 días, ver más abajo). Un supuesto para estimar la TSD en MARK es que los destinos de nidos son independientes (Dinsmore et al. 2002). Por este motivo, se excluyeron del análisis re-intentos de nidificación de las mismas parejas.

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
27
Estimamos la TSD utilizando las condiciones estándar de MARK 6.2, i.e. el seno de la constante del modelo y la función de enlace logit para aquellos modelos que incorporaron covariables (Dinsmore et al. 2002). La TSD para la etapa de huevos fue estimada con 20 intentos de nidificación de los que teníamos información para todas las covariables involucradas en el análisis. De esta muestra, se pudo determinar la fecha de inicio de puesta para 9 nidos encontrados durante la construcción y puesta de huevos. Para nidos encontrados durante la incubación y que sobrevivieron hasta la eclosión (n = 6), estimamos la fecha de inicio de puesta retrotrayendo desde la fecha de eclosión. Para nidos que fueron encontrados y que fallaron durante la incubación (n = 5), se estimó la fecha de inicio de puesta asignando el día en que fue encontrado a la mitad de la incubación (día 7).
A priori se determinaron dos covariables que podrían influir en la supervivencia de los nidos en la etapa de huevos: momento de la temporada en el que se realizó el intento de nidificación (tomando como día 1 el 1ero de octubre) y parasitismo por el Tordo Renegrido. Primero se estimó el modelo general que asume tasa de supervivencia diaria constante en el tiempo y entre los nidos (S(.)); después se construyeron modelos que permiten que la TSD varíe incorporando de a una covariable por vez (momento de la temporada o parasitismo). También se corrió un modelo aditivo incluyendo ambas variables.
Para analizar los factores que influyen en la supervivencia de los nidos durante la etapa de pichones se incluyeron las covariables momento de la temporada y parasitismo por Philornis. No se incluyó la covariable parasitismo de cría porque los pichones de Tordo Renegrido sólo eclosionaron en tres nidos. Para este análisis, se utilizó un subconjunto de 20 intentos de nidificación independientes (pertenecientes a distintas parejas) que alcanzaron la etapa de pichones y de la que se tenía información sobre el parasitismo por Philornis. Se compararon los modelos basándose en el Criterio de Información de Akaike (AIC) corregido para pequeños tamaños de muestras (AICc, Burnham & Anderson 2002). Consideramos el modelo con el valor más bajo de AICc como el mejor ajuste a los datos. Los valores reportados son medias ± ES.
Se obtuvieron estimaciones de la tasa de supervivencia global de los nidos (probabilidad de que el nido sobreviva hasta producir al menos 1 volantón) a partir de los modelos con mayor soporte para cada etapa. La probabilidad de supervivencia de toda la etapa de huevos (puesta + incubación), se obtuvo multiplicando los valores de TSD de los días 1 a 15 días (longitud máxima de esta etapa: tamaño modal de nidada = 3 huevos, inicio de la incubación con la puesta del penúltimo huevo y 13 días de incubación) bajo el modelo que contempla los efectos del parasitismo y del momento de la temporada. Para la etapa de pichones, debido a que el modelo nulo fue el mejor y se supone que la TSD no varía con el tiempo, elevamos la TSD a la 14, que es el número de días que los pichones permanecen en el nido basado en nuestras observaciones de campo (ver Resultados).

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
28
La diferencia entre sexos en la masa corporal se analizó con la prueba t de Student. Las diferencias en el éxito de eclosión y la supervivencia de pichones entre nidos parasitados y no parasitados se analizaron con la prueba U de Mann-Whitney. Estas pruebas estadísticas se realizaron con el programa STATISTICA 7.0 (Statsoft Inc. 2004) considerando un valor de alfa de 0.05.
2.3. Resultados
2.3.1. Construcción del nido, puesta e incubación de huevos
Los primeros intentos de nidificación fueron observados en la primera mitad del mes de octubre y los últimos a mediados de diciembre. La distribución temporal de los intentos de nidificación muestra una forma unimodal y el pico de nidificación (31% de los nidos) ocurrió durante la segunda quincena de noviembre (Figura 2.5). A partir del seguimiento de las parejas anilladas, se pudo determinar que en algunos casos una misma pareja realizó más de un intento de nidificación durante la temporada reproductiva. Se observaron re-intentos de nidificación en 5/16 parejas durante 2011 y en 6/18 parejas en 2012 (9 parejas con un re-intento de nidificación, una con dos, y otra pareja con tres re-intentos de nidificación). El tiempo transcurrido desde la fecha de fracaso del nido hasta el inicio de una nidada de reemplazo fue de 12.4 ± 1.3 días (n = 14, rango 7-26 días). Un 35% de las parejas tuvo re-intentos de nidificación luego de un fracaso pero no hubo segundas nidadas luego de un intento exitoso.
. Figure 2.5. Porcentaje de nidos de Cardenal Amarillo iniciados a lo largo de la temporada reproductiva, sobre la base de 46 nidos muestreados durante 2011 y 2012 en la Reserva Rincón del Socorro y la Estancia La Paz, Provincia de Corrientes, Argentina. La parte gris de la barra indica los primeros intentos de nidificación mientras que la parte blanca indica re-intentos de nidificación.

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
29
El nido tiene forma de copa semi-esférica de 15.5 ± 0.65 cm de diámetro exterior, 10.5 ± 0.46 cm de altura, 8.3 ± 0.27cm de diámetro interno y 5.9 ± 0.36 cm de profundidad (n = 25 nidos). Están construidos con una capa externa de ramas con espinas y líquenes y una capa interna de ramas más finas, pelo de caballo, material vegetal compactado, pasto, semillas e hilos de nylon. La altura media sobre el nivel del suelo fue de 2.1 ± 0.11 m (n = 34 nidos, rango 1.1-4.1 m). Los nidos fueron construidos principalmente en Prosopis affinis (31/46) y Acacia cavens (6/46).
Los huevos son de un color verde azulado con manchas negras (Figura 2.3). En la mayoría de los huevos las manchas se distribuyeron por toda la superficie, pero en algunos las manchas se mostraron concentradas alrededor del polo mayor del huevo. El tamaño medio de los huevos fue de 24.8 ± 0.28 mm de longitud (n = 17 nidadas), 17.9 ± 0.12 mm de ancho (n = 17 nidadas) y una masa de 3.3 ± 0.16 g (n = 12 nidadas). El tamaño modal de nidada fue de 3 ± 0.12 (rango 2-4, n = 17) y no difirió a lo largo de la temporada
reproductiva (Spearman = -0.22, P = 0.18). La incubación fue realizada exclusivamente por la hembra y duró 12.5 ± 1.7 días (n = 13). No observamos pérdida parcial de huevos durante la incubación en nidos no parasitados (n = 21).
2.3.2. Eclosión y crecimiento de pichones
La eclosión fue asincrónica y en la mayoría de los casos los dos primeros huevos (en nidadas de tres) eclosionaron de forma sincrónica mientras que el último huevo eclosionó un día después, aunque también se observaron casos en los que los tres huevos eclosionaron en días consecutivos. Pichones recientemente eclosionados tenían la piel de color naranja con plumón gris en la cabeza y espalda. La boca era roja con el borde de color amarillo claro (Figura 2.4). La masa corporal de los pichones al eclosionar fue de 3 ± 0.25 g (n = 6 nidos y 14 pichones). Ambos padres alimentaron a las crías y los pichones permanecieron en el nido durante 14 días.
Se observó una reducción de nidada en el 85% (6/7) de los nidos que fueron exitosos (produjeron al menos un volantón). En promedio, los nidos exitosos produjeron 1.6 ± 0.2 volantones y la supervivencia de pichones fue de 0.67 ± 0.10 (n = 7).
2.3.3. Distribución de las parejas
Se encontraron 21 parejas de Cardenal Amarillo entre las dos campañas (10 en Estancia Rincón del Socorro y 11 en Estancia La Paz, Figura 2.6). En 2011 se encontraron 12 parejas de las cuales 10 se volvieron a encontrar la temporada siguiente en el mismo territorio. Las restantes parejas no fueron reavistadas.

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
30
Figura 2.6. A la izquierda se muestra la disposición de los territorios de las parejas en la Estancia Rincón del Socorro y a la derecha en la Estancia La Paz. En amarillo se muestran las parejas encontradas por primera vez durante la temporada reproductiva de 2011 y en rojo las parejas encontradas por primera vez en 2012.
En la zona de la Reserva Rincón del Socorro se encontró una densidad de 7 parejas reproductoras en 2.10 km2 con una distancia mínima entre territorios de 240 m y una distancia promedio de 1079 m (año 2012). Sólo una pareja fue exitosa (llegó a producir volantones que abandonaron el nido) en las dos temporadas que duró este estudio. En la Estancia La Paz se encontró una densidad de 11 parejas en 3.74 km2 con una distancia mínima entre territorios de 200 m y una distancia promedio de 612 m (año 2012). En la temporada reproductiva de 2011 cinco parejas fueron exitosas y en 2012 cuatro.
En general, el tamaño promedio del territorio de un cardenal fue de 2.16 ± 0.88 ha (media ± DS). No se detectaron diferencias sexuales en la masa corporal de los cardenales adultos (machos: 47.3 ± 1.5 g, rango de 42 a 62 g, n = 15; hembras: 47.4 ± 0.9, rango de 39.5 a 53 g, n = 14, t = 2.0; p = 0.97).
2.3.4. Efectos del parasitismo
Si bien la causa mas frecuente de fracaso fue la predación, encontramos otros factores que provocaron el fracaso de los nidos. La frecuencia de parasitismo del Tordo Renegrido fue del 33% (11/33 nidos) y la intensidad de parasitismo fue de 1.09 ± 0.09 huevos por nido parasitado (rango 1 - 2, n = 11). El tamaño de los huevos parásitos fue de 25.8 ± 0.30 mm de longitud, 21.0 ± 0.12 mm de ancho y un peso de 5.3 ± 0.10 g (n = 5; Figura 2.7). Las picaduras de huevos en nidos parasitados redujeron el tamaño de nidada de 3.0 ± 0.19 a 1.0 ± 0.45 huevos (prueba de Wilcoxon: Z = 2.4, p = 0.02, n = 11) y, como resultado de estas picaduras, fueron abandonados el 54% de los nidos parasitados. El éxito de eclosión no difirió entre los nidos no parasitados y parasitados (no parasitados: 0.77 ± 0.08, n = 13; parasitados: 0.68 ± 0.11, n = 5; prueba de Mann Whitney, z = 0.74; p = 0.46). Debido a que todos los nidos en los que los pichones de Tordo Renegrido eclosionaron fueron predados (n = 3), no se pudo evaluar si la presencia de un pichón parásito reduce por competencia la supervivencia de los pichones de Cardenal Amarillo.

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
31
Figura 2.7. Parasitismo de Tordo Renegrido (fondo blanco y manchas marrones) en un nido de Cardenal Amarillo (huevos celestes con manchas negras).
La prevalencia del parasitismo de larvas de mosca del género Philornis (Figura 2.8) fue del 22% (4/18 nidos). El primer nido con larvas fue encontrado el 4 de noviembre y el último el 18 de noviembre. Sólo 2 de los 4 nidos parasitados con larvas fueron exitosos (produjeron volantones). La supervivencia de los pichones en nidos con larvas de moscas del género Philornis fue de 0.25 ± 0.32 (n = 4), mientras que en los nidos sin larvas fue de 0.78 ± 0.11 (n = 5) (prueba de U de Mann_Whitney z = 1.84; p = 0.07).
Figura 2.8. Pichones de Cardenal Amarillo con larvas de mosca del género Philornis.
2.3.5. Supervivencia de nidos
El mejor modelo para explicar la supervivencia de los nidos durante la etapa de huevos incorpora los efectos aditivos de momento de la temporada y parasitismo de cría (Tabla 1). Ambas variables mostraron pendientes negativas (βTiempo = -0.074 ± 0.03 y βParasitism = -1.45 ± 0.77) indicando una disminución de la supervivencia de los nidos en relación con el avance de la temporada reproductiva y el parasitismo del Tordo Renegrido (Figura 2.9).

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
32
Con respecto a la supervivencia de nidos durante la etapa de pichones, los modelos que incluyeron los efectos del momento de la temporada y el parasitismo por Philornis no recibieron apoyo; el mejor modelo para explicar los datos fue el que incluye el intercepto y asume una TSD constante (β = 2.98, intervalo de confianza menor = 2.40; intervalo de confianza superior = 3.56).
Figure 2.9. Tasas de supervivencia diarias (TSD) durante la etapa de los huevos (puesta e incubación) de los nidos no parasitados de Cardenal Amarillo (círculos blancos) y aquellos parasitados por el Tordo Renegrido (círculos negros) en función del momento de la temporada (día de la temporada reproductiva en el que el nido comenzó, 1 = 1ero de octubre). Las líneas por arriba y debajo de los círculos indican el error estándar de la DSR (línea discontinua = no parasitados, continua = parasitados).
La probabilidad acumulada de sobrevivir a lo largo del ciclo de nidificación de un nido no parasitado en el comienzo de la temporada reproductiva (1 de octubre) fue 0.497, mientras que la probabilidad de sobrevivir todo el ciclo de nidificación de un nido parasitado iniciado en el momento en que ocurren la mayoría de los intentos de nidificación (20 de noviembre) fue de 0.214.
2.4. Discusión
Este estudio proporciona la primera descripción completa de los principales parámetros reproductivos y factores que influyen en el éxito reproductivo del Cardenal Amarillo. Los resultados muestran que en la provincia de Corrientes la supervivencia de los nidos disminuye a medida que avanza la temporada reproductiva y como resultado del

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
33
parasitismo de cría por el Tordo Renegrido. Además, se reporta por primera vez el parasitismo por larvas de moscas del género Philornis en pichones de cardenales amarillos y su impacto sobre su supervivencia. Estos dos factores no se habían considerado previamente como posibles amenazas para esta especie.
Los Tordos Renegridos son parásitos de cría extremadamente generalistas que utilizan más de 260 hospedadores (Lowther 2013). Son frecuentes en toda América del Sur donde han ampliado su distribución como resultado de la transformación del hábitat (Ortega 1998). Se han expandido hacia el Caribe durante el siglo pasado (Cruz et al. 1989) y en la actualidad se están expandiendo hacia América del Norte (Post et al. 1993, Kluza 1998, Post & Sykes 2011). Hay varios informes de esta especie parasitando paseriformes amenazados como el Saltón Cabecipálido, Atlapetes pallidiceps, considerado en peligro crítico (Oppel et al. 2004), la Mariquita de Puerto Rico, Agelaius xanthomus, considerada en peligro (Wiley et al. 1991, López Ortiz et al. 2002) y el Tordo Amarillo, Xanthopsar flavus (Fraga et al. 1998), categorizado en estado vulnerable. El principal impacto del Tordo Renegrido cuando parasita hospedadores similares o mayores en tamaño es la picadura de los huevos del hospedador (Reboreda et al. 2003). Encontramos evidencias de picaduras en huevos de cardenal en numerosas ocasiones que resultaron en una reducción de nidada y muy a menudo en el abandono del nido. Esto llevó a una menor tasa de supervivencia en los nidos parasitados. No se pudo probar si la competencia con los pichones de Tordo Renegrido genera un efecto negativo sobre la supervivencia de los pichones de Cardenal Amarillo, ya que todos los nidos parasitados fracasaron por predación a los pocos días de la eclosión. El parasitismo por parte de Tordos Renegridos también se ha reportado en la Diuca Común, Diuca diuca, especie estrechamente emparentada con el Cardenal Amarillo, y de manera similar a nuestros resultados, el principal impacto de parasitismo en este hospedador fue la picadura de los huevos provocando luego el abandono del nido (Marin 2011).
De Mársico et al. (2010) demostraron que los Tordos Renegridos exhiben ciertas preferencias por algunas especies dentro de las comunidades de hospedadores, y sólo parasitan con una alta frecuencia (> 50%) una pequeña fracción de los hospedadores disponibles, mientras que no parasitan o parasitan con frecuencias muy bajas (< 25%) a gran parte de los hospedadores disponibles. En nuestro sitio de estudio la frecuencia de parasitismo de los Cardenales Amarillos fue del 33%. Debido a que no tenemos datos sobre frecuencias de parasitismo en otros hospedadores en esta zona, no podemos determinar si esta frecuencia intermedia de parasitismo fue el resultado de una baja densidad de tordos o porque existe una preferencia por otros hospedadores disponibles.
A diferencia de los cardenales comunes, Paroaria coronata, que expulsan los huevos de Tordo Renegrido (Segura & Reboreda 2012), los cardenales amarillos no parecen haber evolucionado defensas antiparasitarias (Rothstein 1990, Krüger 2007). No se

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
34
detectaron evidencias de comportamiento de rechazo de huevos parásitos o de abandono del nido inmediatamente después del evento de parasitismo, aunque los huevos de tordo difieran considerablemente en el color de fondo y en el patrón de manchas de los huevos de los cardenales (Figura 2.7.). Todos los casos de abandono en nidos parasitados se produjeron luego de que la mayoría de los huevos de los cardenales fueran perforados y, por lo tanto, la deserción del nido no puede considerarse una defensa antiparasitaria.
En cuanto a la menor supervivencia de los nidos a medida que avanza la temporada reproductiva, esta tendencia también se ha reportado para especies de aves de pastizal de América del Norte (Grant et al. 2005) y para el yetapá de collar, Alectrurus risora, en el noreste de Argentina (Di Giacomo et al. 2011). Grant et al. (2005) propusieron que esta disminución es el resultado de un aumento en la abundancia local de los depredadores. Sin embargo, debido a que no disponemos de datos sobre la variación estacional de los depredadores no podemos probar esta hipótesis.
Este estudio también muestra que los cardenales amarillos son parasitados por larvas de mosca del género Philornis. La mayoría de los estudios sobre el parasitismo de larvas de moscas muestran que éste reduce considerablemente la supervivencia de los pichones (Dudaniec & Kleindorfer 2006). Se encontró que la prevalencia del parasitismo fue del 22%. En un estudio que también se llevó a cabo en la región del Espinal, en la provincia de Santa Fe, Antoniazzi et al. (2010) encontraron que sólo 7/33 especies de paseriformes tienen una prevalencia de parasitismo de larvas de moscas superior al 20%, lo que indica que el cardenal amarillo sería un huésped preferido de estas moscas. Aunque el número de nidos parasitados con larvas fue bajo, nuestros resultados sugieren que este tipo de parasitismo reduce la supervivencia de los pichones de Cardenal Amarillo y por lo tanto afecta negativamente el éxito reproductivo de esta especie. Como resultado del parasitismo por larvas, y al igual de lo observado para otras especies, el nido es abandonado después de que todos los pichones mueren (Rabuffetti & Reboreda 2007, Segura & Reboreda 2012). Debido a que esto ocurre en una fase avanzada del ciclo de nidificación, luego de que los padres invertieron considerablemente en la actual camada, parecería menos probable que en estos casos trataran de re-nidificar. Los registros más australes de moscas de este género en Argentina están a 35°S (Fraga 1984, Reboreda & Rabuffetti 2007, Segura & Reboreda 2011). Debido a que las principales poblaciones actuales de cardenales amarillos al sur de su distribución se encuentran a latitudes más altas, es probable que no se vean afectadas por el parasitismo de estas larvas. Sin embargo, son necesarios más estudios para estimar mejor el impacto que generan el parasitismo por Tordos Renegridos y por larvas del género Philornis sobre la viabilidad de las poblaciones del Cardenal Amarillo.
En conclusión, se encontró que en la mayoría de los casos, el tamaño de puesta del Cardenal Amarillo fue de tres huevos y no varió a lo largo de la temporada. Asimismo, se encontró que aproximadamente el 35% de las parejas re-nidificó luego de un fracaso,

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
35
observándose hasta cuatro intentos de nidificación durante la temporada reproductiva. Este patrón parece ser relativamente frecuente en aves neotropicales, que tienen altas tasas de depredación de nidos (Martin 1996, Di Giacomo et al. 2011, Segura & Reboreda 2012). La capacidad de re-nidificar varias veces tras el fracaso del nido aumentaría la posibilidad de que una pareja de cardenales amarillos reclute juveniles durante la temporada reproductiva.
2.5. Referencias bibliográficas
Antoniazzi, L. R., D. E. Manzoli, D. Rohrmann, M. J. Saravia, L. Silvestri & P. M. Beldomenico. 2010. Climate variability affects the impact of parasitic flies on Argentinean forest birds. Journal of Zoology 283: 126–134.
Astié, A. A. & J. C. Reboreda. 2006. Costs of egg punctures and Shiny Cowbird parasitism on Creamy-bellied Thrush reproductive success. Auk 123: 23–32.
Briskie, J. V. & S. G. Sealy. 1990. Evolution of short incubation periods in the parasitic cowbirds, Molothrus spp. Auk 107: 789–794.
Burnham, K. P. & D. R. Anderson. 2002. Model selection and multimodel inference: a practical information-theoretic approach. New York: Springer-Verlag.
Cruz, A., J. W. Wiley, T. K. Nakamura & W. Post. 1989. The Shiny Cowbird Molothrus bonariensis in the West Indian region - biogeography and ecological implications. Pp. 519-540. En: C A Wood, ed. Biogeography of the West Indies. Florida: Sandhill Crane Press Gainesville.
De la Peña, M. R. 1981. Notas nidológicas sobre pepiteros, cardenales, etc. (Aves, Emberizidae). Primera Parte. Historia Natural 2: 1–4.
De la Peña, M. R. 1987. Nidos y huevos de aves argentinas. Santa Fe, ed. author.
De Mársico, M. C., B. Mahler, M. Chomnalez, A. G. Di Giacomo & J. C. Reboreda. 2010. Host use by generalist and specialist brood parasitic cowbirds at population and individual levels. Advances in the Study of Behavior 42: 83–121.
Di Giacomo, A. S., Di Giacomo, A. G.and Reboreda, J. C. 2011. Male and female reproductive success in a threatened polygynous species: the Strange-tailed Tyrant, Alectrurus risora. Condor 113: 619–628.
Dinsmore, S. J., G. C. White & F. L. Knopf. 2002. Advanced techniques for modelling nest survival. Ecology 94: 3476–3488.

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
36
Dinsmore, S. J. & J. J. Dinsmore. 2007. Modeling avian nest survival in program MARK. Studies in Avian Biology 34: 73−83.
Dudaniec, R. Y. & S. Kleindorfer. 2006. Effects of the parasitic flies of the genus Philornis (Diptera: Muscidae) on birds. Emu 106: 13–20.
Fessl, B. & S. Tebbich. 2002. Philornis downsi - a recently discovered parasite on the Galapagos Archipelago - a threat for Darwin‘s finches? Ibis 144: 445–451.
Fiorini, V. D., D. T. Tuero & J. C. Reboreda. 2009. Shiny Cowbirds benefits of synchronizing parasitism and puncturing eggs in large and small hosts. Animal Behaviour 77: 561–568.
Fraga, R. M. 1978. The Rufous-collared Sparrow as a host of the Shiny Cowbird. Wilson Bull. 90: 271–284.
Fraga, R. M. 1984. Bay-winged Cowbirds (Molothrus badius) remove ectoparasites from their brood parasites, the Screaming Cowbirds (Molothrus rufoaxillaris). Biotropica 16: 223–226.
Fraga, R. M, G. Pugnali & H. Casañas. 1998. Natural history and conservation status of the endangered Saffron-cowled Blackbird Xanthopsar flavus in Argentina. Bird Conserv. Int. 8: 255–267.
Friedmann, H. 1934. Additional notes on the birds victimized by the Shiny Cowbird (Molothrus bonariensis). Ibis 76: 340–347.
Gosling, L. M. & W. J. Sutherland. 2004. Behaviour and Conservation.Conservation Biology Series 2. Cambridge: Cambridge University Press.
Grant, T. A., T. L. Shaffer, E. M. Madden & P. J. Pietz. 2005. Time-specific variation in passerine nest survival: new insights for old questions. Auk 122: 661–672.
Kattan, G. H. 1995. Mechanisms of short incubation periods in brood parasitic cowbirds. Auk 112: 335–342.
Kluza, D. A. 1998. First record of Shiny Cowbird (Molothrus bonariensis) in Yucatán, Mexico. Wilson Bulletin 110: 429–430.
Kruger, O. 2007. Cuckoos, cowbirds and hosts: adaptations, trade-offs and constraints. Philosophical Transactions of the Royal Society B: Biological Sciences 362: 1873–1886.

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
37
López-Ortiz, R., E. A. Ventosa-Febles, L. R. Reitsma, D. Hengstenberg & W. Deluca. 2002. Increasing nest success in the yellow-shouldered blackbird Agelaius xanthomus in southwest Puerto Rico. Biological Conservation 108: 259–263.
Lowther, P. E. 2013.Lists of victims and hosts of the parasitic cowbirds, version 26 August 2013. The Field Museum,Chicago, IL, <http://fieldmuseum.org/sites/default/files/Molothrus_hosts-26aug2013.pdf > accessed 24 January 2014.
Marin, M. 2011. On the breeding biology of the Diuca Finch (Diuca diuca) in central Chile. Ornitología Neotropical 22: 369–378.
Martin, T. E. & G. R. Geupel. 1993. Nest-monitoring plots: methods for locating nests and monitoring success. Journal of Field Ornithology 64: 507–519.
Martin, T. E. 1996. Life history evolution in tropical and south temperate birds: what do we really know? Journal of Avian Biology. 27: 263–272.
Massoni, V. & J. C. Reboreda. 1998. Costs of parasitism and the lack of defenses on the Yellow-winged Blackbird - Shiny Cowbird system. Behavioural, Ecology and Sociobiology 42: 273–280.
Massoni, V. & J. C. Reboreda. 2002. A neglected cost of brood parasitism: egg punctures by Shiny Cowbirds during inspection of potential host nests. Condor 104: 407–412.
Oppel, S., H. M. Schaefer, V. Schmidt & B. Schröder. 2004. Cowbird parasitism of pale-headed brush-finch Atlapetes pallidiceps: implications for conservation and management. Bird Conservation International 14: 63–75.
Ortega, C. P. 1998. Cowbirds and other brood parasites.Tucson, Arizona: University of Arizona Press.
Peer, B. D. 2006. Egg destruction and egg removal by avian brood parasites: adaptiveness and consequences. Auk 123: 16–22.
Post, W., A. Cruz & D. B. McNair. 1993. The North American invasion pattern of the Shiny Cowbird. Journal of Field Ornithology 64: 32–41.
Post, W. & P. W. Jr. Sykes 2011. Reproductive Status of the Shiny Cowbird in North America. The Wilson Journal of Ornithology 123: 151–154.
Quiroga, M. & J. C. Reboreda. 2012. Lethal and sublethalefects of botfly (Philornis seguryi) parasitism on House Wren nestlings. Condor 114: 197–202.

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
38
Quiroga, M., J. C. Reboreda & A. H. Beltzer. 2012. Host use by botflies Philornis sp. in a passerine community in central Argentina. Revista Mexicana de Biodiversidad 83: 110–116.
Rabuffetti, F. L. & J. C. Reboreda. 2007. Early infestation by botflies (Philornis seguyi) decreases chick survival and nesting success in Chalk-browed Mockingbirds (Mimus saturninus). Auk 124: 898–906.
Reboreda, J. C., M. E. Mermoz, V. Massoni, A. A. Astié, & F. L. Rabuffetti. 2003. Impacto del parasitismo de cría del Tordo Renegrido (Molothrus bonariensis) sobre el éxito reproductivo de sus hospedadores. Hornero 18: 77–88.
Rothstein, S. I. 1990. A model system for coevolution: avian brood parasitism. Annual Review of Ecology and Systematics. 21: 481–508.
Segura, L. N. & J. C. Reboreda. 2011. Botfly parasitism effects on nestling growth and mortality of Red-crested Cardinals. Wilson Journal of Ornithology. 123: 107–115.
Segura, L. N. & J. C. Reboreda. 2012. Nest survival of Red-crested Cardinals increases with nest age in south-temperate forests of Argentina. Journal of Field Ornithology 83: 343–350.
Statsoft, Inc. 2004. STATISTICA data analysis software system, version 7. www.statsoft.com.
Sutherland, W. J. 2000. The Conservation Handbook: Research, Management and Policy. Blackwell Science, Oxford.
Teixeira, D. M. 1999. Myiasis caused by obligatory parasites. Ib. General observations on the biology of species of the genus Philornis Meinert, 1890 (Diptera, Muscidae). Myiasis in man and animals in the Neotropical Region. Bibliographic database.
Tressens, S. G., R. O. Vanni & M. G. López. 2002. Las plantas terrestres. Pp 201-379. En: Arbo, M. M. & Tressens, S. G. eds. Flora del Iberá Corrientes, Argentina: Editorial Universitaria de la Universidad Nacional del Nordeste.
Tuero, D. T., V. D. Fiorini & J. C. Reboreda. 2007. Effects of Shiny Cowbird parasitism on different components of House Wren reproductive success. Ibis 149: 521–527.
Uhazy, L. S. & W. J. Arendt. 1986. Pathogenesis associated with philornis myiasis (Diptera: Muscidae) on nestling Pearly-eyed Thrashers (Aves: Mimidae) in the Luquillo rain forest, PuertoRico. Journal of Wildlife Diseases 22: 224–237.

BIOLOGÍA REPRODUCTIVA Y EFECTOS DEL PARASITISMO
39
White, G. C. & K. P. Burnham. 1999. Program MARK: survival estimation from populations of marked animals. Bird Study (Suppl.) 46: 120–138.
Wiley, J. W., W. Post & A. Cruz. 1991. Conservation of the Yellow-shouldered Blackbird Agelaius xanthomus, an endangered West Indian species. Biological Conservation 55: 119–138.

VARIACIÓN GEOGRÁFICA DEL CANTO
40
CAPÍTULO 3
VARIACIÓN GEOGRÁFICA DEL CANTO
3.1 Introducción
3.1.1. El comportamiento animal en la biología de la conservación
La diversidad biológica incluye a la diversidad de comportamientos, que pueden ser de suma importancia en la evolución, supervivencia y persistencia de las poblaciones y especies. El comportamiento es un elemento crucial del fenotipo de un individuo con el que interactúa con el ambiente biótico y abiótico que lo rodea (Whitehead 2010). Por ello, conocer los patrones de comportamientos de las poblaciones es muy importante para el manejo y conservación de la fauna silvestre (Sutherland 1998). Los comportamientos pueden ser innatos o aprendidos (Cavalli-Sforza & Feldman 1981). Los caracteres aprendidos son adquiridos de otros individuos de la misma generación o de otra generación a través de la transmisión cultural. Este tipo de transmisión no genética puede darse por varios procesos, como imitación, observación, condicionamiento, impronta o enseñanza directa (Cavalli-Sforza & Feldman 1981). La incorporación del estudio de los atributos culturales, además de los innatos, en un contexto de conservación se ha sugerido como una nueva forma de identificar la diversidad y el estado de las poblaciones de animales. Varios autores concuerdan en que los comportamientos de transmisión cultural deben estar integrados a la biología de la conservación (Slabbekoorn & Smith 2002, Whitehead et al. 2004, Laiolo & Jovani 2007) y que su variación en especies amenazadas debe ser conservada. Ryan (2006) propone el concepto de ―unidades culturales‖ para delimitar poblaciones con características particulares del comportamiento.
3.1.2. Adquisición del canto
El canto es el sistema de comunicación más desarrollado en las aves (Catchpole & Slater 2008). Las vocalizaciones son producidas por un órgano especializado llamado siringe (Figura 3.1, Suthers 2004), que sólo se encuentra en este grupo de animales (Greenewalt 1968) y sería análogo a las cuerdas vocales de los mamíferos. Las vocalizaciones emitidas por las aves se dividen en cantos y llamadas. Los cantos, generalmente realizados por los machos en época reproductiva, son más largos y complejos, y tienen dos funciones principales: la atracción de hembras y la defensa del territorio frente a otros machos (Eriksson & Wallin 1986, McDonald 1989, Mountjoy & Lemon 1991). Las llamadas tienden a ser cortas, más simples y no están restringidas a un sexo. Además, pueden tener diversas funciones, como las de contacto o reconocimiento entre miembros de la pareja o familia, la de alerta ante la presencia de predadores u otras fuentes de peligro, o la de

VARIACIÓN GEOGRÁFICA DEL CANTO
41
defensa de territorio y disputas entre machos, entre otras (Brown 1975, Catchpole & Slater 1995).
Figura 3.1. Esquema de la posición del aparato vocal de las aves. Se observa la ―siringe‖, el órgano responsable de producir los sonidos complejos.
El desarrollo del canto varía entre especies. En las aves en general y en los
paseriformes suboscines, el canto es innato y las capacidades para emitir un canto normal están codificadas genéticamente. En los paseriformes oscines, en cambio, la adquisición del canto se realiza a través del aprendizaje (Catchpole & Slater 1995), aunque algunos aspectos, como la duración y el rango de frecuencias, pueden estar determinados genéticamente (Seddon 2005, Mahler & Gil 2009, Valderrama et al. 2013). En los paseriformes oscines, que incluyen al Cardenal Amarillo, el aprendizaje del canto se produce por imitación, principalmente de individuos conespecíficos. Los cantos pasan de generación en generación por transmisión cultural, a menudo entre vecinos (Lachlan & Slater 2003, Nordby et al. 1999), o, más raramente, de padres a hijos (Grant & Grant 1996, Mann & Slater 1995). Esta manera de adquirir los patrones específicos del canto se denomina aprendizaje social (Cavalli-Sforza & Feldman 1981, Boyd & Richerson 1985) y es, probablemente, uno de los ejemplos mejor estudiados de rasgos culturales en animales no humanos (Slater 1986). La transmisión no genética de rasgos vocales a través de generaciones puede ser una poderosa fuente de variación debido a imprecisiones al copiar el canto (Dawkins 1976, Mundinger 1980, Lynch 1996). El aprendizaje vocal imitativo permite la generación y la rápida transmisión de nuevos patrones de estructura vocal (Slabbekoorn & Smith 2002, Slater 1989), que conducen a diferencias en los cantos entre poblaciones de la misma especie (Catchpole & Slater 2008).
Pico Lengua
Tráquea
Siringe Bronquio
Pulmón

VARIACIÓN GEOGRÁFICA DEL CANTO
42
3.1.3. Variación geográfica del canto
Se ha documentado variación geográfica en las vocalizaciones de varios paseriformes (Mundinger 1982, Catchpole & Slater 2008). Estos cantos locales suelen describirse como ―dialectos‖ y ocurren cuando un grupo de machos conspecíficos comparte características vocales que difieren en forma discreta de aquellas de otros grupos (Baker & Cunningham 1985). La variación geográfica en los cantos de las aves evoluciona a través de la interacción entre los mecanismos de aprendizaje y de aislamiento, y está influenciada por los efectos de la deriva cultural, la deriva génica, la selección cultural, la selección natural y la selección sexual (Podos et al. 2004, Podos & Warren 2007, Catchpole & Slater 2008). La acumulación de pequeñas diferencias debido a la copia de imprecisiones durante la transmisión cultural y la menor probabilidad de los individuos distantes para interactuar socialmente a menudo conducen a un patrón de diferenciación vocal llamado "aislamiento por distancia" (Cavalli-Sforza 2000, González & Ornelas 2014).
La deriva cultural puede conducir a la fijación aleatoria de algunos tipos de canto o tipos de nota en poblaciones aisladas (Lynch & Baker 1994, Baker et al. 2001, Koetz et al. 2007) en escalas de tiempo evolutivamente cortas (Baker et al. 2003, Edwards et al. 2005). Este proceso es importante para la biología de la conservación ya que grandes poblaciones continuas se pueden ir reduciendo a poblaciones cada vez más pequeñas y aisladas a través de la pérdida del hábitat y la fragmentación, impactando sobre los rasgos transmitidos culturalmente (Laiolo & Tella 2007, Jamieson & Lacy 2012). Algunos investigadores han observado que el canto de las aves se empobrece como consecuencia de la interrupción en la transmisión cultural (Laiolo & Tella 2005, 2006, Lonsdorf 2007, Parker et al. 2010, 2012). Este empobrecimiento se correlaciona, además, con la viabilidad de las poblaciones (Laiolo 2008, Laiolo et al. 2008), siendo las más pequeñas las que presentan una menor variabilidad en sus cantos y las más susceptibles a la extinción. En un contexto de grandes cambios en el paisaje y de preocupación por la pérdida de diversidad, la comprensión de cómo la fragmentación del paisaje afecta y estructura el comportamiento de las poblaciones naturales es importante para mejorar el conocimiento ecológico y definir estrategias óptimas para la conservación de especies amenazadas (Diniz-Filho & Telles 2002, Manel et al. 2003).
3.1.4. Variación en las características de los dialectos
Los cantos pueden ser caracterizados por una serie de caracteres fácilmente cuantificables, tales como los parámetros de frecuencia, parámetros temporales y el tamaño y estructuración del repertorio (Gil & Gahr 2002). Algunos aspectos del canto están determinados por las restricciones impuestas por la morfología del ave, especialmente por el tamaño corporal y el pico. Las aves de mayor tamaño corporal exhiben frecuencias acústicas más bajas, debido a que sus siringes son más grandes y tienen menor capacidad

VARIACIÓN GEOGRÁFICA DEL CANTO
43
vibratoria de sus membranas (Wallschläger 1980, Ryan & Brenowitz 1985, Tubaro & Mahler 1998). Por otro lado, la forma y tamaño del pico limitan la habilidad para modular el tracto vocal durante la producción del canto, afectando la estructura espectral del sonido, principalmente el ancho de banda y el número de sílabas producidos por unidad de tiempo (Palacios & Tubaro 2000, Podos 2001, Grant & Grant 2002, Podos & Nowicki 2004).Otros aspectos del canto son más variables y pueden variar geográficamente determinando los dilectos. Cuatro hipótesis explican la función y el mantenimiento de los dialectos:
1) Procesos históricos: Los dialectos locales son creados por inexactitudes ocasionales en copiar el canto y se mantienen simplemente por aislamiento geográfico (Andrew 1962, Baptista 1975, King 1972, Payne 1981, Petrinovich et al. 1981).
2) Hipótesis de adaptación local o genética: Los dialectos se mantienen porque sirven como marcadores de población que las hembras locales usan para identificar y por lo tanto para aparearse preferentemente con machos locales (Nottebohm 1969, Baker et al. 1981, MacDougall-Shackleton & MacDougall-Shackleton 2001).
3) Hipótesis de adaptación acústica: Los dialectos surgen y se mantienen debido a que esas vocalizaciones se transmiten mejor a través del hábitat local (Morton 1975, Wiley & Richards 1978). En ambientes cerrados, donde la principal fuente de degradación es el eco, los cantos suelen tener notas largas con baja o lenta modulación de frecuencia, y con una considerable separación temporal entre ellas. Por otro lado, en zonas abiertas, la turbulencia del aire o las fluctuaciones locales en la temperatura y humedad suelen favorecer la producción de notas breves y con menor separación temporal para aumentar la probabilidad de que al menos alguna parte de la señal llegue al receptor.
4) Hipótesis de adaptación social: los dialectos se mantienen a través de la copia de cantos de machos establecidos con el fin de obtener estatus, territorio y pareja (Yoktan et al. 2011).
3.1.5. Objetivos
En este capítulo se estudia la variación geográfica del canto del Cardenal Amarillo. En particular, se estudian las vocalizaciones a lo largo de la distribución y se analiza cómo el aislamiento de las poblaciones a través de la fragmentación del hábitat y la extracción de individuos afecta los patrones del canto. Esta especie, al igual que otras aves, sube a las copas de los árboles para emitir su canto (Domínguez obs. pers.). Este comportamiento puede ser consecuencia de un esfuerzo de los individuos para evitar la interferencia con el ruido de fondo. El ruido causado por el viento (de baja frecuencia) y el de los artrópodos (de alta frecuencia), deja en el medio una banda de frecuencias (generalmente entre 1-4 KHz) con menor interferencia que pueden ser aprovechadas por las aves (Ryan & Brenowitz 1985, Slabbekoorn & Peet 2003).

VARIACIÓN GEOGRÁFICA DEL CANTO
44
3.2 Metodología
3.2.1. Grabación de cantos
Las grabaciones se obtuvieron durante cuatro temporadas reproductivas (septiembre - diciembre de 2011 a 2014) en cuatro poblaciones diferentes que cubren gran parte de la distribución conocida del Cardenal Amarillo en Argentina y Uruguay: 1) Corrientes (C); 2) San Luis (SL); 3) Río Negro (RN); y 4) Uruguay (U; Fig. 3.2). La posibilidad de grabar repetidamente a un mismo individuo fue mínima ya que la mayoría de los cardenales habían sido anillados previamente para identificación individual. Todas las grabaciones se hicieron cerca de las aves (de 5 a 10 m) mientras cantaban con un equipo grabador ZOH4NK H4n. El conjunto de datos incluye de 3 a 8 cantos de alta calidad por individuo de un total de 30 machos. Algunos cantos fueron descartados debido a la mala calidad de la grabación, pero un total de 245 cantos fueron analizados. Los cantos fueron digitalizados con el programa para análisis de sonido "RavenPro" versión 1.4 (Charif et al. 2007) y se visualizaron como espectrogramas con una ventana tipo "Hamming", una frecuencia de muestreo de 256 muestras, y un ancho de banda de 224 Hz. Se eligió la combinación de colores "escala de grises", y los valores de brillo y contraste se mantuvieron a 50% en todos los casos para estandarizar las mediciones.
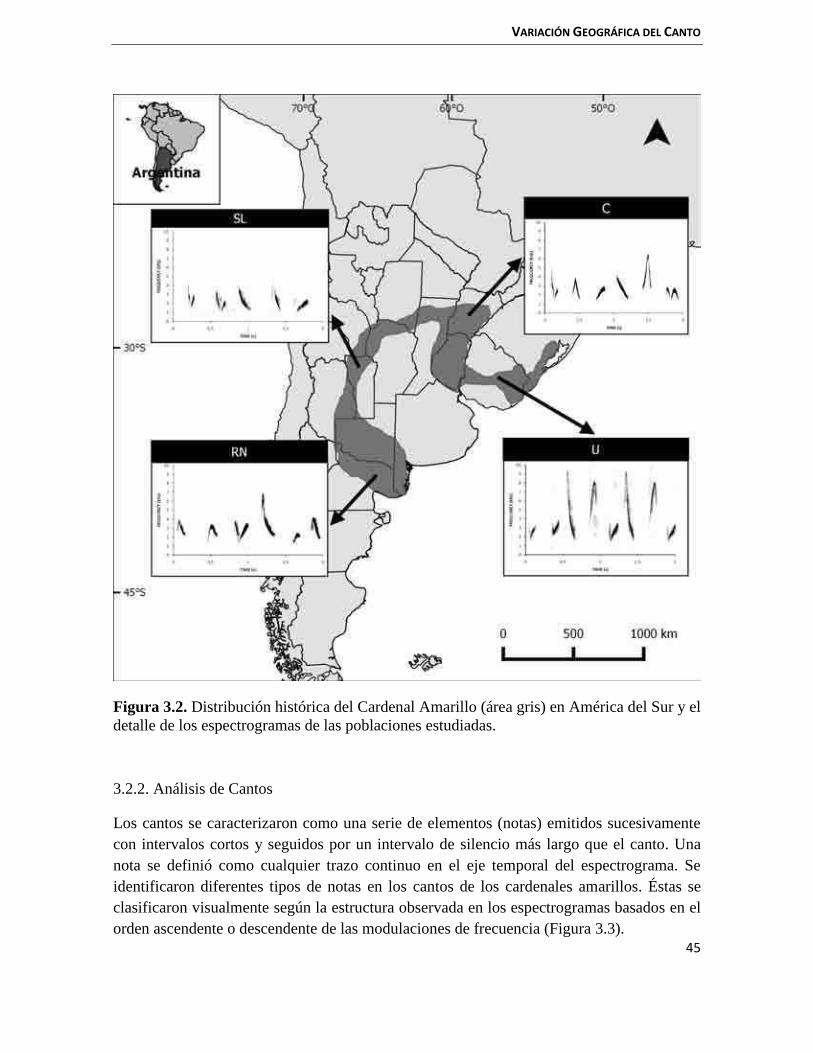
VARIACIÓN GEOGRÁFICA DEL CANTO
45
Figura 3.2. Distribución histórica del Cardenal Amarillo (área gris) en América del Sur y el detalle de los espectrogramas de las poblaciones estudiadas.
3.2.2. Análisis de Cantos
Los cantos se caracterizaron como una serie de elementos (notas) emitidos sucesivamente con intervalos cortos y seguidos por un intervalo de silencio más largo que el canto. Una nota se definió como cualquier trazo continuo en el eje temporal del espectrograma. Se identificaron diferentes tipos de notas en los cantos de los cardenales amarillos. Éstas se clasificaron visualmente según la estructura observada en los espectrogramas basados en el orden ascendente o descendente de las modulaciones de frecuencia (Figura 3.3).

VARIACIÓN GEOGRÁFICA DEL CANTO
46
Figura 3.3. Espectrograma que muestra el repertorio del Cardenal Amarillo con los 5 tipos de notas diferentes (A - E) y las variables medidas sobre los cantos. SD: Duración del canto, NL: Largo de la nota, InI: Intervalo inter-nota, SFMIN: Frecuencia mínima del canto, SFMAX: Frecuencia máxima del canto, SBAND: Ancho de banda de la nota.
En cada espectrograma se midieron 14 variables acústicas (Figura 3.3). Estas variables se pueden clasificar en tres categorías: 1) de frecuencia (medidas en Hz), 2) de tiempo (medidas en seg), y 3) estructurales. 1) Frecuencias máximas y mínimas (SFMAX y SFMIN), ancho de banda máximo (maxSBAND, medido como la diferencia máxima entre las frecuencias máxima y mínima de una nota), ancho de banda mínimo (minSBAND, medido como la diferencia mínima entre la frecuencia máxima y mínima de una nota) y frecuencia enfatizada (SFEMF, correspondiente a la frecuencia con la mayor intensidad en el canto); 2) duración del canto (SD), duración de la nota más larga (maxNL), duración de la nota más corta (minNL), intervalo máximo entre notas (maxInI: duración máxima del silencio entre el final de una nota y el comienzo de la siguiente), intervalo mínimo entre notas (minINI: duración mínima del silencio entre el final de una nota y el comienzo de la siguiente) y proporción de sonido (SP, medida como la diferencia entre la duración del canto y la duración de todos los intervalos de silencio entre notas); 3) número de notas (NN), proporción de notas simples (PSN: se definen como notas ascendentes o descendentes – ver notas C y D en Figura 3.3) y la proporción de notas que inicialmente ascienden (PIA - notas B, C y E en Figura 3.3). Cuando aparecieron armónicos se midió la

VARIACIÓN GEOGRÁFICA DEL CANTO
47
banda de frecuencias con mayor energía. Los valores de estas variables acústicas fueron promediados para los múltiples cantos (3-8) de cada individuo.
3.2.3 Análisis estadísticos
Se estudió la presencia de valores extremos y se analizó la normalidad y homocedasticidad univariada tanto gráfica como analíticamente. Se examinó la asociación entre las variables utilizando matrices de dispersión, coeficientes de correlación de Pearson y análisis de componentes principales (ACP), con el fin de descartar variables muy correlacionadas e incluir sólo las variables independientes en los análisis. Para estudiar las diferencias en los cantos entre las poblaciones de Cardenal Amarillo se realizó un análisis multivariado de la varianza (MANOVA). Para determinar si los cantos pueden ser agrupados de acuerdo a las variables elegidas se llevó a cabo un análisis discriminante utilizando 7 variables cuantitativas y la localidad del canto como la variable de agrupación. Todos los análisis multivariados se realizaron con Infostat (Di Rienzo et al. 2013).
Se evaluó si las diferencias de los cantos entre las poblaciones están correlacionadas con la distancia geográfica realizando una prueba de Mantel. Para construir la matriz de distancias entre cantos, se utilizó la distancia Euclídea entre los centroides de las poblaciones calculada con GenAlEx (Peakall & Smouse 2006, 2012). Las distancias geográficas (no lineales sino reales) se calcularon con Google Earth. El estadístico r de la prueba de Mantel se calculó con GenAlEx aplicando un procedimiento de aleatorización de Monte Carlo con 9999 permutaciones.
3.3 Resultados
Se analizaron los cantos de 30 cardenales amarillos, pertenecientes a cuatro poblaciones diferentes: 10 individuos de Corrientes, 7 de Río Negro, 4 de San Luis y 9 de Uruguay. Se identificaron cinco tipos de notas diferentes en los cantos de los cardenales amarillos (Figura 3.3). Los elementos que componen los cantos son relativamente sencillos con ninguna, una, o tres modulaciones de frecuencia. En promedio, el 33% de las notas del canto son notas simples, es decir, sin modulaciones de frecuencia. En general, el canto está compuesto por seis notas con una duración promedio de 2 segundos y abarca un rango de frecuencias entre 1500 y 4800 Hz (Tabla 3.1).
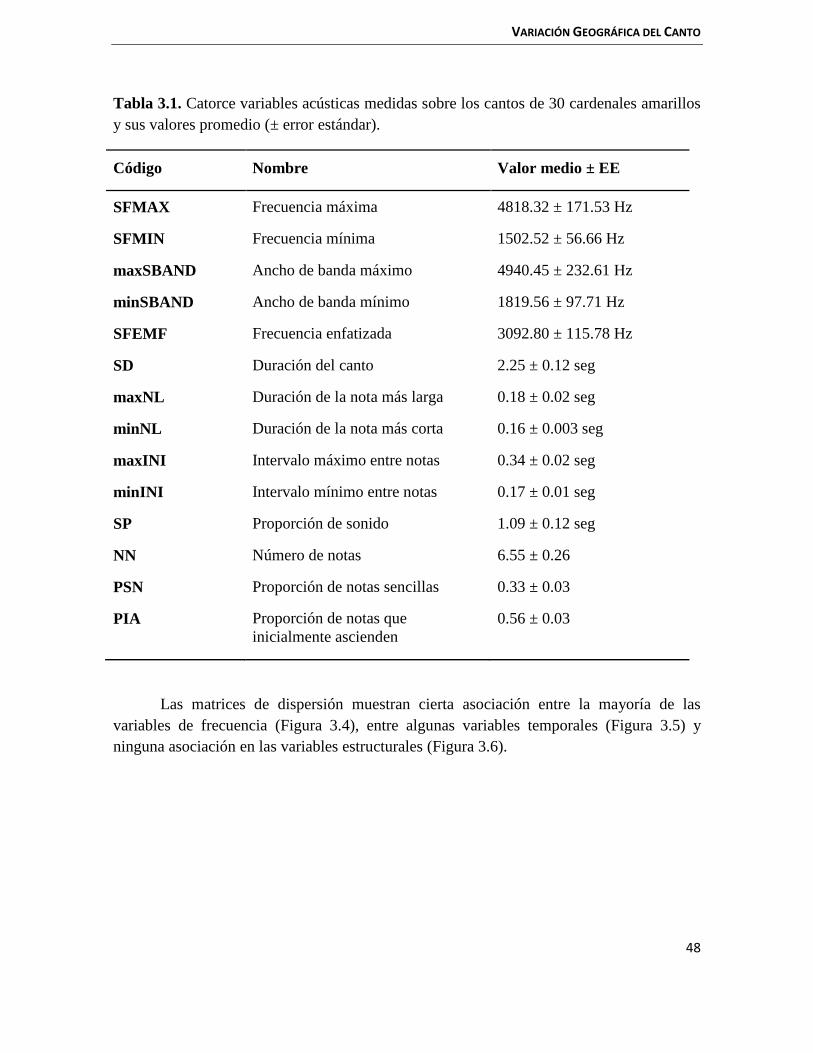
VARIACIÓN GEOGRÁFICA DEL CANTO
48
Tabla 3.1. Catorce variables acústicas medidas sobre los cantos de 30 cardenales amarillos y sus valores promedio (± error estándar).
Código Nombre Valor medio ± EE
SFMAX Frecuencia máxima 4818.32 ± 171.53 Hz
SFMIN Frecuencia mínima 1502.52 ± 56.66 Hz
maxSBAND Ancho de banda máximo 4940.45 ± 232.61 Hz
minSBAND Ancho de banda mínimo 1819.56 ± 97.71 Hz
SFEMF Frecuencia enfatizada 3092.80 ± 115.78 Hz
SD Duración del canto 2.25 ± 0.12 seg
maxNL Duración de la nota más larga 0.18 ± 0.02 seg
minNL Duración de la nota más corta 0.16 ± 0.003 seg
maxINI Intervalo máximo entre notas 0.34 ± 0.02 seg
minINI Intervalo mínimo entre notas 0.17 ± 0.01 seg
SP Proporción de sonido 1.09 ± 0.12 seg
NN Número de notas 6.55 ± 0.26
PSN Proporción de notas sencillas 0.33 ± 0.03
PIA Proporción de notas que inicialmente ascienden
0.56 ± 0.03
Las matrices de dispersión muestran cierta asociación entre la mayoría de las variables de frecuencia (Figura 3.4), entre algunas variables temporales (Figura 3.5) y ninguna asociación en las variables estructurales (Figura 3.6).

VARIACIÓN GEOGRÁFICA DEL CANTO
49
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
SFMAX
SFMIN
SFEMF
maxSBAND
minSBAND
Figura 3.4. Matriz de asociación entre las variables de frecuencia.
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
SL
SP
minNL
maxNL
minInI
maxInI
Figura 3.5. Matriz de asociación entre las variables temporales.

VARIACIÓN GEOGRÁFICA DEL CANTO
50
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Studen
NN
PSN
PIA
Figura 3.6. Matriz de asociación entre las variables estructurales.
Los coeficientes de correlación de Pearson entre variables acústicas fueron significativos para SFMAX, SFMIN y SFEMF, maxINI y minINI (r = 0.72, p < 0.001) y para SD y SP (r = 0.66, p < 0.001). La correlación más alta fue entre SFMAX y SFEMF (r = 0.80, p <0.0001). Las variables estructurales no estuvieron correlacionadas entre sí. Sin embargo, PIA mostró correlación con las variables de frecuencia (0.4 < r < 0.9, P < 0.05). Entre las variables correlacionadas se optó por conservar la de mayor carga en los componentes principales del ACP. minNL y maxSBAND no cumplieron con el supuesto de normalidad (prueba de Shapiro-Wilks, W = 0.75 P < 0.001, W = 0.78 P < 0.001, respectivamente). Teniendo en cuenta las correlaciones entre variables y el cumplimiento del supuesto de normalidad se eliminaron siete variables acústicas. El set de datos final para el MANOVA y el análisis discriminante consistió en 2 variables de frecuencia (SFEMF, minSBAND), 3 variables temporales (SP, maxNL y minINI) y 2 variables estructurales (NN y PSN). El MANOVA mostró variación significativa entre las cuatro poblaciones (Traza de Pillai F21,66 = 2.85; p = 0.0006). Las comparaciones de Hotelling indican que todas las poblaciones difieren entre sí, excepto SL, que sólo difiere de U pero no de las otras dos poblaciones argentinas (Tabla 3.2), aunque esto podría estar relacionado con un bajo tamaño muestral para San Luis.

VARIACIÓN GEOGRÁFICA DEL CANTO
51
Tabla 3.2. Resultados de los contrastes de Hotelling con corrección por Bonferroni de la prueba MANOVA. La tabla muestra los valores medios de las siete variables de canto para las cuatro regiones. Letras iguales significan que no hubo diferencias significativas entre sitios, letras distintas significan que hubo diferencias significativas entre sitios.
Los dos primeros ejes del análisis discriminante explican el 95% de las diferencias entre las poblaciones (Tabla 3.3) y la función discriminante asignó correctamente al 87% de los individuos a su población de origen (4 errores en 30 casos, Tabla 3.4). Esto indica que los cantos de cada población de Cardenal Amarillo son característicos y diferentes de los cantos de otras poblaciones (Figura 3.7). Las variables más importantes que permitieron esta separación fueron SFEMF, minSBAND, PSN y minINI (Tabla 3.3).
-5 -4 -2 -1 1 2 4
Eje Canónico 1
-3
-2
-1
0
1
2
3
Eje
Can
ónic
o 2
SFEMF
minSBAND
NN
PSN
SP
maxNL minInI
SFEMF
minSBAND
NN
PSN
SP
maxNL minInI
Figura 3.7. Análisis discriminante. Los cantos individuales se representan como círculos en los dos primeros ejes canónicos. Las líneas muestran las variables que definen estos ejes y su peso: NN (número de notas), SP (proporción de sonido), PSN (proporción de notas simples), minINI (intervalo internota mínimo), maxNL (máxima longitud de nota), minSBAND (ancho de banda mínimo) y SFEMF (frecuencia enfatizada del canto). Los polígonos incluyen los cantos de los cardenales amarillos de cada área geográfica: Uruguay (rojo), Corrientes (azul), San Luis (verde) y Río Negro (amarillo).
Región \
SFEMF
(Hz) minSBAND
(Hz) NN PSN SP
(seg) maxNL
(seg) minInI
(seg)
n
U 3789.96 2263. 41 7.44 0.47 1.58 0.25 0.14 9 A
C 2922.40 1656.92 6.38 0.35 0.79 0.14 0.13 10 B
SL 2467.08 1377.27 5.85 0.35 0.70 0.16 0.22 4 BC
RN 2797.43 1733.98 6.08 0. 13 1.11 0.18 0.21 7 C
Variable
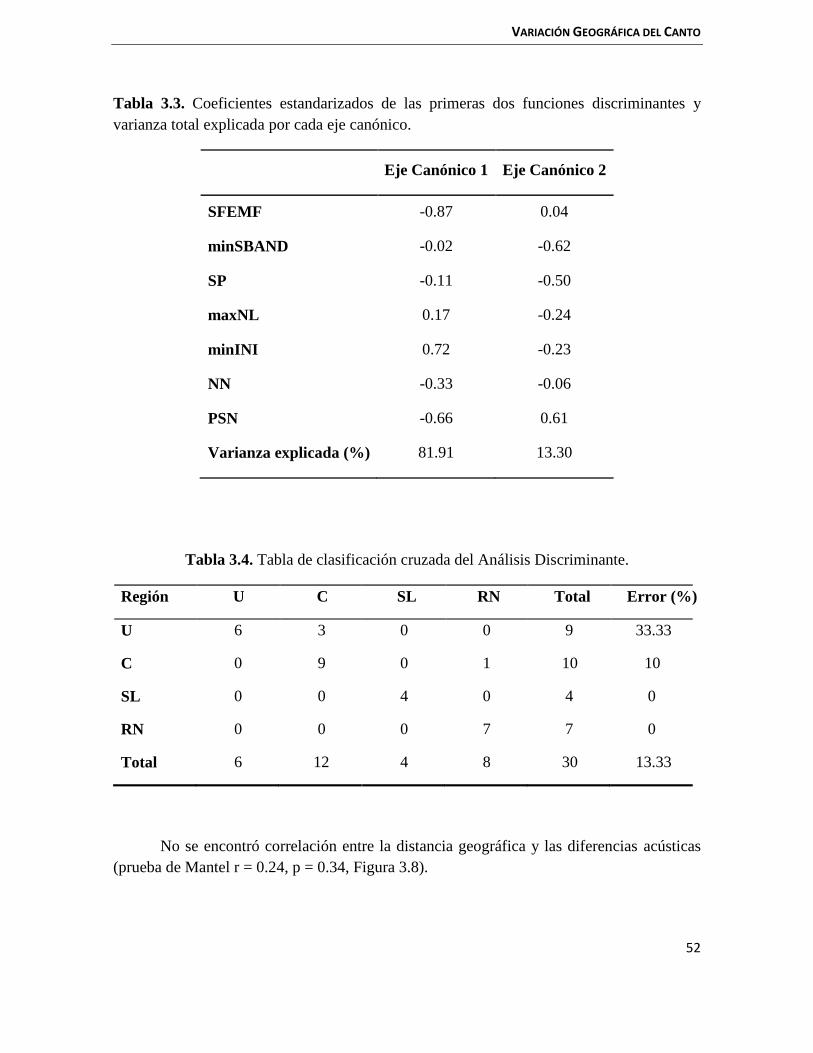
VARIACIÓN GEOGRÁFICA DEL CANTO
52
Tabla 3.3. Coeficientes estandarizados de las primeras dos funciones discriminantes y varianza total explicada por cada eje canónico.
Eje Canónico 1 Eje Canónico 2
SFEMF -0.87 0.04
minSBAND -0.02 -0.62
SP -0.11 -0.50
maxNL 0.17 -0.24
minINI 0.72 -0.23
NN -0.33 -0.06
PSN -0.66 0.61
Varianza explicada (%) 81.91 13.30
Tabla 3.4. Tabla de clasificación cruzada del Análisis Discriminante.
No se encontró correlación entre la distancia geográfica y las diferencias acústicas (prueba de Mantel r = 0.24, p = 0.34, Figura 3.8).
Región U C SL RN Total Error (%)
U 6 3 0 0 9 33.33
C 0 9 0 1 10 10
SL 0 0 4 0 4 0
RN 0 0 0 7 7 0
Total 6 12 4 8 30 13.33

VARIACIÓN GEOGRÁFICA DEL CANTO
53
Figura 3.8. Disimilitud en los cantos (distancia euclídea) versus distancia geográfica (km) entre todos los pares de poblaciones. Mantel r = 0.24, P = 0.34.
3.4 Discusión
El análisis de los cantos del Cardenal Amarillo evidenció que es posible diferenciar a las poblaciones según sus características acústicas. Se encontraron diferencias significativas en las características estructurales, temporales y de frecuencia analizadas entre los cantos de las distintas poblaciones. La frecuencia enfatizada (SFEMF) es la variable que mejor distingue a los individuos de diferentes sitios, siendo los cardenales de Uruguay los que muestran la mayor frecuencia en sus cantos (media = 3789.96 Hz) y los individuos de San Luis el valor más bajo (media = 2467.08 Hz). Los cantos de los cardenales de Uruguay y San Luis no sólo difieren en su frecuencia enfatizada sino que en San Luis los cantos también exhiben los valores más bajos de ancho de banda mínimo, el menor número de notas y la menor proporción de sonido. Los cantos de Uruguay, por el contrario, mostraron los valores más altos de toda la distribución actual de cardenales amarillos para esas mismas variables. Por otro lado, los cantos de los cardenales de Río Negro se caracterizan por tener la menor proporción de notas simples y los de Corrientes, si bien tienen valores intermedios para casi todas las variables, parecen tener un canto con notas más cortas y más seguidas (mínimo valor de maxNL y de minINI). Existe muy poca superposición de cantos en el espacio multivariado y solo ocurre entre los cantos de Uruguay y Corrientes, posiblemente debido a la mayor variabilidad de las características acústicas de los cantos de Uruguay. Las diferencias entre los cantos mostraron una muy buena separación de las poblaciones en el espacio multivariado (Figura 3.7) y una buena clasificación para el análisis discriminante (con sólo 4 errores en 30 casos; Tabla 3.4).

VARIACIÓN GEOGRÁFICA DEL CANTO
54
La existencia de dialectos en los cantos del Cardenal Amarillo a lo largo de su distribución en Argentina y Uruguay no es sorprendente, teniendo en cuenta que las poblaciones de cardenales son residentes y están aisladas entre sí por extensiones de hábitat inadecuado o modificado. No solamente son fieles a sus territorios durante la época reproductiva, sino que también durante la temporada no reproductiva son aves locales y dispersan cortas distancias (Domínguez obs. per.).
La explicación más probable para la existencia de dialectos en esta especie es que la variación se haya generado aleatoriamente en las distintas regiones y que luego las tasas de variación se vieron profundizadas por la fragmentación del ambiente y el aislamiento de las poblaciones. En el caso de que los cardenales estuvieran distribuidos en un espacio continuo, se espera que los más cercanos copien sus patrones de cantos y las características acústicas de regiones cercanas sean más parecidas que las de regiones lejanas. Sin embargo, al desaparecer una población ya sea por la pérdida del hábitat o por la extracción intencional con fines comerciales de todos sus integrantes, algunos tipos de cantos desaparecerían y los cantos remanentes en los extremos de la distribución original ya no podrán copiarse debido a la distancia geográfica que los separa. Las poblaciones aisladas sufrirían una especie de ―cuello de botella cultural‖, presentando una disminución en la variabilidad de las propiedades acústicas de sus cantos. Con el tiempo, y por la acumulación de errores de copiado, estas poblaciones evolucionarían patrones de cantos independientes (Lang & Barlow 1997; Parker et al. 2010) y no se ajustarían a un patrón de aislamiento por distancia.
Si bien la variación acústica también podría ser moldeada por la estructura del hábitat, es poco probable que esto explique las diferencias encontradas entre las poblaciones de cardenales, ya que se encuentran en parches de hábitat con estructuras vegetales similares. A pesar de que las regiones fitogeográficas varían a lo largo del rango de los cardenales amarillos, Chaqueña, del Espinal y del Monte (Cabrera 1976), las diferencias en la cobertura vegetal no parecerían ser tan diferentes como para originar una presión de selección diferencial. Si comparamos los cantos del ambiente de Reserva Quebracho de la Legua donde fueron grabados los cardenales en San Luis, que representa la transición entre el espinal y la ecorregión chaqueña con un 65% de cobertura vegetal (Iglesias et al. 2010), con los del Uruguay, donde el cardenal habita lo que se denominan ―montes abiertos‖ (Chebez 2008, Azpiroz et al. 2012), observamos que concuerda parcialmente con lo esperado bajo la hipótesis de adaptación acústica. En San Luis, el ambiente más cerrado, los cantos presentan la menor frecuencia y la mayor separación entre notas, pero las notas son más cortas, en contraposición a lo esperado para este ambiente. Para estudiar esta hipótesis con más detalle sería necesario cuantificar la cobertura vegetal para todos los ambientes y correlacionar esta variable con las características del canto de cada población.

VARIACIÓN GEOGRÁFICA DEL CANTO
55
La deriva cultural en cantos de poblaciones aisladas fue observada en diversas especies amenazadas. En un estudio realizado en Nueva Zelanda, donde se llevaron a cabo translocaciones con fines de conservación de 20-40 individuos del amenazado Philesturnus rufusater en distintas islas, encontraron que las poblaciones pequeñas translocadas inicialmente tenían una menor diversidad en sus cantos (comparadas con la población fuente) y que un tiempo de aproximadamente 50 años fue suficiente para que aparezcan cantos con características acústicas y repertorio distintos en las poblaciones aisladas (Parker et al. 2012). Un patrón de evolución del canto parecido fue encontrado en poblaciones recientemente translocadas de Callaeas wilsoni (formadas por menos de 20 individuos), que exhibían una variabilidad reducida en sus propiedades acústicas en comparación a la población fuente y que se diferenciadaron en menos de 10 años (Valderrama et al. 2013). Laiolo & Tella (2005, 2006) encontraron evidencias de que la transmisión del canto está limitada por la distribución de los parches de hábitat adecuado para la alondra de Dupont, Chersophilus duponti. En hábitats fragmentados, el grado de similitud en los cantos de los vecinos era mayor; por el contrario, el grado de similitud entre los cantos de inividuos no vecinos era bajo, probablemente debido a la falta de interacciones entre los individuos aislados por barreras de hábitat. Los investigadores concluyen que barreras antropogénicas del hábitat podrían alterar la percepción de las aves sobre la distribución espacial de los rivales, dando lugar a una contracción del rango espacial del nicho acústico del individuo. Aunque se ve profundizada por el aislamiento entre las poblaciones, la variabilidad geográfica del canto es un patrón general en paseriformes oscines (Lougheed & Handford 1992) dado que presentan una rápida adquisición de nuevas variantes (Baker et al. 2003, Grant & Grant 2009). Esto contrasta con otros grupos de aves, donde la transmisión del canto ocurre a nivel genético y presenta poca variabilidad entre poblaciones (McCracken & Sheldon 1997, Mahler & Tubaro 2001, Seddon 2005).
La fragmentación y pérdida del hábitat son algunas de las amenazas antropogénicas más importantes para la viabilidad de las especies en peligro de extinción (Soulé 1986). Estas amenazas también afectan a los comportamientos y podrían actuar más rápido sobre los cantos que se basan en la transmisión cultural. Perder o disminuir el tamaño de una sola población podría comprometer o eliminar una tradición cultural, incluso antes de que se observe una reducción en la diversidad genética o de que se vea amenazada la persistencia de la especie (Laiolo & Jovani 2007). Las poblaciones remanentes de cardenales amarillos son pequeñas, probablemente estén en declive y se distribuyen irregularmente. La identificación de cuatro unidades culturales para el Cardenal Amarillo (U, C, SL y RN) puede ser de importancia para las decisiones de manejo, no solamente para conservar la variabilidad encontrada, sino también en la elaboración de planes de reintroducción o translocación.

VARIACIÓN GEOGRÁFICA DEL CANTO
56
3.5 Referencias bibliográficas
Andrew, R. J. 1962. Evolution of intelligence and vocal mimicking. Science 137: 585–589.
Azpiroz, A. B., M. Alfaro & S. Jiménez. 2012. Lista Roja de las Aves del Uruguay. Una evaluación del estado de conservación de la avifauna nacional con base en los criterios de la Unión Internacional para la Conservación de la Naturaleza. Dirección Nacional de Medio Ambiente, Montevideo.
Baker, M. C., D. B.Thompson, G. L. Sherman & M. A. Cunningham. 1981. The role of male interactions in maintaining population dialect structure. Behavioral Ecology and Sociobiology 8: 65–69.
Baker, M. C. & M. A. Cunningham. 1985. The biology of bird-song dialects. Behavior and Brain Science 8: 85–133.
Baker, M. C., E. M. Baker & M. S. Baker. 2001. Island and island-like effects on vocal repertoire of singing honeyeaters. Animal Behaviour 62: 767–774.
Baker, M. C., M. S. A. Baker & E. M. Baker. 2003. Rapid evolution of a novel song and an increase in repertoire size in an island population of an Australian songbird. Ibis 145: 465–471.
Baptista, L. F. 1975. Song dialects and demes in sedentary populations of the White-crowned Sparrow (Zonotrichia leucophrys nuttalli). University of California Press 105: 1–52.
Boyd, R. & P. J. Richerson. 1985. Culture and the Evolutionary Process. Chicago: University of Chicago Press. Campbell, D. T.
Brown, J. L. 1975. The evolution of behavior. W. W. Norton & company INC, New York,
Cabrera, A. L. 1976. Regiones fitogeográficas argentinas. Pp. 85. En: Enciclopedia argentina de agricultura y jardinería; Tomo 2 fasc. 1. Acme, Buenos Aires.
Catchpole, C. K. & P. J. B. Slater. 1995, Bird song: biological themes and variations, Cambridge University Press, Cambridge, Reino Unido.
Catchpole, C. K. & P. J. B. Slater. 2008. Bird song: biological themes and variations, 2nd edn. Cambridge University Press, Cambridge, Reino Unido.
Cavalli-Sforza, L. L. & M. W. Feldman. 1981. Cultural transmission and evolution: a quantitative approach. Princeton University Press, Princeton, NJ.

VARIACIÓN GEOGRÁFICA DEL CANTO
57
Cavalli-Sforza, L. L. 2000. Genes, Peoples and Languages. North Point Press, New York.
Charif, R. A., C. W. Clarck & K. M. Fristrup. 2007. Raven Pro 1.3 user‘s manual. Cornell Laboratory of Ornithology, Ithaca, NY.
Chebez, J. C. 2008. Los que se van. Fauna argentina amenazada. Tomo 2. Aves. Ed. Albatros, Buenos Aires.
Dawkins, R. 1976. The selfish gene. Oxford University Press, Oxford, United Kingdom.
Diniz-Filho, J. A. F & M. Telles Pires de Campo. 2002. Spatial autocorrelation analysis and the identification of operational units for conservation in continuos populations. Conservation Biology 16: 924–935.
Di Rienzo, J. A., F. Casanoves, M. G. Balzarini, L. Gonzalez, M. Tablada & C. W. Robledo. InfoStat versión 2013. InfoStat Group, Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba, Argentina. URL http://www.infostat.com.ar
Edwards, S. V., S. B. Kingan, J. D. Calkins, C. N. Balakrishnan, W. B. Jennings, W. J. Swanson & M. D. Sorenson. 2005. Speciation in birds: Genes, geography, and sexual selection. Proceedings of the National Academy of Sciences 102: 6550–6557.
Eriksson, D. & L. Wallin. 1986. Male bird song attracts females: a field experiment. Behavioural Ecology and Sociobiology 19: 297–299.
Gil, D. & M. Gahr. 2002. The honesty of bird song: multiple constraints for multiple traits. Trends of Ecology and Evolution 17: 133–141.
González, C. & J. F. Ornelas. 2014. Acoustic Divergence with Gene Flow in a Lekking Hummingbird with Complex Songs. PLoS ONE 9: e109241.
Grant, P. R. & B. R. Grant. 1996. Cultural inheritance of song and its role in the evolution of Darwin‘s finches. Evolution 50: 2471–2487.
Grant, P. R. & B. R. Grant. 2002. Adaptive radiation of Darwin‘s finches. American Scientist 90: 130–139.
Grant, P. R. & B. R. Grant. 2009. The secondary contact phase of allopatric speciation in Darwin's finches. Proceedings of the National Academy of Science 106: 20141–20148.
Greenewalt, C. H. 1968. Bird Song: Acoustics and Physiology. Washington (DC): Smithsonian Institution Press.

VARIACIÓN GEOGRÁFICA DEL CANTO
58
Iglesias, M. R., A. Barchuk & M. P. Grilli. 2010. Dinámica estacional e interanual del NDVI en bosques nativos de zonas áridas argentinas Seasonal and interannual dynamics of NDVI in arid forest of Argentina. Revista de Teledetección 34: 44–54.
Jamieson, I. G. & R. C. Lacy. 2012. Managing genetic issues in reintroduction biology. Pp. 441–476. En Reintroduction Biology: Integrating Science and Management (eds J.G. Ewen, D.P. Armstrong, K.A. Parker & P.J. Seddon), Wiley-Blackwell, Chichester, Reino Unido.
King, J. R. 1972. Variation in the song of the Rufous-collared Sparrow, Zonotrichia capensis, in northwestern Argentina. Zeitschrift für Tierpsychologie 30, 344–373.
Koetz, A. H., D. A. Westcott & B. C. Congdon. 2007. Geographical variation in song frequency and structure: the effects of vicariant isolation, habitat type and body size. Animal Behaviour 74: 1573–1583.
Kroodsma, D. E. 1977. Correlates of song organization among North-American wrens. American Naturalist 111: 995–1008.
Lachlan, R. F. & P. J. B. Slater. 2003. Song learning by chaffinches: how accurate, and from where? Animal Behaviour 65: 957–969.
Laiolo, P. & J. L. Tella. 2005. Habitat fragmentation affects culture transmission: Patterns of song matching in Dupont‘s lark. Journal of Applied Ecology 42: 1183–1193.
Laiolo, P. & J. L. Tella. 2006. Landscape bioacoustics allows detection of the effects of habitat patchiness on population structure. Ecology 87: 1203–14.
Laiolo, P. & R. Jovani. 2007. The emergence of animal culture conservation. Trends in Ecology and Evolution 22: 5.
Laiolo, P. & J. L. Tella. 2007. Erosion of animal cultures in fragmented landscapes. Frontiers in Ecology and the Environment 5: 68–72.
Laiolo, P. 2008. Characterizing the spatial structure of songbird cultures. Ecological Applications 18: 1774–1780.
Laiolo, P., M. Vögeli, D. Serrano & J. L. Tella. 2008. Song diversity predicts the viability of fragmented bird populations. PLoS ONE 3:e1822.
Lang, A. L. & J. C. Barlow. 1997. Cultural evolution in the Eurasian tree sparrow: divergence between introduced and ancestral populations. Condor 99: 413–423.

VARIACIÓN GEOGRÁFICA DEL CANTO
59
Lonsdorf, V. 2007. The role of behavioral research in the conservation of chimpanzees and gorillas. Journal of Applied Animal Welfare Science 10: 71–78.
Lougheed, S. C. & P. Handford. 1992 Vocal dialects and the structure of geographic variation in morphological and allozymic characters in the rufous-collared sparrow, Zonotrichia capensis. Evolution 46: 1443–1456.
Lynch, A. & A. J. Baker. 1994. A population memetics approach to cultural evolution in chaffinch song: differentiation among populations. Evolution 48: 351–359.
Lynch, A. 1996. The population mimetics of birdsong. Pp. 181–197. En Ecology and evolution of acoustic communication in birds (ed. D. Kroodsma & E. Miller), Cornell University Press, Ithaca, Nueva York.
MacDougall-Shackleton, E. A. & S. A. MacDougall-Shackleton. 2001. Cultural and genetic evolution in mountain white-crowned sparrows: song dialects are associated with population structure. Evolution, 55: 2568–2575.
Mahler, B. & P. L. Tubaro. 2001. Relationship between song characters and morphology in New World pigeons. Biological Journal of the Linnean Society 74: 533–539.
Mahler, B. & D. Gil. 2009. The Evolution of Song in the Phylloscopus Leaf Warblers (Aves: Sylviidae): A Tale of Sexual Selection, Habitat Adaptation, and Morphological Constraints. Advances in the study of behavior 40: 35–66.
Manel, S., M. K. Schwartz, G. Luikart & P. Taberlet. 2003. Landscape genetics: combining landscape ecology and population genetics. Trends in Ecology and Evolutiom 18: 189–197.
Mann, N. I. & P. J. B. Slater. 1995. Song tutor choice by zebra finches in aviaries. Animal Behaviour 49: 811–820.
McCracken, K. G. & F. H. Sheldon. 1997. Avian vocalizations and phylogenetic signal. Proceedings of the National Academy of Sciences of the United States of America 94: 3833–3836.
McDonald, M. V. 1989. Function of song in Scott‘s seaside sparrow Ammodramus maritimus peninsulae. Animal Behaviour 38: 468–485.
Morton, E. S. 1975. Ecological sources of selection on avian sounds. Am. Nat. 109: 17–34.
Mountjoy, D. J. & R. E. Lemon. 1991. Song as an attractant for male and female European starlings, and the influence of song complexity on their response. Behavioral Ecology and Sociobiology 28: 97–100.

VARIACIÓN GEOGRÁFICA DEL CANTO
60
Mundinger, P.C. 1980. Animal cultures and a general theory of cultural evolution. Ethology and Sociobiology. 1: 83–223.
Mundinger, P. C. 1982. Pp 147–208. En: Kroodsma DE, Miller EH (eds) Microgeographic and macrogeographic variation in the acquired vocalizations of birds. Ecology and evolution of acoustic communication in birds. Cornell University Press, Ithaca.
Nordby, J. C., S. E. Campbell & M. D. Beecher. 1999. Ecological correlates of song learning in song sparrows. Behavioural Ecology 10: 287–297.
Nottebohm, F. 1969. The song of the chingolo, Zonotrichia capensis, in Argentina: Description and evaluation of a system of dialects. Condor 71: 299–315.
Palacios, M. G. & P. L. Tubaro. 2000. Does beak size affect acoustic frequencies in woodcreepers? The Condor 102: 553–560.
Parker, K. A., M. E. Hauber & D. H. Brunton. 2010. Contemporary cultural evolution of a conspecific recognition signal following serial translocations. Evolution 64: 2431–2441.
Parker, K. A., M. J. Anderson, P. F. Jenkins & D. H. Brunton. 2012. The effects of translocation-induced isolation and fragmentation on the cultural evolution of bird song. Ecology Letters. 15: 778–785.
Payne, R. B. 1981. Population structure and social behavior: Models for testing the ecological significance of song dialects in birds. Pp. 108–120. En ‗‗Natural Selection and Social Behavior: Recent Research and New Theory‘‘ (R. D. Alexander and D. W. Tinkle, Eds.), Chiron Press, New York.
Peakall, R. & P. E. Smouse. 2006. GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes 6: 288–295.
Peakall, R. & P. E. Smouse. 2012. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research—an update. Bioinformatics 28: 2537–2539.
Petrinovich, L., T. L. Patterson & L. F. Baptista. 1981. Song dialects as barriers to dispersal: a reevaluation. Evolution, 35:180–188.
Podos, J. 2001. Correlated evolution of morphology and vocal signal structure in Darwin‘s finches. Nature 409: 185–188.
Podos, J. & S. Nowicki. 2004. Beaks, adaptation, and vocal evolution in Darwin‘s finches. Bioscience 54:501–510.

VARIACIÓN GEOGRÁFICA DEL CANTO
61
Podos, J., S. K. Huber, & B. Taft. 2004. Bird song: the interface of evolution and mechanism. Annual Review of Ecology, Evolution, and Systematics 35: 55–87.
Podos, J. & P. S. Warren. 2007. The evolution of geographic variation in birdsong. Advances in the Study of Behavior 37: 403–458.
Ryan, M. J. & E. A Brenowitz. 1985. The role of body size, phylogeny, and ambient noise in the evolution of bird song. American Naturalist 126: 87–100.
Ryan, S. J. 2006. The role of culture in conservation planning for small or endangered populations. Conserv. Biol. 20: 1321–1324.
Slabbekoorn, H. & T. B. Smith. 2002. Bird song, ecology and speciation. Philosophical Transactions of the Royal Society of London B: Biological Sciences 357: 493–503.
Slabbekoorn, H. & M. Peet. 2003. Birds sing at a higher pitch in urban noise. Nature 424:267.
Slater, P. J. B. 1986. The cultural transmission of bird song. Trends in Ecology and Evolution 1: 94–97.
Slater, P. J. B. 1989. Bird song learning: Causes and consequences. Ethology Ecology & Evolution 1: 19–46.
Seddon, N. 2005. Ecological adaptation and species recognition drives vocal evolution in Neotropical suboscine birds. Evolution 59: 200–215.
Soulé, M. E. 1986. What is conservation biology? BioScience 35: 727–734.
Sutherland, W. J. 1998. The importance of behavioural studies in conservation biology. Animal Behaviour 56: 801–809.
Suthers, R. A. 2004. How birds sing and why it matters. Pp. 272–295. En: Marler, P., Slabbekoorn, H. (Eds.), Nature‘s Music: The Science of Birdsong. Elsevier, San Diego.
Tubaro, P. L. & B. Mahler. 1998. Acoustic frequencies and body mass in new world doves. Condor 100: 54–61.
Valderrama, S. V., L. E. Molles, & J. R. Waas. 2013. Effects of Population Size on Singing Behavior of a Rare Duetting Songbird. Conservation Biology, 27: 210–218.
Wallschäger, D. 1980. Correlation of song frequency and body weight in passerine birds. Experientia 36: 412.

VARIACIÓN GEOGRÁFICA DEL CANTO
62
Wiley, R. H & D. G. Richards. 1978. Physical constraints on acoustic communication in
atmosphereimplications for evolution of animal vocalizations. Behavioural, Ecology
and Sociobiology 3: 69–94.
Whitehead, H., L. Rendell, R. W. Osborne & B. Würsig. 2004. Culture and conservation of non-humans with reference to whales and dolphins: review and new directions. Biological Conservation 120: 427–437.
Whitehead, H. 2010. Conserving and managing animals that learn socially and share cultures. Learning and Behavior 38: 329–336.
Yoktan, K., E. Geffen, A. Ilany, Y. Yom-Tov, A. Naor & N. Leader. 2011. Vocal dialect and genetic subdivisions along a geographic gradient in the orange-tufted sunbird. Behavioural Ecology and Sociobiology 65:1389–1402.

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
63
CAPÍTULO 4
VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
4.1. Introducción
En algunas especies existe variación geográfica de caracteres morfológicos (Dale 2006). Estas diferencias fenotípicas intraespecíficas pueden haber surgido por los efectos combinados de la mutación, deriva genética, migración y eventos históricos relacionados con cambios demográficos sufridos por las poblaciones (Greenberg et al. 1998, Babik et al. 2005, D‘Anatro & Lessa 2006, Fornel et al. 2010). Las poblaciones de especies con dispersión limitada que han ocupado un rango de distribución amplio en hábitats continuos pueden desarrollar un tipo de aislamiento por distancia en relación con su estructura geográfica (Hamilton 1961). En contraposición, existen especies con altas capacidades dispersivas que, por lo general, no muestran variantes geográficas discretas, o bien especies en donde sus poblaciones han permanecido en aislamiento estricto y sin flujo génico durante mucho tiempo, donde las diferencias morfológicas son independientes de la distancia geográfica (Smith & Patton 1984, Slatkin 1987,1993).
Muchas especies exhiben diferenciación gradual a lo largo de su distribución en varias medidas morfológicas y de coloración. El Gorrión Común, Passer domesticus, por ejemplo, fue introducido desde Inglaterra a Estados Unidos y en menos de 50 años se evidenció una diferenciación gradual del tamaño y color (Johnston & Selander 1964, Gould & Johnston 1972, Baker 1980). Otros casos documentados de especies donde se observa un patrón de variación gradual en aspectos de morfología y coloración son el Zorzal Americano, Turdus migratorius (Aldrich & James 1991, James 1991), y el Tordo Sargento, Agelaius phoeniceus (James & NeSmith 1988). También se ha observado variación geográfica de medidas morfológicas para la Tijereta, Tyrannus savana, especies del género Vidua y para el Monarca Colilargo Asiático, Terpsiphone paradisi (Alatalo et al. 1988).
La coloración del plumaje es una de las variables morfológicas más estudiadas y a menudo se observa variación a nivel intraespecífico (Dale 2006). Puede tener un rol importante en la termorregulación, el camuflaje y en la comunicación social de las aves (Saino et al. 2013). La coloración del plumaje es el resultado de pigmentos depositados en las plumas y/o de su microestructura. La mayoría de los colores son el resultado de la combinación de ambos mecanismos, aunque generalmente predomina uno (Andersson & Prager 2006). Los pigmentos son moléculas que absorben y emiten longitudes de onda diferencialmente. El color producido por los pigmentos es determinado por la estructura de la molécula del pigmento y su concentración (Andersson & Prager 2006, McGraw 2006b).

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
64
Entre los principales pigmentos se encuentran los carotenos (colores percibidos por el ojo humano del amarillo al rojo, Hill 2006) y las melaninas (percibidas como colores del castaño al negro, McGraw 2006b). Los carotenos son pigmentos adquiridos con la dieta que no pueden sintetizarse de novo (Brush 1990), por lo que se ha sugerido que podrían ser un indicador honesto de la calidad del portador (Pryke et al. 2001, McGraw 2006a, Lozano 1994, 2001). Los costos asociados a la búsqueda, adquisición y metabolismo de los alimentos, y a la deposición de los pigmentos en las plumas podría reflejar la calidad del individuo en relación con su la calidad genética (Zahavi 1975, Blount 2004), estado nutricional (Lozano 1994), capacidad de forrajeo, capacidad de competir por recursos (Pryke et al. 2001, Griggio et al. 2007) incluyendo alimentos y pareja (Olson y Owens 1998, Hill 2002) o la calidad de cuidado parental que puede proveer (Hill 1991, Massaro et al. 2003). Las melaninas, en cambio, pueden ser sintetizadas por el propio individuos a partir de la tirosina u otro aminoácido esencial y son los pigmentos presentes con mayor frecuencia en las aves y otros animales (McGraw 2006b). Estos pigmentos absorben la mayoría de las longitudes de onda, resultando en colores oscuros. Existen dos categorías generales de pigmentos melánicos, la eumelanina y la feomelanina. La pigmentación melánica está determinada genéticamente y es, por lo tanto, un carácter heredable en aves (Theron et al. 2001) que parecería estar codificada por un solo locus (Mundy 2005). Existen numerosas evidencias que muestran una correlación adaptativa de la variación melánica (Price & Bontranger 2001) aunque variables ambientales y condiciones fisiológicas y hormonales del individuo pueden afectar la producción de melanina (Jawor & Breitwisch 2003).
Los plumajes basados en carotenos y melaninas juegan un importante papel en las interacciones conespecíficas (Andersson 1994), tanto en interacciones intrasexuales (competencia entre los miembros de un sexo para acceder a fertilizaciones, Senar 2006) como intersexuales (evolución de caracteres de un sexo que son preferidos por los miembros del otro sexo, Hill 2006). Muchos estudios comportamentales estudiaron la coloración del plumaje basada en melanina analizando la asociación entre la concentración de este pigmento (o el tamaño de parches producidos por el mismo) y el estatus social o la agresividad del portador (Rohwer & Rohwer 1978, Senar 1999). Por ejemplo, en gorriones el parche melánico es un indicador de dominancia y es utilizado en la competencia intrasexual. Se ha observado que gorriones con parches melánicos más grandes son dominantes sobre los que tienen parches más pequeños (Møller 1987, 1988; Gonzalez et al. 2002). Si la competencia intrasexual se produce en ambos sexos en materia de recursos, pareja, o ambos, la selección debería favorecer la aparición de estos parches en ambos sexos limitando la evolución del dimorfismo sexual. En cuanto a la selección intersexual, se han encontrado evidencias de que la eumelanina también está involucrada en la elección de pareja (Kigma et al. 2008). Se ha observado que el tamaño del babero en los gorriones machos está positivamente asociado al éxito reproductivo (Jensen et al. 2004) y que en

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
65
numerosas especies la elección de pareja está claramente asociada con los parches melánicos (Norris 1990, Roulin 1999, Tarof et al. 2005).
La coloración melánica también puede estar bajo selección natural por su función termorreguladora. El modelo de Gloger (1833) predice una variación geográfica de acuerdo a las diferencias ambientales, donde las plumas tienden a ser de color oscuro en hábitats donde la humedad relativa es alta y pálidas donde la humedad es baja. La generalización de melanismos en climas húmedos también se extiende a mariposas, escarabajos, moscas y vertebrados homeotermos (Mayr 1963).
Diferencias en las presiones de selección sexual o natural a lo largo de la distribución de una especie pueden generar variación geográfica en la coloración (e.g. Toon et al. 2003, Saino et al. 2013). La mayoría de los estudios de coloración en aves se han concentrado únicamente en los machos (Amundsen 2000), posiblemente porque en general los machos son más conspicuos que las hembras. En este capítulo estudiamos la variación geográfica del plumaje y la morfología del Cardenal Amarillo, tanto en machos como en hembras. En el Cardenal Amarillo, ambos sexos tienen un parche melánico, denominado babero, formado por plumas negras que se posicionan debajo del pico, desde la garganta hasta la parte superior del pecho. Además, presentan un parche de coloración arriba del ojo, denominado parche supraciliar, el cual es blanco en las hembras y amarillo en los machos (Figura 4.4). El patrón de coloración y diferenciación morfológica puede tener explicaciones funcionales o no funcionales, aunque el objetivo de este capítulo se limita a conocer la existencia de variación geográfica para estos rasgos en el Cardenal Amarillo.
4.2. Metodología
4.2.1. Colección de datos
Los datos morfológicos fueron tomados de individuos capturados a campo y de pieles de museo. Se capturaron un total de 39 individuos en Corrientes (N = 27), San Luis (N = 6) y La Pampa (N = 6) durante cuatro temporadas reproductivas (septiembre - enero de 2011 a 2014) siguiendo la metodología explicada en el capítulo 2. Además, se midieron 46 pieles de especímenes de museo en los Museos de Ciencias Naturales Bernardino Rivadavia y de La Plata. Los individuos procesados provenían de las provincias de Corrientes (N = 12), San Luis (N = 9), La Pampa (N = 6), sur de Buenos Aires (N = 6) y de la Republica Oriental del Uruguay (N = 13). Los individuos del sur de la provincia de Buenos Aires, muestreados en Villarino, Villa Iris e Igarzábal, pertenecerían a la población asignada a Rio Negro en el capítulo 3. Se midieron once variables morfológicas en los individuos capturados: peso, medido con una balanza de resorte de 60 g marca Pesola AG modelo

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
66
20060, (error ± 0.5 g, Figura 4.1); alto, largo y ancho de pico (± 0.1 mm, Figura 4.2); largo de cresta, medido desde la inserción hasta la punta de la pluma más larga (± 0.1 mm); longitud de tarso, medido siguiendo el protocolo de Taylor y Jamieson (2007; ± 0.1 mm, Figura 4.3); longitud de cola, medida desde la base hasta la punta de las rectrices centrales (± 0.1 mm); largo del ala, medido con una regla del ala desde el vértice flexor del ala hasta el extremo de la primaria más larga, manteniendo la curvatura natural del ala al tomar la medida (± 1 mm); ancho y alto de babero (± 0.1 mm, Figura 4.3A) y largo de parche supraciliar (± 0.1 mm, Figura 4.3B). Todas las variables, excepto peso y largo del ala, fueron medidas con un calibre de reloj marca Measy2000 (error 0.1 mm). El patrón de coloración que se estudia en este capítulo no se enfoca en las diferencias de longitudes de onda reflectadas por el plumaje de los cardenales de distintas regiones sino en el tamaño de los parches de color.
Figura 4.1. Pesado de los cardenales a campo con ―pesola‖ o balanza de resorte.

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
67
Figura 4.2. Medidas de longitud de pico (A) y de tarso (B).
Figura 4.3. Las líneas rojas muestran las medidas morfométricas tomadas sobre los parches del plumaje: A) parche supraciliar (largo total; amarillo en machos); B) babero (ancho y alto). 4.2.2. Análisis Estadístico
Similar a lo realizado para el estudio de las vocalizaciones, se analizó la existencia de individuos ―outliers‖ (valores extremos) y se pusieron a prueba los supuestos de falta de normalidad y homocedasticidad univariada tanto en forma gráfica como analítica. Asimismo, se analizó la existencia de asociación entre variables mediante matrices de diagramas de dispersión y coeficientes de correlación de Pearson.
B
B A
A

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
68
Para determinar si alguna/s de las localidades muestreadas difiere/n significativamente en las medias poblacionales de las variables medidas se llevó a cabo un MANOVA. Para identificar las variables que permiten la separación de los grupos se realizó un Análisis Discriminante y, con una tabla de clasificación cruzada, se evaluó la asignación de los individuos a sus poblaciones de origen. Se excluyeron cuatro variables de ambos análisis: el peso, para el cual no se tenían datos de las pieles de museo, y las tres medidas morfométricas realizadas sobre los parches de coloración del plumaje, que no pudieron ser obtenidas en 7 individuos por la manera en que estaban taxidermizados. Estas cuatro variables fueron analizadas de manera univariada para las muestras disponibles.
Los análisis se realizaron en forma separada para machos y hembras utilizando el paquete estadístico InfoStat (Di Rienzo et al. 2013) y R.
4.3. Resultados
4.3.1. Machos
4.3.1.1. Peso
Esta medida fue tomada únicamente para los individuos capturados a campo: C (n = 15), LP (n = 6) y SL (n =6). No se encontraron diferencias en cuanto al peso para los cardenales amarillos de estas tres provincias (ANOVA F2,24 = 2.91, p > 0.05).
4.3.1.2. Coloración
Las medidas de los parches melánico y supraciliar fueron tomadas tanto en individuos capturados en la naturaleza (n = 24) como en pieles de museo (n = 32) representando las regiones de Uruguay (n = 10), Corrientes (n = 19), San Luis (n = 13), La Pampa (n = 10) y Buenos Aires (n = 5). Las 3 variables de coloración no estaban correlacionadas (Correlaciones de Pearson: -0.03 < r < -0.11; p > 0.05) y sólo se encontraron diferencias entre regiones para el alto de babero (F4,52 = 0.13 p = 0.02), para el que los cardenales de San Luis mostraron menor tamaño que los de Uruguay (Tabla 4.4, Figura 4.5). El ancho del babero (ANOVA F4,52 = 1.15, p > 0.05) y el largo del parche supraciliar (ANOVA F4,52 = 0.19, p > 0.05) no se diferenciaron entre regiones.

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
69
Tabla 4.4. Contraste de Tukey para el ANOVA para alto de babero. Letras iguales denotan diferencias no significativas entre regiones.
Región Media ES n
U 30.54 2.18 10 A
C 28.43 1.58 19 A B
SL 21.49 1.91 13 B
LP 27.59 2.18 10 A B
BS AS 24.54 3.08 5 A B
t version Student version Student version Student version Student version Student version Student vers
Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student vers
Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student vers
Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student vers
Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student vers
Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student vers
Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student vers
Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student vers
Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student vers
Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student vers
Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student vers
Student version Student version Student version Student version Student version Student version Stu
C LP SL U BsAs
Provincia
10
20
30
40
50
Alto B
abero
Figura 4.5. Diagrama de cajas (Box-plot) que muestra los valores del alto del babero para los cardenales amarillos pertenecientes a las 5 regiones estudiadas (C: Corrientes, LP: La Pampa, SL: San Luis, U: Uruguay y BsAs: Buenos Aires). El punto corresponde al valor medio, la línea horizontal dentro de la caja a la mediana. El límite inferior de la caja corresponde al cuartil 1 y el límite superior al cuartil 3. El límite inferior del bigote representa el dato mínimo y el bigote superior al dato máximo. Datos atípicos se muestran como un punto por fuera de la caja.

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
70
4.3.1.3. Análisis multivariados de las variables morfológicas
La matriz de datos para el análisis multivariado incluyó las medidas de 7 variables para 63 individuos: 10 de Uruguay, 22 de Corrientes, 13 de San Luis, 9 de La Pampa y 6 del sur de Buenos Aires. No se encontraron ―outliers‖ y no se detectó colinealidad entre ningún par de variables (Figura 4.5; Correlación de Pearson: 0.001 < r < 0.40; P > 0.05).
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
Student version Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
Student version Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
Student version Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
Student version Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
Student version Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
Student version Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
Student version Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
Student version Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
Student version Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
Student version Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
Student version Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
Student version Student version Student version Student version Student version Student version Student version Student version Student version
t version Student version Student version Student version Student version Student version Student version Student version Student version Student
P Largo
P Alto
P Ancho
Tarso
Ala
Cola
Cresta
Figura 4.5. Matriz de diagrama de dispersión entre siete variables morfométricas medidas sobre los cardenales: largo, alto y ancho del pico (P), largo del tarso, largo del ala, largo de la cola y largo de la cresta.
El MANOVA resultó significativo (Traza de Pillai F4,58 = 2.55, p = 0.0001) y el contraste de Hotelling corregido por Bonferroni evidenció diferencias significativas entre las regiones de Uruguay, Corrientes y San Luis pero una ausencia de diferencias de los individuos de La Pampa con los de Uruguay y San Luis y los de Buenos Aires con los de las demás localidades (Tabla 4.3). Ninguna región se diferenció completamente de todas las demás.

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
71
Tabla 4.3. Resultados de los contrastes de Hotelling corregidos por Bonferroni para el MANOVA.
Región P
Largo P
Alto P
Ancho Tarso Ala Cola Cresta n
U 14.42 11.09 9.94 25.19 92.89 94.86 31.36 10 A
C 15.11 11.76 9.97 26.65 96.80 91.08 28.93 22 B
SL 14.72 10.98 10.21 25.65 90.12 88.86 25.65 13 C
LP 14.07 11.69 10.18 24.89 89.27 87.86 27.48 9 A C
BS AS 14.93 10.52 10.00 25.98 89.40 93.60 28.10 6 A B C
El gráfico de los individuos en función de los primeros dos ejes del Análisis Discriminante refleja este resultado (Figura 4.6), mostrando una tendencia de los individuos de Corrientes a presentar un mayor del largo del ala, y de los de San Luis a presentar una menor cresta, en contraposición a los individuos de Uruguay que muestran crestas más largas (Figura 4.6, Tabla 4.4). A su vez, los individuos de Buenos Aires y La Pampa se solapan ampliamente con los individuos de las demás localidades. La tabla de clasificación cruzada mostró un error de asignación del 35% (Tabla 4.5).

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
72
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
t version Student version Student version Student version Student version Student version Student version Student version Student ver
Student version Student version Student version Student version Student version Student version Student version Student version Stu
-4 -3 -2 -1 1 2 3 4
Eje canónico 1
-3
-2
-1
0
1
2
3
Eje
canó
nico
2
P Largo
P Alto
P Ancho
Tarso
Ala
Cola
Cresta
P Largo
P Alto
P Ancho
Tarso
Ala
Cola
Cresta
Figura 4.6. Biplot del análisis discriminante para los ejes 1 y 2. Los individuos se representan como círculos en los dos primeros ejes canónicos. Las líneas muestran las variables que definen estos ejes y su peso. En verde: Uruguay, azul: Corrientes, rojo: San Luis, amarillo: La Pampa, y en violeta: Buenos Aires.
Tabla 4.4. Coeficientes estandarizados de las primeras dos funciones discriminantes y varianza total explicada por cada eje canónico.
Eje Canónico 1 Eje Canónico 2
P Largo -0.30 0.77
P Alto -0.41 -0.50
P Ancho 0.66 0.09
Tarso -0.39 0.38
Ala -0.84 0.10
Cola -0.05 -0.31
Cresta -0.13 -0.95
Varianza explicada (%) 59.75 28.87

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
73
Tabla 4.5. Tabla de clasificación cruzada del Análisis Discriminante.
Región U C SL LP BSAS Total Error (%)
U 7 0 1 1 1 10 30
C 3 16 0 0 3 22 27.27
SL 0 1 7 1 4 13 46.15
LP 1 1 1 6 0 9 33.33
BSAS 2 0 1 0 3 6 50
Total 13 18 10 8 11 60 35
4.3.2. Hembras
4.3.2.1. Peso
Solamente se obtuvieron datos de peso para hembras de la población de Corrientes (n = 12), razón por la cual no se pudieron realizar comparaciones entre las localidades. El peso promedio de las hembras fue de 47.40 ± 0.9 gr (media ± DE), similar al peso promedio de los machos de esta localidad (47.30 ± 1.5 gr, n = 15).
4.3.2.2. Coloración
Las medidas del babero melánico y del parche supraciliar blanco fueron tomadas tanto en individuos capturados en la naturaleza (N = 10) como en pieles de museo (N = 16) representando las regiones de Uruguay (n = 3), Corrientes (n = 17), San Luis (n = 2), La Pampa (n = 3) y Buenos Aires (n = 3). Los bajos valores muestrales para algunas regiones no permitieron hacer un análisis estadístico, pero un análisis descriptivo donde se grafican los individuos en función de las tres variables de coloración muestra una alta superposición entre las hembras provenientes de diferentes localidades (Figura 4.7).

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
74
Figura 4.7. Gráfica de dispersión 3D donde se muestran las hembras de Cardenal Amarillo de las distintas regiones en función de las tres variables de coloración estudiadas.
4.3.2.3. Análisis multivariados de las variables morfológicas
Las 7 variables morfológicas fueron tomadas tanto en individuos capturados en la naturaleza (N = 12) como en pieles de museo (N = 18) representando las regiones de Uruguay (n = 3), Corrientes (n = 17), San Luis (n = 2), La Pampa (n = 3) y Buenos Aires (n = 5). Dado el bajo número de individuos incluidos en la mayoría de las poblaciones, se realizó un análisis descriptivo. Mediante un análisis de componentes principales se resumieron las siete variables morfológicas en dos ejes, que explicaron el 54 % de la varianza. El gráfico de los individuos en función de estos ejes mostró que también para las variables morfológicas existe una alta superposición de las hembras provenientes de distintas localidades (Figura 4.8).

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
75
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
t version Student version Student version Student version Student version Student version Student version Student version Student versi
Student version Student version Student version Student version Student version Student version Student version Student version Stud
-4 -2 0 2 4
Componente Principal 1
-4
-2
0
2
4
Co
mp
on
en
te P
rin
cip
al 2
P Largo
P Alto
P Ancho
Cola
Ala
Tarso
Cresta
P Largo
P Alto
P Ancho
Cola
Ala
Tarso
Cresta
Figura 4.8. Análisis de Componentes Principales. Los individuos se representan como círculos en los dos primeros ejes. Las líneas muestran las variables que definen estos ejes y su peso. En rosa: Uruguay, amarillo: Corrientes, gris: San Luis, verde: La Pampa, y en azul: Buenos Aires.
4.4. Discusión
Los resultados de este estudio muestran que no existe un diferenciación global para los caracteres morfológicos de los cardenales amarillos en toda su distribución. Observamos que para algunas variables, unas pocas poblaciones se ubican en los extremos de los valores que toman dichas variables, mientras que para las demás poblaciones encontramos una gran varianza a nivel interindividual. Las mayores diferencias se encontraron entre los cardenales machos del este y los del oeste de la distribución. La población de Uruguay mostró los mayores valores para el alto del babero (30.54 mm) y el largo de la cresta (31.36 mm) y el pico más angosto (9.94 mm), en contraposición a San Luis, que mostró los menores valores para alto de babero y largo de cresta (21.49 mm y 21. 49 mm, respectivamente) y el pico más ancho (10.21 mm). A su vez, los individuos de Corrientes mostraron los mayores tamaños estructurales dados por el largo del ala (96.80 mm) y del tarso (26.65 mm). Estas diferencias no fueron evidentes para las hembras de la especie, lo cual podría estar indicando una presencia de selección sexual diferencial en estas localidades, favoreciendo la expresión de estos caracteres en uno de los sentidos (selección direccional). Se ha visto que parches melánicos del plumaje (Senar 1999), así como plumas ornamentales tales como la cresta (Møller & Pomiankowski 1993) son importantes en

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
76
interacciones intra e intersexuales. Es posible que una mayor presión de selección en Uruguay, que podría estar dada por una mayor densidad de individuos o una menor disponibilidad de recursos, favorezca la expresión de estos caracteres de plumaje que podrían reflejar la calidad del individuo en las interacciones con otros machos o la atracción de las hembras. Asimismo, se ha relacionado al tamaño corporal de los machos con presiones de selección sexual que favorecen machos de mayores tamaños (Bosch 1996), tal como se observa en la población de Corrientes.
Por otro lado, se ha visto que diferencias en los nichos alimenticios pueden actuar sobre la morfología de los picos, favoreciendo aquellos que sean más efectivos para la adquisición del alimento (Grant & Grant 2006). Diferencias en los ambientes podrían estar llevando a una divergencia en los tamaños del pico entre los cardenales de Uruguay y San Luis. En Uruguay el ambiente se caracteriza por montes abiertos que son predominantemente húmedos y levemente ondulados, mientras que la region analizada de San Luis se encuentra en la transicion entre el espinal y la ecorregión chaqueña, es más árida y presenta una mayor cobertura vegetal que Uruguay. En caso de que existiera una adaptación local del tamaño del pico al alimento, esperamos que esta diferencia se observe también en las hembras. Aunque no se pudo analizar estadísiticamente, se observa, sin embargo, que las hembras de San Luis presentan picos más angostos que los de Uruguay (11 vs 11.8 mm).
En resumen, los resultados de este capítulo muestran que los indiviudos del Cardenal Amarillo son altamente variables para los caracteres morfológicos y no siguen un patrón de diferenciación geográfica. Solamente encontramos una tendencia a presentar valores extremos para algunos caracteres en algunas de las poblaciones, lo que podría estar relacionado con presiones diferenciales de selección sexual o natural. Sin embargo, esta hipótesis debería ponerse a prueba experimentalmente demostrando que la expresión de estos carcteres confiere una ventaja adaptativa a sus portadores.
4.5. Referencias bibliográficas
Alatalo, R., J. Höglund & A. Lundberg. 1988. Patterns of variation in tail ornament size in Birds. Biological Journal of the Linnean Society 34: 363–374.
Aldrich, J. W. & F. C. James. 1991. Ecogeographic variation in the American Robin (Turdus migratorius). The Auk 108: 230–249.
Amundsen, T. 2000. Why are female birds ornamented?. Trends in Ecology & Evolution 15: 149–155.
Andersson M. 1994. Sexual selection. Princeton University, Press Princeton, Nueva Jersey.

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
77
Andersson, S. & M. Prager. 2006. Quantification of avian coloration. Pp. 41–89. En Hill G. E. & K. J. McGraw [eds.], Bird coloration, Vol. I: Mechanisms and measurements. Harvard University Press, Massachusetts.
Babik, W., W. Branicki, J. Crnobrnja-Isailovi, D. Cogălniceanu, I. Sas, K. Olgun, N. A. Poyarkov, M. Garcia-París & J.W. Arntzen. 2005. Phylogeography of two European newt species: discordance between mtDNA and morphology. Molecular Ecology 14: 2475–2491.
Baker, A. J. 1980. Morphometric differentiation in New Zealand populations of the house sparrow (Passer domesticus). Evolution 34: 638–653.
Blount, J. D. 2004. Carotenoids and lifehistory evolution in animals. Archives
Biochemistry and Biophysics 430: 10–15.
Bosch, M. 1996. Sexual Size Dimorphism and Determination of Sex in Yellow-Legged Gulls. Journal of Field Ornithology 67: 534–541.
Brush, A. H. 1990. Metabolism of carotenoid pigments in birds. The FASEB Journal: 4: 2969–2977.
D‘Anatro, A. & E. P. Lessa. 2006. Geometric morphometric analysis of geographic variation in the Río Negro tuco-tuco, Ctenomys rionegrensis (Rodentia: Ctenomyidae). Zeitschrift für Säugetierkunde 71: 288–298.
Dale, J. 2006. Intraespecific variation in coloration. Pp.3686. En Hill G. E. & K. J. McGraw [eds.], Bird coloration, Vol. II: Function and evolution. Harvard University Press, Cambridge, Massachusetts.
Di Rienzo, J. A., F. Casanoves, M. G. Balzarini, L. Gonzalez, M. Tablada & C. W. Robledo. InfoStat versión 2013. InfoStat Group, Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba, Argentina. URL http://www.infostat.com.ar
Fornel, R., P. Cordeiro-Estrela & R. T. De Freitas. 2010. Skull shape and size variation in Ctenomys minutus (Rodentia: Ctenomyidae) in geographical, chromosomal polymorphism, and environmental contexts. Biological Journal of the Linnean Society 101: 705–720.
Grant, P. R. & B. R. Grant. 2006. Evolution of character displacement in Darwin's finches. Science 313: 224–226.
Gloger, C. 1833. Das abändern der vögel durch einfluss des klimas. Breslau, Germany: A. Schulz.

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
78
Gonzalez, G., G. Sorci, L.C. Smith & F. de Lope. 2002. Social control and physiological cost of cheating in Status signalling male House Sparrows (Passer domesticus). Ethology 108: 289–302.
Gould, S. J., & R. F. Johnston. 1972. Geographic variation. Annual Review of Ecology and Systematics 3: 457–498.
Greenberg, G. R., P. J. Cordero, S. Droege & R. C. Fleischer. 1998. Morphological adaptation with no mitochondrial DNA differentiation in the coastal plain swamp sparrow. The Auk 115: 706–712.
Griggio, M., L. Serra, D. Licheri, A. Monti & A. Pilastro. 2007. Armaments and ornaments in the Rock Sparrow: a possible dual utility of a carotenoid based feather signal. Behavioral Ecology and Sociobiology 61: 423–433.
Hamilton, T. H. 1961. The adaptive significances of intraspecific trends of variation in wing length and body size among bird species. Evolution 15(2): 180–195.
Hill, G.E. 1991. Plumage coloration is a sexually selected indicator of male quality. Nature 350: 337–339.
Hill, G.E. 2002. A red bird in a brown bag: the function and evolution of ornamental plumage coloration in the House Finch. Oxford University Press, Oxford,.
Hill, K. J. 2006. Female mate choice for ornamental coloration. Pp.137200. En Hill G. E. & K. J. McGraw [eds.], Bird coloration, Vol. II: Function and evolution. Harvard University Press, Cambridge, Massachusetts.
James, F. C. & C. NeSmith. 1988. Nongenetic effects in geographic differences among nestling populations of Red-winged Blackbirds. Pp. 1424–1433. En H. Ouellet (ed.) Acta XIX Congressus Internationalis Omithologici, Vol. II.
James, F. C. 1991. Complementary descriptive and experimental studies of clinal variation in birds. American Zoologist 31: 694–706.
Jawor, J.M. & R. Breitwisch. 2003. Melanin ornaments, honesty, and sexual selection. The Auk 120: 249–265.
Jensen, H., B. E. Sæther, T. Ringsby, T. H., J. Tufto, S. Griffith & H. Ellergren. 2004. Lifetime reproductive success in relation to morphology in the house sparrow (Passer domesticus). J. Anim. Ecol. 73: 599–611.
Johnston, R. F. & R. K. Selander. 1964. House Sparrow: Rapid evolution of races in North America. Science 144:548–550.
Kigma, S.A., I. Szentirmai, T. Székely, V. Bókony, M. Bleeker, A. Liker & J. Komdeur.

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
79
2008. Sexual selection and the function of a melanin-based plumage ornament in polygamous Penduline Tits Remiz pendulinus. Behavioral Ecology and Sociobiology 62: 1277–1288.
Lozano, G. A. 1994. Carotenoids, parasites, and sexual selection. Oikos 70: 309–311.
Lozano, G. A. 2001. Carotenoids, immunity, and sexual selection: comparing apples and oranges? American Naturalist 158: 200–203.
Massaro, M., L. D. Davis & J. T. Darby. 2003. Carotenoid-derived ornaments reflect parental quality in male and female yellow-eyed penguins (Megadyptes antipodes). Behavioral Ecology and Sociobiology 55: 169–175.
Mayr, E. 1963. Animal species and evolution. Harvard Univ. Press, Cambridge. McGraw, K. J. & G. E. Hill. 2000. Differential effects of endoparasitism on the expression
of carotenoid- and melanin-based ornamental coloration. Proceedings of the Royal Society of London, Series B 267: 1525–153.
McGraw, K. J. 2006a. Mechanics of carotenoid-based coloration. Pp. 177242. En Hill G.E. & K.J. McGraw [eds.], Bird coloration, Vol. I: Mechanisms and measurements. Harvard University Press, Cambridge, Massachusetts.
McGraw, K. J. 2006b. Mechanics of melanin-based coloration. Pp. 243–294. En Hill G. E. & K. J. McGraw [eds.], Bird coloration, Vol. I: Mechanisms and measurements. Harvard University Press, Cambridge, Massachusetts.
Møller, A. P. 1987. Variation in badge size in male house sparrows Passer domesticus: evidence for status signalling. Animal Behaviour 35: 1637–1644.
Møller, A. P. 1988. Badge size in the house sparrow Passer domesticus. Behavioral Ecology and Sociobiology 22: 373–378.
Møller, A. P. & A. Pomiankowski. 1993. Why have birds got multiple sexual ornaments?. Behavioral Ecology and Sociobiology 32: 167-176.
Mundy, N. I. 2005. A window on the genetics of evolution: MC1R and plumage colouration in birds. Proceedings of the Royal Society of London B: Biological Sciences, 272 (1573): 1633–1640.
Norris, K. J. 1990. Female choice and the evolution of the conspicuous plumage coloration of monogamous male great tits. Behavioral Ecology and Sociobiology 26: 129–138.
Olson, V. A. & I. P. F. Owens. 1998. Costly sexual signals: are carotenoids rare, risky or required? Trends in Ecology and Evolution 13: 510–514.

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
80
Price, T. & A. Bontrager. 2001. Evolutionary genetics: the evolution of plumage patterns. Current Biology, 11: R405–R408.
Pryke, S. R., S. Andersson & M. J. Lawes. 2001. Sexual selection of multiple handicaps in the Redcollared Widowbird: female choice of tail length but not carotenoid display. Evolution 55: 1452–1463.
Rohwer, S. & F. C. Rohwer. 1978. Status signalling in Harris Sparrows: experimental deceptions achieved. Animal Behavior 26: 1012–1022.
Roulin, A. 1999. Nonrandom pairing by male barn owls (Tyto alba) with respect to a female plumage trait. Behavioral Ecology 10: 688–695.
Saino, N., M. Romano, D. Rubolini, R. Ambrosini, M. Caprioli, A. Milzani, A. Constanzo, G. Colombo, L. Canova & K. Wakamatsu. 2013. Viability is associated with melanin-based coloration in the barn swallow (Hirundo rustica). PloS one 8: e60426.
Senar, J. C. 1999. Plumage coloration as a signal of social status. Pp. 1669– 1686. En International Ornithological Congress (eds N.J. Adams & R. Slotow), Vol. 22, BirdLife South Africa, Durban, South Africa.
Senar, J. C. 2006. Color displays as intrasexual signals of aggression and dominance. Pp. 87 136. En Hill G.E y K.J. McGraw [eds.], Bird Coloration, Vol. II: Function and Evolution. Harvard University Press, Cambridge, Massachusetts.
Slatkin, M. 1987. Gene flow and the geographic structure of natural populations. Science 236: 787–792.
Slatkin, M. 1993. Isolation by distance in equilibrium and non-equilibrium populations. Evolution 47: 264–279.
Smith, M. F. & J. L. Patton. 1984. Dynamics of morphological differentiation: temporal impact of gene flow in pocket gophers populations. Evolution 38: 1079–1087.
Tarof, S., P. O. Dunn & L. A. Whittingham. 2005. Dual functions of a melanin-based ornament in the common yellowthroat. Proceedings of the Royal Society of London B 272: 1121–1127.
Taylor, S. S. & I. G. Jamieson. 2007. Factors affecting the survival of founding individuals in translocated New Zealand Saddlebacks Philesturnus carunculatus. Ibis 149: 783–791.
Theron, E., K. Hawkins, E. Bermingham, R. Ricklefs & N. Mundy. 2001. The molecular basis of an avian plumage polymorphism in the wild: a melanocortin-1-receptor point

VARIACIÓN GEOGRÁFICA DE LA MORFOLOGÍA
81
mutation is perfectly associated with the melanic plumage morph of the bananaquit, Coereba flaveola. Current Biology 11: 550–557.
Toon, A., J. Hughes, A. Baker & P. Mather. 2003. Discordance between morphology and genetic structure among three plumage forms of the Australian Magpie. Emu 103: 337–343.
Zahavi, A. 1975. Mate selection - a selection for a handicap. Journal of Theoretical Biology 53: 205–214.

VARIACIÓN GENÉTICA
82
CAPÍTULO 5
VARIACIÓN GENÉTICA
5.1. Introducción
5.1.1. Conservación de la variabilidad genética
Frankel (1974) remarca que la conservación de la variabilidad genética es indispensable para garantizar las condiciones que aseguren la continua evolución de las especies. Conservar la diversidad actual ayuda a asegurar el potencial adaptativo futuro, es decir, la habilidad de una especie a adaptarse a ambientes variables (Mayr 1963). Actualmente, se reconoce que mantener la diversidad genética de especies en peligro de extinción es crítico para asegurar su supervivencia a largo plazo (Hedrick & Kalinowski 2000, Frankham et al. 2002, Spielman et al. 2004). De esta manera, los estudios genéticos contribuyen a definir grupos con prioridad de conservación para mantener la mayor proporción de la variación genética original (Ryan 2006).
5.1.2. Efecto de la fragmentación del hábitat sobre la diversidad genética
La fragmentación del hábitat es el proceso de subdivisión de un hábitat continuo en partes más pequeñas. La causa más importante de la fragmentación del hábitat es la expansión e intensificación de la utilización del suelo para actividades agropecuarias y asentamientos humanos (Burgess & Sharpe 1981). La fragmentación del hábitat se considera la principal amenaza para las aves (Owens & Bennett 2000, Fischer & Lindenmayer 2007, Hoffman et al. 2010, Rands et al. 2010). Usualmente resulta en la reducción del tamaño de las poblaciones y en un aislamiento de las mismas. Si la pérdida de hábitat se agrava con el paso del tiempo, las poblaciones quedan cada vez más separadas, disminuyen el flujo génico y la tasa de recolonización y aumenta la tasa de extinción local (Sutherland 1998).
Una metapoblación se define como un grupo de poblaciones de una especie que ocupan parches de hábitat similar separados por hábitat inadecuado (Levins 1970). Se espera que las poblaciones que quedan en parches más grandes contengan un mayor número de individuos y sean menos propensas a desaparecer. También se espera que parches de hábitat adecuado cercanos a parches ocupados por la especie tengan mayor probabilidad de ser recolonizados (Allendorf & Luikart 2007). El aislamiento entre poblaciones generado por la fragmentación del hábitat hace que se comporten como unidades demográficas independientes ocasionando un aumento de la diferenciación genética entre ellas y una pérdida de la diversidad genética cuando son pequeñas (Frankham 1995).

VARIACIÓN GENÉTICA
83
5.1.3. Los riesgos de las poblaciones pequeñas
Las poblaciones pequeñas y aisladas tienen una mayor probabilidad de extinción (Shaffer 1981, Gilpin & Soule 1986, Lande 1993). Son más susceptibles a variaciones que ocurren al azar, que pueden estar dadas por la estocasticidad ambiental (es decir, la variación al azar en factores abióticos o bióticos, Lande 1993), la estocasticidad demográfica (la variación al azar en la proporción de sexos o en las tasas de mortalidad o reproducción, Gabriel & Burger 1992, Lande 1993) y a factores genéticos tales como la acumulación de mutaciones deletéras, la deriva genética y/o la endogamia (Lande 1994, 1995, Lynch et al. 1995).
La estocasticidad ambiental es una fuerza guiada por fluctuaciones en el ambiente físico (factores ambientales como tormentas o catástrofes) o fluctuaciones bióticas (cambios en las densidades de los parásitos, predadores y/o competidores, Sutherland 1998). La estocasticidad demográfica, por otro lado, es la variación temporal de rasgos demográficos, tales como la razón de sexos y las tasas de nacimientos y mortalidad, que gobiernan la tasa de crecimiento de la población. En una población grande, la variación de estos rasgos genera un menor impacto. Sin embargo, en una población pequeña, las variaciones inusuales de algunos rasgos demográficos pueden ocasionar que la población entera se vea amenazada (Sutherland 1998).
La estocasticidad genética se refleja principalmente en dos mecanismos: la depresión por endogamia y la deriva génica. La primera ocurre cuando individuos emparentados se aparean, aumentando la probabilidad de homocigotas en la descendencia (Allendorf & Luikart 2007). De esta manera, un efecto de la endogamia en las poblaciones pequeñas es la pérdida de variabilidad genética. Además, considerando que algunos alelos recesivos son deletéreos (usualmente enmascarados con un alelo dominante), un aumento en la homocigosis puede incrementar la posibilidad de que la descendencia exprese rasgos recesivos y tenga un fitness bajo. La deriva génica, por otro lado, genera una pérdida o fijación de alelos por azar reduciendo la variación genética y, por lo tanto, el potencial adaptativo de la población (Keller & Waller 2002). En las poblaciones pequeñas, las frecuencias alélicas pueden cambiar de una generación a otra simplemente debido a azar. Ambos procesos dan como resultado una pérdida de la variabilidad genética. Una reducción del fitness individual y de la capacidad de la población a adaptarse a cambios puede conducir, en última instancia, a la extinción local y global (Gilpin & Soulé 1986, Hedrick & Kalinowski 2000, Höglund 2009).
5.1.4. Estructuración de la variabilidad genética
La variación genética existente en una especie es el resultado de varios procesos que actúan a lo largo de su evolución: selección, migración, mutación y deriva genética. Estos procesos tienen distintos efectos sobre la variabilidad. Mientras que la mutación generalmente

VARIACIÓN GENÉTICA
84
incrementa la cantidad de variación, la deriva genética la reduce, y las combinaciones entre dos o más factores pueden generar patrones diversos de variabilidad (Hedrick 2005). La acción de estos procesos en forma independiente en poblaciones aisladas puede redistribuir la variabilidad genética entre las poblaciones espacialmente estructuradas en tan sólo unas pocas generaciones (Harrison & Hastings 1996, Caizergues et al. 2003).
La información sobre la variabilidad genética de las poblaciones de una especie en peligro de extinción es relevante para su conservación (Frankham 2010, Haig et al. 2011). La protección de poblaciones genéticamente distintas ayuda a mantener altos niveles de variabilidad minimizando los riesgos de extinción (Waples 1991). Mientras que para algunas especies de aves amenazadas no se han encontrado evidencias de diferenciación genética asociadas a la fragmentación del hábitat (Ellegren et al. 1999, Bates 2000, Mylecraine et al. 2008, Di Giácomo et al. 2014), posiblemente debido a la distancia de los parches y a la alta capacidad de dispersión de las especies, para muchas otras se ha observado que la fragmentación y consecuente aislamiento de poblaciones puede, en un tiempo relativamente corto, producir un aumento en la diferenciación genética inter-poblacional y una disminución de los niveles de variación local. Por ejemplo, un estudio que analizó la variabilidad genética de cuatro poblaciones naturales de Ñandú Común (Rhea americana) en una de las zonas pampeanas más afectadas por la fragmentación, encontró una importante diferenciación genética entre las mismas (Bouzat 2001). El Gorrión Zacatero Serrano (Xenospiza baileyi) es otra especie de ave amenazada con poblaciones actuales fragmentadas que mostraron estar genéticamente diferenciaciadas, probablemente como consecuencia de su incapacidad de dispersar a través de zonas de hábitat inadecuado (Oliveras de Ita et al. 2012).
El tiempo necesario para detectar las consecuencias de los procesos de fragmentación y pérdida de hábitat sobre la variabilidad genética puede ser de sólo unas pocas generaciones. Por ejemplo, un estudio de fragmentación en la región del Cerrado en Brasil y su asociación con la estructuración genética de las poblaciones remanentes de la Cotorra de Pfrimer (Pyrrhura pfrimeri) sugieren un retraso de 35 años entre la deforestación y su efecto sobre la estructuración genética (Miller et al. 2013). El Carpintero de Cresta Roja (Picoides borealis) era una especie abundante en Norteamérica y en menos de 200 años las poblaciones comenzaron a fragmentarse y a evidenciar gran diferenciación genética (Stangel et al. 1992, Haig et al. 1994). El Urogallo (Tetrao urogallus), por otro lado, tiene una distribución muy fragmentada en algunas zonas de Europa debido al cambio del uso del suelo, pero una distribución continua a lo largo del bosque boreal paleártico. Una investigación documenta que la diferenciación genética aumenta gradualmente desde las zonas de conectividad hacia las que se encuentran en aislamiento (Segelbacher et al. 2003).

VARIACIÓN GENÉTICA
85
5.1.5. Objetivo
El objetivo de este capítulo es conocer cómo la diversidad genética intraespecífica se distribuye geográficamente en las poblaciones naturales del Cardenal Amarillo. Como consecuencia de la fragmentación del ambiente, las poblaciones remanentes de Cardenal Amarillo podrían representar unidades demográficas independientes generando una estructuración genética. Los objetivos específicos son evaluar la variabilidad genética de las poblaciones a partir del uso de marcadores moleculares mitocondriales y nucleares y determinar la diferenciación genética interpoblacional para así identificar las unidades de conservación para esta especie. Ambos marcadores moleculares difieren en sus tasas de mutación y modo de herencia y proporcionan información esencial acerca de la historia de las poblaciones y su demografía (Wayne & Morin 2004).
5.2. Metodología
5.2.1. Colección de datos
Se capturaron 90 individuos adultos durante las temporadas reproductivas 2011 – 2014 siguiendo la metodología descripta en el capítulo 2, que se basa en la utilización de redes de niebla. Las capturas se hicieron en cuatro regiones que representan la mayor parte de la distribución actual de la especie en Argentina (Figura 1.8): 1) Corrientes, donde la captura se realizó en dos sitios, Rincón del Socorro (C1: 28°39'22"S 57°26'4"W) y Estancia La Paz (C2: 29°12'6"S 58°15'41"W); 2) San Luis, en la Reserva Provincial Quebracho de la Legua (SL: 32°25'10"S 66°53'27"W); 3) La Pampa, en los sitios de General Acha (LP1: 37°42'52"S 64°46'14"W), Luro (LP2: 36°38'6"S 64°17'48"W) y Luan Toro (LP3: 36°12'15"S 65°05'58"W); y 4) Río Negro, donde se capturaron individuos en las zonas de General Conesa (RN1: 40°3'9"S 64°8'38"W) y Guardia Mitre (RN2: 40°24'28"S 63°40'54"W). Las áreas de captura dentro de cada provincia están separadas por aproximadamente 100 km. Se tomaron muestras de sangre de los individuos capturados (aproximadamente 50 μl) mediante punción de la vena braquial con una aguja 30G. La sangre fue conservada en 500 μl de buffer de lisis (100mM TRIS, 100mM EDTA, 10mM NaCl, 2% SDS) a temperatura ambiente hasta ser analizada. Además, se obtuvieron plumas de 20 individuos provenientes de la República Oriental del Uruguay mantenidos en cautiverio. Estos individuos se encuentran en cautiverio bajo el Programa para la Cría y Reintroducción del Cardenal Amarillo y a cada uno de ellos se le extrajeron de 5 a 8 plumas para los posteriores análisis genéticos.

VARIACIÓN GENÉTICA
86
5.2.2. Análisis Genéticos
La extracción de ADN a partir de las muestras de sangre y plumas se realizó utilizando un kit de extracción de Qiagen (Hilden, Alemania). Los estudios genéticos se basaron en un fragmento de 736 pb de la región control de ADN mitocondrial (ADNmt) y en 10 marcadores de tipo microsatélite.
Para la amplificación del fragmento de ADNmt, se trabajó con 50-100 ng de ADN genómico como templado, 0.2 µM de cebadores (LCR3 y H1248, Tarr 1995), 0.2 mM de cada dNTP, 2.5 µl de10X PCR buffer (Invitrogen), 2.25 mM MgCl2 y 0.1 U Taq DNA Polymerasa (Invitrogen). El volumen final fue de 25 μl. Las reacciones en cadena de la polimerasa (PCR) se llevaron a cabo en un termociclador marca Labnet, con un ciclo de 2 min a 94°C; luego 35 ciclos de 30 s a 94°C, 30 s a 55°C y 60 s a 72°C, y una extensión final de 8 min a 72°C. Los productos de amplificación fueron secuenciados con ABI Big DyeTM Terminator Chemistry (Applied Biosystems) en el secuenciador ABI 3130 XL (Applied Biosystems, Foster City, CA, USA) del Departamento de Ecología, Genética y Evolución de la Facultad de Ciencias Exactas y Naturales de la Universidad de Buenos Aires.
Las secuencias fueron editadas y alineadas utilizando el programa Bioedit v7.2.0 (Hall 1999). Se obtuvieron medidas de diversidad de este marcador a nivel poblacional: número de haplotipos, sitios polimórficos, número de alelos privados (definidos como haplotipos que ocurrieron en una sola región), la probabilidad de que dos haplotipos al azar sean distintos (diversidad haplotípica H, Nei 1987) y el número promedio de diferencias nucleotídicas por sitio entre dos haplotipos al azar (diversidad nucleotídica Pi, Nei & Li 1979). Estos parámetros fueron calculados utilizando el programa DNAsp 05.10.01 (Librado & Rozas 2009).
Para la amplificación de diez loci microsatélite se utilizaron cebadores diseñados para esta especie (Martins-Ferreira et al. 2010). Se llevaron a cabo PCRs multiplex, agrupando los cebadores en seis sets (set I: C2 y F2, set II: H9, set III: C8 y G10, set IV: H7, set V: E2 y F12, set VI: H6 y H12) optimizando la combinación de colores de la marcación fluorescente (FAM, VIC, PET y NED) para los pesos moleculares de los loci. Las reacciones de amplificación se llevaron a cabo en un volumen final de 25 μL usando de 50 a 100 ng de ADN genómico como templado, 0.2 µM de cada cebador ―forward‖ (marcado con fluorescencia FAM, VIC, PET o NED en el extremo 5´), 0.2 µM de cada cebador ―reverse‖, 0.2 mM de cada dNTP, 2.5 µl de 10X PCR buffer (Invitrogen), 2.25 mM MgCl2 y 0.1 U Taq DNA Polymerase (Invitrogen). Las reacciones de amplificación multiplex se realizaron con el termociclador mencionado previamente con un ciclo de 5 min a 94°C; luego 40 ciclos de 30 s a 94°C, 60 s a 55°C y 45 s a 72°C, y una extensión final de 10 min a 72°C.

VARIACIÓN GENÉTICA
87
Los productos de amplificación con marcación fluorescente fueron dimensionados utilizando el mismo secuenciador mencionado más arriba y los tamaños de los alelos se determinaron con Peak Scanner v1.0 (Applied Biosystems). Los genotipos se analizaron con Microchecker v2.2.3 (Oosterhout et al. 2004) para detectar la presencia de alelos nulos y errores de genotipado. El programa GenAlEx 6 (Peakall & Smouse 2006) se utilizó para calcular las frecuencias alélicas y determinar los alelos privados, mientras que la riqueza alélica se calculó con FSTAT 2.9.3.2 (Goudet 2002). Las heterocigosis observadas y esperadas se calcularon con Arlequin v3.5.1 (Excoffier & Lischer 2010), que también fue utilizado para analizar las desviaciones del equilibrio Hardy-Weinberg (HW) y el desequilibrio por ligamiento. La significancia de las pruebas de desequilibrio por ligamiento se evaluó luego de aplicar una corrección de Bonferroni para comparaciones múltiples (Rice 1989). 5.2.3. Estructuración poblacional genética
Para ambos marcadores, la divergencia genética entre regiones y localidades dentro de cada región (subregiones) se analizó mediante un análisis molecular de la varianza (AMOVA; Excoffier et al., 1992) con Arlequin v3.5.1.
Además, para los marcadores microsatélite se estimó el número de poblaciones (K) mediante un análisis de asignación bayesiano implementado en STRUCTURE v2.3.3 (Prichard et al. 2000). Tomando en cuenta el genotipo multi-locus de cada individuo, el programa asigna probabilísticamente a cada individuo a uno o más grupos genéticos. En este análisis, K es el número de grupos o poblaciones y puede tomar sucesivos valores desde K = 1 hasta un número definido por el usuario. Al final del análisis se determina el valor de K que tiene mayor probabilidad dados los datos (Falush et al. 2003). Se utilizó el modelo que permite ―admixture‖ (i.e. mezcla) entre los grupos genéticos asumiendo frecuencias alélicas correlacionadas para K entre 1 y 6. De esta manera, se permitió que el genoma de un individuo provenga de distintos grupos (mezcla). Esto es recomendable cuando las dimensiones del flujo génico y de la recombinación son desconocidas (Falush et al. 2003). Las simulaciones se llevaron a cabo con un burn-in de 200.000 pasos y un período de recopilación de datos de 1.500.000 iteraciones. Cada simulación se repitió 10 veces. No se proporcionó al programa información previa sobre la población de origen de los individuos. Para seleccionar el número más probable de grupos se consideró la simulación con el valor de probabilidad más alto y el valor máximo del estadístico ΔK según el método de Evanno et al. (2005). Los valores de probabilidad y de ΔK (Earl & von Holdt 2012) se graficaron usando el programa en línea STRUCTURE Harvester y el programa CLUMPP (Jakobsson & Rosenberg 2007) para promediar las matrices de los coeficientes de pertenencia estimados de las múltiples corridas de STRUCTURE utilizando el algoritmo FullSearch. La salida de CLUMPP se visualizó utilizando el programa

VARIACIÓN GENÉTICA
88
DISTRUCT (Rosenberg 2004). El K elegido representa el número más probable de unidades panmícticas que integran la muestra.
Para evaluar si las diferencias genéticas están correlacionadas con la distancia geográfica, se realizó una prueba de Mantel para ambos marcadores genéticos utilizando los valores de FST como medida de distancia genética entre poblaciones y la distancia geográfica (en km). La prueba se realizó con GenAlEx 6 empleando 9999 permutaciones. Las distancias geográficas entre poblaciones adyacentes fueron medidas linealmente con Google Earth y las distancias entre las poblaciones más distantes se calcularon como la suma de las distancias lineales medidas (por ejemplo, la distancia RN_C = (RN_LP) + (LP_SL) + (SL_C)).
5.3. Resultados
5.3.1. Estadística descriptiva
Se obtuvieron secuencias de 736 pb de la región de control del ADNmt de 94 de los 110 cardenales amarillos estudiados. Se encontraron 11 sitios polimórficos en 13 haplotipos diferentes (Tabla 5.1). Dos haplotipos fueron frecuentes, H1 (observado en el 36% de las muestras) y H5 (en 41%), mientras que los otros haplotipos se observaron en frecuencias bajas (Tabla 5.1).

VARIACIÓN GENÉTICA
89
Tabla 5.1. Sitios variables de la región control del ADNmt para los 13 haplotipos encontrados en las 94 muestras de Cardenal Amarillo analizadas a lo largo de su distribución en Argentina y Uruguay.
18 46 163 205 442 454 459 649 658 664 705 715 Frequency
(%)
H1 A A A T A C A A A T G T 36.1
H2 . . . . . . . . G . . . 4.2
H3 . G . . . . . T . . . . 1.1
H4 . . . . . . . . . . A . 1.1
H5 . . . . . . . . . C . . 41.4
H6 . . . . . . G . . . . . 7.4
H7 . . . . . . . . . A . . 1.1
H8 . . . . . . . . . . A C 1.1
H9 . . C . . . . . . C . . 2.1
H10 . . . . . T . . . C . . 1.1
H11 G . . . . . . . . C . . 1.1
H12 . . . C . . . . . . A . 1.1
H13 . . . . G . . . . C . . 1.1

VARIACIÓN GENÉTICA
90
Tabla 5.2. Medidas de diversidad para el marcador mitocondrial.
n Diversidad haplotípica
(H)
Diversidad nucleotídica
(Pi) %
Haplotipos más comunes
Número y frequencia de haplotipos privados
U 23 0.62 1.27 H1: 61% 3 (13%)
C 25 0.36 0.52 H1: 80% 2 (8% )
SL 6 0.33 0.45 H5: 83% 1 (17%)
LP 17 0.49 1.16 H5: 71% 2 (12%)
RN 23 0.37 0.91 H5: 79% 0
U = población de Uruguay, C = población de Corrientes, SL = población de San Luis, LP = población de La Pampa y RN = población de Río Negro.
Los índices de diversidad variaron entre regiones geográficas (Tabla 5.2). Las poblaciones de Uruguay (U) y La Pampa (LP) mostraron los valores más altos de diversidad y las de San Luis (SL) y Corrientes (C) los más bajos. Todas las regiones, a excepción de Río Negro (RN), mostraron haplotipos privados (U: H10, H11 y H12; C: H3 y H4; SL: H13 y LP: H7 y H8) (Figura 5.1).

VARIACIÓN GENÉTICA
91
Figura 5.1. Mapa con la distribución histórica del Cardenal Amarillo (en gris) y las localidades de muestreo en las distintas provincias: Uruguay (U), Corrientes (C), San Luis (SL), La Pampa (LP) and Río Negro (RN). Los puntos blancos indican diferentes áreas de muestreo. Los gráficos muestran la distribución de las frecuencias de haplotipos en cada población: H1 = amarillo, H2 = rojo, H3 = negro, H4 = rosa, H5 = verde, H6 = naranja claro, H7 = rojo oscuro, H8 = luz amarilla, H9 = violeta, H10 = luz azul, H11 = naranja, H12 = violeta claro y H13 = azul.
Se lograron genotipar 102 de los 110 cardenales amarillos. Se encontraron de 5 a 17 alelos por locus microsatélite. No hubo evidencia de deserción de grandes alelos, errores de genotipado o alelos nulos para ningún microsatélite. Las frecuencias alélicas de los loci

VARIACIÓN GENÉTICA
92
microsatélites no se desviaron del equilibrio HW y no hubo evidencia de desequilibrio por ligamiento entre ningún par de loci. El mayor número de alelos privados se encontró en LP (N = 15, Tabla 5.3). A pesar de su número de muestras mayor, las poblaciones de U, C y RN presentaron solo de tres a cinco alelos privados. LP fue, en general, la población con mayores índices de diversidad (Tabla 5.3).
Tabla 5.3. Medidas de diversidad para los microsatélites.
n PA HO ± DE HE ± DE Na Ne AR
U 26 3 0.582 ± 0.089 0.639 ± 0.062 5.6 3.2 4.2
C 26 5 0.485 ± 0.082 0.661 ± 0.037 5.6 3.4 4.2
SL 7 4 0.643 ± 0.091 0.639 ± 0.081 4.8 3.8 4.9
LP 18 15 0.643 ± 0.072 0.748 ± 0.039 7.5 4.6 5.5
RN 25 5 0.500 ± 0.082 0.640 ± 0.051 6.5 3.1 4.4
TOTAL 102 32 0.571 ± 0.037 0.665 ± 0.025 6 3.6
n: número de muestras, PA: alelos privados, HO: heterocigosidad promedio observada, HE: heterocigosidad promedio esperada, Na: Número promedio de alelos en todos los loci, Ne: Número promedio de alelos efectivos en todos los loci, AR: riqueza alélica promedio (basado en el tamaño de muestra de SL, n = 7), DE: desvío estandar.
5.3.2. Estructuración poblacional genética
El análisis de varianza molecular (AMOVA) reveló diferencias significativas entre regiones (ADNmt ɸST= 0.27, p <0.05; microsatélites FST = 0.04, p = 0.004). El mayor porcentaje de variación se produjo dentro de las subregiones (ADNmt 66%, microsatélites 93%), mientras que la fracción más pequeña de la varianza fue debido a la divergencia entre las subregiones dentro de las regiones (ADNmt 7%, microsatélites 3%, Tabla 5.4). La variación entre subregiones dentro de las regiones no fue significativa para el ADNmt (FSC = 0.10, P = 0.078) y significativo para los datos de microsatélites (FSC = 0.04, P < 0.05, Tabla 5.4). Por esta razón, a continuación se investigan las diferencias entre las regiones para los datos de ADN mitocondrial, y también las diferencias entre subregiones dentro de las regiones para los microsatélites.

VARIACIÓN GENÉTICA
93
Tabla 5.4. Análisis molecular de la varianza para ADNmt y microsatélites. Regiones: provincias. Subregiones: localidades dentro de provincias.
Región Grados de libertad
Porcentaje de variación
Estadístico F
P
DNAmt
Entre regiones 4 26.54 0.27 <0.05
Entre subregiones dentro de regiones
4 7.28 0.10 0.078
Dentro de subregiones
85 66.18 0.34 <0.001
Microsatélites
Entre regiones 4 3.74 0.04 <0.05
Entre subregiones dentro de regiones
4 3.38 0.04
<0.05
Dentro de subregiones 195 92.88 0.07 <0.001
Las regiones SL, LP y RN no se diferenciaron en el ADNmt (Tabla 5.5) y por lo tanto, tres grupos pueden ser reconocidos (U, C, SL_LP_RN). Se encontró una fuerte diferenciación entre las poblaciones occidentales (SL_LP_RN) y las orientales (U y C) (ΦSTs = 0.16-0.70, p < 0.005, Tabla 5.5). H1 fue el haplotipo más frecuente en el extremo oriental de la distribución y no se observó en ninguna muestra del oeste, mientras que H5 fue el haplotipo más frecuente en el oeste y fue raro en el este (Figura 5.1, Tabla 5.2). La diferenciación entre U y C fue moderada pero significativa (ΦST = 0.1, p = 0.003).

VARIACIÓN GENÉTICA
94
Tabla 5.5. Índices de fijación (ΦST) por pares de poblaciones obtenidos a partir de un fragmento de 736 pb de la región control de ADNmt. Los valores significativos se resaltan en negrita.
U C SL LP RN
U
C 0.10
SL 0.34 0.70
LP 0.16 0.46 0.06
RN 0.25 0.56 0.04 -0.03
La información nuclear muestra una diferenciación moderada y significativa (valores FST entre pares de poblaciones desde 0.05 hasta 0.1, Tabla 5.6) entre las regiones del este y del oeste, y una diferenciación menor entre las poblaciones del oeste, SL, LP y RN. Las subregiones LP2 y LP3 no se diferenciaron de SL, mientras que LP1 no se diferenció de RN. Las poblaciones de La Pampa quedan así subdivididas en un grupo al norte y otro grupo al sur (Tabla 5.6, Figura 5.1).
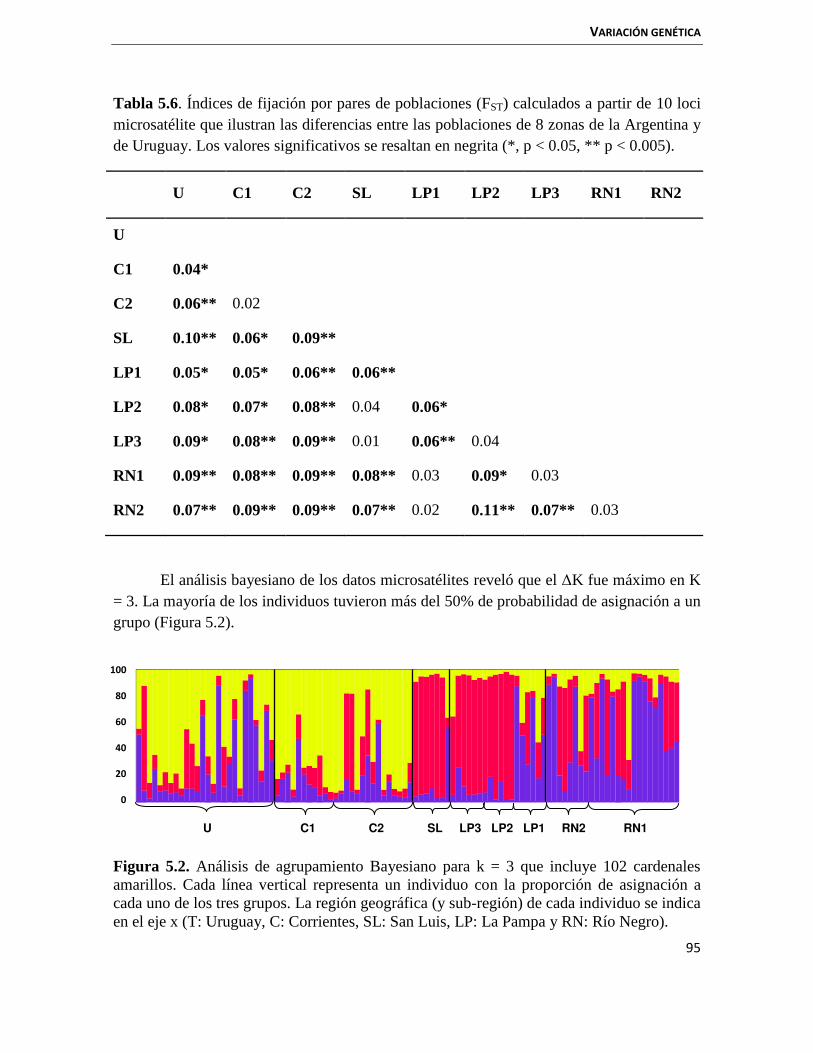
VARIACIÓN GENÉTICA
95
Tabla 5.6. Índices de fijación por pares de poblaciones (FST) calculados a partir de 10 loci microsatélite que ilustran las diferencias entre las poblaciones de 8 zonas de la Argentina y de Uruguay. Los valores significativos se resaltan en negrita (*, p < 0.05, ** p < 0.005).
U C1 C2 SL LP1 LP2 LP3 RN1 RN2
U
C1 0.04*
C2 0.06** 0.02
SL 0.10** 0.06* 0.09**
LP1 0.05* 0.05* 0.06** 0.06**
LP2 0.08* 0.07* 0.08** 0.04 0.06*
LP3 0.09* 0.08** 0.09** 0.01 0.06** 0.04
RN1 0.09** 0.08** 0.09** 0.08** 0.03 0.09* 0.03
RN2 0.07** 0.09** 0.09** 0.07** 0.02 0.11** 0.07** 0.03
El análisis bayesiano de los datos microsatélites reveló que el ΔK fue máximo en K = 3. La mayoría de los individuos tuvieron más del 50% de probabilidad de asignación a un grupo (Figura 5.2).
Figura 5.2. Análisis de agrupamiento Bayesiano para k = 3 que incluye 102 cardenales amarillos. Cada línea vertical representa un individuo con la proporción de asignación a cada uno de los tres grupos. La región geográfica (y sub-región) de cada individuo se indica en el eje x (T: Uruguay, C: Corrientes, SL: San Luis, LP: La Pampa y RN: Río Negro).
U C1 C2 SL LP3 LP2 LP1 RN2 RN1
100
80
60
40
20
0
0

VARIACIÓN GENÉTICA
96
No se encontró correlación significativa entre la distancia geográfica y las diferencias genéticas en los cardenales amarillos (prueba de Mantel para datos mitocondriales r = 0.505, p = 0.138, Figura 5.3; prueba de Mantel para microsatélites r = 0.507, p = 0.097, Figura 5.4).
Figura 5.3. Prueba de Mantel entre la distancia geográfica y genética para el marcador mitoncondrial. Los puntos representan los valores ΦST de a pares y las distancias geográficas entre las cinco poblaciones muestreadas.
Figura 5.4. Prueba de Mantel entre la distancia geográfica y genética para los microsatélites. Los puntos representan los valores FST de a pares y las distancias geográficas entre las cinco poblaciones muestreadas.

VARIACIÓN GENÉTICA
97
5.4. Discusión
Los resultados obtenidos mostraron diferencias en los índices de diversidad genética entre poblaciones. Para el ADNmt las poblaciones con mayor diversidad fueron Uruguay y La Pampa. San Luis y Corrientes mostraron los menores valores para los índices de variabilidad genética medidos. En general, los valores de diversidad haplotípica (H) fueron bajos para todas las poblaciones menos para Uruguay, siguiendo el criterio de Grant & Bowen (1998) que consideran valores altos cuando H 0.5. Una disminución en la variabilidad genética puede ser consecuencia de un cuello de botella o de un bajo número poblacional (Nei, Maruyama & Chakraborty 1974). Dado el reducido número de individuos que se observan en San Luis y la baja cantidad de localidades en los que se puede encontrar al Cardenal Amarillo en la provincia de Corrientes (Domínguez obs. pers.), lo más probable es que ambos factores estén afectando la diversidad genética encontrada en los cardenales. La reducción de las poblaciones por la fragmentación del hábitat y la extracción de individuos para el comercio ilegal generaron un cuello de botella que se refleja en una disminución de la variabilidad genética. Dada la persistencia de las amenazas, las poblaciones no se han recuperado del declive poblacional y aún presentan un bajo número de individuos. Es posible que aquellas poblaciones que presentan una mayor variabilidad genética tengan un mayor tamaño poblacional y no hayan sufrido un cuello de botella tan marcado.
Para los loci microsatélite, también se encontró mayor diversidad en la Provincia de La Pampa, aunque Uruguay mostró menor diversidad para estos marcadores. Si bien no se cuenta con datos de diversidad genética de otras especies de aves amenazadas de Argentina, los niveles encontrados son similares a aquellos presentes en varias especies de guacamayos con problemas de conservación en Brasil. Para el Arara Azul (Anodorhynchus hyacinthinus), el Guacamayo de Spix (Cyanopsitta spixii) y el Guacamayo de Lear (Anodorhynchus leari) los valores de heterocigosis y diversidad nucleotídica fueron similares a los encontrados en el cardenal (Presti et al. 2011, 2015). Los índices de diversidad encontrados también son similares a los de otros tráupidos, como la la tangara bandeada (Neothraupis fasciata, Corrêa et al. 2010).
Ambos marcadores genéticos mostraron estructuración genética en el Cardenal Amarillo. La diferenciación en el marcador nuclear es menor que en el mitocondrial posiblemente debido al tipo de herencia y número efectivo del marcador utilizado. El tamaño efectivo del ADN mitocondrial es menor que el de los microsatélites dado que por cada generación pasa una copia de ADN mitocondrial (por ser haploide y de herencia uniparental) mientras que del nuclear se transmiten dos copias a la siguiente generación. Eso hace que el efecto de la deriva génica pueda ser mayor sobre ADNmt por su menor tamaño efectivo resultando en una mayor diferenciación (Ballard & Whitlock 2004).

VARIACIÓN GENÉTICA
98
Alternativamente, el hecho de que la diferenciación nuclear sea menor que la diferenciación mitocondrial podría ser explicado si las hembras de Cardenal Amarillo son filopátricas y que los machos son el sexo que dispersa, como ocurre en otras especies de aves (Greenwood 1980, Dobson 1982, Dobson & Jones 1985).
Por otro lado, los microsatélites suelen presentar un polimorfismo mayor que el ADN mitocondrial haciéndolos más efectivos para estudios de estructuración poblacional en tiempos evolutivos más recientes (Estoup et al. 2002). Esto puede explicar por qué se encontró un menor número de grupos genéticamente diferenciados para el marcador mitocondrial que para el marcador nuclear. La diferenciación entre las regiones este y oeste en el marcador mitocondrial posiblemente refleje la historia evolutiva de la especie, que podría estar relacionada con una barrera geográfica dada por la cuenca Paraná-Paraguay. Esta barrera ha afectado la estructuración de la diversidad genética en numerosas especies de aves (Short 1975, Campagna et al. 2014). Asimismo, la diferenciación entre Uruguay y Corrientes podría estar influenciada por el río Uruguay. Por otro lado, la continuidad del hábitat en el pasado permitía el flujo génico entre las regiones del sur. Sin embargo, al ser analizadas con los marcadores microsatélite (que brindan información de la historia evolutiva más reciente) aparecen diferenciadas. Esta diferenciación podría ser consecuencia de la fragmentación del ambiente ocurrida en las últimas décadas, que llevó al aislamiento de las poblaciones.
No se encontraron evidencias de aislamiento por distancia, posiblemente debido a la falta de un gradiente ambiental continuo. En la genética de poblaciones, el modelo de aislamiento por distancia (Wright 1943) predice que, en ausencia de selección, hay una correlación significativa entre la distancia geográfica y la diferenciación genética. Bajo este modelo, la similitud genética entre las poblaciones disminuirá a medida que aumenta la distancia geográfica, como resultado del efecto limitante de la distancia geográfica sobre las tasas de flujo génico (Crow & Kimura 1970, Morton 1973, Relenthford 2004). Cualquier desvío a este modelo tiende a indicar fuerzas selectivas o factores históricos como determinantes de la estructura de la población. El flujo génico y la deriva genética suelen ser los principales procesos microevolutivos que configuran la diversidad genética neutra en distintos niveles jerarquicos (Hartl 2000). La diversidad genética del Cardenal Amarillo parece haber sido moldeada por barreras geográficas y por la fragmentación del hábitat, generando un aislamiento de las poblaciones. Un ambiente fragmentado tiende a limitar el flujo génico entre regiones y favorece la aparición de variantes nuevas por deriva génica en cada población aislada.
Los análisis bayesianos apoyan la diferenciación encontrada en el marcador mitocondrial para las poblaciones este y oeste de Argentina. Además, evidencian una diferenciación entre los individuos de la zona sur, que se dividen en dos unidades (Río Negro y sur de La Pampa y norte de La Pampa y San Luis). Sobre la base de la evidencia

VARIACIÓN GENÉTICA
99
colectada, la variabilidad genética del Cardenal Amarillo parece estar estructurada en tres grupos en Argentina. Los cardenales amarillos de la zona este (Corrientes) se comportan como una unidad panmíctica y las poblaciones del sur se subdividen en una población norte y otra población sur.
Nuestros resultados apoyan la delimitación de tres unidades de manejo dentro de la Argentina para el Cardenal Amarillo. Esta información es crucial para la planificación de acciones de conservación tendientes a mantener la mayor proporción de la variabilidad genética existente. Por otro lado, la diferenciación genética entre las regiones permitirá asignar la procedencia de individuos incautados y planificar su liberación en su lugar de origen, siempre que se cumplan las condiciones sanitarias de los mismos. Esta herramienta es sumamente valiosa y ha sido utilizada en otros casos de aves y tortugas que sufren del tráfico ilegal de animales silvestres (Faria et al. 2008, Schwartz & Karl 2008, Fernandes & Caparroz 2013, Presti et al. 2015).
5.5. Referencias bibliográficas
Allendorf, F. W. & G. Luikart. 2007. Conservation and the Genetics of Populations. Blackwell Publishing, Oxford, Estados Unidos.
Ballard, J. W. O., & M. C. Whitlock. 2004. The incomplete natural history of mitochondria. Molecular ecology 13: 729–744.
Bates, J. M. 2000. Allozymic genetic structure and natural habitat fragmentation: data for five species of Amazonian forest birds. Condor 102: 770–783.
Bouzat, J. L. 2001. The population genetic structure of the Greater Rhea (Rhea americana) in an agricultural landscape. Biological Conservation: 99: 277–284.
Burgess, R. L. & D. M. Sharpe. (eds) 1981. Forest island dynamics in man-dominated landscapes. Springer, New York.
Campagna, L., C. Kopuchian , P. L. Tubaro & S. C. Lougheed. 2014. Secondary contact followed by gene flow between divergent mitochondrial lineages of a widespread Neotropical songbird (Zonotrichia capensis). Biological Journal of the Linnean Society 111: 863–868.
Caizergues, A., O. Ratti, P. Helle, L. Rotelli, L. Ellison & J. Rasplus. 2003. Population genetic structure of male black grouse (Tetrao tetrix L.) in fragmented vs. continuous landscapes. Molecular Ecology 12: 2297–2305.

VARIACIÓN GENÉTICA
100
Corrêa, C. L., R. G. Collevatti & R. Caparroz. 2010. Isolation and characterization of microsatellite loci for Neothraupis fasciata, (Emberizidae, Passeriformes) with widely cross amplification in neotropical passerines. Journal of heredity 101 385–389.
Crow, J. F. & M. Kimura. 1970. An introduction to population genetics theory. Minneapolis, MN: Burgess.
Dobson, F. 1982. Competition for mates and predominant juvenile male dispersal in mammals. Animal Behaviour 30: 1183–1192.
Dobson, F. & W. Jones. 1985. Multiple causes of dispersal. American Naturalist 126: 885–58.
Ellegren, H, Carlson A, Stenberg I. 1999. Genetic structure and variability of White-backed Woodpecker (Dendrocopos leucotos) populations in northern Europe. Hereditas 130: 291–299.
Evanno, G., S. Regnaut & J. Goudet. 2005. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Molecular Ecology 14:2611–2620.
Excoffier, L., P. E. Smouse & J. M. Quattro. 1992. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 131: 479–491.
Excoffier, L. & H. E. Lischer. 2010. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Molecular ecology resources 10: 564–567.
Faria, P., N. Guedes, C. Yamashita, P. Martuscelli & C. Miyaki. 2008. Genetic variation and population structure of the endangered Hyacinth Macaw (Anodorhynchus hyacinthinus): implications for conservation. Biodiversity Conservation 17: 765–779.
Fernandes, G. A. & R. Caparroz. 2013. DNA sequence analysis to guide the release of blue and yellow macaws (Ara ararauna, Psittaciformes, Aves) from the illegal trade back to the wild. Molecular Biolgoy Reports 40: 2757–2762.
Frankham, R. 1995. Conservation genetics. Annual review of genetics 29: 305–327.
Frankham, R., J. D. Ballou & D. A. Briscoe. 2002. Introduction to Conservation Genetics. Cambridge University Press, Cambridge.
Frankham, R., J. D. Ballou, & D. A. Briscoe. 2010. Introduction to conservation genetics, 2nd ed. Cambridge, UK: Cambridge University Press.
Frankel, O. H. 1974. Genetic conservation: our evolutionary esponsibility. Genetics 78: 53–65.

VARIACIÓN GENÉTICA
101
Gabriel, W. & R. Burger. 1992. Survival of small populations under demographic stochasticity. Theoretical Population Biology 41: 44–71.
Gilpin, M. E. & M. E. Soule. 1986. Minimum viable populations: processes of species extinction. Pp. 1934. En M. E. Soule (editor) Conservation biology: the science of scarcity and diversity. Sinauer Associates, Sunderland, Mass.
Goudet, J. 2002. fstat (Version 2.9.3.2): A Program to Estimate and Test Gene Diversities and Fixation Indices. Descargado de http://www.unil.ch/izea/softwares/fstat.html.
Greenwood, P. 1980. Mating systems, philopatry and dispersal in birds and mammals. Animal Behaviour 28: 1140–1162.
Haig, S. M., W. M. Bronaugh, R. Crowhurst, J. D'Elia, C. A. Eagles-Smith, C. Epps, B. Knaus, M. P. Miller, M. L. Moses, S. Oyler-McCance, W. D. Robinson & B. Sidlauskas. 2011. Genetic applications in avian conservation. Auk 128: 205–229.
Hall, T. A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 41: 95–98.
Harrison, S. & A. Hastings. 1996. Genetic and evolutionary consequences of metapopulation structure. Trends in Ecology & Evolution 11: 180–183.
Hartl, D. L. 2000. A Primer of Population Genetics. 3rd ed. Sunderland (MA): Sinauer Associates.
Hedrick, P. & S. T. Kalinowski. 2000. Inbreeding depression in conservation biology. Annual Review of Ecology, Evolution, and Systematics. 31: 139–162.
Hedrick, P. W. 2005. Genetics of populations, 3rd Ed. Jones and Bartlett, Boston, MA, Estados Unidos.
Höglund, J. 2009. Evolutionary conservation genetics. Oxford, UK : Oxford University Press.
Keller, L. F. & D. M. Waller. 2002. Inbreeding effects in wild populations. Trends in Ecology & Evolution 17: 230–241.
Lande, R. 1993. Risks of population extinction from demographic and environmental stochasticity and random catastrophes. American Naturalist 142: 911-927.
Lande, R. 1994. Risk of population extinction from fixation of new deleterious mutations. Evolution 48: 1460-1469.
Lande, R. 1995. Mutation and conservation. Conservation Biology 9: 782-791.
Lynch, M., Conery J. & Burger R. 1995. Mutation accumulation and the extinction of small populations. American Naturalist 146: 489-518.

VARIACIÓN GENÉTICA
102
Martins-Ferreira, C., Haddrath O., Baker A. J. & Freitas T. R. O. 2010. Isolation and characterization of 10 microsatellite loci in the Yellow Cardinal Gubernatrix cristata. Molecular Ecology Resources 10: 751-754.
Mayr, E. 1963. Animal Species and Ecolution. Harvard University Press, Massachusetts.
Mi ller, M. P., C. A. Bianchi, T. D. Mullins & S. M. Haig. 2013. Associations between forest fragmentation patterns and genetic structure in Pfrimer‘s Parakeet (Pyrrhura pfrimeri), an endangered endemic to central Brazil‘s dry forests. Conservation Genetics 14: 333–343.
Morin, P. A., G. Luikart & R. K. Wayne. 2004. SNPs in ecology, evolution and conservation. Trends in Ecology & Evolution 19: 208–216.
Morton, N. E. 1973. Isolation by distance. Pp. 76–79. En: Morton N.E., ed. Genetic structure of populations. Honolulu: University Press of Hawaii.
Mylecraine, K. A, N. L. Bulgin, H. L. Gibbs, P. D. Vickery & D. W. Perkins. 2008. Limited genetic structure and evidence for dispersal among populations of the endangered Florida grasshopper sparrow, Ammodramus savannarum floridanus. Conservation Genetics 9: 1633–1638.
Nei, M., T. Maruyama & R. Chakraborty. 1975. The bottleneck effect and genetic variability in populations. Evolution 29: 1–10.
Nei, M. & W. H. Li. 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proceedings of the National Academy of Sciences 76: 5269–5273.
Nei, M. 1987. Molecular evolutionary genetics. Columbia University Press, New York.
Oliveras de Ita, A., K. Oyama, T. B. Smith, R. K. Wayne & B. Mila. 2012. Genetic evidence for recent range fragmentation and severely restricted dispersal in the critically endangered Sierra Madre Sparrow, Xenospiza baileyi. Conservation Genetics 13: 283 – 291.
Oosterhout, C., Hutchinson, W. F., Wills, D. P., & Shipley, P. 2004. MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data. Molecular Ecology Notes 4: 535–538.
Presti, F. T., A. R. Oliveira-Marques, R. Caparroz, C. Biondo & C. Y. Miyaki. 2011. Comparative analysis of microsatellite variability in five macaw species (Psittaciformes, Psittacidae): Application for conservation. Genetics and molecular biology, 34: 348–352.
Presti, F. T., N. M. Guedes, P. T. Antas & C. Y. Miyaki. 2015. Population Genetic Structure in Hyacinth Macaws (Anodorhynchus hyacinthinus) and Identification of the Probable Origin of Confiscated Individuals. Journal of Heredity 106: 491–502.

VARIACIÓN GENÉTICA
103
Pritchard, J. K., M. Stephens & P. Donnelly 2000. Inference of population structure using multilocus genotype data. Genetics 155: 945–959.
Relenthford, J. H. 2004. Global Patterns of Isolation by Distance Based on Genetic and Morphological Data. Human Biology 76: 499–513.
Rice, W. R. 1989. Analyzing tables of statistical tests. Evolution 43: 223–225.
Ryan, J.S. 2006. The Role of Culture in Conservation Planning for Small or Endangered Populations. Conservation Biology. 20: 1321–1324.
Schwartz, T. S. & S. A. Karl. 2008. Population genetic assignment of confiscated gopher tortoises. The Journal of Wildlife Management 72: 254–259.
Segelbacher, G., Höglund, J., & Storch, I. 2003. From connectivity to isolation: genetic consequences of population fragmentation in capercaillie across Europe. Molecular Ecology 12: 1773–1780.
Shaffer, M. L. 1981. Minimum population sizes for species conservation. Bioscience 31: 131–134.
Short, L. L. 1975. A zoogeographic analysis of the South American chaco avifauna. Bulletin of the American Museum of Natural History 154: 3.
Spielman, D., B. W. Brook & R. Frankham. 2004. Most species are not driven to extinction before genetic factors impact them. Proceedings of the National Academy of Sciences 101: 15261–15264.
Stangel, P. W., Lennartz M. R., Smith M. H. 1992. Genetic variation and population structure of red-cockaded woodpeckers. Conservation Biology 6: 283–292.
Sutherland, W. J. 1998. The importance of behavioural studies in conservation biology. Animal Behaviour 56: 801–809.
Tarr, C. L. 1995. Primers for amplification and determination of mitochondrial control region sequences in oscine passerines. Molecular Ecology 4: 527–529.
Waples, R. S. 1991. Genetic methods for estimating the effective size of Cetacean populations. Report of the International Whaling Commission Special Issue 13: 279–300.
Wright, S. 1943. Isolation by distance. Genetics 28: 114.

HIBRIDACIÓN CON LA DIUCA COMÚN
104
CAPÍTULO 6
HIBRIDACIÓN CON LA DIUCA COMÚN
6.1. Introducción
La hibridación interespecífica con descendencia híbrida es un evento que ocurre en menos del 10% de las aves en estado silvestre (Short 1985, Grant & Grant 1992). La hibridación entre especies genera un impacto negativo sobre las especies parentales (Allendorf & Luikart 2007). Por un lado, disminuye el reclutamiento, ya que parte de los individuos dejarán de reproducirse con individuos conespecíficos, y por otro lado, un mayor ―fitness‖ en los híbridos podría generar un desplazamiento de las especies parentales (Allendorf & Luikart 2007). En especies amenazadas, el impacto de la hibridación es aún más crítico al contribuir a la disminución del tamaño poblacional (Carroll & Fox 2008). Algunos ejemplos de especies con problemas de conservación cuya integridad se encontró amenazada por eventos de hibridación son el Pato Zambullidor Cabeza Blanca (Oxyura leucocephala) que en Europa hibrida con el Pato Zambullidor Grande (Oxyura jamaicensis, Peterson et al. 1993) y el Terito Real Negro de Nueva Zelanda (Himantopus novaezelandiae) que hibrida con el Terito Real Común (Himantopus himantopus, Norris & Pain 2002). En dos especies de Macaes actualmente extintas el componente de hibridación pudo haber jugado un rol importante en las últimas etapas. El Macá del Lago Alaotra (Tachybaptus rufolavatus) era endémico de Madagascar y se cree que la hibridación con T. ruficollis, un invasor proveniente de África, pudo haber sido el principal factor que contribuyó a la disminución de esta especie (Hawkins et al. 2000). Al Macá de Atitlán (Podilymbus gigas), en Guatemala, también se lo considera extinto por competencia o hibridación con el Macá Pico Grueso (P. podiceps, Hunter 1988). Algunos investigadores sugieren que la hibridación requiere acciones de manejo en especies amenazadas (Pierce 1984, Nixon 1994, Triggs & Daugherty 1996).
El Cardenal Amarillo hibrida con la Diuca Común Diuca diuca minor (Figura 6.1) en la región sur de su distribución (provincias de Rio Negro, sureste de La Pampa y sur de Buenos Aires) donde se encuentran en simpatría en época reproductiva (Narosky & Yzurieta 1987, Canevari et al. 1991, Narosky & Di Giácomo 1993). Aunque el área de distribución de las dos especies es mayoritariamente alopátrica, existen en simpatría en esta limitada región. La hibridación entre estas especies en estado silvestre ha sido registrada de manera ocasional desde 1987 (Bascarán 1987, Pazos 1995, Bertonatti & López Guerra 1997).

HIBRIDACIÓN CON LA DIUCA COMÚN
105
Figura 6.1. Ejemplar de Diuca Común (Diuca diuca) capturado en zona de simpatría con Cardenal Amarillo.
El género Diuca es el clado hermano del Cardenal Amarillo (Campagna et al. 2011,
Barker et al. 2013, Burns et al. 2014, Figura 6.2) e incluye dos especies, la Diuca Común Diuca diuca y la Diuca Ala Blanca Diuca speculifera. La Diuca Ala Blanca se encuentra en Perú, Bolivia y norte de Chile (Birdlife International 2015). Si bien esta especie fue antiguamente considerada dentro del género Diuca, es una especie poco conocida que no ha sido incluida en las últimas filogenias moleculares y muchos ornitólogos consideran que su morfología, canto y manera de caminar son más parecidos a las especies del género Phrygilus (Jaramillo 2011). Ridgely & Tudor (2009) incluso mencionan que las dos especies de Diuca parecerían no ser cogenéricas. Por su parte, la Diuca Común tiene una amplia distribución que abarca desde el norte de la Patagonia hasta el sur de Bolivia, incluyendo Chile. A diferencia del cardenal, es una especie monomórfica y migratoria que se reproduce en la región sur y oeste de su distribución y en invierno migra hacia el centro y litoral argentino (Figura 6.3). En los últimos años, se ha reportado la existencia de poblaciones reproductivas en Corrientes, donde la especie se ha hecho residente y donde también se ha observado un híbrido de cardenal y diuca (Mahler & Figuerero 2011).
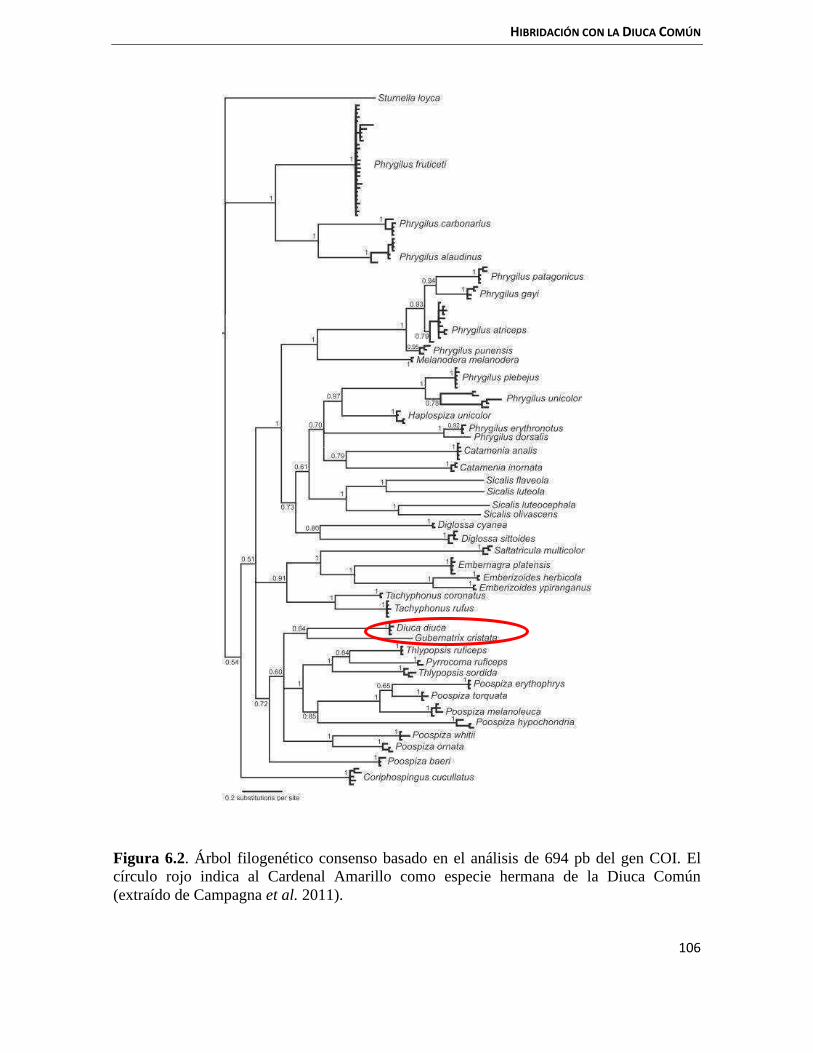
HIBRIDACIÓN CON LA DIUCA COMÚN
106
Figura 6.2. Árbol filogenético consenso basado en el análisis de 694 pb del gen COI. El círculo rojo indica al Cardenal Amarillo como especie hermana de la Diuca Común (extraído de Campagna et al. 2011).

HIBRIDACIÓN CON LA DIUCA COMÚN
107
Figura 6.3. Distribución de la Diuca Común, Diuca diuca (tomada de Birdlife International 2015). En verde se muestra el área donde la especie es nativa y residente y en azul el área donde es nativa pero no reproductora.
El híbrido es similar en su morfología al Cardenal Amarillo pero exhibe una coloración más parecida a la Diuca, con corona, nuca, mejilla, pecho, flancos, alas y cola grises (Figura 6.4). Tiene un copete gris y un babero rufo parecidos a los del Cardenal pero menos prominentes. También exhibe un área auricular gris y un área blanca en la región malar y superciliar. El centro del pecho, abdomen y vientre son blancos y las alas, al igual que en la Diuca Común, tienen 18 remeras a diferencia el Cardenal Amarillo que tiene 16 (Bertonatti & López Guerra 2007).
Figura 6.4. Imágenes de híbridos entre Cardenal Amarillo y Diuca Común. La fotografía a la izquierda corresponde a un individuo en cautiverio y la de la derecha a un individuo juvenil encontrado en la provincia de Corrientes.

HIBRIDACIÓN CON LA DIUCA COMÚN
108
El canto actúa como una importante barrera reproductiva pre-cigótica (Martens 1996). La convergencia en los patrones de canto entre especies podría surgir como consecuencia de la adaptación paralela al mismo hábitat (Wiley 1991) o a través del aprendizaje de cantos heterospecificos (Grant & Grant 1996, 1997, 1998). Cuando ocurre hibridación, la elección hacia los machos heterospecificos parece estar relacionada con la similitud de los cantos (Qvarnstrom et al. 2005) y con la baja disponibilidad de machos conespecíficos (Grant & Grant 1997). Bertonatti & López Guerra (2007) sugieren que la hibridación entre el Cardenal Amarillo y la Diuca Común podría ser consecuencia de la deliberada extracción de la naturaleza de machos de cardenal para proveer el mercado ilegal de animales silvestres.
El objetivo general de este capítulo es estudiar si la hibridación con la Diuca Común (Diuca diuca minor) afectó alguno de los parámetros estudiados en la población sur de cardenales amarillos. A nivel genético, se analiza la presencia de introgresión de alelos de diuca en los cardenales. Este resultado es particularmente importante porque indicaría que los híbridos entre ambas especies son fértiles y se reproducen con una especie parental. Hasta el momento, solo se sabe que existen híbridos pero no se tiene información acerca de su comportamiento reproductivo y, específicamente, su fertilidad. La retrocruza de individuos híbridos con Cardenales Amarillos también podría, en el largo plazo, afectar notoriamente la morfología de esta población. A diferencia de los caracteres heredados, el canto, que es aprendido, puede mostrarse influenciado por los eventos de hibridación aunque los híbridos no sean reproductivos. Es posible que éstos hayan adquirido estructuras vocales de sus padres (diucas comunes) y que otros cardenales copien estas estructuras de los híbridos o diucas, generando unidades de canto nuevas y distintas en la población sur.
6.2. Metodología
6.2.1. Genética
6.2.1.1. Colección de datos
Se capturaron 15 individuos de Diuca Común en las cercanías de General Conesa, Río Negro, durante las temporadas reproductivas 2011 y 2015 siguiendo la metodología con redes de niebla descripta en el capítulo 2. También se obtuvieron muestras de 2 híbridos entre Cardenal Amarillo y Diuca Común mantenidos en cautiverio. Se tomaron muestras de sangre de todos los individuos (aproximadamente 50 μl) mediante punción de la vena braquial con una aguja 30G. La sangre fue conservada en 500 μl de buffer de lisis (100mM TRIS, 100mM EDTA, 10mM NaCl, 2% SDS) a temperatura ambiente hasta ser analizada.

HIBRIDACIÓN CON LA DIUCA COMÚN
109
Además, se utilizó la información genética de los 102 individuos de Cardenal Amarillo utilizada en el capítulo 5.
6.2.1.2. Análisis Genéticos
La extracción de ADN a partir de las muestras de sangre se realizó utilizando un kit de extracción de Qiagen (Hilden, Alemania). Los estudios genéticos se basaron en los 10 marcadores de tipo microsatélite estudiados previamente para los cardenales amarillos, siguiendo las mismas condiciones de amplificación y genotipado.
Además de analizar el genotipo de los híbridos, también se analizó el fragmento de región control de ADN mitocondrial con el mismo par de cebadores y condiciones de PCR utilizados para Cardenal Amarillo. Adicionalmente, se realizó una determinación molecular del sexo. Los dos individuos fueron sexados a partir de la amplificación del gen CHD localizado en los cromosomas sexuales de las aves (Ellegren 1996), que presenta un intrón de distinto tamaño en los cromosomas sexuales Z y W. Las amplificaciones se realizaron a partir de los primers P2 y P8 (Griffiths et al. 1998) por medio de una PCR en un volumen final de 10 µl utilizando 50-100 ng de ADN templado, 0.5 µM de cebadores, 0.25 mM de cada dNTP, 2.5 mM MgCl2 y 0.25 u de Taq-Polymerasa (Invitrogen). Las condiciones de la PCR fueron: desnaturalización inicial a 94°C por 3 min, seguido de 35 ciclos a 94°C por 30 seg, 50°C por 45 seg y 72°C por 60 seg, finalizando con un ciclo de extensión a 72°C por 5 min. Los productos de PCR fueron separados en un gel de agarosa al 3% teñido con GelRed (Biotium).
Los genotipos de las diucas se analizaron con Microchecker v2.2.3 (Oosterhout et al. 2004) para detectar la presencia de alelos nulos. Arlequin v3.5.1 (Excoffier & Lischer 2010) fue utilizado para analizar las desviaciones del equilibrio Hardy-Weinberg (HW) y el desequilibrio por ligamiento. La significancia de las pruebas de desequilibrio por ligamiento se evaluó luego de aplicar una corrección de Bonferroni para comparaciones múltiples (Rice 1989).
Se estimó el número de grupos (K) mediante un análisis de asignación Bayesiano implementado en STRUCTURE v2.3.3 (Prichard et al. 2000) para las muestras de Cardenal Amarillo, Diuca Común e híbridos. Se corrió un modelo que utiliza la información previa sobre la población de origen de cada muestra y que permite ―admixture‖ (i.e. mezcla) entre los grupos genéticos asumiendo frecuencias alélicas independientes para K = 2. Las simulaciones se llevaron a cabo con un burn-in de 200.000 pasos y un período de recopilación de datos de 1.500.000 iteraciones. Cada simulación se repitió 10 veces.

HIBRIDACIÓN CON LA DIUCA COMÚN
110
6.2.2. Cantos
Los cantos de las Diucas Comunes se grabaron en la parte sur de la distribución del Cardenal Amarillo (provincias de La Pampa y Río Negro) donde se regsitró la mayor parte de los eventos de hibridación (Bertonatti & López Guerra 1997). Cantos de Diucas de otras áreas (n = 3) se obtuvieron de la biblioteca de sonido Macaulay Library del Laboratorio de Ornitología de la Universidad de Cornell (números de catálogo 135815 perteneciente a la provincia de Neuquén, 20607 y 20605 ambos de Chubut). Se analizó si existe introgresión cultural entre los cantos de Cardenales Amarillos y Diucas Comunes en las zonas donde se registraron eventos de hibridación. Los cantos se digitalizaron de la manera descripta en el capítulo 3 con el programa Raven Pro 1.4 (Charif et al. 2007) y se analizaron las mismas variables. Se aplicó un análisis de la función discriminante similar al del capítulo 3 con 30 cantos de Cardenal Amarillo y añadiendo 17 muestras Común Diuca como un nuevo grupo. Todos los análisis multivariados se realizaron con Infostat (Di Rienzo et al. 2013).
6.3. Resultados
6.3.1. Genética
Para las muestras de Diuca Común no hubo evidencia de errores de genotipado o alelos nulos para ningún locus polimórfico. Las frecuencias alélicas de los loci microsatélites no se desviaron del equilibrio HW y no hubo evidencia de desequilibrio por ligamiento entre ningún par de loci. Se encontraron de 3 a 14 alelos por locus en las muestras de Diucas. Estos individuos mostraron alelos privados (que están en diuca y no en cardenal) para los 10 loci estudiados (rango 1 – 12 por locus). Para 8 loci, ambas especies compartieron de 2 a 10 alelos.
No se encontraron evidencias de introgresión de alelos provenientes de la Diuca Común en los Cardenales Amarillos, particularmente en los cardenales de la región sur donde ocurren los eventos de hibridación (Figura 6.5). En todos los cardenales la pertenencia del genotipo a la especie fue mayor al 98% y en el caso de las diucas la pertenencia del genotipo a la especie fue del 100%. En el caso de los dos híbridos, hubo un 60% de asignación del genotipo a cardenales y un 40% a diucas (Figura 6.5).
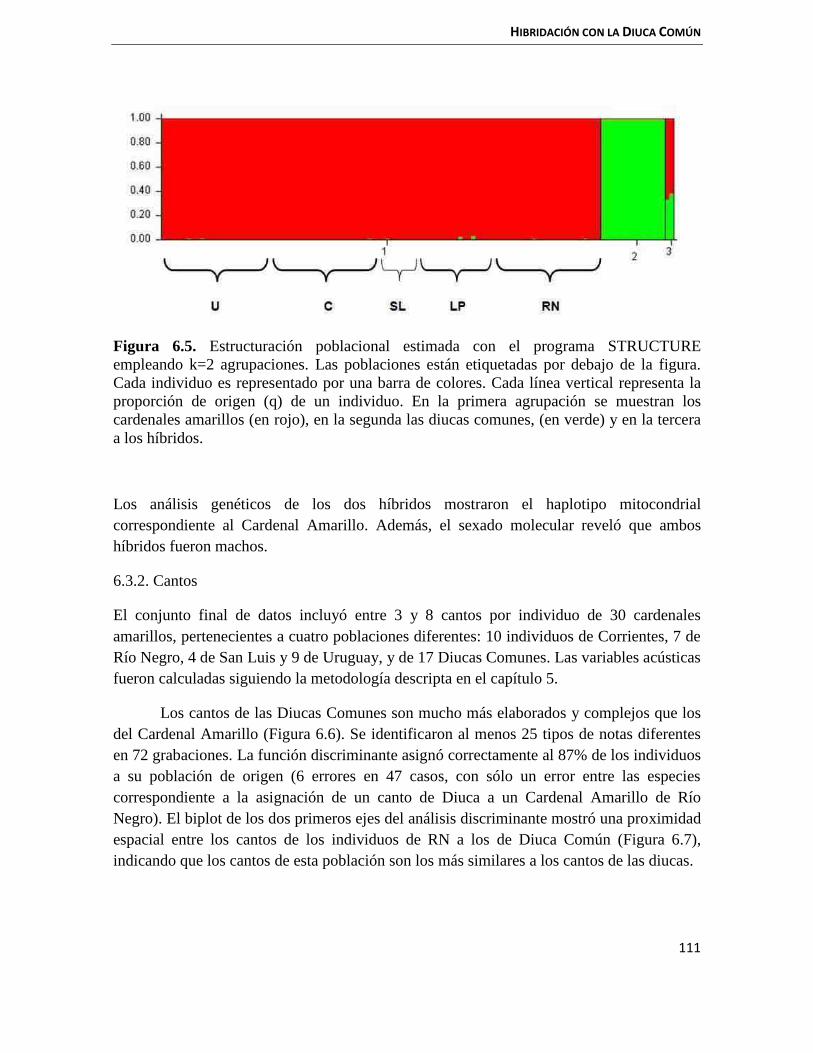
HIBRIDACIÓN CON LA DIUCA COMÚN
111
Figura 6.5. Estructuración poblacional estimada con el programa STRUCTURE empleando k=2 agrupaciones. Las poblaciones están etiquetadas por debajo de la figura. Cada individuo es representado por una barra de colores. Cada línea vertical representa la proporción de origen (q) de un individuo. En la primera agrupación se muestran los cardenales amarillos (en rojo), en la segunda las diucas comunes, (en verde) y en la tercera a los híbridos.
Los análisis genéticos de los dos híbridos mostraron el haplotipo mitocondrial correspondiente al Cardenal Amarillo. Además, el sexado molecular reveló que ambos híbridos fueron machos.
6.3.2. Cantos
El conjunto final de datos incluyó entre 3 y 8 cantos por individuo de 30 cardenales amarillos, pertenecientes a cuatro poblaciones diferentes: 10 individuos de Corrientes, 7 de Río Negro, 4 de San Luis y 9 de Uruguay, y de 17 Diucas Comunes. Las variables acústicas fueron calculadas siguiendo la metodología descripta en el capítulo 5.
Los cantos de las Diucas Comunes son mucho más elaborados y complejos que los del Cardenal Amarillo (Figura 6.6). Se identificaron al menos 25 tipos de notas diferentes en 72 grabaciones. La función discriminante asignó correctamente al 87% de los individuos a su población de origen (6 errores en 47 casos, con sólo un error entre las especies correspondiente a la asignación de un canto de Diuca a un Cardenal Amarillo de Río Negro). El biplot de los dos primeros ejes del análisis discriminante mostró una proximidad espacial entre los cantos de los individuos de RN a los de Diuca Común (Figura 6.7), indicando que los cantos de esta población son los más similares a los cantos de las diucas.
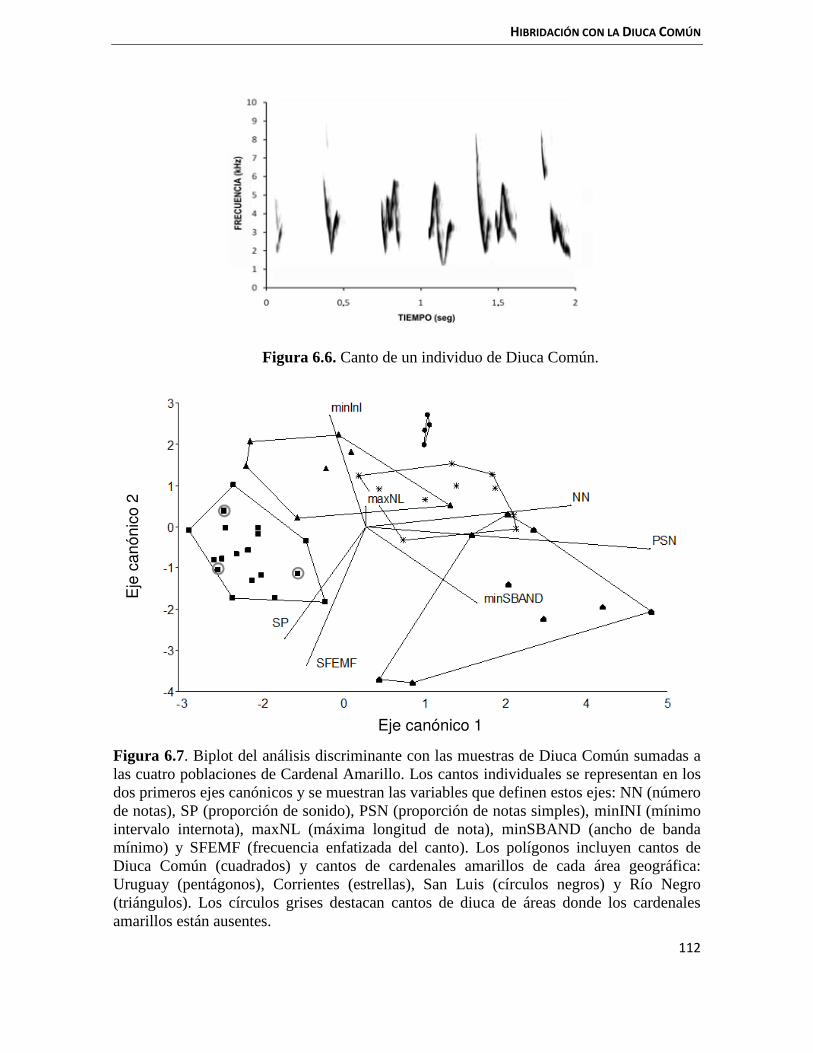
HIBRIDACIÓN CON LA DIUCA COMÚN
112
Figura 6.6. Canto de un individuo de Diuca Común.
Figura 6.7. Biplot del análisis discriminante con las muestras de Diuca Común sumadas a las cuatro poblaciones de Cardenal Amarillo. Los cantos individuales se representan en los dos primeros ejes canónicos y se muestran las variables que definen estos ejes: NN (número de notas), SP (proporción de sonido), PSN (proporción de notas simples), minINI (mínimo intervalo internota), maxNL (máxima longitud de nota), minSBAND (ancho de banda mínimo) y SFEMF (frecuencia enfatizada del canto). Los polígonos incluyen cantos de Diuca Común (cuadrados) y cantos de cardenales amarillos de cada área geográfica: Uruguay (pentágonos), Corrientes (estrellas), San Luis (círculos negros) y Río Negro (triángulos). Los círculos grises destacan cantos de diuca de áreas donde los cardenales amarillos están ausentes.
Eje
ca
nó
nic
o 2
Eje canónico 1

HIBRIDACIÓN CON LA DIUCA COMÚN
113
6.4. Discusión
La hibridación entre especies es un evento raro en la naturaleza pero es aún más raro cuando la hibridación es intergenérica. Además del Cardenal Amarillo y la Diuca Común hay otros ejemplos de especies pertenecientes a géneros distintos que hibridan. Se han reportados híbridos intergenéricos dentro de las familias Pipridae, formada por pequeños pájaros arborícolas de centro y sudamérica como los saltarines, piprites, manaquines y llorones (Parkes 1961), entre especies de la familia Parulidae, como las pequeñas y coloridas reinitas, chipes o bijiritas del Nuevo Mundo (Parkes 1978) y también se ha reportado hibridación intra- e intergenérica entre especies de aves marinas de la familia Sternidae (Hays 1975).
La falta de evidencias de introgresión de alelos de diuca en los cardenales amarillos de la zona sur indicaría que no existen retrocruzas entre los híbridos y las especies parentales o que las tasas de hibridación son muy bajas. Usualmente, la fertilidad de los huevos de individuos híbridos que se reproducen con un individuo de la especie parental es reducida (Short 1985) lo que podría explicar por qué no se encontraron alelos de diuca en las poblaciones silvestres de Cardenal Amarillo. Las experiencias en cautividad indican que es posible cruzar hembras de Cardenal Amarillo con las dos especies de Diuca y que la descendencia es viable (Pazos 1995) pero que la retrocruza y la cruza de machos de Cardenal Amarillo con hembras de Diuca resulta en huevos infértiles o pichones que murieron en los primeros días de vida (Bertonatti & Lopez Guerra 1997). Los híbridos analizados resultaron ser machos, lo cual, a pesar del bajo número muestral, apoyaría la regla de Haldane (1922). Ésta predice que cuando existe algún tipo de incompatibilidad genética entre especies, como la esterilidad o inviabilidad de algún sexo en la progenie híbrida, ésta se produce preferentemente en el sexo heterogamético, que en el caso de las aves corresponde a las hembras cuyo determinismo sexual ZW. La teoría de la dominancia, una versión mejorada de este modelo, propone la evolución rápida de los factores de esterilidad, donde los genes involucrados en la eficacia biológica de los híbridos son parcialmente recesivos y únicamente causarán efectos cuando se hallen en hemizigosis en el sexo heterogamético.Es decir que se espera que se cumpla la regla de Haldane si las incompatibilidades incluyen a loci ligados al cromosoma X (o al Z) de una especie y loci autosómicos de la otra, y si los alelos que causan estas incompatibilidades son parcialmente recesivos (Orr 1993, 1995; Turelli & Orr 1995, 2000; Orr & Turelli 1996, 2001; Zeng 1996). No es sorprendente encontrar que los híbridos entre Cardenal Amarillo y Diuca Común sigan la regla de inviabilidad de Haldane ya que ésta se ha observado en varios grupos incluyendo nematodos, dípteros, ortópteros, heterópteros, lepidópteros, anfibios, reptiles, mamíferos y aves (Laurie 1997).

HIBRIDACIÓN CON LA DIUCA COMÚN
114
La hibridación entre el Cardenal Amarillo y la Diuca Común puede darse por una similitud entre el canto de ambas especies, favorecida por la baja presencia de machos de cardenal en la zona. Se encontró que los cantos de las diucas son más complejos que los cantos de los cardenales amarillos (repertorio diuca = 25 tipos de notas frente al repertorio del cardenal = 5 tipos de notas). La mayor complejidad del canto de diuca podría ser el rasgo seleccionado por las hembras de cardenal, como fue observado para otras especies (Searcy 1992). Las características temporales y de frecuencia de los cantos de cardenal resultaron más similares a los de diuca en las zonas donde la hibridación ocurre. Esto podría ser consecuencia de una influencia del canto de las diucas sobre los patrones de canto de los cardenales amarillos. En la zona donde la presencia de machos de la misma especie es escasa, la elección de los machos de diuca por parte de las hembras de Cardenal Amarillo podría estar relacionada con la similitud de los cantos (Qvarnstrom et al. 2005, Grant & Grant 1997). Podrían ocurrir errores de impronta en el aprendizaje de los patrones de canto en los machos jóvenes de Cardenal Amarillo como consecuencia de la escasez de machos conespecíficos resultando en el copiado de algunas características del canto similar de las diucas. De esta manera, la introgresión cultural podría estar afectando las vocalizaciones de los cardenales. En un estudio realizado en una zona híbrida en Estados Unidos entre Poecile atricapillus y P. carolinensis, se observó que el canto no presentaba signos de introgresión, sino que los individuos híbridos cantaban el canto de alguna de las especies parentales ligeramente modificado, probablemente como consecuencia del proceso de aprendizaje del canto (Sattler et al. 2007). Otro trabajo realizado en una zona híbrida entre Hippolais polyglotta y H. icterina también encontró que los híbridos aprenden el canto de una de las especies parentales (Secondi et al. 2003).
Una explicación alternativa es que la adaptación paralela al mismo ambiente esté impulsando las características de ambos cantos (Wiley 1991). Sin embargo, al analizar los cantos de diucas pertenecientes a diferentes partes de su distribución (ver Figura 6.7), procedentes de hábitats muy diferentes (estepa patagónica), se observa que éstos cayeron dentro del rango de las características de los cantos de las diucas comunes que superponen su distribución con los cardenales amarillos, indicando que las características del canto de las diucas son especie-específicas y no dependientes del ambiente.
En una zona donde Ficedula hypoleuca se encuentra en simpatría con F. albicollis la hibridación entre ambas especies es frecuente. Para ambas especies se observó que los machos con cantos más convergentes con los de la otra especie eran los más propensos a hibridar (Qvarnstrom et al. 2005). No tenemos información acerca de las preferencias de las hembras de cardenal por el canto de las diucas, pero sería interesante analizar si las características del canto de las diucas afecten la probabilidad de hibridación.
Hasta ahora sabemos que las barreras reproductivas entre el Cardenal Amarillo y la Diuca Común no son lo suficientemente fuertes para prevenir la hibridación. Sin embargo, los híbridos detectados en el campo son pocos. Esta escasez puede deberse a una elevada

HIBRIDACIÓN CON LA DIUCA COMÚN
115
tasa de mortalidad y/o una baja eficacia biológica de los híbridos o a que es un evento que ocurre sólo ocasionalmente. Si bien éste constituye el primer estudio que genera conocimientos que nos acercan a conocer un poco más la naturaleza de este fenómeno es necesario reforzarlo aumentando el conocimiento sobre la elección de pareja y el número de híbridos estudiados.
6.5. Referencias bibliográficas
Allendorf, F. W. & G. Luikart. 2007. Hybridization. Pp 421-448. En: Conservation and the Genetics of Populations. Blackwell Publishing, Oxford, Estados Unidos.
Banks, R. C. & N. K. Johnson. 1961. A review of North American hybrid hummingbirds. Condor 63: 3–28.
Bascarán, J. L. 1987. El Cardenal Amarillo. Rev. Asociación Civil de Criadores de Especies Silvestres, Proveedores y Afines (ACCESPA) N° 6, Buenos Aires.
Barker, K., K. Burns, J. Klicka, S. Lanyon & I. Lovette. 2013. Going to extremes: Contrasting rates of diversification in a recent radiation of New World Passerines Birds. Systematic Biology 62: 298–320.
Bertonatti, C. & A. López Guerra. 1997. Hibridación entre Cardenal Amarillo (Gubernatrix cristata) y Diuca Común (Diuca diuca minor) en estado silvestre, en la Argentina. El Hornero 14: 235–242.
BirdLife International. 2015. Species factsheet: Diuca speculifera. Downloaded from http://www.birdlife.org on 18/11/2015.
BirdLife International. 2015. Species factsheet: Diuca diuca. Downloaded from http://www.birdlife.org on 19/11/2015.
Burns, K. J., A. J. Shultz, P. O. Title, N. A. Mason, F. K. Barker, J. Klicka, L. Scott & I. J. Lovette. 2014. Phylogenetics and diversification of tanagers (Passeriformes: Thraupidae), the largest radiation of Neotropical songbirds. Molecular phylogenetics and evolution 75: 41–77.
Campagna, L., K. Geale, P. Handford, D. A. Lijtmaer, P. L. Tubaro & S. C. Lougheed. 2011. A molecular phylogeny of the Sierra-Finches (Phrygilus, Passeriformes): extreme polyphyly in a group of Andean specialists). Molecular Phylogenetics and Evolution 61: 521–533.
Canevari, M., P. Canevari, G. R. Carrizo, G. Harris, J. Rodríguez Mata & R. Straneck.1991. Nueva Guía de las Aves Argentinas. Tomo II, Fundación Acindar, Buenos Aires.

HIBRIDACIÓN CON LA DIUCA COMÚN
116
Carroll, S. P., & C. W. Fox. 2008. Hybridization, Introgression, and the Evolutionary Management of Threatened Species. Pp 130-140. En Conservation biology: evolution in action. Oxford University Press, Oxford, Estados Unidos.
Charif, R. A., C. W. Clarck & K. M. Fristrup. 2007. Raven Pro 1.3 user‘s manual. Cornell Laboratory of Ornithology, Ithaca, NY.
Di Rienzo, J. A., F. Casanoves, M. G. Balzarini, L. Gonzalez, M. Tablada & C. W. Robledo. InfoStat versión 2013. InfoStat Group, Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba, Argentina. URL http://www.infostat.com.ar
Ellegren, H. 1996. First gene on the avian W chromosome (CHD) provides a tag for universal sexing of non-ratite birds. Proceeding of the Royal Society of London B 263: 1635–1641.
Excoffier, L. & H. E. Lischer. 2010. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Molecular ecology resources 10: 564–567.
Grant, P. R. & R. Grant. 1992. Hybridization of bird species. Science 256: 193.
Grant, B. R. & P. R. Grant. 1996. Cultural inheritance of song and its role in the evolution of Darwin‘s finches. Evolution 50: 2471–2487.
Grant, P. R. & R. Grant. 1997. Hybridization, sexual imprinting and mate choice. American Naturalist 149: 1–28.
Griffiths, R., M. C. Double, K. Orr y R. J.G. Dawson. 1998. A DNA test to sex most birds. Molecular Ecology 7: 1071–1075.
Haldane, J. B. 1922. Sex ratio and unisexual sterility in hybrid animals. Journal of genetics 12: 101–109.
Hawkins, F., R. Andriamasimanana, S. Seing & Z. Rabeony. 2000. The sad story of the Alaotra Grebe Tachybaptus rufolavatus. Bulletin of the African Bird Club 7: 115–117.
Hays, H. 1975. Probable Common x Roseate Tern Hybrids. Auk 92 (2): 219–234.
Hunter, L. A. 1988. Status of the endemic atitlan grebe of Guatemala: is it extinct?. The Condor 90: 906–912.
Jaramillo, A. 2011. White winged Diuca finch (Diuca speculifera). En: del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A. & de Juana, E. (eds.) (2014). Handbook of the Birds of the World Alive. Lynx Edicions, Barcelona. (descargado de http://www.hbw.com/node/62049 el 18 de noviembre 2015).

HIBRIDACIÓN CON LA DIUCA COMÚN
117
Laurie, C. C. 1997. The weaker sex is heterogametic: 75 years of Haldane‘s rule. Genetics 147: 937–951.
Mahler, B. & Figuereo C. 2011. Los Cardenales Difíciles. Aves Argentinas Revista de Naturaleza y Conservación 30: 10–11.
Martens, J. 1996. Vocalizations and speciation of Palearctic birds. En: Ecology and Evolution of Acoustic Communication in Birds (D. E. Kroodsma & E. H. Miller, eds), pp. 221–240. Cornell University Press, Ithaca, NY, EEUU.
Narosky, T. & D. Yzurieta. 1987. Guía para la identificación de las Aves de Argentina y Uruguay: 267 y 273. Asociación Omitológica del Plata, Buenos Aires.
Narosky,T. & A. G. Di Giácomo. 1993. Las aves de la Provincia de Buenos Aires: distribución y estatus. Asociación Omitológica del Plata, Vázquez Mazzini Ed. y Literature of Latin America (L.O.L.A.), Buenos Aires.
Nixon, A.J. (1994). Feeding ecology of hybridising parakeets on Mangere Island, Chatham Islands. Notornis, 41(supplement), 5–18.
Norris, K. & D. Pain. 2002. Conserving Bird Biodiversity, General principles and their application. Cambridge University Press, United Kingdom.
Oosterhout, C., Hutchinson, W. F., Wills, D. P., & Shipley, P. 2004. MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data. Molecular Ecology Notes 4: 535–538.
Orr, H. A. 1993. A mathematical model of Haldane‘s rule. Evolution 47: 1606–1611.
Orr, H. A. 1995. The population genetics of speciation: the evolution of hybrid incompatibilities. Genetics 139: 1805–1813.
Orr, H. A. & M. Turelli. 1996. Dominance and Haldane‘s rule. Genetics 143: 613–616.
Orr, H. A. & M. Turelli. 2001. The evolution of postzygotic isolation: accumulating Dobzhansky-Muller incompatibilities. Evolution 55: 1085–1094.
Parkes, K. C. 1961. Intergeneric hybrids in the family Pipridae. Condor 63: 345–350.
Parkes, K. C. 1978. Still another parulid intergeneric hybrid (Mniotilta x Dendroica) and its taxonomic and evolutionary implications. Auk 95: 682–690.
Pazos, J. H. 1995. El Cardenal Amarillo; reproducción en cautiverio. Revista Argentina de Omitología I (1): 34–35.
Peterson, R.T., Mountfort, G. & Hollom, P.A.D. 1993. Birds of Britain and Europe, new edition. Collins Field Guide Series. London: HarperCollins.

HIBRIDACIÓN CON LA DIUCA COMÚN
118
Pierce, R. J. 1984. Plumage, morphology and hybridisation of New Zealand stilts Himantopus spp. Notornis 31: 106–130.
Pritchard, J. K., M. Stephens & P. Donnelly 2000. Inference of population structure using multilocus genotype data. Genetics 155: 945–959.
Qvarnstrom, A., J. Haavie, S. A. Saether, D. Ericsson, & T. Pärt. 2005. Song similarity predicts hybridization in flycatchers. Journal of Evolutionary Biology 19: 1202–1209.
Rice, W. R. 1989. Analyzing tables of statistical tests. Evolution 43: 223–225.
Ridgely, R. S. & G. Tudor. 2009. Field guide to the songbirds of South America: the passerines. University of Texas Press.
Sattler, G. D., P. Sawaya & M. J. Braun. 2007. An assessment of song admixture as an indicator of hybridization in Black-capped Chickadees (Poecile atricapillus) and Carolina Chickadees (P. carolinensis). The Auk 124(3): 926–944.
Searcy, W. A. 1992. Song Repertoire and Mate Choice in Birds. American Zoologist 32: 71–80.
Secondi, J., V. Bretagnolle, C. Compagnon & B. Faivre. 2003. Species-specific song convergence in a moving hybrid zone between two passerines. Biological Journal of the Linnean Society 80: 507–517.
Short, L. L. 1969. Taxonomic aspects ofavian hybridization. Auk 86: 85–105.
Short, L. L. 1985. Hybridization, Zone of Secundary. Pp 296-297. En Campbell, B. & E. Lack (Eds.). A Dictionary of Birds. Buteo Books, Vermillion, South Dakota, Estados Unidos.
Triggs, S. J. & C. H. Daugherty. 1996. Conservation and genetics of New Zealand parakeets. Bird Conservation International 6: 89–101.
Turelli, M. & H. A. Orr. 1995. The dominance theory of Haldane‘s rule. Genetics 140: 389–402.
Turelli, M. & H. A. Orr. 2000. Dominance, epistasis and the genetics of postzygotic isolation. Genetics 154: 1663–1679.
Wiley, R. H. 1991. Associations of song properties with habitats for territorial oscine birds of eastern North America. American Naturalist 138: 973–993.
Zelaya, D. & C. Bertonatti. 1995. Nuestro Libro Rojo: Cardenal Amarillo. Vida Silvestre 43: 21–22, Fundación Vida Silvestre Argentina.
Zeng, L. W. 1996. Resurrecting Muller‘s theory of Haldane‘s rule. Genetics 143: 603–607.

DISCUSIÓN GENERAL Y CONSIDERACIONES FINALES
119
CAPÍTULO 7
DISCUSIÓN GENERAL Y CONSIDERACIONES FINALES
“…in the end, we will conserve what we love. We will love
only what we understand. We will understand only what
we are taught”
Baba Dioum
7.1. Principales resultados
Los resultados de esta tesis brindan información novedosa acerca de la biología del Cardenal Amarillo. Por un lado, se obtuvieron índices reproductivos para la especie y, por otro lado, se pudo determinar la variación geográfica que presenta el Cardenal Amarillo para diversos caracteres. La información publicada en este trabajo podrá ser utilizada para delinear estrategias de conservación exitosas para esta especie de ave amenazada.
7.1.1. Éxito reproductivo
El estudio llevado a cabo durante dos temporadas reproductivas en la provincia de Corrientes permitió conocer el potencial reproductivo de los cardenales amarillos. Las tasas de supervivencia fueron similares a aquellas encontradas para otras especies de aves paseriformes con historias de vida comparables que no se encuentran amenazadas (Grant et al. 2005). Se observó que las parejas son capaces de renidificar luego de un intento fallido e incluso se presenciaron cuatro intentos de una pareja reproductora en una misma temporada. Se encontró que la especie es socialmente monógama y las parejas se mantienen estables a lo largo de la temporada y entre temporadas sucesivas, permaneciendo en el mismo territorio. Los territorios tienen un tamaño promedio de 2.16 ha y pueden encontrarse a una distancia mínima de 200 metros de los vecinos. Esto hace que la capacidad de carga en un área con hábitat adecuado pueda ser de aproximadamente tres parejas por km2. Además, se observó un éxito reproductivo levemente mayor en una población que habita un campo con actividad ganadera sustentable en comparación a la población que habita un área protegida. El tipo de manejo que se lleva a cabo en ese campo involucra cargas no muy altas de ganado y un cuidado del ambiente natural. Esto implica que la conservación del Cardenal Amarillo es compatible con actividades ganaderas que mantengan el ambiente de espinal, particularmente las especies arbóreas que utilizan los cardenales para nidificar.

DISCUSIÓN GENERAL Y CONSIDERACIONES FINALES
120
7.1.2. Amenazas Además de las amenazas conocidas para el Cardenal Amarillo, extracción de individuos para el comercio ilegal y pérdida de hábitat, durante las campañas se encontraron nuevas amenzas que pueden afectar la viabilidad de las poblaciones. Por un lado, se detectó el parasitismo por parte del Tordo Renegrido. El mayor costo del parasitismo de los tordos fue la picadura de los huevos, que resultó en el abandono de los nidos y en consecuencia, disminuyó el éxito reproductivo. Esta amenaza está presente en toda el área de distribución del Cardenal Amarillo y se agrava con el aumento de la abundancia de los tordos favorecida por las actividades agrícola-ganaderas. Por otro lado, se registró el parasitismo de moscas del género Philornis, que provocó una disminución de la supervivencia de los pichones de cardenal. Estas moscas se distribuyen hasta los 35°S, por lo cual las poblaciones más australes de la distribución del Cardenal Amarillo no se encontrarían frente a esta amenaza. La hibridación con la Diuca Común también había sido planteada como una posible amenaza para la supervivencia de las poblaciones del Cardenal Amarillo (Betonatti & López Guerra 2007). Los resultados encontrados en este estudio sugieren que no existen retrocruzas de los híbridos con la especie parental. Si bien sólo se estudiaron dos híbridos, los análisis moleculares revelaron que ambos fueron machos. A pesar de que es necesario analizar un número mayor de híbridos, esto se condice con la regla de Haldane (1922), que predice que cuando existen incompatibilidades genéticas entre especies, como esterilidad o inviabilidad de algún sexo en la progenie híbrida, ésta se produce preferentemente en el sexo heterogamético. Además, los híbridos portaban un haplotipo mitocondrial correspondiente al Cardenal Amarillo indicando que los cruzamientos se darían entre hembras de cardenal y machos de diuca. Esto apoya la idea de que la hibridación puede darse como consecuencia de la baja disponibilidad de cardenales machos en la naturaleza debido a su extracción para abastecer el mercado ilegal de venta de aves silvestres. A pesar de no tener un efecto genético, disminuye el reclutamiento y, la hibridación sumada a la baja presencia de machos podría estar afectando la estructura del canto de los cardenales. La transmisión cultural del canto hace que puedan aparecer variantes nuevas en poco tiempo y podría explicar la similitud de los cantos de los cardenales de Río Negro con los de las diucas.
7.1.3. Unidades de manejo
En los últimos años, un importante número de trabajos ha estudiado la diversidad genética, morfológica y/o comportamental entre poblaciones de diversas especies de aves (e.g. MacDougall-Shackleton & MacDougall-Shackleton 2001, Bulgin et al. 2003, Wright et al. 2005, Alström et al. 2007, Saranathan et al. 2007). En la mayoría de los casos, no se encontró una correlación entre la variación de los caracteres, lo cual está determinado en gran parte por las diferencias en la heredabilidad y los factores evolutivos (selección natural, selección sexual, deriva) que actúan sobre cada uno de ellos. En este caso, no se

DISCUSIÓN GENERAL Y CONSIDERACIONES FINALES
121
encontró una diferenciación clara en las variables morfológicas medidas pero sí en las características acústicas y la variabilidad genética de los cardenales de distintas regiones de la distribución actual, que permiten definir unidades de manejo dentro de esta especie. En base a los caracters genéticos se identificaron cuatro unidades de manejo correspondientes a Uruguay, Corrientes, San Luis y norte de La Pampa, y sur de La Pampa y Rio Negro. Las unidades culturales basadas en las características del canto apoyaron esta delimitación, dado que las vocalizaciones de Uruguay, Corrientes, San Luis y Río Negro diferían entre sí. No se pudieron obtener cantos de los individuos de La Pampa; sin embargo, sería interesante estudiar las características del canto en esa provincia y su correlación con la subestructuración genética. La ausencia de aislamiento por distancia en los caracteres estudiados sugierie que la diversidad (genética y acústica) en el Cardenal Amarillo pudo haber sido moldeada por la fragmentación del hábitat y la extracción deliberada de individuos de la naturaleza. En las poblaciones pequeñas y aisladas la alta probabilidad de fijar variantes por deriva (genética o cultural) genera una mayor diferenciación entre poblaciones.
7.2. Aplicaciones para la conservación de la especie
7.2.1. Conservación de la variabilidad
Las unidades de manejo encontradas en este estudio, delimitadas en base a información genética y de los cantos, son de importancia para delinear estrategias para la conservación de la especie y, sobre todo, asegurar su potencial evolutivo. Perder o disminuir el tamaño de una sola población podría comprometer o eliminar una tradición cultural o variante genética.
Para el caso del Cardenal Amarillo, donde aún quedan poblaciones naturales, se aconseja elaborar un plan de conservación in situ para proteger a la especie en su ambiente natural y permitir la interacción con otros organismos, dejándola evolucionar a largo plazo en su medio natural, en conexión con las condiciones ambientales. La red de poblaciones conservadas debe cubrir toda la variación genética y cultural espacial presente en la especie. Dado el escenario presentado en este estudio, se recomienda conservar las unidades de manejo encontradas en su ambiente natural. No obstante, en el caso de que se llegue al punto de que fuera necesario elaborar proyectos de conservación ex situ, sobre la base de la información genética generada, los programas de cría en cautiverio podrían beneficiarse al identificar potenciales parejas del mismo linaje para evitar problemas como la depresión por exogamia (Ballou et al. 2010).

DISCUSIÓN GENERAL Y CONSIDERACIONES FINALES
122
7.2.2. Manejo de cardenales decomisados
El comercio ilegal de vida silvestre es el tercer mayor tráfico ilícito después de las drogas y las armas. Cada año, de 2 a 5 millones de aves se negocian a nivel mundial para su uso como mascotas, para parques zoológicos y para colecciones privadas (Hemley 1994). La tasa de mortalidad de los animales confiscados ilegalmente generalmente es muy alta debido a las condiciones de captura y de transporte precarias. Aunque muchas aves incautadas no sobreviven, las que se recuperan podrían ser devueltas a su hábitat natural. Generalmente, la procedencia exacta de la mayoría de estos animales se desconoce. Los individuos deben ser liberados en el medio natural con precaución porque la mezcla entre poblaciónes puede inducir a la depresión por exogamia en la población local (Edmands 2007).
Los marcadores moleculares utilizados en este estudio, la región de control del ADN mitocondrial y los microsatélites, son marcadores neutros que no se ven influenciados por la selección natural. Por lo tanto, no brindan ninguna información sobre la naturaleza adaptativa de las diferencias genéticas. Sin embargo, los marcadores neutros han sido ampliamente utilizados para conocer y comparar la variabilidad intraespecífica en especies en peligro de extinción.
Los análisis genéticos también han sido empleados para inferir el origen geográfico de individuos en la investigación biológica (DNA forensics, Ogden et al. 2009). Identificar el origen geográfico de un individuo requiere asignar una muestra a una población en particular, que debe ser genéticamente distinta de otras poblaciones candidatas y se debe contar con información de poblaciones de múltiples regiones. Con la información generada en este trabajo será posible asignar el origen de los individuos incautados para elaborar un plan de liberación en su lugar de origen. Muchos cardenales son decomisados por las autoridades y depositados en centros gubernamentales de recepción. Un estudio similar en guacamayos azules (Anodorhynchus hyacinthinus) en Brasil pudo determinar que cinco individuos mantenidos en cautiverio provenían de la población silvestre localizada en Pantanal (Faria et al. 2008) y sobre la base de un estudio de estructuración genética del Guacamayo Azul y Amarillo (Ara ararauna) se identificaron individuos candidatos a ser liberados al noroeste de Brasil entre los animales de un decomiso (Caparroz et al. 2009, Fernandes & Caparroz 2013).
La información sobre la estructuración genética a nivel poblacional también puede ser relevante para combatir el tráfico ilegal de fauna ya que puede ayudar a identificar el origen geográfico de los individuos confiscados y así planificar acciones policiales preventivas concentradas en zonas sensibles a la captura ilegal (e.g. Presti et al. 2015).

DISCUSIÓN GENERAL Y CONSIDERACIONES FINALES
123
7.3. Referencias bibliográficas Alström, P., U. Olsson, P. C. Rasmussen, C. T. Yao, P. G. Ericson & P. Sundberg. 2007.
Morphological, vocal and genetic divergence in the Cettia acanthizoides complex (Aves: Cettiidae). Zoological Journal of the Linnean Society 149: 437–452.
Ballou J. D., G. Lees, L. J. Faust, S. Lang, C. Lynch, L. B. Lacky & T. J. Foose. 2010. Demographic and genetic management of captive population. Pp 219-262. En: Kleiman D. G. Thompson K. V., Baer C. K., editors. Wild mamals in captivitiy: principles and techniques for zoo management. Chicago: The University of Chicago Press.
Bertonatti, C. & A. López Guerra. 1997. Hibridación entre Cardenal Amarillo (Gubernatrix cristata) y Diuca Común (Diuca diuca minor) en estado silvestre, en la Argentina. El Hornero 14: 235–242.
Bulgin, N. L., H. L. Gibbs, P. Vickery & A. J. Baker. 2003. Ancestral polymorphisms in genetic markers obscure detection of evolutionarily distinct populations in the endangered Florida grasshopper sparrow (Ammodramus savannarum floridanus). Molecular Ecology 12: 831–844.
Caparroz, R., C. Y. Miyaki & A. J. Baker. 2009. Contrasting phylogeographic patterns in mitochondrial DNA and microsatellites: evidence of female philopatry and male-biased gene flow among regional populations of the blue-and-yellow macaw (Psittaciformes: ara ararauna) in Brazil. Auk 126: 359–370.
Edmands, S. 2007. Between a rock and a hard place: evaluating the relative risks of inbreeding and outbreeding for conservation and management. Molecular Ecology 16: 463–475.
Faria, P., N. Guedes, C. Yamashita, P. Martuscelli & C. Miyaki. 2008. Genetic variation and population structure of the endangered Hyacinth Macaw (Anodorhynchus hyacinthinus): implications for conservation. Biodivers Conserv 17:765–779.
Fernándes, G. A. & R. Caparroz. 2013. DNA sequence analysis to guide the release of blue-and-yellow macaws (Ara ararauna, Psittaciformes, Aves) from the illegal trade back into the wild. Molecular Biology Reports 40: 2757–2762.
Frankham, R., J. D. Ballou, M. D. B. Eldridge, R. C. Lacy, K. Ralls, M. R. Dudash & C.B. Fenster. 2011. Predicting the probability of outbreeding depression. Conservation Biology 25: 465–475.

DISCUSIÓN GENERAL Y CONSIDERACIONES FINALES
124
Grant, T. A., Shaffer, T. L., Madden, E. M. and Pietz, P. J. 2005. Time-specific variation in passerine nest survival: new insights for old questions. Auk 122: 661–672.
Haldane, J. B. 1922. Sex ratio and unisexual sterility in hybrid animals. Journal of genetics 12: 101–109.
Hemley, G. 1994. International wildlife trade: a CITES source book. Island Press, Washington.
MacDougall-Shackleton, E. A., & S. A. MacDougall-Shackleton. 2001. Cultural and genetic evolution in mountain white-crowned sparrows: Song dialects are associated with population structure. Evolution 55: 2568–2575.
Ogden, R., N. Dawnay & R. McEwing. 2009. Wildlife DNA forensics—bridging the gap between conservation genetics and law enforcement. Endanger Species Res 9. doi:10.3354/esr00144.
Presti, F. T., N. M. Guedes, P. T. Antas & C. Y. Miyaki. 2015. Population Genetic Structure in Hyacinth Macaws (Anodorhynchus hyacinthinus) and Identification of the Probable Origin of Confiscated Individuals. Journal of Heredity 106: 491–502.
Saranathan, V., D. Hamilton, G. V. Powell, D. E. Kroodsma & R. O. Prum. 2007. Genetic evidence supports song learning in the three-wattled bellbird Procnias tricarunculata (Cotingidae). Molecular Ecology 16: 3689–3702.
Schwartz, T. S. & S. A. Karl 2008. Population genetic assignment of confiscated gopher tortoises. The Journal of Wildlife Management 72: 254–259.
Wright, T. F., A. M. Rodriguez & R. C. Fleischer. 2005. Vocal dialects, sex-biased dispersal, and microsatellite population structure in the parrot Amazona auropalliata. Molecular Ecology 14: 1197–1205.





























